E’ ormai nota l’azione anoressizzante dell’Insulina a livello ipotalamico, come lo è il fatto che gli alimenti con un più elevato indice isulinico apportino un maggior grado di sazietà sebbene ciò non sia una determinante universale. Prova di questo lo sono i casi di soggetti insulino resistenti i quali, per via di tale condizione, vedono meno l’azione dell’ormone peptidico sulla ghiandola in questione e sulla conseguente regolazione del senso di sazietà. In base a queste basiche informazioni verrebbe da pensare che alimenti come latte e derivati, i quali hanno un indice insulinico particolarmente alto, possano essere un vantaggio in una dieta ipocalorica negli individui con un buon grado di insulino sensibilità. Eppure, da un recente studio realizzato da ricercatori presso l’Università di Milano [1] è emerso che il consumo delle semplici Whey può avere un riscontro positivo sul senso di sazietà anche in soggetti obesi, mostrando vie d’azione decisamente più ampie del semplice stimolo insulinico.
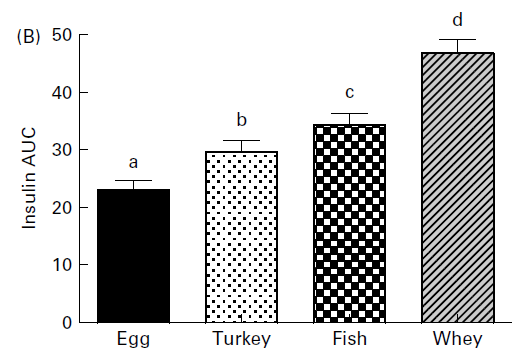
Indice insulinico di quattro diverse fonti proteiche.
Per lo svolgimento dello studio, i ricercatori hanno reclutato 8 giovani donne obese alle quali è stato fatto consumare uno shake in 2 diverse occasioni: nella prima, lo shake era composto da 43g di Maltodestrine, mentre nella seconda era composto da 45g di Whey. L’integratore di Whey utilizzato dai ricercatori (Enervit Gymline Muscle 100% whey protein isolate cacao) era prodotto dall’azienda di integratori italiana Enervit egualmente all’integratore di Maltodestrine (Enervit Maltodestrine Sport).
Quando veniva consumato lo shake contenente i 45g di Whey, i soggetti presi in esame riportavano di avere una diminuita sensazione di fame e una aumentata sensazione di sazietà rispetto alle risposte seguenti il consumo dello shake contenente 43g di Maltodestrine.
Gli esami dei marker ematici hanno evidenziato che il consumo di Whey causava un aumento più elevato nel livello dell’ormone della sazietà GLP-1 (glucagon-like peptide 1) rispetto alle Maltodestrine. Il consumo di Whey ha avuto lo stesso effetto sui livelli di PYY (Peptide YY), un altro ormone della sazietà prodotto nelle cellule L della mucosa intestinale dell’ileo e del colon, ma questo effetto è stato meno pronunciato.
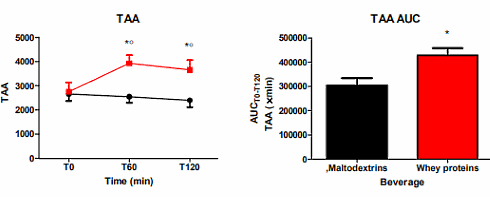
Lo shake contenente Whey ha ovviamente causato un aumento significativo della quantità di aminoacidi nel sangue (TAA nelle figure seguenti).
I ricercatori hanno quindi esaminato nel dettaglio la concentrazione amminoacidica nel sangue dei soggetti presi in esame. Ciò che è emerso è stata una correlazione degli amminoacidi Isoleucina, Leucina, Lisina, Metionina, Fenilalanina, Prolina, Tirosina e Valina con una diminuzione della sensazione di fame, un aumento della sensazione di sazietà e un aumento del livello di GLP-1. I ricercatori sospettano che questi amminoacidi stimolino i recettori nelle cellule L dell’intestino tenue.
I ricercatori affermano infatti che otto specifici amminoacidi (Isoleucina, Leucina, Lisina, Metionina, Fenilalanina, Prolina, Tirosina e Valina) sono implicati negli effetti di soppressione dell’appetito e di stimolazione del GLP-1 dati dall’ingestione di Whey, e che essi possono essere mediatori di tali effetti attraverso il loro legame con i recettori sensibili ai nutrienti espressi dalle cellule L nella parete gastrointestinale.
L’effetto saziante a lungo termine apportato dalle Whey e l’efficacia di un integratore con questo spettro amminoacidico (ovvero come un intervento nutraceutico) somministrato durante un programma alimentare ipocalorico volto alla riduzione del peso corporeo potrebbe meritare di essere ulteriormente studiato in futuro.
Certo, che le proteine avessero un effetto positivo sulla sensazione di fame e sazietà è cosa ben nota, ma questo studio ha pur sempre contribuito ad analizzarne gli effetti in modo approfondito utilizzando una fonte proteica (Whey) economica e che, visti i suoi effetti, può migliorare la compliace alla dieta in restrizione calorica apportando solamente tracce di altri macronutrienti prestandosi perfettamente come base degli spuntini durante la giornata. Teoricamente, per motivi legati all’indice insulinico, l’utilizzo di Whey in un contesto Chetogenico potrebbe non essere la scelta più adatta, anche se non vi sono dati certi riguardo all’impatto reale che un consumo di Whey possa avere sulla chetosi e l’adattamento consequenziale ad essa.
Un’immagine al microscopio elettronico del COVID-19
Il COVID-19 (abbreviazione di coronavirus disease 19), o malattia respiratoria acuta da SARS-CoV-2 o più semplicemente malattia da coronavirus 2019, è una malattia infettiva respiratoria causata dal virus denominato SARS-CoV-2 appartenente alla famiglia dei coronavirus (Virus a RNA). I primi casi sono stati riscontrati in Cina nel dicembre 2019.[1]
La modalità di trasmissione predominante di questo patogeno risulta essere da uomo a uomo, generalmente attraverso particelle di acqua e muco infette (droplet) che le persone liberano starnutendo o tossendo e che poi vengono inspirate da un altro individuo.[2] Una persona infetta può presentare sintomi dopo un periodo di incubazione che può variare tra 2 e 14 giorni circa, durante i quali può comunque essere contagiosa.[3][4]
l COVID-19 colpisce principalmente il tratto respiratorio inferiore e provoca una serie di sintomi descritti come simil-influenzali,[2][5] tra cui febbre, tosse, respiro corto, dolore ai muscoli e stanchezza; nei casi più gravi può verificarsi una polmonite, una sindrome da distress respiratorio acuto, sepsi e shock settico, fino ad arrivare al decesso del paziente. Il trattamento della malattia consiste in genere nel gestire i sintomi clinici poiché, a febbraio 2020, non è stato trovato alcun rimedio efficace, tuttavia sono allo studio alcuni farmaci, tra cui alcuni antivirali, già impiegati con altri agenti patogeni.[2] La malattia venne identificata per la prima volta nei primi giorni del 2020 dalle autorità sanitarie della città di Wuhan, capitale della provincia di Hubei in Cina, tra i pazienti che avevano sviluppato una polmonite senza una causa chiara.[2]
Il 22 gennaio 2020, il Journal of Medical Virology ha pubblicato un rapporto con analisi genomica indicante che i serpenti nell’area di Wuhan sono “il più probabile serbatoio di animali selvatici” per il virus, ma sono necessarie ulteriori ricerche.[6]
Tuttavia, il modo in cui il virus potrebbe adattarsi sia agli ospiti a sangue freddo che a quelli a sangue caldo rimane un mistero, e lo rimarrà dal momento che il patogeno non si adatta ma presenta caratteristiche di adattabilità derivanti da processi entropici.[7] Tra i fautori delle “ibridazioni evoluzioniste” si sono diffuse alcune teorie tra le quali spicca quella secondo cui un “evento di ricombinazione omologa” può aver mescolato un virus “clade A” (virus simili a SARS Bat CoVZC45 e CoVZXC21) con la proteina legante del recettore di un Beta-CoV ancora sconosciuto.[8][9]
A favore di tale ipotesi essi citano le sequenze del betacoronavirus di Wuhan che mostrano somiglianze con i betacoronavirus trovati nei pipistrelli; tuttavia, il virus è geneticamente distinto da altri coronavirus come quello correlato alla sindrome respiratoria acuta grave (SARS) e il coronavirus correlato alla sindrome respiratoria mediorientale da Coronavirus (MERS).[10]
Il SARS-CoV-2 è strettamente correlato al SARS-CoV-1 ( 75%-80%) ma comunque diverso, e la somiglianza è facilmente spiegabile con la famiglia di appartenenza e le già citate mutazioni. Gli istituti di ricerca e per il controllo delle malattie cinesi hanno isolato cinque genomi del nuovo coronavirus, tra cui BetaCoV/Wuhan/IVDC-HB-01/2019, BetaCoV/Wuhan/IVDC-HB-04/2020, BetaCoV/Wuhan/IVDC-HB-05/2019, BetaCoV/Wuhan/WIV04/2019 e BetaCoV/Wuhan/IPBCAMS-WH-01/2019.[10][11][12] La sua sequenza di RNA ha una lunghezza di circa 30 kbp (coppie di basi).[10]
Fatto sta che della reale provenienza del COVID-19 esistono solo, a livello mediatico, ipotesi tra le quali viene annoverata con maggiore sicurezza, per l’appunto, quella dell’origine animale (pipistrello). Ora, i virus che saltano tra le specie sono chiamati zoonotici (notare la parola “zoo” nel nome). Tra i virus zoonotici vi sono quelli dell’influenza [13], i quali appartengono alla famiglia dei coronavirus (vedi anche SARS e MERS) e l’HIV (il virus che causa l’AIDS). Tutti questi causano malattie nell’uomo. Alcuni di loro sono persistiti nella popolazione umana per molto tempo. Fortunatamente, molti “nuovi” virus si esauriscono per causa entropica. Anche i virus si degraderanno geneticamente nel tempo. Mentre si moltiplicano, accumulano mutazioni e, a volte, quelle mutazioni li indeboliscono fino al punto in cui non renderanno più possibile la sopravvivenza del patogeno. Questo però non è sempre il caso, e alcuni virus, come l’HIV o il virus del raffreddore umano (un coronavirus), possono continuare a propagarsi nonostante l’accumulo di mutazioni, probabilmente per un processo degenerativo più lento e meno compromettente per la loro funzionalità. La cosa è dipendente da molti fattori diversi e, cosa molto importante da tenere bene a mente, non esistono due virus uguali.
Abbiamo creato tutti i tipi di reti di sicurezza per prevenire la diffusione dell’infezione e il mondo sta iniziando a reagire più rapidamente alle minacce emergenti. Purtroppo, lo Stato italiano non ha saputo gestire la situazione con i tempi e la prontezza adeguati per arginare sul nascere una pandemia che sta vedendo un aumento dei contagiati di centinaia ogni giorno. Le quarantene, un accurata igiene delle mani e le vaccinazioni fanno tutte parte di quella strategia di contenimento, a seconda della gravità, del rischio e del fatto che abbiamo trovato o meno un modo per vaccinare contro un determinato patogeno. Prendiamo in considerazione la recente epidemia di Ebola in Africa. Sono stati spesi molti milioni di dollari per aiutare quelle persone in quel periodo terribile e ancora una volta è stato prevenuto un focolaio mondiale. L’epidemia di coronavirus, partito dalla Cina, sta attraversando l’intero pianeta. Per fortuna, il tasso di mortalità, che è iniziato a circa il 20%, è diminuito a circa il 3%, probabilmente a causa del miglioramento dei trattamenti medici della malattia. Ma anche un tasso dell’1 o 2% equivarrebbe a molti milioni di morti se sfuggisse completamente al controllo e diventasse comune come, diciamo, il raffreddore. Ma la comunità scientifica ha risposto molto rapidamente, sebbene non ovunque, ed è questo il problema. In breve tempo, sequenze geniche multiple per il virus sono state completate e pubblicate in banche dati pubbliche e microscopi elettronici hanno prodotto immagini di ciò con cui abbiamo a che fare. La velocità in questo è stata senza precedenti. Ciò nonostante, le sperimentazioni per un vaccino specifico sono ancora allo stadio su animali, e si presume che ci vorranno dai 12 ai 18 mesi prima che sia pronto un vaccino e altrettanto tempo prima che venga prodotto su larga scala e distribuito.
Virus dell’influenza A sottotipo H1N1
Se questo focolaio virale segue il corso di quelli precedenti, il COVID-19 potrebbe esaurirsi in un certo, sconosciuto, lasso di tempo. Questo è apparentemente quello che è successo al virus dell’influenza umana H1N1 che colpì il mondo tra il 1918 ed il 1920, uccidendo milioni di persone. Durò per 40 anni prima di scomparire. È stato reintrodotto da un campione di laboratorio conservato nel 1976 ed è durato altri 33 anni prima di scomparire di nuovo durante la pandemia di suina H1N1 del 2009-2010, che non era un virus particolarmente letale. Le versioni successive non avevano la natura letale di quelle precedenti e il fatto che l’H1N1 umano non potesse persistere nella popolazione umana in modo perdurevole è una buona prova del fatto che stava subendo entropia genetica. In effetti, il virus stava subendo oltre 14 mutazioni all’anno mentre era attivo e oltre il 10% del suo genoma era mutato prima che si estinguesse.[14] Ciò corrisponde a delle simulazioni computerizzate.[15]
Ma il COVID-19 non è l’influenza. Inoltre non siamo certi di dove o come questo virus abbia avuto origine, sebbene si insista sulla provenienza da pipistrelli. Ad ogni modo, dovrà essere trattato con molta attenzione e i nostri sistemi sanitari devono trattarlo come una minaccia seria e immediata. Non possiamo aspettare decenni perché l’entropia genetica lo cancelli dalla storia.
Oltre alla pandemia in corso alcuni speculatori e “complottari della domenica” hanno dato il via a chi pubblicava l’assurdità più grande riguardo a presunti metodi naturali per trattare questa emergenza che, vorrei ricordarlo, è mondiale e non è propriamente una semplice influenza o poco più. Dalla Vitamina C, alla A per finire con la D e tisane calde, i peggiori venditori di supercazzole delle italiche lande hanno letteralmente invaso i social convincendo della bontà delle loro affermazioni le povere e deboli menti degli ingenui navigatori. I “Curu” in questione hanno anche affermato che con l’integrazione di Vitamina A si può ridurre la mortalità addirittura al virus Ebola. Sono arrivati a citare alcuni studi a sostegno delle loro tesi sicuri di divenire “inattaccabili” da possibili (e probabili) detrattori. Come spesso capita a coloro che citano studi senza a) valutarne la qualità e b) l’attendibilità contestualizzando l’applicazione delle modalità riportate, i loro “bastioni” cedono senza troppi sforzi ad una lettura attenta della bibliografia citata ed esistente.
Ma diamo un occhiata a questi fantomatici studi. Le speculazioni ai tempi del COVID-19
Il Ragionier Curu di Gran Croc FanTozzi
Il noto “Curu” FanTozzi riporta, tra la bibliografia dei suoi post a tema COVID-19, un (uno!) caso studio con baldanzosa certezza che tale lavoro possa avvalorare le sue tesi. Il “soggettone” tralascia innanzitutto un dato importante: la gerarchia nella piramide delle evidenze scientifiche!. I casi studio occupano uno dei gradini più bassi di tale piramide (vedi immagine seguente), appena sopra il parere degli esperti, vale a dire la base!. Detto ciò, nello studio “rivelatore” , pubblicato su “Viruses” [16], si descrive la somministrazione di 20.000 UI di vitamina A e 2000 UI di vitamina D in un gruppo di bambini dai 2 agli 8 anni che erano stati vaccinati per l’influenza. Il risultato di tale trattamento è stata la semplice ed ovvia dimostrazione che l’integrazione di vitamina A e D può migliorare le risposte immunitarie ai vaccini quando i bambini sono insufficienti con vitamina A e D al basale! Avete capito? “l’integrazione di vitamina A e D può migliorare le risposte immunitarie ai vaccini quando i bambini sono insufficienti con vitamina A e D al basale”! Bene, sembra però che il Fantozzi esponga il risultato in modo tale da vederne la conferma che la supplementazione di vitamina A e D causi universalmente, e indipendentemente dalla condizione nutrizionale del soggetto, una risposta immunitaria migliore. Arriva a affermare che senza un adeguato apporto vitaminico (che, voglio ricordare, corrisponde all’RDA) i vaccini non funzionano. Che scoperta sensazionale, specie nell’area occidentale del mondo che, nonostante sia caratterizzata da una cattiva alimentazione, raramente presenta soggetti carenti in Vitamina A mentre per la Vitamina D il discorso è leggermente diverso.
Il problema che lo studio appena esposto in sintesi, così “tradotto” dallo speculatore, trasmette la errata convinzione che le metodiche integrative da lui proposte possano dare risultati preventivi contro l’attuale, o altre, epidemie. La domanda a questo punto è: quale prova a favore? Nessuna! E’ la solita conclusione riduzionista: se 1+1 fa 2 vorrà dire che aggiungendo un altro 1 avrò un risultato migliore. Peccato però che la biochimica sia un calcolo a più variabili sia per le risposte di base universalmente condivise dagli esseri umani sia per le differenze di risposta individuale.
L’affermazione, o meglio “le affermazioni”, più eclatante è stata a proposito del trattamento con vitamina A (200.000 UI/die) come pratica per ridurre la mortalità nei pazienti affetti dal virus Ebola, pubblicata sul “Journal of Nutrition”. [17] Ora, se esaminiamo lo studio notiamo immediatamente che è stato effettuato in Africa Occidentale, luogo del focolaio del patogeno, caratterizzato da popolazioni con gravi carenze alimentari! Non c’è da stupirsi che il trattamento con vitamina A in soggetti malnutriti abbia garantito un margine di sopravvivenza più alto! Organismi ben nutriti avranno sistemi più efficienti! Non fatevi forviare dalla percentuale di mortalità tra i soggetti trattati (55,0%) e quelli non trattati (71,9%), che è del 16,9% in meno. essa è da ricollegarsi alla prima citata condizione aggiunta di variabili nutrizionali che hanno interessato i soggetti trattati. E, cosa più importante, la riduzione della mortalità non significa immunità al patogeno! Ma, ancora una volta, l’esposizione sensazionalistica del FanTozzi punta al convincimento delle sue teorie alternative, cioè: assumi vitamine come te lo dico io e sarai un super uomo…
La cosa tragicomica è che i post in questione sono caratterizzati per un 90% da attacchi degni del più ardito “anti-vax”. Tutto ciò tra affermazioni ridicole come “mi può pure starnutire in faccia un pipistrello di Whuan” (che inguaribile umorista lol) perché con la “Paleo Dieta” e la Vitamina D diventiamo tutti resistenti a qualsiasi patogeno. L’isteria dei “figli dello scientismo”, relativisti per educazione, complottari per impreparazione e amore cieco per i propri convincimenti che mai vengono messi in dubbio, è dannosa tanto quanto il COVID-19. Perché? Perché convince poveri ingenui a credere ad affermazioni giustificate dalla distorsione di una manciata di studi privi di contestualizzazione! Cosa che, di conseguenza, mette a rischio questi individui e la collettività!
Il noto libro di “Panzerotti”
Avrei potuto citare un altro paio di studi utilizzati come cavallo di battaglia dal FanTozzi ed altri “Curu” come il Panzerotti. Se volete approfondire basta che vi facciate un giro nei loro siti/pagine, la chiave di lettura non cambia comunque. Personalmente preferisco mostrarvi la Verità sul reale potenziale di specifiche vitamine a livello del sistema immunitario e la loro applicazione in soggetti affetti da patologie o virus e come mezzo preventivo.
Vitamine e prevenzione da contagio virale
Vitamina C
Vitamina C (Acido Ascorbico)
Una delle vitamine più “gettonate” del momento è senza dubbio la Vitamina C. Sono ormai anni che vengono diffusi studi sui noti benefici di una adeguata assunzione di tale vitamina legata, per esempio, al suo potenziale antiossidante e alla implicazione nella sintesi di collagene. Ultimamente, però, questa vitamina idrosolubile è stata ammantata di proprietà al limite del miracoloso nel contrastare infezioni virali se utilizzata a dose elevate (>1g/die). Tali affermazioni si basano, come esposto in precedenza, su alcuni studi di basso o medio valore di evidenza. Infatti, se andiamo a consultare attentamente la letteratura scientifica le valutazioni della Vitamina C in questo particolare frangente (sistema immunitario e prevenzione/trattamento da contagio virale) si ridimensionano fortemente sebbene la sua importanza a livello nutrizionale è indiscussa come quella di altri micronutrienti.
Secondo una meta-analisi (fascia alta delle evidenze scientifiche) sul tema della valutazione di dosi di 200mg o più di vitamina C, la sua assunzione non è riuscita a ridurre la frequenza dei raffreddori nella popolazione normale, ma è riuscita a ridurre la durata dei raffreddori (in media dall’8 al 14%).[18] [19] Quando si osservano studi che indagano lo stress fisico estremo (nei maratoneti e sciatori), il rischio di avere un raffreddore è stato dimezzato (come è stato notato in precedenti meta-analisi [20]) È stato evidenziato [21] che le osservazioni fatte da Linus Pauling sulle interazioni della vitamina C con il raffreddore comune potrebbero essere state influenzate da coorti atletiche, come uno degli studi più convincenti di cui ha scritto [22] il quale riguardava i bambini facenti parte di una scuola di sci (PDF tedesco [23]).
La maggior parte della letteratura usa dosaggi compresi tra 200mg e 2g e, sebbene ciò sembri essere inefficace per prevenire o ridurre l’insorgenza del raffreddore comune, sembra ridurne leggermente la durata. Ma ciò sembra manifestarsi nè più e nè meno nelle modalità di fattore concomitante dal momento che sono stati osservati benefici più marcati nelle popolazioni atletiche, dove il rischio può essere dimezzato. Nota bene: “rischio dimezzato” e “fattore concomitante”! Non “immunità assoluta”, non “efficace contro ogni patogeno” e non “composto chiave”!
Un dato interessante riguardante la Vitamina C è rappresentato da uno studio nel quale è stato rilevato che il Mycobacterium tuberculosis resistente ai farmaci (batteri che causano la tubercolosi) è altamente sensibile alla degradazione da parte di tale vitamina , cosa piuttosto singolare in quanto altri batteri testati non ne sono stati interessati. [24-190] Ciò era dovuto a un elevato contenuto di ferro in questo batterio, che è ridotto (da Fe3 + a Fe2 +) e provoca effetti proossidanti dopo aver reagito con l’ossigeno.
Ovviamente si tratta di uno studio nel quale si parla di batteri. Da quanto riportato chiaramente, dovreste essere ormai consci della differenza tra un battere e un virus.
Comunque, non ci sono al momento prove su esseri umani che mostrino un effetto terapeutico nel trattamento della tubercolosi farmaco-resistente a base di Vitamina C, anche se le premesse di un esito positivo non manchino.
Tornando a parlare di COVID-19, attualmente non esiste una sola prova concreta che il supplemento possa aiutare un infetto a debellare tale patogeno.
Gli scienziati in Cina stanno attualmente effettuando test per scoprire gli effetti della Vitamina C sul nuovo Covid-19. Il dosaggio preso in esame è circa 60 volte la quantità giornaliera raccomandata dal SSN.
I ricercatori dell’Ospedale Zhongnan dell’Università di Wuhan hanno somministrato a 120 pazienti affetti dal virus 24g di vitamina C ogni giorno per sette giorni. I risultati devono ancora essere pubblicati e gli addetti ai lavori affermano che non ne conosceremo l’efficacia ancora per qualche tempo (diffidare delle conclusioni sensazionalistiche presenti in rete).[25]
E la Vitamina D?
Uno dei pochi studi degni di nota riguardanti il trattamento/prevenzione di infezioni virali riguarda un esperimento svolto su bambini in età scolastica al fine di misurare l’incidenza dell’influenza stagionale A se questi venivano trattati o meno con Vitamina D3 (1200 IU/die).[26] L’influenza A ha interessato 18 bambini su 167 (10,8%) nel gruppo trattato con Vitamina D3 rispetto ai 31 bambini su 167 (18,6%) del gruppo placebo [rischio relativo (RR), 0,58; IC al 95%: 0,34, 0,99; P = 0,04]. La riduzione dell’influenza A è stata più evidente nei bambini che non avevano assunto in precedenza altri integratori di vitamina D (RR: 0,36; IC al 95%: 0,17, 0,79; P = 0,006), quindi carenti di base, e che hanno iniziato la scuola materna dopo i 3 anni (RR: 0,36; IC al 95%: 0,17, 0,78; P = 0,005), con una condizione immunitaria non ben adattata in seguito a contatto con individui potenzialmente infetti da patogeni di diversa natura. Nei bambini con una precedente diagnosi di asma, si sono verificati attacchi di asma come esito secondario in 2 bambini trattati con vitamina D3 rispetto a 12 bambini trattati con placebo (RR: 0,17; IC al 95%: 0,04, 0,73; P = 0,006).
La conclusione a questo studio è stata un semplice suggerimento che l’integrazione di vitamina D3 durante il periodo invernale possa ridurre l’incidenza dell’influenza A, specialmente in specifici sottogruppi di scolari. Questo processo è stato registrato su https://center.umin.ac.jp come UMIN000001373. In definitiva, nulla di sensazionalistico, di nuovo…
Un sondaggio sui dati della letteratura genera mostra alcune controversie e dubbi sul possibile ruolo della vitamina D nella prevenzione delle infezioni influenzali. Tuttavia, ci sono dati ottenuti in vitro o in vivo che indicano l’attività antivirale della vitamina D in caso di influenza. Oggi la conclusione finale è che il suo significato come agente anti-influenza rimane irrisolto, ma ciò non significa che queste considerazioni siano insensate. È molto importante rendersi conto che l’ampio spettro di attività della vitamina D non esclude tale ruolo.
Alcuni dei seguenti studi non riguardano strettamente l’infezione influenzale, ma anche le malattie respiratorie simil-influenzali delle infezioni del tratto respiratorio (RI) e la polmonite. Tuttavia, il periodo di tempo degli studi, ovvero ottobre-marzo o i mesi invernali, non esclude le infezioni influenzali, più comuni in autunno e in inverno. Secondo Cannell et al. [27], se la vitamina D è uno “stimolo stagionale”, la carenza di vitamina D dovrebbe predisporre i pazienti alle infezioni respiratorie.
Moan et al. [28] hanno confrontato la stagionalità dei decessi per influenza e polmonite in Norvegia con i livelli sierici di vitamina D. Il periodo di studi è stato tra il 1980 e il 2000. La conclusione finale di questi studi è stata che l’alto numero di decessi per influenza e polmonite invernali in Norvegia era correlato ai bassi livelli di vitamina D in questa stagione (anche se, per latitudine, la carenza di Vitamina D è assai diffusa per tutto l’anno). I dati supportano l’ipotesi che la vitamina D agisca da protettore contro l’influenza e la polmonite, sebbene non sia chiaro se richieda alcun aiuto o quale meccanismo domini nella battaglia contro le infezioni virali.
Laaksi et al. [29] hanno condotto uno studio in doppio cieco controllato con placebo (ottobre-marzo) che ha coinvolto 164 giovani finlandesi volontari (18-28 anni di età) sottoposti a addestramento militare. I soggetti sono stati assegnati in modo casuale al gruppo di intervento (n = 80), che ha ricevuto 400UI di vitamina D al giorno, o al gruppo placebo (n = 84). Dopo sei mesi di studio, il gruppo integrato ha mostrato una concentrazione sierica media di 25 (OH) D (± DS) di 71,6 ± 22,9 nM / L (n = 58) e il gruppo placebo ha mostrato 51,3 ± 15,5 nM / L di 25 (OH) D (n = 50) (p <0,001). Il risultato principale considerato è stato il numero di giorni assenti dal servizio a causa di infezione respiratoria. La percentuale di uomini che sono rimasti sani durante il periodo di studio di sei mesi è stata maggiore nel gruppo integrato (51,3%) rispetto al gruppo placebo (35,5%) (p = 0,045). I risultati di cui sopra hanno fornito alcune prove dell’effetto preventivo della supplementazione di vitamina D contro l’infezione del tratto respiratorio e, secondo l’analisi della regressione di Cox, gli autori hanno notato che il rapporto di rischio per assenza dal servizio a causa dell’infezione del tratto respiratorio era inferiore nel gruppo integrato rispetto al gruppo placebo. Come dimostrato dagli autori, sono necessari studi randomizzati controllati con dosi più elevate e popolazioni più grandi per esplorare l’effetto preventivo della supplementazione di vitamina D sull’infezione acuta del tratto respiratorio. Urashima et al hanno presentato risultati promettenti e incoraggianti sull’integrazione con vitamina D per prevenire l’influenza. [30]
Il ruolo positivo della vitamina D nelle infezioni respiratorie e nella funzione polmonare è stato confermato da Berry et al. [31], che utilizzavano dati trasversali di 6789 partecipanti in una coorte di nascita britannica del 1958 a livello nazionale. Gli autori hanno misurato 25 (OH) D, funzionalità polmonare, capacità vitale forzata e infezioni respiratorie dall’età di 45 anni. Hanno mostrato un’associazione lineare tra lo stato della vitamina D e le infezioni stagionali e la funzione polmonare. Ogni aumento di 10nM/L della 25 (OH) D era associato a un rischio inferiore di infezione del 7%. Risultati simili sono stati ottenuti anche in altri studi. Aregbesola et al. [32] hanno studiato il rischio di polmonite ospedalizzata in una popolazione generale di anziani nella Finlandia orientale. Sulla base dello studio, gli autori hanno suggerito un effetto inverso della concentrazione sierica di 25 (OH) D sul rischio di polmonite in via di sviluppo. Jones et al. [33] hanno esaminato 46 campioni di sangue residuo di adulti e bambini, alcuni dei quali hanno avuto infezioni da virus dell’influenza del tratto respiratorio. Sono stati condotti test per la proteina legante il retinolo (RBP), la vitamina D e gli isotipi dell’anticorpo. I risultati hanno mostrato che 44 campioni presentavano insufficienze o carenze di RBP e / o vitamina D. Inoltre, la vitamina D era correlata alle IgM ematiche e alle IgG3, mentre l’RBP era correlata alle IgG4 e alle IgA. È noto che adeguati livelli delle vitamine A e D sono fondamentali per le risposte immunitarie ottimali nelle superfici della mucosa dei topi. Soprattutto, l’IgA è una prima linea di difesa contro i patogeni della mucosa. Quindi, secondo gli autori, i risultati suggeriscono che anche nell’uomo esiste una correlazione tra i livelli di vitamina A e D e il profilo anticorpale. Gli autori suggeriscono che le vitamine possono supportare lo sviluppo delle cellule dendritiche necessarie per la presentazione dell’antigene; Attivazione e homing delle cellule T; Attivazione, divisione e maturazione delle cellule B; e / o la stabilizzazione di cellule differenziate che producono anticorpi. Secondo Jones et al. [33], questi risultati riconoscono la necessità di ulteriori studi sulla correzione della supplementazione di vitamine, in particolare al momento della vaccinazione del virus respiratorio, per migliorare l’efficacia del vaccino e per la protezione contro le malattie delle vie respiratorie.
