Il muscolo scheletrico presenta una notevole complessità biochimica, plasticità fisiologica ed eterogeneità cellulare (Ohlendieck, 2010). È ben noto che l’allenamento contro-resistenza (RT) è la strategia interventistica non farmacologica più potente per ottenere un aumento delle dimensioni del muscolo scheletrico (American College of Sports Medicine, 2009). L’American College of Sports Medicine (2009) raccomanda l’allenamento contro-resistenza nell’intervallo delle 6-12 ripetizioni massime (RM) come ideale per aumentare l’ipertrofia muscolare. Tuttavia, Schoenfeld et al. (2017) hanno concluso che, sulla base delle evidenze attuali, il RT a basso carico (≤60% 1 RM) eseguito fino al cedimento muscolare momentaneo aumenta le dimensioni muscolari in modo simile al RT ad alto carico (>60% 1 RM). Va notato che queste conclusioni sono specifiche per le tecniche di imaging del muscolo intero (cioè ultrasuoni, risonanza magnetica e tomografia computerizzata); non è stato possibile eseguire una meta-analisi per la determinazione istologica diretta (cioè la biopsia muscolare) a causa della mancanza di studi che soddisfacessero i criteri di inclusione della review.
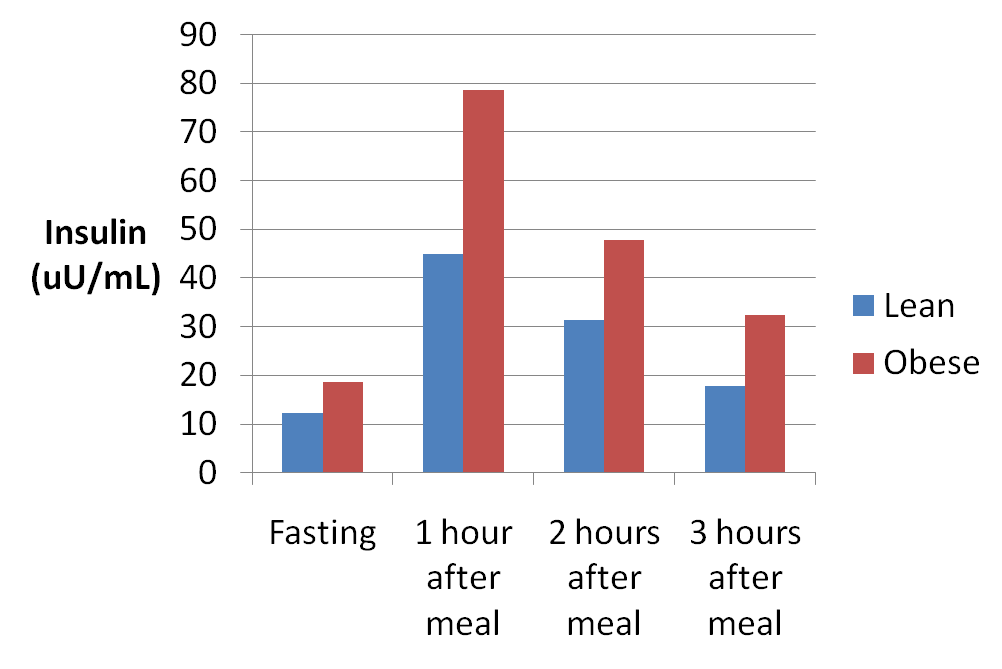
L’evidenza è equivoca per quanto riguarda l’accordo tra le tecniche di imaging del muscolo intero e la determinazione istologica dell’ipertrofia muscolare. Ad esempio, McCall et al. (1996) hanno riportato un aumento del 13% dell’area della sezione trasversale del muscolo (CSA) misurata tramite risonanza magnetica, insieme a un aumento del 10 e del 17% per le fibre muscolari di tipo I e II, rispettivamente. Mentre la meta-analisi di Schoenfeld et al. (2017) ha riportato cambiamenti simili nelle dimensioni del muscolo indipendentemente dagli schemi di carico, resta da chiedersi se l’ipertrofia delle fibre muscolari di tipo I e II sia simile anche tra condizioni di alto e basso carico. È possibile che il RT ad alto carico enfatizzi l’ipertrofia muscolare di tipo II (Campos et al., 2002), mentre il RT a basso carico stimoli una maggiore crescita delle fibre muscolari di tipo I (Ogborn e Schoenfeld, 2014; Grgic et al., 2018).
Diversi studi di elettromiografia (EMG) in acuto mostrano che l’ampiezza EMG è significativamente maggiore con il RT ad alto carico rispetto a quello a basso carico, suggerendo così che sono necessari carichi più elevati per stimolare completamente le unità motorie a soglia più alta associate alle fibre di tipo IIx (Schoenfeld et al., 2014, 2016). Tuttavia, una maggiore ampiezza EMG in una determinata condizione non riflette necessariamente gli adattamenti a lungo termine al RT regolato (per una rassegna dettagliata sull’argomento, si veda Vigotsky et al., 2018). L’unico modo per valutare questo argomento è analizzare gli studi che hanno confrontato direttamente il RT ad alto e basso carico e il suo impatto sull’ipertrofia delle fibre muscolari. Pertanto, il presente articolo riporta le informazioni contenute nella review di Jozo Grgic e Brad J. Schoenfeldsi nella quale sono stati discussi e interpretati gli studi che hanno valutato i cambiamenti delle fibre muscolari che si verificano longitudinalmente con schemi di RT ad alto e basso carico. Pur riconoscendo che esistono diverse fibre di tipo ibrido delineate nel muscolo scheletrico umano, in questo articolo verrà utilizzata la classificazione di base in tipo I, tipo IIa e tipo IIx.
Effetti dell’allenamento contro-resistenza ad alto o basso carico sull’adattamento all’ipertrofia delle fibre muscolari: Finora solo pochi studi hanno analizzato questo argomento (vedi tabella seguente). Jackson et al. (1990) hanno condotto uno studio crossover controbilanciato in cui i partecipanti hanno eseguito prima un mesociclo di 7,5 settimane di RT ad alto carico orientato alla forza muscolare o 7,5 settimane di RT a basso carico orientato alla resistenza muscolare. Dopo un periodo di “washout” di 5,5 settimane (cioè senza RT), i partecipanti sono passati a eseguire la routine alternativa. Dopo le prime 7,5 settimane, si è verificato un aumento delle dimensioni di tutti i tipi di fibre muscolari per entrambi i gruppi di RT. Dopo le seconde 7,5 settimane di RT, i partecipanti che hanno eseguito il mesociclo di forza muscolare come secondo trattamento hanno registrato un’ulteriore crescita delle fibre di tipo I e II. Al contrario, i partecipanti che hanno eseguito il RT orientato alla resistenza muscolare come secondo trattamento hanno mostrato una diminuzione delle dimensioni di tutti i tipi di fibre, con la diminuzione maggiore osservata per le fibre di tipo IIx. Questi risultati suggeriscono che una progressione da carichi bassi a carichi elevati potrebbe portare a guadagni più consistenti.
Sintesi dei risultati degli studi che confrontano l’allenamento ad alto e basso carico e i suoi effetti sugli adattamenti all’ipertrofia delle fibre muscolari.
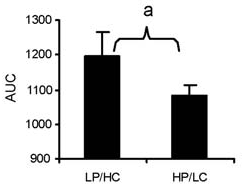
Solo uno studio è stato condotto su adulti anziani. Taaffe et al. (1996) hanno cercato di confrontare gli effetti del RT ad alto carico rispetto a quello a basso carico sulle dimensioni muscolari, utilizzando un intervento della durata di 1 anno. Le donne non allenate si sono allenate 3 giorni alla settimana; il gruppo ad alto carico ha eseguito dieci ripetizioni con carichi esterni dell’80% del 1 RM, mentre il gruppo a basso carico ha eseguito 14 ripetizioni con carichi corrispondenti al 40% del 1 RM. Valutando le variazioni percentuali, il gruppo ad alto carico ha ottenuto una maggiore ipertrofia sia nelle fibre muscolari di tipo I che in quelle di tipo II. In apparenza, questi risultati suggeriscono che il RT ad alto carico è un prerequisito per massimizzare l’ipertrofia in tutti i tipi di fibre. Tuttavia, i ricercatori hanno scelto di equiparare il carico volumetrico totale tra le condizioni, facendo interrompere al gruppo a basso carico ogni serie ben al di sotto dell’affaticamento volitivo. Tali eventi hanno inavvertitamente influenzato i risultati a favore della condizione ad alto carico, dato che un elevato livello di fatica è obbligatorio per ottenere benefici ipertrofici quando ci si allena con carichi inferiori (Morton et al., 2016).
Di fondamentale importanza è il fatto che in entrambi gli studi sopra citati, il programma di RT nei gruppi a basso carico non è stato eseguito fino al cedimento muscolare momentaneo, il che indica una disparità di stimoli di allenamento tra le condizioni di alto e basso carico. L’impiego di un programma di RT a basso carico e la mancata esecuzione di ripetizioni a cedimento muscolare momentaneo possono ostacolare l’adattamento muscolare (Burd et al., 2012). Secondo il principio delle dimensioni di Henneman (1985), le unità motorie più grandi saranno reclutate in sequenza all’aumentare dei requisiti di produzione di forza, portando infine all’attivazione dell’intero pool di unità motorie. È possibile che con l’allenamento a basso carico le unità motorie a soglia più bassa siano sottoposte a carico per un periodo più lungo, il che a sua volta potrebbe aumentare la risposta ipertrofica delle fibre muscolari di tipo I. Questo effetto potrebbe non verificarsi con l’allenamento ad alto carico e, forse per questo motivo, l’ipertrofia preferenziale delle fibre muscolari di tipo II è stata comunemente riportata con il RT (Folland e Williams, 2007).
Campos et al. (2002) sono stati i primi a confrontare il RT ad alto carico con quello a basso carico, con entrambi i gruppi che si allenavano fino al cedimento muscolare momentaneo, utilizzando misure istologiche della crescita muscolare. I ricercatori hanno randomizzato 27 partecipanti non allenati in tre diversi gruppi di carico: programmi di RT ad alto carico (3-5 RM), a carico moderato (9-11 RM) e a basso carico (20-28 RM). Dopo 8 settimane di RT, tutti i tipi di fibre muscolari si sono ipertrofizzati nei gruppi ad alto e moderato carico. Tuttavia, nel gruppo a basso carico non è stato osservato un aumento significativo della fCSA muscolare prima e dopo l’intervento nelle fibre muscolari di tipo I o II. Schuenke et al. (2012) hanno riportato risultati simili a quelli di Campos et al. (2002). Dopo 6 settimane di RT per la parte inferiore del corpo, il gruppo a basso carico (40-60% 1 RM) non ha mostrato aumenti significativi della CSA delle fibre muscolari di tipo I, mentre il gruppo ad alto carico (80-85% 1 RM) ha ottenuto robusti aumenti nelle dimensioni di tutti i tipi di fibre muscolari, con i maggiori guadagni osservati nelle fibre di tipo IIx. Sebbene il tessuto muscolare sia un ottimo esempio di plasticità tissutale e sia in grado di subire drastici cambiamenti nel profilo fenotipico con il RT regolato, il tasso di aumento delle dimensioni delle fibre muscolari nella condizione di carico elevato merita di essere esaminato (Burd et al., 2013). I ricercatori hanno riportato un tasso di crescita delle fibre muscolari dello 0,66% per giorno di allenamento, che è da 4 a 5 volte superiore ai valori presentati nella review di Wernbom et al. (2007). Tuttavia, sebbene non siano così elevati come quelli rilevati da Schuenke et al. (2012), tassi di crescita simili con il RT ad alto carico per la parte inferiore del corpo sono stati rilevati in precedenza in letteratura (Staron et al., 1990; Lamas et al., 2010).
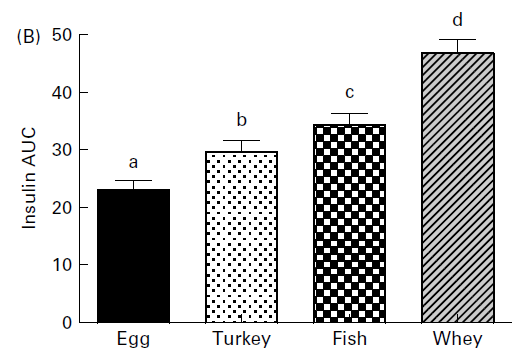
È interessante notare che alcuni studi riportano che il RT a basso carico induce una maggiore risposta ipertrofica nelle fibre muscolari di tipo I. Vinogradova et al. (2013) hanno confrontato il RT ad alto (80-85% di 1 RM) e a basso carico (50% di 1 RM) in un gruppo di giovani uomini non allenati. I risultati hanno indicato che la crescita dei tipi di fibre muscolari è direttamente correlata al carico di allenamento. In particolare, il gruppo ad alto carico ha ottenuto i maggiori aumenti delle dimensioni delle fibre muscolari di tipo II, mentre il gruppo a basso carico ha ottenuto i maggiori aumenti delle dimensioni delle fibre muscolari di tipo I. I ricercatori hanno ipotizzato che i maggiori aumenti delle fibre muscolari di tipo I nel gruppo a basso carico fossero legati a un maggiore stress metabolico (Vinogradova et al., 2013). Lo stress metabolico si riferisce all’accumulo di metaboliti, ad esempio all’aumento del flusso di calcio, del lattato, del potassio e degli ioni idrogeno, ed è un meccanismo ipotizzato per mediare l’ipertrofia muscolare attraverso un maggiore reclutamento di fibre, cambiamenti nella produzione ormonale e/o aumento del volume delle cellule per osmosi e/o re-sintesi glicogeno, tra gli altri (Schoenfeld, 2013). Risultati simili a quelli presentati da Vinogradova et al. (2013) sono stati riportati dallo stesso laboratorio, con un RT a basso carico che porta a un maggiore aumento dell’ipertrofia di tipo I e un RT ad alto carico che aumenta l’ipertrofia delle fibre di tipo II (Netreba et al., 2013).
Utilizzando un within-subject design, Mitchell et al. (2012) hanno assegnato casualmente 18 uomini in modo controbilanciato all’esecuzione di un’estensione unilaterale del ginocchio con una gamba che si allenava ad alto carico (80% di 1 RM) e l’altra a basso carico (30% di 1 RM). Dopo 10 settimane di RT, è stata osservata un’ipertrofia significativa dal basale al post-intervento in tutti i tipi di fibre muscolari per entrambi i gruppi di RT ad alto carico (80% di 1 RM) e a basso carico (30% di 1 RM). Non sono state rilevate differenze statisticamente significative tra le condizioni di carico nella crescita delle fibre muscolari. Tuttavia, è importante sottolineare che il gruppo a basso carico ha ottenuto un aumento del 23% delle dimensioni delle fibre muscolari di tipo I, rispetto a un aumento del 16% delle dimensioni delle fibre muscolari di tipo I osservato nel gruppo ad alto carico. Data l’esiguità del campione impiegato, ciò solleva la possibilità di un errore di tipo 2, in base al quale esistono effettivamente differenze significative tra le condizioni e, considerando l’entità delle differenze, tali risultati possono essere significativi dal punto di vista pratico.
Un limite comune a tutti gli studi citati è l’utilizzo di partecipanti non allenati. È stato dimostrato che gli individui con un RT più elevato sperimentano una risposta nella sintesi proteica attenuata dopo l’esercizio di resistenza, suggerendo un possibile effetto tetto del guadagno muscolare (Damas et al., 2015). Per colmare questa lacuna nella letteratura, Morton et al. (2016) hanno randomizzato giovani uomini con una media di 4 anni di esperienza di RT a un programma di esercizi total body di 12 settimane, utilizzando uno schema ad alto carico (8-12 RM) o a basso carico (20-25 RM). Dopo il protocollo di RT, sono stati osservati tassi di crescita simili in tutti i tipi di fibre muscolari, indipendentemente dalla condizione di carico. Questi risultati isolati suggeriscono che gli individui allenati possono sperimentare una crescita uguale in tutti i tipi di fibre, indipendentemente dallo schema di carico utilizzato.
Come per la maggior parte delle ricerche sulla RT, la stragrande maggioranza degli studi che hanno analizzato questo argomento sono stati di durata relativamente breve; non è chiaro se le potenziali differenze nell’ipertrofia dei tipi di fibre specifiche per un determinato schema di carico possano ampliarsi o ridursi nel tempo. Infine, tutti gli studi inclusi hanno valutato la crescita delle fibre nel muscolo vasto laterale. Come già evidenziato in precedenza, il muscolo vasto laterale è il muscolo più comunemente scelto per le biopsie a causa della sua composizione mista di fibre, della sua accessibilità e della sua allenabilità (Staron et al., 2000). Data la scarsità di dati, l’estrapolazione dei risultati ad altri gruppi muscolari è limitata.
Considerazioni sperimentali e conclusioni generali:
L’esame dei cambiamenti del tipo di fibra attraverso la biopsia muscolare presenta alcune limitazioni intrinseche. In primo luogo, la tecnica bioptica prevede l’estrazione di una piccola quantità di tessuto da un determinato muscolo, che potrebbe non riflettere necessariamente i cambiamenti specifici del tipo di fibra a livello dell’intero muscolo. Inoltre, esistono variazioni nella distribuzione dei tipi di fibre da superficiali a profonde e da prossimali a distali (Blomstrand e Ekblom, 1982), per cui è importante considerare questi fattori quando si estrapolano i risultati in termini pratici. In secondo luogo, il metodo utilizzato per determinare i tipi di fibre varia da uno studio all’altro. La maggior parte della letteratura recensita ha impiegato l’istochimica della mATPasi per delineare la tipizzazione delle fibre, mentre molti altri (Netreba et al., 2013; Vinogradova et al., 2013; Morton et al., 2016) hanno utilizzato anticorpi, che potrebbero avere una maggiore precisione nel delineare le fibre ibride. Infine, le differenze nella misurazione di fCSA possono comportare differenze nelle stime dimensionali, limitando così il confronto tra gli studi.
In conclusione si evince che, quando il RT a basso carico non viene eseguito fino al cedimento muscolare, l’allenamento ad alto carico sembra fornire uno stimolo ipertrofico superiore e quindi una maggiore crescita di tutte le fibre muscolari. Sebbene alcune prove indichino che il RT a basso carico, se effettuata fino al cedimento muscolare, possa indurre una maggiore risposta ipertrofica nelle fibre muscolari di tipo I rispetto al RT ad alto carico e che il RT ad alto carico possa indurre una crescita preferenziale delle fibre muscolari di tipo II, la letteratura rimane alquanto equivoca sull’argomento. In sintesi, attualmente non ci sono prove sufficienti per trarre conclusioni definitive sui cambiamenti che si verificano a livello delle fibre muscolari con i diversi schemi di carico.
Anche in questo caso, sembrerebbe che la formula ” in medio stat virtus” sia nuovamente la più indicata da applicare…
Per accedere alle precedenti parti, dalla prima alla terza, clicca qui, qui e qui.
Disclaimer: quelle che seguono sono informazioni provenienti da casi studio [tranne dove diversamente specificato] e testimonianze di Bodybuilder sull’uso dell’Insulina, alcune delle quali sono palesemente sbagliate. Non prendete assolutamente queste opinioni come consigli.
Insulina – dall’uso clinico al Bodybuinding:
Come abbiamo visto nella prima parte, inizialmente l’Insulina farmaceutica era di origine animale. In questo caso, l’Insulina viene estratta dal pancreas di suino o di mucca (o di entrambi) e preparata per uso medico. Queste preparazioni sono ulteriormente suddivise nelle categorie “standard” e “purificate”, a seconda del livello di purezza e del contenuto non insulinico della soluzione. Con questi preparati c’è sempre la possibilità che contaminanti pancreatici siano presenti nel farmaco.
Nel 1977, Herbert Boyer e i suoi collaboratori Keiichi Itakura e Arthur Riggs al City of Hope National Medical Center descrisse la prima sintesi ed espressione di un gene codificante per un peptide. Nell’agosto del 1978, Boyer produsse Insulina sintetica utilizzando i suoi nuovi batteri transgenici geneticamente modificati, seguita nel 1979 dall’Ormone della Crescita. La Tecnologia del DNA ricombinante faceva il suo debutto e cambiava la storia dell’Insulina per uso medico.
L’Insulina umana biosintetica (insulina umana rDNA), attualmente e maggiormente utilizzata per uso clinico, è prodotta con la tecnologia del DNA ricombinante. L’Insulina umana biosintetica ha una maggiore purezza rispetto all’Insulina animale estrattiva e riduce la formazione di anticorpi. I ricercatori sono riusciti a introdurre il gene dell’Insulina umana nelle piante come un altro metodo per produrre Insulina (“biopharming”) nel cartamo. Si prevede che questa tecnica ridurrà i costi di produzione.
Sono disponibili diversi analoghi dell’Insulina umana. Questi analoghi dell’Insulina sono strettamente correlati alla struttura dell’Insulina umana e sono stati sviluppati per aspetti specifici del controllo glicemico in termini di azione rapida (insuline prandiali) e azione prolungata (insuline basali). Il primo analogo biosintetico dell’insulina è stato sviluppato per l’uso clinico al momento del pasto (insulina prandiale), Humalog (Insulina lispro), è assorbita più rapidamente dopo l’iniezione sottocutanea rispetto all’Insulina normale, con un effetto a 15 minuti dopo l’iniezione. Altri analoghi ad azione rapida sono NovoRapid e Apidra, con profili simili. Tutti vengono assorbiti rapidamente grazie a sequenze aminoacidiche che riducono la formazione di dimeri ed esameri (le insuline monomeriche vengono assorbite più rapidamente). Le insuline ad azione rapida non richiedono l’intervallo iniezione-pasto precedentemente raccomandato per l’Insulina umana e le insuline animali. L’altro tipo è l’Insulina a lunga durata d’azione; la prima di queste è stata Lantus (Insulina glargine). Queste hanno un effetto costante per un periodo prolungato, da 18 a 24 ore. Allo stesso modo, un altro analogo dell’Insulina a lunga durata d’azione (Levemir) si basa su un approccio di acilazione degli acidi grassi. A questo analogo è legata una molecola di acido miristico, che associa la molecola di Insulina all’abbondante albumina sierica, prolungando così l’effetto e riducendo il rischio di ipoglicemia. Entrambi gli analoghi ad azione prolungata devono essere assunti una sola volta al giorno e sono utilizzati nei diabetici di tipo 1 come Insulina basale. È disponibile anche una combinazione di un’Insulina ad azione rapida e di un’Insulina protratta, che consente ai pazienti di ottenere un profilo insulinico simile a quello del rilascio di Insulina da parte dell’organismo. L’Insulina viene utilizzata anche in molte linee cellulari, come CHO-s, HEK 293 o Sf9, per la produzione di anticorpi monoclonali, vaccini virali e prodotti per la terapia genica.
L’Insulina viene solitamente somministrata sotto forma di iniezioni sottocutanee tramite siringhe monouso con aghi, tramite un microinfusore di Insulina o tramite penne da insulina a uso ripetuto con aghi monouso. Sul mercato statunitense è disponibile anche l’Insulina per inalazione.
A differenza di molti farmaci, l’Insulina non può essere assunta per bocca perché, come quasi tutte le altre proteine introdotte nel tratto gastrointestinale, si riduce in frammenti amminoacidici, perdendo tutto il suo potenziale di attività. Sono state condotte alcune ricerche su come proteggere l’Insulina dal tratto digestivo, in modo da poterla somministrare per via orale o sublinguale.
Nel 2021, l’Organizzazione Mondiale della Sanità ha aggiunto l’Insulina al suo modello di elenco di farmaci essenziali.
Complice la descrizione iniziale allettante riportata in letteratura riguardante l’azione dell’Insulina sul metabolismo glucidico e proteico, dove tale peptide veniva descritto come l'”ormone più anabolico”, unita alla maggiore disponibilità di approvvigionamento del farmaco data dalla Tecnologia del DNA ricombinante, nel mondo della cultura fisica di alto livello non mancarono i primi pionieri del suo utilizzo “”off-label”.
Si può stimare che nel giro di 40 anni, vale a dire dagli anni 80 ad oggi, l’uso dell’Insulina nel Bodybuilding abbia subito sia un abuso pratico che teorico. Quello che spero di fare con questa mia piccola opera divulgativa è proprio quello di cambiare questa situazione. Come uomo di scienza con anni di ricerca alle spalle sono qualificato per giudicare le conoscenze dei Bodybuilder. Molti di loro conoscono molto meno di me il funzionamento e l’uso dell’Insulina. Quello che porto sul tavolo sono anni di ricerca nella comunità del bodybuilding e una corposo serie di prospettive diverse di culturisti sull’uso dell’Insulina, raccolte da 20 interviste con utilizzatori di Insulina con un’esperienza che va da mesi a decenni. Tranquilli però, fortunatamente non sono un “classico camicie bianco”, ma uno che analizza con attenzione è sa ammettere quando gli atleti hanno ragione e la ricerca scientifica si sbaglia. È ora quindi di trasformare la “broscience” dell’Insulina in scienza vera e propria e di correggere alcuni miti potenzialmente pericolosi.
Sto scrivendo questo articolo non solo per dimostrare le diverse, e persino contraddittorie, opinioni che i bodybuilder hanno sull’insulina, ma anche per ispirarvi a riflettere lucidamente.
I culturisti, quelli con doti intellettive un minimo sopra la media dei loro colleghi, sanno cose che i medici non potrebbero mai sapere perché hanno un obiettivo diverso e priorità diverse. Ma una cosa è certa, i bodybuilder accorti vogliono praticare il bodybuilding in modo scientificamente informato.
Non saranno presenti i nomi dei culturisti intervistati, dei quali sono stati esaminati i video e di cui sono state raccolte le affermazioni nei forum, per un principio etico che prevede di non rivelare l’identità delle persone che hanno contribuito alla presente ricerca, per evitare qualsiasi rischio di danno alla loro persona e immagine. Pertanto, tutti i nomi sono pseudonimi. È sufficiente dire che sono stati intervistati alcuni individui riconosciuti come esperti mondiali nell’uso dell’Insulina per il bodybuilding. sono stati inclusi anche i bodybuilder medi.
L’Insulina vista dai BodyBuilder:
I bodybuilder hanno tra loro visioni piuttosto diverse riguardo all’Insulina e ai suoi effetti. Per esempio, essi non sono d’accordo all’unanimità su quanto sia anabolizzante l’Insulina. Una minoranza afferma che l’Insulina esogena non è direttamente anabolizzante. Alcuni suggeriscono che l’Insulina induce l’anabolismo solo aumentando l’appetito. Ma, come sappiamo bene, di per se, quest’ultima affermazione ha ben poco senso.
Una piccolissima minoranza di bodybuilder sostiene che l’impatto dell’Insulina esogena sia principalmente, o puramente, cosmetico, in quanto l’Insulina fa apparire il muscolo più “pieno” (piuttosto che aumentare effettivamente le dimensioni del muscolo) aumentando la ritenzione idrica intracellulare.
Al contrario, molti bodybuilder sostengono che l’Insulina sia l’ormone più anabolico.
Alcuni partecipanti hanno descritto l’uso dell’Insulina come il risultato di un aumento muscolare di 3-6kg a settimana rispetto a quello che si potrebbe ottenere con il solo uso di Steroidi Androgeni Anabolizzanti (AAS) o con l’uso combinato di AAS e Ormone della Crescita (GH). Tuttavia, queste affermazioni sono state contestate da alcuni. Vi sono bodybuilder che ritengono che i benefici dell’Insulina non siano sufficienti e non hanno intenzione di utilizzare nuovamente questo peptide. Uno di questi non raccomanda più l’uso dell’Insulina ai suoi clienti.
Mentre alcuni bodybuilder sostengono che l’Insulina da sola sia anabolizzante, altri suggeriscono che essa sia significativamente anabolizzante solo in sinergia con AAS e hGH.
La maggior parte dei bodybuilder concorda sul funzionamento dell’Insulina, anche se le loro spiegazioni sono più o meno scientifiche.
