Nella prima parte di questa disamina sono state passate in rassegna diverse informazioni necessarie per comprendere cosa sono le Incretine e quali sono le loro principali azioni. Si è poi passati a descrivere la classe di farmaci degli Incretino-Mimetici discorrendo sulla loro sintesi, tipologia molecolare [in ordine cronologico in base alla data di commercializzazione] e caratteristiche di azione sia per quanto concerne le attività positive che quelle negative. In conclusione, si è accennato alla consistente diffusione dell’uso “cosmetico” di tali farmaci.
In questa seconda ed ultima parte vedremo come questa relativamente nuova classe di farmaci abbia trovato un certo spazio di diffusione nel Fitness e (sebbene in minor parte) nel BodyBuilding. Verranno discussi i punti di attrattiva e le limitazioni (e rischi) legati al loro uso.
Le “attrattive” degli Incretino-Mimetici:
L’obesità è una grave epidemia che affligge la società del così detto “occidente americanizzato”. I farmaci iniettabili sottocutanei inizialmente concepiti per la gestione del diabete di tipo II, come la Semaglutide e altri agonisti del recettore del GLP-1, stanno rapidamente guadagnando popolarità per i loro effetti sulla perdita di peso. Questi farmaci (Ozempic, Wegovy, Saxenda e Mounjaro) sono onnipresenti sui social media e sono promossi da celebrità di tutte le fasce demografiche. “Viso da Ozempic” e ‘sedere da Ozempic’ sono ormai concetti mainstream che evidenziano i possibili cambiamenti morfologici che si verificano con questi farmaci. Con la diffusione non controllata da personale qualificato dell’uso di questi farmaci, è aumentato anche l’elenco dei potenziali effetti avversi.
Nella prima parte di questa disamina si è constatato che gli Incretino-Mimetici hanno potenzialità che si esprimono su modifiche della composizione corporea e che riguardano principalmente:
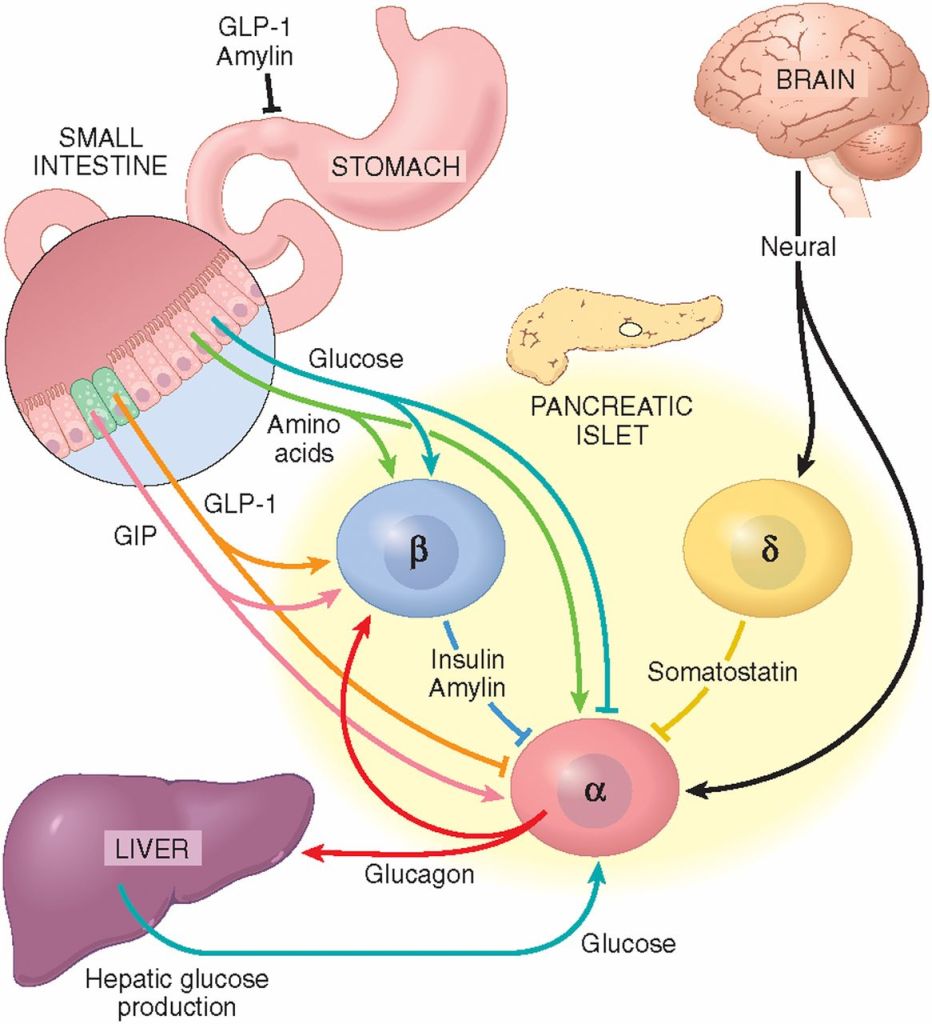
Rallentamento dello svuotamento gastrico con riduzione del senso di fame derivante dal aumento del senso di sazietà;
Gli effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare riducendo il senso di fame e il consumo di cibo;
Indirettamente, per via della stimolazione insulinica, aumento del senso di sazietà dato dal picco insulinico a livello ipotalamico;
Miglioramento del ripartizionamento Kcal attraverso il miglioramento della sensibilità all’insulina.
L'”attrattiva” che ha spinto (e che spinge) diverse persone di diverse classi sociali e di ambo i sessi a prendere in considerazione e concretizzare l’uso di questi farmaci è fondamentalmente ridotta alla riduzione della fame/appetito. Ma questo riguarda la persona “nella media”, la ragazza/donna alla ricerca di rapide soluzioni per l’imminente prova costume o per il servizio fotografico, sfilata ecc…
Nel Bodybuilding questa classe di farmaci ebbe una serie di attrattive comprendenti lo sfruttamento di tutti i punti sopra elencati. E, di conseguenza, si ipotizzarono fasi della preparazione nelle quali applicare tale categoria farmaceutica. Limitandone l’uso in “Cut” per ragioni legate al rischio (seppur limitato in monoterapia) di incorrere in eventi ipoglicemici, l'”attrattiva d’uso” riguardava il potenziale in fasi di “Recomp”; quindi non propriamente “ipocalorico” o, al massimo, leggermente ipocaloriche e non ipoglucidiche.
Ricomposizione corporea, Ripartizionamento calorico e Insulino-Sensibilità:
Ma chiariamo alcuni termini per schiarirci le idee…
Ricomposizione corporea: si intende il raggiungimento di un obiettivo o un risultato desiderato di un regime alimentare e di allenamento (e di farmaci), come il “Cut” o il “Bulk”.
“Cut”: si intende, propriamente, una diminuzione della massa grassa (FM) e il mantenimento della massa muscolare (LBM).
“Bulk”: si intende un aumento della LBM con una concomitante attenuazione dell’aumento della FM.
“Recomp” [comunemente intesa]: è definita come un aumento della LBM e la diminuzione della FM.
La suddivisione terminologica corretta è un concetto funzionale all’obiettivo di tutti gli interventi dietetici razionali.
Il rapporto p (rapporto di ripartizione) descrive le proteine depositate nei tessuti della LBM in relazione all’apporto energetico e, viceversa, le proteine perse dai tessuti della LBM in relazione al deficit energetico.
Il p-ratio comprende i fattori di:
stato ormonale (cioè i livelli assoluti di ormoni chiave noti);
sensibilità all’insulina;
sensibilità alla Leptina.
Esiste un’interazione tra il punto 1 e 3.
La prima chiave è…
Sensibilità all’Insulina: quando si è a dieta (cioè in uno stato di deficit energetico), la resistenza all’Insulina fisiologica è una condizione favorevole all’uso del grasso di deposito limitando l’uso del glucosio da parte del muscolo come substrato energetico, risparmiando il glucosio per il cervello e l’utilizzo degli Acidi Grassi intramuscolare. In condizioni di uscita da un regime ipocalorico, in uno stato di migliorata insulino-sensibilità, l’aumento calorico di una fase di “Bulk” vede, almeno inizialmente, un ripartizionamento delle Kcal al miocita a discapito del adipocita; tale condizione di inverte col protrarsi del regime ipercalorico.
I fattori che influenzano la sensibilità all’insulina includono [1]:
Livelli di grasso corporeo; B.F. % (predittore primario): ↑B.F. ⇒ ↑Acidi Grassi come substrato energetico (risparmiando glucosio e proteine [che possono essere utilizzate dal fegato nella gluconeogenesi]) e detta la segnalazione delle adipochine (cioè gli ormoni secernenti gli adipociti [leptina, TNF-α, IL-…, adiponectina, etc.]) ⇒ ↓ sensibilità all’insulina;
Contrazione muscolare (cioè attività, come ad esempio locomozione, allenamento contro-resistenza) ⇒ ↑ assorbimento di glucosio nella cellula muscolare; traslocazione di GLUT-4 ⇒ ↑ sensibilità all’Insulina;
Dieta: elevata quantità di carboidrati (in ipercalorica), grassi saturi e poche fibre ⇒ ↓ sensibilità all’Insulina;
Stoccaggio del Glicogeno o Supercompensazione [successivo ad una deplezione] ⇒ ↑assorbimento di glucosio e glicogenesi ⇒↑sensibilità Insulinica;
Deplezione di glicogeno (ad esempio, nel periodo successivo a un allenamento intenso, prima di un’alimentazione particolarmente ricca di carboidrati) ⇒ sottoregolazione (deplezione pressoché totale) della disponibilità di glucosio e promozione dell’ossidazione di Acidi Grassi dopo il depauperamento delle scorte di glicogeno muscolare (in media < 700 g negli adulti) ⇒ ↑ Acidi Grassi liberi nel sangue (circolanti) ⇒ ↓ sensibilità all’Insulina;
Fattori genetici in parte modificabili dai farmaci, ad esempio, nei casi di ipogonadismo, l’applicazione TRT inverte chiaramente l’insulino-resistenza nei casi in cui l’eziologia dell’insulino-resistenza è riconducibile alla carenza di Testosterone.
La seconda chiave è…
Struttura molecolare della Leptina
La Leptina: come sappiamo bene, la Leptina è un ormone, più precisamente è una adipochina, secreta principalmente dagli adipociti, che si correla con la %B.F.;↑%B.F. ⇒ ↑Leptina. (i depositi viscerali e quelli sottocutanei hanno rapporti diversi con la Leptina). A una data percentuale di B.F., le donne producono ~2 – 3 volte più Leptina rispetto agli uomini. Le concentrazioni di Leptina cambiano con la restrizione energetica e la sovralimentazione. La Leptina è un segnalatore primario di regolazione dell’accumulo di energia che riflette:
la percentuale di B.F.;
l’assunzione di energia.
Esempio 1: all’inizio di una dieta ipocalorica, la Leptina può diminuire del 50% entro 1 settimana (o meno) – anche se ovviamente il soggetto a dieta non ha perso il 50% di B.F. – quindi, in un primo momento, i cambiamenti della concentrazione di Leptina non sono correlati alla B.F. (piuttosto segnalano l’assunzione di energia).
Dopo il calo iniziale, si assiste a un declino più graduale della Leptina in relazione alla perdita di %B.F. .
Esempio 2: in caso di sovralimentazione, la Leptina subisce un incremento in modo altrettanto rapido (cioè senza relazione con la %B.F., ma in relazione all’assunzione di energia).
A breve termine, la secrezione di Leptina è determinata principalmente dalla disponibilità di glucosio, per cui la riduzione della disponibilità di glucosio nella cellula adiposa (dieta ipocalorica) ⇒ ↓Leptina e viceversa.
Gli effetti specifici dell’ormone Leptina includono effetti sul pancreas e sul fegato, nel muscolo scheletrico ↑FA e ↓AA e l’uso del glucosio come substrato energetico (aumentando la perdita di grasso, promuovendo il risparmio di proteine)… [1]
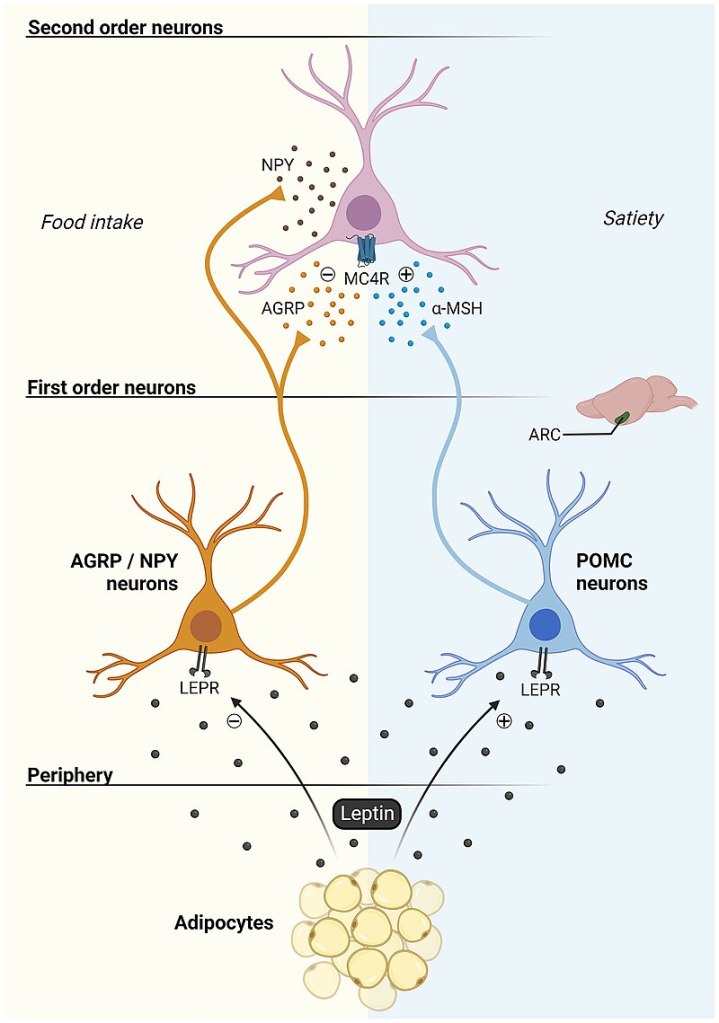
Modello classico Leptina-Melanocortina
In sostanza, il partizionamento (p-ratio) è un concetto che associa la Leptina e la sensibilità all’Insulina come fattori principali che determinano il modo in cui le variazioni dell’apporto calorico e del contenuto di macronutrienti influiscono sul metabolismo (influenzando profondamente la composizione corporea) e sullo stato ormonale. Possiamo modificarlo e migliorarlo, tenendo conto dei tessuti bersaglio e del nostro obiettivo (ad esempio, se Bulk o Cut).
Inoltre, non bisogna confondere il potenziamento dell’insulino-resistenza fisiologica sulla perdita di grasso con l’erronea valutazione che l’insulino-resistenza sia salutare. L’insulino-resistenza, soprattutto in una persona sedentaria, è associata alla sindrome metabolica, al diabete di tipo II, per non parlare del grasso viscerale, ecc.
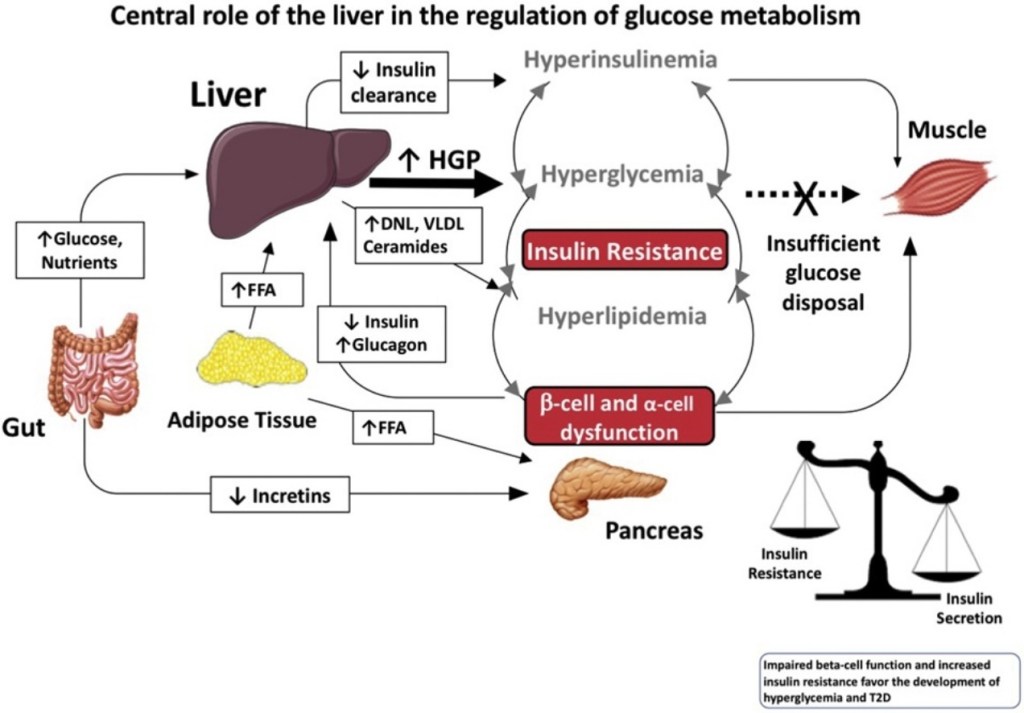
L’insulino-resistenza è uno stato in cui i tessuti dell’organismo (ad esempio, fegato, pancreas, muscolo scheletrico) presentano una scarsa recettività con l’Insulina continuando, se si parla in particolare del fegato, a produrre glucosio in quantità inappropriate. Questo stato di iperglicemia è un effetto piuttosto che la causa dell’insulino-resistenza, anche se i livelli tossici di glucosio degradano la reattività delle isole pancreatiche all’Insulina rappresentando così una delle vie/meccanismi dell’insulino-resistenza, peggiorando la stessa condizione.
Ma tutto questo cosa centra con gli Incretino-Mimetici? Se non ci siete ancora arrivati, calma e capirete …
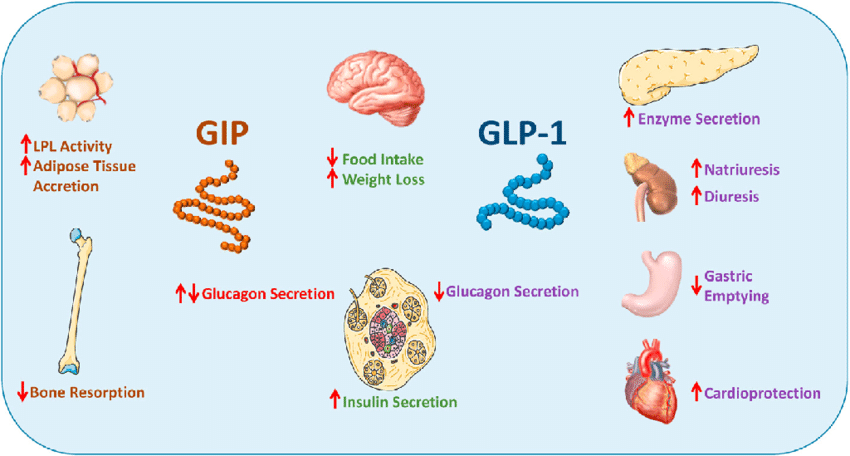
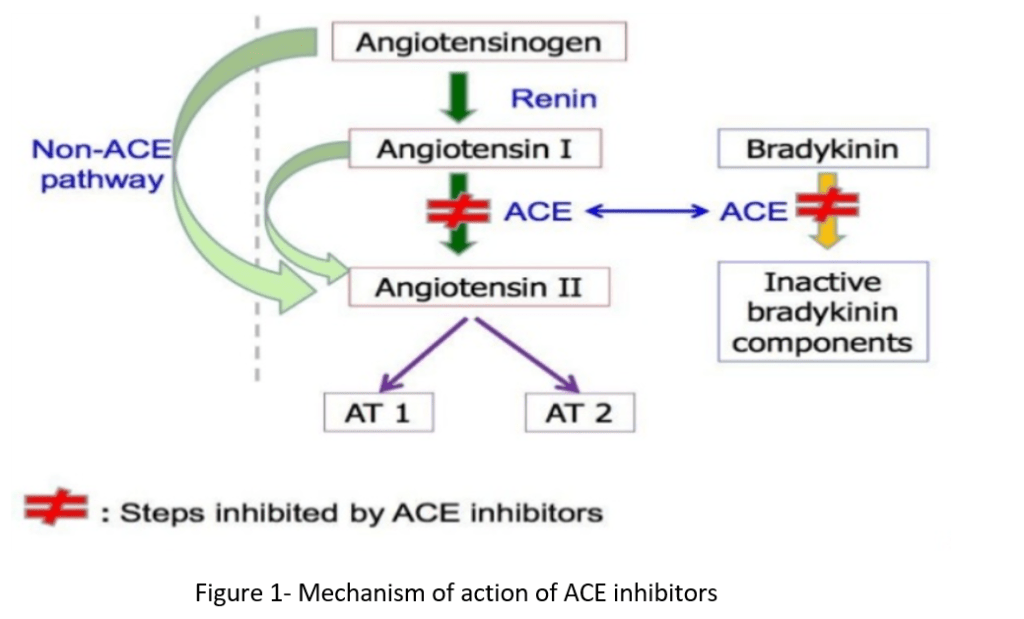
Agonisti del recettore GLP-1 e GIP:
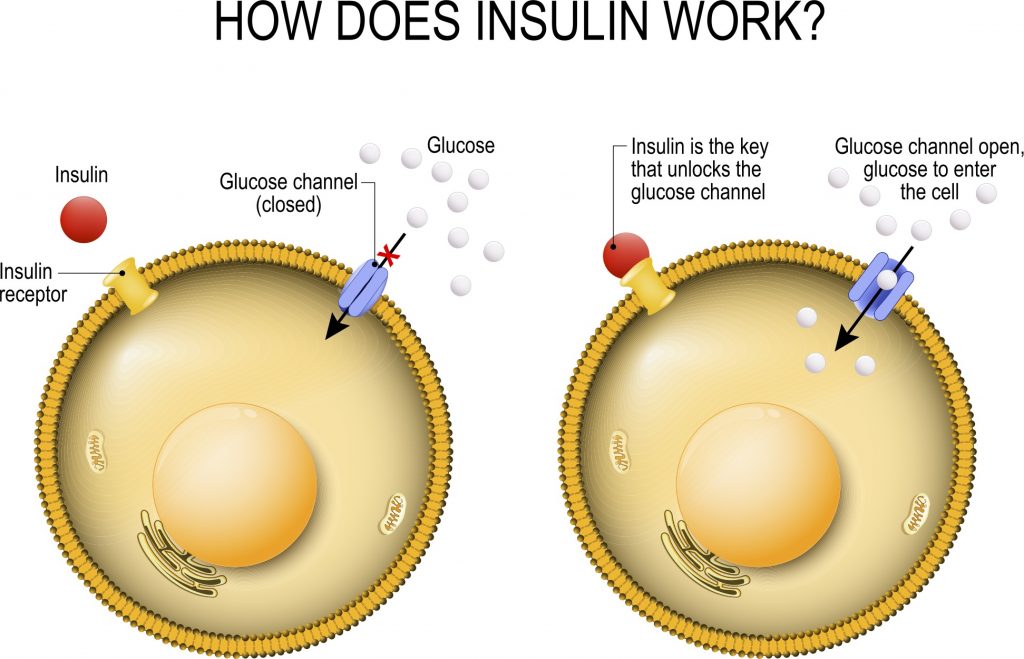
La perdita di grasso si verifica con gli agonisti del GLP-1 e della GIP (Incretino-mimetici) – come la Semaglutide e la Tirzepatide – che sono veri e propri agenti sensibilizzanti dell’Insulina. Tuttavia, non è la sensibilità all’Insulina di per sé che è responsabile della perdita di grasso con questa classe di farmaci – ma piuttosto, come abbiamo già visto, la perdita di grasso avviene grazie agli altri effetti di questi farmaci, come l’alterazione potenziale delle preferenze alimentari, il ritardo dello svuotamento gastrico, il senso di sazietà, che promuovono il controllo dell’appetito e riducono l’assunzione di energia.
Sappiamo che gli agonisti del GLP-1 e del GIP migliorano direttamente la sensibilità all’insulina modulando la secrezione di Insulina – accoppiandola alla presenza di elevate concentrazioni di glucosio. Questa secrezione di Insulina si attenua quando le concentrazioni di glucosio nel sangue diminuiscono e si avvicinano all’euglicemia. Inoltre, anche se indirettamente, riducendo l’assunzione di cibo, questi farmaci determinano una riduzione della %B.F.. La riduzione della percentuale di B.F. dovuta alla riduzione dell’assunzione di cibo riduce le riserve di massa grassa (e quindi gli FFA circolanti), migliorando ulteriormente la sensibilità all’Insulina.
La stragrande maggioranza degli agenti per la perdita di grasso, in quanto agenti lipolitici, favoriscono l’insulino-resistenza. Ad esempio i β-agonisti, non selettivi come l’Efedrina o selettivi come il Clenbuterolo, oppure lo stimolante da banco per eccellenza la caffeina, agendo in modo analogo o aumentano l’azione delle catecolamine (epinefrina e noradrenalina, o adrenalina e noradrelanina) possono portare ad un peggioramento di questa condizione.
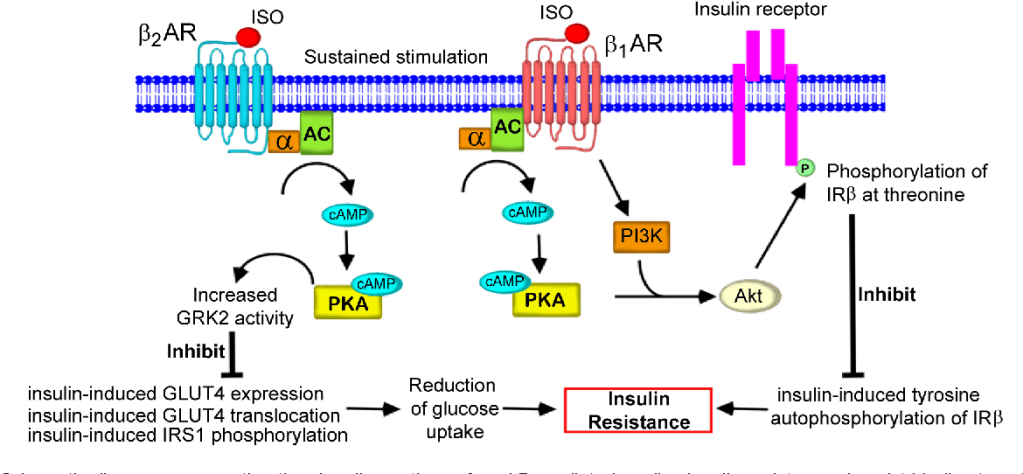
Diagramma schematico che rappresenta la via di segnalazione dell’insulino-resistenza cardiaca mediata dai β-AR. Tale meccanismo interessa (tra gli altri tessuti) anche il muscolo-scheletrico.
Quando tessuti come il fegato e le cellule adipose vedono ridotta l’interazione con l’Insulina, il glucosio non viene ottimamente assorbito dalle cellule. Con un marcato calo del glucosio, il fegato inizia a metabolizzare gli acidi grassi liberi (FFA), aumentando così i livelli di chetoni nel sangue e impedendo che vengano riesterificati nelle cellule adipose (in ipocalorica). Nel fegato e nelle cellule adipose, senza che l’Insulina interagisca ottimamente con questi tessuti, si verifica una soppressione della sintesi/lipogenesi dei grassi (negli adipociti) e della sintesi di lipoproteine a bassissima densità (VLDL) (nel fegato).
Incretino-mimetici, miglioramento della sensibilità all’Insulina e preservazione della massa muscolo-scheletrica:
La classificazione degli incretino-mimetici come agenti di ripartizione calorica:
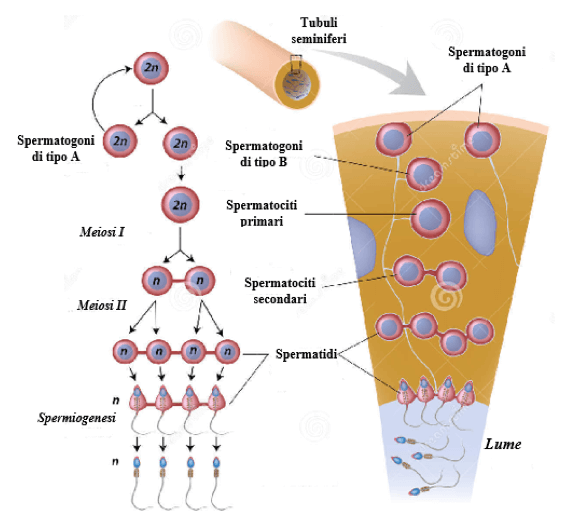
Nel BodyBuilding, il fascino verso questa classe di farmaci si concentra anche sul funzionamento degli agonisti del GLP-1 e della GIP sull’insulino-resistenza, poiché quest’ultima durante la restrizione calorica nel muscolo scheletrico (>60% del peso corporeo, più nei bodybuilder) è un’immagine non proprio esaltante, con le riserve di glicogeno che vengono prima catabolizzate abbastanza rapidamente; poi i trigliceridi intramuscolari (che rappresentano solo l’1% del peso del muscolo idratato, fino al 2% del volume, dato che il grasso è meno denso del muscolo scheletrico, e ~1/3 dell’energia muscolare, dato che il grasso è energeticamente denso) e infine, se necessario, l’organismo utilizzerà gli AA (catabolizzando le proteine muscolari; proteolisi) per ottenere l’energia necessaria a svolgere le attività giornaliere. Questi agenti, quindi, nella misura in cui sono sensibilizzanti per l’Insulina, dovrebbero servire a promuovere il mantenimento della LBM durante il “Cut”.
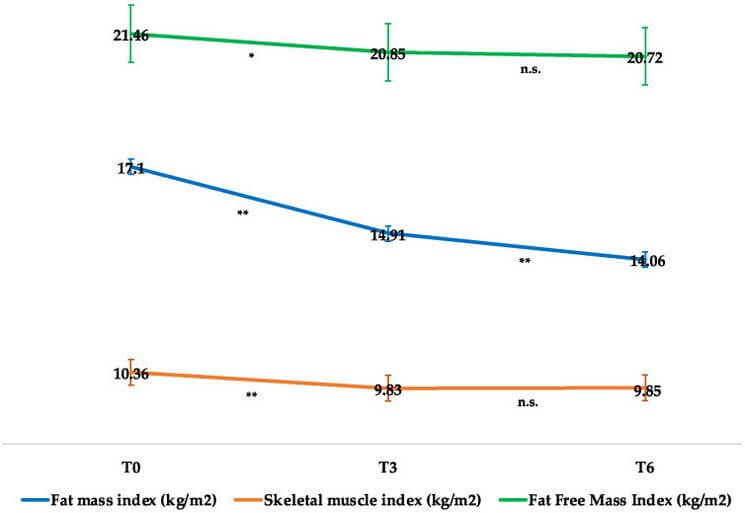
In effetti, come si evince dalla seguente immagine tratta da un articolo di ricerca di Volpe et. al del 2022 [2], la Semaglutide preserva in modo abbastanza efficace la LBM e riduce in modo preferenziale la FM, con riduzioni solo clinicamente insignificanti dell’indice di massa magra (FFMI, kg/m²) e dell’indice della muscolatura scheletrica durante il periodo iniziale di adattamento, che poi si attenua:
In un certo senso, quindi, migliorando il grado di sensibilità all’Insulina dell’equazione della p-ratio, gli incretino-mimetici possono essere classificati come agenti di ripartizione calorica, a grandi linee come il Clenbuterolo, ma invece di promuovere l’insulino-resistenza come i β-agonisti, la migliorano.
* Ovviamente, creare un ambiente significativamente insulino-sensibile in un contesto ipocalorico (soprattutto se ipoglucidico) può mettere l’utilizzatore a maggior rischio (sebbene limitato) di ipoglicemia o calo glicemico borderline con effetti simili allo stato di ipoglicemia (tremore, sudorazione copiosa ecc…).
Per coloro che hanno familiarità con questi concetti, derivanti dalle discussioni sul bodybuilding, può risultare molto confuso il fatto che l’iperglicemia (elevata quantità di glucosio nel sangue) è solo uno dei fattori associati all’insulino-resistenza, ma in realtà non è sinonimo di insulino-resistenza (iperglicemia ≠ insulino-resistenza). Sì, ridurre la glicemia a livelli normali è molto importante per migliorare la sensibilità all’insulina durante l’uso dell’ormone della crescita esogeno (rhGH), perché il glucosio è tossico per le cellule β pancreatiche. Questa glucotossicità a livello delle cellule pancreatiche si traduce in una diminuzione della risposta secretoria dell’insulina all’iperglicemia, alimentando così il fuoco dell’iperglicemia e della glucotossicità, contribuendo all’insulino-resistenza – ma non costituendone l’unica eziologia.
La sensibilità all’Insulina è multifattoriale e comprende componenti sistemiche (ad esempio, QUICKI) e periferiche (ad esempio, GLUT-4) ed è regolata a livello centrale da GLP-1 e GIP. L’iperglicemia è solo uno dei fattori (l’altro è l’Insulina) che funge da proxy dell’insulino-resistenza sistemica. Vi sono altri aspetti, tra cui la tolleranza ai carboidrati [vedi anche capacità di tolleranza del metabolismo glucidico], ecc.
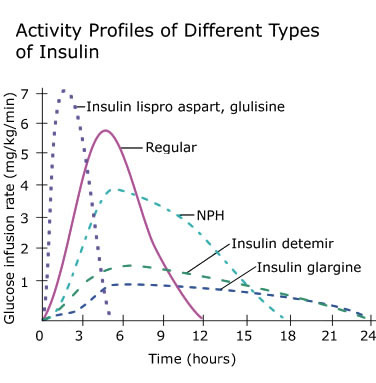
In tema di insulino-resistenza, è utile ricordare che l’Insulina endogena viene secreta in modo pulsatile per regolare il metabolismo glucidico e lipidico, la crescita cellulare ecc…, a differenza del Testosterone che viene secreto in modo più stabile (rilascio graduale nel sangue, ma soggetto a variazioni diurne, ad esempio una maggiore secrezione al mattino rispetto a mezzogiorno/sera). Gli aumenti cronici di Insulina, ad esempio quelli relativi al profilo di rilascio di una bassa dose giornaliera di Insulina Glargine (Lantus), presentano un’area sotto la curva (AUC) relativamente ampia a causa del profilo di rilascio (concentrazioni elevate per lunghi periodi di tempo) rispetto ai normali profili di rilascio dell’Insulina endogena (paragonabili alla farmacocinetica dell’Insulina regolare, ad esempio Actrapid, Novolin o HumuLin -R). L’elevata AUC di Lantus e/o le dosi di Insulina regolare esogena moderatamente elevate e frequenti sono descritte come iperinsulinemia cronica.
Questa resistenza non avviene per feedback negativo a livello delle cellule β.
Al contrario, l’iperinsulinemia cronica che causa l’insulino-resistenza è multifattoriale e comprende:
L’aumento dell’HOMA-IR e la diminuzione del QUICKI (misure biochimiche dell’insulino-resistenza e della sensibilità all’Insulina, rispettivamente).
L’alterata trasduzione del segnale insulinico dovuta alla disfunzione del recettore dell’Insulina (IR) e alla diminuzione dell’autofosforilazione dell’IR, che blocca la traslocazione del GLUT-4 sulla superficie cellulare nelle cellule muscolari e adipose, con conseguente aumento del glucosio nel sangue [3]:
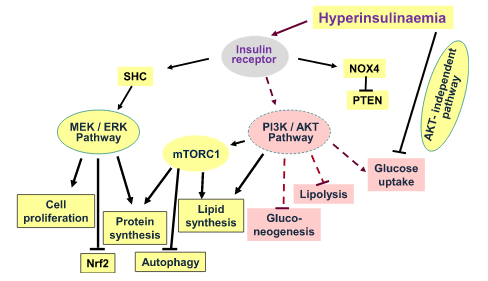
Segnalazione dell’Insulina durante l’insulino-resistenza. Durante l’insulino-resistenza, la segnalazione attraverso le chinasi AKT è parzialmente compromessa. Non tutte le vie AKT-dipendenti sono interessate, così come altre vie di segnalazione, indicando che l’insulino-resistenza è selettiva. Pertanto, l’iperinsulinemia, in presenza di insulino-resistenza, promuove le attività anaboliche delle cellule attraverso la via MEK-ERK e attraverso mTORC1. Sebbene la via PIK/AKT sia compromessa durante l’insulino-resistenza e fornisca solo una traslocazione insufficiente di GLUT4 per l’assorbimento del glucosio e un’attivazione carente di eNOS, sembra esserci un’attivazione normale di mTORC1. Oltre alle conseguenze anaboliche della segnalazione attraverso la via MEK/ERK descritte nella figura, si osserva un’aumentata espressione di ET-1 e PAt-1 (non mostrato), nonché l’inibizione dell’autofagia e del fattore nucleare Nrf2, che compromettono rispettivamente il turnover dei costituenti cellulari e i meccanismi di difesa delle cellule dallo stress radicale. L’iperinsulinemia riduce l’assorbimento del glucosio non solo attraverso lo smorzamento della via PIK/AKT (resistenza all’Insulina), ma anche attraverso altre vie ancora sconosciute.
3. Aumento dei livelli e dell’attività di sn-1,2-diacilglicerolo (DAG) grazie alla sintesi de novo.
Le limitazioni degli Incretino-Mimetici dietro all’iniziale entusiasmo:
Se ci dovessimo basare su quanto esposto fino a questo punto, saremo tutti d’accordo nell’ammantare della nomea di “farmaci prodigiosi per la ricomposizione corporea” tanto la Semaglutide quanto il Tirzepatide e tutti gli altri membri di questa classe. Ma, dal momento che, la conoscenza per essere utile deve essere sufficientemente approfondita, occorre indagare meglio sulle caratteristiche di questi farmaci.
Sappiamo che la Semaglutide è effettivamente associata alla perdita di peso con una differenza media dell’11,85% rispetto al placebo emersa dalle ultime review. Il consolidamento degli studi ha mostrato che nausea, vomito, costipazione e diarrea sono gli eventi avversi più comuni. Nonostante questi effetti sembrano essere per lo più di gravità da lieve a moderata, la loro risoluzione totale era spesso connessa al termine del trattamento.
Dal momento che gli Incretino-mimetici causano una riduzione del senso di fame, in modo anche significativo (dose dipendente), rallentano lo svuotamento gastrico portando anche ad eventi diarroici preceduti da mal assorbimento dei nutrienti, senza sottovalutare il rischio di paresi gastrica, la facilità di perdere massa muscolare in un contesto ipocalorico è molto alta, specie se non supportata da agenti anabolizzanti; l’inserimento di questi ultimi, però, non “tampona” la possibile condizione di mal assorbimento e/o scarsa assunzione proteica/alimentare.
Inoltre, la paresi gastrica è una condizione che un bodybuilder sano di mente cerca di evitare a tutti i costi combattendoci già se sussiste abuso di Insulina e/o hGH.
Trattando la limitazione data da una potenziale eccessiva inappetenza, è giusto specificare un impatto singolare che le Incretine (e gli Incretino-Mimetici) hanno sulle preferenze alimentari. Sappiamo, infatti, che gli alimenti maggiormente palatabili sono tipicamente ricchi di grassi e/o zuccheri e tendono a essere preferiti a quelli a basso contenuto di grassi/zuccheri. L’entità di questa preferenza, tuttavia, può essere influenzata da peptidi intestinali quali la Grelina e il GLP-1. La Grelina e il GLP-1 sono influenzati in modo differenziato dal consumo di alimenti palatabili. Tralasciando la Grelina, le concentrazioni di GLP-1 a digiuno predicono negativamente l’assunzione di alimenti ricchi di zuccheri semplici in un paradigma di distributori automatici, che gli autori dello studio hanno interpretato come prova del fatto che il GLP-1 svolge un ruolo nelle vie di ricompensa che regolano l’assunzione di zuccheri semplici. Diversi studi riportano anche un’alterazione delle preferenze alimentari dopo l’intervento di bypass gastrico, con un allontanamento dalla preferenza per gli zuccheri/grassi elevati. L’assunzione di alimenti appetibili, in particolare di soluzioni zuccherate, è aumentata dalla Grelina, mentre il GLP-1 riduce preferenzialmente l’assunzione di alimenti ad alto contenuto di grassi e zuccheri, almeno dopo una somministrazione acuta. Inoltre, i lavori condotti sull’uomo rivelano che la preferenza per i grassi e gli zuccheri può essere alterata dalla chirurgia bariatrica e contribuire alla perdita di peso, ma non è ancora stato stabilito se questi effetti siano legati a un’alterazione del segnale della Grelina o del GLP-1. Infine, i livelli circolanti di Grelina e GLP-1 possono essere indicativi del consumo di cibo appetibile nell’uomo.[https://www.frontiersin.org/]
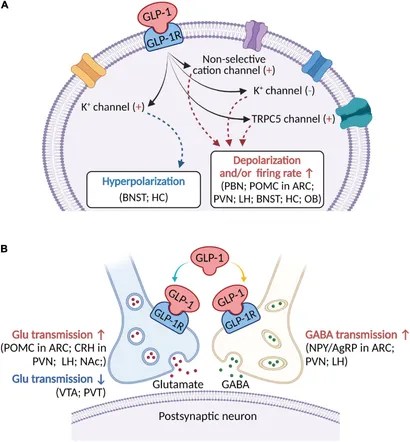
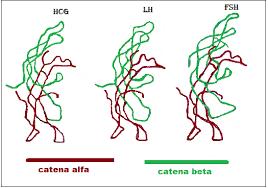
Schema che descrive i principali effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare. (A) Il GLP-1 (compresi i suoi agonisti) si lega al GLP-1R postsinaptico per depolarizzare il potenziale di membrana e/o aumentare la frequenza di sparo nella maggior parte delle regioni cerebrali, ma iperpolarizza il potenziale di membrana in alcune aree cerebrali. Diversi meccanismi ionici, tra cui il canale cationico non selettivo, il canale K+ e il canale TRPC5, possono essere coinvolti nell’attivazione della depolarizzazione o iperpolarizzazione indotta dal GLP-1R. (B) Oltre ai recettori postsinaptici, il GLP-1 agisce sui GLP-1R presinaptici per modulare la neurotrasmissione glutammatergica e GABAergica. ARC, nucleo arcuato; BNST, nucleo del letto della stria terminale; Glu, glutammato; CRH, ormone di rilascio della corticotropina; HC, ippocampo; LH, ipotalamo laterale; NAc, nucleo accumbens; NPY/AgRP, Neuropeptide Y/Peptide legato al gene Agouti; OB, bulbo olfattivo; PBN, nucleo parabrachiale; POMC, proopiomelanocortina; PVN, nucleo paraventricolare; PVT, nucleo talamico paraventricolare; VTA, area tegmentale ventrale.
Di conseguenza, nonostante gli indizi di cui sopra, l’uso di Incretino-Mimetici potrebbe ridurre marcatamente il consumo di cibo indipendentemente dalla fonte anche se, come abbiamo visto, l’effetto anoressizzante del GLP-1 sembra essere a maggior carico della componente alimentare glucidico-lipidica.
Tornando invece sulla questione legata al catabolismo muscolo-scheletrico e l’uso concomitante di agenti anabolizzanti, per ovviare a questo problema nei pazienti trattati con Semaglutide, è stato avviato uno studio clinico di fase 2b, multicentrico, in doppio cieco, controllato con placebo, randomizzato, per la determinazione della dose e per valutare la sicurezza e l’efficacia di Ostarina 3mg, Ostarina 6mg o placebo come trattamento per preservare la massa muscolare e aumentare la perdita di grasso in circa 150 pazienti con obesità sarcopenica o pazienti anziani in sovrappeso (>60 anni di età) trattati con Semaglutide (Wegovy®). L’endpoint primario è la massa corporea magra totale e gli endpoint secondari chiave sono la massa grassa corporea totale e la funzione fisica misurata dal test di salita delle scale a 16 settimane.