Effetti intriganti sono stati presentati da Mamani et al. [34] e Brance et al. [35] nei loro studi. Entrambi gli studi hanno mostrato una correlazione inversa tra il livello di 25 (OH) D e la gravità della CAP (polmonite acquisita in comunità), definita come punteggio CURB65 (confusione, uremia, frequenza respiratoria, bassa pressione sanguigna, ≥65 anni) . Inoltre, secondo Brance et al. [35], concentrazioni più alte di 25 (OH) D sono risultate correlate con un CCI inferiore (indice di comorbidità di Charlson). Nanri et al. [36], in un caso studio di controllo su una coorte di lavoratori in quattro società in Giappone durante la stagione invernale, hanno riscontrato che un rischio di influenza inferiore era associato alla sufficienza di vitamina D (≥30 ng/mL), ma solo tra i partecipanti non vaccinati . In un sottogruppo vaccinato in precedenza contro l’influenza, la concentrazione sierica di 25 (OH) D non era correlata in modo significativo con l’incidenza dell’influenza diagnosticata dal medico.
Esistono dati in vitro e in vivo, nonché dati risultanti da studi sull’uomo, che non dimostrano alcun significativo effetto in dall’integrazione di vitamina D nelle infezioni respiratorie virali.
Gui et al. [37] hanno mostrato l’impatto negativo del trattamento 2D 1α, 25 (OH) sulla risposta immunitaria innata generata dall’infezione da H9N2 nei topi, in particolare nella fase successiva della malattia. Sebbene abbia ridotto il gene influenzale M (codificando la proteina M correlata alla risposta infiammatoria e alla replicazione del virus), IL-6 e IFNβ nelle cellule A549 prima e dopo l’infezione da influenza H9N2, gli autori hanno scoperto che non influiva sulla replicazione del virus in vitro e in vivo. Inoltre, l’effetto del trattamento 1α,25(OH)2D dipendeva dallo stadio della malattia. Come mostrato in vivo, 1α,25(OH)2D a sotto-regolato l’infiammazione polmonare nei topi due giorni dopo l’infezione, ma ha aumentato la risposta infiammatoria da 4 a 6 giorni dopo l’infezione. Allo stesso tempo, l’espressione della citochina antivirale IFNβ era significativamente più alta a due giorni dopo l’infezione e inferiore ai giorni 4 e 8. Questi effetti erano coerenti con il periodo di massima perdita di peso corporeo e il danno polmonare nei topi trattati con calcitriolo. Il motivo dell’attività antinfiammatoria positiva di 1α,25(OH)2D notato nelle cellule A549 e l’effetto opposto riportato nei topi durante la fase successiva dell’infezione non è chiaro. Gli autori indicano due possibili spiegazioni. Il primo è che l’attività della vitamina D in vivo è complessa e ha un impatto su così tanti percorsi e meccanismi che può influenzare un componente di questo sistema, ma non l’altro. In secondo luogo, i virus dell’influenza aviaria come l’H9N2 inducono meccanismi diversi nei topi e nell’uomo. Ciò è in accordo con la conclusione fornita da Grant e Giovanucci [38], che hanno discusso, riguardo ai dati che dimostrano che la soppressione delle citochine proinfiammatorie da parte della vitamina D non ha ridotto il rischio di morte nei topi infettati da virus H5N1, che tale effetto non dovrebbe essere applicato alle infezioni da H1N1 nell’uomo a causa delle differenze nella risposta immunitaria. Anche l’attività antinfiammatoria di 1α,25(OH)2D su A549 infetto da H1N1 è stata dimostrata da Khare et al. [39]. Gli autori hanno osservato che il trattamento 1α,25(OH)2D prima o post-infezione riduceva i livelli di RNA di IL-6 e IL-8 e riduceva i livelli di TNFα, IFNβ e ISG15 indotti dall’infezione. L’1α,25(OH)2D non ha influenzato la clearance virale, analogamente ai risultati riportati da Gui et al. [37], ma ha ridotto l’autofagia e ripristinato l’apoptosi aumentata osservata nell’infezione da H1N1 al suo livello costitutivo.
Negli studi presentati da Jorde et al. [26], la vitamina D sembrava rendere l’infezione influenzale una malattia significativamente più prolungata rispetto ai pazienti che assumevano placebo. Nello studio sono stati inclusi 569 soggetti provenienti da 10 diversi studi clinici. Dei soggetti, 289 sono stati randomizzati a ricevere vitamina D (1111-6800 UI / giorno) e 280 hanno ricevuto placebo. Malattia simil-influenzale è stata segnalata in 38 soggetti nel gruppo vitamina D e 42 nel gruppo placebo. In questi gruppi, 25 e 26 soggetti, rispettivamente, hanno mostrato sintomi clinici dell’influenza secondo i criteri definiti. Nel gruppo vitamina D, la durata della malattia era significativamente più lunga rispetto al gruppo placebo (2-60 giorni contro 2-18 giorni; p = 0,007). Uno dei punti deboli di questo studio, come sottolineato dagli autori, era che lo studio era retrospettivo e si basava su sintomi auto-riportati; quindi, non c’era una diagnosi definita di influenza.
Alcuni risultati contrastanti sono stati presentati da Urashima et al. [40]. Come è stato riportato dagli autori, uno studio randomizzato controllato sugli effetti della supplementazione di vitamina D sulla malattia influenzale durante la pandemia di H1N1 del 2009 ha rivelato che l’influenza A o B si è verificata meno nel gruppo vitamina D che nel gruppo placebo solo durante la prima metà del lo studio. Durante il secondo mese, i risultati del gruppo vitamina D erano simili a quelli del gruppo placebo. Gli autori hanno osservato effetti simili, vale a dire l’azione preventiva della supplementazione di vitamina D solo nella parte iniziale degli studi, negli studi condotti tra gli studenti che hanno ricevuto 2000UI di vitamina D al giorno per due mesi. Come mostrato dall’analisi post-hoc, l’influenza A si è verificata significativamente meno nel gruppo vitaminico (2/148, 1,4%) rispetto al gruppo placebo (8/99, 8,1%), ma solo nel primo mese dello studio. Il vantaggio iniziale è stato perso durante il secondo mese. Questi risultati avrebbero potuto essere correlati con una più lunga integrazione con vitamina D e sono indirettamente coerenti con i risultati di Urashima et al. [30], che non hanno notato l’impatto della vitamina D sull’incidenza di influenza A nei bambini che avevano assunto più di un integratore di vitamina D.
La mancanza di correlazione tra infezioni respiratorie e supplementazione di vitamina D è stata dimostrata da Li-Ng et al. [41], che ha descritto uno studio randomizzato controllato per la prevenzione delle infezioni sintomatiche del tratto respiratorio superiore, condotto durante l’inverno. In totale, 162 adulti hanno ricevuto 2000UI di vitamina D al giorno per 12 settimane. Non vi era alcuna differenza nell’incidenza delle infezioni e nella durata o gravità dei sintomi di infezione delle vie respiratorie tra i gruppi integrato e placebo (48 vs 50 casi, rispettivamente, p = 0,57 e 5,4 ± 4,8 giorni vs. 5,3 ± 3,1 giorni, rispettivamente, p = 0,86). Vale la pena notare che dopo 12 settimane, la concentrazione sierica media di 25 (OH) D nel gruppo integrato era 88,5 ± 23,2 nM / L e nel gruppo placebo era 63,0 ± 25,8 nM / L. È un dato di fatto, la concentrazione sierica di 25 (OH) D nel gruppo integrato non era troppo efficiente; il valore 88,5 ± 23,2 nM / L è posto vicino al limite inferiore definito del livello 25 (OH) D riportato come livello guida (75–200 nM / L). [42] Negli studi condotti nel 2007, gli stessi autori hanno riscontrato una significativa riduzione del raffreddore e dell’influenza nelle donne che assumevano 800 o 2000 UI/giorno di vitamina D. Sorprendentemente, nell’ultimo anno dello studio, le donne integrate con 2000UI erano ancora vitamina D- carenti. [43]. Secondo Aloia e Li-Ng [43], lo studio dovrebbe dovuto usare abbastanza colecalciferolo per aumentare i livelli di 25 (OH) D fino a quelli raggiunti dalla sintesi naturale della pelle in estate, vale a dire ca. 50ng/mL. Una simile mancanza di effetti protettivi della vitamina D è stata riscontrata da Lappe et al. [44] in studi clinici randomizzati in doppio cieco, controllati con placebo, basati sulla popolazione. Tra le donne anziane in postmenopausa sane con un livello medio basale nel siero di 25 (OH) D pari 32,8ng/mL, l’integrazione con vitamina D e calcio rispetto al placebo non ha comportato un rischio significativamente più basso di tutti i tipi di cancro dopo quattro anni.
Nessuna associazione tra vitamina D e risposta sierologica post-vaccino è stata dimostrata nei pazienti con infezione da HIV vaccinati con influenza monovalente A (H1N1) da Crum-Cianflone et al. [45]. Durante uno studio prospettico di coorte di 124 partecipanti (64 infetti da HIV e 64 non infetti), la sieroconversione misurata come aumento> 4 volte del titolo anticorpale è stata raggiunta il giorno 28 post-vaccinazione nel 56% delle persone con infezione da HIV rispetto al 74% di quelli non infetti da HIV, ma la carenza di vitamina D era significativamente diversa tra i due gruppi.
Studi sull’immunizzazione di bambini vaccinati con vaccini antinfluenzali vivi attenuati o inattivati in dipendenza della concentrazione sierica di 25 (OH) D condotta da Lin et al. [46] non ha inoltre fornito una risposta alla domanda: “La vitamina D può agire da adiuvante nei vaccini antinfluenzali?”. I livelli di vitamina D nei titoli di anticorpi sierici e influenzali sono stati misurati prima e 21 giorni dopo la vaccinazione con vaccino antinfluenzale vivo attenuato o inattivato. Sorprendentemente, bassi livelli di vitamina D erano associati a un titolo anticorpale più elevato contro il virus vivo attenuato e questo effetto era specifico del ceppo (p <0,05). Risultati simili sono stati ottenuti da Principi et al. [47] quattro anni prima. Sulla base di uno studio prospettico, randomizzato, in singolo cieco, controllato con placebo su 116 bambini, gli autori hanno indicato che l’integrazione giornaliera con 1000UI di vitamina D per quattro mesi a partire dall’iniezione della prima dose del vaccino trivalente Fluarix contro l’influenza non ha modificato significativamente la risposta anticorpale. Allo stesso modo, Lee et al. [48] non ha osservato alcuna correlazione tra i livelli di 25 (OH) D e il titolo anticorpale post-vaccinazione in uno studio osservazionale retrospettivo condotto su 437 giovani membri sani dell’esercito. Solo 224 di essi (51,3%) hanno dimostrato un aumento del titolo post-vaccinazione antinfluenzale, che non era associato a livelli di 25 (OH) D.
Un riepilogo dei risultati ottenuti da alcuni autori in vitro o in vivo o in studi clinici controllati sull’impatto della vitamina D sulla risposta sierologica ai vaccini anti-influenzali è riportato nella tabella visibile cliccando qui.
Gli autori hanno un approccio critico agli studi clinici che non dimostrano un effetto benefico della vitamina D sul sistema immunitario o indicano la mancanza di alcun effetto di questa vitamina sull’immunizzazione post-vaccino. I principali motivi potenziali di questo risultato, che vengono spesso considerati sono: un tempo troppo breve tra i test di vaccinazione e livello di anticorpi, che potrebbe non essere sufficiente a rivelare l’immunostimolazione; o una dose troppo bassa di vitamina D, dando un livello di concentrazione di 25 (OH) D troppo basso per mediare nei processi di immunizzazione [49]. Ad esempio, Lee et al. [50], nei loro studi, hanno utilizzato i valori limite clinici per insufficienza di 20-30ng/mL e per carenza <20ng/mL, mentre, come dimostrato dagli autori, alcuni esperti hanno dimostrato che è richiesto un livello più alto (≥ 40ng/mL). Alcuni autori ammettono anche che nei loro studi le cellule T non sono state incluse. Pertanto, non è stato esplorato alcun potenziale impatto della vitamina D su questa parte della risposta immunitaria [50, 51]. Il prossimo potenziale motivo è che campioni troppo piccoli vengono utilizzati negli studi o i profili del sistema immunitario nei pazienti con malattie comorbide sono diversi rispetto ai soggetti sani, il che può potenzialmente determinare l’effetto della vitamina D sulla risposta antigene all’influenza, come osservato da Chadha et al. [52]. Inoltre, studi come quanto sopra dovrebbero essere condotti su un gruppo che è rappresentativo di altre popolazioni [50]. Come Lang e Samaras [49] hanno menzionato, ad esempio, il livello di anticorpi presente nel siero preimmunizzato può mascherare il potenziale effetto immunomodulatore della vitamina D sulla risposta immunitaria post-vaccinazione a causa della relazione inversa tra i livelli di anticorpi prima e l’aumento post anticorpale -vaccinazione. Pochi studi hanno incluso soggetti già sottoposti a supplementazione di vitamina D, oppure sono stati condotti in popolazioni selezionate [49,50,53,54].
Il lavoro descritto e fatto da Surman et al. [55-83] ha sollevato la questione molto importante dell’interazione di diversi fattori con conseguente risposta immunitaria. Gli autori hanno riportato nei topi immunizzati con un vaccino contro il virus dell’influenza attenuato che doppie carenze di vitamina A e D hanno ridotto la risposta anticorpale nel tratto respiratorio in misura maggiore rispetto alla carenza di una di queste vitamine. Sebbene l’integrazione con vitamina A abbia avuto un maggiore effetto correttivo rispetto alla vitamina D per la restituzione della sieroprotezione (risposte IgG e IgA), i risultati migliori sono stati con le due vitamine combinate e somministrate al momento della vaccinazione degli animali. Questi studi, sebbene basati su animali, forniscono alcune informazioni utili per la progettazione di studi clinici sull’uomo per il miglioramento dell’efficacia del vaccino antinfluenzale e mostrano che l’approccio all’importanza e il potenziale ruolo dell’integrazione con micronutrienti, come le vitamine, per raggiungere questo scopo l’obiettivo dovrebbe riguardare la loro interazione e sinergia d’azione [56,57].
Lee et al. [58] ha suggerito il ruolo reale della vitamina D nell’immunizzazione contro i virus dell’influenza. È noto che la vitamina D supporta processi antinfiammatori attraverso il suo impatto sulle cellule T. Quindi, secondo gli autori, una misura della maggiore immunità potrebbe non essere il meccanismo d’azione con cui funziona la vitamina D. È possibile che il suo livello più elevato riduca la gravità della risposta infiammatoria provocata dalle infezioni.
Quindi, i risultati presentati non sono troppo entusiasti per quanto riguarda il miglioramento dell’efficacia del vaccino antinfluenzale da parte della vitamina D. Tuttavia, alla luce delle sue fantastiche proprietà, è senza dubbio l’unica vitamina caratterizzata da un così ampio spettro di attività nel sistema immunitario.
L’indagine della letteratura riguardante il ruolo della vitamina D nel sistema immunitario e l’immunizzazione, in particolare contro i virus dell’influenza, non fornisce una risposta inequivocabile e in una sola parola di “sì” o “no” alle domande: “La supplementazione di vitamina D migliora la resistenza dell’ospite all’influenza? ” e “L’integrazione di vitamina D ha un ruolo nella terapia delle malattie da infezione virale?”. Gli autori, come presentati, nonostante la mancanza dei risultati attesi, non escludono il significato di questa vitamina per la battaglia anti-influenza condotta negli organismi umani. Alcuni di essi suggeriscono che gli effetti di 1α,25(OH)2D sull’espressione e la secrezione di chemochine variano tra i patogeni.[59] I risultati degli studi presentati non escludono inoltre la premessa di includere la vitamina D come adiuvante nella direzione del nuovo design del vaccino antinfluenzale. A seguito di Wiwanitkit [60], nonostante l’utilità non confermata della vitamina D come adiuvante per la vaccinazione antinfluenzale, somministrarla contemporaneamente comporta ancora alcuni vantaggi clinici. Secondo Grant e Giovanucci [61], gli integratori di vitamina D o gli alimenti arricchiti dovrebbero essere ulteriormente valutati come una componente forse utile di un programma per ridurre i tassi di mortalità influenzale, specialmente nelle persone anziane.
Gli studi mostrano chiaramente che la vitamina D è senza dubbio parte dei complessi fattori che influenzano la risposta immunitaria. Pertanto, la valutazione dello stato della vitamina D e il mantenimento di livelli sierici ottimali dovrebbero essere considerati in tutti gli adulti e i bambini che invecchiano e i micronutrienti dovrebbero essere considerati uno dei fattori essenziali che migliorano le nostre condizioni di salute in generale e supportano anche la nostra lotta contro le malattie.
E la Vitamina A?
Gli organi immunitari sono organi o tessuti che realizzano la funzione immunitaria e sono luoghi in cui la maggior parte delle cellule immunocompetenti proliferano, si differenziano, maturano, aggregano e rispondono all’immunità. La ricerca ha dimostrato che gli organi immunitari cruciali necessitano di un costante apporto dietetico per mantenere le concentrazioni di Vitamina A (VitA), e in precedenza Acido Retinoico (RA) ha dimostrato sia di promuovere la proliferazione sia di regolare l’apoptosi dei timociti [62,63,64]. Nel timo, la sintesi endogena di retinoidi e retinoidi simili ai glucocorticoidi potrebbero, in effetti, essere coinvolti nella regolazione dei processi di proliferazione e selezione timica, essendo presenti nel timo in quantità funzionalmente efficaci [65]. Nei topi, la VitA porta a un difetto delle risposte immunitarie sia mediate dalle cellule T sia dipendenti dall’anticorpo [66,67]. La VitA può anche inibire il normale processo di apoptosi delle cellule del midollo osseo, che porta ad un aumento del numero di cellule mieloidi nel midollo osseo, nella milza e nel sangue periferico, indicando che la VitA è coinvolta nella regolazione dell’omeostasi del midollo osseo [29]. La VitA probabilmente regola la popolazione del midollo osseo attraverso il recettore dell’acido retinoico (RAR) nel nucleo delle cellule del midollo osseo. Questo legame altera il livello di espressione dei geni dell’apoptosi, come Bcl-2, Fas e altri. I meccanismi specifici con cui questi geni dell’apoptosi regolano l’omeostasi del midollo osseo richiedono ulteriori approfondimenti.
L’acido retinoide svolge ruoli cruciali nella regolazione della differenziazione, della maturazione e della funzione delle cellule del sistema immunitario innato. Le cellule immunitarie innate sono costituite da macrofagi e neutrofili, che danno risposte immediate all’invasione dei patogeni attraverso la fagocitosi e l’attivazione di cellule T killer naturali che svolgono funzioni immunoregolatorie attraverso l’attività citotossica [68,69]. C’è un rapporto che mostra che la VitA è essenziale per il corretto sviluppo e differenziazione dei macrofagi CD169 + del colon [70]. I macrofagi includono principalmente macrofagi M1 che secernono citochine proinfiammatorie e macrofagi M2 che esprimono fattori antinfiammatori. L’all-trans-RA (ATRA) inibisce le reazioni infiammatorie inducendo la differenziazione dei monociti verso il lignaggio dei macrofagi inibendo al contempo il rilascio di fattori infiammatori dai macrofagi, inducendo così i macrofagi M1 nel midollo osseo a trasformarsi in macrofagi M2 [71,72]. L’ATRA agisce sui RAR nel nucleo dei neutrofili, inducendo la differenziazione e l’eterogeneità dei neutrofili attraverso l’attivazione della via di segnalazione mTOR. Questa via migliora le trappole extracellulari e la citotossicità dei neutrofili, consentendo un’uccisione efficiente di più cellule tumorali [73]. Sottoregolando il livello di espressione di IFN-γ e sovraregolando la secrezione di IL-5, l’RA svolge un ruolo regolatorio nella fase iniziale di differenziazione delle cellule T killer naturali [67].
Le cellule dendritiche (DC) sono potenti e versatili cellule presentanti l’antigene e sono sentinelle specializzate del nostro sistema immunitario in grado di orchestrare la risposta immunitaria innata e adattiva [74]. L’ATRA può regolare la differenziazione dei precursori DC [75, 76,77]. I pre-DC residenti nel midollo osseo hanno il potenziale di differenziarsi in DC pre-mucosali (pre-μDC), caratterizzati dall’espressione di recettori intestinali. L’ATRA agisce intrinsecamente a livello cellulare nello sviluppo di pre-μDCs intestinali-tropici per effettuare la differenziazione e favorire la specializzazione dei CD103 + DC intestinali [78]. I pre-DC possono migrare verso la milza, dove possono percepire gli ATRA distorcere la differenziazione verso CD11b + CD8-DC invece che CD11b-CD8α + DC [76]. Il consenso generale sull’effetto di ATRA sulla funzione DC è quello di promuovere un fenotipo antinfiammatorio caratteristico delle DC intestinali [79,80]. Tuttavia, in presenza di IL-15, l’ATRA ha dimostrato di agire come adiuvante nel promuovere la secrezione di citochine pro-infiammatorie IL-12 e IL-23 da parte dei DC [81], e ha proprietà coadiuvanti impreviste che inducono Th1 immunità agli antigeni nutriti. Ciò suggerisce che in condizioni infettive associate all’induzione di IL-15 e IL-6 nella mucosa intestinale, l’ATRA promuoverà anche l’immunità Th17 [82]. Queste osservazioni mettono in guardia contro l’uso di VitA e ATRA per il trattamento di autoimmunità e disturbi infiammatori intestinali associati a livelli elevati di IL-15.
Le cellule T hanno origine da cellule staminali pluripotenti nel midollo osseo. Queste cellule T migrano verso il timo dove si sviluppano in cellule T mature e si spostano verso i tessuti linfoidi periferici interessati. L’intero processo di sviluppo delle cellule T si basa sull’interazione dei recettori di homing delle cellule T con le molecole di adesione endoteliale [83]. L’homing delle cellule T è regolato da varie molecole di adesione che interagiscono con il recettore di homing [83,84,85]. La ricerca ha dimostrato che in condizioni infiammatorie, l’integrina α4β7 e il recettore delle chemochine delle cellule T, CCR9, sono cruciali per la migrazione delle cellule T nell’intestino [83,86]. Dopo aver ricevuto un segnale RA, RARα si lega all’elemento di risposta RA nel gene integrina α4 e regola l’espressione di α4β7. Allo stesso tempo, l’eterodimero di RARα con RXR si lega all’elemento di risposta RAR nella regione del promotore del gene CCR9, svolgendo così un ruolo regolatore aggiuntivo [87,88,89]. Nella lamina propria intestinale, l’AR è un regolatore essenziale per l’homing intestinale delle cellule T CD4 + e CD8 +. La deficienza da VitA ha causato una riduzione della memoria α4β7 (+) / attivato le cellule T negli organi linfoidi e una mancanza di cellule T dalla lamina propria intestinale [84,85]. Sulla base di ciò, la fornitura di ATRA durante la vaccinazione può aumentare la capacità dei vaccini virali basati su cellule T di promuovere le cellule T CD8 + dell’intestino / mucosa, al fine di fornire una maggiore protezione dalla sfida virale della mucosa, e ha anche portato alla formazione di più cellule T CD8 + simili alla memoria centrale specifiche del vaccino in siti sistemici [90,91]. Ulteriori ricerche mostrano che per la sopravvivenza delle cellule T CD8 + [92,93] è necessaria la segnalazione di AR per la sopravvivenza e l’espansione delle cellule T CD8 + e il requisito essenziale è RARα, ma non RARβ o RARγ. L’imaging del corpo intero usando un modello murino di artrite reumatoide ha dimostrato che la segnalazione di AR viene iniziata durante lo sviluppo dell’infiammazione. Inoltre, la segnalazione di RA è limitata al sito di infiammazione sia nel tempo che nello spazio. L’ablazione condizionale della segnalazione di AR nelle cellule T interferisce in modo significativo con la funzione, la migrazione e la polarità dell’effettore delle cellule T CD4 +, indicando il coinvolgimento dell’AR nella migrazione delle cellule T verso l’area dell’infiammazione [94].
Le cellule T regolatorie (Treg) sono una sottopopolazione di cellule T che mantengono la tolleranza immunitaria e regolano la risposta autoimmune [95,96,97,98]. Foxp3 è un fattore di trascrizione essenziale per la differenziazione e la funzione effettrice di Tregs [99,100]. In vivo, ATRA è prodotto principalmente da CD103 + DC nell’intestino [101]. Il fattore di crescita-β (TGF-β) trasforma le citochine convertendo le cellule T native in Treg che prevengono l’autoimmunità. Tuttavia, in presenza di interleuchina-6 (IL-6), è stato anche osservato che il TGF-β promuove la differenziazione dei linfociti T nativi in cellule Th17 che producono citochine IL-17 proinfiammatorie, che promuovono l’autoimmunità e l’infiammazione. L’ATRA, come regolatore chiave delle risposte immunitarie dipendenti dal TGF-β, è in grado di inibire l’induzione indotta da IL-6 delle cellule Th17 proinfiammatorie e promuovere la differenziazione delle cellule Treg anti-infiammatorie [102]. L’ATRA migliora l’espressione di Foxp3 in presenza di TGF-β, inducendo così la differenziazione delle cellule T native in Tregs e inibendo l’espressione di IL-17 [103]. L’ATRA agisce sulla RAR nucleare interagendo con TGF-β per attivare la via di segnalazione ERK1 / 2 e migliorare la modifica dell’istone della regione del promotore Foxp3 e conservare la regione del DNA non codificante. Pertanto, l’ATRA aiuta a mantenere l’espressione del gene Foxp3 e regola la differenziazione e la funzione di Treg [104]. Oltre a indurre la differenziazione delle Treg, è stato riportato che l’ATRA mantiene sia la stabilità delle Treg sia la loro funzione immunoregolatoria [105,106]. Esperimenti in vitro hanno dimostrato che negli ambienti pro-infiammatori, le Treg sono instabili e possono essere trasformate in altre cellule infiammatorie, come le cellule Th17, da citochine come IL-6 e IL-21, favorendo così lo sviluppo dell’infiammazione. Al contrario, l’aggiunta di ATRA inibisce la trasformazione di Tregs in Th17 o altre cellule Th, anche in presenza di IL-6, mantenendo così l’espressione di Foxp3 [105]. L’iniezione locale di Treg non è riuscita a prevenire lo sviluppo di un modello di artrite indotta da collagene, mentre l’iniezione di Treg pretrattati con ATRA ha inibito con successo lo sviluppo di artrite [105]. L’ATRA ha inoltre migliorato la stabilità e la funzionalità delle cellule Treg naturali umane in condizioni infiammatorie [107]. L’ATRA ha impedito la trasformazione di Treg in cellule Th17 e altre cellule infiammatorie inibendo l’espressione di IL-6R sulla superficie cellulare di Treg indotte in modo periferico. Pertanto, ATRA ha migliorato la funzione IL-2, un importante immunomodulatore, e ha promosso la trasformazione delle cellule T native in Treg naturali, inibendo la trasformazione indotta da IL-6 delle cellule T native in cellule Th17 [106]. Inoltre, l’ATRA ha anche la capacità di indurre e promuovere lo sviluppo e la funzione delle cellule Treg indotte nell’uomo. [108]
Sebbene la maggior parte delle prove dimostrino che, a livello farmacologico, la RA inibisce lo sviluppo di cellule infiammatorie e induce o espande le Treg, recenti lavori hanno suggerito che la RA può anche promuovere l’attivazione delle cellule T e le risposte delle cellule T helper a livelli minimi.
Come accennato in precedenza, RA è principalmente prodotto da DC nell’intestino. Alcuni rapporti mostrano che l’AR può essere prodotta anche in altri siti durante una risposta immunitaria in corso [109,110]. Si è già accennato al fatto che la segnalazione di RA è iniziata durante lo sviluppo dell’infiammazione. Allo stesso modo, ci sono prove che dimostrano che l’asse di segnalazione RA – RARα è essenziale per l’immunità adattativa delle cellule T CD4 + poiché le cellule T CD4 + carenti di RARα erano meno efficienti rispetto alle controparti wild-type nell’attivazione policlonale. Inoltre, nelle cellule T carenti di RARα, la fosforilazione di PLCγ ed ERK1 / 2 è stata ridotta e manifesta alterata mobilizzazione Ca2 + e attivazione mTOR / AKT alla stimolazione delle cellule T. Insieme, RARα può regolare le vie di segnalazione a valle dell’impegno del recettore delle cellule T. [111]
A dosi farmacologiche o elevate (10nM o superiori), è stato dimostrato che l’AR inibisce la reazione delle cellule Th17 e induce la generazione di Treg [112] e alte dosi di RA possono compromettere la differenziazione di cellule umane Th17 e Th1 in vitro.[113] Tuttavia, contrariamente alle segnalazioni di RA che inibiscono le risposte Th1 e Th17, alcuni gruppi hanno riferito che l’AR era benefica per la differenziazione delle cellule Th1 e Th17 a basse dosi. In dosi fisiologiche (1nM), l’AR promuove la differenziazione cellulare Th17 in vitro [114,115]. Inoltre, in condizioni di polarizzazione Th1 o Th17, le cellule T carenti di RARa coltivate in vitro non si sono differenziate in cellule Th1 o Th17, supportando il ruolo di RA nella differenziazione delle cellule Th1 e Th17 e i topi carenti in VitA presentano significative risposte Th1 e Th17 in vivo [116,117]. Tutti questi risultati hanno suggerito che l’AR può avere un effetto dose-differenziale sulla differenziazione delle cellule Th17 e Th1 [118]. Il ruolo della VitA / RA sulle cellule Tr1 e Tfh non è ancora chiaro, e merita ulteriori studi per consentire chiarimenti.