In genere i bodybuilder descrivono l’Insulina come una “navetta” o un “mezzo” che trasporta i nutrienti nelle cellule muscolari. L’Insulina viene descritta come una “chiave” che apre le porte delle cellule o come un “autobus” che trasporta i nutrienti. Alcuni bodybuilder citano la letteratura scientifica quando descrivono l’azione dell’Insulina:
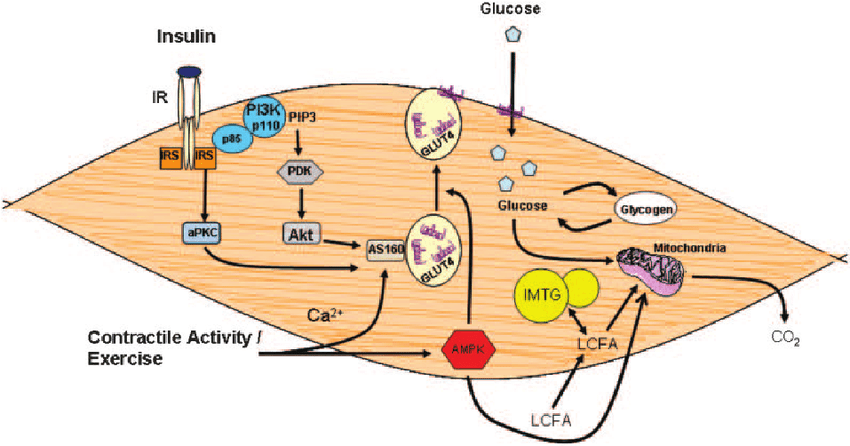
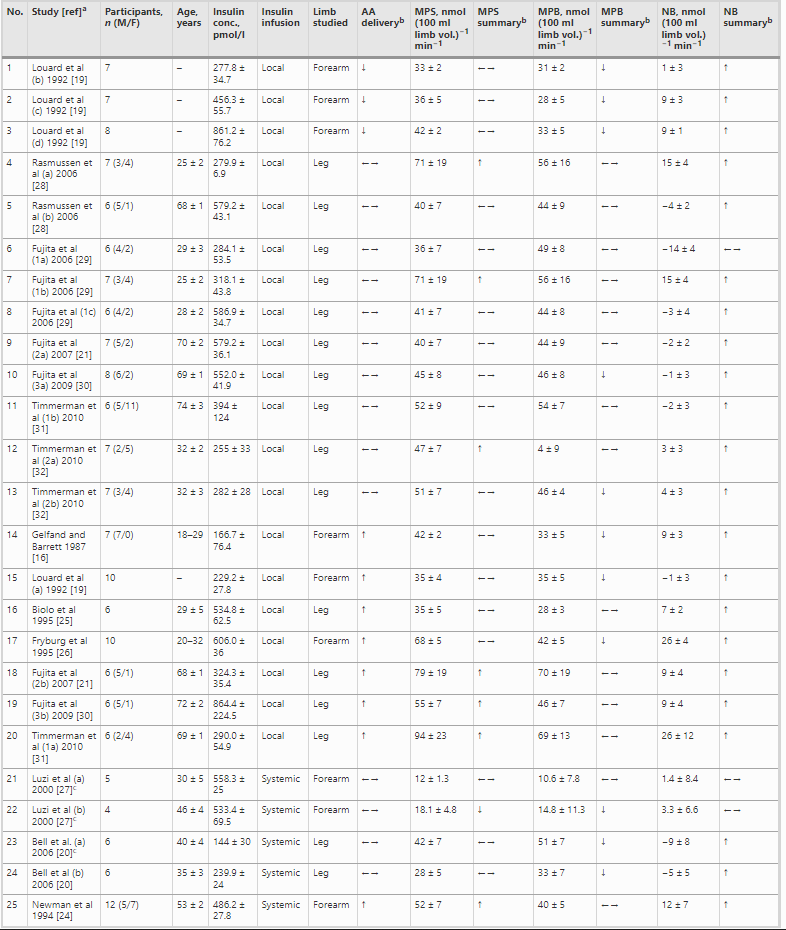
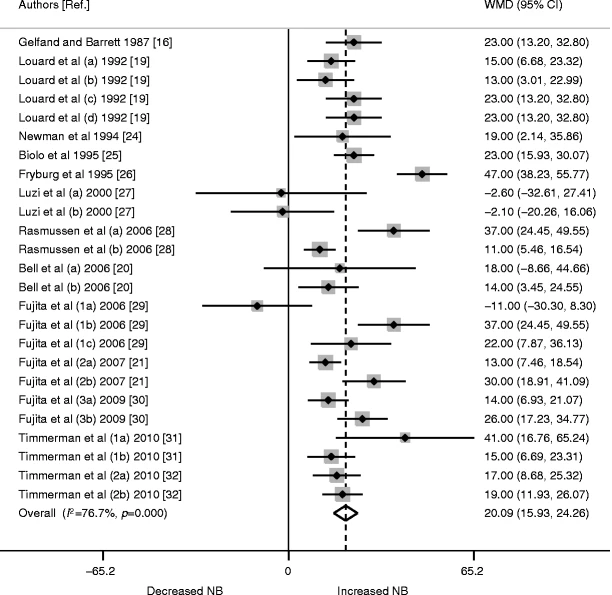
Sembra che l’Insulina abbia almeno un effetto permissivo sulla sintesi proteica, tanto che i suoi livelli basali sono necessari per la normale sintesi proteica miofibrillare (MPS), ma l’aumento dell’Insulina dopo un pasto potrebbe non aumentare la MPS (Greenhaff et al., 2008). Tuttavia, l’Insulina promuove l’anabolismo muscolare (bilancio proteico proattivo) attraverso il suo effetto inibitorio sulla degradazione delle proteine muscolari (MPB) (Deutz e Wolf 2013). Inoltre, l’Insulina può aumentare l’MPS attraverso un maggiore assorbimento di aminoacidi (essenziali) nel muscolo scheletrico, provocato da un aumento del flusso sanguigno associato alla vasodilatazione (Biolo et al., 1995; Fujita et al., 2006; Timmerman et al., 2010). [Chad via e-mail].
Molti concordano sul fatto che:
l’Insulina è molto efficace nel trasportare i nutrienti nelle cellule muscolari, ma anche nelle cellule adipose.
l’Insulina è anabolizzante grazie al suo ruolo nella ripartizione dei nutrienti, in quanto lavora di concerto con GH e IGF-I.
l’Insulina promuove l’anabolismo muscolare grazie al suo effetto inibitorio sulla degradazione delle proteine muscolari.
l’Insulina (così come gli AAS e il GH) promuove la sintesi proteica muscolare solo in presenza di un adeguato apporto di aminoacidi.
l’Insulina svolge un ruolo nel controllo fisiologico della riproduzione, agendo sulla secrezione dell’Ormone di Rilascio delle Gonadotropine (GnRH)/luteinizzante (LH).
L’Insulina è anche descritta da alcuni bodybuilder come anti-catabolica.
Un bodybuilder di alto livello ha dichiarato che l’Insulina dovrebbe essere usata solo se un individuo è carente di Insulina:
A volte è una buona idea prendere l’Insulina solo per aiutare il pancreas a fare il suo lavoro. … se il vostro corpo producesse abbastanza da solo, perché avreste bisogno di assumere Insulina esogena? In altre parole, non apporta alcun beneficio. È utile solo se non si produce abbastanza Insulina. Quindi bisogna innanzitutto stabilire se non si produce abbastanza Insulina. Procuratevi un glucometro e controllate la glicemia. … Quindi l’Insulina dovrebbe essere usata solo se si ha una carenza di insulina, perché si sta usando molto GH o perché si sta mangiando una quantità esorbitante di carboidrati.
Definirei questa ipotesi come “patologica indotta/deficitaria”. Ricordatevi sempre che l’omeostasi organica è regolata da feedback. Ciò significa che l’uso di Insulina esogena causerà una sottoregolazione/inibizione della biosintesi endogena di Insiluna. Di conseguenza, parlare di “pancreas ipoattivi” o “supporto pancreatico” non è in definitiva corretto. In tal caso si parla di una vera e propria terapia ormonale sostitutiva dell’Insulina.
Un bodybuilder ha anche affermato che gli effetti anabolici dell’Insulina sono dovuti alla sua azione osmotica e alla sua capacità di aprire “tutti i recettori del corpo”. Descrizione alquanto particolare ma che può rendere una certa idea di uno degli effetti dell’Insulina.
Mentre alcuni bodybuilder mettono in guardia dall’uso dell’Insulina perché può causare ipoglicemia con conseguente coma ipoglicemico potenzialmente letale, la maggior parte ritiene che i rischi dell’Insulina siano stati sopravvalutati e alcuni suggeriscono che la morte dovuta all’uso di Insulina è estremamente improbabile. Molti pensano che “bisogna essere una testa di cazzo per uccidersi con l’Insulina”.
Sebbene la morte di diversi culturisti di alto livello nel corso della mia ricerca sia stata inizialmente suggerita da membri della comunità dei culturisti come correlata all’Insulina, l’Insulina non è stata implicata nelle cause ufficiali dei loro decessi. Non mi sono imbattuto in un caso confermato di morte o di danni significativi causati dall’uso di Insulina per il bodybuilding, anche se alcuni bodybuilder hanno dichiarato di conoscere qualcuno che è morto a causa dell’uso di Insulina (e uno di loro ha avuto un grave incidente d’auto a causa di una ipoglicemia avuta in autostrada, ma fortunatamente nessuno è rimasto ferito). Nella letteratura medica sono riportati due casi di bodybuilder in coma ipoglicemico (Heidet et al., 2019; Petrovic et al., 2015). Non credo che la mancanza di decessi confermati sia dovuta al fatto che l’Insulina non sia pericolosa, ma più probabilmente perché non è comunemente testata o difficile da rilevare.
Un bodybuilder ha suggerito che le morti premature dovute all’uso di Insulina potrebbero non essere dovute solo all’ipoglicemia, ma ha suggerito che l’aumento dei livelli di Insulina nel corso della vita accorcia la durata della stessa e che quindi i bodybuilder si mettono a rischio in questo senso.
Alcuni bodybuilder sostengono che l’Insulina sia uno dei farmaci più sicuri del loro arsenale, in particolare rispetto al DNP e al Trenbolone. Alcuni suggeriscono addirittura che l’Insulina sia più sicura di qualsiasi AAS.
Mentre tutti i bodybuilder hanno descritto almeno lievi sintomi di ipoglicemia in alcuni momenti del loro utilizzo di Insulina, la maggior parte degli episodi di ipoglicemia si sono verificati durante le prime fasi di utilizzo, quando stavano elaborando il dosaggio dell’Insulina, o sono stati attribuiti alla loro stupidità (ad esempio, dimenticando di mangiare). Tutti i bodybuilder hanno dichiarato che l’ipoglicemia era facilmente gestibile consumando zuccheri.
Molti suggeriscono che per essere competitivi come bodybuilder professionisti è necessario utilizzare l’Insulina. Tuttavia, altri suggeriscono che non è necessario.
L’Insulina fa ormai parte del bodybuilding, ne è parte integrante. È come i denti sbiancati. Tutti sbiancano i denti, tutti hanno denti bianchi e splendenti. Se c’è uno che non li ha, gli si chiede: “Cosa c’è che non va in te?”. … Se non fai l’Insulina, cosa che alcuni professionisti non fanno, alcuni non ne hanno bisogno, allora il tuo aspetto è un po’ diverso da quello degli altri bodybuilder e potresti distinguerti in modo negativo.
Molti attribuiscono il significativo aumento della muscolatura dei mostri di massa all’Insulina e/o all’Ormone della Crescita. Alcuni suggeriscono che l’Insulina abbia rovinato il bodybuilding, poiché l’attenzione si è spostata dall’estetica alle dimensioni a scapito dell’estetica. Altri criticano ulteriormente il look dell’Insulina:
Prima dell’arrivo dell’Insulina, tornando ai primi tempi di Bertil Fox, Tom Platts, Arnold, Sergio, i loro muscoli avevano questo aspetto duro e granitico, sembravano scolpiti nella pietra. Ora ci sono questi ragazzi, certo grandi e stravaganti come i Ramy e tutti questi ragazzi, sono grandi ma non hanno quell’aspetto duro e denso.
Ma altri suggeriscono che Dorian Yates è stato il primo a portare sul palcoscenico del Olympia un fisico potenziato dall’Insulina e viene spesso descritto come se avesse un aspetto granitico.
Alcuni suggeriscono che l’Insulina (e/o l’Ormone della Crescita) provochi la “bolla intestinale” o il “palumboismo” [vedi anche “GH Gut”], e una fonte ha affermato che l’Insulina ha causato l’organomegalia. Per questi motivi alcuni affermano che l’Insulina ha rovinato l’estetica del bodybuilding.
Il dosaggio dell’insulina è molto vario tra i bodybuilder. Ho parlato con culturisti che usano un massimo di 4UI al giorno, e altri che hanno usato un massimo di 360UI al giorno! Anche se lo considero ben poco credibile. Tuttavia, in linea con le precedenti ricerche accademiche che riportavano dosaggi compresi tra 10 e 20 unità al giorno (Dawson e Harrison 1997; Evans 1997; Hildebrandt et al., 2007) e con i sondaggi condotti all’interno della comunità, ho scoperto che la maggior parte dei bodybuilder utilizza dosi che si collocano all’estremità inferiore dello spettro. Tuttavia, una buona parte dei bodybuilder tende a usare più di quanto riportato in precedenza nella letteratura accademica e nella comunità, con una dose giornaliera mediana di 40 unità e una dose mediana di 0,39 unità per chilogrammo di peso corporeo.
La maggior parte dei bodybuilder utilizza una sola forma di insulina (ad azione rapida o ultra-rapida), anche se un quarto degli intervistati (n=20) utilizza anche un’Insulina a lunga durata d’azione.
Ma vediamo nel dettaglio i tipi di Insulina utilizzati:
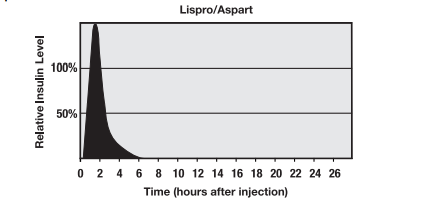
Humalog® (Insulina Lispro): Humalog® è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”.
Novolog® (Insulina Aspart):Novolog è un analogo a breve durata d’azione dell’Insulina umana, creato quando l’aminoacido prolina in posizione B28 viene sostituito con l’acido aspartico. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 15 minuti e l’effetto di picco si raggiunge in 1-3 ore. La durata d’azione totale è compresa tra le 3 e le 5 ore. L’Insulina Aspart viene solitamente utilizzata come supporto a un prodotto contenente insulina a più lunga durata d’azione, fornendo un farmaco a rapida azione che può essere assunto prima o subito dopo i pasti per imitare la risposta insulinica dell’organismo. Molti atleti ritengono che la sua breve finestra di effetto la renda ideale per scopi dopanti, tanto quanto la Lispro, in quanto la maggior parte della sua azione si può concentra nel periodo successivo all’allenamento durante la “finestra anabolica”.
Humulin®-R “Regular” (insulina Inj): Identica all’Insulina umana. Venduta in alcuni mercati anche come Humulin-S® (Solubile), questo prodotto è costituito da cristalli di zinco-insulina disciolti in un liquido chiaro. Non viene aggiunto nulla per rallentare il rilascio di questo prodotto, per cui viene genericamente indicato come insulina umana solubile. Questo farmaco agisce rapidamente e ha una breve durata d’azione. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di 20-30 minuti, e il suo picco d’effetto si raggiunge in 1-3 ore. Ha una durata d’azione totale tra le 5 e le 8 ore. Insieme a Humalog, queste due forme di Insulina sono le scelte più popolari tra gli atleti e i culturisti per scopi dopanti.
Humulin®-N, NPH (Insulina Isofana): Una sospensione cristallina di Insulina con protamina e zinco per ritardarne il rilascio e prolungarne l’azione. L’Insulina Isofana è considerata un’Insulina di lunghezza intermedia. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore e il picco d’effetto si raggiunge in 4-10 ore. La durata totale dell’attività è superiore a 14 ore. Questo tipo di Insulina non è comunemente usata come agente dopante.
Humulin®-L, Lente (sospensione media di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-L è considerata un’insulina di lunghezza d’azione intermedia. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 1-3 ore e l’effetto di picco viene raggiunto in 6-14 ore. Ha una durata totale di attività superiore alle 20 ore. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin®-U, Ultralente (sospensione prolungata di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-U è considerata un’Insulina a lunga durata d’azione. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 6 ore, e l’effetto di picco viene raggiunto in 14-18 ore. La durata totale dell’attività è di 18-24 ore. Questo tipo di insulina non è comunemente usato per scopi dopanti.
Lantus (Insulina Glargine): Analogo a lunga durata d’azione dell’Insulina umana. L’Insulina Glargine viene creata quando l’aminoacido asparagina in posizione A21 viene sostituito con la glicina e vengono aggiunte due arginine al C-terminale della catena B dell’Insulina. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore, e il farmaco è considerato privo di un picco significativo (è stato progettato per un modello di rilascio molto stabile per tutta la durata dell’attività). L’Insulina Glargine ha una durata d’azione compresa tra 20-24 ore nell’organismo dopo l’iniezione sottocutanea. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin® (Miscele) : Sono miscele di Insulina solubile normale per un’azione rapida, e di un’Insulina a lunga durata d’azione o ad azione intermetizzata per un effetto prolungato. Queste miscele sono etichettate con la percentuale di miscela, di solito 10/90, 20/80, 30/70, 40/60 e 50/50. Sono anche disponibili le miscele che utilizzano Humalog come Insulina ad azione rapida.
Indipendentemente dal tipo, tutte le insuline forniscono gli stessi effetti di base, e la durata di azione è la differenziazione primaria. Il trasporto dei nutrienti, l’aumento della sintesi proteica, la diminuzione del catabolismo proteico, l’aumento del IGF-1, l’aumento della biodisponibilità del IGF-1, e una maggiore vasodilatazioni sono i vantaggi più noti.
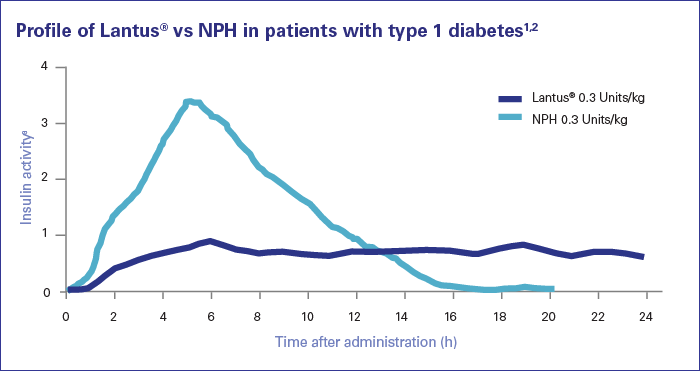
Non è da molto tempo che un tipo di Insulina precedentemente marginale a fini dopanti è diventata di moda tra alcuni culturisti. Sto parlando della Lantus (Insulina Glargine), appunto.
Di tutte le diverse insuline disponibili, la Lantus è probabilmente quella meno utilizzata anche perchè e paradossalmente la più complessa da gestire. Il suo scarso utilizzo nel bodybuilding ha portato, come ovvia conseguenza, ad una scarsità delle informazioni disponibili su di essa.
A differenza delle insuline a breve durata d’azione, che forniscono i benefici di cui sopra per poche ore al giorno, la Lantus continuerà il trasporto dei nutrienti, l’aumento della sintesi proteica, il miglioramento della vasodilatazioni, ecc, per tutto il giorno, anche mentre dormiamo, ed è questo ultimo punto a renderla di non facile gestione. Ma uno degli svantaggi principali della Lantus risiede nella sua possibilità di essere utilizzata solo per brevi periodi di tempo, in quanto l’esposizione continua a livelli elevati di Insulina esogena, e lo sappiamo bene, porterà ad una riduzione della sensibilità all’insulina, la successiva sotto-regolazione dei trasportatori GLUT-4, cose che si vorrebbero evitare sia da un punto di vista della saluta che della crescita muscolare. Così, mentre la Lantus può essere superiore per lo stimolo della crescita muscolare nel breve termine, troviamo che le cose cominciano a pareggiarsi nel lungo periodo, e protocolli come quelli pre-allenamento più comunemente impiegati possono invece essere utilizzati a tempo indeterminato, senza danneggiare eccessivamente la sensibilità all’Insulina a qualsiasi grado significativo. Questo rende la Lantus ideale per dei “blitz”, in cui l’atleta vuole mettere su muscoli il più rapidamente possibile, ma non è adatta per un uso prolungato.
Differenza nella curva di rilascio tra Humulin N e Lantus in pazienti con Diabete di Tipo I.
Una caratteristica decisamente interessante della Lantus è il suo effetto sul IGF-1 ed i suoi recettori. In diversi studi universitari, la Lantus ha dimostrato una maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi. È interessante notare che Levemir, l’unico altro analogo dell’Insulina ad azione prolungata sul mercato, mostra una ridotta affinità di legame ai recettori del IGF-1 umani. Questo mette la Lantus e la Levemir alle estremità opposte dello spettro in termini di affinità di legame. Mentre un aumento di IGF-1 vincolante è visto generalmente come una cosa positiva per la crescita muscolare, è stato il punto focale del dibattito in corso nella comunità medica per parecchi anni, per il fatto che alcuni studi hanno mostrato un aumento del rischio di cancro quando si usa la Lantus. Da allora, altri studi hanno confutato questa nozione, ma il dibattito continua, con la comunità medica riluttante a prendere una posizione in un modo o nell’altro.
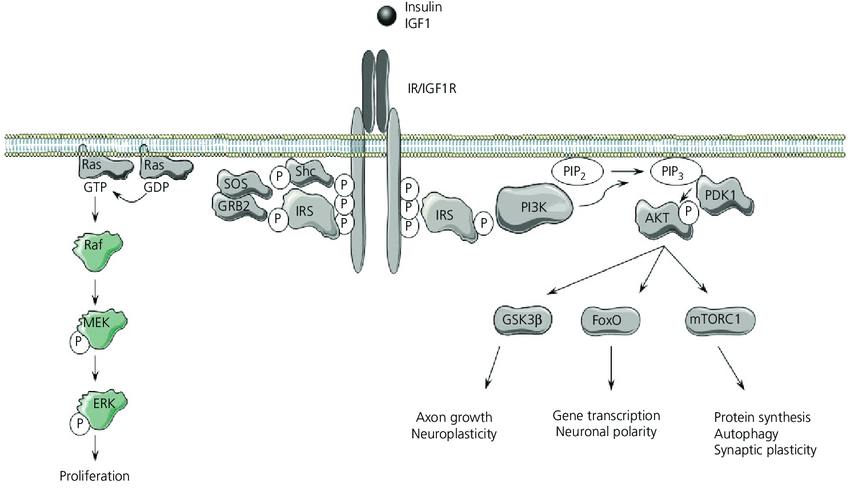
Via di segnalazione del Recettore dell’Insulina (IR) e del Recettore del Fattore di Crescita Insulino-Simile 1 (IGF1R). L’Insulina e l’IGF1 si legano ai loro recettori, inducendo un cambiamento conformazionale e l’autofosforilazione della subunità beta di IR e IGF1R. Successivamente, le proteine substrato del recettore dell’Insulina (IRS) o Shc vengono reclutate e fosforilate. Shc attiva la via della mitogen-activated protein kinase-extracellular signal regulated kinase (MAPK-ERK) e le proteine IRS inducono prevalentemente l’attivazione della via della fosfoinositide 3-chinasi (PI3K)-AKT. In questo caso, l’attivazione di PI3K causa la conversione del fosfatidilinositolo 4,5-bisfosfato (PIP 2 ) in fosfatidilinositolo (3,4,5)-trifosfato (PIP 3 ) e l’attivazione e la fosforilazione di AKT da parte della proteina chinasi 1 dipendente dal fosfoinositide. La regolazione dipendente da AKT della forkhead box O (FoxO), del mammalian target of rapamycin complex 1 (mTORC1) e della glicogeno sintasi chinasi 3b (GSK3b) regola la crescita degli assoni, la trascrizione genica, la sintesi proteica e la plasticità neuronale. MEK, MAPK/ERK chinasi; PDK1, proteina chinasi 1 fosfoinositide-dipendente; SOS, son-of-sevenless. [Adattato da Servier Medical Art di Servier, con licenza Creative CommonsAttribuzione 3.0 Unported].
In definitiva, la Lantus viene solitamente usata come “base” di un protocollo di Insulina affiancata dall’uso del Humalog nei protocolli di Insulina e GH pre o post workout.
UI:CHO ratio e timing di somministrazione
Sappiamo tutti che il rapporto più frequentemente citato tra carboidrati e insulina (UI:CHO ratio) è di 10-15g di carboidrati per 1UI di Insulina. Ma non tutti i bodybuilder si attengono a questo rapporto. Diversi culturisti non hanno stabilito un rapporto fisso tra carboidrati e Insulina, mentre altri hanno utilizzato una gamma di rapporti (5-20g per unità di Insulina) con una media di 9-10g di carboidrati per ogni UI di Insulina.
La maggior parte dei bodybuilder ha assunto l’Insulina durante i pasti, ma considerando che i bodybuilder mangiano spesso, questo non ci dice molto. C’è stato un grande dibattito sul momento più efficace per l’uso dell’Insulina: alcuni suggeriscono che il momento più efficace sia il pre-workout, mentre altri affermano che si tratta di un uso irresponsabile, in quanto è difficile determinare quanti carboidrati verranno ossidati durante l’allenamento e quindi l’uso dell’Insulina nel pre-allenamento potrebbe essere pericoloso. Alcuni bodybuilder usano l’Insulina prima e dopo l’allenamento, altri solo dopo.
Alcuni bodybuilder sostengono che l’uso di Insulina a scopo ricreativo espone i bodybuilder al rischio di sviluppare il diabete. Altri bodybuilder sostengono che l’uso dell’Insulina riduce il rischio di diabete in quanto diminuisce l’impatto delle diete per il bodybuilding. Lasciatemi dire che l’ultima affermazione non ha alcun senso. Il corpo mantiene una condizione di omeostasi attraverso elaborati feedback di controllo. La somministrazione di Insulina esogena causerà un feedback negativo a livello della secrezione endogena pancreatica, e l’Insulina esogena somministrata avrà il medesimo effetto a livello centrale e periferico in un regime alimentare ipercalorico (vedi “dieta per il bodybuilding”) dell’Insulina endogena! E le affermazioni secondo le quali l’uso dell’Insulina esogena sortirebbe un effetto di protezione all’affaticamento pancreatico beh, è un affermazione che non ha basi di riscontro.
La follia del protocollo “No Fat Gain Insulin Program”:
Diversi anni fa riportai un protocollo d’uso dell’Insulina denominato “No Fat Gain Insulin Program”. Questo protocollo “alternativo” fu ideato da Mike Zumpano e Oliver Starr i quali si chiesero se ci poteva essere una strategia che permettesse ai bodybuilder di non aumentare eccessivamente di bf durante l’uso di Insulina. In realtà, la motivazione di base per la quale molti culturisti diventano più grassi che grossi quando nelle loro preparazioni inseriscono l’Insulina è fondamentalmente la “la paura” dell’ipoglicemia. La maggior parte del guadagno di grasso è causato dal consumo eccessivo di carboidrati durante l’uso di Insulina. Una regola di “sicurezza” diffusa con l’uso di Insulina dice che bisogna consumare un minimo di 10-15g di carboidrati per ogni UI di Insulina utilizzata (distribuiti nell’arco di tempo d’azione dell’Insulina utilizzata). Un altro errore commesso da molti bodybuilder e che porta ad un eccesso calorico addizionale è che essi non calcolano le UI in base ai CHO della dieta ma calcolano le UI in base al peso e di conseguenza aggiungono i carboidrati di “sicurezza” a quelli già presenti nel loro programma alimentare. Comunque sia, un Bodybuilder che utilizza 8UI di Insulina 2 volte al giorno, in aggiunta al suo normale apporto di carboidrati, proteine e grassi andrà (con il metodo standard) a consumere una quota addizionale di carboidrati pari a 160g. Difficilmente ci si aspetta che 160g in più di carboidrati, o 640Kcal in più al giorno facciano una differenza significativa su un soggetto che magari mangia 5000 o più calorie al giorno.