Dopo aver completato la parte di determinazione della dose di efficacia dello studio clinico di Fase 2b, si prevede che i partecipanti continueranno in cieco in uno studio clinico di estensione di Fase 2b in cui tutti i pazienti smetteranno di ricevere un GLP-1 RA, ma continueranno ad assumere un placebo, Ostarina 3mg o Ostarina 6mg per ulteriori 12 settimane. Lo studio clinico di estensione di fase 2b valuterà se l’Ostarina è in grado di mantenere la massa muscolare e prevenire l’aumento di grasso e peso che si verifica dopo l’interruzione di un GLP-1 RA.
Lo studio clinico è condotto in 14 centri clinici negli Stati Uniti. È stato raggiunto l’arruolamento completo dei circa 150 pazienti nello studio di fase 2b QUALITY. L’azienda prevede ora che l’ultimo paziente a completare lo studio di fase 2b QUALITY sarà nel dicembre 2024, con i risultati clinici di prima linea per lo studio clinico di fase 2b QUALITY attesi nel gennaio 2025. Inoltre, i risultati principali per lo studio clinico di estensione di Fase 2b in cieco separato possono ora essere attesi nel secondo trimestre solare del 2025.
Ovviamente, queste limitazioni, incisive nel Culturismo, interessano tutti gli Incretino-Mimetici, compresa la Tirzepatide la quale sembrerebbe avere un maggior margine di efficacia e “sicurezza” per quanto riguarda il rischio (seppur limitato in monoterapia) di eventi ipoglicemici.
Il motivo di questa riduzione di rischio ipoglicemico è dovuta alla doppia affinità recettoriale della Tirzepatide la quale, come abbiamo visto, possiede una attività agonista per i recettori del GLP-1 e del GIP. Ed è proprio il legame e l’attivazione di quest’ultimo recettore (GIP) da parte della Tirzepatide a permettere ciò. Infatti, l’attivazione del recettore GIP stimola la secrezione di Glucagone in modo glucosio-dipendente nelle persone sane, con un’attività maggiore in presenza di glicemie più basse. Ciò significa che, raggiunta la soglia euglicemica i livelli di glucosio nel sangue verranno mantenuti più facilmente all’interno di questa soglia per via dell’attività del Glucagone secreto dalle cellule α delle isole di Langerhans.[https://pubmed.ncbi.nlm.nih.gov/]
Sebbene i trials clinici suggeriscano che la Tirzepatide riduca la glucagonemia, un recente studio dimostra che la Tirzepatide è un potente stimolatore della secrezione di Glucagone nelle condizioni sopradette.
Quindi la Tirzepatide è superiore alla Semaglutide?
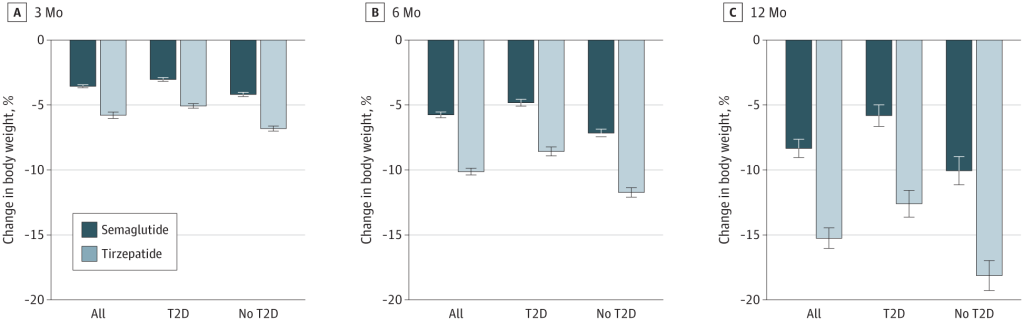
Alcuni studi recenti hanno messo a confronto la Semaglutide e la Tirzepatide per la perdita di peso. Studi di ricerca del 2021, del 2023 e del 2024 suggeriscono che la Tirzepatide può determinare una maggiore perdita di peso rispetto alla Semaglutide.
Variazione percentuale media del peso corporeo a 3, 6 e 12 mesi di trattamento per la popolazione complessiva, i soggetti con diabete di tipo II (TD2) e quelli senza TD2. Le barre rappresentano le variazioni medie del peso corporeo dal basale al punto di riferimento tra la popolazione di pazienti ancora in trattamento, abbinata in base al punteggio di propensione. Barre scure (Semaglutide); Barre chiare (Tirzepatide).
Ma questi studi presentano alcuni importanti limiti.
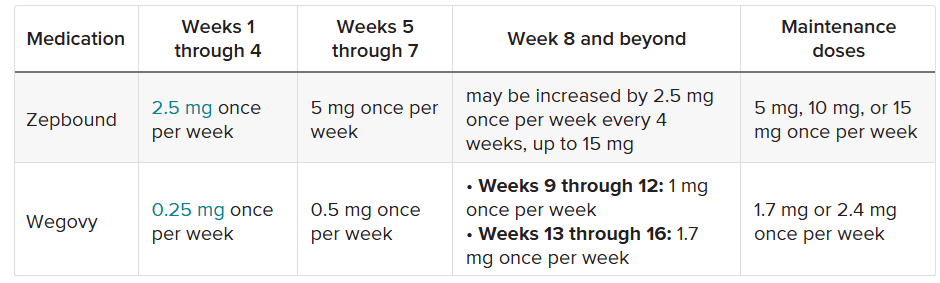
Le dosi di Semaglutide e Tirzepatide somministrate ai partecipanti non erano, per ovvie ragioni, uguali. La Semaglutide e Tirzepatide funzionano in modo leggermente diverso, come ormai sappiamo, e quindi i ricercatori hanno scelto livelli di dose comparabili. Tuttavia, la dose di Tirzepatide era più alta, il che potrebbe aver, anche di poco, influenzato i risultati.
Tabella comparativa tra il dosaggio della Semaglutide [Wegovy] e quella della Tirzepatide [Zepbound] usato negli studi.
Sappiamo altresì che la Tirzepatide ha una maggiore affinità con i recettori GIP rispetto ai recettori GLP-1. Di conseguenza, la ratio della dose di Tirzepatide con quella di Semaglutide risulta maggiormente a carico della prima.
La Tirzepatide è attualmente approvata dalla FDA per l’uso in persone in sovrappeso o con obesità, indipendentemente dal fatto che soffrano o meno di diabete di tipo II. Tuttavia, alcuni studi suggeriscono che la Tirzepatide sia un farmaco che non ha bisogno di essere somministrato in caso di mancanza della condizione diabetica.
Negli studi dove la Tirzepatide è stata somministrata a soggetti obesi, sono comunque stati osservati miglioramenti in tutte le misure cardiometaboliche. Gli eventi avversi più comuni con la Tirzepatide sono i medesimi riscontrati con la Semaglutide o altri membri della stessa famiglia. Essi sono stati di tipo gastrointestinale e la maggior parte di questi sono stati di gravità lieve o moderata e si sono verificati principalmente durante l’aumento della dose. Gli eventi avversi hanno causato l’interruzione del trattamento nel 4,3%, 7,1%, 6,2% e 2,6% dei partecipanti che hanno ricevuto dosi di Tirzepatide da 5, 10 e 15mg e placebo, rispettivamente.
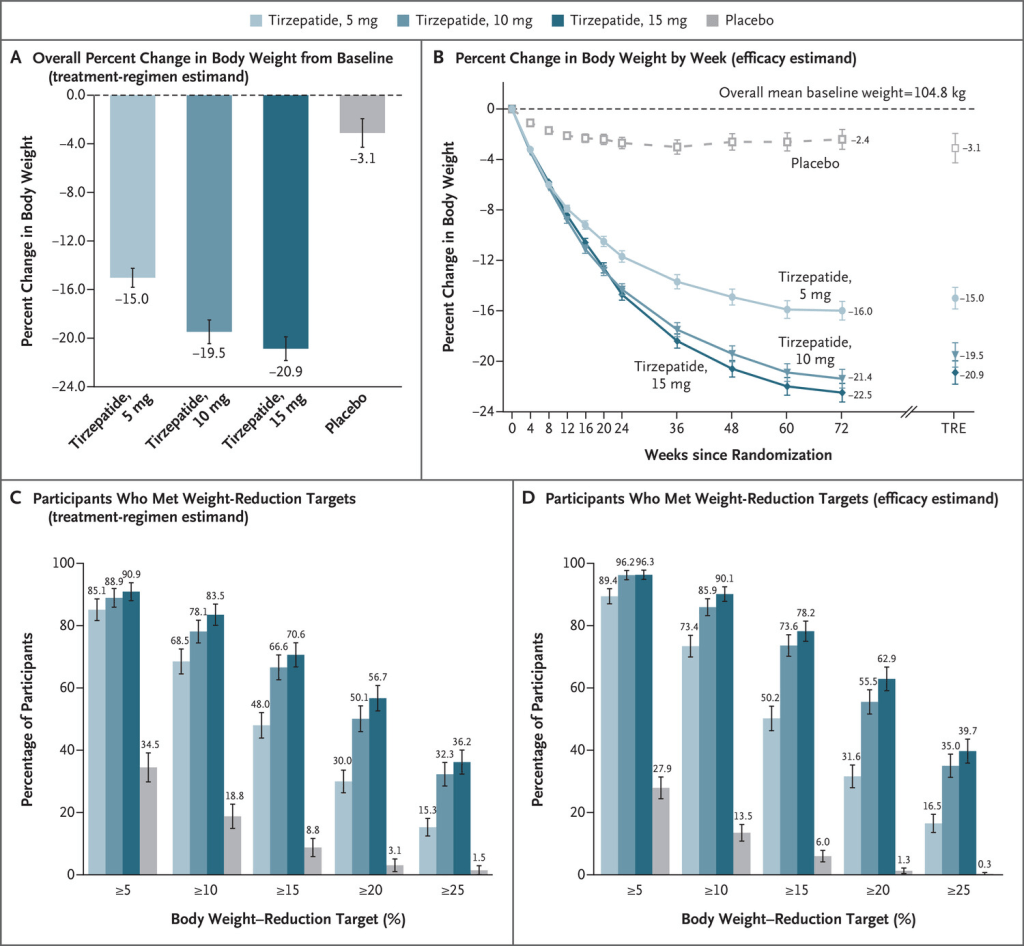
In uno studio di 72 settimane su partecipanti con obesità, 5mg, 10mg o 15mg di Tirzepatide una volta alla settimana hanno fornito riduzioni sostanziali e durature del peso corporeo.
Effetto della Tirzepatide somministrata una volta alla settimana, rispetto al placebo, sul peso corporeo. Le medie dei minimi quadrati sono presentate, se non diversamente specificato. Il pannello A mostra la variazione percentuale del peso corporeo dal basale alla settimana 72, derivata da un modello di analisi della covarianza per la stima del regime di trattamento (TRE). Il pannello B mostra la variazione percentuale del peso corporeo in base alle settimane dalla randomizzazione, derivata da un modello misto per misure ripetute (MMRM) per la stima dell’efficacia; sono riportate anche le stime alla settimana 72 per la stima del regime di trattamento. I riquadri C e D mostrano le percentuali di partecipanti che hanno avuto riduzioni di peso di almeno il 5%, 10%, 15%, 20% e 25% dal basale alla settimana 72. Per il riquadro C, la percentuale è stata calcolata in base al numero di settimane di randomizzazione. Per il pannello C, la percentuale è stata calcolata con l’uso delle regole di Rubin, combinando le percentuali dei partecipanti che hanno raggiunto l’obiettivo nei set di dati imputati. I valori mancanti alla settimana 72 sono stati imputati con MMRM se la mancanza era dovuta esclusivamente a Covid-19 e con imputazione multipla se la mancanza non era dovuta a Covid-19. Per il pannello D, la percentuale di partecipanti che hanno raggiunto gli obiettivi di riduzione del peso è stata ottenuta dividendo il numero di partecipanti che hanno raggiunto i rispettivi obiettivi alla settimana 72 per il numero di partecipanti con un valore al basale e almeno un valore post-base non mancante. I valori mancanti alla settimana 72 sono stati imputati dall’analisi MMRM. Le barre 𝙸 indicano gli intervalli di confidenza al 95%.
Nella pratica su bodybuilder, si sono osservati i minori sides comparati a ottimi risultati su insulino-resistenza e riduzione contenuta della fame con dosaggi settimanali di 2.5mg.
Alla luce di ciò, e in contesto aspecifico, la Tirzepatide mostra un moderato vantaggio gestionale rispetto alla Semaglutide.
Si è parlato di un ipotetico “rebound” di Grelina con incremento della fame e del consumo calorico (con aumento di peso) dopo la cessazione d’uso di Semaglutide o Tirzepatide. Al momento non esistono dati certi che ci indichino un reale collegamento equazionale tra cessazione d’uso di GLP-1 agonisti > picco in cronico della Grelina > iperfagia > aumento ponderale di peso. Sappiamo, però, che le variazioni di Grelina e GLP-1 a 6 mesi dalla cessazione di una dieta ipocalorica non hanno predetto il recupero del peso da 6 a 18 mesi. Ciò significa che, in un soggetto sano, potrebbe si esserci una maggiore attività della Grelina nelle prime settimane dopo cessazione d’uso di un incretino-mimetico (calo soglia ematica del farmaco e stabilizzazione dei livelli endogeni di GLP-1), ma l’aumento del peso successivo potrebbe risultare con maggiore probabilità dalla modifica omeostatica multi-fattoriale la quale, per trovare un nuovo equilibrio, richiede per lo meno un arco di tempo direttamente proporzionale al tempo di trattamento. Si consideri, inoltre, che un anno dopo la sospensione della Semaglutide sottocutanea a 2,4mg una volta alla settimana e dell’intervento sullo stile di vita, i soggetti possono mostrare una riacquisizione di due terzi della perdita di peso precedente, con cambiamenti simili nelle variabili cardiometaboliche. Qualcosa di un possibile rebound grelinico…
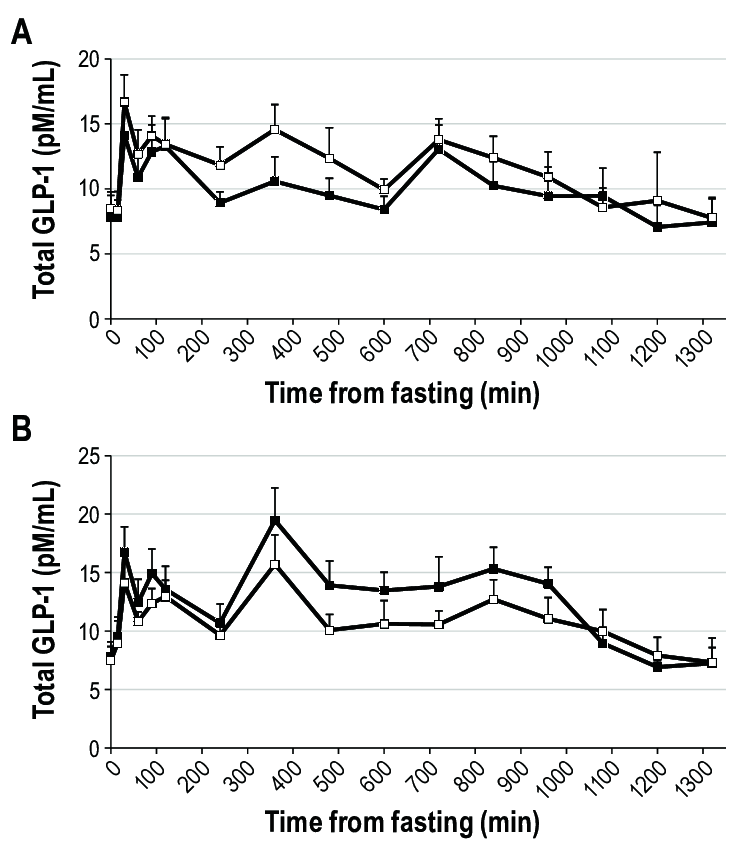
Concentrazione totale di GLP-1 durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli mattutini (P = 0,10) e notturni (P = 0,12) tendevano a essere più bassi e i livelli pomeridiani erano significativamente più bassi (P = 0,016) durante il sonno breve rispetto al sonno abituale nelle donne, mentre negli uomini le concentrazioni pomeridiane di GLP-1 tendevano a essere più alte dopo il sonno breve rispetto al sonno abituale (P = 0,10). I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.Concentrazione di Grelina totale durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli di Grelina mattutina più elevati sono stati osservati dopo il sonno breve rispetto al sonno abituale negli uomini; nelle donne non sono state osservate differenze tra i periodi di sonno. I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.
Conclusioni:
Arrivati alla conclusione di questa disamina abbiamo tutti gli elementi per valutare l’eventuale senso di utilizzo degli Incretino-Mimetici in contesti al di fuori del trattamento del diabete di tipo II o di soggetti obesi.
Le limitazioni date dagli effetti collaterali più comuni, se contestualizzate in una preparazione di un bodybuilder, possono causare non poche problematiche specie nelle vicinanze di una gara; vedi, ad esempio, estrema riduzione dell’assunzione calorica e proteica, mal assorbimento e gonfiore addominale o paresi gastrica.
Sebbene la possibilità di eventi ipoglicemici sia contenuta, e ancor più rara con la Tirzepatide in monoterapia, il rischio, in concomitanza dell’effetto sulla insulino-sensibilità degli AAS o di altre molecole co-somministrate, di questo sides può aumentare in modo sensibile durante una dieta ipocalorica.
In tal sede non ho preso in considerazione i due più preoccupanti, e più rari, effetti collaterali legati all’uso di incretino-mimetici: Tumore Midollare della Tiroide [MTC] e Pancreatite. Quest’ultima può manifestarsi anche con l’uso di AAS, seppur raramente, specie in caso d’uso di molecole aromatizzabili; la presenza di un incretino-mimetico in tali circostanze potrebbe avere un incidenza maggiore nello sviluppo e manifestazione della Pancreatite.[https://jmedicalcasereports.biomedcentral.com/]
In definitiva, i vantaggi potenziali di una una insulino-sensibiltà maggiore iatrogeno-dipendente (visti in precedenza) con l’uso di Incretino-Mimetici è, con i dovuti distinguo complessivi, ottenibile con l’uso di Metformina la quale presenta un margine di sicurezza decisamente più ampio.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Semaglutide and Tirzepatide are More Than Just Weight Loss Drugs [di Type-IIx]
[1] McDonald, L. The Ultimate Diet 2.0: Advanced Cyclical Dieting for Achieving Super Leanness. (2003). Lyle McDonald Publishing.
[2] Volpe S, Lisco G, Racaniello D, Fanelli M, Colaianni V, Vozza A, Triggiani V, Sabbà C, Tortorella C, De Pergola G, Piazzolla G. Once-Weekly Semaglutide Induces an Early Improvement in Body Composition in Patients with Type 2 Diabetes: A 26-Week Prospective Real-Life Study. Nutrients. 2022 Jun 10;14(12):2414. doi: 10.3390/nu14122414.
[3] Kolb H, Kempf K, Röhling M, Martin S. Insulin: too much of a good thing is bad. BMC Med. 2020;18(1):224. Published 2020 Aug 21. doi:10.1186/s12916-020-01688-6
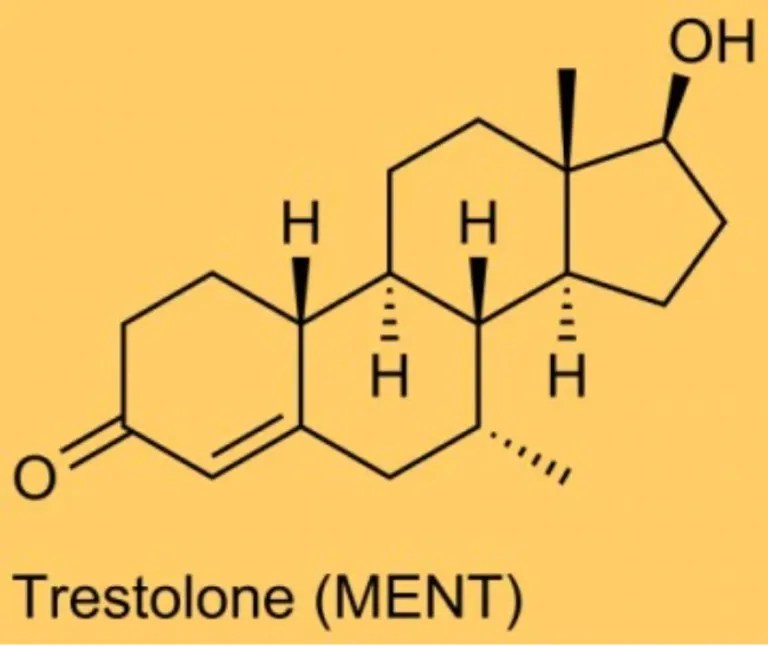
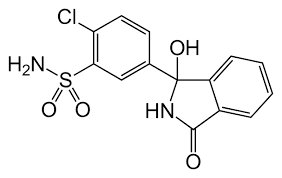
Del Trestolone [MENT; Methylnortestosterone; 7α-methyl-19-nortestosterone; 7α-CH₃-NorT) ho già parlato sia in un articolo che in un video della rubrica “The Lord Of The PEDs”. In entrambi i lavori sono stati esposti in generale pregi e difetti della molecola in questione. In questo articolo, invece, vorrei soffermarmi sulle caratteristiche uniche del Trestolone, le quali, e lo si vedrà, non gli conferiscono particolari e reali vantaggi d’uso nel Bodybuilding, a differenza, per esempio, del Trenbolone, dell’Oxandrolone o del Fluoxymesterone. Piuttosto, le sue peculiarità si combinano in modo sfavorevole per qualsiasi uso pratico nel bodybuilding, attraverso l’influenza sullo sviluppo della ginecomastia, dell’equilibrio dei liquidi e della pressione sanguigna.
Le caratteristiche uniche del Trestolone:
Effetti estrogenici per aromatizzazione al prodotto aromatico più potente del 17β-estradiolo: il 7α-methylestradiolo (7α-ME);
Effetti gestageni dovuti alla forte attivazione del recettore del Progesterone (PR):
Che si combinano sinergicamente per produrre:
1. Effetti ginecomastici (crescita del tessuto mammario negli uomini)
2. Effetti edematosi (ritenzione di liquidi; “gonfiore”) e
3. Effetti ipertensivi (pressione sanguigna elevata, in particolare sistolica, cioè pressione da contrazione cardiaca).
Prima di discutere gli effetti edematosi del Trestolone (ritenzione di liquidi; “gonfiore”), dobbiamo innanzitutto esaminare i fattori che sono alla base della ritenzione di liquidi (effetti estrogenici e gestagenici/progestinici), prima di discutere il modo in cui questi stessi fattori sono alla base dei suoi particolari effetti ipertensivi (aumento della pressione arteriosa, in particolare della pressione sistolica; cioè, aumento della pressione da contrazione cardiaca).
Effetti estrogenici:
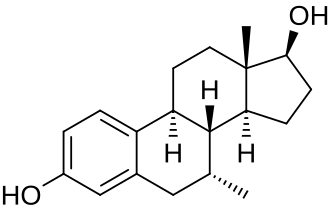
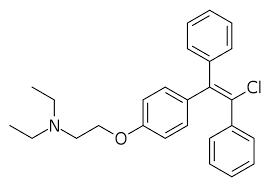
Struttura molecolare del 7α-Methylestradiolo
Il lettore nella media, ormai, dovrebbe essere al corrente sul fatto che il Trestolone aromatizza in 7α-methylestradiolo (7α-ME). [1].
Dalla Teoria delle Potenze Estrogeniche Dipendenti dai Composti (Per-AAS) e Individuali (Per-Utilizzatore) (Modello di Type-IIx), l’estrogenicità si riferisce agli effetti associati all’attivazione di ERα e β (quest’ultima generalmente opposta alla prima), e dipende da Fattori Dipendenti dai Composti (Per-AAS) e Individuali (Per-Utilizzatori) che determinano sia (A) i livelli ematici effettivi che (B) gli effetti a livello tissutale dei prodotti aromatici di un AAS.
Tabella 1: Il più potente di tutti, il 7α-ME produce ½ della risposta di crescita massima (EC₅₀) nel tessuto mammario umano (T47Dco) a una sola attivazione: una concentrazione di 4,4 picomoli (pM). Come il Trenbolone, ma in modo peggiore, esso fa di più con minor quantità.[1]
Tuttavia, in letteratura, viene riportato che il Trestolone possiede solo una debole attività estrogenica data da un tasso di aromatizzazione che sembrerebbe essere insufficiente per scopi sostitutivi, come dimostrato dalla diminuzione della densità minerale ossea negli uomini trattati con questo farmaco per l’ipogonadismo. Dobbiamo, però, anche considerare il tasso di degradazione del 7α-methylestradiolo prodotto, nonché la potenza estrogenica dei metaboliti risultanti che vanno a sommarsi alla potenziale attività estrogenica di altri AAS aromatizzabili co-somministrati [vedi Oxymetholone e la sua attività estrogenica intrinseca e il Methandrostenolone con il suo metabolita estrogenico 17α-methylestradiolo] e l’attività gestatinica propria del Trestolone.
Il prodotto aromatico del Trestolone, il 7α-ME, ha una potenza più che quadrupla (“efficace”, una cosa negativa in questo caso) rispetto al 17β-estradiolo (E2) nelle cellule con presenza di ER.[1] L’efficacia si determina misurando l’effetto, ad esempio la crescita (in questo caso, nelle cellule di cancro al seno). La EC₅₀ (EC50) è determinata dalle concentrazioni alle quali il ligando innesca la crescita e può essere confermata da misurazioni della progressione del ciclo cellulare (cioè l’ingresso nella fase S durante il ciclo cellulare).
L’affinità di legame (IC₅₀) del prodotto aromatico del Trestolone, 7α-ME, è pari al 102% di quella dell’E2, che in letteratura viene tipicamente utilizzato come composto di riferimento per il legame con l’ER, data la sua notevole efficacia, potenza e affinità per il recettore ER-α. [1].
Confrontando il tasso di aromatizzazione tra Trestolone e Nandrolone, Attardi et al. hanno scoperto che, “[a] 180 min, circa il 23% del Trestolone è stato convertito in 7α-ME e circa il 13% del [Nandrolone] in E2”. Poiché il Nandrolone ha un tasso di aromatizzazione del 20% rispetto al Testosterone (T), e che presenta una maggior tendenza alla conversione in Estrone (E1), possiamo dedurre che il Trestolone aromatizza in 7α-ME circa il 35% rispetto al T [che aromatizza in E2], con una potenza quattro volte superiore a quella dell’E2, cioè per provocare la crescita delle cellule del cancro al seno. La semplice moltiplicazione del tasso di aromatizzazione (35%) × EC50(7α-methylestradiolo) × RBA(7α-methylestradiolo) ≈ Il potenziale di crescita del Trestolone nelle cellule con presenza di ER è superiore del 40% rispetto al T. [2]
Differenze strutturali tra una molecola di Estrone (E1) e di Estradiolo (E2)
La deduzione, quindi, supporta le segnalazioni degli utilizzatori secondo cui il Trestolone è potentemente estrogenico. Matematicamente, possiamo affermare che 50mg al giorno di Trestolone Enantato ≈ estrogenico quanto 500mg di Testosterone Enantato alla settimana.
Inoltre, come descritto nella sezione seguente, gli effetti gestageni del Trestolone potenziano notevolmente i suoi effetti ipertensivi ed edematosi (tendenza a trattenere liquidi).
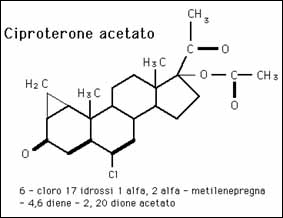
Il Trestolone è un androgeno potentemente progestinico (“ gestagenico”) che possiede il 27,5% della potenza di Androcur™ [Ciproterone Acetato] – un farmaco antiandrogeno e progestinico usato per trattare le patologie androgeno dipendenti, tra cui l’acne, l’irsutismo e il cancro alla prostata – per attivare il recettore del progesterone (PR). [11].
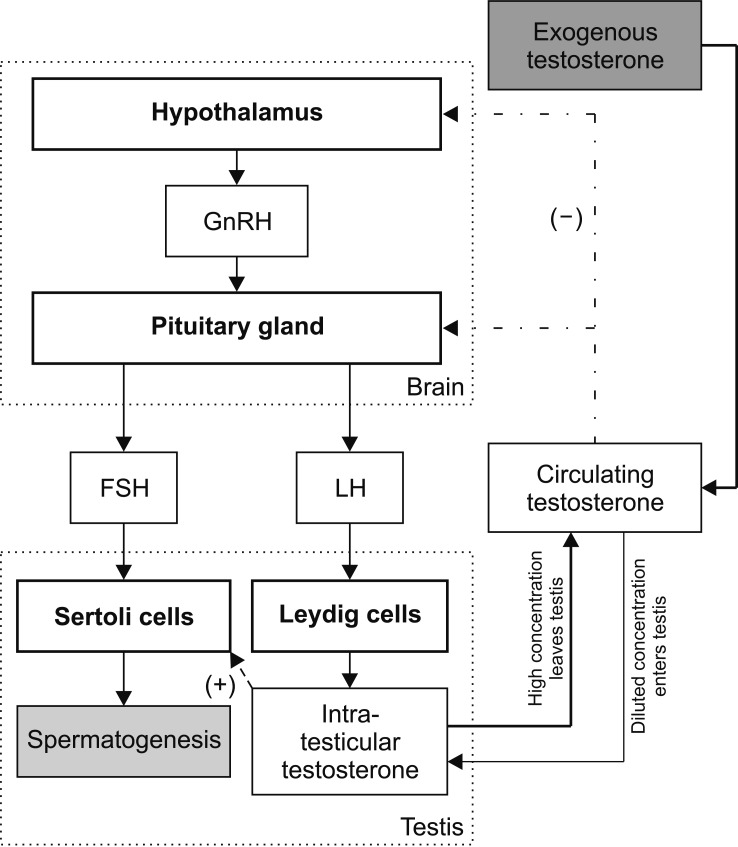
I progestinici contribuiscono alla soppressione dell’asse ipotalamo-ipofisi-gonadi (HPG) disregolando le gonadotropine ipotalamiche attraverso la segnalazione del dendro KNDy, interrompendo la pulsatilità del GnRH e inibendo la secrezione di LH ipofisario [e FSH], inibendo così la sintesi e la secrezione di Testosterone (endogeno). [12][13] I progestinici sintetici utilizzati in ambito contraccettivo traggono la loro efficacia da questa caratteristica. Bebb et al. hanno randomizzato uomini sani a ricevere Testosterone Enantato (100mg settimanali) o lo stesso dosaggio di Testosterone Enantato in combinazione con il progestinico Levonorgestrel, la cui aggiunta ha praticamente soppresso la secrezione di LH e FSH. [14] La diminuzione di LH e FSH può causare ipogonadismo secondario, con conseguente diminuzione del rapporto androgeni/estrogeni (A:E), causando ginecomastia. [15]
Gli effetti dei progestinici sono legati alle loro interazioni con i recettori: recettori degli androgeni (AR) (ad esempio, acne, effetti lipidici); recettori dei glucocorticoidi (GR) (ad esempio, ritenzione di sodio e acqua, gonfiore); o recettori dei mineralocorticoidi (MR) (ad esempio, diminuzione della ritenzione idrica e del peso). I progestinici antiandrogeni possono agire in diversi modi. Possono esercitare un’inibizione competitiva dell’AR, oppure legarsi all’enzima 5-α reduttasi e quindi interagire con la conversione del Testosterone in DHT (il suo metabolita fortemente androgeno)[16].
Il Progesterone e i suoi derivati e i “progestinici-mimici” (ad esempio, il Trestolone) si legano moderatamente all’AR in modo competitivo (cioè antagonista). [17]. I derivati del Progesterone alterano le risposte tissutali mediate da AR e PR, ma non da ER. [17]
Gli estrogeni regolano la sintesi di PR. [18] Inoltre, l’attivazione del PR è stata collegata a una ridotta espressione dell’AR, ostacolando così l’inibizione della crescita del tessuto mammario mediata dagli androgeni osservata in condizioni di normale omeostasi ormonale. [19].
Il Progesterone e i suoi derivati possono ulteriormente ma indirettamente causare ginecomastia potenziando l’effetto dell’E2 sui tessuti mammari. [20].
In sintesi, le caratteristiche discusse – effetti estrogenici e gestagenici del Trestolone – sono alla base dei suoi potenti effetti edematosi e ipertensivi.
Effetti edematosi:
L’edema, o ritenzione di liquidi, è il metro di paragone dell’eccessiva estrogenicità nei bodybuilder che fanno uso di AAS.
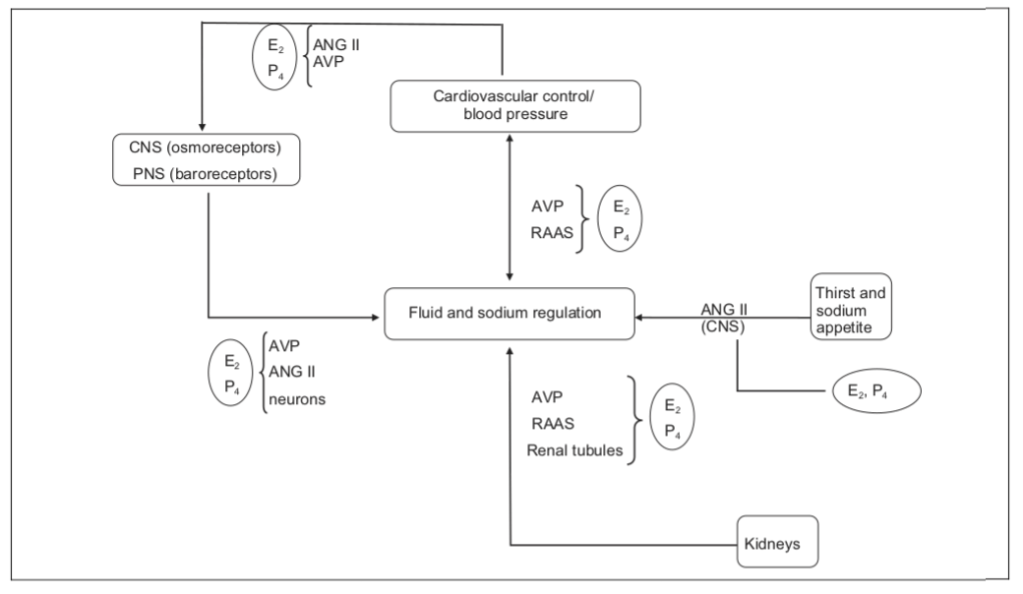
Figura 2: Schema che illustra il controllo complesso dell’equilibrio dei liquidi e del sodio e i molteplici modi in cui l’Estradiolo (E2) e il Progesterone (P4) possono influenzare questi processi. Adattato con il permesso di Stachenfeld. La regolazione dei fluidi e del sodio è controllata da una serie di sistemi complessi, tutti influenzati da estrogeni e Progesterone. Sia il sistema nervoso centrale (SNC) che il sistema nervoso periferico (PNS) contribuiscono alla regolazione dei fluidi; gli estrogeni e i progestinici possono influenzare la regolazione dei fluidi direttamente attraverso il cervello o indirettamente influenzando le azioni dell’Angiotensina II (ANG II) e dell’Arginina Vasopressina (AVP) e dei cambiamenti negli ormoni che regolano il sodio (Aldosterone e Renina [RAAS]). Estradiolo e Progesterone/Progestinici, possono anche aumentare l’Angiotesina II cerebrale mediato nel cervello e aumentare l’importante effetto stimolante di questo ormone sulla sete e sull’assunzione di liquidi. Infine, sia E2 che P4, influenzano la regolazione del sodio e dell’acqua nei tubuli distali del rene. Questo impatto può verificarsi direttamente sui tubuli o sia attraverso l’AVP che il RAAS e contribuire alla ritenzione idrica.
Il Trestolone si sostituisce a E₂ e P₄ per promuovere l’equilibrio dei fluidi attraverso molteplici meccanismi. [10]
Il diagramma qui sopra illustra come il Trestolone agisca in modo molteplice per promuovere la ritenzione di liquidi (edema) e l’ipertensione (aumento della pressione arteriosa sistolica; aumento della pressione da contrazione cardiaca). Il Trestolone è analogo all’E₂ (Estradiolo; E2) e al P₄ (Progesterone) nei confronti del 7α-ME e nella sua potenza del 29,2% nell’attivare la PR come il P₄. [11]
Il Trestolone, quindi, promuove la ritenzione di liquidi agendo su:
1. Il cervello e il sistema nervoso centrale (SNC) per aumentare la sete e il bisogno di assumere sodio attraverso la segnalazione dell’Angiotensina II, e 2. I reni agendo su: 1. gli ormoni antidiuretici, ad esempio l’Arginina Vasopressina (AVP), riducendo la minzione 2. il sistema renina-angiotensina-aldosterone (RAAS), aumentando il bilancio del sodio, e/o 3. i tubuli renali, favorendo la ritenzione di sodio e acqua. [10]
Effetti ipertensivi:
Nella Figura 1 è illustrato anche, in parte, come il Trestolone, al posto dell’E₂ (Estradiolo; E2) e del P₄ (Progesterone), favorisca l’aumento della pressione arteriosa. Lo fa aumentando lo squilibrio dei liquidi (cioè il “gonfiore”), come i suoi effetti edematosi, e agendo sugli osmorecettori del sistema nervoso centrale (SNC) e sui barorecettori del sistema nervoso periferico (SNP), nonché agendo sull’Angiotensina II, sull’AVP e sui neuroni di vari sistemi. [10]
– Aumento della pressione sistolica
Il Trestolone aumenta significativamente la pressione arteriosa sistolica (cioè la pressione da contrazione cardiaca) a una dose settimanale inferiore a 2mg. [21]
La pressione del polso (Pp) è la differenza (mmHg) tra pressione sistolica e diastolica. Una pressione sistolica normale è quindi di 40 mmHg (120 mmHg – 80 mmHg = 40 mmHg).
La pressione sistolica , o del polso (Pp), rappresenta la forza pressoria che il cuore genera ogni volta che si contrae, o la compliance arteriosa (C). Se la pressione del polso è normale a 40 mmHg, una Pp < 25% della pressione sistolica è bassa o ristretta, mentre una Pp > 100 mmHg è alta o estesa.
Una variazione della pressione sistolica (ΔPp) è proporzionale alla variazione del volume (V) (ΔV) ma inversamente proporzionale alla compliance arteriosa (C):
ΔPp = Δ V/C
Poiché la variazione di volume è dovuta al volume della gittata (SV) del sangue espulso dal ventricolo sinistro, possiamo approssimare la pressione del polso come:
Pp = SV/C
Un giovane adulto normale a riposo ha un volume di gittata (SV) di circa 80 mL. La compliance arteriosa (C) è di circa 2mL/mmHg, il che conferma che la pressione normale del polso è di circa 40 mmHg.
Il Trestolone, quindi, induce un aumento della pressione sistolica, aumentando il volume della gittata.
Effetti ematologici:
– Aumento del Ematocrito [HCT]
Il Trestolone, aumentando la ritenzione di sodio e di liquidi, e aumenta il volume plasmatico. Inoltre, aumenta rapidamente l’Emoglobina.
L’Emoglobina (Hb) è una proteina che si lega agli eritrociti (RBC) all’O₂ (Hb 13,5 – 17g/dL [uomini], 12 – 15,5g/dL [donne]).
L’Ematocrito (HCT) rappresenta la % del volume sanguigno occupato dagli eritrociti (RBC) [uomini 41-51%, donne 36-47%].
L’Ematocrito (HCT) è correlato all’Emoglobina (Hb) mediante la formula di base:
Hb (g/dL) × 3 ≈ HCT (%)
Esempio: Hb di 15g/dL ≈ HCT del 45%.
I livelli di emoglobina sono stati significativamente aumentati (149 ± 2,9 g/L → 154 ± 3,3; dimensione dell’effetto: 1,724; %Δ: +3,35%; intervallo di confidenza del 95%) con il Trestolone (~ 2 mg q.w.) a 12 settimane, e un andamento simile (da 0,44 a 0,46; dimensione dell’effetto: 2; %Δ: +4,5%; non significativo) è stato osservato nell’Ematocrito, che però non ha raggiunto la significatività statistica. [21] Nel gruppo Testosterone (~ 120mg q.w.), invece, è stato osservato un aumento progressivo più lento della concentrazione di Emoglobina, che è diventato significativo solo a 48 settimane. Nel gruppo Testosterone si è registrato anche un aumento complessivo significativo dell’Ematocrito, sebbene nessuno dei singoli punti di trattamento fosse significativamente diverso dal pre-trattamento (0,45 ± 0,01). [21]
Conclusioni:
Le caratteristiche di base del Trestolone – la sua potente estro- e gesta- genicità – pongono le basi per i suoi forti effetti edematosi e ipertensivi, in modo tale che i suoi effetti ginecomastici, gli effetti ginecomastici derivanti dall’estrogenicità e gli effetti ginecomastici derivanti dall’antiandrogenicità, non possono essere ignorati. Attraverso il suo metabolita aromatico 7α-ME, sopprime potentemente le gonadotropine (LH, FSH), diminuendo la A:E ratio, sinergizzando con le caratteristiche gestageniche con una potenza pari a quella degli antiandrogeni farmaceutici (ad esempio, Androcur™ [Ciproterone Acetato]), stimolando la crescita direttamente del tessuto mammario attraverso l’ER e indirettamente attraverso l’azione gestagenica e antiandrogena nella modalità del Progesterone (con una potenza di quasi ⅓ per mg). Il Trestolone favorisce in particolare il “gonfiore da ritenzione” attraverso la sua azione sui reni (influenzando negativamente la regolazione della ritenzione di liquidi e sodio) e sul cervello (aumentando la sete e l’ingestione di sodio) e favorisce in particolare l’ipertensione, soprattutto l’aumento della pressione sistolica, aumentando il volume di gittata attraverso l’aumento del volume plasmatico e dell’Ematocrito, cioè la viscosità o lo densità del sangue.
Questo articolo potrebbe prevedibilmente servire da deterrente all’uso di questo agente praticamente inutile (se paragonato alle altre molecole AAS) da parte dei bodybuilder dal momento che presenta alcuni aspetti unici e non accettabili nell’insieme di una corretta valutazione della molecola.
Se lo confrontiamo con il Trenbolone, non abbiamo alcun vantaggio nel suo inserimento sostitutivo: oltre al potenziale neurotossico e cardiotossico, come abbiamo visto, si aggiungono problematiche peculiari date dalla molecola che ne rendono un ipotetico uso nettamente difficile da gestire.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Articles by Type-IIx
[1] Attardi BJ, Pham TC, Radler LC, Burgenson J, Hild SA, Reel JR. Dimethandrolone (7alpha,11beta-dimethyl-19-nortestosterone) and 11beta-methyl-19-nortestosterone are not converted to aromatic A-ring products in the presence of recombinant human aromatase. J Steroid Biochem Mol Biol. 2008;110(3-5):214-222. doi:10.1016/j.jsbmb.2007.11.009
[2] Ryan, Kenneth J. “Biological aromatization of steroids.” Journal of Biological Chemistry 234.2. (1959): 268-272.
