Nella prima parte di questa disamina sono state passate in rassegna diverse informazioni necessarie per comprendere cosa sono le Incretine e quali sono le loro principali azioni. Si è poi passati a descrivere la classe di farmaci degli Incretino-Mimetici discorrendo sulla loro sintesi, tipologia molecolare [in ordine cronologico in base alla data di commercializzazione] e caratteristiche di azione sia per quanto concerne le attività positive che quelle negative. In conclusione, si è accennato alla consistente diffusione dell’uso “cosmetico” di tali farmaci.
In questa seconda ed ultima parte vedremo come questa relativamente nuova classe di farmaci abbia trovato un certo spazio di diffusione nel Fitness e (sebbene in minor parte) nel BodyBuilding. Verranno discussi i punti di attrattiva e le limitazioni (e rischi) legati al loro uso.
Le “attrattive” degli Incretino-Mimetici:
L’obesità è una grave epidemia che affligge la società del così detto “occidente americanizzato”. I farmaci iniettabili sottocutanei inizialmente concepiti per la gestione del diabete di tipo II, come la Semaglutide e altri agonisti del recettore del GLP-1, stanno rapidamente guadagnando popolarità per i loro effetti sulla perdita di peso. Questi farmaci (Ozempic, Wegovy, Saxenda e Mounjaro) sono onnipresenti sui social media e sono promossi da celebrità di tutte le fasce demografiche. “Viso da Ozempic” e ‘sedere da Ozempic’ sono ormai concetti mainstream che evidenziano i possibili cambiamenti morfologici che si verificano con questi farmaci. Con la diffusione non controllata da personale qualificato dell’uso di questi farmaci, è aumentato anche l’elenco dei potenziali effetti avversi.
Nella prima parte di questa disamina si è constatato che gli Incretino-Mimetici hanno potenzialità che si esprimono su modifiche della composizione corporea e che riguardano principalmente:
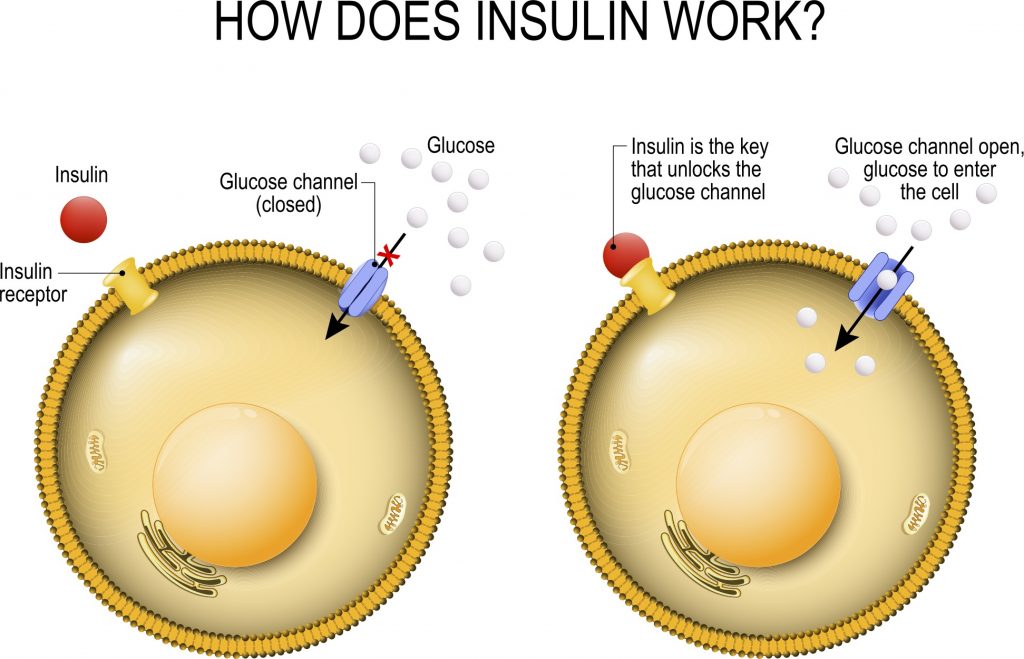
Rallentamento dello svuotamento gastrico con riduzione del senso di fame derivante dal aumento del senso di sazietà;
Gli effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare riducendo il senso di fame e il consumo di cibo;
Indirettamente, per via della stimolazione insulinica, aumento del senso di sazietà dato dal picco insulinico a livello ipotalamico;
Miglioramento del ripartizionamento Kcal attraverso il miglioramento della sensibilità all’insulina.
L'”attrattiva” che ha spinto (e che spinge) diverse persone di diverse classi sociali e di ambo i sessi a prendere in considerazione e concretizzare l’uso di questi farmaci è fondamentalmente ridotta alla riduzione della fame/appetito. Ma questo riguarda la persona “nella media”, la ragazza/donna alla ricerca di rapide soluzioni per l’imminente prova costume o per il servizio fotografico, sfilata ecc…
Nel Bodybuilding questa classe di farmaci ebbe una serie di attrattive comprendenti lo sfruttamento di tutti i punti sopra elencati. E, di conseguenza, si ipotizzarono fasi della preparazione nelle quali applicare tale categoria farmaceutica. Limitandone l’uso in “Cut” per ragioni legate al rischio (seppur limitato in monoterapia) di incorrere in eventi ipoglicemici, l'”attrattiva d’uso” riguardava il potenziale in fasi di “Recomp”; quindi non propriamente “ipocalorico” o, al massimo, leggermente ipocaloriche e non ipoglucidiche.
Ricomposizione corporea, Ripartizionamento calorico e Insulino-Sensibilità:
Ma chiariamo alcuni termini per schiarirci le idee…
Ricomposizione corporea: si intende il raggiungimento di un obiettivo o un risultato desiderato di un regime alimentare e di allenamento (e di farmaci), come il “Cut” o il “Bulk”.
“Cut”: si intende, propriamente, una diminuzione della massa grassa (FM) e il mantenimento della massa muscolare (LBM).
“Bulk”: si intende un aumento della LBM con una concomitante attenuazione dell’aumento della FM.
“Recomp” [comunemente intesa]: è definita come un aumento della LBM e la diminuzione della FM.
La suddivisione terminologica corretta è un concetto funzionale all’obiettivo di tutti gli interventi dietetici razionali.
Il rapporto p (rapporto di ripartizione) descrive le proteine depositate nei tessuti della LBM in relazione all’apporto energetico e, viceversa, le proteine perse dai tessuti della LBM in relazione al deficit energetico.
Il p-ratio comprende i fattori di:
stato ormonale (cioè i livelli assoluti di ormoni chiave noti);
sensibilità all’insulina;
sensibilità alla Leptina.
Esiste un’interazione tra il punto 1 e 3.
La prima chiave è…
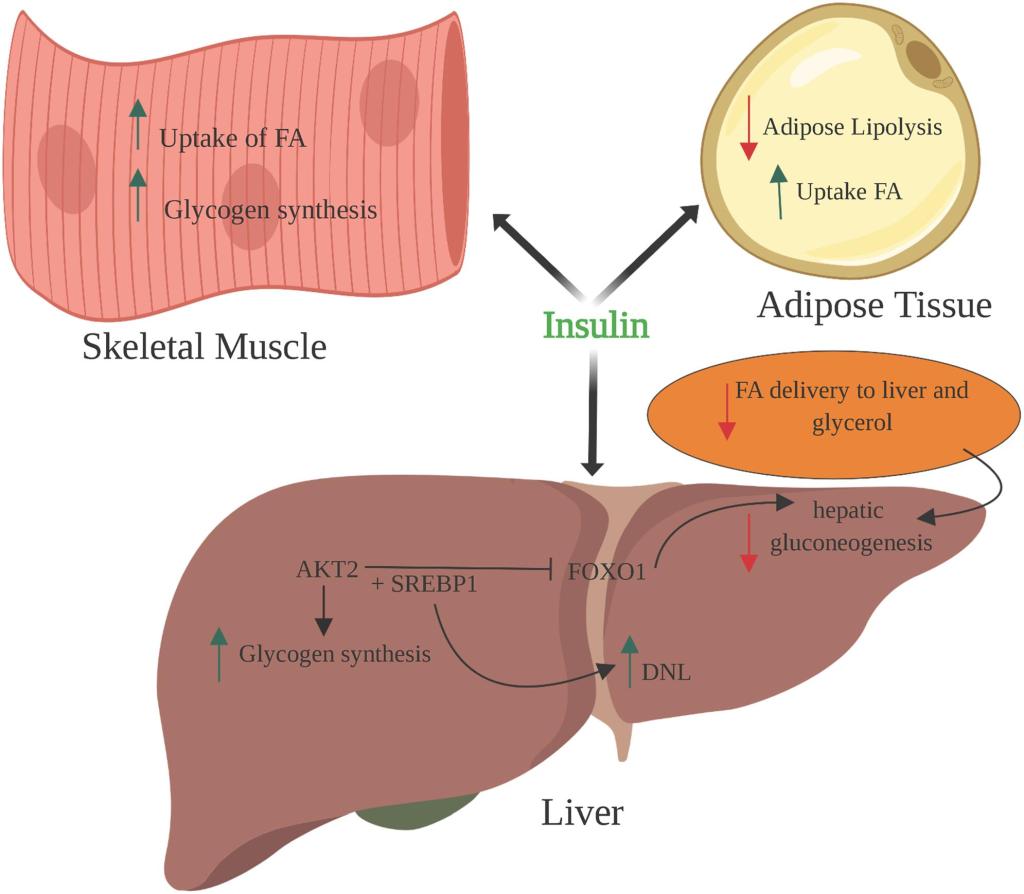
Sensibilità all’Insulina: quando si è a dieta (cioè in uno stato di deficit energetico), la resistenza all’Insulina fisiologica è una condizione favorevole all’uso del grasso di deposito limitando l’uso del glucosio da parte del muscolo come substrato energetico, risparmiando il glucosio per il cervello e l’utilizzo degli Acidi Grassi intramuscolare. In condizioni di uscita da un regime ipocalorico, in uno stato di migliorata insulino-sensibilità, l’aumento calorico di una fase di “Bulk” vede, almeno inizialmente, un ripartizionamento delle Kcal al miocita a discapito del adipocita; tale condizione di inverte col protrarsi del regime ipercalorico.
I fattori che influenzano la sensibilità all’insulina includono [1]:
Livelli di grasso corporeo; B.F. % (predittore primario): ↑B.F. ⇒ ↑Acidi Grassi come substrato energetico (risparmiando glucosio e proteine [che possono essere utilizzate dal fegato nella gluconeogenesi]) e detta la segnalazione delle adipochine (cioè gli ormoni secernenti gli adipociti [leptina, TNF-α, IL-…, adiponectina, etc.]) ⇒ ↓ sensibilità all’insulina;
Contrazione muscolare (cioè attività, come ad esempio locomozione, allenamento contro-resistenza) ⇒ ↑ assorbimento di glucosio nella cellula muscolare; traslocazione di GLUT-4 ⇒ ↑ sensibilità all’Insulina;
Dieta: elevata quantità di carboidrati (in ipercalorica), grassi saturi e poche fibre ⇒ ↓ sensibilità all’Insulina;
Stoccaggio del Glicogeno o Supercompensazione [successivo ad una deplezione] ⇒ ↑assorbimento di glucosio e glicogenesi ⇒↑sensibilità Insulinica;
Deplezione di glicogeno (ad esempio, nel periodo successivo a un allenamento intenso, prima di un’alimentazione particolarmente ricca di carboidrati) ⇒ sottoregolazione (deplezione pressoché totale) della disponibilità di glucosio e promozione dell’ossidazione di Acidi Grassi dopo il depauperamento delle scorte di glicogeno muscolare (in media < 700 g negli adulti) ⇒ ↑ Acidi Grassi liberi nel sangue (circolanti) ⇒ ↓ sensibilità all’Insulina;
Fattori genetici in parte modificabili dai farmaci, ad esempio, nei casi di ipogonadismo, l’applicazione TRT inverte chiaramente l’insulino-resistenza nei casi in cui l’eziologia dell’insulino-resistenza è riconducibile alla carenza di Testosterone.
La seconda chiave è…
Struttura molecolare della Leptina
La Leptina: come sappiamo bene, la Leptina è un ormone, più precisamente è una adipochina, secreta principalmente dagli adipociti, che si correla con la %B.F.;↑%B.F. ⇒ ↑Leptina. (i depositi viscerali e quelli sottocutanei hanno rapporti diversi con la Leptina). A una data percentuale di B.F., le donne producono ~2 – 3 volte più Leptina rispetto agli uomini. Le concentrazioni di Leptina cambiano con la restrizione energetica e la sovralimentazione. La Leptina è un segnalatore primario di regolazione dell’accumulo di energia che riflette:
la percentuale di B.F.;
l’assunzione di energia.
Esempio 1: all’inizio di una dieta ipocalorica, la Leptina può diminuire del 50% entro 1 settimana (o meno) – anche se ovviamente il soggetto a dieta non ha perso il 50% di B.F. – quindi, in un primo momento, i cambiamenti della concentrazione di Leptina non sono correlati alla B.F. (piuttosto segnalano l’assunzione di energia).
Dopo il calo iniziale, si assiste a un declino più graduale della Leptina in relazione alla perdita di %B.F. .
Esempio 2: in caso di sovralimentazione, la Leptina subisce un incremento in modo altrettanto rapido (cioè senza relazione con la %B.F., ma in relazione all’assunzione di energia).
A breve termine, la secrezione di Leptina è determinata principalmente dalla disponibilità di glucosio, per cui la riduzione della disponibilità di glucosio nella cellula adiposa (dieta ipocalorica) ⇒ ↓Leptina e viceversa.
Gli effetti specifici dell’ormone Leptina includono effetti sul pancreas e sul fegato, nel muscolo scheletrico ↑FA e ↓AA e l’uso del glucosio come substrato energetico (aumentando la perdita di grasso, promuovendo il risparmio di proteine)… [1]
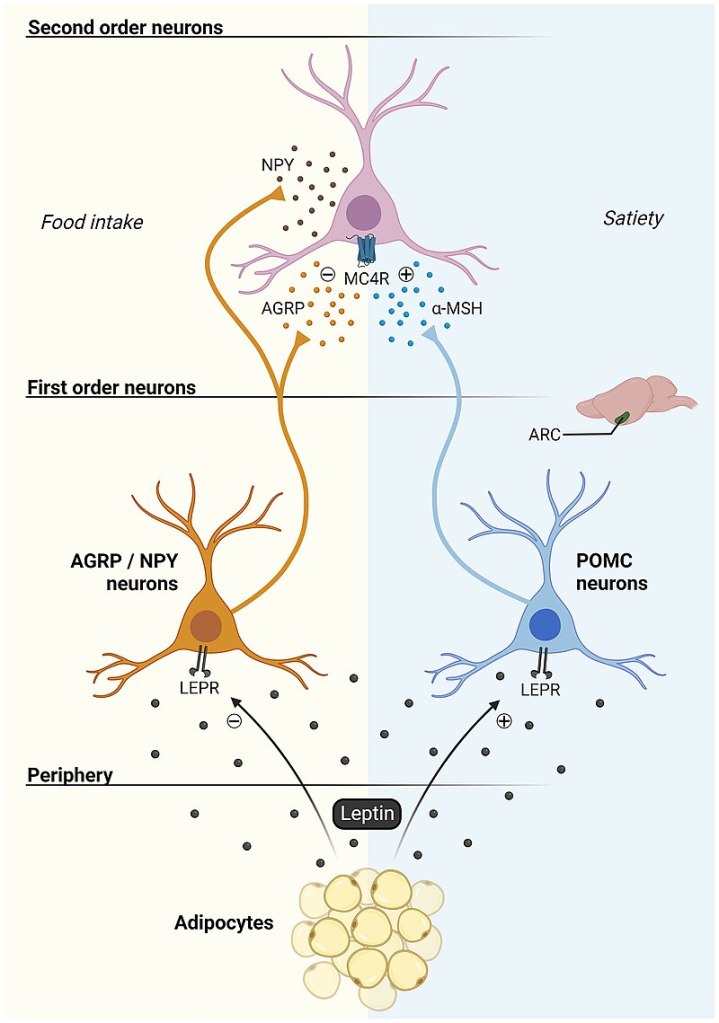
Modello classico Leptina-Melanocortina
In sostanza, il partizionamento (p-ratio) è un concetto che associa la Leptina e la sensibilità all’Insulina come fattori principali che determinano il modo in cui le variazioni dell’apporto calorico e del contenuto di macronutrienti influiscono sul metabolismo (influenzando profondamente la composizione corporea) e sullo stato ormonale. Possiamo modificarlo e migliorarlo, tenendo conto dei tessuti bersaglio e del nostro obiettivo (ad esempio, se Bulk o Cut).
Inoltre, non bisogna confondere il potenziamento dell’insulino-resistenza fisiologica sulla perdita di grasso con l’erronea valutazione che l’insulino-resistenza sia salutare. L’insulino-resistenza, soprattutto in una persona sedentaria, è associata alla sindrome metabolica, al diabete di tipo II, per non parlare del grasso viscerale, ecc.
L’insulino-resistenza è uno stato in cui i tessuti dell’organismo (ad esempio, fegato, pancreas, muscolo scheletrico) presentano una scarsa recettività con l’Insulina continuando, se si parla in particolare del fegato, a produrre glucosio in quantità inappropriate. Questo stato di iperglicemia è un effetto piuttosto che la causa dell’insulino-resistenza, anche se i livelli tossici di glucosio degradano la reattività delle isole pancreatiche all’Insulina rappresentando così una delle vie/meccanismi dell’insulino-resistenza, peggiorando la stessa condizione.
Ma tutto questo cosa centra con gli Incretino-Mimetici? Se non ci siete ancora arrivati, calma e capirete …
Agonisti del recettore GLP-1 e GIP:
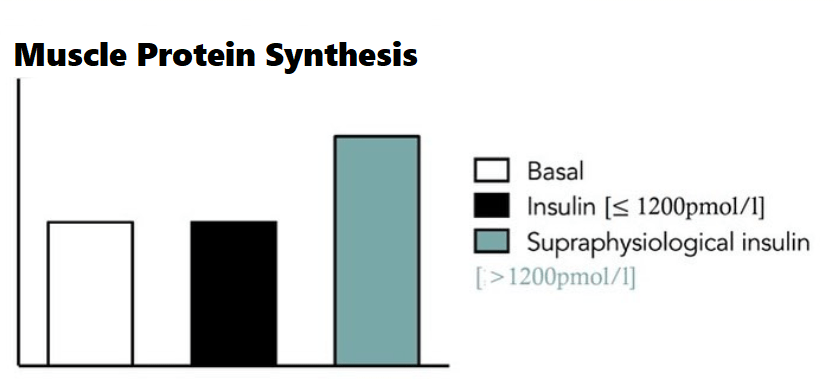
La perdita di grasso si verifica con gli agonisti del GLP-1 e della GIP (Incretino-mimetici) – come la Semaglutide e la Tirzepatide – che sono veri e propri agenti sensibilizzanti dell’Insulina. Tuttavia, non è la sensibilità all’Insulina di per sé che è responsabile della perdita di grasso con questa classe di farmaci – ma piuttosto, come abbiamo già visto, la perdita di grasso avviene grazie agli altri effetti di questi farmaci, come l’alterazione potenziale delle preferenze alimentari, il ritardo dello svuotamento gastrico, il senso di sazietà, che promuovono il controllo dell’appetito e riducono l’assunzione di energia.
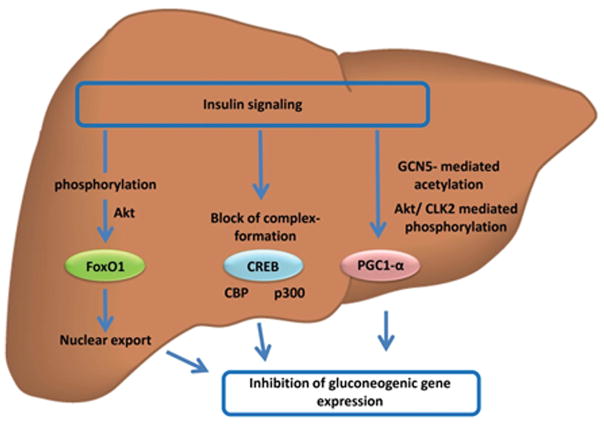
Sappiamo che gli agonisti del GLP-1 e del GIP migliorano direttamente la sensibilità all’insulina modulando la secrezione di Insulina – accoppiandola alla presenza di elevate concentrazioni di glucosio. Questa secrezione di Insulina si attenua quando le concentrazioni di glucosio nel sangue diminuiscono e si avvicinano all’euglicemia. Inoltre, anche se indirettamente, riducendo l’assunzione di cibo, questi farmaci determinano una riduzione della %B.F.. La riduzione della percentuale di B.F. dovuta alla riduzione dell’assunzione di cibo riduce le riserve di massa grassa (e quindi gli FFA circolanti), migliorando ulteriormente la sensibilità all’Insulina.
La stragrande maggioranza degli agenti per la perdita di grasso, in quanto agenti lipolitici, favoriscono l’insulino-resistenza. Ad esempio i β-agonisti, non selettivi come l’Efedrina o selettivi come il Clenbuterolo, oppure lo stimolante da banco per eccellenza la caffeina, agendo in modo analogo o aumentano l’azione delle catecolamine (epinefrina e noradrenalina, o adrenalina e noradrelanina) possono portare ad un peggioramento di questa condizione.
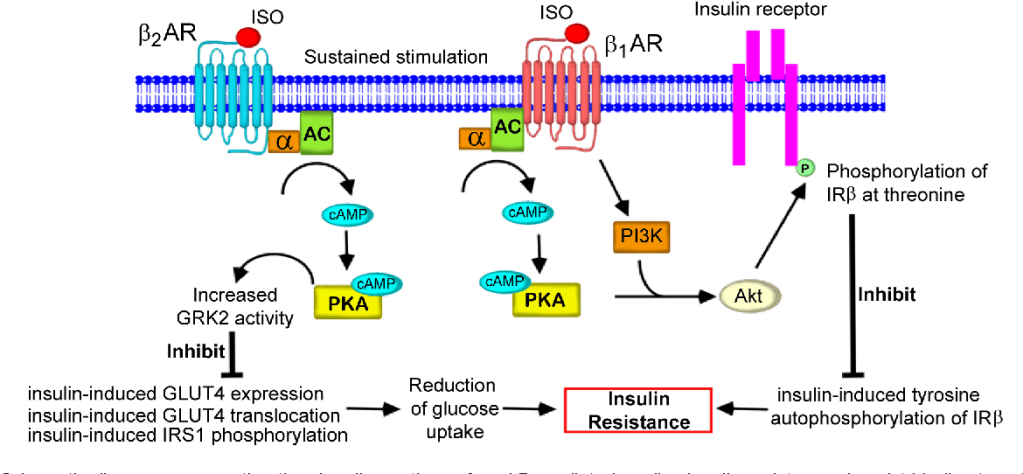
Diagramma schematico che rappresenta la via di segnalazione dell’insulino-resistenza cardiaca mediata dai β-AR. Tale meccanismo interessa (tra gli altri tessuti) anche il muscolo-scheletrico.
Quando tessuti come il fegato e le cellule adipose vedono ridotta l’interazione con l’Insulina, il glucosio non viene ottimamente assorbito dalle cellule. Con un marcato calo del glucosio, il fegato inizia a metabolizzare gli acidi grassi liberi (FFA), aumentando così i livelli di chetoni nel sangue e impedendo che vengano riesterificati nelle cellule adipose (in ipocalorica). Nel fegato e nelle cellule adipose, senza che l’Insulina interagisca ottimamente con questi tessuti, si verifica una soppressione della sintesi/lipogenesi dei grassi (negli adipociti) e della sintesi di lipoproteine a bassissima densità (VLDL) (nel fegato).
Incretino-mimetici, miglioramento della sensibilità all’Insulina e preservazione della massa muscolo-scheletrica:
La classificazione degli incretino-mimetici come agenti di ripartizione calorica:
Nel BodyBuilding, il fascino verso questa classe di farmaci si concentra anche sul funzionamento degli agonisti del GLP-1 e della GIP sull’insulino-resistenza, poiché quest’ultima durante la restrizione calorica nel muscolo scheletrico (>60% del peso corporeo, più nei bodybuilder) è un’immagine non proprio esaltante, con le riserve di glicogeno che vengono prima catabolizzate abbastanza rapidamente; poi i trigliceridi intramuscolari (che rappresentano solo l’1% del peso del muscolo idratato, fino al 2% del volume, dato che il grasso è meno denso del muscolo scheletrico, e ~1/3 dell’energia muscolare, dato che il grasso è energeticamente denso) e infine, se necessario, l’organismo utilizzerà gli AA (catabolizzando le proteine muscolari; proteolisi) per ottenere l’energia necessaria a svolgere le attività giornaliere. Questi agenti, quindi, nella misura in cui sono sensibilizzanti per l’Insulina, dovrebbero servire a promuovere il mantenimento della LBM durante il “Cut”.
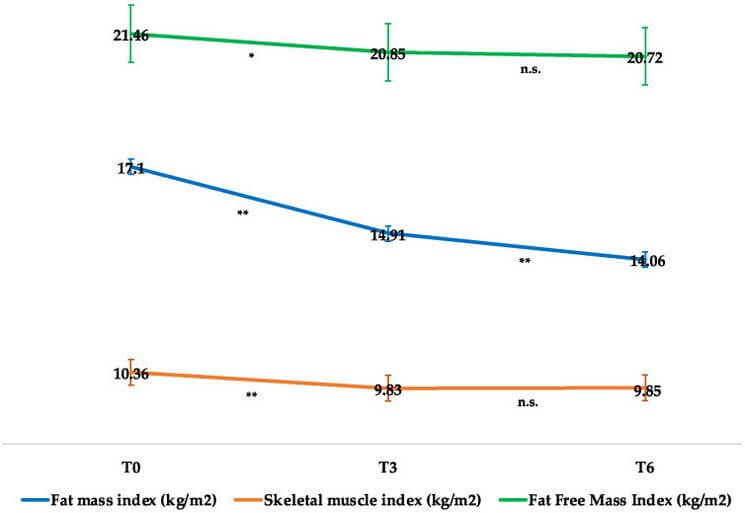
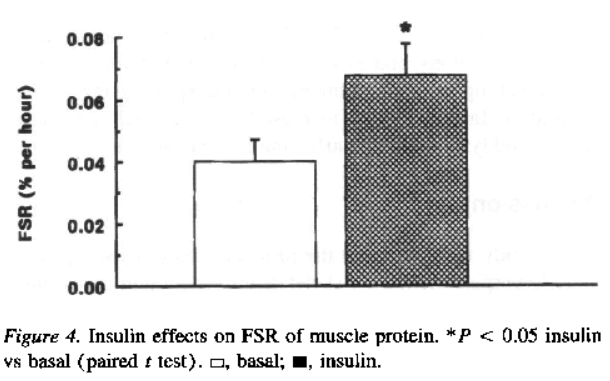
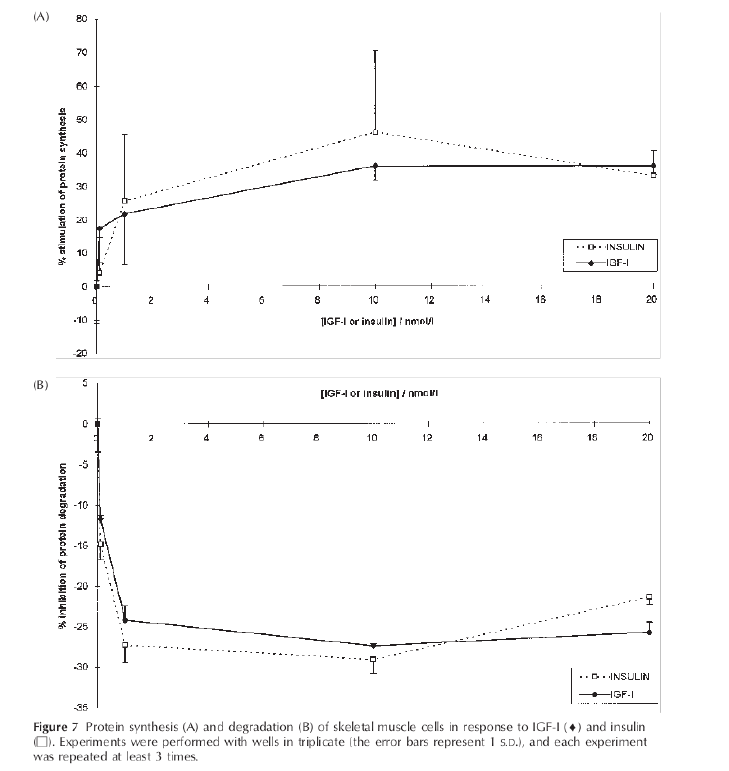
In effetti, come si evince dalla seguente immagine tratta da un articolo di ricerca di Volpe et. al del 2022 [2], la Semaglutide preserva in modo abbastanza efficace la LBM e riduce in modo preferenziale la FM, con riduzioni solo clinicamente insignificanti dell’indice di massa magra (FFMI, kg/m²) e dell’indice della muscolatura scheletrica durante il periodo iniziale di adattamento, che poi si attenua:
In un certo senso, quindi, migliorando il grado di sensibilità all’Insulina dell’equazione della p-ratio, gli incretino-mimetici possono essere classificati come agenti di ripartizione calorica, a grandi linee come il Clenbuterolo, ma invece di promuovere l’insulino-resistenza come i β-agonisti, la migliorano.
* Ovviamente, creare un ambiente significativamente insulino-sensibile in un contesto ipocalorico (soprattutto se ipoglucidico) può mettere l’utilizzatore a maggior rischio (sebbene limitato) di ipoglicemia o calo glicemico borderline con effetti simili allo stato di ipoglicemia (tremore, sudorazione copiosa ecc…).
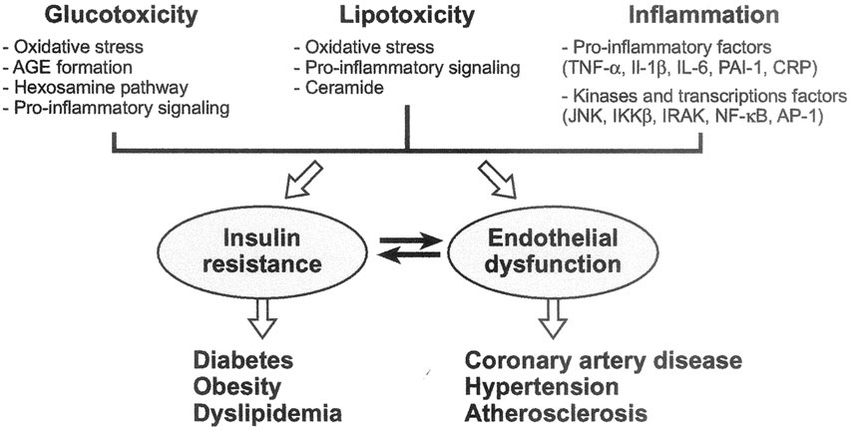
Per coloro che hanno familiarità con questi concetti, derivanti dalle discussioni sul bodybuilding, può risultare molto confuso il fatto che l’iperglicemia (elevata quantità di glucosio nel sangue) è solo uno dei fattori associati all’insulino-resistenza, ma in realtà non è sinonimo di insulino-resistenza (iperglicemia ≠ insulino-resistenza). Sì, ridurre la glicemia a livelli normali è molto importante per migliorare la sensibilità all’insulina durante l’uso dell’ormone della crescita esogeno (rhGH), perché il glucosio è tossico per le cellule β pancreatiche. Questa glucotossicità a livello delle cellule pancreatiche si traduce in una diminuzione della risposta secretoria dell’insulina all’iperglicemia, alimentando così il fuoco dell’iperglicemia e della glucotossicità, contribuendo all’insulino-resistenza – ma non costituendone l’unica eziologia.
La sensibilità all’Insulina è multifattoriale e comprende componenti sistemiche (ad esempio, QUICKI) e periferiche (ad esempio, GLUT-4) ed è regolata a livello centrale da GLP-1 e GIP. L’iperglicemia è solo uno dei fattori (l’altro è l’Insulina) che funge da proxy dell’insulino-resistenza sistemica. Vi sono altri aspetti, tra cui la tolleranza ai carboidrati [vedi anche capacità di tolleranza del metabolismo glucidico], ecc.
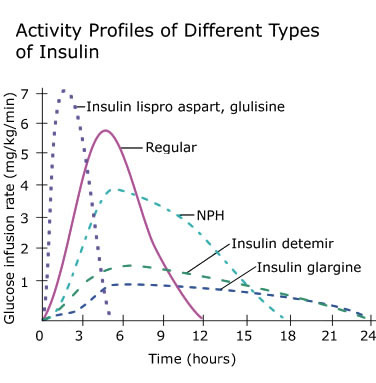
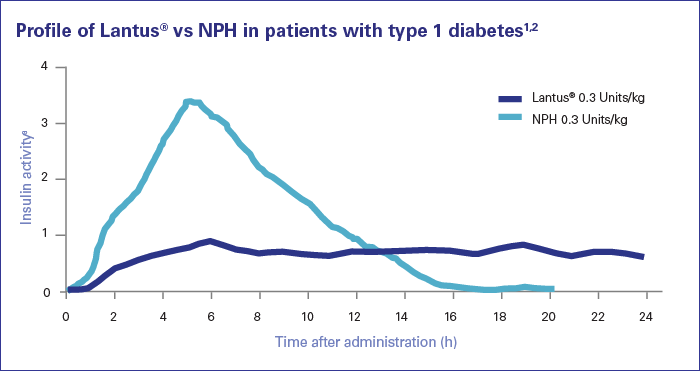
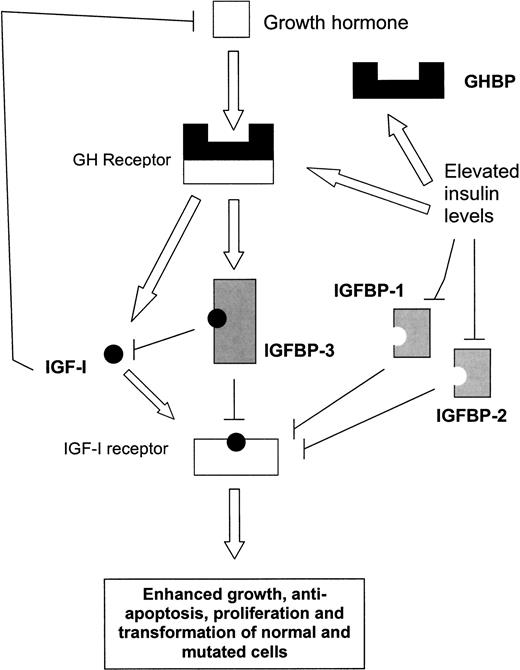
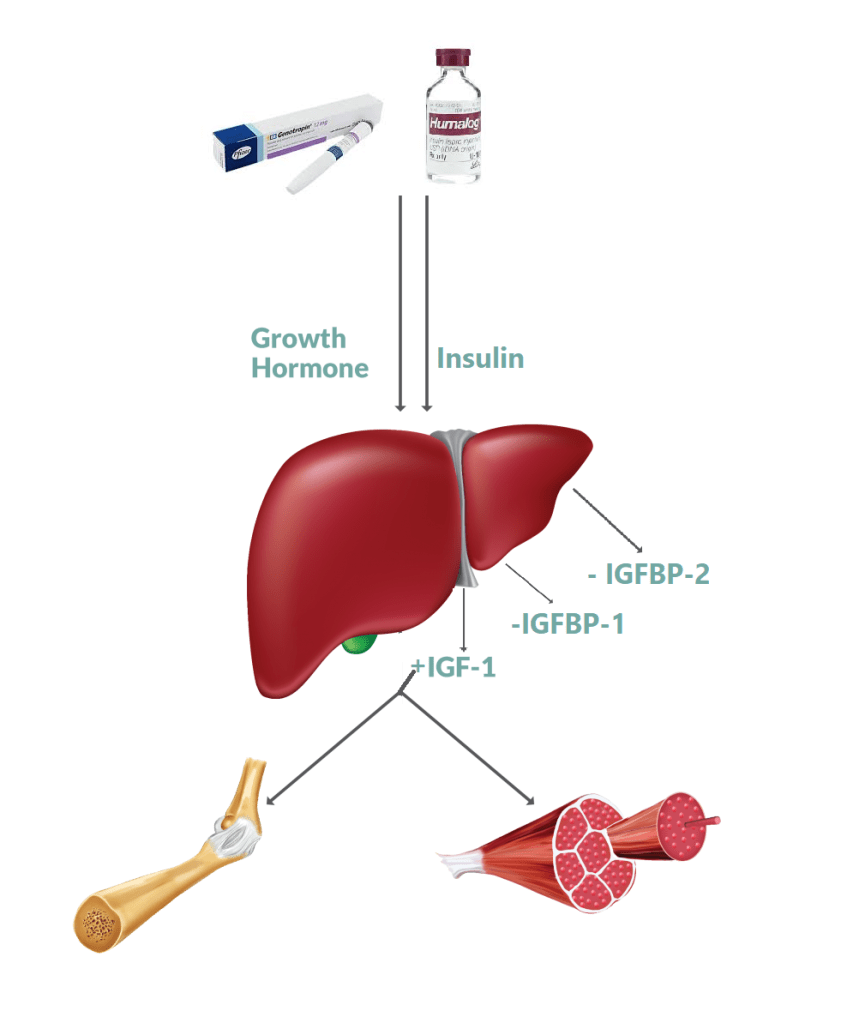
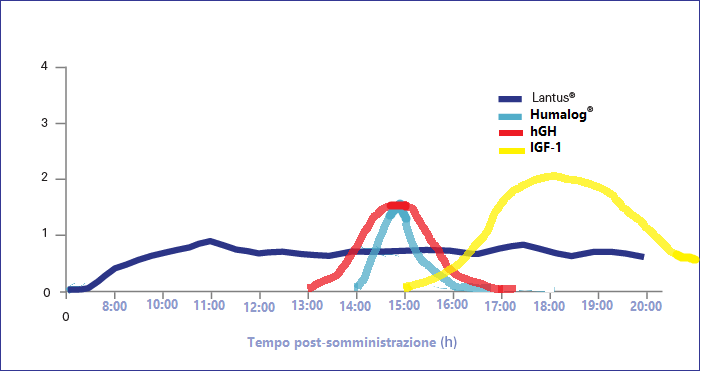
In tema di insulino-resistenza, è utile ricordare che l’Insulina endogena viene secreta in modo pulsatile per regolare il metabolismo glucidico e lipidico, la crescita cellulare ecc…, a differenza del Testosterone che viene secreto in modo più stabile (rilascio graduale nel sangue, ma soggetto a variazioni diurne, ad esempio una maggiore secrezione al mattino rispetto a mezzogiorno/sera). Gli aumenti cronici di Insulina, ad esempio quelli relativi al profilo di rilascio di una bassa dose giornaliera di Insulina Glargine (Lantus), presentano un’area sotto la curva (AUC) relativamente ampia a causa del profilo di rilascio (concentrazioni elevate per lunghi periodi di tempo) rispetto ai normali profili di rilascio dell’Insulina endogena (paragonabili alla farmacocinetica dell’Insulina regolare, ad esempio Actrapid, Novolin o HumuLin -R). L’elevata AUC di Lantus e/o le dosi di Insulina regolare esogena moderatamente elevate e frequenti sono descritte come iperinsulinemia cronica.
Questa resistenza non avviene per feedback negativo a livello delle cellule β.
Al contrario, l’iperinsulinemia cronica che causa l’insulino-resistenza è multifattoriale e comprende:
L’aumento dell’HOMA-IR e la diminuzione del QUICKI (misure biochimiche dell’insulino-resistenza e della sensibilità all’Insulina, rispettivamente).
L’alterata trasduzione del segnale insulinico dovuta alla disfunzione del recettore dell’Insulina (IR) e alla diminuzione dell’autofosforilazione dell’IR, che blocca la traslocazione del GLUT-4 sulla superficie cellulare nelle cellule muscolari e adipose, con conseguente aumento del glucosio nel sangue [3]:
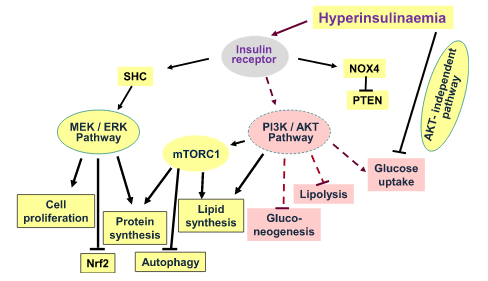
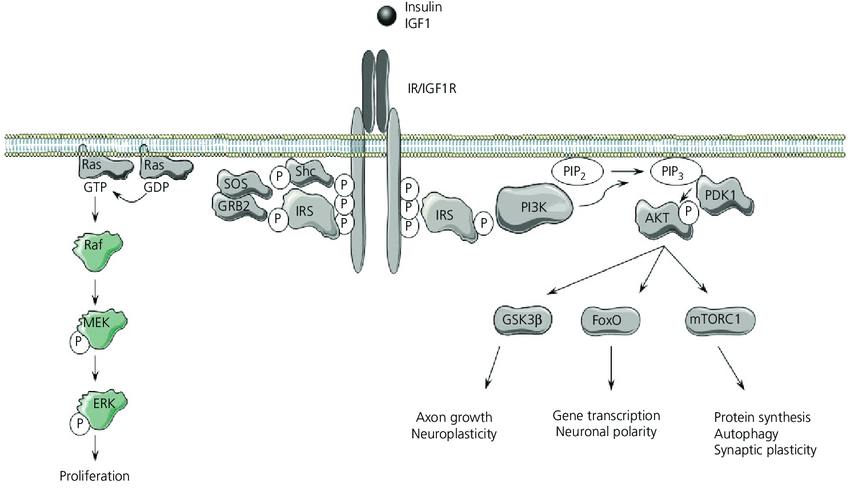
Segnalazione dell’Insulina durante l’insulino-resistenza. Durante l’insulino-resistenza, la segnalazione attraverso le chinasi AKT è parzialmente compromessa. Non tutte le vie AKT-dipendenti sono interessate, così come altre vie di segnalazione, indicando che l’insulino-resistenza è selettiva. Pertanto, l’iperinsulinemia, in presenza di insulino-resistenza, promuove le attività anaboliche delle cellule attraverso la via MEK-ERK e attraverso mTORC1. Sebbene la via PIK/AKT sia compromessa durante l’insulino-resistenza e fornisca solo una traslocazione insufficiente di GLUT4 per l’assorbimento del glucosio e un’attivazione carente di eNOS, sembra esserci un’attivazione normale di mTORC1. Oltre alle conseguenze anaboliche della segnalazione attraverso la via MEK/ERK descritte nella figura, si osserva un’aumentata espressione di ET-1 e PAt-1 (non mostrato), nonché l’inibizione dell’autofagia e del fattore nucleare Nrf2, che compromettono rispettivamente il turnover dei costituenti cellulari e i meccanismi di difesa delle cellule dallo stress radicale. L’iperinsulinemia riduce l’assorbimento del glucosio non solo attraverso lo smorzamento della via PIK/AKT (resistenza all’Insulina), ma anche attraverso altre vie ancora sconosciute.
3. Aumento dei livelli e dell’attività di sn-1,2-diacilglicerolo (DAG) grazie alla sintesi de novo.
Le limitazioni degli Incretino-Mimetici dietro all’iniziale entusiasmo:
Se ci dovessimo basare su quanto esposto fino a questo punto, saremo tutti d’accordo nell’ammantare della nomea di “farmaci prodigiosi per la ricomposizione corporea” tanto la Semaglutide quanto il Tirzepatide e tutti gli altri membri di questa classe. Ma, dal momento che, la conoscenza per essere utile deve essere sufficientemente approfondita, occorre indagare meglio sulle caratteristiche di questi farmaci.
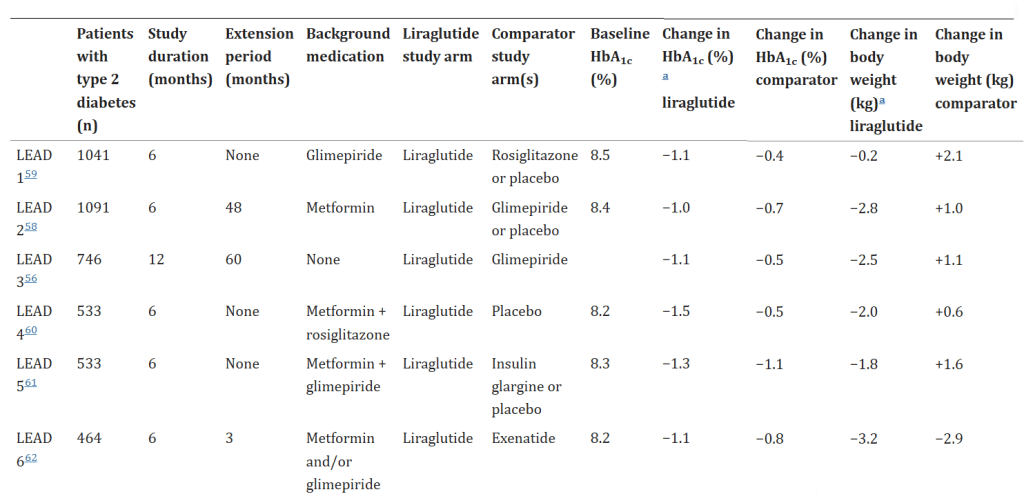
Sappiamo che la Semaglutide è effettivamente associata alla perdita di peso con una differenza media dell’11,85% rispetto al placebo emersa dalle ultime review. Il consolidamento degli studi ha mostrato che nausea, vomito, costipazione e diarrea sono gli eventi avversi più comuni. Nonostante questi effetti sembrano essere per lo più di gravità da lieve a moderata, la loro risoluzione totale era spesso connessa al termine del trattamento.
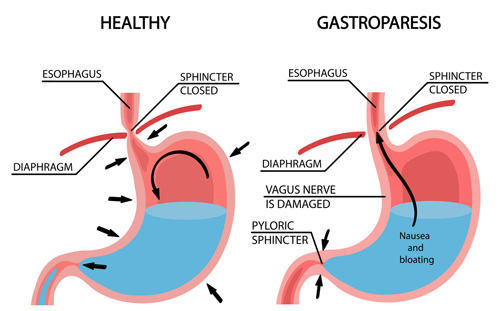
Dal momento che gli Incretino-mimetici causano una riduzione del senso di fame, in modo anche significativo (dose dipendente), rallentano lo svuotamento gastrico portando anche ad eventi diarroici preceduti da mal assorbimento dei nutrienti, senza sottovalutare il rischio di paresi gastrica, la facilità di perdere massa muscolare in un contesto ipocalorico è molto alta, specie se non supportata da agenti anabolizzanti; l’inserimento di questi ultimi, però, non “tampona” la possibile condizione di mal assorbimento e/o scarsa assunzione proteica/alimentare.
Inoltre, la paresi gastrica è una condizione che un bodybuilder sano di mente cerca di evitare a tutti i costi combattendoci già se sussiste abuso di Insulina e/o hGH.
Trattando la limitazione data da una potenziale eccessiva inappetenza, è giusto specificare un impatto singolare che le Incretine (e gli Incretino-Mimetici) hanno sulle preferenze alimentari. Sappiamo, infatti, che gli alimenti maggiormente palatabili sono tipicamente ricchi di grassi e/o zuccheri e tendono a essere preferiti a quelli a basso contenuto di grassi/zuccheri. L’entità di questa preferenza, tuttavia, può essere influenzata da peptidi intestinali quali la Grelina e il GLP-1. La Grelina e il GLP-1 sono influenzati in modo differenziato dal consumo di alimenti palatabili. Tralasciando la Grelina, le concentrazioni di GLP-1 a digiuno predicono negativamente l’assunzione di alimenti ricchi di zuccheri semplici in un paradigma di distributori automatici, che gli autori dello studio hanno interpretato come prova del fatto che il GLP-1 svolge un ruolo nelle vie di ricompensa che regolano l’assunzione di zuccheri semplici. Diversi studi riportano anche un’alterazione delle preferenze alimentari dopo l’intervento di bypass gastrico, con un allontanamento dalla preferenza per gli zuccheri/grassi elevati. L’assunzione di alimenti appetibili, in particolare di soluzioni zuccherate, è aumentata dalla Grelina, mentre il GLP-1 riduce preferenzialmente l’assunzione di alimenti ad alto contenuto di grassi e zuccheri, almeno dopo una somministrazione acuta. Inoltre, i lavori condotti sull’uomo rivelano che la preferenza per i grassi e gli zuccheri può essere alterata dalla chirurgia bariatrica e contribuire alla perdita di peso, ma non è ancora stato stabilito se questi effetti siano legati a un’alterazione del segnale della Grelina o del GLP-1. Infine, i livelli circolanti di Grelina e GLP-1 possono essere indicativi del consumo di cibo appetibile nell’uomo.[https://www.frontiersin.org/]
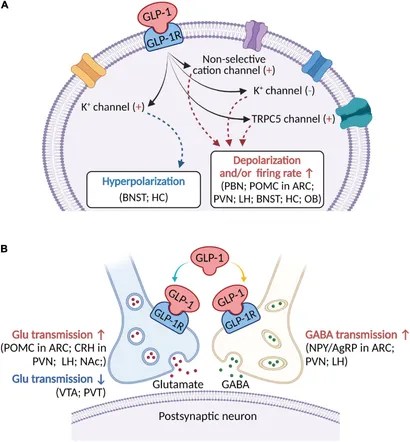
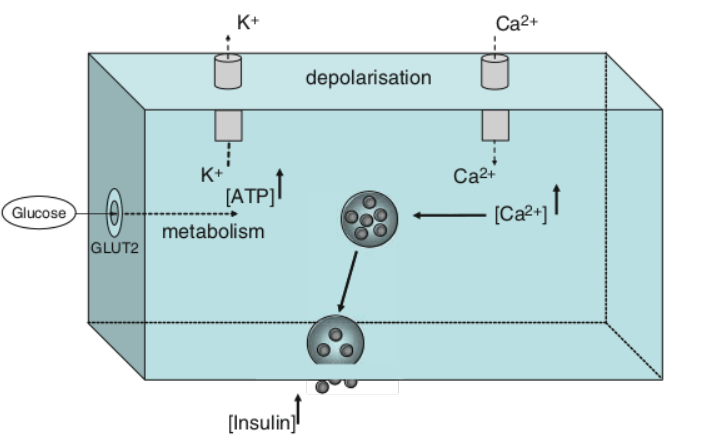
Schema che descrive i principali effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare. (A) Il GLP-1 (compresi i suoi agonisti) si lega al GLP-1R postsinaptico per depolarizzare il potenziale di membrana e/o aumentare la frequenza di sparo nella maggior parte delle regioni cerebrali, ma iperpolarizza il potenziale di membrana in alcune aree cerebrali. Diversi meccanismi ionici, tra cui il canale cationico non selettivo, il canale K+ e il canale TRPC5, possono essere coinvolti nell’attivazione della depolarizzazione o iperpolarizzazione indotta dal GLP-1R. (B) Oltre ai recettori postsinaptici, il GLP-1 agisce sui GLP-1R presinaptici per modulare la neurotrasmissione glutammatergica e GABAergica. ARC, nucleo arcuato; BNST, nucleo del letto della stria terminale; Glu, glutammato; CRH, ormone di rilascio della corticotropina; HC, ippocampo; LH, ipotalamo laterale; NAc, nucleo accumbens; NPY/AgRP, Neuropeptide Y/Peptide legato al gene Agouti; OB, bulbo olfattivo; PBN, nucleo parabrachiale; POMC, proopiomelanocortina; PVN, nucleo paraventricolare; PVT, nucleo talamico paraventricolare; VTA, area tegmentale ventrale.
Di conseguenza, nonostante gli indizi di cui sopra, l’uso di Incretino-Mimetici potrebbe ridurre marcatamente il consumo di cibo indipendentemente dalla fonte anche se, come abbiamo visto, l’effetto anoressizzante del GLP-1 sembra essere a maggior carico della componente alimentare glucidico-lipidica.
Tornando invece sulla questione legata al catabolismo muscolo-scheletrico e l’uso concomitante di agenti anabolizzanti, per ovviare a questo problema nei pazienti trattati con Semaglutide, è stato avviato uno studio clinico di fase 2b, multicentrico, in doppio cieco, controllato con placebo, randomizzato, per la determinazione della dose e per valutare la sicurezza e l’efficacia di Ostarina 3mg, Ostarina 6mg o placebo come trattamento per preservare la massa muscolare e aumentare la perdita di grasso in circa 150 pazienti con obesità sarcopenica o pazienti anziani in sovrappeso (>60 anni di età) trattati con Semaglutide (Wegovy®). L’endpoint primario è la massa corporea magra totale e gli endpoint secondari chiave sono la massa grassa corporea totale e la funzione fisica misurata dal test di salita delle scale a 16 settimane.
Dopo aver completato la parte di determinazione della dose di efficacia dello studio clinico di Fase 2b, si prevede che i partecipanti continueranno in cieco in uno studio clinico di estensione di Fase 2b in cui tutti i pazienti smetteranno di ricevere un GLP-1 RA, ma continueranno ad assumere un placebo, Ostarina 3mg o Ostarina 6mg per ulteriori 12 settimane. Lo studio clinico di estensione di fase 2b valuterà se l’Ostarina è in grado di mantenere la massa muscolare e prevenire l’aumento di grasso e peso che si verifica dopo l’interruzione di un GLP-1 RA.
Lo studio clinico è condotto in 14 centri clinici negli Stati Uniti. È stato raggiunto l’arruolamento completo dei circa 150 pazienti nello studio di fase 2b QUALITY. L’azienda prevede ora che l’ultimo paziente a completare lo studio di fase 2b QUALITY sarà nel dicembre 2024, con i risultati clinici di prima linea per lo studio clinico di fase 2b QUALITY attesi nel gennaio 2025. Inoltre, i risultati principali per lo studio clinico di estensione di Fase 2b in cieco separato possono ora essere attesi nel secondo trimestre solare del 2025.
Ovviamente, queste limitazioni, incisive nel Culturismo, interessano tutti gli Incretino-Mimetici, compresa la Tirzepatide la quale sembrerebbe avere un maggior margine di efficacia e “sicurezza” per quanto riguarda il rischio (seppur limitato in monoterapia) di eventi ipoglicemici.
Il motivo di questa riduzione di rischio ipoglicemico è dovuta alla doppia affinità recettoriale della Tirzepatide la quale, come abbiamo visto, possiede una attività agonista per i recettori del GLP-1 e del GIP. Ed è proprio il legame e l’attivazione di quest’ultimo recettore (GIP) da parte della Tirzepatide a permettere ciò. Infatti, l’attivazione del recettore GIP stimola la secrezione di Glucagone in modo glucosio-dipendente nelle persone sane, con un’attività maggiore in presenza di glicemie più basse. Ciò significa che, raggiunta la soglia euglicemica i livelli di glucosio nel sangue verranno mantenuti più facilmente all’interno di questa soglia per via dell’attività del Glucagone secreto dalle cellule α delle isole di Langerhans.[https://pubmed.ncbi.nlm.nih.gov/]
Sebbene i trials clinici suggeriscano che la Tirzepatide riduca la glucagonemia, un recente studio dimostra che la Tirzepatide è un potente stimolatore della secrezione di Glucagone nelle condizioni sopradette.
Quindi la Tirzepatide è superiore alla Semaglutide?
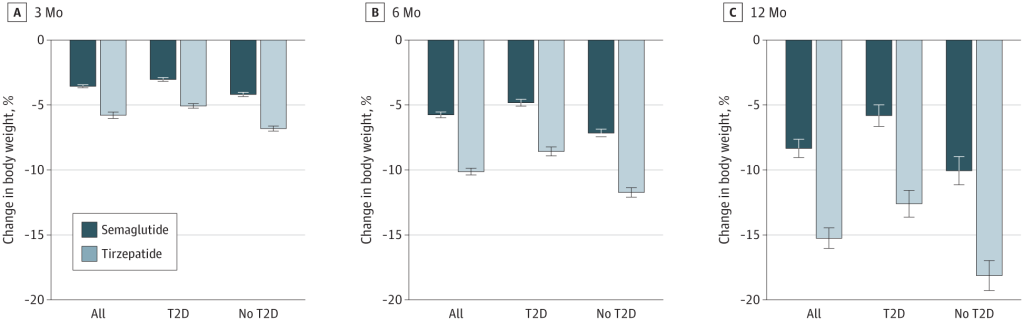
Alcuni studi recenti hanno messo a confronto la Semaglutide e la Tirzepatide per la perdita di peso. Studi di ricerca del 2021, del 2023 e del 2024 suggeriscono che la Tirzepatide può determinare una maggiore perdita di peso rispetto alla Semaglutide.
Variazione percentuale media del peso corporeo a 3, 6 e 12 mesi di trattamento per la popolazione complessiva, i soggetti con diabete di tipo II (TD2) e quelli senza TD2. Le barre rappresentano le variazioni medie del peso corporeo dal basale al punto di riferimento tra la popolazione di pazienti ancora in trattamento, abbinata in base al punteggio di propensione. Barre scure (Semaglutide); Barre chiare (Tirzepatide).
Ma questi studi presentano alcuni importanti limiti.
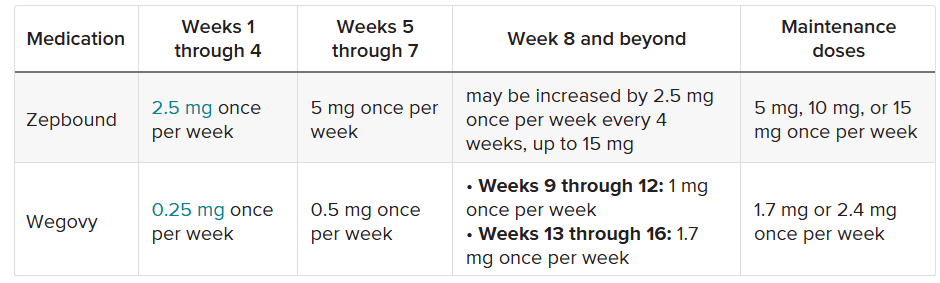
Le dosi di Semaglutide e Tirzepatide somministrate ai partecipanti non erano, per ovvie ragioni, uguali. La Semaglutide e Tirzepatide funzionano in modo leggermente diverso, come ormai sappiamo, e quindi i ricercatori hanno scelto livelli di dose comparabili. Tuttavia, la dose di Tirzepatide era più alta, il che potrebbe aver, anche di poco, influenzato i risultati.
Tabella comparativa tra il dosaggio della Semaglutide [Wegovy] e quella della Tirzepatide [Zepbound] usato negli studi.
Sappiamo altresì che la Tirzepatide ha una maggiore affinità con i recettori GIP rispetto ai recettori GLP-1. Di conseguenza, la ratio della dose di Tirzepatide con quella di Semaglutide risulta maggiormente a carico della prima.
La Tirzepatide è attualmente approvata dalla FDA per l’uso in persone in sovrappeso o con obesità, indipendentemente dal fatto che soffrano o meno di diabete di tipo II. Tuttavia, alcuni studi suggeriscono che la Tirzepatide sia un farmaco che non ha bisogno di essere somministrato in caso di mancanza della condizione diabetica.
Negli studi dove la Tirzepatide è stata somministrata a soggetti obesi, sono comunque stati osservati miglioramenti in tutte le misure cardiometaboliche. Gli eventi avversi più comuni con la Tirzepatide sono i medesimi riscontrati con la Semaglutide o altri membri della stessa famiglia. Essi sono stati di tipo gastrointestinale e la maggior parte di questi sono stati di gravità lieve o moderata e si sono verificati principalmente durante l’aumento della dose. Gli eventi avversi hanno causato l’interruzione del trattamento nel 4,3%, 7,1%, 6,2% e 2,6% dei partecipanti che hanno ricevuto dosi di Tirzepatide da 5, 10 e 15mg e placebo, rispettivamente.
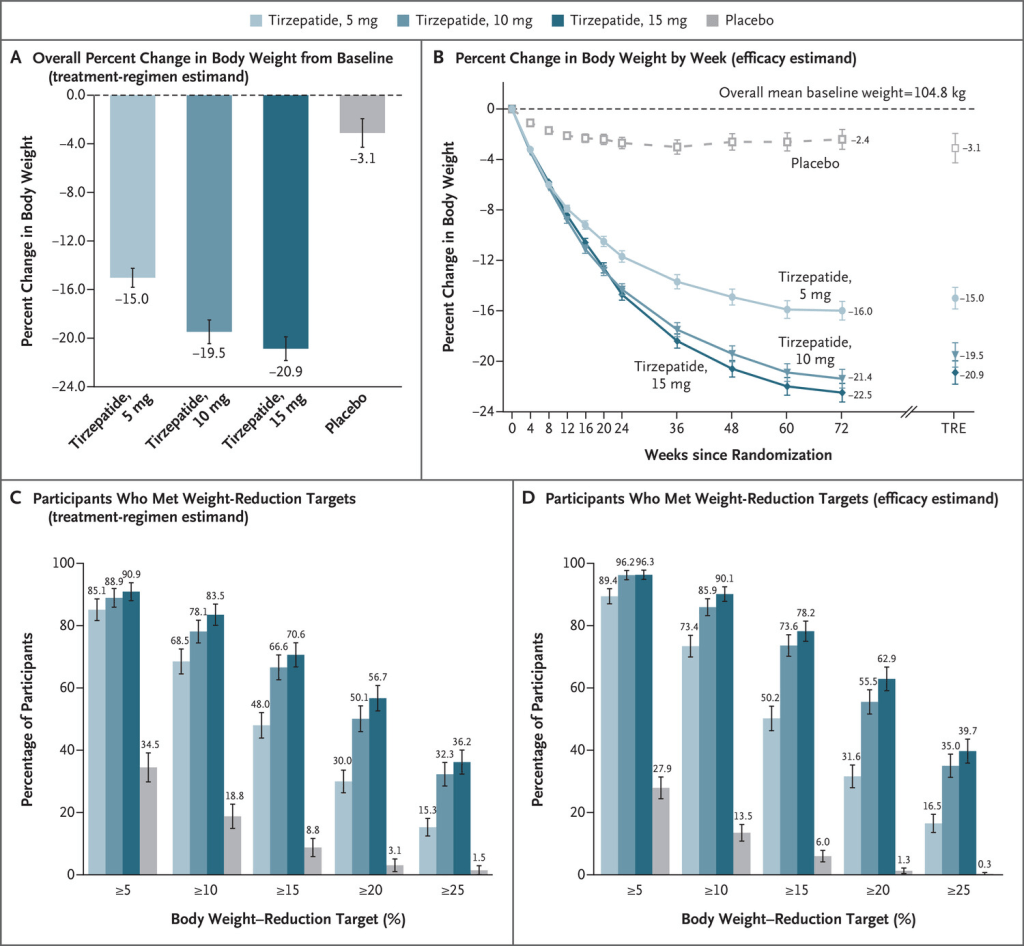
In uno studio di 72 settimane su partecipanti con obesità, 5mg, 10mg o 15mg di Tirzepatide una volta alla settimana hanno fornito riduzioni sostanziali e durature del peso corporeo.
Effetto della Tirzepatide somministrata una volta alla settimana, rispetto al placebo, sul peso corporeo. Le medie dei minimi quadrati sono presentate, se non diversamente specificato. Il pannello A mostra la variazione percentuale del peso corporeo dal basale alla settimana 72, derivata da un modello di analisi della covarianza per la stima del regime di trattamento (TRE). Il pannello B mostra la variazione percentuale del peso corporeo in base alle settimane dalla randomizzazione, derivata da un modello misto per misure ripetute (MMRM) per la stima dell’efficacia; sono riportate anche le stime alla settimana 72 per la stima del regime di trattamento. I riquadri C e D mostrano le percentuali di partecipanti che hanno avuto riduzioni di peso di almeno il 5%, 10%, 15%, 20% e 25% dal basale alla settimana 72. Per il riquadro C, la percentuale è stata calcolata in base al numero di settimane di randomizzazione. Per il pannello C, la percentuale è stata calcolata con l’uso delle regole di Rubin, combinando le percentuali dei partecipanti che hanno raggiunto l’obiettivo nei set di dati imputati. I valori mancanti alla settimana 72 sono stati imputati con MMRM se la mancanza era dovuta esclusivamente a Covid-19 e con imputazione multipla se la mancanza non era dovuta a Covid-19. Per il pannello D, la percentuale di partecipanti che hanno raggiunto gli obiettivi di riduzione del peso è stata ottenuta dividendo il numero di partecipanti che hanno raggiunto i rispettivi obiettivi alla settimana 72 per il numero di partecipanti con un valore al basale e almeno un valore post-base non mancante. I valori mancanti alla settimana 72 sono stati imputati dall’analisi MMRM. Le barre 𝙸 indicano gli intervalli di confidenza al 95%.
Nella pratica su bodybuilder, si sono osservati i minori sides comparati a ottimi risultati su insulino-resistenza e riduzione contenuta della fame con dosaggi settimanali di 2.5mg.
Alla luce di ciò, e in contesto aspecifico, la Tirzepatide mostra un moderato vantaggio gestionale rispetto alla Semaglutide.
Si è parlato di un ipotetico “rebound” di Grelina con incremento della fame e del consumo calorico (con aumento di peso) dopo la cessazione d’uso di Semaglutide o Tirzepatide. Al momento non esistono dati certi che ci indichino un reale collegamento equazionale tra cessazione d’uso di GLP-1 agonisti > picco in cronico della Grelina > iperfagia > aumento ponderale di peso. Sappiamo, però, che le variazioni di Grelina e GLP-1 a 6 mesi dalla cessazione di una dieta ipocalorica non hanno predetto il recupero del peso da 6 a 18 mesi. Ciò significa che, in un soggetto sano, potrebbe si esserci una maggiore attività della Grelina nelle prime settimane dopo cessazione d’uso di un incretino-mimetico (calo soglia ematica del farmaco e stabilizzazione dei livelli endogeni di GLP-1), ma l’aumento del peso successivo potrebbe risultare con maggiore probabilità dalla modifica omeostatica multi-fattoriale la quale, per trovare un nuovo equilibrio, richiede per lo meno un arco di tempo direttamente proporzionale al tempo di trattamento. Si consideri, inoltre, che un anno dopo la sospensione della Semaglutide sottocutanea a 2,4mg una volta alla settimana e dell’intervento sullo stile di vita, i soggetti possono mostrare una riacquisizione di due terzi della perdita di peso precedente, con cambiamenti simili nelle variabili cardiometaboliche. Qualcosa di un possibile rebound grelinico…
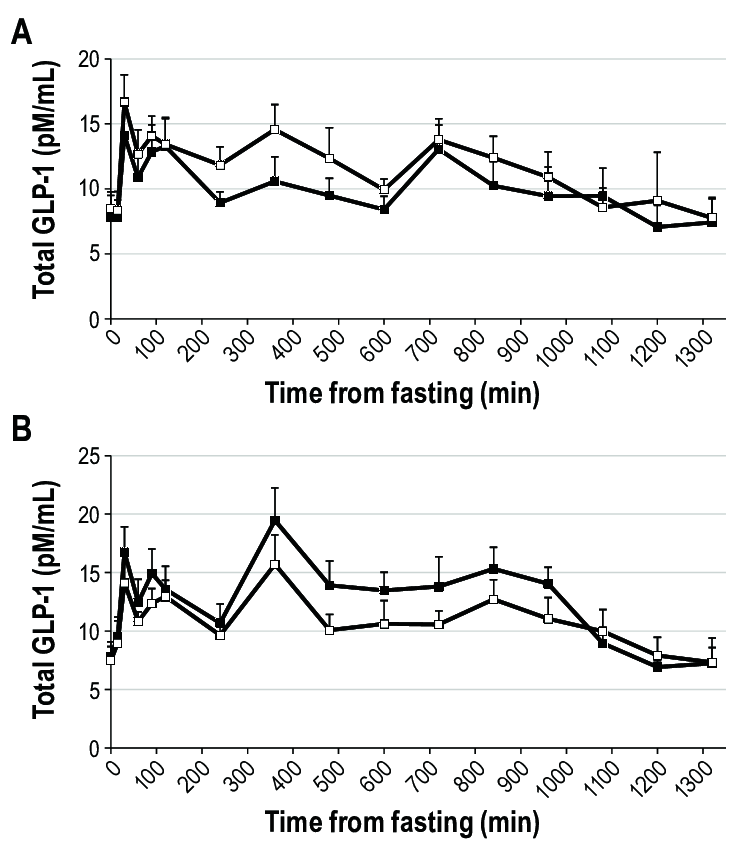
Concentrazione totale di GLP-1 durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli mattutini (P = 0,10) e notturni (P = 0,12) tendevano a essere più bassi e i livelli pomeridiani erano significativamente più bassi (P = 0,016) durante il sonno breve rispetto al sonno abituale nelle donne, mentre negli uomini le concentrazioni pomeridiane di GLP-1 tendevano a essere più alte dopo il sonno breve rispetto al sonno abituale (P = 0,10). I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.Concentrazione di Grelina totale durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli di Grelina mattutina più elevati sono stati osservati dopo il sonno breve rispetto al sonno abituale negli uomini; nelle donne non sono state osservate differenze tra i periodi di sonno. I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.