Anche se la quantità di carboidrati supplementari (10gXUI) comunemente applicata non sembra terribilmente eccessiva, alcuni “pionieri” dei PEDs alla fine degli anni ‘90 erano certi che fosse il motivo principale per cui gli utilizzatori di Insulina guadagnavano quantità sproporzionate di grasso.
Il plasma umano contiene solo circa 5g di carboidrati in uno specifico momento. I diabetici che hanno preso troppa Insulina di solito possono riportare i loro livelli glicemici nel sangue nel range di normalità consumando cinque grammi (solo 20 calorie!) di Destrosio.
Comunque sia, il metodo “alternativo” lo trovarono e fu ribattezzato, come precedentemente accennato, protocollo “No Fat Gain”. Il trucco, se così possiamo definirlo, sarebbe quello di assumere l’Insulina, ma seguendo un dieta Low-Carb. Proprio così, Low. Con un contenuto glucidico di circa 50g al giorno. Oliver Starr, con rudimentali conoscenze in biochimica e fisiologia umana, si chiese se ci fosse qualche altro modo per mantenere la glicemia nel sangue moderata con un alto grado di stabilità. La sua (riduttiva) risposta è stata la gluconeogenesi. Se si guarda su un grafico dei processi biochimici, si può chiaramente vedere che, quando le riserve di glicogeno epatico e muscolare sono esaurite, ma prima che il soggetto vada in chetosi, il corpo comincia a convertire aminoacidi in glucosio per mantenere stabili i livelli di glucosio nel sangue. Questo processo è noto come gluconeogenesi. Come risaputo, seguire una dieta molto povera di carboidrati provoca un esaurimento delle riserve di glicogeno epatico e muscolare. Questo provoca un sovra-regolazione degli enzimi necessari per la conversione rapida ed efficace degli aminoacidi in glucosio. La parola gluconeogenesi significa letteralmente “la nascita di nuovo glucosio.”
La seconda metà dell’ipotesi applicata, ovviamente, è il contenuto proteico della dieta. Se non si mangiano molti carboidrati, l’unico modo con cui il corpo può produrre glucosio è principalmente quello di convertire gli aminoacidi in glucosio. Questo accade in una certa misura ogni volta che si mangiano proteine, tuttavia, quando si mangia una grande quantità di proteine, si viene a creare ancora più glucosio. E’ il livello di glucosio creato dall’eccesso di proteine che dovrebbe impedire il verificarsi di uno stato ipoglicemico in un contesto nel quale si utilizza Insulina esogena con una dieta a basso contenuto di carboidrati.
Per questo protocollo è necessaria l’assunzione di proteine in polvere, perché non c’è modo di riuscire a essere in grado di mangiare la quantità di proteine che si richiedono da cibi interi. Per sostenere il livello di gluconeogenesi supposto per coprire le necessità durante l’utilizzo di Insulina si è proposto che la migliore strategia è quella in cui si consumano 600g di proteine da una combinazione di proteine del siero di latte e caseina, più un pasto solido che contiene da 50 a 100g di proteine, più alcune verdure fibrose a foglia verde. Il resto delle calorie devono provenire da fonti di grassi con scarsissimo o nullo contenuto di carboidrati.
Si può suddividere l’assunzione consumando una bevanda proteica ogni 30 minuti o un’ora, mescolando in un contenitore 3 litri con 100g di proteine e mantenendo una lista di quante volte si svuota ogni giorno.
Bisogna ricordare, però, che in questo protocollo, l’unica cosa che dovrebbe salvaguardare l’atleta (letteralmente) è l’assunzione di proteine. Se si utilizza l’Insulina con tali modalità e non si mantiene un adeguato apporto di proteine, le conseguenze saranno più che spiacevoli, saranno gravissime.
L’approccio teorico all’uso di Insulina in questo protocollo “No Fat Gains” è:
Giorni da 1 a 3: la deplezione di carboidrati. È necessario diminuire i carboidrati al di sotto dei 100g al giorno. Si suggerisce di arrivare a 50g di carboidrati il giorno 3. L’assunzione proteica deve aumentare a 450g al giorno.
Giorni da 4 a 30: le proteine devono essere pari o superiore a 600g al giorno. I carboidrati devono essere mantenuti tra i 100 e i 50g al giorno (50g è meglio) e si dovrebbero utilizzare i grassi affinché si compensi l’equilibrio delle proprie esigenze caloriche giornaliere. Come già detto, si raccomanda l’uso di proteine in polvere di composizione mista (siero di latte e caseina), anche se è possibile utilizzare alcuni cibi interi, se lo si desidera. (Basta tenere a mente che 600g grammi di proteine corrispondono all’incirca a più di 2.5Kg di petto di pollo o di tacchino al giorno)
Partire da una piccola dose di Insulina (4 UI) per poi aumentarla gradualmente (fino anche a 12UI x 3 volte al giorno).
Monitorare regolarmente la glicemia durante il giorno e in specie nel periodo di massima azione dell’Insulina. Se si inizia a perdere la capacità di rimanere svegli, prendere una zolletta di zucchero.
Si, questo protocollo non è soltanto folle ma, cosa fondamentale, è basato su una conoscenza superficiale e con enormi lacune della fisiologia umana. Perchè? Perchè l’Insulina è un regolatore in negativo della gluconeogenesi!
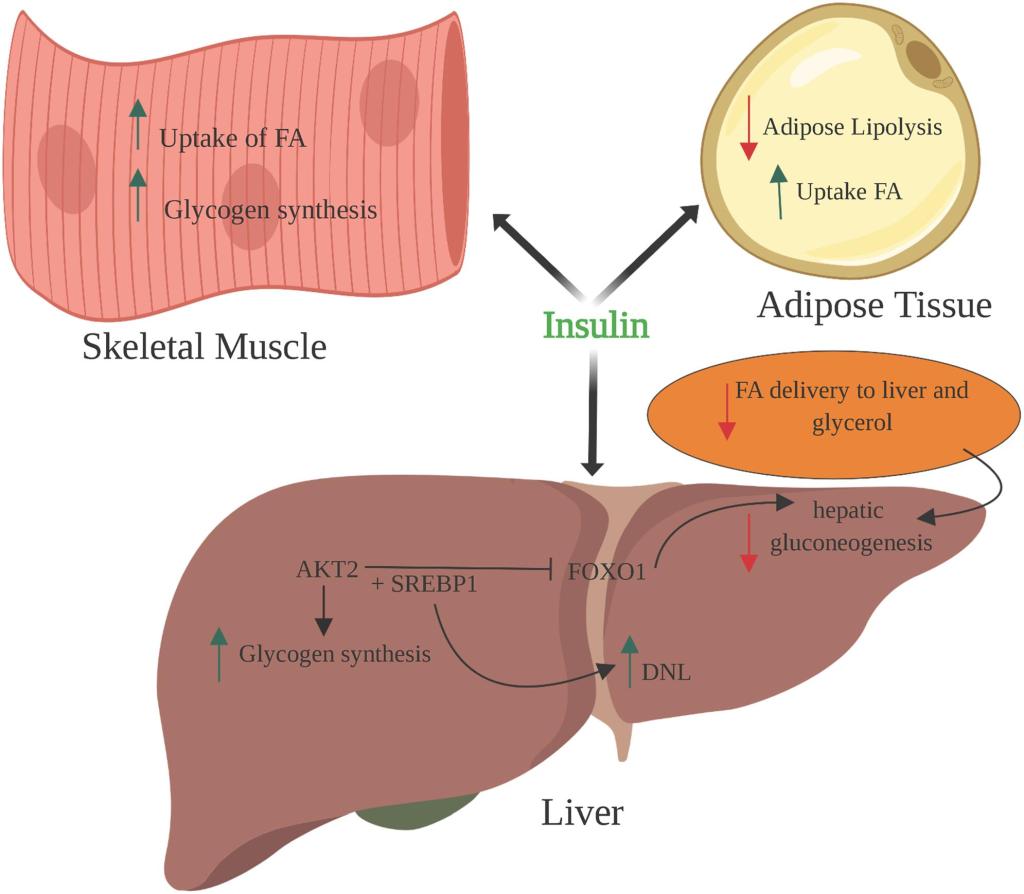
Il ruolo dell’Insulina nella regolazione della produzione epatica di glucosio è ampiamente accettato. Negli individui sani, l’iperinsulinemia fisiologica sopprime la gluconeogenesi del 20%, mentre la glicogenolisi è completamente soppressa. L’iperglicemia da sola sopprime la glicogenolisi epatica con effetti minimi sull’immagazzinamento del glicogeno. Solo la combinazione di iperglicemia e iperinsulinemia ha un effetto significativo sulla sintesi epatica di glicogeno. Pertanto, l’Insulina svolge un ruolo cruciale nel metabolismo epatico del glucosio.
Effetti dell’Insulina sul metabolismo del glucosio e dei lipidi.
Il meccanismo dominante della regolazione insulino-mediata della gluconeogenesi epatica non è però chiaro. L’Insulina esercita un controllo diretto della gluconeogenesi agendo sul fegato, ma influisce anche indirettamente sulla gluconeogenesi agendo su altri tessuti. L’effetto diretto dell’Insulina è stato dimostrato nei cani a digiuno, dove l’Insulina plasmatica portale ha soppresso la produzione epatica di glucosio, anche in assenza di variazioni del glucagone o dei precursori gluconeogenici. Tuttavia, nei modelli murini, l’Insulina è risultata avere effetti più potenti sulla produzione epatica di glucosio in vivo piuttosto che in vitro. Inoltre, è stato dimostrato che gli effetti indiretti dell’Insulina sui tessuti extraepatici sono sufficienti a mantenere il normale metabolismo del glucosio, suggerendo un ruolo importante per la regolazione indiretta della gluconeogenesi da parte dell’Insulina.
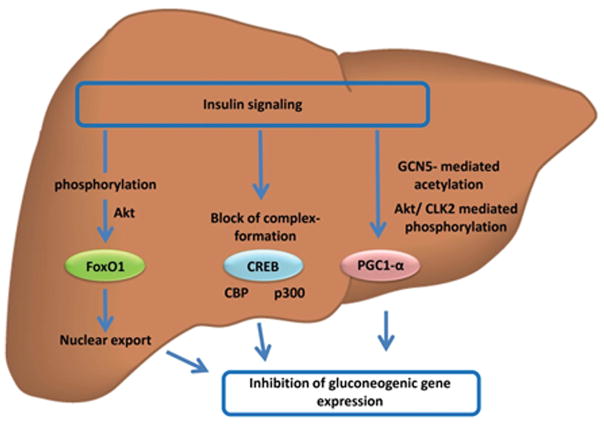
Regolazione dell’espressione genica della gluconeogenesi da parte del segnale insulinico epatico. L’azione dell’Insulina regola l’attività dei fattori di trascrizione che controllano l’espressione dei geni gluconeogenici. La fosforilazione mediata da AKT porta all’esportazione nucleare di FOXO1. La fosforilazione inibitoria di CBP/p300 blocca la formazione del complesso di trascrizione di CREB. La modifica di PGC-1α mediante acetilazione mediata da GCN5 o fosforilazione mediata da AKT/CLK2 riduce l’attività trascrizionale di PGC-1α.
Un sistema di segnalazione insulinica intatto è fondamentale per mantenere i livelli di glucosio nel sangue all’interno di un range glicemico normale e ristretto durante i periodi di digiuno o di eccesso di disponibilità di nutrienti, e questo si ottiene in particolare attraverso la regolazione del flusso metabolico attraverso la via gluconeogenica epatica. Un importante nodo di controllo coinvolge la regolazione trascrizionale dell’espressione dei geni chiave della gluconeogenesi epatica Pck1 e G6pc, che avviene principalmente attraverso il fattore di trascrizione FOXO1 e il recettore nucleare HNF4α e il loro coattivatore trascrizionale PGC-1α. La comprensione di queste vie di regolazione è di estrema importanza per comprendere come un massivo consumo proteico in regime low carb non possa assolutamente garantire una stabilità glicemica in presenza di trattamento con Insulina esogena!
Ma si ingrassa di meno con questo protocollo? Direi di no o, comunque, la differenza è irrisoria… Vorrei ricordare in tal sede che, per quanto dispendioso in termini energetici, in un contesto di eccesso calorico (essenziale in regimi “Bulk”) dove si consumano più proteine del necessario, l’organismo utilizza gli AA che le compongono o come come fonte energetica di scarsa resa [3,3Kcal per 1lt di Ossigeno] o li converte in acidi grassi! Certo, per necessità di deficienza nutrizionale la gluconeogenesi degli AA garantisce una stabilità glicemica, ma in fisiologia e non quando quest’ultima viene marcatamente alterata dall’uso di Insulina esogena!
Per concludere questo paragrafo, è corretto sottolineare che la gluconeogenesi si verifica dopo circa 8 ore di digiuno o scarso apporto glucidico, quando le scorte di glicogeno del fegato iniziano a esaurirsi ed è necessaria una fonte alternativa di glucosio. Inoltre è un processo biochimico piuttosto lento e assolutamente non garante della ben che minima sicurezza in un regime d’uso di Insulina in contesto low carb.
L’uso dell’Insulina pre-workout:
I dibattiti sulla reale efficacia dell’Insulina come agente anabolizzante ha spinto atleti e preparatori a sperimentare protocolli diversi. L’unico di questi che si è avvicinato maggiormente ad una logica d’insieme è ““The Ultimate Insulin Protocol” di Mike Arnold.
Ciò che sta alla base di questo protocollo non è altro che una versione “arricchita” di ciò che sta alla base dell’integrazione “intra-workout”.
Spesso si ragiona sul fatto che durante l’allenamento ci si trova in uno stato fondamentalmente catabolico e dopo in uno stato fondamentalmente anabolico. In realtà però, la sintesi proteica ed i meccanismi di anabolismo e catabolismo sono sempre attivi, con diverse prevalenze, quindi i processi di riparazione tissutale iniziano già nel momento in cui il muscolo viene danneggiato e durante il danneggiamento. Questo è ancor più vero nel momento in cui il workout sarà incentrato su più gruppi muscolari. La presenza di una concentrazione di Insulina esogena in circolo addizionata all’introduzione di macronutrienti ben calibrati (vedi integrazione intra-workout) renderebbe maggiormente incisivo il processo.
Il pump e la pienezza muscolare che si può raggiungere seguendo questo protocollo sono, a detta dei “tester”, impressionanti. Il programma trova la sua “magia” nella sua tempistica e nella sinergia degli ingredienti utilizzati.
Qui di seguito è riportato il protocollo nella sua interezza:
60 minuti pre-workout: *** optional (Uno qualsiasi dei supplementi NO stimolanti sul mercato. Gaspari Nutrition “Vasotropin” è un ottimo prodotto).
45 minuti pre-workout: 15UI di Humulin R.
20 minuti pre-workout: 50g di carboidrati ad alto peso molecolare (ex: Vitargo, Karbolyn, etc). 20g di proteine idrolizzate (es: Hydrowhey, Carnivore). 20g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizata. 2g grammi di Beta-Alanina. 10g di Glutammina. 3g di Taurina. 500mg di Potassio. 1g di Vitamina C.
75 minuti dopo il 1° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 10g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizzata. 2g Beta-Alanina. 10g di Glutammina. 3g di Taurina.
75 minuti dopo il 2° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 3g di Leucina. 5g di Glutammina.
Proteine totali: 60g (esclusi gli amminoacidi in forma libera aggiunti)
Carboidrati totali: 150g (escluse le trace di carboidrati contenute nelle polveri proteiche).
Prima di tutto, nel formulare il rapporto macros/Insulina sopra esposto, Arnold ha aumentato la quantità di carboidrati-proteine al di sopra di ciò che è tipicamente necessario per una UI di Insulina, al fine di tenere conto degli utilizzatori che dimostrano un grado superiore alla media di sensibilità all’Insulina. La maggior parte degli utilizzatori di Insulina, richiedono circa 8g di carboidrati-proteine per UI di Insulina, al fine di chiudere in pareggio e mantenere la normale soglia di glucosio nel sangue. Questo protocollo utilizza un rapporto 14:1 (macros/Insulina), che dovrebbe permettere a praticamente chiunque di utilizzare questo programma mantenendo il glucosio nel sangue all’interno di un range di normalità.
Va notato che questo programma è stato progettato per essere seguito “come è scritto”, soprattutto per quanto riguarda i tempi di assunzione dei nutrienti e le loro quantità. Per gli utilizzatori di Insulina già “navigati” che sanno quali rapporti sono ideali per loro, essi hanno la libertà di ridurre la quantità di macros consumati per UI, se necessario, come determinato dalla valutazione della loro risposta metabolica. Per gli utilizzatori inesperti, la componente nutrizionale del programma dovrebbe essere rispettata come scritta per almeno 2 settimane, a questo punto l’utilizzatore può quindi iniziare a personalizzare il suo rapporto macros/Insulina, se necessario.
La base di questo programma poggia sul tipo specifico di macros utilizzati. Senza di loro, ogni altro componente/aspetto del programma è influenzato negativamente e in alcuni casi rende il tutto sensibilmente limitato negli effetti. Carboidrati ad alto peso molecolare, come il Vitargo o le Ciclodestrine Altamente Ramificate, hanno dimostrato di essere superiori ad altre forme di carboidrati in diversi modi, per esempio un rapido e costante rilascio di glucosio nel sangue e una bassissima osmolaritá in soluzione.
Passando alla componente proteica; le proteine idrolizzate sono molto più velocemente assorbite rispetto ad altri tipi di proteine e sono l’unica proteina che può essere consumata insieme ai carboidrati ad alto peso molecolare senza compromettere il loro assorbimento.
In questo protocollo vi è anche la possibilità di un aggiunta di uno stimolatore del NO a scelta. L’aggiunta di questi stimolatori (vedi, ad esempio, la Citrullina Malato), pur non “necessario”, aumenterà ulteriormente la circolazione e il trasporto dei nutrienti ai muscoli che lavorano, così come contribuiscono ad aumentare il pumping sperimentato durante e dopo l’allenamento. Il Glicerolo monostearato è incluso anche tra gli ingredienti per il suo ruolo di volumizzante muscolare. Questo composto viene spesso utilizzato appena prima della gara, al fine di contribuire al raggiungimento di uno aspetto pieno e asciutto durante l’esibizione. Sono presenti anche dei volumizatori tradizionali, come la Glutammina, la Taurina, la Creatina, e il Potassio.
Al fine di promuovere un maggiore recupero e una maggiore risposta per una crescita muscolare, la temporizzazione dell’assunzione degli shake è stata messa a punto per mantenere un flusso costante dei nutrienti per tutta la vita attiva dell’Insulina. L’Humulin R è stata appositamente scelta per questo scopo, dal momento che con la sua emivita permetterà all’utilizzatore di sfruttare le “finestre” sia intra che post-allenamento. L’Humulin R offre anche un picco di Insulina meno pronunciato, che risulta più facile da gestire per una buona parte degli utilizzatori rispetto ad una versione di Insulina ad azione più rapida, come ad esempio l’Humalog che, però, viene in alcuni casi sostituita alla scelta classica.
Per via del tempo di esposizione all’Insulina limitato con la pratica di questo protocollo, la sensibilità all’Insulina è solo moderatamente influenzata in modo diretto quando si utilizza il programma per circa 5-6 volte alla settimana. Per gli individui che scelgono di utilizzare il presente protocollo per 3-4 volte alla settimana, le alterazioni dirette sulla sensibilità all’Insulina non risulta essere un problema. Per coloro che eseguono il protocollo per 5-6 volte a settimana, possono intervenire in due modi per assicurare una pienamente ottimale sensibilità all’insulina:
L’utilizzatore può fare 2 settimane “off” ogni 4 settimane “on”.
L’individuo può aggiungere Metformina nel suo programma di 3-4 volte a settimana ad un dosaggio di 750mg-1g/die.
Per gli utilizzatori che si apprestano all’uso di questo protocollo per la prima volta, mentre il rapporto macros / Insulina di cui sopra è sufficiente, l’autore consiglia sempre di iniziare con un dosaggio ridotto e aumentarlo poco a poco fino al raggiungimento del pieno dosaggio. Per uno novizio, un dosaggio di 6-8UI è ideale. Questo può essere seguito da una seconda iniezione da 8-10UI, per poi passare ad una terza iniezione di 10-12UI prima di arrivare al dosaggio massimo di 15UI.
Il protocollo in questione non contempla il solo uso di “shake e insulina” ma anche di GH (nel pre-workout a distanza dall’Insulina) e del IGF-1lr3 (nel post workout).
Il protocollo inizia con un’assunzione di GH, circa 30 minuti prima del workout per far si che i livelli plasmatici siano ragionevolmente alti, prima di aggiungere la dose di Insulina. L’idea alla base di questo, è assicurarsi che il GH passi attraverso il fegato mentre si ha una notevole quantità di Insulina in circolo. Questo è il modo in cui produciamo grandi picchi di IGF-1. Dopo l’allenamento, si somministra l’IGF-1LR3.
Il protocollo esemplificativo è il seguente:
30 minuti pre-workout: 6-10UI di GH subq
15 minuti pre-workout: 6-16UI di Novalog subq
10 minuti pre-workout: assumere lo shake #1
Dopo ogni set eseguito: sorseggiare lo shake #2, e terminarlo entro la fine dell’allenamento.
Andare a casa
Somministrare la dose di 100mcg di IGF-1lr3 (per i suoi effetti sulla sensibilità all’Insulina
Assumere lo shake #3
Formulazione degli shake:
Shake 1: 10-20g di EAA (Amino Acidi Essenziali) o PeptoPro, 40-60g Vitargo, 5g Creatina Monoidrata Micronizata, 200mg di Caffeina (migliora la resintesi di glicogeno); anche in questo caso la dose di carboidrati per ogni UI si aggira in media sui 7-10g.
Shake 2: 10-20g EAA o PeptoPro, 50-100g Vitargo, 5g di Carnitina Monoidrata Micronizata.
Shake 3: 2 tazze di albume pastorizzato, 1 tazza di avena istantanea, 1 banana o 1 tazza di mirtilli, Splenda o Stevia.
La somministrazione di Insulina pre-workout trova la sua motivazione d’essere nel fatto che durante l’allenamento con i pesi si viene a creare uno stress meccanico che a sua volta crea una sovra regolazione dello stimolo anabolico. Più comunemente, questo stato iper-anabolizzante viene indicato come “finestra anabolica”; un determinato periodo di tempo che dura dall’inizio dell’allenamento fino a poche ore dopo. Un modo in cui il corpo reagisce all’allenamento con i pesi è attraverso un aumento della sensibilità all’Insulina. Questo accade quando i recettori per l’Insulina, che risiedono sulla superficie della cellula, rispondono al segnale dell’Insulina in modo più efficiente, cosa che ci permette un migliore assorbimento dei nutrienti all’interno della cellula. Oltre a ciò, avviene un aumento della proliferazione dei GLUT-4 sulla superficie cellulare in maniera insulino-indipendente, caratteristica che richiede una minore quantità di Insulina per avere i massimi effetti nell’uptake cellulare.
Infatti, l’allenamento promuove anche il recupero e la crescita a livello intracellulare aumentando la sintesi proteica, la glicogeno sintasi, dei GLUT4 e l’espressione del trasportatore degli aminoacidi, e diminuendo i livelli di Miostatina. In combinazione con un aumento della sensibilità all’Insulina, queste cose non solo si traducono in una crescita accelerata, ma forniscono un effetto di ri-partizionamento dei nutrienti, in cui il cibo che assumiamo e i macronutrienti ivi contenuti vengono indirizzati maggiormente verso il miocita (cellule muscolari), piuttosto che immagazzinati come grasso.
L’Insulina è il complemento perfetto per questa “finestra anabolica” durante l’allenamento, in quanto non solo i nutrienti vengono trasportati alle cellule muscolari, permettendo così al corpo di approfittare di questo stato anabolizzante intensificato, ma l’Insulina agisce anche per molti dei processi di costruzione muscolare, fornendo uno stimolo ipertrofico raddoppiato nel momento in cui il corpo è più sensibile alla risposta del segnale dell’Insulina.
Come anti-catabolico, e lo abbiamo anche visto nella review presente nella terza parte di questa serie di articoli, l’Insulina è decisamente efficace, riducendo sensibilmente la degradazione del tessuto muscolare che si somma attivamente con la ripartizione delle proteine muscolari, e il tempo di recupero risulta ridotto. Questo ha un duplice effetto:
alterazione in positivo del rapporto catabolismo:anabolismo e maggiore supercompensazione (crescita muscolare);
il recupero più rapido si traduce in un aumento della possibile frequenza di allenamento, permettendo ai muscoli di essere stimolati più volte entro un determinato periodo di tempo e, in definitiva, di crescere più rapidamente;
infine, le concentrazioni di Insulina sovrafisiologiche aumentano in modo significativo il volume ematico all’interno nel tessuto muscolare, aggiungendo un pump muscolare generale che di per se stimola la crescita.
Tuttavia, affinché il corpo possa mettere a frutto tutto questo, sono necessari lo stimolo allenante adeguato e i nutrienti giusti che devono essere presenti al momento giusto. A questo punto entra in gioco la nutrizione intra-allenamento presentata in precedenza.
Nonostante questo protocollo abbia rappresentato la migliore applicazione dell’uso di Insulina nel Bodybuilding, esso presenta delle lacune oltre che dei problemi tecnici che riducono la pienezza del potenziale e il margine di “sicurezza”.
Alcuni atleti hanno sperimentato una maggiore tendenza all’ipoglicemia durante i loro workout con protocolli di Insulina pre workout. Questo imprevisto è limitante oltre che decisamente pericoloso, specie durante un workout. E’ ovvio che il controllo della glicemia e il rispetto dei punti base del protocollo riducono le possibilità del verificarsi di casi ipoglicemici. Ma il rischio è comunque maggiore rispetto ad una condizione di trattamento base (assunzione classica ai pasti).
Inoltre, tiene poco in considerazione le reale farmacocinetica dei PEDs utilizzati riducendo le piene ed ottimali interazioni ed effetti additivi.
GH/Insulin Protocol – pre e post-workout:
In risposta alle lacune del protocollo di Arnold che abbiamo appena visto, ho realizzato una versione perfezionata denominata “GH/Insulin Protocol”.
Tale perfezionamento prende in considerazione in modo preciso sia la farmacocinetica che la farmacodinamica incrociata dei componenti Insulina e GH.
Ora, sappiamo che la curva di rilascio del hGH somministrato per via sottocutanea raggiunge un picco iniziale dopo 30 minuti post iniezione per poi attestarsi a 2-3h e subire un calo significativo dopo 4h dalla somministrazione. Lo strascico di IGF-1 perdura per circa 24h sopra il basale.
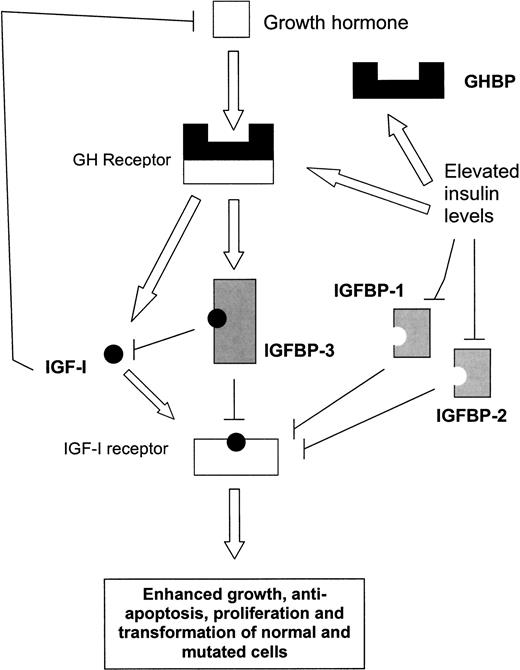
Sappiamo inoltre che l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1;3) l’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1.