La produzione di anticorpi da parte delle cellule B è fondamentale per il mantenimento omeostatico immunologico umorale. Gli anticorpi rappresentano una classe specifica di immunoglobuline. Esperimenti su animali hanno dimostrato che l’aggiunta di cibi ricchi di carotenoidi alle diete dei conigli può aumentare i livelli sierici di IgG, IgM e IgA, migliorando così l’immunità umorale.[119] Ulteriori studi sui ratti hanno rivelato l’associazione tra una scarsità di VitA nella dieta e un aumento del numero di DC, oltre all’espressione significativamente sovraregolata di IL-12, recettore Toll-like 2 e fattore di differenziazione mieloide MyD88 nella mucosa intestinale. Quando i livelli di IgA secretoria diminuiscono, i ratti mostrano una ridotta funzione immunitaria, suggerendo che la VitA è coinvolta nella sintesi delle immunoglobuline e ha un’influenza importante sull’immunità umorale.[120] Un rapporto mostra che la RA è potentemente sinergizzata con i tessuti linfoidi associati all’intestino IL-6 o IL-5 derivati dalla DC per indurre la secrezione di IgA.[121] Uno studio knockout ha dimostrato che l’ablazione di RARα riduce l’espressione di IgA da parte delle cellule B espresse in vivo e in vitro. Ciò indica che RA agisce sulle cellule B direttamente attraverso RARα, che influenza la sintesi e la secrezione di IgA.[122] È anche probabile che l’AR colpisca prima le Treg e quindi moduli indirettamente le cellule B, poiché le Treg hanno un ruolo importante nella regolazione delle risposte delle cellule B.[123]
La stimolazione dell’antigene delle cellule immunitarie attraverso specifici anticorpi IgE provoca una risposta rapida e specifica all’ipersensibilità che è coinvolta nella maggior parte delle condizioni autoimmuni.[124] Le prove mostrano che RA ha un’attività repressiva di IgE in vivo. L’effetto inibitorio di ATRA su IgE riduce principalmente la sintesi e la secrezione di IgE attraverso RARα, e questo effetto inibitorio dipende da IL-10 [125,126,127,128]. Un altro rapporto mostra che il 9-cis-RA esogeno nel contesto di una sensibilizzazione allergica modula profondamente una risposta IgE umorale consolidata, con conseguente riduzione delle risposte IgE specifiche e aumento delle risposte IgA specifiche nei topi, indicando che i retinoidi attivanti RXR svolgono un ruolo importante in la regolazione fisiologica delle IgE dovuta alla sintesi endogena di 9-cis-RA. [124] Questi rendono la VitA una terapia molto promettente per il trattamento della malattia da ipersensibilità mediata da IgE.
Le cellule B regolatorie (Breg) sono una classe di sottoinsiemi di cellule B con funzioni immunomodulanti che sono coinvolte nel mantenimento dell’omeostasi immunitaria e svolgono un ruolo regolatorio essenziale in vari processi immunopatologici.[129,130] L’AR può indurre la differenziazione delle cellule B native in Bregs e stimolare la sintesi di Breg e la secrezione di IL-10 attraverso RARα.[131,132,133,134] Secernendo IL-10, i Breg hanno effetti migliorativi sulla colite, l’artrite e il lupus sperimentali.[131,132,133,134]. Il meccanismo con cui la VitA regola l’attività di Bregs e come migliora la sua funzione immunomodulante non è ancora stato compreso. Saranno necessarie ulteriori ricerche per chiarire questa domanda e per determinare se gli effetti della VitA sui Breg sono stabili.
Come risaputo, la tubercolosi, che è una malattia infettiva cronica causata dal batterio Mycobacterium tuberculosis, è un problema di salute globale. Negli ultimi anni, i risultati terapeutici dei farmaci tradizionalmente utilizzati per il trattamento della tubercolosi sono peggiorati a causa dell’aumento della popolazione batterica resistente ai farmaci. Pertanto, sono necessarie diverse strategie di trattamento.
Studi epidemiologici hanno dimostrato che la popolazione sana ha un livello sierico significativamente più alto di VitA rispetto ai pazienti affetti da tubercolosi.[135, 136, 137] Uno studio di coorte longitudinale sulla tubercolosi ha mostrato che la carenza di VitA è correlata dose-dipendente al verificarsi della tubercolosi.[138] Uno studio in vitro ha dimostrato che l’AR inibisce la crescita di M. tuberculosis e ne riduce il tasso di sopravvivenza quando viene assorbito dai macrofagi.[139] Per il meccanismo di attività batteriocidica della VitA, Wheelwright et al. hanno scoperto che la VitA può indurre l’espressione di NPC2. Nelle cellule knockout del gene NPC2, la stimolazione della VitA non ha mostrato attività batteriocidica sulle cellule infette. Tuttavia, il gene NPC2 è comunemente noto come regolatore del trasporto del colesterolo piuttosto che come fattore regolatorio immunologico. Questo risultato può essere spiegato come segue: il colesterolo è la fonte nutrizionale delle pareti cellulari batteriche della tubercolosi, mentre l’NPC2 facilita il trasporto del colesterolo dai lisosomi, privando quindi i batteri della tubercolosi delle loro esigenze nutrizionali. Senza la capacità di M. tuberculosis di generare pareti cellulari protettive, il lisozima può quindi uccidere efficacemente questo patogeno.[140] Ciò è stato dimostrato in un modello murino di tubercolosi in cui l’aggiunta di ATRA ha migliorato significativamente l’efficacia dei farmaci antitubercolosi tradizionali.[141] Tuttavia, saranno necessarie ulteriori ricerche per chiarire gli effetti positivi degli integratori di VitA sul trattamento della tubercolosi farmaco resistente.
I pazienti con AIDS sono soggetti noti, in generale, per essere carenti di molte vitamine. [142] Poiché varie vitamine hanno il potenziale per migliorare l’immunità dell’organismo e poiché l’AIDS deriva dall’infezione da virus dell’immunodeficienza umana (HIV), si ritiene che lo stress ossidativo abbia un effetto importante sul processo di infezione del virus HIV.[143,144] VitA, VitC e VitE sono antiossidanti naturali e, inibendo lo stress ossidativo dell’organismo, si ipotizza che queste vitamine possano migliorare la progressione dell’AIDS.
Uno studio precedente ha dimostrato che l’infezione da HIV riduce la regolazione di uno stress ossidativo da parte dell’organismo. Tuttavia, un antiossidante esterno, come la VitA, non ha alcun effetto compensativo sulla regolazione della risposta allo stress ossidativo.[145] Inoltre, sebbene le persone con infezione da HIV siano carenti di molte diverse vitamine, l’integrazione di vitamine non ha mostrato benefici clinicamente importanti nei soggetti con HIV.[146] Coerentemente, la VitA non influenza la trasmissione verticale dell’HIV da madre a figlio.[147] Pertanto, la supplementazione di VitA non sembra influenzare l’HIV in sé, ma ciò non significa che i pazienti o i portatori di HIV dovrebbero rifiutare la supplementazione di VitA o di altre vitamine. L’HIV riduce la funzione immunitaria del corpo, rendendo i pazienti sensibili alle malattie infettive, tra cui la tubercolosi, la malaria, l’herpes e altri.[148,149] Come accennato in precedenza, la VitA migliora l’immunità degli organismi ed è stato riportato che riduce l’incidenza della tubercolosi nei pazienti con HIV.[148] Inoltre, l’integrazione in gravidanza e postpartum con un multivitaminico ha migliorato significativamente lo stato ematologico tra le donne con infezione da HIV e i loro bambini e ha ridotto il rischio di anemia.[149] La terapia antiretrovirale è il regime di trattamento più efficace per l’HIV; tuttavia, la sola terapia antiretrovirale non è, ovviamente, sufficiente per migliorare la carenza di micronutrienti. Pertanto, è essenziale integrare VitA, e altre vitamine e micronutrienti durante il trattamento per l’HIV.[150]
L’assunzione giornaliera raccomandata di VitA per i bambini è di 1665UI.[151] La vitamina A, come retinolo, se supera le 20.000UI/ die anche per brevi periodi, può causare intossicazione e, occasionalmente, morte. L’intossicazione da VitA è una sindrome generalizzata, i cui segni e sintomi includono dermatite desquamativa ed edematosa, dolore e dolorabilità ossea, edema delle estremità e del viso, irritabilità, disfunzione epatocellulare e ipercalcemia.[152,153,154,155] Inoltre, l’infiammazione influisce sul metabolismo dei retinoidi. Il retinolo sierico può essere sequestrato nei tessuti, portando a una riduzione dei livelli sierici di retinolo, il che implica che la valutazione dello stato della VitA con l’uso del retinolo sierico durante l’infiammazione può essere problematica.[157]
Mentre l’approccio interdisciplinare continua a svilupparsi nella ricerca, le persone hanno prestato sempre maggiore attenzione al rapporto tra alimentazione e immunità. Inoltre, l’influenza dei micronutrienti sulla funzione immunitaria dell’organismo è stata ampiamente studiata. La VitA ha ruoli sia di promozione che regolatori nel sistema immunitario innato e nell’immunità adattativa; pertanto, può migliorare la funzione immunitaria dell’organismo e fornire una difesa avanzata contro più malattie infettive. Attualmente, l’effetto della VitA sulla funzione immunitaria è stato studiato a livello molecolare e sono in corso ulteriori ricerche sugli effetti terapeutici della VitA sulla prevenzione e la cura di varie malattie infettive. Man mano che crescono prove nel tempo, la VitA svolgerà probabilmente ruoli più critici nella moderna terapia. Ma il livello del dosaggio efficace, richiesto per una buona risposta immunitaria, non necessita di mega-dosi e può benissimo limitarsi a quello raccomandato in una dieta varia ed equilibrata. E, cosa molto importante, l’integrazione con VitA non garantirà alcuna immunità tantomeno nei confronti del nuovo patogeno COVID-19.
Ma le “pitecate” non finiscono qui…
Assumere Glutammina.
La glutammina è nota per essere il principale substrato energetico utilizzato dalle cellule immunitarie chiamate leucociti e contribuisce alla proliferazione di queste cellule,
[158] la ragione per cui la glutammina è il substrato del carburante per i leucociti è la necessità di una fonte di energia più rapida del glucosio (simile alla mucosa intestinale e al midollo osseo).
[158] I leucociti non possono sintetizzare la glutammina da soli e quindi dipendono dalla glutammina fornita da altri tessuti che possiedono l’enzima glutammina sintetasi o dall’assunzione con la dieta. I tassi di crescita dei leucociti sono più alti con una concentrazione di circa 600 umol / L, una concentrazione ben all’interno della normale fisiologia umana.
[159] Per questo motivo la glutammina e il suo uso supplementare tendono ad essere praticamente limitati ai tempi in cui la sintesi o l’assunzione viene soppressa o reindirizzata, come una malattia critica o un prolungato esercizio cardiovascolare.
[160]
In uno studio su bambini affetti da disturbi gastrointestinali con eppisodi diarroici (preesistenti al trattamento) sono stati trattati giornalmente con 0,3g di Glutammina per ogni kg di peso corporeo.[161] I risultati dello studio hanno evidenziato che l’integrazione orale di Glutammina non ha avuto alcun effetto sulla conta dei Leucociti o dei Linfociti nel sangue dei soggetti trattati.
In conclusione, l’integrazione con Glutammina non ha avuto influenza significativa sull’immunità in sé.
Assumere Zinco
L’IL-2 è noto per subire riduzioni in presenza di una carenza di Zinco [161] e di essere ripristinato a livelli ottimali con un assunzione adeguata del minerale in questione.[162] La produzione ex vivo di IL-2 (valutata mediante induzione dell’mRNA di IL-2 da cellule immunitarie stimolate) è aumentata con l’integrazione di Zinco a 45mg/die per un anno, nonostante le concentrazioni basali di IL-2 non siano particolarmente influenzate.[163]
È noto che la carenza di Zinco porta ad un ridotto numero di cellule T e successivamente a un’immunità umorale e mediata dalle cellule depressa.[164]
Una meta-analisi di 15 studi, tra cui sono state prese in esame 1360 persone, ha evidenziato che lo Zinco, sotto forma di losanghe (gluconato) o sciroppo (solfato), è stato associato con una durata e gravità inferiori del raffreddore comune quando assunto entro 24 ore dall’esordio di quest’ultimo che, dopo una settimana, aveva un rapporto di intensità e presenza, nonché di ricaduta, pari ad una ratio di 0,45 (meno della metà del rischio). [165]
L’integrazione di 45mg di Zinco nelle persone anziane (con concentrazioni di Zinco nel siero inferiori rispetto ai giovani di controllo) per la durata di un anno è associata a un tasso ridotto di infezioni del tratto respiratorio superiore (riduzione del 50% che non è stata significativa) e infezioni generali (occorrenza dell’88% ridotta al 29%).[163] L’integrazione prolungata di Zinco nell’arco di cinque mesi è associata a una ridotta incidenza di malattia (RR 0,64) sebbene con un’alta variabilità.[165]
Una dose elevata di Zinco, in soggetti deficitari, in risposta al raffreddore comune (non assunta preventivamente, ma solo all’inizio dell’infezione) sembra essere efficace nel ridurre la durata e la gravità della malattia.
Quindi, l’integrazione giornaliera di Zinco come preventivo, con diverse variabili dettate per lo più dalle concentrazioni ematiche di base del minerale, sembra ridurre il tasso di infezione da Rinovirius (virus del raffreddore). Soggetti con una buona assunzione di Zinco non mostrano significativi miglioramenti nella percentuale di infezione virale, ben che meno in riferimento al COVID-19.
Bere ogni 15 minuti
Alcuni hanno affermato che dovremmo garantire che la nostra bocca e la nostra gola rimangano umide e non si secchino mai.
Alcuni individui suggeriscono di bere un sorso d’acqua ogni 15 minuti al fine di trasportare il virus dalla cavità orale allo stomaco dove (secondo la loro fantasiosa teoria) il virus verrà eliminato dall’acido gastrico. Ovviamente l’acqua non impedirà di eliminatre il COVID-19.
Il dott. William Schaffner, un esperto di malattie infettive alla Vanderbilt University, ha dichiarato: “Mettiamo sempre in guardia chiunque, che sia sano o malato, di mantenere l’assunzione di liquidi e mantenere umide le mucose.
Ti fa sentire sicuramente meglio; ma non vi è alcuna chiara indicazione che ciò possa proteggere direttamente da complicazioni legate al contatto con il patogeno.
Radersi la barba
Un grafico dei Centers for Disease Control and Prevention che raccomandava agli uomini di radersi la barba per proteggersi dal COVID-19 è risultato essere una manipolazione fatta ad arte utilizzando frammenti di grafici emessi dall’organo di controllo sopra riportato.
Esso, oltretutto, risale al 2017, prima dello scoppio dell’epidemia di COVID-19, ed era destinato a professionisti che indossano respiratori per la protezione durante lo svolgimento delle loro mansioni.
Da allora il CDC ha affermato di non raccomandare l’uso di routine di respiratori o maschere per il viso al di fuori delle impostazioni del luogo di lavoro.
Viene detto che il personale di sesso maschile del SSN al Southampton General Hospital è stato recentemente invitato a radersi per aiutare a limitare la diffusione del virus.
Questo perché, secondo il responsabile della salute e della sicurezza, una barba lunga “rende impossibile ottenere una buona aderenza della maschera sul viso”.
Da qui la presunta correlazione tra infezione da COVID-19 e barba lunga.
Quindi, la rimozione della barba non impedisce la diffusione del virus.
Gli asciugamani riscaldati uccidono il COVID-19
Sono circolate voci secondo cui l’uso di asciugamani riscaldati da aria calda per 30 secondi potrebbe causare la neutralizzazione del virus presente nelle mani.
Ma l’OMS afferma che questo non è vero e non rappresenta un mezzo per ridurre la contaminazione virale.
Infatti, le raccomandazioni sono ben chiare: “per proteggersi dal nuovo coronavirus, è necessario lavarsi spesso le mani con un gel a base alcolica o con acqua e sapone”.
“Una volta pulite le mani, asciugarle accuratamente utilizzando asciugamani di carta o un asciugatore ad aria calda.”
Il collutorio
Il collutorio non può proteggerti dalle infezioni virali.
Non ci sono prove che l’uso del collutorio possa proteggere dall’infezione da COVID-19 .
Alcuni collutori possono eliminare alcuni microbi per alcuni minuti nella saliva. Tuttavia, ciò non significa che protegga dall’infezione del nuovo patogeno.
Mangiare aglio previene l’infezione
Sebbene il regolare consumo di buone quantità d’aglio possa contribuire ad abbassare la pressione, non impedirà al consumatore di essere infettato dal COVID-19.
“L’aglio è un alimento sano che può avere alcune proprietà antimicrobiche”, afferma l’OMS.
“Tuttavia, non ci sono prove nell’attuale epidemia che il consumo di aglio abbia protetto le persone dal contagio del nuovo coronavirus.”
I giovani non contraggono il COVID-19
Ormai lo sappiamo fin troppo bene, anche i giovani sono a rischio di contagio del COVID-19, nonostante i modelli statistici mostrino che gli anziani hanno un tasso di contagio più elevato.
“Le persone di tutte le età possono essere infettate dal nuovo coronavirus (2019-nCoV)”, afferma l’OMS.
“Le persone anziane e le persone con condizioni mediche preesistenti (come l’asma, il diabete, le malattie cardiache) sembrano essere più vulnerabili a diventare gravemente malati con il nuovo virus.
L’OMS consiglia alle persone di tutte le età di prendere provvedimenti per proteggersi dal virus, ad esempio seguendo una buona igiene delle mani e una buona igiene respiratoria.
Lampade UV e COVID-19
L’OMS ha esortato le persone a non usare le lampade UV per sterilizzare le mani o altre aree della pelle nel tentativo di eliminare il COVID-19.
L’organizzazione ha dichiarato: “Le lampade UV non devono essere utilizzate per sterilizzare le mani o altre aree della pelle poiché le radiazioni UV possono causare irritazione alla pelle”.
Spargersi l’alcool denaturato sul corpo e/o ingerirlo elimina il COVID-19
C’è chi consiglia di cospargersi il corpo di alcol o cloro per eliminare il virus quando questo è già presente nell’organismo.
In realtà, cospargersi, o peggio ingerire, tali sostanze può essere molto dannoso e non solo per i vestiti ma anche per le mucose (cioè occhi, bocca).
Bisogna essere consapevoli del fatto che sia l’alcol che il cloro possono essere utili per disinfettare le superfici, ma devono essere utilizzati secondo le raccomandazioni appropriate e non per usi impropri (come quello consigliato in seguito all’isteria da COVID-19).
Lettere e pacchi dalla Cina non sono sicuri
Contrariamente alle ipotesi, è effettivamente sicuro ricevere pacchi dalla Cina.
“Le persone che ricevono pacchi dalla Cina non sono a rischio di contrarre il nuovo coronavirus”, afferma l’OMS.
“Da precedenti analisi, sappiamo che i coronavirus non sopravvivono a lungo su oggetti, come lettere o pacchi.”
I vaccini contro la polmonite debellano il COVID-19
Secondo l’OMS, è falso affermare che i vaccini contro la polmonite possano proteggere dal COVID-19.
I vaccini per il COVID-19 sono ancora in fase di sperimentazione su animali e difficilmente saranno pronti in tempo per frenare l’attuale epidemia.
“I vaccini contro la polmonite, come il vaccino pneumococcico e il vaccino contro l’influenza Haemophilus di tipo B (Hib), non forniscono protezione contro il nuovo coronavirus”, afferma l’OMS.
“Il virus è così nuovo e diverso che ha bisogno del proprio vaccino. I ricercatori stanno tentando di sviluppare un vaccino contro il 2019-nCoV e l’OMS sta supportando i loro sforzi”.
“Sebbene questi vaccini non siano efficaci contro il 2019-nCoV, la vaccinazione contro le malattie respiratorie è altamente raccomandata per proteggere la salute.”
Il risciacquo delle cavità nasali con una soluzione salina previene l’infezione
Non ci sono prove che il risciacquo regolare del naso con soluzione salina abbia protetto le persone dall’infezione del nuovo coronavirus, afferma l’OMS.
“Esistono prove limitate che il risciacquo regolare del naso con soluzione salina possa aiutare le persone a riprendersi più rapidamente dal comune raffreddore”, aggiunge l’organizzazione.
“Tuttavia, il risciacquo regolare del naso non ha dimostrato di prevenire le infezioni respiratorie.”
L’olio di sesamo neutralizza il virus
L’olio di sesamo è un alimento base nella cucina asiatica, ma è tutto ciò per cui è buono usarlo.
“L’olio di sesamo non uccide il nuovo coronavirus”, avverte l’OMS.
“Esistono alcuni disinfettanti chimici che possono uccidere il 2019-nCoV sulle superfici.
“Questi includono disinfettanti a base di candeggina/cloro, solventi, etanolo al 75%, acido peracetico e cloroformio”.
“Tuttavia, hanno un impatto minimo o nullo sul virus se applicati sulla pelle o sotto il naso. Può persino essere pericoloso applicare questi prodotti chimici sulla pelle.”
Gli antibiotici contro il COVID-19
Il COVID-19 è un virus e, pertanto, gli antibiotici non devono essere usati come mezzo di prevenzione o trattamento per il semplice fatto che NON FUNZIONANO SUI VIRUS MA SOLO SUI BATTERI!
Quali comportamenti utili adottare?
Lavarsi spesso ed accuratamente le mani (per minimo 20 secondi). Si raccomanda di mettere a disposizione in tutti i locali pubblici, palestre, supermercati, farmacie e altri luoghi di aggregazione, soluzioni idroalcoliche per il lavaggio delle mani. Per i privati cittadini che devono spostarsi per motivi lavorativi e/o di approvvigionamento alimentare e/o di prodotti di necessità è consigliato l’uso di un disinfettante a base alcolica da potersi realizzare in casa secondo le indicazioni dell’OMS di seguito riportate.
Per 1lt:
– 833ml di Alcol Etilico al 96%;
– 42ml di Acqua Ossigenata al 3%;
-15ml di Glicerina (Glicerolo) al 98%;
-Acqua distillata oppure bollita è raffreddata quanto basta per arrivare ad 1lt;
-Mettere in una o più bottigliette.
Evitare il contatto ravvicinato con persone che soffrono di infezioni respiratorie acute.
Evitare abbracci, strette di mano, creare aggregazione o aggregarsi ad altre persone .
Mantenimento, nei contatti sociali, di una distanza interpersonale di almeno un metro.
Igiene respiratoria (starnutire e/o tossire in un fazzoletto evitando il contatto delle mani con le secrezioni respiratorie).
Evitare l’uso promiscuo di bottiglie e bicchieri, in particolare durante l’attività sportiva.
Non toccarsi occhi, naso e bocca con le mani.
Coprirsi bocca e naso se si starnutisce o tossisce senza usare la mano ma il braccio all’altezza del gomito.
Non assumere farmaci antivirali autoprescritti.
Pulire le superfici con disinfettanti a base di cloro o alcol.
Usare la mascherina solo se si sospetta di essere malati o se si presta assistenza a persone malate. La mascherina non è un mezzo preventivo.
Ma quanto resta sulle superfici e in sospensione nell’aria il COVID-19?
4 ore sul rame;
fino a 24 ore sul cartone;
fino a 72 ore su plastica (polipropilene) e acciaio inox;
in sospensione nell’aria fino a 3 ore. [166]
Ma il Tocilizumab?
E’ notizia recente che il trattamento di alcuni pazienti affetti da COVID-19 con Tocilizumab, un farmaco comunemente utilizzato per il trattamento dell’artrite reumatoide, abbia avuto esito positivo nel arrestare la cascata di eventi infiammatori derivanti dal patogeno. L’Aifa ha approvato il protocollo della sperimentazione scientifica del Tolicizumab, e Napoli e Modena sono i capofila. Sono partiti con priorità immediata su 250 pazienti in Italia.
Al momento della stesura di questo articolo sono in totale 11 i pazienti trattati a Napoli con il farmaco off label a seguito di una sperimentazione nata dalla collaborazione tra il direttore della Uoc di oncologia dell’azienda ospedaliera dei Colli, Vincenzo Montesarchio, e il direttore dell’unità di oncologia melanoma, immunoterapia oncologica e terapie innovative dell’istituto Pascale di Napoli, Paolo Ascierto.
E’ stata rilasciata la notizia che quattro dei pazienti in rianimazione stanno meglio, e per il primo paziente si attendono le valutazioni del rianimatore prima che si possa procedere ad estubarlo. Per tre dei quattro pazienti a cui è stato somministrato il Tocilizumab a partire da sabato c’è stata un’ottima risposta. Non erano in rianimazione ma erano ricoverati in condizioni severe. Risultati incoraggianti arrivano anche dall’ospedale di Cosenza, dove due pazienti hanno risposto molto bene al trattamento. In Puglia, in alcuni pazienti selezionati, contagiati dal COVID-19, è già iniziata la somministrazione del farmaco Tocilizumab.
Il farmaco utilizzato per la cura dell’artrite reumatoide e altre malattie croniche autoimmuni, il Tocilizumab, si sta dimostrando efficace nel trattamento della polmonite interstiziale causata dal Covid-19, anche su altri pazienti curati in Italia e non solo sui primi casi trattati a Napoli. Nella maggior parte dei casi si registra un rapido miglioramento tanto che l’azienda produttrice, la Roche, in pochissimo tempo ha già dispensato 600 trattamenti ai vari ospedali che lo chiedevano.
L’idea è nata perché l’immunoterapia viene usata nei tumori e alcuni effetti collaterali vengono trattati con il Tocilizumab. Il meccanismo che sta alla base del distress del Covid -19 è molto simile a quello dei trattamenti oncologici per questo motivo si è incominciato ad usarlo sui pazienti infetti dal nuovo patogeno. Il confronto con i medici cinesi è stato essenziale dal momento che essi avevano già usato il farmaco in 21 pazienti e in 20 si era registrato un miglioramento importante.
Il farmaco agisce sulla complicanza di questo virus cioè sull’infiammazione importante che il Covid-19 crea. Insomma diminuisce l’iperattività del sistema immunitario che è quella che causa l’insufficienza respiratoria. I medici in Cina hanno riportato che in due settimane dal trattamento i pazienti trattati sono stati dimessi e sono tornati a casa. Chiaramente segue per loro un periodo di convalescenza nel proprio domicilio. Ma è comunque un buon risultato. Ovviamente, l’isolamento contenitivo è la prima misura importante, l’unica che in questo momento può in qualche modo ridurre il numero dei contagiati. Ed è quello che hanno applicato severamente in Cina e che ha dato i suoi risultati positivi.
Prima che corriate per strada a stappare lo champagne fatto col bicarbonato che avete avanzato da capodanno, sappiate che stiamo comunque parlando di un farmaco sperimentale per questo trattamento e che non previene l’infezione da COVID-19 ne debella il patogeno già presente nell’organismo ospite. E’ stato testato solamente sugli ammalati molto gravi per arginare la risposta infiammatoria massiva causa delle complicazioni respiratorie. Si tratta quindi di un promettente farmaco per il trattamento dei sintomi che però, a mio avviso, si dovrà ancora attendere qualche tempo prima di poterlo considerare a tutti gli effetti, a 360°, un efficiente mezzo per il trattamento e cura (coadiuvato da antiretrovirali appositi) per i pazienti affetti da COVID-19.
Il costo del farmaco varia da un minimo di 211,77€ per il ROACTEMRA INF FL 4ML 20MG/ML a un massimo indicativo di 1.481,61€ per il ROACTEMRA SC 4SIR 162MG 0,9ML. Ovviamente non è disponibile alcuna formulazione generica dal momento che il brevetto in possesso della Roche non è scaduto.
Ma l’SR9009, abbassando l’infiammazione riducendo le IL-6 del 72%, non potrebbe avere un potenziale applicativo nel trattamento dei sintomi da contagio del COVID-19?
SR9009
Una risposta affrettata sarebbe senza ombra di dubbio “si”, ma non sono solito dare giudizi prematuri so molecole sperimentali prive di sufficiente letteratura. Sebbene la molecola in questione, che giova ricordare è un agonista dei Rev-ErbA (agisce aumentando la repressione dei geni regolati da parte dei Rev-ErbA), si sia dimostrata promettente in un recente studio su topi [167] nel quale si osservavano i suoi effetti in seguito ad infarto (dimostrando l’assenza di tessuto cicatriziale, di danno cardiaco e insufficienza cardiaca), per via della sua azione modulatoria sul inflammosoma Nlrp3, un complesso proteico che scatena una risposta infiammatoria esagerata e patologica, la sua sperimentazione non sembra volta verso ulteriori applicazioni.
Dal momento che si tratta di una frontiera applicativa molto delicata, si sconsiglia caldamente l’auto sperimentazione!
Cosa concludere?
Beh, senz’altro, dopo questa approfondita disamina, se dotati di un minimo di capacità cognitiva, dovreste essere in grado di riconoscere l’inutilità di base nell’assunzione di dosi elevate di vitamine al fine di sviluppare una presunta resistenza immunitaria tale da rendervi immuni dal COVID-19 o altro patogeno altamente contagioso e potenzialmente letale. Sono consapevole del fatto che diverse persone, fin troppe, continueranno comunque a voler credere ciecamente a delle “belle” menzogne piuttosto che concretizzare il problema e adoperarsi a seguire linee guida per l’igiene pubblica confermate da valida, e ricca, letteratura scientifica. Nessuno mette in dubbio l’importanza di una corretta alimentazione, come più volte sottolineato, per garantire una capacità sistemica/immunitaria ottimale al fine di affrontare con una percentuale maggiore di esito positivo eventuali infezioni virali. Ma da qui ad affermare che mega dosi di Vitamina C, D ed A rendano il soggetto trattato praticamente immune dagli agenti patogeni beh, non solo rappresenta una distorsione dei dati scientifici reali a nostra disposizione ma comporta un vero e proprio atto criminale a danno della collettività!