Conclusioni:
Arrivati alla conclusione di questa disamina abbiamo tutti gli elementi per valutare l’eventuale senso di utilizzo degli Incretino-Mimetici in contesti al di fuori del trattamento del diabete di tipo II o di soggetti obesi.
Le limitazioni date dagli effetti collaterali più comuni, se contestualizzate in una preparazione di un bodybuilder, possono causare non poche problematiche specie nelle vicinanze di una gara; vedi, ad esempio, estrema riduzione dell’assunzione calorica e proteica, mal assorbimento e gonfiore addominale o paresi gastrica.
Sebbene la possibilità di eventi ipoglicemici sia contenuta, e ancor più rara con la Tirzepatide in monoterapia, il rischio, in concomitanza dell’effetto sulla insulino-sensibilità degli AAS o di altre molecole co-somministrate, di questo sides può aumentare in modo sensibile durante una dieta ipocalorica.
In tal sede non ho preso in considerazione i due più preoccupanti, e più rari, effetti collaterali legati all’uso di incretino-mimetici: Tumore Midollare della Tiroide [MTC] e Pancreatite. Quest’ultima può manifestarsi anche con l’uso di AAS, seppur raramente, specie in caso d’uso di molecole aromatizzabili; la presenza di un incretino-mimetico in tali circostanze potrebbe avere un incidenza maggiore nello sviluppo e manifestazione della Pancreatite.[https://jmedicalcasereports.biomedcentral.com/]
In definitiva, i vantaggi potenziali di una una insulino-sensibiltà maggiore iatrogeno-dipendente (visti in precedenza) con l’uso di Incretino-Mimetici è, con i dovuti distinguo complessivi, ottenibile con l’uso di Metformina la quale presenta un margine di sicurezza decisamente più ampio.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Semaglutide and Tirzepatide are More Than Just Weight Loss Drugs [di Type-IIx]
[1] McDonald, L. The Ultimate Diet 2.0: Advanced Cyclical Dieting for Achieving Super Leanness. (2003). Lyle McDonald Publishing.
[2] Volpe S, Lisco G, Racaniello D, Fanelli M, Colaianni V, Vozza A, Triggiani V, Sabbà C, Tortorella C, De Pergola G, Piazzolla G. Once-Weekly Semaglutide Induces an Early Improvement in Body Composition in Patients with Type 2 Diabetes: A 26-Week Prospective Real-Life Study. Nutrients. 2022 Jun 10;14(12):2414. doi: 10.3390/nu14122414.
[3] Kolb H, Kempf K, Röhling M, Martin S. Insulin: too much of a good thing is bad. BMC Med. 2020;18(1):224. Published 2020 Aug 21. doi:10.1186/s12916-020-01688-6
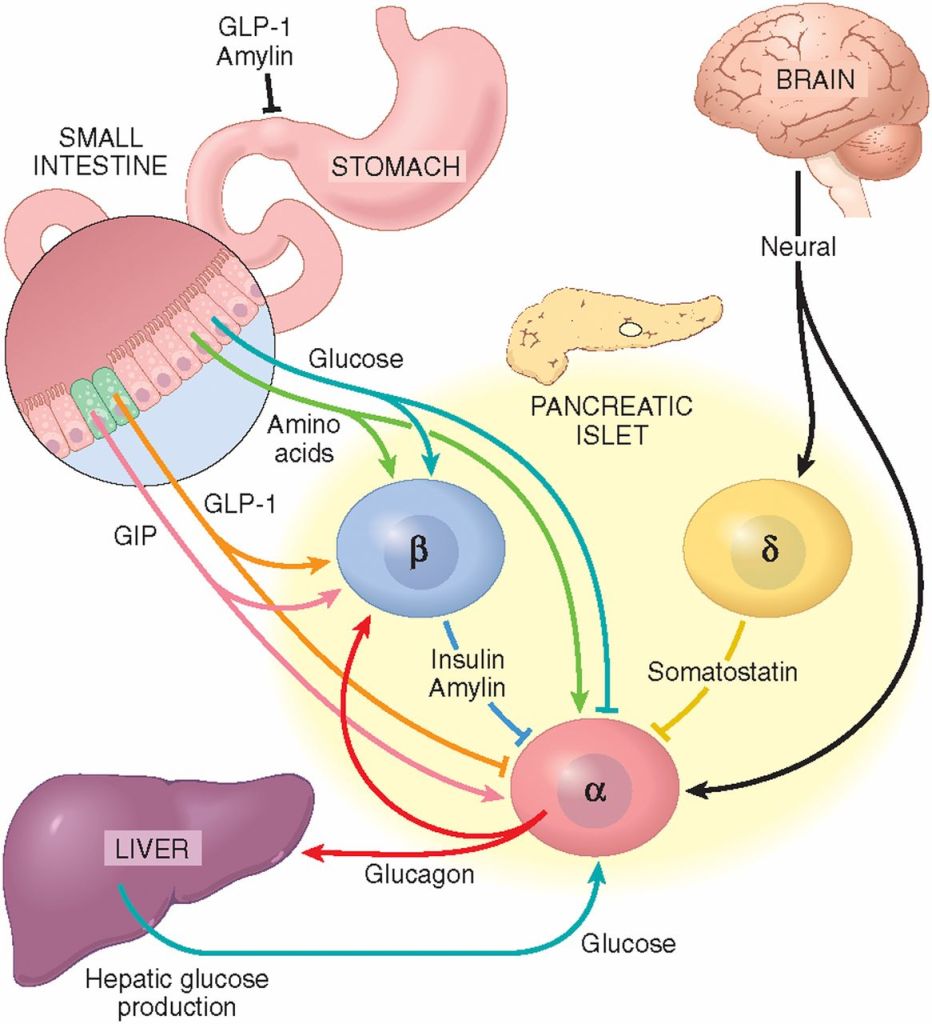
Gli incretino-mimetici sono una recente classe di farmaci antidiabete che prevede la modulazione del sistema delle Incretine. Si legano e attivano i recettori del peptide glucagone-simile-1 (GLP-1) sulle beta-cellule pancreatiche, dando inizio alla secrezione e alla sintesi di Insulina. Gli incretino-mimetici di ultima generazione hanno attività sia sul recettore del GLP-1 sia su quello del Polipeptide Inibitore Gastrico (GIP), un ormone inibitore della famiglia delle secretine (Incretine) con ruolo principale, essendo un’Incretina, di stimolare la secrezione di Insulina.
Poiché questi composti non hanno attività insulinotropica a concentrazioni di glucosio inferiori, il rischio di ipoglicemia – una nota carenza degli attuali trattamenti antidiabete – è basso. Inoltre, è stato dimostrato che gli incretino-mimetici sono associati a effetti benefici sui fattori di rischio cardiovascolare, come la perdita di peso, la riduzione della pressione sanguigna e le modifiche del profilo lipidico. Ciò nonostante, gli effetti avversi che possono causare non sono trascurabili e la loro diffusione per uso “Off-Label” ne ha mostrato le limitazioni sia in soggetti obesi che in utilizzatori per finalità estetiche.
La disamina che seguirà sarà divisa in due parti preposte a spiegare, a partire dalla loro genesi, il funzionamento delle incretine per poi passare in rassegna le caratteristiche di quelle che sono le forme farmaceutiche di incretino-mimetici utilizzate in medicina e in ambito “Off-Label/ricreativo”.
In questa prima parte vedremo nel dettaglio le incretine endogene e le molecole incretino-mimentiche presenti nel mercato…
Ormoni Incretinici [Incretine]:
Joel Habener
Negli anni ’70, Jens Juul Holst e Joel Habener iniziarono la ricerca sull’ormone GLP-1, inizialmente in relazione alla malattia dell’ulcera duodenale.[1] Esaminarono gli ormoni secreti durante l’alimentazione e li testarono su pancreas di maiale, portando alla scoperta della notevole potenza del GLP-1 nel 1988. Il loro lavoro, che in seguito ha contribuito in modo significativo al trattamento del diabete e dell’obesità, è valso a loro e a Daniel Drucker il premio della Fondazione Warren Alpert del 2021.[1] La ricerca è proseguita e nel 1993 Michael Nauck è riuscito a infondere il GLP-1 in persone affette da diabete di tipo II, stimolando l’Insulina e inibendo il Glucagone e portando la glicemia a livelli normali. Tuttavia, il trattamento dei pazienti diabetici con gli ormoni GLP-1 ha provocato notevoli effetti collaterali, inducendo i ricercatori finanziati da Novo Nordisk a cercare di sviluppare un composto adatto all’uso terapeutico.[1]
Nel 1998 un team di ricercatori di Novo Nordisk, guidato dalla scienziata Lotte Bjerre Knudsen, ha sviluppato la Liraglutide, un agonista del recettore del peptide glucagone-simile-1 con potenziale di utilizzo per il trattamento del diabete di tipo II.[2]
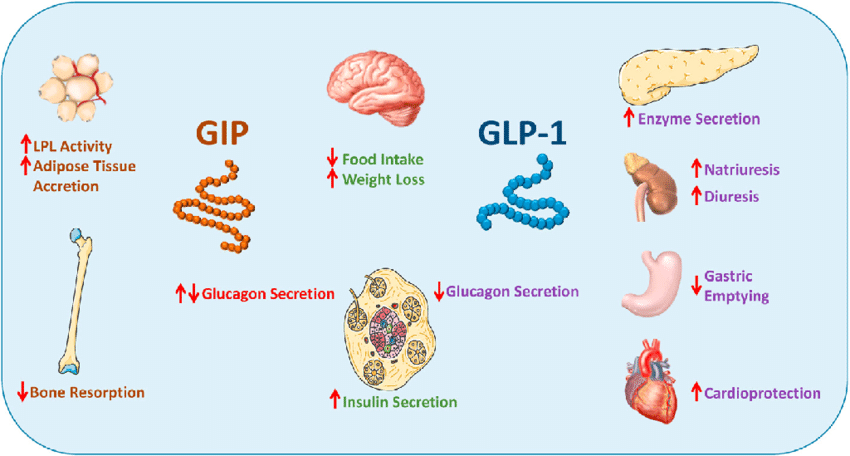
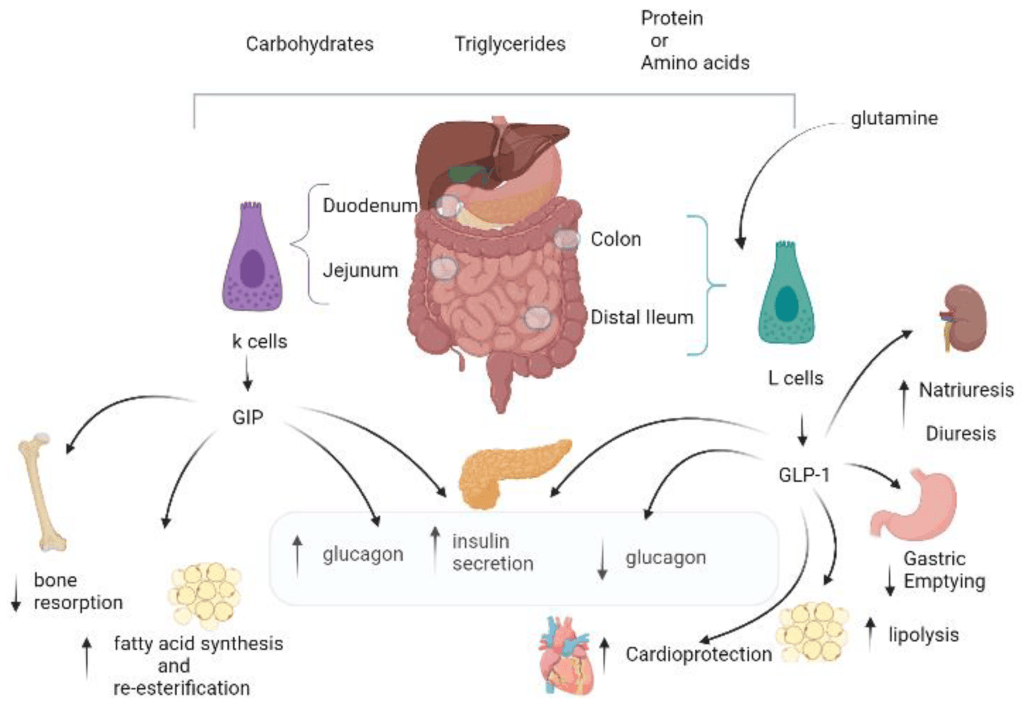
Gli ormoni incretinici GLP-1 e il GIP sono ormoni peptidici intestinali rilasciati in risposta all’ingestione di cibo.(3) L’effetto più importante del GLP-1 e del GIP è la loro capacità di potenziare la secrezione di Insulina indotta dal Glucosio da parte del pancreas – il cosiddetto effetto incretinico. Nei soggetti sani l’effetto incretinico rappresenta fino al 70% dell’Insulina secreta in risposta all’ingestione di Glucosio.(4) Il GLP-1 è un polipeptide di 30 aminoacidi sintetizzato dal proglucagone nelle cellule endocrine L distribuite principalmente nella mucosa della parte distale dell’intestino tenue e del colon. Il GIP è un polipeptide di 42 aminoacidi rilasciato dalle cellule endocrine K presenti nella mucosa del duodeno e della parte superiore del digiuno.(5) Mentre il GLP-1 viene rapidamente degradato (dall’enzima ubiquitario dipeptidil peptidasi-4 (DPP-4)) in circolo con un’emivita apparente di 1 – 1,5 minuti,(6) il GIP viene degradato più lentamente, con un’emivita per l’ormone intatto di 7 minuti.(7) L’ormone aumenta la secrezione di Insulina all’inizio del pasto, ma non ha alcuna attività insulinotropica a concentrazioni di glucosio inferiori (meno di 4mM), non favorendo così l’ipoglicemia. Il GLP-1 aumenta anche la biosintesi dell’Insulina e l’espressione genica della stessa. Inoltre, esercita azioni trofiche e protettive sulle beta-cellule(8) e inibisce fortemente la secrezione pancreatica di Glucagone in modo glucosio-dipendente.(9) Al contrario, è stato dimostrato che la GIP stimola la secrezione di Glucagone. Gli ormoni esercitano il loro effetto insulinotropico attraverso recettori accoppiati a proteine G sulle beta-cellule pancreatiche.(10) Oltre agli effetti sul pancreas endocrino, entrambi gli ormoni hanno diverse altre funzioni. I recettori del GLP-1 si trovano in varie regioni del cervello (11) e, una volta attivati, promuovono il senso di sazietà che, in combinazione con l’inibizione della motilità gastrointestinale indotta dal GLP-1 (mediata dal nervo vago (12) ), riduce l’assunzione di cibo e, consequenzialmente, il peso corporeo. I recettori del GLP-1 si trovano anche nel cuore (13) e la maggior parte dei dati suggerisce che il GLP-1 esercita effetti protettivi sul miocardio. È stato inoltre riscontrato che il GLP-1 riduce l’aumento postprandiale dei trigliceridi e abbassa la concentrazione di acidi grassi liberi nell’uomo.(14) Infine, studi su animali e sull’uomo indicano che il GLP-1 ha proprietà natriuretiche e diuretiche attraverso la modulazione dello scambio renale Na+/H+ (15) – un meccanismo che potrebbe servire a ridurre la pressione sanguigna. La GIP non sembra avere effetti fisiologici sul tratto gastrointestinale, sull’appetito o sull’assunzione di cibo, ma può avere un ruolo nel metabolismo dei lipidi (16) e delle ossa.(17)
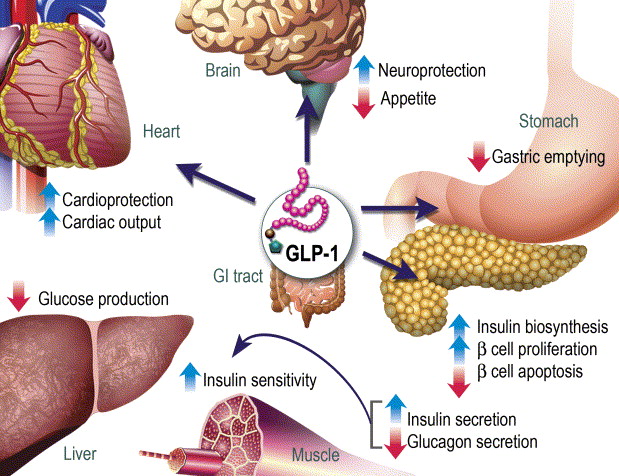
Azioni del GLP-1 nei tessuti periferici. Il GLP-1 agisce direttamente sul pancreas endocrino, sul cuore, sullo stomaco e sul cervello, mentre le azioni su fegato e muscoli sono indirette.
Nei pazienti con diabete di tipo II l’effetto incretinico è gravemente ridotto.(18, 19) Questo tratto fisiopatologico gioca probabilmente un ruolo centrale nell’incapacità di questi pazienti di secernere una quantità di Insulina sufficiente a prevenire l’iperglicemia in seguito all’assunzione di glucosio per via orale.(20-30) L’attenuata secrezione postprandiale (21) e la diminuita potenza insulinotropica del GLP-1 (22), in combinazione con l’abolizione dell’effetto insulinotropico del GIP (23) , sembrano essere responsabili del ridotto effetto incretinico nei pazienti con diabete di tipo II. Poiché l’effetto insulinotropico del solo GLP-1 (e non del GIP) è conservato nei pazienti con diabete di tipo II, sono state sviluppate modalità di trattamento antidiabete basate sull’effetto di questo peptide. È interessante notare che l’infusione endovenosa (iv) di GLP-1 nativo è in grado di normalizzare la glicemia nei pazienti con diabete di tipo 2, (24) ma a causa della breve emivita del GLP-1, la somministrazione terapeutica del GLP-1 nativo non è praticabile. Pertanto, per sfruttare le azioni benefiche del GLP-1 nel diabete di tipo II, sono stati sviluppati agonisti stabili a lunga durata d’azione del recettore del GLP-1 (incretino-mimetici). Nella sezione seguente verranno descritti gli incretino-mimetici disponibili, le loro caratteristiche e applicazioni.
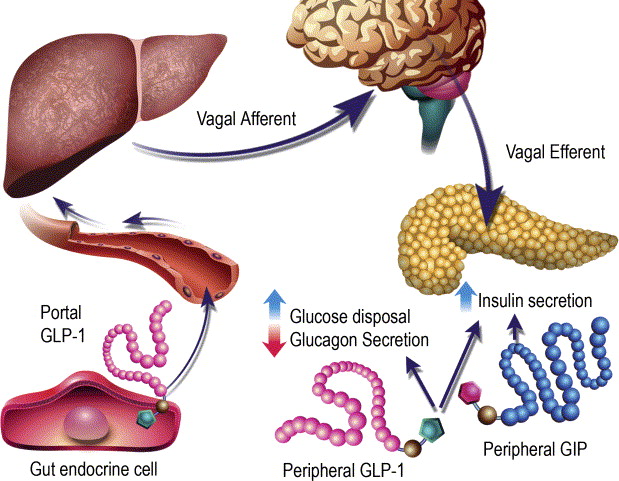
Ruoli contrastanti di GLP-1 e GIP sull’omeostasi del glucosio. Il GLP-1 secreto nella vena porta attiva un sensore del glucosio portale che segnala, tramite afferenze vagali, il sistema nervoso centrale e, a sua volta, le efferenze vagali aumentano la secrezione di Insulina. Sia il GLP-1 che il GIP attivano anche direttamente la secrezione di Insulina attraverso il legame con i loro recettori distinti sulle cellule β dell’isolotto.
Farmaci Incretino-mimetici:
Exenatide[approvato nel 2005/2012]
L’Exenatide, il primo di questa classe di farmaci, è stato introdotto sul mercato negli Stati Uniti nel 2005 e in Europa nel 2007 con il nome commerciale Byetta® (Amylin Pharmaceuticals/Eli Lilly).
L’Exenatide è un peptide di 39 aminoacidi; è una versione sintetica dell’exendin-4, un peptide presente nel veleno del “mostro di Gila”.
L’ Exenatide è stata isolata per la prima volta nel 1992 presso il Veterans Administration Medical Center di New York City.[25]
L’Exenatide si lega al recettore umano intatto del Peptide Glucagone-Simile-1 (GLP-1R) in modo simile al GLP-1; l’Exenatide ha un’omologia aminoacidica del 50% con il GLP-1 e ha un’emivita più lunga in vivo.[26]
Si ritiene che l’Exenatide faciliti il controllo del glucosio in almeno cinque modi:
L’Exenatide aumenta la risposta del Pancreas[27] (cioè aumenta la secrezione di Insulina) in risposta al consumo dei pasti; il risultato è il rilascio di una quantità di Insulina maggiore e più appropriata che aiuta a ridurre l’aumento della glicemia dovuto al consumo di cibo. Una volta che i livelli di glucosio nel sangue si avvicinano ai valori normali, la risposta del Pancreas alla produzione di Insulina si riduce; altri farmaci (come la rInsulina ) sono efficaci nell’abbassare la glicemia, ma possono “superare” il loro obiettivo e causare un abbassamento eccessivo della glicemia, provocando la pericolosa condizione di ipoglicemia.
L’Exenatide sopprime anche il rilascio di Glucagone da parte del Pancreas in risposta al pasto, impedendo al fegato di produrre eccessivamente glucosio quando non è necessario e prevenendo così l’iperglicemia (livelli elevati di glucosio nel sangue).
L’Exenatide contribuisce a rallentare lo svuotamento gastrico e quindi a ridurre la velocità di comparsa nel sangue del glucosio derivato dai pasti.
L’Exenatide ha un effetto sottile ma prolungato di riduzione dell’appetito e di promozione della sazietà attraverso i recettori ipotalamici (recettori diversi da quelli dell’Amilina). La maggior parte delle persone che utilizzano l’Exenatide perdono lentamente peso e, in genere, la perdita di peso maggiore è ottenuta dalle persone più in sovrappeso all’inizio della terapia con l’Exenatide. Gli studi clinici hanno dimostrato che l’effetto di riduzione del peso continua allo stesso ritmo per 2,25 anni di uso continuato. Se suddivisi in quartili di perdita di peso, il 25% più alto registra una sostanziale perdita di peso, mentre il 25% più basso non registra alcuna perdita o un lieve aumento di peso.
L’Exenatide riduce il contenuto di grasso nel fegato. L’accumulo di grasso nel fegato o la malattia del fegato grasso non alcolico (NAFLD) è fortemente correlata a diversi disturbi metabolici, in particolare a un basso livello di HDL e a Trigliceridi elevati, presenti nei pazienti con diabete di tipo II. È emerso che l’Exenatide ha ridotto il grasso epatico nei topi[28], nei ratti[29] e nell’uomo.[30]
I principali effetti collaterali del Exenatide (che condivide con tutte le molecole apopartenenti alla sua “famiglia”) sono nausea e vomito da lievi a moderati e transitori. L’incidenza dell’ipoglicemia associata al trattamento è bassa (31) – apparentemente dovuta agli effetti insulinotropi e glucagonostatici del GLP-1, dipendenti dal glucosio. Tuttavia, in combinazione con altri ipoglicemizzanti l’incidenza aumenta e dipende dalla dose di questi. Nella maggior parte degli studi con Exenatide gli episodi ipoglicemici minori sono definiti come glucosio plasmatico <3,3mM; negli studi LEAD sono definiti come glucosio plasmatico <3,1 mM. Negli studi che utilizzano Exenatide in combinazione con ipoglicemizzanti il rischio di episodi ipoglicemici minori è riportato tra il 15% e il 36%.(32) Nello studio Exenatide/Insulina Glargine l’1,5% dei pazienti ha sperimentato un’ipoglicemia grave.(33) Non c’è stata differenza tra i gruppi e l’incidenza è stata simile nei due gruppi.
Circa il 40% dei pazienti trattati con Exenatide in studi a lungo termine, controllati con placebo, ha sviluppato anticorpi contro il peptide durante le prime 30 settimane di trattamento.(32) L’esatto impatto degli anticorpi a lungo termine deve essere stabilito. L’Exenatide non è raccomandata durante la gravidanza o l’allattamento a causa della mancanza di dati sufficienti. L’Exenatide non deve essere utilizzata in pazienti con insufficienza renale, poiché viene eliminata principalmente nei reni attraverso la filtrazione glomerulare (31) e sono stati segnalati casi di insufficienza renale acuta. Nessun dato indica l’inibizione o l’induzione degli enzimi di metabolizzazione dei farmaci del citocromo P450. Dal 2005 la Food and Drug Administration (FDA) statunitense ha ricevuto 170 segnalazioni di pancreatite in pazienti trattati con Exenatide e ha ricevuto segnalazioni di pancreatite acuta, alcune delle quali erano casi gravi di pancreatite emorragica o necrotizzante in pazienti che assumevano Exenatide. Negli studi LEAD sono state osservate in totale (al 2010) 9 segnalazioni di pancreatite, nei pazienti trattati con Exanatide. E’ tutt’ora poco chiaro il nesso causale tra pancreatite ed Exenatide. Tuttavia, si raccomanda di interrompere il trattamento con incretino-mimetici in caso di sospetta pancreatite.(34) È stato suggerito che i risultati sul cancro nei roditori siano causati da un meccanismo non genotossico e specifico mediato dal recettore GLP-1 a cui i roditori sono particolarmente sensibili. La rilevanza per l’uomo è probabilmente insignificante dal punto di vista clinico, ma sono necessari ulteriori studi per chiarire i potenziali meccanismi alla base dello sviluppo del tumore delle cellule C nei pazienti trattati con analoghi del GLP-1 e le loro possibili implicazioni cliniche.(35)
Nel marzo 2013, la FDA ha pubblicato una Drug Safety Communication in cui annunciava l’avvio di indagini sui mimetici dell’incretina a causa dei risultati ottenuti da ricercatori accademici.[36] Poche settimane dopo, l’Agenzia europea per i medicinali ha avviato un’indagine simile sugli agonisti del GLP-1 e sugli inibitori della DPP-4.[37]
53 cause consolidate contro i produttori di “prodotti GLP-1/DPP-4” sono state archiviate nel 2015.[38]
Nel 2016 è stato pubblicato un lavoro che dimostra che è in grado di invertire l’alterata segnalazione del calcio nelle cellule epatiche steatotiche, che a sua volta potrebbe essere associata a un corretto controllo del glucosio.[39]
È in fase di valutazione per il trattamento del morbo di Parkinson.[40] Uno studio clinico di fase 3, iniziato nel gennaio 2020, ha avuto la sua data di completamento il 30 giugno 2024 (NCT04232969).[41]
L’Exenatide si presenta come soluzione parenterale destinata all’iniezione. Può essere somministrata nel sottocute dell’addome, del braccio o della coscia. Per migliorare la tollerabilità gastrointestinale, la terapia con Exenatide deve essere iniziata a 5mcg per dose somministrata due volte al giorno per almeno un mese. La dose di Exenatide può poi essere aumentata a 10mcg due volte al giorno.(31) La soddisfazione dei pazienti che hanno utilizzato Exenatide è stata valutata in un paio di studi. L’aggiunta di Exenatide a Metformina e altri ipoglicemizzanti ha determinato un miglioramento significativo della soddisfazione per il trattamento e della qualità di vita correlata alla salute dei pazienti dal basale a 26 settimane.(42) Il miglioramento è stato simile per i pazienti trattati con Insulina Glargine.
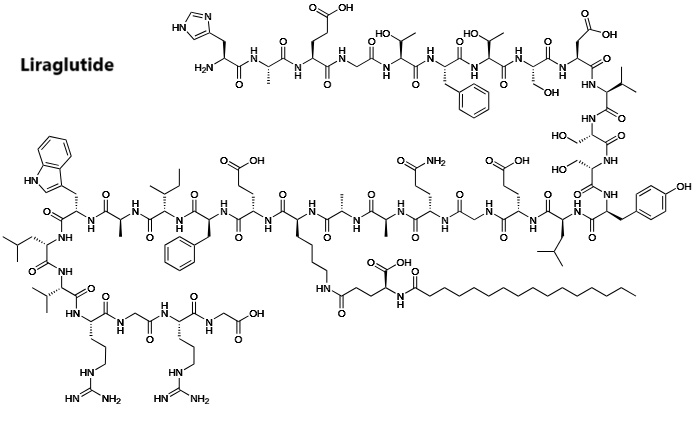
Liraglutide [approvato nel 2010]
La Liraglutide è un analogo acilato del GLP-1 umano e presenta il 97% di omologia di sequenza con il GLP-1 nativo. L’analogo è prodotto con la tecnologia del DNA ricombinante nel lievito.(43) Ha un effetto sul recettore del GLP-1 simile a quello descritto per l’Exenatide. Un elevato grado di legame con le proteine plasmatiche causa una minore suscettibilità al metabolismo da parte della DPP-4 e l’emivita dopo la somministrazione di Liraglutide è di circa 13 ore.(44) Questo profilo d’azione prolungato rende Liraglutide adatto alla somministrazione una volta al giorno. Non ci sono differenze clinicamente significative nella farmacocinetica di Liraglutide tra soggetti di sesso maschile e femminile, soggetti di razza diversa o soggetti anziani e giovani.(45)
La Liraglutide, venduta tra l’altro con i marchi Victoza e Saxenda, è un farmaco antidiabetico utilizzato per il trattamento del diabete di tipo II e dell’obesità cronica.[46][47] Si tratta di una terapia di seconda linea per il diabete dopo la terapia di prima linea con la Metformina.[46][48] Non sono chiari i suoi effetti sugli esiti di salute a lungo termine, come le malattie cardiache e l’aspettativa di vita.[46][49] Viene somministrata mediante iniezione sotto cutanea.[46]
La Liraglutide è stata approvata per uso medico nell’Unione Europea nel 2009 e negli Stati Uniti nel 2010.[50][51] Nel 2021 è stato il 166° farmaco più comunemente prescritto negli Stati Uniti, con oltre 3 milioni di prescrizioni.[52][53]
L’azione prolungata della Liraglutide si ottiene attaccando una molecola di acido grasso in una posizione della molecola GLP-1-(7-37), consentendole di auto-associarsi e di legarsi all’albumina nel tessuto sottocutaneo e nel flusso sanguigno. Il GLP-1 attivo viene quindi rilasciato dall’albumina a un ritmo lento e costante. Il legame con l’albumina determina inoltre una degradazione più lenta e un’eliminazione renale ridotta rispetto a quella del GLP-1-(7-37).[54]
A) La struttura molecolare del GLP-1 umano. B) La struttura molecolare di Exenatide (il colore grigio indica le differenze di struttura rispetto al GLP-1 umano). C) La struttura molecolare della Liraglutide (il colore grigio indica le differenze di struttura rispetto al GLP-1 umano).
Come abbiamo visto, La Liraglutide è un agonista acilato del recettore del GLP-1, derivato dal GLP-1-(7-37) umano, una forma meno comune di GLP-1 endogeno.
Riduce l’iperglicemia correlata ai pasti (per 24 ore dopo la somministrazione) aumentando la secrezione di Insulina (solo) quando richiesto dall’aumento dei livelli di glucosio, ritardando lo svuotamento gastrico e sopprimendo la secrezione prandiale di Glucagone.[54][55]
Quindi, la Liraglutide provoca il rilascio di Insulina nelle cellule beta pancreatiche in presenza di una glicemia elevata. Questa secrezione di Insulina si attenua quando le concentrazioni di glucosio diminuiscono e si avvicinano all’euglicemia (livello normale di glucosio nel sangue). Diminuisce inoltre la secrezione di Glucagone in modo glucosio-dipendente e ritarda lo svuotamento gastrico. A differenza del GLP-1 endogeno, la Liraglutide è stabile contro la degradazione metabolica da parte delle peptidasi, con un’emivita plasmatica di 13 ore.[56][54]
Nei pazienti ad alto rischio cardiovascolare, è stato dimostrato che la Liraglutide riduce il rischio di morte per cause cardiovascolari, infarto miocardico non fatale o ictus non fatale. Le linee guida dell’ADA considerano attualmente Liraglutide una terapia farmacologica di prima linea per il diabete di tipo II (di solito insieme alla Metformina), in particolare per i pazienti con malattie cardiovascolari aterosclerotiche o obesità.[57] Una revisione Cochrane del 2011 ha dimostrato una riduzione dell’HbA1c dello 0,24% in più con Liraglutide. Del 24% in più con Liraglutide a 1,8 mg rispetto a Insulina Glargine, 0,33% in più rispetto a Exenatide 10mcg due volte al giorno, Sitagliptin e Rosiglitazone. In uno studio randomizzato e controllato (RCT) che ha confrontato Liraglutide, Insulina Glargine, Glimepiride e Sitagliptin (tutti aggiunti alla Metformina) con un follow-up di cinque anni, Insulina Glargine e Liraglutide sono risultate modestamente più efficaci nel raggiungimento e nel mantenimento dell’HbA1c target,[58] senza alcuna differenza negli esiti delle malattie microvascolari e cardiovascolari.[59]
La Liraglutide può anche essere utilizzata insieme alla dieta e all’esercizio fisico per la gestione cronica del peso negli adulti.[46] La Liraglutide ha portato a una perdita di peso maggiore rispetto ad alcuni precedenti analoghi del peptide glucagone-simile,[60] ma è meno efficace della dose standard di Semaglutide per la perdita di peso.[61][62]
In un recente studio pubblicato nel settembre 2024, Liraglutide ha aiutato i bambini di età compresa tra i 6 e i 12 anni a ridurre l’indice di massa corporea del 7,4% in uno studio di 56 settimane.[63] Se da un lato lo studio ha mostrato i potenziali benefici del farmaco, dall’altro solleva preoccupazioni riguardo all’uso di farmaci contro l’obesità in bambini così piccoli.[64] Novo Nordisk, l’azienda innovatrice che commercializza Liraglutide, ha chiesto alle autorità di regolamentazione statunitensi ed europee di estendere l’approvazione di Saxenda anche a questa fascia d’età più giovane, dato che attualmente è approvato solo per adolescenti e adulti.[65]
aUso di 1.8mg di Liraglutide.
Come per l’Exenatide, la Liraglutide ha un effetto significativo sul peso corporeo, come dimostrano i dati relativi a Liraglutide somministrata a 1,8mg/die. Liraglutide ha ridotto il peso corporeo medio o è stato neutro rispetto al placebo o ai comparatori attivi, in monoterapia (66) e in combinazione con uno (67) o due (68) agenti antidiabete orali. Lo studio LEAD 662 ha esaminato il profilo lipidico con Exenatide e Liraglutide. Sono state osservate riduzioni significative maggiori dei trigliceridi (-0,4 vs -0,2 mM) e degli acidi grassi liberi (-0,17 vs -0,10 mM) nel gruppo Liraglutide. Entrambi i composti hanno causato una riduzione significativa della pressione arteriosa (pressione sistolica -2,2 mmHg e pressione diastolica -1,5 mmHg) senza differenze significative tra i due composti.
Tra gli effetti collaterali si annoverano ipoglicemia, nausea, vertigini, dolore addominale e dolore nel sito di iniezione.[46] Gli effetti collaterali gastrointestinali tendono a essere più forti all’inizio del periodo di trattamento e si attenuano con il tempo.[60] Altri effetti collaterali gravi possono includere angioedema, pancreatite, malattie della cistifellea e problemi renali. L’uso in gravidanza e durante l’allattamento non è sicuro.[46] Una black box warning avverte che nei ratti trattati con Liraglutide sono stati osservati tumori midollari della tiroide, ma è “Sconosciuto se Liraglutide causi tumori delle cellule C della tiroide, incluso il carcinoma midollare della tiroide (MTC), nell’uomo, poiché la rilevanza per l’uomo di tali tumori nei roditori non è stata determinata.”[46]
A proposito del MTC, a esposizioni otto volte superiori a quelle utilizzate nell’uomo, la Liraglutide ha causato un aumento statisticamente significativo dei tumori alla tiroide nei ratti. La rilevanza clinica di questi risultati è sconosciuta.[69] Negli studi clinici, il tasso di tumori alla tiroide nei pazienti trattati con Liraglutide è stato di 1,3 per 1000 anni-paziente (4 persone) rispetto a 1,0 per 1000 pazienti (1 persona) nei gruppi di confronto. L’unica persona nel gruppo di confronto e quattro delle cinque persone nel gruppo Liraglutide avevano marcatori sierici (calcitonina elevata) suggestivi di una malattia preesistente al basale.[69]
L’FDA ha dichiarato che la calcitonina sierica, un biomarcatore del carcinoma midollare della tiroide, era leggermente aumentata nei pazienti con Liraglutide, ma ancora nei limiti della norma, e che era necessario un monitoraggio continuo per 15 anni in un registro dei tumori.[70]
Un altro effetto collaterale preoccupante è rappresentato dalla possibilità (sebbene rara) di sviluppare pancreatite.
Nel 2013, un gruppo della Johns Hopkins ha riportato un’associazione con apparenza statisticamente significativa tra l’ospedalizzazione per pancreatite acuta e un precedente trattamento con derivati del GLP-1 (come la precedentemente vista Exenatide) e inibitori della DPP-4 (come il Sitagliptin).[71] In risposta, la FDA degli Stati Uniti e l’Agenzia Europea per i Medicinali hanno condotto una revisione di tutti i dati disponibili in merito alla possibile connessione tra i mimetici dell’Incretina e la pancreatite o il cancro al pancreas. In una lettera congiunta del 2014 al New England Journal of Medicine, le agenzie hanno concluso che “Un’analisi congiunta dei dati di 14.611 pazienti con diabete di tipo II provenienti da 25 studi clinici nel database di sitagliptin non ha fornito alcuna prova convincente di un aumento del rischio di pancreatite o di cancro al pancreas” e “Entrambe le agenzie concordano sul fatto che le affermazioni relative a un’associazione causale tra i farmaci a base di Incretine e la pancreatite o il cancro al pancreas, espresse di recente nella letteratura scientifica e nei media, non sono coerenti con i dati attuali”. L’FDA e l’EMA non hanno ancora raggiunto una conclusione definitiva su tale relazione causale. Sebbene la totalità dei dati esaminati fornisca rassicurazioni, la pancreatite continuerà a essere considerata un rischio associato a questi farmaci finché non saranno disponibili ulteriori dati; entrambe le agenzie continuano a indagare su questo segnale di sicurezza”[72].
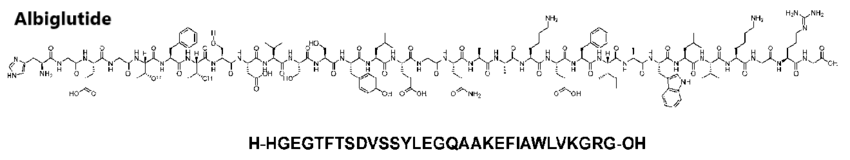
Albiglutide [approvato nel 2014]
L’Albiglutide (nome commerciale Eperzan in Europa e Tanzeum negli Stati Uniti) è un farmaco agonista del GLP-1 commercializzato da GlaxoSmithKline (GSK) per il trattamento del diabete di tipo II.
L’Albiglutide è un peptide composto da 645 aminoacidi proteinogenici con 17 ponti disolfuro. Gli aminoacidi 1-30 e 31-60 costituiscono due copie di GLP-1 umano modificato, in cui l’alanina in posizione 2 è stata scambiata con una glicina per migliorare la resistenza alla DPP-4.[73] La sequenza rimanente è costituita da albumina umana.
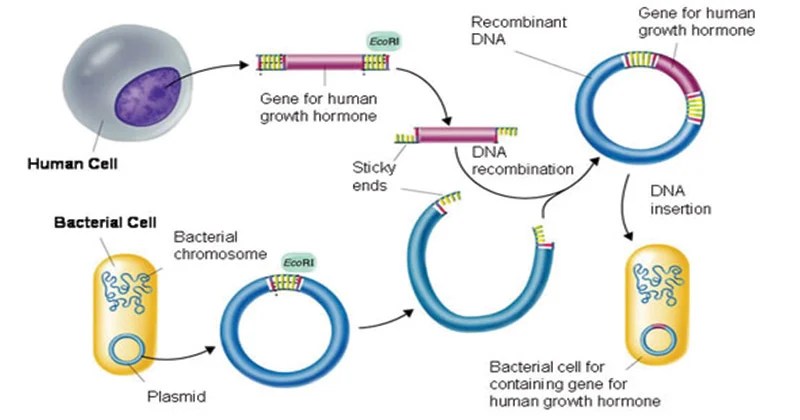
Viene bioingegnerizzata nel lievito Saccharomyces cerevisiae utilizzando la tecnologia del DNA ricombinante.[74]
Fasi applicative della tecnologia del DNA ricombinante.
Il farmaco è stato brevettato dalla Human Genome Sciences e sviluppato in collaborazione con GSK.[75]
La GSK ha presentato domanda di approvazione alla FDA statunitense il 14 gennaio 2013 e all’Agenzia europea per i medicinali (EMA) il 7 marzo 2013. Nel marzo 2014, GSK ha ricevuto dalla Commissione Europea l’approvazione per la commercializzazione di Albiglutide con il nome di Eperzan.[76] Nell’aprile 2014, la FDA statunitense ha approvato Albiglutide con il nome di Tanzeum.[77]
Nell’agosto 2017, GSK annunciò l’intenzione di ritirare il farmaco dal mercato mondiale entro luglio 2018 per motivi economici.[78]
L’Albiglutide, come gli altri incretino-mimetici, è stato utilizzato per il trattamento del diabete di tipo II negli adulti. Può essere utilizzato da solo (se la terapia con Metformina è inefficace o non tollerata) o in combinazione con altri farmaci antidiabetici, comprese le forme di Insulina.[74]
Secondo un’analisi del 2015, l’Albiglutide è meno efficace di altri agonisti del GLP-1 per la riduzione dell’HbA1c e la perdita di peso. Sembra inoltre avere meno effetti collaterali rispetto alla maggior parte degli altri farmaci di questa classe, ad eccezione delle reazioni nel sito di iniezione che sono più comuni con Albiglutide rispetto, ad esempio, a Liraglutide.[79]
Dopo l’iniezione sottocutanea, l’Albiglutide raggiunge le massime concentrazioni ematiche dopo tre-cinque giorni. Le concentrazioni allo stato stazionario vengono raggiunte dopo tre-cinque settimane. Essendo resistente alla dipeptidil peptidasi-4 (DPP-4),[73] l’enzima che scompone il GLP-1, l’Albiglutide ha un’emivita biologica di cinque (da quattro a sette) giorni, notevolmente più lunga rispetto agli analoghi del GLP-1 più vecchi, l’Exenatide e la Liraglutide. [80][81] Ciò consente una somministrazione una volta alla settimana,[74] a differenza della Liraglutide ma come la forma a rilascio prolungato dell’Exenatide.
L’Albiglutide agisce come agonista del recettore GLP-1, il che lo rende un tipo di incretino-mimetico. Questo provoca un aumento della secrezione di insulina, soprattutto in presenza di glucosio elevato nel sangue, e rallenta anche lo svuotamento gastrico.[74]
La differenza nel meccanismo d’azione del Albiglutide con gli altri agonisti del recettore del GLP-1, dipende dalla sua struttura la quale rende difficile l’attraversamento della barriera emato-encefalica. Ciò significa che non influisce sul sistema nervoso centrale come altri agonisti del recettore del GLP-1 e potrebbe essere responsabile della limitata perdita di peso osservata con questo farmaco.[81]
Dulaglutide[approvato nel 2014]
La Dulaglutide, venduta tra l’altro con il nome commerciale Trulicity,[8] è un farmaco utilizzato per il trattamento del diabete di tipo II in combinazione con la dieta e l’esercizio fisico.[83][84] È inoltre approvato negli Stati Uniti per la riduzione degli eventi cardiovascolari avversi maggiori negli adulti con diabete di tipo II che presentano una malattia cardiovascolare conclamata o molteplici fattori di rischio cardiovascolare.[85]
Come per gli altri incretino-mimetici visti in precedenza, la Dulaglutide si lega ai recettori del GLP-1, rallentando lo svuotamento gastrico e aumentando la secrezione di Insulina da parte delle cellule β pancreatiche. Contemporaneamente, il peptide riduce l’elevata secrezione di Glucagone inibendo le cellule α del pancreas, poiché è noto che il Glucagone è elevato in modo inappropriato nei pazienti diabetici.
Più precisamente, la Dulaglutide è un agonista del recettore del GLP-1 costituito da GLP-1(7-37) legato covalentemente a un frammento Fc di IgG4 umana.
La sicurezza e l’efficacia della Dulaglutide sono state valutate in sei studi clinici in cui 3.342 soggetti con diabete di tipo II hanno ricevuto Dulaglutide. I soggetti che hanno ricevuto Dulaglutide hanno registrato un miglioramento del controllo glicemico, osservato con riduzioni del livello di HbA1c.[86]
La Food and Drug Administration (FDA) statunitense ha approvato la Dulaglutide con una strategia di valutazione e mitigazione dei rischi (REMS),[86] e ha concesso l’approvazione di Trulicity a Eli Lilly and Company.[86] La REMS consiste in una serie di misure che Eli Lilly adotterà per sensibilizzare i medici sul rischio di pancreatite e sul potenziale rischio di carcinoma midollare della tiroide associato al farmaco.[87]
Nel 2020, la FDA ha approvato due dosi più elevate del farmaco, 3,0mg e 4,5mg, sulla base dei risultati dello studio AWARD-11 che hanno dimostrato una migliore riduzione del glucosio e benefici sul peso.[88]
Il peptide è indicato per gli adulti con diabete di tipo II come aggiunta alla dieta e all’esercizio fisico per migliorare il controllo glicemico. La Dulaglutide non è indicato nel trattamento di soggetti con diabete di tipo I o di pazienti con chetoacidosi diabetica perché questi problemi sono dovuti all’incapacità delle isole pancreatiche di produrre Insulina e una delle azioni della Dulaglutide è quella di stimolare le isole funzionanti a produrre più Insulina. La Dulaglutide può essere utilizzata da solo o in combinazione con altri farmaci per il diabete di tipo II, in particolare Metformina, Sulfoniluree, Tiazolidinedioni e Insulina da assumere contemporaneamente ai pasti.[89]
Il programma di sperimentazione clinica di fase 3 del farmaco ha dimostrato riduzioni dell’emoglobina A1c di circa l’1% con le dosi di 0,75mg e 1,5mg del farmaco, insieme a una perdita di peso media di circa 5Kg. Le dosi più elevate da 3,0mg e 4,5mg, approvate nel 2020, hanno dimostrato riduzioni dell’emoglobina A1c più vicine all’1,5% e una perdita di peso leggermente superiore.[90]
DPP-4
Una meta-analisi del 2017 non ha supportato l’ipotesi che il trattamento con agonisti del GLP-1 o inibitori della DPP-4 aumenti la mortalità per tutte le cause nei diabetici di tipo II.[91]
La Dulaglutide viene assorbita lentamente dopo l’iniezione sottocutanea. In uno studio farmacocinetico condotto su 20 adulti sani, la Cmax si è verificata entro 24-48 ore dalla somministrazione. La biodisponibilità assoluta media di Dulaglutide dopo iniezioni sottocutanee di dosi singole da 0,75mg e 1,5mg è stata rispettivamente del 65% e del 47%. L’emivita media della Dulaglutide somministrato a varie dosi è stata di circa 3,75 giorni (89,9 ore). Questa emivita prolungata consente la somministrazione una volta alla settimana. Le informazioni di prescrizione indicano un’emivita di circa 5 giorni.
Gli effetti collaterali più comuni includono disturbi gastrointestinali, come dispepsia, inappetenza, nausea, vomito, dolore addominale, diarrea.[92] Alcuni pazienti possono manifestare reazioni avverse gravi: pancreatite acuta (i sintomi includono dolore addominale persistente e grave, che talvolta si irradia alla schiena ed è accompagnato da vomito), ipoglicemia, insufficienza renale (che talvolta può richiedere l’emodialisi). Il rischio di ipoglicemia aumenta se il farmaco è usato in combinazione con Sulfoniluree o Insulina.[93][94] Esiste anche un rischio potenziale di carcinoma midollare della tiroide associato all’uso del farmaco.[87]
Lixisenatide [approvato nel 2016]
La Lixisenatide (nome commerciale Lyxumia nell’Unione Europea e Adlyxin negli Stati Uniti e prodotto da Sanofi) è un agonista del recettore GLP-1 iniettabile una volta al giorno per il trattamento del diabete di tipo II.
È stato sintetizzato dalla danese Zealand Pharma A/S;[95] nel 2003 Zealand lo ha concesso in licenza a Sanofi, che ha sviluppato il farmaco.[96] La Lixisenatide è stata approvata dalla Commissione europea nel febbraio 2013.
La Lixisenatide è un peptide composto da 44 aminoacidi, con un gruppo amidico sul suo terminale C.[97]
E’ stata descritta come “des-38-prolina-exendin-4 (Heloderma suspectum)-(1-39)-peptidilpenta-L-lisil-L-lisinamide”, ovvero è derivata dai primi 39 aminoacidi della sequenza del peptide exendin-4, isolato dal veleno del “mostro di Gila”, omettendo la Prolina in posizione 38 e aggiungendo sei residui di Lisina. La sua sequenza completa è:
La Lixisenatide, appartenendo alla classe dei farmaci agonisti del GLP-1, come per i precedentemente trattati composti agisce rallentando lo svuotamento gastrico e aumentando la secrezione di Insulina da parte delle cellule β pancreatiche.
I risultati di una ricerca condotta da McClean PL et al. hanno dimostrato che la Liraglutide e la Lixisenatide sono promettenti come potenziali trattamenti farmacologici della malattia di Alzheimer AD. La Lixisenatide è risultata ugualmente efficace a una dose inferiore rispetto alla Liraglutide in alcuni dei parametri misurati dopo dieci settimane di iniezioni intraperitoneali giornaliere di Liraglutide (2,5 o 25 nmol/kg) o Lixisenatide (1 o 10 nmol/kg) o soluzione fisiologica in topi APP/PS1 a un’età in cui le placche amiloidi si erano già formate. Analizzando la plasticità sinaptica nell’ippocampo, l’LTP è stato fortemente aumentato nei topi APP/PS1 da entrambi i farmaci, con maggiore efficacia con la Lixisenatide. La riduzione del numero di sinapsi osservata nei topi APP/PS1 è stata evitata dai due farmaci. Il carico di placche amiloidi e il carico di placche Congo rosso positivo a nucleo denso nella corteccia sono stati ridotti da entrambi i farmaci a tutte le dosi. Anche la risposta infiammatoria cronica (attivazione microgliale) è stata ridotta da tutti i trattamenti.[98]
Cai HY et al. hanno dimostrato in uno studio che la lixisenatide è in grado di ridurre le placche amiloidi, i grovigli neurofibrillari e la neuroinfiammazione negli ippocampi di topi femmina APP/PS1/tau di 12 mesi; l’attivazione della via di segnalazione PKA-CREB e l’inibizione della p38-MAPK potrebbero essere i meccanismi importanti nella funzione neuroprotettiva della lixisenatide. Pertanto, la lixisenatide potrebbe avere il potenziale per essere sviluppata come nuova terapia per l’AD. [99] Liu Wet al hanno trovato risultati interessanti confrontando exendin-4 (10 nmol/kg), liraglutide (25 nmol/kg) e lixisenatide (10 nmol/kg): è emerso che exendin-4 non ha mostrato effetti protettivi alla dose scelta, mentre sia liraglutide che lixisenatide hanno mostrato effetti nel prevenire la compromissione motoria indotta da MPTP (Rotarod, locomozione in campo aperto, test di catalessi), la riduzione dei livelli di tirosina idrossilasi (TH) (sintesi di dopamina) nella substantia nigra e nei gangli della base, una riduzione della molecola di segnalazione pro-apoptotica BAX e un aumento della molecola di segnalazione anti-apoptotica B-cell lymphoma-2. I risultati precedenti dimostrano che sia la liraglutide che la lixisenatide sono superiori all’exendin-4 ed entrambi i farmaci sono promettenti come nuovo trattamento della malattia di Parkinson.[100]
Un altro studio condotto da Kerry Hunter et al. ha analizzato gli agonisti del recettore GLP-1 liraglutide e lixisenatide. Sono state valutate le cinetiche di attraversamento della barriera ematoencefalica (BBB), l’attivazione del GLP-1R attraverso la misurazione dei livelli di cAMP e gli effetti fisiologici nel cervello sulla proliferazione delle cellule staminali neuronali e sulla neurogenesi. Entrambi i farmaci sono stati in grado di attraversare la BBB. La lixisenatide ha attraversato la BBB a tutte le dosi testate (2,5, 25 o 250 nmol/kg ip.) quando misurate 30 minuti dopo l’iniezione e a 2,5-25 nmol/kg ip. 3 ore dopo l’iniezione. La lixisenatide ha anche aumentato la neurogenesi nel cervello. La liraglutide ha attraversato la BBB a 25 e 250 nmol/kg ip. ma nessun aumento è stato rilevato a 2,5 nmol/kg ip. 30 minuti dopo l’iniezione, e a 250 nmol/kg ip. a 3 ore dopo l’iniezione. Liraglutide e lixisenatide hanno aumentato i livelli di cAMP nel cervello, con lixisenatide più efficace. I risultati precedenti suggeriscono che questi nuovi analoghi dell’incretina attraversano la BBB mostrando attività fisiologica e neurogenesi nel cervello, il che li rende buoni candidati per essere utilizzati come trattamento delle malattie neurodegenerative.[101]
Anche la Lixisenatide è utilizzata come coadiuvante della dieta e dell’esercizio fisico per il trattamento del diabete di tipo II.[97] Nell’Unione Europea il suo uso è limitato all’integrazione della terapia Insulinica.[102][103] Al 2017 non è chiaro se influisca sul rischio di morte di una persona.[104]
Viene fornito in un autoiniettore contenente quattordici dosi e viene iniettato per via sottocutanea.[97]
La Lixisenatide non deve essere utilizzata da persone che hanno problemi di svuotamento gastrico.[97] La Lixisenatide ritarda lo svuotamento gastrico, il che può modificare la velocità con cui altri farmaci assunti oralmente esplicano la loro efficacia.[97]
Dopo la somministrazione sottocutanea nell’uomo, la Lixisenatide mostra una farmacocinetica lineare e un’emivita di eliminazione dipendente dall’assorbimento di 2-3 ore.
La dose iniziale di Lixisenatide è di 10mcg una volta al giorno, per 14 giorni. La dose di mantenimento è successivamente di 20mcg una volta al giorno nell’ora che precede il primo pasto della giornata o il pasto serale.
In circa lo 0,1% dei casi le persone hanno avuto reazioni anafilattiche alla lixisenatide e in circa lo 0,2% dei casi il farmaco ha causato pancreatite.[97] L’uso con insulina o sulfonilurea può causare ipoglicemia.[97] In alcuni casi, persone senza malattie renali hanno avuto lesioni renali acute e in alcune persone con malattie renali esistenti la condizione è peggiorata. Poiché la Lixisenatide è un peptide, le persone possono sviluppare una risposta immunitaria nei suoi confronti che finirà per rendere il farmaco inefficace; le persone che hanno sviluppato anticorpi contro la Lixisenatide tendono ad avere una maggiore infiammazione nel sito di iniezione.[97]
Almeno il 5% delle persone ha avuto nausea, vomito, diarrea, mal di testa o vertigini dopo l’assunzione di Lixisenatide.[97]
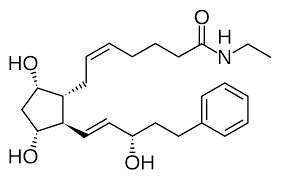
Semaglutide [approvata nel 2017]
La Semaglutide è chimicamente simile al GLP-1 umano.[105-41] Mancano i primi sei aminoacidi del GLP-1.[105] Le sostituzioni sono effettuate nelle posizioni 8 e 34 del GLP-1 (posizioni 2 e 28 della Semaglutide), dove l’Alanina e la Lisina sono sostituite rispettivamente dall’acido 2-aminoisobutirrico e dall’Arginina. La sostituzione dell’Alanina impedisce la degradazione chimica da parte della dipeptidil peptidasi-4.[106] La Lisina in posizione 26 del GLP-1 (posizione 20 del Semaglutide) ha una lunga catena attaccata, che termina con una catena di 17 atomi di carbonio e un gruppo carbossilico.[106] Ciò aumenta il legame del farmaco con le proteine trasportatrici nel sangue (albumina), consentendo una più lunga presenza nella circolazione sanguigna.[106]
L’emivita del Semaglutide nel sangue è di circa sette giorni (165-184 ore).
Come per gli altri incretino-mimetici, la Semaglutide è un agonista del recettore del GLP -1.[107][108][109] Il farmaco riduce i livelli di glucosio nel sangue. Sembra inoltre che aumenti la crescita delle cellule β pancreatiche, responsabili della produzione e del rilascio di Insulina.[110][111] Inoltre, inibisce la produzione di Glucagone, l’ormone che aumenta la glicogenolisi (rilascio dei carboidrati immagazzinati dal fegato) e la Gluconeogenesi (sintesi di nuovo glucosio). Riduce l’assunzione di cibo abbassando l’appetito e rallentando la digestione nello stomaco e suo svuotamento,[112] contribuendo a ridurre il peso corporeo.[113][114]
Effetti di svuotamento gastrico degli agonisti del recettore del glucagone peptide-1 ad azione breve rispetto a quelli ad azione prolungata (GLP-1RA). (A) I GLP-1RA a breve durata d’azione sopprimono lo svuotamento gastrico, prolungando la presenza di cibo nello stomaco e nella parte superiore dell’intestino tenue; il ridotto flusso transpilorico provoca un ritardo nell’assorbimento intestinale del glucosio e una diminuzione della secrezione insulinica postprandiale. I GLP-1RA a breve durata d’azione possono anche sopprimere direttamente la secrezione di glucagone. (B) I GLP-1RA a lunga durata d’azione non influenzano significativamente la motilità gastrica, a causa della tachifilassi. Invece, i GLP-1RA ad azione prolungata esercitano maggiormente il loro effetto attraverso il pancreas, aumentando la secrezione di insulina e inibendo la secrezione di glucagone attraverso il rilascio paracrino di somatostatina. Agendo sul sistema nervoso centrale, sia i GLP-1RA a più breve (A) che a più lunga durata d’azione (B) aumentano la sazietà e possono anche indurre la nausea. Adattato da Meier. Adattato su autorizzazione di Macmillan Publishers Ltd: Nature Reviews Endocrinology 2012;8(12):728-42, copyright 2012.
Nel giugno 2008 è stato avviato uno studio clinico di fase II al fine di esaminare la Semaglutide come terapia per il diabete da somministrare una volta alla settimana, come alternativa ad azione prolungata alla Liraglutide .[115][116] Gli studi clinici sono iniziati nel gennaio 2016 e si sono conclusi nel maggio 2017.[117][118]
Nel giugno 2021, una versione iniettabile a dosaggio più elevato, venduta con il marchio Wegovy, è stata approvata dalla Food and Drug Administration (FDA) statunitense come farmaco anti-obesità per la gestione del peso a lungo termine negli adulti.[119-15] Nel novembre 2021, il Comitato per i Medicinali per Uso Umano (CHMP) dell’Agenzia Europea per i Medicinali (EMA) ha raccomandato di concedere a Novo Nordisk A/S l’autorizzazione all’immissione in commercio di Wegovy[120]. Nel gennaio 2022, Wegovy è stato approvato per uso medico nell’Unione Europea.[121]
Nel gennaio 2023, l’etichetta di Rybelsus è stata aggiornata per indicare che può essere utilizzato come trattamento di prima linea per gli adulti con diabete di tipo 2.[122]
Nel marzo 2021, in uno studio di fase III randomizzato, in doppio cieco, 1.961 adulti con un indice di massa corporea pari o superiore a 30 sono stati assegnati, in un rapporto 2:1, a un trattamento con Semaglutide sottocutaneo una volta alla settimana o placebo, più un intervento sullo stile di vita. Gli studi si sono svolti in 129 siti in 16 Paesi di Asia, Europa, Nord America e Sud America. La variazione percentuale media del peso corporeo alla settimana 68 è stata di -14,9% nel gruppo Semaglutide contro -2,4% con placebo, per una differenza di trattamento stimata di -12,4 punti percentuali (95% CI, da -13,4 a -11,5).[123][124][125][126]
Una revisione dei trattamenti anti-obesità del 2022 ha rilevato che il Semaglutide e la Tirzepatide (che ha un meccanismo d’azione sovrapponibile) erano più promettenti dei precedenti farmaci anti-obesità, anche se meno efficaci della chirurgia bariatrica.[127]
Nel marzo 2023, un funzionario di Novo Nordisk ha dichiarato che i pazienti che utilizzano la Semaglutide per perdere peso possono riacquistare il peso originario entro 5 anni dall’interruzione del trattamento.[128]
Nel marzo 2024, l’FDA ha esteso l’indicazione di Semaglutide (Wegovy) per ridurre il rischio di morte cardiovascolare, infarto e ictus in adulti con malattie cardiovascolari e obesità o sovrappeso. L’efficacia e la sicurezza di questa nuova indicazione sono state studiate in uno studio multinazionale, multicentrico, in doppio cieco, controllato con placebo, che ha assegnato in modo casuale oltre 17.600 partecipanti a ricevere Semaglutide (Wegovy) o placebo.[129] I partecipanti di entrambi i gruppi hanno ricevuto anche un trattamento medico standard (ad es, Semaglutide (Wegovy) ha ridotto significativamente il rischio di eventi cardiovascolari avversi maggiori (morte cardiovascolare, infarto e ictus), che si sono verificati nel 6,5% dei partecipanti che hanno ricevuto Semaglutide (Wegovy) rispetto all’8% dei partecipanti che hanno ricevuto placebo.[129]
Una meta-analisi del 2014 ha rilevato che la Semaglutide può essere efficace nell’abbassare gli enzimi epatici (transaminite) e nel migliorare alcune caratteristiche radiologicamente osservate della malattia epatica steatotica associata a disfunzione metabolica.[130]
Nel luglio 2023, l’Agenzia islandese per i medicinali ha segnalato due casi di pensieri suicidi e un caso di autolesionismo tra i consumatori del farmaco, inducendo a valutare la sicurezza di Ozempic,[131] Wegovy, Saxenda e altri farmaci simili.[132] Nel gennaio 2024, una revisione preliminare condotta dalla FDA ha confermato che non sono state trovate prove che suggeriscano che il farmaco causi pensieri o azioni suicide.[133][134]
La Semaglutide ha dimostrato di poter ridurre l’interesse per il consumo di alcol tra gli utilizzatori. Gli scienziati ipotizzano che il Semaglutide possa influenzare le regioni cerebrali coinvolte nella dipendenza e nella regolazione dell’appetito, sebbene i meccanismi esatti siano ancora in fase di studio. La ricerca sugli animali ha indicato che farmaci simili alla Semaglutide possono ridurre l’assunzione di alcolici.[135]
La Semaglutide e farmaci simili, come la Dulaglutide e la Liraglutide, sono stati utilizzati per trattare il disturbo da alimentazione incontrollata (BED), in quanto possono minimizzare i pensieri ossessivi sul cibo e gli impulsi ad abbuffarsi.[136][137] Alcuni utilizzatori di questi farmaci hanno riferito di aver ridotto in modo significativo quello che è colloquialmente noto come “food noise” (pensieri costanti e inarrestabili di mangiare nonostante non si abbia fisicamente fame), che può essere un fattore di BED.[138][139]
Attualmente, la Semaglutide indicata come coadiuvante della dieta e dell’esercizio fisico per migliorare il controllo glicemico negli adulti con diabete di tipo II.[140][141]
La formulazione a dosi più elevate di Semaglutide è indicata come coadiuvante della dieta e dell’esercizio fisico per la gestione del peso a lungo termine negli adulti con obesità (indice di massa corporea (IMC) iniziale ≥ 30 kg/m2) o in sovrappeso (IMC iniziale ≥ 27 kg/m2) e con almeno una comorbidità correlata al peso.[142]
Nel marzo 2024, la Food and Drug Administration (FDA) statunitense ha ampliato l’indicazione di Semaglutide (Wegovy), in combinazione con una dieta a ridotto contenuto calorico e un aumento dell’attività fisica, per ridurre il rischio di morte cardiovascolare, infarto e ictus in adulti obesi o in sovrappeso con malattie cardiovascolari.[143]
La dose iniziale è di 0,25mg di Semaglutide una volta alla settimana. Dopo 4 settimane, la dose deve essere aumentata a 0,5 mg una volta alla settimana. Dopo almeno 4 settimane con una dose da 0,5 mg una volta alla settimana, la dose può essere aumentata a 1 mg una volta alla settimana per migliorare ulteriormente il controllo glicemico. Dopo almeno 4 settimane con una dose da 1 mg una volta alla settimana, la dose può essere aumentata a 2 mg una volta alla settimana per migliorare ulteriormente il controllo glicemico.
Semaglutide 0,25mg non è una dose di mantenimento. Non sono raccomandate dosi superiori a 2 mg alla settimana.
Quando Ozempic viene aggiunto alla terapia in atto a base di Metformina e/o Tiazolidinedione o dell’ inibitore del cotrasportatore sodio-glucosio (SGLT2), la dose di Metformina e/o Tiazolidinedione o dell’inibitore SGLT2 può essere mantenuta senza variazioni.
Quando Ozempic viene aggiunto alla terapia in atto con Sulfanilurea o con un’insulina, è necessario considerare una riduzione della dose di Sulfanilurea o di insulina per ridurre il rischio di ipoglicemia (vedere paragrafi 4.4 e 4.8).
Non è necessario automonitorare la glicemia per aggiustare la dose di Ozempic. L’auto-monitoraggio della glicemia è necessario per correggere la dose di Sulfanilurea e insulina, in particolare quando si inizia Ozempic e si riduce l’insulina. Si raccomanda un approccio graduale alla riduzione dell’insulina.