[3] Navarro, V. M., Gottsch, M. L., Chavkin, C., Okamura, H., Clifton, D. K., and Steiner, R. A. (2009). Regulation of Gonadotropin-Releasing Hormone Secretion by Kisspeptin/Dynorphin/Neurokinin B Neurons in the Arcuate Nucleus of the Mouse. Journal of Neuroscience, 29(38), 11859–11866. doi:10.1523/jneurosci.1569-09.2009
[4] Girmus, R. L., and Wise, M. E. (1992). Progesterone Directly Inhibits Pituitary Luteinizing Hormone Secretion in an Estradiol-dependent Manner1. Biology of Reproduction, 46(4), 710–714. doi:10.1095/biolreprod46.4.710
[5] Bebb, R. A., Anawalt, B. D., Christensen, R. B., Paulsen, C. A., Bremner, W. J., and Matsumoto, A. M. (1996). Combined administration of levonorgestrel and testosterone induces more rapid and effective suppression of spermatogenesis than testosterone alone: a promising male contraceptive approach. The Journal of Clinical Endocrinology and Metabolism, 81(2), 757–762. doi:10.1210/jcem.81.2.8636300
[6] Sitruk-Ware, R. (2004). Pharmacological profile of progestins. Maturitas, 47(4), 277–283. doi:1
0.1016/j.maturitas.2004.01.001
[7] Fang, H., Tong, W., Branham, W. S., Moland, C. L., Dial, S. L., Hong, H., … Sheehan, D. M. (2003). Study of 202 Natural, Synthetic, and Environmental Chemicals for Binding to the Androgen Receptor. Chemical Research in Toxicology, 16(10), 1338–1358. doi:10.1021/tx030011g
[8] Eyster, K. M. (Ed.). (2016). Estrogen Receptors. Methods in Molecular Biology. doi:10.1007/978-1-4939-3127-9
[9] Sansone, A., Romanelli, F., Sansone, M., Lenzi, A., and Di Luigi, L. (2016). Gynecomastia and hormones. Endocrine, 55(1), 37–44. doi:10.1007/s12020-016-0975-9
[10] Zhou, J., Ng, S., Adesanya-Famuiya, O., Anderson, K., and Bondy, C. A. (2000). Testosterone inhibits estrogen-induced mammary epithelial proliferation and suppresses estrogen receptor expression. The FASEB Journal, 14(12), 1725–1730. doi:10.1096/fj.99-0863com
[11] Houtman, C. J., Sterk, S. S., van de Heijning, M. P. M., Brouwer, A., Stephany, R. W., van der Burg, B., and Sonneveld, E. (2009). Detection of anabolic androgenic steroid abuse in doping control using mammalian reporter gene bioassays. Analytica Chimica Acta, 637(1-2), 247–258. doi:10.1016/j.aca.2008.09.037
[12] Navarro, V. M., Gottsch, M. L., Chavkin, C., Okamura, H., Clifton, D. K., and Steiner, R. A. (2009). Regulation of Gonadotropin-Releasing Hormone Secretion by Kisspeptin/Dynorphin/Neurokinin B Neurons in the Arcuate Nucleus of the Mouse. Journal of Neuroscience, 29(38), 11859–11866. doi:10.1523/jneurosci.1569-09.2009
[13] Girmus, R. L., and Wise, M. E. (1992). Progesterone Directly Inhibits Pituitary Luteinizing Hormone Secretion in an Estradiol-dependent Manner1. Biology of Reproduction, 46(4), 710–714. doi:10.1095/biolreprod46.4.710
[14] Bebb, R. A., Anawalt, B. D., Christensen, R. B., Paulsen, C. A., Bremner, W. J., and Matsumoto, A. M. (1996). Combined administration of levonorgestrel and testosterone induces more rapid and effective suppression of spermatogenesis than testosterone alone: a promising male contraceptive approach. The Journal of Clinical Endocrinology and Metabolism, 81(2), 757–762. doi:10.1210/jcem.81.2.8636300
[16] Sitruk-Ware, R. (2004). Pharmacological profile of progestins. Maturitas, 47(4), 277–283. doi:10.1016/j.maturitas.2004.01.001
[17] Fang, H., Tong, W., Branham, W. S., Moland, C. L., Dial, S. L., Hong, H., … Sheehan, D. M. (2003). Study of 202 Natural, Synthetic, and Environmental Chemicals for Binding to the Androgen Receptor. Chemical Research in Toxicology, 16(10), 1338–1358. doi:10.1021/tx030011g
[18] Eyster, K. M. (Ed.). (2016). Estrogen Receptors. Methods in Molecular Biology. Doi:10.1007/978-1-4939-3127-9
[19] Sansone, A., Romanelli, F., Sansone, M., Lenzi, A., and Di Luigi, L. (2016). Gynecomastia and hormones. Endocrine, 55(1), 37–44. doi:10.1007/s12020-016-0975-9
[20] Zhou, J., Ng, S., Adesanya-Famuiya, O., Anderson, K., and Bondy, C. A. (2000). Testosterone inhibits estrogen-induced mammary epithelial proliferation and suppresses estrogen receptor expression. The FASEB Journal, 14(12), 1725–1730. doi:10.1096/fj.99-0863com
[21] Walton, M. J., Kumar, N., Baird, D. T., Ludlow, H., & Anderson, R. A. (2007). 7 -Methyl-19-Nortestosterone (MENT) vs Testosterone in Combination With Etonogestrel Implants for Spermatogenic Suppression in Healthy Men. Journal of Andrology, 28(5), 679–688. doi:10.2164/jandrol.107.002683
La pressione alta, nota anche come ipertensione, è una delle cause più frequenti (che contribuiscono) di morte e complicazioni cardiovascolari nel mondo. Quando viene misurata, si divide in pressione sistolica (il numero superiore) e pressione diastolica (il numero inferiore). La pressione arteriosa sistolica è la pressione più alta raggiunta durante la contrazione del cuore, mentre la pressione diastolica è la pressione più bassa raggiunta durante il rilassamento del cuore. Una tipica lettura della pressione arteriosa potrebbe essere 120/80 mmHg, ovvero una pressione sistolica di 120 mmHg e una pressione diastolica di 80 mmHg. (L’unità di misura, millimetri di mercurio [Hg], risale a quando la pressione sanguigna veniva ancora misurata con manometri contenenti mercurio).
Per quantificare quanto sia grave l’ipertensione, diamo un’occhiata a un documento storico pubblicato su Lancet nel 2002 che, secondo Google Scholar, è stato citato ben 12.000 volte [1]. In questo lavoro, i ricercatori hanno riunito i dati dei singoli pazienti provenienti da 61 studi prospettici osservazionali. Questo studio comprendeva circa un milione di adulti senza precedenti malattie vascolari al basale. Per questo motivo, hanno avuto a disposizione dati davvero straordinari su cui lavorare e da cui trarre conclusioni.
Che cosa hanno dimostrato i dati? Hanno dimostrato che la mortalità per malattie coronariche e ictus aumenta con una pressione sistolica superiore a 115 mmHg e una pressione diastolica superiore a 75 mmHg. Ogni aumento di 20 mmHg della pressione arteriosa sistolica e di 10 mmHg della pressione arteriosa diastolica oltre questi valori raddoppia la mortalità per coronaropatia e ictus. In altre parole, chi ha una pressione arteriosa sistolica di 135 mmHg ha il doppio del rischio di morire per malattia coronarica o ictus rispetto a chi ha una pressione arteriosa sistolica di 115 mmHg. Si tratta di una differenza notevole. Questa relazione tra pressione arteriosa e mortalità, ad esempio per ictus, è illustrata nell’immagine sottostante:
Si noti che anche la probabilità di morire per ictus aumenta fortemente con l’aumentare dell’età. Il che ha senso, ovviamente. Sebbene non siano molte le persone che muoiono di ictus a 40 anni, è molto più comune negli anziani. Pertanto, l’aumento del rischio relativo di ipertensione diventa più rilevante con l’aumentare dell’età, poiché il rischio assoluto è molto più elevato.
Oltre a questo evidente aumento della mortalità a causa di eventi cardiovascolari, l’ipertensione provoca alterazioni strutturali e funzionali di diversi organi, danneggiandoli. Il danno agli organi bersaglio comprende, oltre al cuore e alla vascolarizzazione, il cervello, gli occhi e i reni. Il danno agli organi bersaglio può manifestarsi, oltre che con eventi cardiovascolari fatali e non fatali, con retinopatia, demenza, ischemia, albuminuria, glomerulopatia e ipertrofia ventricolare sinistra [2].
È chiaro che la pressione arteriosa elevata è dannosa per la salute.
Come influiscono gli steroidi anabolizzanti sulla pressione sanguigna?
Per rispondere alla domanda su come gli steroidi anabolizzanti influenzino la pressione arteriosa, si possono effettuare due tipi di studi. Un tipo di studio è costituito dagli studi prospettici interventistici. Questi, in sostanza, sono i più affidabili. Si prende un gruppo di persone, si somministra loro uno steroide anabolizzante e le si segue nel tempo per vedere cosa succede alla loro pressione sanguigna. Inoltre, si può includere un gruppo di controllo/placebo con cui confrontare i risultati (e se si randomizzano i soggetti si ottiene uno studio randomizzato-controllato). Sebbene questi studi siano sicuramente i migliori in termini di qualità delle prove, soffrono di un grosso inconveniente: non imitano correttamente l’uso reale, poiché i dosaggi sono inferiori a quelli utilizzati dalla maggior parte delle persone che fanno uso di steroidi anabolizzanti in modo illecito.
Detto questo, diamo un’occhiata a queste prove. Li ho riassunti nella tabella sottostante per fornire una buona panoramica:
Effetto degli steroidi anabolizzanti sulla pressione arteriosa in studi prospettici interventistici. ↑ significa un aumento statisticamente significativo, ? significa che non sono stati eseguiti test statistici, * significa rispetto al basale, † significa rispetto alla variazione nel gruppo placebo. Abbreviazioni: BP, pressione sanguigna; TE, testosterone enantato; Dbol, metandienone; ND, nandrolone decanoato.
L’unico studio che ha dimostrato un aumento statisticamente significativo della pressione arteriosa (sistolica) è stato quello di Freed et al [5]. In questo caso, sollevatori di pesi esperti hanno ricevuto 10mg o 25mg di Methandienone (Dianabol) al giorno per 6 settimane in doppio cieco controllato con placebo. La pressione arteriosa sistolica è aumentata significativamente di circa 9 mmHg. La pressione diastolica ha mostrato un leggero aumento di circa 4 mmHg, ma non è stato statisticamente significativo.
Gli altri studi non hanno eseguito test statistici [3,4] o non hanno rilevato cambiamenti statisticamente significativi rispetto al basale [6] o rispetto al cambiamento nel gruppo placebo [7, 8].
Come si può notare anche osservando i dosaggi, questi erano piuttosto bassi e non possono essere considerati rappresentativi dell’uso di steroidi anabolizzanti che si fa regolarmente in ambito del culturismo e simili. Per avere un’idea più precisa, si potrebbe ricorrere a studi prospettici osservazionali. In questi studi gli utilizzatori di AAS vengono seguiti nel tempo autosomministrando il proprio ciclo di AAS. Naturalmente, questi studi presentano anche degli inconvenienti. Uno di questi è il “policonsumo”. Non tutti gli steroidi anabolizzanti possono influire allo stesso modo sulla pressione arteriosa e, quando i consumatori di AAS li cumulano, è difficile dire quale steroide anabolizzante possa esserne responsabile. Per non parlare del fatto che è molto probabile che un consumatore di AAS stia somministrando steroidi anabolizzanti diversi da quelli che pensa di somministrare a causa di un’etichettatura errata. [9]. Inoltre, i consumatori di AAS potrebbero associarli a diversi altri tipi di farmaci, come l’rhGH, tiroidei, beta-agonisti e, al giorno d’oggi, la vasta gamma di farmaci sperimentali per il miglioramento delle prestazioni, il cui uso è in aumento.
Consideriamo anche brevemente alcuni studi prospettici osservazionali. Hartgens et al. hanno osservato gli effetti degli AAS autosomministrati per un periodo di 8 settimane in un piccolo gruppo di atleti di forza [10]. Prima dello studio, i soggetti, in media, non avevano fatto uso di AAS per quasi 8 mesi. Il dosaggio medio era relativamente basso, circa 400mg a settimana, il che mi fa pensare a quanto sia stato accurato. In ogni caso, la pressione arteriosa sistolica è aumentata da 131 a 139 mmHg. Il gruppo di controllo ha registrato un aumento da 129 a 134 mmHg. Quindi la variazione media rispetto al gruppo di controllo è stata di +3 mmHg. È stato osservato un piccolo aumento di 2 mmHg della pressione arteriosa diastolica, mentre il gruppo di controllo non ha registrato alcuna variazione. In ogni caso, le differenze non erano statisticamente significative.
Se si parte dal presupposto che gli AAS possono influenzare la pressione sanguigna e che questo fenomeno è completamente reversibile dopo la cessazione dell’uso, è possibile utilizzare un disegno di studio leggermente diverso. In altre parole, si potrebbe prendere un gruppo di utilizzatori mentre fanno uso di AAS, misurare la loro pressione sanguigna e poi misurarla di nuovo dopo un certo periodo di tempo, quando hanno smesso di usare gli AAS. Questo è esattamente il tipo di approccio che altri due gruppi hanno utilizzato [11, 12].
Uno di questi ha valutato tre gruppi: soggetti sedentari, bodybuilder che non fanno uso di AAS e bodybuilder che ne fanno uso [11]. I cicli di AAS duravano in media 8 settimane e, purtroppo, i dosaggi non possono essere ricavati con precisione dallo studio. Ciononostante, sembrano essere bassi. Subito dopo i cicli, la pressione sanguigna misurava 141/84 mmHg e 9 settimane dopo la cessazione dell’uso era 140/83 mmHg. Come riferimento, i bodybuilder che non ne facevano uso avevano una pressione sanguigna di 136/87 e i soggetti sedentari di 139/85 mmHg.
Palatini et al. hanno effettuato misurazioni della pressione arteriosa nelle 24 ore in un piccolo gruppo di consumatori di AAS [12]. I cicli duravano in media 9 settimane e il dosaggio era di circa 500mg settimanali. La pressione arteriosa era di 128/83 mmHg alla fine dei cicli e di 129/84 mmHg circa 12 settimane dopo la cessazione.
Nello studio HAARLEM, 100 consumatori di steroidi anabolizzanti sono stati seguiti nel tempo mentre si autosomministravano AAS [9]. Il dosaggio medio, basato sulle informazioni riportate sull’etichetta, era di 898mg a settimana, rendendo così il loro ciclo di AAS abbastanza rappresentativo dell’uso comune da parte dei bodybuilder. Le misurazioni sono state effettuate prima, durante, 3 mesi dopo la fine del ciclo e 1 anno dopo l’inizio del ciclo. I dati non pubblicati di questo studio hanno dimostrato un aumento di 7 mmHg della pressione sanguigna sistolica e di 3 mmHg della pressione sanguigna diastolica durante l’uso di steroidi anabolizzanti rispetto al basale [DL Smit, comunicazione personale]. Queste misurazioni sono tornate al valore basale dopo il ciclo. Data la dimensione relativamente ampia del campione di 100 utilizzatori di AAS e la natura osservazionale prospettica di questo studio, questa è attualmente la migliore stima della misura in cui gli AAS potrebbero influenzare la pressione sanguigna ai dosaggi comunemente utilizzati dai bodybuilder.
Nel complesso, si può concludere con cautela che i dosaggi sovrafisiologici di AAS possono aumentare transitoriamente la pressione arteriosa sistolica di circa 5-10 mmHg durante l’uso. È difficile dire in che misura questo aggravi il rischio cardiovascolare. Ma potremmo trarre qualche indizio da altri dati presenti nella letteratura scientifica. Ne parlerò nel prossimo articolo, in cui tratterò dei farmaci per abbassare la pressione sanguigna.ù
[altri] PEDs e pressione arteriosa:
Le evidenze nella ricerca e i dati aneddotici hanno mostrato un effetto ipertensivo legato all’uso di β-Agonisti sia non selettivi che selettivi. In particolare, è stato osservato che l’allele Gly16 del recettore adrenergico β-2 AR associato all’ipertensione. Questo effetto è stato osservato sia in trattamento con Salbutamolo che con Clenbuterolo, ed è responsivo ad alterazioni maggiori in base al dosaggio utilizzato [https://pubmed.ncbi.nlm.nih.gov/10373227/].
Anche l’Insulina può aumentare la pressione arteriosa attraverso diversi meccanismi. Per esempio, portando all’aumento del riassorbimento renale di sodio, all’attivazione del sistema nervoso simpatico, all’alterazione del trasporto ionico transmembrana e all’ipertrofia dei vasi di resistenza. Ovviamente si tratta di casi emersi, o possibili, in condizione di IR o alterazione subclinica del metabolismo glucidico e dell’attività biochimica dell’Insulina. Non è raro che bodybuilder in fase di Off-Season, con abuso di Insulina esogena e/o GH, ma anche in situazioni di non uso del peptide, presentino alterazioni pressorie correlati a sensibili aumenti di peso: tale causa vede anche la condizione di IR come co-fattore peggiorativo [https://pubmed.ncbi.nlm.nih.gov/].
In letteratura viene riportato che in seguito a somministrazione di rhGH si manifesti ritenzione idrica, e che essa sia un effetto collaterale concreto e documentato. Infatti, la maggior parte dei dati indicano che i pazienti adulti con deficit di hGH sono disidratati, cioè non hanno un volume d’acqua positivo nel corpo, e presentano una bassa concentrazione di acqua extracellulare nel plasma. Quando viene avviata la terapia sostitutiva del GH in questi pazienti i loro fluidi corporei vengono ripristinati alla normalità. La capacità di ritenzione dei fluidi del GH dovrebbe quindi essere considerata in ambito clinico come una normalizzazione fisiologica desiderabile dell’omeostasi dei liquidi corporei piuttosto che un effetto collaterale sgradevole [https://www.ncbi.nlm.nih.gov/pubmed/10592455]. Ovviamente, nel Bodybuilding le cose cambiano nettamente come l’incidenza quantitativa e anche indiretta sulla pressione vascolare di questo effetto sulla ritenzione idrica dato dall’uso di rhGH.
Si è ipotizzato che l’ormone alterato dall’uso di rhGH e che causa la ritenzione idrica possa essere l’Aldosterone. Nel qual caso, un diuretico antagonista come lo Spironolattone aiuterebbe. Il problema può essere risolto con del Lasix, il Furosemide (non è un consiglio!) , ma dal momento che l’esperienza sul campo non ha mostrato risoluzione al problema con queste pratiche, la domanda non ha così trovato una risposta chiara.
Da sinistra: Spironolattone e Furosemide
Un altra ipotesi indica una correlazione tra ritenzione idrica da rhGH e un aumento dell’ADH (Ormone Antidiuretico, conosciuto anche come vasopressina). Uno studio giunge alla conclusione che il hGH aumenta l’ADH [https://www.ncbi.nlm.nih.gov/pubmed/2405233], come effetto che trova la sua causa nella attivazione del sistema renina-angiotensina.
Il GH esogeno aumenta la Somatostatina, e dato che il rene possiede recettori specifici per la Somatostatina questi possono attivare il sistema renina-angiotensina [https://www.ncbi.nlm.nih.gov/pubmed/2405233]. Ciò può causare la ritenzione idrica da GH che può essere inibita da un ACE-inibitore.
I ricercatori, per vederci più chiaro, hanno studiato gli effetti di un preparato biosintetico autentico del hGH (bio-hGH) sul metabolismo del sodio e l’attività del sistema renina-angiotensina. Questa preparazione è stata somministrata a 6 giovani uomini ad un dosaggio di 0,2 U / kg / die per via sottocutanea per cinque giorni consecutivi. E’ stata effettuata la raccolta delle urine nelle ventiquattro ore per la misurazione dell’escrezione di sodio e l’osmolalità, ed è stato prelevato il sangue per quantificare i cambiamenti del sodio, dell’osmolalità, dell’attività della renina plasmatica (PRA), dell’aldosterone, e delle concentrazioni della arginina vasopressina (AVP). La somministrazione di Bio-hGH ha determinato un calo nelle 24 ore dell’escrezione urinaria di sodio (197 +/- 38 a 42 +/- 20 mmol, media +/- SD, P meno di 0,005), una riduzione del volume delle urine (1.652 + / – 182-848 +/- 348 mL, P inferiore a 0,05), ma non l’osmolalità. Il PRA è aumentato in modo significativo di 1.118 +/- 73 a 3.608 +/- 1.841 fmol angiotensina 1 L / s (P meno di 0,005), il che è stato associato con un aumento di sette volte nella concentrazione plasmatica (52 +/- 12-402 + / – 99 pg / mL, P meno di 0,001). L’osmolalità del plasma e le concentrazioni di AVP non sono cambiate in modo significativo. I risultati mostrano che la ritenzione di sodio indotta dal Bio-GH comporta l’attivazione del sistema renina-angiotensina. Questo meccanismo può spiegare in parte l’insorgenza dell’espansione del volume plasmatico e l’ipertensione e suggerisce un rischio di ritenzione di liquidi e, eventualmente, l’ipertensione nei soggetti trattati con dosi sovrafisiologiche di bio-hGH per il trattamento della bassa statura [http://www.dtic.mil/dtic/tr/fulltext/u2/611818.pdf].
Quindi, il rhGH provoca un rialzo del ADH(Ormone Antidiuretico). L’ADH è un costrittore coronarico molto potente che costringente vascolare/venoso. dato questo aumento del ADH, abbiamo la ritenzione idrica aumentata, e ciò provoca un aumento della pressione sanguinea. L’aumento del ADH causato dal rhGH è dose-dipendente.
L’uso della Clonidina è indicato se si soffre di pressione alta causata dal ADH.
Ovviamente, la somministrazione di rhGH, soprattutto se in concomitanza con AAS fortemente aromatizzabili peggiora considerevolmente la situazione collegandosi anche all’Insulino Resistenza e all’azione di alcuni AAS con i recettori mineralocorticoidi e sul danno endoteliale di questi.
La Clondina viene consigliata principalmente perché agisce direttamente sul ADH è la ritenzione idrica causata da questo ormone. Tuttavia, alcuni assumono il Furosemide (una molecola tutt’altro che sicura) per via della sua maggior potenza; ma non sembra lavorare tanto efficacemente nel complesso. Anche lo Spironolattone mostra una sua efficacia in tale circostanza con il suo effetto di diminuzione dell’attività dell’Aldosterone; riducendo quindi l’edema.
Ma la Clonidina risulta essere la prima scelta per contrastare l’effetto di un aumento del ADH dovuto alla somministrazione di rhGH esogeno. La sua somministrazione è solitamente indicata prima di dormire.
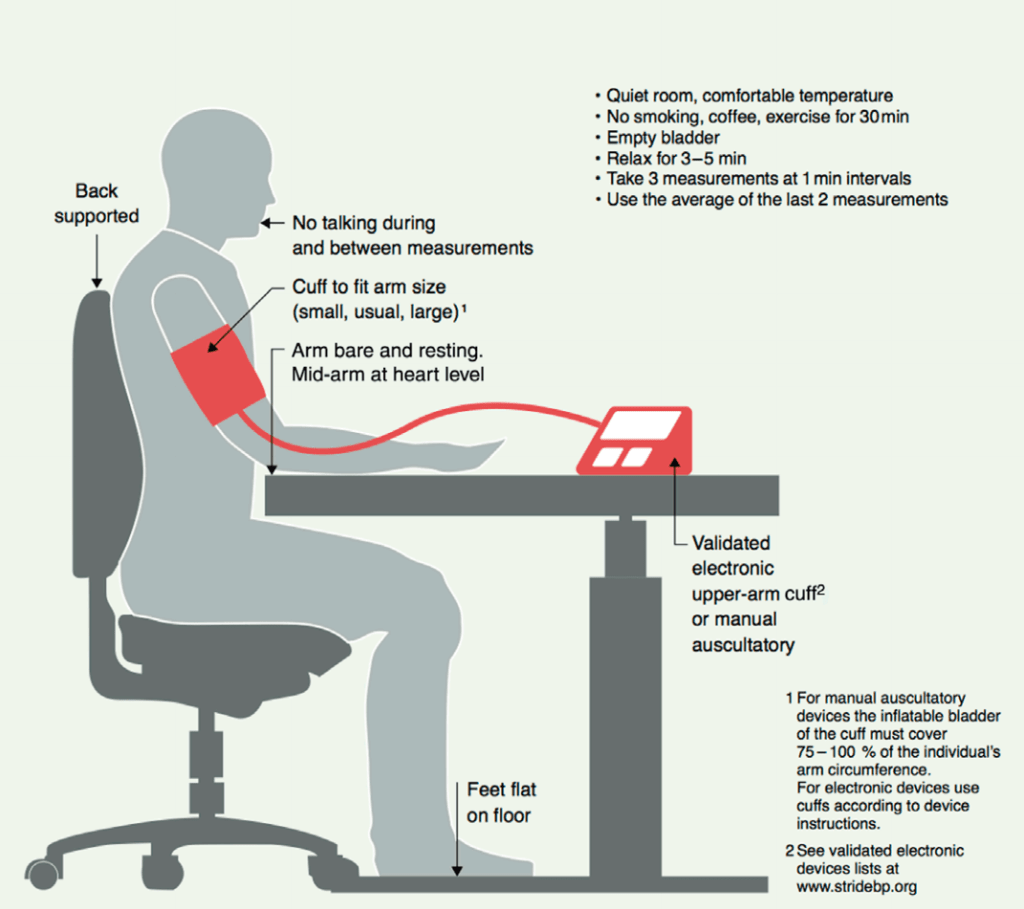
Come misurare la pressione arteriosa
In primo luogo, naturalmente, è necessario un dispositivo per misurare la pressione sanguigna a casa. Consiglio vivamente di utilizzare un apparecchio elettronico automatico che la misuri a livello della parte superiore del braccio. Questi dispositivi sono affidabili e richiedono la minima abilità, per cui c’è la minima possibilità di sbagliare la misurazione. Sono facilmente reperibili e costano circa 50 dollari. Valgono bene l’investimento. La maggior parte dei miei clienti utilizza dispositivi Omron, ma sono certo che esistono molte altre marche che producono dispositivi eccellenti.
Inoltre, assicuratevi che il misuratore di pressione sia dotato di un bracciale di dimensioni adeguate al vostro braccio. Di solito, i dispositivi per la misurazione della pressione arteriosa sono dotati di un bracciale di misura M, adatto a braccia con una circonferenza massima di 31-33 cm. Naturalmente, questa non è la circonferenza del braccio per i bicipiti flessi, ma la circonferenza quando il braccio è leggermente piegato senza essere flesso. La maggior parte degli utilizzatori di AAS ha braccia più grandi. Nella maggior parte dei casi è appropriato un bracciale di taglia L, che si adatta a braccia con circonferenza fino a 41-43 cm. Se siete molto grandi, potreste aver bisogno della taglia XL, che si adatta a braccia con circonferenza fino a circa 51-53 cm.
Un bracciale di dimensioni adeguate è importante perché, se troppo piccolo, potrebbe sovrastimare la pressione arteriosa. Ciò è ben illustrato in uno studio che ha esaminato le differenze di pressione arteriosa tra un bracciale di taglia M e uno di taglia L in 193 bodybuilder che partecipavano al Campionato Nazionale Messicano di Bodybuilding e Fitness [13]. Coloro che avevano braccia troppo grandi per il bracciale di taglia M (>33 cm) avevano una pressione sistolica più alta di 8,2 mmHg con questo bracciale rispetto al bracciale di taglia L. La pressione diastolica era più alta di 1,6 mmHg.
Anche un altro studio, condotto su individui obesi, ha sottolineato l’importanza di un bracciale di dimensioni adeguate [14]. Per ogni aumento di 5 cm della circonferenza del braccio oltre i 35 cm, si registrava un aumento di 2-5 mmHg della pressione arteriosa sistolica e di 1-3 mmHg della pressione arteriosa diastolica.
Ora che siete pronti a misurarla, come dovete fare? La Società Internazionale dell’Ipertensione ha un’ottima immagine che lo illustra [15], diamo un’occhiata:
Assicuratevi di non dover fare pipì, di non fumare (ovviamente non fumate, giusto?), di non aver bevuto caffè/caffeina o di aver fatto esercizio fisico 30 minuti prima della misurazione e di mettervi in una stanza tranquilla e confortevole per rilassarvi un paio di minuti. Siete seduti su una sedia che sostiene adeguatamente la vostra schiena dietro una scrivania. Si indossa il bracciale e si appoggia il braccio sulla scrivania, appoggiandolo completamente con la parte centrale del braccio all’altezza del cuore. Si appoggiano i piedi sul pavimento, non si accavallano le gambe e si batte il più forte possibile per far sì che il dispositivo misuri la pressione sanguigna. Ripetete la misurazione due volte con una piccola pausa tra l’una e l’altra e voilà. Prendete la media delle ultime due misurazioni e annotatela da qualche parte (la maggior parte degli apparecchi elettronici per la misurazione della pressione arteriosa ha anche una funzione di memoria, quindi potete evitare di scriverla).
Con quale frequenza si dovrebbero effettuare queste misurazioni? Le linee guida della Società Europea dell’Ipertensione per il monitoraggio domiciliare della pressione arteriosa raccomandano di farlo inizialmente almeno 3 e preferibilmente 7 giorni prima di considerare il trattamento della pressione arteriosa [16]. Le misurazioni dovrebbero essere effettuate sia al mattino che alla sera. Dopo questo periodo iniziale, è sufficiente misurarla circa una volta alla settimana.
Quando iniziare a trattare la pressione arteriosa
Dopo aver letto quanto fino a questo punto esposto, potreste pensare di dover trattare la pressione arteriosa quando è superiore a 115/75 mmHg. Tuttavia, una recente revisione sistematica e meta-analisi ha rilevato che, nella prevenzione primaria, l’abbassamento della pressione arteriosa riduce la mortalità e il rischio di malattie cardiovascolari solo se la pressione sistolica al basale è pari o superiore a 140 mmHg [17]. Se era inferiore a quella al basale, gli autori non sono riusciti a trovare alcun beneficio per quanto riguarda la mortalità o il rischio di malattie cardiovascolari. In effetti, questo è anche il motivo per cui la Società Europea dell’Ipertensione classifica l’ipertensione come una pressione arteriosa sistolica in ufficio pari o superiore a 140 mmHg e/o una pressione arteriosa diastolica pari o superiore a 90 mmHg [16]. L’ipertensione è definita come il livello di pressione arteriosa al quale i benefici del trattamento (con interventi sullo stile di vita o con farmaci) superano inequivocabilmente i rischi del trattamento, come documentato da studi clinici.
Va sottolineato che questa soglia di 140/90 mmHg riguarda le misurazioni della pressione arteriosa effettuate in ufficio. Queste sono di solito leggermente più alte rispetto alle misurazioni della pressione sanguigna effettuate a casa. Pertanto, la soglia per le misurazioni domiciliari della pressione arteriosa è definita come un valore medio di 135/85 mmHg [16].
Il trattamento dell’ipertensione produce chiari benefici clinici. Una meta-analisi mostra che ogni riduzione di 10 mmHg della pressione arteriosa sistolica riduce il rischio di eventi cardiovascolari maggiori del 20%, di malattia coronarica del 17%, di ictus del 27%, di insufficienza cardiaca del 28% e di mortalità per tutte le cause del 13% [18]. Sfortunatamente, il trattamento dell’ipertensione non annulla completamente tutti i rischi osservati nei grandi studi osservazionali. Due probabili ragioni sono: 1) l’ipertensione per periodi prolungati può danneggiare in modo irreversibile alcuni organi, e il trattamento non può annullare i danni subiti; 2) l’ipertensione spesso è associata a diverse altre comorbilità che possono influire sull’esito (ad esempio, l’obesità). Il primo motivo può essere affrontato iniziando il trattamento il prima possibile quando necessario, mentre il secondo non è particolarmente applicabile a un aumento della pressione sanguigna indotto da farmaci (come nel caso degli AAS). Tuttavia, non bisogna assolutamente dimenticare che, al di là del modesto aumento della pressione arteriosa, gli AAS hanno un impatto negativo sulla salute. Quindi, la correzione della pressione arteriosa, ovviamente, non annulla completamente i rischi per la salute degli AAS, così come non risolve i rischi per la salute di altre comorbidità che spesso vanno di pari passo con l’ipertensione.
Trattamento dell’ipertensione: modifiche allo stile di vita
Proprio come nella popolazione generale, ci possono essere alcuni cambiamenti nello stile di vita da adottare per ridurre la pressione sanguigna prima di ricorrere ai farmaci per abbassarla. Tuttavia, in alcuni casi i farmaci devono essere utilizzati immediatamente insieme ai cambiamenti dello stile di vita. La Società Europea dell’Ipertensione raccomanda di iniziare immediatamente il trattamento farmacologico nei soggetti con un rischio elevato o molto elevato di malattie cardiovascolari, malattie renali o danni agli organi mediati dall’ipertensione. Raccomanda inoltre un trattamento farmacologico immediato in tutti i pazienti che hanno una pressione arteriosa pari o superiore a 160/100 mmHg. In questi casi, vi invito a non usare steroidi anabolizzanti e a rivolgervi al vostro medico per un trattamento. Sconsiglio vivamente l’uso di steroidi anabolizzanti se questo è il vostro caso.
Detto questo, ecco alcuni cambiamenti nello stile di vita. Uno ovvio è quello di smettere di fumare, se lo fate. Non tanto per abbassare la pressione sanguigna, ma semplicemente perché il fumo aumenta enormemente il rischio di malattie cardiovascolari. Probabilmente la lettura di questo articolo non vi farà smettere di fumare (se solo fosse così facile, no?), ma volevo solo informarvi.
Una strategia efficace consiste nel ridurre il sodio alimentare, cioè il sale. Diverse linee di evidenza hanno costantemente implicato l’assunzione di sale nella dieta con il rischio cardiovascolare [19]. Una meta-analisi Cochrane del 2013 di 34 studi randomizzati e controllati ha dimostrato una riduzione della pressione arteriosa di 4,2/2,1 mmHg per ogni riduzione di 4,4 g/die di assunzione di sale (=1,8 g di sodio) [20]. Di conseguenza, la Società Europea dell’Ipertensione raccomanda di limitare l’assunzione di sale a 5 g al giorno (= 2 g di sodio) [16]. Tuttavia, l’assunzione di sale con la dieta mostra una curva a U per quanto riguarda il rischio di eventi cardiovascolari e di morte. Ciò significa che, mentre la riduzione dell’assunzione di sale diminuisce questo rischio, esso ricomincia ad aumentare al di sotto di una certa dose giornaliera. Una recente meta-analisi ha rilevato che, rispetto a un’assunzione di 7 o più g di sodio al giorno, 4-5 g al giorno comportano un rischio inferiore di eventi cardiovascolari e morte [21]. Allo stesso modo, anche un’assunzione di meno di 3 g al giorno mostrava un rischio maggiore rispetto a un’assunzione di 4-5 g di sodio al giorno. (Gli studi hanno esaminato l’escrezione urinaria di sodio come proxy dell’assunzione di sodio. Questa è eccellente come proxy, quindi in questo articolo faccio finta che siano la stessa cosa). Non è del tutto chiaro quale sia la causa, poiché 3 g di sodio al giorno sono sufficienti a coprire il fabbisogno giornaliero. La Società Europea dell’Ipertensione si aggrappa all’effetto di abbassamento della pressione arteriosa come decisivo per ridurla ulteriormente. Sentitevi liberi di farlo, ma credo che sia più pragmatico attenersi ai 4-5 g al giorno, a meno che non abbiate già un apporto inferiore, e lasciarlo così. E poi realizzare un’ulteriore riduzione della pressione arteriosa, se necessario, con ulteriori interventi sullo stile di vita o con farmaci. Infine, una cosa che non potrò mai sottolineare abbastanza: controllate il contenuto di sale di tutto ciò che mangiate. Potreste rimanere sorpresi dal contenuto di sale di alcuni prodotti che consumate.
Se bevete molto alcol, è ovviamente consigliabile moderare il consumo di alcol (o astenersi dal farlo). L’effetto di abbassamento della pressione sanguigna è molto modesto (riduzione di ~1,2/0,7 mmHg [22]), ma l’alcol fa male alla salute (cardiovascolare) a prescindere. La raccomandazione è di limitare il consumo a 14 unità a settimana per gli uomini (8 a settimana per le donne; 1 unità equivale a 125 ml di vino o 250 ml di birra) [16]. A parte questo, aggiungete un po’ di esercizio aerobico alla vostra routine, se non l’avete già fatto, e assicuratevi di non ingrassare. Anche questo aiuta.
Trattamento dell’ipertensione: i farmaci
Lercanidipina; un calcio-antagonista appartenente al sottogruppo dei diidropiridinici
Per il trattamento dell’ipertensione sono disponibili diversi farmaci. Esistono cinque classi principali di farmaci raccomandati per questo scopo: ACE-inibitori, bloccanti del recettore dell’angiotensina (ARB), beta-bloccanti, calcio-antagonisti (CCB) e diuretici tiazidici. Esistono alcune piccole differenze per quanto riguarda gli esiti specifici per causa tra questi farmaci. Tuttavia, gli esiti cardiovascolari maggiori e la mortalità sono complessivamente simili e pertanto tutti sono raccomandati dalla Società Europea dell’Ipertensione come trattamento di prima linea [16]. Anche la Società Internazionale dell’Ipertensione raccomanda questi farmaci come trattamento di prima linea, ad eccezione dei beta-bloccanti [15]. Ogni classe di farmaci ha le proprie controindicazioni. Ad esempio, gli atleti e i pazienti fisicamente attivi sono indicati come possibile controindicazione all’uso dei beta-bloccanti, mentre una frequenza cardiaca inferiore a 60 bpm è indicata come controindicazione assoluta sia per i beta-bloccanti che per alcuni calcio-antagonisti (le non diidropiridine) [16]. I beta-bloccanti vengono generalmente aggiunti al trattamento quando esiste un’indicazione specifica per il loro utilizzo. Inoltre, i diuretici tiazidici sono preferiti ai tiazidici. In definitiva, la terapia si riduce ai calcio-antagonisti diidropiridinici, agli ACE-inibitori, agli ARB e ai diuretici tiazidici. Secondo la mia esperienza, i consumatori di AAS hanno un accesso relativamente più facile a questi ultimi tre, ma non ai calcio-antagonisti. (Naturalmente, a meno che non vengano prescritti dal medico, ma in quel caso si fa quello che prescrive il medico). Pertanto, mi concentrerò su queste tre modalità di trattamento.
Captopril; prima molecola sintetizzata della famiglia degli ACE II-inibitori
Sia gli ACE-inibitori che gli ARB si agganciano al cosiddetto sistema renina-angiotensina-aldosterone (RAAS). Si tratta di un sistema ormonale che svolge un ruolo estremamente importante nella regolazione del volume del sangue, degli elettroliti e della resistenza vascolare sistemica. Come tale, costituisce un bersaglio molto interessante per il trattamento dell’ipertensione. Questo sistema ormonale funziona come segue. I reni rilasciano un enzima chiamato renina ogni volta che rilevano un calo della pressione sanguigna. Questo enzima, a sua volta, converte una proteina prodotta dalla leva, l’angiotensinogeno, in angiotensina I. Si tratta di un piccolo peptide che costituisce il substrato per l’enzima di conversione dell’angiotensina (ACE), che taglia altri due aminoacidi da questo peptide producendo angiotensina II. L’angiotensina II è responsabile della vasocostrizione, soprattutto nelle arteriole. Di conseguenza, aumenta la pressione arteriosa, chiudendo così il circuito avviato dal rilevamento di una diminuzione della pressione arteriosa da parte dei reni. Inoltre, l’angiotensina II inibisce il processo di escrezione di acqua (diuresi) e sodio (natriuresi) da parte dei reni. Questo effetto si ottiene in parte evocando il rilascio di aldosterone da parte dei surreni. L’aldosterone è un ligando per i recettori dei mineralocorticoidi (MR) situati nei reni. L’attivazione del MR provoca la ritenzione di acqua e sodio e la secrezione di potassio. Il RAAS è schematizzato di seguito:
Ora che sapete come funziona il RAAS, sapete anche come funzionano gli ACE-inibitori e gli ARB. Gli ACE inibitori inibiscono l’enzima ACE (naturalmente, è scritto nel nome). In questo modo inibiscono la formazione di angiotensina II a partire dall’angiotensina I. Allo stesso modo, gli ARB – bloccanti del recettore dell’angiotensina – assicurano che l’angiotensina II non possa svolgere la sua azione bloccando il recettore a cui l’angiotensina II dovrebbe legarsi.
Telmisartan; appartenente alla famiglia degli ARB
Sia gli ACE-inibitori che gli ARB hanno effetti simili sulla riduzione della pressione sanguigna. Le meta-analisi Cochrane hanno rilevato una riduzione della pressione arteriosa sistolica di 8 mmHg e della pressione arteriosa diastolica di 5 mmHg nel trattamento dell’ipertensione primaria in entrambi [23, 24]. La metà della dose massima giornaliera raccomandata dal produttore ha ottenuto un effetto di abbassamento della pressione sanguigna pari al 90% della dose massima nel caso degli ACE-inibitori e all’80% della dose massima nel caso degli ARB. Di conseguenza, l’aumento del dosaggio di questi farmaci di solito porta solo a riduzioni molto modeste della pressione arteriosa. Diventa quindi più interessante associarli a un diuretico tiazidico. Tuttavia, vorrei sottolineare una cosa: se avete bisogno di più farmaci per abbassare sufficientemente la pressione sanguigna, vi esorto a farlo sotto la supervisione di un medico.
Sebbene entrambi i farmaci siano abbastanza sicuri e ben tollerati in generale, come ogni farmaco possono avere effetti collaterali. Questi includono ipotensione/vertigini di prima dose, insufficienza renale acuta, iperkaliemia, tosse, eruzioni cutanee, disturbi del gusto (disgeusia), epatotossicità e angioedema per gli ACE-inibitori [25]. L’ipotensione da prima dose si riferisce all’improvviso calo della pressione sanguigna che può verificarsi nelle prime fasi del trattamento. Tenetene conto in caso di situazioni come la guida, ecc. Questo effetto è esacerbato quando si è disidratati (per la competizione o quando si utilizza un diuretico per qualsiasi motivo). Non utilizzare il farmaco in questi casi. L’insufficienza renale acuta si verifica in alcuni pazienti, ma di solito non comporta alcun segno clinico. Non è permanente: una volta smesso, la funzione renale torna normale. Anche in questo caso la disidratazione è un ulteriore fattore di rischio. A questo scopo, è necessario misurare la creatinina sierica nel tempo (o altri marcatori, forse più affidabili, utilizzati per stimare la velocità di filtrazione glomerulare [GFR]).
L’iperkaliemia (eccesso di potassio nel sangue) è abbastanza rara da sviluppare se questo è l’unico farmaco in uso, ma la combinazione con altri farmaci risparmiatori di potassio, un apporto molto elevato di potassio dalla dieta o un’insufficienza renale esistente possono aumentare il rischio. Per questo motivo, oltre alla misurazione della funzionalità renale, è necessario misurare anche gli elettroliti sierici. Ridurre il dosaggio (se possibile) se si è iperkaliemici, o passare a diuretici tiazidici.
Idroclorotiazide; farmaco appartenente alla famiglia dei diuretici triazidici
L’effetto collaterale più caratteristico degli ACE-inibitori è forse la tosse secca e irritante. Si verifica all’incirca in 1 persona su 10 [26], e sembra essere nettamente maggiore negli asiatici [27]. Occasionalmente, alcune persone sviluppano anche un’eruzione cutanea a causa degli ACE-inibitori [28]. Anche in questo caso, è necessario ridurre il dosaggio e talvolta il passaggio da un ACE-inibitore a un altro risolve il problema (in particolare il passaggio dal captopril). In alcuni casi molto rari, sembrano verificarsi colestasi, epatite colestatica o lesioni epatocellulari [29]. Ma in letteratura esistono solo alcuni limitati casi di questo tipo.
Un ultimo effetto collaterale che vorrei sottolineare è l’angioedema: l’accumulo di liquido sotto la pelle o le membrane mucose. Questo può includere il viso, la mucosa orale, la lingua, le labbra e anche la faringe e la laringe [30]. A seconda del luogo in cui si verifica, può causare una situazione di pericolo di vita bloccando le vie respiratorie. L’incidenza di questo fenomeno, tuttavia, è piuttosto bassa. Una meta-analisi ha rilevato un’incidenza dello 0,3% rispetto allo 0,07% del placebo [31]. Questo effetto collaterale non deve necessariamente verificarsi nelle fasi iniziali di utilizzo, ma può talvolta manifestarsi anche dopo anni di utilizzo. La raccomandazione è di interrompere completamente l’uso di qualsiasi ACE-inibitore quando si verifica l’angioedema e di non utilizzarlo mai più.