Lasciate perdere gli improvvisati virologi e tutti coloro che, millantando rivelazioni tenute nascoste dai “gomblotti”, si definiscono “divulgatori scientifici” dimostrando soltanto una criminosa fantasia generata dal “markettismo” e da un modus relativista di interpretare dati scientifici sufficientemente chiari e, spesso, rilegati ai gradini più bassi della piramide delle evidenze scientifiche.
STATE A CASA E RIDUCETE LE USCITE, NON CHE LA VICINANZA CON GLI ESTRANEI, AL MINIMO INDISPENSABILE! SEBBENE SIANO GLI ANZIANI (MEDIA 80.3 ANNI) A RISCHIARE MAGGIORMENTE UN ESITO NEGATIVO DALL’INFEZIONE DA COVID-19, QUESTO NON AUTORIZZA NESSUNO A DIVENIRE UNA “BOMBA VIRALE”, UN “UNTORE”, CAPACE DI METTERE A RISCHIO LA FASCIA PIU’ DEBOLE DELLA POPOLAZIONE (COMPRESI PAZIENTI CON PATOLOGIE GIA’ PRESISTENTI E DI DIVERSE FASCE D’ETA’)!
Nota: ho evitato in tale sede di riportare le MIE conclusioni, maturate in seguito a giorni e notti di studio incessante, sulle possibilità di contagio e i modelli matematici che mostrano la media del numero di infetti una volta raggiunto il picco. La mia decisione è dettata 1) dal desiderio di non causare ulteriore panico e 2) dal fatto che sono molto cauto prima di esporre una teoria che potrebbe condizionare fortemente il comportamento della collettività.
Gabriel Bellizzi
Riferimenti:
1- World Health Organization (WHO).
2- Erika Edwards, How does coronavirus spread?.
3- WHO COVID-19 situation report 29, su World Health Organization, 19 febbraio 2020.
4- Q&A on coronaviruses (COVID-19): How long is the incubation period for COVID-19?
5- Australian Government Department of Health, Novel coronavirus (2019-nCoV).
6- Novel coronavirus named ‘Covid-19’: WHO.
7- Homologous recombination within the spike glycoprotein of the newly identified coronavirus may boost cross‐species transmission from snake to human.
8- Ewen Callaway e David Cyranoski, Why snakes probably aren’t spreading the new China virus, in Nature, 23 gennaio 2020.
9- Megan Multeni, No, the Wuhan Virus Is Not a ‘Snake Flu’.
10- Zheng-Li Shi, Peng Zhou, Xing-Lou Yang, Xian-Guang Wang, Ben Hu, Lei Zhang, Wei Zhang, Hao-Rui Si, Yan Zhu, Bei Li e Chao-Lin Huang, Discovery of a novel coronavirus associated with the recent pneumonia outbreak in humans and its potential bat origin, in bioRxiv.
11- Wei Ji, Wei Wang, Xiaofang Zhao, Junjie Zai e Xingguang Li, Homologous recombination within the spike glycoprotein of the newly identified coronavirus may boost cross‐species transmission from snake to human, in Journal of Medical Virology.
12- Coronavirus, su who.int.
13- Ma, W., Kahn, R.E., and Richt, J.A., The pig as a mixing vessel for influenza viruses: human and veterinary implications, J. Mol. Genet. Med. 3(1):158–166, 2008.
14- A coronavirus (similar to the new coronavirus), the cause of Middle East respiratory syndrome.
15- Carter, R.W., and Sanford, J.C., A new look at an old virus: mutation accumulation in the human H1N1 influenza virus since 1918, Theoretical Biology and Medical Modelling 9:42, 2012.
26- Vitamin D supplementation did not prevent influenza-like illness as diagnosed retrospectively by questionnaires in subjects participating in randomized clinical trials.
Jorde R, Witham M, Janssens W, Rolighed L, Borchhardt K, de Boer IH, Grimnes G, Hutchinson MS. Scand J Infect Dis. 2012 Feb; 44(2):126-32.
27- Jakóbisiak M. Immunologia. 2nd ed. Wydawnictwo Naukowe PWN; Warsaw, Poland: 1995. Główne komponenty i zasadnicze cechy odpowiedzi immunologicznej; pp. 28–36.
28- nfluenza, solar radiation and vitamin D. Moan J, Dahlback A, Ma L, Juzeniene A
Dermatoendocrinol. 2009 Nov; 1(6):307-9.
29- Vitamin D supplementation for the prevention of acute respiratory tract infection: a randomized, double-blinded trial among young Finnish men. Laaksi I, Ruohola JP, Mattila V, Auvinen A, Ylikomi T, Pihlajamäki H. J Infect Dis. 2010 Sep 1; 202(5):809-14.
30- Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Urashima M, Segawa T, Okazaki M, Kurihara M, Wada Y, Ida H. Am J Clin Nutr. 2010 May; 91(5):1255-60.
31- Vitamin D status has a linear association with seasonal infections and lung function in British adults. Berry DJ, Hesketh K, Power C, Hyppönen E. Br J Nutr. 2011 Nov; 106(9):1433-40.
32- Serum 25-hydroxyvitamin D3 and the risk of pneumonia in an ageing general population. Aregbesola A, Voutilainen S, Nurmi T, Virtanen JK, Ronkainen K, Tuomainen TP. J Epidemiol Community Health. 2013 Jun; 67(6):533-6.
33- Retinol binding protein and vitamin D associations with serum antibody isotypes, serum influenza virus-specific neutralizing activities and airway cytokine profiles.
Jones BG, Oshansky CM, Bajracharya R, Tang L, Sun Y, Wong SS, Webby R, Thomas PG, Hurwitz JL. Clin Exp Immunol. 2016 Feb; 183(2):239-47.
34- Association between serum concentration of 25-hydroxyvitamin D and community-acquired pneumonia: a case-control study. Mamani M, Muceli N, Ghasemi Basir HR, Vasheghani M, Poorolajal J. Int J Gen Med. 2017; 10():423-429.
35- Serum 25-hydroxyvitamin D levels in hospitalized adults with community-acquired pneumonia. Brance ML, Miljevic JN, Tizziani R, Taberna ME, Grossi GP, Toni P, Valentini E, Trepat A, Zaccardi J, Moro J, Finuci Curi B, Tamagnone N, Ramirez M, Severini J, Chiarotti P, Consiglio F, Piñeski R, Ghelfi A, Kilstein J, Street E, Moretti D, Oliveto V, Mariño M, Manera J, Brun LR Clin Respir J. 2018 Jul; 12(7):2220-2227.
36- Association of serum 25-hydroxyvitamin D with influenza in case-control study nested in a cohort of Japanese employees. Nanri A, Nakamoto K, Sakamoto N, Imai T, Akter S, Nonaka D, Mizoue T. Clin Nutr. 2017 Oct; 36(5):1288-1293.
37- Effects of calcitriol (1, 25-dihydroxy-vitamin D3) on the inflammatory response induced by H9N2 influenza virus infection in human lung A549 epithelial cells and in mice. Gui B, Chen Q, Hu C, Zhu C, He G. Virol J. 2017 Jan 23; 14(1):10.
38- The possible roles of solar ultraviolet-B radiation and vitamin D in reducing case-fatality rates from the 1918-1919 influenza pandemic in the United States. Grant WB, Giovannucci E. Dermatoendocrinol. 2009 Jul; 1(4):215-9.
39- Calcitriol [1, 25[OH]2 D3] pre- and post-treatment suppresses inflammatory response to influenza A (H1N1) infection in human lung A549 epithelial cells. Khare D, Godbole NM, Pawar SD, Mohan V, Pandey G, Gupta S, Kumar D, Dhole TN, Godbole MM. Eur J Nutr. 2013 Jun; 52(4):1405-15.
40- Effects of vitamin D supplements on influenza A illness during the 2009 H1N1 pandemic: a randomized controlled trial. Urashima M, Mezawa H, Noya M, Camargo CA Jr. Food Funct. 2014 Sep; 5(9):2365-70.
41- A randomized controlled trial of vitamin D3 supplementation for the prevention of symptomatic upper respiratory tract infections. Li-Ng M, Aloia JF, Pollack S, Cunha BA, Mikhail M, Yeh J, Berbari N. Epidemiol Infect. 2009 Oct; 137(10):1396-404.
42- Lorenc R.S., Karczmarewicz E., Kryśkiewicz E., Płudowski P. Vitamin D provision and supplementation standards. Stand. Med. Pediatr. 2012;9:595–604.
43- Re: epidemic influenza and vitamin D. Aloia JF, Li-Ng M. Epidemiol Infect. 2007 Oct; 135(7):1095-6; author reply 1097-8.
44- Effect of Vitamin D and Calcium Supplementation on Cancer Incidence in Older Women: A Randomized Clinical Trial. Lappe J, Watson P, Travers-Gustafson D, Recker R, Garland C, Gorham E, Baggerly K, McDonnell SL. JAMA. 2017 Mar 28; 317(12):1234-1243.
45- Vitamin D levels and influenza vaccine immunogenicity among HIV-infected and HIV-uninfected adults. Crum-Cianflone NF, Won S, Lee R, Lalani T, Ganesan A, Burgess T, Agan BK. Vaccine. 2016 Sep 22; 34(41):5040-5046.
46- Are children’s vitamin D levels and BMI associated with antibody titers produced in response to 2014-2015 influenza vaccine? Lin CJ, Martin JM, Cole KS, Zimmerman RK, Susick M, Moehling KK, Levine MZ, Spencer S, Flannery B, Nowalk MP. Hum Vaccin Immunother. 2017 Jul 3; 13(7):1661-1665.
47- Principi N., Marchisio P., Terranova L., Zampiero A., Baggi E., Daleno C., Tirelli S., Pelucchi C., Esposito S. Impact of vitamin D administration on immunogenicity of trivalent inactivated influenza vaccine in previously unvaccinated children. Hum. Vaccines Immunother. 2013;9:969–974. doi: 10.4161/hv.23540.
48- Lee R.U., Won S.H., Hansen C., Crum-Cianflone N.F. 25-hydroxyvitamin D, influenza vaccine response and healthcare encounters among a young adult population. PLoS ONE. 2018;13:e0192479. doi: 10.1371/journal.pone.0192479.
49- Lang P.O., Samaras D. Aging adults and seasonal influenza: Does the vitamin D status (H)arm the body? J. Aging Res. 2011;2012 doi: 10.1155/2012/806198.
50- Lee R.U., Won S.H., Hansen C., Crum-Cianflone N.F. 25-hydroxyvitamin D, influenza vaccine response and healthcare encounters among a young adult population. PLoS ONE. 2018;13:e0192479. doi: 10.1371/journal.pone.0192479.
51- Sundaram M.E., Talbot H.K., Zhu Y., Griffin M.R., Spencer S., Shay D.K., Coleman L.A. Vitamin D is not associated with serologic response to influenza vaccine in adults over 50 years old. Vaccine. 2013;31:2057–2061. doi: 10.1016/j.vaccine.2013.02.028.
52- Chadha M.K., Fakih M., Muindi J., Tian L., Mashtare T., Johnson C.S., Trump D. Effect of 25-hydroxyvitamin D status on serological response to influenza vaccine in prostate cancer patients. Prostate. 2011;71:368–372. doi: 10.1002/pros.21250.
53- Sadarangani S.P., Ovsyannikova I.G., Goergen K., Grill D.E., Poland G.A. Vitamin D, leptin and impact on immune response to seasonal influenza A/H1N1 vaccine in older persons. Hum. Vaccines Immunother. 2016;12:691–698. doi: 10.1080/21645515.2015.1097015.
54- Galant K., Barg E., Kazanowska B. Vitamin D and metabolic, autoimmunologic and neoplasm diseases. Pediatr. Endocrinol. Diabetes Metab. 2016;24:31–37. doi: 10.18544/PEDM-22.01.0048.
55- Surman S.L., Penkert R.R., Jones B.G., Sealy R.E., Hurwitz J.L. Vitamin supplementation at the time of immunization with a cold-adapted influenza virus vaccine corrects poor mucosal antibody responses in mice deficient for vitamins A and D. Clin. Vaccine Immunol. 2016;23 doi: 10.1128/CVI.00739-15.
56- Lee R.U., Won S.H., Hansen C., Crum-Cianflone N.F. 25-hydroxyvitamin D, influenza vaccine response and healthcare encounters among a young adult population. PLoS ONE. 2018;13:e0192479. doi: 10.1371/journal.pone.0192479.
57- Surman S.L., Penkert R.R., Jones B.G., Sealy R.E., Hurwitz J.L. Vitamin supplementation at the time of immunization with a cold-adapted influenza virus vaccine corrects poor mucosal antibody responses in mice deficient for vitamins A and D. Clin. Vaccine Immunol. 2016;23 doi: 10.1128/CVI.00739-15.
58- Lee R.U., Won S.H., Hansen C., Crum-Cianflone N.F. 25-hydroxyvitamin D, influenza vaccine response and healthcare encounters among a young adult population. PLoS ONE. 2018;13:e0192479. doi: 10.1371/journal.pone.0192479.
59- Greiller C.L., Martineau A.R. Modulation of the immune response to respiratory viruses by vitamin D. Nutrients. 2015;7:4240–4270. doi: 10.3390/nu7064240.
60- Wiwanitkit V. Vitamin D and influenza vaccination. Hum. Vaccines Immunother. 2013;9 doi: 10.4161/hv.23847.
61- Grant W.B., Giovannucci E. The possible roles of solar ultraviolet-B radiation and vitamin D in reducing case-fatality rates from the 1918–1919 influenza pandemic in the United States. Dermato-Endocrinol. 2009;1:215–219. doi: 10.4161/derm.1.4.9063.
62- Riabroy N., Tanumihardjo S.A. Oral doses of α-retinyl ester track chylomicron uptake and distribution of vitamin A in a male piglet model for newborn infants. J. Nutr. 2014;144:1188–1195. doi: 10.3945/jn.114.191668.
63- Kiss I., Rühl R., Szegezdi E., Fritzsche B., Toth B., Pongrácz J., Perlmann T., Fésüs L., Szondy Z. Retinoid receptor-activating ligands are produced within the mouse thymus during postnatal development. Eur. J. Immunol. 2008;38:147–155. doi: 10.1002/eji.200737342.
64- Kuwata T., Wang I.M., Tamura T., Ponnamperuma R.M., Levine R., Holmes K.L., Morse H.C., De Luca L.M., Ozato K. Vitamin A deficiency in mice causes a systemic expansion of myeloid cells. Blood. 2000;95:3349.
65- Kiss I., Rühl R., Szegezdi E., Fritzsche B., Toth B., Pongrácz J., Perlmann T., Fésüs L., Szondy Z. Retinoid receptor-activating ligands are produced within the mouse thymus during postnatal development. Eur. J. Immunol. 2008;38:147–155. doi: 10.1002/eji.200737342.
66- van Bennekum A.M., Wong Yen Kong L.R., Gijbels M.J., Tielen F.J., Roholl P.J., Brouwer A., Hendriks H.F. Mitogen response of B cells, but not T cells, is impaired in adult vitamin A-deficient rats. J. Nutr. 1991;121:1960–1968.
67- Darwiche N., Celli G., Sly L., Lancillotti F., De Luca L.M. Retinoid status controls the appearance of reserve cells and keratin expression in mouse cervical epithelium. Cancer Res. 1993;53:2287–2299.
68- Chang H.K., Hou W.S. Retinoic acid modulates interferon-γ production by hepatic natural killer T cells via phosphatase 2A and the extracellular signal-regulated kinase pathway. J. Interferon Cytokine Res. 2015;35:200–212.
69- Wynn T.A., Vannella K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity. 2016;44:450–462.
70- Hiemstra I.H., Beijer M.R., Veninga H., Vrijland K., Borg E.G., Olivier B.J., Mebius R.E., Kraal G., den Haan J.M. The identification and developmental requirements of colonic CD169+ macrophages. Immunology. 2014;142:269. doi: 10.1111/imm.12251. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
71- Pereira W.F., Ribeiro-Gomes F.L., Guillermo L.V., Vellozo N.S., Montalvão F., Dosreis G.A., Lopes M.F. Myeloid-derived suppressor cells help protective immunity to Leishmania major infection despite suppressed T cell responses. J. Leukoc. Biol. 2011;90:1191–1197.
72- Vellozo N.S., Pereiramarques S.T., Cabralpiccin M.P., Filardy A.A., Ribeirogomes F.L., Rigoni T.S., Dosreis G.A., Lopes M.F. All-trans retinoic acid promotes an m1- to m2-phenotype shift and inhibits macrophage-mediated immunity to leishmania major. Front. Immunol. 2017;8:1560.
73- Shrestha S., Kim S.Y., Yun Y.J., Kim J.K., Lee J.M., Shin M., Song D.K., Hong C.W. Retinoic acid induces hypersegmentation and enhances cytotoxicity of neutrophils against cancer cells. Immunol. Lett. 2017;182:24–29.
74- Worbs T., Hammerschmidt S.I., Förster R. Dendritic cell migration in health and disease. Nat. Rev. Immunol. 2017;17:30–48.
75- Beijer M.R., Molenaar R., Goverse G., Mebius R.E., Kraal G., den Haan J.M. A crucial role for retinoic acid in the development of Notch-dependent murine splenic CD8−CD4− and CD4+ dendritic cells. Eur. J. Immunol. 2013;43:1608–1616.
76- Klebanoff C.A., Spencer S.P., Torabi-Parizi P., Grainger J.R., Roychoudhuri R., Ji Y., Sukumar M., Muranski P., Scott C.D., Hall J.A., et al. Retinoic acid controls the homeostasis of pre-cDC-derived splenic and intestinal dendritic cells. J. Exp. Med. 2013;210:1961–1976.
77- Duriancik D.M., Hoag K.A. Vitamin A deficiency alters splenic dendritic cell subsets and increases CD8+Gr-1+ memory T lymphocytes in C57BL/6J mice. Cell Immunol. 2010;265:156–163.
78- Zeng R., Bscheider M., Lahl K., Lee M., Butcher E.C. Generation and transcriptional programming of intestinal dendritic cells: Essential role of retinoic acid. Mucosal. Immunol. 2016;9:183–193.
79- Sun C.M., Hall J.A., Blank R.B., Bouladoux N., Oukka M., Mora J.R., Belkaid Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 Treg cells via retinoic acid. J. Exp. Med. 2007;204:1775–1785.
80- Coombes J.L., Siddiqui K.R., Arancibia-Cárcamo C.V., Hall J., Sun C.M., Belkaid Y., Powrie F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-β– and retinoic acid–dependent mechanism. J. Exp. Med. 2007;204:1757–1764.
81- Scott C.L., Aumeunier A.M., Mowat A.M. Intestinal CD103+, dendritic cells: Master regulators of tolerance? Trends Immunol. 2011;32:412–419.
82- DePaolo R.W., Abadie V., Tang F., Fehlner-Peach H., Hall J.A., Wang W., Marietta E.V., Kasarda D.D., Waldmann T.A., Murray J.A., et al. Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens. Nature. 2011;471:220–224.
83- Mora J.R., von Andrian U.H. T-cell homing specificity and plasticity: New concepts and future challenges. Trends Immunol. 2006;27:235–243.
84- Iwata M., Hirakiyama A., Eshima Y., Kagechika H., Kato C., Song S.Y. Retinoic acid imprints gut-homing specificity on T cells. Immunity. 2004;21:527–538.
85- Svensson M., Johansson-Lindbom B., Zapata F., Jaensson E., Austenaa L.M., Blomhoff R., Agace W.W. Retinoic acid receptor signaling levels and antigen dose regulate gut homing receptor expression on CD8+ T cells. Mucosal. Immunol. 2008;1:38–48.
86- Hosoe N., Miura S., Watanabe C., Tsuzuki Y., Hokari R., Oyama T., Fujiyama Y., Nagata H., Ishii H. Demonstration of functional role of TECK/CCL25 in T lymphocyte-endothelium interaction in inflamed and uninflamed intestinal mucosa. Am. J. Physiol. Gastrointest. Liver Physiol. 2004;286:458–466.
87- Denucci C.C., Pagán A.J., Mitchell J.S., Shimizu Y. Control of α4β7 Integrin Expression and CD4 T Cell Homing by the β1 Integrin Subunit. J. Immunol. 2010;184:2458–2467.
88- Kang S.G., Park J., Cho J.Y., Ulrich B., Kim C.H. Complementary roles of retinoic acid and TGF-β1 in coordinated expression of mucosal integrins by T cells. Mucosal. Immunol. 2011;4:66–82.
89- Ohoka Y., Yokota A., Takeuchi H., Maeda N., Iwata M. Retinoic acid-induced CCR9 expression requires transient TCR stimulation and cooperativity between NFATc2 and the retinoic acid receptor/retinoid X receptor complex. J. Immunol. 2011;186:733–744.
90- Tan X., Sande J.L., Pufnock J.S., Blattman J.N., Greenberg P.D. Retinoic acid as a vaccine adjuvant enhances CD8+ T cell response and mucosal protection from viral challenge. J. Virol. 2011;85:8316–8327.
91- Allie S.R., Zhang W., Tsai C.Y., Noelle R.J., Usherwood E.J. Critical role for all-trans retinoic acid for optimal effector and effector memory CD8 T cell differentiation. J. Immunol. 2013;190:2178–2187.
92- Guo Y., Pino-Lagos K., Ahonen C.A., Bennett K.A., Wang J., Napoli J.L., Blomhoff R., Sockanathan S., Chandraratna R.A., Dmitrovsky E., et al. A retinoic acid–rich tumor microenvironment provides clonal survival cues for tumor-specific CD8+ T cells. Cancer Res. 2012;72:5230–5239.
93- Guo Y., Lee Y.C., Brown C., Zhang W., Usherwood E., Noelle R.J. Dissecting the Role of Retinoic Acid Receptor Isoforms in the CD8 Response to Infection. J. Immunol. 2014;192:3336–3344.
94- Pino-Lagos K., Guo Y., Brown C., Alexander M.P., Elgueta R., Bennett K.A., De Vries V., Nowak E., Blomhoff R., Sockanathan S., et al. A retinoic acid–dependent checkpoint in the development of CD4+ T cell–mediated immunity. J. Exp. Med. 2011;208:1767–1775.
95- Tao X.J., Ma J.L., Zhang Y.H., Yu J.N., Cai L., Wang J.H., Zheng S.G. Neutralization of IL-4 and IFN-γ Facilitates inducing TGF-β-induced CD4+Foxp3+ Regulatory Cells. Int. J. Biomed. Sci. 2008;4:52–57.
96- Zheng S.G., Gray J.D., Ohtsuka K., Yamagiwa S., Horwitz D.A. Generation ex vivo of TGF-beta-producing regulatory T cells from CD4+CD25− precursors. J. Immunol. 2002;169:4183–4189.
97- Zheng S.G., Wang J.H., Gray J.D., Soucier H., Horwitz D.A. Natural and induced CD4+CD25+ cells educate CD4+CD25− cells to develop suppressive activity: The role of IL-2, TGF-beta, and IL-10. J. Immunol. 2004;172:5213–5221.
98- Zheng S.G., Wang J., Wang P., Gray J.D., Horwitz D.A. IL-2 is essential for TGF-beta to convert naive CD4+CD25− cells to CD25+Foxp3+ regulatory T cells and for expansion of these cells. J. Immunol. 2007;178:2018–2027.
99- Hori S., Nomura T., Sakaguchi S. Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science. 2003;299:1057–1061.
100- Ma J., Liu Y., Li Y., Gu J., Liu J., Tang J., Wang J., Ryffel B., Shen Y., Brand D., et al. Differential role of all-trans retinoic acid in promoting the development of CD4+ and CD8+ regulatory T cells. J. Leukoc. Biol. 2014;95:275–283.
101- Liu Z.M., Wang K.P., Ma J., Zheng S.G. The role of all-trans retinoic acid in the biology of Foxp3+ regulatory T cells. Cell Mol. Immunol. 2015;12:553–557.
102- Mucida D., Park Y., Kim G., Turovskaya O., Scott I., Kronenberg M., Cheroutre H. Reciprocal th17 and regulatory t cell differentiation mediated by retinoic acid. Science. 2007;317:256–260.
103- Schambach F., Schupp M., Lazar M.A., Reiner S.L. Activation of retinoic acid receptor-α favours regulatory T cell induction at the expense of IL-17-secreting T helper cell differentiation. Eur. J. Immunol. 2007;37:2396–2399.
104- Lu L., Ma J., Li Z., Lan Q., Chen M., Liu Y., Xia Z., Wang J., Han Y., Shi W., et al. All-Trans Retinoic Acid Promotes TGF-β-Induced Tregs via Histone Modification but Not DNA Demethylation on Foxp3 Gene Locus. PLoS ONE. 2011;6:e24590.
105- Zhou X., Kong N., Wang J., Fan H., Zou H., Horwitz D., Brand D., Liu Z., Zheng S.G. Cutting edge: All-trans retinoic acid sustains the stability and function of natural regulatory T cells in an inflammatory milieu. J. Immunol. 2010;185:2675–2679.
106- Kwok S.K., Park M.K., Cho M.L., Oh H.J., Park E.M., Lee D.G., Lee J., Kim H.Y., Park S.H. Retinoic acid attenuates rheumatoid inflammation in mice. J. Immunol. 2012;189:1062–1067.
107- Lu L., Lan Q., Li Z., Zhou X., Gu J., Li Q., Wang J., Chen M., Liu Y., Shen Y., et al. Critical role of all-trans retinoic acid in stabilizing human natural regulatory T cells under inflammatory conditions. Proc. Natl. Acad. Sci. USA. 2014;111:3432–3440.
108- Lu L., Zhou X.H., Wang J.L., Zheng S.G. Correction: Characterization of Protective Human CD4+CD25+ FOXP3+ Regulatory T Cells Generated with IL-2, TGF-β and Retinoic Acid. PLoS ONE. 2010;5:e15150.
109- Hammerschmidt S.I., Ahrendt M., Bode U., Wahl B., Kremmer E., Forster R., Pabst O. Stromal mesenteric lymph node cells are essential for the generation of gut-homing T cells in vivo. J. Exp. Med. 2008;205:2483–2490.
110- Molenaar R., Greuter M., van der Marel A.P., Roozendaal R., Martin S.F., Edele F., Huehn J., Forster R., O’Toole T., Jansen W., et al. Lymph node stromal cells support dendritic cell-induced gut-homing of T cells. Immunology. 2009;183:6395–6402.
111- Hall J.A., Cannons J.L., Grainger J.R., Dos Santos L.M., Hand T.W., Naik S., Wohlfert E.A., Chou D.B., Oldenhove G., Robinson M., et al. Essential role for retinoic acid in the promotion of CD4+ T cell effector responses via retinoic acid receptor alpha. Immunity. 2011;34:435–447.
112- Xiao S., Jin H., Korn T., Liu S.M., Oukka M., Lim B., Kuchroo V.K. Retinoic acid increases Foxp3+ regulatory T cells and inhibits development of Th17 cells by enhancing TGF-β-driven Smad3 signaling and inhibiting IL-6 and IL-23 receptor expression. Immunology. 2008;181:2277–2284.
113- Rampal R., Awasthi A., Ahuja V. Retinoic acid-primed human dendritic cells inhibit Th9 cells and induce Th1/Th17 cell differentiation. J. Leukoc. Biol. 2016;100:111–120.
114- Takahashi H., Kanno T., Nakayamada S., Hirahara K., Sciume G., Muljo S.A., Kuchen S., Casellas R., Wei L., Kanno Y., et al. TGF-[beta] and retinoic acid induce the microRNA miR-10a, which targets Bcl-6 and constrains the plasticity of helper T cells. Nat. Immunol. 2012;13:587–595.
115- Wang C., Kang S.G., HogenEsch H., Love P.E., Kim C.H. Retinoic acid determines the precise tissue tropism of inflammatory Th17 cells in the intestine. J. Immunol. 2010;184:5519–5526.
116- Wang C., Kang S.G., HogenEsch H., Love P.E., Kim C.H. Retinoic acid determines the precise tissue tropism of inflammatory Th17 cells in the intestine. J. Immunol. 2010;184:5519–5526.
117- Cha H.R., Chang S.Y., Chang J.H., Kim J.O., Yang J.Y., Kim C.H., Kweon M.N. Downregulation of Th17 cells in the small intestine by disruption of gut flora in the absence of retinoic acid. Immunology. 2010;184:6799–6806.
118- Raverdeau M., Mills K.H. Modulation of T cell and innate immune responses by retinoic Acid. J. Immunol. 2014;192:2953–2958.
119- Ghodratizadeh S., Kanbak G., Beyramzadeh M., Dikmen Z.G., Memarzadeh S., Habibian R. Effect of carotenoid β-cryptoxanthin on cellular and humoral immune response in rabbit. Vet. Res. Commun. 2014;38:59–62.
120- Yang Y., Yuan Y., Tao Y., Wang W. Effects of vitamin A deficiency on mucosal immunity and response to intestinal infection in rats. Nutrition. 2011;27:227–232.
121- Mora J.R., Iwata M., Eksteen B., Song S.Y., Junt T., Senman B., Otipoby K.L., Yokota A., Takeuchi H., Ricciardi-Castagnoli P., et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science. 2006;314:1157–1160.
122- Pantazi E., Marks E., Stolarczyk E., Lycke N., Noelle R.J., Elgueta R. Cutting Edge: Retinoic Acid Signaling in B Cells Is Essential for Oral Immunization and Microflora Composition. J. Immunol. 2015;195:1368–1371.
123- Xu A., Liu Y., Chen W., Wang J., Xue Y., Huang F., Rong L., Lin J., Liu D., Yan M., et al. TGF-β-induced regulatory T cells directly suppress B cell responses through a non-cytotoxic mechanism. J. Immunol. 2016;196:3631–3641.
124- Heine G., Hollstein T., Treptow S., Radbruch A., Worm M. 9-cis retinoic acid modulates the type I allergic immune response. J. Allergy. Clin. Immunol. 2018;141:650–658.
125- Seo G.Y., Lee J.M., Jang Y.S., Kang S.G., Yoon S.I., Ko H.J., Lee G.S., Park S.R., Nagler C.R., Kim P.H. Mechanism underlying the suppressor activity of retinoic acid on IL4-induced IgE synthesis and its physiological implication. Cell Immunol. 2017;322:49–55.
126- Dilillo D.J., Matsushita T., Tedder T.F. B10 cells and regulatory B cells balance immune responses during inflammation, autoimmunity, and cancer. Ann. N. Y. Acad. Sci. 2010;1183:38–57.