Similmente agli altri incretino-mimetici, possibili effetti avversi con l’uso di questo peptide includono nausea, diarrea, vomito, costipazione, dolore addominale, cefalea, affaticamento, indigestione/bruciore di stomaco, vertigini, distensione addominale, eruttazioni, ipoglicemia (basso livello di glucosio nel sangue) nelle persone con diabete di tipo II (ma non limitato ad esse), flatulenza, gastroenterite e malattia da reflusso gastroesofageo (GERD). In passato è stato sospettato di causare pancreatite e può causare gastroparesi e ostruzione intestinale.[144]Tra le persone a cui è stato prescritto un agonista del recettore del GLP-1, lo 0,1% ha ricevuto una diagnosi di gastroparesi. L’1% ha ricevuto una diagnosi di gastroparesi almeno sei mesi dopo, il che equivale a un aumento del 52% del rischio di diagnosi di gastroparesi durante l’assunzione di un farmaco di questa classe.[145] Una meta-analisi del 2019 non ha indicato un rischio significativamente elevato di pancreatite acuta.[146]Secondo il sistema di segnalazione degli eventi avversi dell’FDA (FAERS), più di 150 pazienti che assumevano Ozempic hanno riportato ileo o ostruzioni intestinali dopo l’assunzione del farmaco.[147]
Confronto visivo tra stomaco sano e stomaco con gastroparesi.
L’etichetta dell’FDA statunitense per il Semaglutide contiene un boxed warning per i tumori della tiroide a cellule C nei roditori.[148] Non è noto se il Semaglutide causi tumori della tiroide a cellule C, incluso il carcinoma midollare della tiroide, nell’uomo.[149]
Tirzepatide[approvato nel 2022]
La Tirzepatide è un farmaco antidiabetico utilizzato per il trattamento del diabete di tipo II [150][151][152][153] e per la perdita di peso.[154][155] La Tirzepatide viene somministrata tramite iniezioni sottocutanee.[150][151] Viene venduta con i marchi Mounjaro per il trattamento del diabete,[150] e Zepbound per la perdita di peso.[154] La Tirzepatide è un agonista del recettore del GIP e del GLP-1.[154]
La sintesi della Tirzepatide è stata divulgata per la prima volta nei brevetti depositati da Eli Lilly and Company.[156] Questa utilizza la sintesi standard di peptidi in fase solida, con un gruppo protettivo allilossicarbonilico sulla Lisina in posizione 20 della catena lineare degli amminoacidi, consentendo una serie finale di trasformazioni chimiche in cui l’ammina della catena laterale di tale Lisina viene derivatizzata con il frammento contenente lipidi.
Per questo composto sono stati riportati processi di produzione su larga scala.[157]
La Tirzepatide è un analogo dell’ormone GIP umano con una porzione diacidica grassa C20, utilizzata per ottimizzare l’assorbimento e il metabolismo del composto.[158] La sezione diacidica grassa (acido eicosanedioico) è legata tramite un acido glutammico e due unità di acido (2-(2-aminoetossi)etossico)acetico alla catena laterale del residuo di Lisina. Questa disposizione consente un’emivita molto più lunga, prolungando il tempo tra una dose e l’altra, grazie alla sua elevata affinità con l’albumina.[159]
Quindi, la Tirzepatide è un polipeptide lineare di 39 aminoacidi che è stato modificato chimicamente mediante lipidazione per migliorarne l’assorbimento nelle cellule e la stabilità al metabolismo.[158] Ha completato la sperimentazione di fase III a livello globale nel 2021.[160][161]
La Tirzepatide ha un’affinità maggiore per i recettori GIP rispetto ai recettori GLP-1 e questo comportamento da doppio agonista ha dimostrato di produrre una maggiore riduzione dell’iperglicemia rispetto a un agonista selettivo dei recettori GLP-1.[162] Studi di segnalazione hanno riportato che la Tirzepatide imita le azioni del GIP naturale sul recettore GIP. [Studi di segnalazione hanno riportato che la Tirzepatide imita le azioni del GIP naturale sul recettore del GIP.[163] Tuttavia, sul recettore del GLP-1, la Tirzepatide mostra una predilezione per la generazione di cAMP (un messaggero associato alla regolazione del metabolismo del glicogeno, degli zuccheri e dei lipidi), piuttosto che per il reclutamento della β-arrestina. Questa combinazione di preferenza verso il recettore GIP e di proprietà di segnalazione distinte del GLP-1 suggerisce che questo agonismo distorto aumenta la secrezione di Insulina.[163] È stato riportato che la Tirzepatide aumenta i livelli di adiponectina, un’adipochina coinvolta nella regolazione del metabolismo del glucosio e dei lipidi, con un aumento massimo del 26% rispetto al basale dopo 26 settimane, al dosaggio di 10mg.[162]
GIP e GLP-1: somiglianze e differenze. La GIP è secreta dalle cellule K dell’intestino tenue prossimale (duodeno e digiuno), mentre il GLP-1 è secreto dalle cellule L dell’intestino tenue e crasso (ileo distale e colon), in seguito all’introduzione di carboidrati, trigliceridi, proteine o aminoacidi. Un’importante eccezione è rappresentata dalla glutammina, che è uno stimolatore specifico del GLP-1. GIP e GLP-1 determinano la secrezione di Insulina in modo dipendente dal glucosio. Le azioni del GLP-1 e del GIP sulla secrezione di Glucagone sono diverse: il GLP-1 sopprime il Glucagone durante l’iperglicemia, ma non in presenza di una normale concentrazione di glucosio plasmatico a digiuno, mentre il GIP può stimolare la secrezione di Glucagone a digiuno, durante l’ipoglicemia e l’iperglicemia. Per quanto riguarda il tessuto adiposo, il GLP-1 stimola la lipolisi, mentre il GIP determina un accumulo di grasso corporeo.
Negli studi preliminari finanziati dall’industria che hanno confrontato la Tirzepatide con la Semaglutide, la Tirzepatide ha mostrato un miglioramento minore delle riduzioni (2,01%-2,30% a seconda del dosaggio) nei test dell’emoglobina glicata rispetto alla Semaglutide (1,86%). [164] Una dose di 10mg si è dimostrata efficace anche nel ridurre l’insulino-resistenza, con una riduzione di circa l’8% rispetto al basale, misurata utilizzando l’HOMA2-IR (calcolato con l’Insulina a digiuno).[162] I livelli a digiuno delle proteine che legano l’IGF, come IGFBP1 e IGFBP2, sono aumentati in seguito al trattamento con Tirzepatide, aumentando la sensibilità all’Insulina.[162]
IGFBP1
Una meta-analisi del 2021 ha mostrato che, nell’arco di un anno di utilizzo clinico, la Tirzepatide è risultata superiore a Dulaglutide, Semaglutide, Degludec e Insulina glargine per quanto riguarda l’efficacia glicemica e la riduzione dell’obesità.[165]
In uno studio di fase III, in doppio cieco, randomizzato e controllato, sostenuto da Eli Lilly, adulti non diabetici con un indice di massa corporea pari o superiore a 30, o pari o superiore a 27 e almeno una complicazione correlata al peso, escluso il diabete, sono stati randomizzati a ricevere Tirzepatide sottocutanea una volta alla settimana (5mg, 10mg o 15mg) o placebo. La variazione percentuale media del peso alla settimana 72 è stata di -15,0% (intervallo di confidenza [IC] al 95%, da -15,9 a -14,2) con dosi settimanali di Tirzepatide di 5mg, -19,5% (IC al 95%, da -20,4 a -18,5) con dosi di 10mg e -20,9% (IC al 95%, da -21,8 a -19,9) con dosi di 15mg. La variazione di peso nel gruppo placebo è stata del -3,1% (95% CI, da -4,3 a -1,9).[166][167][168]
La Tirzepatide è stata approvata per uso medico nell’Unione Europea nel settembre 2022.[169][170]
La dose iniziale di Tirzepatide è 2,5mg una volta a settimana. Dopo 4 settimane, la dose deve essere aumentata a 5mg una volta a settimana. Se necessario, è possibile aumentare la dose con incrementi di 2,5mg dopo un minimo di 4 settimane con la dose in uso.
Le dosi di mantenimento raccomandate sono 5mg, 10mg e 15mg.
La dose massima è 15mg una volta a settimana.
Quando Tirzepatide viene aggiunto alla terapia esistente con Metformina e/o inibitore del co- trasportatore di sodio-glucosio 2 (SGLT2i), può essere mantenuta la dose in uso di Metformina e/o SGLT2i.
Quando Tirzepatide viene aggiunto alla terapia esistente con una sulfonilurea e/o Insulina, si può considerare una riduzione della dose di sulfonilurea o Insulina per ridurre il rischio di ipoglicemia. L’automonitoraggio della glicemia è necessario per aggiustare la dose di sulfonilurea e Insulina. Si raccomanda un approccio graduale per la riduzione dell’Insulina.
Gli studi preclinici, di fase I e clinici di fase II hanno indicato che la Tirzepatide presenta effetti avversi simili a quelli di altri agonisti del recettore GLP-1, come visto in precedenza. Questi effetti si verificano in gran parte a livello del tratto gastrointestinale.[171] I più frequentemente osservati sono nausea, diarrea e vomito, la cui incidenza è aumentata con l’entità del dosaggio (cioè la probabilità è maggiore quanto più alta è la dose). Anche il numero di pazienti che hanno interrotto l’assunzione di Tirzepatide è aumentato con l’aumentare del dosaggio: i pazienti che assumevano 15mg avevano un tasso di interruzione del 25% rispetto al 5,1% dei pazienti che assumevano 5mg e all’11,1% di quelli che assumevano Dulaglutide.[172] In misura leggermente minore, i pazienti hanno anche riferito una riduzione dell’appetito.[171] Altri effetti collaterali segnalati sono stati dispepsia, costipazione, dolore addominale, vertigini e ipoglicemia.[173][174]
Uso off-label e “ricreativo”:
Oltre ai loro usi medici, gli agonisti del GLP-1 hanno visto una massiva diffusione in ambito della perdita di peso a fini “estetici” nel Fitness e in parte nel BodyBuilding, resa popolare da influencer e celebrità.[175] I venditori del mercato nero offrono online prodotti non autorizzati che si spacciano per agonisti del GLP-1. Questa pratica è illegale sia negli Stati Uniti che in Europa, ma alcuni acquirenti si rivolgono a rivenditori non autorizzati perché non hanno la possibilità di farsi prescrivere legalmente il farmaco.[176][177][178][179][180] Gli acquirenti, ovviamente, corrono rischi dovuti a farmaci contraffatti o di qualità inferiore venduti da soggetti non autorizzati.[181]
L’uso, le modalità di applicazione e le limitazioni degli incretino-mimetici in campo “cosmetico” saranno riportate nella seconda parte…
Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev. 2007;87:1409–1439. [PubMed] [Google Scholar]
Nauck MA, Homberger E, Siegel EG, et al. Incretin effects of increasing glucose loads in man calculated from venous insulin and C-peptide responses. J Clin Endocrinol Metab. 1986;63:492–498. [PubMed] [Google Scholar]
Brown JC, Mutt V, Pederson RA. Further purification of a polypeptide demonstrating enterogastrone activity. J Physiol. 1970;209:57–64. [PMC free article] [PubMed] [Google Scholar]
Deacon CF, Johnsen AH, Holst JJ. Degradation of glucagon-like peptide-1 by human plasma in vitro yields an N-terminally truncated peptide that is a major endogenous metabolite in vivo. J Clin Endocrinol Metab. 1995;80:952–957. [PubMed] [Google Scholar]
Vilsboll T, Agerso H, Lauritsen T, et al. The elimination rates of intact GIP as well as its primary metabolite, GIP 3–42, are similar in type 2 diabetic patients and healthy subjects. Regul Pept. 2006;137:168–172. [PubMed] [Google Scholar]
Drucker DJ. The biology of incretin hormones. Cell Metab. 2006;3:153–165. [PubMed] [Google Scholar]
Orskov C, Holst JJ, Poulsen SS, Kirkegaard P. Pancreatic and intestinal processing of proglucagon in man. Diabetologia. 1987;30:874–881. [PubMed] [Google Scholar]
Orci L, Bordi C, Unger RH, Perrelet A. Glucagon- and glicentin-producing cells. In: Lefebvre PJ, editor. Glucagon. Berlin: Springer Verlag; 1983. pp. 57–79. [Google Scholar]
Goke R, Larsen PJ, Mikkelsen JD, Sheikh SP. Distribution of GLP-1 binding sites in the rat brain: evidence that exendin-4 is a ligand of brain GLP-1 binding sites. Eur J Neurosci. 1995;7:2294–2300. [PubMed] [Google Scholar]
Flint A, Raben A, Astrup A, Holst JJ. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J Clin Invest. 1998;101:515–520. [PMC free article] [PubMed] [Google Scholar]
Gros R, You X, Baggio LL, et al. Cardiac function in mice lacking the glucagon- like peptide-1 receptor. Endocrinology. 2003;144:2242–2252. [PubMed] [Google Scholar]
Meier JJ, Gethmann A, Gotze O, et al. Glucagon-like peptide 1 abolishes the postprandial rise in triglyceride concentrations and lowers levels of non-esterified fatty acids in humans. Diabetologia. 2006;49:452–458. [PubMed] [Google Scholar]
Carraro-Lacroix LR, Malnic G, Girardi AC. Regulation of Na+/H+ exchanger NHE3 by glucagon-like peptide 1 receptor agonist exendin-4 in renal proximal tubule cells. Am J Physiol Renal Physiol. 2009;297:F1647–F1655. [PubMed] [Google Scholar]
Gault VA, O’Harte FP, Flatt PR. Glucose-dependent insulinotropic polypeptide (GIP): anti-diabetic and anti-obesity potential? Neuropeptides. 2003;37:253–263. [PubMed] [Google Scholar]
Tsukiyama K, Yamada Y, Yamada C, et al. Gastric inhibitory polypeptide as an endogenous factor promoting new bone formation after food ingestion. Mol Endocrinol. 2006;20:1644–1651. [PubMed] [Google Scholar]
Knop FK, Vilsboll T, Hojberg PV, et al. Reduced incretin effect in type 2 diabetes: cause or consequence of the diabetic state? Diabetes. 2007;56:1951–1959. [PubMed] [Google Scholar]
Nauck M, Stockmann F, Ebert R, Creutzfeldt W. Reduced incretin effect in type 2 (non-insulin-dependent) diabetes. Diabetologia. 1986;29:46–52. [PubMed] [Google Scholar]
Bagger JO, Knop FK, Lund A, Vestergaard H, Holst JJ, Vilsboll T. Impaired Regulation of the Incretin Effect in Patients with Type 2 Diabetes Mellitus. Diabetes. 2010;58 (Suppl 1):A369. [PubMed] [Google Scholar]
Vilsboll T, Krarup T, Deacon CF, Madsbad S, Holst JJ. Reduced postprandial concentrations of intact biologically active glucagon-like peptide 1 in type 2 diabetic patients. Diabetes. 2001;50:609–613. [PubMed] [Google Scholar]
Kjems LL, Holst JJ, Volund A, Madsbad S. The influence of GLP-1 on glucose-stimulated insulin secretion: effects on beta-cell sensitivity in type 2 and nondiabetic subjects. Diabetes. 2003;52:380–386. [PubMed] [Google Scholar]
Nauck MA, Heimesaat MM, Orskov C, Holst JJ, Ebert R, Creutzfeldt W. Preserved incretin activity of glucagon-like peptide 1 [7–36 amide] but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J Clin Invest. 1993;91:301–307. [PMC free article] [PubMed] [Google Scholar]
Nauck MA, Kleine N, Orskov C, Holst JJ, Willms B, Creutzfeldt W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7–36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia. 1993;36:741–744. [PubMed] [Google Scholar]
European medicines Agency. European Public Assessment Report Byetta. 2006. EMEA/H/C/698. [Google Scholar]
Amori RE, Lau J, Pittas AG. Efficacy and safety of incretin therapy in type 2 diabetes: systematic review and meta-analysis. JAMA. 2007;298:194–206. [PubMed] [Google Scholar]
Heine RJ, Van Gaal LF, Johns D, Mihm MJ, Widel MH, Brodows RG. Exenatide versus insulin glargine in patients with suboptimally controlled type 2 diabetes: a randomized trial. Ann Intern Med. 2005;143:559–569. [PubMed] [Google Scholar]
Cvetkovic RS, Plosker GL. Exenatide: a review of its use in patients with type 2 diabetes mellitus (as an adjunct to metformin and/or a sulfonylurea) Drugs. 2007;67:935–954. [PubMed] [Google Scholar]
Knudsen LB, Nielsen PF, Huusfeldt PO, et al. Potent derivatives of glucagon-like peptide-1 with pharmacokinetic properties suitable for once daily administration. J Med Chem. 2000;43:1664–1669. [PubMed] [Google Scholar]
Elbrond B, Jakobsen G, Larsen S, et al. Pharmacokinetics, pharmacodynamics, safety, and tolerability of a single-dose of NN2211, a long-acting glucagon-like peptide 1 derivative, in healthy male subjects. Diabetes Care. 2002;25:1398–1404. [PubMed] [Google Scholar]
Damholt B, Golor G, Wierich W, Pedersen P, Ekblom M, Zdravkovic M. An open-label, parallel group study investigating the effects of age and gender on the pharmacokinetics of the once-daily glucagon-like peptide-1 analogue liraglutide. J Clin Pharmacol. 2006;46:635–641. [PubMed] [Google Scholar]
O’Neil PM, Birkenfeld AL, McGowan B, Mosenzon O, Pedersen SD, Wharton S, et al. (August 2018). “Efficacy and safety of semaglutide compared with liraglutide and placebo for weight loss in patients with obesity: a randomised, double-blind, placebo and active controlled, dose-ranging, phase 2 trial”. Lancet. 392 (10148): 637–649. doi:10.1016/S0140-6736(18)31773-2. PMID30122305. S2CID52041320.
Fox CK, Barrientos-Pérez M, Bomberg EM, Dcruz J, Gies I, Harder-Lauridsen NM, et al. (September 2024). “Liraglutide for Children 6 to <12 Years of Age with Obesity – A Randomized Trial”. The New England Journal of Medicine. doi:10.1056/NEJMoa2407379. PMID39258838.
“News details”. Novo Nordisk. Retrieved 12 September 2024.
Garber A, Henry R, Ratner R, et al. Liraglutide versus glimepiride monotherapy for type 2 diabetes (LEAD-3 Mono): a randomised, 52-week, phase III, double-blind, parallel-treatment trial. Lancet. 2009;373:473–481. [PubMed] [Google Scholar]
Marre M, Shaw J, Brandle M, et al. Liraglutide, a once-daily human GLP-1 analogue, added to a sulphonylurea over 26 weeks produces greater improvements in glycaemic and weight control compared with adding rosiglitazone or placebo in subjects with Type 2 diabetes (LEAD-1 SU) Diabet Med. 2009;26:268–278. [PMC free article] [PubMed] [Google Scholar]
Buse JB, Rosenstock J, Sesti G, et al. Liraglutide once a day versus exenatide twice a day for type 2 diabetes: a 26-week randomised, parallel-group, multinational, open-label trial (LEAD-6) Lancet. 2009;374:39–47. [PubMed] [Google Scholar]
Parks M, Rosebraugh C (March 2010). “Weighing risks and benefits of liraglutide–the FDA’s review of a new antidiabetic therapy”. The New England Journal of Medicine. 362 (9): 774–7. doi:10.1056/NEJMp1001578. PMID20164475.
Singh S, Chang HY, Richards TM, Weiner JP, Clark JM, Segal JB (April 2013). “Glucagonlike peptide 1-based therapies and risk of hospitalization for acute pancreatitis in type 2 diabetes mellitus: a population-based matched case-control study”. JAMA Internal Medicine. 173 (7): 534–539. doi:10.1001/jamainternmed.2013.2720. PMID23440284. S2CID425632.
Tibble CA, Cavaiola TS, Henry RR (May 2013). “Longer acting GLP-1 receptor agonists and the potential for improved cardiovascular outcomes: a review of current literature”. Expert Review of Endocrinology & Metabolism. 8 (3): 247–259. doi:10.1586/eem.13.20. PMID30780817. S2CID73313508.
Monami M, Dicembrini I, Nardini C, Fiordelli I, Mannucci E (February 2014). “Glucagon-like peptide-1 receptor agonists and pancreatitis: a meta-analysis of randomized clinical trials”. Diabetes Research and Clinical Practice. 103 (2): 269–275. doi:10.1016/j.diabres.2014.01.010. PMID24485345. S2CID33922845.
Christensen M, Knop FK, Holst JJ, Vilsboll T (August 2009). “Lixisenatide, a novel GLP-1 receptor agonist for the treatment of type 2 diabetes mellitus”. IDrugs. 12 (8): 503–13. PMID19629885.
McClean PL, Hölscher C (November 2014). “Lixisenatide, a drug developed to treat type 2 diabetes, shows neuroprotective effects in a mouse model of Alzheimer’s disease”. Neuropharmacology. 86: 241–58. doi:10.1016/j.neuropharm.2014.07.015. PMID25107586. S2CID24550291.
Cai HY, Yang JT, Wang ZJ, Zhang J, Yang W, Wu MN, Qi JS (January 2018). “Lixisenatide reduces amyloid plaques, neurofibrillary tangles and neuroinflammation in an APP/PS1/tau mouse model of Alzheimer’s disease”. Biochemical and Biophysical Research Communications. 495 (1): 1034–1040. doi:10.1016/j.bbrc.2017.11.114. PMID29175324.
Clinical trial number NCT00696657 for “A Randomised Controlled Clinical Trial in Type 2 Diabetes Comparing Semaglutide to Placebo and Liraglutide” at ClinicalTrials.gov
“Wegovy : Pending EC decision”. European Medicines Agency. 11 November 2021. Archived from the original on 13 November 2021. Retrieved 13 November 2021. Text was copied from this source which is copyright European Medicines Agency. Reproduction is authorized provided the source is acknowledged.
^ Abd El Aziz M, Cahyadi O, Meier JJ, Schmidt WE, Nauck MA (April 2020). “Incretin-based glucose-lowering medications and the risk of acute pancreatitis and malignancies: a meta-analysis based on cardiovascular outcomes trials”. Diabetes, Obesity & Metabolism. 22 (4): 699–704. doi:10.1111/dom.13924. PMID31750601.
US patent 9474780, Bokvist BK, Coskun T, Cummins RC, Alsina-Fernandez J, “GIP and GLP-1 co-agonist compounds”, issued 2016-10-25, assigned to Eli Lilly and Co
Ahangarpour M, Kavianinia I, Harris PW, Brimble MA (January 2021). “Photo-induced radical thiol-ene chemistry: a versatile toolbox for peptide-based drug design”. Chemical Society Reviews. 50 (2). Royal Society of Chemistry: 898–944. doi:10.1039/d0cs00354a. PMID33404559. S2CID230783854.
“Mounjaro EPAR”. European Medicines Agency (EMA). 18 July 2022. Archived from the original on 12 December 2022. Retrieved 2 January 2023. Text was copied from this source which is copyright European Medicines Agency. Reproduction is authorized provided the source is acknowledged.
DISCLAIMER: Il presente articolo è a solo scopo educativo, di intrattenimento e informativo. Non rappresenta in alcun modo una forma di incitamento all’uso/abuso di sostanze dopanti. L’autore ed il sito, per tanto, è esentato da qualsiasi responsabilità dipendente dalla libera scelta individuale.
Introduzione:
L’uso dell’eritropoietina (EPO) per migliorare le prestazioni atletiche, soprattutto nel ciclismo agonistico, è una questione controversa da oltre vent’anni. Nonostante la sua diffusione e le controversie che ne derivano, mancano ancora prove scientifiche solide che ne dimostrino l’efficacia nel migliorare le prestazioni dei ciclisti ben allenati.
Jules Heuberger e il suo team del Centre for Human Drug Research nei Paesi Bassi si sono posti l’obiettivo di affrontare scientificamente proprio questa domanda: L’EPO migliora effettivamente le prestazioni dei ciclisti esperti? Hanno condotto uno studio in doppio cieco, randomizzato e controllato con placebo – considerato il “gold standard” della ricerca scientifica – con dosaggi di EPO che rispecchiano l’uso reale nel ciclismo agonistico. Questo articolo ne condivide i risultati.
Ma partiamo con ordine…
Caratteristiche e azioni dell’Eritropoietina (EPO)
Paul Carnot
Nel 1905, Paul Carnot propose l’idea che un ormone regolasse la produzione di globuli rossi. Dopo aver condotto esperimenti su conigli sottoposti a salasso, Carnot e la sua studentessa laureata Clotilde-Camille Deflandre[1] attribuirono un aumento dei globuli rossi nei conigli trattati a un fattore emotropico chiamato emopoietina. Eva Bonsdorff e Eeva Jalavisto chiamarono la sostanza emopoietica “eritropoietina”. K.R. Reissman e Allan J. Erslev hanno dimostrato che una certa sostanza, circolante nel sangue, è in grado di stimolare la produzione di globuli rossi e di aumentare l’ematocrito. Questa sostanza è stata purificata e confermata come eritropoietina.[2][3]
Nel 1977, Goldwasser e Kung hanno purificato l’EPO.[4] L’EPO pura ha permesso di identificare parzialmente la sequenza aminoacidica e di isolare il gene.[2] L’EPO sintetica è stata utilizzata per la prima volta con successo per correggere l’anemia nel 1987.[5] Nel 1985, Lin et al. hanno isolato il gene dell’eritropoietina umana da una libreria genomica di fagi e l’hanno utilizzato per produrre l’EPO.[6] Nel 1989, la Food and Drug Administration (FDA) statunitense ha approvato l’ormone Epogen per l’uso in alcune anemie.[7][8]
Gregg L. Semenza e Peter J. Ratcliffe hanno studiato il gene dell’EPO e la sua regolazione ossigeno-dipendente. Insieme a William Kaelin Jr. hanno ricevuto il Premio Nobel 2019 per la Fisiologia o la Medicina per la loro scoperta del fattore inducibile dell’ipossia (HIF), che regola il gene dell’EPO, così come altri geni, in risposta all’ipossia.[9]
L’Eritropoietina (/ɪˌrɪθroʊˈpɔɪ. ɪtɪn, -rə-, -pɔɪˈɛtɪn, -ˈiːtɪn/; [10][11][12] EPO), nota anche come eritropoetina, ematopoietina o emopoietina, è una citochina glicoproteica secreta principalmente dai reni in risposta all’ipossia cellulare; stimola la produzione di globuli rossi (eritropoiesi) nel midollo osseo. Bassi livelli di EPO (circa 10mU/mL) sono costantemente secreti in quantità sufficiente a compensare il normale ricambio dei globuli rossi. Le cause comuni di ipossia cellulare che determinano livelli elevati di EPO (fino a 10.000mU/mL) comprendono qualsiasi anemia e l’ipossiemia dovuta a malattie polmonari croniche e alla bocca.
L’Eritropoietina è prodotta dai fibroblasti interstiziali del rene in stretta associazione con il capillare peritubulare e il tubulo contorto prossimale. Viene prodotta anche nelle cellule perisinusoidali del fegato. La produzione epatica predomina nel periodo fetale e perinatale; la produzione renale predomina nell’età adulta. È omologa della trombopoietina.
rhEPO
L’Eritropoietina esogena, l’Eritropoietina umana da DNA ricombinante (rhEPO), viene prodotta con la tecnologia del DNA ricombinante in coltura cellulare e viene chiamata collettivamente agenti stimolanti l’eritropoiesi (ESA): due esempi sono l’epoetina alfa e l’epoetina beta. Gli ESA sono utilizzati nel trattamento dell’anemia nella malattia renale cronica, dell’anemia nella mielodisplasia e dell’anemia da chemioterapia oncologica. I rischi della terapia includono morte, infarto miocardico, ictus, tromboembolismo venoso e recidiva del tumore. Il rischio aumenta quando il trattamento con EPO aumenta i livelli di emoglobina oltre 11g/dL fino a 12g/dL: questo è da evitare.
GATA2
Come accennato, i livelli di eritropoietina nel sangue sono piuttosto bassi in assenza di anemia, circa 10mU/mL. Tuttavia, in caso di stress ipossico, la produzione di EPO può aumentare fino a 1000 volte, raggiungendo 10.000mU/mL di sangue. Negli adulti, l’EPO è sintetizzata principalmente dalle cellule interstiziali nel letto capillare peritubulare della corteccia renale, con quantità aggiuntive prodotte nel fegato,[13][14][15] e nei periciti del cervello.[16] Si ritiene che la regolazione si basi su un meccanismo di feedback che misura l’ossigenazione del sangue e la disponibilità di ferro.[17] I fattori di trascrizione per l’EPO sintetizzati costitutivamente, noti come fattori inducibili dall’ipossia, sono idrossilati e digeriti proteosomicamente in presenza di ossigeno e ferro. Durante la normossia, GATA2 inibisce la regione promotrice dell’EPO. I livelli di GATA2 diminuiscono durante l’ipossia e permettono di promuovere la produzione di EPO.[18]
PGC-1α
La produzione di eritropoietina può essere indotta da HIF-2α e da PGC-1α.[19] L’eritropoietina attiva anche questi fattori, dando luogo a un ciclo di feedback positivo.[19]
È stato dimostrato che l’eritropoietina esercita i suoi effetti legandosi al recettore dell’eritropoietina (EpoR).[20][21] L’EPO si lega al recettore dell’eritropoietina sulla superficie dei progenitori dei globuli rossi e attiva una cascata di segnalazione JAK2. Questo avvia le vie di STAT5, PIK3 e Ras MAPK. Ciò determina la differenziazione, la sopravvivenza e la proliferazione delle cellule eritroidi.[22] Vengono inoltre espressi SOCS1, SOCS3 e CIS, che agiscono come regolatori negativi del segnale delle citochine.[23]
L’espressione del recettore dell’eritropoietina ad alto livello è localizzata nelle cellule progenitrici eritroidi. Sebbene sia stato riferito che i recettori dell’EPO si trovano in una serie di altri tessuti, come il cuore, il muscolo, il rene e il tessuto nervoso periferico/centrale, questi risultati sono confusi dalla non specificità dei reagenti, come gli anticorpi anti-EpoR.[24] In esperimenti controllati, un recettore funzionale dell’EPO non viene rilevato in questi tessuti.[25] Nel flusso sanguigno, gli stessi globuli rossi non esprimono il recettore dell’eritropoietina, quindi non possono rispondere all’EPO. Tuttavia, è stata segnalata una dipendenza indiretta della longevità dei globuli rossi nel sangue dai livelli plasmatici di eritropoietina, un processo definito neocitolisi.[26] Inoltre, vi sono prove inconfutabili che l’espressione del recettore dell’EPO è regolata in modo elevato nelle lesioni cerebrali.[27]
L’eritropoietina è un ormone essenziale per la produzione di globuli rossi. Senza di essa, l’eritropoiesi definitiva non ha luogo. In condizioni di ipossia, il rene produrrà e secernerà eritropoietina per aumentare la produzione di globuli rossi, mirando alle sottopopolazioni di CFU-E, proeritroblasti ed eritroblasti basofili nella differenziazione. L’eritropoietina ha un effetto primario sui progenitori e sui precursori dei globuli rossi (che si trovano nel midollo osseo degli esseri umani), promuovendo la loro sopravvivenza attraverso la protezione di queste cellule dall’apoptosi, o morte cellulare.
IL-6
L’eritropoietina è il fattore eritropoietico primario che coopera con vari altri fattori di crescita (ad esempio, IL-3, IL-6, glucocorticoidi e SCF) coinvolti nello sviluppo della linea eritroide da progenitori multipotenti. Le cellule eritroidi a formazione di unità di esplosione (BFU-E) iniziano a esprimere il recettore per l’eritropoietina e sono sensibili all’eritropoietina. Lo stadio successivo, l’unità formante colonie eritroidi (CFU-E), esprime la massima densità di recettori per l’eritropoietina ed è completamente dipendente dall’eritropoietina per l’ulteriore differenziazione. Anche i precursori dei globuli rossi, i proeritroblasti e gli eritroblasti basofili, esprimono il recettore dell’eritropoietina e ne sono quindi influenzati.
È stato riferito che l’eritropoietina ha una serie di azioni che vanno oltre la stimolazione dell’eritropoiesi, tra cui l’ipertensione dipendente dalla vasocostrizione, la stimolazione dell’angiogenesi e la promozione della sopravvivenza cellulare attraverso l’attivazione dei recettori dell’EPO, con conseguenti effetti anti-apoptotici sui tessuti ischemici. Questa proposta è tuttavia controversa, in quanto numerosi studi non hanno dimostrato alcun effetto.[28] È inoltre incoerente con i bassi livelli di recettori dell’EPO su queste cellule. Gli studi clinici condotti su esseri umani con tessuti ischemici cardiaci, neurali e renali non hanno dimostrato gli stessi benefici osservati negli animali. Inoltre, alcuni studi hanno dimostrato un effetto neuroprotettivo sulla neuropatia diabetica, ma questi dati non sono stati confermati da studi clinici condotti sui nervi peroneo profondo, peroneo superficiale, tibiale e surale.[29]
Come sappiamo, le eritropoietine disponibili come agenti terapeutici sono prodotte con la tecnologia del DNA ricombinante in coltura cellulare e comprendono Epogen/Procrit (epoetina alfa) e Aranesp (darbepoetina alfa); sono utilizzate per il trattamento dell’anemia derivante da malattie renali croniche,[30] dell’anemia indotta dalla chemioterapia in pazienti affetti da cancro, da malattie infiammatorie intestinali (morbo di Crohn e colite ulcerosa)[31] e da mielodisplasia dovuta al trattamento del cancro (chemioterapia e radiazioni). I foglietti illustrativi includono avvertenze relative all’aumento del rischio di morte, infarto del miocardio, ictus, tromboembolismo venoso e recidiva del tumore, in particolare quando viene utilizzato per aumentare i livelli di emoglobina a più di 11g/dL – 12g/dL.[32]
L’EPO è altamente glicosilata (40% del peso molecolare totale), con un’emivita nel sangue di circa 5 ore. L’emivita dell’EPO può variare tra le versioni endogene e quelle ricombinanti. L’ulteriore glicosilazione o altre alterazioni dell’EPO attraverso la tecnologia ricombinante hanno portato a un aumento della stabilità dell’EPO nel sangue (richiedendo così iniezioni meno frequenti).
EPO come PEDs
Come farmaco per il miglioramento delle prestazioni, l’EPO è stato vietato dall’inizio degli anni ’90, ma un primo test non è stato disponibile fino alle Olimpiadi estive del 2000. Prima che questo test fosse disponibile, alcuni atleti sono stati sanzionati dopo aver confessato di aver fatto uso di EPO, ad esempio nel caso Festina, quando fu trovata un’auto con prodotti dopanti per la squadra ciclistica Festina.
A questo punto, però, è necessario comprendere le questioni di fondo relative all’uso dell’EPO nello sport e, in particolare, il suo impatto sulle prestazioni ciclistiche. Molti atleti agonisti e osservatori ritengono che l’EPO migliori le prestazioni atletiche aumentando la produzione di globuli rossi e l’apporto di ossigeno ai muscoli. Come abbiamo visto precedentemente, infatti, la rHuEPO clinica viene utilizzata per trattare l’anemia, aumentando il numero di globuli rossi nei pazienti, il che implica che potrebbe fare lo stesso per gli atleti. Questa logica ha contribuito al suo status di sostanza vietata dalla maggior parte delle agenzie antidoping, a partire proprio dal Comitato Olimpico Internazionale nel 1990.
Tuttavia, nonostante questa convinzione diffusa e il suo uso illecito tra gli atleti, è difficile trovare prove scientifiche concrete a sostegno dei suoi effetti di miglioramento delle prestazioni nei ciclisti ben allenati.
Mentre molti si concentrano sul potenziale dell’EPO di aumentare la massa dei globuli rossi e la capacità di trasportare ossigeno, altri, più previdenti, ritengono che il suo uso improprio possa portare a gravi effetti negativi. Sono state rilasciate innumerevoli dichiarazioni relative a complicazioni cardiovascolari come ipertensione, trombosi e aumento del rischio di ictus, nonché a disturbi ematologici come la policitemia.
Inoltre, alcuni avvertono che la somministrazione o il dosaggio improprio dell’EPO possono provocare uno squilibrio nella produzione di globuli rossi, portando a livelli pericolosi di ematocrito e viscosità, che a loro volta possono aumentare il rischio di coaguli di sangue e altri eventi cardiovascolari. Tuttavia, come abbiamo detto in questa serie, i presunti pericoli dell’EPO nel ciclismo di prestazione sono stati oggetto di analisi e studi, che hanno dimostrato che le agenzie sportive si sono spesso basate su affermazioni non comprovate.
Sebbene alcuni considerino l’EPO tra i migliori farmaci per il miglioramento delle prestazioni nel ciclismo, esistono studi che ne confermano la potenziale pericolosità.
Sebbene l’entità dei rischi associati all’uso dell’eritropoietina (EPO) negli atleti rimanga incerta, i dati provenienti da revisioni della letteratura e da studi condotti su soggetti sani e allenati forniscono indicazioni sui potenziali pericoli.
Pressione sanguigna sistolica
In uno studio, i ricercatori hanno notato un notevole aumento della pressione arteriosa sistolica, sia a riposo che durante l’esercizio submassimale, in seguito alla somministrazione di EPO. L’aumento della pressione arteriosa può predisporre gli atleti a complicazioni cardiovascolari, tra cui l’ipertensione e l’aumento del rischio di eventi trombotici.
Eventi trombotici
Le evidenze delle revisioni della letteratura evidenziano anche un’elevata incidenza di eventi trombotici nei pazienti trattati con dosi elevate di rHuEPO rispetto a quelli che ricevono un placebo. Tuttavia, è importante notare che questi studi hanno tipicamente utilizzato dosi significativamente superiori a quelle comunemente utilizzate negli studi sulle prestazioni di resistenza. Fattori come l’aumento della viscosità del sangue, l’aumento della coagulazione, l’attivazione endoteliale, la reattività piastrinica e l’infiammazione possono contribuire a questi eventi avversi.
Impatto con l’esercizio fisico
È inoltre importante considerare i cambiamenti fisiologici indotti dall’esercizio fisico acuto. Questi impatti includono riduzioni del volume plasmatico e del volume sanguigno accompagnate da un aumento dell’ematocrito, potenzialmente in grado di esacerbare il rischio di eventi trombotici negli atleti di resistenza, soprattutto in condizioni di disidratazione e ipertermia.
Non conclusività delle prove
Sebbene questi risultati suggeriscano potenziali pericoli associati all’uso di EPO negli atleti, mancano prove conclusive. Non è possibile trarre conclusioni definitive senza studi di ricerca ben progettati che analizzino specificamente gli effetti dell’EPO sulle prestazioni e sulla sicurezza dei ciclisti d’élite.
Heuberger et al. [33] hanno progettato uno studio randomizzato, in doppio cieco e controllato con placebo per affrontare le incertezze che circondano gli effetti e la sicurezza dell’uso dell’EPO in ciclisti ben allenati. Questo studio si propone di analizzare in modo rigoroso l’impatto di NeoRecormon, una forma di eritropoietina umana ricombinante sintetica, sulle prestazioni e sui parametri di sicurezza nei ciclisti d’élite.
Lo studio si è concentrato sugli effetti su ciclisti ben allenati, comprendendo 48 soggetti sani e stabili dal punto di vista medico, reclutati tramite pubblicità sui media e associazioni ciclistiche. Questi soggetti sono stati selezionati per rappresentare l’élite del ciclismo, con un alto livello di forma fisica ed esperienza di allenamento. Il protocollo dello studio prevedeva un periodo di 8 settimane durante il quale i partecipanti sarebbero stati assegnati in modo casuale a ricevere NeoRecormon o un placebo.
NeoRecormon è stato somministrato a dosi di 2000, 5000 o tra 6000 e 10.000 UI alla settimana. L’obiettivo era quello di raggiungere l’intervallo prefissato, con aggiustamenti necessari in base ai risultati dell’emoglobina (Hb) o dell’ematocrito (Ht).
Lo studio ha avuto una durata totale di 129 giorni, con un periodo di trattamento di 8 settimane. Questo lasso di tempo ha permesso di valutare in modo completo le prestazioni e la sicurezza dopo la somministrazione di NeoRecormon. Prima e dopo il periodo di intervento sono state condotte valutazioni dettagliate delle metriche di prestazione, tra cui resistenza, potenza e utilizzo dell’ossigeno.
Obiettivi primari
L’obiettivo dello studio era esplorare gli effetti di NeoRecormon sulle prestazioni ciclistiche in ciclisti ben allenati. L’obiettivo è stato raggiunto con diversi mezzi, tra cui valutazioni separate delle prestazioni in test da sforzo, in condizioni di gara e la misurazione dei marcatori ematologici tramite il Passaporto Biologico dell’Atleta. Sono state effettuate anche misurazioni del flusso sanguigno per valutare le risposte fisiologiche alla somministrazione di EPO.
Obiettivi secondari
Gli obiettivi secondari comprendevano un’ulteriore esplorazione degli effetti del NeoRecormon in un contesto di gara su strada, per facilitare la determinazione della sua capacità tra gli integratori e i farmaci che migliorano le prestazioni ciclistiche. Altri obiettivi comprendevano una valutazione completa del suo profilo di sicurezza in ciclisti ben allenati e una valutazione dei metodi di rilevamento del doping per l’uso del NeoRecormon.
I risultati dello studio
La valutazione della sicurezza del trattamento con rHuEPO in ciclisti ben allenati ha rivelato risultati rassicuranti. I segni vitali come il peso, la frequenza cardiaca e la pressione sanguigna erano simili tra i due gruppi di trattamento, mentre gli eventi avversi osservati erano da lievi a moderati e comparabili tra i gruppi. In particolare, non sono stati segnalati eventi avversi gravi (di grado 3 o peggiore) in nessuno dei due gruppi.
Mentre alcuni marcatori della funzione endoteliale hanno mostrato un leggero aumento con il trattamento con rHuEPO, suggerendo un potenziale aumento della trombogenicità, non ci sono stati segni clinici di effetti avversi associati alla somministrazione di rHuEPO.
In termini di miglioramento delle prestazioni, il trattamento con rHuEPO ha portato a miglioramenti nei test di laboratorio di esercizio massimale, con conseguente aumento della resistenza e delle prestazioni. Tuttavia, i suoi effetti sui test di esercizio submassimale e sulle prestazioni nelle corse su strada non sono stati rilevabili. Nel complesso, i risultati dello studio sono stati meno pronunciati rispetto alle affermazioni spesso riportate nella letteratura popolare e nei resoconti aneddotici. Ciò sottolinea l’importanza di una ricerca basata sull’evidenza per valutare l’efficacia e la sicurezza di interventi di miglioramento delle prestazioni come l’EPO.
L’assenza di effetti significativi di miglioramento delle prestazioni osservati nello studio può essere attribuita a diversi fattori:
Differenze contestuali
Gli effetti dell’EPO possono essere più pronunciati in eventi a più tappe come il Tour de France, in cui la resistenza e il recupero giocano un ruolo critico, rispetto a gare di un solo giorno come quella del Mont Ventoux inclusa nello studio. La durata e l’intensità degli eventi possono influenzare la rilevabilità degli effetti dell’EPO.
Dimensione dello studio e potenza statistica
La dimensione del campione dello studio potrebbe essere stata insufficiente per rilevare sottili differenze nei risultati delle prestazioni, in particolare nel contesto della corsa su strada. La complessità della misurazione delle prestazioni nelle competizioni ciclistiche reali e la variabilità inerente alle prestazioni dei singoli atleti possono aver limitato la potenza statistica dello studio nel rilevare effetti significativi.
Entità ridotta dei benefici
È possibile che i benefici della rHuEPO sulle prestazioni, pur essendo presenti, siano minori di quanto si tende a sostenere o a credere, il che li rende difficili da distinguere in assenza di campioni più ampi o di tecniche di misurazione più sensibili.
Conclusioni sulla sicurezza dell’EPO e sul miglioramento delle prestazioni
Sulla base dei risultati dello studio controllato in doppio cieco, la sicurezza dell’uso dell’EPO per migliorare le prestazioni ciclistiche rimane un argomento di dibattito e di cautela. Sebbene non siano state osservate differenze significative negli eventi avversi tra i gruppi EPO e placebo, lo studio ha rivelato un aumento preoccupante dei marcatori endoteliali, in particolare E-selectina e P-selectina, associati a trombogenicità e infiammazione.
Questi risultati suggeriscono un potenziale aumento del rischio cardiovascolare associato al trattamento con rHuEPO, che potrebbe non essere stato adeguatamente colto a causa della bassa incidenza di eventi cardiovascolari negli atleti sani. La limitata potenza dello studio nel rilevare tali rischi sottolinea la necessità di ulteriori ricerche con campioni di dimensioni maggiori e periodi di follow-up più lunghi.
Dato l’uso diffuso e non controllato della rHuEPO tra gli atleti, non si può escludere il rischio potenziale di eventi cardiovascolari. Pertanto, sebbene l’EPO possa offrire benefici per le prestazioni, il suo uso deve essere affrontato con cautela e gli atleti devono essere consapevoli dei potenziali rischi associati alla sua somministrazione.
Per il futuro, è indispensabile condurre studi su larga scala con periodi di follow-up prolungati per valutare in modo completo la sicurezza e l’efficacia dell’uso dell’EPO nei ciclisti ben allenati. È necessario implementare politiche e regolamenti basati sull’evidenza per mitigare i potenziali rischi associati all’abuso di sostanze che migliorano le prestazioni, evitando consapevolmente di usare iperboli o esagerazioni per demonizzare una sostanza.
Comprendere il protocollo di dosaggio dell’EPO
I ricercatori del CHDR hanno progettato e attuato con cura un protocollo di dosaggio dell’EPO [33], con l’obiettivo di replicare le pratiche note nel ciclismo professionistico, garantendo al contempo la sicurezza dei partecipanti e il rispetto degli standard etici. I partecipanti assegnati al gruppo rHuEPO (eritropoietina umana ricombinante) hanno ricevuto otto dosi totali durante il periodo di studio.
Il regime di dosaggio prevedeva la somministrazione di una dose media di rHuEPO, sotto forma di NeoRecormon, di 5000 UI per partecipante a settimana durante le prime 4 settimane dello studio. Successivamente, la dose è stata aumentata a 7000 UI a settimana per le restanti 4 settimane. Per mitigare il rischio di parametri ematologici eccessivi, ai partecipanti che hanno superato un aumento del 15% dell’emoglobina rispetto al basale o che hanno raggiunto una concentrazione di ematocrito superiore al 52% sono state somministrate iniezioni di placebo in cinque occasioni.
La dose media di rHuEPO somministrata per tutto il periodo di studio è stata di 48.000 UI, pari a una media di 6000 UI a settimana. Questa strategia di dosaggio ha determinato un aumento sostanziale della concentrazione di emoglobina, con un incremento medio del 12% fino a 10,2 mmol/L, e un aumento del 16% dei livelli di ematocrito, che ha raggiunto il 50%. Al contrario, i partecipanti al gruppo placebo hanno mostrato concentrazioni di emoglobina ed ematocrito relativamente stabili per tutta la durata dello studio.
I diari dei partecipanti sono stati tenuti con una documentazione meticolosa, contribuendo a confermare l’aderenza al regime di integrazione prescritto per tutto il periodo dello studio, garantendo coerenza e affidabilità nella somministrazione delle dosi di rHuEPO.
In seguito, approfondiremo il razionale di questa strategia di dosaggio, le sue implicazioni per le prestazioni ciclistiche e il contesto più ampio delle linee guida per il dosaggio dell’EPO sotto controllo medico.
Un esempio di protocollo di 8 settimane
Un esempio di protocollo di 8 settimane per la somministrazione di NeoRecormon potrebbe essere il seguente:
Settimana 1-4: Dosaggio di NeoRecormon da 2000 a 10.000 UI alla settimana (aggiustato in base alle misurazioni di Hb e Ht).
Settimana 5-8: continui aggiustamenti del dosaggio di NeoRecormon secondo le necessità, con regolare monitoraggio dei parametri ematologici.
Integrazione giornaliera: 50 mg di vitamina C e 200 mg di ferro per ottimizzare l’assorbimento e la salute generale.
I dettagli del dosaggio di NeoRecormon spiegati
Il farmaco sperimentale utilizzato nello studio era l’eritropoietina umana ricombinante (rHuEPO) NeoRecormon, contenente il principio attivo Epoëtine beta. NeoRecormon è stato somministrato per via sottocutanea (nel tessuto adiposo sotto la pelle) ai partecipanti rispettando uno schema di dosaggio attentamente studiato, nella speranza di ottimizzare i parametri ematologici riducendo al minimo i rischi potenziali.
Il protocollo di dosaggio di NeoRecormon si è basato su uno schema decisionale completo, ideato per guidare i ricercatori nell’aggiustamento del dosaggio in base alle caratteristiche e alle risposte dei singoli partecipanti, in particolare alla concentrazione di emoglobina (Hb) e ai livelli di ematocrito (Ht). Questo albero decisionale è stato un aiuto visivo per facilitare il processo decisionale in tempo reale per quanto riguarda gli aggiustamenti durante il periodo di trattamento di 8 settimane.
L’albero decisionale delineava vari scenari basati sulle misurazioni di Hb e Ht prima di ogni somministrazione di NeoRecormon o placebo. Se l’Ht di un partecipante superava il 52%, indicando un alto rischio di complicazioni ematologiche, la somministrazione del dosaggio veniva prontamente interrotta per ridurre i rischi potenziali. Al contrario, se i livelli di Ht erano inferiori al 52%, l’albero decisionale indirizzava i ricercatori a valutare la concentrazione di Hb del partecipante per determinare il dosaggio appropriato di NeoRecormon.
Ecco le fasi dell’albero decisionale in dettaglio:
Se il livello di Ht raggiungeva un valore superiore o uguale al 52%, il doping veniva interrotto. Se i livelli di Hb scendono al di sotto di circa 1,15x, anche il dosaggio si interrompe.
Se i livelli di Hb erano superiori o uguali a 1,10x, il dosaggio sarebbe rimasto a 2000IU/settimana.
Se la situazione non si fosse evoluta prima di 5 settimane, il dosaggio sarebbe arrivato a 5000IU/settimana; se la situazione non si fosse evoluta dopo 5 settimane o più, il dosaggio sarebbe stato aumentato a più o uguale a 6000IU/settimana, con un dosaggio massimo di 10.000IU/settimana.
Il dosaggio di NeoRecormon variava da 2.000 a 10.000 UI alla settimana, con la flessibilità di aggiustare questo intervallo in base alla risposta di ciascun partecipante al trattamento. Questo intervallo di dosaggio è stato concepito per garantire l’efficacia nell’innalzare i livelli di Hb e Ht all’interno dell’intervallo target, riducendo al minimo il rischio di effetti avversi.
Valutazione dei benefici e dei rischi
NeoRecormon è un farmaco registrato con un profilo di sicurezza noto, che lo rende adatto all’uso in contesti di ricerca. Tuttavia, i ricercatori hanno riconosciuto la possibilità di effetti collaterali, tra cui reazioni anafilattoidi, anche se con un basso tasso di incidenza di ≤1 su 10.000 casi. Per questo motivo, tutte le somministrazioni del farmaco in studio sono state condotte in un ambiente clinico sotto stretta supervisione medica, contribuendo a mitigare i rischi.
Metodologia di monitoraggio
I partecipanti sono stati monitorati attentamente per tutta la durata dello studio e le loro condizioni mediche sono state valutate regolarmente per garantire la sicurezza e il benessere. Gli aggiustamenti del dosaggio sono stati effettuati, se necessario, in base ai parametri ematologici e alle risposte individuali al trattamento, guidati dall’algoritmo dell’albero decisionale.
Rispettando il protocollo di dosaggio prescritto, seguendo accuratamente l’albero decisionale e implementando rigorose misure di sicurezza, i ricercatori miravano a ottimizzare l’efficacia e la sicurezza della somministrazione di NeoRecormon nel migliorare le prestazioni ciclistiche dei partecipanti.
Valutazione della sicurezza e del rischio dell’uso di NeoRecormon
NeoRecormon, un’eritropoietina umana ricombinante (rHuEPO), è comunemente utilizzato in ambito clinico per il trattamento di diverse condizioni mediche, tra cui l’anemia associata a malattie renali croniche e alla chemioterapia del cancro. Il suo profilo di sicurezza è stato ampiamente studiato, con linee guida di dosaggio ben stabilite per garantire l’efficacia (in questi contesti medici) riducendo al minimo i rischi potenziali.
Il rischio più comunemente associato a NeoRecormon riguarda il suo potenziale aumento dei livelli di ematocrito e di emoglobina che, se elevati eccessivamente, possono portare a complicazioni come trombosi, ipertensione ed eventi cardiovascolari. Tuttavia, se somministrato entro gli intervalli di dosaggio raccomandati, NeoRecormon è generalmente considerato sicuro ed efficace per gli scopi medici previsti. La ricerca ha anche dimostrato che l’aumento delle prestazioni dell’EPO probabilmente non è così pericoloso come le organizzazioni antidoping vorrebbero far credere al pubblico.
Confronto con un programma di pre-donazione di sangue autologo
Gli effetti dell’uso di NeoRecormon nei pazienti che partecipano a programmi di pre-donazione di sangue autologo assomigliano molto a quelli dei volontari sani. In entrambi gli scenari, è stato riconosciuto un aumento della produzione di globuli rossi. Il Riassunto delle Caratteristiche del Prodotto (SmPC) per NeoRecormon in un programma di pre-donazione di sangue autologo specifica una dose massima raccomandata di 1200 UI/kg a settimana per somministrazione sottocutanea, equivalente a 90.000 UI per un individuo di 75 kg.
In questo protocollo di dosaggio dell’EPO, che prevede dosi pianificate da 2.000 a 10.000 UI alla settimana, i dosaggi di NeoRecormon sono ben al di sotto del limite massimo raccomandato. Pertanto, il rischio associato alla somministrazione di NeoRecormon nello studio è considerato piccolo e accettabile.
Integrazione obbligatoria
Per sostenere gli effetti fisiologici della somministrazione di NeoRecormon e ridurre i rischi potenziali, ai partecipanti è stata prescritta un’integrazione giornaliera obbligatoria di 50 mg di vitamina C (acido ascorbico) e 200 mg di ferro (ferrofumarato) per tutto il periodo di trattamento di 8 settimane. Questi integratori contribuiscono a ottimizzare il metabolismo del ferro, l’eritropoiesi e la salute generale durante la terapia con EPO.
Supervisione medica e misure di Harm Reduction
La supervisione medica durante la somministrazione di NeoRecormon controlla da vicino i parametri ematologici, i segni vitali e lo stato di salute generale dei partecipanti, con controlli dettagliati eseguiti prima di ogni dose, il che significa 8 controlli durante tutto il processo. I partecipanti hanno sempre ricevuto le iniezioni in un ambiente clinico sotto la diretta supervisione di un professionista sanitario, garantendo una corretta somministrazione e la gestione immediata di eventuali reazioni avverse.
Le misure di Harm Reduction includono:
Valutazione settimanale dei parametri ematologici per individuare e prevenire aumenti eccessivi dei livelli di Hb e Ht.
Fornitura di un’integrazione obbligatoria di vitamina C e ferro per supportare l’eritropoiesi e minimizzare il rischio di carenza di ferro.
Interruzione tempestiva del dosaggio di NeoRecormon se i parametri ematologici superano soglie predefinite o se si verificano reazioni avverse.
Esami del sangue effettuati
Durante lo studio, i partecipanti sono stati inoltre sottoposti a esami del sangue settimanali per monitorare i parametri ematologici, tra cui:
Concentrazione di emoglobina
Livelli di ematocrito
Conteggio dei globuli rossi
Conteggio delle piastrine
Profilo di coagulazione
Questi esami del sangue hanno fornito informazioni cruciali per valutare la sicurezza e l’efficacia della somministrazione di NeoRecormon e per guidare gli aggiustamenti del dosaggio, se necessario.
Implicazioni per i medici dello sport e per i pazienti che praticano l’automedicazione
Per i medici sportivi che si trovano di fronte a pazienti che si curano da soli con l’EPO o che ne considerano l’uso per migliorare le prestazioni, questa ricerca fornisce indicazioni preziose su ciò che costituisce un protocollo di dosaggio supervisionato da un medico con le giuste misure di sicurezza. Lo studio sottolinea l’importanza di livelli di dosaggio accurati, di un monitoraggio regolare dei parametri ematologici e dell’aderenza ai regimi di integrazione per ridurre i rischi potenziali e ottimizzare i benefici.
Criteri per i dosaggi iniziali, per le modifiche o per l’interruzione del trattamento
I criteri che hanno portato alla direzione degli aggiustamenti sono stati:
Parametri ematologici di base (livelli di Hb e Ht)
Risposta ai dosaggi iniziali (per esempio, tasso di aumento di Hb e Ht)
Valori di soglia per le concentrazioni di Hb e Ht per evitare aumenti eccessivi
Comparsa di reazioni avverse o di sintomi suggestivi di complicazioni ematologiche
Questi criteri hanno guidato i ricercatori nell’individualizzazione dei regimi di trattamento e nel garantire la sicurezza dei partecipanti durante lo studio.
Rischi ed effetti collaterali monitorati
I ricercatori hanno monitorato attentamente i partecipanti per individuare potenziali rischi ed effetti collaterali, tra cui:
Aumento eccessivo dei livelli di Hb e Ht con conseguenti complicazioni ematologiche (ad es. trombosi, ipertensione).
Reazioni anafilattoidi o risposte allergiche alle iniezioni di NeoRecormon.
Carenza di ferro o disturbi del metabolismo del ferro dovuti all’aumento dell’eritropoiesi.
La valutazione regolare dei parametri ematologici e la valutazione clinica hanno permesso di individuare e gestire precocemente gli eventi avversi, riducendo al minimo il loro impatto sulla sicurezza e sul benessere dei partecipanti.
Conclusioni:
Uno studio del 2007 ha dimostrato che l’EPO ha un effetto significativo sulle prestazioni di esercizio.[chiarisci][https://link.springer.com/article/] Uno studio del 2017 ha dimostrato che allo sforzo submassimale gli effetti dell’EPO non erano distinguibili da quelli del placebo. Si afferma che “[Allo] sforzo [submassimale]…[la potenza media] non differiva tra i gruppi”. Tuttavia, “alla potenza massima [da sforzo] era più alta nel gruppo rHuEPO rispetto al gruppo placebo”. Quindi, anche se non c’erano differenze a livelli inferiori di sforzo, allo sforzo massimale il gruppo EPO ha comunque ottenuto risultati migliori rispetto al gruppo placebo.[https://www.thelancet.com/]
Ma attraverso l’approfondimento di questo articolo abbiamo compreso che il protocollo di dosaggio qui discusso fa luce sull’uso potenziale dell’EPO per migliorare le prestazioni ciclistiche sotto supervisione, e con questo articolo è stato dimostrato che potrebbe essere efficace in determinati contesti. Questo protocollo è un’ottima risorsa per chi sta valutando l’uso dell’EPO e sottolinea l’importanza di aderire a pratiche scientificamente valide per ottimizzare i risultati dando priorità alla sicurezza.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Carnot P, Deflandre C (1906). “Sur l’activite hematopoietique du serum au cours de la regeneration du sang”. Compt. Rend. Acad. Sci. 143: 384–386.
Eschbach JW, Egrie JC, Downing MR, Browne JK, Adamson JW (January 1987). “Correction of the anemia of end-stage renal disease with recombinant human erythropoietin. Results of a combined phase I and II clinical trial”. The New England Journal of Medicine. 316 (2): 73–8. doi:10.1056/NEJM198701083160203. PMID3537801.
^ Livnah O, Johnson DL, Stura EA, Farrell FX, Barbone FP, You Y, Liu KD, Goldsmith MA, He W, Krause CD, Pestka S, Jolliffe LK, Wilson IA (November 1998). “An antagonist peptide-EPO receptor complex suggests that receptor dimerization is not sufficient for activation”. Nature Structural Biology. 5 (11): 993–1004. doi:10.1038/2965. PMID9808045. S2CID24052881.
Hosseini-Zare MS, Dashti-Khavidaki S, Mahdavi-Mazdeh M, Ahmadi F, Akrami S (July 2012). “Peripheral neuropathy response to erythropoietin in type 2 diabetic patients with mild to moderate renal failure”. Clinical Neurology and Neurosurgery. 114 (6): 663–7. doi:10.1016/j.clineuro.2012.01.007. PMID22296650. S2CID19516031.
Liu S, Ren J, Hong Z, Yan D, Gu G, Han G, Wang G, Ren H, Chen J, Li J (February 2013). “Efficacy of erythropoietin combined with enteral nutrition for the treatment of anemia in Crohn’s disease: a prospective cohort study”. Nutrition in Clinical Practice. 28 (1): 120–7. doi:10.1177/0884533612462744. PMID23064018.
Heuberger, J. A. A. C., Rotmans, J. I., Gal, P., Stuurman, F. E., van ’t Westende, J., Post, T. E., Daniels, J. M. A., Moerland, M., van Veldhoven, P. L. J., de Kam, M. L., Ram, H., de Hon, O., Posthuma, J. J., Burggraaf, J., & Cohen, A. F. (2017). Effects of erythropoietin on cycling performance of well trained cyclists: A double-blind, randomised, placebo-controlled trial. The Lancet Haematology, 4(8), e374–e386. https://doi.org/10.1016/S2352-3026(17)30105-9
I reni sono responsabili, tra l’altro, del filtraggio del sangue e della produzione di urina. Lo fanno creando un filtrato dalle grandi quantità di sangue che li attraversano. Di solito, più di un litro di sangue passa attraverso i reni ogni minuto. Se si sottrae la frazione non fluida, lasciando quindi il plasma sanguigno, questo si traduce in circa 625 ml di plasma sanguigno che passa attraverso i reni ogni minuto. Circa un quinto di questo fluido viene filtrato attraverso i capillari glomerulari (vedi figura sotto) in ogni singolo nefrone di cui sono composti i reni. Il nefrone è l’unità funzionale del rene. Ciascuno di essi è in grado di filtrare il sangue e di produrre l’urina. Un rene è composto all’incirca da 1 milione di nefroni, ma la percentuale varia notevolmente da una persona all’altra [1]. Il fluido che viene filtrato attraverso i capillari glomerulari viene catturato in un “sacco” chiamato capsula di Bowman. La velocità con cui questo fluido, o filtrato glomerulare, viene catturato collettivamente nella capsula di Bowman da tutti i nefroni al minuto è definita velocità di filtrazione glomerulare (eGFR). Negli adulti sani è di circa 125mL/min (il 20% dei 625mL/min di cui sopra).
Circa 625 mL/min di flusso di plasma renale (RPF) passano attraverso i reni, di cui 125 mL/min vengono catturati dalla capsula di Bowman. Di conseguenza, quasi tutto questo viene riassorbito (REAB; 124 mL/min), portando a una produzione di urina di circa 1 mL/min. Immagine tratta da Guyton and Hall Textbook of Medical Physiology 13a edizione.
Stima della velocità di filtrazione glomerulare (eGFR)
La stima del eGFR viene utilizzata come indicatore della funzione renale. Il metodo migliore per farlo è utilizzare una sostanza che viene filtrata liberamente dal glomerulo e non viene né secreta, né riassorbita, né sintetizzata, né metabolizzata dal rene. Pertanto, qualsiasi quantità di sostanza venga filtrata dal glomerulo viene escreta anche nelle urine. Pertanto, l’eGFR può essere ricavato con precisione dalle misurazioni delle urine e dalla conoscenza della quantità somministrata. Il gold standard per misurarlo è l’utilizzo di una sostanza chiamata inulina. È poco utilizzata nella pratica perché è costosa, la maggior parte dei laboratori non è in grado di dosarla e per una valutazione più accurata è necessaria un’endovena con diversi campioni di sangue e la cateterizzazione della vescica. Tutto sommato, non è molto pratico.
Per questo motivo, l’eGFR viene spesso stimato in base alla concentrazione di creatinina nel siero. La creatinina non viene riassorbita o metabolizzata dai reni e viene filtrata liberamente a livello del glomerulo. Inoltre, l’apporto dal tessuto muscolare scheletrico è costante (essendo un prodotto di degradazione della creatina), per cui non è necessario somministrarla per via endovenosa, a differenza dell’inulina. Tuttavia, può verificarsi una significativa secrezione tubulare di creatinina [2]. Pertanto, pur non essendo assolutamente perfetta, queste proprietà della creatinina la rendono comunque utile per ricavare l’eGFR. Sono state stabilite diverse formule che possono fornire una stima del eGFR in base alla sua concentrazione. Tutte si basano sul presupposto che livelli di creatinina più elevati implicano una minore eliminazione di creatinina, ovvero una diminuzione del eGFR.
La formula attualmente raccomandata nella pratica clinica è l’equazione della Chronic Kidney Disease Epidemiology Collaboration (CKD-EPI) [3]. In precedenza, veniva comunemente utilizzata la Modification of Diet in Renal Disease (MDRD). L’equazione CKD-EPI tiene conto, oltre che della concentrazione di creatinina sierica, del sesso, dell’età e del gruppo etnico ed è corretta per un’area di superficie corporea di 1,73 m2. In questo modo si ottengono valori di eGFR con un’unità di misura di mL/min/1,73m2. Un eGFR normale o elevato è considerato superiore a 90 [3].
Problemi con l’eGFR basato sulla creatinina negli utilizzatori di AAS
La stima del eGFR basata sulla concentrazione di creatinina nel siero è notoriamente inaffidabile nei soggetti muscolosi. Poiché il muscolo è il principale sito di immagazzinamento della creatina nell’organismo, i soggetti muscolosi hanno una maggiore quantità di creatina nel corpo. Di conseguenza, anche il tasso di produzione di creatinina è più elevato. Di conseguenza, con tassi di clearance simili, anche i livelli di creatinina nel siero saranno più elevati. Di conseguenza, queste formule sottostimano il vero eGFR. Un altro problema che si presenta in questa popolazione è quello causato dall’integrazione di creatina. L’integrazione di creatina è una pratica comune tra i soggetti che si allenano contro-resistenza ed è efficace nell’aumentare le riserve corporee di creatina [4]. Di conseguenza, la produzione di creatinina è in assoluto più elevata. Inoltre, la creatina etil-estere in particolare può portare a un forte aumento dei livelli di creatinina nel siero [5, 6]. Il motivo più probabile è che la creatina etil-estere viene rapidamente degradata in creatinina nell’organismo dopo l’ingestione [7]. Anche l’ingestione di carne cotta può aumentare transitoriamente i livelli di creatinina nel siero per diverse ore [8]. Inoltre, ci sono prove che indicano che l’uso di steroidi anabolizzanti potrebbe aumentare la biosintesi della creatina. La creatina viene sintetizzata con un meccanismo a due fasi, come illustrato di seguito:
La reazione catalizzata dall’AGAT, che forma l’acido guanidinoacetico, è la fase limitante della sintesi della creatina [9]. È stato riscontrato che la somministrazione di uno steroide anabolizzante (17α-metil testosterone) aumenta l’espressione di AGAT [10]. Inoltre, ha aumentato l’escrezione di acido guanidinoacetico nelle urine del 70%. L’insieme di questi dati suggerisce fortemente che gli steroidi anabolizzanti, almeno quelli biodisponibili per via orale, stimolano la biosintesi della creatina. Di conseguenza, potrebbero aumentare l’accumulo di creatina e quindi influenzare anche il tasso assoluto di produzione di creatinina. Infine, la maggior parte dei consumatori di steroidi anabolizzanti consuma anche una dieta ad alto contenuto proteico. È stato riscontrato che una dieta ad alto contenuto proteico aumenta la eGFR [11]. Si noti che non si tratta di una sovrastima del eGFR, ma di un leggero aumento del eGFR reale.
Riassumendo, i seguenti fattori possono influenzare i livelli di creatinina sierica e quindi l’eGFR stimato senza influenzare effettivamente l’eGFR reale:
Essere più muscolosi
Integrazione di creatina (in particolare di creatina etil-estere)
Aver mangiato carne cotta nelle ore precedenti la misurazione
Assunzione di steroidi anabolizzanti (per via orale).