L’aspetto positivo degli ARB è che non causano tosse come gli ACE-inibitori [32] e non sembrano aumentare il rischio di angioedema [31]. Tuttavia, rimane l’aumento del rischio di ipotensione, iperkaliemia e disfunzione renale [32]. In ogni caso, il suo tasso di aderenza è superiore a quello di qualsiasi altra classe di farmaci antipertensivi [33]. Alla luce di ciò e del fatto che una recente meta-analisi non ha riscontrato differenze tra ACE-inibitori e ARB in termini di riduzione della pressione arteriosa, eventi fatali per qualsiasi causa e cause cardiovascolari, infarti miocardici fatali e non fatali e ictus [34], gli ARB sembrano la scelta più probabile tra i due, se disponibili.
Benazepril
Gli ACE-inibitori comunemente prescritti sono Benazepril, Captopril, Enalapril, Fosinopril, Lisinopril, Perindopril, Quinapril, Ramipril e Zofenopril. Gli ARB comunemente prescritti sono Candesartan, Eprosartan, Irbesartan, Losartan, Olmesartan, Telmisartan e Valsartan. Le revisioni Cochrane che ho citato in precedenza non hanno rilevato alcun ACE-inibitore con prestazioni migliori o peggiori rispetto agli altri, e lo stesso vale per gli ARB. Potrebbero esserci delle eccezioni in alcune popolazioni (ad esempio i diabetici), ma in generale si tratta di quello che si riesce a reperire. Gli ACE-inibitori generalmente più prescritti sono il Captopril, l’Enalapril e il Lisinopril, mentre gli ARB più prescritti sono il Valsartan, il Candesartan, il Telmisartan e il Losartan. In generale, la metà della dose massima raccomandata dal produttore è una buona dose iniziale. Eseguire un esame del sangue prima e un mese dopo l’inizio (o dopo un aumento della dose) e, se tutto risulta normale, ogni sei mesi. Includere creatinina, eGFR ed elettroliti. (Esistono molte linee guida sul monitoraggio, ma non c’è molto consenso al riguardo [35]). Tenete presente che sono necessarie circa 4-6 settimane per ottenere l’effetto completo del trattamento. L’ideale è raggiungere una pressione arteriosa inferiore o uguale a 130/80 mmHg (ma superiore a 120 mmHg).
I diuretici tiazidici inibiscono l’azione dei simpatizzanti sodio-cloruro nel lume del tubulo distale dei nefroni. I nefroni sono gli elementi costitutivi dei reni, l’unità di base del funzionamento. Ognuno di essi (e i reni ne contengono diverse centinaia di migliaia) contribuisce in minima parte alla funzione di filtraggio cumulativa dei reni. Guardate l’immagine qui sotto per avere un’idea di come si presenta.
Il sangue viene filtrato attraverso un gruppo di capillari “specializzati” chiamati glomeruli e il filtrato viene poi catturato in un sacco simile a una tazza che lo circonda, chiamato capsula di Bowman. Il filtrato entra quindi nel tubulo renale per essere trasformato in urina. Durante questo percorso, varie sostanze vengono riassorbite dal filtrato nel sangue e secrete dal sangue nel filtrato.
I simpaticatori su cui agiscono i diuretici tiazidici trasportano il sodio e il cloruro fuori dal filtrato. Pertanto, bloccando questo simpatizzante, nel filtrato rimane più cloruro di sodio e quindi viene espulsa più acqua. In altre parole, i diuretici tiazidici devono la loro azione diuretica (escrezione di acqua) all’azione natriuretica (escrezione di sodio).
Come nel caso di qualsiasi diuretico che porti a un aumento delle concentrazioni di sodio nella parte distale del tubulo distale, si verifica una perdita di potassio. Questo accade perché il sodio viene assorbito in misura maggiore e finisce per essere scambiato con il potassio (che viene quindi secreto nel filtrato). Pertanto, contrariamente a quanto avviene con gli ARB e gli ACE-inibitori, può verificarsi un’ipopotassiemia (un livello troppo basso di potassio nel sangue). Se si sviluppa un’ipopotassiemia, di solito è lieve, ma evidenzia la necessità di analizzare gli elettroliti. Un’ipokaliemia lieve (3,0-3,5 mmol/L) raramente provoca sintomi. Quindi non la noterete. Tuttavia, se diventa più grave (<2,5-3,0 mmol/L), si possono sviluppare sintomi come debolezza generalizzata, affaticamento e costipazione [36]. Livelli di potassio molto bassi possono anche evocare aritmie cardiache. Nel contesto dei bodybuilder che fanno uso di PED, diversi altri composti possono contribuire allo sviluppo dell’ipokaliemia. Tra questi vi sono i beta-agonisti, giàcitati in precedenza, come il clenbuterolo, ma anche dosi elevate di insulina. Ciò può causare uno spostamento transitorio di potassio dal compartimento extracellulare a quello intracellulare. Sebbene tale spostamento duri solo un paio d’ore, l’effetto può essere molto drastico nel contesto di un’ipopotassiemia esistente. Inoltre, anche l’assunzione di caffeina può contribuire profondamente. Per questo motivo, è bene controllare anche gli integratori pre-allenamento che si utilizzano, poiché si può osservare una diminuzione di 0,26 mmol/L 4 ore dopo una dose di 180 mg di caffeina e un aumento ancora maggiore di 0,44 mmol/L dopo 360 mg [37]. In ogni caso, se i risultati del sangue mostrano una lieve ipokaliemia, è possibile aumentare l’apporto di potassio con la dieta.
Come con qualsiasi natriuretico, si può sviluppare iponatriemia (bassi livelli di sodio). Il rischio è basso, ma può essere, ovviamente, molto pericoloso [38, 39]. Bisogna fare attenzione a nausea, mal di testa, crampi muscolari, affaticamento, disturbi dell’andatura, vomito, sensazione di confusione e difficoltà di pensiero. Anche bere molta acqua può contribuire a sviluppare l’iponatriemia. Non bevete quindi di proposito litri e litri di acqua. In alcuni casi possono verificarsi anche ipomagnesiemia e ipercalcemia.
Misurare creatinina, eGFR ed elettroliti prima e una settimana dopo l’inizio del trattamento. Se tutto risulta normale, effettuare una seconda misurazione entro 4-8 settimane, dopodiché è possibile ripeterla ogni 6-12 mesi [40].
Clortalidone; appartenente alla famiglia dei diuretici Sulfonamidi
Utilizzare preferibilmente Clortalidone o Indapamide e iniziare con un dosaggio basso, ad esempio 12,5 mg di Clortalidone al giorno o 1,25 mg di Indapamide al giorno (anche se di solito sono disponibili in formato da 2,5 mg senza interruzione). Dopo che la seconda misurazione del sangue è risultata normale e la pressione arteriosa non è inferiore a 130/80 mmHg, si può prendere in considerazione un ulteriore aumento della dose (si tratta di raddoppiare la dose a 25 mg e 2,5 mg per Clortalidone e Indapamide, rispettivamente). Ripetere gli esami del sangue una settimana dopo l’aumento della dose.
Conclusioni:
La pressione alta è un killer silenzioso. La maggior parte delle persone non la sente, ma aumenta drasticamente la mortalità dovuta a malattie coronariche o ictus. Ogni aumento di 20 mmHg della pressione arteriosa sistolica oltre i 115 mmHg e ogni aumento di 10 mmHg della pressione arteriosa diastolica oltre i 75 mmHg è associato a un raddoppio della mortalità per queste cause. Inoltre, l’ipertensione per un periodo di tempo prolungato provoca danni agli organi bersaglio. Questo non riguarda solo il cuore e la vascolarizzazione, ma anche altri organi come il cervello, gli occhi e i reni. I dati della letteratura indicano che l’uso di steroidi anabolizzanti ad alti dosaggi può potenzialmente aumentare leggermente la pressione sanguigna. Una cifra indicativa è di circa 5-10 mmHg per la pressione arteriosa sistolica e la metà per quella diastolica. Naturalmente, le persone possono discostarsi da questi intervalli. Ciò può dipendere o meno dal dosaggio, dai tipi di AAS utilizzati e dall’uso concomitante di farmaci ausiliari che possono influire. Fortunatamente, questi aumenti sono transitori e reversibili dopo la cessazione dell’uso.
Ora sapete come misurare correttamente la pressione arteriosa, quando è necessario trattarla e come trattarla. Il trattamento va effettuato quando la pressione è superiore a 135/85mmHg, misurata a casa, e va effettuato con ARB, ACE-inibitori o diuretici tiazidici sotto stretto controllo medico. In generale, opterei per gli ARB, che di solito sono meglio tollerati. Come nota finale di questo articolo, vi invito ancora una volta a chiedere al vostro medico di curare la vostra pressione arteriosa piuttosto che farlo da soli.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Prospective Studies Collaboration. “Age-specific relevance of usual blood pressure to vascular mortality: a meta-analysis of individual data for one million adults in 61 prospective studies.” The Lancet 360.9349 (2002): 1903-1913.
Cohuet, G., and H. Struijker-Boudier. “Mechanisms of target organ damage caused by hypertension: therapeutic potential.” Pharmacology & therapeutics 111.1 (2006): 81-98.
Giorgi, Anthony, Robert P. Weatherby, and Peter W. Murphy. “Muscular strength, body composition and health responses to the use of testosterone enanthate: a double blind study.” Journal of science and medicine in sport 2.4 (1999): 341-355.
Zmuda, Joseph M., et al. “The effect of testosterone aromatization on high-density lipoprotein cholesterol level and postheparin lipolytic activity.” Metabolism 42.4 (1993): 446-450.
Freed, D. L., et al. “Anabolic steroids in athletics: crossover double-blind trial on weightlifters.” Br Med J 2.5969 (1975): 471-473.
Glazer, Gary, and Anthony L. Suchman. “Lack of demonstrated effect of nandrolone on serum lipids.” Metabolism 43.2 (1994): 204-210.
Hartgens, F., E. C. Cheriex, and H. Kuipers. “Prospective echocardiographic assessment of androgenic-anabolic steroids effects on cardiac structure and function in strength athletes.” International journal of sports medicine 24.05 (2003): 344-351.
Kuipers, H., et al. “Influence of anabolic steroids on body composition, blood pressure, lipid profile and liver functions in body builders.” International journal of sports medicine 12.04 (1991): 413-418.
Smit, Diederik L., et al. “Baseline characteristics of the HAARLEM study: 100 male amateur athletes using anabolic androgenic steroids.” Scandinavian journal of medicine & science in sports 30.3 (2020): 531-539.
Hartgens, F., E. C. Cheriex, and H. Kuipers. “Prospective echocardiographic assessment of androgenic-anabolic steroids effects on cardiac structure and function in strength athletes.” International journal of sports medicine 24.05 (2003): 344-351.
De Piccoli, B., et al. “Anabolic steroid use in body builders: an echocardiographic study of left ventricle morphology and function.” International journal of sports medicine 12.04 (1991): 408-412.
Palatini, Paolo, et al. “Cardiovascular effects of anabolic steroids in weight‐trained subjects.” The Journal of Clinical Pharmacology 36.12 (1996): 1132-1140.
Prospective Studies Collaboration. “Age-specific relevance of usual blood pressure to vascular mortality: a meta-analysis of individual data for one million adults in 61 prospective studies.” The Lancet 360.9349 (2002): 1903-1913.
Fonseca-Reyes, Salvador, et al. “Differences and effects of medium and large adult cuffs on blood pressure readings in individuals with muscular arms.” Blood pressure monitoring 14.4 (2009): 166-171.
Fonseca-Reyes, Salvador, et al. “Effect of standard cuff on blood pressure readings in patients with obese arms. How frequent are arms of a ‘large circumference’?.” Blood pressure monitoring 8.3 (2003): 101-106.
Unger, Thomas, et al. “2020 International Society of Hypertension global hypertension practice guidelines.” Hypertension 75.6 (2020): 1334-1357.
Parati, Gianfranco, et al. “European Society of Hypertension practice guidelines for home blood pressure monitoring.” Journal of human hypertension 24.12 (2010): 779-785.
Brunström, Mattias, and Bo Carlberg. “Association of blood pressure lowering with mortality and cardiovascular disease across blood pressure levels: a systematic review and meta-analysis.” JAMA internal medicine 178.1 (2018): 28-36.
Ettehad, Dena, et al. “Blood pressure lowering for prevention of cardiovascular disease and death: a systematic review and meta-analysis.” The Lancet 387.10022 (2016): 957-967.
He, Feng J., and Graham A. MacGregor. “Salt, blood pressure and cardiovascular disease.” Current opinion in cardiology 22.4 (2007): 298-305.
He, Feng J., Jiafu Li, and Graham A. MacGregor. “Effect of longer‐term modest salt reduction on blood pressure.” Cochrane database of systematic reviews 4 (2013).
Mente, Andrew, et al. “Associations of urinary sodium excretion with cardiovascular events in individuals with and without hypertension: a pooled analysis of data from four studies.” The Lancet 388.10043 (2016): 465-475.
Cushman, William C., et al. “Prevention and Treatment of Hypertension Study (PATHS): effects of an alcohol treatment program on blood pressure.” Archives of internal medicine 158.11 (1998): 1197-1207.
Heran, Balraj S., et al. “Blood pressure lowering efficacy of angiotensin converting enzyme (ACE) inhibitors for primary hypertension.” Cochrane Database of Systematic Reviews 4 (2008).
Heran, Balraj S., et al. “Blood pressure lowering efficacy of angiotensin receptor blockers for primary hypertension.” Cochrane Database of Systematic Reviews 4 (2008).
Alderman, Christopher P. “Adverse effects of the angiotensin-converting enzyme inhibitors.” Annals of Pharmacotherapy 30.1 (1996): 55-61.
Bangalore, Sripal, Sunil Kumar, and Franz H. Messerli. “Angiotensin-converting enzyme inhibitor associated cough: deceptive information from the Physicians’ Desk Reference.” The American journal of medicine 123.11 (2010): 1016-1030.
Tseng, Daniel S., et al. “Angiotensin-converting enzyme-related cough among Chinese-Americans.” The American journal of medicine 123.2 (2010): 183-e11.
Weber, Michael A. “Safety issues during antihypertensive treatment with angiotensin converting enzyme inhibitors.” The American journal of medicine 84 (1988): 16-23.
Campo, Paloma, et al. “Angioedema induced by angiotensin-converting enzyme inhibitors.” Current opinion in allergy and clinical immunology 13.4 (2013): 337-344.
Makani, Harikrishna, et al. “Meta-analysis of randomized trials of angioedema as an adverse event of renin–angiotensin system inhibitors.” The American journal of cardiology 110.3 (2012): 383-391.
Caldeira, Daniel, Cláudio David, and Cristina Sampaio. “Tolerability of angiotensin-receptor blockers in patients with intolerance to angiotensin-converting enzyme inhibitors.” American Journal of Cardiovascular Drugs 12.4 (2012): 263-277.
Kronish, Ian M., et al. “Meta-analysis: impact of drug class on adherence to antihypertensives.” Circulation 123.15 (2011): 1611-1621.
Dimou, Chrisa, et al. “A systematic review and network meta-analysis of the comparative efficacy of angiotensin-converting enzyme inhibitors and angiotensin receptor blockers in hypertension.” Journal of human hypertension 33.3 (2019): 188-201.
McDowell, Sarah E., et al. “A practical guide to monitoring for adverse drug reactions during antihypertensive drug therapy.” Journal of the Royal Society of Medicine 106.3 (2013): 87-119.
Gennari, F. John. “Hypokalemia.” New England Journal of Medicine 339.7 (1998): 451-458.
Passmore, A. P., G. B. Kondowe, and G. D. Johnston. “Caffeine and hypokalemia.” Annals of internal medicine 105.3 (1986): 468-468.
Rodenburg, Eline M., et al. “Thiazide-associated hyponatremia: a population-based study.” American journal of kidney diseases 62.1 (2013): 67-72.
Spital, Aaron. “Diuretic-induced hyponatremia.” American journal of nephrology 19.4 (1999): 447-452.
Gli effetti regolatori degli androgeni sull’ematopoiesi sono stati riconosciuti fin dall’inizio del XX secolo. La castrazione dei ratti maschi causa anemia (1) che è reversibile dopo il trattamento con androgeni (2). Un classico studio clinico di Vahlquist ha fornito prove indirette che gli uomini hanno livelli di ematocrito (Hct) intrinsecamente più elevati rispetto alle donne; è stato osservato che le donne in pre-menopausa non hanno livelli di Hct più elevati rispetto alle donne in post-menopausa, mentre hanno un aumento dell’Hct in risposta all’integrazione di ferro (3). Dati aneddotici provenienti da atleti agonisti suggeriscono che l’abuso di androgeni può migliorare le prestazioni, in parte attraverso l’aumento del VO2max (capacità di trasporto di ossigeno nel sangue mediata dall’Hb), anche se a spese di un aumento del rischio di trombosi arteriosa e venosa (4). Allo stesso modo, le donne affette da endocrinopatie iperandrogeniche, come l’iperplasia surrenale congenita e la sindrome di Cushing, possono presentare un’eritrocitosi relativa (5, 6). I risultati storici di cui sopra confermano che gli androgeni stimolano l’eritropoiesi della midollare.
È noto che i parametri ematologici e l’eritropoiesi sono influenzati dall’uso di steroidi androgeni anabolizzanti (AAS), anche se poco si sa in relazione a dosi sovra-fisiologiche di questa classe di farmaci sui dettagli meccanicistici di tale processo.
Si è visto che la somministrazione di Testosterone stimola la produzione di EPO, cosa che è stata osservata sia negli uomini ipogonadici che nei volontari sani di sesso maschile, mentre non esiste alcuna correlazione tra le concentrazioni endogene di Testosterone e di EPO durante la pubertà maschile.[7] Il meccanismo alla base dell’aumento dell’EPO indotto dagli androgeni non è stato compreso e, per quanto ne so, i livelli di EPO non sono stati studiati in uomini sani che utilizzano dosi elevate di AAS, in base allo stato ipogonadico. Molti consumatori di AAS presentano ipogonadismo ipogonadotropo (HH), definito ipogonadismo indotto da AAS (ASIH).[8]
Uno studio recente indica che la frazione reticolocitaria ad alta fluorescenza (HFR) è sensibile all’assunzione di Testosterone nelle donne,19 ma l’associazione dell’HFR all’esposizione sovra-fisiologica agli AAS negli uomini non è stata studiata. È possibile che dosi sovra-fisiologiche di AAS esercitino effetti diversi sulle frazioni reticolocitarie.
Sappiamo però che vi è una risposta nell’aumento dei processi eritrocitari in seguito all’assunzione di AAS off-label. Ciò è risultato di grado soggettivo con alcuni utilizzatori che mostrano una risposta minore mentre altri ne mostrano una sensibilmente elevata anche con dosaggi tipici di una TRT [100mg/week di Testosterone Enantato = 72mg di Testosterone effettivo/slegato dall’estere]. La variabile di stimolo eritrocitario sembra variare anche in modo dipendente dal/dagli AAS utilizzato/i con una maggiore risposta a carico del Boldenone e dell’Oxymetholone, ma non solo.
Alcune testimonianze riportate da preparatori e provenienti dai controlli ematici di diversi bodybuilder, suggeriscono che il Methandrostenolone (Dianabol), nonostante sia molto semplicemente una forma metilata in C-17 del Boldenone, al contrario di quest’ultimo eserciti un azione negativa, o sottoregolativa, dell’ematopoiesi e, nel dettaglio, dell’ematopoiesi.
In questo articolo analizzerò quanto attualmente si conosce dei processi di interazione degli androgeni (e AAS esogeni) sulla ematopoiesi e eritropoiesi e quali potrebbero essere le ragioni per le quali è stata osservata la risposta “paradossale” del Methandrostenolone su tali processi (“Dbol Hematocrit Theory”).
Androgeni ed eritropoiesi:
Studi su animali e sull’uomo hanno suggerito un effetto stimolatorio diretto e indiretto degli androgeni sull’eritropoiesi, anche se l’esatto meccanismo di tale relazione rimane vagamente compreso. La somministrazione di androgeni determina un aumento della massa cellulare eritroide, delle unità formanti colonie di eritrociti (CFU-E) e della produzione e secrezione di eritropoietina (EPO) (6), mentre la privazione di androgeni causa una riduzione degli indici di globuli rossi a causa della ridotta proliferazione dei precursori eritroidi del midollo (9).
Gli androgeni vengono convertiti in 17-cheto-steroidi in grado di aumentare la sintesi di mRNA nel nucleo, causando la differenziazione delle cellule del midollo osseo da non responsive all’EPO a responsive all’EPO (6). Inoltre, gli androgeni aumentano l’assorbimento di glucosio con conseguente glicolisi e trascrizione genica e sintesi di mRNA negli eritroidi (10-11).
Il Testosterone può aumentare l’Hct inibendo la secrezione di Epcidina, il principale peptide regolatore del ferro, con conseguente aumento del ferro biodisponibile (12), ma può anche aumentare l’incorporazione del ferro nei globuli rossi (13) e migliorare la sopravvivenza di questi ultimi (14). Infine, il riscontro di un aumento dei livelli di IGF-1 nei soggetti che ricevono androgeni ha suggerito un potenziale legame tra gli androgeni e la proliferazione e la differenziazione delle cellule progenitrici eritroidi guidata da IGF-1 (15, 16).
L’effetto del Testosterone sull’eritropoiesi è più pronunciato durante la pubertà, con l’Hb prepuberale che è simile nei ragazzi e nelle ragazze, ma aumenta nei ragazzi dopo i 13 anni in tandem con l’aumento delle concentrazioni di Testosterone(6, 17). I ragazzi con pubertà ritardata hanno livelli di Hb simili a quelli dei ragazzi e delle ragazze prepuberi e il trattamento con Testosterone normalizza i livelli di emoglobina a quelli osservati nella pubertà maschile tardiva (18, 19).
Giova ricordare che prima dello sviluppo della terapia con EPO alla fine degli anni ’80, gli androgeni erano l’unica opzione per il trattamento dell’anemia legata alla Malattia Renale Cronica (CKD) negli uomini. I pazienti con CKD possono presentare una riduzione della densità minerale ossea, della massa muscolare, dei livelli di energia, della qualità della vita e della funzione sessuale, con esiti cardiovascolari avversi, soprattutto in quelli con diabete concomitante (20); sebbene queste caratteristiche abbiano probabilmente un’origine multifattoriale, si verificano anche in caso di ipogonadismo. Il basso livello di Testosterone è prevalente nei pazienti con CKD e può contribuire all’anemia renale (21, 22). Fino a due terzi degli uomini in emodialisi (HD) presentano livelli sierici di Testosterone nell’intervallo ipogonadico, a causa di anomalie a tutti i livelli dell’asse Ipotalamo-Ipofisi-Testicolo (23-24). Dati non pubblicati di una coorte di 113 pazienti in HD e 85 in pre-dialisi (preD) allo stadio 4 e 5 della CKD hanno riportato livelli di Testosterone subnormali nel 76% dei maschi in pre-dialisi e nell’80% dei maschi in HD, con una significativa correlazione inversa della dose di Dα con il T totale (R -0,253; p <0,01) e il T libero (-0,29; <0,01) (25).
Il fatto che l’ipogonadismo sia una causa consolidata di anemia e di ridotta responsività all’EPO negli uomini con CKD può suggerire un possibile ruolo della terapia con Testosterone come aggiunta o alternativa all’EPO in alcuni uomini con anemia correlata alla CKD (26). Ciò è particolarmente rilevante in alcuni sistemi sanitari in cui i pazienti con CKD non possono permettersi la terapia con EPO, che potrebbe causare anemia con necessità di trasfusioni di sangue, soprattutto se manca l’evidenza che l’EPO migliori la morbilità o la mortalità nella CKD (27).
L’anemia negli uomini anziani può aumentare il rischio di morbilità e mortalità. In uno studio retrospettivo su uomini di età superiore ai 65 anni ricoverati con infarto miocardico acuto, livelli di Hct più bassi erano associati a un aumento della mortalità a 30 giorni, mentre il trattamento dell’anemia migliorava i tassi di mortalità (28). Un’osservazione simile di elevata mortalità è stata riportata in una coorte di pazienti anemici che presentavano un’insufficienza cardiaca di nuova insorgenza (29). L’ipogonadismo, o il calo ponderale dei livelli di androgeni legati all’invecchiamento possono entrambi causare anemia, sarcopenia e osteoporosi (30), con studi longitudinali e trasversali che mostrano costantemente un calo dei livelli sierici di Testosterone negli uomini che invecchiano.
Alcuni studi hanno esaminato i cambiamenti dell’Hb nei pazienti sottoposti a terapia sostitutiva con Testosterone; alcuni hanno sperimentato specificamente la terapia con Testosterone con l’obiettivo primario di migliorare l’anemia, con un accumulo di evidenze. In uno studio randomizzato in doppio cieco controllato con placebo di Dhinsda et al., la terapia con Testosterone ha soppresso l’Epcidina con un marcato aumento dell’Hb, dell’EPO e dell’espressione dei recettori della ferroportina e della transferrina in pazienti ipogonadici con diabete di tipo 2 (31). Gli studi con Testosterone finanziati dal National Institute of Health (NIH) hanno esaminato gli effetti della TRT in uomini ipogonadici di età superiore ai 65 anni (32). Utilizzando un design in doppio cieco, controllato con placebo, 12 mesi di trattamento quotidiano con gel di Testosterone hanno aumentato i livelli di Hb di almeno 1,0g/dL in ~52% degli uomini con ipogonadismo e una causa nota di anemia rispetto al placebo. Inoltre, nei 64 uomini anziani con anemia inspiegabile, l’Hb è migliorata di almeno 1,0g/dL nel 54% degli uomini rispetto al 15% del gruppo placebo. Tuttavia, la carenza di androgeni (AD) è tipicamente trascurata nelle linee guida sull’indagine dell’anemia (33).
È importante notare che la maggior parte delle linee guida cliniche raccomanda di valutare l’Hct e l’antigene prostatico specifico (PSA) prima di iniziare la terapia sostitutiva con Testosterone (34). Dopo tutto l’influenza degli androgeni sulla emopoiesi/eritropoiesi era già piuttosto chiara fin dagli studi della fine degli anni 40′. In definitiva, in ambito terapeutico, il trattamento con androgeni ha anche un potenziale per trattare l’anemia della CKD e negli uomini ipogonadici come aggiunta all’EPO.
Ma per gli utilizzatori off-label lo stimolo eritropoietico dato dagli AAS può diventare un problema non di poca rilevanza.
Dosi sovrafisiologiche di AAS e risposta emopoietica/eritrocitaria:
Come già discusso in un mio precedente articolo trattante l’uso di AAS off-label ed eritrocitosi/policitemia, ho fatto notare che la somministrazione di Testosterone per 20 settimane mostra un aumento dose-dipendente (fino a 600mg di Testosterone Enantato alla settimana; pari a 432mg di Testosterone effettivo) dell’emoglobina e dell’ematocrito, soprattutto negli uomini più anziani, mentre l’EPO non lo fa [35]. E come già esposto, la somministrazione di Testosterone porta alla soppressione dell’Epcidina sierica in uomini giovani e anziani [36]. Il dosaggio del Testosterone (fino a 600mg di Testosterone Enantato a settimana) è altamente correlato con l’ampiezza di questa soppressione. In sintesi, gli androgeni aumentano l’ematocrito/emoglobina attraverso un aumento iniziale dei livelli di EPO e una contemporanea diminuzione dei livelli di Epcidina, che poi scendono gradualmente ai livelli di base di fronte all’aumento dell’ematocrito/emoglobina: un nuovo set point EPO/emoglobina. I meccanismi d’azione responsabili e il loro contributo relativo a questo fenomeno sono ancora da stabilire.
Con dosaggi fino a 600mg di Testosterone Enantato alla settimana, l’emoglobina ha mostrato un aumento di 1,42g/dL nei giovani uomini dopo 20 settimane [37]. Ciò si traduce in un aumento dell’ematocrito di poco superiore al 4%.
Fortunatamente, sembra esserci un limite alla misura in cui gli AAS possono aumentare l’ematocrito. Grazie allo studio HAARLEM, sono stati osservati 100 consumatori di steroidi anabolizzanti seguiti nel tempo mentre si autosomministravano AAS, il cui dosaggio medio, basato sulle informazioni riportate sull’etichetta, era di 898mg a settimana, rendendo così il loro ciclo di AAS abbastanza rappresentativo dell’uso comune da parte dei bodybuilder. Le misurazioni sono state effettuate prima, durante, 3 mesi dopo la fine del ciclo e 1 anno dopo l’inizio del ciclo. I ricercatori hanno riscontrato un aumento del 3% dell’ematocrito dei soggetti dello studio al termine del ciclo. Questo dato è in linea con l’aumento del 4% osservato nello studio nei giovani uomini. L’autore principale ha fatto sapere che l’aumento dell’ematocrito sembra stabilizzarsi a un dosaggio di androgeni di circa 500mg a settimana. Infine, c’è da aggiungere che i soggetti dello studio non hanno effettuato donazioni di sangue, quindi questo non è stato un fattore confondente.
Naturalmente, questi risultati presentano alcune variazioni. Alcuni rispondono agli AAS con un aumento dell’ematocrito maggiore di altri. Tuttavia, livelli molto elevati di ematocrito sembrano essere rari, come si può vedere nei grafici a scatola e baffi dei partecipanti allo studio HAARLEM (T0 = subito prima del ciclo di AAS, T1 = alla fine, T2 = 3 mesi dopo la cessazione dell’uso, T3 = 1 anno dopo l’inizio del ciclo):
Trattamento consuetudinario:
Forse il trattamento migliore consiste nel ridurre notevolmente il dosaggio (ben al di sotto dei 500mg settimanali) o nell’interrompere del tutto l’uso di AAS [tornando in fisiologia controllata]. In questo modo si abbasserà l’ematocrito, con un effetto completo dopo un paio di mesi, e si annullerà il rischio. Tuttavia, questo non è probabilmente il metodo più gradito per contrastare questo problema.
Pratica comunemente diffusa consiste nell’assunzione di un basso dosaggio di CardioAspirina (Acido Acetilsalicilico con gastroprotettore). Sebbene non influisca sui livelli di ematocrito, è ampiamente utilizzata per la prevenzione delle malattie cardiovascolari [38]. Più precisamente, è utilizzata nella prevenzione secondaria delle malattie cardiovascolari. Previene la coagulazione del sangue inibendo un enzima chiamato ciclossigenasi (COX) nei trombociti. Se da un lato riduce il rischio di trombosi, dall’altro aumenta il rischio di emorragie. Parliamo anche di emorragie interne, come l’ictus emorragico. I benefici devono quindi essere attentamente soppesati rispetto ai rischi del suo utilizzo. Attualmente, le linee guida europee sulla prevenzione delle malattie cardiovascolari nella pratica clinica ne sconsigliano l’uso nella prevenzione primaria (anche se questo potrebbe cambiare per alcune popolazioni, come i diabetici) [39]. Oltretutto, nelle settimane successive all’interruzione del farmaco vi è un aumento del rischio di eventi trombotici [40, 41]. Pertanto, anche l’assunzione e la sospensione frequente del farmaco sono sconsigliate.
Nota: prima di un eventuale uso, è caldamente consigliato, oltre il parere medico, un controllo accurato dei fattori che regolano la coagulazione: tempo di tromboplastinaparziale attivata (aPTT), tempo di protrombina (PT) Fibrinogeno, D-Dimero, Antitrombina e Inibitore C1 Esterasi.
Struttura cristallina della Nattochinasi.
Anche la Nattochinasi, un enzima digestivo (una proteasi alcalina) presente nel natto, un alimento tradizionale giapponese fermentato, viene comunemente utilizzata per la prevenzioni di eventi trombotici da parte di utilizzatori di AAS con alterazione del Ematocrito. Ma non ci sono prove sufficienti sull’uomo che utilizzano la Nattochinasi isolatamente e che valutano la formazione di trombi per raccomandarne l’uso come farmaco anti-clottico, anche se sembra esserci qualche promessa. Come per la CardioAspirina, la necessità di utilizzo andrebbe valutata per via esami dei fattori della coagulazione.
Un modo per ridurre efficacemente l’ematocrito è senza dubbio la flebotomia (salasso). Un modo per farlo è la donazione di sangue a una banca del sangue. Tuttavia, molti Paesi (e giustamente) limitano il numero di volte in cui è possibile farlo ogni anno. Vi sono paesi dove è limitato a cinque volte l’anno. Questo potrebbe non essere sufficiente a mantenere i valori nel range desiderato, dato che uno studio ha rilevato livelli di emoglobina persistentemente elevati in occasione di visite ripetute in un numero elevato di pazienti TRT che hanno donato il sangue [42]. Se la donazione di sangue non è sufficiente, si può sempre consultare un medico generico per eseguire una flebotomia terapeutica a intervalli più frequenti.
Occorre ricordare che ad ogni donazione di sangue/salasso si perde Ferro. Di conseguenza, si corre il rischio di esaurire le proprie riserve di Ferro e, consequenzialmente, anche l’emoglobina rimarrà molto bassa e si diventerà temporaneamente anemici. È possibile contrastare questo fenomeno integrando il Ferro, ma questo riduce drasticamente il tempo necessario all’organismo per recuperare i livelli di emoglobina/ematocrito [43]. In uno studio è stato utilizzato un dosaggio di 37,5mg di Ferro elementare al giorno. Pertanto, a intervalli più frequenti, è consigliabile un controllo con analisi del sangue. Inoltre, va ricordato che una flebotomia non monitorata nei tempi di prelievo può portare a rebound dell’ematocrito con peggioramento del quadro clinico ematico.
La “Dbol Hematocrit Theory”:
Come accennato nell’introduzione, alcuni preparatori hanno osservato una “anomalia” con la somministrazione di Methandrostenolone. Questa “anomalia” consisteva in una sensibile riduzione dell’Ematocrico e in una certa misura una alterazione della conta dei globuli bianchi, in special modo dei Linfociti.
La risposta osservata si è verificata in condizioni di monoterapia, quindi priva anche di una base TRT. Ciò è stato fatto per evitare che soggetti sensibili all’aumento del Ematocrito subissero un interazione negativa anche da dosi terapeutiche di Testosterone.
Esami ematici di confronto tra pre-trattamento [sinistra] e post-trattamento con Methandrostenolone. Come si può osservare, l’Ematocrito dal 54% è passato al 47%, gli Eritrociti dal 5,8 T/l al 5,1 T/l, l’Emoglobina da 18,7g/dL a 15,8g/dL e i Leucociti da 7,6 G/l a 4,8 G/l. Fonte documentazione Alberto Prevedi .
Alcuni di voi diranno “ma come? Senza una base di Testosterone il soggetto andrebbe in uno stato di malessere psicofisico dipendente dalla riduzione marcata di Estradiolo e DHT!”. Questa affermazione è parzialmente vera ma:
Sebbene il Methandrostenolone sia un derivato 17α-Metilato del Boldenone , sembra soggetto ad una maggiore conversione in 17α-Methylestradiolo. A causa della presenza del suo gruppo metilico C17α, il 17α-Methylestradiolo non può essere disattivato mediante ossidazione del gruppo ossidrile C17β, con conseguente miglioramento della stabilità metabolica e della potenza rispetto al 17β-Estradiolo.[44];
L’uso di DHT esogeno o di analogo metilato in C-1 sopperisce generalmente alla bassa 5α-riduzione senza interferire con la finalità del protocollo su Ematocrito.
A questo punto la domanda è: “Quali sono i meccanismi attraverso i quali l’uso monoterapico del Methandrostenolone porta ad una riduzione dell’Ematocrito e ad una risposta paradossale del Emopoiesi?”
Al momento possiamo solo ipotizzare utilizzando l’attuale conoscenza in nostro possesso riguardo ai meccanismi androgeni su Emopoiesi e Eritropoiesi. Le ipotesi in merito sono le seguenti:
Ipotesi della riduzione attività e concentrazioni di 17-Ketosteroidi: Sappiamo che il Methandrostenolone viene metabolizzato nel fegato mediante 6β-idrossilazione, 3α- e 3β-ossidazione, 5β-riduzione, 17-epimerizzazione e coniugazione tra le altre reazioni. Non mi è stato possibile reperire materiale in riferimento riguardante eventuali metaboliti 17-ketosteroidi del Methandrostenolone. Sapendo che, gli androgeni convertendo in 17-Ketosteroidi sono in grado di aumentare la sintesi di mRNA nel nucleo, causando la differenziazione delle cellule del midollo osseo da non responsive all’EPO a responsive all’EPO. Una eventuale riduzione dei metaboliti di conversione ed interconversione dei 17-Ketosteroidi [vedi enzimi 17β-Hydroxysteroide dehydrogenasi;17β-HSD] dati da soppressione/sottoregolazione di sintesi e scarsa affinità enzimatica del Methandrostenolone potrebbero esserne la causa. Mentre per quanto riguarda la riduzione dei Leucociti, essa potrebbe essere riconducibile ad una attività immunosoppressiva osservata anche con l’uso di altri AAS.
Ipotesi della attività di legame antagonista: si potrebbe ipotizzare che il Methandrostenolone possa agire come una molecola antagonista/agonista dei recettori cellulari del midollo emopoietico portando ad una riduzione dell’attività dei metaboliti 17-ketosteroidi riducendo sia l’Eritropoiesi che la formazione di globuli bianchi e piastrine [derivanti da un unica cellula staminale emopoietica pluripotente]. Il problema è che se ciò fosse vero, la regolazione del segnale indotto dall’attività antagonista/agonista sembra essere maggiore per la risposta eritrocitaria e leucocitaria.
Ipotesi del metabolita “sintetico” 17-ketosteroideo: si potrebbe ipotizzare che il Methandrostenolone converta in una forma non ancora identificata di 17-ketosteroide avente effetto agonista/antagonista a livello delle cellule staminali emopoietiche multinucleate. Si ricordi, infatti, che di recente (2010) è stato identificato un metabolita del Methandrostenolone nei campioni di urina fino a 19 giorni dopo la sua somministrazione. Il metabolita in questione è il 17beta-idrossimetil-17 alfa-metil-18-norandrosta-1,4,13-trien-3-one (20OH-NorMD), scoperto tramite LC-MS/MS e GC -SM. L’enzima CYP21 e, in misura minore, anche il CYP3A4 possono catalizzare questa idrossilazione dello steroide. Non escluderei in assoluto, quindi, la presenza di un metabolita non ancora identificato[45].
Struttura base 17-Ketosteroide.
E’ bene ricordare che il Boldenone, precursore strutturale del Methandrostenolone, mostra in corso di trattamento un aumento significativo della conta eritrocitaria totale e dei valori di emoglobina ed ematocrito, mentre gli indici medi di emoglobina corpuscolare e di concentrazione media di emoglobina corpuscolare sembrano diminuire. Il leucogramma, similmente all’effetto notato con il Methandrostenolone, mostra leucopenia, linfopenia e granulocitosi rispetto al controllo negli studi su animali e in alcuni casi studio. Il metabolita androsta–1,4–dien–17 beta-ol-3-one sembra avere un ipotetico ruolo significativo negli effetti emopoietici del Boldenone insieme ad altri steroidi strettamente correlati al 17β-boldenone. E ricordiamoci che le 17β-idrossisteroide deidrogenasi (17β-HSD, HSD17B) (EC 1.1.1.51), anche 17-chetosteroide reduttasi (17-KSR), il gruppo di alcol ossidoreduttasi che catalizzano la riduzione dei 17-Ketosteroidi e la deidrogenazione dei 17β-idrossisteroidi nella steroidogenesi e nel metabolismo degli steroidi, sono implicate nel metabolismo dei 17-ketosteroidi che, a loro volta, sono direttamente correlati alla eritropoiesi/emopoiesi.[46][47]
I dosaggi di Methandrostenolone generalmente utilizzati per tale scopo si attestano nel range dei 10-15mg/die. In questo modo l’atleta, in un discreto numero di casi, non è obbligato a cessare completamente l’utilizzo di AAS sfruttando il forte potenziale anticatabolico del Methandrostenolone con finalità stabilizzative. In corso di terapia il soggetto si sottopone a flebotomia con regolarità in funzione delle risposte al trattamento valutate per via esami ematici.
Nota: durante il periodo di trattamento, l’uso di hCG non è indicato dal momento che l’aumento nella biosintesi testicolare di Testosterone potrebbe alterare la risposta di controllo del Eritropoiesi/Emopoiesi con conseguente non ottenimento della risposta terapeutica ricercata.
Le limitazione di applicazione della monoterapia con Methandrostenolone per il trattamento della eritrocitosi sono le seguenti:
Stato dislipidemico marcato [alterazione marcata dei livelli di HDL-C (<25mg/dL), LDL-C (>150mg/dL), Trigliceridi (>80mg/dL);
coesistenza della prima citata dislipidemia ematica con livelli di Omocisteine > 13µmol/L;
alterazioni marcate delle Transaminasi e/o dei risultati della elettroforesi proteica;
eGFR < 60, Creatininuria da raccolta urine nelle 24h >2000mg/dL oppure da campione di urine al mattino >300mg/dL, Creatininemia 2mg/dL, Cistatina C >1mg/Lt con valutazione della elettroforesi proteica.
Conclusioni:
L’identificazione del fattore, o dei fattori, implicati nella risposta sottoregolativa sulla Eritropoiesi/Emopoiesi in monoterapia con Methandrostenolone è assai lungi dall’essere con sufficiente sicurezza scoperta. Ad oggi, però, le ipotesi sopra esposte rappresentano le possibilità con la probabilità maggiore di riscontro positivo con eventuali futuri studi specifici.
Si rammenta che ogni cosa esposta concernente la “Dbol Hematocrit Theory” è puramente ipotetico-teorica, non rappresenta quindi una pratica scientificamente avvalorata ne tantomeno un consiglio terapico. Chiunque decida liberamente di utilizzare le informazioni presentate per improvvisarsi ricercatore e/o cavia, lo fa prendendosene la piena responsabilità d’esito.
In conclusione, ringrazio i preparatori (colleghi) che mi hanno fornito esami di confronto e dati raccolti in anni di osservazione attenta degli esami ematici. In particolare modo ringrazio Alberto Prevedi che mi ha fornito gli esami esposti in questo articolo.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
1.Steinglass P, Gordon AS, Charipper HA. Effect of castration and sex hormones on blood of the rat. Proc Soc Exp Biol Med. (1941) 48:169–77. doi: 10.3181/00379727-48-13259
2. Crafts RC. Effects of hypophysectomy, castration and testosterone propionate on hemopoiesis in the adult male rat. Endocrinology. (1946) 39:401–13. doi: 10.1210/endo-39-6-401
3. Vahlquist B. The cause of the sexual differences in erythrocyte, hemoglobin and serum iron levels in human adults. Blood. (1950) 5:874–5. doi: 10.1182/blood.V5.9.874.874
4. Hartgens F, Kuipers S. Effects of androgenic-anabolic steroids in athletes. Sports Med. (2004) 34:513–54. doi: 10.2165/00007256-200434080-00003
5. McCullagh EP, Jones R. A note on the effect of certain androgens upon red blood cell count and upon glucose tolerance. Cleve Clin Q. (1941) 8:79–84. doi: 10.3949/ccjm.8.2.79
6. Shahani S, Braga-Basaria M, Maggio M, Basaria S. Androgens and erythropoiesis: past and present. J Endocrinol Invest. (2009) 32:704–16. doi: 10.1007/BF03345745
7. Shimoda K, Shide K, Kamezaki K, Okamura T, Harada N, Kinukawa N, et al. The effect of anabolic steroids on anemia in myelofibrosis with myeloid metaplasia: retrospective analysis of 39 patients in Japan. Int J Hematol. (2007) 85:338–43. doi: 10.1532/IJH97.06135
8.Jaime-Pérez JC, Colunga-Pedraza PR, Gómez-Ramírez CD, Gutiérrez-Aguirre CH, Cantú-Rodríguez OG, Tarín-Arzaga LC, et al. Danazol as first-line therapy for aplastic anemia. Ann Hematol. (2011) 90:523–7. doi: 10.1007/s00277-011-1163-x
9.Gagliano-Jucá T, Pencina KM, Ganz T, Travison TG, Kantoff PW, Nguyen PL, et al. Mechanisms responsible for reduced erythropoiesis during androgen deprivation therapy in men with prostate cancer. Am J Physiol Endocrinol Metab. (2018) 315:E1185–93. doi: 10.1152/ajpendo.00272.2018
10.Larner J. Intermediary Metabolism and Its Regulation. EnglewoodCliffs, NJ: Prentice Hall Inc. (1971). p. 228.