127- Yanaba K., Bouaziz J.D., Haas K.M., Poe J.C., Fujimoto M., Tedder T.F. A regulatory B cell subset with a unique CD1dhi CD5+ phenotype controls T cell-dependent inflammatory responses. Immunity. 2008;28:639–650.
128- Yanaba K., Bouaziz J.D., Matsushita T., Tsubata T., Tedder T.F. The Development and Function of Regulatory B Cells Expressing IL-10 (B10 cells) Requires Antigen Receptor Diversity and TLR Signals. J. Immunol. 2009;182:7459–7472.
129- Wolf S.D., Dittel B.N., Hardardottir F., Janeway C.A., Jr. Experimental autoimmune encephalomyelitis induction in genetically B cell-deficient mice. J. Exp. Med. 1996;184:2271–2278.
130- Fillatreau S., Sweenie C.H., McGeachy M.J., Gray D., Anderton S.M. B cells regulate autoimmunity by provision of IL-10. Nat. Immunol. 2002;3:944–950.
131- Di Caro V., Phillips B., Engman C., Harnaha J., Trucco M., Giannoukakis N. Retinoic acid-producing, ex-vivo-generated human tolerogenic dendritic cells induce the proliferation of immunosuppressive B lymphocytes. Clin. Exp. Immunol. 2013;174:302–317.
132- Iwata Y., Matsushita T., Horikawa M., Dilillo D.J., Yanaba K., Venturi G.M., Szabolcs P.M., Bernstein S.H., Magro C.M., Williams A.D., et al. Characterization of a rare IL-10-competent B-cell subset in humans that parallels mouse regulatory B10 cells. Blood. 2011;117:530–541.
133- Mauri C., Gray D., Mushtaq N., Londei M. Prevention of arthritis by interleukin 10-producing B cells. J. Exp. Med. 2003;197:489–501.
134- Blair P.A., Noreña L.Y., Flores-Borja F., Rawlings D.J., Isenberg D.A., Ehrenstein M.R., Mauri C. CD19+ CD24hi CD38hi B cells exhibit regulatory capacity in healthy individuals but are functionally impaired in systemic Lupus Erythematosus patients. Immunity. 2010;32:129–140.
135- Ramachandran G., Santha T., Garg R., Baskaran D., Iliayas S.A., Venkatesan P., Fathima R., Narayanan P.R. Vitamin A levels in sputum-positive pulmonary tuberculosis patients in comparison with household contacts and healthy ‘normals’ Int. J. Tuberc. Lung Dis. 2004;8:1130–1133.
136- Mugusi F.M., Rusizoka O., Habib N., Fawzi W. Vitamin A status of patients presenting with pulmonary tuberculosis and asymptomatic HIV-infected individuals, Dar es Salaam, Tanzania. Int. J. Tuberc. Lung Dis. 2003;7:804–807.
137- Qrafli M., El Kari K., Aguenaou H., Bourkadi J.E., Sadki K., El Mzibri M. Low plasma vitamin A concentration is associated with tuberculosis in Moroccan population: A preliminary case control study. BMC Res. Notes. 2017;10:421.
138- Aibana O., Franke M.F., Huang C.C., Galea J.T., Calderon R., Zhang Z., Becerra M.C., Smith E.R., Ronnenberg A.G., Contreras C., et al. Impact of Vitamin A and Carotenoids on the Risk of Tuberculosis Progression. Clin. Infect. Dis. 2017;65:900–909.
139- Anand P.K., Kaul D., Sharma M. Synergistic action of vitamin D and retinoic acid restricts invasion of macrophages by pathogenic mycobacteria. J. Microbiol. Immunol. Infect. 2008;41:17–25.
140- Wheelwright M., Kim E.W., Inkeles M.S., De Leon A., Pellegrini M., Krutzik S.R., Liu P.T. All-trans retinoic acid triggered antimicrobial activity against Mycobacterium tuberculosis is dependent on NPC2. J. Immunol. 2014;192:2280–2290.
141- Mourik B.C., Leenen P.J., de Knegt G.J., Huizinga R., van der Eerden B.C., Wang J., Krois C.R., Napoli J.L., Bakker-Woudenberg I.A., de Steenwinkel J.E. Immunotherapy added to antibiotic treatment reduces relapse of disease in a mouse model of tuberculosis. Am. J. Respir. Cell Mol. Biol. 2017;56:233–241.
142- Drain P.K., Kupka R., Mugusi F., Fawzi W.W. Micronutrients in HIV-positive persons receiving highly active antiretroviral therapy. Am. J. Clin. Nutr. 2007;85:333–345.
143- Chin J. The AIDS Pandemic: The collision of epidemiology with political correctness. Drugs Alcohol. Today. 2007;7:46–47.
144- Schreck R., Rieber P., Baeuerle P.A. Reactive oxygen intermediates as apparently widely used messengers in the activation of the NF-kappa B transcription factor and HIV-1. EMBO J. 1991;10:2247–2258.
145- Makinde O., Rotimi K., Ikumawoyi V., Adeyemo T., Olayemi S. Effect of vitamin A and vitamin C supplementation on oxidative stress in HIV and HIV-TB co-infection at Lagos University Teaching Hospital (LUTH) Nigeria. Afr. Health Sci. 2017;17:308–314.
146- Visser M.E., Durao S., Sinclair D., Irlam J.H., Siegfried N. Micronutrient supplementation in adults with HIV infection. Cochrane Database Syst. Rev. 2017;5:CD003650.
147- Wiysonge C.S., Ndze V.N., Kongnyuy E.J., Shey M.S. Vitamin A supplements for reducing mother-to-child HIV transmission. Cochrane Database Syst. Rev. 2017;9:CD003648.
148- Campa A., Baum M.K., Bussmann H., Martinez S.S., Farahani M., van Widenfelt E., Moyo S., Makhema J., Essex M., Marlink R. The effect of micronutrient supplementation on active TB incidence early in HIV infection in Botswana. Nutr. Diet. Suppl. 2017;2017:37–45.
149- Fawzi W.W., Msamanga G.I., Kupka R., Spiegelman D., Villamor E., Mugusi F., Wei R., Hunter D. Multivitamin supplementation improves hematologic status in HIV-infected women and their children in Tanzania. Am. J. Clin. Nutr. 2007;85:1335–1343.
150- Shivakoti R., Christian P., Yang W.T., Gupte N., Mwelase N., Kanyama C., Pillay S., Samaneka W., Santos B., Poongulali S., et al. Prevalence and risk factors of micronutrient deficiencies pre- and post-antiretroviral therapy (ART) among a diverse multicountry cohort of HIV-infected adults. Clin. Nutr. 2016;35:183–189.
151- Rytter M.J., Kolte L., Briend A., Friis H., Christensen V.B. The Immune System in Children with Malnutrition—A Systematic Review. PLoS ONE. 2014;9:e105017.
152- Oliver T.K. Chronic vitamin A intoxication. Am. J. Dis. Child. 1953;95:57–68.
153- Bush M.E., Dahms B.B. Fatal hypervitaminosis A in a neonate. Arch. Pathol. Lab Med. 1984;108:838–842.
154- James M.B., Leonard J.C., Fraser J.J., Jr., Stuemky J.H. Hypervitaminosis A: A case report. Pediatrics. 1982;69:112–115.
155- Lippe B., Hensen L., Mendoza G., Finerman M., Welch M. Chronic vitamin A intoxication: A multisystem disease that could reach epidemic proportions. Am. J. Dis. Child. 1981;135:634–636.
156- Rubin L.P., Ross A.C., Stephensen C.B., Bohn T., Tanumihardjo S.A. Metabolic effects of inflammation on vitamin A and carotenoids in humans and animal models. Adv. Nutr. 2017;8:197–212.
157- Yang H.M., Mao M., Wan C. Vitamin A for treating measles in children. Cochrane Database Syst. Rev. 2005;5:85–86.
158- Ardawi MS, Newsholme EA. Glutamine metabolism in lymphocytes of the rat.
Biochem J (1983).
159- Parry-Billings M, et al. Does glutamine contribute to immunosuppression after major burns. Lancet (1990).
160- Kim H. Glutamine as an immunonutrient. Yonsei Med J (2011).
162- Mantzoros CS, et al. Zinc may regulate serum leptin concentrations in humans.
J Am Coll Nutr (1998)
163- Prasad AS, et al. Zinc supplementation decreases incidence of infections in the elderly: effect of zinc on generation of cytokines and oxidative stress. Am J Clin Nutr (2007).
164- The Dynamic Link between the Integrity of the Immune System and Zinc Status.
165- Singh M, Das RR. Zinc for the common cold. Cochrane Database Syst Rev (2011).
166- National Institutes of Health (Nih).
167- Nature Communications Biology, da Tami Martino della Università di Guelph, in Ontario, Canada. 10 ottobre 2019.
Con l’attuale pandemia di COVID-19 il web è letteralmente invaso da una pestilenza parallela che rischia di far diventare quella virale ancor peggiore di come la mala gestione statale l’ha già fatta diventare. Chi, in questi giorni di isteria collettiva, non si è imbattuto in uno o più post dove l’autore millantava prodigiosi effetti protettivi dati dalla somministrazione di mega dosi di Vitamina C, A e D? Postando in allegato tanto di letteratura scientifica? Ben pochi immagino. Purtroppo, sono molto pochi coloro che si prendono l’onere di approfondire tali studi citati e sono ancora meno coloro che sono in grado di capirli e contestualizzarli. Chi scrive questi post è per lo più sprovvisto della più elementare conoscenza di base in virologia per poter trattare l’argomento in modo non solo esaustivo ma anche, e soprattutto, credibile. Ed ecco il motivo che mi spinge ora a scrivere il presente articolo: per spiegare in modo più fruibile possibile cos’è un virus e fin dove la ricerca ci può dare, ad oggi, direttive veramente utili per prevenire e trattare una contaminazione virale, smontando le “perle guresche” che rischiano di far sottovalutare un problema lievitato a causa, per l’appunto, di una sottovalutazione eccessiva fatta di ignoranza all’italiana e paragoni impropri.
Virus: loro scoperta e caratteristiche
Un Virus (la parola “virus” deriva dalla parola latina vīrus, che significa “tossina” o “veleno”) è un microrganismo acellulare con caratteristiche di parassita obbligato, in quanto si replica esclusivamente all’interno delle cellule di altri organismi. Infatti, i Virus possono infettare tutti i tipi di forme di vita, dagli animali, alle piante, ai microrganismi (compresi batteri e archeobatteri)[1] e anche altri Virus.[2][3]
Nel 1892 Dmitri Ivanovsky descrisse in un articolo un agente patogeno non batterico in grado di infettare le piante di tabacco, da cui in seguito, sei anni più tardi, Martinus Beijerinck scoprì il Virus del mosaico del tabacco nel 1898.[4] Già tempo prima, Louis Pasteur riuscì a determinare l’agente eziologico per la rabbia e previde l’esistenza di un patogeno troppo piccolo per essere rilevato utilizzando un microscopio.[5] Nel 1884, il microbiologo francese Charles Chamberland inventò un filtro (conosciuto oggi come il filtro Chamberland o filtro di Chamberland-Pasteur) con pori più piccoli dei batteri. Così, egli poteva passare una soluzione contenente batteri attraverso esso e quindi rimuoverli completamente dalla soluzione.[6]
Dmitri Ivanovsky e l’agente patogeno di sua scoperta (Virus del mosaico del Tabacco).
Gli esperimenti di Dmitri Ivanovsky hanno dimostrato che estratti di foglie da piante di tabacco schiacciate e infettate rimangono infettive dopo la filtrazione. Ivanovsky suggerì che l’infezione poteva essere causata da una tossina prodotta da batteri, ma non approfondì la questione.[7] Al momento si pensava che tutti gli agenti infettivi potessero essere trattenuti dai filtri e coltivati su un terreno nutritivo.[8] Nel 1898, il microbiologo olandese Martinus Beijerinck ripeté gli esperimenti e si convinse che la soluzione filtrata conteneva una nuova forma di agente infettivo.[9] Egli osservò che questo agente si moltiplicava solo nelle cellule che si stavano dividendo, ma poiché i suoi esperimenti non mostrarono che fosse fatto di particelle, lo chiamò contagium vivum fluidum (solubile germe di vita), e reintrodusse la parola virus.[7] Beijerinck sostenne che i virus fossero liquidi in natura, una teoria più tardi screditata da Wendell Meredith Stanley, che dimostrò che erano delle particelle.[7] Nello stesso anno Friedrich Loeffler e Paolo Frosch isolarono il primo virus animale, l’agente dell’afta epizootica (aphthovirus), attraverso un filtro simile.[10]
Nel XX secolo, il batteriologo inglese Frederick Twort scoprì un gruppo di virus capaci di infettare i batteri, oggi chiamati batteriofagi[8] (o comunemente fagi) e il microbiologo franco-canadese Félix d’Herelle descrisse un virus che, quando aggiunto a batteri su agar, produrrebbe aree di batteri morti. I fagi furono considerati come un potenziale trattamento per alcune malattie come il tifo e il colera, ma tale idea fu messa da parte grazie all’introduzione della penicillina. Lo studio di fagi, tuttavia, ha permesso di capire alcuni aspetti dell’espressione genica e un meccanismo utile per introdurre geni estranei nei batteri.
Alla fine del XIX secolo, i virus vennero classificati secondo la loro infettività, la loro capacità di essere filtrati e i requisiti per i loro ospiti viventi. Inizialmente i virus vennero coltivati solo nelle piante e negli animali. Nel 1906, Ross Granville Harrison introdusse un metodo per la coltivazione nella linfa, e, nel 1913, E. Steinhardt, C. Israeli, and R. A. Lambert utilizzarono questo metodo per allevare i virus destinati a diventare vaccini in frammenti di tessuto corneale di una cavia.[11] Nel 1928, HB Maitland e MC Maitland fecero moltiplicare i virus vaccino in sospensioni di reni di galline. Il loro metodo incominciò a essere ampiamente adottato a partire dal 1950, quando il poliovirus venne coltivato su larga scala per la produzione del vaccino antipoliomielite.[12]
Ernest William Goodpasture
Nel 1931 il patologo statunitense Ernest William Goodpasture fece crescere il virus dell’influenza e molti altri virus nelle uova di galline fecondate.[13-23] Nel 1949, John Franklin Enders, Thomas Weller e Frederick Robbins fecero crescere il virus della poliomielite in cellule in coltura su embrioni umani, il primo virus a essere coltivato senza l’uso di tessuti animali solidi o uova. Questo lavoro permise a Jonas Salk di realizzare un efficace vaccino antipolio.[14-24]
Nel 1935, dopo l’invenzione del microscopio elettronico evvenuta nel 1931 grazie al lavoro degli ingegneri tedeschi Ernst Ruska e Max Knoll [15], il biochimico e virologo americano Wendell Meredith Stanley analizzò il virus del mosaico del tabacco e dimostrò che era per la maggior parte costituito da proteine.[16] Poco tempo dopo, fu possibile separare in questo virus le proteine dall’RNA.[17] Le prime immagini a raggi X di diffrazione del virus cristallizzato, furono ottenute da Bernal e Fankuchen nel 1941. Sulla base delle loro foto, Rosalind Franklin scoprì, nel 1955, la struttura completa del virus.[18] Nello stesso anno, Heinz Fraenkel-Conrat e Robley Williams dimostrarono che il virus del mosaico del tabacco purificato e il suo rivestimento proteico sono in grado di assemblare da soli dei virus funzionali, suggerendo che questo semplice meccanismo è probabilmente stato il mezzo attraverso cui i virus vengono composti all’interno delle loro cellule ospiti.[19]
Luc Antoine Montagnier
La seconda metà del XX secolo è stato un periodo d’oro per lo studio dei virus, e buona parte delle circa 5000 specie è stata scoperta nel corso di questi anni.[20] Nel 1957 fu scoperto l’arteriviridae equino, la causa della diarrea virale bovina (una pestivirus). Nel 1963 Baruch Blumberg scoprì il virus dell’epatite B,[21] e nel 1965, Howard Temin descrisse il primo retrovirus. La trascrittasi inversa, l’enzima che i retrovirus utilizzano per fare copie di DNA del loro RNA, è stata descritta per la prima volta nel 1970, indipendentemente da Howard Martin Temin e da David Baltimore.[22] Nel 1983, il gruppo guidato da Luc Montagnier, presso l’Istituto Pasteur in Francia, isolò per primo il retrovirus oggi chiamato HIV.[23]
Come precedentemente accennato, i virus sono tutti parassiti endocellulari obbligati. All’esterno delle cellule ospiti sono costituiti da un virione, formato da una capsula proteica (detta capside) contenente l’acido nucleico. I virus degli Eucarioti possono possedere anche una membrana che avvolge il capside detta peplos o pericapside. Talvolta tra il capside e il peplos presentano un ulteriore strato proteico che prende il nome di tegumento. I virioni non possiedono metabolismo: vengono quindi trasportati passivamente finché non trovano una cellula da infettare. L’infezione di una cellula ospite richiede il legame con proteine specifiche di membrana.
Nelle cellule infettate i virus perdono la loro individualità strutturale: consistono negli acidi nucleici e nei loro prodotti che assumono il controllo di parte dell’attività biosintetica cellulare al fine di produrre nuovi virioni.
In alternativa, alcuni virus possono inserire fisicamente il loro genoma in quello dell’ospite in modo che sia replicato insieme con esso. Il genoma virale inserito in quello dell’ospite, detto provirus, riprende la sua individualità e produce nuovi virioni in caso di danneggiamento della cellula ospite.
Attenzione: sebbene virus e batteri possano provocare malattie, sono assai diversi tra loro! Sebbene abbiano alcune caratteristiche comuni, come la capacità di trasmettersi da un individuo a un altro, e possano indurre, talvolta, patologie simili, sono molto diversi tra loro. I batteri sono organismi costituiti da una sola cellula (unicellulari) procarioti (ossia privi di nucleo cellulare), ma assimilabili a organismi viventi completi, poiché sono dotati di tutte le strutture e gli enzimi necessari per espletare le funzioni metaboliche fondamentali, potendo quindi vivere in modo indipendente e riprodursi autonomamente, per divisione cellulare. I virus sono, mediamente, 100 volte più piccoli dei batteri (al punto da risultare invisibili al microscopio ottico), e non possiedono né una struttura cellulare completa, né tutti gli enzimi necessari per la sopravvivenza e la replicazione del proprio acido nucleico, che può essere costituito da DNA o RNA (in questo secondo caso vengono chiamati “retrovirus” e l’esponente più noto del gruppo è l’HIV). Inoltre, i virus non posseggono ribosomi, che sono gli organelli cellulari indispensabili per produrre tutte le proteine esistenti in natura, quindi anche quelle che formano il capside virale, corrispondente all’involucro del virus. Ciò rende i virus incapaci di riprodursi autonomamente e fa sì che, per potersi moltiplicare e causare infezioni, debbano entrare in una cellula ospite “completa” (che può essere una cellula animale o vegetale oppure una cellula batterica) e sfruttarne le strutture e gli enzimi di cui sono carenti. I virus che infettano e sfruttano cellule batteriche sono chiamati “batteriofagi”.
Vorrei rammentare, quindi, che gli antibiotici non hanno alcun effetto sui virus!
L’origine dei virus non è tuttavia chiara anche perché non formano fossili nel senso classico del termine; tecniche molecolari sono state utilizzate per confrontare il DNA o l’RNA dei virus e si sono dimostrate molto utili per indagare su come si potevano presentare in principio. [24] Inoltre, il materiale genetico virale può occasionalmente passare nella linea germinale degli organismi ospitanti, per cui i discendenti ospitanti avranno il virus integrato nel proprio genoma. Ciò fornisce una preziosa fonte di informazioni per i virologi per poter risalire ai “virus primari”. Attualmente, vi sono tre ipotesi principali che mirano a spiegare le origini dei virus:[25][26]
Ipotesi regressiva
Questa teoria presenta due diverse varianti espositive. La prima, prettamente evoluzionista, afferma che i virus potrebbero essere stati una volta delle piccole cellule parassitarie di cellule più grandi. Nel corso del tempo, i geni non richiesti dalla loro natura parassitaria sono andati persi. La seconda, molto simile alla prima, ricerca la variazione genetica sotto la luce dell’entropia, ovvero della perdita di informazioni geniche dei sistemi virali primari, ed è la più accreditabile rispetto alla precedente versione sebbene si discosti di poche essenziali puntualizzazioni. Comunque sia, i batteri rickettsia e clamidia sono le cellule che, come i virus, sono in grado di riprodursi solo all’interno di cellule ospiti viventi. Essi prestano sostegno a questa ipotesi per il fatto che la loro dipendenza da parassitismo probabilmente causò loro la perdita dei geni, oppure la perdita di informazioni geniche ne è stato l’imput, che gli potevano permettere di sopravvivere al di fuori di una cellula. Questa ipotesi è chiamata anche di degenerazione[27][28] o di riduzione.[29]
Ipotesi dell’origine cellulare
Questa, talvolta chiamata ipotesi del vagabondaggio[27][30] o ipotesi di fuga,[29] consiste nel credere, e sottolineo “credere”, che alcuni virus si siano evoluti da frammenti di DNA o RNA che sono “sfuggiti” da geni di un organismo più grande. Il DNA fuggito avrebbe potuto provenire da plasmidi (frammenti di DNA che possono muoversi tra le cellule) o trasposoni/retrotrasposoni (molecole di DNA che si replicano e si muovono da diverse posizioni all’interno dei geni della cellula).[31] Una volta chiamati “geni che saltano”, i trasposoni sono esempi di elementi genetici mobili che potrebbero essere l’origine di alcuni virus perché hanno delle sequenze analoghe. Essi sono stati scoperti nel mais da Barbara McClintock nel 1950.[32] Ipotesi della coevoluzione
La teoria più eccentrica e meno scientifica (nel senso più stretto del termine) è senz’altro quella chiamata anche ipotesi del primo virus[29] e propone che i virus si siano evoluti da molecole complesse di proteine e acidi nucleici contemporaneamente alla prima apparizione sulla Terra della cellula e sarebbero rimasti dipendenti dalla vita cellulare per miliardi di anni. I viroidi sono molecole di RNA che non sono classificati come virus poiché mancano di un rivestimento proteico. Tuttavia, essi hanno caratteristiche che sono comuni a diversi virus e sono spesso chiamati agenti subvirali.[33] I viroidi sono importanti patogeni delle piante.[34] Loro non codificano proteine ma interagiscono con la cellula ospite e utilizzano le sue strutture per replicarsi.[35] Il virus dell’epatite delta degli esseri umani ha un genoma RNA simile ai viroidi, ma è dotato di un rivestimento proteico derivato dal virus dell’epatite B e non può produrne uno suo. È, quindi, un virus difettoso. Sebbene il genoma del virus dell’epatite delta possa replicarsi indipendentemente all’interno di una cellula ospite, esso richiede la presenza del virus dell’epatite B che gli fornisca un rivestimento proteico in modo che possa essere trasmesso alle nuove cellule.[36] In modo analogo, il virofago Sputnik dipende dai mimivirus, e infetta il protozoo Acanthamoeba castellanii.[37] Questi virus, che dipendono dalla presenza di altre specie di virus nella cellula ospite, sono chiamati “satelliti” e possono rappresentare intermedi evolutivi dei viroidi e dei virus.[38][39]
In passato, vi sono stati problemi con tutte queste ipotesi: l’ipotesi regressiva non spiega il motivo per cui anche il più piccolo dei parassiti cellulari non assomigli ai virus in alcun modo. L’ipotesi della fuga non spiega la presenza di capsidi complessi e altre strutture di particelle virali. L’ipotesi del primo virus viola la stessa definizione di virus che richiede una cellula ospite.[29] I virus sono ora riconosciuti come molto antichi e con una data d’origine antecedente alla divergenza della vita nei tre domini.[40] Questa scoperta ha portato i virologi moderni a riconsiderare e rivalutare le tre ipotesi classiche.[40-51]
L’evidenza di un mondo ancestrale delle cellule RNA[41] e le analisi al computer di sequenze di DNA virale stanno fornendo una migliore comprensione delle relazioni evolutive tra i diversi virus e potranno aiutare a identificare gli antenati dei virus moderni. Fino a oggi, queste analisi non hanno potuto dimostrare quale di queste ipotesi sia più corretta.[41] Tuttavia, sembra improbabile che tutti i virus attualmente conosciuti abbiano un antenato comune e probabilmente molti virus sono nati in passato con diversi meccanismi.[42]
Qualunque sia la loro origine primordiale, vi è una divergenza di opinioni sulla questione se i virus siano una forma di vita o strutture organiche che interagiscono con gli organismi viventi. Nonostante possiedano geni, essi non hanno una struttura cellulare, aspetto che viene spesso visto, da un certo filone di ricercatori, come l’unità base della vita. I virus non hanno nemmeno un proprio metabolismo e necessitano della cellula ospite per realizzare nuovi prodotti. Essi, pertanto, non possono naturalmente riprodursi al di fuori di una cellula ospite.[43] Tuttavia specie batteriche come la rickettsia e la clamidia sono considerati organismi viventi nonostante abbiano la stessa limitazione.[44][45] Forme di vita accettate come tali utilizzano la divisione cellulare per riprodursi, mentre i virus si assemblano spontaneamente all’interno delle cellule. Si differenziano dalla crescita autonoma dei cristalli per il fatto che ereditano mutazioni genetiche e sono soggetti alla selezione naturale. Sebbene la cerchia degli evoluzionisti abbia affermato che tali caratteristiche di “auto-assemblaggio” dei virus nelle cellule ospiti comporta implicazioni per lo studio dell’origine della vita, in quanto, secondo loro ipotesi, conferisce ulteriore credito all’ipotesi che la vita possa aver avuto origine da molecole organiche auto-assemblate.[1] In realtà, tale caratteristica continua ad avvalorare il fenomeno entropico dei geni e spinge ad ipotizzare che i virus siano “mezzi di controllo” specifici interagenti con le forme di vita e soggetti a degenerazione sistemica del genoma con la conseguente comparsa di forme virali mutate e potenzialmente dannose in alcuni casi e a diversa entità.
I virus mostrano una grande diversità di forme e dimensioni, chiamate morfologie. In generale, i virus sono molto più piccoli dei batteri. La maggior parte dei virus che sono stati studiati hanno un diametro tra 20 e 300 nanometri. Alcuni filovirus arrivano fino a una lunghezza totale a 1400 nm, ma il loro diametro è solo di circa 80 nm.[46] Per questo motivo maggior parte dei virus non possono essere visti con un microscopio ottico e dunque per visualizzare i virioni è necessario utilizzare il microscopio elettronico.[47]
Una particella virale completa, nota come un virione, è costituita da acido nucleico circondato da un rivestimento protettivo formato da proteine chiamato capside. Questo è realizzato da subunità proteiche identiche chiamati capsomeri.[48] I virus possono avere una “sacca” lipidica derivata dalla membrana della cellula ospite. Il capside è costituito da proteine codificate dal genoma virale e la sua forma serve come base per la distinzione morfologica.[49][50] Perché le subunità proteiche virali codificate si possano auto-assemblare per formare il capside, in generale si richiede la presenza del genoma virale. Virus complessi codificano proteine che aiutano la costruzione del loro capside. Le proteine associate all’acido nucleico sono note come nucleoproteine e l’associazione di proteine del capside virale con l’acido nucleico virale è chiamato nucleocapside. Il capside e l’intera struttura del virus possono essere fisicamente sondati attraverso il microscopio a forza atomica.[51][52]
In generale, i virus si possono distinguere in quattro tipologie morfologiche:
Elicoidale
Virus Elicoidale
Questi virus sono composti da un unico tipo di capsomero accatastato intorno a un asse centrale, formando una struttura elicoidale, che può avere una cavità centrale. Questa disposizione comporta virioni a forma di bastoncello o filamentosi: questi possono essere brevi ed estremamente rigidi o lunghi e molto flessibili. Il materiale genetico, in generale, RNA a singolo filamento, ma in alcuni casi anche DNA a singolo filamento, è vincolato nell’elica proteica dalle interazioni tra l’acido nucleico caricato negativamente e le cariche positive poste sulla proteina. In generale, la lunghezza di un capside elicoidale è correlato alla lunghezza dell’acido nucleico in esso contenuto e il diametro dipende dalla dimensione e disposizione dei capsomeri. Il virus del mosaico del tabacco è un esempio ben analizzato di virus elicoidale.[53]
Icosaedrica
Virus a struttura Icosaedrica
La maggior parte dei virus che colpiscono gli animali sono di morfologia icosaedrica o quasi sferica con simmetria icosaedrica. Un icosaedro regolare è un modo ottimale per formare un guscio chiuso da sub-unità identiche. Il numero minimo di capsomeri identici richiesto è dodici, ciascuno composto da cinque subunità identiche. Molti virus, come il rotavirus, possiedono più di dodici capsomeri e sembrano sferici pur mantenendo questa simmetria. I capsomeri agli apici sono circondati da altri cinque capsomeri e sono chiamati pentoni. I capsomeri sulle facce triangolari sono circondati da altri sei e sono chiamati esoni.[54] Gli esoni sono essenzialmente pianeggianti, mentre i pentoni che formano i 12 vertici sono curvilinei. La stessa proteina può agire come subunità sia nei pentoni sia negli esoni o possono essere composti da proteine differenti.