Interazioni biologiche a livello ipofisario ed epatico tra Insulina, Ormone della Crescita (GH), Fattore di Crescita Insulino-Simile-I (IGF-I) e Proteine Leganti il Fattore di Crescita Insulino-Simile (IGFBPs). Le frecce aperte indicano la stimolazione e le linee nere sottili l’inibizione. Livelli elevati di Insulina (a destra) possono aumentare indirettamente la biodisponibilità di IGF-I (cerchi pieni) sopprimendo la produzione di IGFBP-1 e, in misura minore, di IGFBP-2 (simboli ombreggiati). A sua volta, l’aumento della biodisponibilità di IGF-I può aumentare l’effetto di feedback negativo sul GH (quadrato aperto), portando a una riduzione della secrezione di GH e a una minore produzione epatica di IGF-I e IGFBP-3. Tuttavia, livelli elevati di Insulina possono anche aumentare il numero e l’attività dei recettori epatici del GH (barra aperta), riflessi da un aumento dei livelli di Proteina Legante l’Ormone della Crescita (GHBP) circolante. Questo effetto può portare a un aumento della produzione epatica di IGF-I e IGFBP-3 regolata dal GH, con un aumento maggiore dei livelli di IGF-I circolante rispetto a quelli di IGFBP-3. Pertanto, insieme a fattori genetici, ormonali e ambientali, l’entità relativa di questi due effetti opposti dell’Insulina sulla produzione di IGF-I potrebbe determinare i livelli di IGF-I circolante. Nel tempo, un’eccessiva biodisponibilità di IGF-I potrebbe aumentare il rischio di cancro del colon-retto, favorendo la sopravvivenza di cellule trasformate e mutate che normalmente andrebbero incontro ad apoptosi.
Quindi, dal momento che l’obbiettivo è quello di creare un ambiente non fisiologicamente riproducibile senza alterazioni iatrogene al fine di ottenere la massima risposta anabolica dal protocollo, il primo punto di congiunzione, o meglio la chiave di volta del protocollo, deve essere l’incrocio del picco di hGH con quello dell’Insulina. Per fare ciò le modalità di somministrazione dovrebbero contemplare primariamente l’iniezione di hGH pre-workout e quella di Insulina (preferibilmente Humalog per via del picco raggiunto entro 15 minuti dalla somministrazione) nel post-workout. Un vantaggio aggiuntivo di questa modifica è la riduzione del rischio ipoglicemico durante la sessione di allenamento.
E no, il vantaggio dello shake intra-workout non viene perso dal momento che la ripartizione calorica è di per se ottimale per via di meccanismi insulino-indipendenti dati dall’attività contro-resistenza (vedi aumento dei GLUT-4 sulla superficie cellulare in seguito all’attività muscolare). Si veda anche l’assorbimento dilazionato dei composti facenti parte della soluzione ingerita e costituente lo shake intra-workout.
I punti chiave del protocollo sono i seguenti:
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8, in questo ultimo caso divise in una dose da 4UI pre workout e due da 2UI appena sveglio e prima di dormire, a secondo della modalità d’uso del hGH; se somministrato giornalmente o a giorni alterni. Vedi a tal proposito l’articolo dedicato alla somministrazione di hGH a giorni alterni];
1h pre-workout possibilità di assumere tra i 25 ed i 50mg di Sildenafil seguiti a 30 minuti dall’inizio del workout dall’assunzione di 8g di Citrullina Malato;
consumare uno shake intra-workout contenente di base 0,5-1g di carboidrati ad alto peso molecolare, 0,25g di proteine idrolizzate e 5g di Creatina Monoidrato;
somministrazione dell’Insulina (Humalog) post-workout [le UI vanno calibrate in base ai CHO consumati nel pasto post-workout; in linea di massima 1UI ogni 10Kg di peso]. L’Humalog può essere sostituita con l’Humulin-R se per la risposta del soggetto in quanto a tolleranza è migliore.
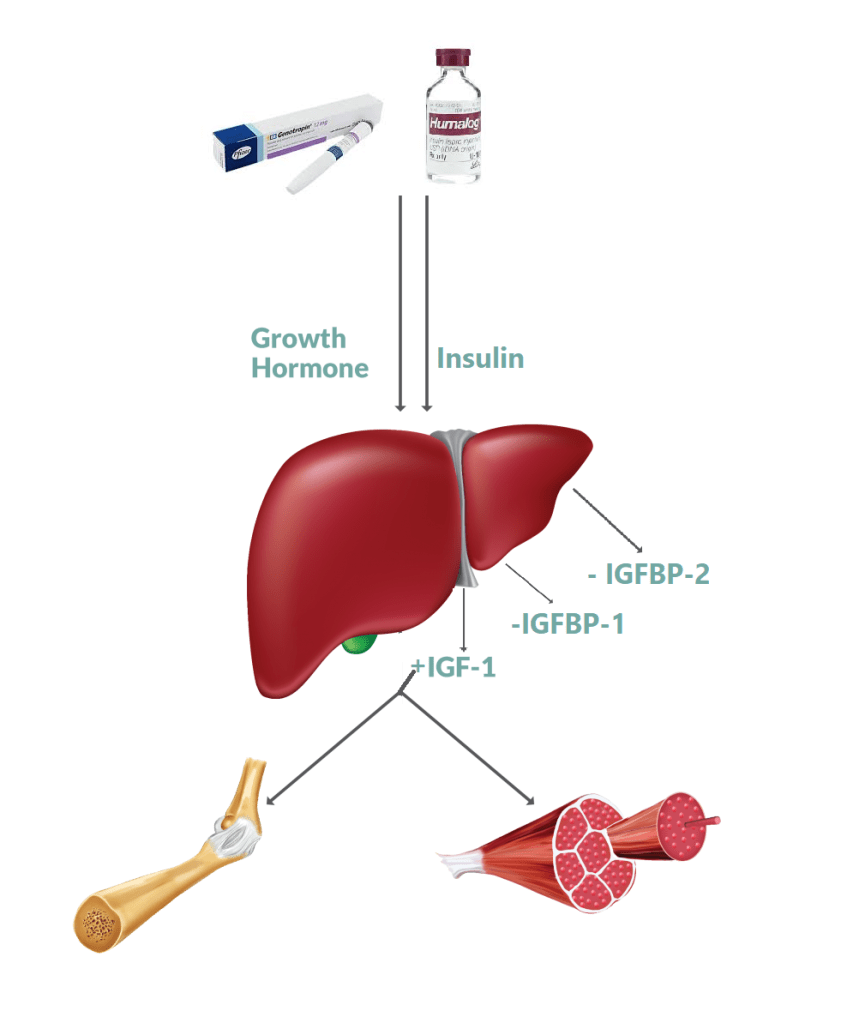
Schema esemplificativo dell’azione incrociata tra Insulina e GH a livello epatico e sue consequenziali principali aree di influenza ricercate.
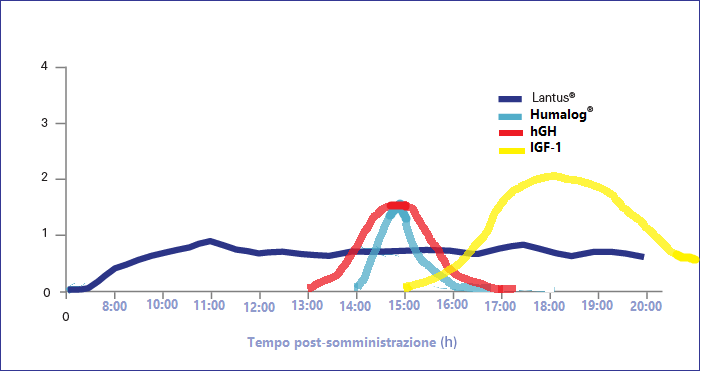
Esiste una variante del suddetto protocollo la quale contempla l’uso di due tipi di Insulina: una base (Lantus) e una post-workout (Humalog):
calcolare la dose totale di Insulina giornaliera con la formula 1UI ogni 10Kg di peso;
dividere il totale della dose a metà tra Insulina Glargine (Lantus) e Insulina lispro (Humalog) o Humulin-R;
somministrazione di Insulina glargina (Lantus) dividendo la dose giornaliera in 2: la prima, pari al 65% della dose totale, al mattino in concomitanza con il primo pasto e la seconda, pari al 35% della dose totale, 12h dopo la prima somministrazione;
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8 a secondo della modalità d’uso del hGH];
somministrare la dose di Insulina lispro o Humulin-R nel post-workout.
L’uso di una base di Insulina rappresentata dalla Insulina glargina ha il potenziale di aumentare ulteriormente l’espressione del IGF-1 e, di conseguenza, la risultante anabolizzante del protocollo. Ma, ovviamente, i rischi di ipoglicemia con questa modifica protocollare sono maggiori.
Grafico esemplificativo delle curve ematiche di Insulina, hGH e IGF-1 nel protocollo con Insulina Glargine di base.
Il monitoraggio regolare della glicemia e il rispetto dei punti base del protocollo evitano con un buon margine il verificarsi di casi ipoglicemici. Quindi, il glucometro, che sia classico o con sensore, è essenziale in questi casi.
Repetita iuvant: QUESTO PROTOCOLLO COME I PRECEDENTEMENTE PRESENTATI, E L’USO DI INSULINA, SONO APPANNAGGIO DI ATLETI AVANZATI E MONITORATI DA PERSONALE QUALIFICATO! NULLA DI CIO’ CHE E’ STATO DESCRITTO DEVE ESSERE PRESO COME UNA PRESCRIZIONE MEDICA O UN CONSIGLIO!
Conclusioni “dopo un lungo viaggio”:
Siamo ora giunti alla conclusione di questa serie di articoli dedicati all’Insulina e al suo centesimo anniversario.
Durante questo lungo percorso abbiamo imparato a conoscere meglio questo affascinante e mal compreso peptide. Abbiamo visto come è stato scoperto, abbiamo compreso l’enorme passo avanti nella medicina e nella tutela della vita umana che la sua scoperta ha rappresentato, abbiamo compreso come essa sia fisiologicamente regolata e quali sono le sue reali azioni a livello sistemico ed abbiamo imparato a separare i luoghi comuni che vi aleggiano intorno dai fatti.
Con la terza parte molti sono rimasti delusi nel constatare che in fisiologia l’Insulina abbia un effetto prettamente anticatabolico, effetto preminente che mantiene anche se somministrata esogenamente entro i range fisiologici [<1.200ng/dl]. Altresì gli umori sono migliorati quando siamo venuti a conoscenza del fatto che non solo il dosaggio fa la differenza tra preminenza anticatabolica e anabolica ma anche la sensibilità.
Con la recente constatazione che il risultato dell’equazione “Insulina/anabolismo” cambi drasticamente in positivo se vi si aggiunge la variabile del hGH, abbiamo imparato che l’unico uso minimamente sensato dell’Insulina per il miglioramento della massa muscolo-scheletrica è in associazione con il peptide ipofisario con le ultime due modalità esposte.
Ci tengo però a precisare che l’uso dell’Insulina “off-label” dovrebbe rappresentare la componente più marginale nella carriera di un culturista. Vale a dire che se ne può benissimo fare a meno, in specie quando si è semplici amatori o agonisti di piccoli o medi circuiti competitivi. Non complicatevi la vita.
Fate tesoro delle nozioni che vi ho esposto affinché il confine della conoscenza si espanda e prevalga su quello dell’ignoranza.
Per accedere alla prima e alla seconda parte clicca qui e qui.
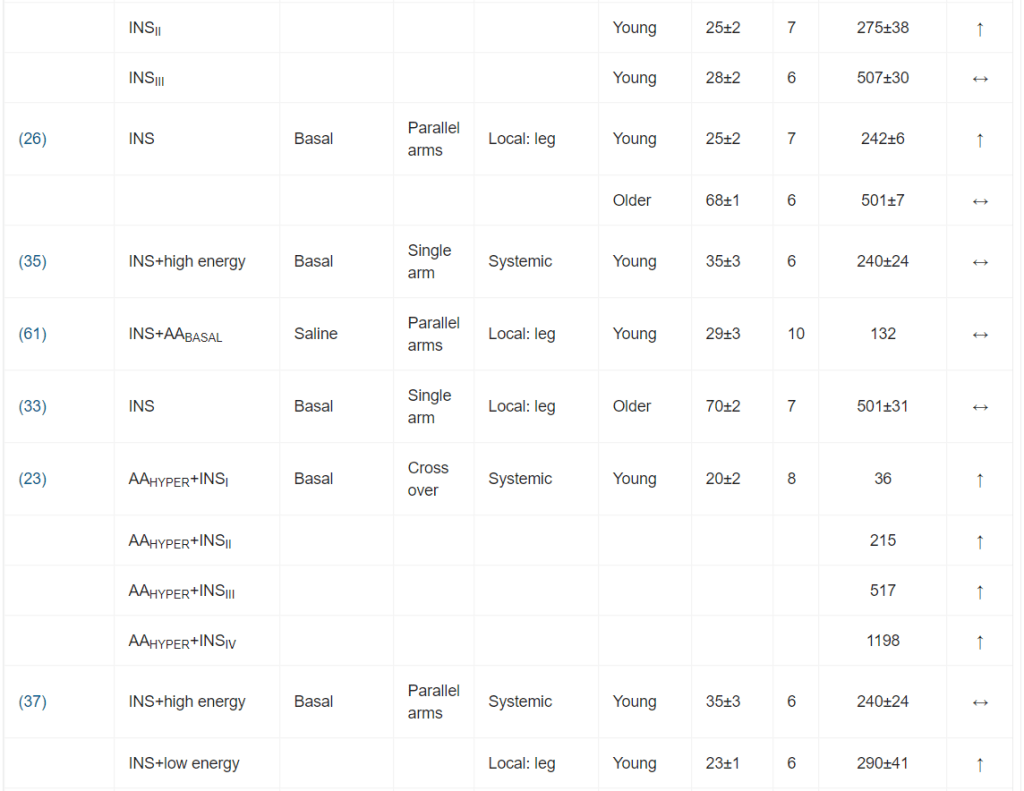
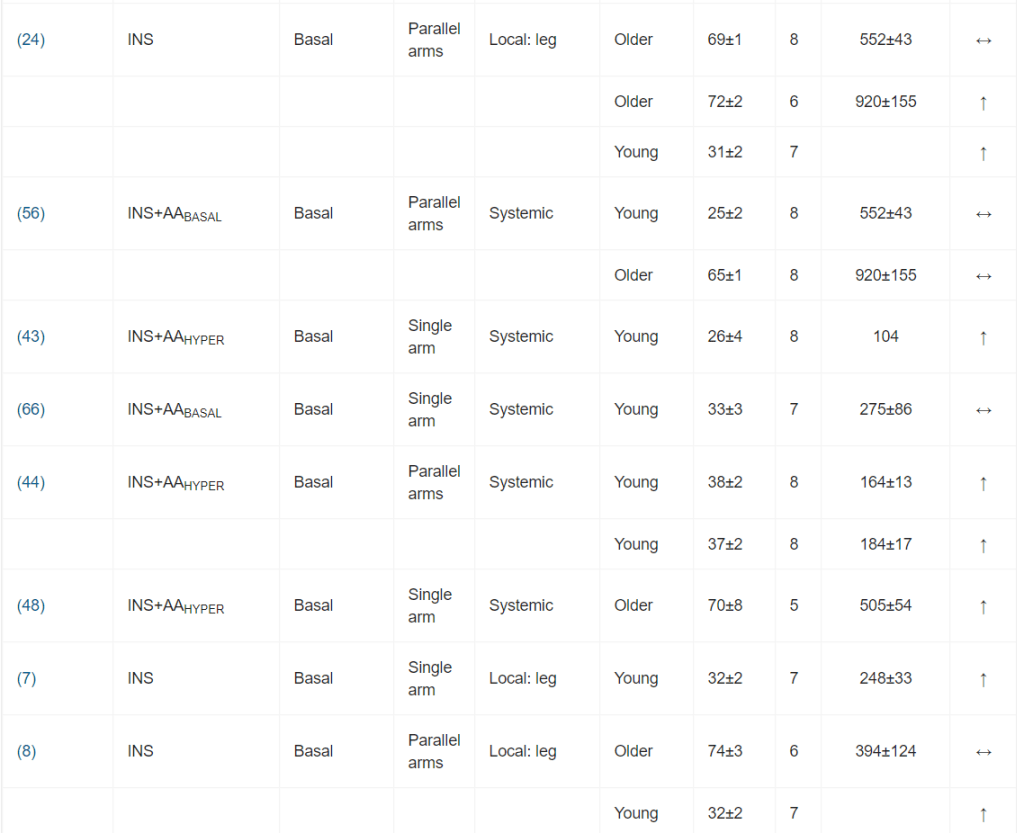
Il ruolo dell’Insulina nella regolazione della sintesi e della degradazione delle proteine del muscolo-scheletrico umano:
Nell’uomo, le proteine costituiscono circa il 15% del peso corporeo [1]. Sono il principale macronutriente che compone il muscolo-scheletrico, che a sua volta contiene circa il 30-45% delle proteine totali del corpo e contribuisce al 20-35% del turnover proteico dell’intero organismo. È stato dimostrato che sia gli aminoacidi (AA) che l’Insulina svolgono un ruolo cruciale nella regolazione delle variazioni diurne del turnover proteico del muscolo scheletrico [2] e che gli squilibri tra i tassi di sintesi proteica muscolare (MPS) e di degradazione delle proteine muscolari (MPB) hanno importanti conseguenze sulle dimensioni, sulla qualità e sulla funzione del muscolo [3]. La “sarcopenia” descrive la perdita di massa e forza muscolare scheletrica che si verifica con l’avanzare dell’età [4]. Il processo di invecchiamento stesso è caratterizzato dall’incipiente sviluppo della sarcopenia, in cui è stato segnalato un costante declino della massa magra (e della funzione associata) di circa l’1% all’anno oltre i 60 anni di età [5]. A causa della fragilità associata, la sarcopenia porta a una diminuzione della qualità della vita e della salute, caratterizzata da scarsa mobilità, sedentarietà, aumento del rischio di cadute e scarso recupero dalle malattie [6, 7].
Turnover Proteico schematico e diversi destini metabolici degli aminoacidi nel muscolo-scheletrico.
I dati provenienti da studi epidemiologici e sperimentali hanno riportato che il diabete di tipo II è correlato a una scarsa forza e funzione muscolare, con un tasso accelerato di declino della qualità e della forza muscolare negli individui anziani fino al 30% [8]. Alla luce della crescente prevalenza del diabete e delle sequele metaboliche della sarcopenia legata all’età, è aumentato l’interesse per i meccanismi con cui il diabete di tipo II esacerba il declino della massa muscolare legato all’età. Inoltre, poiché il muscolo scheletrico è un sito importante per lo smaltimento del glucosio, la riduzione quantitativa del volume del muscolo appendicolare potrebbe potenzialmente influire negativamente sullo smaltimento e sul metabolismo del glucosio [9]. Una maggiore comprensione dei fattori endocrini che regolano la massa muscolare è quindi importante per il controllo glicemico e per contrastare la sarcopenia.
Molti pazienti con diabete di tipo II necessitano di Insulina per raggiungere gli obiettivi ottimali di glucosio, poiché la capacità di produrre Insulina endogena da parte delle cellule beta pancreatiche diminuisce progressivamente [10]. Tuttavia, la terapia insulinica è associata a un aumento di peso [11, 12], soprattutto di massa grassa [13], anche se non correlate all’ormone in se come abbiamo visto nella seconda parte, che aumenta l’insulino-resistenza e rende necessario l’uso di dosi più elevate di Insulina a scapito di un ulteriore aumento di peso. L’esatto ruolo dell’Insulina nel metabolismo del muscolo scheletrico umano, tuttavia, continua a far discutere. Sebbene gli studi sugli animali abbiano riportato che l’Insulina promuove la MPS, questi studi sono stati condotti principalmente su animali in crescita [14, 15]. Il ruolo dell’Insulina nel muscolo scheletrico umano adulto è più complesso e soggetto all’interazione tra altri fattori come la disponibilità di AA, il flusso sanguigno muscolare e il reclutamento microvascolare [16, 17]. Ciò ha portato a diversi studi che riportano conclusioni opposte per quanto riguarda la relazione tra Insulina e turnover proteico del muscolo scheletrico umano [16-22]. Andiamo quindi a tentare di chiarire il ruolo dell’Insulina nella regolazione del metabolismo muscolare nell’uomo.
Vie di segnalazione insulinica del muscolo scheletrico. Il muscolo scheletrico sano è in grado di alternare l’uso dei carboidrati nei periodi di abbondanza (aumento dell’Insulina) e dei lipidi nei periodi di scarsità calorica (diminuzione dell’Insulina). L’utilizzo di substrati carboidratici e lipidici può essere potenziato anche durante i periodi di elevata richiesta metabolica dell’esercizio fisico. IR 5 recettore dell’Insulina; IRS 5 substrato del recettore dell’Insulina; PI3K 5 fosfatidilinositolo-3-chinasi; PDK 5 proteina chinasi fosfoinositide-dipendente; aPKC 5 proteina chinasi C atipica; PIP3 5 fosfatidilinositolo 3,4,5-trifosfato; Akt 5 proteina chinasi B; AS160 5 substrato 160 di Akt; GLUT4 5 trasportatore di glucosio insulino-sensibile; IMTG 5 trigliceridi intramiocellulari; LCFA 5 acidi grassi a catena lunga; AMPK 5 proteina chinasi attivata dall’AMP.
Per la realizzazione di una interessante review sistematica e meta-analisi riguardante il ruolo dell’Insulina sulla MPS e MPB sono stati cercati studi in lingua inglese pubblicati tra il 1946 e il novembre 2013. Sono stati selezionati gli articoli sottoposti a revisione paritaria che indagavano il ruolo dell’Insulina sulle MPS e/o sulla MPB. Per la review sistematica sono stati selezionati tutti gli studi sperimentali che riportavano cambiamenti nel metabolismo delle proteine muscolari nell’uomo in risposta a interventi con Insulina, indipendentemente dal metodo di valutazione. Sia il metodo a due pool (compartimenti) che quello a tre pool sono stati utilizzati per riportare il metabolismo proteico muscolare, ed entrambi forniscono cambiamenti qualitativamente comparabili nel metabolismo proteico dal sangue e dall’arricchimento intracellulare di fenilalanina [21]. Nel modello a due pool (arteria e vena), la fenilalanina entra ed esce dall’arto rispettivamente attraverso l’arteria e la vena. La velocità di scomparsa della fenilalanina dall’arteria è utilizzata per stimare l’MPS e deriva dalle misurazioni della velocità di MPB e del bilancio netto (NB); MPB è determinato dalla velocità di comparsa della fenilalanina in vena (cioè la diluizione dell’arricchimento del tracciante attraverso l’arto), mentre NB è semplicemente la differenza di concentrazione della fenilalanina attraverso l’arto. Nel modello a tre bacini (arteria, vena e muscolo), la fenilalanina entra ed esce dall’arto come sopra. Il flusso unidirezionale di fenilalanina libera dall’arteria al compartimento intramuscolare è determinato dalla velocità di trasporto verso l’interno. La velocità di comparsa intracellulare della fenilalanina definisce la velocità di rilascio dal MPB. Poiché la fenilalanina non viene ossidata dal muscolo scheletrico, il tasso di utilizzo intracellulare corrisponde al tasso di utilizzo per la MPS [22].
Negli studi esaminati, il modello a due pool è stato il metodo analitico più comunemente utilizzato e la fenilalanina è stata il tracciante AA più comunemente usato. Pertanto, per consentire un’analisi quantitativa comparabile degli studi eleggibili per la review sistematica e per evitare un’eterogeneità significativa, la meta-analisi ha incluso solo gli studi che hanno utilizzato il modello a due pool per analizzare i dati di fenilalanina, rispetto ad altri metodi analitici o ad altri traccianti di AA (ad esempio, la leucina). I dati a tre pool sono stati inclusi nella review sistematica. A causa di un’ampia sovrapposizione tra gli studi che riportavano dati a due e tre pool (n = 10) e del numero significativamente inferiore di studi che riportavano esclusivamente dati a tre pool (n = 5), non è stata eseguita una meta-analisi dei dati a tre pool.
Sono stati esaminati tutti gli studi che rispondevano ai criteri di inclusione. L’esito primario era la variazione di MPS e/o MPB in risposta all’intervento insulinico. I dati pubblicati sono stati estratti dagli studi e sono state calcolate le medie. A causa dei metodi di misurazione simili tra gli studi inclusi, sono stati utilizzati modelli a effetti casuali per calcolare le differenze medie ponderate (WMD), gli CI al 95% e i valori di p corrispondenti. L’eterogeneità tra gli studi è stata valutata utilizzando la statistica I 2, che descrive la percentuale di variazione totale tra gli studi che è il risultato dell’eterogeneità piuttosto che del caso [23]. Poiché gli AA sono il substrato principale per la sintesi proteica, sono state effettuate analisi di sottogruppo in base ai diversi livelli di apporto di AA al muscolo e quindi alla quantità disponibile (aumentata, invariata o diminuita) per il metabolismo proteico. Un’altra analisi di sottogruppo è stata eseguita con studi che coinvolgevano popolazioni con diabete. In questi studi, l’apporto di AA non è cambiato. È stata condotta un’analisi di meta-regressione per verificare le differenze nelle stime in pool tra i sottogruppi e per verificare se le stime in pool differissero in base ad altre covarianti (ad esempio, i livelli di concentrazione di Insulina raggiunti, l’età o la massa corporea magra).
Il bias di pubblicazione è stato valutato esaminando un funnel plot in funzione della dimensione dell’effetto. I test statistici per le meta-analisi sono stati eseguiti utilizzando il pacchetto statistico STATA 13.0 (StataCorp, College Station, TX, USA).
Dopo la rimozione dei duplicati, sono stati recuperati 646 articoli dalla ricerca e dalle liste di riferimento degli articoli selezionati (Fig. seguente). Lo screening del titolo e dell’abstract ha portato all’esclusione di 455 articoli a causa dell’irrilevanza (ad esempio, studi in vitro, studi sul metabolismo delle proteine epatiche) e di altri 87 articoli perché gli studi erano stati condotti su muscolo scheletrico animale. In totale sono stati identificati 104 articoli potenzialmente rilevanti, che sono stati valutati in modo più approfondito. Di questi, altri 60 articoli sono stati esclusi. I principali motivi di esclusione sono stati: (1) gli studi valutavano il ruolo degli interventi nutrizionali e non dell’Insulina in sé; e (2) gli articoli erano revisioni piuttosto che studi di ricerca. Un totale di 44 articoli che comprendevano 75 studi soddisfaceva i criteri per la review sistematica. Di questi 44 articoli, 13 (contenenti 25 studi) sono stati inclusi nella meta-analisi, in quanto rappresentavano il gruppo più numeroso che conteneva dati quantitativamente comparabili. Tutti i 25 studi hanno utilizzato la fenilalanina come tracciante AA, hanno riportato la MPS/MPB in unità di nmol (100ml leg vol.)-1 min-1 e hanno utilizzato l’approccio a due pool (equilibrio arterovenoso) per stimare le variabili di esito.
Diagramma di flusso che rappresenta il processo di reperimento degli articoli e delinea gli articoli che hanno soddisfatto i criteri per la revisione sistematica e la meta-analisi. Solo gli studi che utilizzavano il modello a due pool e i traccianti di fenilalanina sono stati inclusi nella meta-analisi.