11.Molinari PF, Neri LL. Effect of a single oral dose of oxymetholone on the metabolism of human erythrocytes. Exp Hematol. (1978) 6:648–54.
12.Bachman E, Feng R, Travison T, Li M, Olbina G, Ostland V, et al. Testosterone suppresses hepcidin in men: a potential mechanism for testosterone-induced erythrocytosis. J Clin Endocrinol Metab. (2010) 95:4743–7. doi: 10.1210/jc.2010-0864
13. Naets JP, Wittek M. The mechanism of action of androgens on erythropoiesis. Ann N Y Acad Sci. (1968) 149:366–76.
14. Rishpon-Meyerstein N, Kilbridge T, Simone J, Fried W. The effect of testosterone on erythropoietin levels in anaemic patients. Blood. (1968) 31:453–60. doi: 10.1182/blood.V31.4.453.453
15. Hagenfeldt Y, Linde K, Sjoberg HE, Zumkeller W, Arver S. Testosterone increases serum 1,25-dihydroxyvitamin D and insulin like growth factor-I (IGF-1) in hypogonadal men. Int J Androl. (1992) 15:93–102. doi: 10.1111/j.1365-2605.1992.tb01118.x
17. Shahidi NT. Androgens and erythropoiesis. N Engl J Med. (1973) 289:72–80. doi: 10.1056/NEJM197307122890205
18. Krabbe S, Christensen T, Worm J, Christiansen C, Transbøl I. Relationship between haemoglobin and serum testosterone in normal children and adolescents and in boys with delayed puberty. Acta Paediatr Scand. (1978) 67:655–8. doi: 10.1111/j.1651-2227.1978.tb17818.x
19. Hero M, Wickman S, Hanhijärvi R, Siimes MA, Dunkel L. Pubertal upregulation of erythropoiesis in boys is determined primarily by androgen. J Pediatr. (2005) 146:245–52. doi: 10.1016/j.jpeds.2004.09.002
20. Tong PC, Kong AP, So WY, Ng MH, Yang X, Ng MC, et al. Hematocrit, independent of chronic kidney disease, predicts adverse cardiovascular outcomes in Chinese patients with type 2 diabetes. Diabetes Care. (2006) 29:2439–44. doi: 10.2337/dc06-0887
21. Karakitsos D, Patrianakos AP, De Groot E, Boletis J, Karabinis A, Kyriazis J, et al. Androgen deficiency and endothelial dysfunction in men with end-stage kidney disease receiving maintenance hemodialysis. Am J Nephrol. (2006) 26:536–43. doi: 10.1159/000097816
22. Carrero JJ, Qureshi AR, Nakashima A, Arver S, Parini P, Lindholm B, et al. Prevalence and clinical implications of testosterone deficiency in men with end-stage renal disease. Nephrol Dial Transplant. (2011) 26:184–90. doi: 10.1093/ndt/gfq397
23. Handelsman DJ. Hypothalamic-pituitary gonadal dysfunction in renal failure, dialysis, and renal transplantation. Endocr Rev. (1985) 6:151–82. doi: 10.1210/edrv-6-2-151
24.National Kidney Foundation. K/DOQI clinical practice guidelines for anemia of chronic kidney disease. Am J Kidney Dis. (2000) 37:182–238. doi: 10.1016/S0272-6386(01)70008-X
25. Kanagasundaram N, Shipley T, Wright R, Playford M, Peaston R, Moochhala S, et al. The Prevalence of Androgen Deficiency in Male Patients With Advanced CKD – An Evaluation in Pre-dialysis and Haemodialysis Population. (2010). Available online at: http://www.renalarchive.org/FetchDoc.aspx?id=6309 (accessed June 26, 2019).
26.DeLong M, Logan JL, Yong KC, Lien YH. Renin angiotensin blockade reduces serum free testosterone in middle-aged men on haemodialysis and correlates with erythropoietin resistance. Nephrol Dial Transplant. (2005) 20:585–90. doi: 10.1093/ndt/gfh638
27. Pfeffer MA, Burdmann EA, Chen CY, Cooper ME, de Zeeuw D, Eckardt KU, et al. A trial of darbepoetin alfa in type 2 diabetes and chronic kidney disease. N Engl J Med. (2009) 361:2019–32. doi: 10.1056/NEJMoa0907845
28.Wu WC, Rathore SS, Wang Y, Radford MJ, Krumholz HM. Blood transfusion in elderly patients with acute myocardial infarction. N Engl J Med. (2001) 345:1230–6. doi: 10.1056/NEJMoa010615
29. Ezekowitz JA, McAlister FA, Armstrong PW. Anemia is common in heart failure and is associated with poor outcomes: insights from a cohort of 12,065 patients with new-onset heart failure. Circulation. (2003) 107:223–5. doi: 10.1161/01.CIR.0000052622.51963.FC
30. Wu FC, Tajar A, Beynon JM, Pye SR, Silman AJ, Finn JD, et al. Identification of late-onset hypogonadism in middle-aged and elderly men. N Engl J Med. (2010) 363:123–35. doi: 10.1056/NEJMoa0911101
31.Dhindsa S, Ghanim H, Batra M, Kuhadiya ND, Abuaysheh S, Green K, et al. Effect of testosterone on hepcidin, ferroportin, ferritin and iron binding capacity in patients with hypogonadotropic hypogonadism and type 2 diabetes. Clin Enocrinol. (2016) 85:772–28. doi: 10.1111/cen.13130
32.Roy CN, Snyder PJ, Stephens-Shields AJ, Artz AS, Bhasin S, Cohen HJ, et al. Association of testosterone levels with anemia in older men: a controlled clinical trial. JAMA Intern Med. (2017) 177:480–90. doi: 10.1001/jamainternmed.2016.9540
33. Al-Sharefi A, Quinton R. Re:male hypogonadism, a treatable yet forgotten cause of unexplained anaemia. REMARQ commentary on Strauder et al. Anemia at older age: etiologies, clinical implications, and management. Blood. (2018) 131:505–14. doi: 10.1182/blood-2017-07-746446
34. Bhasin S, Brito JP, Cunningham GR, Hayes FJ, Hodis HN, Matsumoto AM, et al. Testosterone therapy in men with hypogonadism: an endocrine scoety clinical practice guidelines. J Clin Endocrinol Metab. (2018) 103:715–44. doi: 10.1210/jc.2018-00229
35.Coviello, Andrea D., et al. “Effects of graded doses of testosterone on erythropoiesis in healthy young and older men.” The Journal of Clinical Endocrinology & Metabolism 93.3 (2008): 914-919.
36.Bachman, Eric, et al. “Testosterone suppresses hepcidin in men: a potential mechanism for testosterone-induced erythrocytosis.” The Journal of Clinical Endocrinology & Metabolism 95.10 (2010): 4743-4747.
37.Bhasin, Shalender, et al. “Testosterone dose-response relationships in healthy young men.” American Journal of Physiology-Endocrinology And Metabolism (2001).
38.Baigent, Colin, et al. “Aspirin in the primary and secondary prevention of vascular disease: collaborative meta-analysis of individual participant data from randomised trials.” Lancet 373.9678 (2009): 1849-1860.
39.F. Hobbs, M. Piepoli, A. Hoes, S. Agewall, C. Albus, C. Brotons, A. Catapano, M. Cooney, U. Corra, B. Cosyns, et al. 2016 european guidelines on cardiovascular disease prevention in clinical practice. European Heart Journal, 37(29):2315–2381, 2016.
40.M. Lordkipanidzé, J. G. Diodati, and C. Pharand. Possibility of a rebound phenomenon following antiplatelet therapy withdrawal: a look at the clinical and pharmacological evidence. Pharmacology & therapeutics, 123(2):178–186, 2009.
41.L. A. G. Rodríguez, L. C. Soriano, C. Hill, and S. Johansson. Increased risk of stroke after discontinuation of acetylsalicylic acid a uk primary care study. Neurology, pages WNL–0b013e31820d62b5, 2011
42.B. Chin-Yee, A. Lazo-Langner, T. Butler-Foster, C. Hsia, and I. Chin-Yee. Blood donation and testosterone replacement therapy. Transfusion, 57(3):578–581, 2017
43.Kiss, Joseph E., et al. “Oral iron supplementation after blood donation: a randomized clinical trial.” Jama 313.6 (2015): 575-583.
44.Thieme D, Hemmersbach P (18 December 2009). Doping in Sports. Springer Science & Business Media. pp. 470
46.Labrie F, Luu-The V, Lin SX, Labrie C, Simard J, Breton R, Bélanger A (January 1997). “The key role of 17 beta-hydroxysteroid dehydrogenases in sex steroid biology”. Steroids. 62 (1): 148–58.
In questo articolo mi concentrerò sugli effetti degli androgeni sulla libido negli uomini (nel prossimo articolo analizzerò gli effetti degli androgeni sulla libido nelle donne). Verrà inoltre presentato il Nandrolone come controesempio al generale aumento della libido da parte degli androgeni e verranno enumerati e discussi i meccanismi putativi che ostacolano la libido e la funzione sessuale. Infine, verrà discusso il neurotrasmettitore dopamina e la sua influenza sul sistema eccitatorio, in particolare la sua relazione con la soppressione della libido da parte del Nandrolone. Si ipotizza un nuovo meccanismo in base al quale il Nandrolone contribuisce probabilmente a sopprimere la libido attraverso un aumento del metabolismo della dopamina.
Negli uomini, gli androgeni endogeni Testosterone (T) e il suo prodotto della 5α-reduttasi, il DHT, nonché l’Estradiolo, esercitano un chiaro effetto organizzatore e attivatore sul comportamento sessuale, compreso il desiderio sessuale (libido) che è riconducibile allo spermarca (la comparsa della prima eiaculazione) e alla maturazione (la pubertà nel maschio è associata a un aumento di 18 volte delle concentrazioni di Testosterone endogeno). [1].
Gli uomini si differenziano ampiamente dalle donne per quanto riguarda la libido. Pochi sosterrebbero, nonostante alcune variazioni e l’esistenza di relativi outlier, che gli uomini non siano caratteristicamente più virili delle donne. Eppure, nonostante un ampio consenso tra i ricercatori biologi e biomedici, così come tra i non addetti ai lavori, sul fatto che il Testosterone sia implicato nelle differenze di sesso nella libido e che anzi migliori la libido, in letteratura si trovano prove sorprendentemente contrastanti su questo argomento, in particolare per quanto riguarda le differenze nelle e tra le donne. Gli effetti particolari degli androgeni sulla libido nelle donne sono discussi nella seconda parte di questa serie.
Definizioni
Libido: desiderio o interesse sessuale che nasce dall’eccitazione e dalla risposta centrale, che si manifesta con pensieri sul sesso accompagnati da una risposta genitale. [1].
Funzione sessuale: concetto più ampio che comprende le misure della libido (ad esempio, il Male Sexual Health Questionnaire: desiderio sessuale [comprehensive]), come un aspetto o una componente.
Gli steroidi anabolizzanti-androgeni (AAS) o gli androgeni migliorano, con differenze tra molecole, la funzione sessuale (agendo sul SNC e sui tessuti genitali). [2].
Effetto soglia
È chiaro che la TRT (terapia sostitutiva del Testosterone) somministrata terapeuticamente a un uomo ipogonadico allevierà quasi certamente i sintomi della scarsa libido, in assenza di una patologia organica della funzione sessuale.
Concettualmente, la visione medica prevalente è che le concentrazioni di Testosterone negli uomini sono soggette a un effetto soglia, con una linea di base stabilita di concentrazioni normali di Testosterone totale (TT) e Testosterone libero (fT) (normale TT 450 – 1.000 ng/dL e fT 1 – 2% di TT), al di sotto della quale prevalgono gli effetti negativi sulla libido e sulla funzione sessuale; e al di sopra della quale si manifestano pochi cambiamenti comportamentali.
La TRT per alleviare i sintomi dell’ipogonadismo (compresa la scarsa libido) mostra effetti chiari e consistenti, aumentando generalmente la frequenza dei rapporti sessuali e della masturbazione. Tuttavia, è necessario fare importanti distinzioni tra i pazienti che utilizzano la TRT. In primo luogo, la TRT viene spesso prescritta a uomini anziani, altrimenti sani e con relazioni stabili, il che aumenta naturalmente le opportunità di rapporti sessuali. La TRT viene prescritta più frequentemente in culture che hanno una visione liberale della masturbazione e ne riportano onestamente la frequenza. In effetti, gli uomini differiscono da una cultura all’altra per quanto riguarda la manifestazione della libido come comportamento masturbatorio [3]; e la popolazione dei consumatori di AAS o androgeni (ad esempio, per obiettivi di miglioramento del fisico o delle prestazioni) può essere qualitativamente diversa da quella della TRT. Pertanto, le misure dei cambiamenti della libido causati dagli androgeni devono necessariamente essere misurate in modo da non essere influenzate da questi fattori culturali o socio-relazionali.
Esiste davvero un “effetto tetto” ai livelli endogeni, al di sopra del quale gli androgeni non hanno alcun effetto sulla libido?
È interessante notare che le prove suggeriscono fortemente l’assenza di un limite superiore teorico, o tetto massimo, per gli effetti comportamentali degli androgeni. I dati relativi agli uomini normali e sani (la popolazione a cui si rivolge la visione medica prevalente) suggeriscono che non esiste un limite superiore teorico, o almeno che, se tale limite esiste, è di gran lunga superiore anche ai livelli endogeni normali di androgeni circolanti:
Anderson e colleghi hanno dimostrato che una dose settimanale di 200mg di Testosterone Enantato aumenta l’interesse sessuale in uomini normali e sani. In particolare, i risultati hanno mostrato un aumento dei punteggi della Sottoscala 2 della Sexual Experience Scale, che misura l’entità in cui un individuo cerca o permette (piuttosto che evita o rifiuta) stimoli sessuali di tipo audiovisivo o immaginario; si tratta, quindi, di un indice di interesse sessuale indipendente dall’interazione con un partner (maggiore validità rispetto alla frequenza del coito, poiché la disponibilità di un partner romantico influenza la frequenza delle attività sessuali; la frequenza della masturbazione è influenzata culturalmente; ecc.) [4].
Su e colleghi hanno dimostrato che una dose giornaliera di 240mg di Metiltestosterone ha aumentato l’eccitazione sessuale su scala analogica visiva (VAS) in uomini normali e sani. [5].
Moss e colleghi hanno dimostrato che gli atleti maschi che fanno uso di androgeni si impegnano in una maggiore frequenza di rapporti sessuali e raggiungono un numero più elevato di eiaculazioni settimanali (tutti i soggetti avevano la disponibilità di un partner sessuale) rispetto alle loro controparti che non fanno uso di androgeni. [6].
Funzione del recettore degli androgeni nel SNC (maschi)
I modelli di knockout del recettore degli androgeni (ARKO) sono uno strumento utile per studiare la funzione dei T/androgeni. Nei tessuti del SNC, l’eliminazione del recettore AR (espressione nulla) provoca nei roditori maschi un comportamento privo di attività sessuale e aggressività. [7].
Ciò ha implicazioni teleologiche: non è solo un costrutto sociale che ci si aspetta che gli uomini, entro i limiti della società, siano l’inseguitore romantico (chiedere un appuntamento, piuttosto che lo faccia la donna); e forse anche che mostrino aggressività in camera da letto (essere un gentiluomo per le strade, ma una bestia tra le lenzuola). È probabile che le pressioni ambientali e l’interesse della specie umana per la riproduzione si basino sul fatto che gli uomini affermino il loro ruolo sessuale nella competizione per le compagne e che questa competizione sia necessariamente legata alla prestanza fisica. Come spesso accade, la società interagisce con la biologia, definendo i confini dell’aggressività maschile e della ricerca sessuale.
L’ipotesi della desensibilizzazione (che verrà approfondita nel prossimo articolo) descrive un modello teorico che trae inferenze su alcuni probabili processi biologici ed epigenetici che determinano la desensibilizzazione comportamentale agli androgeni nei maschi a causa dell’aumento perinatale del Testosterone (e, di conseguenza, la manifestazione di comportamenti potenzialmente disadattivi se tale desensibilizzazione viene abbandonata, in età adulta, quando i livelli di T aumentano di 18 volte). In particolare, Bancroft e colleghi sostengono che “l’esposizione a livelli di T sostanzialmente più elevati durante lo sviluppo fetale e anche durante le prime settimane postnatali [il picco perinatale] potrebbe essere responsabile della desensibilizzazione del SNC agli effetti del T nel maschio. Tale desensibilizzazione agirebbe presumibilmente a livello genomico piuttosto che nella fase recettoriale dell’azione ormonale… e a breve termine, sia l’esposizione al T che al DHT determina una sovraregolazione dell’AR. Una conseguenza di tale desensibilizzazione nel maschio consisterebbe nelle variazioni geneticamente determinate nella reattività dei recettori del SNC al T che verrebbero “appiattite”, consentendo livelli molto più elevati di T dalla pubertà in poi senza iperstimolazione dei meccanismi del SNC.” [1].
La diminuzione della libido Nandrolone-correlata:
Un farmaco in particolare, utilizzato clinicamente e terapeuticamente negli uomini e nelle donne, che rappresenta un controesempio all’apparente aumento della libido da parte degli androgeni è il Nandrolone. Infatti, è spesso associato a una riduzione della libido. [8]. Una logica non dichiarata – forse poco rassicurante – del suo uso negli uomini con HIV, piuttosto che il Testosterone, è quella di ridurre la libido (e quindi di ridurre le interazioni sessuali tra uomini gay per ridurre la diffusione della malattia).
Hulsbæk e colleghi hanno somministrato il Nandrolone in dosi diverse a tre gruppi: (1) uomini il cui Testosterone totale (TT) era ≥11 nmol/L (100 mg di Nandrolone Decanoato ogni tre settimane), (2) uomini il cui TT era <11 nmol/L (200mg di Nandrolone Decanoato ogni tre settimane) e (3) donne (50mg ogni tre settimane), per 12 settimane. I risultati non hanno mostrato alcuna incidenza di aumento della libido da parte del Nandrolone Decanoato (0/9 donne e 0/3 uomini; 1 incidenza di aumento della libido è stata riportata nel gruppo placebo, a cui è stato somministrato un veicolo inattivo a base di olio senza androgeni). [9].
Prolattina
È stato affermato che è improbabile che il Nandrolone diminuisca la libido aumentando la Prolattina. Mentre alti livelli di Prolattina negli uomini, come nel caso degli adenomi che secernono Prolattina, sono associati a sintomi di ipogonadismo (cioè bassa libido) e persino galattorrea (lattazione), è improbabile che il Nandrolone aumenti la Prolattina (almeno alle dosi utilizzate nella pratica comune). In generale, gli androgeni aromatizzabili (ad esempio, Testosterone [10], MENT [11]) mostrano una tendenza (una tendenza, piuttosto che un effetto significativo) ad aumentare la Prolattina sierica come conseguenza dei loro prodotti aromatici (cioè gli estrogeni) che agiscono come fattori stimolanti la secrezione di Prolattina dall’ipofisi anteriore [10]. Il Nandrolone a dosi inferiori non sembra avere effetti significativi sulla Prolattina sierica (probabilmente a causa di livelli di E2 inferiori alla norma) [12].
Al contrario, gli androgeni non aromatizzabili (ad esempio, Trenbolone, Oxandrolone, ecc.) probabilmente riducono la Prolattina sierica. Questa è un’osservazione empirica basata sui risultati di analisi del sangue umano, nonché la base di un’ipotesi prevalente, a conoscenza di questo autore, avanzata per la prima volta da De Las Heras e colleghi nel 1979. [13].:
Poiché è stato riportato che la secrezione di Prolattina nel ratto maschio è pulsatile (17), l’analisi delle differenze tra i livelli basali di Prolattina basata su una singola determinazione può essere fuorviante. Una possibilità alternativa è che alcuni androgeni siano in realtà inibitori della secrezione di Prolattina. Nei nostri studi, i valori più bassi tra tutti i gruppi sono stati ottenuti negli animali trattati con Diidrotestosterone o Androstanediolo, anche se le differenze non hanno mai raggiunto la significatività. Nolin et al. (11) hanno riportato che il Diidrotestosterone ha soppresso in modo significativo i livelli di Prolattina in ratti femmina intatti.
Tra i fattori che probabilmente influenzano l’incapacità del Nandrolone di aumentare – e persino di diminuire – la libido vi sono [8]:
Estrogeni*: Il nandrolone tende a determinare livelli di estrogeni sub-normali negli uomini a dosi terapeutiche fino a 200mg settimanali. Dopo 6 settimane, l’Estradiolo sierico (E2) si è ridotto a 11 ± 9pg/mL con una dose settimanale di 100mg di Nandrolone Decanoato e a 14 ± 4pg/mL con una dose settimanale di 200mg di Nandrolone Decanoato in uomini normali [14]. L’influenza degli estrogeni e dell’Estradiolo sulla libido non è stata stabilita, né negli uomini né nelle donne; tuttavia, alcune indicazioni suggeriscono una curva a forma di U inversa rispetto alle concentrazioni di Estradiolo e alla libido (con concentrazioni troppo basse e troppo alte che causano riduzioni della libido).
5α-riduzione a DHN, un androgeno indebolito: Il Nandrolone, a differenza del Testosterone che viene convertito in DHT, più potente, nel SNC e nei tessuti sessuali dalla 5α-reduttasi, diminuisce la sua potenza androgena in questi organi bersaglio producendo DHN (5α-diidronandrolone). È stato dimostrato che gli inibitori della 5α-reduttasi (ad esempio, Dutasteride, Finasteride) che riducono la 5α-riduzione del T in DHT sono associati a una riduzione della libido negli uomini sani. [15].
Feedback negativo (inibizione) delle gonadotropine: Il Nandrolone, privo del gruppo metile C-19 del Testosterone, presenta un’omologia più ampia per la superfamiglia dei recettori nucleari, compreso il Recettore del Progesterone (PR), dati gli effetti di questa modifica sulle sue proprietà stereochimiche e sulla sua forma conformazionale. Pertanto, contribuisce a disregolare la secrezione di gonadotropine regolata dal GnRH ipotalamico, che comprende l’interazione tra la secrezione dell’Ormone Luteinizzante (LH) e dell’Ormone Follicolo-Stimolante (FSH) dall’ipotalamo [regolatori positivi], della globulina legante gli ormoni sessuali (SHBG) e dell’inibina dalle cellule del Sertoli [regolatori negativi] e del T dalle cellule di Leydig [regolatore negativo], in modo più pluripotente rispetto al Testosterone. Il Nandrolone serve a disregolare questo sistema di regolazione agendo per: –esercitare un feedback negativo sull’ipofisi (secrezione di LH) attraverso la sua aromatizzazione, anche se ridotta, in Estradiolo e maggiormente in Estrone -rallentare la frequenza degli impulsi dell’ormone di rilascio delle gonadotropine (GnRH) ipotalamico mediante un’azione androgena ed estrogenica -disregolare la regolazione ipotalamica del T e delle gonadotropine attraverso la segnalazione/pulsatilità del dendro KNDy come analogo del progestinico o del Progesterone -aumentano l’espressione della Prolattina ad alte dosi (attraverso il suo prodotto aromatico, l’Estrone e, in misura minore, l’Estradiolo) – l’elevata Prolattina sierica ha un ruolo terziario nella riduzione della libido (principalmente agendo sull’ipotalamo). [16].
Aumento del metabolismo della Dopamina (cioè della sua degradazione netta): A conoscenza di questo autore, si tratta di un nuovo meccanismo putativo, mai proposto prima, per il modo in cui il Nandrolone può contribuire alla riduzione della libido: il Nandrolone aumenta l’acido omovanillico (HVA) nel siero dell’uomo [14], riflettendo il suo metabolismo (cioè la sua degradazione), ed è probabilmente correlato alla riduzione del numero di recettori della Dopamina, L’acido omovanillico (HVA) sierico è cambiato significativamente con il Nandrolone Decanoato a 100mg settimanali (+17,6 ± 7,7 pmol/L) e con il Nandrolone Decanoato a 300mg settimanali (+11,0 ± 3,3 pmol/L), ma non nei gruppi di Testosterone Enantato a 100mg o di Testosterone Enantato a 300mg settimanali [14]. Quindi, anche a dosaggi clinici terapeutici, esiste un effetto del Nandrolone sul metabolismo della Dopamina. Gli stimoli erotici di natura audiovisiva riducono l’HVA [17], riflettendo l’aumento dell’attività dopaminergica associata all’eccitazione sessuale. Ne consegue quindi che, se l’attività dopaminergica è parte integrante della libido e delle manifestazioni della risposta genitale e dei pensieri sul sesso derivanti dall’eccitazione centrale (riflessa dalla diminuzione dell’HVA), gli effetti del Nandrolone sul metabolismo della Dopamina (riflessi dall’aumento dell’HVA) potrebbero determinare una riduzione dell’eccitazione sessuale.
*Il ruolo esatto dell’Estradiolo in ogni area della funzione sessuale maschile, compresa la libido, la funzione erettile e la spermatogenesi, è difficile da determinare con esattezza. Un complesso equilibrio di Testosterone, Estradiolo, Aromatasi ed ER nei testicoli, nel pene e nel cervello conferma un’interazione ormonale indispensabile e altamente regolata degli estrogeni nell’uomo. Gli ER e l’Aromatasi condividono le posizioni topografiche con i feromoni nel cervello, rendendo chiaro che gli estrogeni contribuiscono allo sviluppo sessuale precoce e al comportamento sessuale in età adulta. Gli estrogeni possono sostenere la libido e influenzare la quantità di recettori della serotonina nel cervello, modulando l’umore, lo stato mentale, la cognizione e le emozioni. La funzione erettile è influenzata negativamente dall’esposizione agli estrogeni nelle prime fasi dello sviluppo del pene e l’esposizione all’Estradiolo nel pene maturo porta a un aumento della permeabilità vascolare con conseguente aumento della disfunzione erettile. La disfunzione erettile dovuta a una maggiore esposizione all’Estradiolo è indipendente dal livello di Testosterone. Inoltre, la spermatogenesi dipende in qualche misura dall’Estradiolo, poiché tutte le cellule coinvolte nel processo di produzione dello sperma contengono Aromatasi ed esprimono ER. Infine, i livelli di Estradiolo devono essere presi in considerazione quando si trattano uomini con TRT, poiché i livelli di estradiolo inferiori a 5ng/dl sono correlati a una diminuzione della libido.
Inoltre, l’Estrone, che rappresenta il maggior metabolita della aromatizzazione del Nandrolone, essendo meno potente del Estradiolo (circa il 4% dell’attività estrogenica del E2) nelle attività tissutali, aggrava la condizione di riduzione della libido e della funzionalità erettile negli utilizzatori di questo progestinico .
Dopamina e libido
Il sistema eccitatorio stimola la libido, mentre il sistema inibitorio stimola la ricompensa sessuale, la sedazione e la sazietà. Il nucleo del sistema eccitatorio si trova nei sistemi cerebrali della Dopamina (DA) (incertoipotalamico e mesolimbico) che collegano l’ipotalamo e i sistemi limbici e comprende le Melanocortine (MC), l’Ossitocina (OT) e la Noradrenalina (NE). Il sistema inibitorio contiene i sistemi cerebrali degli oppioidi, degli Endocannabinoidi (ECB) e della Serotonina (5-HT), che si attivano durante i periodi di inibizione sessuale e bloccano il sistema eccitatorio. [18].
I farmaci che stimolano l’attivazione della DA ipotalamica o che bloccano il rilascio di ECB o 5-HT e/o il legame postsinaptico possono stimolare la libido. [18].
Gli ormoni steroidei attivano i meccanismi di eccitazione sessuale dirigendo la sintesi di enzimi e recettori per i sistemi neurochimici interattivi di DA, NE, MC e OT, che agiscono nelle regioni cerebrali ipotalamiche e limbiche per stimolare l’eccitazione sessuale, l’attenzione e i comportamenti. L’attivazione di questi sistemi neurochimici eccitatori smorza l’influenza dei meccanismi inibitori, quali:
gli oppioidi endogeni rilasciati nella corteccia, nel sistema limbico, nell’ipotalamo e nel mesencefalo durante un orgasmo o una ricompensa sessuale (che inducono un periodo refrattario e una diminuzione dell’espressione dell’AR nelle regioni ipotalamiche e limbiche)
ECB che mediano la sedazione e
5-HT, che viene elevata in queste regioni per indurre refrattarietà e sazietà sessuale. [18].
L’Estradiolo (E2) facilita il rilascio di DA e il Testosterone (T) potenzia la sintesi di Ossido Nitrico che controlla il rilascio di DA nei ratti (86-88). [18]. Pertanto, gli ormoni steroidei endogeni sembrano porre le basi – [un effetto priming] – per un aumento della sintesi e del rilascio di DA durante i periodi in cui la risposta sessuale potrebbe essere potenziata. [18].
Il lavoro comportamentale (ad esempio, il corteggiamento negli esseri umani o l’attraversamento di griglie elettrificate per raggiungere una compagna nei ratti maschi) per acquisire la ricompensa sessuale, considerato analogo alla libido nella ricerca osservazionale sugli animali, è ridotto dalla castrazione, indicando che l’azione degli steroidi gonadici nel cervello è necessaria per lo sviluppo e/o il mantenimento di questo comportamento strumentale. Il lavoro comportamentale per acquisire la ricompensa sessuale (cioè la libido) è analogamente ridotto da lesioni all’amigdala basolaterale, una regione cerebrale che concentra gli steroidi, e dalla somministrazione di un antagonista della Dopamina al nucleo accumbens all’interno del sistema limbico. Questo comportamento strumentale viene ripristinato nei ratti maschi con lesioni all’amigdala basale dall’infusione di anfetamina nel nucleo accumbens, indicando che il rilascio mesolimbico di DA è parte integrante della libido. [18].
Esiste un nesso preciso tra gli ormoni steroidei (ad esempio, androgeni ed estrogeni), l’attività della Dopamina e la libido. Gli steroidi sessuali endogeni innescano la sintesi e il rilascio di Dopamina nei centri cerebrali chiave per sostenere le funzioni sessuali e la Dopamina stimola il sistema eccitatorio centrale per governare la libido e il conseguente comportamento sessuale.
Conclusione
Gli androgeni endogeni (Testosterone e DHT) esercitano un chiaro effetto sulla libido negli uomini; effetto nel quale è implicato anche l’Estradiolo. Gli androgeni sovrafisiologici generalmente aumentano la libido negli uomini (anche in quelli normali e sani), ma le modifiche chimiche degli androgeni possono influire sul fatto che determinati androgeni esercitino un effetto di aumento o addirittura di soppressione della libido. In generale, l’eccezione conferma la regola per quanto riguarda l’affermazione che gli androgeni tendono ad aumentare la libido negli uomini.
Il Testosterone, in quanto ormone sessuale maschile primario, ha la funzione biologica di controllare l’espressione del comportamento sessuale e aggressivo maschile, che deve necessariamente essere collegato alla prestanza fisica per quanto riguarda i vantaggi adattativi e competitivi per promuovere la sopravvivenza della specie umana.
I meccanismi dell’influenza degli androgeni sulla libido coinvolgono il neurotrasmettitore Dopamina e la sua attivazione del sistema eccitatorio, nonché gli effetti indiretti degli estrogeni, l’amplificazione periferica e la diminuzione, le gonadotropine dell’Asse Ipotalamo-Ipofisi-Gonadi, gli effetti sul metabolismo della Dopamina e persino gli effetti terziari sulla libido della Prolattina.
Per gli uomini è semplice: il Testosterone governa chiaramente la funzione sessuale e la libido (con una certa influenza dell’aromatizzazione in Estradiolo, in particolare nel SNC e nel cervello).
Nella prossimo articolo, analizzeremo come le donne siano più sfaccettate nelle dinamiche del loro ambiente ormonale e le ramificazioni dei cambiamenti ormonali del ciclo ovulatorio-mestruale; l’influenza predominante degli affetti (umore, benessere ed energia) sulla libido femminile; i problemi che derivano dalla difficoltà intrinseca di disgiungere gli effetti degli androgeni da quelli degli estrogeni nelle donne; l’ipotesi della desensibilizzazione e il suo potere esplicativo nel descrivere le differenze negli effetti degli androgeni sulla libido nelle donne rispetto agli uomini e tra le donne, nonché altri fattori che influenzano la libido e che sono correlati alle difficoltà nell’assegnare la causalità agli effetti degli androgeni esogeni sulla libido femminile.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Fonte studi articolo Type-IIx
[1] Bancroft, J. Androgens and sexual function in men and women. In: Bremner, W., Bagattel, C., eds. Androgens in health and disease. Totowa: Humana Press.
[2] Isidori, A. M., Buvat, J., Corona, G., Goldstein, I., Jannini, E. A., Lenzi, A., … Maggi, M. (2014). A Critical Analysis of the Role of Testosterone in Erectile Function: From Pathophysiology to Treatment—A Systematic Review. European Urology, 65(1), 99–112. doi:10.1016/j.eururo.2013.08.048
[3] Anderson, R. A., Martin, C. W., Kung, A. W. C., Everington, D., Pun, T. C., Tan, K. C. B., … Baird, D. T. (1999). 7α-Methyl-19-Nortestosterone Maintains Sexual Behavior and Mood in Hypogonadal Men1. The Journal of Clinical Endocrinology & Metabolism, 84(10), 3556–3562. doi:10.1210/jcem.84.10.6028
[4] Anderson, R. A., Bancroft, J., & Wu, F. C. (1992). The effects of exogenous testosterone on sexuality and mood of normal men. The Journal of Clinical Endocrinology & Metabolism, 75(6), 1503–1507. doi:10.1210/jcem.75.6.1464655
[5] Su, T.-P. (1993). Neuropsychiatric Effects of Anabolic Steroids in Male Normal Volunteers. JAMA: The Journal of the American Medical Association, 269(21), 2760. doi:10.1001/jama.1993.03500210060032
[6] Moss, H. B., Panzak, G. L., & Tarter, R. E. (1993). Sexual functioning of male anabolic steroid abusers. Archives of Sexual Behavior, 22(1), 1–12. doi:10.1007/bf01552908
[7] Sato, T., Matsumoto, T., Kawano, H., Watanabe, T., Uematsu, Y., Sekine, K., … Kato, S. (2004). Brain masculinization requires androgen receptor function. Proceedings of the National Academy of Sciences, 101(6), 1673–1678. doi:10.1073/pnas.0305303101
[8] Pan, M. M., & Kovac, J. R. (2016). Beyond testosterone cypionate: evidence behind the use of nandrolone in male health and wellness. Translational Andrology and Urology, 5(2), 213–219. doi:10.21037/tau.2016.03.03
[9] Hulsbæk, S., Bandholm, T., Ban, I., Foss, N. B., Jensen, J.-E. B., Kehlet, H., & Kristensen, M. T. (2021). Feasibility and preliminary effect of anabolic steroids in addition to strength training and nutritional supplement in rehabilitation of patients with hip fracture: a randomized controlled pilot trial (HIP-SAP1 trial). BMC Geriatrics, 21(1). doi:10.1186/s12877-021-02273-z
[10] Sodi, R., Fikri, R., Diver, M., Ranganath, L., & Vora, J. (2005). Testosterone replacement-induced hyperprolactinaemia: case report and review of the literature. Annals of Clinical Biochemistry, 42(2), 153–159. doi:10.1258/0004563053492784
[11] Segaloff, A., Weeth, J. B., Cuningham, M., & Meyer, K. K. (1964). Hormonal therapy in cancer of the breast.XXIII. Effect of 7α-methyl-19-nortestosterone acetate and testosterone propionate on clinical course and hormonal excretion. Cancer, 17(10), 1248–1253. doi:10.1002/1097-0142(196410)17:10<1248::aid-cncr2820171005>3.0.co;2-a
[12] Maeda Y, Nakanishi T, Ozawa K, Kijima Y, Nakayama I, Shoji T, Sasaoka T. Anabolic steroid-associated hypogonadism in male hemodialysis patients. Clin Nephrol. 1989 Oct;32(4):198-201. PMID: 2805460.
[13] De Las Heras, F., & Negro-vilar, A. (1979). Effect of Aromatizable Androgens and Estradiol on Prolactin Secretion in Prepuberal Male Rats. Archives of Andrology, 2(2), 135–139. doi:10.3109/01485017908987305
[14] Hannan, C.J., Friedl, K., Zold, A., Kettler, T., & Plymate, S. (1991). Psychological and serum homovanillic acid changes in men administered androgenic steroids. Psychoneuroendocrinology, 16(4), 335–343. doi:10.1016/0306-4530(91)90019-p
[15] Lee S, Lee YB, Choe SJ, Lee WS. Adverse Sexual Effects of Treatment with Finasteride or Dutasteride for Male Androgenetic Alopecia: A Systematic Review and Meta-analysis. Acta Derm Venereol. 2019 Jan 1;99(1):12-17. doi:10.2340/00015555-3035
[17] Meston, C. M., & McCall, K. M. (2005). Dopamine and Norepinephrine Responses to Film-Induced Sexual Arousal in Sexually Functional and Sexually Dysfunctional Women. Journal of Sex & Marital Therapy, 31(4), 303–317. doi:10.1080/00926230590950217
[18] Pfaus, J. G. (2009). Pathways of Sexual Desire. The Journal of Sexual Medicine, 6(6), 1506–1533. doi:10.1111/j.1743-6109.2009.01309.x
E’ solo di recente che si è iniziato a trattare in maniera più dettagliata dei potenziali effetti collaterali neuronali degli AAS. Infatti, ci si è sempre e solo concentrati sull’impatto che queste molecole possono avere, per esempio, sulla lipidemia ematica, sulla funzionalità epatica e renale , tralasciando tutte quelle alterazioni che possono emergere a livello comportamentale e plastico-cerebrale, che sono certamente da non sottovalutare. Trattai già dell’attività neurosteroidea, vi rimando quindi alla lettura di quell’articolo per ulteriori approfondimenti.
Vista la scarsa diffusioni di valide informazioni sulla attività neuronale degli AAS, un mito comune che circola nella comunità del bodybuilding e dei pazienti in TRT è che il Testosterone sia intrinsecamente neuroprotettivo e che sia in questo unico rispetto a tutti gli altri steroidi androgeni anabolizzanti.
Ma il motivo per cui il Testosterone è neuroprotettivo è semplicemente perché esso aromatizza ad un tasso tale da fornire una quantità di Estradiolo sufficiente a bilanciare l’Androgenicità esercitata dallo stesso Testosterone e dal DHT.
I dati ottenuti in modelli di roditori che utilizzano cellule corticali lo suggeriscono in modo molto convincente, mostrando come l’inibitore dell’aromatasi Anastrozolo (Arimidex) elimini completamente gli effetti neuroprotettivi del Testosterone [1].
AAS sintetici e Testosterone a confronto:
Sono stati pubblicati molti studi che suggeriscono quanto gli AAS sintetici siano peggiori rispetto al Testosterone per quanto riguarda le malattie cardiovascolari, la neurotossicità e una miriade di altri esiti organici deleteri.
Ritengo che una parte significativa di questi dati sia in parte estremizzata, data la mancanza di osservazioni di risposta seguenti alla somministrazione di estrogeni esogeni. In definitiva, il Testosterone è l’Androgeno più sicuro a dosaggi fisiologici. Il DHEA segue il grado favorevole di sicurezza, ma questo è un altro discorso.
Tuttavia, si ipotizza che molti AAS non siano così pericolosi come siamo portati a credere. Attenzione però, con questo non si intende assolutamente dire che essi abbiano un ottimale margine di sicurezza.
L’ipotesi è che alcune molecole non sono intrinsecamente più pericolose del Testosterone, ma è la loro mancanza di aromatizzazione, di 5-α riduzione o di diversa affinità per i recettori fuori bersaglio a renderli più pericolosi.
L’effetto sui Recettori degli Estrogeni (ER) e l’affinità dell’AAS per l’attività dell’Enzima Aromatasi sono i fattori principali che incidono sulla validità di un ormone in un contesto di monoterapia.
I derivati del DHT non possono essere convertiti dall’Enzima Aromatasi in un estrogeno come l’Estradiolo.
Il Nandrolone (19-nortestosterone) e i suoi derivati (19-nor) hanno ciascuno la propria affinità (o mancanza di affinità) per i ER e l’interazione con l’Aromatasi, che spesso si traduce in livelli di estrogeni inferiori alla media (esistono eccezioni come il Trestolone/MENT il quale si converte nel potente 7α-methyl-Estradiolo).
In sostanza, credo che alcuni AAS possano risultare significativamente più cardiotossici e neurotossici nei dati perché vengono sempre utilizzati da soli con una quantità insufficiente di estrogeni per bilanciare l’attività androgenica.
L’importanza di mantenere livelli ottimali di Estradiolo:
Livelli correttamente bilanciati di estrogeni sono necessari per la libido, la qualità dell’erezione, la vasodilatazione, la salute cardiovascolare, la salute del cervello, la salute delle ossa e molte altre funzioni sistemiche.
Nelle donne il rischio di malattie cardiovascolari aumenta notevolmente dopo la menopausa.
Non è una coincidenza che la maggior parte delle donne che sviluppano malattie cardiache lo facciano dopo che la produzione di estrogeni è scesa a livelli inferiori a quelli maschili.
Se non si dispone di una quantità sufficiente di estrogeni rispetto ai livelli di androgeni nell’organismo, i livelli di cardiotossicità e neurotossicità saranno significativamente più alti di quelli che si avrebbero se si mantenessero livelli ottimali di estrogeni.
Dal punto di vista del Bodybuilding, gli estrogeni sono necessari per ottimizzare la crescita muscolare, l’insulino-sensibilità e la sintesi di IGF-1 e fattori di crescita/segnalazione cellulare.
Per questo motivo gli AAS altamente aromatizzabili possono indirettamente determinare un maggiore potenziale di crescita e sono spesso classificati come composti “Bulking”.
Aneddoticamente, molti bodybuilder riferiscono di essere cresciuti al massimo durante l’Off-Season, quando i loro livelli di estrogeni erano sufficientemente alti.
La logica di inserimento di una “base” di Testosterone:
Il Testosterone non ha una selettività tissutale e, in realtà, è un modesto “costruttore muscolare” milligrammo per milligrammo rispetto ad altri AAS sintetici sviluppati negli anni successivi alla sua scoperta.
Per quanto riguarda la ritenzione di azoto, sulla carta non è superiore a molti AAS.
Tuttavia, il Testosterone aromatizza in Estradiolo a un ritmo strettamente regolato, è bioidentico e il corpo sa esattamente cosa fare con esso.
Inoltre, il corpo sa quanto Testosterone legare con le SHBG, quanto liberarne e mettere a disposizione dei tessuti, nonché quanto 5α ridurne a DHT per antagonizzare l’attivazione dei Recettori degli Estrogeni nel caso in cui questa vada fuori controllo.
Dal punto di vista del bodybuilding, il Testosterone è sottovalutato sotto molti aspetti.
Tuttavia, in un contesto di salute generale, longevità e bodybuilding, il Testosterone non può essere battuto a dosaggi terapeutici.
Utilizzando una base di Testosterone o una fonte di estrogeni sufficiente, le carenze di altri agenti anabolizzanti possono essere attenuate in una certa misura, motivo per cui il Testosterone è la base della maggior parte dei cicli di SARM steroidei e non steroidei.
Bilancio tra Testosterone, DHT ed Estradiolo:
La steroidogenesi nell’organismo si esplica come un’enorme orchestra volta alla regolazione di innumerevoli funzioni. Essa è molto più complessa della semplice sintesi di Testosterone, estrogeni e DHT.
La steroidogenesi umana, con le principali classi di ormoni steroidei, i singoli steroidi e le vie enzimatiche.[Häggström M, Richfield D (2014). “Diagram of the pathways of human steroidogenesis”. WikiJournal of Medicine.] I cambiamenti nella struttura molecolare da un precursore sono evidenziati in bianco.
Anche a livello acuto, l’equilibrio tra androgeni ed estrogeni nell’organismo è strettamente regolato e viene attuato per garantire una ottimizzazione della salute.
Questo equilibrio diventa sempre più disfunzionale con l’età, uno stile di vita scorretto, un’alimentazione scorretta, una cattiva igiene del sonno e numerosi altri fattori.