Ovoidale
Virus a struttura ovoidale
Si tratta di una forma a icosaedro allungata di circa cinque volte lungo l’asse ed è tipica delle teste dei batteriofagi. Questa struttura è composta da un cilindro con una capsula a entrambe le estremità.[55]
Involucro
Virus con involucro
Alcune specie di virus si avvolgono in una forma modificata di una delle membrane cellulari, o la membrana esterna che circonda la cellula ospite infettata o le membrane interne come la membrana nucleare o il reticolo endoplasmatico, ottenendo così un doppio strato lipidico esterno noto come pericapside o involucro virale. Questa membrana è costellata da proteine codificate dal genoma virale e dal genoma ospite; la stessa membrana lipidica e qualsiasi carboidrato presente provengono interamente dall’ospite. Il virus influenzale e quello dell’HIV usano questa strategia. L’infettività della maggior parte di questi virus dipende dal loro involucro.[56]
Complesso
Virus complesso
Questi virus possiedono un capside che non è né puramente elicoidale né puramente icosaedrico e che può possedere strutture extra come code proteiche o una parete esterna complessa. Alcuni batteriofagi, come, il batteriofago t4 hanno una struttura complessa costituita da una testa icosaedrica associata a una coda elicoidale, che può avere una piastra di base esagonale con una coda proteica sporgente. Tale struttura a coda agisce come una siringa molecolare, attaccandosi al batterio ospite e per poi iniettare il genoma virale nella cellula.[57]
Il genoma virale è associato con le proteine all’interno di una struttura centrale del disco, conosciuta come nucleoide. Il nucleoide è circondato da una membrana e due corpi laterali dalla funzione ad oggi sconosciuta. Il virus ha un involucro esterno con uno strato molto spesso di proteine costellato sulla sua superficie. L’intero virione è leggermente pleimorfico, dalla forma ovoidale.[58]
Un’enorme varietà di strutture genomiche possono essere riscontrate tra le specie virali; come gruppo, essi contengono una diversità genomica strutturale maggiore rispetto alle piante, agli animali, agli archeobatteri o ai batteri. Vi sono milioni di diversi tipi di virus,[6] anche se solo circa 5.000 tipi sono stati descritti in dettaglio.[5] A settembre 2015, il database NCBI dei genoma dei virus contava più di 75.000 sequenze genomiche complete[59], ma senza dubbio ve ne sono molte di più ancora da scoprire.[60]
Un virus può essere dotato di un genoma a DNA o uno a RNA e pertanto vengono denominati rispettivamente virus a DNA o virus a RNA; la stragrande maggioranza sono a RNA. I virus delle piante tendono ad avere genomi composti da un singolo filamento di RNA mentre i batteriofagi spesso hanno un genoma a DNA a doppia elica.[61]
I genomi virali possono essere circolari, come nel poliomavirus, o lineari come negli adenovirus. Il tipo di acido nucleico è irrilevante per la forma del genoma. Tra virus a RNA e certi virus a DNA, il genoma è spesso diviso in parti distinte, nel qual caso esso è chiamato “segmentato”. Per i virus a RNA, ogni segmento spesso codifica per una sola proteina e si trovano solitamente in un unico capside. Tuttavia, non è necessario che tutti i segmenti siano nello stesso virione perché il virus sia infettivo, come dimostrato dal virus del mosaico del bromus e da molti altri virus delle piante.[46]
Un genoma virale, indipendentemente dal tipo di acido nucleico, è quasi sempre a singolo filamento o doppio filamento. Genomi a singolo filamento consistono di un acido nucleico spaiato. Genomi a doppio filamento sono costituiti da due acidi nucleici complementari accoppiati. Le particelle virali di alcune famiglie di virus, come quelli appartenenti agli Hepadnaviridae, contengono un genoma che è parzialmente a doppio filamento e parzialmente a singolo filamento.[61]
Per la maggior parte dei virus con genoma a RNA e alcuni con genomi a DNA a singolo filamento, i singoli filamenti sono classificati a senso-positivo (chiamato filamento positivo) o senso-negativo (chiamato filamento negativo), a seconda se sono complementari all’RNA messaggero virale (mRNA). L’RNA virale a senso positivo è nello stesso senso dell’mRNA virale e quindi almeno una parte di esso può essere tradotto immediatamente dalla cellula ospite. L’RNA virale a senso negativo è complementare all’mRNA e quindi deve essere convertito in RNA a senso-positivo tramite una RNA polimerasi RNA-dipendente prima della traduzione. La nomenclatura dei virus DNA con un genoma a un solo senso ssDNA è simile alla nomenclatura RNA, in quanto il filamento stampo per l’mRNA virale è complementare a esso (-) e il filamento codificante è una copia (+).[46] Tuttavia, diversi tipi di virus ssDNA e ssRNA hanno genomi che sono ambisenso e che la trascrizione può verificarsi fuori da entrambi i filamenti in un doppio filamento replicativo intermedio. Gli esempi includono i geminivirus, che sono i virus ssDNA delle piante e gli arenavirus che sono i virus ssRNA degli animali.[62]
La dimensione del genoma varia notevolmente tra le specie. I genomi virali più piccoli, i circoviruses ssDNA della famiglia dei circoviridae, codificano solo per due proteine e hanno una dimensione del genoma di due sole kilobase;[63] il più grande dei pandoravirus possono avere dimensioni del genoma di circa due megabasi che codificano per circa 2500 proteine.[64]
In generale, i virus a RNA hanno dimensioni del genoma più piccoli rispetto ai virus a DNA a causa di un più alto tasso di errore durante la replica e hanno un limite di dimensione massima superiore. Al di là di questo limite, gli errori nel genoma durante la replica rendono il virus inutile o non infettivo. Per compensare questo, i virus a RNA spesso hanno un genoma segmentato: il genoma risulta pertanto diviso in molecole più piccole, riducendo così la possibilità che un errore in un singolo componente inabiliti l’intero genoma. Al contrario, i virus a DNA, in genere, possiedono genomi più grandi a causa della alta fedeltà dei loro enzimi di replicazione.[65] I virus a DNA a singolo filamento sono un’eccezione a questa regola, tuttavia, i tassi di mutazione di questi genomi possono avvicinarsi al caso estremo dei virus ssRNA.[66]
I virus, al pari degli altri sistemi biologici, subiscono mutazioni genetiche attraverso diversi meccanismi. Tra questi, un processo chiamato deriva antigenica le cui basi individuali nel DNA o RNA mutano altre basi ma sempre all’interno della tipologia virale di appartenenza. La maggior parte di queste mutazioni puntiformi sono “silenziose”, ovvero non cambiano la proteina che il gene codifica. Ma altri possono conferire diversità adattative quali la resistenza ai farmaci antivirali.[67-84][68-85] Lo spostamento antigenico si verifica quando vi è un cambiamento importante nel genoma del virus. Questo può essere il risultato di una ricombinazione o un riassortimento dei nucleotidi già presenti. Quando questo accade con i virus influenzali, possono verificarsi le pandemie, come quella che stiamo attualmente subendo.[69-86] I virus a RNA spesso esistono come “quasispecie” o sciami di virus della stessa specie, ma con una sequenza nucleotidica leggermente diversa del genoma. Tali quasispecie sono un obiettivo primario per la selezione naturale scientificamente intesa, ossia una riduzione delle informazioni che determina ad un certo punto la scomparsa di un ceppo virale o, in precedenza, la sua modifica di attività.[70]
Sequenze di genomi conferiscono vantaggi adattativi in ambienti specifici; diversi ceppi di un virus con un genoma segmentato possono mischiare e combinare i geni e quindi produrre prole virus con caratteristiche uniche. Questo è chiamato riassortimento.[71]
La ricombinazione genetica è il processo mediante il quale un filamento di DNA viene rotto per poi essere unito al termine di una diversa molecola di DNA. Questo può verificarsi quando i virus infettano cellule simultaneamente e studi riguardanti l’evoluzione virale hanno dimostrato che la ricombinazione è stata molto frequente nelle specie studiate.[72] La ricombinazione è comune a entrambi i virus, sia a RNA sia a DNA.[73][74]
I virus non sono in grado di riprodursi attraverso la divisione cellulare poiché non sono cellule. Per tanto sfruttano il metabolismo e le risorse di una cellula ospite per produrre copie multiple di sé che si assemblano nella cellula.
Il ciclo di vita dei virus varia molto attraverso le diverse specie, ma vi sono sei fasi fondamentali:[75]
L’attaccamento è un legame tra le proteine del capside virale e specifici recettori che si trovano sulla superficie cellulare sulla cellula ospite. La presenza di dati recettori sulla superficie determinano la gamma degli ospiti di un virus. Ad esempio, l’HIV infetta una gamma limitata di leucociti umani. Questo perché le sue proteine di superficie, gp120, interagiscono specificamente con la molecola CD4, un recettore delle chemochine, che frequentemente si trova sulla superficie dei linfociti T. Questo meccanismo si è evoluto in modo da favorire quei virus che infettano solo le cellule in cui sono in grado di replicarsi. Il recettore attaccato può indurre la proteina dell’involucro virale a subire cambiamenti che si traducono nella fusione tra le membrane virali e cellulari, o cambiamenti alle proteine senza involucro di superficie del virus che non gli permettono di entrare.
L’ingressosegue l’attaccamento: virioni entrano nella cellula ospite mediante endocitosi mediata da recettori o attraverso la fusione delle membrane. Questo è spesso chiamato ingresso del virus. L’infezione delle cellule delle piante e dei funghi è diversa da quella delle cellule animali. Le piante hanno una parete cellulare rigida formata da cellulosa, mentre i funghi una di chitina, quindi la maggior parte dei virus possono entrare all’interno di queste cellule solo dopo aver perpetrato una rottura alla parete cellulare.[76] Tuttavia, quasi tutti i virus delle piante (come il virus mosaico del tabacco) possono anche passare direttamente da cellula a cellula, sotto forma di complessi a singolo filamento nucleoproteici, attraverso i pori chiamati plasmodesmi.[77] I batteri, come le piante, hanno pareti cellulari che un virus deve violare per infettare la cellula. Tuttavia, dato che le pareti delle cellule batteriche sono molto meno spesse delle pareti cellulari delle piante, per via delle loro dimensioni molto più piccole, alcuni virus possiedono evoluti meccanismi che iniettano il loro genoma della cellula batterica attraverso la parete cellulare, mentre il capside virale rimane all’esterno.[78]
L’uncoating è un processo in cui il capside virale viene rimosso: questo può essere dovuto alla degradazione da parte degli enzimi virali o agli enzimi ospiti o per semplice dissociazione; il risultato finale è la liberazione dell’acido nucleico genomico virale.
La replicazionedei virus coinvolge principalmente la moltiplicazione del genoma. La replica consiste nella sintesi dell’RNA messaggero (mRNA) virale dai geni “early” (con eccezioni per i virus RNA a senso positivo), la sintesi proteica virale, il possibile montaggio delle proteine virali, quindi la replicazione del genoma virale. Questo può essere seguito, per i virus più complessi con genomi più grandi, da parte di uno o più cicli di sintesi di mRNA.
Assemblaggio, dopo l’auto-assemblaggio struttura-mediata delle particelle virali, alcune modifiche delle proteine si verifica spesso. Nei virus come l’HIV, questa modifica (a volte chiamato la maturazione) avviene dopo che il virus è stato rilasciato dalla cellula ospite.[79]
Rilascio, i virus possono essere rilasciati dalla cellula ospite per lisi, un processo che uccide la cellula rompendo la sua membrana e la parete cellulare, se presenti: questo avviene in molti batteri e di alcuni virus animali. Alcuni virus subiscono un ciclo lisogenico in cui il genoma virale è incorporato mediante la ricombinazione genetica in un luogo specifico dei cromosomi dell’ospite. Il genoma virale è quindi noto come “provirus” o, nel caso di un batteriofagi “profago”.[80-97] Ogni volta che l’ospite si divide, anche il genoma virale viene replicato. Il genoma virale è per lo più silente all’interno dell’ospite. Tuttavia, a un certo punto, il provirus o profago possono dar luogo al virus attivo, che può lisare le cellule ospiti.[81] I virus capsulati (ad esempio, l’HIV) di solito vengono rilasciati dalla cellula ospite per gemmazione. Durante questo processo il virus acquisisce il suo involucro, che è una parte modificata del plasma dell’ospite o della membrana interna.[82]
Il materiale genetico all’interno delle particelle del virus e il metodo con cui il materiale viene replicato, varia notevolmente tra i diversi tipi di virus.
Virus a DNA
La replicazione del genoma della maggior parte dei virus a DNA avviene nel nucleo della cellula. Se la cellula possiede il recettore appropriato sulla sua superficie, questi virus entrano, a volte, nella cellula per fusione diretta con la membrana cellulare (per esempio, l’herpes virus) o, più spesso. per endocitosi mediata da recettori. La maggior parte dei virus a DNA dipendono interamente dal DNA della cellula ospite e dai suoi strumenti di sintesi e lavorazione dell’RNA. Tuttavia, virus dotati di genomi più grandi, possono codificare gran parte di questi stessi strumenti. Negli eucarioti, il genoma virale deve attraversare la membrana nucleare della cellula per accedere ai meccanismi di replicazione, mentre nei batteri deve solo entrare nella cellula.[83]
Virus a DNA
Virus a RNA
La replicazione di solito avviene nel citoplasma. I virus a RNA possono essere classificati in quattro gruppi differenti in base alle modalità di replicazione. La polarità (se può o non può essere utilizzata direttamente dai ribosomi per produrre proteine) dei virus a singolo filamento di RNA determina in gran parte il meccanismo di replicazione; l’altro criterio principale è se il materiale genetico è a singolo o doppio filamento. Tutti i virus a RNA utilizzano i propri enzimi RNA replicasi per creare copie dei loro genomi.[84]
Virus a RNA
Virus a trascrizione inversa
Infezione da virus a trascrizione inversa
I virus a trascrizione inversa, possiedono ssRNA (Retroviridae, Metaviridae, Pseudoviridae) o dsDNA (Caulimoviridae, Hepadnaviridae) nelle loro particelle. Quelli con genomi a RNA (retrovirus), utilizzano un intermedio del DNA per replicare, mentre quelli con genomi di DNA (pararetroviruses) utilizzano un intermedio di RNA durante la replicazione del genoma. Entrambi i tipi utilizzano una trascrittasi inversa, o un enzima DNA polimerasi (RNA-dipendente) per effettuare la conversione dell’acido nucleico. I retrovirus integrano il DNA prodotto dalla trascrizione inversa nel genoma dell’ospite come provirus come parte del processo di replica; tuttavia, i pararetroviruses non lo fanno, anche se copie del genoma di particolari pararetroviruses infiltrati nei vegetali possono dar luogo a virus infettivo.[85] Essi sono suscettibili di farmaci antivirali che inibiscono l’enzima della trascrittasi inversa, ad esempio, la zidovudina e la lamivudina. Un esempio del primo tipo è l’HIV, che è un retrovirus. Esempi del secondo tipo sono il l’hepadnaviridae, che comprende il virus dell’epatite B.[86]
Vi è un’ampia serie di effetti strutturali e biochimici che i virus producono sulla cellula ospite.[87] Questi sono chiamati effetti citopatici.[88] La maggior parte delle infezioni da virus alla fine provocano la morte della cellula ospite. Le cause di morte sono la lisi cellulare, le alterazioni della membrana superficie della cellula e l’apoptosi.[89-106] Spesso la morte delle cellule è causata dalla cessazione delle sue normali attività a causa della soppressione da parte delle proteine specifiche del virus, che non tutti sono componenti della particella virale.[90]
Alcuni virus non provocano cambiamenti apparenti alla cellula infettata. Le cellule in cui il virus è latente e inattivo mostrano pochi segni di infezione e spesso vivono normalmente.[91] Ciò provoca infezioni persistenti e il virus rimane spesso in sospeso per molti mesi o anni. Ciò avviene frequentemente, ad esempio, con l’herpes virus.[92][93] Alcuni virus, come il virus di Epstein-Barr, possono indurre le cellule a proliferare senza provocare malignità,[94] mentre altri, come il papillomavirus, sono cause ormai stabilite di insorgenza di tumori.[95]
I virus sono di gran lunga le più abbondanti entità biologiche sulla Terra e superano tutte le altre messe insieme.[96] Essi sono in grado di infettare tutti i tipi di vita cellulare, tra cui animali, piante, batteri e funghi.[5] Tuttavia, le diverse tipologie di virus possono infettare solo una gamma limitata di ospiti e molti sono specie-specifici. Alcuni, come il virus del vaiolo, ad esempio, possono infettare solo una specie, in questo caso l’uomo,[97-114] e pertanto si dice che hanno una gamma di ospiti potenziali ristretta. Altri virus, come il virus della rabbia, possono infettare diverse specie di mammiferi e si dice che hanno una gamma di ospiti vasta.[98] I virus che infettano le piante sono innocui per gli animali e la maggior parte dei virus che infettano gli altri animali sono innocui per gli esseri umani.[99] La gamma dei possibili ospiti di alcuni batteriofagi è limitata a un unico ceppo di batteri e può possono essere usati per rintracciare la fonte di focolai di infezioni, tramite un metodo chiamato fagotipizzazione.[100]
La classificazione dei virus cerca di descrivere la loro diversità nominandoli e raggruppandoli sulla base di somiglianze. Nel 1962, André Lwoff, Robert W. Horne e Paul Tournier per primi svilupparono un sistema di classificazione dei virus, in base al sistema gerarchico di Linneo.[101] Questo approccio si basa sul phylum, sulla classe, sull’ordine, sulla famiglia, sul genere e sulla specie. I virus sono stati raggruppati in base alle loro proprietà condivise (non quelle dei loro ospiti) e al tipo di acido nucleico che forma i loro genomi.[102] In seguito l’International Committee on Taxonomy of Viruses è stato costituito. Tuttavia, i virus non sono stati classificati sulla base del phylum o sulla classe, poiché con le piccole dimensioni del genoma e l’alto tasso di mutazione, rende difficile determinare la loro ascendenza sopra l’ordine. Così la classificazione di Baltimore è utilizzata per integrare la gerarchia più tradizionale.
L’International Committee on Taxonomy of Viruses (ICTV) ha sviluppato l’attuale sistema di classificazione e ha formulato le linee guida che attribuiscono un peso maggiore ad alcune proprietà virali per mantenere l’uniformità della famiglia. È stata istituita un tassonomia unificata (un sistema universale di classificazione per i virus). Il rapporto lCTV 9[103] definisce il concetto di specie virale come il gruppo più basso nella gerarchia della ramificazione dei virus.[104] Tuttavia, attualmente solo una piccola parte dei virus è stata studiata, con analisi di campioni provenienti da esseri umani che hanno dimostrato che il 20% circa delle sequenze virali recuperate non sono mai state viste prima, mentre campioni provenienti dall’ambiente, come dall’acqua di mare e sedimenti oceanici hanno rivelato la presenza di una grande maggioranza di sequenze totalmente nuove.[105] La struttura tassonomica generale è la seguente:
Ordine (-virales)
Famiglia (-viridae)
Sottofamiglia (-virinae)
Genere (-virus)
Specie (-virus)
Nell’attuale (2019) tassonomia ICTV, sono stati istituiti 7 ordini: Caudovirales, Herpesvirales, Ligamenvirales, Mononegavirales, Nidovirales, Picornavirales e Tymovirales. Il comitato non distingue formalmente tra sottospecie, varietà e isolati. In totale vi sono 7 ordini, 103 famiglie, 22 sottofamiglie, 455 generi, circa 2.827 specie e oltre 4.000 tipi non ancora classificati.[106][107]
Il biologo premio Nobel David Baltimore ha ideato il sistema di classificazione di Baltimore.[108] La moderna classificazione dei virus utilizza una combinazione del sistema ICTV e del sistema di Baltimore.[109][110][111]
La classificazione dei virus di Baltimore è basato sul meccanismo di produzione di mRNA. I virus devono generare mRNA dai loro genomi per produrre proteine e replicarsi, ma diversi meccanismi sono utilizzati per raggiungere questo obiettivo in ogni famiglia di virus. I genomi virali possono essere a singolo filamento (ss) o doppio filamento (ds), RNA o DNA, e possono o non possono utilizzare la trascrittasi inversa (RT).
Inoltre, i virus ssRNA possono essere sia senso (+) sia antisenso (-). Questa classificazione pone i virus in sette gruppi:
I: virus dsDNA (Papillomaviridae, Herpesviridae, Poxviridae)
II: virus ssDNA (“senso” +) DNA (ad esempio, Parvovirus)
III: virus dsRNA (ad esempio Reoviridae)
IV: virus (+)ssRNA (senso +) RNA (es Picornaviridae, Togaviridae)
V: virus (−)ssRNA (antisenso -) RNA (es Hepadnaviridae, Rhabdoviridae)
VI: virus ssRNA-RT (senso +) di RNA con intermedio DNA nel ciclo di vita (ad esempio Retrovirus)
VII: virus dsDNA-RT (ad esempio Hepadnaviridae)
Come esempio di classificazione virale, il virus della varicella, il varicella zoster (VZV), appartiene all’ordine Herpesvirales, famiglia Herpesviridae, sottofamiglia Alphaherpesvirinae e genere Varicellovirus. VZV è nel gruppo I della classificazione Baltimore perché è un virus dsDNA che non utilizza la trascrittasi inversa.
Esempi di comuni malattie umane causate da virus includono il raffreddore comune, l’influenza, la varicella e l’herpes labiale. Vi sono anche gravi condizioni, come l’AIDS, la malattia da virus Ebola, l’influenza aviaria e la SARS, che sono causate da virus. La capacità relativa di un virus di causare una malattia è descritta in termini della sua virulenza. Altre patologie sono tutt’oggi indagate per scoprire se hanno un virus come agente causale, come ad esempio se vi sia una possibile connessione tra l’herpesvirus umano 6 (HHV-6) e malattie neurologiche come la sclerosi multipla e la sindrome da affaticamento cronico.[112] Vi sono accesi dibattiti sulla possibilità che il bornavirus, ritenuto causa di malattie neurologiche nei cavalli, possa essere anche responsabile per alcune malattie psichiatriche negli esseri umani.[113]
I virus hanno differenti meccanismi per cui causano una malattia in un organismo, i quali dipendono in larga misura dalla specie virale. Meccanismi a livello cellulare comprendono principalmente la lisi cellulare, che comporta la rottura della cellula e la sua conseguente morte. Negli organismi pluricellulari, se un numero abbastanza consistente di cellule muoiono, l’intero organismo incomincerà a subirne gli effetti. Sebbene i virus causino la perdita dell’omeostasi, con conseguente insorgenza di una malattia, possono esisterne di relativamente innocui all’interno di un organismo. Un esempio potrebbe includere la capacità del virus dell’herpes simplex, responsabile dell’herpes labiale, di rimanere in uno stato inattivo nel corpo umano, una condizione denominata di “latenza”[114] che è una caratteristica dei virus herpes, tra cui il virus di Epstein-Barr, che provoca la febbre ghiandolare, e del virus della varicella-zoster, causa della varicella e dell’herpes zoster. La maggior parte delle persone al mondo, nella loro vita, sono state infettate con almeno uno di questi tipi di herpes virus.[115] Tuttavia, questi virus latenti possono talvolta essere utili, infatti la presenza del virus può aumentare l’immunità contro alcuni batteri patogeni, come lo Yersinia pestis.[116]
Alcuni virus possono causare un’infezione permanente o cronica, in cui i virus continuano a replicarsi nel corpo, nonostante i meccanismi di difesa dell’ospite.[117] Questo è comune nei virus dell’epatite B e nelle infezioni da virus dell’epatite C. Le persone con una infezione cronica sono note come vettori, in quanto fungono da serbatoi del virus infettivo.[118] Nelle popolazioni con un’alta percentuale di portatori, la malattia si dice che sia endemica.[119]
L’epidemiologia virale è una branca della scienza medica che si occupa di studiare la trasmissione e controllare le infezioni da virus negli esseri umani. La trasmissione di virus può essere verticale, cioè da madre a figlio, o orizzontale, il che significa che da persona a persona. Esempi di trasmissione verticale comprendono il virus dell’epatite B e l’HIV, in cui il bambino nasce già infettato con il virus.[120] Un altro, più raro, esempio è il virus della varicella zoster, che, pur causando infezioni relativamente lievi negli esseri umani, può essere fatale per il feto e il neonato.[121]
La trasmissione orizzontale è il meccanismo più comune di diffusione del virus nelle popolazioni. La trasmissione può verificarsi quando vi è scambio di fluidi corporei durante un rapporto sessuale, come nel caso dell’HIV; o quando vi è contaminazione di sangue infetto come durante una trasfusione o con lo scambio di aghi, come spesso si riscontra nella trasmissione dell’epatite C; lo scambio di saliva dalla bocca, per esempio può portare alla trasmissione del virus di Epstein-Barr; il norovirus può trasmettersi con l’ingestione di cibo o acqua contaminati; il virus dell’influenza per inalazione; mentre insetti vettori, come le zanzare, trasmettono all’organismo ospite alcuni virus come quello della febbre dengue. Il tasso o la velocità di trasmissione delle infezioni virali dipende da vari fattori che includono la densità della popolazione, il numero dei soggetti suscettibili (cioè quelli che non immuni),[122] la qualità dell’assistenza sanitaria e il tempo.[123]
L’epidemiologia tenta di interrompere la catena delle infezioni nelle popolazioni durante le epidemie di malattie virali.[124] Le misure di controllo utilizzate si basano sulla conoscenza di come il virus si trasmette. È fondamentale trovare la fonte, o le fonti, del focolaio, in modo da identificare il virus. Una volta che è stato identificato, la catena di trasmissione può talvolta essere interrotta grazie a vaccini. Quando essi non sono disponibili, misure di igiene e la disinfezione possono essere rimedi efficaci. Spesso, le persone infette vengono isolate dal resto della comunità e quelli che sono stati esposti al virus vengono posti in quarantena.[125] Per controllare l’epidemia di afta epizootica nel bestiame avvenuta nel 2001 in Gran Bretagna, migliaia di bovini sono stati abbattuti.[126] Le infezioni virali degli esseri umani e degli altri animali hanno periodi di incubazione durante i quali l’infezione provoca segni o sintomi.[127] I periodi di incubazione per le malattie virali variano da pochi giorni a settimane, ma sono noti per la maggior parte delle infezioni.[127] Con una piccola sovrapposizione, al termine del periodo di incubazione vi è il periodo di contagiosità, in cui un individuo infetto può trasmettere la malattia a un altro.[127] Anche questo periodo è noto per molte infezioni virali e la conoscenza della durata di entrambi i periodi è importante nel controllo dei focolai.[128] Quando un focolaio provoca una percentuale insolitamente alta di casi in una popolazione o in una comunità o una regione, questi casi prendono il nome di epidemie. Se i focolai sono geograficamente distribuiti, essi sono chiamati pandemie.[129]
Le popolazioni dei nativi americani furono decimate dalle malattie contagiose, in particolare dal vaiolo, portate dai coloni europei in America. Non è chiaro quanti siano deceduti a causa delle malattie importare dagli stranieri dopo l’arrivo di Colombo, ma le stime indicano un possibile 70% della popolazione indigena.[senza fonte] Tutto ciò ha contribuito in modo significativo ai tentativi europei di spostare e sottomettere la popolazione nativa.[130]
Immagine al microscopio elettronico del virus dell’influenza A sottotipo H1N1 (ricreato in laboratorio), “virus della spagnola”
Una pandemia è un’epidemia mondiale. La pandemia influenzale del 1918 (chiamata “influenza spagnola”) che durò fino al 1919, fu catalogata come pandemia di livello 5, causata da un virus influenzale particolarmente aggressivo e mortale. Le vittime furono spesso giovani adulti sani, in contrasto con la maggior parte dei focolai di influenza che colpiscono prevalentemente i pazienti pediatrici, gli anziani o gli individui già indeboliti.[131] Le stime più datate indicano che questa pandemia abbia ucciso tra le 40 e le 50 milioni di persone,[132] mentre ricerche più recenti suggeriscono che tale dato potrebbe arrivare fino a 100 milioni di persone, il 5% della popolazione mondiale del 1918.[133]
La maggior parte dei ricercatori ritiene che l’origine del virus HIV sia nell’Africa sub-sahariana e che sia nato nel corso del XX secolo.[134] Anche la sua diffusione è oramai considerata una pandemia, con una stima di 38,6 milioni di persone infettate in tutto il mondo.[135] Le stime del Programma delle Nazioni Unite per l’AIDS/HIV(UNAIDS) e dell’Organizzazione Mondiale della Sanità (OMS) indicando che l’AIDS (la condizione dovuta al virus HIV) abbia ucciso più di 25 milioni di persone dal 5 giugno 1981, quando è stato riconosciuto, rendendola una delle epidemie più devastanti della storia.[136] Nel 2007 sono state stimate 2,7 milioni di nuove infezioni da HIV e 2 milioni di decessi correlati alla condizione.[137]
Diversi agenti patogeni virali altamente letali sono membri dei Filoviridae. I filovirus sono virus simili a filamenti che causano febbre emorragica, e comprendono l’Ebolavirus e il Marburgvirus. Il virus di Marburg, scoperto nel 1967, ha attirato l’attenzione dei media nel mese di aprile 2005 per un focolaio in Angola.[138] Il virus della malattia da virus Ebola ha causato epidemie intermittenti con alti tassi di mortalità, dal 1976 quando è stato identificato. Quella peggiore e più recente è stata l’epidemia di in Africa Occidentale del 2014.[139]
Si è stabilito che i virus siano una causa di tumore negli esseri umani e in altre specie. Tuttavia, i tumori virali si verificano solo in una minoranza di persone (o animali) infette. I virus causa di tumore possono far parte di diverse famiglie di virus, compresi i virus a RNA e DNA e quindi non esiste un unico tipo di “oncovirus” (un termine ormai obsoleto). Lo sviluppo della neoplasia è determinato da una serie di fattori come ad esempio l’immunità dell’ospite[140] e le sue mutazioni.[141] I virus ritenuti in grado di provocare tumori negli umani comprendono alcuni genotipi di papillomavirus umano, il virus dell’epatite B, il virus dell’epatite C, il virus di Epstein-Barr, l’herpesvirus associato al sarcoma di Kaposi e il virus T-linfotropico dell’uomo. Il virus causa di tumore umano più recentemente scoperto è un poliomavirus (poliomavirus delle cellule di Merkel) che causa la maggior parte dei casi di una rara forma di tumore della pelle, chiamato carcinoma a cellule di Merkel.[142]
Papilloma Virus umano
I virus dell’epatite possono sviluppare una infezione virale cronica che può portare al tumore del fegato.[143][144] L’infezione da virus T-linfotropico dell’uomo può portare a paraparesi spastica tropicale e la leucemia a cellule T dell’adulto.[145] I papillomavirus umani sono una causa consolidata di tumori della cervice uterina, della pelle, dell’ano e del pene.[146] Gli Herpesvirus umano 8 provoca il sarcoma di Kaposi, mentre il virus di Epstein-Barr causa il linfoma di Burkitt, il linfoma di Hodgkin, disordini linfoproliferativi delle cellule B e il carcinoma nasofaringeo.[147] Il poliomavirus delle cellule di Merkel è strettamente correlato al Simian virus 40 e al poliomavirus del topo; entrambi vengono utilizzati, da oltre 50 anni, come modelli animali per i virus tumorali.[148]
La prima linea di difesa dell’organismo contro i virus è il sistema immunitario innato. Questo comprende le cellule e gli altri meccanismi che difendono l’ospite da infezioni in modo non specifico. Ciò significa che le cellule del sistema innato riconoscono e rispondono agli agenti patogeni in modo generico ma, a differenza del sistema immunitario adattativo, esso non conferisce un’immunità duratura.[149]
L’interferenza dell’RNA è un importante difesa innata contro i virus.[150] Molti virus adottano una strategia di replica che coinvolge l’RNA a doppio filamento (dsRNA). Quando un virus infetta una cellula, rilascia la sua molecola di RNA che si lega immediatamente a un complesso proteico chiamato ribonucleasi III che taglia l’RNA in pezzi più piccoli. Un percorso biochimico, il complesso RISC, si attiva assicurando la sopravvivenza cellulare degradando l’mRNA virale. I rotavirus si sono evoluti per evitare questo meccanismo di difesa evitando di tagliare completamente l’RNA all’interno della cellula ma rilasciando una nuova produzione di mRNA attraverso i pori del capside interno della particella. In questo modo il loro dsRNA genomico resta protetto all’interno del nucleo del virione.[151][152]
Quando il sistema immunitario adattativo di un vertebrato incontra un virus, produce anticorpi specifici che si legano a esso e spesso sono in grado di renderlo non infettivo. Ciò è chiamata immunità umorale. Due tipi di anticorpi sono importanti. I primi, chiamati IgM, sono altamente efficaci nel neutralizzare i virus, ma vengono prodotti dalle cellule del sistema immunitario solo per poche settimane. I secondi, chiamati IgG, vengono prodotti di continuo. La presenza di IgM nel sangue dell’ospite viene utilizzato per diagnosticare la presenza di una infezione acuta, mentre le IgG indicano un’infezione passata.[153] Quando i test di immunità vengono effettuati, vengono misurati gli anticorpi IgG.[154]
Gli anticorpi possono continuare a essere un meccanismo di difesa efficace anche dopo che i virus sono riusciti a ottenere l’ingresso nella cellula ospite. Una proteina presente nelle cellule, chiamata TRIM21, è in grado di legarsi agli anticorpi sulla superficie della particella virale. Questo innesca la successiva distruzione del virus da parte degli enzimi del proteosoma della cellula.[155]
Una seconda difesa dei vertebrati contro i virus, si chiama immunità cellulo-mediata e coinvolge le cellule immunitarie conosciute come cellule T. Le cellule del corpo mostrano costantemente brevi frammenti delle loro proteine sulla superficie della cellula e se una cellula T riconosce un frammento virale sospetto, la cellula ospite viene distrutta dalle cellule T killer e le cellule T virus-specifiche proliferano. Le cellule, come i macrofagi sono specialisti in questa presentazione dell’antigene.[156] La produzione di interferone è anch’esso un importante meccanismo di difesa dell’ospite. Questo è un ormone prodotto dal corpo quando vi è la presenza di virus. Il suo ruolo nel sistema immunitario è complesso; ma sostanzialmente mette fine alla riproduzione del virus, uccidendo la cellula infetta e le sue vicine.[157]
Virus del HIV mentre entra nelle cellule.