MPS e Insulina
Per l’inclusione nella meta-analisi sono stati identificati 13 articoli [16, 19-21, 24-32], contenenti 25 studi sperimentali che hanno utilizzato diverse concentrazioni di insulina (Tabella 1); tutti hanno analizzato l’effetto dell’insulina sia sulla MPS che sulla MPB (Tabella 1). In totale sono stati inclusi 173 individui in questi studi. Un totale di 13 studi ha coinvolto giovani adulti e tre di questi 13 studi hanno coinvolto individui con diabete. Otto studi hanno coinvolto persone anziane e sane, mentre per quattro studi non erano disponibili dati sull’età. L’età media dei partecipanti variava da 18 a 74 anni. La maggior parte degli studi (20 studi) ha utilizzato l’infusione locale di insulina intra-arteriosa per limitare lo sviluppo di ipoglicemia sistemica e un’infusione obbligatoria di glucosio, anche per limitare l’ipoaminoacidemia [33]. Questo aspetto è di fondamentale importanza, soprattutto quando si utilizzano concentrazioni di insulina sovrafisiologiche. Le concentrazioni sistemiche di insulina variavano tra 62,5 e 861,2 pmol/l. L’apporto di AA (concentrazione di AA nell’arteria × flusso sanguigno arterioso) è stato mantenuto in 14 studi e aumentato in otto studi. Tuttavia, come conseguenza diretta della somministrazione sistemica di insulina (vedi discussione sotto), gli AA circolanti sono diminuiti in tre studi.
Caratteristiche degli studi inclusi nella meta-analisi, ordinati per modalità di somministrazione dell’insulina e di erogazione degli AA. I valori sono medie ± SEM, se non diversamente specificato. aI numeri accanto al nome dell’autore distinguono studi diversi dello stesso autore; le lettere indicano interventi o caratteristiche dei partecipanti diversi all’interno di uno stesso studio. bRispetto al basale cPartecipanti con diabete
I dati della meta-analisi sono stati raggruppati da 13 studi (25 studi o confronti) che hanno coinvolto 173 individui. La WMD per la MPS era 3,90 (95% CI -0,74, 8,55; p = 0,71). L’analisi degli studi basati sulla somministrazione di AA ha rivelato un aumento della MPS (WMD 13,44 [95% CI 4,07, 22,81], p < 0,01) negli studi in cui la somministrazione di AA era aumentata (otto studi, 63 individui). Tuttavia, la MPS non è cambiata significativamente quando la somministrazione di AA è stata ridotta (tre studi; 22 partecipanti; WMD 1,57 [95% CI -3,97, 7,12], p = 0,58) o mantenuta al basale (11 studi; 73 partecipanti; WMD 2,00 [95% CI -5,28, 9,28], p = 0,59). Gli studi che hanno coinvolto individui con diabete (tre studi, 15 individui) hanno mostrato riduzioni significative della MPS in risposta all’Insulina, anche se l’apporto di AA è stato mantenuto (WMD -6,67 [95% CI -12,69, -0,66], p < 0,05).
Forest plot della meta-analisi a effetti casuali della WMD (95% CI) sull’effetto dell’insulina sulla MPS. Nel complesso non è stato osservato un aumento significativo della MPS (p = 0,710). Quando si è stratificato per l’apporto di AA, è stato osservato un aumento della MPS quando l’AA è stato aumentato (p < 0,01), ma non è stata osservata alcuna differenza quando l’apporto di AA è stato mantenuto (p = 0,59) o ridotto (p = 0,58). Nei soggetti con insulino-resistenza o diabete (IR), la MPS era significativamente ridotta (p < 0,05) nonostante il mantenimento dell’apporto di AA. I numeri accanto al nome dell’autore differenziano i diversi studi dello stesso autore; le lettere indicano interventi diversi o caratteristiche dei partecipanti all’interno di uno stesso studio.
All’analisi di meta-regressione, la dimensione della stima (WMD) era significativamente diversa tra i sottogruppi basati sulla disponibilità di AA (p = 0,001).
L’I 2 per l’effetto complessivo dell’Insulina sulla MPS era del 49% (p = 0,003). Questa significativa eterogeneità moderata sembrava essere dovuta principalmente all’eterogeneità all’interno del sottogruppo con aumento degli AA (I 2 59%; p = 0,018). Gli altri sottogruppi hanno mostrato valori di p non significativi per l’eterogeneità, suggerendo una maggiore coerenza tra questi studi rispetto ai dati complessivi della MPS (sottogruppi: consegna AA mantenuta: I 2 21%; p = 0,241; diminuzione del rilascio di AA: I 2 0%; p = 0,811; individui con diabete: I 2 0%; p = 0,605).
MPB e Insulina
I dati sono stati raggruppati dagli stessi 25 studi come per la MPS. La WMD per la MPB era di -15,46 (95% CI -19,74, -11,18; p < 0,0001; Fig. 3). La disponibilità di AA non ha avuto un impatto significativo sulla dimensione stimata della MPB (p = 0,754). L’I 2 per l’effetto complessivo dell’Insulina sulla MPB era del 13% (p = 0,282), indicando un’eterogeneità non significativa (Fig. 3).
Forest plot della meta-analisi a effetti casuali della WMD (95% CI) sull’effetto dell’Insulina sulla MPB. Nel complesso è stata osservata una riduzione significativa della MPB (p < 0,0001). I numeri accanto al nome dell’autore differenziano i diversi studi dello stesso autore; le lettere indicano interventi o caratteristiche dei partecipanti diversi all’interno di uno stesso studio.
Insulina e bilancio netto delle proteine [NB]
In un’ulteriore analisi in pool di tutti i 25 studi, è stato riscontrato che l’Insulina aumenta significativamente l’assorbimento di proteine NB (WMD 20,09 [95% CI 15,93, 24,26], p < 0,0001).
Forest plot della meta-analisi a effetti casuali della WMD (95% CI) sull’effetto dell’Insulina sulla cinetica delle proteine NB. L’insulina ha avuto un forte effetto positivo sull’assorbimento di proteine NB nel complesso (p < 0,0001). Gli studi sono ordinati cronologicamente. I numeri accanto al nome dell’autore differenziano i diversi studi dello stesso autore; le lettere indicano diversi interventi o caratteristiche dei partecipanti all’interno di uno stesso studio.
Analisi di meta-regressione di altre variabili
È stata condotta un’analisi di meta-regressione per verificare se altre variabili confondenti di interesse avessero un effetto sulla WMD (ad esempio, la concentrazione di insulina infusa, l’età e la massa corporea magra, se disponibili). Le differenze nelle concentrazioni di Insulina infusa non hanno avuto alcun effetto su MPS (p = 0,955), MPB (p = 0,713) o NB (p = 0,621). Non vi è stato alcun effetto nemmeno per le differenze di età (p = 0,480, p = 0,159 e p = 0,610, rispettivamente) o per le variazioni della massa corporea magra (p = 0,433, p = 0,936 e p = 0,617, rispettivamente).
Bias di pubblicazione e altri dati
I diagrammi a imbuto dell’effetto dell’insulina su MPS e MPB rispetto a SE non hanno mostrato alcun bias di pubblicazione (vedi materiale supplementare elettronico [ESM]). Tra gli articoli esaminati, 15 studi hanno utilizzato dati a tre pool [21, 25, 28, 30-35]. I dati a due pool di dieci di questi studi sono stati inclusi nella meta-analisi, che nel complesso ha riportato risultati simili con visualizzazioni quantitativamente diverse. Tutti e cinque gli studi che hanno riportato esclusivamente dati a tre pool hanno dimostrato che l’Insulina ha migliorato l’assorbimento di proteine NB [25, 33-35]. L’aumento dell’NB in questi studi è stato determinato principalmente da una riduzione della MPB [34] o da un aumento della MPS [25, 33-35]. Nessuno di questi studi prevedeva una riduzione della disponibilità di AA. Due degli studi hanno riportato un aumento della MPS in uno stato iperinsulinemico indotto sperimentalmente in soggetti apparentemente insulino-resistenti con gravi ustioni trattati in un’unità di alta dipendenza [33, 35].
Caratteristiche degli studi che riportano dati su tre pool. I valori sono medie ± SEM, se non diversamente specificato. aI numeri accanto al nome dell’autore distinguono studi diversi dello stesso autore; le lettere indicano interventi o caratteristiche dei partecipanti diversi all’interno di uno stesso studio. bRispetto al basale.
Discussioni conclusive
L’infusione sistemica di Insulina porta a ipoglicemia e a una ridotta disponibilità di AA (ipoaminoacidemia) per la sintesi proteica. Per ovviare a queste conseguenze, glucosio e AA vengono co-infusi per mantenere la glicemia target e la disponibilità di AA. L’infusione locale di Insulina intra-arteriosa sembra limitare l’effetto dell’ipoglicemia sistemica e dell’ipoaminoacidemia, evitando così la necessità di una co-infusione obbligatoria di glucosio (o AA) [33].
La meta-analisi di questi 25 studi non ha mostrato alcun effetto significativo dell’Insulina sulla MPS. L’analisi dei sottogruppi, tuttavia, ha rivelato che negli individui sani l’effetto dell’Insulina sulla MPS diventa significativo solo quando viene aumentata la somministrazione di AA al muscolo scheletrico. Questi risultati sono stati replicati da altri ricercatori in studi in cui la coinfusione di AA e Insulina ha aumentato con successo l’apporto di AA al muscolo [30, 31, 36-42]. In uno studio di Fujita et al, l’esercizio fisico per 45 minuti ha aumentato con successo l’apporto di AA e la MPS rispetto ai controlli non allenati, sebbene l’esercizio fisico di per sé abbia effetti anabolici acuti sulla MPS muscolare [21]. In alcuni studi, tuttavia, l’aumento dell’apporto di AA non ha prodotto un aumento di MPS indotto dall’Insulina [16, 43]. Ciò è probabilmente dovuto al fatto che l’aumento dell’apporto di AA era minimo e ottenuto principalmente attraverso l’aumento del flusso sanguigno, piuttosto che attraverso un aumento della concentrazione di AA. Gli anziani mostrano una resistenza all’effetto anabolico dell’Insulina rispetto alle loro controparti più giovani, probabilmente attraverso meccanismi legati alla disfunzione endoteliale, alla ridotta perfusione tissutale e all’attenuazione della segnalazione anabolica, piuttosto che a una ridotta tolleranza al glucosio [30]. Tuttavia, in presenza di un maggiore apporto di AA, l’Insulina sembra conservare il suo effetto anabolico negli anziani sani. Questo sembra essere il caso sia che l’aumento dell’apporto di AA al muscolo sia ottenuto tramite concentrazioni fisiologiche [24, 38] o sovrafisiologiche [30] di Insulina, farmacologicamente con nitroprussiato di sodio [31] o tramite esercizio fisico [21].
L’aumento delle concentrazioni di Insulina nell’intervallo postprandiale non sembra influire sulla MPS. Uno studio precedente ha riportato che, con aumenti incrementali delle concentrazioni di AA, la MPS ha risposto positivamente a concentrazioni di Insulina di 139,0-194,5 pmol/l, aumentando del 22% rispetto al basale e del 72% quando sono state somministrate concentrazioni di AA più elevate [37]. D’altra parte, un altro studio ha riportato che, in presenza di concentrazioni fisse di AA, l’aumento della concentrazione di Insulina da 34,7 pmol/l a 500,0 pmol/l non ha prodotto ulteriori incrementi significativi della MPS [39]. L’Insulina non ha avuto alcun effetto sulla MPS quando l’apporto di AA è rimasto invariato rispetto al basale. Questi risultati sono supportati anche da altri studi [17, 20, 28-31, 34, 44]. In tutti gli studi sull’uomo sano in cui la disponibilità di AA è stata ridotta, la MPS si è ridotta o è rimasta invariata [18, 19, 37, 45, 46], anche in presenza di concentrazioni sovrafisiologiche di Insulina [18].
La meta-analisi dei 25 studi ha dimostrato che l’Insulina esercita la sua regolazione della massa muscolare magra principalmente attraverso un effetto anticatabolico nella riduzione della MPB. Questo è più evidente se si considerano i dati sulla NB, che hanno mostrato un effetto positivo sulla massa muscolare. Pertanto, le capacità proanaboliche dell’Insulina sono prevalentemente guidate dalla sua capacità di attenuare la MPB scheletrica, piuttosto che da un effetto positivo sulla MPS. Questo risultato è in accordo con le valutazioni di altri ricercatori [16, 29].
La riduzione della MPB indotta dall’Insulina sembra essere più potente quando gli AA sono scarsi. Questi risultati sono coerenti con altri studi che hanno riportato una riduzione della risposta all’Insulina [30, 32, 37, 39, 42, 44], ad eccezione di tre studi che non hanno osservato alcun cambiamento significativo nella MPB [45, 47, 48]. Ciò potrebbe essere dovuto alla resistenza anabolica all’Insulina in una popolazione di studio relativamente anziana e alla presenza di diabete mellito. È interessante notare che l’inibizione massima della MPB da parte dell’Insulina si verifica in risposta a incrementi molto modesti della concentrazione di Insulina (cioè a 104,2 pmol/l) [44].
È stato riferito che il diabete attenua l’effetto positivo dell’Insulina sulle MPB in presenza di una somministrazione prolungata di AA [17, 20, 49]. Tuttavia, in risposta a un trattamento intensivo a lungo termine con Insulina s.c., Halvatsiotis et al. non hanno riscontrato differenze nelle MPS mitocondriali, sarcoplasmatiche o miste rispetto ai controlli sani che non ricevevano Insulina [50]. Non è noto se l’aumento dell’assunzione di AA nei pazienti diabetici trattati con Insulina possa portare a un aumento della massa muscolare. Tuttavia, dato il ruolo facilitante dell’Insulina nel mantenimento della massa muscolare, in particolare in presenza di una maggiore disponibilità di AA, si può ipotizzare la necessità di consigliare ai pazienti con diabete in trattamento insulinico di aumentare l’assunzione di AA per sfruttare gli effetti positivi dell’Insulina sul metabolismo muscolare. Nei pazienti gravemente malati, dove ci si aspetta una significativa resistenza all’Insulina, sono stati osservati aumenti della MPS, ma solo quando sono state utilizzate concentrazioni sovrafisiologiche di Insulina [33, 51]. In uno studio condotto su individui affetti da obesità [40] e insufficienza cardiaca [42], è stato dimostrato che la MPB si riduce in risposta all’Insulina. Questa riduzione, tuttavia, è stata significativamente inferiore a quella osservata nei controlli sani. È evidente che sono necessari ulteriori studi per comprendere appieno il ruolo dell’insulino-resistenza nella regolazione di MPS e MPB nel diabete di tipo II.
Comunque, la review qui esposta presenta diverse limitazioni. I diversi metodi di stima del metabolismo proteico del muscolo scheletrico e la mancanza di dati primari disponibili hanno reso difficile eseguire una valutazione quantitativa completa mediante meta-analisi per tutti gli studi che soddisfacevano i criteri di inclusione della review sistematica. Riconosciamo inoltre che l’uso di più studi per ogni pubblicazione significa che il pooling degli studi non è del tutto indipendente. Inoltre, poiché si tratta di una meta-analisi di studi sperimentali, non è stato possibile effettuare una valutazione completa dei bias, come normalmente si fa nelle meta-analisi di studi controllati randomizzati [52] (ad esempio, generazione della sequenza e occultamento dell’allocazione per controllare i bias di selezione; cecità per controllare eventuali bias di performance, dati di esito incompleti, bias di segnalazione selettiva e altre fonti di bias), poiché nessuno degli studi ha utilizzato questi metodi per l’allocazione dei pazienti. Inoltre, poiché tutti gli studi erano di dimensioni simili, l’analisi del funnel plot potrebbe non essere completamente affidabile nell’informarci di eventuali bias di pubblicazione.
In sintesi, questa review sistematica e meta-analisi suggerisce che il ruolo principale dell’Insulina nell’anabolismo del muscolo scheletrico umano è di tipo facilitativo ed è influenzato dalla velocità di somministrazione degli AA. In situazioni in cui l’apporto di AA è invariato, sono necessarie concentrazioni sovrafisiologiche di Insulina per ottenere l’anabolismo del muscolo scheletrico. Tuttavia, il ruolo dell’Insulina nel ridurre la MPB è chiaramente evidente nella maggior parte degli studi. Questo effetto è attenuato nelle persone anziane e in quelle con resistenza all’Insulina. Questa resistenza è probabilmente legata a un’alterata segnalazione insulinica del metabolismo proteico muscolare e alla disfunzione endoteliale, piuttosto che all’intolleranza al glucosio. Sono necessarie ulteriori prove per tradurre questi risultati in strategie per massimizzare la massa muscolare nei pazienti con diabete insulino-trattato.
Effetto dell’Insulina esogena sull’aumenta del tasso di sintesi proteica muscolare:
L’Insulina è ben nota come ormone chiave responsabile dell’aumento dell’accumulo endogeno di carboidrati e grassi. Tuttavia, il suo ruolo nel metabolismo delle proteine è più controverso. Studi in vitro hanno dimostrato che l’Insulina stimola la sintesi proteica muscolare mediante l’attivazione diretta del meccanismo di traduzione attraverso la via PI3K→Akt→mTORC1 (53,54,55,56,57,58,59,60,61,62,63). L’Insulina può anche influenzare il metabolismo proteico in vivo grazie alle sue proprietà vasoattive. L’aumento postprandiale dell’Insulina circolante stimola la vasodilatazione endotelio-dipendente in virtù della sua azione sull’ossido nitrico sintasi endoteliale, con conseguente maggiore reclutamento capillare, aumento del volume microvascolare e flusso sanguigno nutritivo al tessuto muscolare scheletrico (64). Si potrebbe ipotizzare che la maggiore perfusione postprandiale aumenti l’esposizione del tessuto muscolare ai nutrienti e ai fattori di crescita e aumenti la sintesi proteica muscolare. Tuttavia, se l’Insulina abbia un effetto stimolante sulla sintesi proteica muscolare postprandiale nell’uomo è stato oggetto di un ampio dibattito come, tra l’altro, abbiamo appena visto (65, 66). Molti ritengono che le concentrazioni di Insulina in circolo siano semplicemente permissive, anziché modulatorie, per consentire un aumento della sintesi proteica muscolare in soggetti giovani e sani (66). In particolare, si ritiene che sia necessaria solo una piccola quantità di Insulina per “innescare” il sistema e che sia il successivo aumento della disponibilità di aminoacidi a guidare la risposta della sintesi proteica muscolare post-prandiale (67). Tuttavia, si ipotizza che gli anziani siano più resistenti all’effetto dell’Insulina sulla sintesi proteica muscolare, un difetto associato alla disfunzione endoteliale (68, 69, 70). Questa nuova review sistematica esamina la letteratura esistente sull’effetto proposto dell’aumento dei livelli di Insulina circolante sulla sintesi proteica muscolare in vivo nell’uomo e cerca di definire se tale effetto differisce tra giovani e anziani.
Attivazione della via del mammalian target of rapamycin (mTOR) nella sintesi proteica muscolare da parte di leucina e fattori anabolizzanti. Insulina e IGF-1: fattore di crescita insulino-simile, PKB/Akt: protein chinasi B, AMPK: adenosina monofasfato protein chinasi, mTOR: mammalian target of rapamycin, p70S6K: proteina ribosomiale S6 chinasi, 4E-BP1: proteina legante il fattore di iniziazione eucariotica 4E, eIF4G: fattore di iniziazione eucariotica 4G.
È stata eseguita una review sistematica secondo le linee guida PRISMA (71). In breve, nel gennaio 2014 è stata eseguita una ricerca computerizzata della letteratura utilizzando il database PubMed (http://www.ncbi.nlm.gov/pubmed/) e cercando a mano le liste di riferimento degli studi identificati e le principali review della letteratura. Sono stati utilizzati i seguenti termini di ricerca: Insulina; iperinsulinismo; muscolo; gamba; avambraccio; miofibrillare; anabol; sintesi proteica e accrescimento proteico e le funzioni booleane AND e OR. La ricerca elettronica finale è stata effettuata l’8 agosto 2014.
Tipi di studi Studi clinici che studiano la somministrazione di Insulina a persone sane. Gli studi sono stati limitati a quelli scritti in lingua inglese. Non sono state imposte restrizioni sulla data di pubblicazione.
Tipi di partecipanti Sono stati presi in considerazione partecipanti sani di qualsiasi età che ricevevano Insulina esogena e sono stati stratificati in giovani adulti sani (età media del gruppo tra 18 e 65 anni) o adulti anziani (età media del gruppo ≥65 anni). Per studiare l’effetto dell’età sulla sintesi proteica muscolare insulino-mediata di per sé, sono stati esclusi i soggetti con qualsiasi co-morbilità apparente, incluso il diabete.
Tipi di intervento Questa review è stata limitata agli studi che hanno esaminato la somministrazione di Insulina esogena.
Tipi di misure di esito La misura di esito primaria è la valutazione qualitativa della sintesi proteica muscolare, ossia un aumento significativo o nessun effetto. Gli studi inclusi hanno valutato la sintesi proteica muscolare come misurata dal tasso di scomparsa dei precursori (leg Rd) nel metodo del bilancio arteriovenoso a due vasche, dall’utilizzo intracellulare dei precursori (Fo,m) nel metodo del bilancio arteriovenoso a tre vasche o dal tasso di sintesi frazionale (FSR) nel modello precursore-prodotto (28). Queste misure tendevano a raggiungere valutazioni qualitative simili della sintesi proteica muscolare (dati non mostrati), pertanto non è stata fatta alcuna distinzione tra i modelli per l’interpretazione dell’effetto riportato dell’Insulina sulla sintesi proteica muscolare.
La valutazione dell’idoneità è stata eseguita individualmente da due autori (J Trommelen e B B L Groen). Le divergenze tra i revisori sono state risolte per consenso. I titoli e gli abstract identificati dalla strategia di ricerca sono stati vagliati per la rilevanza, definita dal rispetto di tutti i seguenti criteri: i) soggetti umani, ii) il disegno dello studio era un trial clinico, iii) l’intervento includeva la somministrazione di insulina esogena in almeno uno dei bracci dello studio, iv) valutava la sintesi proteica muscolare mista (Rd, Fo,m o FSR) e v) l’accessibilità al testo completo. Due autori (J Trommelen e B B L Groen) hanno estratto individualmente i dati dagli studi inclusi. Le divergenze tra i revisori sono state risolte per consenso. Da ogni studio incluso sono state estratte informazioni su: i) caratteristiche dei soggetti, tra cui età e numero; ii) modello di valutazione della sintesi proteica muscolare (dati non mostrati); iii) tipo di intervento, tra cui dose, co-intervento, compartimento di infusione e gruppi di confronto; e iv) esito dello studio, tra cui effetto dell’insulina esogena sulla sintesi proteica muscolare, raggruppato in aumento significativo o nessun effetto.
Gli studi sono stati esaminati con una tabulazione completa dei risultati di tutti gli studi inclusi. A causa dell’eterogeneità clinica dei disegni sperimentali, non è stato possibile condurre una meta-analisi. Inoltre, è noto che le differenze nei metodi sperimentali introducono variabilità nella sintesi proteica muscolare, complicando l’analisi quantitativa tra gli studi (si rimanda a Smith et al. (72) per una rassegna su questo argomento). Pertanto, per gli studi inclusi è stata determinata una valutazione qualitativa della sintesi proteica muscolare, ossia un aumento significativo o nessun effetto. Sulla base di questi dati, abbiamo costruito diversi modelli in cui i bracci di studio sono stati esclusi sulla base di motivazioni (biologiche). Nel modello 1 sono stati esclusi i bracci di studio con iperamminoacidemia concomitante. Gli aminoacidi possono stimolare in modo indipendente la sintesi proteica muscolare e il rilascio endogeno di insulina, rendendo impossibile distinguere tra l’effetto dell’insulina e quello dell’infusione di aminoacidi (73, 74). Il modello 2 esclude inoltre i bracci di studio con ipoaminoacidemia insulino-mediata. È stato suggerito che l’effetto dell’insulina sulla sintesi proteica muscolare sia mediato da cambiamenti indotti dall’insulina nell’apporto di aminoacidi al muscolo (75, 76, 77, 78). L’infusione sistemica di insulina induce ipoaminoacidemia, che può limitare l’apporto di aminoacidi al muscolo. Pertanto, in questo modello, sono stati esclusi i bracci di studio che hanno permesso ai livelli di aminoacidi di scendere al di sotto del valore basale. Il modello 3 esclude inoltre i bracci di studio che raggiungono concentrazioni di insulina sovrafisiologiche. Ciò è stato fatto per differenziare tra livelli sovrafisiologici che possono essere raggiunti solo con la somministrazione di Insulina (>1200 pmol/l) e livelli fisiologici alla portata della produzione endogena in risposta a un pasto misto (≤1200 pmol/l). Il modello 4 esclude inoltre i bracci di studio in soggetti anziani, perché è stato suggerito che gli anziani sono più resistenti alle proprietà stimolanti proposte dall’insulina sulla sintesi proteica muscolare (79, 80).
La ricerca nel database PubMed ha fornito un totale di 2021 citazioni. Dal totale di 2025 citazioni, sono stati scartati 1980 studi perché, dopo aver esaminato gli abstract, non soddisfacevano i criteri di inclusione. Il testo completo delle restanti 45 citazioni è stato esaminato in modo più approfondito. Dopo un’attenta lettura del testo integrale, altri dieci studi non soddisfacevano i criteri di inclusione descritti. In totale, 40 studi hanno soddisfatto i criteri di ammissibilità e sono stati inclusi nella revisione sistematica.
Diagramma di flusso dell’identificazione dello studio.AABASAL, somministrazione di aminoacidi per mantenere i livelli basali; AAHYPER, somministrazione di aminoacidi per raggiungere l’iperamminoacidemia; INS, Insulina; INSSUPRA, somministrazione di Insulina per raggiungere i livelli sovrafisiologici; MPS, sintesi proteica muscolare; ↑, aumento significativo; ↔, nessun effetto significativo.
È stata osservata un’elevata eterogeneità metodologica. I design degli studi includevano studi a braccio singolo, a bracci paralleli e crossover. I gruppi di confronto variavano notevolmente, includendo nessun intervento (condizioni basali), soluzione fisiologica, dosaggi insulinici alternativi o protocolli di somministrazione di aminoacidi. I principali criteri di ammissibilità prevedevano un buono stato di salute. La maggior parte degli studi riportava un esame anamnestico preliminare ed esami del sangue standard (principalmente per la valutazione della tolleranza al glucosio). I dosaggi di insulina esogena applicati hanno portato a concentrazioni plasmatiche di insulina che variavano da livelli a digiuno (36 pmol/l) a livelli sovrafisiologici (81 078 pmol/l). Sono stati comunemente utilizzati protocolli di infusione sia sistemici che locali. I protocolli di infusione locale di Insulina utilizzavano l’avambraccio o la gamba come compartimento. Il co-intervento variava notevolmente; il più comune era la coinfusione di aminoacidi.
Sono stati identificati 40 studi da includere nella review, che variavano notevolmente nel design sperimentale. Numerosi studi hanno applicato disegni di ricerca che comprendevano più bracci sperimentali con interventi separati e molti dei quali hanno riportato risultati opposti, rendendo difficile trarre conclusioni a livello di studio. Inoltre, molti degli studi identificati includevano co-interventi, come la somministrazione di farmaci o protocolli di esercizio, che influenzano i risultati. Pertanto, la sintesi dei dati è stata effettuata a livello di braccio di studio.
Dai 40 studi selezionati, dopo l’esclusione degli interventi non correlati ai pasti (cioè la co-somministrazione di farmaci), è stato identificato un totale di 66 bracci di studio che includevano il trattamento insulinico. Di questi 66 bracci di studio, 34 hanno riscontrato un effetto insulino-stimolante sulla sintesi proteica muscolare, mentre 32 non hanno riscontrato tale effetto. Dodici bracci di studio consistevano in dati riutilizzati da altri studi inclusi (81, 82, 83, 84, 85, 86, 87, 88). Dopo la deduplicazione, è stato trovato un totale di 54 bracci di studio unici, di cui 28 hanno riportato un aumento dei tassi di sintesi proteica muscolare, mentre 26 non lo hanno fatto.
Diagramma di flusso dei modelli stepwise che riportano la presenza o l’assenza di un aumento della sintesi proteica muscolare in risposta all’Insulina esogena. MPS, sintesi proteica muscolare; ↑, aumento significativo; ↔, nessun effetto significativo.