Tuttavia, come detto precedentemente, l’organismo sa esattamente cosa fare con il Testosterone, come creare una quantità ottimale di estrogeni e quanto Testosterone ridurre in DHT per contrastare l’eccesso di estrogeni e sostenere le caratteristiche sessuali secondarie maschili.
Quando si confrontano i dati clinici su un AAS sintetico con il Testosterone in un contesto di monoterapia, bisogna considerare che questi studi utilizzano l’AAS sintetico da solo, non con estrogeni esogeni o qualsiasi ormone supplementare che potrebbe essere necessario per bilanciare la sua androgenicità, la mancanza di attività estrogenica e/o l’interazione con l’Aromatasi.
Ovviamente, se si prende un composto che non aromatizza a sufficienza in estrogeni e lo si confronta con l’androgeno bioidentico che il nostro corpo sa aromatizzare e 5α ridurre in modo perfettamente bilanciato, si può solo immaginare quale sarà la scelta migliore data dall’osservazione comparativa in termini di cardiotossicità e neurotossicità.
La neurotossicità di Testosterone, Nandrolone e Stanozololo:
Confrontando l’effetto del Testosterone con quello del 19-nortestosterone (Nandrolone) e dello Stanozololo (Winstrol) sulla neurotossicità, si vede chiaramente che sono gli estrogeni a proteggere i neuroni del cervello, non il Testosterone in se.
Strutture molecolari di Testosterone, Nandrolone e Stanozololo
Nello studio del 2007 di Rosamaria Orlando et al., un dosaggio fisiologico di Testosterone risultava essere neuroprotettivo [1]. Il Testosterone amplificava la neurotossicità solo a dosaggi sovrafisiologici.
L’effetto neuroprotettivo di un dosaggio fisiologico di Testosterone è stato completamente eliminato quando è stato co-somministrato l’inibitore dell’Aromatasi Anastrozolo (Arimidex), suggerendo che la tossicità intrinseca del Testosterone come androgeno è controbilanciata solo dalla sua aromatizzazione in 17β-estradiolo.
Struttura molecolare di Estradiolo
A differenza del Testosterone, il Nandrolone non presenta un tasso di aromatizzazione sufficiente in termini assoluti (quantità) e specifici (tipo di estrogeno) mentre lo Stanozololo non subisce a nessun grado l’aromatizzazione.
Come prevedibile, il Nandrolone e lo Stanozololo sono risultati entrambi neurotossici a ogni singola dose valutata, indipendentemente dalla co-somministrazione o meno di Anastrozolo.
L’antiandrogeno Flutamide è stato in grado di attenuare la neurotossicità di tutti e tre gli androgeni, rafforzando così ulteriormente il fatto che dosaggi fisiologici di androgeni senza una quantità sufficiente di estrogeni a controbilanciarne gli effetti, o dosaggi sovrafisiologici di androgeni possono facilitare la morte neuronale.
Nessuno degli steroidi androgeni anabolizzanti di questo studio è risultato tossico in assenza di NMDA (recettore N-metil-D-aspartato), suggerendo quindi che il meccanismo attraverso il quale gli androgeni non controbilanciati facilitano la morte neuronale è una maggiore vulnerabilità agli insulti eccitotossici.
Rappresentazione grafica semplificata della struttura del recettore NMDA
Effetto neuroprotettivo del Testosterone a dosaggi fisiologici senza Anastrozolo:
A dosaggi fisiologici senza la presenza di un inibitore dell’Aromatasi, il Testosterone ha dimostrato di avere un effetto neuroprotettivo.
Spesso si ritiene che sia l’androgeno stesso (Testosterone) a proteggere il cervello. Tuttavia, l’inibitore dell’Aromatasi Anastrozolo ha eliminato completamente tutti gli effetti neuroprotettivi del Testosterone allo stesso dosaggio fisiologico.
L’Anastrozolo ha esacerbato la neurotossicità a ogni singolo dosaggio di Testosterone quando è stato co-somministrato.
Ciò suggerisce che il Testosterone non è un androgeno unico e con attività che neuroprotettiva maggiore rispetto a tutti gli altri AAS, ma che è la sua aromatizzazione in estrogeni a essere neuroprotettiva.
Neurotossicità del Testosterone a dosaggi soprafisiologici con e senza Anastrozolo:
A dosaggi sovrafisiologici il Testosterone ha dimostrato di esacerbare la neurotossicità. Sebbene la sua aromatizzazione in estrogeni prevenga comunque una quantità significativa di morte neuronale, possiamo vedere chiaramente che le concentrazioni sovrafisiologiche di Testosterone esacerbano la neurotossicità in ogni caso e che i livelli sovrafisiologici di estrogeni non forniscono un aumento dose-dipendente della neuroprotezione.
Quindi, i dati suggeriscono che le concentrazioni fisiologiche di Testosterone facilitano la neuroprotezione cerebrale attraverso l’aromatizzazione in estrogeni, ma c’è una soglia per questa neuroprotezione e le concentrazioni sovrafisiologiche non sono comunque neurologicamente salutari.
La neurotossicità del Nandrolone è indipendente dall’uso di Anastrozolo:
Il Nandrolone ha esacerbato la neurotossicità a tutti i dosaggi, indipendentemente dal fatto che sia stata valutata una concentrazione bassa o alta.
Inoltre, la co-somministrazione di Anastrozolo non ha avuto alcun impatto sulla neurotossicità del Nandrolone in questo modello, a qualsiasi dosaggio.
Ciò suggerisce che il Nandrolone non aromatizza in estrogeni ad un tasso sufficiente, né attiva i recettori degli estrogeni da solo ad un grado soddisfacente per fornire gli effetti neuroprotettivi di livelli sani di estrogeni.
Sarebbe probabilmente necessaria una fonte di estrogeni da co-somministrare con il Nandrolone per poterlo considerare una valida alternativa alla monoterapia in un contesto di HRT o di ciclo con un certo margine di “sicurezza”.
La neurotossicità dello Stanozololo è indipendente dall’uso di Anastrozolo:
Lo Stanozololo ha esacerbato la neurotossicità a tutti i dosaggi, indipendentemente dal fatto che si valutasse una concentrazione bassa o alta.
Inoltre, la co-somministrazione di Anastrozolo non ha avuto alcun impatto sulla neurotossicità dello Stanozololo in questo modello, a qualsiasi dosaggio.
Sappiamo già che lo Stanozololo non è soggetto ad aromatizzazione.
Anche in questo caso, i dati suggeriscono che sarebbe necessaria una fonte di estrogeni da co-somministrare con lo Stanozololo per poterlo considerare una valida alternativa alla monoterapia in un contesto di HRT o di ciclo con un certo margine di “sicurezza”.
Attenuazione della neurotossicità con co-somministrazione di antiandrogeni:
Gli androgeni non controbilanciati da una quantità sufficiente di estrogeni sono cardiotossici e neurotossici.
Per questo motivo la Flutamide (un anti-androgeno) è stata in grado di eliminare la neurotossicità del Nandrolone e dello Stanozololo.
Gli antiandrogeni hanno una risposta dose-dipendente proprio come gli AAS, quindi si verifica una competizione tra gli antiandrogeni e gli androgeni per il legame e l’attivazione del Recettore degli Androgeni (AR).
Gli antiandrogeni agiscono come antagonisti competitivi dell’AR o come steroidi sintetici di fortuna, anche se con un’androgenicità significativamente ridotta o pressoché assente.
In pratica, a seconda dell’antiandrogeno utilizzato, o enzimatico (vedi inibitori della 5α-reduttasi come la Finasteride) o competitivo (vedi Bicalutamide, Flutamide ecc…), essi agiranno riducendo l’attività degli androgeni a livello sistemico (orali) o topico (soluzione da applicare).
L’efficacia dell’antiandrogeno nell’inibire il legame degli androgeni con l’AR si basa sull’affinità di legame, sulla costante di legame, sull’emivita, sul dosaggio utilizzato e su una miriade di altri fattori.
La Flutamide è un antagonista selettivo del recettore degli androgeni non-steroideo, competitivo e “silenzioso” dell’AR. Si tratta di un antiandrogeno primitivo e di livello inferiore rispetto agli sviluppi più recenti della medicina, tuttavia è ancora efficace nell’impedire agli androgeni di legarsi ai recettori bersaglio specifici.
Per questo motivo la Flutamide è stata in grado di eliminare completamente la neurotossicità del Nandrolone e dello Stanozololo a tutti i dosaggi. Impedendo al Nandrolone e allo Stanozololo di legarsi ai recettori degli androgeni, essi non sono più in grado di innescare la trascrizione e, quindi, manifestare i loro effetti nei tessuti.
I dati relativi alla Flutamide e all’Anastrozolo rafforzano il fatto che il Nandrolone non converte in estrogeni ad un tasso sufficiente da fornire la neuroprotezione necessaria per evitare la morte neuronale. Con o senza inibitore dell’Aromatasi, il Nandrolone aggrava la neurotossicità allo stesso modo.
Senza un inibitore dell’Aromatasi ma con un antiandrogeno, la Neurotossicità del Nandrolone viene eliminata completamente. Ed è probabile che lo stesso valga anche per la cardiotossicità intrinseca del Nandrolone.
Quanto detto vale, prevedibilmente, anche per lo Stanozololo, che non è soggetto ad aromatizzazione.
È qui che tutti gli studi che dimostrano quanto il Nandrolone abbia un influenza negativa per il cuore e il cervello vengono messi in discussione, poiché gli esiti negativi riscontrati nei dati potrebbero non essere stati così drastici se fosse stata co-somministrata una fonte di estrogeni.
Lo stesso dosaggio di Flutamide non è stato in grado di compensare completamente la neurotossicità del Testosterone a dosaggi sovrafisiologici senza la presenza di Anastrozolo.
Quando i dosaggi di Testosterone superano le concentrazioni fisiologiche, la vulnerabilità alla neurotossicità e alla cardiotossicità sale vertiginosamente.
Una quantità eccessiva di qualsiasi cosa nell’organismo è dannosa, e il Testosterone non è esente da questo problema solo perché è l’ormone bioidentico che produciamo naturalmente e che è soggetto ad aromatizzazione in estrogeni.
L’aromatizzazione degli androgeni in estrogeni regola la neurotossicità:
Riflettendo sui dati con e senza inibitore dell’Aromatasi, possiamo vedere chiaramente che sono gli estrogeni ad esercitare la neuroprotezione, non il Testosterone.
Molti pensano erroneamente che il Testosterone sia un androgeno unico che si lega all’AR in qualche modo speciale per proteggere il cervello e che gli altri AAS lo danneggino. Ma i dati ci mostrano che le cose siano proprio così nette.
Infatti, i dati mostrino chiaramente che con la co-somministrazione di Anastrozolo l’effetto neuroprotettivo viene annullato, mentre senza Anastrozolo si ha un effetto neuroprotettivo.
Se si dispone di una quantità sufficiente di estrogeni per bilanciare gli androgeni nel corpo, si ottiene un livello stabile e ottimale di neuroprotezione, che si riflette nel modo in cui il nostro corpo regola l’aromatizzazione endogena degli androgeni.
Ma se si ha un livello sovrafisiologico di androgeni o si inibisce l’Aromatasi così da impedire la sintesi di una quantità di estrogeni necessari per svolgere le funzioni organiche, la neurotossicità aumenta indipendentemente dal fatto che si stia valutando il Testosterone e non un AAS sintetico non bioidentico. E tutto ciò si ricollega alla logica dell’utilizzo di una base di Testosterone durante il ciclo o di una fonte sufficiente di estrogeni esogeni in caso di carenza.
Questo rafforza anche il fatto che gli inibitori dell’aromatasi sono più che altro deleteri se usati senza una reale necessità.
Sarebbe opportuno fare tutto il possibile per evitare l’uso di inibitori dell’Aromatasi.
Se si ha bisogno di un inibitore dell’Aromatasi, è spesso probabile che ciò sia legato, per esempio, ad un dosaggio troppo alto di Testosterone (o altri AAS soggetti ad aromatizzazione), o ad una percentuale di grasso troppo alta (più grasso = più Aromatasi). Altre volte ciò può dipendere da un polimorfismo genetico che determina un metabolismo degli ormoni sessuali alterato o da una sovraespressione epigenetico-dipendente alla modificata omeostasi ormonale.
In definitiva, la probabilità che vi sia il bisogno di un AI per gestire una dose terapeutica di Testosterone, se i soprariportati punti sono ottimizzati, non è così probabile.
In un contesto diBodybuilding con dosaggi sovrafisiologici, ritengo inoltre che nella maggior parte dei casi (quindi non in tutti) l’uso di un AI solo per poter utilizzare una dose “troppo alta” di Testosterone sia una strategia sbagliata.
È opportuno assumere estrogeni esogeni per prevenire la neurotossicità e la cardiotossicità?
Il fatto che siano gli estrogeni a fornire protezione neurologica, e non il Testosterone, non significa assolutamente che si debba iniziare ad assumere pillole anticoncezionali come fossero caramelle. Gli estrogeni non controbilancianti nell’organismo sono a loro volta cancerogeni. C’è un motivo per cui i primi trattamenti per il cancro al seno sono stati SERM e gli AI. Inoltre, gli estrogeni non forniscono neuroprotezione in modo dipendente dalla dose.
C’è un punto di equilibrio per ogni cosa nell’organismo, e una quantità eccessiva di qualsiasi cosa può essere dannosa. Un pretrattamento di 4 giorni con basse concentrazioni 0,01 μM (10 nM) di 17β-estradiolo è stato sostanzialmente neuroprotettivo contro la tossicità NMDA.
Tuttavia, è possibile notare chiaramente dal grafico che non si è verificata una diminuzione dose-dipendente della neurotossicità.
La neuroprotezione è stata significativamente inferiore con 1μM di 17β-estradiolo rispetto al dosaggio molto più basso di 0,01 μM di 17β-estradiolo.
L’organismo ha un sistema strettamente regolato in cui è necessaria una certa quantità di estrogeni per le funzioni fisiologiche. Una quantità eccessiva di estrogeni senza una quantità sufficiente di androgeni può provocare lo sviluppo di tumori, ginecomastia e diversi altri problemi.
Troppo pochi estrogeni ed eccessivi androgeni possono causare malattie cardiovascolari, morte neuronale e altrettanti problemi.
Quindi, cosa concludere?
Quindi, se si utilizza un androgeno senza una quantità sufficiente di estrogeni opposti per bilanciarlo, non solo si inibisce la crescita muscolare e la perdita di grasso, ma si mette il corpo in uno stato di salute che si deteriora ancora più rapidamente di quello in cui si sarebbe già trovato semplicemente a causa dei livelli di androgeni sovrafisiologici.
Se si utilizza una dose di Testosterone da TRT, sarebbe meglio non inibire inutilmente l’Aromatasi.
Inoltre, se si utilizza un AAS che o non è un potente substrato per l’aromatasi o non è soggetto ad essa (e questo interessa tutti i SARM steroidei e non steroidei), sarebbe meglio aggiungere al ciclo una base di Testosterone o una fonte di estrogeni adeguata. La co-somministrazione di DHEA non è garante di una risultante estrogenica ematica adeguata per via di variabili enzimatiche legate alla conversione dell’androgeno surrenalico a Androstenedione e Estradiolo.
La cosa interessante da valutare sarebbe se tutti gli AAS precedentemente descritti dalla letteratura riportata come deleteri per il cuore e il cervello sarebbero ancora descritti come tali se un dosaggio adeguato di estrogeni esogeni venisse usato insieme ad essi negli studi corrispondenti.
Questo apre sicuramente nuove prospettive per potenziali alternative alla HRT.
Il BodyBuilding si differenzia dagli sport di prestazione perché il giorno della gara gli atleti vengono giudicati in base all’aspetto piuttosto che alle capacità atletiche. I bodybuilder posano sul palco dove vengono giudicati per la muscolatura, la definizione e la simmetria. Nel corso di una stagione, i bodybuilder attraversano tre fasi diverse: la fase di crescita muscolare (Off-Season), la dieta per la competizione (preparazione alla gara) e la gara stessa. La maggior parte della letteratura riguarda la fase di dieta pre-gara e la peak week.[1]
Tuttavia, la letteratura scientifica sulle raccomandazioni alimentari per i bodybuilder durante la Off-Season è carente. Si tratta di una lacuna importante, poiché la maggior parte della carriera di un bodybuilder si svolge in questa fase, in cui l’obiettivo è aumentare la massa muscolare riducendo al minimo l’aumento eccessivo della massa grassa. I bodybuilder sono noti per avere atteggiamenti rigidi nei confronti della selezione degli alimenti, della frequenza dei pasti, dei tempi di alimentazione e dell’integrazione [2]. Storicamente, le informazioni sull’alimentazione e l’integrazione sono state trasmesse dalle riviste di bodybuilding e dai concorrenti di successo, ma recentemente sono emerse più informazioni attraverso Internet e i forum [3,4]. Di conseguenza, molte delle strategie alimentari utilizzate dai bodybuilder non hanno un solido supporto scientifico e la letteratura scientifica dimostra che alcune di queste strategie, tra cui l’uso massiccio di farmaci, ma anche di integratori più in generale, possono essere ovviamente dannosi per la salute [5,6,7].
Poiché i bodybuilder trascorrono la maggior parte del loro tempo in Off-Season, è evidente la necessità di raccomandazioni nutrizionali e di supplementazione, sia OTC che PEDs, il più possibile “sicure” e basate sull’evidenza per questa popolazione. È stato inoltre dimostrato che alcuni bodybuilder, e non soltanto i concorrenti di alto livello nel bodybuilding “Natural”, potrebbero essere interessati a informazioni basate sull’evidenza [8]. Con il supporto della review realizzata e pubblicata da Juma Iraki et al. che tratta del Off-Season a livello alimentare e integrativo, lo scopo di questo articolo sarà quello di riportare quanto evidenziato dalla letteratura scientifica sugli argomenti relativi all’alimentazione e all’integrazione alimentare e supplementazione PEDs rilevanti per i bodybuilder nella Off-Season e di fornire raccomandazioni pratiche sull’assunzione di energia, macronutrienti, frequenza dei pasti, tempistica dei nutrienti, integratori alimentari e PEDs .
Transizione dalla dieta pre-gara/peak week alla dieta in Off-Season – Reverse Diet Vs. Recovery Diet:
Il primo step che il bodybuilder si trova davanti è la gestione del passaggio da una dieta ipocalorica ad una ipercalorica. Ed è in questo frangente che emergono due strategie simili all’apparenza ma in realtà diverse: la “Recovery Diet” e la “Reverse Diet”.
Ora, molto semplicemente, la “Recovery Diet” consiste in un graduale aumento calorico ma di consistenza tale che l’atleta esca dalla condizione di ipocalorica nel giro di due settimane circa. Con la “Reverse Diet”, invece, abbiamo sempre un graduale aumento calorico ma caratterizzato da una ridotta consistenza dello stesso (si parla di circa 100Kcal/die a settimana). In questo caso specifico, il bodybuilder rimarrebbe in ipocalorica per diverse settimane con possibile emersione di problemi psicofisici legati al protrarsi dello stato stressorio.
Quindi, con il termine “Recovery Diet” ci riferiamo ad uno schema alimentare avente l’obiettivo generale di RECUPERARE da un periodo di dieta cronica sperimentato durante la preparazione alla gara. La “Recovery Diet” incoraggia i bodybuilder a guadagnare il 5-10% del loro peso di gara nelle prime 4-8 settimane successive all’evento. Questo con l’intento di accelerare l’aumento di grasso corporeo e far rientrare il soggetto in un range di grasso corporeo “sano”, fisiologico, il prima possibile. In seguito, si consiglia agli atleti di rallentare il ritmo di aumento del peso e di mantenere un surplus controllato, con un aumento medio dello 0,5-1% del peso corporeo al mese passando pienamente nella Off-Season. Questo fino a quando non raggiungono un punto in cui un ulteriore aumento di peso è considerato improduttivo. Con il termine “Reverse Diet” ci si riferisce ad una strategia la quale può ancora essere attuata con discreti vantaggi per aiutare un agonista a recuperare dopo il contest. Tuttavia, se rispettata e seguita correttamente, piccoli aumenti di cibo di ~100 Kcal/die a settimana potrebbero comunque protrarre il deficit calorico del soggetto, prolungando così il periodo di dieta ipocalorica. Sebbene questa possa essere una strategia utile in alcune circostanze, ad esempio durante l’avvicinamento alla competizione, le modalità di applicazione non permettono un recupero di una bf salubre in tempi ottimali. È risaputo che un bodybuilder in condizioni di picco non è necessariamente al massimo della salute, e questo è in gran parte correlato al livello di grasso corporeo. Accettare un certo aumento di grasso avrà effetti positivi su tutti gli aspetti della Off-Season come le prestazioni in allenamento, i marcatori ormonali, la disponibilità di energia, la qualità del sonno e, inoltre, sarà vantaggioso sulla longevità complessiva dello sport praticato.
In definitiva, se si parte da body fat estremamente basse, tipiche da gara, allora la “Recovery Diet” è la scelta migliore per shiftare dal regime ipocalorico che ha caratterizzato il periodo di preparazione alla gara a quello ipercalorico del Off-Season. Discorso diverso se ci troviamo di fronte ad un soggetto amatoriale, con una body fat del 8-10% arrivato al termine del percorso di “Cut”. In questo caso la “Reverse Diet” è la scelta più funzionale permettendo un controllo migliore degli incrementi calorici evitando che la massa grassa sfori eccessivamente e che il lavoro precedentemente svolto in “Cut” venga facilmente e totalmente compromesso. Anche “ibridazioni” con aumenti settimanali di 45-50g di CHO die possono essere applicati con buoni risultati.
Energia:
Durante la Off-Season, l’obiettivo principale di un bodybuilder è quello di aumentare la massa muscolare riducendo al minimo l’aumento della massa grassa attraverso l’uso di allenamenti contro-resistenza e il mantenimento di un bilancio energetico positivo. Per valutare con precisione il fabbisogno energetico dei bodybuilder durante la bassa stagione, è necessario considerare il volume, la frequenza e l’intensità dell’allenamento. Durante la fase off-season, è stato riportato che i bodybuilder si allenano alla resistenza 5-6 volte a settimana, esercitando ogni gruppo muscolare 1-2 volte a settimana [9]. È stato inoltre riferito che seguono una routine di allenamento ad alto volume con 4-5 esercizi per gruppo muscolare, eseguendo 3-6 serie per esercizio, 7-12 ripetizioni massime (RM) per ogni serie con 1-2 minuti di riposo tra le serie. La durata della sessione di allenamento è stata indicata in ~40-90 minuti. Tuttavia, i piani di allenamento possono variare notevolmente da atleta ad atleta. È necessario valutare anche l’apporto calorico medio dei bodybuilder. Nella fase off-season, l’apporto energetico è di solito sostanzialmente più elevato rispetto alla fase di dieta: tra i bodybuilder maschi è stato riportato un apporto medio di ~3800 kcal/giorno durante la fase off-season e di ~2400 kcal/giorno durante la fase di dieta [2].
Bilancio energetico positivo:
È stato dimostrato che un bilancio energetico positivo ha un importante effetto anabolico, anche in assenza di allenamento contro-resistenza [10]. Tuttavia, la combinazione di un bilancio energetico positivo con l’allenamento contro-resistenza rappresenta il metodo più efficace per garantire che gli effetti anabolici siano diretti all’aumento della massa muscolo-scheletrica [11,12]. L’entità del surplus energetico ideale per guadagnare massa muscolare limitando l’accumulo di tessuto adiposo può variare in base allo stato di allenamento. Nei soggetti non allenati, è stato dimostrato che un surplus energetico sostanziale di circa 2.000 kcal, combinato con l’allenamento contro-resistenza, fornisce un robusto aumento di peso, in cui il contributo della massa magra (LBM) può raggiungere il 100% [12]. Tuttavia, nei soggetti allenati, un surplus energetico sostanziale potrebbe non essere necessario o vantaggioso. Uno studio condotto su atleti d’élite ha esaminato l’effetto delle indicazioni dietetiche sui cambiamenti della composizione corporea tra gli atleti d’élite quando l’allenamento contro-resistenza è stato combinato con diverse entità di surplus energetico. Un gruppo con un peso corporeo medio di 75kg ha consumato energia ad libitum (2964 kcal) per raggiungere un surplus molto ridotto, mentre un secondo gruppo con un peso corporeo medio di 71kg ha ricevuto una consulenza dietetica e ha consumato ~600 kcal in più rispetto al gruppo ad libitum [13].
Entrambi i gruppi hanno seguito lo stesso programma di allenamento contro-resistenza di 4 giorni alla settimana per un periodo di 8-12 settimane. I ricercatori hanno ipotizzato che il gruppo ipercalorico avrebbe avuto un aumento maggiore del peso corporeo e della LBM. Sebbene il gruppo ipercalorico abbia ottenuto un aumento maggiore della LBM rispetto a quelli che mangiavano ad libitum, questo non ha raggiunto la significatività statistica (1,7kg contro 1,2kg, rispettivamente). Inoltre, rispetto al gruppo che mangiava a sazietà, hanno registrato un aumento significativamente maggiore della massa grassa (1,1kg contro 0,2kg, rispettivamente). I ricercatori hanno concluso che un surplus di 200-300 kcal al giorno negli atleti altamente allenati potrebbe essere più appropriato di 500 kcal per minimizzare il rischio di inutili aumenti di grasso corporeo. I soggetti non allenati, più lontani dal loro tetto genetico di massa muscolare, possono essere in grado di aumentare i muscoli a un ritmo più veloce rispetto agli individui allenati.
Il tasso di crescita muscolare può rallentare con l’avanzare dell’età [14]. Pertanto, un maggiore surplus energetico può essere più vantaggioso per i bodybuilder alle prime armi, mentre i bodybuilder avanzati potrebbero trarre maggiore beneficio da diete ipercaloriche conservative per limitare inutili aumenti di grasso corporeo. Studi precedenti hanno raccomandato ai bodybuilder di consumare una dieta leggermente ipercalorica, con un aumento dell’apporto energetico di circa il 15% rispetto al mantenimento nella Off-Season [15]. Tuttavia, ciò non tiene conto della storia di allenamento e del livello di esperienza del singolo bodybuilder. Poiché la capacità di aumentare la massa muscolare è limitata, un surplus aggressivo può portare a un inutile aumento del grasso corporeo, che aumenterebbe la durata o la gravità dei successivi periodi di preparazione alle gare, aumentando di conseguenza la durata o la gravità della scarsa disponibilità energetica. Pertanto, il numero di calorie che un bodybuilder consuma al di sopra del livello di mantenimento può essere stabilito in base al livello di esperienza e poi regolato in base al tasso di aumento di peso e ai cambiamenti nella composizione corporea. Dato che i bodybuilder spesso aumentano rapidamente di peso dopo una gara, potrebbe essere utile avere un obiettivo di aumento di peso per settimana e regolarsi di conseguenza [16,17].
Tuttavia, come detto precedentemente, inizialmente, dopo la gara, potrebbe essere utile un aumento di peso più rapido per aiutare a riportare il concorrente a uno stato di salute sia psicologico che fisiologico, prima che il tasso di aumento di peso venga rallentato per limitare l’accumulo eccessivo di tessuto adiposo. Nella letteratura scientifica si raccomanda di puntare a un aumento di peso di circa 0,25-0,5 kg a settimana per cercare di aumentare la LBM e ridurre al minimo l’aumento della massa grassa [14,18]. Per un bodybuilder avanzato, un potenziale aumento di 2kg di peso corporeo su base mensile potrebbe essere eccessivo e comportare un’inutile accumulazione di grasso corporeo; pertanto, questo tasso dovrebbe essere considerato con cautela. Sulla base delle prove attuali, potrebbe essere opportuno raccomandare ai bodybuilder di consumare una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) nella Off-Season e raccomandare ai bodybuilder avanzati di puntare all’estremità inferiore di questa raccomandazione, o addirittura di essere più conservativi se si verificano aumenti sostanziali della massa grassa. Dato che i bodybuilder consumano in media 45 kcal/kg durante la bassa stagione, il surplus raccomandato equivale a circa 42-48 kcal/kg [2]. Potrebbe essere utile puntare a un aumento di peso di circa 0,25-0,5% del peso corporeo a settimana, regolando al contempo l’apporto energetico in base alle variazioni della composizione corporea. Inoltre, potrebbe essere più appropriato considerare le variazioni di peso medie settimanali basate su pesate giornaliere (o più volte alla settimana) per limitare gli errori delle fluttuazioni giornaliere del peso che possono verificarsi durante la settimana. Una volta determinato il surplus calorico, il passo successivo sarà quello di distribuire le calorie tra proteine, grassi e carboidrati.
Proteine:
Il turnover proteico del muscolo scheletrico è il rapporto tra la sintesi proteica muscolare (MPS) e la degradazione proteica muscolare (MPB). L’ipertrofia del muscolo scheletrico richiede un equilibrio netto in cui la MPS supera la MPB. L’esercizio contro-resistenza fornisce lo stimolo di tensione iniziale che induce l’ipertrofia risultante dall’aumento cumulativo della MPS dopo l’esercizio cronico [19]; tuttavia, l’aumento della massa grassa (FFM) può essere limitato se l’apporto proteico giornaliero è insufficiente [20]. Oltre alla quantità totale consumata al giorno, i ricercatori hanno ipotizzato che la qualità delle proteine possa aumentare il guadagno muscolare indotto dall’allenamento contro-resistenza [21]. Pertanto, entrambi questi argomenti saranno discussi nelle sezioni seguenti.
Introito proteico giornaliero:
Mentre l’attuale RDA per le proteine negli individui sani sedentari è di 0,8 g/kg, in una meta-analisi del 2018 di Morton e colleghi [22] è stato osservato che il doppio di questa quantità massimizza l’ipertrofia indotta dall’allenamento contro-resistenza. Inoltre, gli autori hanno osservato che “potrebbe essere prudente raccomandare ~2,2g di proteine/kg/die per coloro che cercano di massimizzare i guadagni di FFM indotti dall’allenamento contro-resistenza”, poiché 2,2g/kg era l’estremità superiore del limite di confidenza [22] e le differenze individuali impongono che alcuni atleti abbiano un fabbisogno proteico più elevato di altri [23]. Inoltre, la raccomandazione “meglio prevenire che curare” è probabilmente sicura, vista l’assenza di danni apparenti in studi di 1-2 anni tra i sollevatori che consumavano apporti proteici di almeno 2,2 g/kg [24,25]. Infine, la media e il limite superiore di confidenza del 95% per il fabbisogno proteico utilizzando la tecnica di ossidazione degli aminoacidi con indicatore tra i bodybuilder maschi nei giorni di non allenamento sono stati riportati rispettivamente come 1,7 e 2,2g/kg [26], che è simile al fabbisogno tra le donne quando è normalizzato alla FFM [27].
Tuttavia, è stato riportato che i bodybuilder consumano fino a 4,3g/kg di proteine al giorno tra i soggetti di sesso maschile e 2,8g/kg tra quelli di sesso femminile, superando di gran lunga queste raccomandazioni [2]. Le linee guida precedentemente fornite per i bodybuilder nella Off-Season erano di consumare il 25-30% del loro apporto energetico dalle proteine [15]. Potrebbe essere ragionevole opporsi all’indicazione di raccomandazioni basate su percentuali dell’apporto energetico totale, poiché un individuo con un peso non particolarmente elevato ma con un alto fabbisogno energetico potrebbe finire per consumare proteine che superano di gran lunga quelle necessarie e quindi richieste. Inoltre, questo può portare a un’assunzione insufficiente di carboidrati e grassi se l’atleta mira a un apporto calorico specifico. Pertanto, potrebbe essere più appropriato raccomandare un fabbisogno proteico basato sul peso corporeo. Pertanto, i bodybuilder dovrebbero consumare un minimo di 1,6g/kg di proteine nella Off-Season, anche se un obiettivo più vicino a 2,2 g/kg potrebbe garantire una risposta ottimizzata in modo più coerente in una maggiore percentuale di atleti.
E per i “Doped”? Dovremo ormai sapere che la fisiologia di base è la medesima per ogni individuo con le consuete variabili. Detto ciò, l’uso di PEDs va si ad alterare la fisiologia ma in questo specifico ambito, ossia introito proteico per massimizzare lo stimolo ipertrofico, hanno una azione di perfezionamento dell'”economia proteica cellulare”: in parole più semplici, sembra che l’uso di AAS porti ad una migliore resa nell’utilizzo degli amminoacidi scissi e assorbiti dalle proteine alimentari. Di conseguenza, a parità di apporto proteico, la veicolazione degli amminoacidi a scopo plastico è maggiore come minore è l’attività catabolica. Ciò significa che abusare delle proteine, in special modo durante una fase ipercalorica, perchè si è sotto AAS potrebbe risultare più inutile di quanto non lo sia in contesto “Natural”.
Infine, ed è necessario sottolinearlo, tra i bodybuilder che lottano con la fame in Off-Season e che di conseguenza assumono quantità caloriche che portano a un aumento di peso più rapido e all’accumulo di grasso in eccesso, un apporto proteico più elevato può essere utile (se non controindicato per motivi clinici). In uno studio condotto da Antonio e colleghi, i partecipanti ad allenamenti contro-resistenza che consumavano più proteine (4,4g/kg al giorno) e più calorie hanno guadagnato una quantità simile di FFM, ma non hanno guadagnato ulteriore grasso corporeo rispetto al gruppo che consumava meno proteine e meno calorie [28]. Allo stesso modo, in uno studio di follow-up, un gruppo che consumava 3,4g/kg di proteine al giorno ha guadagnato una quantità simile di FFM, ma ha perso una percentuale maggiore di grasso corporeo rispetto a un gruppo a basso contenuto proteico, ancora una volta, nonostante un apporto energetico più elevato [29]. Gli autori di questi studi sulla “vita libera” hanno ipotizzato che i loro risultati fossero dovuti a un aumento della termogenesi indotta dalla dieta attraverso protocolli alimentari ad alto contenuto proteico. Tuttavia, ciò è in contrasto con uno studio di Bray e colleghi del 2012 sul reparto metabolico, più strettamente controllato, in cui il contenuto proteico della dieta influenzava la percentuale di massa corporea acquisita, mentre la massa corporea totale era dettata dal solo contenuto energetico della dieta [30].
Pertanto, mentre la termogenesi indotta dalla dieta potrebbe essere significativamente più elevata con assunzioni di proteine nell’intervallo di 3 g/kg o superiore, la perdita di grasso o la mancanza di aumento di peso osservata da Antonio e colleghi, nonostante un apporto energetico più elevato, potrebbe con più probabilità riflettere l’effetto saziante di assunzioni proteiche molto elevate che diminuiscono l’assunzione calorica effettiva, piuttosto che un aumento della sola termogenesi.
Qualità delle Proteine:
Gli aminoacidi essenziali (EAA) sono gli unici aminoacidi necessari per stimolare il processo di MPS [31]. Sebbene tutti gli aminoacidi forniscano i “mattoni” necessari per la sintesi di nuovi tessuti, l’aminoacido Leucina in particolare sembra essere particolarmente importante come “innesco metabolico” della MPS [32]. È stato suggerito che una concentrazione sufficiente di Leucina è necessaria per raggiungere una “soglia di Leucina” che è richiesta per stimolare al massimo la MPS [33]. In breve, dal punto di vista della costruzione muscolare, le fonti proteiche che innescano una consistente risposta della MPS (quantità sufficiente di Leucina) e forniscono i mattoni essenziali per la costruzione di nuovo tessuto muscolare (contengono l’intero spettro di aminoacidi essenziali in abbondanza) possono essere considerate di “qualità superiore”.
Sebbene l’effetto meccanicistico della Leucina sulle MPS esuli dallo scopo di questo articolo, si invitano i lettori a leggere una rassegna che tratta questo argomento in dettaglio [34]. In generale, su una base di grammo per grammo, le fonti proteiche di origine animale contengono in genere più Leucina ed EAA, anche se ci sono eccezioni degne di nota. Le proteine della soia, uno dei più comuni integratori proteici di origine vegetale, contengono tutti gli EAA, ma in una quantità inferiore per grammo rispetto alle proteine del latte e quindi, in uno studio, hanno prodotto un aumento minore delle MPS rispetto al siero di latte dopo un’ingestione acuta [35]. È interessante notare che in questo stesso studio la soia ha prodotto un aumento maggiore delle MPS rispetto alla caseina, anch’essa una proteina casearia di “alta qualità”, presumibilmente a causa della più lenta velocità di digestione della caseina [35]. Rammentate sempre la differenza tra risposta “acuta” e “cronica”. Per l’appunto, ciò significa che, sebbene il contenuto di Leucina e di EAA di una fonte proteica debba essere preso in considerazione, la risposta acuta alla MPS non è l’unica variabile legata all’ipertrofia a lungo termine. Infatti, una proteina di alta qualità ma “lenta” come la caseina produce inizialmente una risposta MPS di minore ampiezza. Tuttavia, la caseina (e altre proteine a lenta digestione) può produrre un’area MPS sotto la curva simile o maggiore se osservata longitudinalmente rispetto a una fonte proteica “veloce” come il siero di latte, che determina un aumento iniziale maggiore e poi una brusca riduzione [36].
Inoltre, la risposta acuta della MPS a un determinato tipo di proteina non deve essere vista in una prospettiva riduzionista. Nel mondo reale si consumano quotidianamente più porzioni di varie fonti proteiche, rendendo probabilmente superflue alcune di queste distinzioni nel profilo aminoacidico e nella cinetica di digestione. Infatti, in una meta-analisi che ha confrontato i cambiamenti longitudinali della composizione corporea con diversi tipi di integratori proteici, non sono state riscontrate differenze significative tra i partecipanti che consumavano soia rispetto al siero di latte, ad altre proteine del latte o alle proteine isolate del manzo [37].
Come dimostrato in uno studio che ha messo a confronto gruppi che consumavano proteine dopo l’allenamento (in aggiunta a una dieta già composta dal 25% di proteine), sia che venissero forniti 48g di proteine del siero del latte (contenenti 5,5g di Leucina), sia che venissero forniti 48g di proteine del riso (contenenti 3,8g di Leucina), non è stato osservato alcun impatto sui cambiamenti della composizione corporea tra i gruppi dopo otto settimane [38]. Pertanto, se consumate in quantità sufficienti (soprattutto se si considera l’apporto proteico totale giornaliero), la qualità delle proteine di un singolo pasto è meno preoccupante. Tuttavia, se si volesse consumare una dieta dominata da fonti proteiche di origine vegetale, esistono alternative alla soia e al riso. Ad esempio, le proteine isolate del pisello sono ricche di EAA e di Leucina. In uno studio di 12 settimane, un gruppo che consumava 50g di proteine isolate di pisello al giorno ha registrato un aumento maggiore dello spessore muscolare indotto dall’allenamento di resistenza rispetto al placebo, non significativamente diverso da un gruppo che consumava 50g di siero di latte [39].
Pertanto, nel contesto delle indicazioni di questo articolo, la qualità delle proteine può essere un problema solo se si utilizza la fascia bassa delle linee guida sulle proteine (1,6g/kg) o se si consuma una dieta a base prevalentemente vegetale. In entrambi i casi, potrebbe essere utile integrare con fonti proteiche ricche di Leucina e di EAA, a seconda delle preferenze alimentari (ad esempio, proteine del latte o del pisello se si è vegani), per garantire la risposta attesa della MPS all’assunzione di proteine.
Grassi:
Il grasso è un nutriente fondamentale per molte funzioni dell’organismo. Tuttavia, non si sa molto dell’effetto dei grassi alimentari sull’ipertrofia del muscolo scheletrico. È stato riportato che l’assunzione di grassi alimentari tra i bodybuilder varia dall’8 al 33% delle calorie totali [2]. Sebbene i trigliceridi intramuscolari possano fungere da substrato energetico durante l’allenamento di resistenza, non sono un fattore limitante poiché i substrati derivano principalmente da processi anaerobici [40]. Di interesse per il bodybuilder, è dimostrato che negli atleti allenati contro-resistenza [41] e nei giocatori di hockey [42] le diete a basso contenuto di carboidrati (30-45% dell’energia o meno) possono influire sul rapporto Testosterone libero/Cortisolo (fTC), il che potrebbe avere un impatto negativo sul recupero. D’altra parte, la riduzione dei grassi alimentari nelle diete isocaloriche da ~30-40% a ~15-25% ha portato a riduzioni significative ma modeste dei livelli di Testosterone [43,44,45,46].
Tuttavia, non è chiaro se le variazioni di Testosterone all’interno di intervalli normali influenzino in modo significativo l’aumento della massa muscolare [47]. Nonostante la possibilità che i livelli di testosterone possano essere più elevati quando si consuma una percentuale maggiore di energia proveniente dai grassi alimentari, i cambiamenti effettivi nella massa muscolare durante gli studi longitudinali di individui allenati alla resistenza che seguono diete “chetogeniche” ad alto contenuto di grassi sono stati costantemente inferiori rispetto ad approcci moderati o a basso contenuto di grassi con ampi carboidrati [48,49,50,51]. Non è ancora stato chiarito se ciò sia dovuto a cambiamenti nella capacità di esercizio, ad alterazioni del rapporto fTC o a qualche altro meccanismo legato alla componente ad alto contenuto di grassi o a basso contenuto di carboidrati della dieta.
Tuttavia, ciò indica che forse si dovrebbe consumare una proporzione più moderata di grassi nella dieta, piuttosto che un apporto basso o alto. In letteratura sono state proposte raccomandazioni del 15-20% e del 20-30% delle calorie provenienti dai grassi alimentari [15,52]. Tuttavia, sono necessarie ulteriori ricerche per stabilire l’effetto e la quantità ottimale di grassi alimentari per favorire l’ipertrofia muscolare.
Sulla base delle evidenze attuali, può essere prudente raccomandare che i grassi alimentari rappresentino il 20-35% delle calorie, in linea con le raccomandazioni dell’American College of Sports Medicine per gli atleti [53], che nella maggior parte dei casi corrispondono a circa 0,5-1,5 g/kg/giorno. Inoltre, va notato che un apporto sufficiente di proteine e carboidrati non deve essere compromesso da un’elevata assunzione di grassi nella dieta.
Anche la qualità dei grassi, come gli essenziali omega 3 e gli omega 6, potrebbe essere importante per i bodybuilder. Se l’apporto di questi acidi grassi è sufficiente, non è necessario integrarli con una dieta di alta qualità contenente buone fonti di acidi grassi. Tuttavia, per alcuni potrebbe essere difficile assumere le quantità ottimali. Per questo motivo, l’argomento verrà trattato in modo più approfondito nella sezione dedicata agli integratori alimentari.
Carboidrati:
A differenza delle proteine e dei grassi, i carboidrati sono considerati non essenziali per la dieta umana perché l’organismo è in grado di produrre il glucosio necessario ai tessuti attraverso la gluconeogenesi [54]. Tuttavia, l’assunzione di carboidrati ha un ruolo importante nella dieta del bodybuilder come regolatore degli ormoni tiroidei e come contributo al fabbisogno di micronutrienti [55,56]. Inoltre, una dieta a basso contenuto di carboidrati potrebbe limitare la rigenerazione dell’adenosina trifosfato (ATP) e limitare la capacità dei muscoli di contrarsi con una forza elevata [57,58]. Durante l’esercizio ad alta intensità, il glicogeno muscolare è il principale contributore di substrato energetico ed è stato dimostrato che la glicolisi fornisce circa l’80% del fabbisogno di ATP di una serie di flessioni del gomito se portata al cedimento muscolare [59]. Nonostante ciò, parte del glicogeno utilizzato durante questo tipo di esercizio può essere risintetizzato dal lattato, il che potrebbe ridurre il fabbisogno di carboidrati. È stato inoltre dimostrato che l’allenamento contro-resistenza riduce il glicogeno muscolare del 24-40% in una singola sessione [59,60].
La quantità esaurita può variare in base alla durata, all’intensità e al lavoro svolto, ma l’allenamento tipico del bodybuilding con ripetizioni più elevate e carichi moderati sembra causare la maggiore riduzione delle scorte di glicogeno muscolare [61]. Inoltre, è stato suggerito che quando le scorte di glicogeno sono troppo basse (~70 mmol/kg), ciò può inibire il rilascio di calcio e accelerare l’insorgenza della fatica muscolare [62]. Un basso livello di glicogeno muscolare riduce significativamente il numero di ripetizioni eseguite quando si eseguono tre serie di Squat all’80% di 1RM [57].