Non tutte le infezioni da virus producono una risposta immunitaria protettiva in questo modo. L’HIV elude il sistema immunitario cambiando continuamente la sequenza aminoacidica delle proteine sulla superficie del virione. Questa tecnica è nota come “mutazione di fuga”. Questi virus persistono nell’eludere il sistema immunitario, attraverso il sequestro e blocco della presentazione dell’antigene, la resistenza alle citochine, l’evasione delle attività delle cellule natural killer, evitare l’apoptosi e la sostituzione antigenica.[158] Altri virus, chiamati virus neurotropi, sfruttano la diffusione neurale in cui il sistema immunitario potrebbe non essere in grado di raggiungerli.
Poiché i virus utilizzano vie metaboliche vitali all’interno cellule ospiti per replicarsi, essi risultano difficili da eliminare senza usare farmaci che causino effetti tossici alle cellule dell’ospite. Gli approcci medici più efficaci per le malattie virali sono le vaccinazioni, in grado di fornire l’immunità alle infezioni, e i farmaci antivirali che interferiscono selettivamente con la replicazione del virus.
Nota: non posso di certo essere definito un “Full-Vax” ma nemmeno un folle riduzionista opposto “No-Vax”. Sono piuttosto uno “Smart-Vax”.
La vaccinazione è un modo economico ed efficace per prevenire le infezioni da virus. I vaccini sono stati usati per prevenire le infezioni virali molto prima della scoperta dei virus reali. Il loro utilizzo ha portato a un drastico calo della morbilità (l’ammalarsi) a alla mortalità (morte) associata a infezioni virali, come la poliomielite, il morbillo, la parotite e la rosolia.[159] Le infezioni di vaiolo sono state debellate.[160] A oggi, 2015, vi è la disponibilità di vaccini per oltre tredici infezioni virali che colpiscono gli esseri umani,[161] e molti altri vengono utilizzati per prevenire le infezioni virali degli animali.[162]
I vaccini possono essere costituiti da virus vivi, attenuati o morti, o da proteine virali (antigeni).[163] Le forme di vaccini vivi, contengono virus indeboliti che non causano la malattia ma, ciò nonostante, conferiscono l’immunità. I vaccini vivi possono essere pericolosi quando somministrati a persone con una debole immunità (individui immunocompromessi) poiché in queste persone il virus indebolito può provocare la malattia originale.[164] La biotecnologia e le tecniche di ingegneria genetica vengono utilizzate per la produzione di vaccini di subunità. Questi vaccini utilizzano solo le proteine del capside del virus. Il vaccino contro l’epatite B è un esempio di questo tipo di vaccino.[165] I vaccini di subunità sono sicuri per i pazienti immunocompromessi perché non possono in nessun modo provocare la malattia.[166] Il vaccino contro il virus della febbre gialla, un ceppo vivo attenuato e chiamato 17D, è probabilmente il più sicuro e il più efficace vaccino mai prodotto.[167]
Un mezzo di contrasto all’infezione virale è rappresentatpo dai farmaci antivirali. Essi sono spesso analoghi nucleosidici (costruzioni di DNA falso), che i virus erroneamente incorporano nei loro genomi durante la replicazione. Il ciclo di vita del virus viene bloccata perché il DNA di nuova sintesi risulta inattivo. Infatti in questi analoghi mancano i gruppi ossidrilici, che, insieme con gli atomi di fosforo, si collegano insieme per formare il la forte “spina dorsale” della molecola di DNA. Ciò prende il nome di interruzione della catena del DNA.[168] Esempi di analoghi nucleosidici sono l’aciclovir per il Virus dell’herpes simplex e la lamivudina per le infezioni da virus HIV e epatite B. L’aciclovir è uno dei più antichi farmaci antivirali e più frequentemente prescritti.[169] Altri farmaci antivirali utilizzano come bersaglio diverse fasi del ciclo di vita del virus. L’HIV dipende da un enzima proteolitico chiamato HIV-1 proteasi per farlo diventare completamente infettivo. Vi è un vasto gruppo di farmaci, chiamati inibitori della proteasi, che inattivano questo enzima.
L’epatite C è causata da un virus a RNA. Nel 80% delle persone infette la malattia risulta cronica e in assenza di trattamento restano infetti per tutta la loro vita. Tuttavia, vi è ora un trattamento efficace che utilizza un analogo nucleosidico, la ribavirina, combinato con l’interferone.[170] Il trattamento dei portatori cronici del virus dell’epatite B utilizzando una strategia simile utilizzando la lamivudina.[171]
Nota: i virus possono essere considerati regolatori delle forme biologiche “semplici” e “complesse”.
Un virus batteriofago regola il microbiota intestinale.
Una caratteristica dei virus, che la maggior parte delle persone non conosce, è che molti di essi hanno azione benefica. Lo ammetto, ogni volta che viene detto per molti è un vero è oroprio shock, ma in effetti le cose stanno così: la maggior parte dei virus ha azione benefica. Provate solo a considerare il fatto che vi sono più virus nell’intestino che batteri! In effetti, la popolazione virale (chiamata “viroma”[172]) svolge un ruolo importante nella regolazione del numero e dei tipi di batteri nel corpo.[173] Senza di essi, potremmo essere rapidamente consumati dai piccoli batteri affamati che vivono nel nostro intestino.
Avete presente quando nuotate nel mare? Ecco, è un po’ come nuotare in una zuppa batterica altamente concentrata. Ci sono molti batteri nell’acqua del mare, di molte specie diverse. Ma, come nell’intestino, ci sono più virus che batteri e probabilmente i primi svolgono un ruolo nel mantenere e bilanciare la popolazione batterica nelle acque marine. Alcuni si pongono se vi sarebbe vita negli oceani/mari se non vi fossero virus. Ed è questo il motivo per il quale in precedenza ho denominato i virus come “agenti regolatori” che influenzano altre entità biologiche.
L’esempio della “zuppa batterico-virale” può essere applicata anche ad un lago. Infatti, in esso possono aver nuotato anatre, cigni o oche. Di conseguenza vi si possono trovare virus dell’influenza. In effetti, gli uccelli acquatici trasportano tutti i possibili tipi di virus influenzale, compresi quelli che non infettano l’uomo. Questi virus vengono introdotti nell’acqua quando gli uccelli defecano. Ma la presenza del virus di solito non produce malattie in questi uccelli, [174] o in un soggetto umano, anche se entrano in contatto con occhi, orecchie, naso e bocca. Un evoluzionista potrebbe dire che il motivo per cui gli uccelli non si ammalano (di solito) è perché la convivenza prolungata nel tempo del ceppo virale e dell’animale ha portato ad una condizione collaborativa, dove in virus usa l’ospite semplicemente come mezzo per sopravvivere senza causarne malessere o il decesso. Dal punto di vista dell’intelligence design , il virus dell’influenza ha probabilmente un ruolo benefico per gli uccelli, oppure ha semplicemente subito mutazioni degenerative che lo rendono controllabile dall’organismo ospite in questione.
Dal momento che le nostre cellule posseggono molte delle stesse cose di cui sono composti i virus (rivestimenti proteici, DNA e RNA, disponiamo di meccanismi per spostare il DNA in diverse parti del genoma, ecc), alcuni virus, come già accennato, potrebbero aver avuto origine dalle normali operazioni cellulari.[175] Le componenti sono tutte lì, a volte esse sono assemblate in cose che sembrano quasi virus. Basterebbero alcune modifiche non ben calibrate e l’assemblaggio potrebbe sfuggire di mano e “diventare virale”.
Ma non tutti i virus sono genoma-simili. Molti virus che causano malattie sembrano progettati per fare ciò. Quindi, la domanda che sorge spontanea è: da dove vengono ? Bene, se esiste un virus progettato per infettare un batterio, una cellula di topo o di una persona, probabilmente ci sono controlli e equilibri in quel sistema. Se uno di questi controlli fallisce, il virus potrebbe essere in grado di riprodursi molto più velocemente di quanto originariamente era stato progettato di fare. Ciò comporterebbe una malattia. Pertanto, un virus “benefico” potrebbe essere in grado di mutare in un virus pericoloso, sempre un virus ma con effetti differenti per mutazione. Potrebbero essere necessarie alcune piccole mutazioni, come forse un cambiamento in un fattore di riconoscimento cellulare che impedisce alle cellule ospiti di rilevare e quindi regolare il virus.
Adesso che abbiamo sufficienti e reali informazioni sui Virus possiamo iniziare a trattare l’argomento Covid-19 per comprendere la prima epidemia virale mondiale del XXI Secolo, caratteristiche del patogeno e cosa realmente possiamo fare per prevenirne il contagio e quali sono le attuali possibilità di trattamento degli infetti.
Continua…
Gabriel Bellizzi
Riferimenti:
1- Koonin EV, Senkevich TG, Dolja VV, The ancient Virus World and evolution of cells, in Biol. Direct, vol. 1, 2006, p. 29.
2- Bernard La Scola, Christelle Desnues e Isabelle Pagnier, The virophage as a unique parasite of the giant mimivirus, in Nature, vol. 455, nº 7209, pp. 100–104.
3- Ed Yong, The virophage – a virus that infects other viruses, su National Geographic – Phenomena, 8 luglio 2008. URL consultato il 12 marzo 2016.
4- Nigel J. Dimmock, Andrew J. Easton e Keith N. Leppard, Introduction to Modern Virology, VI, Blackwell Publishing, 2007.
5- Bordenave G, Louis Pasteur (1822–1895), in Microbes and Infection / Institut Pasteur, vol. 5, nº 6, 2003, pp. 553–60.
6- eri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
7- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998.
8- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
9- Nigel J. Dimmock, Andrew J. Easton e Keith N. Leppard, Introduction to Modern Virology, VI, Blackwell Publishing, 2007.
10- F. Fenner, History of Virology: Vertebrate Viruses, in Brian W. J. Mahy e Marc H. V. Van Regenmortel (a cura di), Desk Encyclopedia of General Virology, 1ª ed., Oxford, UK, Academic Press, 2009, pp. 15-19.
11- Steinhardt E, Israeli C, Lambert R.A., Studies on the cultivation of the virus of vaccinia, in J. Inf Dis., vol. 13, nº 2, 1913, pp. 294–300.
12- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998.
13- Goodpasture EW, Woodruff AM, Buddingh GJ, The cultivation of vaccine and other viruses in the chorioallantoic membrane of chick embryos, in Science, vol. 74, nº 1919, 1931, pp. 371–372.
14- Rosen, FS, Isolation of poliovirus—John Enders and the Nobel Prize, in New England Journal of Medicine, vol. 351, nº 15, 2004, pp. 1481–83.
15- From Nobel Lectures, Physics 1981–1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co., Singapore.
In 1887, Buist visualised one of the largest, Vaccinia virus, by optical microscopy after staining it. Vaccinia was not known to be a virus at that time. (Buist J.B. Vaccinia and Variola: a study of their life history Churchill, London).
16- Stanley WM, Loring HS, The isolation of crystalline tobacco mosaic virus protein from diseased tomato plants, in Science, vol. 83, nº 2143, 1936, p. 85.
17- Stanley WM, Lauffer MA, Disintegration of tobacco mosaic virus in urea solutions, in Science, vol. 89, nº 2311, 1939, pp. 345–347.
18- Creager AN, Morgan GJ, After the double helix: Rosalind Franklin’s research on Tobacco mosaic virus, in Isis, vol. 99, nº 2, 2008, pp. 239–72.
19- Nigel J. Dimmock, Andrew J. Easton e Keith N. Leppard, Introduction to Modern Virology, VI, Blackwell Publishing, 2007.
20- Norrby E, Nobel Prizes and the emerging virus concept, in Arch. Virol., vol. 153, nº 6, 2008, pp. 1109–23.
21- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998, ISBN 0-340-66316-2.
22- Temin HM, Baltimore D, RNA-directed DNA synthesis and RNA tumor viruses, in Adv. Virus Res., vol. 17, 1972, pp. 129–86.
23- Barré-Sinoussi, F. et al., Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS), in Science, vol. 220, nº 4599, 1983, pp. 868–871.
24- Sanjuán R, Nebot MR, Chirico N, Mansky LM, Belshaw R, Viral mutation rates, in Journal of Virology, vol. 84, nº 19, October 2010, pp. 9733–48.
25- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
26- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998.
27- Nigel J. Dimmock, Andrew J. Easton e Keith N. Leppard, Introduction to Modern Virology, VI, Blackwell Publishing, 2007.
28- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998.
29- Mahy WJ & Van Regenmortel MHV (eds), Desk Encyclopedia of General Virology, Oxford, Academic Press, 2009, pp. 24.
30- Leslie Collier, Albert Balows e Max Sussman, Virology, in Brian H Mahy e Leslie Collier (a cura di), Topley and Wilson’s Microbiology and Microbial Infections, vol. 1, 9ª ed., Arnold, 1998.
31- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
32- B. McClintock, The origin and behavior of mutable loci in maize, in Proc Natl Acad Sci U S A., vol. 36, nº 6, 1950, pp. 344–55.
33- Nigel J. Dimmock, Andrew J. Easton e Keith N. Leppard, Introduction to Modern Virology, VI, Blackwell Publishing, 2007.
34- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
35- Tsagris EM, de Alba AE, Gozmanova M, Kalantidis K, Viroids, in Cell. Microbiol., vol. 10, nº 11, 2008, pp. 2168.
36- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
37- Tsagris EM, de Alba AE, Gozmanova M, Kalantidis K, Viroids, in Cell. Microbiol., vol. 10, nº 11, 2008, pp. 2168,
38- Teri Shors, Understanding Viruses, Jones and Bartlett Publishers, 2008.
39- a Scola B, Desnues C, Pagnier I, Robert C, Barrassi L, Fournous G, Merchat M, Suzan-Monti M, Forterre P, Koonin E, Raoult D, The virophage as a unique parasite of the giant mimivirus, in Nature, vol. 455, nº 7209, 2008, pp. 100–4.
40- Mahy WJ & Van Regenmortel MHV (eds), Desk Encyclopedia of General Virology, Oxford, Academic Press, 2009, pp. 28.
41- Mahy WJ & Van Regenmortel MHV (eds), Desk Encyclopedia of General Virology, Oxford, Academic Press, 2009, pp. 26.
42- Dimmock, pp. 15-16.
43- Holmes EC, Viral evolution in the genomic age, in PLoS Biol., vol. 5, nº 10, 2007, pp. e278.
44- Wimmer E, Mueller S, Tumpey TM, Taubenberger JK, Synthetic viruses: a new opportunity to understand and prevent viral disease, in Nature Biotechnology, vol. 27, nº 12, 2009, pp. 1163–72.
45- Horn M, Chlamydiae as symbionts in eukaryotes, in Annual Review of Microbiology, vol. 62, 2008, pp. 113–31.
46- Collier, pp. 33-55.
47- Collier, pp. 33-37.
48- Collier, p. 40.
49- Caspar DL, Klug A, Physical principles in the construction of regular viruses, in Cold Spring Harb. Symp. Quant. Biol., vol. 27, 1962, pp. 1–24.
50- Crick FH, Watson JD, Structure of small viruses, in Nature, vol. 177, nº 4506, 1956, pp. 473–5.
51- M.R. Falvo, S. Washburn, R. Superfine, M. Finch, F.P. Brooks Jr, V. Chi, R.M. Taylor 2nd, Manipulation of individual viruses: friction and mechanical properties, in Biophysical Journal, vol. 72, nº 3, 1997, pp. 1396–1403.
52- Yu. G. Kuznetsov, A. J. Malkin, R. W. Lucas, M. Plomp, A. McPherson, Imaging of viruses by atomic force microscopy, in J Gen Virol, vol. 82, nº 9, 2001, pp. 2025–2034.
53- Collier, p. 37.
54- Collier, pp. 40, 42.
55- Casens, S., Desk Encyclopedia of General Virology, Boston, Academic Press, 2009, pp. 167–174.
56- Collier, pp. 42-43.
57- Rossmann MG, Mesyanzhinov VV, Arisaka F, Leiman PG, The bacteriophage T4 DNA injection machine, in Current Opinion in Structural Biology, vol. 14, nº 2, 2004, pp. 171–80.
58- Long GW, Nobel J, Murphy FA, Herrmann KL, Lourie B, Experience with electron microscopy in the differential diagnosis of smallpox, in Appl Microbiol, vol. 20, nº 3, 1970, pp. 497–504.
59- NCBI Viral Genome database, su ncbi.nlm.nih.gov.
60- Pennisi E, Microbiology. Going viral: exploring the role of viruses in our bodies [collegamento interrotto], in Science, vol. 331, nº 6024, 2011, pp. 1513.
61- Collier, pp. 96-99.
62- Carter, p. 72.
63- Belyi VA, Levine AJ e Skalka AM, Sequences from ancestral single-stranded DNA viruses in vertebrate genomes: the parvoviridae and circoviridae are more than 40 to 50 million years old, in J. Virol., vol. 84, nº 23, 2010, pp. 12458–62.
64- Philippe N, Legendre M, Doutre G, Couté Y, Poirot O, Lescot M, Arslan D, Seltzer V, Bertaux L, Bruley C, Garin J, Claverie JM e Abergel C, Pandoraviruses: amoeba viruses with genomes up to 2.5 Mb reaching that of parasitic eukaryotes, in Science, vol. 341, nº 6143, 2013, pp. 281–6.
65- Pressing J, Reanney DC, Divided genomes and intrinsic noise, in J Mol Evol, vol. 20, nº 2, 1984, pp. 135–46.
66- Duffy S, Holmes EC, Validation of high rates of nucleotide substitution in geminiviruses: phylogenetic evidence from East African cassava mosaic viruses, in The Journal of General Virology, vol. 90, Pt 6, 2009, pp. 1539–47.
67- Sandbulte MR, Westgeest KB, Gao J, Xu X, Klimov AI, Russell CA, Burke DF, Smith DJ, Fouchier RA, Eichelberger MC, Discordant antigenic drift of neuraminidase and hemagglutinin in H1N1 and H3N2 influenza viruses, in Proceedings of the National Academy of Sciences of the United States of America, vol. 108, nº 51, December 2011, pp. 20748–53.
68- Moss RB, Davey RT, Steigbigel RT, Fang F, Targeting pandemic influenza: a primer on influenza antivirals and drug resistance, in The Journal of Antimicrobial Chemotherapy, vol. 65, nº 6, June 2010, pp. 1086–93.
69- Hampson AW, Mackenzie JS, The influenza viruses, in Med. J. Aust., vol. 185, 10 Suppl, 2006, pp. S39–43.
70- Metzner KJ, Detection and significance of minority quasispecies of drug-resistant HIV-1, in J HIV Ther, vol. 11, nº 4, 2006, pp. 74–81.
71- Goudsmit.
72- Worobey M, Holmes EC, Evolutionary aspects of recombination in RNA viruses, in J. Gen. Virol., 80 ( Pt 10), 1999, pp. 2535–43.
73- Lukashev AN, Role of recombination in evolution of enteroviruses, in Rev. Med. Virol., vol. 15, nº 3, 2005, pp. 157–67.
74- Umene K, <171::AID-RMV243>3.0.CO;2-A Mechanism and application of genetic recombination in herpesviruses, in Rev. Med. Virol., vol. 9, nº 3, 1999, pp. 171–82.
75- Collier, pp. 75-91.
76- Dimmock, p. 70.
77- Boevink P, Oparka KJ, Virus-host interactions during movement processes, in Plant Physiol., vol. 138, nº 4, 2005, pp. 1815–21.
78- Dimmock, p. 71.
79- Barman S, Ali A, Hui EK, Adhikary L, Nayak DP, Transport of viral proteins to the apical membranes and interaction of matrix protein with glycoproteins in the assembly of influenza viruses, in Virus Res., vol. 77, nº 1, 2001, pp. 61–9.
80- Shors, pp. 60, 597.
81- Dimmock, Capitolo 15, Mechanisms in virus latentcy, pp.243–259.
82- Dimmock, pp. 185-187.
83- Shors, p. 54; Collier, p. 78.
84- Collier, p. 79.
85- Staginnus C, Richert-Pöggeler KR, Endogenous pararetroviruses: two-faced travelers in the plant genome, in Trends in Plant Science, vol. 11, nº 10, 2006, pp. 485–91.
86- Collier, pp. 88-89.
87- Collier, pp. 115-146.
88- Collier, p. 115.
89- Roulston A, Marcellus RC, Branton PE, Viruses and apoptosis, in Annu. Rev. Microbiol., vol. 53, 1999, pp. 577–628.
90- Alwine JC, Modulation of host cell stress responses by human cytomegalovirus, in Curr. Top. Microbiol. Immunol., vol. 325, 2008, pp. 263–79.
91- Sinclair J, Human cytomegalovirus: Latency and reactivation in the myeloid lineage, in J. Clin. Virol., vol. 41, nº 3, 2008, pp. 180–5.
92- Jordan MC, Jordan GW, Stevens JG, Miller G, Latent herpesviruses of humans, in Annals of Internal Medicine, vol. 100, nº 6, 1984, pp. 866–80.
93- Sissons JG, Bain M, Wills MR, Latency and reactivation of human cytomegalovirus, in J. Infect., vol. 44, nº 2, 2002, pp. 73–7.
94- Barozzi P, Potenza L, Riva G, Vallerini D, Quadrelli C, Bosco R, Forghieri F, Torelli G, Luppi M, B cells and herpesviruses: a model of lymphoproliferation, in Autoimmun Rev, vol. 7, nº 2, 2007, pp. 132–6.
95- Subramanya D, Grivas PD, HPV and cervical cancer: updates on an established relationship, in Postgrad Med, vol. 120, nº 4, 2008, pp. 7–13.
96- Crawford, p. 16.
97- Shors, p. 388.
98- Shors, p. 353.
99- Dimmock, p. 272.
100- Baggesen DL, Sørensen G, Nielsen EM, Wegener HC, Phage typing of Salmonella Typhimurium – is it still a useful tool for surveillance and outbreak investigation?, in Eurosurveillance, vol. 15, nº 4, 2010, pp. 19471.
101- Lwoff A, Horne RW, Tournier P, A virus system, in C. R. Hebd. Seances Acad. Sci., vol. 254, 1962, pp. 4225–7.
102- Lwoff A, Horne R, Tournier P, A system of viruses, in Cold Spring Harb. Symp. Quant. Biol., vol. 27, 1962, pp. 51–5.
103- King AMQ, Lefkowitz E, Adams MJ, Carstens EB, Virus Taxonomy: Ninth Report of the International Committee on Taxonomy of Viruses, Elsevier, 2011, p. 6.
104- MJ. Adams, EJ. Lefkowitz, AM. King e EB. Carstens, Recently agreed changes to the International Code of Virus Classification and Nomenclature., in Archives of Virology, vol. 158, nº 12, Dec 2013, pp. 2633–9.
105- As defined therein, “A species is the lowest taxonomic level in the hierarchy approved by the ICTV. A species is a monophyletic group of viruses whose properties can be distinguished from those of other species by multiple criteria.”
106- Delwart EL, Viral metagenomics, in Rev. Med. Virol., vol. 17, nº 2, 2007, pp. 115–31.
107- Virus Taxonomy 2013. International Committee on Taxonomy of Viruses. Retrieved on 24 December 2011.
108- ICTV Master Species List 2013 v2 Archiviato il 2 ottobre 2015 in Internet Archive.
This Excel file contains the official ICTV Master Species list for 2014 and lists all approved virus taxa. This is version 11 of the MSL published on 24 August 2014. Retrieved on 24 December 2014
109- Baltimore D, The strategy of RNA viruses, in Harvey Lect., 70 Series, 1974, pp. 57–74.
110- van Regenmortel MH, Mahy BW, Emerging issues in virus taxonomy, in Emerging Infect. Dis., vol. 10, nº 1, 2004, pp. 8–13.
111- Mayo MA, Developments in plant virus taxonomy since the publication of the 6th ICTV Report. International Committee on Taxonomy of Viruses, in Arch. Virol., vol. 144, nº 8, 1999, pp. 1659–66.
112- Komaroff AL, Is human herpesvirus-6 a trigger for chronic fatigue syndrome?, in J. Clin. Virol., 37 Suppl 1, 2006, pp. S39–46.
113- Chen C, Chiu Y, Wei F, Koong F, Liu H, Shaw C, Hwu H, Hsiao K, High seroprevalence of Borna virus infection in schizophrenic patients, family members and mental health workers in Taiwan, in Mol Psychiatry, vol. 4, nº 1, 1999, pp. 33–8.
114- Margolis TP, Elfman FL, Leib D, et al., Spontaneous reactivation of herpes simplex virus type 1 in latently infected murine sensory ganglia, in J. Virol., vol. 81, nº 20, 2007, pp. 11069–74.
115- Whitley RJ, Roizman B, Herpes simplex virus infections, in Lancet, vol. 357, nº 9267, 2001, pp. 1513–8.
116- Barton ES, White DW, Cathelyn JS, et al., Herpesvirus latency confers symbiotic protection from bacterial infection, in Nature, vol. 447, nº 7142, 2007, pp. 326–9, Bibcode:2007Natur.447..326B.
117- Bertoletti A, Gehring A, Immune response and tolerance during chronic hepatitis B virus infection, in Hepatol. Res., 37 Suppl 3, 2007, pp. S331–8.
118- Rodrigues C, Deshmukh M, Jacob T, Nukala R, Menon S, Mehta A, Significance of HBV DNA by PCR over serological markers of HBV in acute and chronic patients, in Indian journal of medical microbiology, vol. 19, nº 3, 2001, pp. 141–4.
119- Nguyen VT, McLaws ML, Dore GJ, Highly endemic hepatitis B infection in rural Vietnam, in Journal of Gastroenterology and Hepatology, vol. 22, nº 12, 2007, p. 2093.
120- Fowler MG, Lampe MA, Jamieson DJ, Kourtis AP, Rogers MF, Reducing the risk of mother-to-child human immunodeficiency virus transmission: past successes, current progress and challenges, and future directions, in Am. J. Obstet. Gynecol., vol. 197, 3 Suppl, 2007, pp. S3–9.
121- Sauerbrei A, Wutzler P, The congenital varicella syndrome, in Journal of perinatology : official journal of the California Perinatal Association, vol. 20, 8 Pt 1, 2000, pp. 548–54.
122- Garnett GP, Role of herd immunity in determining the effect of vaccines against sexually transmitted disease, in J. Infect. Dis., 191 Suppl 1, 2005, pp. S97–106.
123- Platonov AE, (The influence of weather conditions on the epidemiology of vector-borne diseases by the example of West Nile fever in Russia), in Vestn. Akad. Med. Nauk SSSR, nº 2, 2006, pp. 25–9.
124- Shors, p. 198.
125- Shors, pp. 199, 209.
126- Shors, p. 19.
127- Shors, pp. 193–194.
128- Shors, p. 194.
129- Shors, pp. 192–193.
130- Ranlet P, The British, the Indians, and smallpox: what actually happened at Fort Pitt in 1763?, in Pa Hist, vol. 67, nº 3, 2000, pp. 427–41.
Van Rijn K, “Lo! The poor Indian!” colonial responses to the 1862–63 smallpox epidemic in British Columbia and Vancouver Island, in Can Bull Med Hist, vol. 23, nº 2, 2006, pp. 541–60.
Patterson KB, Runge T, Smallpox and the Native American, in Am. J. Med. Sci., vol. 323, nº 4, 2002, pp. 216–22.