Detto questo, se si tiene conto del fatto che l’eGFR sarà alterato da questi fattori, si possono comunque osservare variazioni dell’eGFR nel tempo. Supponendo di mantenere tutto abbastanza costante, queste variazioni possono essere indicative di cambiamenti nella velocità di filtrazione glomerulare.
Gli AAS influenzano l’eGFR basato sulla creatinina
Pochi studi hanno misurato l’effetto degli steroidi anabolizzanti, in particolare del Testosterone Enantato, sui livelli di creatinina sierica. Bhasin et al. hanno riportato un lieve aumento da 1,0mg/dL a 1,1mg/dL in uomini normali che ricevevano 600mg di Testosterone Enantato settimanalmente in associazione a esercizi contro-resistenza [12]. Tuttavia, uno studio successivo dello stesso gruppo non ha rilevato cambiamenti significativi nei livelli di creatinina sierica in giovani uomini sani che ricevevano dosi graduate di Testosterone Enantato da 25 a 600mg settimanali per una durata di 20 settimane [13]. Lo stesso gruppo, sempre con un design di studio simile, ma in uomini più anziani, ha riscontrato un aumento da 1,03 a 1,17mg/dL negli uomini che ricevevano 600mg settimanali e da 1,12 a 1,19mg/dL nel gruppo che riceveva 125mg settimanali (che è limite di dosaggio della terapia sostitutiva del Testosterone [TRT]) [14]. Anche uno studio che ha fornito il proormone orale 1-androsterone al dosaggio di 330mg al giorno per 4 settimane ha rilevato un aumento da 1,1mg/dL a 1,3mg/dL dei livelli di creatinina sierica [15]. Gli autori hanno anche calcolato l’eGFR, che è sceso da 88,3 a 71,9ml/min/1,73m2. Non è chiaro se questi aumenti della creatinina sierica riflettano un’effettiva diminuzione del eGFR o se siano semplicemente artefatti derivanti dai problemi relativi all’eGFR basato sulla creatinina, come sottolineato in precedenza. In particolare, non sono note disfunzioni o malattie renali causate dagli steroidi anabolizzanti, ad eccezione di alcuni casi riportati.
Alternative all’eGFR basato sulla creatinina
Nei casi in cui vi siano chiare ragioni per sospettare che l’eGFR basato sui livelli di creatinina sierica sia impreciso, si possono utilizzare alcuni metodi alternativi. Uno di questi si basa sulla misurazione dei livelli di cistatina C nel siero. L’idea è più o meno simile a quella della misurazione della creatinina. La differenza principale è che la cistatina C è prodotta da tutte le cellule (nucleate) a un tasso relativamente costante. Tuttavia, una differenza importante è che una certa metabolizzazione della sostanza avviene nei tubuli. Inoltre, mentre inizialmente si pensava che non fosse influenzata dal sesso, dall’età o dalla massa muscolare, le prove che si stanno accumulando suggeriscono che in realtà lo sia. Diversi studi hanno rilevato che è influenzato da sesso, età, razza, peso, altezza, composizione corporea e stato di fumatore [16, 17, 18]. Tuttavia, uno studio ha concluso che la cistatina C potrebbe rappresentare un’alternativa più adeguata per valutare la funzione renale nei soggetti con massa muscolare più elevata quando si sospetta una lieve compromissione renale [19]. È probabile che l’eGFR basato sulla cistatina C possa fornire un’immagine più chiara del vero eGFR di quanto non faccia l’eGFR basato sulla creatinina nei soggetti allenati contro-resistenza che fanno o non fanno uso di steroidi anabolizzanti. Infine, i dati suggeriscono che la combinazione delle due misurazioni potrebbe addirittura fornire un quadro ancora più accurato di una delle due da sola nella malattia renale cronica [20]. Tuttavia, questi dati non sono stati verificati in modo specifico nei soggetti muscolosi/bodybuilder.
Negli studi di ricerca vengono utilizzati anche marcatori più affidabili come lo Iotalamato e lo Ioexolo. Entrambi possono essere utilizzati con una singola iniezione in bolo, ma richiedono misurazioni plasmatiche multiple. Tuttavia, sono entrambi poco costosi e forniscono stime migliori rispetto all’eGFR basato sulla creatinina/cistatina C. L’Iotalamato, per quanto ne so, è il meno utilizzato nella pratica clinica ed è radioattivo (lo Ioexolo non lo è). Lo Iohexolo comporta un piccolo rischio di nefrotossicità e di reazione allergica (soprattutto ad alte dosi). Lo menziono più per completezza che per altro, in quanto non è qualcosa che dovrebbe essere usato di routine.
Glomerulosclerosi focale segmentale (FSGS) e rilevamento del danno renale con la misurazione delle urine
In letteratura ci sono pochissime segnalazioni di steroidi anabolizzanti dannosi per i reni. Un articolo degno di nota che riporta disfunzioni renali nei consumatori di steroidi anabolizzanti è quello del 2010 di Herlitz et al. [21]. Descrive 10 pazienti provenienti dagli archivi del loro laboratorio di patologia renale in un periodo di 10 anni. I pazienti erano tutti culturisti con una lunga storia di uso di steroidi anabolizzanti. Sono state prelevate biopsie renali che hanno rivelato una glomerulosclerosi focale segmentaria (FSGS) in nove di loro, e quattro di loro presentavano anche glomerulomegalia. In uno dei pazienti non sono stati riscontrati segni di FSGS, ma solo di glomerulomegalia. Che cos’è la FSGS? È un termine un po’ generico per indicare un gruppo di malattie che portano a lesioni glomerulari, mediate da diversi insulti diretti o inerenti al podocita (le cellule che formano la superficie esterna dei capillari glomerulari) [22]. O forse, per meglio dire, è un reperto istologico che non indica necessariamente una malattia specifica. In ogni caso, nella FSGS i podociti iniziano a cambiare forma, diventando più o meno appiattiti (effacement). A un certo punto, il podocita muore e si stacca dalla membrana basale. Poiché i podociti sono cosiddetti “differenziati terminali”, non possono andare incontro a divisione cellulare (proliferare). Pertanto, queste cellule vengono sostituite da tessuto connettivo (sclerosi). Ovviamente il tessuto connettivo non funziona come i podociti e quindi la funzione di filtraggio del glomerulo è compromessa.
Questo può manifestarsi con la perdita di proteine nelle urine. Che non dovrebbero esserci (a parte alcune tracce). I bodybuilder di questo studio hanno perso grandi quantità di proteine nelle urine (in media circa 10 grammi al giorno). In prospettiva, di solito non dovrebbe trattarsi di più di qualche milligrammo. Uno dei bodybuilder era addirittura in grado di produrre il proprio frullato proteico, visto che ha fatto la pipì con ben 26 g di proteine al giorno. In particolare, anche la creatinina sierica era marcatamente elevata in questi soggetti. Mentre l’intervallo di normalità va da 0,9mg/dL a 1,3mg/dL, questi soggetti presentavano in media livelli di creatinina sierica pari a 3,0mg/dL, con uno che raggiungeva l’incredibile valore di 7,8mg/dL. Ovviamente, queste grandi deviazioni sono chiaramente causate da una diminuzione della funzione renale.
Ci sono alcune cose che vorrei sottolineare in questo articolo. Uno è che questi bodybuilder non erano bodybuilder medi. Il loro IMC medio era di 35 kg/m2. Erano dannatamente enormi. Quattro di loro hanno ammesso di aver fatto uso di steroidi anabolizzanti in combinazione con il GH e uno di loro con l’Insulina. Inoltre, sei avevano anche l’ipertensione. Di seguito è riportata la foto di uno di loro:
Sono riusciti a effettuare un follow-up di otto dei soggetti dopo la sospensione degli steroidi anabolizzanti (a tutti, tranne uno, erano stati prescritti anche dei farmaci, per lo più inibitori del sistema Renina-Angiotensina-Aldosterone [RAAS]). Al follow-up, sono stati osservati grandi miglioramenti nella proteinuria e miglioramenti variabili nei livelli di creatinina sierica. In particolare, un paziente è ricaduto nell’uso di steroidi anabolizzanti e ha visto aumentare di nuovo in modo considerevole l’escrezione di proteine nelle urine.
Sebbene sia difficile affermare che tutto questo possa essere il risultato dell’uso di steroidi anabolizzanti, sembra probabile che in alcuni rari casi l’uso cronico eccessivo di steroidi anabolizzanti possa portare a questo fenomeno. Anche perché non se ne parla molto in letteratura, nonostante i milioni e milioni di consumatori di steroidi anabolizzanti sparsi per il mondo. Tuttavia, potrebbe esserci una significativa sottostima, in quanto forme più lievi di danno renale potrebbero passare inosservate, anche con le misurazioni di routine dell’eGFR. La diagnosi precoce può essere ottenuta con il test delle proteine nelle urine, che raramente viene effettuato senza indicazione.
Per questo motivo, si potrebbe raccomandare di effettuare le misurazioni delle urine con una certa regolarità. Ad esempio, annualmente o semestralmente. Lievi aumenti di albumina nelle urine dovrebbero indurre a ripetere l’esame, poiché possono derivare, ad esempio, da un’infezione o dall’esercizio fisico, senza essere causati da un vero e proprio danno renale. In caso di elevazioni persistenti o elevate, è necessario avviare un ulteriore follow-up e, idealmente, interrompere l’uso di steroidi anabolizzanti.
Nota: la ricerca in atto ha ipotizzato che l’uso di AAS sia adittivo al possibile emergere di disfunzioni renali. I risultati di alcuni studi indicano infatti che un’elevata assunzione di proteine, l’uso di AAS, in particolare gli schemi, tra cui il Boldenone Undecylenato, e altri farmaci con un certo “carico renale”, aumentano l’ecogenicità corticale, lo spessore del parenchima renale e il volume renale nei bodybuilder.
Interazione di rInsulina e rhGH sulla funzionalità renale
L’insulino-resistenza è una caratteristica comune nei bodybuilder che usano per lunghi periodi di tempo protocolli di hGH/Insulina. L’IR è comune nei pazienti con malattia renale cronica (CKD), anche in assenza di diabete (DeFronzo et al., 1981; Shinohara et al., 2002; Becker et al., 2005; Kobayashi et al., 2005; Landau et al., 2011), ed è un fattore di rischio per la progressione della CKD (Fox et al., 2004). La sua prevalenza nella CKD varia dal 30 al 50% e dipende principalmente dal metodo di misurazione adottato (Spoto et al., 2016). L’insulino-resistenza può essere rilevata nelle fasi iniziali, quando l’eGFR è ancora nel range di normalità, suggerendo un ruolo potenziale nell’innescare la CKD (Fliser et al., 1998). Un ampio studio basato sulla coorte Atherosclerosis Risk in Communities (ARICs) ha confermato che lo sviluppo della CKD aumenta in stretto parallelismo con il numero di criteri della sindrome metabolica misurati negli adulti non diabetici, e questa relazione rimane significativa anche dopo aver controllato lo sviluppo di diabete e ipertensione (Kurella et al., 2005). L’insulino-resistenza è stata anche associata a una prevalente CKD e a un rapido declino della funzione renale in individui asiatici anziani e non diabetici (Cheng et al., 2012) e alla microalbuminuria nella popolazione generale (Mykkänen et al, 1998) e in pazienti con T1DM (Yip et al., 1993; Ekstrand et al., 1998) e T2DM (Groop et al., 1993), indicando che questa relazione è indipendente dal diabete (Mykkänen et al., 1998; Chen et al., 2003, 2004). Il meccanismo proposto per cui l’IR contribuisce al danno renale prevede il peggioramento dell’emodinamica renale attraverso l’attivazione del sistema nervoso simpatico (Rowe et al., 1981), la ritenzione di sodio, la diminuzione dell’attività della Na+, K+-ATPasi e l’aumento del GFR (Gluba et al., 2013).
L’eziologia dell’IR nella CKD è multifattoriale e dipende da fattori di rischio classici e specifici della CKD, come l’inattività fisica, l’infiammazione e lo stress ossidativo, le alterazioni delle adipochine, la carenza di vitamina D, l’acidosi metabolica, l’anemia e le tossine microbiche (Spoto et al., 2016).
L’emodialisi a lungo termine ha un effetto positivo sull’IR (DeFronzo et al., 1978), ma ci sono pochi dati clinici sull’effetto della dialisi peritoneale.
Oltre a essere un fattore di rischio per l’insorgenza e la progressione della CKD, l’IR è anche coinvolta nell’aumento del rischio cardiovascolare (CV) in questa popolazione. L’IR può essere responsabile dell’ipertensione arteriosa attraverso la stimolazione diretta del RAAS (Nickenig et al., 1998), l’attivazione del sistema simpatico (Sowers et al., 2001) e la sottoregolazione del sistema dei peptidi natriuretici (Sarzani et al., 1999).
Similmente a quanto accade nei pazienti acromegalici, livelli cronicamente alti di rhGH possono essere associati a ipertrofia renale nell’uomo [Kamenický P et al. 2014]. In uno studio caso-controllo, la lunghezza del rene valutata mediante ecografia renale è risultata significativamente aumentata di circa 5cm (55%) e 2cm (20%) rispettivamente nei pazienti acromegalici attivi e controllati [Auriemma RS et al. 2010]. Le dimensioni del rene si normalizzano rapidamente entro 3-6 mesi nei pazienti acromegalici sottoposti a chirurgia transfenoidale [Zhang Z et al. 2018]. Mancano studi sistematici sull’istologia renale nei pazienti acromegalici. Rari casi, in cui i pazienti acromegalici sono stati sottoposti a biopsia renale a causa della sindrome nefrosica o della proteinuria persistente, hanno rivelato una glomerulosclerosi focale segmentaria [Takai M et al. 2001]. In un paziente acromegalico che presentava proteinuria di gamma nefrosica e glomerulosclerosi focale segmentaria alla biopsia renale, la proteinuria si è rapidamente normalizzata dopo l’asportazione del tumore, ma è ritornata 4 mesi dopo, rispondendo però al trattamento con Prednisolone [Wang R et al. 2021]. Nei pazienti acromegalici sottoposti a biopsia renale è stata notata solo un’ipertrofia moderata o non glomerulare.
I pazienti acromegalici presentano un’iperfiltrazione glomerulare caratterizzata da un aumento di circa il 15% del eGFR e del RPF rispetto ai soggetti sani, che è reversibile nella maggior parte dei pazienti, ma non in tutti, con la rimozione chirurgica degli adenomi ipofisari [Fujio S et al. 2016]. Si ritiene che l’iperfiltrazione glomerulare persistente contribuisca allo sviluppo di albuminuria nei pazienti acromegalici sottoposti a chirurgia tardiva [Grunenwald S et al. 2011]. Nello studio Baldelli, la microalbuminuria è stata riportata nel 55% dei pazienti acromegalici e associata a ipertensione, alterata tolleranza al glucosio e diabete [Baldelli R et al. 2008].
Similmente a quanto osservato con gli abusatori di rhGH, i pazienti acromegalici mostrano un aumento dell’acqua corporea totale e del sodio e possono presentare un edema evidente. Questi cambiamenti sono legati alle proprietà di ritenzione di sodio del GH e dell’IGF-1 attraverso l’ENaC nei tubuli distali renali e possono essere invertiti se i pazienti sono sottoposti a un trattamento efficace del tumore che produce GH [Kamenický P et al. 2020]. L’acqua corporea totale (56% contro 50% del peso corporeo) ed extracellulare (20% contro 15% del peso corporeo), così come il sodio scambiabile, sono risultati aumentati nei pazienti acromegalici rispetto ai soggetti sani, mentre non sono state rilevate differenze nel contenuto di acqua intracellulare [Ikkos D et al. 1954]. Anche il volume plasmatico è risultato aumentato in questi pazienti [Hirsch EZ et al. 1969]. Le conseguenze cliniche di queste alterazioni sono l’ipertensione arteriosa, l’ipertrofia ventricolare sinistra e l’insufficienza cardiaca congestizia, che contribuiscono all’aumento complessivo della mortalità nei pazienti non trattati. È importante notare che l’ipertensione arteriosa è associata a un esito inferiore in questi pazienti [Vila G et al. 2020]. Inoltre, i pazienti acromegalici diabetici presentano un’ipertrofia ventricolare sinistra più pronunciata rispetto ai pazienti non diabetici [Nemes A et al. 2020].
I pazienti acromegalici spesso presentano una lieve iperfosfatemia nonostante l’aumento del eGFR, a causa dell’aumento del TmP/eGFR, che può essere utilizzato come misura completa dello stato della malattia e può essere invertito con il trattamento [Xie T et al. 2020]. I meccanismi sottostanti includono l’up-regulation del cotrasportatore Na-Pi 2a nei tubuli prossimali renali indotta dall’IGF-1 e un maggiore assorbimento intestinale di fosfato, dovuto all’aumento della sintesi di calcitriolo indotto dal GH. I pazienti mostrano spesso concentrazioni sieriche verso l’intervallo superiore di normalità in associazione a ipercalciuria [Manroa P et al. 2014]. Questi risultati sono molto probabilmente correlati alla sintesi di calcitriolo indotta dal GH, con conseguente aumento dell’assorbimento intestinale di calcio, poiché i livelli di calcitriolo tendono a essere elevati in questi pazienti. Inoltre, nei pazienti acromegalici è stato dimostrato un maggiore assorbimento di calcio nei reni, molto probabilmente legato alla stimolazione indotta dal calcitriolo dell’espressione di TRPV5 nei tubuli renali distali [Suzuki Y et al. 2008]. Si ritiene che l’alterato metabolismo del calcio contribuisca all’aumento della fragilità scheletrica osservato nei pazienti acromegalici [Mazziotti G et al. 2013].
Conclusioni:
Sebbene non vi siano prove certe della correlazione tra patologie renali e AAS, questi ultimi hanno mostrato di poter causare peggioramenti della funzionalità renale anche solo in modo transitorio. La loro azione addittiva con altre molecole e loro alterazione del contesto metabolico (vedi abuso di rInsulina e rhGH con conseguente peggioramento dell’IR) può essere in parte la causa delle problematiche renali osservati in diversi bodybuilder Enhanced, specie di alto livello. Gli studi svolti su animali hanno mostrato possibili attività nefrotossiche in particolari AAS come, ad esempio, il Boldenone. La ricerca, seppur in piccolo, continua e un giorno potremmo avere le idee più chiare sulla reale correlazione tra AAS (e PEDs) e malattie renali.
Bertram, John F., et al. “Human nephron number: implications for health and disease.” Pediatric nephrology 26.9 (2011): 1529-1533.
Shemesh, Ovadia, et al. “Limitations of creatinine as a filtration marker in glomerulopathic patients.” Kidney international 28.5 (1985): 830-838.
Eknoyan, Garabed, et al. “KDIGO 2012 clinical practice guideline for the evaluation and management of chronic kidney disease.” Kidney Int 3.1 (2013): 5-14.
Harris, Roger C., Karin Söderlund, and Eric Hultman. “Elevation of creatine in resting and exercised muscle of normal subjects by creatine supplementation.” Clinical science 83.3 (1992): 367-374.
Williamson, Lydia, and David New. “How the use of creatine supplements can elevate serum creatinine in the absence of underlying kidney pathology.” Case Reports 2014 (2014): bcr2014204754.
Velema, M. S., and W. De Ronde. “Elevated plasma creatinine due to creatine ethyl ester use.” Neth J Med 69.2 (2011): 79-81.
Gufford, Brandon T., et al. “pH-dependent stability of creatine ethyl ester: relevance to oral absorption.” Journal of dietary supplements 10.3 (2013): 241-251.
Preiss, David J., et al. “The influence of a cooked-meat meal on estimated glomerular filtration rate.” Annals of clinical biochemistry 44.1 (2007): 35-42.
Walker, James B. “Creatine: biosynthesis, regulation, and function.” Adv Enzymol Relat Areas Mol Biol 50.177 (1979): 2.
Hoberman, Henry D., Ethan AH Sims, and William W. Engstrom. “The effect of methyltestosterone on the rate of synthesis of creatine.” Journal of Biological Chemistry 173.1 (1948): 111-116.
Martin, William F., Lawrence E. Armstrong, and Nancy R. Rodriguez. “Dietary protein intake and renal function.” Nutrition & metabolism 2.1 (2005): 1-9.
Bhasin, Shalender, et al. “The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men.” New England Journal of Medicine 335.1 (1996): 1-7.
Bhasin, Shalender, et al. “Testosterone dose-response relationships in healthy young men.” American Journal of Physiology-Endocrinology And Metabolism (2001).
Bhasin, Shalender, et al. “Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle.” The Journal of Clinical Endocrinology & Metabolism 90.2 (2005): 678-688.
Granados, Jorge, et al. “Prohormone supplement 3β-hydroxy-5α-androst-1-en-17-one enhances resistance training gains but impairs user health.” Journal of applied physiology 116.5 (2014): 560-569.
Knight, Eric L., et al. “Factors influencing serum cystatin C levels other than renal function and the impact on renal function measurement.” Kidney international 65.4 (2004): 1416-1421.
Groesbeck, Darcy, et al. “Age, gender, and race effects on cystatin C levels in US adolescents.” Clinical Journal of the American Society of Nephrology 3.6 (2008): 1777-1785.
Macdonald, Jamie, et al. “GFR estimation using cystatin C is not independent of body composition.” American journal of kidney diseases 48.5 (2006): 712-719.
Baxmann, Alessandra Calábria, et al. “Influence of muscle mass and physical activity on serum and urinary creatinine and serum cystatin C.” Clinical Journal of the American Society of Nephrology 3.2 (2008): 348-354.
Stevens, Lesley A., et al. “Estimating GFR using serum cystatin C alone and in combination with serum creatinine: a pooled analysis of 3,418 individuals with CKD.” American journal of kidney diseases 51.3 (2008): 395-406.
Herlitz, Leal C., et al. “Development of focal segmental glomerulosclerosis after anabolic steroid abuse.” Journal of the American Society of Nephrology 21.1 (2010): 163-172.
D’Agati, Vivette D., Frederick J. Kaskel, and Ronald J. Falk. “Focal segmental glomerulosclerosis.” New England Journal of Medicine 365.25 (2011): 2398-2411.
Nella 2° parte abbiamo analizzato le caratteristiche e funzioni biochimiche della Glutammina. In questa terza parte, invece, andremo ad analizzare due AA legati tra loro per via metabolica, la L-Citrullina e la L-Arginina.
Dalla L-Citrullina alla L-Arginina – Biologia e principali attività:
Il composto organico Citrullina è un α-amminoacido (formula H2NC(O)NH(CH 2)3CH(NH2)CO2H. ).[1] Sebbene sia stato nominato e descritto dai gastroenterologi fin dalla fine del XIX secolo, è stato isolato per la prima volta dall’anguria nel 1914 dai ricercatori giapponesi Yotaro Koga e Ryo Odake [2] [3] e ulteriormente codificato da Mitsunori Wada dell’Università Imperiale di Tokyo nel 1930.[4] La L-Citrullina è un composto aminoacidico non proteico (non viene utilizzato per formare proteine strutturali come gli enzimi) e, a differenza della L-Arginina, non è ampiamente presente in tutte le proteine. Si trova in concentrazioni particolarmente alte nell’anguria (da cui deriva il suo nome, dato che i cocomeri sono conosciuti come Citrullus vulgaris[1]), dove si trova in media a 2,1mg/g di peso umido (anche se i numeri assoluti variano)[2] e si è notato che il consumo di anguria aumenta in modo acuto sia l’Arginina plasmatica che la Citrullina (3.3 kg di anguria equivalgono a 10g di L-Arginina supplementare)[3][4] e di aumentare l’Arginina e l’Ornitina a digiuno del 12-22% in seguito al consumo di 780-1560g al giorno.[5]
Altre fonti alimentari di L-Citrullina sono i meloni, i meloni amari, le zucchine, le zucche e i cetrioli.[6]
La Citrullina viene sintetizzata nell’organismo attraverso una delle due vie: riciclata dall’Arginina (la conversione dell’arginina in ossido nitrico lascia la citrullina come sottoprodotto)[7][8] o prodotta dall’azoto (e da una parte del carbonio) contenuto nella L-glutammina,[9] dove l’enzima ornitina transcarbamilasi utilizza sia l’Ornitina che il carbamoilfosfato (che richiede la Clutammina) per produrre Citrullina negli enterociti.[10][11]
Sembra che la via dell’Arginina sia responsabile di circa il 10% della Citrullina circolante, mentre la via della Glutammina ne rappresenta il 90%;[6] la riduzione dei livelli plasmatici di Glutammina può ridurre la Citrullina plasmatica.[12]
Per quanto riguarda il ciclo dell’urea (uno dei meccanismi alla base del 10%), la L-Arginina viene convertita in L-Ornitina tramite l’enzima arginasi (cedendo urea come cofattore)[13][14] e da qui l’Ornitina (utilizzando il carbamoilfosfato come cofattore) viene sottoposta all’enzima Ornitina carbamoiltransferasi per produrre L-Citrullina. In questo senso, la via metabolica dall’Arginina alla Citrullina (attraverso l’Ornitina) provoca un aumento dell’urea e una concomitante diminuzione dell’ammoniaca, utilizzata dall’enzima carbamoilfosfato sintasi per creare carbamoilfosfato.[15] Se necessario, l’arginina può essere convertita direttamente in L-Citrullina attraverso un enzima arginina deiminasi per produrre, anziché richiedere, ammoniaca.[16]
Il ciclo si forma quando la citrullina si lega con l’L-aspartato (correlato all’acido D-aspartico come isomero) per formare l’arginosuccinato attraverso l’enzima arginosuccinato sintasi, quindi l’enzima arginosuccinato lisasi degrada l’arginosuccinato in arginina libera e fumarato; l’arginina rientra quindi nel ciclo dell’urea. [Il fumarato può semplicemente entrare nel ciclo TCA (Krebs) come intermedio energetico,[17] e la citrullina regola negativamente l’enzima arginasi.[18]
Anche la conversione della citrullina in L-arginosuccinato e la successiva conversione in L-arginina è coinvolta nel ciclo dell’ossido nitrico piuttosto che nel ciclo dell’urea, con l’unica differenza che l’arginina si converte direttamente in citrullina (cedendo una molecola di ossido nitrico) piuttosto che essere convertita indirettamente tramite l’ornitina.[18][19]
Come accennato, l’Arginina entra prima in contrata con il metabolismo intestinale e splancnico, in cui una certa quantità di essa viene consumata dagli enterociti o interconvertita in L-citrullina o L-ornitina. Oltre all’elevato utilizzo dell’arginina da parte del fegato, anche l’assorbimento intestinale dell’arginina è scarso in condizioni normali e aumenta in varie patologie.[20] Sembra che una quantità minima di L-arginina arrivi ai tessuti sistemici rispetto agli altri aminoacidi del ciclo dell’urea, dato che la L-ornitina supplementare raggiunge una concentrazione sierica doppia rispetto alla L-arginina e la L-citrullina 9,3 volte superiore. Ciò sembra direttamente correlato al grado di metabolismo epatico e intestinale.[21][22][23]
L’Arginina alimentare rappresenta il 40-60% dell’arginina sierica, come evidenziato da un calo equivalente durante i periodi di assenza di arginina. Il tasso di conversione della L-citrullina in L-arginina non sembra influenzato dall’assunzione con la dieta.[24]
La citrullina di per sé è più che altro un sottoprodotto del metabolismo dell’arginina (ciclo dell’ossido nitrico) e dell’ornitina (ciclo dell’urea) e viene semplicemente riconvertita in arginina tramite l’arginosuccinato. Detto questo, l’integrazione di citrullina influisce positivamente anche sulle concentrazioni di arginina e ornitina, quindi anche la loro bioattività è rilevante.
L’arginina può essere convertita in L-citrullina attraverso gli enzimi dell’ossido nitrico sintasi (NOS), di cui esistono forme endoteliali (eNOS) e neuronali specifiche (nNOS), nonché una forma inducibile (iNOS) che risponde ai segnali infiammatori. La conversione dell’arginina attraverso gli enzimi NOS produce ossido nitrico come sottoprodotto più importante, e la Citrullina è vista come un sottoprodotto.[25] La Citrullina può poi riconvertirsi in L-arginina attraverso l’arginosuccinato, ma la L-ornitina non è coinvolta nella via dell’ossido nitrico.
La L-Citrullina viene assorbita nell’intestino in misura molto maggiore rispetto alla sua controparte L-Arginina e determina un livello plasmatico più elevato di L-Arginina attraverso il ciclo Arginina/Ornitina/Citrullina.[26] Viene assorbita attraverso numerosi trasportatori sodio-dipendenti.[27]
È stato osservato che l’integrazione orale di citrullina nell’uomo a 0,18 g/kg raddoppia l’arginina plasmatica[28], cosa che è stata replicata altrove[29], insieme a un aumento equivalente delle concentrazioni di ornitina[29], ma questi raddoppi di arginina e ornitina sono associati a un aumento di 6-11 volte della citrullina plasmatica[28][29].
Una singola dose di 6 g di citrullina malato (0,08 g/kg) in atleti prima dell’esercizio fisico ha fatto registrare aumenti della citrullina plasmatica (aumento del 173%), dell’ornitina (aumento del 152%) e dell’arginina (aumento del 123%) quando misurata dopo l’esercizio fisico, valori che si sono normalizzati con 3 ore di riposo.[30] Questa stessa dose è stata notata altrove per aumentare la citrullina plasmatica e l’arginina in misura simile.[31]
È interessante notare che gli studi sopra citati che hanno utilizzato 0,18 g/kg di citrullina hanno rilevato un aumento di 6-11 volte della citrullina a fronte di un mero raddoppio dell’arginina e dell’ornitina[28][29], mentre lo studio successivo che ha utilizzato 6 g (calcolati come 0,08 g/kg) ha registrato un aumento molto minore della citrullina, ma ha comunque più che raddoppiato sia l’arginina che l’ornitina. [30] Ciò è stato osservato anche in uno studio dose-risposta che ha utilizzato da 2 a 15 g di citrullina, in cui la citrullina nel plasma ha seguito una dipendenza lineare dalla dose, mentre sia l’arginina che l’ornitina hanno avuto una dipendenza minore dalla dose.[29] Gli autori hanno ipotizzato che, dato che l’aumento dell’arginina è stato inferiore a quello previsto e che la citrullina sierica è il principale predittore della sintesi dell’arginina[19], ciò indichi il raggiungimento di una fase di limitazione della velocità nei reni.
È stato osservato che la citrullina non influenza i livelli sierici degli aminoacidi a catena ramificata a riposo,[21] ma può accelerare la deplezione dei BCAA indotta dall’esercizio fisico prolungato (aumentandone l’utilizzo come carburante).[20]
Con l’integrazione di citrullina è stata notata una riduzione della glutammina (13% dopo 0,18 g/kg di citrullina per 7 giorni)[21], anche se un altro studio ha rilevato che l’uso acuto di 6 g di citrullina (0,08 g/kg) non ha alterato le concentrazioni di glutammina.[20]
Gli altri aminoacidi testati (acido glutammico, acido aspartico, asparagina, alanina, lisina, triptofano, fenilalanina, L-tirosina, istidina) sono per lo più inalterati.[20]
Circa l’83% della citrullina ingerita per via orale sembra essere assorbita dai reni[26][27][28] dove viene convertita in L-arginina nei tubuli prossimali (attraverso gli enzimi arginosuccinato sintasi e arginosuccinato liasi[29]); Questa conversione della citrullina in arginina (sia da citrullina supplementare che da quella prodotta come sottoprodotto della creazione di ossido nitrico da parte dell’arginina) rappresenta il 5-15% dell’arginina circolante. [11][30]
Il meccanismo principale con cui l’integrazione di arginina (e, per estensione, di L-citrullina) influisce sulla salute del sangue è quello di essere il substrato per gli enzimi dell’ossido nitrico sintasi (NOS) per la produzione di ossido nitrico, che poi segnala attraverso i recettori ciclici solubili della guanilina la produzione di cGMP. La produzione di ossido nitrico e la successiva produzione di cGMP intracellulare sono alla base di buona parte dei benefici dell’arginina.
Gli enzimi NOS si presentano in tre isoforme principali: [32][33] la NOS inducibile (iNOS), che viene creata in risposta a fattori di stress infiammatori, la NOS neuronale (nNOS), che è stata scoperta per la prima volta nei neuroni e si trova anche nelle terminazioni motorie dei muscoli scheletrici, e la NOS endoteliale (eNOS), che inizialmente si pensava si trovasse solo nell’endotelio, ma è piuttosto diffusa[34], compreso il tessuto cerebrale.[35][36]
Gli enzimi NOS lavorano in dimeri uniti testa a testa e i meccanismi catalitici dipendono da questa dimerizzazione, oltre che dall’eme, dalla tetraidrobiopterina, dalla calmodulina, dal NADPH (come donatore di elettroni) e da FMN e FAD.[37][38][39] Di conseguenza, gli enzimi NOS (tutte e tre le isoforme) sono flavoproteine che richiedono NADPH. [40][41][42] La loro struttura e funzione è complessa (esaminata qui[43]), ma esiste un sito di legame di base per l’arginina e gli elettroni donati dal NADPH fanno sì che l’arginina si converta in citrullina, rilasciando come sottoprodotto l’ossido nitrico; l’iNOS utilizza esclusivamente e l’eNOS può anche utilizzare un intermedio radicale libero chiamato Nω-idrossi-L-arginina (L-NOHA), che si degrada in citrullina e ossido nitrico in presenza di H2O2.[32][44]
L’aumento dell’ossido nitrico (solitamente misurato attraverso le concentrazioni plasmatiche di nitrato/nitrito, citrullina o cGMP urinario) sembra essere aumentato con la L-arginina in persone affette da ipertensione essenziale,[45] arterotrombosi,[46] e diabete di tipo II. [47] Gli studi condotti su atleti altrimenti sani che assumono L-arginina sono piuttosto contrastanti; ci sono casi in cui i biomarcatori del metabolismo dell’ossido nitrico sono aumentati[48] mentre altri studi non notano alcuna modifica.[49][50][51] Non sorprende che i benefici associati all’ossido nitrico non si verifichino quando i biomarcatori dell’ossido nitrico non sono aumentati.
L’inaffidabilità dell’aumento dell’ossido nitrico da parte dell’arginina può essere dovuta al fatto che le concentrazioni fisiologiche di arginina (40-100µM nello spazio extracellulare[52] e forse fino a 800µM a livello intracellulare[53]) sono sufficienti a saturare intrinsecamente l’ossido nitrico sintasi endoteliale (eNOS) (di solito si dichiara una Km di 3µM[54][55], ma a volte viene misurata fino a 29μM[56). Ciò implica che l’enzima è già al massimo dell’efficacia e che un’ulteriore integrazione non aumenta il tasso di conversione (a causa di un arretrato di arginina nel siero); l’osservazione che l’arginina aumenta ancora l’ossido nitrico a volte (anche se in modo inaffidabile) è indicata come il paradosso della L-arginina.[57][58]
Questa teoria è in linea con le osservazioni secondo cui a volte il metabolismo dell’ossido nitrico non viene influenzato nonostante aumenti fino al 300% dell’arginina plasmatica.[59]
Uno studio ha osservato un aumento transitorio della produzione di ossido nitrico che sembra essere più simile a quello di un agonista che di un substrato[60] e successivamente è stato scoperto che l’arginina ha la capacità di attivare i recettori α2-adrenergici,[61] che possono stimolare direttamente l’ossido nitrico senza richiedere la conversione in citrullina attraverso la NOS. Tuttavia, l’arginina è risultata piuttosto debole (superata dall’agmatina)[61] ma questo meccanismo non è ancora stato escluso.
Inoltre, l’arginina extracellulare sembra essere un fattore determinante per il rilascio di ossido nitrico[56] (il trasportatore CAT1 che trasporta l’arginina è altamente associato alla eNOS[62] e l’inibizione dell’afflusso extracellulare impedisce l’attivazione della eNOS[63]), mentre la concentrazione intracellulare di arginina non sembra essere associata. [58] Poiché il trasporto è necessario, ma l’arginina intracellulare non è di per sé necessaria, si ritiene che la colocalizzazione di CAT1 con eNOS[62][64] possa svolgere un ruolo nella stimolazione dell’attività di eNOS.
ADMA
L’ADMA è un metabolita metilato dell’arginina e sembra agire in opposizione all’arginina inibendo le azioni dell’enzima NOS e la conseguente produzione di ossido nitrico. Livelli eccessivi di ADMA possono essere causati da fattori di stress ossidativo che diminuiscono l’attività dell’enzima che lo degrada, mentre la riduzione dell’ADMA provoca una vasodilatazione dovuta alla produzione di ossido nitrico.
Sebbene la maggior parte delle evidenze suggerisca che l’ADMA non aumenta con l’integrazione di L-arginina (questi studi notano che il rapporto arginina:ADMA è aumentato a causa dell’aumento dell’arginina plasmatica), ci sono prove limitate che suggeriscono un aumento che richiede ulteriori indagini.[38]
L’integrazione orale di arginina (anche la citrullina si applica in questo caso perché aumenta l’arginina plasmatica) è in grado di aumentare il flusso sanguigno nelle persone con flusso sanguigno ridotto e, sebbene abbia il potenziale di ridurre la pressione sanguigna, sembra un po’ inaffidabile e può verificarsi solo negli ipertesi. Esistono prove contrastanti sugli effetti dell’integrazione di arginina sul flusso sanguigno in persone con resistenza periferica o cladicazione intermittente, con studi a breve termine che notano un beneficio e studi a più lungo termine che notano un’alterazione.[65][66]
L’integrazione di citrullina sembra ridurre la pressione sanguigna e migliorare il flusso sanguigno in situazioni in cui il flusso sanguigno è altrimenti ostacolato o la pressione sanguigna è più alta del normale, ma la citrullina non ha effetti di riduzione unidirezionali; può essere inefficace in persone normotese a riposo.[41]
In atleti allenati a cui sono stati somministrati 6 g di citrullina malato prima di un test ciclistico prolungato (137 km), l’aumento dell’ormone della crescita indotto dall’esercizio sembra essere aumentato; quando è stato misurato subito dopo l’esercizio, il gruppo con citrullina aveva concentrazioni di GH più elevate del 66,8%, che (dopo 3 ore di riposo) si sono attenuate al 28%.[20] Altrove, dosi di 2-15 g di citrullina non sono riuscite a influenzare l’ormone della crescita a riposo, se misurate nell’arco di 8 ore.[22]
Le concentrazioni di IGF-1 dopo 0,18 g/kg di citrullina per 7 giorni non sono state influenzate in modo significativo.
Durante l’esercizio fisico, sebbene uno studio che ha utilizzato 3 g di L-arginina (associata a 2.200 mg di L-ornitina e 12 mg di vitamina B12) per 3 settimane abbia rilevato un aumento del 35,7% della secrezione di ormone della crescita indotta dall’esercizio fisico (che si è normalizzata entro un’ora)[67], altri studi notano il contrario; è stato osservato che l’integrazione di arginina determina un minore picco di ormone della crescita durante l’esercizio fisico rispetto all’esercizio fisico da solo[68][69] e che, sebbene sembri influenzare maggiormente i giovani rispetto agli anziani[70], si dice che influisca su entrambi i gruppi di età. [L’entità della soppressione (supponendo che il 100% sia il valore di base) è stata notata intorno a un aumento del 300-500% visto con l’esercizio fisico, attenuato al 200%.[68]
È possibile che un aumento eccessivo dell’ormone della crescita stimoli un feedback autogeno, il che spiegherebbe come gli individui più anziani siano meno sensibili a questa soppressione, in quanto hanno intrinsecamente meno picchi di GH dovuti all’esercizio fisico rispetto ai giovani.[71] Infine, poiché i picchi dell’ormone della crescita si normalizzano comunque nel giro di poche ore[67][71], non si sa esattamente quanto sia preoccupante questa soppressione (dato che le concentrazioni di ormone della crescita nelle 24 ore sono più rilevanti).
A riposo, l’integrazione di 5-9 g di L-arginina è in grado di provocare un aumento delle concentrazioni di picco dell’ormone della crescita (aumento del 34,4-120%), mentre 13 g sono risultati inefficaci a causa della sofferenza intestinale che impedisce l’assorbimento della L-arginina.[36]
Negli studi che misurano la secrezione di GH nelle 24 ore, non sono state riscontrate alterazioni significative con la somministrazione di 2 g due volte al giorno[72] o con dosi acute di 5 g.[73] Ciò è potenzialmente legato a un noto fenomeno di feedback autonomo dell’ormone della crescita,[69][74][75] e un effetto modulatorio simile sull’ormone della crescita si riscontra anche durante la restrizione del sonno (in cui una riduzione del rilascio di ormone della crescita indotto dal sonno viene compensato durante le ore di luce). È stato osservato che l’arginina ad alte dosi (250mg/kg di arginina aspartato al giorno, circa 17,5g di arginina) aumenta l’impulso di GH nel sonno a onde lente di circa il 60%, pur non avendo un’influenza sufficiente sulle concentrazioni di GH durante la veglia.[76] Non è chiaro come questo grande aumento influisca sulle concentrazioni di ormone della crescita nell’intera giornata.
L’integrazione di arginina nei ratti è in grado di aumentare il nitrato urinario post-esercizio, indicativo della produzione di ossido nitrico.[77] Aumenti nella produzione di ossido nitrico (nitrato urinario o sierico) sono stati confermati anche nell’uomo in seguito ad assunzione orale o infusione endovenosa.[78]
Non sempre si riscontra un aumento della produzione di ossido nitrico (anche nonostante l’aumento dell’arginina plasmatica), suggerendo che l’attività dell’enzima NOS potrebbe essere un fattore limitante. Per quanto riguarda gli studi in acuto (assunzione di una singola dose di L-arginina prima dell’esercizio), 3 g di arginina (sotto forma di AAKG) non hanno apportato benefici all’allenamento con i pesi,[79] 6 g di L-arginina per 3 giorni non hanno modificato i risultati del cicloergometro in atleti di judo, mentre un protocollo simile in ciclisti allenati ha rilevato un miglioramento del tempo di esaurimento (25,8%).[80]
Alcuni studi hanno utilizzato una forma di arginina nota come GAKIC (Glycine L-Arginine α-Ketoisocaproic acid) e hanno rilevato un aumento della potenza media durante gli sprint di 10s su cicloergometro (con 11,2 g di GAKIC)[81] e un aumento del 10,5+/-0. Questi studi, tuttavia, sono confusi sia dall’inclusione della glicina sia da quella del metabolita della leucina, l’acido α-chetoisocaproico.
Per quanto riguarda gli studi più cronici, l’integrazione di L-arginina (come asparato) con 2,8 o 5,7 g di arginina al giorno per 4 settimane non è riuscita a modificare le prestazioni o altri biomarcatori[82]; anche uno studio precedente, condotto per 2 settimane con una metodologia simile, ha fallito.[83] Nel complesso, quando si esaminano le revisioni o le meta-analisi sull’argomento L-arginina e prestazioni sportive, si nota che è promettente, ma manca un consenso sufficiente per raccomandarla come ergogenico.[84]
Si ritiene che la citrullina sia un agente pro-erettile in quanto è un precursore dell’arginina, e l’arginina è il substrato da cui viene prodotto l’ossido nitrico che può poi indurre il cGMP (attraverso la via NO/cGMP/VEGF);[65] un aumento del cGMP è anche l’effetto finale degli inibitori della PDE5 come il viagra o l’icariina dall’erba cornuta.[66]
Negli uomini con disfunzione erettile, valutata come debolezza dell’erezione (valutata con il punteggio di durezza yerettile[67]), la somministrazione di 1.500 mg di citrullina al giorno (due dosi da 750 mg) per un mese è stata in grado di apportare benefici alla metà dei 24 pazienti valutati (valutati come “molto soddisfatti” del trattamento), mentre il miglioramento del placebo è stato solo dell’8,3%.[68]
La citrullina sembra interagire con il metabolismo dei BCAA nell’organismo, anche se gli studi sull’uomo sembrano avere risultati diversi a seconda del contesto dello studio.
La citrullina può mediare positivamente la segnalazione della leucina attraverso mTOR, il che suggerisce teoricamente una sinergia. L’applicazione di questa combinazione ai sollevatori di pesi non è ancora stata studiata, quindi il sinergismo è attualmente solo un’ipotesi piuttosto che un fatto dimostrato.
Il nitrato è un piccolo donatore di ossido nitrico che costituisce il principale bioattivo del succo di barbabietola. Il nitrito sierico (forma ridotta del nitrato) sembra aumentare durante l’esercizio fisico in seguito al consumo di 6 g di citrullina malato, che si ritiene sia un indicatore dell’aumento della produzione di ossido nitrico.[20]
I farmaci a base di statine possono aumentare l’espressione dell’enzima che media la conversione dell’arginina in ossido nitrico e per questo motivo è possibile che vi sia un sinergismo per tutto ciò che riguarda l’ossido nitrico. Questo non è ancora stato testato in un sistema vivente.
L-Citrullina come sostituto alla L-Arginina?
L’integrazione di L-citrullina è stata definita un’alternativa alla L-arginina, in quanto ne aggira lo scarso assorbimento e si converte in L-arginina nei reni. La L-citrullina tecnicamente segue aumenti dose-dipendenti della L-arginina sierica fino a 15 g, ma la dose orale più alta di citrullina assunta ha ritorni sempre minori (cioè per ogni 5 g in più di citrullina ingerita si aggiunge meno arginina al siero).[85]
È stato osservato che la citrullina orale a 0,18 g/kg raddoppia approssimativamente l’arginina plasmatica (aumento del 100%)[86][87] o è leggermente superiore (123%) con 0,08 g/kg.[88] Poiché le dosi più elevate di L-citrullina hanno una minore conversione in arginina[85], è improbabile che la L-citrullina supplementare possa essere utilizzata per superare l’arginina per l’aumento acuto dell’arginina sierica.
Gli studi che hanno confrontato direttamente la L-arginina con la L-citrullina hanno osservato che entrambe aumentano la Cmax a livelli comparabili a dosi orali simili (Cmax di 79+/-8μM per 3 g di citrullina e 84+/-9μM per l’arginina), ma la citrullina risulta in un’AUC complessiva maggiore (48,7% in più rispetto all’arginina). [Questo potrebbe essere dovuto al fatto che, anche fino all’ingestione di 15g di citrullina, non si verifica un aumento significativo dell’escrezione di citrullina.[85] L’assenza di un aumento dell’eliminazione di L-citrullina dal sangue nonostante l’integrazione consentirebbe di avere un pool di L-citrullina disponibile per la conversione su richiesta in L-arginina.
Citrullina Malato:
La Citrullina Malato (CM), una combinazione di L-citrullina e acido malico, è stata pubblicizzata come un aiuto ergogenico (che aumenta l’energia) per l’allenamento contro-resistenza e l’esercizio ad alta intensità.
Come abbiamo visto, la L-citrullina è un precursore dell’ossido nitrico (NO), un vasodilatatore che può migliorare l’apporto di sangue e ossigeno ai muscoli durante l’esercizio. Tuttavia, le prove finora disponibili suggeriscono che il miglioramento del flusso sanguigno non è il meccanismo attivo degli effetti ergogenici del CM. Il meccanismo potrebbe invece essere dovuto alla capacità della citrullina di favorire l’eliminazione dell’ammoniaca durante l’esercizio ad alta intensità, alla capacità del malato di aumentare la produzione di ATP, a un aumento dell’espressione genica o a una maggiore efficienza della navetta malato-aspartato.
La maggior parte delle ricerche condotte finora ha utilizzato una dose acuta di 8 grammi di CM un’ora prima dell’esercizio. Sebbene l’assunzione di CM un’ora prima dell’esercizio rimanga la migliore raccomandazione, alcuni dati suggeriscono che dosi maggiori, fino a 15 grammi, potrebbero essere più benefiche.
È stato dimostrato che l’ingestione di una serie di dosi di CM (2-15 grammi) non ha effetti negativi sui marker ematologici. Sebbene la sicurezza di un’integrazione di CM a lungo termine richieda ulteriori indagini, le ricerche condotte finora indicano che la CM è ben tollerata nella maggior parte degli individui.
Ricerche preliminari hanno suggerito che 8 grammi di CM ingeriti un’ora prima dell’esercizio fisico aumentano la resistenza muscolare (ripetizioni fino al cedimento) in uomini e donne. Tuttavia, ricerche più recenti hanno suggerito che il CM potrebbe non avere un beneficio sulle prestazioni nell’allenamento contro-resistenza, potenzialmente a causa di variazioni nei tempi e nei dosaggi.[89]
Arginina AKG:
La differenza principale tra la L-arginina e Arginina AKG è che la L-arginina è un aminoacido non essenziale che l’organismo è in grado di produrre, mentre l’arginina AKG è un sale della L-arginina e dell’acido α-chetoglutarato. Inoltre, la L-arginina regola il flusso sanguigno attraverso la produzione di ossido nitrico, mentre l’Arginina AKG dovrebbe potenzialmente aumentare il flusso sanguigno, l’energia e il recupero.
Nella nutrizione sportiva, l’AKG è stato utilizzato come integratore per migliorare la sintesi proteica muscolare e diminuire la disgregazione muscolare, ed è quindi utilizzato dagli atleti per migliorare la composizione corporea.[90][91] L’integrazione di AKG potrebbe anche migliorare le prestazioni atletiche. Uno studio ha rilevato che un integratore di arginina e alfa-chetoglutarato (AAKG) ha migliorato la forza nella panca, ma non la capacità aerobica. Sono necessarie ulteriori ricerche per sostenere le affermazioni sull’AKG come aiuto ergogenico.[92]
L’AKG viene utilizzato anche per il recupero da interventi chirurgici o traumi, perché è un precursore dell’aminoacido glutammina. Sebbene la glutammina sia un aminoacido non essenziale, viene talvolta definita “condizionatamente essenziale” perché la quantità di glutammina di cui l’organismo ha bisogno per il recupero dopo un trauma significativo può superare la quantità che l’organismo è in grado di produrre. In questo caso, un integratore di AKG può aiutare il processo di recupero.[93][94]
L’AKG è stato proposto come integratore per la longevità; alcune ricerche condotte su vermi tondi, ratti e topi suggeriscono che potrebbe aumentare la durata della vita e ritardare l’insorgenza di malattie legate all’età, anche se gli studi clinici dovranno confermare questi risultati.[95][96]
Nelle persone con malattie renali croniche, in particolare in quelle sottoposte a dialisi, la somministrazione di AKG in combinazione con il calcio ha migliorato i biomarcatori della funzione renale.[97][98]
In uno studio è stato rilevato che l’AKG aumenta l’espressione di involucrina, filaggrina e serina palmitoil transferasi. Queste molecole sono tutte importanti per la struttura dello strato esterno della pelle e per l’idratazione dello strato esterno della pelle, quindi l’uso di AKG per via topica potrebbe migliorare l’aspetto della pelle.[99][100]
Nella ricerca, i dosaggi utilizzati variano da 3,6 g a 6 g, con dosaggi più elevati nelle persone che hanno subito ustioni, ma non è ancora stata stabilita una dose giornaliera raccomandata.[101] Poiché gli effetti sono dose-dipendenti, trovare una raccomandazione di dosaggio accurata sarà una parte importante della ricerca in corso.[102]
Sicurezza e tossicità:
La citrullina sembra essere ben tollerata dai ratti in dosi fino a 3 g/kg di peso corporeo[58][46]. Negli esseri umani, 15 g di citrullina assunti acutamente non sembrano causare diarrea o disturbi intestinali[22], il che è notevolmente diverso rispetto all’ornitina e all’arginina che possono causare diarrea a dosaggi di 10 g se assunti in bolo[74][75] a causa del limitato assorbimento di questi aminoacidi che poi procedono verso il colon causando diarrea osmotica.[74]
Il limite di sicurezza osservato, ovvero la dose più alta in cui si può essere relativamente sicuri che non si verifichino effetti collaterali nel corso della vita, è stato suggerito in 20 g di arginina al giorno in forma di integratore (al di sopra dell’assunzione di cibo).[103] Dosi più elevate sono state testate e ben tollerate, ma non esistono prove che suggeriscano la loro sicurezza in tutte le popolazioni nel corso della vita.
La L-arginina ha un tasso di assorbimento gastrointestinale piuttosto scarso. Può inoltre agire come assorbente, rilasciando acqua ed elettroliti nel lume intestinale attraverso la stimolazione dell’ossido nitrico e inducendo disturbi gastrici e diarrea.[12] Questo fenomeno è noto come diarrea osmolitica e tende a verificarsi a dosi orali superiori a 10 g circa, se assunte in bolo.[36]
Si pensa che ciò avvenga attraverso la stimolazione della produzione di ossido nitrico, poiché la D-Arginina (incapace di produrre NO) non produce diarrea[104] e l’ossido nitrico stesso è noto per essere un meccanismo attraverso il quale molti lassativi osmolitici funzionano.[105]
Singoli boli di 5-9 g di L-arginina senza cibo non sembrano causare disturbi intestinali come dosi superiori a 10 g,[36] suggerendo che, almeno per uno stomaco vuoto, il dosaggio di 9 g è un limite superiore.
Conclusioni:
L’Integrazione di L-citrullina si è dimostrata più redditizia tra costi e benefici (vedi assorbimento intestinale) rispetto alla L-arginina. L’uso alternativo di Citrullina Malato può portare ad eventi gastrointestinali più frequenti rispetto alla semplice forma L-citrullina. Nonostante la ridotta biodisponibilità orale della L-arginina, questa può essere mixata con L-citrullina per un effetto additivo, anche se non rappresenta un vero e proprio vantaggio proprio di tale abbinamento.
L’assunzione di L-arginina e/o L-citrullina vede la sua miglior tempistica prima dell’allenamento al fine di aumentare il flusso sanguigno ai distretti allenati, per effetto della vasodilatazione NO indotta.
Ciò si traduce in:
Aumento dell’apporto di nutrienti e ossigeno al tessuto muscolare abbinato ad un effetto di pulizia dalle molecole di scarto, come l’acido lattico;
Esaltazione dilatatoria sul reticolo venoso sottocutaneo, che migliora la qualità estetica in definizione.
Bonus: l’abbinamento con estratto di barbabietola notoriamente ricco di nitrati.
Come effetto diretto dell’introduzione nell’organismo di estratto di barbabietola abbiamo un aumento della sintesi di ossido nitrico (NOs), dovuta, per l’appunto, ai nitrati (NO3-) contenuti in questo vegetale, convertiti rapidamente in nitriti (NO2-2) tramite enzimi che si trovano fin dal tratto orofaringeo, gastrointestinale e tracheo-bronchiale. Dato ciò, la sintesi di NO sfrutta un percorso non usuale come quello della L-Arginina, ma coadiuvante a questa e alla L-Citrullina.
Le concentrazioni di nitrati raggiungono il picco dopo circa un’ora dalla sua ingestione, per ritornare ai livelli basali dopo quasi 24h, mentre gli effetti della L-Arginina permangono per almeno 75-80 minuti, per poi iniziare a tornare ai livelli basali.
“Citrulline – Compound Summary”. PubChem Compound. USA: National Center for Biotechnology Information. 16 September 2004. Identification. Retrieved 1 May 2012.
Chin RM, Fu X, Pai MY, Vergnes L, Hwang H, Deng G, Diep S, Lomenick B, Meli VS, Monsalve GC, Hu E, Whelan SA, Wang JX, Jung G, Solis GM, Fazlollahi F, Kaweeteerawat C, Quach A, Nili M, Krall AS, Godwin HA, Chang HR, Faull KF, Guo F, Jiang M, Trauger SA, Saghatelian A, Braas D, Christofk HR, Clarke CF, Teitell MA, Petrascheck M, Reue K, Jung ME, Frand AR, Huang JThe metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR.Nature.(2014-Jun-19)
Dell’Insulina ho già parlato abbondantemente non molto tempo fa attraverso la serie di quattro articoli dedicati a questo peptide nella ricorrenza dei cento anni dalla sua scoperta e prima applicazione in medicina (per approfondire clicca qui, qui, qui e qui). Nella quarta e ultima parte della prima citata serie di articoli, mi concentrai sulle pratiche protocollari dell’Insulina utilizzate in campo culturistico e, tra queste, vi era anche il protocollo di GH/Insulina nelle sue varianti applicative. Vi starete sicuramente chiedendo del perchè io scriva nuovamente di un argomento già trattato, oltretutto di recente. Beh, la risposta è abbastanza semplice: è un argomento che richiede per complessità degli approfondimenti aggiuntivi. Soprattutto alla luce delle “leggende da spogliatoio” e della limitatezza dei divulgatori “naturnazi”.
Come mi è già capitato di precisare, la visione dell’Insulina come ormone puramente anticatabolico è mentalmente riduttivo tanto quanto affermare che, di per se, l’Insulina sia “l’ormone anabolico per eccellenza”. Ed ecco perchè sopra ho specificato come sia necessario chiarire alcuni aspetti in contrapposizione al settarismo “magna e spigni” e “orcojo natty”. Sono volutamente ironico e provocatorio, ovviamente.
Giova anche ripetere che non sono l’unico a ritenere che la narrazione secondo la quale l’Insulina sia prettamente anti-catabolica sia simile alla visione precedente a quella di Bhasin (2001), secondo cui gli AAS non avrebbero effetti anabolici nel muscolo-scheletrico. Attualmente, la comunità (o, per meglio dire, “cupola”) scientifica vuole far credere che l’Insulina esogena non sia particolarmente anabolizzante nel muscolo-scheletrico umano. Le nozioni e i dati provenienti dai miei appunti sull’Insulina, in parte già riportati negli articoli dedicati prima citati, sono una forte contestazione di questo pregiudizio istituzionale (analogo alla visione pre-Bhasin sugli AAS). Il punto cieco istituzionale deriva dalla mancanza di studi sull’uso dell’Insulina esogena con:
dosaggio sovrafisiologico;
somministrazione sistemica;
iperamminoacidemia;
associazione a rhGH sovrafisiologico (e, più spesso, a AAS).
Ed è proprio su questi punti che si diramerà la mia disamina in questo articolo: valutazione teorico/pratica della co-somministrazione di GH e Insulina in condizioni sovra-fisiologiche.
Il motivo? Oltre le affermazioni arbitrarie e riduttive di alcuni divulgatori, vi è anche il risultato emerso dallo studio di Fryburg et al. i quali proposero che il rhGH e l’Insulina esogena lavorassero in modo incrociato, che la crescita fosse una conseguenza del turnover delle proteine muscolari piuttosto che dell’azione ormonale ipo/iperglicemica. La co-somministrazione di livelli endogeni (quindi fisiologici) di Insulina e rhGH sembrò dimostrare semplicemente una attenuazione degli effetti anabolici proteici del GH e che non fosse additiva. Ovviamente questa conclusione era limitata da: 1) il contesto di “picco ormonale” e della sua curva mantenuta in fisiologia e, cosa da non trascurare, 2) è stata trascurata l’importanza della disponibilità intracellulare di AA sulla sintesi proteica.
Ma andiamo per ordine…
Breve ripasso su azione dell’Insulina e meccanismi nell’ipertrofia muscolare:
L’effetto primario dell’Insulina nel muscolo scheletrico è sul gradiente elettrochimico di transmembrana, dove induce un’iperpolarizzazione nelle cellule muscolari attivando direttamente la pompa Na⁺-K⁺-ATPasi (46). [Biolo, G. et al. (1995)]. L’Insulina aumenta anche il numero di pompe Na+/K+-ATPasi nella membrana, determinando uno spostamento intracellulare del potassio e causando ipopotassiemia (basso livello di K/potassio) nello spazio extracellulare del sangue (siero).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
L’Insulina stimola l’espressione genica di MHCα (isoforma lenta) [Toniolo, L. (2005).] nel muscolo scheletrico e dell’albumina nel fegato (Dillman, 1988).
L’Insulina sembra stimolare in egual misura le fibre di tipo I e II [Albers et al. 2014]
Il muscolo scheletrico è un tessuto eterogeneo composto da diversi tipi di fibre. Gli studi suggeriscono che il metabolismo del glucosio mediato dall’insulina è diverso tra i vari tipi di fibre muscolari. Abbiamo ipotizzato che le differenze siano dovute all’espressione/regolazione specifica di elementi di segnalazione dell’insulina e/o di enzimi metabolici. Sono stati preparati pool di fibre di tipo I e II da biopsie dei muscoli del vasto laterale di soggetti magri, obesi e diabetici di tipo 2 prima e dopo un clamp iperinsulinemico-euglicemico. Le fibre di tipo I rispetto a quelle di tipo II presentano livelli proteici più elevati di recettore dell’insulina, GLUT4, esochinasi II, glicogeno sintasi (GS) e piruvato deidrogenasi-E1α (PDH-E1α) e un contenuto proteico inferiore di Akt2, TBC1 domain family member 4 (TBC1D4) e TBC1D1. Nelle fibre di tipo I rispetto a quelle di tipo II, la risposta di fosforilazione all’insulina era simile (TBC1D4, TBC1D1 e GS) o ridotta (Akt e PDH-E1α). Le risposte di fosforilazione all’Insulina aggiustate per il livello di proteine non erano diverse tra i tipi di fibre. Indipendentemente dal tipo di fibra, la segnalazione dell’insulina era simile (TBC1D1, GS e PDH-E1α) o ridotta (Akt e TBC1D4) nel muscolo di pazienti con diabete di tipo 2 rispetto a soggetti magri e obesi. Concludiamo che le fibre muscolari umane di tipo I rispetto a quelle di tipo II hanno una maggiore capacità di gestione del glucosio ma una sensibilità simile alla fosforegolazione da parte dell’Insulina.[Albers et al. 2014]
Livelli proteici più elevati di IRβ (+16%), esochinasi II impegna il glicogeno nella cellula, GLUT4 trasporta il glucosio nella cellula e complesso II della catena di trasporto degli elettroni ciclo mitocondriale/Kreb sono stati riscontrati nelle fibre di tipo I rispetto a quelle di tipo II.
Akt, mTOR: il contenuto proteico di Akt2 era più basso (-27%) nelle fibre di tipo I rispetto a quelle di tipo II. Gli aumenti medi della fosforilazione di [Akt] sotto stimolazione insulinica (ipertrofia) sono stati rispettivamente di 5,8 e 3,5 volte nelle fibre di tipo I e di 6,1 e 3,7 volte nelle fibre di tipo II. La risposta relativa all’insulina è stata simile tra i tipi di fibre.
Le fibre umane di tipo I hanno una maggiore abbondanza di trasporto (+29% GLUT4), fosforilazione (+470% HKII) e ossidazione (+35% complesso ETC II e +34% complesso piruvato deidrogenasi) del glucosio e di sintesi del glicogeno (+35%) rispetto alle fibre di tipo II.
Le fibre di tipo I possiedono una maggiore capacità di immagazzinamento del glicogeno.
Le differenze apparenti tra i tipi di fibre nella fosforilazione stimolata dall’insulina di Akt, NDRG1 (a valle di mTOR [queste sono le nostre vie dell’ipertrofia])… sono state eliminate quando sono state aggiustate per… l’abbondanza proteica. Questi risultati suggeriscono una sensibilità simile delle fibre muscolari di tipo I e di tipo II alla regolazione da parte dell’insulina delle proteine analizzate. [Albers et al. 2014]
Questi dati [Albers et al. 2014] supportano l’idea che, piuttosto che promuovere in modo preferenziale l’anabolismo proteico muscolare nelle fibre a contrazione lenta, entrambe le fibre muscolari di tipo I e II siano ugualmente sensibili agli effetti anabolici proteici dell’insulina esogena.
L’iperinsulinemia nei soggetti normali diminuisce acutamente le concentrazioni plasmatiche di aminoacidi come conseguenza del deposito netto di proteine. [Biolo, G., & Wolfe, R. R. (1993)].
Una volta secreta dalle beta-cellule pancreatiche, l’insulina circola nel flusso sanguigno con un’emivita di circa 12 minuti. Numerosi tessuti e organi esprimono il recettore dell’insulina e si attivano diverse azioni (Sonksen 2001), alcune delle quali sono di importanza generale e di particolare interesse per gli sport d’élite.
L’effetto centrale dell’insulina, in concerto con altri ormoni come il glucagone o la somatostatina, è il controllo dei livelli di glucosio nel sangue. La secrezione di insulina in risposta a concentrazioni elevate di glucosio (ad esempio, postprandiale) inibisce la produzione epatica di glucosio, abbassando così i livelli di glucosio nel sangue. Il glucagone, invece, composto da 29 residui aminoacidici, è un ormone controregolatore dell’insulina. Aumenta i livelli di glucosio plasmatico in risposta all’ipoglicemia indotta dall’insulina e svolge un ruolo importante nell’omeostasi del glucosio aumentando la gluconeogenesi e diminuendo la glicolisi. Il terzo ormone pancreatico rilevante per il controllo dei livelli di glucosio nel sangue è la somatostatina, un peptide di 14 residui che esercita effetti inibitori sulla secrezione di insulina ma non sulla sua biosintesi. L’intero meccanismo della sua azione non è ancora stato chiarito, ma gli studi suggeriscono un effetto paracrino che inibisce l’esocitosi dell’insulina dalle cellule adiacenti alle cellule D produttrici di somatostatina (Reichlin 1983).
Tuttavia, gli effetti dell’insulina sull’intero organismo sono molteplici e complessi. L’insulina provoca, ad esempio, la traslocazione del GLUT-4 (il trasportatore di glucosio che si trova prevalentemente nel muscolo scheletrico e nel tessuto adiposo) dalle vescicole intracellulari alla membrana cellulare e, quindi, aumenta la velocità di ingresso del glucosio per una determinata concentrazione nel tessuto bersaglio. Un eccesso di glucosio trasferito nelle cellule stimola successivamente la formazione di glicogeno (Halse et al. 2001; Yeaman et al. 2001), un fatto di notevole interesse negli sport di resistenza, dove la quantità di glicogeno immagazzinata nelle cellule muscolari può influenzare le prestazioni atletiche. Inoltre, il metabolismo proteico (muscolare) è significativamente influenzato dalle proprietà chaloniche (Rooyackers e Nair 1997; Sonksen 2001) e stimolanti dell’insulina (Biolo et al. 1995; Biolo e Wolfe 1993; Tipton e Wolfe 2001; Wolfe 2000, 2005). Grazie all’effetto anticatabolico dell’insulina, la disgregazione proteica è significativamente ridotta e consente la conservazione degli elementi muscolari contrattili. Inoltre, in numerosi studi che hanno dimostrato le proprietà anaboliche dell’insulina, sono stati osservati effetti di stimolazione della sintesi, utilizzando ad esempio analoghi marcati con isotopi stabili. L’anabolismo, tuttavia, è fortemente dipendente dalla disponibilità di aminoacidi (Fujita et al.2006; Garlick e Grant 1988; Zhang et al.1999).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
Giova anche sottolineare i punti chiave dell’effetto dell’Insulina sul metabolismo delle proteine:
aumenta la velocità di trasporto di alcuni aminoacidi nei tessuti
aumenta il tasso di sintesi proteica nel muscolo, nel tessuto adiposo, nel fegato e in altri tessuti
diminuisce il tasso di degradazione delle proteine nel muscolo (e forse anche in altri tessuti)
diminuisce la velocità di formazione dell’urea. Questi effetti dell’insulina servono a favorire la sintesi di carboidrati, grassi e proteine.[Newsholme, E., & Dimitriadis, G. (2001).]
Un ormone anabolico può indurre l’ipertrofia del muscolo scheletrico attraverso:
l’aumento della MPS o
la riduzione della proteolisi.
Inoltre, il trasporto transmembrana degli AA (AA trafficking) può essere sotto controllo ormonale.