Sulla base di questi dati, sono stati costruiti diversi modelli che escludevano i bracci di studio sulla base di un razionale (biologico), come descritto in precedenza. Nel modello 1, sono stati esclusi i bracci di studio in cui la somministrazione di insulina era combinata con co-interventi di aminoacidi che aumentavano gli aminoacidi plasmatici oltre i livelli basali e non avevano un gruppo di confronto per correggere l’aumento degli aminoacidi plasmatici (89, 90, 91, 92, 93, 94, 95, 96, 97, 98). Questo criterio ha escluso 17 bracci di studio, che hanno tutti riportato un aumento della sintesi proteica muscolare.
Il modello 2 escludeva inoltre i bracci di studio in cui i livelli di aminoacidi potevano scendere al di sotto dei livelli basali (99, 100, 101, 102, 102, 103, 104, 105). Questo criterio ha escluso altri dieci bracci di studio rispetto al modello 1, nessuno dei quali ha riportato un effetto sulla sintesi proteica muscolare.
Il modello 3 ha inoltre escluso i bracci di studio in cui è stata raggiunta una concentrazione sovrafisiologica di insulina (106). Questo criterio ha escluso altri due bracci di studio rispetto al modello 2, che hanno entrambi riportato che l’insulina aumenta la sintesi proteica muscolare.
Il modello 4 ha inoltre escluso i bracci di studio in soggetti anziani (107, 108, 109,110, 111). Questo criterio ha escluso altri quattro interventi rispetto al modello 3, uno dei quali ha riportato un effetto insulino-stimolante sulla sintesi proteica muscolare, mentre gli altri tre non lo hanno fatto. Dopo queste esclusioni finali, il modello 4 includeva un totale di 21 bracci di studio, otto dei quali riportavano un aumento della sintesi proteica muscolare, mentre 13 non lo facevano.
Discussione finale
Questa review sistematica ha esaminato la letteratura riguardante l’effetto proposto, legato all’età, della somministrazione di Insulina esogena sui tassi di sintesi proteica muscolare in vivo nell’uomo. Sebbene siano stati condotti numerosi studi per valutare l’impatto della somministrazione di Insulina esogena sulla sintesi proteica muscolare, i dati non supportano un ruolo stimolatorio della somministrazione di Insulina esogena sui tassi di sintesi proteica muscolare in vivo nell’uomo.
Gli aminoacidi sono ben noti per la loro capacità indipendente di stimolare la sintesi proteica muscolare (112, 113). Pertanto, è stato costantemente dimostrato che la somministrazione di Insulina e aminoacidi aumenta la sintesi proteica muscolare (114, 115, 116). Senza un adeguato gruppo di controllo con un grado simile di iperamminoacidemia, è impossibile differenziare le proprietà anaboliche proposte dalla somministrazione di Insulina e aminoacidi. Come previsto, tutti i 17 bracci di studio che combinavano la somministrazione di Insulina e aminoacidi, esclusi dalle analisi in base a questo criterio, hanno riportato un aumento della sintesi proteica muscolare. Va notato che uno stato di iperinsulinemia e iperamminoacidemia concomitanti riflette le condizioni fisiologiche successive all’ingestione di un pasto misto. Tre studi hanno esaminato se la somministrazione di Insulina esogena possa aumentare ulteriormente la sintesi proteica muscolare in condizioni di iperamminoacidemia, e tutti non hanno rilevato un effetto incrementale (60, 75, 83). Questi risultati suggeriscono che l’iperinsulinemia e l’iperamminoacidemia concomitanti aumentano la sintesi proteica muscolare ma, almeno nei soggetti giovani e sani, questo effetto sembra interamente attribuito all’iperamminoacidemia.
La somministrazione di Insulina per via endovenosa è seguita da una riduzione dose-dipendente dei livelli plasmatici di aminoacidi, con gli aminoacidi a catena ramificata più sensibili all’aumento dei livelli circolanti di Insulina (90). Questa ipoaminoacidemia indotta dall’Insulina è il riflesso di un aumento dell’assorbimento di aminoacidi dal plasma in combinazione con l’azione inibitoria proposta dall’aumento dei livelli di Insulina sulla proteolisi endogena (80, 70). È stato suggerito che il proposto effetto positivo della somministrazione di Insulina esogena sulla sintesi proteica muscolare sia mediato dall’aumento del flusso sanguigno indotto dall’Insulina e dal conseguente maggiore apporto di aminoacidi al muscolo. Un calo delle concentrazioni circolanti di aminoacidi può limitare l’apporto di aminoacidi al muscolo e di conseguenza limitare la capacità dell’Insulina di stimolare la sintesi proteica muscolare. Per evitare questo calo dei livelli di aminoacidi, diversi studi hanno applicato infusioni i.v. di aminoacidi in combinazione con la somministrazione di Insulina o hanno infuso Insulina esogena localmente nell’arteria femorale o brachiale. Nel modello 2, sono stati esclusi dieci bracci di studio che hanno mostrato un’ipoaminoacidemia indotta dall’Insulina. Nessuno di questi dieci bracci di studio ha riscontrato un effetto insulino-stimolante dell’Insulina sulla sintesi proteica muscolare, sostenendo il razionale che l’ipoaminoacidemia indotta dall’Insulina possa ovviare alle proprietà stimolanti proposte dalla somministrazione di Insulina sull’apporto di aminoacidi al muscolo e sul conseguente aumento della sintesi proteica muscolare.
È stato dimostrato che l’Insulina aumenta la sintesi proteica muscolare in vitro (117, 118, 119). Tuttavia, dagli studi in vivo sull’uomo emergono molte discrepanze sugli effetti positivi proposti dall’Insulina esogena sui tassi di sintesi proteica muscolare. È stato suggerito che l’apparente discrepanza sia attribuita alle concentrazioni di Insulina sovrafisiologiche più che decuplicate (14.000 pmol/l o superiori) applicate nei modelli in vitro rispetto agli aumenti più fisiologici dei livelli di Insulina plasmatica (fino a 1.200 pmol/l) applicati nella maggior parte degli studi umani in vivo (53, 60). Uno studio ha somministrato Insulina esogena localmente nell’avambraccio per ottenere livelli locali di Insulina sovrafisiologica superiori a 50.000pmol/l, mentre si bloccavano gli aminoacidi a livelli arteriosi o venosi basali, e ha riportato un aumento dei tassi di sintesi proteica muscolare (60). I loro risultati suggeriscono che i livelli di Insulina sovrafisiologici possono stimolare efficacemente la sintesi proteica muscolare.
È stato ipotizzato che gli anziani siano più resistenti agli stimoli anabolici, come gli aumenti delle concentrazioni plasmatiche circolanti di Insulina e aminoacidi, rispetto agli adulti più giovani (106). La resistenza anabolica alla somministrazione di aminoacidi e/o Insulina nella popolazione anziana potrebbe essere attribuita a un’alterazione dell’apporto di aminoacidi al muscolo stimolato dall’Insulina (58). Di conseguenza, si è ipotizzato che la somministrazione di Insulina esogena possa aumentare la sintesi proteica muscolare negli adulti più anziani (più resistenti all’Insulina). Dopo aver stratificato i dati in giovani e anziani, è stato identificato uno studio che ha riportato un effetto positivo della somministrazione di Insulina esogena sulla sintesi proteica muscolare nei soggetti anziani (54), mentre tre bracci di studio non hanno osservato tale effetto (112, 113). È interessante notare che l’aumento della sintesi proteica muscolare stimolato dall’Insulina nei soggetti anziani è stato osservato solo in presenza di livelli locali di Insulina relativamente elevati, superiori a 900 pmol/l (54). Dosaggi inferiori di somministrazione locale di Insulina, con livelli plasmatici locali di Insulina di ∼500 pmol, non sembravano aumentare la sintesi proteica muscolare negli anziani. Questi dati suggeriscono che gli anziani potrebbero essere più resistenti alle proprietà anaboliche dell’Insulina, una resistenza che potrebbe essere superata con concentrazioni di Insulina più elevate.
Nel modello 4 sono stati applicati criteri di esclusione rigorosi per escludere i fattori che potrebbero modulare l’effetto stimolante proposto dalla somministrazione di Insulina esogena negli adulti sani, tra cui:
iperamminoacidemia concomitante;
ipoaminoacidemia indotta dall’Insulina;
concentrazioni di insulina sovrafisiologiche e
soggetti più anziani e più resistenti all’Insulina.
Sono stati identificati otto bracci di studio che hanno riportato un aumento della sintesi proteica muscolare stimolata dall’Insulina, mentre 14 bracci di studio non hanno osservato un aumento dei tassi di sintesi proteica muscolare stimolata dall’Insulina in soggetti giovani e sani. Un sottogruppo del modello 4 comprende otto bracci di studio in cui l’Insulina viene infusa localmente nella gamba, di cui cinque bracci di studio hanno riportato un aumento del tasso di sintesi proteica muscolare, mentre tre non lo hanno fatto. Tre di questi bracci di studio provengono da uno studio dose-risposta, in cui la somministrazione di Insulina a bassa dose (0,05 mU/min×100 ml di gamba) e ad alta dose (0,30 mU/min×100 ml di gamba) non ha aumentato il tasso di sintesi proteica muscolare, mentre la dose intermedia (0,15 mU/min×100 ml di gamba) sì (55). Questa dose intermedia ha aumentato i tassi di sintesi proteica muscolare in tutti e cinque i bracci di studio in cui è stata applicata (7, 24, 28, 54, 55), mentre non è stato osservato alcun aumento dei tassi di sintesi proteica muscolare nei tre bracci di studio che hanno applicato un dosaggio alternato (55, 61). Ciò suggerisce un effetto dose-risposta a forma di U della somministrazione di Insulina esogena sulla sintesi proteica muscolare, dove una dose di ∼0,15mU/min×100 ml di gamba può stimolare la sintesi proteica muscolare. È interessante notare che la somministrazione di Insulina esogena non sembra stimolare la sintesi proteica muscolare quando viene infusa localmente nell’avambraccio. In questo sottogruppo del modello 4, tutti e sei gli interventi non hanno riportato alcun aumento dei tassi di sintesi proteica muscolare, nonostante un’ampia gamma di protocolli di dosaggio studiati (58, 62, 63). Nel tentativo di delineare ulteriormente i risultati, i dati sono stati esaminati per individuare altri potenziali fattori modulanti. La presenza o l’assenza di un effetto stimolante in questi studi non poteva essere attribuita a differenze nei livelli di Insulina circolante o alla scelta dei traccianti aminoacidici.
In tutti i lavori presentati, l’Insulina esogena è stata somministrata con un approccio basato sul clamp. Questo approccio può avere dei limiti, in quanto si potrebbe ipotizzare che il forte aumento postprandiale del livello di Insulina circolante abbia una funzione regolatoria per attivare vari processi fisiologici che facilitano l’aumento postprandiale del tasso di sintesi proteica muscolare. Questi cambiamenti temporali nella secrezione di Insulina, nell’apporto e nell’assorbimento di aminoacidi e nella segnalazione intramiocellulare devono essere strettamente regolati per sostenere la risposta anabolica postprandiale. Inoltre, è stato notato che potrebbero esserci differenze nella rilevanza dei livelli di Insulina circolante sulla modulazione della sintesi di varie (serie di) proteine nel tessuto muscolare scheletrico (60).
Dai dati presentati nell’attuale review sistematica, si può concludere che:
la somministrazione esogena di Insulina e aminoacidi aumenta efficacemente la sintesi proteica muscolare; tuttavia, questo effetto è attribuito all’iperaminoacidemia;
l’Insulina esogena somministrata per via sistemica induce ipoaminoacidemia, che ovvia a qualsiasi effetto insulino-stimolatorio sulla sintesi proteica muscolare;
l’Insulina esogena che determina livelli di Insulina sovrafisiologici superiori a 50.000 pmol/l può stimolare efficacemente la sintesi proteica muscolare anche se i livelli di soglia minima sono di 1.200pmol/l;
l’Insulina esogena può avere un effetto ridotto sulla sintesi proteica muscolare negli adulti più anziani a causa della resistenza anabolica legata all’età;
l’Insulina esogena somministrata in range fisiologici per via sistemica non aumenta la sintesi proteica muscolare nei giovani adulti sani.
In definitiva, in base ai dati raccolti dalla letteratura esistente, gli autori della review concludono che la somministrazione di Insulina esogena non aumenta i tassi di sintesi proteica muscolare negli adulti sani, giovani o anziani se non superando la soglia fisiologica.
Di conseguenza, ci troviamo di fronte ad un peptide con funzione prevalentemente anticatabolica e soggetto nella sua efficacia al grado di insulino-sensibilità tissutale?… Così sembrerebbe, specie in contesto fisiologico endogeno e indotto esogenamente. Ma se all’equazione ci aggiungessimo altre variabili che influenzino la risposta di base fisiologica e che non interessino il semplice dosaggio di Insulina utilizzato?…
Continua…
Gabriel Bellizzi
Riferimenti:
Bier DM (1989) Intrinsically difficult problems: the kinetics of body proteins and amino acids in man. Diabetes Metab Rev 5:111–132ArticleCASPubMedGoogle Scholar
Prod’homme M, Rieu I, Balage M, Dardevet D, Grizard J (2004) Insulin and amino acids both strongly participate to the regulation of protein metabolism. Curr Opin Clin Nutr Metab Care 7:71–77ArticlePubMedGoogle Scholar
McNurlan MA, Garlick PJ (1989) Influence of nutrient intake on protein turnover. Diabetes Metab Rev 5:165–189ArticleCASPubMedGoogle Scholar
Cruz-Jentoft AJ, Landi F, Schneider SM et al (2014) Prevalence and intervention for sarcopenia in ageing adults: a systematic review. Report of the International Sarcopenia Initiative (EWGSOP and IWGS). Age Ageing 43:748–759PubMed CentralArticlePubMedGoogle Scholar
Atherton PJ, Etheridge T, Watt PW et al (2010) Muscle full effect after oral protein: time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am J Clin Nutr 92:1080–1088ArticleCASPubMedGoogle Scholar
Wolfson L, Judge J, Whipple R, King M (1995) Strength is a major factor in balance, gait, and the occurrence of falls. J Gerontol A Biol Sci Med Sci 50:64–67PubMedGoogle Scholar
Tinetti ME, Williams CS (1997) Falls, injuries due to falls, and the risk of admission to a nursing home. N Engl J Med 337:1279–1284ArticleCASPubMedGoogle Scholar
Park SW, Goodpaster BE, Lee JS et al (2009) Excessive loss of skeletal muscle mass in older adults with type 2 diabetes. Diabetes Care 32:1993–1997PubMed CentralArticlePubMedGoogle Scholar
Mavros Y, Kay S, Anderberg KA et al (2013) Changes in insulin resistance and HbA1c are related to exercise-mediated changes in body composition in older adults with type 2 diabetes: interim outcomes from the GREAT2DO trial. Diabetes Care 36:2372–2379PubMed CentralArticleCASPubMedGoogle Scholar
Prospective UK (1995) Diabetes Study Group. UK Prospective Diabetes Study 16. Overview of 6 years’ therapy of type II diabetes: a progressive disease. Diabetes 44:1249–1258ArticleGoogle Scholar
Koro CE, Bowlin SJ, Bourgeois N, Fedder DO (2004) Glycemic control from 1988 to 2000 among U.S. adults diagnosed with type 2 diabetes: a preliminary report. Diabetes Care 27:17–20ArticlePubMedGoogle Scholar
Biesenbach G, Raml A, Alsaraji N (2006) Weight gain and insulin requirement in type 2 diabetic patients during the first year after initiating insulin therapy dependent on baseline BMI. Diabetes Obes Metab 8:669–673ArticleCASPubMedGoogle Scholar
Packianathan IC, Fuller NJ, Peterson DB, Wright A, Coward WA, Finer N (2005) Use of a reference four-component model to define the effects of insulin treatment on body composition in type 2 diabetes: the ‘Darwin study’. Diabetologia 48:222–229ArticleCASPubMedGoogle Scholar
Pain VM, Albertse EC, Garlick PJ (1983) Protein metabolism in skeletal muscle, diaphragm, and heart of diabetic rats. Am J Physiol 245:E604–E610CASPubMedGoogle Scholar
Garlick PJ, Grant I (1988) Amino acid infusion increases the sensitivity of muscle protein synthesis in vivo to insulin. Effect of branched-chain amino acids. Biochem J 254:579–584PubMed CentralArticleCASPubMedGoogle Scholar
Gelfand RA, Barrett EJ (1987) Effect of physiologic hyperinsulinemia on skeletal muscle protein synthesis and breakdown in man. J Clin Invest 80:1–6PubMed CentralArticleCASPubMedGoogle Scholar
Pacy PJ, Nair KS, Ford C, Halliday D (1989) Failure of insulin infusion to stimulate fractional muscle protein synthesis in type I diabetic patients; anabolic effect of insulin and decreased proteolysis 38:612–624
Denne SC, Liechty EA, Liu YM, Brechtel G, Baron AD (1991) Proteolysis in skeletal muscle and whole body in response to euglycemic hyperinsulinemia in normal adults. Am J Physiol 261:E809–E814CASPubMedGoogle Scholar
Louard RJ, Fryburg DA, Gelfand RA, Barrett EJ (1992) Insulin sensitivity of protein and glucose metabolism in human forearm skeletal muscle. J Clin Invest 90:2348–2354PubMed CentralArticleCASPubMedGoogle Scholar
Bell JA, Volpi E, Fujita S et al (2006) Skeletal muscle protein anabolic response to increased energy and insulin is preserved in poorly controlled type 2 diabetes. J Nutr 136:1249–1255PubMed CentralCASPubMedGoogle Scholar
Fujita S, Rasmussen BB, Cadenas JG et al (2007) Aerobic exercise overcomes the age-related insulin resistance of muscle protein metabolism by improving endothelial function and Akt/mammalian target of rapamycin signaling. Diabetes 56:1615–1622PubMed CentralArticleCASPubMedGoogle Scholar
Biolo G, Fleming RY, Maggi SP, Wolfe RR (1995) Transmembrane transport and intracellular kinetics of amino acids in human skeletal muscle. Am J Physiol 268:E75–E84CASPubMedGoogle Scholar
Newman E, Heslin MJ, Wolf RF, Pisters PW, Brennan MF (1994) The effect of systemic hyperinsulinemia with concomitant amino acid infusion on skeletal muscle protein turnover in the human forearm. Metabolism 43:70–78ArticleCASPubMedGoogle Scholar
Biolo G, Declan Fleming RY, Wolfe RR (1995) Physiologic hyperinsulinemia stimulates protein synthesis and enhances transport of selected amino acids in human skeletal muscle. J Clin Invest 95:811–819PubMed CentralArticleCASPubMedGoogle Scholar
Fryburg DA, Jahn LA, Hill SA, Oliveras DM, Barrett EJ (1995) Insulin and insulin-like growth factor-I enhance human skeletal muscle protein anabolism during hyperaminoacidemia by different mechanisms. J Clin Invest 96:1722–1729PubMed CentralArticleCASPubMedGoogle Scholar
Luzi L, Piceni Sereni L, Spessot M et al (2000) Postabsorptive muscle protein metabolism in type 1 diabetic patients after pancreas transplantation. Acta Diabetol 37:219–224ArticleCASPubMedGoogle Scholar
Rasmussen BB, Fujita S, Wolfe RR et al (2006) Insulin resistance of muscle protein metabolism in aging. FASEB J 20:768–769PubMed CentralCASPubMedGoogle Scholar
Fujita S, Rasmussen BB, Cadenas JG, Grady JJ, Volpi E (2006) Effect of insulin on human skeletal muscle protein synthesis is modulated by insulin-induced changes in muscle blood flow and amino acid availability. Am J Physiol Endocrinol Metab 291:E745–E754PubMed CentralArticleCASPubMedGoogle Scholar
Fujita S, Glynn EL, Timmerman KL, Rasmussen BB, Volpi E (2009) Supraphysiological hyperinsulinaemia is necessary to stimulate skeletal muscle protein anabolism in older adults: evidence of a true age-related insulin resistance of muscle protein metabolism. Diabetologia 52:1889–1898PubMed CentralArticleCASPubMedGoogle Scholar
Timmerman KL, Lee JL, Fujita S et al (2010) Pharmacological vasodilation improves insulin-stimulated muscle protein anabolism but not glucose utilization in older adults. Diabetes 59:2764–2771PubMed CentralArticleCASPubMedGoogle Scholar
Timmerman KL, Lee JL, Dreyer HC et al (2010) Insulin stimulates human skeletal muscle protein synthesis via an indirect mechanism involving endothelial-dependent vasodilation and mammalian target of rapamycin complex 1 signaling. J Clin Endocrinol Metab 95:3848–3857PubMed CentralArticleCASPubMedGoogle Scholar
Gore DC, Herndon DN, Wolfe RR (2005) Comparison of peripheral metabolic effects of insulin and metformin following severe burn injury. J Trauma Inj Infect Crit Care 59:316–323ArticleCASGoogle Scholar
Biolo G, Williams BD, Fleming RY, Wolfe RR (1999) Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes 48:949–957ArticleCASPubMedGoogle Scholar
Ferrando AA, Chinkes DL, Wolf SE, Matin S, Herndon DN, Wolfe RR (1999) A submaximal dose of insulin promotes net skeletal muscle protein synthesis in patients with severe burns. Ann Surg 229:11–18PubMed CentralArticleCASPubMedGoogle Scholar
Hillier TA, Fryburg DA, Jahn LA, Barrett EJ, Teresa A (1998) Extreme hyperinsulinemia unmasks insulin’s effect to stimulate protein synthesis in the human forearm. Am J Physiol Endocrinol Metab 247:E1067–E1074Google Scholar
Nygren J, Nair KS (2003) Differential regulation of protein dynamics in splanchnic and skeletal muscle beds by insulin and amino acids in healthy human subjects. Diabetes 52:1377–1385ArticleCASPubMedGoogle Scholar
Guillet C, Prod’homme M, Balage M et al (2004) Impaired anabolic response of muscle protein synthesis is associated with S6K1 dysregulation in elderly humans. FASEB J 18:1586–1587CASPubMedGoogle Scholar
Greenhaff PL, Karagounis LG, Peirce N et al (2008) Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am J Physiol Endocrinol Metab 295:595–604ArticleGoogle Scholar
Guillet C, Delcourt I, Rance M et al (2009) Changes in basal and insulin and amino acid response of whole body and skeletal muscle proteins in obese men. J Clin Endocrinol Metab 94:3044–3050ArticleCASPubMedGoogle Scholar
Smith GI, Atherton P, Reed DN et al (2009) No major sex differences in muscle protein synthesis rates in the postabsorptive state and during hyperinsulinemia-hyperaminoacidemia in middle-aged adults. J Appl Physiol 107:1308–1315PubMed CentralArticleCASPubMedGoogle Scholar
Toth MJ, LeWinter MM, Ades PA, Matthews DE (2010) Impaired muscle protein anabolic response to insulin and amino acids in heart failure patients: relationship with markers of immune activation. Clin Sci (Lond) 119:467–476
Heslin MJ, Newman E, Wolf RF, Pisters PW, Brennan MF (1992) Effect of hyperinsulinemia on whole body and skeletal muscle leucine carbon kinetics in humans. Am J Physiol 262:E911–E918CASPubMedGoogle Scholar
Wilkes EA, Selby AL, Atherton PJ et al (2009) Blunting of insulin inhibition of proteolysis in legs of older subjects may contribute to age-related sarcopenia. Am J Clin Nutr 90:1343–1350ArticleCASPubMedGoogle Scholar
Arfvidsson B, Zachrisson H, Moller-Loswick AC, Hyltander A, Sandstrom R, Lundholm K (1991) Effect of systemic hyperinsulinemia on amino acid flux across human legs in postabsorptive state. Am J Physiol 260:E46–E52CASPubMedGoogle Scholar
Barazzoni R, Short KR, Asmann Y, Coenen-Schimke JM, Robinson MM, Nair KS (2012) Insulin fails to enhance mTOR phosphorylation, mitochondrial protein synthesis, and ATP production in human skeletal muscle without amino acid replacement. Am J Physiol Endocrinol Metab 303:E1117–E1125PubMed CentralArticleCASPubMedGoogle Scholar
Wolf RF, Heslin MJ, Newman E, Pearlstone DB, Gonenne A, Brennan MF (1992) Growth hormone and insulin combine to improve whole-body and skeletal muscle protein kinetics. Surgery 112:284–291, discussion 291–292CASPubMedGoogle Scholar
Bell JA, Fujita S, Volpi E, Cadenas JG, Rasmussen BB (2005) Short-term insulin and nutritional energy provision do not stimulate muscle protein synthesis if blood amino acid availability decreases. Am J Physiol Endocrinol Metab 289:E999–E1006PubMed CentralArticleCASPubMedGoogle Scholar
Charlton MR, Balagopal P, Nair KS (1997) Skeletal muscle myosin heavy chain synthesis in type 1 diabetes. Diabetes 46:1336–1340ArticleCASPubMedGoogle Scholar
Halvatsiotis P, Short KR, Bigelow M, Nair KS (2002) Synthesis rate of muscle proteins, muscle functions, and amino acid kinetics in type 2 diabetes. Diabetes 51:2395–2404ArticleCASPubMedGoogle Scholar
Sakurai Y, Aarsland A, Herndon DN et al (1995) Stimulation of muscle protein synthesis by long-term insulin infusion in severely burned patients. Ann Surg 222:283–297PubMed CentralArticleCASPubMedGoogle Scholar
Wang C, Mamza J, Idris I (2015) Biphasic vs basal bolus insulin regimen in type 2 diabetes: a systematic review and meta-analysis of randomized controlled trials. Diabet Med 32:585–594
Cuthbertson D, Smith K, Babraj J, Leese G, Waddell T, Atherton P, Wackerhage H, Taylor PM, Rennie MJ. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB Journal 2005 19 422–424. (doi:10.1096/fj.04-2640fje).
Kinsella K & He W. An aging world: 2008. In International Population Reports, pp P95/09-91. U.S. Department of Health and Human Services, 2009. http://www.census.gov/prod/2009pubs/p95-09-1.pdf.
Volpi E, Sheffield-Moore M, Rasmussen BB, Wolfe RR. Basal muscle amino acid kinetics and protein synthesis in healthy young and older men. Journal of the American Medical Association 2001 286 1206. (doi:10.1001/jama.286.10.1206).
Koopman R, van Loon LJC. Aging, exercise, and muscle protein metabolism. Journal of Applied Physiology 2009 106 2040. (doi:10.1152/japplphysiol.91551.2008).
Katsanos CS, Kobayashi H, Sheffield-Moore M, Aarsland A, Wolfe RR. Aging is associated with diminished accretion of muscle proteins after the ingestion of a small bolus of essential amino acids. American Journal of Clinical Nutrition 2005 82 1065–1073.
Timmerman KL, Lee JL, Dreyer HC, Dhanani S, Glynn EL, Fry CS, Drummond MJ, Sheffield-Moore M, Rasmussen BB, Volpi E. Insulin stimulates human skeletal muscle protein synthesis via an indirect mechanism involving endothelial-dependent vasodilation and mammalian target of rapamycin complex 1 signaling. Journal of Clinical Endocrinology and Metabolism 2010 95 3848–3857. (doi:10.1210/jc.2009-2696).
Frayn KN, Maycock PF. Regulation of protein metabolism by a physiological concentration of insulin in mouse soleus and extensor digitorum longus muscles. Effects of starvation and scald injury. Biochemical Journal 1979 184 323.
Fulks RM, Li JB, Goldberg AL. Effects of insulin, glucose, and amino acids on protein turnover in rat diaphragm. Journal of Biological Chemistry 1975 250 290–298.
Jefferson L, Rannels D, Munger B, Morgan H. Insulin in the regulation of protein turnover in heart and skeletal muscle. Federation Proceedings 1974 33 1098–1104.
Jefferson L, Li J, Rannels S. Regulation by insulin of amino acid release and protein turnover in the perfused rat hemicorpus. Journal of Biological Chemistry 1977 252 1476.
Li J, Higgins J, Jefferson L. Changes in protein turnover in skeletal muscle in response to fasting. American Journal of Physiology. Endocrinology and Metabolism 1979 236 E222.
Lundholm K, Edström S, Ekman L, Karlberg I, Walker P, Schersten T. Protein degradation in human skeletal muscle tissue: the effect of insulin, leucine, amino acids and ions. Clinical Science 1981 60 319.