Sessa R, Palagiano C, Scifoni MG, di Pietro M, Del Piano M, The major epidemic infections: a gift from the Old World to the New?, in Panminerva Med, vol. 41, nº 1, 1999, pp. 78–84.
Bianchine PJ, Russo TA, The role of epidemic infectious diseases in the discovery of America, in Allergy Proc, vol. 13, nº 5, 1992, pp. 225–32, DOI:10.2500/108854192778817040, PMID 1483570. URL consultato il 19 dicembre 2014.
Hauptman LM, Smallpox and American Indian; Depopulation in Colonial New York, in N Y State J Med, vol. 79, nº 12, 1979, pp. 1945–9.
Fortuine R, Smallpox decimates the Tlingit (1787), in Alaska Med, vol. 30, nº 3, 1988, p. 109.
131- Collier, pp. 409-415.
132- Patterson KD, Pyle GF, The geography and mortality of the 1918 influenza pandemic, in Bull Hist Med., vol. 65, nº 1, 1991, pp. 4–21.
133- Johnson NP, Mueller J, Updating the accounts: global mortality of the 1918–1920 “Spanish” influenza pandemic, in Bull Hist Med, vol. 76, nº 1, 2002, pp. 105–15.
134- Gao F, Bailes E, Robertson DL, et al., Origin of HIV-1 in the Chimpanzee Pan troglodytes troglodytes, in Nature, vol. 397, nº 6718, 1999, pp. 436–441.
135- Shors, p. 447.
136- Mawar N, Saha S, Pandit A, Mahajan U, The third phase of HIV pandemic: social consequences of HIV/AIDS stigma & discrimination & future needs (PDF), in Indian J. Med. Res., vol. 122, nº 6, 2005, pp. 471–84.
137- Status of the global HIV epidemic (PDF), UNAIDS, 2008.
138- Towner JS, Khristova ML, Sealy TK, et al., Marburgvirus genomics and association with a large hemorrhagic fever outbreak in Angola, in J. Virol., vol. 80, nº 13, 2006, pp. 6497–516.
139- World Health Organisation report, 24 September 2014.
140- Einstein MH, Schiller JT, Viscidi RP, Strickler HD, Coursaget P, Tan T, Halsey N, Jenkins D, Clinician’s guide to human papillomavirus immunology: knowns and unknowns, in The Lancet Infectious Diseases, vol. 9, nº 6, 2009, pp. 347–56.
141- Shuda M, Feng H, Kwun HJ, Rosen ST, Gjoerup O, Moore PS, Chang Y, T antigen mutations are a human tumor-specific signature for Merkel cell polyomavirus, in Proceedings of the National Academy of Sciences of the United States of America, vol. 105, nº 42, 2008, pp. 16272–7.
142- Pulitzer MP, Amin BD, Busam KJ, Merkel cell carcinoma: review, in Advances in Anatomic Pathology, vol. 16, nº 3, 2009, pp. 135–44.
143- Koike K, Hepatitis C virus contributes to hepatocarcinogenesis by modulating metabolic and intracellular signalling pathways, in J. Gastroenterol. Hepatol., 22 Suppl 1, 2007, pp. S108–11.
144- Hu J, Ludgate L, HIV-HBV and HIV-HCV coinfection and liver cancer development, in Cancer Treat. Res., vol. 133, 2007, pp. 241–52.
145- Bellon M, Nicot C, Telomerase: a crucial player in HTLV-I-induced human T-cell leukemia, in Cancer genomics & proteomics, vol. 4, nº 1, 2007, pp. 21–5.
146- Schiffman M, Castle PE, Jeronimo J, Rodriguez AC, Wacholder S, Human papillomavirus and cervical cancer, in Lancet, vol. 370, nº 9590, 2007, pp. 890–907.
147- Klein E, Kis LL, Klein G, Epstein-Barr virus infection in humans: from harmless to life endangering virus-lymphocyte interactions, in Oncogene, vol. 26, nº 9, 2007, pp. 1297–305.
148- zur Hausen H, Novel human polyomaviruses—re-emergence of a well known virus family as possible human carcinogens, in International Journal of Cancer. Journal International Du Cancer, vol. 123, nº 2, 2008, pp. 247–50.
149- Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters, Molecular Biology of the Cell; Fourth Edition, New York and London, Garland Science, 2002.
150- Ding SW, Voinnet O, Antiviral immunity directed by small RNAs, in Cell, vol. 130, nº 3, 2007, pp. 413–26.
151- Patton JT, Vasquez-Del Carpio R, Spencer E, Replication and transcription of the rotavirus genome, in Curr. Pharm. Des., vol. 10, nº 30, 2004, pp. 3769–77.
152- Jayaram H, Estes MK, Prasad BV, Emerging themes in rotavirus cell entry, genome organization, transcription and replication, in Virus Res., vol. 101, nº 1, 2004, pp. 67–81.
153- Greer S, Alexander GJ, Viral serology and detection, in Baillieres Clin. Gastroenterol., vol. 9, nº 4, 1995, pp. 689–721.
154- Matter L, Kogelschatz K, Germann D, Serum levels of rubella virus antibodies indicating immunity: response to vaccination of subjects with low or undetectable antibody concentrations, in J. Infect. Dis., vol. 175, nº 4, 1997, pp. 749–55.
155- Mallery DL, McEwan WA, Bidgood SR, Towers GJ, Johnson CM, James LC, McEwan, Bidgood, Towers, Johnson e James, Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21), in Proceedings of the National Academy of Sciences of the United States of America, vol. 107, nº 46, novembre 2010, pp. 19985–90.
156- Cascalho M, Platt JL, Novel functions of B cells, in Crit. Rev. Immunol., vol. 27, nº 2, 2007, pp. 141–51.
157- Le Page C, Génin P, Baines MG, Hiscott J, Interferon activation and innate immunity, in Rev Immunogenet, vol. 2, nº 3, 2000, pp. 374–86.
158- Hilleman MR, Strategies and mechanisms for host and pathogen survival in acute and persistent viral infections, in Proc. Natl. Acad. Sci. U.S.A., 101 Suppl 2, 2004, pp. 14560–6.
159- Asaria P, MacMahon E, Measles in the United Kingdom: can we eradicate it by 2010?, in BMJ, vol. 333, nº 7574, 2006, pp. 890–5.
160- Lane JM, Mass vaccination and surveillance/containment in the eradication of smallpox, in Curr. Top. Microbiol. Immunol., vol. 304, 2006, pp. 17–29.
161- Arvin AM, Greenberg HB, New viral vaccines, in Virology, vol. 344, nº 1, 2006, pp. 240–9.
162- Pastoret PP, Schudel AA, Lombard M, Conclusions—future trends in veterinary vaccinology, in Rev. – Off. Int. Epizoot., vol. 26, nº 2, 2007, pp. 489–94, 495–501, 503–9.
163- Palese P, Making better influenza virus vaccines?, in Emerging Infect. Dis., vol. 12, nº 1, 2006, pp. 61–5.
164- Thomssen R, Live attenuated versus killed virus vaccines, in Monographs in allergy, vol. 9, 1975, pp. 155–76.
165- McLean AA, Development of vaccines against hepatitis A and hepatitis B, in Rev. Infect. Dis., vol. 8, nº 4, 1986, pp. 591–8.
166- Casswall TH, Fischler B, Vaccination of the immunocompromised child, in Expert review of vaccines, vol. 4, nº 5, 2005, pp. 725–38.
167- Barnett ED, Wilder-Smith A, Wilson ME, Yellow fever vaccines and international travelers, in Expert Rev Vaccines, vol. 7, nº 5, 2008, pp. 579–87.
168- Magden J, Kääriäinen L, Ahola T, Inhibitors of virus replication: recent developments and prospects, in Appl. Microbiol. Biotechnol., vol. 66, nº 6, 2005, pp. 612–21.
169- Mindel A, Sutherland S, Genital herpes — the disease and its treatment including intravenous acyclovir, in J. Antimicrob. Chemother., 12 Suppl B, 1983, pp. 51–9.
170- Witthöft T, Möller B, Wiedmann KH, et al., Safety, tolerability and efficacy of peginterferon alpha-2a and ribavirin in chronic hepatitis C in clinical practice: The German Open Safety Trial, in J. Viral Hepat., vol. 14, nº 11, 2007, pp. 788–96.
171- Rudin D, Shah SM, Kiss A, Wetz RV, Sottile VM, Interferon and lamivudine vs. interferon for hepatitis B e antigen-positive hepatitis B treatment: meta-analysis of randomized controlled trials, in Liver Int., vol. 27, nº 9, 2007, pp. 1185–93.
173- Francis, J.W., Ingle, M., and Wood, T.C., Bacteriophages as beneficial regulators of the mammalian Microbiome, Proc. Int. Conf. Creationism 8:152–157, 2018.
174- Barber, M.R. et al., Association of RIG-I with innate immunity of ducks to influenza, PNAS 107(13):5913–5918, 2010.
175- Terborg, P., The ‘VIGE-first hypothesis–how easy it is to swap cause and effect, J. Creation 27(3):105–112, 2013. (VIGE = Variation-Inducing Genetic Element).
La Melatonina (chimicamente N-acetil-5-metossitriptammina; formula bruta o molecolare C13H16N2O2) è un ormone prodotto dalla ghiandola pineale (o epifisi), ghiandola posta alla base del cervello. Agisce sull’ipotalamo e ha la principale funzione di regolare il ciclo sonno-veglia. Oltre che negli esseri umani, essa è prodotta da specie animali, piante (Fitomelatonina) e microorganismi.[1][2]
Nel 1917, McCord e Allen avevano notato (J. exp. zool.[3], 1917) che l’estratto delle ghiandole pineali delle mucche schiariva la pelle delle rane.[4][5] Si noti, infatti, che la Melatonina è coinvolta nel meccanismo con il quale alcuni anfibi e rettili cambiano il colore della propria pelle e pertanto fu scoperta in relazione a questo suo ruolo biologico.[6][7] Quindi, la sostanza responsabile (Melatonina) fu isolata nel 1958 dall’urina di topo, dal professore di dermatologia Aaron B. Lerner, insieme a suoi colleghi dell’Università di Yale, in ricerche mosse dall’aspettativa che la sostanza presente nella ghiandola pineale potesse essere utile nel trattamento di affezioni della pelle.[8] Questi stessi ricercatori coniarono il nome Melatonina.[9]
Verso la metà degli anni settanta, Lynch et al. dimostrarono[10] che la produzione di Melatonina da parte della ghiandola pineale umana segue un ritmo circadiano. La scoperta delle proprietà antiossidanti della Melatonina è del 1993.[11] Il primo brevetto per il suo impiego a basse dosi nel trattamento dell’insonnia fu accordato a Richard Wurtman del MIT nel 1995.[12] Nello stesso periodo la Melatonina apparve spesso sulla stampa, anche e soprattutto in quella a diffusione non settoriale, come possibile trattamento per varie malattie.[13] Famoso è l’impiego della Melatonina nel “Metodo Di Bella” insieme ad altre molecole biologiche.[14]
In questo articolo il mio intento è quello di analizzare i potenziali effetti sulla Leptina e il tessuto adiposo in generale legati alla supplementazione di Melatonina.
Leptina
Uno studio sull’uomo nel quale è stata utilizzata Melatonina integrativa e sono state fatte misurazioni della Leptina sierica, ha osservato che, in una popolazione di 11 persone con ulcere allo stomaco idiopatica, la Leptina è aumentata da 6,2-7,0ng/ml a 12,2-16,2ng/ml dopo 7 giorni, mantenendo questo livello fino a 21 giorni dopo l’assunzione di 5 mg di Melatonina due volte al giorno, mattina e sera.[15] La stessa dose (10 mg) nelle persone con epatopatia grassa non alcolica per 28 giorni con Leptina elevata ha causato un ulteriore aumento di quest’ultima del 33%.[16] Questi effetti sono stati osservati anche nei ratti a 25mcg/mL miscelati nell’acqua potabile (circa 500mcg al giorno) per 9 settimane con una dieta ad alto contenuto di grassi (35% di grassi, 35% di carboidrati) o un basso contenuto di grassi (4% di grassi, 60 % di carboidrati), in cui l’area sotto la curva della Leptina era aumentata, ma solo se misurata dalla mattina presto alla sera presto, senza differenze significative in qualsiasi momento della sera.[17] Un altro studio sui ratti che utilizzava una dose inferiore di 10mcg/mL (alla fine era di 35mcg al giorno) sempre disciolti in acqua ha anche riscontrato influenze sulla Leptina circolante, in cui i livelli erano aumentati a circa il 150% rispetto al gruppo di controllo (dati derivati dal grafico) dopo un mese. Questo studio ha anche riscontrato un aumento dello Zinco circolante. [18] Risultati simili sono stati osservati in casi di eccessiva somministrazione di Melatonina (3 mg/kg nei topi tramite I.V), in cui la Leptina è aumentata al 127% rispetto al controllo per 6 mesi. [19]
Desametasone
Quando si esaminano le cellule adipose isolate (dove viene prodotta la maggior parte della Leptina), la quantità di Leptina secreta non viene significativamente aumentata quando incubata con Melatonina 1nM.[18] Tuttavia, ciò può essere dovuto all’incubazione con la sola Melatonina, poiché altri studi che accoppiano la Melatonina con l’Insulina mostrano che la Melatonina può aumentare la secrezione di leptina indotta dall’Insulina, poiché nessuna delle due ha indotto la secrezione di leptina in vitro da sola, mentre la combinazione ha causato un aumento della secrezione del 120% e del 50% quella del contenuto di mRNA. L’aggiunta di Desametasone alla combinazione ha aumentato questi livelli rispettivamente al 250% e al 100%. [20] La Melatonina è stata in grado di sopprimere una marcata sotto regolazione indotta dal Adenosina Monofosfato Ciclico (cAMP) del rilascio di Leptina e svolgere un ruolo sinergico nell’attivazione del recettore dell’Insulina e del suo bersaglio, la proteina chinasi B (Akt). I suoi effetti furono soppressi quando venne impedita l’azione del MT1.[20] Questi effetti sono stati successivamente replicati dallo stesso gruppo di ricerca, con la stessa potenza, quando gli adipociti sono stati incubati con un protocollo di “12 ore on/12 ore off ” per imitare il ritmo circadiano.[21]
Processo lipogenetico
L’aumento della Leptina può essere un meccanismo di feedback negativo dovuto alla segnalazione della Melatonina. La Melatonina è stata studiata per le sue interazioni con l’obesità, poiché i ratti privi di ghiandole pineale che secerne la Melatonina (ratti che hanno subito una pinealectomia) sperimentano un aumento della lipogenesi e una riduzione della lipolisi. [22] L’abbinamento di una mancanza di secrezione e sintesi di Melatonina con l’aumento di peso suggerisce che la Melatonina può essere anti-obesogenica (riduce l’aumento di grasso) o induttiva della perdita di grasso.
La Melatonina sembra essere in qualche modo un regolatore negativo della fisiologia degli adipociti, essendo in grado di influenzare la differenziazione delle cellule staminali mesenchimali (MSC) lontano dagli adipociti e promuovere la crescita delle cellule osteogeniche secondaria all’inibizione del Recettore Attivato dai Proliferatori Perossisomiali (PPAR) [23] mentre sopprime la proliferazione di adipociti maturi 3T3-L1, secondari alla soppressione dell’attività trascrizionale C/EBPbeta. [24] Se trattata in preadipociti 3T3-L1, la Melatonina è in grado di indurre proliferazione. Questo sembra agire attraverso l’attivazione del recettore MT1. [25]
GLUT-4
In un modello di adipociti PAZ6 (la linea cellulare dei preadipociti bruni umani)
[26]), è stato riscontrato che esisteva l’mRNA per entrambi i recettori della Melatonina e, tramite i recettori MT2, sopprimeva la traslocazione dei GLUT4 e l’assorbimento del glucosio di circa il 25% in 14 giorni di incubazione, ma non riusciva a ridurre significativamente l’attività dopo un giorno. [26] In questo studio, gli adipociti bruni e bianchi sono stati entrambi testati e sebbene quelli bianchi avessero meno MT1 rispetto agli adipociti bruni, quelli bianchi non esprimevano MT2. [26] Uno studio che utilizzava Luzindole (un agonista per lo più degli MT2 ma con una certa affinità con gli MT1) ha dimostrato che era meno efficace della Melatonina nel creare questi effetti, supportando la mancanza o una relativa assenza di MT2 attivo sugli adipociti bianchi.[25] L’attivazione dei recettori della Melatonina sembra essere associata alla soppressione dell’Adenil Ciclasi e ad una diminuzione dei livelli di Cortisolo-indotta dell’Aromatasi (cAMP). [25] [24] Questa riduzione dei livelli di cAMP (associata alla proteina Gi accoppiata ai recettori della Melatonina) può sopprimere la lipolisi indotta dalla stimolazione beta-2 adrenergica. [27]
Superossido Dismutasi (SOD)
Quando si studia l’ossidazione, l’incubazione dei preadipociti con la Melatonina è associata ad un aumento dei livelli di Rame, Zinco, Manganese e Superossido Dismutasi (SOD). È stato osservato un aumento della catalasi dopo un’incubazione di 24 ore. Queste tendenze si sono invertite dopo 48 ore di incubazione.
Quando i ratti vengono trattati con 500mcg di Melatonina al giorno attraverso l’acqua potabile e contemporaneamente vengono sottoposti ad una dieta ricca di grassi (35%), il tasso di aumento di peso viene attenuato, indipendentemente dalle variazioni delle calorie. [17] Questo è stato replicato con 0,4mcg/mL, dove è stata registrata una riduzione del 7% del peso corporeo e una massa adiposa intra-addominale inferiore del 16%. [28] Quando 5mg di Melatonina vengono somministrati 2 ore prima di coricarsi a un campione di persone con sindrome metabolica, è stata osservata una piccola ma statisticamente significativa riduzione dell’IMC (da 29 a 28,8) nell’arco di due mesi, che si correla con miglioramenti della pressione sanguigna e profilo antiossidante.[29]
Fino a questo punto le informazioni esposte sono senza dubbio interessanti e applicabili in diversi contesti, dal soggetto obeso/sovrappeso all’atleta in regime ipocalorico prolungato (vedi calo della Leptina e sue conseguenze sistemiche in condizioni di ipoalimentazione). Ma gli effetti collaterali? Ovviamente esistono sebbene hanno un incidenza piuttosto contenuta se la supplementazione si limita ai dosaggi comunemente indicati (1-10mg/die max).
Quindi, la Melatonina sembra causare pochissimi effetti collaterali come osservato nei test sul breve termine, fino a tre mesi, e a basse dosi [vedi soglia variabile sopra esposta]. Due reviews sistematiche non hanno riscontrato effetti avversi della Melatonina esogena in numerosi studi clinici e studi comparativi hanno riscontrato che gli effetti collaterali quali mal di testa, vertigini, nausea e sonnolenza sono stati riportati in egual misura sia per la Melatonina che per il placebo. [30] [31] La Melatonina a rilascio prolungato è sicura con un uso a lungo termine fino a 12 mesi. [32] Sebbene non sia raccomandato per un uso a lungo termine oltre a questo lasso di tempo, la Melatonina a basso dosaggio è generalmente più sicura e un’alternativa migliore, rispetto a molte prescrizioni farmaceutiche da banco e non per indurre il sonno con necessità d’uso per un lungo periodo di tempo e con rischio di sviluppare dipendenza. Le basse dosi di Melatonina sono generalmente sufficienti per produrre un effetto ipnotico nella maggior parte delle persone. Dosi più elevate non sembrano produrre un effetto più forte, ma sembrano invece provocare sonnolenza per un periodo di tempo più lungo.[33]
La Melatonina può causare nausea, irritabilità il giorno successivo e irritabilità. [34] Negli anziani, il suo uso può causare una riduzione del flusso sanguigno e ipotermia. [35] [necessità di aggiornamento] Nei disturbi autoimmuni, l’evidenza è in conflitto se l’integrazione di Melatonina può migliorare o esacerbare i sintomi dovuti all’immunomodulazione. [36] [37] [necessità di aggiornamento]
La Melatonina può abbassare i livelli dell’ormone follicolo-stimolante. [38] Gli effetti della Melatonina sulla riproduzione umana rimangono poco chiari. [39]
In quelli che assumono warfarin, alcune prove suggeriscono che potrebbe esistere una potente interazione farmacologica, aumentando l’effetto anticoagulante del warfarin e il rischio di sanguinamento. [40]
Comunque sia, in base ai dati ad oggi disponibili, la supplementazione di Melatonina al fine di coadiuvare la perdita di peso in soggetti obesi o sovrappeso, o per mantenere livelli più alti di Leptina in periodi prolungati di ipocalorica (con tutti i vantaggi del caso, specie in un Natural), necessita di un dosaggio tra i 5 ed i 10mg al giorno somministrati in una o due dosi giornaliere. Ovviamente si tratta di una pratica attualmente sperimentale sebbene i presupposti per una sua azione a grado soddisfacente non sono così scarse.
Gabriel Bellizzi
Riferimenti:
1- Caniato R et al., Melatonin in plants, in Adv. Exp. Med. Biol., Advances in Experimental Medicine and Biology, vol. 527, 2003, pp. 593–7, DOI:10.1007/978-1-4615-0135-0_68, ISBN 978-0-306-47755-3, PMID 15206778.
2- Paredes SD et al., Phytomelatonin: a review, in J. Exp. Bot., vol. 60, nº 1, 2009, pp. 57–69, DOI:10.1093/jxb/ern284, PMID 19033551.
3- The journal of experimental zoology.
4- Paul M. Coates, Marc R. Blackman; Gordon M. Cragg; Mark Levine; Joel Moss; Jeffrey D. White, Encyclopedia of dietary supplements, CRC Press, 2005, p. 819, ISBN 978-0-8247-5504-1. URL consultato il 16 febbraio 2012.
5- Paul M. Coates, M. Coates Paul, Marc Blackman, Marc R. Blackman, Gordon M. Cragg, Mark Levine, Jeffrey D. White, Joel Moss e Mark A. Levine, Encyclopedia of Dietary Supplements, su books.google.it. URL consultato il 12 giugno 2018.
«However, the finding did indicate that the pineal contained e compound with at least some biological activity, and it provided a way of identifying tue active compound, using assays based on the ability of purified extracts to aggregate the melanin granules in the frog’s pigment cells. In 1958, Lerner et al.[2] discovered the compound’s chemical structure to be 5-methoxy-N-acetyltryptamine and named melatonin.».
6- Ana Maria Caliman Filadelfi, Castrucci, Ana Maria de Lauro, Comparative aspects of the pineal/melatonin system of poikilothermic vertebrates, in Journal of Pineal Research, vol. 20, nº 4, 1996, pp. 175–86, DOI:10.1111/j.1600-079X.1996.tb00256.x, PMID 8836950.
7- David Sugden, Davidson, Kathryn; Hough, Kate A;Teh, Muy-Teck, Melatonin, Melatonin Receptors and Melanophores: A Moving Story, in Pigment Cell Research, vol. 17, nº 5, 2004, pp. 454–60, DOI:10.1111/j.1600-0749.2004.00185.x, PMID 15357831.
8- Hing-Sing Yu, Russel J. Reiter, Melatonin: Biosynthesis, Physiological Effects, and Clinical Applications, CRC Press, 1993, p. 550, ISBN 978-0-8493-6900-1.
9- A.B. Lerner, Case, J.D. Takahashi, Y., Isolation of melatonin and 5-methoxyindole-3-acetic acid from bovine pineal glands, in The Journal of biological chemistry, vol. 235, 1960, pp. 1992–7, PMID 14415935.
10- H. Lynch, Wurtman, R. Moskowitz, M. Archer, M. Ho, M., Daily rhythm in human urinary melatonin, in Science, vol. 187, nº 4172, 1975, pp. 169–71, DOI:10.1126/science.1167425, PMID 1167425.
11- D. X. Tan, Chen, L. D. Poeggeler, B. Manchester, L. C. Reiter, R. J., Melatonin: a potent, endogenous hydroxyl radical scavenger, in Endocrine J, vol. 1, 1993, pp. 57-60.
12- Methods of inducing sleep using melatonin, Patent Genius. URL consultato l’11 giugno 2018 (archiviato dall’url originale il 4 febbraio 2018).
«Brevetto USA #5449683».
13- J. Arendt, Melatonin: Characteristics, Concerns, and Prospects, in Journal of Biological Rhythms, vol. 20, nº 4, 2005, pp. 291-303, DOI:10.1177/0748730405277492, PMID 16077149.
«There is very little evidence in the short term for toxicity or undesirable effects in humans. The extraordinary “hype” of the miraculous powers of melatonin in the recent past did a disservice to acceptance of its genuine benefits».
15- Celinski K, et al. Melatonin or L-tryptophan accelerates healing of gastroduodenal ulcers in patients treated with omeprazole.
J Pineal Res (2011)
16- Gonciarz M, et al. Plasma insulin, leptin, adiponectin, resistin, ghrelin, and melatonin in nonalcoholic steatohepatitis patients treated with melatonin.
J Pineal Res (2012)
17- Ríos-Lugo MJ, et al. Melatonin effect on plasma adiponectin, leptin, insulin, glucose, triglycerides and cholesterol in normal and high fat-fed rats.
J Pineal Res (2010)
18- Song YM, Chen MD. Effects of melatonin administration on plasma leptin concentration and adipose tissue leptin secretion in mice.
Acta Biol Hung (2009)
19- Baltaci AK, Mogulkoc R. Pinealectomy and melatonin administration in rats: their effects on plasma leptin levels and relationship with zinc.
Acta Biol Hung (2007)
20- Alonso-Vale MI, et al. Melatonin enhances leptin expression by rat adipocytes in the presence of insulin.
Am J Physiol Endocrinol Metab (2005)
21- Alonso-Vale MI, et al. Intermittent and rhythmic exposure to melatonin in primary cultured adipocytes enhances the insulin and dexamethasone effects on leptin expression.
J Pineal Res (2006)
22- Borges-Silva CN, et al. Reduced lipolysis and increased lipogenesis in adipose tissue from pinealectomized rats adapted to training.
J Pineal Res (2005)
23- Zhang L, et al. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARγ expression and enhancing Runx2 expression.
J Pineal Res (2010)
24- Alonso-Vale MI, et al. Adipocyte differentiation is inhibited by melatonin through the regulation of C/EBPbeta transcriptional activity.
J Pineal Res (2009)
25- Zwirska-Korczala K, et al. Influence of melatonin on cell proliferation, antioxidative enzyme activities and lipid peroxidation in 3T3-L1 preadipocytes–an in vitro study.
J Physiol Pharmacol (2005)
26- Kazantzis M, et al. PAZ6 cells constitute a representative model for human brown pre-adipocytes.
Front Endocrinol (Lausanne) (2012)
27- Zalatan F, Krause JA, Blask DE. Inhibition of isoproterenol-induced lipolysis in rat inguinal adipocytes in vitro by physiological melatonin via a receptor-mediated mechanism.
Endocrinology (2001)
28- Wolden-Hanson T, et al. Daily melatonin administration to middle-aged male rats suppresses body weight, intraabdominal adiposity, and plasma leptin and insulin independent of food intake and total body fat.
Endocrinology (2000)
29- Koziróg M, et al. Melatonin treatment improves blood pressure, lipid profile, and parameters of oxidative stress in patients with metabolic syndrome.
J Pineal Res (2011)
30- Buscemi N, Vandermeer B, Hooton N, Pandya R, Tjosvold L, Hartling L, et al. (December 2005). “The efficacy and safety of exogenous melatonin for primary sleep disorders. A meta-analysis”. Journal of General Internal Medicine. 20 (12): 1151–8. doi:10.1111/j.1525-1497.2005.0243.x. PMC 1490287. PMID 16423108.
31- Buscemi N, Vandermeer B, Hooton N, Pandya R, Tjosvold L, Hartling L, et al. (February 2006). “Efficacy and safety of exogenous melatonin for secondary sleep disorders and sleep disorders accompanying sleep restriction: meta-analysis”. BMJ. 332 (7538): 385–93. doi:10.1136/bmj.38731.532766.F6. PMC 1370968. PMID 16473858.
32- Lyseng-Williamson KA (November 2012). “Melatonin prolonged release: in the treatment of insomnia in patients aged ≥55 years”. Drugs & Aging. 29 (11): 911–23. doi:10.1007/s40266-012-0018-z. PMID 23044640.
33- Database of Abstracts of Reviews of Effects (DARE): Quality-assessed Reviews [Internet]. York (UK): Centre for Reviews and Dissemination (UK); 1995. Optimal dosages for melatonin supplementation therapy in older adults: a systematic review of current literature. 2014.
34- Brent Bauer, M.D. “Melatonin side effects: What are the risks?”. Mayo Clinic. Retrieved 17 August 2011.
35- Zhdanova IV, Wurtman RJ, Regan MM, Taylor JA, Shi JP, Leclair OU (October 2001). “Melatonin treatment for age-related insomnia”. The Journal of Clinical Endocrinology and Metabolism. 86 (10): 4727–30. doi:10.1210/jc.86.10.4727. PMID 11600532.
36- Morera AL, Henry M, de La Varga M (2001). “[Safety in melatonin use]” [Safety in melatonin use]. Actas Espanolas de Psiquiatria (in Spanish). 29 (5): 334–7. PMID 11602091.
37- Terry PD, Villinger F, Bubenik GA, Sitaraman SV (January 2009). “Melatonin and ulcerative colitis: evidence, biological mechanisms, and future research”. Inflammatory Bowel Diseases. 15 (1): 134–40. doi:10.1002/ibd.20527. PMID 18626968.
38- Juszczak M, Michalska M (2006). “[The effect of melatonin on prolactin, luteinizing hormone (LH), and follicle-stimulating hormone (FSH) synthesis and secretion]” [The effect of melatonin on prolactin, luteinizing hormone (LH), and follicle-stimulating hormone (FSH) synthesis and secretion]. Postepy Higieny I Medycyny Doswiadczalnej (in Polish). 60: 431–8. PMID 16921343.
39- Srinivasan V, Spence WD, Pandi-Perumal SR, Zakharia R, Bhatnagar KP, Brzezinski A (December 2009). “Melatonin and human reproduction: shedding light on the darkness hormone”. Gynecological Endocrinology. 25 (12): 779–85. doi:10.3109/09513590903159649. PMID 19905996.
40- Shane-McWhorter L. Melatonin. Merck Manual Professional Version. October 2018. Accessed 22 April 2019.