L’Insulina induce l’ipertrofia del muscolo scheletrico attraverso tutti e tre questi aspetti del metabolismo proteico muscolare:
profondamente (“il tasso di sintesi frazionale delle proteine muscolari era superiore del 65% (P = 0,02) durante l’infusione di insulina”) [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)].
in misura modesta (nelle proteine non miofibrillari del muscolo scheletrico)
influenzando l’attività di almeno quattro distinti sistemi di trasporto degli AA (non è un mediatore primario dell’azione dell’insulina sull’ipertrofia del muscolo scheletrico):
Primariamente: trasporta gli AA con catene laterali polari corte, come l’Alanina e la Glicina; non limita il ritmo della sintesi proteica.
ASC
Nᵐ, sostanzialmente
Xc [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)]
In pazienti gravemente ustionati l’infusione di Insulina (10 – 12 UI/ora, paziente di 70 kg) ↑MPS senza influenzare la MPB.[Hadley JS et al.2002]. Nessun cambiamento nell’assorbimento degli AA nel muscolo scheletrico, suggerendo che gli effetti anabolici muscolari derivino da un più efficiente riutilizzo degli AA generati dalla proteolisi.[Hadley JS et al.2002]
La sintesi proteica del muscolo scheletrico indotta dall’Insulina è modulata dalle variazioni di:
del flusso sanguigno muscolare e
disponibilità di AA. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)]. In altre parole, l’insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in mancanza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)].
Adesso è giunto il momento di aprire una parentesi sul GH…
Effetti dell’Ormone della Crescita e del IGF-1 sulla sintesi proteica:
Struttura molecolare di hGH
La maggior parte degli studi svolti sul GH, quando considerati nel loro insieme, suggeriscono che il peptide in questione sia anabolico. Più specificamente, il GH è anabolico perché stimola la sintesi proteica di tutto il corpo con o nessun effetto, o un effetto soppressivo, sui tassi di degradazione proteica. [Møller N et asl. 2007] Tuttavia, quando si approfondisce l’argomento, le cose diventano un po’ meno chiare dal momento che i risultati degli studi tendono ad essere diversi. I diversi risultati sono un riflesso diretto dell’ immensa complessità del GH.
Il GH esplica i suoi effetti sulla sintesi proteica legandosi prima con il suo recettore specifico (GHR) e successivamente aumentando la trascrizione del gene muscolare attraverso i percorsi di segnalazione a valle, in definitiva attivando la segnalazione del mTOR. [Hayashi AA et al.2007] Questi effetti si manifestano in acuto, spesso si verificano in pochi minuti e sono di natura simile all’Insulina, usando molte delle stesse vie anaboliche. [Costoya JA et al.1999] La rapida comparsa di questi cambiamenti metabolici legati alle proteine suggerisce che essi siano direttamente causati dal GH e non secondariamente mediati tramite l’IGF-1 [Copeland KC et al.1994]. L’impatto del GH sulla proteolisi, d’altro canto, è molto probabilmente di natura indiretta. A detta di tutti, ciò ha più a che fare con i suoi effetti inibitori sull’Insulina, che è stata vista avere effetti diretti sulla proteolisi.[Umpleby AM et al.]
È stato dimostrato che l’IGF-1 inibisce allo stesso modo la disgregazione proteica a livello sistemico [Fryburg DA et al.1994], il che avrebbe senso a causa della stretta correlazione con il GH. Quando gli amminoacidi e l’Insulina vengono somministrati ai soggetti esaminati, già sottoposti a somministrazione di IGF-1, è stato dimostrato, sia negli uomini che negli animali, che i tassi di sintesi proteica aumentano a livello sistemico [Jacob R et al.1996]. Vale la pena notare che l’IGF-1 è bifasico nel senso che quando è somministrato ad alto dosaggio e, di conseguenza, i livelli serici diventano elevati, il suo comportamento cambia passando da un azione “GH-simile” ad una “insulino-simile”.
Per riassumere, il GH è molto adatto per prevenire la degradazione proteica, e lo fa in una vasta gamma di condizioni alimentari a ristretto apporto calorico. Tuttavia, in presenza di un apporto energetico sufficiente (e di AA), il suo comportamento cambia. L’effetto principale del GH sul metabolismo proteico è volto dapprima a creare un ambiente con una ossidazione amminoacidica ridotta [Buijs MM et al.2002] e successivamente ad aumentare la sintesi proteica sistemica. [Gibney J et al.2005]
L’Ormone della Crescita è noto per aumentare i livelli di IGF-1 circolante così come la sintesi locale di IGF-1, in modo autocrino. Entrambe queste azioni giocano un ruolo fondamentale nella regolazione della massa muscolare e, quindi, in conseguenza di ciò, risulta utile comprendere meglio come la secrezione di GH porta ad un aumento dei livelli endocrino e autocrino di IGF-1.
La stragrande maggioranza dell’Ormone della Crescita negli adulti sani è secreto dalla ghiandola pituitaria e, più specificamente, dalle cellule somatotrope nel lobo anteriore mediate dal fattore di trascrizione Prophet of Pit-1 (PROP1).[Hemchand K et al.2011] Il GH può anche essere sintetizzato localmente in molti tessuti come il cervello, le cellule immunitarie, il tessuto mammario, i denti e la placenta che sono tutti al di fuori della regolazione dell’ipofisi. [Waters MJ et al.1999] Questo supporta l’idea secondo cui il GH abbia ruoli autocrini oltre ai suoi già consolidati ruoli endocrini.
Il GH circolante si lega al GHR, recettore dell’Ormone della Crescita appartenente alla superfamiglia delle proteine transmembrana presenti in tutte le cellule del corpo e che include il recettore della Prolattina e un certo numero di recettori delle citochine [Zou L et al. 1997]. I livelli sulla superficie cellulare, o la densità recettoriale, dei GHR sono il determinante principale dell’affinità di legame del GH con le cellule. La traslocazione del GHR, cioè il recettore che si sposta dal nucleo di una cellula alla sua membrana esterna, è direttamente inibita dall’IGF-1 – che è uno dei molti meccanismi di feedback che esistono tra questi ormoni strettamente correlati. Mediante l’inibizione della traslocazione dei GHR, l’IGF-1 contribuisce direttamente ad abbassare la reattività di queste cellule a uno stimolo esterno di GH. [Leung KC et al.1997]
I GHR esistono sulle membrane cellulari come omodimeri preformati e inattivi. Questo significa che il GHR ha due dimeri identici del recettore della proteina, e questi omodimeri saranno sempre accoppiati al JAK2 quando sono privi di attività enzimatica. Questo accoppiamento al JAK2 provoca un’azione inibitoria complessiva sul recettore. [Sawada T et al.2017] In altre parole, il GHR rimane dormiente finché non viene attivato come parte del processo di legame GH/GHR. Quando una molecola di GH si lega al GHR, si verifica un cambiamento strutturale all’interno del GHR che si traduce in movimento effettivo dei domini intracellulari del recettore separatamente l’uno dall’altro. Questo smorza quell’azione inibitoria data dal JAK2 e consente loro di attivarsi l’un l’altro.[Brown RJ et al.2005]
Successivamente, la molecola di GH si lega sequenzialmente a uno dei due omodimeri di GHR e il completamento di questo processo di legame facilita le interazioni con il secondo omodimero. Dopo questo, i domini intracellulari di questo dimero-GHR appena formato subiscono una rotazione effettiva. La rotazione del nuovo dimero-GHR consente ai domini chinasi del JAK2 di essere in contatto l’uno con l’altro, consentendo loro di transactivare e ciascuno successivamente si lega a una molecola di JAK2. [Lanning NJ et al.2006] Ciascuna molecola di JAK2 eseguirà quindi la fosforilazione incrociata (attivazione) dei residui di Tirosina, e sono proprio questi residui che formano “punti di attracco” per molte delle diverse molecole di segnalazione che costituiscono le vie di segnalazione a valle, e alla fine portano all’espressione genica. [Brooks AJ et al.2010] Uno dei più importanti percorsi a valle di maggior interesse per l’argomento trattato è il percorso JAK-STAT. Questo percorso è di vitale importanza sia per la trascrizione epatica di IGF-1 dal GH che per molti dei processi anabolici mediati dal GH all’interno del tessuto muscolare.
IGF-1:
Struttura molecolare di IGF-1
Gli IGF sono una famiglia di peptidi, in gran parte dipendenti dal GH, che mediano molte delle azioni di stimolo della crescita date dal GH.[Cohen P. et al.2006] Il fegato è il principale responsabile di tutta la produzione endocrina di IGF-1, con circa il 75% della sintesi a carico epatico sotto la regolazione del GH.[Laron Z. et al. 2001] Ciò presuppone che ci sia un apporto macro-calorico sufficiente e livelli elevati di Insulina portale.[D’Ercole AJ et al.1984] La sintesi autocrina di IGF-1 è anche regolata dal GH, in aggiunta ad altri fattori autocrini dipendenti dal tessuto.[Gunawardane K et al.2000]
Alla famiglia degli IGF appartengono oltre dieci proteine strutturalmente simili tra cui IGF-1, IGF-2, Insulina, Relaxina e Pro-Insulina.[Lu C et al.2005] Sono tutti altamente omologhi sia nella struttura che nella funzione e gli effetti metabolici dell’IGF-1 sono stati definiti “insulino-simili” proprio a causa delle somiglianze e dei percorsi che condividono l’uno con l’altro. L’IGF-1 ha un’omologia di sequenza aminoacidica superiore al 50% con l’Insulina e il recettore del IGF-1 ha un’omologia della sequenza aminoacidica del 60% con il recettore dell’Insulina.[Samani AA et al.2006] A causa di queste somiglianze strutturali, i membri della famiglia IGF possono spesso legarsi con i recettori nativi in modo “incrociato”.[Kim JJ et al.2002] Per riassumere brevemente queste relazioni di legame, la molecola di IGF-1 si lega con il recettore del IGF-1 con un elevata affinità, tuttavia sia l’IGF-2 che l’Insulina possono legarsi al recettore del IGF-1, ma con una affinità significativamente inferiori. L’IGF-2 si lega al recettore del IGF-2 con una elevata affinità, e l’IGF-1 si lega a questo recettore con un’affinità inferiore mentre l’Insulina non presenta alcun legame con esso.
La famiglia dei recettori IGF ha densità che variano significativamente in base ai tipi di cellule in cui sono presenti.[Clemmons DR. et al. 2012] Questo è uno dei motivi per cui l’Insulina e l’IGF-1 possono avere diverse azioni metaboliche nonostante siano strutturalmente simili. Cellule come gli epatociti e gli adipociti hanno molti più recettori dell’Insulina rispetto ai recettori del IGF-1. Al contrario, le cellule muscolari lisce vascolari situate nei vasi sanguigni hanno un numero significativamente più elevato di recettori del IGF-1 rispetto ai recettori dell’Insulina.
E’ necessario comprendere che la famiglia dei recettori IGF è anche attivata dalla Tirosina Chinasi che, come ora sappiamo, porta alla fosforilazione dei substrati, all’attivazione delle vie cellulari e infine all’espressione genica e alla sintesi proteica.[Cohen P. et al.2006] L’attivazione del recettore del IGF-1 sembra essere indipendente dall’isoforma da cui è stato prodotto l’IGF-1. Inoltre, si noti che entrambi i tipi di recettori IGF sono stati trovati nelle cellule muscolari umane.[Shimizu M et al.1986]
Nel flusso sanguineo, l’IGF-1 esiste principalmente in forma legata a proteine leganti l’IGF (IGFBP). La superfamiglia IGFBP comprende sei proteine ad alta affinità che vanno dal IGFBP-1 al IGFBP-6, nonché un certo numero di proteine a bassa affinità denominate proteine legate all’IGFBP.[Hwa V et al.1999] Quasi il 95% di tutto l’IGF-1 circolante esiste in forma legata, con circa il 75% legato specificamente con l’IGFBP-3.[Firth SM et al.2002] Una piccola frazione di IGF-1 (normalmente inferiore al 5%) può anche esistere in forma libera, e queste molecole non legate agiscono come regolatore negativo della secrezione di GH. Gli IGFBP possono legarsi con l’IGF-1 e l’IGF-2, ma non con l’Insulina. [Bach LA et al.1993] L’IGF-1 legato esiste più comunemente in un complesso ternario da 150-kDa mentre è nel circolo ematico. Questo complesso ternario è costituito da una molecola di IGF-1, dal IGFBP-3 e dalla subunità labile acida (ALS) – sebbene possa esistere in un complesso binario con altri IGFBP.[Duan C. et al.1996] Questi complessi servono a scopi come l’aumento della biodisponibilità degli IGF circolanti, estendendo la loro emivita serica, trasportando gli IGF alle cellule bersaglio e modulando l’interazione degli IGF con i loro rispettivi recettori di membrana posti sulla superficie delle cellule.[Hwa V et al.1996] Ad esempio, nel plasma, il complesso ternario stabilizza IGF-1, aumentando significativamente la sua emivita da meno di 5 minuti a oltre 16 ore in alcuni casi.[Firth SM et al. 2002]
Struttura molecolare IGFBP-1
Gli IGFB sembrano normalmente inibire l’azione degli IGF, e questo perché competono con i recettori IGF per l’affinità di legame con gli IGF.[Collett-Solberg PF et al.2000] Tuttavia, non è sempre così, poiché gli IGFBP sono anche in grado di potenziare le azioni dell’IGF, potenzialmente facilitando la consegna dell’IGF al recettore.[Wetterau LA et al. 1999] Sebbene esista un’interazione piuttosto complessa, basti ricordare che il ruolo principale degli IGFBP è quello di trasportare gli IGF dal flusso ematico ai tessuti periferici. Una volta che ciò è avvenuto, gli IGFBP vengono rilasciati dai complessi binari e ternari mediante proteolisi o tramite legame alla matrice extracellulare del recettore del IGF-1.[Parker A et al.1999] Una volta rilasciate, le molecole di IGF-1 diventano libere, attive e possono quindi esplicare la loro azione.[Monzavi R et al.2002]
Una volta nei tessuti, gli IGFBP modulano le azioni dell’IGF in quanto hanno una maggiore affinità per il sito recettore rispetto all’IGF stesso [Velloso CP et al.2008], tuttavia essi possono anche esercitare effetti indipendenti dall’IGF.[Jones JI et al.1995] Alcuni degli effetti diretti del IGFBP che sono già stati chiariti includono l’inibizione della crescita, l’induzione diretta dell’apoptosi e la modulazione degli effetti dei fattori di crescita non-IGF.[Cohen P. et al.2006]
Additività Insulina/rhGH sovrafisiologici:
Come ho già riportato in passato, la somministrazione di Insulina esogena capace di creare un livello ematico oltre i 1.200pmol/lt ha effetti significativi e tangibili sulla sintesi proteica muscolare. Ma non si esaurisce tutto a meri e limitati numeri statistici.
La sintesi proteica nel muscolo-scheletrico indotta dall’Insulina è modulata dalle variazioni di:
1- del flusso sanguigno muscolare e
2- disponibilità di AA. [Fujita, S. et al. (2006).]. In altre parole, l’Insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in assenza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S. et al. (2006)].
l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1, riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1. L’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1. Il discorso precedentemente fatto sulle IGFB deve essere compreso correttamente alla luce di modifiche sostanziali in concentrazione (di IGF-1) e sua frazione legata (minoritaria ma presente).
Quindi, l’Insulina può aumentare la biodisponibilità di IGF-I inibendo la produzione di IGFBP-1 e aumentando l’attività proteolitica di IGFBP-3, che riduce l’affinità di IGF-I per IGFBP-3. [Hadley JS et al. 2002]. Esemplificato in formula:
IGF-I (libero) ↑AA e assorbimento del glucosio e ripartizione delle proteine.[Hadley JS et al. 2002]
L’Insulina dirige la MPS attraverso l’attivazione diretta della via PI3K/Akt/mTORC1 e grazie all’aumento del flusso sanguigno muscolare tramite proprietà vasoattive.[Trommelen et al. 2015]
Variabile della disponibilità intracellulare di AA:
Come già alcuni di voi sapranno, la prima fase della sintesi proteica avviene nel nucleo delle cellule e comporta la trascrizione di un gene specifico in molecole di mRNA. Gli mRNA sono trasportati nel citosol, dove si associano ai ribosomi per la traduzione della sequenza di basi in una sequenza di AA. Il processo di traduzione può essere suddiviso in tre fasi: iniziazione, allungamento e terminazione della catena peptidica. Molti ribosomi possono associarsi a molecole di mRNA, formando poliribosomi. Un poliribosoma può sintetizzare diverse catene peptidiche da un singolo mRNA. Gli AA liberi nel citoplasma non vengono utilizzati direttamente per la sintesi proteica. Il processo di traduzione prevede il legame degli AA a specifiche molecole di RNA di trasferimento (tRNA), formando gli aminoacil-tRNA. Gli aminoacil-tRNA costituiscono pool molto piccoli che si trasformano rapidamente. [Biolo, G., & Wolfe, R. R. (1993)]
È stato dimostrato che l’insulina stimola sia i processi di trascrizione che di traduzione di proteine specifiche (Kimball e Jefferson, 1988). È stato dimostrato che l’espressione genica (riflessa dai livelli di mRNA) di molte proteine è stimolata dall’insulina, tra cui l’albumina nel fegato e l’MHC α nel muscolo scheletrico (Dillman, 1988). Il contenuto di mRNA riflette il potenziale ultimo di sintesi proteica nella cellula (tessuto), ma potrebbe non essere direttamente correlato al tasso di traduzione e quindi di sintesi delle proteine. L’insulina migliora anche il processo di traduzione nel muscolo scheletrico, stimolando l’iniziazione della catena peptidica (Jefferson et al, 1974; Fulks e Goldberg, 1975). Pertanto, da una base molecolare, ci si aspetterebbe che in vivo l’insulina aumenti in generale la capacità di sintesi proteica (contenuto di mRNA) e stimoli la traduzione e la produzione di proteine specifiche, come quelle muscolari. [Biolo, G., & Wolfe, R. R. (1993)]
Esistono due meccanismi attraverso i quali l’aumento dell’AA intracellulare può stimolare la sintesi proteica muscolare. In primo luogo, un effetto di massa dovuto agli AA esogeni forniti; il tRNA necessario per la traduzione degli AA in proteine è disponibile nel muscolo in quantità superiori al pool di AA disponibile. Di conseguenza, l’aumento della disponibilità di AA spingerà la MPS semplicemente caricando più tRNA. [Tipton, K. D., & Wolfe, R. R. (2001).].
È anche possibile che alcuni AA, singoli o gruppi di AA (ad esempio, i BCAA), segnalino l’avvio del processo di traduzione (ribosomiale). Rennie et al. hanno dimostrato che i singoli AA essenziali somministrati in dosi massicce (cioè riempiendo lo spazio AV e lo spazio intracellulare) stimolano la MPS, mentre gli AA non essenziali non hanno questo effetto. Inoltre, abbiamo recentemente dimostrato che non è necessario includere gli AA non essenziali in una soluzione che simula l’anabolismo muscolare in volontari a riposo . [Tipton, K. D., & Wolfe, R. R. (2001).].
Questi studi sono coerenti con l’ipotesi che uno o più degli AA essenziali possano agire come segnale per l’avvio delle MPS. Tuttavia, la stimolazione della sintesi proteica attraverso l’azione di massa non può essere esclusa solo sulla base di queste evidenze. È possibile che siano presenti sufficienti AA non essenziali e che l’aggiunta di grandi quantità di AA essenziali fornisca una carica di tRNA sufficiente a guidare la sintesi proteica. [Tipton, K. D., & Wolfe, R. R. (2001)].
Il punto di vista di Wolfe: L’insulina sembra aumentare l’efficienza della reincorporazione degli AA derivanti dalla disgregazione delle proteine (intracellulare) nelle proteine di nuova sintesi. [Biolo et al. (1995)] [Wolfe, R. R. (2000)].
Disponibilità di AA come segnale per attivare la traduzione
Sistema ubiquitina-proteosoma (sistema MPB predominante nel muscolo a riposo)
I fattori di iniziazione eucariotici (eIF), in particolare eIF4E, sono componenti importanti del controllo dell’iniziazione della traduzione. Quando eIF4E forma un complesso con eIF4G, viene promossa l’iniziazione della traduzione. Il legame di eIF4E con eIF4G è normalmente impedito perché eIF4E è legato a una proteina vincolante, eIF4E-BP1, la cui fosforilazione libera eIF4E. L’apporto di AA favorisce la sua fosforilazione, permettendo così alla sintesi proteica di procedere . In altre parole, la formazione del complesso attivo eIF4E-eIF4G aumenta in risposta alla somministrazione di proteine. [Tipton, K. D., & Wolfe, R. R. (2001)].
La disponibilità di AA come chalonico per la proteolisi muscolare
Anche il pool intracellulare di AA sembra seguire un effetto di massa per evitare la disgregazione delle proteine muscolari.
Un aumento degli AA intracellulari inibisce la MPB solo in determinate condizioni. Ad esempio, l’iperamminoacidemia a riposo ha un effetto minimo o nullo sulla MPB. Tuttavia, quando gli AA vengono somministrati dopo la RT, quando la MPB sarebbe normalmente elevata, non si verifica alcun aumento della MPB. Questa inibizione condizionale della MPB è coerente con l’idea che questa relazione tra disponibilità di AA e riduzione della MPB si verifichi principalmente quando il pool intracellulare di AA disponibili si esaurisce. Ad esempio, se la risposta iniziale all’esercizio fisico è un aumento della MPS, il pool di AA si riduce, portando alla MPB per mantenere il pool intracellulare di AA. [Tipton, K. D., & Wolfe, R. R. (2001)].
In alternativa, poiché esistono diverse vie di MPB, è possibile che in seguito all’esercizio fisico la via lisosomiale diventi predominante nel mediare l’accelerazione della MPB, e che questa via sia reattiva alla disponibilità di AA [e all’insulina esogena!] [Tipton, K. D., & Wolfe, R. R. (2001)].
…L’unico effetto probabile dei carboidrati e dei grassi sul bilancio proteico netto è l’influenza ormonale come substrati [ad esempio, l’influenza dei CHO sulla secrezione di Insulina]. [Tipton, K. D., & Wolfe, R. R. (2001)].
Trasporto transmembrana AA
Alanina +48%, Lisina +75%, Leucina +22%. [Biolo et al. (1995)].
L’alanina è uno dei principali substrati del sistema A, un sistema sodio-dipendente che mantiene ripidi gradienti transmembrana delle concentrazioni di AA ed è regolato dall’insulina [Biolo et al. (1995)] . Inoltre, la sintesi di alanina nella cellula muscolare aumenta a causa dell’incremento insulino-mediato dell’assorbimento di glucosio e della produzione intracellulare di piruvato nel muscolo. [Biolo et al. (1995)].
La lisina è un AA cationico che viene trasportato dal sistema sodio-indipendente y⁺ che è fortemente influenzato dal potenziale elettrochimico della membrana cellulare. Poiché l’insulina induce un’iperpolarizzazione nelle cellule muscolari scheletriche attivando direttamente la pompa Na⁺-K⁺-ATPae, l’accelerazione del trasporto della lisina può essere secondaria all’effetto primario dell’insulina sul gradiente elettrochimico di transmembrana. [Biolo et al. (1995)].
Gli AA a catena ramificata (leucina, isoleucina e valina) e aromatici (fenilalanina e tirosina) vengono trasportati preferibilmente attraverso il sistema L. Questo sistema, indipendente dal sodio, non è in grado di generare elevati gradienti transmembrana per i suoi substrati. È stato dimostrato che le caratteristiche cinetiche del sistema L non sono influenzate dall’insulina. [Biolo et al. (1995)].
L’azione dell’insulina sulla leucina è particolarmente modulata dall’aumento del flusso sanguigno muscolare! La leucina è soggetta a ossidazione e transaminazione che devono essere sottratte dal tasso di scomparsa nel muscolo per ottenere i tassi di sintesi proteica con la tecnica del bilancio arteriovenoso. [Rooyackers et al. (1997).].
In presenza di un adeguato apporto di AA, il sistema degradativo più importante nel muscolo è il sistema dell’ubiquitina indipendente dall’ATP. [Biolo et al. (1995)]. Questo sistema non è sensibile all’insulina [Biolo et al. (1995)]. L’insulina sembra avere un ruolo solo nella regolazione dell’attività dei lisosomi [Biolo et al. (1995)]. I lisosomi (organelli) non sono coinvolti nella degradazione delle proteine miofibrillari in condizioni normali, ma solo in presenza di bassi livelli di insulina o di una ridotta disponibilità di AA. [Biolo et al. (1995)].
La riduzione della degradazione proteica del muscolo scheletrico indotta dall’insulina si verifica in proteine non miofibrillari (soprattutto epatiche). [Rooyackers et al. (1997)].
E’ utile inoltre ricordare che il GH aumenta sia l’assorbimento degli AA dal lume intestinale che a livello cellulare nel muscolo-scheletrico, azione, quest’ultima, che va a sommarsi a quella esercitata da IGF-1 e Insulina.
Conclusioni:
Qual conclusione allora?
Oggettivamente, parlare di protocolli di Insulina/hGH ha senso quando l’atleta in questione è un avanzato, con una carriera “Natty” alle spalle ad OC e una esperienza ben controllata e graduale con i PEDs. Ma andando oltre a ciò che dovrebbe essere scontato, da quanto riportato in questo articolo, si può ben capire che essa rappresenti una pratica con un margine di guadagno in termini ipertrofici decisamente significativo.
I punti chiave degli effetti consequenziali del protocollo Insulina/hGH sono:
Aumento della sensibilità epatica al GH per via dell’attività insulinica di picco;
risposta massiva consequenziale sulla sintesi e rilascio sistemico di IGF-I;
riduzione delle proteine di trasporto IGFBP-1 e 3 insulino-correlato;
aumento della frazione libera e attiva di IGF-I.
I limiti della presente disamina non sono pochi basandosi in gran parte su dati teorici ed estrapolazioni aneddotiche tratte da piccoli casi studio. Ma ciò nonostante è sufficiente a far comprendere, almeno a livello base, come le modifiche dell’omeostasi attraverso l’uso di farmaci possa dare risultanti anche di molto distanti da quelle ottenute in contesti fisiologici.
Inoltre, se volessimo trovare un altro limite al presente lavoro, non sono stati trattati gli effetti sommativi dati dalla co-assunzione di AAS. Ciò è ovviamente legato al fatto di non volere disperdere l’attenzione del lettore dal concetto fondamentale trattato: la validità dei protocolli Insulina/hGH.
Sicuramente, i vantaggi del protocollo Insulina/hGH vanno ben oltre il banale e poco sensato assunto secondo il quale l’Insulina esogena vada semplicemente a compensare in un certo senso la ridotta sensibilità all’Insulina GH correlata.
Nella seconda parte di questa serie di articoli dedicati alla alopecia androgenetica (e, quindi, anche AAS-correlata) ho discusso le modalità di trattamento più convenzionali per tale condizione , ovvero le versioni orali e topiche di Finasteride e Minoxidil con accenni alla Dutasteride. In questo articolo ne illustrerò alcune di più nuove o sperimentali, come gli Antagonisti topici del Recettore degli Androgeni, la terapia con plasma ricco di piastrine (PRP), i modulatori del segnale di Wnt e le Prostaglandine.
Antagonisti del Recettore degli Androgeni:
L’obbiettivo ottenuto con gli Antagonisti del Recettore degli Androgeni è simile a quello degli inibitori della 5α-reduttasi, come la Finasteride e la Futasteride: ridurre l’azione androgenica. Il meccanismo è tuttavia diverso. Gli inibitori della 5α-reduttasi bloccano la conversione del Testosterone nel più potente androgeno Diidrotestosterone (DHT). In questo modo, l’effetto androgeno del Testosterone non viene amplificato nel tessuto del cuoio capelluto. Gli Antagonisti del Recettore degli Androgeni bloccano l’azione androgena impedendo agli androgeni di legarsi al loro recettore. In questo modo, la loro azione viene bloccata a livello del Recettore degli Androgeni stesso, e quindi si rivolge praticamente a tutti gli androgeni piuttosto che a quello specifico, come nel caso degli inibitori della 5α-reduttasi. Il problema è che i suoi effetti devono rimanere localizzati al cuoio capelluto. Bloccare l’azione complessiva degli androgeni in altri tessuti, come quello muscolare, è decisamente indesiderato.
Rappresentazione grafica semplificata dell’attività degli inibitori della 5α-reduttasi e degli Antagonisti del Recettore degli Androgeni.
Uno di questi farmaci, attualmente in fase di sperimentazione clinica, è il Clascoterone (Breezula). La ricerca è condotta dall’azienda farmaceutica Cassiopea S.p.A. . Ricerche in vitro su cellule di papilla dermica umana hanno dimostrato che il composto è efficace nell’inibire l’azione degli androgeni [1]. Lo fa in misura maggiore rispetto all’Enzalutamide, un altro antagonista del recettore degli androgeni utilizzato nel trattamento del cancro alla prostata, e in misura paragonabile alla Finasteride. L’affinità per il AR è relativamente bassa, circa 100 volte inferiore all’affinità del DHT per il AR [2]. Questo non è un vero problema, si può rimediare semplicemente assicurandosi che le cellule del follicolo pilifero siano esposte a una concentrazione sufficientemente alta della molecola. Tuttavia, ci si chiede quale sia la sua affinità per altri recettori steroidei, come quello dei glucocorticoidi. Se non ha una specificità sufficientemente elevata per il Recettore degli Androgeni, si possono ottenere effetti fuori bersaglio legandosi a questi altri recettori. A sua volta, questo può portare a effetti collaterali. Anche questo non è necessariamente un problema se l’esposizione sistemica è minima o inesistente.
Nell’agosto 2020, la FDA ha approvato il Clascoterone crema 1 % (Winlevi) per il trattamento dell’acne vulgaris in pazienti di età pari o superiore a 12 anni [3]. È quindi in linea con le aspettative utili al fine di ottenere l’approvazione anche per l’alopecia androgenetica. Infatti, nel 2019 è stato completato uno studio di fase 2 su 404 uomini per il trattamento dell’alopecia androgenetica (EudraCT #2016-003733-23).
I soggetti sono stati trattati con una soluzione da 1mL di Clascoterone al 2,5, 5,0 o 7,5% da applicare due volte al giorno, oppure 0,0 (veicolo) e 7,5% una volta al giorno, o veicolo due volte al giorno, per un anno. Sebbene i risultati non siano stati pubblicati nella letteratura scientifica, possono essere consultati online nel registro degli studi clinici dell’UE. Il numero totale di peli nell’area è aumentato in modo significativo rispetto al gruppo con soluzione veicolante in tutti i gruppi di trattamento. (Soprattutto perché nel gruppo con soluzione veicolante si è verificata una diminuzione significativa del numero totale di capelli, che riflette la progressione dell’alopecia androgenetica). È interessante notare che le valutazioni della crescita dei capelli sono state simili tra tutti i gruppi, anche se un aumento è stato riportato con una frequenza leggermente maggiore nei gruppi di trattamento (dal 56,1 al 61,8% dei soggetti rispetto al 50,0% del gruppo con il solo veicolo). Gli eventi avversi sono stati simili tra i gruppi.
L’esposizione sistemica della crema all’1% utilizzata per il trattamento dell’acne è minima [4]. I dati relativi alla soluzione topica non sono purtroppo disponibili nella letteratura pubblicata. Gli effetti collaterali sessuali non sono stati monitorati nel loro studio, quindi è difficile ricavare una potenziale esposizione sistemica sulla base di questi risultati.
Come nota finale: è interessante vedere che nel 2016 è stato completato uno studio in cui una soluzione di clascoterone è stata confrontata con una soluzione di minoxidil al 5% o un placebo per il trattamento dell’alopecia androgenetica (NCT02279823). I risultati non sono mai stati pubblicati nella letteratura scientifica. Le ragioni possono essere molteplici, ma forse la più ovvia, dal punto di vista di un’azienda farmaceutica, è: risultati deludenti. Ho l’impressione che non abbia fatto molto bene rispetto al minoxidil.
Un altro antagonista del recettore degli androgeni che sta facendo il giro di internet è RU58841 (noto anche come PSK-3841 o HMR-3841). Nel 2004 era in fase di sperimentazione II, ma da allora lo sviluppo del farmaco è stato interrotto. All’epoca era oggetto di ricerca da parte di Proskelia, l’unità francese del gruppo ProStrakan. Proskelia è stata poi acquisita da Galapagos nel 2006. È importante notare che i risultati degli studi clinici non sono mai stati riportati in letteratura. Si dice che ciò sia dovuto a motivi finanziari. Questo sembra plausibile, Proskelia era un’azienda relativamente piccola (visto che è stata acquisita per 16,5 milioni di dollari nel 2006). I costi degli studi clinici di fase 3 sono molto elevati. Si parla di almeno qualche migliaio di dollari per soggetto (in media costano diverse decine di migliaia di dollari per soggetto). Se si moltiplica questa cifra per i 1000-2000 soggetti necessari per una sperimentazione di questo tipo, diventa subito evidente che molto probabilmente hanno dovuto fare affidamento sugli investitori per realizzarla. Ciononostante, se Galapagos fosse stata interessata a questo composto, avrebbe potuto facilmente finanziare uno studio di fase 3. Va ricordato che la ragione principale per cui i farmaci non entrano nella fase 3 è la mancanza di efficacia o di sicurezza.
Mostra un’elevata affinità per il recettore degli androgeni umani, leggermente inferiore a quella del testosterone (il che è notevole, dato che la maggior parte degli antagonisti ha un’affinità sostanzialmente inferiore) [5]. Sono stati pubblicati alcuni dati di studi su animali. Mostra un’efficacia simile a quella della finasteride nei macachi dalla coda monca [6]. In topi nudi femmina condizionati con testosterone, lo xenotrapianto di tessuto del cuoio capelluto di uomini calvi ha mostrato risultati più favorevoli rispetto ai controlli [7]. Onestamente, questi studi sono preclinici per un motivo: forniscono solo un’indicazione sul fatto che potrebbe essere interessante o meno proseguire con gli studi clinici. Non forniscono altre informazioni, quindi li cito solo per completezza. Senza dati di sperimentazione clinica non si può dire molto su questo composto.
RU58841
Un’ultima osservazione che vorrei fare è che è stato suggerito che RU58841 può influenzare il recettore degli androgeni in modo allosterico [8]. Ciò significa che influisce sulla sua funzione legandosi a un sito diverso da quello di legame con il ligando (dove si legherebbero gli androgeni). Questo ha un’implicazione pratica molto importante. Se c’è un legame competitivo, la sua efficacia dipende dalla concentrazione di altri ligandi (come il DHT). In caso di legame allosterico, ciò non avviene, per cui il suo effetto è indipendente dalle concentrazioni di ligandi, il che sarebbe ideale per i consumatori di steroidi anabolizzanti, in quanto le dosi sovrafisiologiche utilizzate non influirebbero sulla sua efficacia. Purtroppo non sono in grado di accedere allo studio originale che ipotizza questa caratteristica.
Fluridil
Un altro antagonista topico dei recettori degli androgeni è il Fluridil, noto anche come topilutamide e venduto con il nome commerciale di Eucapil. È approvato per uso cosmetico nella Repubblica Ceca. È stato pubblicato uno studio clinico su piccola scala, ma i risultati non sembrano promettenti [9]. 43 soggetti con alopecia androgenetica sono stati randomizzati a ricevere una soluzione topica di Fluridil al 2% o un placebo per 9 mesi. Il conteggio dei capelli in fase anagen o telogen è stato effettuato a 0, 3, 6 e 9 mesi. Mentre nel gruppo del Fluridil si è registrato un aumento maggiore dei peli in fase anagen e una diminuzione maggiore dei peli in fase telogen rispetto al placebo a 3 mesi, non c’è stata alcuna differenza significativa a 9 mesi. (È piuttosto deludente. Studi futuri (che a questo punto non mi aspetto) potrebbero chiarire se si tratta di una peculiarità dello studio o meno. Si potrebbe ipotizzare che sia necessaria una maggiore concentrazione di soluzione di fluridil perché sia efficace.
Terapia con Plasma Ricco di Piastrine (PRP):
Credo che una piccola introduzione sul Plasma Ricco di Piastrine (PRP) sia necessaria. Che cos’è in realtà? In sostanza, è un concentrato di sangue con un’alta presenza di piastrine e la rimozione dei globuli rossi. Viene prodotto mediante un processo chiamato centrifugazione differenziale [9]. Il PRP prodotto contiene una concentrazione di piastrine da 2 a 8 volte superiore a quella del sangue intero. La concentrazione ottenuta dipende dal dispositivo e dal metodo utilizzato. Di solito, per preparare il PRP si prelevano circa 30ml di sangue.
Le piastrine sono importanti per la coagulazione, ma contengono anche una serie di fattori di crescita e citochine [10]. Queste molecole di segnalazione sono il motivo per cui vengono impiegate in diversi campi medici, tra cui la dermatologia, ad esempio nel trattamento dell’alopecia androgenetica. Le piastrine rilasciano questi fattori di crescita e citochine al momento dell’attivazione, che può avvenire dopo l’iniezione nel cuoio capelluto da parte dell’organismo oppure aggiungendo sali di calcio o trombina alle piastrine prima dell’iniezione. Si ritiene che questi fattori di crescita agiscano sulle cellule del follicolo pilifero, esercitando così il loro effetto benefico nel trattamento dell’alopecia (androgenetica).
Poiché non esiste una procedura standardizzata per l’applicazione del PRP, gli studi possono dimostrare risultati diversi a seguito di procedure PRP differenti. Sebbene manchino prove valide, si ritiene che la pre-attivazione delle piastrine prima dell’iniezione e la preparazione del PRP mediante il cosiddetto protocollo a doppia centrifuga portino a risultati migliori.
Una meta-analisi del 2020 ha valutato gli effetti della terapia con PRP come trattamento dell’alopecia androgenetica [11]. Ha incluso 30 studi randomizzati e controllati per l’analisi qualitativa e 5 di questi hanno potuto essere utilizzati per l’analisi quantitativa. La terapia con PRP è risultata efficace nell’aumentare la densità e lo spessore dei capelli. Di fatto, è apparsa più efficace del Minoxidil e della Finasteride. Mentre una meta-analisi del 2017 ha rilevato che la Finasteride e il Minoxidil topico al 5% aumentano la densità dei capelli rispettivamente di 18 e 15 capelli per cm quadrato, la terapia con PRP ha portato a un aumento medio di 33 capelli per cm quadrato.
Gli eventi avversi sono stati riportati nella metà degli studi e si sono limitati a dolore, eritema (rossore) ed edema locale, sanguinamento puntuale, mal di testa transitorio, sonnolenza, ematomi e sensibilità del cuoio capelluto. Non sono stati segnalati eventi avversi gravi. Nel complesso, la terapia con PRP è molto promettente.
Modulatori della via Wnt/β-catenina:
La via Wnt/β-catenina è coinvolta in numerosi processi cellulari. E, come si è scoperto, la via è anche coinvolta nella crescita e nello sviluppo del follicolo pilifero [12, 13]. La via canonica prevede il legame di una proteina Wnt a Frizzled, il suo recettore sulla superficie cellulare, e al suo co-recettore Proteina legata al recettore LDL (LRP) [14]. In assenza di segnalazione Wnt, la β-catenina viene continuamente degradata, mentre con l’attivazione da parte di una proteina Wnt, la β-catenina inizia ad accumularsi nel citosol. La β-catenina trasloca poi nel nucleo dove stimolerà la trascrizione dei geni bersaglio di Wnt.
Via canonica Wnt/β-catenina: Stato “WNT ON”: le proteine WNT, legandosi ai recettori frizzled e al co-recettore LRP, agiscono per sopprimere l’attività della glicogeno sintasi chinasi-3β (GSK-3β). ZNRF3 promuove la degradazione dei recettori WNT che funzionano come soppressori tumorali. Ciò impedisce la fosforilazione delle molecole a valle, consentendo l’associazione della β-catenina con Tcf/Lef nel nucleo e il conseguente aumento della proliferazione cellulare. Stato “WNT OFF”: in assenza del ligando WNT, il complesso di distruzione della β-catenina (contrassegnato dal riquadro tratteggiato), un complesso terziario formato da axina, APC, CK1α e GSK 3β, fosforila la β-catenina, che successivamente va incontro a degradazione proteasomica.
Un attivatore di Wnt attualmente in fase di studio per il trattamento dell’alopecia androgenetica è il SM04554 (noto anche come Dalosirvat). È stato sviluppato da Biosplice Therapeutics (precedentemente nota come Samumed) e sono stati registrati e completati 3 studi clinici: NCT02275351, NCT02503137, NCT03742518. Questi numeri NCT possono essere consultati sul sito www.clinicaltrials.gov per visualizzarne i dettagli. Lo studio registrato con il numero NCT03742518 è uno studio di fase II/III con 675 partecipanti che sono stati randomizzati in tre gruppi. Un gruppo ha utilizzato una soluzione di SM04554 allo 0,15% una volta al giorno, un altro ha utilizzato una soluzione allo 0,25% una volta al giorno e il terzo gruppo ha ricevuto una soluzione veicolo. Lo studio è durato 48 settimane e si è concluso il 31 dicembre 2020. Purtroppo non sono ancora stati pubblicati i risultati dello studio clinico nella letteratura scientifica. Tuttavia, facendo qualche ricerca su Google, è possibile trovare alcune diapositive utilizzate durante una presentazione al Congresso Internazionale di Dermatologia e Cosmetologia (INDERCOS) nel marzo 2019. In esse vengono presentati alcuni risultati di uno studio di fase II, tra cui questa diapositiva:
I partecipanti hanno ricevuto l’intervento per 90 giorni, dopodiché è stato effettuato un follow-up 45 giorni dopo. Ciò che mi colpisce è che la soluzione allo 0,25% ha fatto molto peggio della soluzione allo 0,15% e che la soluzione allo 0,15% ha iniziato a funzionare solo dopo aver terminato la somministrazione. (In ogni caso, i risultati non mi entusiasmano. E sospetto che anche la sperimentazione di fase III non sia andata molto bene. Se si utilizza la Wayback Machine per dare un’occhiata al sito web di Biosplice Therapeutics, si può vedere che SM04554 è ancora elencato nell’agosto 2021. Se si guarda oggi, il farmaco è scomparso e non è più presente nemmeno nella pagina della pipeline. Hanno rinunciato al farmaco?
Quindi, dov’è finito il SM04554?
Nel complesso, i modulatori della via di segnalazione Wnt sono promettenti, ma forse dovremo aspettare ancora un po’ prima di vedere il primo di questa classe di farmaci approvato dalla FDA.
Prostaglandine:
Le Prostaglandine sono emerse come importanti regolatori del ciclo del follicolo pilifero (poiché alcuni farmaci basati su di esse, di cui parlerò più avanti, si sono rivelati in grado di provocare una crescita localizzata dei capelli/ipertricosi). Sono sintetizzate dall’acido grasso arachidonico. In particolare, la Prostaglandina D2 (PGD2) è ritenuta responsabile dell’inibizione della crescita dei capelli nell’alopecia androgenetica [15]. La PGD2 è il prodotto di una reazione catalizzata dall’enzima Prostaglandina D2 Sintasi (PTGDS), il cui substrato è la Prostaglandina H2 (PGH2). La PGH2 è sintetizzata direttamente dall’Acido Arachidonico, una reazione catalizzata da un enzima ciclossigenasi (COX). Quindi, in poche parole:
Acido Arachidonico (COX)-> PGH2 (PTGDS)-> PGD2
Al contrario, la Prostaglandina F2α (PGF2α) e la Prostaglandina E2 (PGE2) stimolano la crescita dei capelli [16]. Sia la PGF2α che la PGE2 derivano anche dalla PGH2. Il primo sintetizzato dalla PGF2α Sintasi e il secondo dalla PGE2 Sintasi.
Bimatoprost
Anche l’industria farmaceutica ha esplorato quest’area di ricerca per il trattamento dell’alopecia androgenetica. Alcuni farmaci che sono stati sviluppati sono una soluzione topica di Bimatoprost (un analogo della PGE2) e una soluzione topica di Latanoprost (un analogo della PGF2α). Entrambi i farmaci sono stati originariamente utilizzati per trattare l’ipertensione oculare o il glaucoma, in quanto abbassano la pressione oculare. Tuttavia, si è scoperto per caso che provocano la crescita dei peli delle ciglia (ipertricosi). Alcuni studi clinici su piccola scala hanno valutato i loro effetti e sembrano promettenti [17, 18].
Setipiprant
Un altro farmaco è il Setipiprant, che agisce come Antagonista Selettivo del Recettore della Prostaglandina D2. Il farmaco è attualmente oggetto di studio da parte di Allergan Aesthetics e nell’ottobre 2021 sono stati pubblicati i risultati di uno studio di fase 2 [19]. I partecipanti hanno ricevuto il Setipiprant orale due volte al giorno (1g x 2 volte), 1mg di Finasteride una volta al giorno o un placebo, per 24 settimane. Sfortunatamente, però, non è stato possibile ottenere risultati migliori rispetto al placebo.
Penso che in futuro sentiremo ancora parlare di Prostaglandine (topiche), o di farmaci che potrebbero inibire la produzione di PGD2 inibendo l’enzima PTGDS (o di farmaci che stimolano la produzione di PGF2α o PGE2 stimolando i rispettivi enzimi che li sintetizzano).
Conclusione:
In questi tre articoli abbiamo imparato a conoscere una condizione (alopecia androgenetica) che interessa a diverso grado circa il 70% degli uomini ed il 40% delle donne. Abbiamo visto quali sono i farmaci approvati per il suo trattamento e quelli sperimentali più promettenti. Ora, sappiamo anche che con l’uso di dosi sovrafisiologiche di AAS riducono fortemente l’impatto apportato dagli inibitori della 5α-reduttasi i quali, comunque, interessano per lo più il Testosterone essendo il substrato principale per le 5α-reduttasi. Discorso diverso potrebbe essere fatto per ciò che concerne l’uso di soluzioni topiche contenenti Antagonisti del Recettore degli Androgeni. Ma, ad oggi, su questo punto non possiamo fare altro che analizzare la letteratura e ipotizzare.
Avrei sicuramente potuto parlare del potenziale effetto del TB-500 sulla sostanziale crescita di nuovi peli nella barba. La letteratura scientifica indica anche che la Timosina beta-4 attiva le cellule staminali nei follicoli (Questo è potenzialmente rilevante, poiché il TB-500 è, approssimativamente, un frammento della Timosina beta-4). È stato osservato che le applicazioni topiche giornaliere di TB-500 accelerano la crescita dei capelli. È stato osservato anche che i capelli risultanti sono più spessi, più scuri e più densi. Nel complesso, i risultati ottenuti su animali [topi e ratti] suggeriscono che, oltre ai suoi noti effetti angiogenici e di guarigione delle ferite, la Timosina β4 ( e potenzialmente il TB500) possa essere un modulatore naturale della crescita dei capelli che agisce stimolando la migrazione delle cellule staminali, la produzione di proteasi e la differenziazione. Attenzione però! Qui non si parla di una “resurrezione” del bulbo miniaturizzato e morto tipico della alopecia adrogenetica. Si parla di follicoli derivanti da sviluppo di cellule staminali, indi “nuovi”. Ma tutto questo rappresenta, ad oggi, poco più di una pura ipotesi nell’uomo.
L’unica soluzione, se la “rasata a 0” non è contemplata, è ovviamente il trapianto. Ma ciò non toglie che esso possa risultare migliorato da una combinazione di fattori iatrogeni, come quelli descritti in questo lavoro.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Rosette, Caridad, et al. “Cortexolone 17α-propionate (clascoterone) is an androgen receptor antagonist in dermal papilla cells in vitro.” Journal of drugs in dermatology: JDD 18.2 (2019): 197-201.
Celasco, Giuseppe, et al. “Biological profile of cortexolone 17a-propionate (CB-03-01), a new topical and peripherally selective androgen antagonist.” Arzneimittelforschung 54.12 (2004): 881-886.
Dhillon, Sohita. “Clascoterone: first approval.” Drugs (2020): 1-6.
Mazzetti, Alessandro, et al. “Pharmacokinetic profile, safety, and tolerability of clascoterone (cortexolone 17-alpha propionate, CB-03-01) topical cream, 1% in subjects with acne vulgaris: an open-label phase 2a study.” Journal of drugs in dermatology: JDD 18.6 (2019): 563-563.
Battmann, T., et al. “RU 58841, a new specific topical antiandrogen: a candidate of choice for the treatment of acne, androgenetic alopecia and hirsutism.” The Journal of Steroid Biochemistry and Molecular Biology 48.1 (1994): 55-60.
Uno, H., et al. “Follicular regrowth with 5 α-reductase inhibitor (finasteride) or androgen receptor blocker (RU58841) in the bald scalp of the stumptailed macaque.” Journal of Investigative Dermatology 4.104 (1995): 658.
De Brouwer, B., et al. “A controlled study of the effects of RU58841, a non‐steroidal antiandrogen, on human hair production by balding scalp grafts maintained on testosterone‐conditioned nude mice.” British Journal of Dermatology 137.5 (1997): 699-702.
Poulos, Georgann A., and Paradi Mirmirani. “Investigational medications in the treatment of alopecia.” Expert opinion on investigational drugs 14.2 (2005): 177-184.
Dhurat, Rachita, and M. S. Sukesh. “Principles and methods of preparation of platelet-rich plasma: a review and author’s perspective.” Journal of cutaneous and aesthetic surgery 7.4 (2014): 189.
Alves, Rubina, and Ramon Grimalt. “A review of platelet-rich plasma: history, biology, mechanism of action, and classification.” Skin appendage disorders 4.1 (2018): 18-24.
Evans, Adam G., et al. “Platelet-rich plasma as a therapy for androgenic alopecia: a systematic review and meta-analysis.” Journal of Dermatological Treatment (2020): 1-14.
Beaudoin, Gerard MJ, et al. “Hairless triggers reactivation of hair growth by promoting Wnt signaling.” Proceedings of the National Academy of Sciences 102.41 (2005): 14653-14658.
Lei, Ming-Xing, Cheng-Ming Chuong, and Randall B. Widelitz. “Tuning Wnt signals for more or fewer hairs.” Journal of Investigative Dermatology 133.1 (2013): 7-9.
Clevers, Hans, and Roel Nusse. “Wnt/β-catenin signaling and disease.” Cell 149.6 (2012): 1192-1205.
Garza, Luis A., et al. “Prostaglandin D2 inhibits hair growth and is elevated in bald scalp of men with androgenetic alopecia.” Science translational medicine 4.126 (2012): 126ra34-126ra34.
Johnstone, Murray A., and Daniel M. Albert. “Prostaglandin-induced hair growth.” Survey of ophthalmology 47 (2002): S185-S202.
Blume-Peytavi, Ulrike, et al. “A randomized double-blind placebo-controlled pilot study to assess the efficacy of a 24-week topical treatment by latanoprost 0.1% on hair growth and pigmentation in healthy volunteers with androgenetic alopecia.” Journal of the American Academy of Dermatology 66.5 (2012): 794-800.
Barrón-Hernández, Yevher Lorena, and Antonella Tosti. “Bimatoprost for the treatment of eyelash, eyebrow and scalp alopecia.” Expert opinion on investigational drugs 26.4 (2017): 515-522.
DuBois, Janet, et al. “Setipiprant for Androgenetic Alopecia in Males: Results from a Randomized, Double-Blind, Placebo-Controlled Phase 2a Trial.” Clinical, Cosmetic and Investigational Dermatology 14 (2021): 1507.
Attualmente gli unici trattamenti approvati per l’ipogonadismo o la carenza di Testosterone sono la Terapia Sostitutiva con Testosterone (TRT) e la terapia con gonadotropina corionica umana (hCG). Tra le due, la TRT è sicuramente quella più comunemente prescritta. Uno dei motivi è rappresentato dal fatto che l’hCG è inefficace nell’ipogonadismo primario, un tipo di ipogonadismo in cui la causa è l’insufficienza testicolare. Questo esclude circa il 15% dei casi di carenza di Testosterone [1]. Altri motivi possono essere il fatto che l’hCG richiede iniezioni frequenti (di solito tre volte alla settimana) ed è più costoso di alcune alternative alla TRT.
Un problema centrale della TRT non affiancata dall’uso regolare di hCG (o abbinamento dell’hCG con l’hMG) è che sopprime la spermatogenesi e quindi porta all’infertilità in un numero considerevole di uomini. Inoltre, le dimensioni dei testicoli diminuiscono. Per gli uomini che desiderano preservare la fertilità e le dimensioni dei testicoli, la TRT in modalità priva di hCG è ovviamente un candidato non ideale. Sebbene questo aspetto sia meno importante per gli uomini più anziani che possono beneficiare della TRT, in quanto è meno probabile che abbiano in programma di avere figli, è un problema importante per gli uomini giovani che desiderano trattare l’ipogonadismo.
Come discusso nel mio precedente articolo sulla fertilità durante l’uso di AAS o in TRT, il Testosterone sopprime la secrezione di LH e FSH, con conseguente inibizione della spermatogenesi. Parte di questa soppressione è mediata dalla conversione del Testosterone in Estradiolo. Si potrebbero quindi aumentare i livelli di Testosterone annullando l’effetto soppressivo dell’Estradiolo sull’ipotalamo e, conseguentemente, sull’ipofisi. In effetti, l’uso di modulatori selettivi dei recettori degli estrogeni (SERM) – che esercitano un’azione antagonista sui recettori degli estrogeni nell’ipotalamo e nell’ipofisi – porta a un forte aumento di LH, FSH e Testosterone negli uomini con ipogonadismo secondario [2]. Allo stesso modo, l’uso di inibitori dell’Aromatasi – che impediscono al Testosterone di essere convertito in Estradiolo dall’azione dell’Enzima Aromatasi – porta a un aumento di LH, FSH e Testosterone negli uomini trattati [3]. Una conseguenza di ciò è che la spermatogenesi può essere preservata sebbene l’uso di AI sia maggiormente deleterio per il profilo lipidico ematico.
Enclomifene nell’Ipogonadismo Secondario:
Per inserire tra le opzioni terapeutiche per il trattamento dell’ipogonadismo secondario l’Enclomifene, un’azienda farmaceutica, la Repros Therapeutics Inc. ha tentato di farlo approvare dalla FDA. Prima di continuare a parlare di come si è svolto il processo, vorrei fornire alcune informazioni sul SERM in questione: l’Enclomifene Citrato (nome commerciale Androxal, successivamente ribattezzato EnCyzix).
Negli anni ’60 è stato scoperto che un farmaco chiamato clomifene citrato induce l’ovulazione. In quanto tale, poteva essere utilizzato come modalità di trattamento per promuovere la fertilità in caso di anovulazione o oligovulazione. Già all’epoca si sapeva che il clomifene agisce aumentando il rilascio di gonadotropine (LH e FSH) [4]. Per questo motivo, i ricercatori hanno iniziato a valutarne l’effetto anche negli uomini sulla spermatogenesi e sul testosterone. Nei decenni successivi, numerosi studi hanno dimostrato la sua efficacia nello stimolare la produzione di testosterone negli uomini ipogonadici. Tuttavia, il clomifene non è stato approvato dalla FDA per il trattamento dell’ipogonadismo. Tuttavia, viene prescritto off-label per questa indicazione e la linea guida 2018 per la valutazione e la gestione della carenza di testosterone dell’American Urological Association ne sostiene condizionatamente l’uso come alternativa alla TRT [5].
Un problema legato al trattamento con Clomifene è che, nonostante il significativo aumento dei livelli di Testosterone, i dati sul suo effetto sulla riduzione dei sintomi dell’ipogonadismo sono contrastanti [6]. Studi su larga scala e di buona qualità potrebbero chiarire questi aspetti e forse fare luce su quali pazienti potrebbero trarre i maggiori benefici dal suo utilizzo. Poiché il brevetto del farmaco è scaduto da tempo e vengono prodotti farmaci generici, le aziende farmaceutiche non sono molto attratte dagli investimenti. Pertanto, questi studi potrebbero non venir mai realizzati.
Come accade per molti altri farmaci, anche il Clomifene è una miscela racemica. Ciò significa che è costituito da una molecola di tipo “levogiro” e una di tipo “destrogiro”. In genere solo uno di questi stereoisomeri, come vengono chiamati, è il composto attivo. E ciò dà come risultato che si adatta meglio al recettore su cui agisce. Come un guanto si adatta solo a una mano e non all’altra, il tipo “levogiro” è più efficace nel legarsi a un recettore “levogiro” rispetto allo stereoisomero “destrogiro”. Il Clomifene è costituito dagli stereoisomeri Zuclomifene (nell’immagine sotto a sinistra) e, come alcuni di voi già sapranno, l’Enclomifene (nell’immagine sotto a destra):
Da sinistra: Zuclomifene e Enclomifene.
In generale, lo Zuclomifene è considerato un agonista del recettore degli estrogeni, mentre l’Enclomifene è considerato un potente antagonista degli estrogeni [7]. L’Enclomifene può quindi essere considerato lo stereoisomero attivo del Clomifene. L’idea dell’Enclomifene privo dello stereoisomero Zuclomifene, quindi, è quella di avere qualcosa di più efficace e sicuro del Clomifene. Tuttavia, l’aspetto più importante è che Repros Therapeutics Inc. potrebbe brevettarne l’uso terapeutico per il trattamento dell’ipogonadismo maschile.
Per richiedere l’approvazione della FDA, l’azienda farmaceutica ha dovuto condurre alcuni studi clinici. Il primo studio pubblicato comprendeva solo 12 uomini e non era in cieco [8]. In altre parole, sia i partecipanti che i ricercatori sapevano quale trattamento stavano ricevendo gli uomini. I partecipanti erano uomini con ipogonadismo secondario trattati in precedenza con Testosterone topico. Sono stati randomizzati a ricevere nuovamente Testosterone topico o Enclomifene (25mg al giorno).
Dopo sei mesi di trattamento, i livelli di Testosterone erano praticamente gli stessi tra i gruppi: 545ng/dL (18,9nmol/L) nel gruppo che riceveva il gel e 525ng/dL (18,2nmol/L) nel gruppo che riceveva l’Enclomifene. Anche i livelli di Testosterone libero sono aumentati e sono rimasti praticamente invariati tra i gruppi. Inoltre, e naturalmente, il numero di spermatozoi è stato ridotto negli uomini che ricevevano Testosterone, con numeri intorno ai 20milioni/mL. Inoltre, come previsto, il numero di spermatozoi è aumentato negli uomini che hanno ricevuto l’Enclomifene, con una media di circa 150milioni/mL.
Sono stati condotti un paio di studi clinici successivi. Forse il più interessante è stato quello pubblicato nel 2016, rivolto a uomini ipogonadici obesi [9]. Il documento comprende due studi paralleli randomizzati, in doppio cieco, a doppio braccio e controllati con placebo. Si tratta di un’affermazione che lascia a bocca aperta e credo che il termine “doppio cieco” richieda qualche spiegazione. Nel precedente studio di cui mi sono occupato, ho detto che era di natura non cieca. Quindi i partecipanti e i ricercatori sapevano quale trattamento stava ricevendo ciascun soggetto. Di solito, quando si confrontano due farmaci diversi, si possono semplicemente mettere in cieco i soggetti (e i ricercatori) dando ai gruppi capsule, o pastiglie, o altro identici. Tuttavia, il gel di Testosterone è un gel, mentre l’Enclomifene è una compressa da inghiottire. Quindi non è possibile farlo. Per poter effettuare uno studio come questo in cieco, è necessario somministrare a entrambi i gruppi sia le pastiglie che il gel. Quindi un gruppo riceve un gel placebo e l’Enclomifene, mentre l’altro gruppo riceve un gel di Testosterone e una compressa placebo. Ovvero, doppio braccio. (E poiché lo studio era controllato con placebo, un gruppo ha ricevuto un gel e una compressa placebo).
I due studi descritti in questo articolo hanno utilizzato lo stesso protocollo e l’aspetto forse più interessante è stata la dimensione del campione: 256 soggetti in totale! Finalmente si è capito qualcosa. L’intervento è durato 16 settimane e i soggetti del gruppo Enclomifene hanno ricevuto 12,5mg al giorno e sono stati trattati fino a 25mg al giorno se i livelli di Testosterone non erano aumentati ad almeno 450ng/dL (15,6nmol/L) alla quarta settimana. La dose è stata aumentata per la metà dei soggetti che ricevevano l’Enclomifene. A questo punto le cose iniziano a farsi interessanti: sebbene metà dei soggetti sia stata modificata nel dosaggio alla quarta settimana, non è successo assolutamente nulla con la concentrazione media di Testosterone:
E, in effetti, alla fine dell’intervento, la media del gruppo era appena al di sotto del valore limite di 450ng/dL (15,6nmol/L) per l’up-titration. Infine, 29 degli 85 uomini del gruppo Enclomifene non hanno visto il loro Testosterone aumentare al di sopra del valore limite di ipogonadismo di 300ng/dL (10,4nmol/L) dopo 16 settimane di trattamento. Inoltre, i ricercatori hanno fatto un LAVORO ORRIBILE nel trattare correttamente il gruppo che utilizzava il gel di Testosterone, come si può vedere dalla concentrazione media di Testosterone di quel gruppo. Quasi come se l’avessero fatto apposta per far sì che il gruppo Enclomifene facesse meglio in alcune misurazioni… (anche se si tratta di uno studio a doppio braccio, è comunque possibile istruire i pazienti in modo scorretto con l’applicazione del gel).
È importante notare che gli unici endpoint erano i livelli di Testosterone, LH e FSH e la concentrazione di sperma. Non sono stati analizzati endpoint clinicamente rilevanti, come il desiderio sessuale, la funzione erettile, la stanchezza/vitalità, ecc. A quanto pare, nemmeno negli altri studi (pubblicati). O, forse, sono stati analizzati, ma semplicemente non sono stati riportati nei risultati dello studio perché erano deludenti. E penso che potrebbe essere stata la seconda ipotesi, visto che la FDA non ha approvato il farmaco per il trattamento dell’ipogonadismo secondario, a causa della mancanza di un miglioramento sintomatico misurabile [10]. Anche l’equivalente della FDA nell’UE, l’EMA, ha rifiutato l’autorizzazione all’immissione in commercio dell’Enclomifene qualche tempo dopo, con preoccupazioni simili:
“Il CHMP [Comitato per i Medicinali per Uso Umano] ha osservato che, sebbene gli studi abbiano mostrato un aumento dei livelli di Testosterone con EnCyzix [Enclomifene], non hanno esaminato se EnCyzix migliorasse sintomi quali la densità e resistenza ossea, l’aumento di peso, l’impotenza e la libido. Inoltre, il farmaco comporta un rischio di tromboembolismo venoso (problemi dovuti alla formazione di coaguli di sangue nelle vene)”.
E il Clomifene mostra in realtà risultati molto simili, anche mg per mg, a quelli dell’Enclomifene. Non riassumerò qui l’intera letteratura sul Clomifene, ma prendiamo ad esempio uno studio di Katz et al. in cui 86 giovani uomini ipogonadici hanno ricevuto il Clomifene Citrato a 25mg o 50mg a giorni alterni per una media di 19 mesi e hanno visto aumentare il Testosterone totale del 152% (da 192 ng/dL a 485 ng/dL) [11]. In particolare, il Testosterone libero è aumentato di ben il 332%. Se consideriamo un altro studio condotto su uomini obesi, il Testosterone è aumentato del 98% (da 303ng/dL a 599ng/dL) con 25mg al giorno [12]. In termini di aumento del Testosterone, l’Enclomifene non sembra avere un vantaggio rispetto al Clomifene (non sono riuscito a trovare uno studio di confronto testa a testa).
Conclusioni:
Quindi, per concludere, purtroppo non esiste ancora un’alternativa approvata dalla FDA oltre all’hCG o alla TRT per il trattamento dell’ipogonadismo. E con ciò, gli uomini ipogonadici che cercano un trattamento saranno vincolati alle iniezioni di hCG ( e spesso anche hMG) se desiderano preservare la fertilità durante la TRT. Forse i SERM (attuali) sono solo un vicolo cieco, poiché il loro antagonismo con gli estrogeni contrasta anche gli effetti positivi. Infatti, come hanno dimostrato elegantemente Finkelstein et al., l’aggiunta di un inibitore dell’Aromatasi a un gel di Testosterone ha un impatto negativo sul grasso corporeo e sulla funzione sessuale [13]. Avrebbero dovuto inserire anche l’aumento della neurotossicità e cardiotossicità da carenza di Estradiolo, oltre a stati depressivi e condizioni annesse.
Gabriel Bellizzi
Riferimenti:
Tajar, Abdelouahid, et al. “Characteristics of secondary, primary, and compensated hypogonadism in aging men: evidence from the European Male Ageing Study.” The Journal of Clinical Endocrinology & Metabolism 95.4 (2010): 1810-1818.
Wheeler, Karen M., et al. “Clomiphene citrate for the treatment of hypogonadism.” Sexual medicine reviews 7.2 (2019): 272-276.
De Ronde, Willem, and Frank H. de Jong. “Aromatase inhibitors in men: effects and therapeutic options.” Reproductive Biology and Endocrinology 9.1 (2011): 1-7.
Jungck, Edwin C., et al. “Effect of clomiphene citrate on spermatogenesis in the human: a preliminary report.” Obstetrical & Gynecological Survey 19.3 (1964): 520.
Mulhall, John P., et al. “Evaluation and management of testosterone deficiency: AUA guideline.” The Journal of urology 200.2 (2018): 423-432.
Scovell, Jason M., and Mohit Khera. “Testosterone replacement therapy versus clomiphene citrate in the young hypogonadal male.” European urology focus 4.3 (2018): 321-323.
Fontenot, Gregory K., Ronald D. Wiehle, and Joseph S. Podolski. “Differential effects of isomers of clomiphene citrate on reproductive tissues in male mice.” BJU Int 117.2 (2016): 344-50.
Kaminetsky, Jed, et al. “Oral enclomiphene citrate stimulates the endogenous production of testosterone and sperm counts in men with low testosterone: comparison with testosterone gel.” The journal of sexual medicine 10.6 (2013): 1628-1635.
Kim, Edward D., Andrew McCullough, and Jed Kaminetsky. “Oral enclomiphene citrate raises testosterone and preserves sperm counts in obese hypogonadal men, unlike topical testosterone: restoration instead of replacement.” BJU international 117.4 (2016): 677-685.
Earl, Joshua A., and Edward D. Kim. “Enclomiphene citrate: A treatment that maintains fertility in men with secondary hypogonadism.” Expert review of endocrinology & metabolism 14.3 (2019): 157-165.
Katz, Darren J., et al. “Outcomes of clomiphene citrate treatment in young hypogonadal men.” BJU International-British Journal of Urology 110.4 (2012): 573.
Pelusi, Carla, et al. “Clomiphene citrate effect in obese men with low serum testosterone treated with metformin due to dysmetabolic disorders: a randomized, double-blind, placebo-controlled study.” PLoS One 12.9 (2017): e0183369.
Finkelstein, Joel S., et al. “Gonadal steroids and body composition, strength, and sexual function in men.” New England Journal of Medicine 369.11 (2013): 1011-1022.