Tuttavia, è stato dimostrato che il consumo di una dieta contenente 7,7 g/kg/die di carboidrati per 48 ore prima di una sessione di allenamento non ha un effetto maggiore sulle prestazioni rispetto a 0,37g/kg/die quando si eseguono 15 serie a 15RM di esercizi per la parte inferiore del corpo [63]. Analogamente, un altro studio ha rilevato che una dieta con il 70% di carboidrati rispetto a una dieta con il 50% di carboidrati non ha un effetto maggiore sulle prestazioni durante l’esercizio sopramassimale; tuttavia, una dieta composta dal 25% di carboidrati ha ridotto significativamente le prestazioni [64].
Inoltre, visti gli effetti negativi a lungo termine sulla massa muscolare osservati di recente in studi su popolazioni allenate alla resistenza che seguono diete chetogeniche [49,51], potrebbe essere prudente per i bodybuilder assicurarsi semplicemente un apporto sufficiente di carboidrati, visti questi risultati disparati. Pertanto, mentre le diete a moderato e alto contenuto di carboidrati sono probabilmente appropriate per il bodybuilding, le diete a bassissimo contenuto di carboidrati possono essere dannose per l’allenamento.
Nei bodybuilder maschi, sono stati riportati apporti medi di carboidrati pari a 5,3g/kg/giorno durante la Off-Season [2]. Tuttavia, non sono state stabilite le quantità ottimali di carboidrati per i bodybuilder. In letteratura sono state proposte raccomandazioni per gli sport di forza, tra cui il bodybuilding, con assunzioni di 4-7g/kg/giorno e 5-6g/kg [15,65]. I carboidrati sembrano essere importanti per il bodybuilder, ma per ottenere benefici possono essere necessarie solo quantità moderate. Pertanto, dopo aver destinato le calorie alle proteine (1,6-2,2g/kg/die) e ai grassi (0,5-1,5g/kg/die), le restanti calorie dovrebbero essere destinate ai carboidrati. Tuttavia, sulla base delle prove attuali, potrebbe essere ragionevole consumare quantità sufficienti di carboidrati nell’intervallo ≥3-5g/kg/giorno, se possibile.
Sono necessarie ulteriori ricerche tra i bodybuilder per stabilire se l’assunzione abituale di carboidrati, superiore o inferiore a quella osservata, possa produrre ulteriori benefici. La Tabella sottostante riassume le raccomandazioni per le calorie e i macronutrienti.
Raccomandazioni dietetiche per i bodybuilder in Off-Season.
Distribuzione e timing dei nutrienti:
Si dice che i bodybuilder consumino in media sei pasti al giorno [66]; tuttavia, non esistono studi che esaminino specificamente quale possa essere la frequenza ottimale dei pasti per questa popolazione [65]. Questa elevata frequenza dei pasti si basa sulla convinzione di un maggiore stato di anabolismo e persino di un migliore utilizzo dei nutrienti durante il giorno, che potrebbe tradursi in un miglioramento della composizione corporea.
Il concetto di temporizzazione dell’assunzione di proteine per massimizzare l’ipertrofia comprende diverse strategie di dosaggio. La prima a comparire in letteratura è stata il consumo di proteine in prossimità dell’allenamento contro-resistenza. I picchi di MPS sono più elevati in questo periodo quando si consumano proteine; pertanto, questa strategia è stata proposta per migliorare l’efficienza della riparazione e del rimodellamento del muscolo scheletrico [31]. Inoltre, a causa dell'”effetto muscolo pieno”, per cui un ulteriore apporto di proteine non aumenta la MPS finché non è trascorso un tempo sufficiente, distribuire uniformemente l’assunzione di proteine tra più pasti è un’altra strategia studiata per massimizzare la MPS totale giornaliera [67]. Infine, il consumo prima di andare a letto di proteine a lenta digestione (come la caseina) per evitare periodi catabolici prolungati durante il sonno è la strategia proposta più di recente per migliorare il bilancio proteico netto giornaliero [68], sebbene si sia dimostrata inutile nel perseguire il fine o, per lo meno, non molto diversa dalla risultante di una assunzione di isolate in un contesto alimentare con parità nel totale proteico giornaliero. Ciascuna di queste tre strategie sarà discussa in seguito.
Dosaggio proteico:
Il periodo post-allenamento consente un picco della MPS più elevato quando si consumano proteine [31] e per raggiungere il picco di MPS può essere necessaria un’adeguata dose di Leucina “soglia” [32]. Diversi studi hanno esaminato il dosaggio proteico necessario per massimizzare la MPS dopo l’allenamento [69,70,71]. In uno studio sono stati consumati 0, 5, 10, 20 o 40g di proteine d’uovo intere dopo l’esercizio contro-resistenza della parte inferiore del corpo, con 20g che stimolavano al massimo la MPS [69]. Risultati simili sono stati riscontrati anche in un altro studio, in cui 20 g di siero di latte sono stati sufficienti a stimolare al massimo i tassi post-assorbitivi di MPS sia a riposo che dopo un lavoro unilaterale delle gambe all’80% del 1RM [70]. Inoltre, 40g di siero di latte non hanno prodotto ulteriori aumenti di MPS in questo studio e hanno portato all’ossidazione amminoacidica e alla produzione di urea.
Tuttavia, uno studio recente ha rilevato che, durante l’esecuzione di esercizi contro-resistenza per tutto il corpo al 75% del 1RM, 40g di siero di latte hanno prodotto una risposta MPS significativamente più elevata rispetto a 20g [71]. Esiste quindi una relazione tra il volume di tessuto muscolare danneggiato e stimolato e l’assunzione adeguata di proteine. È interessante notare che gli autori di una meta-analisi del 2013 hanno osservato che, nonostante gli studi con traccianti a breve termine mostrassero risposte nella MPS maggiori quando le proteine venivano consumate nella “finestra anabolica” post-allenamento, negli studi longitudinali sull’allenamento non è stato riscontrato alcun effetto significativo sull’ipertrofia quando si controllava l’apporto proteico totale giornaliero, indipendentemente dal fatto che le proteine fossero consumate all’interno della “finestra anabolica” o al di fuori di essa [72].
Nutrient Timing:
Analogamente, i ricercatori di uno studio tracciante a breve termine che ha esaminato il dosaggio delle proteine nel corso di 12 ore hanno riportato una maggiore area sotto la curva della MPS quando sono state consumate quattro dosi di proteine del siero di latte da 20g ogni tre ore rispetto a due dosi da 40g a distanza di sei ore e otto dosi da 10g ogni ora e mezza [73]. In teoria, data la soglia oltre la quale le proteine supplementari consumate in una singola seduta non contribuiscono ulteriormente alla MPS [69] e a causa del “periodo refrattario” postprandiale durante il quale la MPS non può essere nuovamente stimolata al massimo [67], si potrebbe concludere che un bodybuilder dovrebbe raggiungere, ma non superare, questa dose soglia ogni poche ore per massimizzare l’ipertrofia a lungo termine. Tuttavia, gli autori di una review sistematica del 2018 sugli integratori proteici, comprendente 34 studi randomizzati e controllati, hanno riportato guadagni di massa magra simili tra i gruppi che utilizzavano un programma di dosaggio con i pasti (che comportava un minor numero di dosi di proteine di entità elevata) e tra i pasti (che comportava un maggior numero di dosi di proteine di entità moderata) [74].
È interessante notare che i dati che esaminano l’alimentazione proteica notturna mostrano uno distacco simile tra gli studi meccanicistici a breve termine e gli interventi di allenamento a lungo termine. Nel 2012 è stata condotta la prima ricerca che esaminava la risposta acuta all’alimentazione notturna con caseina [68]. Gli autori hanno riportato che 40g di caseina consumati prima di andare a letto sono stati digeriti, assorbiti e hanno stimolato la MPS e migliorato l’equilibrio proteico dell’intero corpo durante il periodo notturno in misura maggiore rispetto al placebo. Negli anni successivi sono stati pubblicati altri studi in acuto che hanno confermato [75] e riconfermato questi risultati in una popolazione più anziana [76]. Nel 2015, gli autori del primo studio longitudinale hanno riportato un aumento della forza e dell’ipertrofia in un gruppo a cui era stato somministrato un supplemento proteico notturno rispetto a un gruppo placebo [77].
Tuttavia, la quantità totale di proteine giornaliere non è stata equiparata, in quanto il gruppo con proteine notturne ha consumato 1,9g/kg/giorno, mentre il gruppo placebo ha consumato solo 1,3g/kg. È importante notare che in entrambi gli unici studi longitudinali con corrispondenza proteica che hanno confrontato l’integrazione notturna di caseina con i gruppi che hanno assunto l’integrazione prima, non sono state riportate differenze significative nell’aumento della FFM tra i gruppi [78,79]. Pertanto, la domanda è la stessa per ogni strategia di distribuzione: perché ci sono ripetuti distacchi tra gli studi meccanicistici a breve termine sulle MPS e le ricerche a lungo termine che esaminano l’effettiva ipertrofia? La risposta potrebbe risiedere nei metodi utilizzati negli studi sulla MPS, in quanto i partecipanti sono a digiuno, ricevono solo proteine in polvere in isolamento, spesso viene loro somministrato del siero di latte (che viene digerito molto rapidamente) e vengono osservati per brevi periodi. Questi contesti di laboratorio determinano tempi di digestione e cinetiche degli aminoacidi diversi da quelli che si verificano nel “mondo reale”. In particolare, in queste condizioni di laboratorio i livelli di base degli aminoacidi nel corpo sono più bassi del normale e la digestione e il successivo apporto di aminoacidi al muscolo sono più rapidi.
In condizioni di vita libera, le proteine vengono consumate principalmente da fonti alimentari intere, più volte al giorno e insieme ad altri alimenti, il che ritarda lo svuotamento gastrico. Per questi motivi, gli aminoacidi vengono titolati nel flusso sanguigno in modo più lento e costante; pertanto, in condizioni normali, le scorte sono quasi sempre prontamente disponibili [80]. Pertanto, l’efficacia della “finestra anabolica” e persino delle strategie di distribuzione delle proteine potrebbe non tradursi nella pratica. Inoltre, le limitazioni specifiche del laboratorio si estendono anche agli studi sull’alimentazione notturna. Si consideri, ad esempio, che 26g di proteine provenienti da una bistecca magra determinano un aumento sostenuto della MPS che dura almeno sei ore (l’intero periodo di tempo studiato) [81].
Inoltre, 26g sono solo il ~37% della dose di proteine contenuta in media in una cena americana [82], che richiederebbe più tempo per essere digerita a causa della maggiore porzione di proteine e dell’aggiunta di fibre, lipidi e altri nutrienti che ritarderebbero ulteriormente la digestione [80]. Pertanto, il tipico pasto finale potrebbe già soddisfare lo scopo di un frullato di caseina. Detto questo, nonostante queste discrepanze tra MPS e risultati della composizione corporea, non c’è nulla di male nel tentare queste strategie, soprattutto se attuate in modo pragmatico e senza introdurre ulteriori oneri logistici nel proprio programma quotidiano.
Pertanto, potrebbe essere prudente consigliare ai bodybuilder di suddividere l’assunzione giornaliera di 1,6-2,2 g/kg di proteine in più pasti contenenti ciascuno ~0,40-0,55g/kg [80] e di fare in modo che uno di questi pasti avvenga entro 1-2 ore prima o dopo l’allenamento, mentre un’alimentazione costituita da una fonte proteica e non proteica venga consumata 1-2 ore prima di dormire. Ad esempio, un bodybuilder di 90 kg potrebbe consumare 40-50g di proteine alle 8-9 del mattino per la colazione, allenarsi alle 11, consumare 40-50g di proteine alle 12-13 per il pranzo/post-allenamento, 40-50g di proteine a cena tra le 17-18, e poi un pasto finale di 40-50g di proteine non contenenti fonti proteiche grasse alle 21-10 prima di andare a letto entro le 23.
I carboidrati consumati prima dell’allenamento sono spesso una strategia utilizzata dagli atleti per migliorare le prestazioni negli esercizi ad alta intensità. La completa risintesi del glicogeno può essere raggiunta entro 24 ore da un allenamento che depaupera il glicogeno se si consumano quantità sufficienti di carboidrati [83]. Tuttavia, solo il 24-40% del glicogeno muscolare viene esaurito dopo un allenamento contro-resistenza [59,60]. Pertanto, una quantità di ≥3-5g/kg di carboidrati al giorno sarebbe probabilmente sufficiente per la risintesi del glicogeno. Questo elevato apporto giornaliero di carboidrati probabilmente riduce anche l’impatto della tempistica dei carboidrati pre-allenamento sulle prestazioni dell’esercizio.
Inoltre, per i bodybuilder che non hanno bisogno di enfatizzare il rifornimento di glicogeno, le proteine aumentano la MPS post-allenamento a livelli massimi anche senza l’aggiunta di carboidrati [86,87]. Anche se il consumo di carboidrati nel post-allenamento non è certo dannoso, è improbabile che questo favorisca l’ipertrofia a lungo termine, come discusso in precedenti review [1,88]. Pertanto, è meglio concentrarsi sul consumo di un’adeguata quantità di carboidrati giornalieri e basare la distribuzione dei carboidrati intorno all’allenamento sulle preferenze personali.
Supplementazione OTC:
In un recente sondaggio condotto tra i bodybuilder, è stato riportato che tutti i partecipanti assumevano integratori alimentari [9]. Gli integratori alimentari più comuni erano: integratori di proteine (86%), creatina (68%), aminoacidi a catena ramificata (67%), glutammina (42%), vitamine (40%), olio di pesce (37%) e prodotti contenenti caffeina/efedrina (24%).
Sebbene gli integratori proteici siano molto popolari tra i bodybuilder, vengono utilizzati prevalentemente come gli alimenti interi per raggiungere gli obiettivi proteici. Pertanto, non verranno discussi in dettaglio. I lettori sono invitati a leggere la posizione dell’ISSN su questo argomento [89]. Inoltre, la trattazione di tutti gli integratori comunemente utilizzati dai bodybuilder esula dallo scopo di questo articolo. L’attenzione si concentrerà piuttosto sugli integratori alimentari che potrebbero potenzialmente produrre un effetto ergogenico e sugli integratori che possono garantire un apporto sufficiente di micronutrienti e acidi grassi essenziali.
Creatina Monoidrato:
La Creatin-fosfato si trova in alte concentrazioni nel muscolo scheletrico e cardiaco, dove agisce come fonte di energia [90]. La Creatina può essere ottenuta anche attraverso la dieta nei soggetti che consumano carne; tuttavia, le concentrazioni di Creatina nella carne si riducono con la cottura [91].
Numerosi studi hanno osservato un aumento della massa e della forza muscolare in seguito a fasi di carico di Creatina, in genere di 20g al giorno per circa una settimana, spesso seguite da fasi di mantenimento di 2-3g di Creatina al giorno [92]. Tuttavia, la fase di carico potrebbe non essere necessaria. È stato dimostrato che la saturazione della Creatina muscolare dopo un’integrazione di 3g di Creatina Monoidrato per 28 giorni è simile al consumo di Creatina Monoidrato dopo la tipica fase di carico [93].
La maggior parte degli individui non raggiunge i 3g giornalieri con la dieta e può essere necessaria un’integrazione. Esistono numerose forme di Creatina negli integratori in commercio, tra le quali la Creatina Monoidrato è la più studiata. Le versioni più recenti di Creatina, come la kre-alkalyn [94] e la Creatina etil-estere [95], non si sono dimostrate superiori alla Creatina Monoidrato, nonostante abbiano in genere un prezzo più elevato. Pertanto, si raccomanda il consumo di 3-5g di Creatina Monoidrato al giorno. La tempistica di assunzione della Creatina non sembra avere importanza, poiché la saturazione delle riserve di Creatin-fosfato richiede circa 28 giorni per raggiungere le concentrazioni massime quando si consumano 3g al giorno e non ha un effetto in acuto [93].
Caffeina:
Uno degli integratori alimentari più utilizzati dai bodybuilder sono gli stimolanti, in particolare la Caffeina [9]. Oltre ad aumentare l’eccitazione [96], la Caffeina può ridurre il dolore e lo sforzo percepito durante l’esercizio [97] e migliora la gestione del Calcio, aumentando la potenza [98]. Studi sull’allenamento contro-resistenza hanno rilevato che la Caffeina riduce la fatica e aumenta la forza [99,100]. Tuttavia, non tutti gli studi hanno dimostrato un effetto ergogenico sull’allenamento contro-resistenza [101]. Gli studi che hanno dimostrato un effetto ergogenico hanno utilizzato dosaggi elevati di caffeina (5-6 mg/kg), che sono al limite superiore di quello che è considerato un dosaggio sicuro [99,100]. Tuttavia, può essere consigliabile consumare il dosaggio minimo efficace per individuo, poiché l’assunzione regolare può generare tolleranza [102]. A causa dell’effetto acuto della Caffeina, è consigliabile assumerla circa 1 ora prima dell’esercizio fisico [99]. Tuttavia, l’emivita della Caffeina è di circa 3-9 ore; pertanto, può essere consigliabile consumare la Caffeina all’inizio della giornata per favorire un sonno sano se l’esercizio fisico viene svolto più tardi nel corso della giornata [103]. Sono necessarie ulteriori ricerche per trovare un consenso sull’uso della Caffeina nell’allenamento contro-resistenza, ma sulla base delle prove attuali un dosaggio di 5-6 mg/kg consumato prima dell’esercizio potrebbe produrre un effetto ergogenico sulle prestazioni nell’allenamento contro-resistenza.
Beta-Alanina:
È stato dimostrato che l’ingestione di 4-6 g di beta-alanina aumenta i livelli di carnosina muscolare [104]. La carnosina agisce come tampone del pH nel muscolo scheletrico e può ritardare l’inizio dell’affaticamento muscolare durante l’esercizio ad alta intensità [105]. Una meta-analisi ha concluso che la beta-alanina potrebbe produrre effetti ergogenici durante l’esercizio ad alta intensità della durata di 60-240 secondi [104]. Inoltre, non sono stati riscontrati effetti benefici negli esercizi di durata inferiore a 60 secondi. La maggior parte degli studi inclusi nella meta-analisi riguardava l’esercizio di resistenza.
Tuttavia, è dimostrato che l’integrazione di beta-alanina può migliorare la resistenza muscolare negli atleti allenati alla resistenza [105] e può migliorare la composizione corporea [106]. Sono necessari ulteriori studi per esaminare l’effetto ergogenico della beta-alanina sulla composizione corporea e sulle prestazioni. Tuttavia, dato che i bodybuilder si allenano spesso con più di 10 ripetizioni per serie e spesso includono tecniche di intensità come drop set, pause di riposo, myo reps e altre, la beta-alanina potrebbe apportare un beneficio alla resistenza di queste serie [9].
Pertanto, potrebbe essere ragionevole per un bodybuilder consumare 3-5 g di beta alanina al giorno durante le fasi di allenamento ad alte ripetizioni o nelle fasi di allenamento in cui si incorporano diverse tecniche di intensità che prolungano la durata di un set. Come la creatina monoidrato, la beta-alanina non ha un effetto acuto, in quanto le concentrazioni di carnosina muscolare richiedono circa 4 settimane per raggiungere concentrazioni tali da produrre un effetto ergogenico, a condizione che se ne consumi una quantità sufficiente al giorno [104].
Citrullina Malato:
Recentemente, la Citrullina Malato ha guadagnato popolarità tra i bodybuilder. Il potenziale effetto ergogenico è dovuto all’aumento del flusso ematico al muscolo, alla produzione di ATP e alla potenziale capacità della Citrullina Malato di agire come agente tampone [107]. È stato dimostrato che il consumo di 8g di Citrullina Malato aumenta le ripetizioni fino al cedimento del 50% [107,108,109,110], riduce l’indolenzimento muscolare del 40% [107] e migliora la forza massimale e la potenza anaerobica [111].
Tuttavia, non tutti gli studi hanno osservato effetti ergogenici del consumo di Citrullina Malato. Due studi recenti non hanno mostrato un miglioramento delle prestazioni, un aumento della risposta del gonfiore muscolare dovuto all’allenamento, un’attenuazione della fatica o un aumento dell’attenzione e dell’energia in seguito all’integrazione di Citrullina Malato in uomini allenati contro-resistenza a livello amatoriale [112,113].
Una recente meta-analisi di Trexler et al. ha analizzato 12 studi sullla CM per le prestazioni di forza e potenza [114]. Sebbene abbiano riscontrato solo una piccola dimensione dell’effetto (0,20), hanno concluso che questo potrebbe essere rilevante per gli atleti di alto livello in cui i risultati delle competizioni si decidono su margini ridotti, come i culturisti agonisti di alto livello. Si consiglia di assumere la Citrullina Malato circa 60 minuti prima dell’esercizio fisico per consentire un assorbimento sufficiente.
Sono necessarie ulteriori ricerche per determinare l’efficacia della Citrullina Malato nell’esercizio contro-resistenza. Allo stato attuale, i dati indicano un effetto benefico o neutro sulle prestazioni. Pertanto, sulla base delle prove attuali, 8g al giorno di Citrullina Malato consumati prima dell’esercizio potrebbero avere dei benefici interessanti per i bodybuilder.
Alfa-GPC:
L’Alfa-GPC (alfa-glicerofosfocolina o colina alfoscerato) è un fosfolipide contenente colina. Quando viene ingerita, l’Alfa-GPC viene metabolizzata in colina e glicerolo-1-fosfato. La colina è un precursore dell’acetilcolina, un neurotrasmettitore coinvolto nella memoria, nell’attenzione e nella contrazione dei muscoli scheletrici. Il glicerolo-1-fosfato serve a sostenere le membrane cellulari.[https://pubmed.ncbi.nlm.]
L’Alfa-GPC sembra attraversare facilmente la barriera emato-encefalica e viene assorbito rapidamente. Attualmente è il miglior colinergico per aumentare i livelli plasmatici e cerebrali di colina.[https://pubmed.ncbi.nlm.]
L’integrazione orale di Alfa-GPC è interessante soprattutto per scopi nootropici o di potenziamento cognitivo. Esistono numerosi studi sui roditori che supportano questo effetto, ma non è ancora stato dimostrato negli esseri umani altrimenti sani. Negli anziani affetti da demenza lieve o moderata – che comporta un’alterazione della neurotrasmissione colinergica – l’Alfa-GPC migliora i sintomi cognitivi (ad esempio, disturbi della memoria e dell’attenzione).[https://pubmed.ncbi.nlm] L’Alfa-GPC può anche migliorare l’efficacia degli inibitori dell’acetilcolinesterasi (cioè i farmaci che aumentano la disponibilità di acetilcolina rallentandone la degradazione), utilizzati per il trattamento della malattia di Alzheimer.[https://pubmed.ncbi.nlm.]
Gli atleti sono un’altra popolazione che può trarre beneficio dall’integrazione di Alfa-GPC. Prove preliminari suggeriscono che l’alfa-GPC aumenta la potenza del salto verticale.[https://jissn.biomedcentral.com][https://pubmed.ncbi.nlm.] Inoltre, uno studio pilota ha riportato che l’Alfa-GPC ha aumentato il picco di forza nella panca, ma non la potenza di picco o il tasso di sviluppo della forza.[Ziegenfuss T, Landis J, Hofheins JJ Int Soc Sports Nutr.] Attualmente non è chiaro se l’Alfa-GPC aumenti la forza isometrica, ma i dati empirici e aneddotici sono incoraggianti [https://pubmed.ncbi.nlm.]
L’integrazione di un dosaggio pari a 600mg di Alpha-GPC prima di un test di potenza (spinte su panca) ha riportato un miglioramento della potenza del 14% rispetto al placebo quando assunta 45 minuti prima dell’attività; si trattava di uno studio pilota.[http://www.jissn.com] In media si è notato che il dosaggio di Alfa-GPC efficacie per trarre miglioramenti nella forza è nel range dei 300-600mg 45-30 minuti prima della seduta allenante.
Multi Vitaminico-Multi Minerale:
Storicamente, i bodybuilder hanno utilizzato diete restrittive che eliminano alimenti o interi gruppi di alimenti. Di conseguenza, sono comuni numerose carenze di vitamine e minerali. Nei bodybuilder a dieta sono state osservate carenze di Calcio, vitamina D, Zinco, Ferro e altre ancora [115,116,117]. Tuttavia, la maggior parte della letteratura sulle pratiche alimentari dei bodybuilder risale agli anni ’80 e ’90; pertanto, sono necessari dati più recenti [2].
Più di recente, le pratiche alimentari dei bodybuilder che seguono una dieta tradizionale restrittiva sono state confrontate con quelle degli agonisti che utilizzano un approccio dietetico basato sui macronutrienti, in cui nessun alimento o gruppo alimentare è off limits [118]. Non sorprende che i concorrenti che utilizzano un approccio dietetico più flessibile presentino meno carenze di micronutrienti. In particolare, la vitamina E, la vitamina K e le proteine sono risultate significativamente inferiori nelle donne che utilizzavano approcci dietetici rigidi rispetto a quelle che utilizzavano approcci più flessibili. Nel presente articolo, specie se si parla di Off-Season, si raccomanda di utilizzare un approccio dietetico flessibile, in cui nessun alimento o gruppo viene eliminato dalla dieta.
In questo modo, è meno probabile che si verifichino carenze di micronutrienti, soprattutto se si considera che le atlete in Off-Season hanno a disposizione una maggiore quantità di calorie rispetto a quelle a dieta per un contest, il che dovrebbe consentire loro di incorporare una maggiore varietà di alimenti.
Ciononostante, può essere consigliabile raccomandare un integratore multivitaminico/minerale a basso dosaggio (≤100% RDA) come misura di sicurezza per prevenire eventuali carenze di micronutrienti, sottolineando al contempo il consumo di una buona varietà di alimenti al giorno per soddisfare il fabbisogno di micronutrienti.
Omega 3 (EPA-DHA):
Gli acidi grassi polinsaturi con un doppio legame a tre atomi di distanza dal gruppo metilico terminale sono noti come ω-3 o acidi grassi omega-3 (O3). Un basso apporto di O3 nelle diete occidentali rispetto ad altre fonti di grassi alimentari (come gli acidi grassi omega-6) è associato a un peggioramento della salute multispettrale negli studi epidemiologici [119]. Pertanto, è interessante concentrarsi specificamente sulle modifiche della dieta per fornire acidi eicosapentaenoici e docosaesaenoici (EPA e DHA) – la carenza alimentare più comune nel mondo occidentale; ma vale la pena notare che la misurazione, l’interazione e l’effetto di O3 e acidi grassi omega-6 in relazione alla salute non sono chiari e vanno oltre lo scopo di questo articolo. Per una rassegna si rimanda ad altra pubblicazione [120].
Oltre alla salute, c’è interesse per i potenziali effetti anabolici degli integratori di EPA e DHA [121], che di solito vengono forniti attraverso l’olio di pesce o, in alcuni casi, l’olio di alghe. Tuttavia, ci sono dati contrastanti sulla capacità dell’olio di pesce di aumentare la risposta della sintesi proteica muscolare all’ingestione di proteine. Mentre un articolo di revisione del 2014 ha evidenziato una serie di studi secondo cui l’olio di pesce può aumentare la risposta [122], uno studio recente non ha rilevato alcun effetto sulla risposta della MPS a una sessione di allenamento contro-resistenza e all’ingestione di proteine dopo l’allenamento [123]. Inoltre, i dati sull’ipertrofia longitudinale sono pochi [124] e gli studi sulle prestazioni dell’allenamento contro-resistenza sono contrastanti [125] e in gran parte non applicabili o difficili da valutare a causa dell’uso di partecipanti non allenati o di allenamenti non standardizzati ed ecologicamente non realistici rispetto al bodybuilding.
In una recente review che affronta specificamente la questione se gli integratori di O3 possano o meno aumentare l’ipertrofia [126], gli autori hanno concluso che attualmente non ci sono prove sufficienti per fare tale affermazione. Sebbene siano necessarie ulteriori ricerche prima di poter raccomandare l’integrazione di O3 (o di alterazioni della dieta) a fini di costruzione muscolare, i benefici per la salute dell’integrazione di O3 sono degni di nota. Ad esempio, recenti meta-analisi hanno riportato che l’integrazione di olio di pesce riduce i sintomi della depressione [127], diminuisce il rischio di morte cardiaca [128], riduce la pressione sanguigna [129] e diminuisce la circonferenza vita [130]. Pertanto, gli atleti estetici possono prendere in considerazione l’integrazione giornaliera di olio di pesce (o di alghe) (1.5-2.5g di EPA/DHA) per la salute generale e multi spettro, ma sono necessari studi futuri per formulare raccomandazioni relative alle prestazioni nel bodybuilding.
Acido Arachidonico (AA):
L’Acido Arachidonico (AA) è l’acido grasso omega-6 più rilevante dal punto di vista biologico e, nella membrana lipidica di una cellula, è l’acido grasso che viene confrontato con i due acidi grassi dell’olio di pesce (EPA e DHA) nella costituzione di un rapporto omega-3:6. Dati recenti suggeriscono un’assunzione giornaliera di 50-250mg di Acido Arachidonico[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] con alcune fonti che stimano livelli fino a 500mg al giorno;[https://www.ncbi.nlm.] l’assunzione di Acido Arachidonico sembra essere inferiore nei vegetariani[https://www.ncbi.nlm.].
Si ritiene che l’Acido Arachidonico sia importante per il metabolismo del muscolo scheletrico, poiché si pensa che i fosfolipidi della membrana del sarcoplasma riflettano la dieta,[https://www.ncbi.nlm.][https://www.ncbi.nlm.] l’allenamento stesso sembra alterare il contenuto di fosfolipidi del muscolo (indipendentemente dalla composizione delle fibre muscolari[https://www.ncbi.nlm.] e associato a un rapporto omega 6:3 più basso[https://www.ncbi.nlm.][https://www.ncbi.nlm.]) e gli eicosanoidi dell’Acido Arachidonico interagiscono con la sintesi proteica muscolare attraverso i loro recettori.
L’Acido Arachidonico segnala la sintesi proteica muscolare attraverso una via dipendente dalla COX-2 (che suggerisce il coinvolgimento delle prostaglandine)[https://www.ncbi.nlm.] che è associata ad aumenti sia della prostaglandina E2 (PGE2) che del PGF(2α),[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] anche se l’incubazione con PGE2 o PGF(2α) isolati non sembra replicare pienamente gli effetti ipertrofici dell’Acido Arachidonico. [https://www.ncbi.nlm.] PGE2 e PGF(2α) sono indotti anche dall’esercizio fisico (nello specifico, dallo stiramento delle cellule muscolari in vitro[https://www.ncbi.nlm.]) ed è stato osservato sia nel siero[https://pubmed.ncbi.nlm.][https://www.ncbi.nlm.] che a livello intramuscolare (quadruplicato, da 0,95+/-0,26ng/mL a 3,97+/-0. La capacità del riflesso da stiramento di aumentare le concentrazioni di PGE2 e PGF(2α)[https://www.ncbi.nlm.] potrebbe essere dovuta semplicemente al fatto che lo stiramento aumenta l’attività delle COX2.[https://www.ncbi.nlm.][https://www.ncbi.nlm.]
Va notato che l’integrazione di 1.500mg di Acido Arachidonico (rispetto a una dieta di controllo contenente 200mg dello stesso) per 49 giorni ha aumentato la secrezione di PGE2 da parte di cellule immunitarie stimolate (del 50-100%) in giovani uomini altrimenti sani,[https://www.ncbi.nlm.] ma la rilevanza di questo studio per il muscolo scheletrico non è nota. Questo studio ha anche osservato che, senza stimolazione, non c’erano differenze significative tra i gruppi.[https://www.ncbi.nlm.] Altrove, è stata osservata una tendenza all’aumento delle concentrazioni sieriche di PGE2 a riposo in uomini allenati a cui sono stati somministrati 1.000mg di Acido Arachidonico per 50 giorni.[https://www.ncbi.nlm.]
L’Acido Arachidonico, attraverso gli eicosanoidi noti come PGF(2α) e PGE2, stimola la sintesi proteica muscolare. Sono prodotti a partire dall’Acido Arachidonico, ma normalmente non formano i rispettivi eicosanoidi per la costruzione del muscolo finché la cellula non viene stimolata da un fattore di stress (come il riflesso di stiramento di una cellula muscolare) che ne induce la produzione.
Il recettore per il PGF(2α) (recettore FP) sembra essere sovraregolato dagli inibitori della COX1 (l’acetaminofene utilizzato in questo studio)[https://www.ncbi.nlm.] e si ritiene che una maggiore segnalazione del PGF(2α) sia alla base del miglioramento della sintesi proteica muscolare osservato nei soggetti anziani con farmaci antinfiammatori. La supplementazione di Acido Arachidonico non sembra influenzare la quantità di recettori FP nei giovani;[https://www.ncbi.nlm.] mentre l’esercizio fisico stesso può aumentare il contenuto di recettori EP3, né gli inibitori della COX1[https://www.ncbi.nlm.] né l’Acido Arachidonico[https://www.ncbi.nlm.] sembrano influenzarlo ulteriormente.
Tuttavia, è stato riscontrato che l’uso di inibitori della COX2 (nei giovani) sopprime l’aumento di PGF(2α) indotto dall’esercizio fisico (Ibuprofene e Acetaminofene)[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e di PGE2,[https://www.ncbi.nlm.] il che si pensa sia dovuto al fatto che la conversione da PGH2 in questi metaboliti dipende dall’attività della COX2.
Poiché la produzione di questi eicosanoidi dipende dall’enzima COX2, si ritiene che l’inibizione di questo enzima riduca gli effetti anabolizzanti dell’esercizio fisico se assunto prima dello stesso.
L’acido arachidonico (così come l’EPA dall’olio di pesce) non ha compromesso l’assorbimento del glucosio nelle cellule muscolari isolate e 10μM di acido grasso sono in grado di attenuare la resistenza all’Insulina indotta dai grassi saturi; [https://pubmed.ncbi.nlm.] un fenomeno osservato con i grassi saturi a 18 o più catene di carbonio[https://www.ncbi.nlm.] che non sembra applicarsi agli acidi grassi polinsaturi di uguale lunghezza di catena[https://www.ncbi.nlm.][https://www.ncbi.nlm.] ed è probabilmente legato all’aumento delle ceramidi intracellulari[https://www.ncbi.nlm.] che compromettono la segnalazione di Akt[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e riducono l’assorbimento di glucosio mediato da GLUT4 con l’Insulina.[https://www.ncbi.nlm.]
L’Acido Arachidonico e i grassi polinsaturi omega-3 sono entrambi associati a una migliore sensibilità all’Insulina delle cellule muscolari, che potrebbe essere secondaria alla riduzione dei livelli di grassi saturi nella membrana lipidica e quindi alla riduzione delle concentrazioni intracellulari di ceramidi. È possibile che ciò non sia correlato agli eicosanoidi o al rapporto omega-3:6.
In 31 uomini allenati, sottoposti a un programma di sollevamento pesi e a una dieta standardizzata (500kcal in eccesso con 2g/kg di proteine) con 1g di Acido Arachidonico al giorno o placebo, l’integrazione per 50 giorni è sembrata aumentare la potenza di picco (7,1%) e la potenza media (3,6%) al test di Wingate, ma non è riuscita a influenzare positivamente la massa muscolare o le misure di potenza del sollevamento pesi (bench press e leg press).[https://www.ncbi.nlm.]
Attualmente non ci sono prove sufficienti per raccomandare una dose ideale di integrazione di Acido Arachidonico, ma aneddoticamente si usa un dosaggio di circa 1.500 mg da assumere 45 minuti prima dell’allenamento per un periodo medio di 8 settimane. Non è certo che si tratti di una dose ottimale o che sia necessaria la tempistica.
Va inoltre notato che per le persone affette da patologie infiammatorie croniche, come l’artrite reumatoide o le malattie infiammatorie intestinali, la dose ideale di Acido Arachidonico può essere in realtà una sua restrizione dietetica. Nei casi di malattie infiammatorie, l’integrazione di Acido Arachidonico è probabilmente controindicata.
Raccomandazioni per gli integratori alimentari e il dosaggio per i bodybuilder in Off-Season:
Creatina Monoidrato= 3-5g/die;
Beta-Alanina= 3-5g/die;
Citrullina Malato= 8g/pre-workout;
Alfa-GPC= 300-600mg/pre-workout;
Caffeina= 5-6mg/Kg/pre-workout (media standard tra 200 e 600mg/die);
Multi Vitaminico – Multi Minerale= ≤100% RDA/die;
Omega 3 (EPA-DHA)= 1.5-2.5g/die;
Acido Arachidonico= 1.5g/pre-workout.
Supplementazione PEDs:
Una cosa occorre premettere prima di procedere con la descrizione delle molecole più utilizzate nel contesto della Off-Season: non esistono PEDs esclusivamente confinabili in uno dei contesti della programmazione di un bodybuilder. Esiste il grado di versatilità il quale sta ad indicare quanto una molecola possa essere gestita con facilità in situazioni preparatorie differenti. Esistono molecole che per caratteristiche possono dare vantaggi maggiori in Off-Season/Bulk per via di alcune loro caratteristiche che in altro contesto, per esempio il pre-contest, risulterebbero più complesse da gestire. Ma questo non significa che tali molecole siano generalemnte da considerarsi “off-limitz” in un altra fase della preparazione annuale.
Premesso ciò, l’attenzione in questo paragrafo si concentrerà sui principali PEDs usati in Off-Season.
Tra tutti gli AAS, il Testosterone è quello che non ha bisogno di particolari presentazioni. Si tratta dell’ormone sessuale maschile per antonomasia. Nell’uomo, il Testosterone svolge un ruolo fondamentale nello sviluppo dei tessuti riproduttivi maschili, come i testicoli e la prostata, oltre a promuovere le caratteristiche sessuali secondarie, come l’aumento della massa muscolare e ossea e la crescita dei peli. Inoltre, in entrambi i sessi, il Testosterone è coinvolto nella salute e nel benessere, compresi gli stati d’animo, il comportamento e la prevenzione dell’osteoporosi in cooperazione con l’Estradiolo. Livelli insufficienti di Testosterone negli uomini possono portare ad anomalie, tra cui la fragilità e la perdita ossea.
In generale, il Testosterone promuove la sintesi proteica e quindi la crescita dei tessuti dotati di recettori per gli androgeni. Il Testosterone può essere descritto come avente effetti virilizzanti e anabolizzanti (anche se queste descrizioni categoriali sono in qualche modo arbitrarie, poiché vi è una grande sovrapposizione reciproca tra di essi).
Gli effetti anabolizzanti comprendono la crescita della massa e della forza muscolare, l’aumento della densità e della resistenza ossea e la stimolazione della crescita lineare e della maturazione ossea.
Gli effetti androgeni comprendono la maturazione degli organi sessuali, in particolare del pene, e la formazione dello scroto nel feto, e dopo la nascita (di solito nella pubertà) l’approfondimento della voce, la crescita dei peli del viso (come la barba) e dei peli ascellari. Molti di questi effetti rientrano nella categoria dei caratteri sessuali secondari maschili.
Al principio degli anni 30 del novecento avvenne la sintesi chimica del Testosterone, quando Butenandt e G. Hanisch pubblicarono un articolo che descriveva “Un metodo per preparare il Testosterone dal colesterolo”. Solo una settimana dopo, il terzo gruppo, Ruzicka e A. Wettstein, annunciò una domanda di brevetto in un documento “Sulla preparazione artificiale dell’ormone testicolare Testosterone (Androsten-3-one-17-ol).” Ruzicka e Butenandt ricevettero il premio Nobel per la chimica nel 1939 per il loro lavoro.
Gli studi clinici sull’uomo, che prevedevano dosi PO (per via orale) di Methyltestosterone o iniezioni di Testosterone Propionato, iniziarono già nel 1937. Il Testosterone Propionato è menzionato in una lettera all’editore della rivista Strength and Health nel 1938; questo è il primo riferimento noto a un AAS in una rivista statunitense di sollevamento pesi o Bodybuilding.
Lo sviluppo delle proprietà di costruzione muscolare del Testosterone proseguì negli anni ’40, in Unione Sovietica e nei paesi del blocco orientale come la Germania dell’Est, dove sono stati utilizzati programmi di AAS per migliorare le prestazioni dei sollevatori di pesi olimpici e di altri dilettanti già prima degli anni ’50. In risposta al successo dei sollevatori di pesi russi, il medico della squadra olimpica statunitense John Ziegler lavorò con un equipe di chimici per sviluppare un AAS con effetti androgeni ridotti. Ma questa è un altra storia.
L’uso del Testosterone nello sport si diffuse tra gli anni ’50 e gli anni ’60. Le forme utilizzate nei primi tempi erano il Testosterone in sospensione e il Testosterone Propionato, che rappresentano con il Methyltestosterone (Testosterone metilato in C-17) le forme più datate dell’ormone in questione (1935).
In ambito culturistico, il Testosterone rappresenta un AAS sufficientemente versatile in maniera dose-dipendente e sensibilità-dipendente dal momento che il dosaggio dovrebbe essere tarato in base alle risposte metaboliche soggettive alle quali è soggetto l’ormone (vedi, ad esempio, aromatizzazione in estrogeni). Questo ultimo punto è di estrema importanza al fine di evitare l’uso/abuso di AI (Inibitori dell’Aromatasi) e/o SERM (Modulatori Selettivi del Recettore degli Estrogeni). Oltre a peggiorare potenzialmente il quadro lipidico, sommandosi all’azione degli AAS utilizzati, essi riducono l’espressione epatica di IGF-1 cosa che può ridurre la risposta anabolizzante del protocollo PEDs. Nei soggetti caratterizzati da una elevata sensibilità all’attività estrogenica, le procedure applicate vedono: 1) l’uso di Raloxifene o Tamoxifene (SERM) a dosi sufficienti a impedire la comparsa o il peggioramento di una ginecomastia in stadio iniziale già presente e non ancora asportata chirurgicamente 2) l’uso di dosi fisiologiche di Testosterone come base onde evitare la comparsa di stati letargici, affaticabilità, disfunzioni sessuali ecc 3) l’uso di un “mix” composto da Testosterone e Boldenone (vedi in seguito) tale da poter usufruire della bassa e diversa sensibilità all’azione dell’Enzima Aromatasi su quest’ultimo riuscendo ad avere un controllo estrogenico teoricamente migliore (Testosterone e Boldenone mostrano qualità anabolizzanti intrinseche simili).
In un contesto Off-Season, quindi, vista l’importanza della presenza di un buon livello di Estradiolo sia sul complesso degli effetti anabolizzanti ricercati sia per la sua attività sessuale e cerebrale, il Testosterone andrebbe inizialmente calibrato sul soggetto e nel caso affiancato da dosi altrettanto ben tarate di SERM la dove ne risultasse un reale bisogno.
L’uso di un estere che garantisca un rilascio graduale della molecola (vedi Enantato o Cypionato) risulta la scelta migliore al fine di creare una soglia ematica stabile e esente da picchi e cali che possono risultare controproducenti a livello psicofisico. Tenere sempre in considerazione l’emivita di una molecola è uno dei punti fondamentali per sfruttarla al meglio. Nel caso degli esteri sopra citati, una divisione del dosaggio settimanale in due somministrazioni uguali distanziate da quattro-cinque giorni l’una dall’altra risulta una pratica ottimale allo scopo di creare una soglia ematica stabile.
I dosaggi comunemente utilizzati, parlando di molecole esterificate, vanno da 200mg ad 1g a settimana. Per quanto riguarda il Testosterone in sospensione, le dosi comunemente utilizzate vanno dai 175mg ai 700mg a settimana.
Il Boldenone [1,4-androstadiene-3-one,17b-ol], commercializzato con il nome di Equipoise, Ganabol, Equigan, Ultragan, e Boldane, è uno steroide anabolizzante-androgeno spesso legato all’estere Undecylenato. Strutturalmente molto simile al Testosterone, il Boldenone differisce da questo per il doppio legame tra C1 e C2.
La Ciba brevettò il Boldenone nel 1949. Successivamente, negli anni ’50 e ’60, sviluppò diversi esteri sperimentali del farmaco. Uno di questi era il Boldenone Undecilenato, che fu introdotto per uso clinico con il marchio Parenabol e fu utilizzato alla fine degli anni ’60 e all’inizio degli anni ’70. Tuttavia, fu sospeso prima della fine degli anni ’70. Ad oggi l’uso del Boldenone è legale in alcuni paesi in campo veterinario.