Jefferson L, Koehler J, Morgan H. Effect of insulin on protein synthesis in skeletal muscle of an isolated perfused preparation of rat hemicorpus. PNAS 1972 69 816. (doi:10.1073/pnas.69.4.816).
Stirewalt WS, Low R, Slaiby JM. Insulin sensitivity and responsiveness of epitrochlearis and soleus muscles from fed and starved rats. Recognition of differential changes in insulin sensitivities of protein synthesis and glucose incorporation into glycogen. Biochemical Journal 1985 227 355.
Kimball SR, Vary TC, Jefferson LS. Regulation of protein synthesis by insulin. Annual Review of Physiology 1994 56 321–348. (doi:10.1146/annurev.ph.56.030194.001541).
Phillips SM. Insulin and muscle protein turnover in humans: stimulatory, permissive, inhibitory, or all of the above? American Journal of Physiology. Endocrinology and Metabolism 2008 295 E731. (doi:10.1152/ajpendo.90569.2008).
Timmerman KL, Volpi E. Amino acid metabolism and regulatory effects in aging. Current Opinion in Clinical Nutrition and Metabolic Care 2008 11 45. (doi:10.1097/MCO.0b013e3282f2a592).
Greenhaff PL, Karagounis L, Peirce N, Simpson EJ, Hazell M, Layfield R, Wackerhage H, Smith K, Atherton P, Selby A. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. American Journal of Physiology. Endocrinology and Metabolism 2008 295 E595–E604. (doi:10.1152/ajpendo.90411.2008).
Fujita S, Glynn EL, Timmerman KL, Rasmussen BB, Volpi E. Supraphysiological hyperinsulinaemia is necessary to stimulate skeletal muscle protein anabolism in older adults: evidence of a true age-related insulin resistance of muscle protein metabolism. Diabetologia 2009 52 1889–1898. (doi:10.1007/s00125-009-1430-8).
Meneilly G, Elliot T, Bryer-Ash M, Floras J. Insulin-mediated increase in blood flow is impaired in the elderly. Journal of Clinical Endocrinology and Metabolism 1995 80 1899.
Rasmussen BB, Fujita S, Wolfe RR, Mittendorfer B, Roy M, Rowe VL, Volpi E. Insulin resistance of muscle protein metabolism in aging. FASEB Journal 2006 20 768–769. (doi:10.1096/fj.05-4607fje).
Moher D, Liberati A, Tetzlaff J, Altman DG. Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. PLoS Medicine 2009 6 e1000097. (doi:10.1371/journal.pmed.1000097).
Biolo G, Declan Fleming RY, Wolfe RR. Physiologic hyperinsulinemia stimulates protein synthesis and enhances transport of selected amino acids in human skeletal muscle. Journal of Clinical Investigation 1995 95 811–819. (doi:10.1172/JCI117731).
Smith GI, Patterson BW, Mittendorfer B. Human muscle protein turnover – why is it so variable? Journal of Applied Physiology 2011 110 480–491. (doi:10.1152/japplphysiol.00125.2010).
Bennet W, Connacher A, Scrimgeour C, Smith K, Rennie M. Increase in anterior tibialis muscle protein synthesis in healthy man during mixed amino acid infusion: studies of incorporation of [1-13C]leucine. Clinical Science 1989 76 447.
Kimball S. The role of nutrition in stimulating muscle protein accretion at the molecular level. Biochemical Society Transactions 2007 35 1298–1301. (doi:10.1042/BST0351298).
Fujita S, Rasmussen BB, Cadenas JG, Drummond MJ, Glynn EL, Sattler FR, Volpi E. Aerobic exercise overcomes the age-related insulin resistance of muscle protein metabolism by improving endothelial function and Akt/mammalian target of rapamycin signaling. Diabetes 2007 56 1615–1622. (doi:10.2337/db06-1566).
Louard RJ, Fryburg DA, Gelfand RA, Barrett EJ. Insulin sensitivity of protein and glucose metabolism in human forearm skeletal muscle. Journal of Clinical Investigation 1992 90 2348–2354. (doi:10.1172/JCI116124).
Bell JA, Volpi E, Fujita S, Cadenas JG, Sheffield-Moore M, Rasmussen BB. Skeletal muscle protein anabolic response to increased energy and insulin is preserved in poorly controlled type 2 diabetes. Journal of Nutrition 2006 136 1249–1255.
Newman E, Heslin MJ, Wolf RF, Pisters PW, Brennan MF. The effect of systemic hyperinsulinemia with concomitant amino acid infusion on skeletal muscle protein turnover in the human forearm. Metabolism 1994 43 70–78. (doi:10.1016/0026-0495(94)90159-7).
Drummond MJ, Bell JA, Fujita S, Dreyer HC, Glynn EL, Volpi E, Rasmussen BB. Amino acids are necessary for the insulin-induced activation of mTOR/S6K1 signaling and protein synthesis in healthy and insulin resistant human skeletal muscle. Clinical Nutrition 2008 27 447–456. (doi:10.1016/j.clnu.2008.01.012).
Biolo G, Williams BD, Fleming R, Wolfe R. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes 1999 48 949–957. (doi:10.2337/diabetes.48.5.949).
Zanetti M, Barazzoni R, Kiwanuka E, Tessari P. Effects of branched-chain-enriched amino acids and insulin on forearm leucine kinetics. Clinical Science 1999 97 437–448. (doi:10.1042/CS19990163).
Hillier TA, Fryburg DA, Jahn LA, Barrett EJ. Extreme hyperinsulinemia unmasks insulin’s effect to stimulate protein synthesis in the human forearm. American Journal of Physiology 1998 274 E1067–E1074.
Nygren J, Nair KS. Differential regulation of protein dynamics in splanchnic and skeletal muscle beds by insulin and amino acids in healthy human subjects. Diabetes 2003 52 1377–1385. (doi:10.2337/diabetes.52.6.1377).
Guillet C, Prod’homme M, Balage M, Gachon P, Giraudet C, Morin L, Grizard J, Boirie Y. Impaired anabolic response of muscle protein synthesis is associated with S6K1 dysregulation in elderly humans. FASEB Journal 2004 18 1586–1587.
Guillet C, Delcourt I, Rance M, Giraudet C, Walrand S, Bedu M, Duche P, Boirie Y. Changes in basal and insulin and amino acid response of whole body and skeletal muscle proteins in obese men. Journal of Clinical Endocrinology and Metabolism 2009 94 3044–3050. (doi:10.1210/jc.2008-2216).
Smith GI, Atherton P, Reeds DN, Mohammed BS, Jaffery H, Rankin D, Rennie MJ, Mittendorfer B. No major sex differences in muscle protein synthesis rates in the postabsorptive state and during hyperinsulinemia–hyperaminoacidemia in middle-aged adults. Journal of Applied Physiology 2009 107 1308–1315. (doi:10.1152/japplphysiol.00348.2009).
Chevalier S, Goulet ED, Burgos SA, Wykes LJ, Morais JA. Protein anabolic responses to a fed steady state in healthy aging. Journals of Gerontology. Series A, Biological Sciences and Medical Sciences 66A 2011 681–688. (doi:10.1093/gerona/glr036).
Smith GI, Atherton P, Reeds DN, Mohammed BS, Rankin D, Rennie MJ, Mittendorfer B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: a randomized controlled trial. American Journal of Clinical Nutrition 2011 93 402–412. (doi:10.3945/ajcn.110.005611).
Tessari P, Barazzoni R, Zanetti M. Differences in estimates of forearm protein synthesis between leucine and phenylalanine tracers following unbalanced amino acid infusion. Metabolism 1999 48 1564–1569. (doi:10.1016/S0026-0495(99)90246-9).
Toth MJ, LeWinter MM, Ades PA, Matthews DE. Impaired muscle protein anabolic response to insulin and amino acids in heart failure patients: relationship to markers of immune activation. Clinical Science 2010 119 467. (doi:10.1042/CS20100110).
Denne SC, Liechty EA, Liu YM, Brechtel G, Baron AD. Proteolysis in skeletal muscle and whole body in response to euglycemic hyperinsulinemia in normal adults. American Journal of Physiology. Endocrinology and Metabolism 1991 261 E809–E814.
Tessari P, Inchiostro S, Biolo G, Vincenti E, Sabadin L. Effects of acute systemic hyperinsulinemia on forearm muscle proteolysis in healthy man. Journal of Clinical Investigation 1991 88 27–33. (doi:10.1172/JCI115287).
Arfvidsson B, Zachrisson H, Moller-Loswick AC, Hyltander A, Sandstrom R, Lundholm K. Effect of systemic hyperinsulinemia on amino acid flux across human legs in postabsorptive state. American Journal of Physiology. Endocrinology and Metabolism 1991 260 E46–E52.
McNurlan MA, Essen P, Thorell A, Calder AG, Anderson SE, Ljungqvist O, Sandgren A, Grant I, Tjader I, Ballmer PE et al.. Response of protein synthesis in human skeletal muscle to insulin: an investigation with l-[2H5]phenylalanine. American Journal of Physiology 1994 267 E102–E108.
Meek SE, Persson M, Ford GC, Nair KS. Differential regulation of amino acid exchange and protein dynamics across splanchnic and skeletal muscle beds by insulin in healthy human subjects. Diabetes 1998 47 1824–1835. (doi:10.2337/diabetes.47.12.1824).
Bell JA, Fujita S, Volpi E, Cadenas JG, Rasmussen BB. Short-term insulin and nutritional energy provision do not stimulate muscle protein synthesis if blood amino acid availability decreases. American Journal of Physiology. Endocrinology and Metabolism 2005 289 E999–E1006. (doi:10.1152/ajpendo.00170.2005).
Fujita S, Rasmussen BB, Cadenas JG, Grady JJ, Volpi E. Effect of insulin on human skeletal muscle protein synthesis is modulated by insulin-induced changes in muscle blood flow and amino acid availability. American Journal of Physiology. Endocrinology and Metabolism 2006 291 E745–E754. (doi:10.1152/ajpendo.00271.2005).
Wilkes EA, Selby AL, Atherton PJ, Patel R, Rankin D, Smith K, Rennie MJ. Blunting of insulin inhibition of proteolysis in legs of older subjects may contribute to age-related sarcopenia. American Journal of Clinical Nutrition 2009 90 1343–1350. (doi:10.3945/ajcn.2009.27543).
Moller-Loswick AC, Zachrisson H, Hyltander A, Korner U, Matthews DE, Lundholm K. Insulin selectively attenuates breakdown of nonmyofibrillar proteins in peripheral tissues of normal men. American Journal of Physiology. Endocrinology and Metabolism 1994 266 E645–E652.
Fryburg DA, Jahn LA, Hill SA, Oliveras DM, Barrett EJ. Insulin and insulin-like growth factor-I enhance human skeletal muscle protein anabolism during hyperaminoacidemia by different mechanisms. Journal of Clinical Investigation 1995 96 1722–1729. (doi:10.1172/JCI118217).
Fukagawa N, Minaker K, Young V, Rowe J. Insulin dose-dependent reductions in plasma amino acids in man. American Journal of Physiology. Endocrinology and Metabolism 1986 250 E13–E17.
Boirie Y, Short KR, Ahlman B, Charlton M, Nair KS. Tissue-specific regulation of mitochondrial and cytoplasmic protein synthesis rates by insulin. Diabetes 2001 50 2652. (doi:10.2337/diabetes.50.12.2652).
Chow LS, Albright RC, Bigelow ML, Toffolo G, Cobelli C, Nair KS. Mechanism of insulin’s anabolic effect on muscle: measurements of muscle protein synthesis and breakdown using aminoacyl-tRNA and other surrogate measures. American Journal of Physiology. Endocrinology and Metabolism 2006 291 E729–E736. (doi:10.1152/ajpendo.00003.2006).
Gelfand RA, Barrett EJ. Effect of physiologic hyperinsulinemia on skeletal muscle protein synthesis and breakdown in man. Journal of Clinical Investigation 1987 80 1. (doi:10.1172/JCI113033).
Fryburg DA, Barrett EJ, Louard RJ, Gelfand RA. Effect of starvation on human muscle protein metabolism and its response to insulin. American Journal of Physiology. Endocrinology and Metabolism 1990 259 E477–E482.
Heslin MJ, Newman E, Wolf RF, Pisters PW, Brennan MF. Effect of hyperinsulinemia on whole body and skeletal muscle leucine carbon kinetics in humans. American Journal of Physiology 1992 262 E911–E918.
Newman E, Heslin MJ, Wolf RF, Pisters PW, Brennan MF. The effect of insulin on glucose and protein metabolism in the forearm of cancer patients. Surgical Oncology 1992 1 257–267. (doi:10.1016/0960-7404(92)90086-Z).
Short KR, Bigelow ML, Nair KS. Short-term prednisone use antagonizes insulin’s anabolic effect on muscle protein and glucose metabolism in young healthy people. American Journal of Physiology. Endocrinology and Metabolism 2009 297 E1260–E1268. (doi:10.1152/ajpendo.00345.2009).
Barazzoni R, Short KR, Asmann Y, Coenen-Schimke JM, Robinson MM, Nair KS. Insulin fails to enhance mTOR phosphorylation, mitochondrial protein synthesis, and ATP production in human skeletal muscle without amino acid replacement. American Journal of Physiology. Endocrinology and Metabolism 2012 303 E1117–E1125. (doi:10.1152/ajpendo.00067.2012).
Le mistificazioni e i luoghi comuni sull’Insulina nel mondo del Fitness e BodyBuilding:
I luoghi comuni completamente o in buona parte errati sull’Insulina abbondano nel settore del Fitness e del BodyBuilding. Uno di questi luoghi comuni riguarda un’elevata assunzione di carboidrati e la sua presunta correlazione con livelli cronicamente (e sottolineo cronicamente) elevati di Insulina, i quali porterebbero il soggetto a ingrassare dal momento che la lipogenesi supererà costantemente la lipolisi (ricordate che l’aumento di grasso può avvenire solo se il tasso di lipogenesi supera quello di lipolisi). Tuttavia, nelle persone sane l’Insulina aumenta solo in risposta ai pasti. Ciò significa che la lipogenesi supera la lipolisi solo nelle ore successive al pasto (il cosiddetto periodo postprandiale). Durante i periodi di digiuno (come i periodi prolungati tra un pasto e l’altro o quando si dorme), la lipolisi supera la lipogenesi (cioè si ossidano i grassi). Nell’arco delle 24 ore, tutto si equilibra (a patto che non si assumano più calorie di quante se ne consumino), il che significa che non si ingrassa. Ecco un grafico che mostra come funziona:
Dopo i pasti, il grasso viene depositato con l’aiuto dell’Insulina. Tuttavia, tra i pasti e durante il sonno, il grasso viene perso. Il bilancio dei grassi sarà pari a zero nell’arco delle 24 ore se l’apporto energetico corrisponde al dispendio energetico.
Questo è ovviamente solo un grafico approssimativo, ma l’area rossa rappresenta la lipogenesi che si verifica in risposta a un pasto. L’area blu rappresenta la lipolisi che si verifica in risposta al digiuno tra i pasti e durante il sonno. Nell’arco delle 24 ore, questi valori si bilanciano, a patto che non si assumano più calorie di quante se ne consumino. Questo è vero anche se l’assunzione di carboidrati è elevata. Inoltre, se l’apporto energetico è inferiore al dispendio energetico, una dieta ad alto contenuto di carboidrati comporta una perdita di peso come qualsiasi altra dieta.
Insulina, HSL e ASP
Un altro luogo comune sull’Insulina riguarda la sua necessità per l’accumulo di grasso. Peccato che non sia così. Il corpo ha modi per immagazzinare e trattenere i grassi anche quando l’Insulina è bassa. Ad esempio, nelle cellule adipose è presente un enzima chiamato lipasi ormonosensibile (HSL). L’HSL aiuta a scomporre i Trigliceridi di deposito in acidi grassi liberi. L’Insulina sopprime l’attività dell’HSL e quindi la scomposizione dei Trigliceridi. Questo ha portato le persone a puntare il dito contro i carboidrati come causa dell’aumento di grasso.
Tuttavia, i grassi sopprimono l’HSL anche quando i livelli di Insulina sono bassi. Ciò significa che non è possibile perdere grasso anche quando l’apporto di carboidrati è basso, se si esagera con le calorie. Se non si mangiassero carboidrati ma 5.000 calorie di grassi, non si riuscirebbe comunque a perdere grasso anche se l’Insulina non sarebbe elevata. Questo perché l’elevato apporto di grassi sopprime l’HSL. Questo significa anche che, se si segue una dieta a basso contenuto di carboidrati, per perdere peso è necessario mangiare meno calorie di quelle che si consumano.
Struttura della ASP (Proteina Stimolante l’Acilazione)
Ora, qualcuno potrebbe dire: “Provate a consumare 5.000 calorie di olio d’oliva e vedrete che risultati otterrete”. Beh, 5000 calorie di olio d’oliva non sono molto appetibili, quindi è ovvio che probabilmente non si riuscirà nell’impresa, soprattutto con regolarità. La stessa cosa accadrebbe consumando 5.000 calorie di puro zucchero da tavola.
Molti pensano che l’Insulina sia legata solo e soltanto al consumo di Carboidrati (eh Sears, quanti danni hai fatto…). Questo è probabilmente il più grande luogo comune in circolazione. I carboidrati hanno una cattiva reputazione a causa del loro effetto sull’Insulina, ma anche le proteine stimolano la secrezione di Insulina. Anzi, possono essere uno stimolo per l’Insulina altrettanto maggiore di quello dei carboidrati. Uno studio ha confrontato gli effetti di due pasti diversi sull’Insulina. Un pasto conteneva 21g di proteine e 125g di carboidrati. L’altro pasto conteneva 75g di proteine e 75g di carboidrati. Entrambi i pasti contenevano 675 calorie. Ecco un grafico della risposta Insulinica:
Confronto della risposta Insulinica tra un pasto a basso contenuto di proteine e alto contenuto di carboidrati e un pasto ad alto contenuto di proteine e basso contenuto di carboidrati.
Ecco un grafico della risposta glicemica:
Confronto della risposta glicemica a un pasto a basso contenuto di proteine e ad alto contenuto di carboidrati e a un pasto ad alto contenuto di proteine e basso contenuto di carboidrati.
Si può notare che, nonostante la risposta glicemica fosse molto più alta nel pasto con più carboidrati, la risposta insulinica non era più alta. In realtà, la risposta insulinica era leggermente più alta dopo il pasto ad alto contenuto proteico, anche se non era statisticamente significativa.
Alcuni potrebbero obiettare che la condizione di “basso contenuto di carboidrati” non era veramente tale perché conteneva 75g di carboidrati. Ma non è questo il punto. Il punto è che la condizione ad alto contenuto di carboidrati aveva quasi il DOPPIO dei carboidrati, con una risposta di glucosio più elevata, ma la secrezione di Insulina era leggermente inferiore. Le proteine erano altrettanto potenti nello stimolare l’Insulina quanto i carboidrati.
Risposta insulinica a pasti ad alto contenuto di proteine e di carboidrati.
Come potete notare nel grafico si osserva la tendenza a un picco di Insulina più rapido con la variabile ad alto contenuto proteico, con una risposta media di 45uU/mL a 20 minuti dal pasto, rispetto a circa 30uU/mL nella variabile ad alto contenuto di carboidrati.
Tornando al discorso accennato in precedenza, questa tendenza a una risposta insulinica più elevata era associata a una tendenza a una maggiore soppressione dell’appetito. I soggetti tendevano ad avere meno fame e più sazietà dopo il pasto ad alto contenuto proteico:
Confronto tra pasti a basso contenuto di proteine e ad alto contenuto di carboidrati e pasti ad alto contenuto di proteine e a basso contenuto di carboidrati e loro effetti su fame e sazietà.
Ecco i risultati di un altro studio che ha confrontato gli effetti di 4 diversi tipi di proteine sulla risposta insulinica a un pasto. Questo studio è interessante perché sono stati preparati frullati con le diverse proteine (si, hanno usato anche frullati di tonno). I frullati contenevano solo 11g di carboidrati e 51g di proteine. Ecco la risposta insulinica ai diversi frullati:
Risposta insulinica a 4 differenti fonti proteiche.
Si può notare che tutte queste proteine hanno prodotto una risposta insulinica, nonostante il fatto che i carboidrati nel frullato fossero bassi. La risposta insulinica è stata diversa anche tra le proteine, con il siero di latte che ha prodotto la risposta insulinica più elevata.
Ora, qualcuno potrebbe pensare che la risposta sia dovuta alla gluconeogenesi (un processo attraverso il quale il fegato converte le proteine in glucosio). L’idea è che le proteine vengano convertite in glucosio e che quindi aumentino i livelli di Insulina. Come ho già detto, si sostiene che questo comporta una risposta insulinica molto più lenta e prolungata, poiché il fegato impiega tempo a trasformare le proteine in glucosio. Tuttavia, non è così, perché la risposta insulinica è stata rapida, con un picco di 30 minuti e un rapido calo a 60 minuti:
Risposta insulinica alle diverse fonti proteiche.
Questa rapida risposta insulinica non era dovuta a variazioni della glicemia. Infatti, le proteine del siero del latte, che hanno provocato la maggiore risposta insulinica, hanno causato un calo della glicemia:
Risposta glicemica alle diverse fonti proteiche.
La risposta insulinica è stata associata alla soppressione dell’appetito. Infatti, le proteine del siero del latte, che avevano la risposta insulinica più alta, hanno causato la maggiore soppressione dell’appetito. Ecco un grafico che mostra l’apporto calorico dei soggetti quando hanno pranzato 4 ore dopo aver bevuto il frullato:
Apporto calorico di un pranzo consumato 4 ore dopo il consumo di varie proteine.
I soggetti hanno mangiato quasi 150 calorie in meno a pranzo quando hanno assunto proteine del siero di latte, che hanno anche provocato la maggiore risposta insulinica. In effetti, è stata riscontrata una fortissima correlazione inversa tra l’Insulina e l’assunzione di cibo (una correlazione di -0,93).
Ecco i dati di un altro studio che ha esaminato la risposta insulinica a un pasto che conteneva 485 calorie, 102g di proteine, 18g di carboidrati e quasi nessun grasso:
Risposta insulinica a un pasto ad alto contenuto proteico e a basso contenuto di carboidrati in persone magre e obese.
Si può notare che la risposta insulinica era esagerata nei soggetti obesi, probabilmente a causa della resistenza all’insulina. Ecco un grafico della risposta della glicemia ematica. Si può notare che non c’era alcuna relazione tra la risposta del glucosio e l’Insulina, come nello studio discusso in precedenza.
Risposta della glicemia a un pasto ad alto contenuto proteico e a basso contenuto di carboidrati in persone magre e obese.
Il fatto è che le proteine sono un potente stimolatore della secrezione di Insulina, e questa secrezione di Insulina non è correlata a variazioni della glicemia o della gluconeogenesi da parte delle proteine. In effetti, uno studio ha rilevato che la carne di manzo stimola la secrezione di Insulina tanto quanto il riso integrale. La risposta glicemica di 38 alimenti diversi poteva spiegare solo il 23% della variabilità della secrezione insulinica in questo studio. Quindi, dietro la secrezione di Insulina c’è molto di più dei soli carboidrati.
Va ricordato perché il Glucagone viene rilasciato in risposta alle proteine. Poiché le proteine stimolano la secrezione di Insulina, se non si consumano carboidrati con le proteine, esse causano un rapido calo della glicemia. Il Glucagone impedisce questo rapido calo di zuccheri nel sangue stimolando il fegato a produrre glucosio.
Adesso sappiamo che le proteine alimentari possono causare picchi di Insulina proprio come i carboidrati alimentari, e questi picchi non sono legati alla gluconeogenesi delle proteine (cioè alla loro conversione in zucchero). Sappiamo anche che questi picchi sono in parte responsabili della soppressione dell’appetito causata dalle proteine alimentari (grazie agli effetti dell’Insulina sul cervello che inibiscono l’appetito).
Picchi insulinici e aumento del peso
Vorrei approfondire un altro luogo comune riguardante l’Insulina e che interessa i rapidi picchi dell’ormone. Essi sono importanti nella regolazione della glicemia. E’ necessario quindi discutere le fasi della secrezione di Insulina. La secrezione di Insulina da parte del pancreas avviene in due fasi. La prima fase avviene molto rapidamente: il pancreas percepisce l’aumento del glucosio e l’Insulina viene rilasciata entro 1-2 minuti dall’aumento della glicemia. Questa risposta in fase rapida è il risultato del rilascio da parte del pancreas dell’Insulina immagazzinata. In genere si conclude entro 10 minuti. È stato riscontrato che questa risposta di fase rapida è compromessa nelle persone con alterata tolleranza al glucosio (persone che hanno risposte glicemiche ai pasti più elevate del normale e livelli di zucchero nel sangue a digiuno più elevati, ma che non sono diabetiche). Questa risposta in fase rapida è completamente assente nelle persone affette da diabete di tipo II.
Esiste una seconda fase che continua finché il glucosio è elevato. Questo rilascio di Insulina avviene attraverso la liberazione dell’Insulina immagazzinata e la creazione di nuova Insulina (l’Insulina viene creata da un precursore chiamato proinsulina, come abbiamo visto nella prima parte). Quando si infonde glucosio nel sangue di persone sane e di diabetici di tipo II, si ottengono risposte insuliniche di questo tipo:
Risposta insulinica alla somministrazione di glucosio per via endovenosa in persone sane e in diabetici di tipo II.
Si può notare che i diabetici mancano completamente della risposta di fase rapida che è presente negli individui sani.
Esiste un farmaco chiamato Exenatide (Byetta), che si è rivelato in grado di ripristinare questa risposta insulinica di fase rapida nei diabetici:
Risposte insuliniche di diabetici di tipo II e di individui sani, a cui è stato somministrato glucosio per via endovenosa. I cerchi rappresentano la risposta insulinica dei diabetici di tipo II quando viene loro somministrato un placebo. I quadrati rappresentano la risposta insulinica dei diabetici alla somministrazione di Exenatide. Si può notare che l’Exenatide ripristina la risposta insulinica in fase rapida. I cerchi neri rappresentano la risposta insulinica dei soggetti sani.
Il ripristino della risposta insulinica in fase rapida migliora la regolazione della glicemia nei diabetici:
Risposta glicemica a un pasto in diabetici di tipo II. I cerchi rappresentano i soggetti in trattamento con placebo. I triangoli e i cerchi scuri rappresentano i soggetti che assumono Exenatide. Si può notare che la glicemia è rimasta costante nei soggetti che assumevano Exenatide, mentre è aumentata gradualmente nei soggetti che assumevano il placebo.
Nel grafico sopra riportato si può notare che la glicemia è rimasta costante in risposta a un pasto nei soggetti che assumevano l’Exenatide, mentre è aumentata nel tempo nei soggetti che assumevano il placebo.
A molti piace attribuire la colpa dell’obesità e dell’aumento di peso all’Insulina, ma l’Exenatide, che ripristina i picchi di Insulina nei diabetici di tipo II, fa perdere peso:
Effetto del Exenatide (Byetta) sul peso corporeo.
Parte di questa perdita di peso è dovuta a un miglioramento del senso di sazietà. L’Exenatide è un farmaco che imita gli effetti di un ormone chiamato peptide glucagone-simile-1 (GLP-1). Il GLP-1 è un ormone che stimola l’Insulina a livello intestinale (noto come Incretina). Il GLP-1 potenzia la secrezione di Insulina, aumenta la sintesi di Insulina, aumenta l’espressione genica dell’Insulina e inibisce la secrezione di Glucagone (l’ormone antagonista dell’Insulina). Eppure l’Exenatide, che imita il GLP-1 e contribuisce a stimolare la secrezione di Insulina, fa perdere peso.