Il BodyBuilding si differenzia dagli sport di prestazione perché il giorno della gara gli atleti vengono giudicati in base all’aspetto piuttosto che alle capacità atletiche. I bodybuilder posano sul palco dove vengono giudicati per la muscolatura, la definizione e la simmetria. Nel corso di una stagione, i bodybuilder attraversano tre fasi diverse: la fase di crescita muscolare (Off-Season), la dieta per la competizione (preparazione alla gara) e la gara stessa. La maggior parte della letteratura riguarda la fase di dieta pre-gara e la peak week.[1]
Tuttavia, la letteratura scientifica sulle raccomandazioni alimentari per i bodybuilder durante la Off-Season è carente. Si tratta di una lacuna importante, poiché la maggior parte della carriera di un bodybuilder si svolge in questa fase, in cui l’obiettivo è aumentare la massa muscolare riducendo al minimo l’aumento eccessivo della massa grassa. I bodybuilder sono noti per avere atteggiamenti rigidi nei confronti della selezione degli alimenti, della frequenza dei pasti, dei tempi di alimentazione e dell’integrazione [2]. Storicamente, le informazioni sull’alimentazione e l’integrazione sono state trasmesse dalle riviste di bodybuilding e dai concorrenti di successo, ma recentemente sono emerse più informazioni attraverso Internet e i forum [3,4]. Di conseguenza, molte delle strategie alimentari utilizzate dai bodybuilder non hanno un solido supporto scientifico e la letteratura scientifica dimostra che alcune di queste strategie, tra cui l’uso massiccio di farmaci, ma anche di integratori più in generale, possono essere ovviamente dannosi per la salute [5,6,7].
Poiché i bodybuilder trascorrono la maggior parte del loro tempo in Off-Season, è evidente la necessità di raccomandazioni nutrizionali e di supplementazione, sia OTC che PEDs, il più possibile “sicure” e basate sull’evidenza per questa popolazione. È stato inoltre dimostrato che alcuni bodybuilder, e non soltanto i concorrenti di alto livello nel bodybuilding “Natural”, potrebbero essere interessati a informazioni basate sull’evidenza [8]. Con il supporto della review realizzata e pubblicata da Juma Iraki et al. che tratta del Off-Season a livello alimentare e integrativo, lo scopo di questo articolo sarà quello di riportare quanto evidenziato dalla letteratura scientifica sugli argomenti relativi all’alimentazione e all’integrazione alimentare e supplementazione PEDs rilevanti per i bodybuilder nella Off-Season e di fornire raccomandazioni pratiche sull’assunzione di energia, macronutrienti, frequenza dei pasti, tempistica dei nutrienti, integratori alimentari e PEDs .
Transizione dalla dieta pre-gara/peak week alla dieta in Off-Season – Reverse Diet Vs. Recovery Diet:
Il primo step che il bodybuilder si trova davanti è la gestione del passaggio da una dieta ipocalorica ad una ipercalorica. Ed è in questo frangente che emergono due strategie simili all’apparenza ma in realtà diverse: la “Recovery Diet” e la “Reverse Diet”.
Ora, molto semplicemente, la “Recovery Diet” consiste in un graduale aumento calorico ma di consistenza tale che l’atleta esca dalla condizione di ipocalorica nel giro di due settimane circa. Con la “Reverse Diet”, invece, abbiamo sempre un graduale aumento calorico ma caratterizzato da una ridotta consistenza dello stesso (si parla di circa 100Kcal/die a settimana). In questo caso specifico, il bodybuilder rimarrebbe in ipocalorica per diverse settimane con possibile emersione di problemi psicofisici legati al protrarsi dello stato stressorio.
Quindi, con il termine “Recovery Diet” ci riferiamo ad uno schema alimentare avente l’obiettivo generale di RECUPERARE da un periodo di dieta cronica sperimentato durante la preparazione alla gara. La “Recovery Diet” incoraggia i bodybuilder a guadagnare il 5-10% del loro peso di gara nelle prime 4-8 settimane successive all’evento. Questo con l’intento di accelerare l’aumento di grasso corporeo e far rientrare il soggetto in un range di grasso corporeo “sano”, fisiologico, il prima possibile. In seguito, si consiglia agli atleti di rallentare il ritmo di aumento del peso e di mantenere un surplus controllato, con un aumento medio dello 0,5-1% del peso corporeo al mese passando pienamente nella Off-Season. Questo fino a quando non raggiungono un punto in cui un ulteriore aumento di peso è considerato improduttivo. Con il termine “Reverse Diet” ci si riferisce ad una strategia la quale può ancora essere attuata con discreti vantaggi per aiutare un agonista a recuperare dopo il contest. Tuttavia, se rispettata e seguita correttamente, piccoli aumenti di cibo di ~100 Kcal/die a settimana potrebbero comunque protrarre il deficit calorico del soggetto, prolungando così il periodo di dieta ipocalorica. Sebbene questa possa essere una strategia utile in alcune circostanze, ad esempio durante l’avvicinamento alla competizione, le modalità di applicazione non permettono un recupero di una bf salubre in tempi ottimali. È risaputo che un bodybuilder in condizioni di picco non è necessariamente al massimo della salute, e questo è in gran parte correlato al livello di grasso corporeo. Accettare un certo aumento di grasso avrà effetti positivi su tutti gli aspetti della Off-Season come le prestazioni in allenamento, i marcatori ormonali, la disponibilità di energia, la qualità del sonno e, inoltre, sarà vantaggioso sulla longevità complessiva dello sport praticato.
In definitiva, se si parte da body fat estremamente basse, tipiche da gara, allora la “Recovery Diet” è la scelta migliore per shiftare dal regime ipocalorico che ha caratterizzato il periodo di preparazione alla gara a quello ipercalorico del Off-Season. Discorso diverso se ci troviamo di fronte ad un soggetto amatoriale, con una body fat del 8-10% arrivato al termine del percorso di “Cut”. In questo caso la “Reverse Diet” è la scelta più funzionale permettendo un controllo migliore degli incrementi calorici evitando che la massa grassa sfori eccessivamente e che il lavoro precedentemente svolto in “Cut” venga facilmente e totalmente compromesso. Anche “ibridazioni” con aumenti settimanali di 45-50g di CHO die possono essere applicati con buoni risultati.
Energia:
Durante la Off-Season, l’obiettivo principale di un bodybuilder è quello di aumentare la massa muscolare riducendo al minimo l’aumento della massa grassa attraverso l’uso di allenamenti contro-resistenza e il mantenimento di un bilancio energetico positivo. Per valutare con precisione il fabbisogno energetico dei bodybuilder durante la bassa stagione, è necessario considerare il volume, la frequenza e l’intensità dell’allenamento. Durante la fase off-season, è stato riportato che i bodybuilder si allenano alla resistenza 5-6 volte a settimana, esercitando ogni gruppo muscolare 1-2 volte a settimana [9]. È stato inoltre riferito che seguono una routine di allenamento ad alto volume con 4-5 esercizi per gruppo muscolare, eseguendo 3-6 serie per esercizio, 7-12 ripetizioni massime (RM) per ogni serie con 1-2 minuti di riposo tra le serie. La durata della sessione di allenamento è stata indicata in ~40-90 minuti. Tuttavia, i piani di allenamento possono variare notevolmente da atleta ad atleta. È necessario valutare anche l’apporto calorico medio dei bodybuilder. Nella fase off-season, l’apporto energetico è di solito sostanzialmente più elevato rispetto alla fase di dieta: tra i bodybuilder maschi è stato riportato un apporto medio di ~3800 kcal/giorno durante la fase off-season e di ~2400 kcal/giorno durante la fase di dieta [2].
Bilancio energetico positivo:
È stato dimostrato che un bilancio energetico positivo ha un importante effetto anabolico, anche in assenza di allenamento contro-resistenza [10]. Tuttavia, la combinazione di un bilancio energetico positivo con l’allenamento contro-resistenza rappresenta il metodo più efficace per garantire che gli effetti anabolici siano diretti all’aumento della massa muscolo-scheletrica [11,12]. L’entità del surplus energetico ideale per guadagnare massa muscolare limitando l’accumulo di tessuto adiposo può variare in base allo stato di allenamento. Nei soggetti non allenati, è stato dimostrato che un surplus energetico sostanziale di circa 2.000 kcal, combinato con l’allenamento contro-resistenza, fornisce un robusto aumento di peso, in cui il contributo della massa magra (LBM) può raggiungere il 100% [12]. Tuttavia, nei soggetti allenati, un surplus energetico sostanziale potrebbe non essere necessario o vantaggioso. Uno studio condotto su atleti d’élite ha esaminato l’effetto delle indicazioni dietetiche sui cambiamenti della composizione corporea tra gli atleti d’élite quando l’allenamento contro-resistenza è stato combinato con diverse entità di surplus energetico. Un gruppo con un peso corporeo medio di 75kg ha consumato energia ad libitum (2964 kcal) per raggiungere un surplus molto ridotto, mentre un secondo gruppo con un peso corporeo medio di 71kg ha ricevuto una consulenza dietetica e ha consumato ~600 kcal in più rispetto al gruppo ad libitum [13].
Entrambi i gruppi hanno seguito lo stesso programma di allenamento contro-resistenza di 4 giorni alla settimana per un periodo di 8-12 settimane. I ricercatori hanno ipotizzato che il gruppo ipercalorico avrebbe avuto un aumento maggiore del peso corporeo e della LBM. Sebbene il gruppo ipercalorico abbia ottenuto un aumento maggiore della LBM rispetto a quelli che mangiavano ad libitum, questo non ha raggiunto la significatività statistica (1,7kg contro 1,2kg, rispettivamente). Inoltre, rispetto al gruppo che mangiava a sazietà, hanno registrato un aumento significativamente maggiore della massa grassa (1,1kg contro 0,2kg, rispettivamente). I ricercatori hanno concluso che un surplus di 200-300 kcal al giorno negli atleti altamente allenati potrebbe essere più appropriato di 500 kcal per minimizzare il rischio di inutili aumenti di grasso corporeo. I soggetti non allenati, più lontani dal loro tetto genetico di massa muscolare, possono essere in grado di aumentare i muscoli a un ritmo più veloce rispetto agli individui allenati.
Il tasso di crescita muscolare può rallentare con l’avanzare dell’età [14]. Pertanto, un maggiore surplus energetico può essere più vantaggioso per i bodybuilder alle prime armi, mentre i bodybuilder avanzati potrebbero trarre maggiore beneficio da diete ipercaloriche conservative per limitare inutili aumenti di grasso corporeo. Studi precedenti hanno raccomandato ai bodybuilder di consumare una dieta leggermente ipercalorica, con un aumento dell’apporto energetico di circa il 15% rispetto al mantenimento nella Off-Season [15]. Tuttavia, ciò non tiene conto della storia di allenamento e del livello di esperienza del singolo bodybuilder. Poiché la capacità di aumentare la massa muscolare è limitata, un surplus aggressivo può portare a un inutile aumento del grasso corporeo, che aumenterebbe la durata o la gravità dei successivi periodi di preparazione alle gare, aumentando di conseguenza la durata o la gravità della scarsa disponibilità energetica. Pertanto, il numero di calorie che un bodybuilder consuma al di sopra del livello di mantenimento può essere stabilito in base al livello di esperienza e poi regolato in base al tasso di aumento di peso e ai cambiamenti nella composizione corporea. Dato che i bodybuilder spesso aumentano rapidamente di peso dopo una gara, potrebbe essere utile avere un obiettivo di aumento di peso per settimana e regolarsi di conseguenza [16,17].
Tuttavia, come detto precedentemente, inizialmente, dopo la gara, potrebbe essere utile un aumento di peso più rapido per aiutare a riportare il concorrente a uno stato di salute sia psicologico che fisiologico, prima che il tasso di aumento di peso venga rallentato per limitare l’accumulo eccessivo di tessuto adiposo. Nella letteratura scientifica si raccomanda di puntare a un aumento di peso di circa 0,25-0,5 kg a settimana per cercare di aumentare la LBM e ridurre al minimo l’aumento della massa grassa [14,18]. Per un bodybuilder avanzato, un potenziale aumento di 2kg di peso corporeo su base mensile potrebbe essere eccessivo e comportare un’inutile accumulazione di grasso corporeo; pertanto, questo tasso dovrebbe essere considerato con cautela. Sulla base delle prove attuali, potrebbe essere opportuno raccomandare ai bodybuilder di consumare una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) nella Off-Season e raccomandare ai bodybuilder avanzati di puntare all’estremità inferiore di questa raccomandazione, o addirittura di essere più conservativi se si verificano aumenti sostanziali della massa grassa. Dato che i bodybuilder consumano in media 45 kcal/kg durante la bassa stagione, il surplus raccomandato equivale a circa 42-48 kcal/kg [2]. Potrebbe essere utile puntare a un aumento di peso di circa 0,25-0,5% del peso corporeo a settimana, regolando al contempo l’apporto energetico in base alle variazioni della composizione corporea. Inoltre, potrebbe essere più appropriato considerare le variazioni di peso medie settimanali basate su pesate giornaliere (o più volte alla settimana) per limitare gli errori delle fluttuazioni giornaliere del peso che possono verificarsi durante la settimana. Una volta determinato il surplus calorico, il passo successivo sarà quello di distribuire le calorie tra proteine, grassi e carboidrati.
Proteine:
Il turnover proteico del muscolo scheletrico è il rapporto tra la sintesi proteica muscolare (MPS) e la degradazione proteica muscolare (MPB). L’ipertrofia del muscolo scheletrico richiede un equilibrio netto in cui la MPS supera la MPB. L’esercizio contro-resistenza fornisce lo stimolo di tensione iniziale che induce l’ipertrofia risultante dall’aumento cumulativo della MPS dopo l’esercizio cronico [19]; tuttavia, l’aumento della massa grassa (FFM) può essere limitato se l’apporto proteico giornaliero è insufficiente [20]. Oltre alla quantità totale consumata al giorno, i ricercatori hanno ipotizzato che la qualità delle proteine possa aumentare il guadagno muscolare indotto dall’allenamento contro-resistenza [21]. Pertanto, entrambi questi argomenti saranno discussi nelle sezioni seguenti.
Introito proteico giornaliero:
Mentre l’attuale RDA per le proteine negli individui sani sedentari è di 0,8 g/kg, in una meta-analisi del 2018 di Morton e colleghi [22] è stato osservato che il doppio di questa quantità massimizza l’ipertrofia indotta dall’allenamento contro-resistenza. Inoltre, gli autori hanno osservato che “potrebbe essere prudente raccomandare ~2,2g di proteine/kg/die per coloro che cercano di massimizzare i guadagni di FFM indotti dall’allenamento contro-resistenza”, poiché 2,2g/kg era l’estremità superiore del limite di confidenza [22] e le differenze individuali impongono che alcuni atleti abbiano un fabbisogno proteico più elevato di altri [23]. Inoltre, la raccomandazione “meglio prevenire che curare” è probabilmente sicura, vista l’assenza di danni apparenti in studi di 1-2 anni tra i sollevatori che consumavano apporti proteici di almeno 2,2 g/kg [24,25]. Infine, la media e il limite superiore di confidenza del 95% per il fabbisogno proteico utilizzando la tecnica di ossidazione degli aminoacidi con indicatore tra i bodybuilder maschi nei giorni di non allenamento sono stati riportati rispettivamente come 1,7 e 2,2g/kg [26], che è simile al fabbisogno tra le donne quando è normalizzato alla FFM [27].
Tuttavia, è stato riportato che i bodybuilder consumano fino a 4,3g/kg di proteine al giorno tra i soggetti di sesso maschile e 2,8g/kg tra quelli di sesso femminile, superando di gran lunga queste raccomandazioni [2]. Le linee guida precedentemente fornite per i bodybuilder nella Off-Season erano di consumare il 25-30% del loro apporto energetico dalle proteine [15]. Potrebbe essere ragionevole opporsi all’indicazione di raccomandazioni basate su percentuali dell’apporto energetico totale, poiché un individuo con un peso non particolarmente elevato ma con un alto fabbisogno energetico potrebbe finire per consumare proteine che superano di gran lunga quelle necessarie e quindi richieste. Inoltre, questo può portare a un’assunzione insufficiente di carboidrati e grassi se l’atleta mira a un apporto calorico specifico. Pertanto, potrebbe essere più appropriato raccomandare un fabbisogno proteico basato sul peso corporeo. Pertanto, i bodybuilder dovrebbero consumare un minimo di 1,6g/kg di proteine nella Off-Season, anche se un obiettivo più vicino a 2,2 g/kg potrebbe garantire una risposta ottimizzata in modo più coerente in una maggiore percentuale di atleti.
E per i “Doped”? Dovremo ormai sapere che la fisiologia di base è la medesima per ogni individuo con le consuete variabili. Detto ciò, l’uso di PEDs va si ad alterare la fisiologia ma in questo specifico ambito, ossia introito proteico per massimizzare lo stimolo ipertrofico, hanno una azione di perfezionamento dell'”economia proteica cellulare”: in parole più semplici, sembra che l’uso di AAS porti ad una migliore resa nell’utilizzo degli amminoacidi scissi e assorbiti dalle proteine alimentari. Di conseguenza, a parità di apporto proteico, la veicolazione degli amminoacidi a scopo plastico è maggiore come minore è l’attività catabolica. Ciò significa che abusare delle proteine, in special modo durante una fase ipercalorica, perchè si è sotto AAS potrebbe risultare più inutile di quanto non lo sia in contesto “Natural”.
Infine, ed è necessario sottolinearlo, tra i bodybuilder che lottano con la fame in Off-Season e che di conseguenza assumono quantità caloriche che portano a un aumento di peso più rapido e all’accumulo di grasso in eccesso, un apporto proteico più elevato può essere utile (se non controindicato per motivi clinici). In uno studio condotto da Antonio e colleghi, i partecipanti ad allenamenti contro-resistenza che consumavano più proteine (4,4g/kg al giorno) e più calorie hanno guadagnato una quantità simile di FFM, ma non hanno guadagnato ulteriore grasso corporeo rispetto al gruppo che consumava meno proteine e meno calorie [28]. Allo stesso modo, in uno studio di follow-up, un gruppo che consumava 3,4g/kg di proteine al giorno ha guadagnato una quantità simile di FFM, ma ha perso una percentuale maggiore di grasso corporeo rispetto a un gruppo a basso contenuto proteico, ancora una volta, nonostante un apporto energetico più elevato [29]. Gli autori di questi studi sulla “vita libera” hanno ipotizzato che i loro risultati fossero dovuti a un aumento della termogenesi indotta dalla dieta attraverso protocolli alimentari ad alto contenuto proteico. Tuttavia, ciò è in contrasto con uno studio di Bray e colleghi del 2012 sul reparto metabolico, più strettamente controllato, in cui il contenuto proteico della dieta influenzava la percentuale di massa corporea acquisita, mentre la massa corporea totale era dettata dal solo contenuto energetico della dieta [30].
Pertanto, mentre la termogenesi indotta dalla dieta potrebbe essere significativamente più elevata con assunzioni di proteine nell’intervallo di 3 g/kg o superiore, la perdita di grasso o la mancanza di aumento di peso osservata da Antonio e colleghi, nonostante un apporto energetico più elevato, potrebbe con più probabilità riflettere l’effetto saziante di assunzioni proteiche molto elevate che diminuiscono l’assunzione calorica effettiva, piuttosto che un aumento della sola termogenesi.
Qualità delle Proteine:
Gli aminoacidi essenziali (EAA) sono gli unici aminoacidi necessari per stimolare il processo di MPS [31]. Sebbene tutti gli aminoacidi forniscano i “mattoni” necessari per la sintesi di nuovi tessuti, l’aminoacido Leucina in particolare sembra essere particolarmente importante come “innesco metabolico” della MPS [32]. È stato suggerito che una concentrazione sufficiente di Leucina è necessaria per raggiungere una “soglia di Leucina” che è richiesta per stimolare al massimo la MPS [33]. In breve, dal punto di vista della costruzione muscolare, le fonti proteiche che innescano una consistente risposta della MPS (quantità sufficiente di Leucina) e forniscono i mattoni essenziali per la costruzione di nuovo tessuto muscolare (contengono l’intero spettro di aminoacidi essenziali in abbondanza) possono essere considerate di “qualità superiore”.
Sebbene l’effetto meccanicistico della Leucina sulle MPS esuli dallo scopo di questo articolo, si invitano i lettori a leggere una rassegna che tratta questo argomento in dettaglio [34]. In generale, su una base di grammo per grammo, le fonti proteiche di origine animale contengono in genere più Leucina ed EAA, anche se ci sono eccezioni degne di nota. Le proteine della soia, uno dei più comuni integratori proteici di origine vegetale, contengono tutti gli EAA, ma in una quantità inferiore per grammo rispetto alle proteine del latte e quindi, in uno studio, hanno prodotto un aumento minore delle MPS rispetto al siero di latte dopo un’ingestione acuta [35]. È interessante notare che in questo stesso studio la soia ha prodotto un aumento maggiore delle MPS rispetto alla caseina, anch’essa una proteina casearia di “alta qualità”, presumibilmente a causa della più lenta velocità di digestione della caseina [35]. Rammentate sempre la differenza tra risposta “acuta” e “cronica”. Per l’appunto, ciò significa che, sebbene il contenuto di Leucina e di EAA di una fonte proteica debba essere preso in considerazione, la risposta acuta alla MPS non è l’unica variabile legata all’ipertrofia a lungo termine. Infatti, una proteina di alta qualità ma “lenta” come la caseina produce inizialmente una risposta MPS di minore ampiezza. Tuttavia, la caseina (e altre proteine a lenta digestione) può produrre un’area MPS sotto la curva simile o maggiore se osservata longitudinalmente rispetto a una fonte proteica “veloce” come il siero di latte, che determina un aumento iniziale maggiore e poi una brusca riduzione [36].
Inoltre, la risposta acuta della MPS a un determinato tipo di proteina non deve essere vista in una prospettiva riduzionista. Nel mondo reale si consumano quotidianamente più porzioni di varie fonti proteiche, rendendo probabilmente superflue alcune di queste distinzioni nel profilo aminoacidico e nella cinetica di digestione. Infatti, in una meta-analisi che ha confrontato i cambiamenti longitudinali della composizione corporea con diversi tipi di integratori proteici, non sono state riscontrate differenze significative tra i partecipanti che consumavano soia rispetto al siero di latte, ad altre proteine del latte o alle proteine isolate del manzo [37].
Come dimostrato in uno studio che ha messo a confronto gruppi che consumavano proteine dopo l’allenamento (in aggiunta a una dieta già composta dal 25% di proteine), sia che venissero forniti 48g di proteine del siero del latte (contenenti 5,5g di Leucina), sia che venissero forniti 48g di proteine del riso (contenenti 3,8g di Leucina), non è stato osservato alcun impatto sui cambiamenti della composizione corporea tra i gruppi dopo otto settimane [38]. Pertanto, se consumate in quantità sufficienti (soprattutto se si considera l’apporto proteico totale giornaliero), la qualità delle proteine di un singolo pasto è meno preoccupante. Tuttavia, se si volesse consumare una dieta dominata da fonti proteiche di origine vegetale, esistono alternative alla soia e al riso. Ad esempio, le proteine isolate del pisello sono ricche di EAA e di Leucina. In uno studio di 12 settimane, un gruppo che consumava 50g di proteine isolate di pisello al giorno ha registrato un aumento maggiore dello spessore muscolare indotto dall’allenamento di resistenza rispetto al placebo, non significativamente diverso da un gruppo che consumava 50g di siero di latte [39].
Pertanto, nel contesto delle indicazioni di questo articolo, la qualità delle proteine può essere un problema solo se si utilizza la fascia bassa delle linee guida sulle proteine (1,6g/kg) o se si consuma una dieta a base prevalentemente vegetale. In entrambi i casi, potrebbe essere utile integrare con fonti proteiche ricche di Leucina e di EAA, a seconda delle preferenze alimentari (ad esempio, proteine del latte o del pisello se si è vegani), per garantire la risposta attesa della MPS all’assunzione di proteine.
Grassi:
Il grasso è un nutriente fondamentale per molte funzioni dell’organismo. Tuttavia, non si sa molto dell’effetto dei grassi alimentari sull’ipertrofia del muscolo scheletrico. È stato riportato che l’assunzione di grassi alimentari tra i bodybuilder varia dall’8 al 33% delle calorie totali [2]. Sebbene i trigliceridi intramuscolari possano fungere da substrato energetico durante l’allenamento di resistenza, non sono un fattore limitante poiché i substrati derivano principalmente da processi anaerobici [40]. Di interesse per il bodybuilder, è dimostrato che negli atleti allenati contro-resistenza [41] e nei giocatori di hockey [42] le diete a basso contenuto di carboidrati (30-45% dell’energia o meno) possono influire sul rapporto Testosterone libero/Cortisolo (fTC), il che potrebbe avere un impatto negativo sul recupero. D’altra parte, la riduzione dei grassi alimentari nelle diete isocaloriche da ~30-40% a ~15-25% ha portato a riduzioni significative ma modeste dei livelli di Testosterone [43,44,45,46].
Tuttavia, non è chiaro se le variazioni di Testosterone all’interno di intervalli normali influenzino in modo significativo l’aumento della massa muscolare [47]. Nonostante la possibilità che i livelli di testosterone possano essere più elevati quando si consuma una percentuale maggiore di energia proveniente dai grassi alimentari, i cambiamenti effettivi nella massa muscolare durante gli studi longitudinali di individui allenati alla resistenza che seguono diete “chetogeniche” ad alto contenuto di grassi sono stati costantemente inferiori rispetto ad approcci moderati o a basso contenuto di grassi con ampi carboidrati [48,49,50,51]. Non è ancora stato chiarito se ciò sia dovuto a cambiamenti nella capacità di esercizio, ad alterazioni del rapporto fTC o a qualche altro meccanismo legato alla componente ad alto contenuto di grassi o a basso contenuto di carboidrati della dieta.
Tuttavia, ciò indica che forse si dovrebbe consumare una proporzione più moderata di grassi nella dieta, piuttosto che un apporto basso o alto. In letteratura sono state proposte raccomandazioni del 15-20% e del 20-30% delle calorie provenienti dai grassi alimentari [15,52]. Tuttavia, sono necessarie ulteriori ricerche per stabilire l’effetto e la quantità ottimale di grassi alimentari per favorire l’ipertrofia muscolare.
Sulla base delle evidenze attuali, può essere prudente raccomandare che i grassi alimentari rappresentino il 20-35% delle calorie, in linea con le raccomandazioni dell’American College of Sports Medicine per gli atleti [53], che nella maggior parte dei casi corrispondono a circa 0,5-1,5 g/kg/giorno. Inoltre, va notato che un apporto sufficiente di proteine e carboidrati non deve essere compromesso da un’elevata assunzione di grassi nella dieta.
Anche la qualità dei grassi, come gli essenziali omega 3 e gli omega 6, potrebbe essere importante per i bodybuilder. Se l’apporto di questi acidi grassi è sufficiente, non è necessario integrarli con una dieta di alta qualità contenente buone fonti di acidi grassi. Tuttavia, per alcuni potrebbe essere difficile assumere le quantità ottimali. Per questo motivo, l’argomento verrà trattato in modo più approfondito nella sezione dedicata agli integratori alimentari.
Carboidrati:
A differenza delle proteine e dei grassi, i carboidrati sono considerati non essenziali per la dieta umana perché l’organismo è in grado di produrre il glucosio necessario ai tessuti attraverso la gluconeogenesi [54]. Tuttavia, l’assunzione di carboidrati ha un ruolo importante nella dieta del bodybuilder come regolatore degli ormoni tiroidei e come contributo al fabbisogno di micronutrienti [55,56]. Inoltre, una dieta a basso contenuto di carboidrati potrebbe limitare la rigenerazione dell’adenosina trifosfato (ATP) e limitare la capacità dei muscoli di contrarsi con una forza elevata [57,58]. Durante l’esercizio ad alta intensità, il glicogeno muscolare è il principale contributore di substrato energetico ed è stato dimostrato che la glicolisi fornisce circa l’80% del fabbisogno di ATP di una serie di flessioni del gomito se portata al cedimento muscolare [59]. Nonostante ciò, parte del glicogeno utilizzato durante questo tipo di esercizio può essere risintetizzato dal lattato, il che potrebbe ridurre il fabbisogno di carboidrati. È stato inoltre dimostrato che l’allenamento contro-resistenza riduce il glicogeno muscolare del 24-40% in una singola sessione [59,60].
La quantità esaurita può variare in base alla durata, all’intensità e al lavoro svolto, ma l’allenamento tipico del bodybuilding con ripetizioni più elevate e carichi moderati sembra causare la maggiore riduzione delle scorte di glicogeno muscolare [61]. Inoltre, è stato suggerito che quando le scorte di glicogeno sono troppo basse (~70 mmol/kg), ciò può inibire il rilascio di calcio e accelerare l’insorgenza della fatica muscolare [62]. Un basso livello di glicogeno muscolare riduce significativamente il numero di ripetizioni eseguite quando si eseguono tre serie di Squat all’80% di 1RM [57].
Tuttavia, è stato dimostrato che il consumo di una dieta contenente 7,7 g/kg/die di carboidrati per 48 ore prima di una sessione di allenamento non ha un effetto maggiore sulle prestazioni rispetto a 0,37g/kg/die quando si eseguono 15 serie a 15RM di esercizi per la parte inferiore del corpo [63]. Analogamente, un altro studio ha rilevato che una dieta con il 70% di carboidrati rispetto a una dieta con il 50% di carboidrati non ha un effetto maggiore sulle prestazioni durante l’esercizio sopramassimale; tuttavia, una dieta composta dal 25% di carboidrati ha ridotto significativamente le prestazioni [64].
Inoltre, visti gli effetti negativi a lungo termine sulla massa muscolare osservati di recente in studi su popolazioni allenate alla resistenza che seguono diete chetogeniche [49,51], potrebbe essere prudente per i bodybuilder assicurarsi semplicemente un apporto sufficiente di carboidrati, visti questi risultati disparati. Pertanto, mentre le diete a moderato e alto contenuto di carboidrati sono probabilmente appropriate per il bodybuilding, le diete a bassissimo contenuto di carboidrati possono essere dannose per l’allenamento.
Nei bodybuilder maschi, sono stati riportati apporti medi di carboidrati pari a 5,3g/kg/giorno durante la Off-Season [2]. Tuttavia, non sono state stabilite le quantità ottimali di carboidrati per i bodybuilder. In letteratura sono state proposte raccomandazioni per gli sport di forza, tra cui il bodybuilding, con assunzioni di 4-7g/kg/giorno e 5-6g/kg [15,65]. I carboidrati sembrano essere importanti per il bodybuilder, ma per ottenere benefici possono essere necessarie solo quantità moderate. Pertanto, dopo aver destinato le calorie alle proteine (1,6-2,2g/kg/die) e ai grassi (0,5-1,5g/kg/die), le restanti calorie dovrebbero essere destinate ai carboidrati. Tuttavia, sulla base delle prove attuali, potrebbe essere ragionevole consumare quantità sufficienti di carboidrati nell’intervallo ≥3-5g/kg/giorno, se possibile.
Sono necessarie ulteriori ricerche tra i bodybuilder per stabilire se l’assunzione abituale di carboidrati, superiore o inferiore a quella osservata, possa produrre ulteriori benefici. La Tabella sottostante riassume le raccomandazioni per le calorie e i macronutrienti.
Raccomandazioni dietetiche per i bodybuilder in Off-Season.
Distribuzione e timing dei nutrienti:
Si dice che i bodybuilder consumino in media sei pasti al giorno [66]; tuttavia, non esistono studi che esaminino specificamente quale possa essere la frequenza ottimale dei pasti per questa popolazione [65]. Questa elevata frequenza dei pasti si basa sulla convinzione di un maggiore stato di anabolismo e persino di un migliore utilizzo dei nutrienti durante il giorno, che potrebbe tradursi in un miglioramento della composizione corporea.
Il concetto di temporizzazione dell’assunzione di proteine per massimizzare l’ipertrofia comprende diverse strategie di dosaggio. La prima a comparire in letteratura è stata il consumo di proteine in prossimità dell’allenamento contro-resistenza. I picchi di MPS sono più elevati in questo periodo quando si consumano proteine; pertanto, questa strategia è stata proposta per migliorare l’efficienza della riparazione e del rimodellamento del muscolo scheletrico [31]. Inoltre, a causa dell'”effetto muscolo pieno”, per cui un ulteriore apporto di proteine non aumenta la MPS finché non è trascorso un tempo sufficiente, distribuire uniformemente l’assunzione di proteine tra più pasti è un’altra strategia studiata per massimizzare la MPS totale giornaliera [67]. Infine, il consumo prima di andare a letto di proteine a lenta digestione (come la caseina) per evitare periodi catabolici prolungati durante il sonno è la strategia proposta più di recente per migliorare il bilancio proteico netto giornaliero [68], sebbene si sia dimostrata inutile nel perseguire il fine o, per lo meno, non molto diversa dalla risultante di una assunzione di isolate in un contesto alimentare con parità nel totale proteico giornaliero. Ciascuna di queste tre strategie sarà discussa in seguito.
Dosaggio proteico:
Il periodo post-allenamento consente un picco della MPS più elevato quando si consumano proteine [31] e per raggiungere il picco di MPS può essere necessaria un’adeguata dose di Leucina “soglia” [32]. Diversi studi hanno esaminato il dosaggio proteico necessario per massimizzare la MPS dopo l’allenamento [69,70,71]. In uno studio sono stati consumati 0, 5, 10, 20 o 40g di proteine d’uovo intere dopo l’esercizio contro-resistenza della parte inferiore del corpo, con 20g che stimolavano al massimo la MPS [69]. Risultati simili sono stati riscontrati anche in un altro studio, in cui 20 g di siero di latte sono stati sufficienti a stimolare al massimo i tassi post-assorbitivi di MPS sia a riposo che dopo un lavoro unilaterale delle gambe all’80% del 1RM [70]. Inoltre, 40g di siero di latte non hanno prodotto ulteriori aumenti di MPS in questo studio e hanno portato all’ossidazione amminoacidica e alla produzione di urea.
Tuttavia, uno studio recente ha rilevato che, durante l’esecuzione di esercizi contro-resistenza per tutto il corpo al 75% del 1RM, 40g di siero di latte hanno prodotto una risposta MPS significativamente più elevata rispetto a 20g [71]. Esiste quindi una relazione tra il volume di tessuto muscolare danneggiato e stimolato e l’assunzione adeguata di proteine. È interessante notare che gli autori di una meta-analisi del 2013 hanno osservato che, nonostante gli studi con traccianti a breve termine mostrassero risposte nella MPS maggiori quando le proteine venivano consumate nella “finestra anabolica” post-allenamento, negli studi longitudinali sull’allenamento non è stato riscontrato alcun effetto significativo sull’ipertrofia quando si controllava l’apporto proteico totale giornaliero, indipendentemente dal fatto che le proteine fossero consumate all’interno della “finestra anabolica” o al di fuori di essa [72].
Nutrient Timing:
Analogamente, i ricercatori di uno studio tracciante a breve termine che ha esaminato il dosaggio delle proteine nel corso di 12 ore hanno riportato una maggiore area sotto la curva della MPS quando sono state consumate quattro dosi di proteine del siero di latte da 20g ogni tre ore rispetto a due dosi da 40g a distanza di sei ore e otto dosi da 10g ogni ora e mezza [73]. In teoria, data la soglia oltre la quale le proteine supplementari consumate in una singola seduta non contribuiscono ulteriormente alla MPS [69] e a causa del “periodo refrattario” postprandiale durante il quale la MPS non può essere nuovamente stimolata al massimo [67], si potrebbe concludere che un bodybuilder dovrebbe raggiungere, ma non superare, questa dose soglia ogni poche ore per massimizzare l’ipertrofia a lungo termine. Tuttavia, gli autori di una review sistematica del 2018 sugli integratori proteici, comprendente 34 studi randomizzati e controllati, hanno riportato guadagni di massa magra simili tra i gruppi che utilizzavano un programma di dosaggio con i pasti (che comportava un minor numero di dosi di proteine di entità elevata) e tra i pasti (che comportava un maggior numero di dosi di proteine di entità moderata) [74].
È interessante notare che i dati che esaminano l’alimentazione proteica notturna mostrano uno distacco simile tra gli studi meccanicistici a breve termine e gli interventi di allenamento a lungo termine. Nel 2012 è stata condotta la prima ricerca che esaminava la risposta acuta all’alimentazione notturna con caseina [68]. Gli autori hanno riportato che 40g di caseina consumati prima di andare a letto sono stati digeriti, assorbiti e hanno stimolato la MPS e migliorato l’equilibrio proteico dell’intero corpo durante il periodo notturno in misura maggiore rispetto al placebo. Negli anni successivi sono stati pubblicati altri studi in acuto che hanno confermato [75] e riconfermato questi risultati in una popolazione più anziana [76]. Nel 2015, gli autori del primo studio longitudinale hanno riportato un aumento della forza e dell’ipertrofia in un gruppo a cui era stato somministrato un supplemento proteico notturno rispetto a un gruppo placebo [77].
Tuttavia, la quantità totale di proteine giornaliere non è stata equiparata, in quanto il gruppo con proteine notturne ha consumato 1,9g/kg/giorno, mentre il gruppo placebo ha consumato solo 1,3g/kg. È importante notare che in entrambi gli unici studi longitudinali con corrispondenza proteica che hanno confrontato l’integrazione notturna di caseina con i gruppi che hanno assunto l’integrazione prima, non sono state riportate differenze significative nell’aumento della FFM tra i gruppi [78,79]. Pertanto, la domanda è la stessa per ogni strategia di distribuzione: perché ci sono ripetuti distacchi tra gli studi meccanicistici a breve termine sulle MPS e le ricerche a lungo termine che esaminano l’effettiva ipertrofia? La risposta potrebbe risiedere nei metodi utilizzati negli studi sulla MPS, in quanto i partecipanti sono a digiuno, ricevono solo proteine in polvere in isolamento, spesso viene loro somministrato del siero di latte (che viene digerito molto rapidamente) e vengono osservati per brevi periodi. Questi contesti di laboratorio determinano tempi di digestione e cinetiche degli aminoacidi diversi da quelli che si verificano nel “mondo reale”. In particolare, in queste condizioni di laboratorio i livelli di base degli aminoacidi nel corpo sono più bassi del normale e la digestione e il successivo apporto di aminoacidi al muscolo sono più rapidi.
In condizioni di vita libera, le proteine vengono consumate principalmente da fonti alimentari intere, più volte al giorno e insieme ad altri alimenti, il che ritarda lo svuotamento gastrico. Per questi motivi, gli aminoacidi vengono titolati nel flusso sanguigno in modo più lento e costante; pertanto, in condizioni normali, le scorte sono quasi sempre prontamente disponibili [80]. Pertanto, l’efficacia della “finestra anabolica” e persino delle strategie di distribuzione delle proteine potrebbe non tradursi nella pratica. Inoltre, le limitazioni specifiche del laboratorio si estendono anche agli studi sull’alimentazione notturna. Si consideri, ad esempio, che 26g di proteine provenienti da una bistecca magra determinano un aumento sostenuto della MPS che dura almeno sei ore (l’intero periodo di tempo studiato) [81].
Inoltre, 26g sono solo il ~37% della dose di proteine contenuta in media in una cena americana [82], che richiederebbe più tempo per essere digerita a causa della maggiore porzione di proteine e dell’aggiunta di fibre, lipidi e altri nutrienti che ritarderebbero ulteriormente la digestione [80]. Pertanto, il tipico pasto finale potrebbe già soddisfare lo scopo di un frullato di caseina. Detto questo, nonostante queste discrepanze tra MPS e risultati della composizione corporea, non c’è nulla di male nel tentare queste strategie, soprattutto se attuate in modo pragmatico e senza introdurre ulteriori oneri logistici nel proprio programma quotidiano.
Pertanto, potrebbe essere prudente consigliare ai bodybuilder di suddividere l’assunzione giornaliera di 1,6-2,2 g/kg di proteine in più pasti contenenti ciascuno ~0,40-0,55g/kg [80] e di fare in modo che uno di questi pasti avvenga entro 1-2 ore prima o dopo l’allenamento, mentre un’alimentazione costituita da una fonte proteica e non proteica venga consumata 1-2 ore prima di dormire. Ad esempio, un bodybuilder di 90 kg potrebbe consumare 40-50g di proteine alle 8-9 del mattino per la colazione, allenarsi alle 11, consumare 40-50g di proteine alle 12-13 per il pranzo/post-allenamento, 40-50g di proteine a cena tra le 17-18, e poi un pasto finale di 40-50g di proteine non contenenti fonti proteiche grasse alle 21-10 prima di andare a letto entro le 23.
I carboidrati consumati prima dell’allenamento sono spesso una strategia utilizzata dagli atleti per migliorare le prestazioni negli esercizi ad alta intensità. La completa risintesi del glicogeno può essere raggiunta entro 24 ore da un allenamento che depaupera il glicogeno se si consumano quantità sufficienti di carboidrati [83]. Tuttavia, solo il 24-40% del glicogeno muscolare viene esaurito dopo un allenamento contro-resistenza [59,60]. Pertanto, una quantità di ≥3-5g/kg di carboidrati al giorno sarebbe probabilmente sufficiente per la risintesi del glicogeno. Questo elevato apporto giornaliero di carboidrati probabilmente riduce anche l’impatto della tempistica dei carboidrati pre-allenamento sulle prestazioni dell’esercizio.
Inoltre, per i bodybuilder che non hanno bisogno di enfatizzare il rifornimento di glicogeno, le proteine aumentano la MPS post-allenamento a livelli massimi anche senza l’aggiunta di carboidrati [86,87]. Anche se il consumo di carboidrati nel post-allenamento non è certo dannoso, è improbabile che questo favorisca l’ipertrofia a lungo termine, come discusso in precedenti review [1,88]. Pertanto, è meglio concentrarsi sul consumo di un’adeguata quantità di carboidrati giornalieri e basare la distribuzione dei carboidrati intorno all’allenamento sulle preferenze personali.
Supplementazione OTC:
In un recente sondaggio condotto tra i bodybuilder, è stato riportato che tutti i partecipanti assumevano integratori alimentari [9]. Gli integratori alimentari più comuni erano: integratori di proteine (86%), creatina (68%), aminoacidi a catena ramificata (67%), glutammina (42%), vitamine (40%), olio di pesce (37%) e prodotti contenenti caffeina/efedrina (24%).
Sebbene gli integratori proteici siano molto popolari tra i bodybuilder, vengono utilizzati prevalentemente come gli alimenti interi per raggiungere gli obiettivi proteici. Pertanto, non verranno discussi in dettaglio. I lettori sono invitati a leggere la posizione dell’ISSN su questo argomento [89]. Inoltre, la trattazione di tutti gli integratori comunemente utilizzati dai bodybuilder esula dallo scopo di questo articolo. L’attenzione si concentrerà piuttosto sugli integratori alimentari che potrebbero potenzialmente produrre un effetto ergogenico e sugli integratori che possono garantire un apporto sufficiente di micronutrienti e acidi grassi essenziali.
Creatina Monoidrato:
La Creatin-fosfato si trova in alte concentrazioni nel muscolo scheletrico e cardiaco, dove agisce come fonte di energia [90]. La Creatina può essere ottenuta anche attraverso la dieta nei soggetti che consumano carne; tuttavia, le concentrazioni di Creatina nella carne si riducono con la cottura [91].
Numerosi studi hanno osservato un aumento della massa e della forza muscolare in seguito a fasi di carico di Creatina, in genere di 20g al giorno per circa una settimana, spesso seguite da fasi di mantenimento di 2-3g di Creatina al giorno [92]. Tuttavia, la fase di carico potrebbe non essere necessaria. È stato dimostrato che la saturazione della Creatina muscolare dopo un’integrazione di 3g di Creatina Monoidrato per 28 giorni è simile al consumo di Creatina Monoidrato dopo la tipica fase di carico [93].
La maggior parte degli individui non raggiunge i 3g giornalieri con la dieta e può essere necessaria un’integrazione. Esistono numerose forme di Creatina negli integratori in commercio, tra le quali la Creatina Monoidrato è la più studiata. Le versioni più recenti di Creatina, come la kre-alkalyn [94] e la Creatina etil-estere [95], non si sono dimostrate superiori alla Creatina Monoidrato, nonostante abbiano in genere un prezzo più elevato. Pertanto, si raccomanda il consumo di 3-5g di Creatina Monoidrato al giorno. La tempistica di assunzione della Creatina non sembra avere importanza, poiché la saturazione delle riserve di Creatin-fosfato richiede circa 28 giorni per raggiungere le concentrazioni massime quando si consumano 3g al giorno e non ha un effetto in acuto [93].
Caffeina:
Uno degli integratori alimentari più utilizzati dai bodybuilder sono gli stimolanti, in particolare la Caffeina [9]. Oltre ad aumentare l’eccitazione [96], la Caffeina può ridurre il dolore e lo sforzo percepito durante l’esercizio [97] e migliora la gestione del Calcio, aumentando la potenza [98]. Studi sull’allenamento contro-resistenza hanno rilevato che la Caffeina riduce la fatica e aumenta la forza [99,100]. Tuttavia, non tutti gli studi hanno dimostrato un effetto ergogenico sull’allenamento contro-resistenza [101]. Gli studi che hanno dimostrato un effetto ergogenico hanno utilizzato dosaggi elevati di caffeina (5-6 mg/kg), che sono al limite superiore di quello che è considerato un dosaggio sicuro [99,100]. Tuttavia, può essere consigliabile consumare il dosaggio minimo efficace per individuo, poiché l’assunzione regolare può generare tolleranza [102]. A causa dell’effetto acuto della Caffeina, è consigliabile assumerla circa 1 ora prima dell’esercizio fisico [99]. Tuttavia, l’emivita della Caffeina è di circa 3-9 ore; pertanto, può essere consigliabile consumare la Caffeina all’inizio della giornata per favorire un sonno sano se l’esercizio fisico viene svolto più tardi nel corso della giornata [103]. Sono necessarie ulteriori ricerche per trovare un consenso sull’uso della Caffeina nell’allenamento contro-resistenza, ma sulla base delle prove attuali un dosaggio di 5-6 mg/kg consumato prima dell’esercizio potrebbe produrre un effetto ergogenico sulle prestazioni nell’allenamento contro-resistenza.
Beta-Alanina:
È stato dimostrato che l’ingestione di 4-6 g di beta-alanina aumenta i livelli di carnosina muscolare [104]. La carnosina agisce come tampone del pH nel muscolo scheletrico e può ritardare l’inizio dell’affaticamento muscolare durante l’esercizio ad alta intensità [105]. Una meta-analisi ha concluso che la beta-alanina potrebbe produrre effetti ergogenici durante l’esercizio ad alta intensità della durata di 60-240 secondi [104]. Inoltre, non sono stati riscontrati effetti benefici negli esercizi di durata inferiore a 60 secondi. La maggior parte degli studi inclusi nella meta-analisi riguardava l’esercizio di resistenza.
Tuttavia, è dimostrato che l’integrazione di beta-alanina può migliorare la resistenza muscolare negli atleti allenati alla resistenza [105] e può migliorare la composizione corporea [106]. Sono necessari ulteriori studi per esaminare l’effetto ergogenico della beta-alanina sulla composizione corporea e sulle prestazioni. Tuttavia, dato che i bodybuilder si allenano spesso con più di 10 ripetizioni per serie e spesso includono tecniche di intensità come drop set, pause di riposo, myo reps e altre, la beta-alanina potrebbe apportare un beneficio alla resistenza di queste serie [9].
Pertanto, potrebbe essere ragionevole per un bodybuilder consumare 3-5 g di beta alanina al giorno durante le fasi di allenamento ad alte ripetizioni o nelle fasi di allenamento in cui si incorporano diverse tecniche di intensità che prolungano la durata di un set. Come la creatina monoidrato, la beta-alanina non ha un effetto acuto, in quanto le concentrazioni di carnosina muscolare richiedono circa 4 settimane per raggiungere concentrazioni tali da produrre un effetto ergogenico, a condizione che se ne consumi una quantità sufficiente al giorno [104].
Citrullina Malato:
Recentemente, la Citrullina Malato ha guadagnato popolarità tra i bodybuilder. Il potenziale effetto ergogenico è dovuto all’aumento del flusso ematico al muscolo, alla produzione di ATP e alla potenziale capacità della Citrullina Malato di agire come agente tampone [107]. È stato dimostrato che il consumo di 8g di Citrullina Malato aumenta le ripetizioni fino al cedimento del 50% [107,108,109,110], riduce l’indolenzimento muscolare del 40% [107] e migliora la forza massimale e la potenza anaerobica [111].
Tuttavia, non tutti gli studi hanno osservato effetti ergogenici del consumo di Citrullina Malato. Due studi recenti non hanno mostrato un miglioramento delle prestazioni, un aumento della risposta del gonfiore muscolare dovuto all’allenamento, un’attenuazione della fatica o un aumento dell’attenzione e dell’energia in seguito all’integrazione di Citrullina Malato in uomini allenati contro-resistenza a livello amatoriale [112,113].
Una recente meta-analisi di Trexler et al. ha analizzato 12 studi sullla CM per le prestazioni di forza e potenza [114]. Sebbene abbiano riscontrato solo una piccola dimensione dell’effetto (0,20), hanno concluso che questo potrebbe essere rilevante per gli atleti di alto livello in cui i risultati delle competizioni si decidono su margini ridotti, come i culturisti agonisti di alto livello. Si consiglia di assumere la Citrullina Malato circa 60 minuti prima dell’esercizio fisico per consentire un assorbimento sufficiente.
Sono necessarie ulteriori ricerche per determinare l’efficacia della Citrullina Malato nell’esercizio contro-resistenza. Allo stato attuale, i dati indicano un effetto benefico o neutro sulle prestazioni. Pertanto, sulla base delle prove attuali, 8g al giorno di Citrullina Malato consumati prima dell’esercizio potrebbero avere dei benefici interessanti per i bodybuilder.
Alfa-GPC:
L’Alfa-GPC (alfa-glicerofosfocolina o colina alfoscerato) è un fosfolipide contenente colina. Quando viene ingerita, l’Alfa-GPC viene metabolizzata in colina e glicerolo-1-fosfato. La colina è un precursore dell’acetilcolina, un neurotrasmettitore coinvolto nella memoria, nell’attenzione e nella contrazione dei muscoli scheletrici. Il glicerolo-1-fosfato serve a sostenere le membrane cellulari.[https://pubmed.ncbi.nlm.]
L’Alfa-GPC sembra attraversare facilmente la barriera emato-encefalica e viene assorbito rapidamente. Attualmente è il miglior colinergico per aumentare i livelli plasmatici e cerebrali di colina.[https://pubmed.ncbi.nlm.]
L’integrazione orale di Alfa-GPC è interessante soprattutto per scopi nootropici o di potenziamento cognitivo. Esistono numerosi studi sui roditori che supportano questo effetto, ma non è ancora stato dimostrato negli esseri umani altrimenti sani. Negli anziani affetti da demenza lieve o moderata – che comporta un’alterazione della neurotrasmissione colinergica – l’Alfa-GPC migliora i sintomi cognitivi (ad esempio, disturbi della memoria e dell’attenzione).[https://pubmed.ncbi.nlm] L’Alfa-GPC può anche migliorare l’efficacia degli inibitori dell’acetilcolinesterasi (cioè i farmaci che aumentano la disponibilità di acetilcolina rallentandone la degradazione), utilizzati per il trattamento della malattia di Alzheimer.[https://pubmed.ncbi.nlm.]
Gli atleti sono un’altra popolazione che può trarre beneficio dall’integrazione di Alfa-GPC. Prove preliminari suggeriscono che l’alfa-GPC aumenta la potenza del salto verticale.[https://jissn.biomedcentral.com][https://pubmed.ncbi.nlm.] Inoltre, uno studio pilota ha riportato che l’Alfa-GPC ha aumentato il picco di forza nella panca, ma non la potenza di picco o il tasso di sviluppo della forza.[Ziegenfuss T, Landis J, Hofheins JJ Int Soc Sports Nutr.] Attualmente non è chiaro se l’Alfa-GPC aumenti la forza isometrica, ma i dati empirici e aneddotici sono incoraggianti [https://pubmed.ncbi.nlm.]
L’integrazione di un dosaggio pari a 600mg di Alpha-GPC prima di un test di potenza (spinte su panca) ha riportato un miglioramento della potenza del 14% rispetto al placebo quando assunta 45 minuti prima dell’attività; si trattava di uno studio pilota.[http://www.jissn.com] In media si è notato che il dosaggio di Alfa-GPC efficacie per trarre miglioramenti nella forza è nel range dei 300-600mg 45-30 minuti prima della seduta allenante.
Multi Vitaminico-Multi Minerale:
Storicamente, i bodybuilder hanno utilizzato diete restrittive che eliminano alimenti o interi gruppi di alimenti. Di conseguenza, sono comuni numerose carenze di vitamine e minerali. Nei bodybuilder a dieta sono state osservate carenze di Calcio, vitamina D, Zinco, Ferro e altre ancora [115,116,117]. Tuttavia, la maggior parte della letteratura sulle pratiche alimentari dei bodybuilder risale agli anni ’80 e ’90; pertanto, sono necessari dati più recenti [2].
Più di recente, le pratiche alimentari dei bodybuilder che seguono una dieta tradizionale restrittiva sono state confrontate con quelle degli agonisti che utilizzano un approccio dietetico basato sui macronutrienti, in cui nessun alimento o gruppo alimentare è off limits [118]. Non sorprende che i concorrenti che utilizzano un approccio dietetico più flessibile presentino meno carenze di micronutrienti. In particolare, la vitamina E, la vitamina K e le proteine sono risultate significativamente inferiori nelle donne che utilizzavano approcci dietetici rigidi rispetto a quelle che utilizzavano approcci più flessibili. Nel presente articolo, specie se si parla di Off-Season, si raccomanda di utilizzare un approccio dietetico flessibile, in cui nessun alimento o gruppo viene eliminato dalla dieta.
In questo modo, è meno probabile che si verifichino carenze di micronutrienti, soprattutto se si considera che le atlete in Off-Season hanno a disposizione una maggiore quantità di calorie rispetto a quelle a dieta per un contest, il che dovrebbe consentire loro di incorporare una maggiore varietà di alimenti.
Ciononostante, può essere consigliabile raccomandare un integratore multivitaminico/minerale a basso dosaggio (≤100% RDA) come misura di sicurezza per prevenire eventuali carenze di micronutrienti, sottolineando al contempo il consumo di una buona varietà di alimenti al giorno per soddisfare il fabbisogno di micronutrienti.
Omega 3 (EPA-DHA):
Gli acidi grassi polinsaturi con un doppio legame a tre atomi di distanza dal gruppo metilico terminale sono noti come ω-3 o acidi grassi omega-3 (O3). Un basso apporto di O3 nelle diete occidentali rispetto ad altre fonti di grassi alimentari (come gli acidi grassi omega-6) è associato a un peggioramento della salute multispettrale negli studi epidemiologici [119]. Pertanto, è interessante concentrarsi specificamente sulle modifiche della dieta per fornire acidi eicosapentaenoici e docosaesaenoici (EPA e DHA) – la carenza alimentare più comune nel mondo occidentale; ma vale la pena notare che la misurazione, l’interazione e l’effetto di O3 e acidi grassi omega-6 in relazione alla salute non sono chiari e vanno oltre lo scopo di questo articolo. Per una rassegna si rimanda ad altra pubblicazione [120].
Oltre alla salute, c’è interesse per i potenziali effetti anabolici degli integratori di EPA e DHA [121], che di solito vengono forniti attraverso l’olio di pesce o, in alcuni casi, l’olio di alghe. Tuttavia, ci sono dati contrastanti sulla capacità dell’olio di pesce di aumentare la risposta della sintesi proteica muscolare all’ingestione di proteine. Mentre un articolo di revisione del 2014 ha evidenziato una serie di studi secondo cui l’olio di pesce può aumentare la risposta [122], uno studio recente non ha rilevato alcun effetto sulla risposta della MPS a una sessione di allenamento contro-resistenza e all’ingestione di proteine dopo l’allenamento [123]. Inoltre, i dati sull’ipertrofia longitudinale sono pochi [124] e gli studi sulle prestazioni dell’allenamento contro-resistenza sono contrastanti [125] e in gran parte non applicabili o difficili da valutare a causa dell’uso di partecipanti non allenati o di allenamenti non standardizzati ed ecologicamente non realistici rispetto al bodybuilding.
In una recente review che affronta specificamente la questione se gli integratori di O3 possano o meno aumentare l’ipertrofia [126], gli autori hanno concluso che attualmente non ci sono prove sufficienti per fare tale affermazione. Sebbene siano necessarie ulteriori ricerche prima di poter raccomandare l’integrazione di O3 (o di alterazioni della dieta) a fini di costruzione muscolare, i benefici per la salute dell’integrazione di O3 sono degni di nota. Ad esempio, recenti meta-analisi hanno riportato che l’integrazione di olio di pesce riduce i sintomi della depressione [127], diminuisce il rischio di morte cardiaca [128], riduce la pressione sanguigna [129] e diminuisce la circonferenza vita [130]. Pertanto, gli atleti estetici possono prendere in considerazione l’integrazione giornaliera di olio di pesce (o di alghe) (1.5-2.5g di EPA/DHA) per la salute generale e multi spettro, ma sono necessari studi futuri per formulare raccomandazioni relative alle prestazioni nel bodybuilding.
Acido Arachidonico (AA):
L’Acido Arachidonico (AA) è l’acido grasso omega-6 più rilevante dal punto di vista biologico e, nella membrana lipidica di una cellula, è l’acido grasso che viene confrontato con i due acidi grassi dell’olio di pesce (EPA e DHA) nella costituzione di un rapporto omega-3:6. Dati recenti suggeriscono un’assunzione giornaliera di 50-250mg di Acido Arachidonico[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] con alcune fonti che stimano livelli fino a 500mg al giorno;[https://www.ncbi.nlm.] l’assunzione di Acido Arachidonico sembra essere inferiore nei vegetariani[https://www.ncbi.nlm.].
Si ritiene che l’Acido Arachidonico sia importante per il metabolismo del muscolo scheletrico, poiché si pensa che i fosfolipidi della membrana del sarcoplasma riflettano la dieta,[https://www.ncbi.nlm.][https://www.ncbi.nlm.] l’allenamento stesso sembra alterare il contenuto di fosfolipidi del muscolo (indipendentemente dalla composizione delle fibre muscolari[https://www.ncbi.nlm.] e associato a un rapporto omega 6:3 più basso[https://www.ncbi.nlm.][https://www.ncbi.nlm.]) e gli eicosanoidi dell’Acido Arachidonico interagiscono con la sintesi proteica muscolare attraverso i loro recettori.
L’Acido Arachidonico segnala la sintesi proteica muscolare attraverso una via dipendente dalla COX-2 (che suggerisce il coinvolgimento delle prostaglandine)[https://www.ncbi.nlm.] che è associata ad aumenti sia della prostaglandina E2 (PGE2) che del PGF(2α),[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] anche se l’incubazione con PGE2 o PGF(2α) isolati non sembra replicare pienamente gli effetti ipertrofici dell’Acido Arachidonico. [https://www.ncbi.nlm.] PGE2 e PGF(2α) sono indotti anche dall’esercizio fisico (nello specifico, dallo stiramento delle cellule muscolari in vitro[https://www.ncbi.nlm.]) ed è stato osservato sia nel siero[https://pubmed.ncbi.nlm.][https://www.ncbi.nlm.] che a livello intramuscolare (quadruplicato, da 0,95+/-0,26ng/mL a 3,97+/-0. La capacità del riflesso da stiramento di aumentare le concentrazioni di PGE2 e PGF(2α)[https://www.ncbi.nlm.] potrebbe essere dovuta semplicemente al fatto che lo stiramento aumenta l’attività delle COX2.[https://www.ncbi.nlm.][https://www.ncbi.nlm.]
Va notato che l’integrazione di 1.500mg di Acido Arachidonico (rispetto a una dieta di controllo contenente 200mg dello stesso) per 49 giorni ha aumentato la secrezione di PGE2 da parte di cellule immunitarie stimolate (del 50-100%) in giovani uomini altrimenti sani,[https://www.ncbi.nlm.] ma la rilevanza di questo studio per il muscolo scheletrico non è nota. Questo studio ha anche osservato che, senza stimolazione, non c’erano differenze significative tra i gruppi.[https://www.ncbi.nlm.] Altrove, è stata osservata una tendenza all’aumento delle concentrazioni sieriche di PGE2 a riposo in uomini allenati a cui sono stati somministrati 1.000mg di Acido Arachidonico per 50 giorni.[https://www.ncbi.nlm.]
L’Acido Arachidonico, attraverso gli eicosanoidi noti come PGF(2α) e PGE2, stimola la sintesi proteica muscolare. Sono prodotti a partire dall’Acido Arachidonico, ma normalmente non formano i rispettivi eicosanoidi per la costruzione del muscolo finché la cellula non viene stimolata da un fattore di stress (come il riflesso di stiramento di una cellula muscolare) che ne induce la produzione.
Il recettore per il PGF(2α) (recettore FP) sembra essere sovraregolato dagli inibitori della COX1 (l’acetaminofene utilizzato in questo studio)[https://www.ncbi.nlm.] e si ritiene che una maggiore segnalazione del PGF(2α) sia alla base del miglioramento della sintesi proteica muscolare osservato nei soggetti anziani con farmaci antinfiammatori. La supplementazione di Acido Arachidonico non sembra influenzare la quantità di recettori FP nei giovani;[https://www.ncbi.nlm.] mentre l’esercizio fisico stesso può aumentare il contenuto di recettori EP3, né gli inibitori della COX1[https://www.ncbi.nlm.] né l’Acido Arachidonico[https://www.ncbi.nlm.] sembrano influenzarlo ulteriormente.
Tuttavia, è stato riscontrato che l’uso di inibitori della COX2 (nei giovani) sopprime l’aumento di PGF(2α) indotto dall’esercizio fisico (Ibuprofene e Acetaminofene)[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e di PGE2,[https://www.ncbi.nlm.] il che si pensa sia dovuto al fatto che la conversione da PGH2 in questi metaboliti dipende dall’attività della COX2.
Poiché la produzione di questi eicosanoidi dipende dall’enzima COX2, si ritiene che l’inibizione di questo enzima riduca gli effetti anabolizzanti dell’esercizio fisico se assunto prima dello stesso.
L’acido arachidonico (così come l’EPA dall’olio di pesce) non ha compromesso l’assorbimento del glucosio nelle cellule muscolari isolate e 10μM di acido grasso sono in grado di attenuare la resistenza all’Insulina indotta dai grassi saturi; [https://pubmed.ncbi.nlm.] un fenomeno osservato con i grassi saturi a 18 o più catene di carbonio[https://www.ncbi.nlm.] che non sembra applicarsi agli acidi grassi polinsaturi di uguale lunghezza di catena[https://www.ncbi.nlm.][https://www.ncbi.nlm.] ed è probabilmente legato all’aumento delle ceramidi intracellulari[https://www.ncbi.nlm.] che compromettono la segnalazione di Akt[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e riducono l’assorbimento di glucosio mediato da GLUT4 con l’Insulina.[https://www.ncbi.nlm.]
L’Acido Arachidonico e i grassi polinsaturi omega-3 sono entrambi associati a una migliore sensibilità all’Insulina delle cellule muscolari, che potrebbe essere secondaria alla riduzione dei livelli di grassi saturi nella membrana lipidica e quindi alla riduzione delle concentrazioni intracellulari di ceramidi. È possibile che ciò non sia correlato agli eicosanoidi o al rapporto omega-3:6.
In 31 uomini allenati, sottoposti a un programma di sollevamento pesi e a una dieta standardizzata (500kcal in eccesso con 2g/kg di proteine) con 1g di Acido Arachidonico al giorno o placebo, l’integrazione per 50 giorni è sembrata aumentare la potenza di picco (7,1%) e la potenza media (3,6%) al test di Wingate, ma non è riuscita a influenzare positivamente la massa muscolare o le misure di potenza del sollevamento pesi (bench press e leg press).[https://www.ncbi.nlm.]
Attualmente non ci sono prove sufficienti per raccomandare una dose ideale di integrazione di Acido Arachidonico, ma aneddoticamente si usa un dosaggio di circa 1.500 mg da assumere 45 minuti prima dell’allenamento per un periodo medio di 8 settimane. Non è certo che si tratti di una dose ottimale o che sia necessaria la tempistica.
Va inoltre notato che per le persone affette da patologie infiammatorie croniche, come l’artrite reumatoide o le malattie infiammatorie intestinali, la dose ideale di Acido Arachidonico può essere in realtà una sua restrizione dietetica. Nei casi di malattie infiammatorie, l’integrazione di Acido Arachidonico è probabilmente controindicata.
Raccomandazioni per gli integratori alimentari e il dosaggio per i bodybuilder in Off-Season:
Creatina Monoidrato= 3-5g/die;
Beta-Alanina= 3-5g/die;
Citrullina Malato= 8g/pre-workout;
Alfa-GPC= 300-600mg/pre-workout;
Caffeina= 5-6mg/Kg/pre-workout (media standard tra 200 e 600mg/die);
Multi Vitaminico – Multi Minerale= ≤100% RDA/die;
Omega 3 (EPA-DHA)= 1.5-2.5g/die;
Acido Arachidonico= 1.5g/pre-workout.
Supplementazione PEDs:
Una cosa occorre premettere prima di procedere con la descrizione delle molecole più utilizzate nel contesto della Off-Season: non esistono PEDs esclusivamente confinabili in uno dei contesti della programmazione di un bodybuilder. Esiste il grado di versatilità il quale sta ad indicare quanto una molecola possa essere gestita con facilità in situazioni preparatorie differenti. Esistono molecole che per caratteristiche possono dare vantaggi maggiori in Off-Season/Bulk per via di alcune loro caratteristiche che in altro contesto, per esempio il pre-contest, risulterebbero più complesse da gestire. Ma questo non significa che tali molecole siano generalemnte da considerarsi “off-limitz” in un altra fase della preparazione annuale.
Premesso ciò, l’attenzione in questo paragrafo si concentrerà sui principali PEDs usati in Off-Season.
Tra tutti gli AAS, il Testosterone è quello che non ha bisogno di particolari presentazioni. Si tratta dell’ormone sessuale maschile per antonomasia. Nell’uomo, il Testosterone svolge un ruolo fondamentale nello sviluppo dei tessuti riproduttivi maschili, come i testicoli e la prostata, oltre a promuovere le caratteristiche sessuali secondarie, come l’aumento della massa muscolare e ossea e la crescita dei peli. Inoltre, in entrambi i sessi, il Testosterone è coinvolto nella salute e nel benessere, compresi gli stati d’animo, il comportamento e la prevenzione dell’osteoporosi in cooperazione con l’Estradiolo. Livelli insufficienti di Testosterone negli uomini possono portare ad anomalie, tra cui la fragilità e la perdita ossea.
In generale, il Testosterone promuove la sintesi proteica e quindi la crescita dei tessuti dotati di recettori per gli androgeni. Il Testosterone può essere descritto come avente effetti virilizzanti e anabolizzanti (anche se queste descrizioni categoriali sono in qualche modo arbitrarie, poiché vi è una grande sovrapposizione reciproca tra di essi).
Gli effetti anabolizzanti comprendono la crescita della massa e della forza muscolare, l’aumento della densità e della resistenza ossea e la stimolazione della crescita lineare e della maturazione ossea.
Gli effetti androgeni comprendono la maturazione degli organi sessuali, in particolare del pene, e la formazione dello scroto nel feto, e dopo la nascita (di solito nella pubertà) l’approfondimento della voce, la crescita dei peli del viso (come la barba) e dei peli ascellari. Molti di questi effetti rientrano nella categoria dei caratteri sessuali secondari maschili.
Al principio degli anni 30 del novecento avvenne la sintesi chimica del Testosterone, quando Butenandt e G. Hanisch pubblicarono un articolo che descriveva “Un metodo per preparare il Testosterone dal colesterolo”. Solo una settimana dopo, il terzo gruppo, Ruzicka e A. Wettstein, annunciò una domanda di brevetto in un documento “Sulla preparazione artificiale dell’ormone testicolare Testosterone (Androsten-3-one-17-ol).” Ruzicka e Butenandt ricevettero il premio Nobel per la chimica nel 1939 per il loro lavoro.
Gli studi clinici sull’uomo, che prevedevano dosi PO (per via orale) di Methyltestosterone o iniezioni di Testosterone Propionato, iniziarono già nel 1937. Il Testosterone Propionato è menzionato in una lettera all’editore della rivista Strength and Health nel 1938; questo è il primo riferimento noto a un AAS in una rivista statunitense di sollevamento pesi o Bodybuilding.
Lo sviluppo delle proprietà di costruzione muscolare del Testosterone proseguì negli anni ’40, in Unione Sovietica e nei paesi del blocco orientale come la Germania dell’Est, dove sono stati utilizzati programmi di AAS per migliorare le prestazioni dei sollevatori di pesi olimpici e di altri dilettanti già prima degli anni ’50. In risposta al successo dei sollevatori di pesi russi, il medico della squadra olimpica statunitense John Ziegler lavorò con un equipe di chimici per sviluppare un AAS con effetti androgeni ridotti. Ma questa è un altra storia.
L’uso del Testosterone nello sport si diffuse tra gli anni ’50 e gli anni ’60. Le forme utilizzate nei primi tempi erano il Testosterone in sospensione e il Testosterone Propionato, che rappresentano con il Methyltestosterone (Testosterone metilato in C-17) le forme più datate dell’ormone in questione (1935).
In ambito culturistico, il Testosterone rappresenta un AAS sufficientemente versatile in maniera dose-dipendente e sensibilità-dipendente dal momento che il dosaggio dovrebbe essere tarato in base alle risposte metaboliche soggettive alle quali è soggetto l’ormone (vedi, ad esempio, aromatizzazione in estrogeni). Questo ultimo punto è di estrema importanza al fine di evitare l’uso/abuso di AI (Inibitori dell’Aromatasi) e/o SERM (Modulatori Selettivi del Recettore degli Estrogeni). Oltre a peggiorare potenzialmente il quadro lipidico, sommandosi all’azione degli AAS utilizzati, essi riducono l’espressione epatica di IGF-1 cosa che può ridurre la risposta anabolizzante del protocollo PEDs. Nei soggetti caratterizzati da una elevata sensibilità all’attività estrogenica, le procedure applicate vedono: 1) l’uso di Raloxifene o Tamoxifene (SERM) a dosi sufficienti a impedire la comparsa o il peggioramento di una ginecomastia in stadio iniziale già presente e non ancora asportata chirurgicamente 2) l’uso di dosi fisiologiche di Testosterone come base onde evitare la comparsa di stati letargici, affaticabilità, disfunzioni sessuali ecc 3) l’uso di un “mix” composto da Testosterone e Boldenone (vedi in seguito) tale da poter usufruire della bassa e diversa sensibilità all’azione dell’Enzima Aromatasi su quest’ultimo riuscendo ad avere un controllo estrogenico teoricamente migliore (Testosterone e Boldenone mostrano qualità anabolizzanti intrinseche simili).
In un contesto Off-Season, quindi, vista l’importanza della presenza di un buon livello di Estradiolo sia sul complesso degli effetti anabolizzanti ricercati sia per la sua attività sessuale e cerebrale, il Testosterone andrebbe inizialmente calibrato sul soggetto e nel caso affiancato da dosi altrettanto ben tarate di SERM la dove ne risultasse un reale bisogno.
L’uso di un estere che garantisca un rilascio graduale della molecola (vedi Enantato o Cypionato) risulta la scelta migliore al fine di creare una soglia ematica stabile e esente da picchi e cali che possono risultare controproducenti a livello psicofisico. Tenere sempre in considerazione l’emivita di una molecola è uno dei punti fondamentali per sfruttarla al meglio. Nel caso degli esteri sopra citati, una divisione del dosaggio settimanale in due somministrazioni uguali distanziate da quattro-cinque giorni l’una dall’altra risulta una pratica ottimale allo scopo di creare una soglia ematica stabile.
I dosaggi comunemente utilizzati, parlando di molecole esterificate, vanno da 200mg ad 1g a settimana. Per quanto riguarda il Testosterone in sospensione, le dosi comunemente utilizzate vanno dai 175mg ai 700mg a settimana.
Il Boldenone [1,4-androstadiene-3-one,17b-ol], commercializzato con il nome di Equipoise, Ganabol, Equigan, Ultragan, e Boldane, è uno steroide anabolizzante-androgeno spesso legato all’estere Undecylenato. Strutturalmente molto simile al Testosterone, il Boldenone differisce da questo per il doppio legame tra C1 e C2.
La Ciba brevettò il Boldenone nel 1949. Successivamente, negli anni ’50 e ’60, sviluppò diversi esteri sperimentali del farmaco. Uno di questi era il Boldenone Undecilenato, che fu introdotto per uso clinico con il marchio Parenabol e fu utilizzato alla fine degli anni ’60 e all’inizio degli anni ’70. Tuttavia, fu sospeso prima della fine degli anni ’70. Ad oggi l’uso del Boldenone è legale in alcuni paesi in campo veterinario.
Se qualcuno volesse usare 500mg di Testosterone, ma non potrebbe usare un tale dosaggio dal momento che presenta particolare difficoltà nella gestione estrogenica in specie senza l’uso di AI come Exemestane o Anastrozolo, una conclusione a cui molti superficialmente sono giunti è che si potrebbe semplicemente usare il Boldenone al dosaggio sopra citato per ridurre della metà l’attività estrogenica, ma comunque supportare un’adeguata produzione di Estradiolo. Ma quando si approfondisci l’ipotesi e la si testa sul campo, è davvero così che stanno le cose? In realtà no, o, comunque, la media delle variabili di risposta spinge a confermare una maggiore validità nel “mixare” Testosterone e Boldenone coprendo la dose base calcolata in precedenza, e con variazione di percentuale T:B ratio da 1:1 a 2:1.
Comunque, oltre a rappresentare genericamente una discreta molecola sia in in preparazione alla gara che in Off-Season, I dosaggi utilizzati si settano nel range tra i 200mg ed i 500mg a settimana, spesso abbinato ad una dose variabile (vedi sopra) di Testosterone.
Il Nandrolone, noto anche come 19-nortestosterone, è uno Steroide Androgeno Anabolizzante (AAS) utilizzato sotto forma di molecola legata a esteri come quello Decanoato (nome commerciale Deca-Durabolin) e il Fenilpropionato (nome commerciale Durabolin). Gli esteri del Nandrolone sono utilizzati nel trattamento di anemie, cachessia (sindrome da deperimento), osteoporosi, cancro al seno e per altre indicazioni mediche.
Il Nandrolone è stato sintetizzato per la prima volta nel 1950. È stato introdotto per la prima volta nel mercato farmaceutico, come Nandrolone Fenilpropionato, nel 1959, e poi come Nandrolone Decanoato nel 1962, seguito da ulteriori esteri.
Il Nandrolone ha una bassa affinità di interazione con l’Enzima Aromatasi convertendo in Estrone, un estrogeno molto meno potente dell’Estradiolo, circa 10 volte meno attivo, e, come tale, è un estrogeno relativamente debole. In una condizione di somministrazione del Nandrolone senza una base di Testosterone, i livelli di Estradiolo calerebbero marcatamente a favore di un aumento del Estrone il quale non potrebbe però sostituire nelle diverse attività tissutali il prima citato E2. Le conseguenze negative si verificherebbero dall’attività sessuale all’attività neurosteroidea.
Infatti, un effetto da non sottovalutare con l’uso di Nandrolone è il suo impatto sul SNC. L’impatto del Nandrolone sul Sistema Nervoso Centrale è stato osservato scientificamente. Nello studio intitolato “The Impact of Nandrolone Decanoate on the Central Nervous System” vengono descritti chiaramente i numerosi effetti psicologici di questa molecola. Essi comprendono e influenzano:
1- Aggressività 2- Ansia, paura e stress 3- Ricompensa e dipendenza 4- Apprendimento, memoria e capacità di lavoro 5- Locomozione e attività fisica 6- Effetti sulla HPAA (Asse Ipotalamo-Pituitaria-Surrene) 7- Effetto sui neurotrasmettitori: Recettore Acido γ-Aminobutirrico Tipo A (GABAA); Recettori 5-idrossitriptamina (5-HT) e 5-HT; Recettori della Dopamina e Recettori Oppioidi.
Questo effetto, unito alla modesta potenzialità anabolizzante se confrontata con altre molecole anche della stessa famiglia, fa pendere l’ago della bilancia verso gli svantaggi d’uso piuttosto che i vantaggi. Sebbene vi sia un rapporto tra Testosterone e Nandrolone finalizzato a ridurre la comparsa di questi effetti avversi (ratio T:N = 2:1) su un buon numero di soggetti risulta dare comunque problemi rilevanti.
Il suo uso principale in Off-Season comprende dosaggi medi tra i 200mg ed i 400mg a settimana, con un adeguato rapporto con il Testosterone. Se utilizzato a fini di recupero articolare viene usato a dosaggi di 100mg a settimana, e con tali dosaggi difficilmente emergono i problemi sopra elencati a patto che ci sia una base di Testosterone.
Il Drostanolone, noto anche come 2α-metil-5α-diidrotestosterone (2α-metil-DHT) o come 2α-metil-5α-androstan-17β-ol-3-one, è uno steroide androstano sintetico e un derivato del DHT. Si tratta nello specifico di DHT con un gruppo metile in posizione C2α. La forma esterificata Drostanolone Propionato è stata usata in passato nel trattamento del cancro al seno nelle donne per via della sua attività antiestrogenica. Questa azione il Drostanolone la esplica sia agendo come antagonista del recettore degli estrogeni e sia come inibitore dell’Enzima Aromatasi. Ed è proprio per questo motivo che una molecola generalmente relegata all’uso in “Cut” o pre-gara trova un suo uso funzionale in Off-Season. La sua attività AI è comunque moderata ma sufficiente in un buon numero di soggetti per evitare l’aggiunta di SERM e/o AI di altro genere. L’attività AI moderata sembra non incidere negativamente in modo sensibile sull’Asse GH/IGF1.
L’effetto miotrofico risulta simile a quello osservato con il Methenolone, in generale moderatamente inferiore al Testosterone. I dosaggi utilizzati in Off-Season per il controllo estrogenico sono nel range dei 200-400mg a settimana (diviso in due iniezioni distanziate da 4-5 giorni) per l’estere Enantato, mentre per il Propionato 150-350mg a settimana (dosi a giorni alterni o giornaliere).
Il Trenbolone, noto anche come 19-nor-δ9,11-testosterone o come estra-4,9,11-trien-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone) sintetizzato per la prima volta nel 1963. Si tratta nello specifico di Nandrolone con due doppi legami aggiuntivi nel nucleo steroideo. Gli esteri del Trenbolone, che hanno un estere in posizione C17β, includono il Trenbolone Acetato, il Trenbolone Enantato, Il Trenbolone Hexahydrobenzylcarbonato e il Trenbolone Undecanoato. Il Trenbolone Acetato (marchi Finajet, Finaplix, e altri) e il Trenbolone Hexahydrobenzylcarbonato (marchi Parabolan, Hexabolan), sono o sono stati commercializzati per uso veterinario e clinico nell’uomo. Il Trenbolone Acetato è utilizzato in medicina veterinaria nel bestiame per aumentare la crescita muscolare e l’appetito degli animali, mentre il Trenbolone Hexahydrobenzylcarbonato è stato utilizzato in passato a livello clinico nell’uomo, ma ora non è più commercializzato.
Si tratta di uno degli AAS più versatili in assoluto, con un ottima resa tanto in preparazione alla gara quanto in Off-Season. L’enorme potenziale anabolizzante del Trenbolone, così come dei suoi analoghi, è stato riportato fin dagli anni ’60. La sua diffusione nel Bodybuilding è iniziata circa negli anni ’80 del secolo scorso. La sua elevata potenzialità miotrofica, lipolitica e di spinta mentale lo resero in poco tempo estremamente popolare tra i culturisti.
In Off-Season viene utilizzato nelle sue forme eseterificate Enantato e Hexahydrobenzylcarbonato a dosaggi nell’ordine dei 100-400mg a settimana (divisa in due somministrazioni distanziate l’una dall’altra da 4-5 giorni), sebbene il trend d’oltre oceano è arrivato a dosaggi decisamente eccessivi e nell’ordine del grammo. Per l’esetere Acetato i dosaggi medi vanno da 150mg a 350mg a settimana con dosaggi a giorni alterni o giornalieri.
E’ necessario ricordare ai lettori che gli effetti collaterali a livello del SNC possono verificarsi in alcuni punti come nel caso del Nandrolone sebbene i vantaggi rendano il Trenbolone più bilanciato tra sides e vantaggi.
Il Trestolone, noto anche come 7α-metil-19-nortestosterone (MENT) o come 7α-metilestr-4-en-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone). È una forma modificata del Nandrolone con un gruppo metile in posizione C7α. Tra gli AAS strettamente correlati vi sono il 7α-metil-19-norandrostenedione (MENT dione, trestione), un pro-ormone androgeno del Trestolone, e il Dimetandrolone (7α, 11β-dimetil-19-nortestosterone), il derivato metilato C11β del Trestolone, nonché il Mibolerone (7α,17α-dimetil-19-nortestosterone) e il Dimetiltrienolone (7α,17α-dimetil-δ9,11-19-nortestosterone). Anche il progestinico Tibolone (7α-metil-17α-etinil-δ5(10)-19-nortestosterone) è strettamente correlato al Trestolone.
Il Trestolone è stato descritto per la prima volta nel 1963. Tuttavia, non è stato successivamente studiato fino al 1990. Lo sviluppo del Trestolone per un potenziale uso nella contraccezione ormonale maschile e nella terapia sostitutiva degli androgeni è stato avviato nel 1993 ed è proseguito in seguito. Non sembra che siano stati condotti ulteriori sviluppi dal 2013. Il Trestolone è stato sviluppato dal Population Council, un’organizzazione non governativa senza scopo di lucro dedicata alla salute riproduttiva.
Come AAS, il Trestolone è un agonista del recettore degli androgeni (AR), analogamente agli androgeni come il Testosterone e il Diidrotestosterone (DHT). Questo AAS presenta spiccate proprietà anticortisolemiche sia attraverso l’inibizione enzimatica sia per attività antagonista recettoriale. Il Trestolone non è un substrato per la 5α-reduttasi e quindi non è potenziato o inattivato nei cosiddetti tessuti “androgeni” come la pelle, i follicoli piliferi e la ghiandola prostatica. Come tale, ha un elevato rapporto tra attività anabolica e androgena, analogamente ad altri derivati del Nandrolone. Il Trestolone è un substrato per l’Aromatasi e quindi produce come metabolita l’estrogeno 7α-metilestradiolo. Tuttavia, il Trestolone ha solo una debole attività estrogenica e una quantità che sembrerebbe essere insufficiente per scopi terapici sostitutivi, come evidenziato dalla diminuzione della densità minerale ossea negli uomini trattati con esso per l’ipogonadismo.
Il potenziale anabolizzante del Trestolone ha mostrato un grado di superiorità miotrofica rispetto al Trenbolone. Le sue caratteristiche ne fanno prediligere l’uso in Off-Season/Bulk. I dosaggi utilizzati con la forma Acetato sono nell’ordine dei 150-350mg a settimana con una cadenza nelle somministrazioni a giorni alterni. Sebbene sia più rara da reperire, la forma Enantato è utilizzato nel range dei 200-400mg a settimana divisi in somministrazioni ogni 4-5 giorni.
L’Oxymetholone, noto anche come 2-idrossimetilene-17α-metil-4,5α-diidrotestosterone (2-idrossimetilene-17α-metil-DHT) o come 2-idrossimetilene-17α-metil-5α-androstan-17β-olo-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del DHT. L’Oxymetholone è stato descritto per la prima volta in un articolo del 1959 da scienziati della Syntex. È stato introdotto per uso medico dalla Syntex e dalla Imperial Chemical Industries nel Regno Unito con il marchio Anapolon nel 1961. L’Oxymetholone è stato introdotto anche con i marchi Adroyd (Parke-Davis) nel 1961 e Anadrol (Syntex) nel 1962. Il farmaco è stato commercializzato negli Stati Uniti nei primi anni ’60.
Come altri AAS, l’Oxymetholone è un agonista del recettore degli androgeni (AR). Non è un substrato per la 5α-reduttasi (dal momento che è già 5α-ridotto) ed è uno substrato scarso per il 3α-idrossisteroide deidrogenasi (3α-HSD), e quindi mostra un alto rapporto di attività anabolizzante rispetto all’effetto androgenico.
Data la sua derivanza dal DHT, l’Oxymetholone non è un substrato per l’Enzima Aromatasi e quindi non può essere aromatizzato in metaboliti estrogenici. Tuttavia, caratteristica unica tra i derivati del DHT, l’Oxymetholone è comunque associato a un’estrogenicità relativamente elevata ed è noto per avere il potenziale di produrre effetti collaterali estrogenici come ginecomastia (anche se non comune) e ritenzione idrica. È stato suggerito che questo può essere una conseguenza del legame diretto a l’attivazione del recettore degli estrogeni da parte dell’Oxymetholone (estrogenicità intrinseca). L’Oxymetholone non possiede alcuna attività progestinica significativa. Per via della caratteristica attività estrogenica intrinseca, con l’uso di Oxymetholone è spesso necessario l’uso di un SERM onde avere un controllo sulla aumentata attività estrogenica.
A causa della sua struttura 17α-alchilata, l’Oxymetholone è epatotossico. L’uso a lungo termine del farmaco può causare una varietà di disturbi gravi, tra cui l’epatite, il cancro al fegato e la cirrosi; pertanto si raccomandano test periodici di funzionalità epatica per coloro che assumono l’Oxymetholone a fini terapeutici. Questa molecola ha ottenuto, infatti, la nomea di essere uno tra gli AAS più epatotossici. Ciò deriva da i dosaggi comunemente, ed erroneamente, utilizzati in contesto culturistico. Si parla di dosaggi che facilmente sforano i 150mg/die.
Osservazioni e esaminazione di diversi referti di esami ematici hanno evidenziato una soglia di “vantaggio/svantaggio” a favore del primo con un dosaggio calcolato con la formula 1mg/Kg. Genericamente, però, il dosaggio standard e conservativo si attesta nel range dei 50-100mg/die per non più di 28 giorni consecutivi, al fine di ridurre l’impatto negativo sul fegato e lipidemia.
Il Methandrostenolone, noto anche come 17α-metil-δ1-testosterone o come 17α-metilandrost-1,4-dien-17β-ol-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del Testosterone. È una modifica del Testosterone con un gruppo metile in posizione C17α e un doppio legame aggiuntivo tra le posizioni C1 e C2. Il farmaco è anche il derivato 17α-metilato del Boldenone (δ1-testosterone) e l’analogo δ1 del Methyltestosterone (17α-metiltestosterone).
Il Methandrostenolone è stato descritto per la prima volta nel 1955. È stato sintetizzato dai ricercatori dei laboratori CIBA di Basilea, in Svizzera. La CIBA depositò un brevetto statunitense nel 1957 e iniziò a commercializzare il farmaco sotto il nome di Dianabol nel 1958 negli Stati Uniti. Inizialmente veniva prescritto alle vittime di ustioni e agli anziani. Tra i primi utilizzatori vi furono i giocatori dell’Oklahoma University e l’allenatore dei San Diego Chargers Sid Gillman, che somministrò il Dianabol alla sua squadra a partire dal 1963.
Anche se il primo a somministrare il Methandrostenolone agli atleti fu il Dr. John Ziegler, personaggio che ebbe non poca importanza nella storia dell’uso degli AAS negli Stati Uniti. Ziegler contribuì a facilitare l’adozione degli AAS in generale, e del Dianabol in particolare, da parte degli atleti americani. Ziegler fu la prima persona a somministrare il Dianabol agli atleti competitivi poco dopo la sua introduzione da parte della CIBA nel 1958. Ebbe accesso al laboratorio CIBA a Summit (New Jersey) nel corso degli anni 50’ e somministrava già ai pesisti il Testosterone Propionato per “scopi di ricerca”. Da li il passo fu breve per la diffusione a macchia d’olio di questo AAS tra i culturisti.
Data la sua principale modifica strutturale, ossia la metilazione in C-17, il Methandrostenolone mostra un aumentata stabilità del legame recettoriale aumentando così l’affinità sia al AR sia, successivamente all’aromatizzazione nel suo metabolita 17-Methylestradiolo, per i recettori estrogenici rendendo il composto molto più estrogenico del Testosterone. Tale caratteristiche migliora però il potenziale proliferativo dei AR e l’influenza positiva sulla sintesi di IGF-1. Da non dimenticare è il suo significativo impatto anticortisolemico.
Trattandosi di una molecola con una discreta tendenza all’aromatizzazione, il suo uso tipico la vede inserita nelle fasi Off-Season. Il calcolo del dosaggio, per via dati aneddotici e osservativi raccolti, lo si ottiene con la formula 5mg/12Kg di peso corporeo. Trattandosi di un composto orale metilato in C-17 se ne scoraggia l’utilizzo oltre i 28 giorni consecutivi onde ridurre l’impatto negativo su fegato e lipidemia. Data la sua emivita di circa 4h, il dosaggio giornaliero dovrebbe essere diviso in più assunzioni distribuite durante l’arco della giornata.
L’Ormone della Crescita (GH) o Somatotropina, noto anche come Ormone della Crescita Umano (hGH o HGH), è un ormone peptidico che stimola la crescita, la riproduzione e la rigenerazione cellulare nell’uomo e in altri animali. È quindi importante per lo sviluppo umano. Il GH stimola anche la produzione di IGF-1 e aumenta la concentrazione di glucosio e acidi grassi liberi nel sangue. È un tipo di mitogeno specifico solo per i recettori di alcuni tipi di cellule. Il GH è un polipeptide a catena singola di 191 aminoacidi che viene sintetizzato, immagazzinato e secreto dalle cellule somatotrope nelle ali laterali dell’ipofisi anteriore.
Una forma ricombinante di hGH, chiamata Somatropina, viene utilizzata come farmaco da prescrizione per il trattamento dei disturbi della crescita nei bambini e della carenza di Ormone della Crescita negli adulti. Molte delle funzioni dell’hGH rimangono sconosciute.
Nel suo ruolo di agente anabolizzante, l’hGH è stato utilizzato dagli sportivi agonisti almeno dal 1982, quando la sola forma disponibile era quella derivata dall’Ipofisi dei cadaveri, ed è stato vietato dal CIO e dall’NCAA. L’analisi tradizionale delle urine non è in grado di rilevare il doping con HGH, pertanto il divieto è stato applicato solo all’inizio degli anni 2000, quando sono stati sviluppati test del sangue in grado di distinguere tra hGH naturale e artificiale.
In ambiente bodybuilding, l’hGH viene utilizzato in Off-Season (dai soggetti meglio informati) a dosaggi nel range delle 4-8UI al giorno o 8-16UI a giorni alterni. La somministrazione in concomitanza con l’uso di Insulina ha mostrato effetti sinergici molto evidenti che trovano la loro origine nel miglioramento della sintesi di IGF-1 e della sua frazione libera quindi attiva. Ricordo inoltre che l’uso di hGH può causare una sottoregolazione della funzionalità tiroidea per via del feedback negativo causato da un aumento della conversione del T4 in T3 per azione del GH. L’uso di T4, nel periodo d’uso in Off-Season, è in alcuni casi una necessità.
Il Fattore di Crescita Insulino-Simile 1 (IGF-1), chiamato anche Somatomedina C, è un ormone dalla struttura molecolare simile a quella dell’insulina che svolge un ruolo importante nella crescita infantile e ha effetti anabolici negli adulti. L’IGF-1 è costituito da 70 aminoacidi in una singola catena con tre ponti disolfuro intramolecolari.
L’IGF-1 è prodotto principalmente dal fegato. La produzione è stimolata dall’Ormone della Crescita (GH). La maggior parte dell’IGF-1 è legata a una delle 6 proteine di legame (IGF-BP). L’IGFBP-1 è regolato dall’Insulina. L’IGF-1 viene prodotto durante tutta la vita; i tassi più alti di produzione di IGF-1 si verificano durante la crescita puberale. I livelli più bassi si verificano nell’infanzia e nella vecchiaia.
L’IGF-1 lega e attiva il proprio recettore, l’IGF-1R, attraverso l’espressione sulla superficie cellulare delle tirosin-chinasi recettoriali (RTK) e segnala ulteriormente attraverso molteplici cascate di trasduzione intracellulare. L’IGF-1R è l’induttore che svolge un ruolo critico nella modulazione degli effetti metabolici dell’IGF-1 per la senescenza e la sopravvivenza cellulare. L’IGF-1 è responsabile di stimolare la crescita di tutti i tipi di cellule e di provocare effetti metabolici significativi. Un importante effetto metabolico dell’IGF-1 è la sua capacità di segnalare alle cellule che sono disponibili nutrienti sufficienti per l’ipertrofia e la divisione cellulare. Questi segnali consentono inoltre all’IGF-1 di inibire l’apoptosi cellulare e di aumentare la produzione di proteine cellulari. I recettori dell’IGF-1 sono ubiquitari, il che consente che i cambiamenti metabolici causati dall’IGF-1 si verifichino in tutti i tipi di cellule. Gli effetti metabolici dell’IGF-1 sono di vasta portata e possono coordinare il metabolismo delle proteine, dei carboidrati e dei grassi in una varietà di tipi di cellule diverse. La regolazione degli effetti metabolici dell’IGF-1 sui tessuti bersaglio è coordinata anche con altri ormoni, come l’Ormone della Crescita e l’Insulina.
L’IGF-1 da DNA ricombinante è disponibile principalmente in due diversi formati/varianti, lr3 e DES. È importante ricordare che, a prescindere dalla variante, tutti funzionano a livello sistemico nell’organismo e che, nonostante la somministrazione dell’ormone per via intramuscolare direttamente in un muscolo specifico, non genererà una crescita localizzata misurabile.
Ovviamente tralascerò di descrivere l’IGF-1 bioidentico commercializzato come Mecasermina dal momento che la sua farmacocinetica è identica a quella del IGF-1 endogeno. Dirò soltanto che mediamente viene utilizzato in dosi giornaliere nel range tra 60-1.000mcg post-workout. L’emivita di questa forma di IGF-1 è di circa 5.8h.
IGF-1 LR3: Questa forma è la variante di IGF-1 più comune e molto popolare sul mercato e utilizzata da bodybuilder e atleti di altre discipline. Contiene IGF-1 bioidentico costituito dalla catena originale di 70 aminoacidi, ma con 13 aminoacidi in più all’estremità N, per un totale di 83 aminoacidi. Possiede anche una seconda modifica, in cui un’Arginina si trova in 3a posizione invece dell’Acido Glutammico originale. Il risultato di queste modifiche è che l’IGF-1 continua a svolgere la sua attività originaria sul recettore dell’IGF-1 nei tessuti corporei e ha un’affinità di legame molto bassa per le proteine leganti l’IGF menzionate in precedenza. Inoltre, presenta una vita attiva significativamente più lunga, di circa 20-30 ore, rispetto a quella dell’IGF-1 di 12-15 ore. L’insieme di questi fattori ha dimostrato che l’LR3 ha un’efficacia circa tre volte superiore a quella dell’IGF-1.
I dosaggi medi utilizzati per questa forma sono nel range dei 40-80mcg/die. A causa della sua lunga vita attiva nell’organismo, la variante LR3 non dovrebbe essere somministrata più di una volta al giorno per il semplice fatto che non risulta necessario. Nei giorni di allenamento, il dosaggio di IGF-1 è solitamente somministrato subito dopo l’allenamento. La scelta è a discrezione dell’utilizzatore, in quanto può essere benissimo somministrato sia prima che dopo (solo prima dell’allenamento o solo dopo l’allenamento). E’ possibile comunque dividere il dosaggio giornaliero in due somministrazioni nell’arco della giornata, il dosaggio giornaliero completo può essere diviso quindi a metà tra i due (ad esempio, 20mcg prima dell’allenamento e 20mcg dopo l’allenamento, per un totale di 40mcg al giorno). Nei giorni di non allenamento, può essere somministrato in qualsiasi momento della giornata.
IGF-1 DES: Conosciuto anche come DES(1-3)IGF-1, questa è la forma di IGF-1 comunemente conosciuta come ad azione molto rapida e di solito è la meno preferita tra le due. Le sue modifiche rispetto alla molecola originale di IGF-1 sono tali da farle mancare i primi 3 aminoacidi all’N terminale, il che conferisce all’IGF-1 DES un totale di 67 aminoacidi nella sua catena rispetto ai 70 originali. Questa modifica garantisce all’IGF-1 DES una ridotta affinità di legame per le proteine leganti l’IGF menzionate in precedenza, oltre a una maggiore forza di legame e potenziale miotrofico, circa dieci volte superiore a quella dell’IGF-1 originale e cinque volte superiore a quella dell’IGF-1 LR3. A differenza dell’IGF-1 LR3, l’IGF-1 DES ha un’emivita molto più breve, di circa 20-30 minuti. Grazie alla sua attività più rapida e alla maggiore forza/potenza, la variante DES dell’IGF-1 è comunemente ritenuta in grado di ottenere una crescita muscolare localizzata nel sito in cui viene iniettata. Sebbene ciò sia in parte vero, gli studi hanno dimostrato che, come l’IGF-1 in generale, agisce a livello sistemico una volta raggiunti i capillari e il flusso sanguigno. Quindi l’effetto localizzato è minimo e non significativamente differente dall’effetto sistemico.
Il dosaggio della variante DES è leggermente più variabile rispetto a quello del LR3. Per l’IGF-1 DES, il dosaggio varia da 50 a 150 mcg al giorno. A causa della sua emivita molto più breve rispetto alla variante LR3, è possibile utilizzare dosaggi più elevati con una ipotetica riduzione del rischio di effetti a lungo termine sull’organismo, anche se è necessario usare comunque cautela. Può essere utilizzato nello stesso modo dell’IGF-1 LR3 post-workout, ed è infatti comunemente usato in questo modo a causa della sua breve emivita.
Entrambe le forme di IGF-1 possono essere somministrate per via intramuscolare o sottocutanea. L’uso di una delle due forme non deve superare la durata di 30 giorni prima di una pausa di almeno 2 settimane, anche se fare pause più lunghe di 2 settimane tra un ciclo di IGF-1 e l’altro è l’opzione migliore. Questo non solo per ridurre il rischio di effetti sulla salute a lungo termine, ma anche per garantire che i recettori dell’IGF-1 tornino ad un grado di sensibilità ottimale e, quindi, a “rispondere” correttamente dopo un ciclo.
L’insulina è un ormone peptidico prodotto dalle cellule beta delle isole pancreatiche. Regola il metabolismo dei carboidrati, dei grassi e delle proteine promuovendo l’assorbimento del glucosio dal sangue nelle cellule del fegato, dei grassi e dei muscoli scheletrici. In questi tessuti il glucosio assorbito viene convertito in glicogeno attraverso la glicogenesi o in grassi (trigliceridi) attraverso la lipogenesi o, nel caso del fegato, in entrambi. La produzione e la secrezione di glucosio da parte del fegato sono fortemente inibite da alte concentrazioni di Insulina nel sangue. L’Insulina circolante influisce anche sulla sintesi di proteine in un’ampia varietà di tessuti. È quindi un ormone anabolico e anticatabolico, che promuove la conversione di piccole molecole nel sangue in grandi molecole all’interno delle cellule. Bassi livelli di Insulina nel sangue hanno l’effetto opposto, favorendo un diffuso catabolismo, soprattutto del grasso corporeo di riserva.
La maggior parte dei bodybuilder utilizza una sola forma di Insulina (ad azione rapida o ultra-rapida), anche se alcuni utilizzano anche un’Insulina a lunga durata d’azione o in monoterapia insulinica o in conbinazione con le forme ad azione rapida o ultra-rapida.
L’Humalog® (Insulina Lispro) è senza dubbio la forma di Insulina più diffusa tra i bodybuilder insieme all’Humulin-R. L’Humalog è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”. Proprio al fine di potenziare la “finestra anabolica”, l’Humalog viene usata in concomitanza del GH il quale viene somministrato in una tempistica tale che i due picchi di rilascio (curva ematica massima) si “incrocino” andando a creare un affetto additivo di potenziamento della sintesi epatica di IGF-1 e della sua attività per via della riduzione dei trasportatori IGFBP.
Tuttavia, l’uso di una base insulinica composta da Insuline Glargine (Lantus), con una vita attiva di 24-26.5h, la quale sembra avere effetti di maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi, viene da alcuni inserita nei protocolli Off-Season.
I dosaggi di Insulina non andrebbero calcolati in modo distaccato dal piano alimentare e dal suo contenuto glucidico. Se il margine di “sicurezza” indica un assunzione di 10-15g di Carboidrati per UI di Insulina, questi non dovrebbero essere addizionati al piano alimentare già tarata in surplus calorico. Il calcolo delle unità dovrebbe essere tarato sul quantitativo glucidico della dieta e sul rapporto con il peso corporeo dell’atleta. Facciamo un semplice esempio: Soggetto di 90Kg = formula 1UI ogni 10Kg di peso = 9UI massime somministrabili per pasto e in base alla vita attiva della forma utilizzata = assicurarsi che il pasto appena successivo alla somministrazione dell’Insulina a questo dosaggio sia pari o superiore ai 90g di Carboidrati. Il monitoraggio della glicemia attraverso un glucometro è ovviamente d’obbligo in un protocollo di Insulina.
Nota: tali informazioni esposte non rappresentano in nessun modo un parere medico ne tanto meno una prescrizione e/o incentivo all’uso di sostanze dopanti e illegali. Le descrizioni presentate per i PEDs solitamente più utilizzati in Off-Season sono sintetiche sia per motivi di “Off Topic” sia per ragioni legate alla loro descrizione approfondita in altri articoli presenti nel database di questo sito. In queste pubblicazioni potrete trovare informazioni inerenti anche agli affetti collaterali connessi ad un uso/abuso “off-label” dei diversi PEDs.
Conclusioni:
Per concludere e fare una sintesi delle nozioni esposte in questo articolo, dobbiamo ricordarci che i bodybuilder in Off-Season dovrebbero concentrarsi sul consumo di una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) con l’obiettivo di guadagnare ~0,25-0,5% del peso corporeo a settimana per un “Natural”, mentre nel caso di un “Doped” la soglia può spostarsi tra l’1-2% con variabili connesse a risposte genetiche differenziali e anzianità nella carriera culturistica (principiante, intermedio e avanzato). In ogni caso, in una fetta maggioritaria di praticanti, ai bodybuilder avanzati si consiglia di essere più prudenti con il surplus calorico e il tasso di aumento di peso settimanale. L’assunzione di proteine nella dieta è raccomandata a 1,6-2,2 g/kg/giorno, con particolare attenzione a una quantità sufficiente di proteine a ogni pasto (0,40-0,55 g/kg/pasto) e a una distribuzione uniforme nell’arco della giornata (3-6 pasti). Per i “Doped”, in alcuni casi, l’introito proteico può essere portato, con minimi vantaggi in contesto ipercalorico, a 2,5g/Kg con le medesime linee guida di suddivisione per numero di pasti. I grassi alimentari devono essere consumati a livelli moderati, né troppo bassi né troppo alti (0,5-1,5 g/kg/die), per evitare un rapporto fTC sfavorevole e per prevenire riduzioni dei livelli di testosterone. Nei “Doped” l’obbiettivo con i lipidi è principalmente quello di assumerne una dose necessaria, e altamente qualitativa, al fine di assimilare vitamine liposolubili, per substrato strutturale, per sintesi di eicosanoidi (vedi assunzione EPA, DHA e AA), protezioni epidermide e capelli; di conseguenza attenersi ad un dosaggio medio pari a 35-50g/die. Dopo che le calorie sono state distribuite tra Proteine e Grassi, le restanti calorie dovrebbero provenire dai Carboidrati, assicurandosi di consumarne una quantità sufficiente (≥3-5 g/kg/giorno). Si possono ottenere benefici maggiori consumando proteine (0,40-0,55 g/kg/pasto) in prossimità delle sessioni di allenamento (1-2 ore prima dell’esercizio ed entro 1-2 ore dopo l’esercizio). È opportuno prendere in considerazione la Creatina Monoidrato (3-5 g/giorno) e la Caffeina (5-6 mg/kg), in quanto possono produrre effetti ergogenici per i bodybuilder. Inoltre, Beta-Alanina (3-5 g/die) e Citrullina Malato (8 g/die) sono integratori alimentari che possono essere presi in considerazione in quanto potenzialmente utili per i bodybuilder, a seconda dei regimi di allenamento individuali. I bodybuilder che non sono in grado di assumere un apporto sufficiente di micronutrienti e acidi grassi essenziali nella loro dieta dovrebbero prendere in considerazione l’integrazione di questi nutrienti per evitare carenze. Il limite principale di questo articolo è la mancanza di studi su larga scala e a lungo termine sui bodybuilder durante la Off-Season. Sono necessarie ulteriori ricerche su questa popolazione per ottimizzare la nutrizione e le raccomandazioni sugli integratori alimentari.
4. Philen R.M., Ortiz D.I., Auerbach S.B., Falk H. Survey of Advertising for Nutritional Supplements in Health and Bodybuilding Magazines. JAMA. 1992;268:1008. doi: 10.1001/jama.1992.03490080082029. [PubMed] [CrossRef] [Google Scholar]
5. Giampreti A., Lonati D., Locatelli C., Rocchi L., Campailla M.T. Acute neurotoxicity after yohimbine ingestion by a bodybuilder. [(accessed on 25 March 2019)];Clin. Toxicol. 2009 47:827–829. doi: 10.1080/15563650903081601. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19640235 [PubMed] [CrossRef] [Google Scholar]
7. Della Guardia L., Cavallaro M., Cena H. The risks of self-made diets: The case of an amateur bodybuilder. J. Int. Soc. Sports Nutr. 2015;12:5. doi: 10.1186/s12970-015-0077-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Hackett D.A., Johnson N.A., Chow C.-M. Training Practices and Ergogenic Aids Used by Male Bodybuilders. J. Strength Cond. Res. 2013;27:1609–1617. doi: 10.1519/JSC.0b013e318271272a. [PubMed] [CrossRef] [Google Scholar]
10. Forbes G.B., Brown M.R., Welle S.L., Lipinski B.A. Deliberate overfeeding in women and men: Energy cost and composition of the weight gain. Br. J. Nutr. 1986;56:1–9. doi: 10.1079/BJN19860080. [PubMed] [CrossRef] [Google Scholar]
11. Kreider R.B., Klesges R., Harmon K., Ramsey L., Bullen D., Wood L., Almada A., Grindstaff P., Li Y. Effects of Ingesting Supplements Designed to Promote Lean Tissue Accretion on Body Composition during Resistance Training. Int. J. Sport Nutr. 1996;6:234–246. doi: 10.1123/ijsn.6.3.234. [PubMed] [CrossRef] [Google Scholar]
12. Rozenek R., Ward P., Long S., Garhammer J. Effects of high-calorie supplements on body composition and muscular strength following resistance training. J. Sports Med. Phys. Fit. 2002;42:340–347. [PubMed] [Google Scholar]
13. Garthe I., Raastad T., Refsnes P.E., Sundgot-Borgen J. Effect of nutritional intervention on body composition and performance in elite athletes. Eur. J. Sport Sci. 2013;13:295–303. doi: 10.1080/17461391.2011.643923. [PubMed] [CrossRef] [Google Scholar]
14. American College og Sports Medicine American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. [(accessed on 25 March 2019)];Med. Sci. Sport. Exerc. 2009 41:687–708. doi: 10.1249/MSS.0b013e3181915670. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19204579 [PubMed] [CrossRef] [Google Scholar]
15. Lambert C.P., Frank L.L., Evans W.J., Lambert D.C.P. Macronutrient Considerations for the Sport of Bodybuilding. Sports Med. 2004;34:317–327. doi: 10.2165/00007256-200434050-00004. [PubMed] [CrossRef] [Google Scholar]
16. Walberg-Rankin J., Edmonds C.E., Gwazdauskas F.C. Diet and Weight Changes of Female Bodybuilders Before and After Competition. Int. J. Sport Nutr. 1993;3:87–102. doi: 10.1123/ijsn.3.1.87. [PubMed] [CrossRef] [Google Scholar]
17. Lamar-Hildebrand N., Saldanha L., Endres J. Dietary and exercise practices of college-aged female bodybuilders. J. Am. Diet. Assoc. 1989;89:1308–1310. [PubMed] [Google Scholar]
18. Houston M.E. Gaining Weight: The Scientific Basis of Increasing Skeletal Muscle Mass. Can. J. Appl. Physiol. 1999;24:305–316. doi: 10.1139/h99-024. [PubMed] [CrossRef] [Google Scholar]
19. Phillips S.M. A Brief Review of Critical Processes in Exercise-Induced Muscular Hypertrophy. Sports Med. 2014;44:71–77. doi: 10.1007/s40279-014-0152-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Campbell B.I., Aguilar D., Conlin L., Vargas A., Schoenfeld B.J., Corson A., Gai C., Best S., Galvan E., Couvillion K. Effects of High Versus Low Protein Intake on Body Composition and Maximal Strength in Aspiring Female Physique Athletes Engaging in an 8-Week Resistance Training Program. Int. J. Sport Nutr. Exerc. Metab. 2018;28:580–585. doi: 10.1123/ijsnem.2017-0389. [PubMed] [CrossRef] [Google Scholar]
22. Morton R.W., Murphy K.T., McKellar S.E., Schoenfeld B.J., Henselmans M., Helms E., Aragon A.A., Devries M.C., Banfield L., Krieger J.W., et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. [(accessed on 25 March 2019)];Br. J. Sports Med. 2018 52:376–384. doi: 10.1136/bjsports-2017-097608. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28698222 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Houltham S.D., Rowlands D.S. A snapshot of nitrogen balance in endurance-trained women. Appl. Physiol. Nutr. Metab. 2014;39:219–225. doi: 10.1139/apnm-2013-0182. [PubMed] [CrossRef] [Google Scholar]
25. Antonio J., Ellerbroek A., Silver T., Vargas L., Peacock C. The effects of a high protein diet on indices of health and body composition—A crossover trial in resistance-trained men. J. Int. Soc. Sports Nutr. 2016;13:8. doi: 10.1186/s12970-016-0114-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Bandegan A., Courtney-Martin G., Rafii M., Pencharz P.B., Lemon P.W. Indicator Amino Acid–Derived Estimate of Dietary Protein Requirement for Male Bodybuilders on a Nontraining Day Is Several-Fold Greater than the Current Recommended Dietary Allowance. J. Nutr. 2017;147:850–857. doi: 10.3945/jn.116.236331. [PubMed] [CrossRef] [Google Scholar]
27. Malowany J.M., West D.W.D., Williamson E., Volterman K.A., Sawan S.A., Mazzulla M., Moore D.R. Protein to Maximize Whole-Body Anabolism in Resistance-trained Females after Exercise. Med. Sci. Sports Exerc. 2019;51:798–804. doi: 10.1249/MSS.0000000000001832. [PubMed] [CrossRef] [Google Scholar]
28. Antonio J., Peacock C.A., Ellerbroek A., Fromhoff B., Silver T. The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. J. Int. Soc. Sports Nutr. 2014;11:19. doi: 10.1186/1550-2783-11-19. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
29. Antonio J., Ellerbroek A., Silver T., Orris S., Scheiner M., Gonzalez A., Peacock C.A. A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women—A follow-up investigation. J. Int. Soc. Sports Nutr. 2015;12:39. doi: 10.1186/s12970-015-0100-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Bray G.A., Smith S.R., de Jonge L., Xie H., Rood J., Martin C.K., Most M., Brock C., Mancuso S., Redman L.M. Effect of dietary protein content on weight gain, energy expenditure, and body composition during overeating: A randomized controlled trial. [(accessed on 25 March 2019)];JAMA. 2012 307:47–55. doi: 10.1001/jama.2011.1918. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22215165 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Tipton K.D., Ferrando A.A., Phillips S.M., Doyle D., Wolfe R.R. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am. J. Physiol. Metab. 1999;276:628–634. doi: 10.1152/ajpendo.1999.276.4.E628. [PubMed] [CrossRef] [Google Scholar]
32. Rieu I., Balage M., Sornet C., Giraudet C., Pujos E., Grizard J., Mosoni L., Dardevet D. Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia. J. Physiol. 2006;575:305–315. doi: 10.1113/jphysiol.2006.110742. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Burd N.A., Tang J.E., Moore D.R., Phillips S.M. Exercise training and protein metabolism: Influences of contraction, protein intake, and sex-based differences. [(accessed on 25 March 2019)];J. Appl. Physiol. 2008 106:1692–1701. doi: 10.1152/japplphysiol.91351.2008. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19036897 [PubMed] [CrossRef] [Google Scholar]
34. Drummond M.J., Dreyer H.C., Fry C.S., Glynn E.L., Rasmussen B.B. Nutritional and contractile regulation of human skeletal muscle protein synthesis and mTORC1 signaling. J. Appl. Physiol. 2009;106:1374–1384. doi: 10.1152/japplphysiol.91397.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Tang J.E., Moore D.R., Kujbida G.W., Tarnopolsky M.A., Phillips S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009;107:987–992. doi: 10.1152/japplphysiol.00076.2009. [PubMed] [CrossRef] [Google Scholar]
36. Kanda A., Nakayama K., Sanbongi C., Nagata M., Ikegami S., Itoh H. Effects of Whey, Caseinate, or Milk Protein Ingestion on Muscle Protein Synthesis after Exercise. Nutrients. 2016;8:339. doi: 10.3390/nu8060339. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Messina M., Lynch H., Dickinson J.M., Reed K.E. No Difference Between the Effects of Supplementing With Soy Protein Versus Animal Protein on Gains in Muscle Mass and Strength in Response to Resistance Exercise. Int. J. Sport Nutr. Exerc. Metab. 2018;28:674–685. doi: 10.1123/ijsnem.2018-0071. [PubMed] [CrossRef] [Google Scholar]
38. Joy J.M., Lowery R.P., Wilson J.M., Purpura M., De Souza E.O., Mc Wilson S., Kalman D.S., Dudeck J.E., Jäger R. The effects of 8 weeks of whey or rice protein supplementation on body composition and exercise performance. Nutr. J. 2013;12:86. doi: 10.1186/1475-2891-12-86. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
39. Babault N., Paizis C., Deley G., Guérin-Deremaux L., Saniez M.-H., Lefranc-Millot C., Allaert F.A. Pea proteins oral supplementation promotes muscle thickness gains during resistance training: A double-blind, randomized, Placebo-controlled clinical trial vs. Whey protein. J. Int. Soc. Sports Nutr. 2015;12:1692. doi: 10.1186/s12970-014-0064-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Tesch P.A. Glycogen and triglyceride utilization in relation to muscle metabolic characteristics in men performing heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1990;61:5–10. [PubMed] [Google Scholar]
41. Lane A.R., Duke J.W., Hackney A.C. Influence of dietary carbohydrate intake on the free testosterone: Cortisol ratio responses to short-term intensive exercise training. [(accessed on 25 March 2019)];Eur. J. Appl. Physiol. 2010 108:1125–1131. doi: 10.1007/s00421-009-1220-5. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20091182 [PubMed] [CrossRef] [Google Scholar]
42. Tegelman R., Aberg T., Pousette A., Carlström K. Effects of a diet regimen on pituitary and steroid hormones in male ice hockey players. [(accessed on 25 March 2019)];Int. J. Sports Med. 1992 13:420–430. doi: 10.1055/s-2007-1021292. Available online: https://www.ncbi.nlm.nih.gov/pubmed/1387870 [PubMed] [CrossRef] [Google Scholar]
43. Dorgan J.F., Judd J.T., Longcope C., Brown C., Schatzkin A., Clevidence B.A., Campbell W.S., Nair P.P., Franz C., Kahle L., et al. Effects of dietary fat and fiber on plasma and urine androgens and estrogens in men: A controlled feeding study. Am. J. Clin. Nutr. 1996;64:850–855. doi: 10.1093/ajcn/64.6.850. [PubMed] [CrossRef] [Google Scholar]
44. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Decrease of serum total and free testosterone during a low-fat high-fibre diet. J. Steroid Biochem. 1983;18:369–370. doi: 10.1016/0022-4731(83)90117-6. [PubMed] [CrossRef] [Google Scholar]
45. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Diet and serum sex hormones in healthy men. J. Steroid Biochem. 1984;20:459–464. doi: 10.1016/0022-4731(84)90254-1. [PubMed] [CrossRef] [Google Scholar]
46. Wang C., Catlin D.H., Starcevic B., Heber D., Ambler C., Berman N., Lucas G., Leung A., Schramm K., Lee P.W.N., et al. Low-Fat High-Fiber Diet Decreased Serum and Urine Androgens in Men. J. Clin. Endocrinol. Metab. 2005;90:3550–3559. doi: 10.1210/jc.2004-1530. [PubMed] [CrossRef] [Google Scholar]
47. Morton R.W., Sato K., Gallaugher M.P.B., Oikawa S.Y., McNicholas P.D., Fujita S., Phillips S.M. Muscle Androgen Receptor Content but Not Systemic Hormones Is Associated With Resistance Training-Induced Skeletal Muscle Hypertrophy in Healthy, Young Men. Front. Physiol. 2018;9:9. doi: 10.3389/fphys.2018.01373. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Tinsley G.M., Willoughby D.S. Fat-Free Mass Changes During Ketogenic Diets and the Potential Role of Resistance Training. Int. J. Sport Nutr. Exerc. Metab. 2016;26:78–92. doi: 10.1123/ijsnem.2015-0070. [PubMed] [CrossRef] [Google Scholar]
49. Vargas S., Romance R., Petro J.L., Bonilla D.A., Galancho I., Espinar S., Kreider R.B., Benítez-Porres J. Efficacy of ketogenic diet on body composition during resistance training in trained men: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2018;15:31. doi: 10.1186/s12970-018-0236-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
50. Kephart W.C., Pledge C.D., Roberson P.A., Mumford P.W., Romero M.A., Mobley C.B., Martin J.S., Young K.C., Lowery R.P., Wilson J.M., et al. The Three-Month Effects of a Ketogenic Diet on Body Composition, Blood Parameters, and Performance Metrics in CrossFit Trainees: A Pilot Study. Sports. 2018;6:1. doi: 10.3390/sports6010001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
51. Greene D.A., Varley B.J., Hartwig T.B., Chapman P., Rigney M. A Low-Carbohydrate Ketogenic Diet Reduces Body Mass Without Compromising Performance in Powerlifting and Olympic Weightlifting Athletes. [(accessed on 26 March 2019)];J. Strength Cond. Res. 2018 32:3373–3382. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30335720 [PubMed] [Google Scholar]
52. Bird S. Strength Nutrition: Maximizing Your Anabolic Potential. Strength Cond. J. 2010;32:80–86. doi: 10.1519/SSC.0b013e3181d5284e. [CrossRef] [Google Scholar]
53. American Dietetic Association. Dietitians of Canada. American College of Sports Medicine. Rodriguez N.R., Di Marco N.M., Langley S. American College of Sports Medicine position stand. Nutrition and athletic performance. [(accessed on 26 March 2019)];Med. Sci. Sports Exerc. 2009 41:709–731. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19225360 [PubMed] [Google Scholar]
54. Chung S.T., Chacko S.K., Sunehag A.L., Haymond M.W. Measurements of Gluconeogenesis and Glycogenolysis: A Methodological Review. Diabetes. 2015;64:3996–4010. doi: 10.2337/db15-0640. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Azizi F. Effect of dietary composition on fasting-induced changes in serum thyroid hormones and thyrotropin. Metabolism. 1978;27:935–942. doi: 10.1016/0026-0495(78)90137-3. [PubMed] [CrossRef] [Google Scholar]
56. Mathieson R.A., Walberg J.L., Gwazdauskas F.C., Hinkle D.E., Gregg J.M. The effect of varying carbohydrate content of a very-low-caloric diet on resting metabolic rate and thyroid hormones. Metabolism. 1986;35:394–398. doi: 10.1016/0026-0495(86)90126-5. [PubMed] [CrossRef] [Google Scholar]
57. Leveritt M., Abernethy P.J. Effects of Carbohydrate Restriction on Strength Performance. J. Strength Cond. Res. 1999;13:52–57. [Google Scholar]
58. Jacobs I., Kaiser P., Tesch P. Muscle strength and fatigue after selective glycogen depletion in human skeletal muscle fibers. Graefe’s Arch. Clin. Exp. Ophthalmol. 1981;46:47–53. doi: 10.1007/BF00422176. [PubMed] [CrossRef] [Google Scholar]
59. Ray S., Sale D.G., Lee P., Garner S., MacDougall J.D., McCartney N. Muscle Substrate Utilization and Lactate Production During Weightlifting. Can. J. Appl. Physiol. 1999;24:209–215. [PubMed] [Google Scholar]
60. Tesch P.A., Colliander E.B., Kaiser P. Muscle metabolism during intense, heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1986;55:362–366. doi: 10.1007/BF00422734. [PubMed] [CrossRef] [Google Scholar]
63. Mitchell J.B., DiLauro P.C., Pizza F.X., Cavender D.L. The Effect of Preexercise Carbohydrate Status on Resistance Exercise Performance. Int. J. Sport Nutr. 1997;7:185–196. doi: 10.1123/ijsn.7.3.185. [PubMed] [CrossRef] [Google Scholar]
64. Lima-Silva A.E., Silva-Cavalcante M.D., Oliveira R.S., Kiss M.A., Pires F.O., Bertuzzi R., Bishop D. Effects of a low- or a high-carbohydrate diet on performance, energy system contribution, and metabolic responses during supramaximal exercise. Appl. Physiol. Nutr. Metab. 2013;38:928–934. doi: 10.1139/apnm-2012-0467. [PubMed] [CrossRef] [Google Scholar]
65. Vega F., Jackson R. Dietary habits of bodybuilders and other regular exercisers. Nutr. Res. 1996;16:3–10. doi: 10.1016/0271-5317(95)02054-3. [CrossRef] [Google Scholar]
66. Chappell A.J., Simper T., Barker M.E. Nutritional strategies of high level natural bodybuilders during competition preparation. J. Int. Soc. Sports Nutr. 2018;15:4. doi: 10.1186/s12970-018-0209-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
67. Atherton P.J., Etheridge T., Watt P.W., Wilkinson D., Selby A., Rankin D., Smith K., Rennie M.J. Muscle full effect after oral protein: Time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am. J. Clin. Nutr. 2010;92:1080–1088. doi: 10.3945/ajcn.2010.29819. [PubMed] [CrossRef] [Google Scholar]
68. Res P.T., Groen B., Pennings B., Beelen M., Wallis G.A., Gijsen A.P., Senden J.M., Van Loon L.J. Protein ingestion before sleep improves postexercise overnight recovery. [(accessed on 25 March 2019)];Med. Sci. Sports Exerc. 2012 44:1560–1569. doi: 10.1249/MSS.0b013e31824cc363. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22330017 [PubMed] [CrossRef] [Google Scholar]
69. Moore D.R., Robinson M.J., Fry J.L., Tang J.E., Glover E.I., Wilkinson S.B., Prior T., Tarnopolsky M.A., Phillips S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2009 89:161–168. doi: 10.3945/ajcn.2008.26401. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19056590 [PubMed] [CrossRef] [Google Scholar]
70. Witard O.C., Jackman S.R., Breen L., Smith K., Selby A., Tipton K.D. Muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2014 99:86–95. doi: 10.3945/ajcn.112.055517. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24257722 [PubMed] [CrossRef] [Google Scholar]
71. Macnaughton L.S., Wardle S.L., Witard O.C., McGlory C., Hamilton D.L., Jeromson S., Lawrence C.E., Wallis G.A., Tipton K.D. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol. Rep. 2016;4:e12893. doi: 10.14814/phy2.12893. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
72. Schoenfeld B.J., Aragon A.A., Krieger J.W. The effect of protein timing on muscle strength and hypertrophy: A meta-analysis. J. Int. Soc. Sports Nutr. 2013;10:53. doi: 10.1186/1550-2783-10-53. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Areta J.L., Burke L.M., Ross M.L., Camera D.M., West D.W.D., Broad E.M., Jeacocke N.A., Moore D.R., Stellingwerff T., Phillips S.M., et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J. Physiol. 2013;591:2319–2331. doi: 10.1113/jphysiol.2012.244897. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Hudson J.L., Bergia R.E., Campbell W.W. Effects of protein supplements consumed with meals, versus between meals, on resistance training–induced body composition changes in adults: A systematic review. Nutr. Rev. 2018;76:461–468. doi: 10.1093/nutrit/nuy012. [PubMed] [CrossRef] [Google Scholar]
75. Trommelen J., Kouw I.W.K., Holwerda A.M., Snijders T., Halson S.L., Rollo I., Verdijk L.B., Van Loon L.J.C. Pre-sleep dietary protein-derived amino acids are incorporated in myofibrillar protein during post-exercise overnight recovery. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2018 1:457–467. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28536184 [Google Scholar]
76. Kouw I.W., Holwerda A.M., Trommelen J., Kramer I.F., Bastiaanse J., Halson S.L., Wodzig W.K., Verdijk L.B., Van Loon L.J. Protein Ingestion before Sleep Increases Overnight Muscle Protein Synthesis Rates in Healthy Older Men: A Randomized Controlled Trial. [(accessed on 25 March 2019)];J. Nutr. 2017 147:2252–2261. doi: 10.3945/jn.117.254532. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28855419 [PubMed] [CrossRef] [Google Scholar]
77. Snijders T., Res P.T., Smeets J.S., Van Vliet S., Van Kranenburg J., Maase K., Kies A.K., Verdijk L.B., Van Loon L.J. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. [(accessed on 25 March 2019)];J. Nutr. 2015 145:1178–1184. doi: 10.3945/jn.114.208371. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25926415 [PubMed] [CrossRef] [Google Scholar]
78. Joy J.M., Vogel R.M., Broughton K.S., Kudla U., Kerr N.Y., Davison J.M., Wildman R.E.C., DiMarco N.M. Daytime and nighttime casein supplements similarly increase muscle size and strength in response to resistance training earlier in the day: A preliminary investigation. J. Int. Soc. Sports Nutr. 2018;15:24. doi: 10.1186/s12970-018-0228-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Antonio J., Ellerbroek A., Peacock C., Silver T. Casein Protein Supplementation in Trained Men and Women: Morning versus Evening. Int. J. Exerc. Sci. 2017;10:479–486. [PMC free article] [PubMed] [Google Scholar]
80. Schoenfeld B.J., Aragon A.A. How much protein can the body use in a single meal for muscle-building? Implications for daily protein distribution. J. Int. Soc. Sports Nutr. 2018;15:10. doi: 10.1186/s12970-018-0215-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
81. Pennings B., Groen B.B., Van Dijk J.-W., De Lange A., Kiskini A., Kuklinski M., Senden J.M., Van Loon L.J. Minced beef is more rapidly digested and absorbed than beef steak, resulting in greater postprandial protein retention in older men. Am. J. Clin. Nutr. 2013;98:121–128. doi: 10.3945/ajcn.112.051201. [PubMed] [CrossRef] [Google Scholar]
82. Kim I.Y., Schutzler S., Schrader A., Spencer H.J., Azhar G., Ferrando A.A., Wolfe R.R. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2016 310:73–80. doi: 10.1152/ajpendo.00365.2015. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26530155 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Jentjens R., Jeukendrup A.E. Determinants of Post-Exercise Glycogen Synthesis During Short-Term Recovery. Sports Med. 2003;33:117–144. doi: 10.2165/00007256-200333020-00004. [PubMed] [CrossRef] [Google Scholar]
84. Biolo G., Williams B.D., Fleming R.Y., Wolfe R.R. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes. 1999;48:949–957. doi: 10.2337/diabetes.48.5.949. [PubMed] [CrossRef] [Google Scholar]
85. Greenhaff P.L., Karagounis L.G., Peirce N., Simpson E.J., Hazell M., Layfield R., Wackerhage H., Smith K., Atherton P., Selby A., et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am. J. Physiol. Metab. 2008;295:E595–E604. doi: 10.1152/ajpendo.90411.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
86. Glynn E.L., Fry C.S., Timmerman K.L., Drummond M.J., Volpi E., Rasmussen B.B., Leroy J.L., Gadsden P., De Cossío T.G., Gertler P. Addition of Carbohydrate or Alanine to an Essential Amino Acid Mixture Does Not Enhance Human Skeletal Muscle Protein Anabolism123. J. Nutr. 2013;143:307–314. doi: 10.3945/jn.112.168203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Koopman R., Beelen M., Stellingwerff T., Pennings B., Saris W.H.M., Kies A.K., Kuipers H., Van Loon L.J.C. Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am. J. Physiol. Metab. 2007;293:E833–E842. doi: 10.1152/ajpendo.00135.2007. [PubMed] [CrossRef] [Google Scholar]
88. Aragon A.A., Schoenfeld B.J. Nutrient timing revisited: Is there a post-exercise anabolic window? J. Int. Soc. Sports Nutr. 2013;10:5. doi: 10.1186/1550-2783-10-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
89. Jäger R., Kerksick C.M., Campbell B.I., Cribb P.J., Wells S.D., Skwiat T.M., Purpura M., Ziegenfuss T.N., Ferrando A.A., Arent S.M., et al. International Society of Sports Nutrition position stand: Protein and exercise. [(accessed on 25 March 2019)];J. Int. Soc. Sport. Nutr. 2017 4:20. doi: 10.1186/s12970-017-0177-8. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28642676 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Darrabie M.D., Arciniegas A.J.L., Mishra R., Bowles D.E., Jacobs D.O., Santacruz L. AMPK and substrate availability regulate creatine transport in cultured cardiomyocytes. Am. J. Physiol. Metab. 2011;300:870–876. doi: 10.1152/ajpendo.00554.2010. [PubMed] [CrossRef] [Google Scholar]
91. Purchas R., Busboom J., Wilkinson B. Changes in the forms of iron and in concentrations of taurine, carnosine, coenzyme Q10, and creatine in beef longissimus muscle with cooking and simulated stomach and duodenal digestion. Meat Sci. 2006;74:443–449. doi: 10.1016/j.meatsci.2006.03.015. [PubMed] [CrossRef] [Google Scholar]
92. Branch J.D. Effect of Creatine Supplementation on Body Composition and Performance: A Meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2003;13:198–226. doi: 10.1123/ijsnem.13.2.198. [PubMed] [CrossRef] [Google Scholar]
93. Hultman E., Söderlund K., Timmons J.A., Cederblad G., Greenhaff P.L. Muscle creatine loading in men. [(accessed on 25 March 2019)];J. Appl. Physiol. Soc. 1996 81:232–237. doi: 10.1152/jappl.1996.81.1.232. Available online: https://www.ncbi.nlm.nih.gov/pubmed/8828669 [PubMed] [CrossRef] [Google Scholar]
94. Jagim A.R., Oliver J.M., Sanchez A., Galvan E., Fluckey J., Riechman S., Greenwood M., Kelly K., Meininger C., Rasmussen C., et al. A buffered form of creatine does not promote greater changes in muscle creatine content, body composition, or training adaptations than creatine monohydrate. J. Int. Soc. Sports Nutr. 2012;9:43. doi: 10.1186/1550-2783-9-43. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
95. Spillane M., Schoch R., Cooke M., Harvey T., Greenwood M., Kreider R., Willoughby D.S., Cooke M. The effects of creatine ethyl ester supplementation combined with heavy resistance training on body composition, muscle performance, and serum and muscle creatine levels. J. Int. Soc. Sports Nutr. 2009;6:6. doi: 10.1186/1550-2783-6-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Childs E., De Wit H., Wit H. Subjective, behavioral, and physiological effects of acute caffeine in light, nondependent caffeine users. Psychopharmacology. 2006;185:514–523. doi: 10.1007/s00213-006-0341-3. [PubMed] [CrossRef] [Google Scholar]
97. Bellar D., Kamimori G.H., Glickman E.L. The Effects of Low-Dose Caffeine on Perceived Pain During a Grip to Exhaustion Task. J. Strength Cond. Res. 2011;25:1225–1228. doi: 10.1519/JSC.0b013e3181d9901f. [PubMed] [CrossRef] [Google Scholar]
98. Davis J.K., Green J.M. Caffeine and anaerobic performance: Ergogenic value and mechanisms of action. [(accessed on 25 March 2019)];Sport. Med. 2009 39:813–832. doi: 10.2165/11317770-000000000-00000. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19757860 [PubMed] [CrossRef] [Google Scholar]
99. Wickwire P.J., McLester J.R., Gendle S., Hudson G., Pritchett R.C., Laurent C.M., Green J.M. Effects of Caffeine on Repetitions to Failure and Ratings of Perceived Exertion during Resistance Training. Int. J. Sports Physiol. Perform. 2007;2:250–259. [PubMed] [Google Scholar]
100. Duncan M.J., Oxford S.W. The effect of caffeine ingestion on mood state and bench press performance to failure. [(accessed on 25 March 2019)];J. Strength Cond. Res. 2001 25:178–185. doi: 10.1519/JSC.0b013e318201bddb. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22124354 [PubMed] [CrossRef] [Google Scholar]
101. Williams A.D., Cribb P.J., Cooke M.B., Hayes A. The Effect of Ephedra and Caffeine on Maximal Strength and Power in Resistance-Trained Athletes. J. Strength Cond. Res. 2008;22:464–470. doi: 10.1519/JSC.0b013e3181660320. [PubMed] [CrossRef] [Google Scholar]
102. Tarnopolsky M.A., Atkinson S.A., MacDougall J.D., Sale D.G., Sutton J.R. Physiological responses to caffeine during endurance running in habitual caffeine users. Med. Sci. Sports Exerc. 1989;21:418–424. doi: 10.1249/00005768-198908000-00013. [PubMed] [CrossRef] [Google Scholar]
103. Blanchard J., Sawers S.J.A. The absolute bioavailability of caffeine in man. Eur. J. Clin. Pharmacol. 1983;24:93–98. doi: 10.1007/BF00613933. [PubMed] [CrossRef] [Google Scholar]
104. Hobson R.M., Saunders B., Ball G., Harris R.C., Sale C. Effects of β-alanine supplementation on exercise performance: A meta-analysis. Amino Acids. 2012;43:25–37. doi: 10.1007/s00726-011-1200-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
105. Hoffman J., Ratamess N.A., Ross R., Kang J., Magrelli J., Neese K., Faigenbaum A.D., Wise J.A. Beta-alanine and the hormonal response to exercise. [(accessed on 25 March 2019)];Int. J. Sports Med. 2008 29:952–958. doi: 10.1055/s-2008-1038678. Available online: https://www.ncbi.nlm.nih.gov/pubmed/18548362 [PubMed] [CrossRef] [Google Scholar]
106. Hoffman J., Ratamess N., Kang J., Mangine G., Faigenbaum A., Stout J. Effect of creatine and β-alanine supplementation on performance and endocrine responses in strength/power athletes. [(accessed on 25 March 2019)];Int. J. Sport Nutr. Exerc. Metab. 2006 16:430–446. doi: 10.1123/ijsnem.16.4.430. Available online: https://www.ncbi.nlm.nih.gov/pubmed/17136944 [PubMed] [CrossRef] [Google Scholar]
108. Wax B., Kavazis A.N., Weldon K., Sperlak J. Effects of Supplemental Citrulline Malate Ingestion During Repeated Bouts of Lower-Body Exercise in Advanced Weightlifters. J. Strength Cond. Res. 2015;29:786–792. doi: 10.1519/JSC.0000000000000670. [PubMed] [CrossRef] [Google Scholar]
109. Wax B., Kavazis A.N., Luckett W. Effects of Supplemental Citrulline-Malate Ingestion on Blood Lactate, Cardiovascular Dynamics and Resistance Exercise Performance in Trained Males. [(accessed on 25 March 2019)];J. Diet. 2016 13:269–282. doi: 10.3109/19390211.2015.1008615. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25674699 [PubMed] [CrossRef] [Google Scholar]
110. Glenn J.M., Gray M., Wethington L.N., Stone M.S., Stewart R.W., Jr., Moyen N.E. Acute citrulline malate supplementation improves upper- and lower-body submaximal weightlifting exercise performance in resistance-trained females. [(accessed on 25 March 2019)];Eur. J. Nutr. 2017 56:775–784. doi: 10.1007/s00394-015-1124-6. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26658899 [PubMed] [CrossRef] [Google Scholar]
111. Glenn J.M., Gray M., Jensen A., Stone M.S., Vincenzo J.L. Acute citrulline-malate supplementation improves maximal strength and anaerobic power in female, masters athletes tennis players. Eur. J. Sport Sci. 2016;16:1–9. doi: 10.1080/17461391.2016.1158321. [PubMed] [CrossRef] [Google Scholar]
112. Gonzalez A.M., Spitz R.W., Ghigiarelli J.J., Sell K.M., Mangine G.T. Acute Effect of Citrulline Malate Supplementation on Upper-Body Resistance Exercise Performance in Recreationally Resistance-Trained Men. J. Strength Cond. Res. 2018;32:3088–3094. doi: 10.1519/JSC.0000000000002373. [PubMed] [CrossRef] [Google Scholar]
113. Farney T.M., Bliss M.V., Hearon C.M., Salazar D.A. The Effect of Citrulline Malate Supplementation On Muscle Fatigue Among Healthy Participants. J. Strength Cond. Res. 2017:1. doi: 10.1519/JSC.0000000000002356. [PubMed] [CrossRef] [Google Scholar]
114. Trexler E.T., Persky A.M., Ryan E.D., Schwartz T.A., Stoner L., Smith-Ryan A.E. Acute Effects of Citrulline Supplementation on High-Intensity Strength and Power Performance: A Systematic Review and Meta-Analysis. Sports Med. 2019;49:707–718. doi: 10.1007/s40279-019-01091-z. [PubMed] [CrossRef] [Google Scholar]
115. Kleiner S.M., Bazzarre T.L., Litchford M.D. Metabolic profiles, diet, and health practices of championship male and female bodybuilders. J. Am. Diet. Assoc. 1990;90:962–967. [PubMed] [Google Scholar]
116. Kleiner S.M., Bazzarre T.L., Ainsworth B.E. Nutritional Status of Nationally Ranked Elite Bodybuilders. Int. J. Sport Nutr. 1994;4:54–69. doi: 10.1123/ijsn.4.1.54. [PubMed] [CrossRef] [Google Scholar]
117. Sandoval W.M., Heyward V.H. Food Selection Patterns of Bodybuilders. Int. J. Sport Nutr. 1991;1:61–68. doi: 10.1123/ijsn.1.1.61. [PubMed] [CrossRef] [Google Scholar]
118. Ismaeel A., Weems S., Willoughby D.S. A Comparison of the Nutrient Intakes of Macronutrient-Based Dieting and Strict Dieting Bodybuilders. Int. J. Sport Nutr. Exerc. Metab. 2018;28:502–508. doi: 10.1123/ijsnem.2017-0323. [PubMed] [CrossRef] [Google Scholar]
119. Nelson J.R., Raskin S. The eicosapentaenoic acid:arachidonic acid ratio and its clinical utility in cardiovascular disease. Postgrad. Med. 2019;131:268–277. doi: 10.1080/00325481.2019.1607414. [PubMed] [CrossRef] [Google Scholar]
120. Harris W.S. The Omega-6: Omega-3 ratio: A critical appraisal and possible successor. [(accessed on 15 June 2019)];Prostaglandins Leukot Essent Fatty Acids. 2018 132:34–40. doi: 10.1016/j.plefa.2018.03.003. Available online: https://www.ncbi.nlm.nih.gov/m/pubmed/29599053/ [PubMed] [CrossRef] [Google Scholar]
121. Tachtsis B., Camera D., Lacham-Kaplan O. Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration. Nutrients. 2018;10:309. doi: 10.3390/nu10030309. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
122. Di Girolamo F.G., Situlin R., Mazzucco S., Valentini R., Toigo G., Biolo G. Omega-3 fatty acids and protein metabolism: Enhancement of anabolic interventions for sarcopenia. [(accessed on 15 June 2019)];Curr. Opin. Clin. Nutr. Metab Care. 2014 17:145–150. doi: 10.1097/MCO.0000000000000032. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24500439 [PubMed] [CrossRef] [Google Scholar]
123. McGlory C., Wardle S.L., Macnaughton L.S., Witard O.C., Scott F., Dick J., Bell J.G., Phillips S.M., Galloway S.D.R., Hamilton D.L., et al. Fish oil supplementation suppresses resistance exercise and feeding-induced increases in anabolic signaling without affecting myofibrillar protein synthesis in young men. Physiol. Rep. 2016;4:e12715. doi: 10.14814/phy2.12715. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
124. Crestani D.M., Bonin E.F.R., Barbieri R.A., Zagatto A.M., Higino W.P., Milion F. Chronic supplementation of omega-3 can improve body composition and maximal strength, but does not change the resistance to neuromuscular fatigue. [(accessed on 15 June 2019)];Sport Sci. Health. 2017 13:259–265. doi: 10.1007/s11332-016-0322-9. Available online: https://link.springer.com/article/10.1007/s11332-016-0322-9 [CrossRef] [Google Scholar]
125. Lewis E.J.H., Radonic P.W., Wolever T.M.S., Wells G.D. 21 days of mammalian omega-3 fatty acid supplementation improves aspects of neuromuscular function and performance in male athletes compared to olive oil placebo. J. Int. Soc. Sports Nutr. 2015;12:28. doi: 10.1186/s12970-015-0089-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
126. Rossato L.T., Schoenfeld B.J., De Oliveira E.P. Is there sufficient evidence to supplement omega-3 fatty acids to increase muscle mass and strength in young and older adults? Clin. Nutr. 2019 doi: 10.1016/j.clnu.2019.01.001. [PubMed] [CrossRef] [Google Scholar]
127. Mocking R.J.T., Harmsen I., Assies J., Koeter M.W.J., Ruhé H.G., Schene A.H. Meta-analysis and meta-regression of omega-3 polyunsaturated fatty acid supplementation for major depressive disorder. Transl. Psychiatry. 2016;6:e756. doi: 10.1038/tp.2016.29. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
128. Maki K.C., Palacios O.M., Bell M., Toth P.P. Use of supplemental long-chain omega-3 fatty acids and risk for cardiac death: An updated meta-analysis and review of research gaps. J. Clin. Lipidol. 2017;11:1152–1160.e2. doi: 10.1016/j.jacl.2017.07.010. [PubMed] [CrossRef] [Google Scholar]
129. Miller P.E., Van Elswyk M., Alexander D.D. Long-Chain Omega-3 Fatty Acids Eicosapentaenoic Acid and Docosahexaenoic Acid and Blood Pressure: A Meta-Analysis of Randomized Controlled Trials. Am. J. Hypertens. 2014;27:885–896. doi: 10.1093/ajh/hpu024. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
130. Du S., Jin J., Fang W., Su Q. Does Fish Oil Have an Anti-Obesity Effect in Overweight/Obese Adults? A Meta-Analysis of Randomized Controlled Trials. PLoS ONE. 2015;10:e0142652. doi: 10.1371/journal.pone.0142652. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Per accedere alle precedenti parti, dalla prima alla terza, clicca qui, qui e qui.