Il fatto è che i rapidi picchi di Insulina di per sé non sono un male. Le proteine causano rapidi picchi di Insulina, ma le proteine riducono l’appetito e aiutano a perdere peso. Il GLP-1 e i farmaci come l’Exenatide contribuiscono ai picchi di Insulina, ma riducono l’appetito e fanno perdere peso. Il problema è che le persone confondono i picchi di Insulina con i picchi di glucosio nel sangue. È ormai assodato che un rapido aumento e una rapida diminuzione del glucosio nel sangue possono contribuire alla fame. Poiché i rapidi aumenti della glicemia causano anche rapidi aumenti dell’Insulina, le persone finiscono per incolpare l’Insulina (e gli effetti dei carboidrati ad alto indice glicemico sull’Insulina) del problema.
Differenza del possibile impatto tra somministrazione esogena e secrezione endogena di Insulina sul peso corporeo
Un altro luogo comune sull’Insulina correla l’aumento di peso dei soggetti diabetici trattati con tale ormone agli aumenti di peso dei soggetti sani. Evidentemente queste persone non conoscono l’Amilina.
Molecola di Amilina
L’Amilina è un ormone che viene secreto dal pancreas contemporaneamente all’Insulina. L’Amilina riduce l’appetito e stimola la lipolisi (la scomposizione dei Trigliceridi in acidi grassi liberi).
I diabetici di tipo I non producono Amilina e nei diabetici di tipo II la secrezione di Amilina è compromessa. È stato riscontrato che la Pramlintide, un farmaco che imita gli effetti dell’Amilina, produce una perdita di peso nei diabetici.
Queste informazioni dimostrano che gli effetti dell’iniezione di Insulina in un diabetico non possono essere paragonati agli effetti delle variazioni fisiologiche dell’Insulina in un non diabetico, eppure molte persone fanno erroneamente questo confronto come se fossero simili.
Prodotti lattiero-caseari e secrezione insulinica
Una delle premesse che alcune persone fanno è che i carboidrati stimolino l’accumulo di grasso stimolando la secrezione di Insulina. Ma abbiamo già visto come questa premessa sia errata. In particolare, abbiamo visto come anche le proteine stimolino la secrezione di Insulina (a volte tanto quanto i carboidrati), ma non favoriscano l’aumento di peso o di grasso. Vi ho anche mostrato come il farmaco Exenatide ripristini la secrezione di Insulina in fase rapida nei diabetici, pur favorendo la perdita di peso.
Nonostante le risposte glicemiche basse, i latticini creano risposte insuliniche molto elevate. Per esempio, in uno studio, i latticini hanno creato risposte insuliniche simili o superiori a quelle del pane bianco, nonostante la risposta glicemica per alcuni dei latticini fosse del 60% inferiore a quella del pane bianco. In questo studio, i ricercatori hanno confrontato le risposte glicemiche e insulinemiche tra pane bianco, una miscela a basso contenuto di glutine/lattosio, una miscela ad alto contenuto di glutine/lattosio, merluzzo con aggiunta di lattosio, latte, proteine del siero del latte con aggiunta di lattosio e formaggio con aggiunta di lattosio. Tutte le variabili contenevano 25g di carboidrati e 18,2g di proteine, tranne il pane bianco e le miscele a basso contenuto di glutine/lattosio, che contenevano 25g di carboidrati e 2,8g di proteine. Pertanto, il lattosio era il carboidrato in tutte le condizioni, ad eccezione del pane bianco.
Osservando l’area dell’Insulina sotto la curva (AUC) per le varie condizioni, si può notare che i prodotti lattiero-caseari hanno effettivamente creato risposte insuliniche maggiori rispetto al pane bianco, pur avendo quantità simili di carboidrati:
Risposta insulinica a prodotti lattiero-caseari e pane bianco.
È evidente che non è il lattosio il responsabile della maggiore risposta insulinica, perché le miscele glutine/lattosio e merluzzo/lattosio hanno dato risposte insuliniche simili o inferiori a quelle del pane bianco.
Anche la risposta glicemica non è responsabile della maggiore risposta insulinica. Infatti, la risposta glicemica era più bassa in tutte le condizioni rispetto al pane bianco, con il latte che creava la risposta glicemica più bassa ma la terza risposta insulinica più alta:
Risposta della glicemia ai latticini rispetto al pane bianco.
L’indice insulinogenico, che mette in relazione la quantità di secrezione di Insulina con la risposta del glucosio nel sangue, era significativamente più alto nei prodotti lattiero-caseari, indicando che i prodotti lattiero-caseari stimolavano una secrezione di Insulina molto maggiore di quella che ci si aspetterebbe in base alla risposta del glucosio nel sangue:
indice insulinogenico dei prodotti lattiero-caseari rispetto al pane bianco.
Questo non è l’unico studio che mostra gli effetti insulinemici dei latticini. In precedenza si è visto come le proteine del siero di latte, una proteina casearia, creino la risposta insulinica più elevata rispetto alle proteine non casearie. In uno studio condotto su diabetici di tipo II, l’inclusione di proteine del siero di latte in un pasto ha aumentato la risposta insulinica del 31-57%, mentre la risposta glicemica si è ridotta fino al 21%. In un altro studio, l’aggiunta di 400ml di latte a un pasto a base di pane ha aumentato la risposta insulinica del 65%, nonostante non vi sia stata alcuna variazione nella risposta glicemica. Nello stesso studio, l’aggiunta di 200 o 400ml di latte a un pasto a base di spaghetti ha aumentato la risposta insulinica del 300%; anche in questo caso, la risposta glicemica non ha subito variazioni. In effetti, bere latte con il pasto di spaghetti ha creato una risposta insulinica simile a quella del pane bianco.
Ecco i risultati di un altro studio che mostra gli indici glicemici e insulinemici del latte rispetto al pane bianco:
Un altro motivo per cui i latticini stimolano la secrezione di Insulina è il loro effetto su un ormone chiamato polipeptide insulinotropico glucosio-dipendente (GIP). Come il GLP-1, di cui ho scritto precedentemente, il GIP è un’incretina. Ciò significa che è un ormone prodotto dall’intestino che stimola la secrezione di Insulina. I latticini stimolano una maggiore produzione di GIP. Nello studio di cui ho parlato in precedenza, che ha messo a confronto il siero di latte, il latte e il formaggio con il pane bianco, il siero di latte e il formaggio hanno prodotto risposte alla GIP superiori del 21-67% rispetto al pane bianco:
Risposta del polipeptide insulinotropico glucosio-dipendente (GIP) agli alimenti a base di latte rispetto al pane bianco.
I dati sopra riportati illustrano uno dei problemi dell’ipotesi carboidrati/insulina… essa presuppone che i carboidrati siano lo stimolo principale della secrezione insulinica. Tuttavia, è chiaro che anche gli aminoacidi e le incretine svolgono un ruolo significativo nella secrezione di Insulina. E come ho sottolineato, la risposta glicemica di un alimento spiega solo il 23% della variazione della risposta insulinica. Pertanto, la secrezione di Insulina è molto più importante della risposta glicemica dovuta al consumo di carboidrati.
Sebbene si tratti di studi osservazionali, i risultati di studi controllati su animali ed esseri umani sono simili. In effetti, gli studi sugli animali mostrano un minore aumento di peso quando vengono nutriti con prodotti caseari. Nei topi, l’integrazione di yogurt determina un minore aumento di peso e di grasso rispetto ai controlli che seguono una dieta isocalorica. In un altro studio, i topi transgenici hanno perso peso con una dieta ipocalorica. I topi sono stati poi lasciati mangiare ad libitum (cioè quanto volevano). I topi alimentati con prodotti caseari hanno riacquistato meno grasso e peso durante la rialimentazione. In un terzo studio, l’assunzione di prodotti lattiero-caseari, ma non di un integratore di calcio, ha ridotto l’aumento di peso e il grasso corporeo nei topi alimentati con una dieta ad alto contenuto di grassi. In un quarto studio, le proteine dei latticini hanno attenuato l’aumento di grasso nei roditori alimentati con una dieta ad alto contenuto di grassi e zuccheri. In un quinto studio, una dieta a base di latticini ha attenuato l’aumento di peso settimanale nei ratti Sprague-Dawley.
Naturalmente, si tratta di studi sugli animali. E per gli esseri umani? In uno studio, i latticini a basso contenuto di grassi non hanno favorito l’aumento di peso, mentre quelli ad alto contenuto di grassi sì. È possibile che l’aumento di peso in questo studio sia stato causato semplicemente dall’eccesso di calorie e non dall’Insulina? In un altro studio, l’aumento dell’assunzione di latticini non ha influito sulla composizione corporea. In un terzo studio, l’aumento dell’assunzione di latticini non ha compromesso la perdita di peso. In uno studio di un anno, l’aumento dell’assunzione di latticini non ha influito sulle variazioni della massa grassa. In un follow-up di 6 mesi, un’elevata assunzione di latticini ha predetto livelli inferiori di massa grassa. In uno studio di 9 mesi, l’aumento dell’assunzione di latticini non ha influito sul mantenimento del peso, ma il gruppo ad alto contenuto di latticini ha evidenziato una maggiore ossidazione dei grassi.
Ora dovrebbe essere più che chiaro il fatto che le prove sono schiaccianti sulla questione che i prodotti lattiero-caseari non favoriscono l’aumento di peso e anzi lo inibiscono per via dell’effetto saziante. Questo nonostante il fatto che i latticini producano una risposta insulinica molto ampia, pari o superiore a quella di molti alimenti ad alto contenuto di carboidrati. Pertanto risulta chiaro che l’ipotesi carboidrati/insulina è errata.
Per concludere la serie di luoghi comuni sull’Insulina vi citerò quello che desterà maggiore incredulità…
Molti pensano che le cellule abbiano bisogno di Insulina per utilizzare il glucosio nel circolo ematico. Una delle prove a sostegno di questa tesi è rappresentata dal soggetto diabetico di tipo I. Quando un diabetico di tipo I non ha Insulina, la glicemia sale alle stelle. Questo perché, a quanto pare, il glucosio non riesce a entrare nelle cellule.
Tuttavia, lo scenario sopra descritto non è quello che si verifica in un diabetico di tipo I a cui è stata tolta l’Insulina. Il glucosio può entrare nelle cellule senza problemi. In realtà sta succedendo qualcos’altro. Un articolo pubblicato sul Journal of Anasthesia descrive in modo esauriente come l’Insulina sia stata fraintesa nel suo ruolo di regolazione della glicemia ematica.
Nel 1916, Sir Edward Schafer, professore di fisiologia del quale ho già parlato nella prima parte, pubblicò un libro intitolato “The Endocrine Organs”. In questo libro ipotizzò l’esistenza di quella che oggi chiamiamo Insulina:
I risultati dell’asportazione del pancreas e dell’innesto del pancreas si spiegano meglio ipotizzando che il tessuto delle isole produca un Autacoide che passa nel flusso sanguigno e agisce sul metabolismo dei carboidrati e sull’immagazzinamento dei carboidrati in modo tale da evitare un indebito accumulo di glucosio nel sangue. In via provvisoria sarà opportuno riferirsi a questa ipotetica sostanza come Insulina.
L’insulina sarebbe stata scoperta 5 anni dopo. Schafer ipotizzò anche che l’Insulina fosse creata da un precursore inattivo:
Va tuttavia precisato che non è ancora stato determinato se la sostanza attiva sia prodotta come tale nel pancreas o se esista come pro-insulina che viene convertita altrove in un autacoide attivo.
La pro-insulina fu scoperta quasi 50 anni dopo. Schafer era davvero un uomo in anticipo sui tempi.
Schafer evitava di usare il termine “ormone” per descrivere l’Insulina. Utilizzò invece i termini “autacoide” e “chalone”. Un autacoide è una sostanza con azione eccitatoria, cioè stimola l’azione del corpo. Un autacoide può essere considerato simile al pedale dell’acceleratore dell’auto; si preme il pedale e si stimola l’auto ad andare più veloce. Il chalone è una sostanza ad azione inibitoria, che rallenta le cose nel corpo. Il chalone può essere considerato simile al freno dell’auto. Schafer ipotizzò correttamente che l’Insulina agisse sia come autacoide che come chalone nell’organismo. Egli riteneva inoltre che l’Insulina agisse come chalone molto più che come autacoide nell’organismo. In altre parole, riteneva che le funzioni inibitorie dell’Insulina fossero molto più importanti di quelle eccitatorie o stimolatorie. Molti anni dopo si sarebbe dimostrato corretto.
Tuttavia, prima che a Schafer venisse data ragione, si verificò l'”età nera dell’endocrinologia”. Si tratta del periodo di tempo compreso tra il 1950 e il 1980, in cui gli scienziati estrapolavano conclusione che andavano ben oltre le loro effettive scoperte. Prendevano i dati di studi su animali o in vitro (ricerche condotte in provetta o in coltura) e poi ipotizzavano che la stessa cosa avvenisse nell’uomo in vivo (all’interno del corpo). Gli scritti come “Good Calories, Bad Calories” di Gary Taubes si basano pesantemente sulla ricerca di questo periodo, nonostante il fatto che gran parte di ciò che si pensava all’epoca sia stato ribaltato da ricerche migliori, o almeno modificato in modo significativo.
L’età nera dell’endocrinologia è quella che ha portato alla convinzione, oggi errata, che l’Insulina sia necessaria alle cellule per assorbire il glucosio. Gli esperimenti condotti negli anni Cinquanta dimostrarono che l’Insulina era in grado di stimolare l’assorbimento del glucosio da parte di campioni di muscolo e di grasso di ratto. Questi dati sono stati trasposti all’uomo e si è ipotizzato erroneamente che la mancanza di Insulina impedisca al glucosio di entrare nelle cellule e che quindi la glicemia salga a livelli pericolosi. Questo pensiero errato è stato insegnato nei libri di testo e nei corsi universitari di tutto il mondo per molti anni, dando vita a un vero e proprio dogma. Purtroppo è molto difficile superare una convinzione fortemente radicata, anche se negli anni ’70 è stato dimostrato che questo concetto di Insulina è sbagliato, continua a essere insegnato ancora oggi.
L’ipotesi errata secondo la quale la sospensione dell’Insulina provochi un’elevata glicemia perché “il glucosio non riesce a entrare nelle cellule” si basava sul presupposto che l’Insulina sia necessaria per l’assorbimento del glucosio da parte delle cellule, piuttosto che l’Insulina si limiti a migliorare l’assorbimento del glucosio. Quello che gli scienziati degli anni ’50 non hanno notato è che i tessuti possono assumere notevoli quantità di glucosio anche in assenza di Insulina.
Il glucosio entra nelle cellule attraverso una famiglia di trasportatori. Un trasportatore primario nelle cellule muscolari e adipose lo conosciamo più o meno tutti, si tratta del GLUT-4. L’Insulina stimola il GLUT-4 a spostarsi dall’interno della cellula alla superficie cellulare, dove il glucosio può legarsi al trasportatore GLUT-4 ed entrare nella cellula. Tuttavia, sulla superficie cellulare sono presenti numerosi trasportatori di glucosio, anche in assenza di Insulina. In effetti, ci sono abbastanza trasportatori sulla superficie cellulare per consentire alla cellula di ottenere abbastanza glucosio per sostenere il suo fabbisogno energetico. Pertanto, il trasporto di glucosio nelle cellule non è mai veramente dipendente dall’Insulina. L’Insulina favorisce l’assorbimento del glucosio nelle cellule, ma non è necessaria. Infatti, quando si elimina il Recettore dell’Insulina nei topi in modo che l’Insulina non possa stimolare l’assorbimento del glucosio nelle cellule muscolari o adipose (pur mantenendo intatto il Recettore dell’Insulina in altre cellule come il cervello e il fegato), gli animali non diventano diabetici e presentavano valori della glicemia ematica normali.
Gli studi sui traccianti metabolici ci hanno permesso di capire come funziona l’Insulina nell’uomo in vivo. Quando si toglie l’Insulina a un diabetico di tipo I, il glucosio nel sangue sale bruscamente. Tuttavia, non è perché il glucosio non riesce a entrare nelle cellule. In realtà, l’assorbimento del glucosio nelle cellule aumenta. Questo perché la concentrazione di glucosio nel sangue è talmente superiore a quella cellulare che il glucosio deve spostarsi all’interno delle cellule (ricordate che sulla superficie delle cellule ci sono già abbastanza trasportatori di glucosio anche in assenza di Insulina). Allora perché il glucosio nel sangue sale così tanto? Ricordiamo che la quantità di glucosio nel sangue è in funzione sia della quantità di glucosio che entra nel sangue (velocità di comparsa), sia della quantità di glucosio che esce dal sangue (velocità di scomparsa). In un diabetico a digiuno e senza Insulina, tutto il glucosio proviene dal fegato. Ricordiamo che il fegato contribuisce a mantenere i livelli di zucchero nel sangue a digiuno rilasciando glucosio; questo glucosio proviene sia dalla gluconeogenesi (la formazione di glucosio da fonti non glucidiche, come le proteine) sia dalla glicogenolisi (la degradazione del glicogeno immagazzinato nel fegato). L’Insulina agisce come un freno (un chalone, come lo ha definito il dottor Schafer) su questi processi. Pertanto, in assenza di Insulina, si verificano fenomeni di gluconeogenesi e glicogenolisi incontrollati. La glicemia elevata in un diabetico non controllato è quindi causata da una sovrapproduzione di glucosio da parte del fegato, non perché il glucosio non riesca a entrare nelle cellule.
Infatti, poiché l’Insulina non è presente, molti processi si attuano a ritmi elevati, completamente sregolati. L’Insulina normalmente inibisce la produzione di chetoni da parte del fegato; senza l’Insulina che rallenta la produzione di chetoni, questi ultimi vengono prodotti a ritmi elevati, dando luogo alla chetoacidosi diabetica. Ecco perché l’iperglicemia e la chetoacidosi si verificano contemporaneamente. Senza Insulina, si ha anche un’accelerazione della proteolisi (la scomposizione delle proteine) e della lipolisi (la scomposizione dei grassi). Gli aminoacidi elevati nel sangue forniscono ulteriore substrato al fegato per continuare a produrre grandi quantità di glucosio. Gli acidi grassi elevati forniscono al fegato il substrato per continuare a produrre grandi quantità di chetoni.
L’Insulina è quindi come un vigile urbano o un semaforo a un incrocio. Aiuta a rallentare e a controllare il traffico. Senza un semaforo o un vigile urbano, le auto attraversano l’incrocio senza controllo e si verificano incidenti stradali. Allo stesso modo, senza Insulina nell’organismo, la gluconeogenesi, la glicolisi, la proteolisi, la chetogenesi e la lipolisi procedono a ritmi elevati senza che nulla possa fermarle. Il risultato finale è l’iperglicemia, la chetoacidosi e infine la morte.
Quando si inietta l’Insulina in un diabetico non controllato, si frenano tutti i processi menzionati in precedenza. Si inibisce la produzione di glucosio da parte del fegato e la glicemia si abbassa. Poiché non c’è più iperglicemia, l’assorbimento di glucosio nelle cellule diminuisce. La lipolisi viene inibita, quindi la concentrazione di acidi grassi liberi scende quasi a zero. Poiché non ci sono più acidi grassi liberi per la produzione di chetoni, la produzione di chetoni rallenta. Anche la proteolisi viene inibita.
Gli studi sui traccianti metabolici hanno dimostrato ciò che Schafer aveva ipotizzato più di un secolo fa… che il ruolo principale dell’Insulina nell’organismo è inibitorio piuttosto che eccitatorio. Sebbene l’Insulina abbia certamente funzioni eccitatorie, non è principalmente un “ormone di stoccaggio” come molti sostengono. L’Insulina non è necessaria alle cellule per assorbire e immagazzinare il glucosio. Certamente ne favorisce l’assorbimento, ma c’è una grande differenza tra il favorire l’assorbimento e l’essere necessaria per l’assorbimento.
Naturalmente, questa ricerca ci dice solo cosa succede quando l’Insulina è presente rispetto a quando non è presente. Che dire della situazione normale di una persona sana, che ingerisce un pasto e vede un aumento del glucosio nel sangue? Cosa succede per riportare il glucosio alla normalità? E cosa succede in una persona diabetica di tipo II in questa situazione?
Un classico studio sui traccianti metabolici ha seguito cosa succede al glucosio quando viene assunto per via orale. In questo studio, ai diabetici di tipo II e ai soggetti sani di controllo è stato somministrato 1g di glucosio per chilogrammo di peso corporeo (quasi mezzo grammo per libbra). Utilizzando dei traccianti metabolici, i ricercatori hanno determinato non solo dove andava il glucosio, ma anche cosa succedeva alla produzione di glucosio da parte del fegato. I ricercatori hanno anche misurato i livelli di Insulina nel sangue.
Come prevedibile, il glucosio orale ha provocato un aumento del glucosio nel sangue e un corrispondente aumento dell’Insulina. Nelle persone sane, la produzione di glucosio da parte del fegato è stata drasticamente soppressa dall’aumento dell’Insulina. Infatti, la produzione di glucosio è diminuita del 70-80% a 75-105 minuti dall’ingestione del glucosio. Dopo 3,5 ore, la produzione di glucosio era ancora soppressa del 50%. Pertanto, uno dei modi in cui l’Insulina aiuta a controllare la glicemia dopo un pasto è quello di comunicare al fegato di smettere di produrre glucosio. Questo ha senso: non si vuole che il fegato produca glucosio quando il glucosio entra nel flusso sanguigno dall’apparato digerente.
Anche la produzione epatica di glucosio è stata soppressa nei diabetici di tipo II. Tuttavia, questa soppressione è stata compromessa. Tuttavia, questa soppressione era compromessa nei diabetici di circa il 40%. Si tratta di un caso di resistenza all’Insulina nel fegato dei diabetici; il fegato non risponde all’Insulina come dovrebbe (ricordiamo che l’Insulina sopprime la produzione epatica di glucosio) e quindi produce troppo glucosio. Nel documento gli autori affermano che:
…si può concludere che la sovrapproduzione di glucosio è un importante fattore determinante dell’iperglicemia diabetica, sia nello stato postassorbitivo che in quello postprandiale.
La produzione di glucosio da parte del fegato ci racconta solo metà della storia. Se da un lato l’Insulina inibisce la produzione di glucosio da parte del fegato, dall’altro aumenta la capacità dei tessuti di prelevare il glucosio dal sangue (ricordate che la migliora). Le cellule assorbono il glucosio in due modi… attraverso l’azione di massa del glucosio (cioè il gradiente di concentrazione, in cui la concentrazione di glucosio nel sangue è talmente superiore a quella delle cellule che il glucosio si sposta all’interno delle cellule) e attraverso la stimolazione dell’Insulina. In questo studio, l’assorbimento del glucosio nelle cellule era compromesso nei diabetici. Poiché il movimento del glucosio nelle cellule attraverso l’azione di massa è simile tra i diabetici e i soggetti sani, l’assorbimento ridotto del glucosio nei diabetici era dovuto all’insulino-resistenza nelle cellule. L’assorbimento del glucosio da parte dei tessuti era compromesso di circa il 27% nei diabetici.
Da questa ricerca emerge chiaramente che la risposta glicemica elevata di un diabetico di tipo II è dovuta sia a un’alterata risposta del fegato all’Insulina (per cui la produzione di glucosio è superiore a quella che dovrebbe essere), sia a un’alterata risposta delle cellule ad assorbire il glucosio dal sangue. Tuttavia, se si considerano le percentuali, la risposta alterata del fegato è maggiore di quella delle cellule. Un’altra ricerca pubblicata nello stesso anno ha mostrato una pari resistenza all’Insulina nel fegato e in altri tessuti, sebbene questa ricerca sia stata condotta con soggetti a digiuno. In quello studio, così come in quello di cui abbiamo parlato, è stata riscontrata una correlazione molto forte tra l’iperglicemia a digiuno e la produzione epatica di glucosio; ciò indica che, a digiuno, è la sovrapproduzione di glucosio da parte del fegato il fattore più importante nel causare l’iperglicemia in un diabetico. Da questa ricerca si evince anche che l’assorbimento di glucosio a digiuno è in realtà aumentato, non diminuito, nei diabetici di tipo II (proprio come nei diabetici di tipo I non controllati). Pertanto, l’iperglicemia a digiuno dei diabetici di tipo II e dei diabetici di tipo I non controllati è dovuta a una sovrapproduzione di glucosio da parte del fegato, non perché “il glucosio non riesce a entrare nelle cellule”. In un diabetico di tipo II, in risposta a un pasto, l’assorbimento del glucosio nelle cellule è compromesso, ma la resistenza all’Insulina nel fegato svolge comunque un ruolo importante.
Ciò che risulta chiaro da tutte queste ricerche è che le principali funzioni dell’Insulina nell’organismo sono inibitorie, agendo da freno su molti processi corporei. Sebbene l’Insulina stimoli l’immagazzinamento del glucosio e di altri nutrienti, questa funzione non è altrettanto importante di quella inibitoria. Pertanto, l’Insulina dovrebbe essere considerata più un vigile urbano che un ormone di stoccaggio.
Siete sconvolti ma consapevoli, oppure continuerete a vagare nel relativismo?
La conclusione è che l’Insulina non è il male assoluto, anzi, e le leggende e luoghi comuni che le ruotano intorno nel Fitness e BodyBuilding sono in definitiva sbagliati!
Per riassumere i punti chiave volti a demistificare i luoghi comuni sull’Insulina:
L’Insulina sopprime l’appetito, non lo aumenta.
Una dieta ad alto contenuto di carboidrati non causa livelli di Insulina cronicamente elevati.
Le proteine sono insulinemiche e, in alcuni casi, possono essere insulinemiche quanto i carboidrati.
Contrariamente a quanto si crede, il Glucagone non “annulla” la soppressione della lipolisi da parte dell’Insulina quando si ingeriscono proteine.
Gli effetti insulinemici delle proteine sono dovuti a un effetto stimolante diretto sul pancreas e non alla conversione delle proteine in glucosio.
La combinazione di proteine e carboidrati può produrre una maggiore secrezione di Insulina rispetto all’uno o all’altro da solo, eppure le diete ad alto contenuto di proteine e moderato-alto contenuto di carboidrati sono molto efficaci per la perdita di peso.
È stato dimostrato che le diete ad alto contenuto di carboidrati producono una perdita di peso quando le persone sono in deficit energetico.
I latticini sono estremamente insulinemici, tanto quanto il pane bianco, eppure non favoriscono l’aumento di peso in assenza di un surplus energetico. Questo dato è supportato da un numero molto elevato di studi, tra cui studi su animali, studi osservazionali e studi controllati randomizzati.
L’Insulina non è necessaria per l’accumulo di grasso
I livelli di Insulina non sono predittivi di aumento o perdita di peso nella maggior parte degli studi prospettici.
L’Exenatide ripristina il rilascio di Insulina in fase rapida nei diabetici, ma provoca una perdita di peso
Gli effetti dell’iniezione di Insulina non possono essere paragonati al normale rilascio fisiologico di Insulina, poiché l’Amilina è co-secreta con l’Insulina dal pancreas
L’Insulina funziona principalmente come ormone inibitorio piuttosto che come ormone di accumulo, agendo come freno su molti importanti processi fisiologici
Un diabetico di tipo I senza Insulina diventa iperglicemico a causa della sovrapproduzione di glucosio da parte del fegato, non perché il glucosio non riesca a entrare nelle cellule.
L’Insulina favorisce l’assorbimento del glucosio nelle cellule, ma non è necessaria per questo.
L’Insulina regola la glicemia dopo un pasto sia impedendo al fegato di produrre glucosio, sia favorendo l’assorbimento del glucosio nelle cellule.
A digiuno, l’Insulina regola la glicemia controllando la produzione di glucosio da parte del fegato e non influenzando l’assorbimento del glucosio nelle cellule.
Non ci si può limitare a considerare gli effetti temporanei dell’Insulina sulla lipolisi e sull’accumulo di glucosio. Bisogna considerare ciò che accade nell’arco delle 24 ore; il grasso corporeo non aumenta se non c’è un surplus energetico complessivo.
Tutto chiaro ora? Ecco, bravi, adesso però smettete di fare gli ortoressici isulinofobici!
Continua…
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Gli studi utilizzati per realizzare questo articolo provengono dalla serie di articoli scritti e pubblicati da James Krieger sul suo sito weightology.net e intitolati “INSULIN: AN UNDESERVED BAD REPUTATION”.