Disclaimer: quelle che seguono sono informazioni provenienti da casi studio [tranne dove diversamente specificato] e testimonianze di Bodybuilder sull’uso dell’Insulina, alcune delle quali sono palesemente sbagliate. Non prendete assolutamente queste opinioni come consigli.
Insulina – dall’uso clinico al Bodybuinding:
Come abbiamo visto nella prima parte, inizialmente l’Insulina farmaceutica era di origine animale. In questo caso, l’Insulina viene estratta dal pancreas di suino o di mucca (o di entrambi) e preparata per uso medico. Queste preparazioni sono ulteriormente suddivise nelle categorie “standard” e “purificate”, a seconda del livello di purezza e del contenuto non insulinico della soluzione. Con questi preparati c’è sempre la possibilità che contaminanti pancreatici siano presenti nel farmaco.
Nel 1977, Herbert Boyer e i suoi collaboratori Keiichi Itakura e Arthur Riggs al City of Hope National Medical Center descrisse la prima sintesi ed espressione di un gene codificante per un peptide. Nell’agosto del 1978, Boyer produsse Insulina sintetica utilizzando i suoi nuovi batteri transgenici geneticamente modificati, seguita nel 1979 dall’Ormone della Crescita. La Tecnologia del DNA ricombinante faceva il suo debutto e cambiava la storia dell’Insulina per uso medico.
L’Insulina umana biosintetica (insulina umana rDNA), attualmente e maggiormente utilizzata per uso clinico, è prodotta con la tecnologia del DNA ricombinante. L’Insulina umana biosintetica ha una maggiore purezza rispetto all’Insulina animale estrattiva e riduce la formazione di anticorpi. I ricercatori sono riusciti a introdurre il gene dell’Insulina umana nelle piante come un altro metodo per produrre Insulina (“biopharming”) nel cartamo. Si prevede che questa tecnica ridurrà i costi di produzione.
Sono disponibili diversi analoghi dell’Insulina umana. Questi analoghi dell’Insulina sono strettamente correlati alla struttura dell’Insulina umana e sono stati sviluppati per aspetti specifici del controllo glicemico in termini di azione rapida (insuline prandiali) e azione prolungata (insuline basali). Il primo analogo biosintetico dell’insulina è stato sviluppato per l’uso clinico al momento del pasto (insulina prandiale), Humalog (Insulina lispro), è assorbita più rapidamente dopo l’iniezione sottocutanea rispetto all’Insulina normale, con un effetto a 15 minuti dopo l’iniezione. Altri analoghi ad azione rapida sono NovoRapid e Apidra, con profili simili. Tutti vengono assorbiti rapidamente grazie a sequenze aminoacidiche che riducono la formazione di dimeri ed esameri (le insuline monomeriche vengono assorbite più rapidamente). Le insuline ad azione rapida non richiedono l’intervallo iniezione-pasto precedentemente raccomandato per l’Insulina umana e le insuline animali. L’altro tipo è l’Insulina a lunga durata d’azione; la prima di queste è stata Lantus (Insulina glargine). Queste hanno un effetto costante per un periodo prolungato, da 18 a 24 ore. Allo stesso modo, un altro analogo dell’Insulina a lunga durata d’azione (Levemir) si basa su un approccio di acilazione degli acidi grassi. A questo analogo è legata una molecola di acido miristico, che associa la molecola di Insulina all’abbondante albumina sierica, prolungando così l’effetto e riducendo il rischio di ipoglicemia. Entrambi gli analoghi ad azione prolungata devono essere assunti una sola volta al giorno e sono utilizzati nei diabetici di tipo 1 come Insulina basale. È disponibile anche una combinazione di un’Insulina ad azione rapida e di un’Insulina protratta, che consente ai pazienti di ottenere un profilo insulinico simile a quello del rilascio di Insulina da parte dell’organismo. L’Insulina viene utilizzata anche in molte linee cellulari, come CHO-s, HEK 293 o Sf9, per la produzione di anticorpi monoclonali, vaccini virali e prodotti per la terapia genica.
L’Insulina viene solitamente somministrata sotto forma di iniezioni sottocutanee tramite siringhe monouso con aghi, tramite un microinfusore di Insulina o tramite penne da insulina a uso ripetuto con aghi monouso. Sul mercato statunitense è disponibile anche l’Insulina per inalazione.
A differenza di molti farmaci, l’Insulina non può essere assunta per bocca perché, come quasi tutte le altre proteine introdotte nel tratto gastrointestinale, si riduce in frammenti amminoacidici, perdendo tutto il suo potenziale di attività. Sono state condotte alcune ricerche su come proteggere l’Insulina dal tratto digestivo, in modo da poterla somministrare per via orale o sublinguale.
Nel 2021, l’Organizzazione Mondiale della Sanità ha aggiunto l’Insulina al suo modello di elenco di farmaci essenziali.
Complice la descrizione iniziale allettante riportata in letteratura riguardante l’azione dell’Insulina sul metabolismo glucidico e proteico, dove tale peptide veniva descritto come l'”ormone più anabolico”, unita alla maggiore disponibilità di approvvigionamento del farmaco data dalla Tecnologia del DNA ricombinante, nel mondo della cultura fisica di alto livello non mancarono i primi pionieri del suo utilizzo “”off-label”.
Si può stimare che nel giro di 40 anni, vale a dire dagli anni 80 ad oggi, l’uso dell’Insulina nel Bodybuilding abbia subito sia un abuso pratico che teorico. Quello che spero di fare con questa mia piccola opera divulgativa è proprio quello di cambiare questa situazione. Come uomo di scienza con anni di ricerca alle spalle sono qualificato per giudicare le conoscenze dei Bodybuilder. Molti di loro conoscono molto meno di me il funzionamento e l’uso dell’Insulina. Quello che porto sul tavolo sono anni di ricerca nella comunità del bodybuilding e una corposo serie di prospettive diverse di culturisti sull’uso dell’Insulina, raccolte da 20 interviste con utilizzatori di Insulina con un’esperienza che va da mesi a decenni. Tranquilli però, fortunatamente non sono un “classico camicie bianco”, ma uno che analizza con attenzione è sa ammettere quando gli atleti hanno ragione e la ricerca scientifica si sbaglia. È ora quindi di trasformare la “broscience” dell’Insulina in scienza vera e propria e di correggere alcuni miti potenzialmente pericolosi.
Sto scrivendo questo articolo non solo per dimostrare le diverse, e persino contraddittorie, opinioni che i bodybuilder hanno sull’insulina, ma anche per ispirarvi a riflettere lucidamente.
I culturisti, quelli con doti intellettive un minimo sopra la media dei loro colleghi, sanno cose che i medici non potrebbero mai sapere perché hanno un obiettivo diverso e priorità diverse. Ma una cosa è certa, i bodybuilder accorti vogliono praticare il bodybuilding in modo scientificamente informato.
Non saranno presenti i nomi dei culturisti intervistati, dei quali sono stati esaminati i video e di cui sono state raccolte le affermazioni nei forum, per un principio etico che prevede di non rivelare l’identità delle persone che hanno contribuito alla presente ricerca, per evitare qualsiasi rischio di danno alla loro persona e immagine. Pertanto, tutti i nomi sono pseudonimi. È sufficiente dire che sono stati intervistati alcuni individui riconosciuti come esperti mondiali nell’uso dell’Insulina per il bodybuilding. sono stati inclusi anche i bodybuilder medi.
L’Insulina vista dai BodyBuilder:
I bodybuilder hanno tra loro visioni piuttosto diverse riguardo all’Insulina e ai suoi effetti. Per esempio, essi non sono d’accordo all’unanimità su quanto sia anabolizzante l’Insulina. Una minoranza afferma che l’Insulina esogena non è direttamente anabolizzante. Alcuni suggeriscono che l’Insulina induce l’anabolismo solo aumentando l’appetito. Ma, come sappiamo bene, di per se, quest’ultima affermazione ha ben poco senso.
Una piccolissima minoranza di bodybuilder sostiene che l’impatto dell’Insulina esogena sia principalmente, o puramente, cosmetico, in quanto l’Insulina fa apparire il muscolo più “pieno” (piuttosto che aumentare effettivamente le dimensioni del muscolo) aumentando la ritenzione idrica intracellulare.
Al contrario, molti bodybuilder sostengono che l’Insulina sia l’ormone più anabolico.
Alcuni partecipanti hanno descritto l’uso dell’Insulina come il risultato di un aumento muscolare di 3-6kg a settimana rispetto a quello che si potrebbe ottenere con il solo uso di Steroidi Androgeni Anabolizzanti (AAS) o con l’uso combinato di AAS e Ormone della Crescita (GH). Tuttavia, queste affermazioni sono state contestate da alcuni. Vi sono bodybuilder che ritengono che i benefici dell’Insulina non siano sufficienti e non hanno intenzione di utilizzare nuovamente questo peptide. Uno di questi non raccomanda più l’uso dell’Insulina ai suoi clienti.
Mentre alcuni bodybuilder sostengono che l’Insulina da sola sia anabolizzante, altri suggeriscono che essa sia significativamente anabolizzante solo in sinergia con AAS e hGH.
La maggior parte dei bodybuilder concorda sul funzionamento dell’Insulina, anche se le loro spiegazioni sono più o meno scientifiche.
In genere i bodybuilder descrivono l’Insulina come una “navetta” o un “mezzo” che trasporta i nutrienti nelle cellule muscolari. L’Insulina viene descritta come una “chiave” che apre le porte delle cellule o come un “autobus” che trasporta i nutrienti. Alcuni bodybuilder citano la letteratura scientifica quando descrivono l’azione dell’Insulina:
Sembra che l’Insulina abbia almeno un effetto permissivo sulla sintesi proteica, tanto che i suoi livelli basali sono necessari per la normale sintesi proteica miofibrillare (MPS), ma l’aumento dell’Insulina dopo un pasto potrebbe non aumentare la MPS (Greenhaff et al., 2008). Tuttavia, l’Insulina promuove l’anabolismo muscolare (bilancio proteico proattivo) attraverso il suo effetto inibitorio sulla degradazione delle proteine muscolari (MPB) (Deutz e Wolf 2013). Inoltre, l’Insulina può aumentare l’MPS attraverso un maggiore assorbimento di aminoacidi (essenziali) nel muscolo scheletrico, provocato da un aumento del flusso sanguigno associato alla vasodilatazione (Biolo et al., 1995; Fujita et al., 2006; Timmerman et al., 2010). [Chad via e-mail].
Molti concordano sul fatto che:
l’Insulina è molto efficace nel trasportare i nutrienti nelle cellule muscolari, ma anche nelle cellule adipose.
l’Insulina è anabolizzante grazie al suo ruolo nella ripartizione dei nutrienti, in quanto lavora di concerto con GH e IGF-I.
l’Insulina promuove l’anabolismo muscolare grazie al suo effetto inibitorio sulla degradazione delle proteine muscolari.
l’Insulina (così come gli AAS e il GH) promuove la sintesi proteica muscolare solo in presenza di un adeguato apporto di aminoacidi.
l’Insulina svolge un ruolo nel controllo fisiologico della riproduzione, agendo sulla secrezione dell’Ormone di Rilascio delle Gonadotropine (GnRH)/luteinizzante (LH).
L’Insulina è anche descritta da alcuni bodybuilder come anti-catabolica.
Un bodybuilder di alto livello ha dichiarato che l’Insulina dovrebbe essere usata solo se un individuo è carente di Insulina:
A volte è una buona idea prendere l’Insulina solo per aiutare il pancreas a fare il suo lavoro. … se il vostro corpo producesse abbastanza da solo, perché avreste bisogno di assumere Insulina esogena? In altre parole, non apporta alcun beneficio. È utile solo se non si produce abbastanza Insulina. Quindi bisogna innanzitutto stabilire se non si produce abbastanza Insulina. Procuratevi un glucometro e controllate la glicemia. … Quindi l’Insulina dovrebbe essere usata solo se si ha una carenza di insulina, perché si sta usando molto GH o perché si sta mangiando una quantità esorbitante di carboidrati.
Definirei questa ipotesi come “patologica indotta/deficitaria”. Ricordatevi sempre che l’omeostasi organica è regolata da feedback. Ciò significa che l’uso di Insulina esogena causerà una sottoregolazione/inibizione della biosintesi endogena di Insiluna. Di conseguenza, parlare di “pancreas ipoattivi” o “supporto pancreatico” non è in definitiva corretto. In tal caso si parla di una vera e propria terapia ormonale sostitutiva dell’Insulina.
Un bodybuilder ha anche affermato che gli effetti anabolici dell’Insulina sono dovuti alla sua azione osmotica e alla sua capacità di aprire “tutti i recettori del corpo”. Descrizione alquanto particolare ma che può rendere una certa idea di uno degli effetti dell’Insulina.
Mentre alcuni bodybuilder mettono in guardia dall’uso dell’Insulina perché può causare ipoglicemia con conseguente coma ipoglicemico potenzialmente letale, la maggior parte ritiene che i rischi dell’Insulina siano stati sopravvalutati e alcuni suggeriscono che la morte dovuta all’uso di Insulina è estremamente improbabile. Molti pensano che “bisogna essere una testa di cazzo per uccidersi con l’Insulina”.
Sebbene la morte di diversi culturisti di alto livello nel corso della mia ricerca sia stata inizialmente suggerita da membri della comunità dei culturisti come correlata all’Insulina, l’Insulina non è stata implicata nelle cause ufficiali dei loro decessi. Non mi sono imbattuto in un caso confermato di morte o di danni significativi causati dall’uso di Insulina per il bodybuilding, anche se alcuni bodybuilder hanno dichiarato di conoscere qualcuno che è morto a causa dell’uso di Insulina (e uno di loro ha avuto un grave incidente d’auto a causa di una ipoglicemia avuta in autostrada, ma fortunatamente nessuno è rimasto ferito). Nella letteratura medica sono riportati due casi di bodybuilder in coma ipoglicemico (Heidet et al., 2019; Petrovic et al., 2015). Non credo che la mancanza di decessi confermati sia dovuta al fatto che l’Insulina non sia pericolosa, ma più probabilmente perché non è comunemente testata o difficile da rilevare.
Un bodybuilder ha suggerito che le morti premature dovute all’uso di Insulina potrebbero non essere dovute solo all’ipoglicemia, ma ha suggerito che l’aumento dei livelli di Insulina nel corso della vita accorcia la durata della stessa e che quindi i bodybuilder si mettono a rischio in questo senso.
Alcuni bodybuilder sostengono che l’Insulina sia uno dei farmaci più sicuri del loro arsenale, in particolare rispetto al DNP e al Trenbolone. Alcuni suggeriscono addirittura che l’Insulina sia più sicura di qualsiasi AAS.
Mentre tutti i bodybuilder hanno descritto almeno lievi sintomi di ipoglicemia in alcuni momenti del loro utilizzo di Insulina, la maggior parte degli episodi di ipoglicemia si sono verificati durante le prime fasi di utilizzo, quando stavano elaborando il dosaggio dell’Insulina, o sono stati attribuiti alla loro stupidità (ad esempio, dimenticando di mangiare). Tutti i bodybuilder hanno dichiarato che l’ipoglicemia era facilmente gestibile consumando zuccheri.
Molti suggeriscono che per essere competitivi come bodybuilder professionisti è necessario utilizzare l’Insulina. Tuttavia, altri suggeriscono che non è necessario.
L’Insulina fa ormai parte del bodybuilding, ne è parte integrante. È come i denti sbiancati. Tutti sbiancano i denti, tutti hanno denti bianchi e splendenti. Se c’è uno che non li ha, gli si chiede: “Cosa c’è che non va in te?”. … Se non fai l’Insulina, cosa che alcuni professionisti non fanno, alcuni non ne hanno bisogno, allora il tuo aspetto è un po’ diverso da quello degli altri bodybuilder e potresti distinguerti in modo negativo.
Molti attribuiscono il significativo aumento della muscolatura dei mostri di massa all’Insulina e/o all’Ormone della Crescita. Alcuni suggeriscono che l’Insulina abbia rovinato il bodybuilding, poiché l’attenzione si è spostata dall’estetica alle dimensioni a scapito dell’estetica. Altri criticano ulteriormente il look dell’Insulina:
Prima dell’arrivo dell’Insulina, tornando ai primi tempi di Bertil Fox, Tom Platts, Arnold, Sergio, i loro muscoli avevano questo aspetto duro e granitico, sembravano scolpiti nella pietra. Ora ci sono questi ragazzi, certo grandi e stravaganti come i Ramy e tutti questi ragazzi, sono grandi ma non hanno quell’aspetto duro e denso.
Ma altri suggeriscono che Dorian Yates è stato il primo a portare sul palcoscenico del Olympia un fisico potenziato dall’Insulina e viene spesso descritto come se avesse un aspetto granitico.
Alcuni suggeriscono che l’Insulina (e/o l’Ormone della Crescita) provochi la “bolla intestinale” o il “palumboismo” [vedi anche “GH Gut”], e una fonte ha affermato che l’Insulina ha causato l’organomegalia. Per questi motivi alcuni affermano che l’Insulina ha rovinato l’estetica del bodybuilding.
Il dosaggio dell’insulina è molto vario tra i bodybuilder. Ho parlato con culturisti che usano un massimo di 4UI al giorno, e altri che hanno usato un massimo di 360UI al giorno! Anche se lo considero ben poco credibile. Tuttavia, in linea con le precedenti ricerche accademiche che riportavano dosaggi compresi tra 10 e 20 unità al giorno (Dawson e Harrison 1997; Evans 1997; Hildebrandt et al., 2007) e con i sondaggi condotti all’interno della comunità, ho scoperto che la maggior parte dei bodybuilder utilizza dosi che si collocano all’estremità inferiore dello spettro. Tuttavia, una buona parte dei bodybuilder tende a usare più di quanto riportato in precedenza nella letteratura accademica e nella comunità, con una dose giornaliera mediana di 40 unità e una dose mediana di 0,39 unità per chilogrammo di peso corporeo.
La maggior parte dei bodybuilder utilizza una sola forma di insulina (ad azione rapida o ultra-rapida), anche se un quarto degli intervistati (n=20) utilizza anche un’Insulina a lunga durata d’azione.
Ma vediamo nel dettaglio i tipi di Insulina utilizzati:
Humalog® (Insulina Lispro): Humalog® è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”.
Novolog® (Insulina Aspart):Novolog è un analogo a breve durata d’azione dell’Insulina umana, creato quando l’aminoacido prolina in posizione B28 viene sostituito con l’acido aspartico. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 15 minuti e l’effetto di picco si raggiunge in 1-3 ore. La durata d’azione totale è compresa tra le 3 e le 5 ore. L’Insulina Aspart viene solitamente utilizzata come supporto a un prodotto contenente insulina a più lunga durata d’azione, fornendo un farmaco a rapida azione che può essere assunto prima o subito dopo i pasti per imitare la risposta insulinica dell’organismo. Molti atleti ritengono che la sua breve finestra di effetto la renda ideale per scopi dopanti, tanto quanto la Lispro, in quanto la maggior parte della sua azione si può concentra nel periodo successivo all’allenamento durante la “finestra anabolica”.
Humulin®-R “Regular” (insulina Inj): Identica all’Insulina umana. Venduta in alcuni mercati anche come Humulin-S® (Solubile), questo prodotto è costituito da cristalli di zinco-insulina disciolti in un liquido chiaro. Non viene aggiunto nulla per rallentare il rilascio di questo prodotto, per cui viene genericamente indicato come insulina umana solubile. Questo farmaco agisce rapidamente e ha una breve durata d’azione. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di 20-30 minuti, e il suo picco d’effetto si raggiunge in 1-3 ore. Ha una durata d’azione totale tra le 5 e le 8 ore. Insieme a Humalog, queste due forme di Insulina sono le scelte più popolari tra gli atleti e i culturisti per scopi dopanti.
Humulin®-N, NPH (Insulina Isofana): Una sospensione cristallina di Insulina con protamina e zinco per ritardarne il rilascio e prolungarne l’azione. L’Insulina Isofana è considerata un’Insulina di lunghezza intermedia. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore e il picco d’effetto si raggiunge in 4-10 ore. La durata totale dell’attività è superiore a 14 ore. Questo tipo di Insulina non è comunemente usata come agente dopante.
Humulin®-L, Lente (sospensione media di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-L è considerata un’insulina di lunghezza d’azione intermedia. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 1-3 ore e l’effetto di picco viene raggiunto in 6-14 ore. Ha una durata totale di attività superiore alle 20 ore. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin®-U, Ultralente (sospensione prolungata di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-U è considerata un’Insulina a lunga durata d’azione. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 6 ore, e l’effetto di picco viene raggiunto in 14-18 ore. La durata totale dell’attività è di 18-24 ore. Questo tipo di insulina non è comunemente usato per scopi dopanti.
Lantus (Insulina Glargine): Analogo a lunga durata d’azione dell’Insulina umana. L’Insulina Glargine viene creata quando l’aminoacido asparagina in posizione A21 viene sostituito con la glicina e vengono aggiunte due arginine al C-terminale della catena B dell’Insulina. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore, e il farmaco è considerato privo di un picco significativo (è stato progettato per un modello di rilascio molto stabile per tutta la durata dell’attività). L’Insulina Glargine ha una durata d’azione compresa tra 20-24 ore nell’organismo dopo l’iniezione sottocutanea. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin® (Miscele) : Sono miscele di Insulina solubile normale per un’azione rapida, e di un’Insulina a lunga durata d’azione o ad azione intermetizzata per un effetto prolungato. Queste miscele sono etichettate con la percentuale di miscela, di solito 10/90, 20/80, 30/70, 40/60 e 50/50. Sono anche disponibili le miscele che utilizzano Humalog come Insulina ad azione rapida.
Indipendentemente dal tipo, tutte le insuline forniscono gli stessi effetti di base, e la durata di azione è la differenziazione primaria. Il trasporto dei nutrienti, l’aumento della sintesi proteica, la diminuzione del catabolismo proteico, l’aumento del IGF-1, l’aumento della biodisponibilità del IGF-1, e una maggiore vasodilatazioni sono i vantaggi più noti.
Non è da molto tempo che un tipo di Insulina precedentemente marginale a fini dopanti è diventata di moda tra alcuni culturisti. Sto parlando della Lantus (Insulina Glargine), appunto.
Di tutte le diverse insuline disponibili, la Lantus è probabilmente quella meno utilizzata anche perchè e paradossalmente la più complessa da gestire. Il suo scarso utilizzo nel bodybuilding ha portato, come ovvia conseguenza, ad una scarsità delle informazioni disponibili su di essa.
A differenza delle insuline a breve durata d’azione, che forniscono i benefici di cui sopra per poche ore al giorno, la Lantus continuerà il trasporto dei nutrienti, l’aumento della sintesi proteica, il miglioramento della vasodilatazioni, ecc, per tutto il giorno, anche mentre dormiamo, ed è questo ultimo punto a renderla di non facile gestione. Ma uno degli svantaggi principali della Lantus risiede nella sua possibilità di essere utilizzata solo per brevi periodi di tempo, in quanto l’esposizione continua a livelli elevati di Insulina esogena, e lo sappiamo bene, porterà ad una riduzione della sensibilità all’insulina, la successiva sotto-regolazione dei trasportatori GLUT-4, cose che si vorrebbero evitare sia da un punto di vista della saluta che della crescita muscolare. Così, mentre la Lantus può essere superiore per lo stimolo della crescita muscolare nel breve termine, troviamo che le cose cominciano a pareggiarsi nel lungo periodo, e protocolli come quelli pre-allenamento più comunemente impiegati possono invece essere utilizzati a tempo indeterminato, senza danneggiare eccessivamente la sensibilità all’Insulina a qualsiasi grado significativo. Questo rende la Lantus ideale per dei “blitz”, in cui l’atleta vuole mettere su muscoli il più rapidamente possibile, ma non è adatta per un uso prolungato.
Differenza nella curva di rilascio tra Humulin N e Lantus in pazienti con Diabete di Tipo I.
Una caratteristica decisamente interessante della Lantus è il suo effetto sul IGF-1 ed i suoi recettori. In diversi studi universitari, la Lantus ha dimostrato una maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi. È interessante notare che Levemir, l’unico altro analogo dell’Insulina ad azione prolungata sul mercato, mostra una ridotta affinità di legame ai recettori del IGF-1 umani. Questo mette la Lantus e la Levemir alle estremità opposte dello spettro in termini di affinità di legame. Mentre un aumento di IGF-1 vincolante è visto generalmente come una cosa positiva per la crescita muscolare, è stato il punto focale del dibattito in corso nella comunità medica per parecchi anni, per il fatto che alcuni studi hanno mostrato un aumento del rischio di cancro quando si usa la Lantus. Da allora, altri studi hanno confutato questa nozione, ma il dibattito continua, con la comunità medica riluttante a prendere una posizione in un modo o nell’altro.
Via di segnalazione del Recettore dell’Insulina (IR) e del Recettore del Fattore di Crescita Insulino-Simile 1 (IGF1R). L’Insulina e l’IGF1 si legano ai loro recettori, inducendo un cambiamento conformazionale e l’autofosforilazione della subunità beta di IR e IGF1R. Successivamente, le proteine substrato del recettore dell’Insulina (IRS) o Shc vengono reclutate e fosforilate. Shc attiva la via della mitogen-activated protein kinase-extracellular signal regulated kinase (MAPK-ERK) e le proteine IRS inducono prevalentemente l’attivazione della via della fosfoinositide 3-chinasi (PI3K)-AKT. In questo caso, l’attivazione di PI3K causa la conversione del fosfatidilinositolo 4,5-bisfosfato (PIP 2 ) in fosfatidilinositolo (3,4,5)-trifosfato (PIP 3 ) e l’attivazione e la fosforilazione di AKT da parte della proteina chinasi 1 dipendente dal fosfoinositide. La regolazione dipendente da AKT della forkhead box O (FoxO), del mammalian target of rapamycin complex 1 (mTORC1) e della glicogeno sintasi chinasi 3b (GSK3b) regola la crescita degli assoni, la trascrizione genica, la sintesi proteica e la plasticità neuronale. MEK, MAPK/ERK chinasi; PDK1, proteina chinasi 1 fosfoinositide-dipendente; SOS, son-of-sevenless. [Adattato da Servier Medical Art di Servier, con licenza Creative CommonsAttribuzione 3.0 Unported].
In definitiva, la Lantus viene solitamente usata come “base” di un protocollo di Insulina affiancata dall’uso del Humalog nei protocolli di Insulina e GH pre o post workout.
UI:CHO ratio e timing di somministrazione
Sappiamo tutti che il rapporto più frequentemente citato tra carboidrati e insulina (UI:CHO ratio) è di 10-15g di carboidrati per 1UI di Insulina. Ma non tutti i bodybuilder si attengono a questo rapporto. Diversi culturisti non hanno stabilito un rapporto fisso tra carboidrati e Insulina, mentre altri hanno utilizzato una gamma di rapporti (5-20g per unità di Insulina) con una media di 9-10g di carboidrati per ogni UI di Insulina.
La maggior parte dei bodybuilder ha assunto l’Insulina durante i pasti, ma considerando che i bodybuilder mangiano spesso, questo non ci dice molto. C’è stato un grande dibattito sul momento più efficace per l’uso dell’Insulina: alcuni suggeriscono che il momento più efficace sia il pre-workout, mentre altri affermano che si tratta di un uso irresponsabile, in quanto è difficile determinare quanti carboidrati verranno ossidati durante l’allenamento e quindi l’uso dell’Insulina nel pre-allenamento potrebbe essere pericoloso. Alcuni bodybuilder usano l’Insulina prima e dopo l’allenamento, altri solo dopo.
Alcuni bodybuilder sostengono che l’uso di Insulina a scopo ricreativo espone i bodybuilder al rischio di sviluppare il diabete. Altri bodybuilder sostengono che l’uso dell’Insulina riduce il rischio di diabete in quanto diminuisce l’impatto delle diete per il bodybuilding. Lasciatemi dire che l’ultima affermazione non ha alcun senso. Il corpo mantiene una condizione di omeostasi attraverso elaborati feedback di controllo. La somministrazione di Insulina esogena causerà un feedback negativo a livello della secrezione endogena pancreatica, e l’Insulina esogena somministrata avrà il medesimo effetto a livello centrale e periferico in un regime alimentare ipercalorico (vedi “dieta per il bodybuilding”) dell’Insulina endogena! E le affermazioni secondo le quali l’uso dell’Insulina esogena sortirebbe un effetto di protezione all’affaticamento pancreatico beh, è un affermazione che non ha basi di riscontro.
La follia del protocollo “No Fat Gain Insulin Program”:
Diversi anni fa riportai un protocollo d’uso dell’Insulina denominato “No Fat Gain Insulin Program”. Questo protocollo “alternativo” fu ideato da Mike Zumpano e Oliver Starr i quali si chiesero se ci poteva essere una strategia che permettesse ai bodybuilder di non aumentare eccessivamente di bf durante l’uso di Insulina. In realtà, la motivazione di base per la quale molti culturisti diventano più grassi che grossi quando nelle loro preparazioni inseriscono l’Insulina è fondamentalmente la “la paura” dell’ipoglicemia. La maggior parte del guadagno di grasso è causato dal consumo eccessivo di carboidrati durante l’uso di Insulina. Una regola di “sicurezza” diffusa con l’uso di Insulina dice che bisogna consumare un minimo di 10-15g di carboidrati per ogni UI di Insulina utilizzata (distribuiti nell’arco di tempo d’azione dell’Insulina utilizzata). Un altro errore commesso da molti bodybuilder e che porta ad un eccesso calorico addizionale è che essi non calcolano le UI in base ai CHO della dieta ma calcolano le UI in base al peso e di conseguenza aggiungono i carboidrati di “sicurezza” a quelli già presenti nel loro programma alimentare. Comunque sia, un Bodybuilder che utilizza 8UI di Insulina 2 volte al giorno, in aggiunta al suo normale apporto di carboidrati, proteine e grassi andrà (con il metodo standard) a consumere una quota addizionale di carboidrati pari a 160g. Difficilmente ci si aspetta che 160g in più di carboidrati, o 640Kcal in più al giorno facciano una differenza significativa su un soggetto che magari mangia 5000 o più calorie al giorno.
Anche se la quantità di carboidrati supplementari (10gXUI) comunemente applicata non sembra terribilmente eccessiva, alcuni “pionieri” dei PEDs alla fine degli anni ‘90 erano certi che fosse il motivo principale per cui gli utilizzatori di Insulina guadagnavano quantità sproporzionate di grasso.
Il plasma umano contiene solo circa 5g di carboidrati in uno specifico momento. I diabetici che hanno preso troppa Insulina di solito possono riportare i loro livelli glicemici nel sangue nel range di normalità consumando cinque grammi (solo 20 calorie!) di Destrosio.
Comunque sia, il metodo “alternativo” lo trovarono e fu ribattezzato, come precedentemente accennato, protocollo “No Fat Gain”. Il trucco, se così possiamo definirlo, sarebbe quello di assumere l’Insulina, ma seguendo un dieta Low-Carb. Proprio così, Low. Con un contenuto glucidico di circa 50g al giorno. Oliver Starr, con rudimentali conoscenze in biochimica e fisiologia umana, si chiese se ci fosse qualche altro modo per mantenere la glicemia nel sangue moderata con un alto grado di stabilità. La sua (riduttiva) risposta è stata la gluconeogenesi. Se si guarda su un grafico dei processi biochimici, si può chiaramente vedere che, quando le riserve di glicogeno epatico e muscolare sono esaurite, ma prima che il soggetto vada in chetosi, il corpo comincia a convertire aminoacidi in glucosio per mantenere stabili i livelli di glucosio nel sangue. Questo processo è noto come gluconeogenesi. Come risaputo, seguire una dieta molto povera di carboidrati provoca un esaurimento delle riserve di glicogeno epatico e muscolare. Questo provoca un sovra-regolazione degli enzimi necessari per la conversione rapida ed efficace degli aminoacidi in glucosio. La parola gluconeogenesi significa letteralmente “la nascita di nuovo glucosio.”
La seconda metà dell’ipotesi applicata, ovviamente, è il contenuto proteico della dieta. Se non si mangiano molti carboidrati, l’unico modo con cui il corpo può produrre glucosio è principalmente quello di convertire gli aminoacidi in glucosio. Questo accade in una certa misura ogni volta che si mangiano proteine, tuttavia, quando si mangia una grande quantità di proteine, si viene a creare ancora più glucosio. E’ il livello di glucosio creato dall’eccesso di proteine che dovrebbe impedire il verificarsi di uno stato ipoglicemico in un contesto nel quale si utilizza Insulina esogena con una dieta a basso contenuto di carboidrati.
Per questo protocollo è necessaria l’assunzione di proteine in polvere, perché non c’è modo di riuscire a essere in grado di mangiare la quantità di proteine che si richiedono da cibi interi. Per sostenere il livello di gluconeogenesi supposto per coprire le necessità durante l’utilizzo di Insulina si è proposto che la migliore strategia è quella in cui si consumano 600g di proteine da una combinazione di proteine del siero di latte e caseina, più un pasto solido che contiene da 50 a 100g di proteine, più alcune verdure fibrose a foglia verde. Il resto delle calorie devono provenire da fonti di grassi con scarsissimo o nullo contenuto di carboidrati.
Si può suddividere l’assunzione consumando una bevanda proteica ogni 30 minuti o un’ora, mescolando in un contenitore 3 litri con 100g di proteine e mantenendo una lista di quante volte si svuota ogni giorno.
Bisogna ricordare, però, che in questo protocollo, l’unica cosa che dovrebbe salvaguardare l’atleta (letteralmente) è l’assunzione di proteine. Se si utilizza l’Insulina con tali modalità e non si mantiene un adeguato apporto di proteine, le conseguenze saranno più che spiacevoli, saranno gravissime.
L’approccio teorico all’uso di Insulina in questo protocollo “No Fat Gains” è:
Giorni da 1 a 3: la deplezione di carboidrati. È necessario diminuire i carboidrati al di sotto dei 100g al giorno. Si suggerisce di arrivare a 50g di carboidrati il giorno 3. L’assunzione proteica deve aumentare a 450g al giorno.
Giorni da 4 a 30: le proteine devono essere pari o superiore a 600g al giorno. I carboidrati devono essere mantenuti tra i 100 e i 50g al giorno (50g è meglio) e si dovrebbero utilizzare i grassi affinché si compensi l’equilibrio delle proprie esigenze caloriche giornaliere. Come già detto, si raccomanda l’uso di proteine in polvere di composizione mista (siero di latte e caseina), anche se è possibile utilizzare alcuni cibi interi, se lo si desidera. (Basta tenere a mente che 600g grammi di proteine corrispondono all’incirca a più di 2.5Kg di petto di pollo o di tacchino al giorno)
Partire da una piccola dose di Insulina (4 UI) per poi aumentarla gradualmente (fino anche a 12UI x 3 volte al giorno).
Monitorare regolarmente la glicemia durante il giorno e in specie nel periodo di massima azione dell’Insulina. Se si inizia a perdere la capacità di rimanere svegli, prendere una zolletta di zucchero.
Si, questo protocollo non è soltanto folle ma, cosa fondamentale, è basato su una conoscenza superficiale e con enormi lacune della fisiologia umana. Perchè? Perchè l’Insulina è un regolatore in negativo della gluconeogenesi!
Il ruolo dell’Insulina nella regolazione della produzione epatica di glucosio è ampiamente accettato. Negli individui sani, l’iperinsulinemia fisiologica sopprime la gluconeogenesi del 20%, mentre la glicogenolisi è completamente soppressa. L’iperglicemia da sola sopprime la glicogenolisi epatica con effetti minimi sull’immagazzinamento del glicogeno. Solo la combinazione di iperglicemia e iperinsulinemia ha un effetto significativo sulla sintesi epatica di glicogeno. Pertanto, l’Insulina svolge un ruolo cruciale nel metabolismo epatico del glucosio.
Effetti dell’Insulina sul metabolismo del glucosio e dei lipidi.
Il meccanismo dominante della regolazione insulino-mediata della gluconeogenesi epatica non è però chiaro. L’Insulina esercita un controllo diretto della gluconeogenesi agendo sul fegato, ma influisce anche indirettamente sulla gluconeogenesi agendo su altri tessuti. L’effetto diretto dell’Insulina è stato dimostrato nei cani a digiuno, dove l’Insulina plasmatica portale ha soppresso la produzione epatica di glucosio, anche in assenza di variazioni del glucagone o dei precursori gluconeogenici. Tuttavia, nei modelli murini, l’Insulina è risultata avere effetti più potenti sulla produzione epatica di glucosio in vivo piuttosto che in vitro. Inoltre, è stato dimostrato che gli effetti indiretti dell’Insulina sui tessuti extraepatici sono sufficienti a mantenere il normale metabolismo del glucosio, suggerendo un ruolo importante per la regolazione indiretta della gluconeogenesi da parte dell’Insulina.
Regolazione dell’espressione genica della gluconeogenesi da parte del segnale insulinico epatico. L’azione dell’Insulina regola l’attività dei fattori di trascrizione che controllano l’espressione dei geni gluconeogenici. La fosforilazione mediata da AKT porta all’esportazione nucleare di FOXO1. La fosforilazione inibitoria di CBP/p300 blocca la formazione del complesso di trascrizione di CREB. La modifica di PGC-1α mediante acetilazione mediata da GCN5 o fosforilazione mediata da AKT/CLK2 riduce l’attività trascrizionale di PGC-1α.
Un sistema di segnalazione insulinica intatto è fondamentale per mantenere i livelli di glucosio nel sangue all’interno di un range glicemico normale e ristretto durante i periodi di digiuno o di eccesso di disponibilità di nutrienti, e questo si ottiene in particolare attraverso la regolazione del flusso metabolico attraverso la via gluconeogenica epatica. Un importante nodo di controllo coinvolge la regolazione trascrizionale dell’espressione dei geni chiave della gluconeogenesi epatica Pck1 e G6pc, che avviene principalmente attraverso il fattore di trascrizione FOXO1 e il recettore nucleare HNF4α e il loro coattivatore trascrizionale PGC-1α. La comprensione di queste vie di regolazione è di estrema importanza per comprendere come un massivo consumo proteico in regime low carb non possa assolutamente garantire una stabilità glicemica in presenza di trattamento con Insulina esogena!
Ma si ingrassa di meno con questo protocollo? Direi di no o, comunque, la differenza è irrisoria… Vorrei ricordare in tal sede che, per quanto dispendioso in termini energetici, in un contesto di eccesso calorico (essenziale in regimi “Bulk”) dove si consumano più proteine del necessario, l’organismo utilizza gli AA che le compongono o come come fonte energetica di scarsa resa [3,3Kcal per 1lt di Ossigeno] o li converte in acidi grassi! Certo, per necessità di deficienza nutrizionale la gluconeogenesi degli AA garantisce una stabilità glicemica, ma in fisiologia e non quando quest’ultima viene marcatamente alterata dall’uso di Insulina esogena!
Per concludere questo paragrafo, è corretto sottolineare che la gluconeogenesi si verifica dopo circa 8 ore di digiuno o scarso apporto glucidico, quando le scorte di glicogeno del fegato iniziano a esaurirsi ed è necessaria una fonte alternativa di glucosio. Inoltre è un processo biochimico piuttosto lento e assolutamente non garante della ben che minima sicurezza in un regime d’uso di Insulina in contesto low carb.
L’uso dell’Insulina pre-workout:
I dibattiti sulla reale efficacia dell’Insulina come agente anabolizzante ha spinto atleti e preparatori a sperimentare protocolli diversi. L’unico di questi che si è avvicinato maggiormente ad una logica d’insieme è ““The Ultimate Insulin Protocol” di Mike Arnold.
Ciò che sta alla base di questo protocollo non è altro che una versione “arricchita” di ciò che sta alla base dell’integrazione “intra-workout”.
Spesso si ragiona sul fatto che durante l’allenamento ci si trova in uno stato fondamentalmente catabolico e dopo in uno stato fondamentalmente anabolico. In realtà però, la sintesi proteica ed i meccanismi di anabolismo e catabolismo sono sempre attivi, con diverse prevalenze, quindi i processi di riparazione tissutale iniziano già nel momento in cui il muscolo viene danneggiato e durante il danneggiamento. Questo è ancor più vero nel momento in cui il workout sarà incentrato su più gruppi muscolari. La presenza di una concentrazione di Insulina esogena in circolo addizionata all’introduzione di macronutrienti ben calibrati (vedi integrazione intra-workout) renderebbe maggiormente incisivo il processo.
Il pump e la pienezza muscolare che si può raggiungere seguendo questo protocollo sono, a detta dei “tester”, impressionanti. Il programma trova la sua “magia” nella sua tempistica e nella sinergia degli ingredienti utilizzati.
Qui di seguito è riportato il protocollo nella sua interezza:
60 minuti pre-workout: *** optional (Uno qualsiasi dei supplementi NO stimolanti sul mercato. Gaspari Nutrition “Vasotropin” è un ottimo prodotto).
45 minuti pre-workout: 15UI di Humulin R.
20 minuti pre-workout: 50g di carboidrati ad alto peso molecolare (ex: Vitargo, Karbolyn, etc). 20g di proteine idrolizzate (es: Hydrowhey, Carnivore). 20g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizata. 2g grammi di Beta-Alanina. 10g di Glutammina. 3g di Taurina. 500mg di Potassio. 1g di Vitamina C.
75 minuti dopo il 1° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 10g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizzata. 2g Beta-Alanina. 10g di Glutammina. 3g di Taurina.
75 minuti dopo il 2° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 3g di Leucina. 5g di Glutammina.
Proteine totali: 60g (esclusi gli amminoacidi in forma libera aggiunti)
Carboidrati totali: 150g (escluse le trace di carboidrati contenute nelle polveri proteiche).
Prima di tutto, nel formulare il rapporto macros/Insulina sopra esposto, Arnold ha aumentato la quantità di carboidrati-proteine al di sopra di ciò che è tipicamente necessario per una UI di Insulina, al fine di tenere conto degli utilizzatori che dimostrano un grado superiore alla media di sensibilità all’Insulina. La maggior parte degli utilizzatori di Insulina, richiedono circa 8g di carboidrati-proteine per UI di Insulina, al fine di chiudere in pareggio e mantenere la normale soglia di glucosio nel sangue. Questo protocollo utilizza un rapporto 14:1 (macros/Insulina), che dovrebbe permettere a praticamente chiunque di utilizzare questo programma mantenendo il glucosio nel sangue all’interno di un range di normalità.
Va notato che questo programma è stato progettato per essere seguito “come è scritto”, soprattutto per quanto riguarda i tempi di assunzione dei nutrienti e le loro quantità. Per gli utilizzatori di Insulina già “navigati” che sanno quali rapporti sono ideali per loro, essi hanno la libertà di ridurre la quantità di macros consumati per UI, se necessario, come determinato dalla valutazione della loro risposta metabolica. Per gli utilizzatori inesperti, la componente nutrizionale del programma dovrebbe essere rispettata come scritta per almeno 2 settimane, a questo punto l’utilizzatore può quindi iniziare a personalizzare il suo rapporto macros/Insulina, se necessario.
La base di questo programma poggia sul tipo specifico di macros utilizzati. Senza di loro, ogni altro componente/aspetto del programma è influenzato negativamente e in alcuni casi rende il tutto sensibilmente limitato negli effetti. Carboidrati ad alto peso molecolare, come il Vitargo o le Ciclodestrine Altamente Ramificate, hanno dimostrato di essere superiori ad altre forme di carboidrati in diversi modi, per esempio un rapido e costante rilascio di glucosio nel sangue e una bassissima osmolaritá in soluzione.
Passando alla componente proteica; le proteine idrolizzate sono molto più velocemente assorbite rispetto ad altri tipi di proteine e sono l’unica proteina che può essere consumata insieme ai carboidrati ad alto peso molecolare senza compromettere il loro assorbimento.
In questo protocollo vi è anche la possibilità di un aggiunta di uno stimolatore del NO a scelta. L’aggiunta di questi stimolatori (vedi, ad esempio, la Citrullina Malato), pur non “necessario”, aumenterà ulteriormente la circolazione e il trasporto dei nutrienti ai muscoli che lavorano, così come contribuiscono ad aumentare il pumping sperimentato durante e dopo l’allenamento. Il Glicerolo monostearato è incluso anche tra gli ingredienti per il suo ruolo di volumizzante muscolare. Questo composto viene spesso utilizzato appena prima della gara, al fine di contribuire al raggiungimento di uno aspetto pieno e asciutto durante l’esibizione. Sono presenti anche dei volumizatori tradizionali, come la Glutammina, la Taurina, la Creatina, e il Potassio.
Al fine di promuovere un maggiore recupero e una maggiore risposta per una crescita muscolare, la temporizzazione dell’assunzione degli shake è stata messa a punto per mantenere un flusso costante dei nutrienti per tutta la vita attiva dell’Insulina. L’Humulin R è stata appositamente scelta per questo scopo, dal momento che con la sua emivita permetterà all’utilizzatore di sfruttare le “finestre” sia intra che post-allenamento. L’Humulin R offre anche un picco di Insulina meno pronunciato, che risulta più facile da gestire per una buona parte degli utilizzatori rispetto ad una versione di Insulina ad azione più rapida, come ad esempio l’Humalog che, però, viene in alcuni casi sostituita alla scelta classica.
Per via del tempo di esposizione all’Insulina limitato con la pratica di questo protocollo, la sensibilità all’Insulina è solo moderatamente influenzata in modo diretto quando si utilizza il programma per circa 5-6 volte alla settimana. Per gli individui che scelgono di utilizzare il presente protocollo per 3-4 volte alla settimana, le alterazioni dirette sulla sensibilità all’Insulina non risulta essere un problema. Per coloro che eseguono il protocollo per 5-6 volte a settimana, possono intervenire in due modi per assicurare una pienamente ottimale sensibilità all’insulina:
L’utilizzatore può fare 2 settimane “off” ogni 4 settimane “on”.
L’individuo può aggiungere Metformina nel suo programma di 3-4 volte a settimana ad un dosaggio di 750mg-1g/die.
Per gli utilizzatori che si apprestano all’uso di questo protocollo per la prima volta, mentre il rapporto macros / Insulina di cui sopra è sufficiente, l’autore consiglia sempre di iniziare con un dosaggio ridotto e aumentarlo poco a poco fino al raggiungimento del pieno dosaggio. Per uno novizio, un dosaggio di 6-8UI è ideale. Questo può essere seguito da una seconda iniezione da 8-10UI, per poi passare ad una terza iniezione di 10-12UI prima di arrivare al dosaggio massimo di 15UI.
Il protocollo in questione non contempla il solo uso di “shake e insulina” ma anche di GH (nel pre-workout a distanza dall’Insulina) e del IGF-1lr3 (nel post workout).
Il protocollo inizia con un’assunzione di GH, circa 30 minuti prima del workout per far si che i livelli plasmatici siano ragionevolmente alti, prima di aggiungere la dose di Insulina. L’idea alla base di questo, è assicurarsi che il GH passi attraverso il fegato mentre si ha una notevole quantità di Insulina in circolo. Questo è il modo in cui produciamo grandi picchi di IGF-1. Dopo l’allenamento, si somministra l’IGF-1LR3.
Il protocollo esemplificativo è il seguente:
30 minuti pre-workout: 6-10UI di GH subq
15 minuti pre-workout: 6-16UI di Novalog subq
10 minuti pre-workout: assumere lo shake #1
Dopo ogni set eseguito: sorseggiare lo shake #2, e terminarlo entro la fine dell’allenamento.
Andare a casa
Somministrare la dose di 100mcg di IGF-1lr3 (per i suoi effetti sulla sensibilità all’Insulina
Assumere lo shake #3
Formulazione degli shake:
Shake 1: 10-20g di EAA (Amino Acidi Essenziali) o PeptoPro, 40-60g Vitargo, 5g Creatina Monoidrata Micronizata, 200mg di Caffeina (migliora la resintesi di glicogeno); anche in questo caso la dose di carboidrati per ogni UI si aggira in media sui 7-10g.
Shake 2: 10-20g EAA o PeptoPro, 50-100g Vitargo, 5g di Carnitina Monoidrata Micronizata.
Shake 3: 2 tazze di albume pastorizzato, 1 tazza di avena istantanea, 1 banana o 1 tazza di mirtilli, Splenda o Stevia.
La somministrazione di Insulina pre-workout trova la sua motivazione d’essere nel fatto che durante l’allenamento con i pesi si viene a creare uno stress meccanico che a sua volta crea una sovra regolazione dello stimolo anabolico. Più comunemente, questo stato iper-anabolizzante viene indicato come “finestra anabolica”; un determinato periodo di tempo che dura dall’inizio dell’allenamento fino a poche ore dopo. Un modo in cui il corpo reagisce all’allenamento con i pesi è attraverso un aumento della sensibilità all’Insulina. Questo accade quando i recettori per l’Insulina, che risiedono sulla superficie della cellula, rispondono al segnale dell’Insulina in modo più efficiente, cosa che ci permette un migliore assorbimento dei nutrienti all’interno della cellula. Oltre a ciò, avviene un aumento della proliferazione dei GLUT-4 sulla superficie cellulare in maniera insulino-indipendente, caratteristica che richiede una minore quantità di Insulina per avere i massimi effetti nell’uptake cellulare.
Infatti, l’allenamento promuove anche il recupero e la crescita a livello intracellulare aumentando la sintesi proteica, la glicogeno sintasi, dei GLUT4 e l’espressione del trasportatore degli aminoacidi, e diminuendo i livelli di Miostatina. In combinazione con un aumento della sensibilità all’Insulina, queste cose non solo si traducono in una crescita accelerata, ma forniscono un effetto di ri-partizionamento dei nutrienti, in cui il cibo che assumiamo e i macronutrienti ivi contenuti vengono indirizzati maggiormente verso il miocita (cellule muscolari), piuttosto che immagazzinati come grasso.
L’Insulina è il complemento perfetto per questa “finestra anabolica” durante l’allenamento, in quanto non solo i nutrienti vengono trasportati alle cellule muscolari, permettendo così al corpo di approfittare di questo stato anabolizzante intensificato, ma l’Insulina agisce anche per molti dei processi di costruzione muscolare, fornendo uno stimolo ipertrofico raddoppiato nel momento in cui il corpo è più sensibile alla risposta del segnale dell’Insulina.
Come anti-catabolico, e lo abbiamo anche visto nella review presente nella terza parte di questa serie di articoli, l’Insulina è decisamente efficace, riducendo sensibilmente la degradazione del tessuto muscolare che si somma attivamente con la ripartizione delle proteine muscolari, e il tempo di recupero risulta ridotto. Questo ha un duplice effetto:
alterazione in positivo del rapporto catabolismo:anabolismo e maggiore supercompensazione (crescita muscolare);
il recupero più rapido si traduce in un aumento della possibile frequenza di allenamento, permettendo ai muscoli di essere stimolati più volte entro un determinato periodo di tempo e, in definitiva, di crescere più rapidamente;
infine, le concentrazioni di Insulina sovrafisiologiche aumentano in modo significativo il volume ematico all’interno nel tessuto muscolare, aggiungendo un pump muscolare generale che di per se stimola la crescita.
Tuttavia, affinché il corpo possa mettere a frutto tutto questo, sono necessari lo stimolo allenante adeguato e i nutrienti giusti che devono essere presenti al momento giusto. A questo punto entra in gioco la nutrizione intra-allenamento presentata in precedenza.
Nonostante questo protocollo abbia rappresentato la migliore applicazione dell’uso di Insulina nel Bodybuilding, esso presenta delle lacune oltre che dei problemi tecnici che riducono la pienezza del potenziale e il margine di “sicurezza”.
Alcuni atleti hanno sperimentato una maggiore tendenza all’ipoglicemia durante i loro workout con protocolli di Insulina pre workout. Questo imprevisto è limitante oltre che decisamente pericoloso, specie durante un workout. E’ ovvio che il controllo della glicemia e il rispetto dei punti base del protocollo riducono le possibilità del verificarsi di casi ipoglicemici. Ma il rischio è comunque maggiore rispetto ad una condizione di trattamento base (assunzione classica ai pasti).
Inoltre, tiene poco in considerazione le reale farmacocinetica dei PEDs utilizzati riducendo le piene ed ottimali interazioni ed effetti additivi.
GH/Insulin Protocol – pre e post-workout:
In risposta alle lacune del protocollo di Arnold che abbiamo appena visto, ho realizzato una versione perfezionata denominata “GH/Insulin Protocol”.
Tale perfezionamento prende in considerazione in modo preciso sia la farmacocinetica che la farmacodinamica incrociata dei componenti Insulina e GH.
Ora, sappiamo che la curva di rilascio del hGH somministrato per via sottocutanea raggiunge un picco iniziale dopo 30 minuti post iniezione per poi attestarsi a 2-3h e subire un calo significativo dopo 4h dalla somministrazione. Lo strascico di IGF-1 perdura per circa 24h sopra il basale.
Sappiamo inoltre che l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1;3) l’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1.
Interazioni biologiche a livello ipofisario ed epatico tra Insulina, Ormone della Crescita (GH), Fattore di Crescita Insulino-Simile-I (IGF-I) e Proteine Leganti il Fattore di Crescita Insulino-Simile (IGFBPs). Le frecce aperte indicano la stimolazione e le linee nere sottili l’inibizione. Livelli elevati di Insulina (a destra) possono aumentare indirettamente la biodisponibilità di IGF-I (cerchi pieni) sopprimendo la produzione di IGFBP-1 e, in misura minore, di IGFBP-2 (simboli ombreggiati). A sua volta, l’aumento della biodisponibilità di IGF-I può aumentare l’effetto di feedback negativo sul GH (quadrato aperto), portando a una riduzione della secrezione di GH e a una minore produzione epatica di IGF-I e IGFBP-3. Tuttavia, livelli elevati di Insulina possono anche aumentare il numero e l’attività dei recettori epatici del GH (barra aperta), riflessi da un aumento dei livelli di Proteina Legante l’Ormone della Crescita (GHBP) circolante. Questo effetto può portare a un aumento della produzione epatica di IGF-I e IGFBP-3 regolata dal GH, con un aumento maggiore dei livelli di IGF-I circolante rispetto a quelli di IGFBP-3. Pertanto, insieme a fattori genetici, ormonali e ambientali, l’entità relativa di questi due effetti opposti dell’Insulina sulla produzione di IGF-I potrebbe determinare i livelli di IGF-I circolante. Nel tempo, un’eccessiva biodisponibilità di IGF-I potrebbe aumentare il rischio di cancro del colon-retto, favorendo la sopravvivenza di cellule trasformate e mutate che normalmente andrebbero incontro ad apoptosi.
Quindi, dal momento che l’obbiettivo è quello di creare un ambiente non fisiologicamente riproducibile senza alterazioni iatrogene al fine di ottenere la massima risposta anabolica dal protocollo, il primo punto di congiunzione, o meglio la chiave di volta del protocollo, deve essere l’incrocio del picco di hGH con quello dell’Insulina. Per fare ciò le modalità di somministrazione dovrebbero contemplare primariamente l’iniezione di hGH pre-workout e quella di Insulina (preferibilmente Humalog per via del picco raggiunto entro 15 minuti dalla somministrazione) nel post-workout. Un vantaggio aggiuntivo di questa modifica è la riduzione del rischio ipoglicemico durante la sessione di allenamento.
E no, il vantaggio dello shake intra-workout non viene perso dal momento che la ripartizione calorica è di per se ottimale per via di meccanismi insulino-indipendenti dati dall’attività contro-resistenza (vedi aumento dei GLUT-4 sulla superficie cellulare in seguito all’attività muscolare). Si veda anche l’assorbimento dilazionato dei composti facenti parte della soluzione ingerita e costituente lo shake intra-workout.
I punti chiave del protocollo sono i seguenti:
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8, in questo ultimo caso divise in una dose da 4UI pre workout e due da 2UI appena sveglio e prima di dormire, a secondo della modalità d’uso del hGH; se somministrato giornalmente o a giorni alterni. Vedi a tal proposito l’articolo dedicato alla somministrazione di hGH a giorni alterni];
1h pre-workout possibilità di assumere tra i 25 ed i 50mg di Sildenafil seguiti a 30 minuti dall’inizio del workout dall’assunzione di 8g di Citrullina Malato;
consumare uno shake intra-workout contenente di base 0,5-1g di carboidrati ad alto peso molecolare, 0,25g di proteine idrolizzate e 5g di Creatina Monoidrato;
somministrazione dell’Insulina (Humalog) post-workout [le UI vanno calibrate in base ai CHO consumati nel pasto post-workout; in linea di massima 1UI ogni 10Kg di peso]. L’Humalog può essere sostituita con l’Humulin-R se per la risposta del soggetto in quanto a tolleranza è migliore.
Schema esemplificativo dell’azione incrociata tra Insulina e GH a livello epatico e sue consequenziali principali aree di influenza ricercate.
Esiste una variante del suddetto protocollo la quale contempla l’uso di due tipi di Insulina: una base (Lantus) e una post-workout (Humalog):
calcolare la dose totale di Insulina giornaliera con la formula 1UI ogni 10Kg di peso;
dividere il totale della dose a metà tra Insulina Glargine (Lantus) e Insulina lispro (Humalog) o Humulin-R;
somministrazione di Insulina glargina (Lantus) dividendo la dose giornaliera in 2: la prima, pari al 65% della dose totale, al mattino in concomitanza con il primo pasto e la seconda, pari al 35% della dose totale, 12h dopo la prima somministrazione;
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8 a secondo della modalità d’uso del hGH];
somministrare la dose di Insulina lispro o Humulin-R nel post-workout.
L’uso di una base di Insulina rappresentata dalla Insulina glargina ha il potenziale di aumentare ulteriormente l’espressione del IGF-1 e, di conseguenza, la risultante anabolizzante del protocollo. Ma, ovviamente, i rischi di ipoglicemia con questa modifica protocollare sono maggiori.
Grafico esemplificativo delle curve ematiche di Insulina, hGH e IGF-1 nel protocollo con Insulina Glargine di base.
Il monitoraggio regolare della glicemia e il rispetto dei punti base del protocollo evitano con un buon margine il verificarsi di casi ipoglicemici. Quindi, il glucometro, che sia classico o con sensore, è essenziale in questi casi.
Repetita iuvant: QUESTO PROTOCOLLO COME I PRECEDENTEMENTE PRESENTATI, E L’USO DI INSULINA, SONO APPANNAGGIO DI ATLETI AVANZATI E MONITORATI DA PERSONALE QUALIFICATO! NULLA DI CIO’ CHE E’ STATO DESCRITTO DEVE ESSERE PRESO COME UNA PRESCRIZIONE MEDICA O UN CONSIGLIO!
Conclusioni “dopo un lungo viaggio”:
Siamo ora giunti alla conclusione di questa serie di articoli dedicati all’Insulina e al suo centesimo anniversario.
Durante questo lungo percorso abbiamo imparato a conoscere meglio questo affascinante e mal compreso peptide. Abbiamo visto come è stato scoperto, abbiamo compreso l’enorme passo avanti nella medicina e nella tutela della vita umana che la sua scoperta ha rappresentato, abbiamo compreso come essa sia fisiologicamente regolata e quali sono le sue reali azioni a livello sistemico ed abbiamo imparato a separare i luoghi comuni che vi aleggiano intorno dai fatti.
Con la terza parte molti sono rimasti delusi nel constatare che in fisiologia l’Insulina abbia un effetto prettamente anticatabolico, effetto preminente che mantiene anche se somministrata esogenamente entro i range fisiologici [<1.200ng/dl]. Altresì gli umori sono migliorati quando siamo venuti a conoscenza del fatto che non solo il dosaggio fa la differenza tra preminenza anticatabolica e anabolica ma anche la sensibilità.
Con la recente constatazione che il risultato dell’equazione “Insulina/anabolismo” cambi drasticamente in positivo se vi si aggiunge la variabile del hGH, abbiamo imparato che l’unico uso minimamente sensato dell’Insulina per il miglioramento della massa muscolo-scheletrica è in associazione con il peptide ipofisario con le ultime due modalità esposte.
Ci tengo però a precisare che l’uso dell’Insulina “off-label” dovrebbe rappresentare la componente più marginale nella carriera di un culturista. Vale a dire che se ne può benissimo fare a meno, in specie quando si è semplici amatori o agonisti di piccoli o medi circuiti competitivi. Non complicatevi la vita.
Fate tesoro delle nozioni che vi ho esposto affinché il confine della conoscenza si espanda e prevalga su quello dell’ignoranza.