Se qualcuno volesse usare 500mg di Testosterone, ma non potrebbe usare un tale dosaggio dal momento che presenta particolare difficoltà nella gestione estrogenica in specie senza l’uso di AI come Exemestane o Anastrozolo, una conclusione a cui molti superficialmente sono giunti è che si potrebbe semplicemente usare il Boldenone al dosaggio sopra citato per ridurre della metà l’attività estrogenica, ma comunque supportare un’adeguata produzione di Estradiolo. Ma quando si approfondisci l’ipotesi e la si testa sul campo, è davvero così che stanno le cose? In realtà no, o, comunque, la media delle variabili di risposta spinge a confermare una maggiore validità nel “mixare” Testosterone e Boldenone coprendo la dose base calcolata in precedenza, e con variazione di percentuale T:B ratio da 1:1 a 2:1.
Comunque, oltre a rappresentare genericamente una discreta molecola sia in in preparazione alla gara che in Off-Season, I dosaggi utilizzati si settano nel range tra i 200mg ed i 500mg a settimana, spesso abbinato ad una dose variabile (vedi sopra) di Testosterone.
Il Nandrolone, noto anche come 19-nortestosterone, è uno Steroide Androgeno Anabolizzante (AAS) utilizzato sotto forma di molecola legata a esteri come quello Decanoato (nome commerciale Deca-Durabolin) e il Fenilpropionato (nome commerciale Durabolin). Gli esteri del Nandrolone sono utilizzati nel trattamento di anemie, cachessia (sindrome da deperimento), osteoporosi, cancro al seno e per altre indicazioni mediche.
Il Nandrolone è stato sintetizzato per la prima volta nel 1950. È stato introdotto per la prima volta nel mercato farmaceutico, come Nandrolone Fenilpropionato, nel 1959, e poi come Nandrolone Decanoato nel 1962, seguito da ulteriori esteri.
Il Nandrolone ha una bassa affinità di interazione con l’Enzima Aromatasi convertendo in Estrone, un estrogeno molto meno potente dell’Estradiolo, circa 10 volte meno attivo, e, come tale, è un estrogeno relativamente debole. In una condizione di somministrazione del Nandrolone senza una base di Testosterone, i livelli di Estradiolo calerebbero marcatamente a favore di un aumento del Estrone il quale non potrebbe però sostituire nelle diverse attività tissutali il prima citato E2. Le conseguenze negative si verificherebbero dall’attività sessuale all’attività neurosteroidea.
Infatti, un effetto da non sottovalutare con l’uso di Nandrolone è il suo impatto sul SNC. L’impatto del Nandrolone sul Sistema Nervoso Centrale è stato osservato scientificamente. Nello studio intitolato “The Impact of Nandrolone Decanoate on the Central Nervous System” vengono descritti chiaramente i numerosi effetti psicologici di questa molecola. Essi comprendono e influenzano:
1- Aggressività 2- Ansia, paura e stress 3- Ricompensa e dipendenza 4- Apprendimento, memoria e capacità di lavoro 5- Locomozione e attività fisica 6- Effetti sulla HPAA (Asse Ipotalamo-Pituitaria-Surrene) 7- Effetto sui neurotrasmettitori: Recettore Acido γ-Aminobutirrico Tipo A (GABAA); Recettori 5-idrossitriptamina (5-HT) e 5-HT; Recettori della Dopamina e Recettori Oppioidi.
Questo effetto, unito alla modesta potenzialità anabolizzante se confrontata con altre molecole anche della stessa famiglia, fa pendere l’ago della bilancia verso gli svantaggi d’uso piuttosto che i vantaggi. Sebbene vi sia un rapporto tra Testosterone e Nandrolone finalizzato a ridurre la comparsa di questi effetti avversi (ratio T:N = 2:1) su un buon numero di soggetti risulta dare comunque problemi rilevanti.
Il suo uso principale in Off-Season comprende dosaggi medi tra i 200mg ed i 400mg a settimana, con un adeguato rapporto con il Testosterone. Se utilizzato a fini di recupero articolare viene usato a dosaggi di 100mg a settimana, e con tali dosaggi difficilmente emergono i problemi sopra elencati a patto che ci sia una base di Testosterone.
Il Drostanolone, noto anche come 2α-metil-5α-diidrotestosterone (2α-metil-DHT) o come 2α-metil-5α-androstan-17β-ol-3-one, è uno steroide androstano sintetico e un derivato del DHT. Si tratta nello specifico di DHT con un gruppo metile in posizione C2α. La forma esterificata Drostanolone Propionato è stata usata in passato nel trattamento del cancro al seno nelle donne per via della sua attività antiestrogenica. Questa azione il Drostanolone la esplica sia agendo come antagonista del recettore degli estrogeni e sia come inibitore dell’Enzima Aromatasi. Ed è proprio per questo motivo che una molecola generalmente relegata all’uso in “Cut” o pre-gara trova un suo uso funzionale in Off-Season. La sua attività AI è comunque moderata ma sufficiente in un buon numero di soggetti per evitare l’aggiunta di SERM e/o AI di altro genere. L’attività AI moderata sembra non incidere negativamente in modo sensibile sull’Asse GH/IGF1.
L’effetto miotrofico risulta simile a quello osservato con il Methenolone, in generale moderatamente inferiore al Testosterone. I dosaggi utilizzati in Off-Season per il controllo estrogenico sono nel range dei 200-400mg a settimana (diviso in due iniezioni distanziate da 4-5 giorni) per l’estere Enantato, mentre per il Propionato 150-350mg a settimana (dosi a giorni alterni o giornaliere).
Il Trenbolone, noto anche come 19-nor-δ9,11-testosterone o come estra-4,9,11-trien-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone) sintetizzato per la prima volta nel 1963. Si tratta nello specifico di Nandrolone con due doppi legami aggiuntivi nel nucleo steroideo. Gli esteri del Trenbolone, che hanno un estere in posizione C17β, includono il Trenbolone Acetato, il Trenbolone Enantato, Il Trenbolone Hexahydrobenzylcarbonato e il Trenbolone Undecanoato. Il Trenbolone Acetato (marchi Finajet, Finaplix, e altri) e il Trenbolone Hexahydrobenzylcarbonato (marchi Parabolan, Hexabolan), sono o sono stati commercializzati per uso veterinario e clinico nell’uomo. Il Trenbolone Acetato è utilizzato in medicina veterinaria nel bestiame per aumentare la crescita muscolare e l’appetito degli animali, mentre il Trenbolone Hexahydrobenzylcarbonato è stato utilizzato in passato a livello clinico nell’uomo, ma ora non è più commercializzato.
Si tratta di uno degli AAS più versatili in assoluto, con un ottima resa tanto in preparazione alla gara quanto in Off-Season. L’enorme potenziale anabolizzante del Trenbolone, così come dei suoi analoghi, è stato riportato fin dagli anni ’60. La sua diffusione nel Bodybuilding è iniziata circa negli anni ’80 del secolo scorso. La sua elevata potenzialità miotrofica, lipolitica e di spinta mentale lo resero in poco tempo estremamente popolare tra i culturisti.
In Off-Season viene utilizzato nelle sue forme eseterificate Enantato e Hexahydrobenzylcarbonato a dosaggi nell’ordine dei 100-400mg a settimana (divisa in due somministrazioni distanziate l’una dall’altra da 4-5 giorni), sebbene il trend d’oltre oceano è arrivato a dosaggi decisamente eccessivi e nell’ordine del grammo. Per l’esetere Acetato i dosaggi medi vanno da 150mg a 350mg a settimana con dosaggi a giorni alterni o giornalieri.
E’ necessario ricordare ai lettori che gli effetti collaterali a livello del SNC possono verificarsi in alcuni punti come nel caso del Nandrolone sebbene i vantaggi rendano il Trenbolone più bilanciato tra sides e vantaggi.
Il Trestolone, noto anche come 7α-metil-19-nortestosterone (MENT) o come 7α-metilestr-4-en-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone). È una forma modificata del Nandrolone con un gruppo metile in posizione C7α. Tra gli AAS strettamente correlati vi sono il 7α-metil-19-norandrostenedione (MENT dione, trestione), un pro-ormone androgeno del Trestolone, e il Dimetandrolone (7α, 11β-dimetil-19-nortestosterone), il derivato metilato C11β del Trestolone, nonché il Mibolerone (7α,17α-dimetil-19-nortestosterone) e il Dimetiltrienolone (7α,17α-dimetil-δ9,11-19-nortestosterone). Anche il progestinico Tibolone (7α-metil-17α-etinil-δ5(10)-19-nortestosterone) è strettamente correlato al Trestolone.
Il Trestolone è stato descritto per la prima volta nel 1963. Tuttavia, non è stato successivamente studiato fino al 1990. Lo sviluppo del Trestolone per un potenziale uso nella contraccezione ormonale maschile e nella terapia sostitutiva degli androgeni è stato avviato nel 1993 ed è proseguito in seguito. Non sembra che siano stati condotti ulteriori sviluppi dal 2013. Il Trestolone è stato sviluppato dal Population Council, un’organizzazione non governativa senza scopo di lucro dedicata alla salute riproduttiva.
Come AAS, il Trestolone è un agonista del recettore degli androgeni (AR), analogamente agli androgeni come il Testosterone e il Diidrotestosterone (DHT). Questo AAS presenta spiccate proprietà anticortisolemiche sia attraverso l’inibizione enzimatica sia per attività antagonista recettoriale. Il Trestolone non è un substrato per la 5α-reduttasi e quindi non è potenziato o inattivato nei cosiddetti tessuti “androgeni” come la pelle, i follicoli piliferi e la ghiandola prostatica. Come tale, ha un elevato rapporto tra attività anabolica e androgena, analogamente ad altri derivati del Nandrolone. Il Trestolone è un substrato per l’Aromatasi e quindi produce come metabolita l’estrogeno 7α-metilestradiolo. Tuttavia, il Trestolone ha solo una debole attività estrogenica e una quantità che sembrerebbe essere insufficiente per scopi terapici sostitutivi, come evidenziato dalla diminuzione della densità minerale ossea negli uomini trattati con esso per l’ipogonadismo.
Il potenziale anabolizzante del Trestolone ha mostrato un grado di superiorità miotrofica rispetto al Trenbolone. Le sue caratteristiche ne fanno prediligere l’uso in Off-Season/Bulk. I dosaggi utilizzati con la forma Acetato sono nell’ordine dei 150-350mg a settimana con una cadenza nelle somministrazioni a giorni alterni. Sebbene sia più rara da reperire, la forma Enantato è utilizzato nel range dei 200-400mg a settimana divisi in somministrazioni ogni 4-5 giorni.
L’Oxymetholone, noto anche come 2-idrossimetilene-17α-metil-4,5α-diidrotestosterone (2-idrossimetilene-17α-metil-DHT) o come 2-idrossimetilene-17α-metil-5α-androstan-17β-olo-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del DHT. L’Oxymetholone è stato descritto per la prima volta in un articolo del 1959 da scienziati della Syntex. È stato introdotto per uso medico dalla Syntex e dalla Imperial Chemical Industries nel Regno Unito con il marchio Anapolon nel 1961. L’Oxymetholone è stato introdotto anche con i marchi Adroyd (Parke-Davis) nel 1961 e Anadrol (Syntex) nel 1962. Il farmaco è stato commercializzato negli Stati Uniti nei primi anni ’60.
Come altri AAS, l’Oxymetholone è un agonista del recettore degli androgeni (AR). Non è un substrato per la 5α-reduttasi (dal momento che è già 5α-ridotto) ed è uno substrato scarso per il 3α-idrossisteroide deidrogenasi (3α-HSD), e quindi mostra un alto rapporto di attività anabolizzante rispetto all’effetto androgenico.
Data la sua derivanza dal DHT, l’Oxymetholone non è un substrato per l’Enzima Aromatasi e quindi non può essere aromatizzato in metaboliti estrogenici. Tuttavia, caratteristica unica tra i derivati del DHT, l’Oxymetholone è comunque associato a un’estrogenicità relativamente elevata ed è noto per avere il potenziale di produrre effetti collaterali estrogenici come ginecomastia (anche se non comune) e ritenzione idrica. È stato suggerito che questo può essere una conseguenza del legame diretto a l’attivazione del recettore degli estrogeni da parte dell’Oxymetholone (estrogenicità intrinseca). L’Oxymetholone non possiede alcuna attività progestinica significativa. Per via della caratteristica attività estrogenica intrinseca, con l’uso di Oxymetholone è spesso necessario l’uso di un SERM onde avere un controllo sulla aumentata attività estrogenica.
A causa della sua struttura 17α-alchilata, l’Oxymetholone è epatotossico. L’uso a lungo termine del farmaco può causare una varietà di disturbi gravi, tra cui l’epatite, il cancro al fegato e la cirrosi; pertanto si raccomandano test periodici di funzionalità epatica per coloro che assumono l’Oxymetholone a fini terapeutici. Questa molecola ha ottenuto, infatti, la nomea di essere uno tra gli AAS più epatotossici. Ciò deriva da i dosaggi comunemente, ed erroneamente, utilizzati in contesto culturistico. Si parla di dosaggi che facilmente sforano i 150mg/die.
Osservazioni e esaminazione di diversi referti di esami ematici hanno evidenziato una soglia di “vantaggio/svantaggio” a favore del primo con un dosaggio calcolato con la formula 1mg/Kg. Genericamente, però, il dosaggio standard e conservativo si attesta nel range dei 50-100mg/die per non più di 28 giorni consecutivi, al fine di ridurre l’impatto negativo sul fegato e lipidemia.
Il Methandrostenolone, noto anche come 17α-metil-δ1-testosterone o come 17α-metilandrost-1,4-dien-17β-ol-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del Testosterone. È una modifica del Testosterone con un gruppo metile in posizione C17α e un doppio legame aggiuntivo tra le posizioni C1 e C2. Il farmaco è anche il derivato 17α-metilato del Boldenone (δ1-testosterone) e l’analogo δ1 del Methyltestosterone (17α-metiltestosterone).
Il Methandrostenolone è stato descritto per la prima volta nel 1955. È stato sintetizzato dai ricercatori dei laboratori CIBA di Basilea, in Svizzera. La CIBA depositò un brevetto statunitense nel 1957 e iniziò a commercializzare il farmaco sotto il nome di Dianabol nel 1958 negli Stati Uniti. Inizialmente veniva prescritto alle vittime di ustioni e agli anziani. Tra i primi utilizzatori vi furono i giocatori dell’Oklahoma University e l’allenatore dei San Diego Chargers Sid Gillman, che somministrò il Dianabol alla sua squadra a partire dal 1963.
Anche se il primo a somministrare il Methandrostenolone agli atleti fu il Dr. John Ziegler, personaggio che ebbe non poca importanza nella storia dell’uso degli AAS negli Stati Uniti. Ziegler contribuì a facilitare l’adozione degli AAS in generale, e del Dianabol in particolare, da parte degli atleti americani. Ziegler fu la prima persona a somministrare il Dianabol agli atleti competitivi poco dopo la sua introduzione da parte della CIBA nel 1958. Ebbe accesso al laboratorio CIBA a Summit (New Jersey) nel corso degli anni 50’ e somministrava già ai pesisti il Testosterone Propionato per “scopi di ricerca”. Da li il passo fu breve per la diffusione a macchia d’olio di questo AAS tra i culturisti.
Data la sua principale modifica strutturale, ossia la metilazione in C-17, il Methandrostenolone mostra un aumentata stabilità del legame recettoriale aumentando così l’affinità sia al AR sia, successivamente all’aromatizzazione nel suo metabolita 17-Methylestradiolo, per i recettori estrogenici rendendo il composto molto più estrogenico del Testosterone. Tale caratteristiche migliora però il potenziale proliferativo dei AR e l’influenza positiva sulla sintesi di IGF-1. Da non dimenticare è il suo significativo impatto anticortisolemico.
Trattandosi di una molecola con una discreta tendenza all’aromatizzazione, il suo uso tipico la vede inserita nelle fasi Off-Season. Il calcolo del dosaggio, per via dati aneddotici e osservativi raccolti, lo si ottiene con la formula 5mg/12Kg di peso corporeo. Trattandosi di un composto orale metilato in C-17 se ne scoraggia l’utilizzo oltre i 28 giorni consecutivi onde ridurre l’impatto negativo su fegato e lipidemia. Data la sua emivita di circa 4h, il dosaggio giornaliero dovrebbe essere diviso in più assunzioni distribuite durante l’arco della giornata.
L’Ormone della Crescita (GH) o Somatotropina, noto anche come Ormone della Crescita Umano (hGH o HGH), è un ormone peptidico che stimola la crescita, la riproduzione e la rigenerazione cellulare nell’uomo e in altri animali. È quindi importante per lo sviluppo umano. Il GH stimola anche la produzione di IGF-1 e aumenta la concentrazione di glucosio e acidi grassi liberi nel sangue. È un tipo di mitogeno specifico solo per i recettori di alcuni tipi di cellule. Il GH è un polipeptide a catena singola di 191 aminoacidi che viene sintetizzato, immagazzinato e secreto dalle cellule somatotrope nelle ali laterali dell’ipofisi anteriore.
Una forma ricombinante di hGH, chiamata Somatropina, viene utilizzata come farmaco da prescrizione per il trattamento dei disturbi della crescita nei bambini e della carenza di Ormone della Crescita negli adulti. Molte delle funzioni dell’hGH rimangono sconosciute.
Nel suo ruolo di agente anabolizzante, l’hGH è stato utilizzato dagli sportivi agonisti almeno dal 1982, quando la sola forma disponibile era quella derivata dall’Ipofisi dei cadaveri, ed è stato vietato dal CIO e dall’NCAA. L’analisi tradizionale delle urine non è in grado di rilevare il doping con HGH, pertanto il divieto è stato applicato solo all’inizio degli anni 2000, quando sono stati sviluppati test del sangue in grado di distinguere tra hGH naturale e artificiale.
In ambiente bodybuilding, l’hGH viene utilizzato in Off-Season (dai soggetti meglio informati) a dosaggi nel range delle 4-8UI al giorno o 8-16UI a giorni alterni. La somministrazione in concomitanza con l’uso di Insulina ha mostrato effetti sinergici molto evidenti che trovano la loro origine nel miglioramento della sintesi di IGF-1 e della sua frazione libera quindi attiva. Ricordo inoltre che l’uso di hGH può causare una sottoregolazione della funzionalità tiroidea per via del feedback negativo causato da un aumento della conversione del T4 in T3 per azione del GH. L’uso di T4, nel periodo d’uso in Off-Season, è in alcuni casi una necessità.
Il Fattore di Crescita Insulino-Simile 1 (IGF-1), chiamato anche Somatomedina C, è un ormone dalla struttura molecolare simile a quella dell’insulina che svolge un ruolo importante nella crescita infantile e ha effetti anabolici negli adulti. L’IGF-1 è costituito da 70 aminoacidi in una singola catena con tre ponti disolfuro intramolecolari.
L’IGF-1 è prodotto principalmente dal fegato. La produzione è stimolata dall’Ormone della Crescita (GH). La maggior parte dell’IGF-1 è legata a una delle 6 proteine di legame (IGF-BP). L’IGFBP-1 è regolato dall’Insulina. L’IGF-1 viene prodotto durante tutta la vita; i tassi più alti di produzione di IGF-1 si verificano durante la crescita puberale. I livelli più bassi si verificano nell’infanzia e nella vecchiaia.
L’IGF-1 lega e attiva il proprio recettore, l’IGF-1R, attraverso l’espressione sulla superficie cellulare delle tirosin-chinasi recettoriali (RTK) e segnala ulteriormente attraverso molteplici cascate di trasduzione intracellulare. L’IGF-1R è l’induttore che svolge un ruolo critico nella modulazione degli effetti metabolici dell’IGF-1 per la senescenza e la sopravvivenza cellulare. L’IGF-1 è responsabile di stimolare la crescita di tutti i tipi di cellule e di provocare effetti metabolici significativi. Un importante effetto metabolico dell’IGF-1 è la sua capacità di segnalare alle cellule che sono disponibili nutrienti sufficienti per l’ipertrofia e la divisione cellulare. Questi segnali consentono inoltre all’IGF-1 di inibire l’apoptosi cellulare e di aumentare la produzione di proteine cellulari. I recettori dell’IGF-1 sono ubiquitari, il che consente che i cambiamenti metabolici causati dall’IGF-1 si verifichino in tutti i tipi di cellule. Gli effetti metabolici dell’IGF-1 sono di vasta portata e possono coordinare il metabolismo delle proteine, dei carboidrati e dei grassi in una varietà di tipi di cellule diverse. La regolazione degli effetti metabolici dell’IGF-1 sui tessuti bersaglio è coordinata anche con altri ormoni, come l’Ormone della Crescita e l’Insulina.
L’IGF-1 da DNA ricombinante è disponibile principalmente in due diversi formati/varianti, lr3 e DES. È importante ricordare che, a prescindere dalla variante, tutti funzionano a livello sistemico nell’organismo e che, nonostante la somministrazione dell’ormone per via intramuscolare direttamente in un muscolo specifico, non genererà una crescita localizzata misurabile.
Ovviamente tralascerò di descrivere l’IGF-1 bioidentico commercializzato come Mecasermina dal momento che la sua farmacocinetica è identica a quella del IGF-1 endogeno. Dirò soltanto che mediamente viene utilizzato in dosi giornaliere nel range tra 60-1.000mcg post-workout. L’emivita di questa forma di IGF-1 è di circa 5.8h.
IGF-1 LR3: Questa forma è la variante di IGF-1 più comune e molto popolare sul mercato e utilizzata da bodybuilder e atleti di altre discipline. Contiene IGF-1 bioidentico costituito dalla catena originale di 70 aminoacidi, ma con 13 aminoacidi in più all’estremità N, per un totale di 83 aminoacidi. Possiede anche una seconda modifica, in cui un’Arginina si trova in 3a posizione invece dell’Acido Glutammico originale. Il risultato di queste modifiche è che l’IGF-1 continua a svolgere la sua attività originaria sul recettore dell’IGF-1 nei tessuti corporei e ha un’affinità di legame molto bassa per le proteine leganti l’IGF menzionate in precedenza. Inoltre, presenta una vita attiva significativamente più lunga, di circa 20-30 ore, rispetto a quella dell’IGF-1 di 12-15 ore. L’insieme di questi fattori ha dimostrato che l’LR3 ha un’efficacia circa tre volte superiore a quella dell’IGF-1.
I dosaggi medi utilizzati per questa forma sono nel range dei 40-80mcg/die. A causa della sua lunga vita attiva nell’organismo, la variante LR3 non dovrebbe essere somministrata più di una volta al giorno per il semplice fatto che non risulta necessario. Nei giorni di allenamento, il dosaggio di IGF-1 è solitamente somministrato subito dopo l’allenamento. La scelta è a discrezione dell’utilizzatore, in quanto può essere benissimo somministrato sia prima che dopo (solo prima dell’allenamento o solo dopo l’allenamento). E’ possibile comunque dividere il dosaggio giornaliero in due somministrazioni nell’arco della giornata, il dosaggio giornaliero completo può essere diviso quindi a metà tra i due (ad esempio, 20mcg prima dell’allenamento e 20mcg dopo l’allenamento, per un totale di 40mcg al giorno). Nei giorni di non allenamento, può essere somministrato in qualsiasi momento della giornata.
IGF-1 DES: Conosciuto anche come DES(1-3)IGF-1, questa è la forma di IGF-1 comunemente conosciuta come ad azione molto rapida e di solito è la meno preferita tra le due. Le sue modifiche rispetto alla molecola originale di IGF-1 sono tali da farle mancare i primi 3 aminoacidi all’N terminale, il che conferisce all’IGF-1 DES un totale di 67 aminoacidi nella sua catena rispetto ai 70 originali. Questa modifica garantisce all’IGF-1 DES una ridotta affinità di legame per le proteine leganti l’IGF menzionate in precedenza, oltre a una maggiore forza di legame e potenziale miotrofico, circa dieci volte superiore a quella dell’IGF-1 originale e cinque volte superiore a quella dell’IGF-1 LR3. A differenza dell’IGF-1 LR3, l’IGF-1 DES ha un’emivita molto più breve, di circa 20-30 minuti. Grazie alla sua attività più rapida e alla maggiore forza/potenza, la variante DES dell’IGF-1 è comunemente ritenuta in grado di ottenere una crescita muscolare localizzata nel sito in cui viene iniettata. Sebbene ciò sia in parte vero, gli studi hanno dimostrato che, come l’IGF-1 in generale, agisce a livello sistemico una volta raggiunti i capillari e il flusso sanguigno. Quindi l’effetto localizzato è minimo e non significativamente differente dall’effetto sistemico.
Il dosaggio della variante DES è leggermente più variabile rispetto a quello del LR3. Per l’IGF-1 DES, il dosaggio varia da 50 a 150 mcg al giorno. A causa della sua emivita molto più breve rispetto alla variante LR3, è possibile utilizzare dosaggi più elevati con una ipotetica riduzione del rischio di effetti a lungo termine sull’organismo, anche se è necessario usare comunque cautela. Può essere utilizzato nello stesso modo dell’IGF-1 LR3 post-workout, ed è infatti comunemente usato in questo modo a causa della sua breve emivita.
Entrambe le forme di IGF-1 possono essere somministrate per via intramuscolare o sottocutanea. L’uso di una delle due forme non deve superare la durata di 30 giorni prima di una pausa di almeno 2 settimane, anche se fare pause più lunghe di 2 settimane tra un ciclo di IGF-1 e l’altro è l’opzione migliore. Questo non solo per ridurre il rischio di effetti sulla salute a lungo termine, ma anche per garantire che i recettori dell’IGF-1 tornino ad un grado di sensibilità ottimale e, quindi, a “rispondere” correttamente dopo un ciclo.
L’insulina è un ormone peptidico prodotto dalle cellule beta delle isole pancreatiche. Regola il metabolismo dei carboidrati, dei grassi e delle proteine promuovendo l’assorbimento del glucosio dal sangue nelle cellule del fegato, dei grassi e dei muscoli scheletrici. In questi tessuti il glucosio assorbito viene convertito in glicogeno attraverso la glicogenesi o in grassi (trigliceridi) attraverso la lipogenesi o, nel caso del fegato, in entrambi. La produzione e la secrezione di glucosio da parte del fegato sono fortemente inibite da alte concentrazioni di Insulina nel sangue. L’Insulina circolante influisce anche sulla sintesi di proteine in un’ampia varietà di tessuti. È quindi un ormone anabolico e anticatabolico, che promuove la conversione di piccole molecole nel sangue in grandi molecole all’interno delle cellule. Bassi livelli di Insulina nel sangue hanno l’effetto opposto, favorendo un diffuso catabolismo, soprattutto del grasso corporeo di riserva.
La maggior parte dei bodybuilder utilizza una sola forma di Insulina (ad azione rapida o ultra-rapida), anche se alcuni utilizzano anche un’Insulina a lunga durata d’azione o in monoterapia insulinica o in conbinazione con le forme ad azione rapida o ultra-rapida.
L’Humalog® (Insulina Lispro) è senza dubbio la forma di Insulina più diffusa tra i bodybuilder insieme all’Humulin-R. L’Humalog è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”. Proprio al fine di potenziare la “finestra anabolica”, l’Humalog viene usata in concomitanza del GH il quale viene somministrato in una tempistica tale che i due picchi di rilascio (curva ematica massima) si “incrocino” andando a creare un affetto additivo di potenziamento della sintesi epatica di IGF-1 e della sua attività per via della riduzione dei trasportatori IGFBP.
Tuttavia, l’uso di una base insulinica composta da Insuline Glargine (Lantus), con una vita attiva di 24-26.5h, la quale sembra avere effetti di maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi, viene da alcuni inserita nei protocolli Off-Season.
I dosaggi di Insulina non andrebbero calcolati in modo distaccato dal piano alimentare e dal suo contenuto glucidico. Se il margine di “sicurezza” indica un assunzione di 10-15g di Carboidrati per UI di Insulina, questi non dovrebbero essere addizionati al piano alimentare già tarata in surplus calorico. Il calcolo delle unità dovrebbe essere tarato sul quantitativo glucidico della dieta e sul rapporto con il peso corporeo dell’atleta. Facciamo un semplice esempio: Soggetto di 90Kg = formula 1UI ogni 10Kg di peso = 9UI massime somministrabili per pasto e in base alla vita attiva della forma utilizzata = assicurarsi che il pasto appena successivo alla somministrazione dell’Insulina a questo dosaggio sia pari o superiore ai 90g di Carboidrati. Il monitoraggio della glicemia attraverso un glucometro è ovviamente d’obbligo in un protocollo di Insulina.
Nota: tali informazioni esposte non rappresentano in nessun modo un parere medico ne tanto meno una prescrizione e/o incentivo all’uso di sostanze dopanti e illegali. Le descrizioni presentate per i PEDs solitamente più utilizzati in Off-Season sono sintetiche sia per motivi di “Off Topic” sia per ragioni legate alla loro descrizione approfondita in altri articoli presenti nel database di questo sito. In queste pubblicazioni potrete trovare informazioni inerenti anche agli affetti collaterali connessi ad un uso/abuso “off-label” dei diversi PEDs.
Conclusioni:
Per concludere e fare una sintesi delle nozioni esposte in questo articolo, dobbiamo ricordarci che i bodybuilder in Off-Season dovrebbero concentrarsi sul consumo di una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) con l’obiettivo di guadagnare ~0,25-0,5% del peso corporeo a settimana per un “Natural”, mentre nel caso di un “Doped” la soglia può spostarsi tra l’1-2% con variabili connesse a risposte genetiche differenziali e anzianità nella carriera culturistica (principiante, intermedio e avanzato). In ogni caso, in una fetta maggioritaria di praticanti, ai bodybuilder avanzati si consiglia di essere più prudenti con il surplus calorico e il tasso di aumento di peso settimanale. L’assunzione di proteine nella dieta è raccomandata a 1,6-2,2 g/kg/giorno, con particolare attenzione a una quantità sufficiente di proteine a ogni pasto (0,40-0,55 g/kg/pasto) e a una distribuzione uniforme nell’arco della giornata (3-6 pasti). Per i “Doped”, in alcuni casi, l’introito proteico può essere portato, con minimi vantaggi in contesto ipercalorico, a 2,5g/Kg con le medesime linee guida di suddivisione per numero di pasti. I grassi alimentari devono essere consumati a livelli moderati, né troppo bassi né troppo alti (0,5-1,5 g/kg/die), per evitare un rapporto fTC sfavorevole e per prevenire riduzioni dei livelli di testosterone. Nei “Doped” l’obbiettivo con i lipidi è principalmente quello di assumerne una dose necessaria, e altamente qualitativa, al fine di assimilare vitamine liposolubili, per substrato strutturale, per sintesi di eicosanoidi (vedi assunzione EPA, DHA e AA), protezioni epidermide e capelli; di conseguenza attenersi ad un dosaggio medio pari a 35-50g/die. Dopo che le calorie sono state distribuite tra Proteine e Grassi, le restanti calorie dovrebbero provenire dai Carboidrati, assicurandosi di consumarne una quantità sufficiente (≥3-5 g/kg/giorno). Si possono ottenere benefici maggiori consumando proteine (0,40-0,55 g/kg/pasto) in prossimità delle sessioni di allenamento (1-2 ore prima dell’esercizio ed entro 1-2 ore dopo l’esercizio). È opportuno prendere in considerazione la Creatina Monoidrato (3-5 g/giorno) e la Caffeina (5-6 mg/kg), in quanto possono produrre effetti ergogenici per i bodybuilder. Inoltre, Beta-Alanina (3-5 g/die) e Citrullina Malato (8 g/die) sono integratori alimentari che possono essere presi in considerazione in quanto potenzialmente utili per i bodybuilder, a seconda dei regimi di allenamento individuali. I bodybuilder che non sono in grado di assumere un apporto sufficiente di micronutrienti e acidi grassi essenziali nella loro dieta dovrebbero prendere in considerazione l’integrazione di questi nutrienti per evitare carenze. Il limite principale di questo articolo è la mancanza di studi su larga scala e a lungo termine sui bodybuilder durante la Off-Season. Sono necessarie ulteriori ricerche su questa popolazione per ottimizzare la nutrizione e le raccomandazioni sugli integratori alimentari.
4. Philen R.M., Ortiz D.I., Auerbach S.B., Falk H. Survey of Advertising for Nutritional Supplements in Health and Bodybuilding Magazines. JAMA. 1992;268:1008. doi: 10.1001/jama.1992.03490080082029. [PubMed] [CrossRef] [Google Scholar]
5. Giampreti A., Lonati D., Locatelli C., Rocchi L., Campailla M.T. Acute neurotoxicity after yohimbine ingestion by a bodybuilder. [(accessed on 25 March 2019)];Clin. Toxicol. 2009 47:827–829. doi: 10.1080/15563650903081601. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19640235 [PubMed] [CrossRef] [Google Scholar]
7. Della Guardia L., Cavallaro M., Cena H. The risks of self-made diets: The case of an amateur bodybuilder. J. Int. Soc. Sports Nutr. 2015;12:5. doi: 10.1186/s12970-015-0077-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Hackett D.A., Johnson N.A., Chow C.-M. Training Practices and Ergogenic Aids Used by Male Bodybuilders. J. Strength Cond. Res. 2013;27:1609–1617. doi: 10.1519/JSC.0b013e318271272a. [PubMed] [CrossRef] [Google Scholar]
10. Forbes G.B., Brown M.R., Welle S.L., Lipinski B.A. Deliberate overfeeding in women and men: Energy cost and composition of the weight gain. Br. J. Nutr. 1986;56:1–9. doi: 10.1079/BJN19860080. [PubMed] [CrossRef] [Google Scholar]
11. Kreider R.B., Klesges R., Harmon K., Ramsey L., Bullen D., Wood L., Almada A., Grindstaff P., Li Y. Effects of Ingesting Supplements Designed to Promote Lean Tissue Accretion on Body Composition during Resistance Training. Int. J. Sport Nutr. 1996;6:234–246. doi: 10.1123/ijsn.6.3.234. [PubMed] [CrossRef] [Google Scholar]
12. Rozenek R., Ward P., Long S., Garhammer J. Effects of high-calorie supplements on body composition and muscular strength following resistance training. J. Sports Med. Phys. Fit. 2002;42:340–347. [PubMed] [Google Scholar]
13. Garthe I., Raastad T., Refsnes P.E., Sundgot-Borgen J. Effect of nutritional intervention on body composition and performance in elite athletes. Eur. J. Sport Sci. 2013;13:295–303. doi: 10.1080/17461391.2011.643923. [PubMed] [CrossRef] [Google Scholar]
14. American College og Sports Medicine American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. [(accessed on 25 March 2019)];Med. Sci. Sport. Exerc. 2009 41:687–708. doi: 10.1249/MSS.0b013e3181915670. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19204579 [PubMed] [CrossRef] [Google Scholar]
15. Lambert C.P., Frank L.L., Evans W.J., Lambert D.C.P. Macronutrient Considerations for the Sport of Bodybuilding. Sports Med. 2004;34:317–327. doi: 10.2165/00007256-200434050-00004. [PubMed] [CrossRef] [Google Scholar]
16. Walberg-Rankin J., Edmonds C.E., Gwazdauskas F.C. Diet and Weight Changes of Female Bodybuilders Before and After Competition. Int. J. Sport Nutr. 1993;3:87–102. doi: 10.1123/ijsn.3.1.87. [PubMed] [CrossRef] [Google Scholar]
17. Lamar-Hildebrand N., Saldanha L., Endres J. Dietary and exercise practices of college-aged female bodybuilders. J. Am. Diet. Assoc. 1989;89:1308–1310. [PubMed] [Google Scholar]
18. Houston M.E. Gaining Weight: The Scientific Basis of Increasing Skeletal Muscle Mass. Can. J. Appl. Physiol. 1999;24:305–316. doi: 10.1139/h99-024. [PubMed] [CrossRef] [Google Scholar]
19. Phillips S.M. A Brief Review of Critical Processes in Exercise-Induced Muscular Hypertrophy. Sports Med. 2014;44:71–77. doi: 10.1007/s40279-014-0152-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Campbell B.I., Aguilar D., Conlin L., Vargas A., Schoenfeld B.J., Corson A., Gai C., Best S., Galvan E., Couvillion K. Effects of High Versus Low Protein Intake on Body Composition and Maximal Strength in Aspiring Female Physique Athletes Engaging in an 8-Week Resistance Training Program. Int. J. Sport Nutr. Exerc. Metab. 2018;28:580–585. doi: 10.1123/ijsnem.2017-0389. [PubMed] [CrossRef] [Google Scholar]
22. Morton R.W., Murphy K.T., McKellar S.E., Schoenfeld B.J., Henselmans M., Helms E., Aragon A.A., Devries M.C., Banfield L., Krieger J.W., et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. [(accessed on 25 March 2019)];Br. J. Sports Med. 2018 52:376–384. doi: 10.1136/bjsports-2017-097608. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28698222 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Houltham S.D., Rowlands D.S. A snapshot of nitrogen balance in endurance-trained women. Appl. Physiol. Nutr. Metab. 2014;39:219–225. doi: 10.1139/apnm-2013-0182. [PubMed] [CrossRef] [Google Scholar]
25. Antonio J., Ellerbroek A., Silver T., Vargas L., Peacock C. The effects of a high protein diet on indices of health and body composition—A crossover trial in resistance-trained men. J. Int. Soc. Sports Nutr. 2016;13:8. doi: 10.1186/s12970-016-0114-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Bandegan A., Courtney-Martin G., Rafii M., Pencharz P.B., Lemon P.W. Indicator Amino Acid–Derived Estimate of Dietary Protein Requirement for Male Bodybuilders on a Nontraining Day Is Several-Fold Greater than the Current Recommended Dietary Allowance. J. Nutr. 2017;147:850–857. doi: 10.3945/jn.116.236331. [PubMed] [CrossRef] [Google Scholar]
27. Malowany J.M., West D.W.D., Williamson E., Volterman K.A., Sawan S.A., Mazzulla M., Moore D.R. Protein to Maximize Whole-Body Anabolism in Resistance-trained Females after Exercise. Med. Sci. Sports Exerc. 2019;51:798–804. doi: 10.1249/MSS.0000000000001832. [PubMed] [CrossRef] [Google Scholar]
28. Antonio J., Peacock C.A., Ellerbroek A., Fromhoff B., Silver T. The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. J. Int. Soc. Sports Nutr. 2014;11:19. doi: 10.1186/1550-2783-11-19. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
29. Antonio J., Ellerbroek A., Silver T., Orris S., Scheiner M., Gonzalez A., Peacock C.A. A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women—A follow-up investigation. J. Int. Soc. Sports Nutr. 2015;12:39. doi: 10.1186/s12970-015-0100-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Bray G.A., Smith S.R., de Jonge L., Xie H., Rood J., Martin C.K., Most M., Brock C., Mancuso S., Redman L.M. Effect of dietary protein content on weight gain, energy expenditure, and body composition during overeating: A randomized controlled trial. [(accessed on 25 March 2019)];JAMA. 2012 307:47–55. doi: 10.1001/jama.2011.1918. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22215165 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Tipton K.D., Ferrando A.A., Phillips S.M., Doyle D., Wolfe R.R. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am. J. Physiol. Metab. 1999;276:628–634. doi: 10.1152/ajpendo.1999.276.4.E628. [PubMed] [CrossRef] [Google Scholar]
32. Rieu I., Balage M., Sornet C., Giraudet C., Pujos E., Grizard J., Mosoni L., Dardevet D. Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia. J. Physiol. 2006;575:305–315. doi: 10.1113/jphysiol.2006.110742. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Burd N.A., Tang J.E., Moore D.R., Phillips S.M. Exercise training and protein metabolism: Influences of contraction, protein intake, and sex-based differences. [(accessed on 25 March 2019)];J. Appl. Physiol. 2008 106:1692–1701. doi: 10.1152/japplphysiol.91351.2008. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19036897 [PubMed] [CrossRef] [Google Scholar]
34. Drummond M.J., Dreyer H.C., Fry C.S., Glynn E.L., Rasmussen B.B. Nutritional and contractile regulation of human skeletal muscle protein synthesis and mTORC1 signaling. J. Appl. Physiol. 2009;106:1374–1384. doi: 10.1152/japplphysiol.91397.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Tang J.E., Moore D.R., Kujbida G.W., Tarnopolsky M.A., Phillips S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009;107:987–992. doi: 10.1152/japplphysiol.00076.2009. [PubMed] [CrossRef] [Google Scholar]
36. Kanda A., Nakayama K., Sanbongi C., Nagata M., Ikegami S., Itoh H. Effects of Whey, Caseinate, or Milk Protein Ingestion on Muscle Protein Synthesis after Exercise. Nutrients. 2016;8:339. doi: 10.3390/nu8060339. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Messina M., Lynch H., Dickinson J.M., Reed K.E. No Difference Between the Effects of Supplementing With Soy Protein Versus Animal Protein on Gains in Muscle Mass and Strength in Response to Resistance Exercise. Int. J. Sport Nutr. Exerc. Metab. 2018;28:674–685. doi: 10.1123/ijsnem.2018-0071. [PubMed] [CrossRef] [Google Scholar]
38. Joy J.M., Lowery R.P., Wilson J.M., Purpura M., De Souza E.O., Mc Wilson S., Kalman D.S., Dudeck J.E., Jäger R. The effects of 8 weeks of whey or rice protein supplementation on body composition and exercise performance. Nutr. J. 2013;12:86. doi: 10.1186/1475-2891-12-86. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
39. Babault N., Paizis C., Deley G., Guérin-Deremaux L., Saniez M.-H., Lefranc-Millot C., Allaert F.A. Pea proteins oral supplementation promotes muscle thickness gains during resistance training: A double-blind, randomized, Placebo-controlled clinical trial vs. Whey protein. J. Int. Soc. Sports Nutr. 2015;12:1692. doi: 10.1186/s12970-014-0064-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Tesch P.A. Glycogen and triglyceride utilization in relation to muscle metabolic characteristics in men performing heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1990;61:5–10. [PubMed] [Google Scholar]
41. Lane A.R., Duke J.W., Hackney A.C. Influence of dietary carbohydrate intake on the free testosterone: Cortisol ratio responses to short-term intensive exercise training. [(accessed on 25 March 2019)];Eur. J. Appl. Physiol. 2010 108:1125–1131. doi: 10.1007/s00421-009-1220-5. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20091182 [PubMed] [CrossRef] [Google Scholar]
42. Tegelman R., Aberg T., Pousette A., Carlström K. Effects of a diet regimen on pituitary and steroid hormones in male ice hockey players. [(accessed on 25 March 2019)];Int. J. Sports Med. 1992 13:420–430. doi: 10.1055/s-2007-1021292. Available online: https://www.ncbi.nlm.nih.gov/pubmed/1387870 [PubMed] [CrossRef] [Google Scholar]
43. Dorgan J.F., Judd J.T., Longcope C., Brown C., Schatzkin A., Clevidence B.A., Campbell W.S., Nair P.P., Franz C., Kahle L., et al. Effects of dietary fat and fiber on plasma and urine androgens and estrogens in men: A controlled feeding study. Am. J. Clin. Nutr. 1996;64:850–855. doi: 10.1093/ajcn/64.6.850. [PubMed] [CrossRef] [Google Scholar]
44. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Decrease of serum total and free testosterone during a low-fat high-fibre diet. J. Steroid Biochem. 1983;18:369–370. doi: 10.1016/0022-4731(83)90117-6. [PubMed] [CrossRef] [Google Scholar]
45. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Diet and serum sex hormones in healthy men. J. Steroid Biochem. 1984;20:459–464. doi: 10.1016/0022-4731(84)90254-1. [PubMed] [CrossRef] [Google Scholar]
46. Wang C., Catlin D.H., Starcevic B., Heber D., Ambler C., Berman N., Lucas G., Leung A., Schramm K., Lee P.W.N., et al. Low-Fat High-Fiber Diet Decreased Serum and Urine Androgens in Men. J. Clin. Endocrinol. Metab. 2005;90:3550–3559. doi: 10.1210/jc.2004-1530. [PubMed] [CrossRef] [Google Scholar]
47. Morton R.W., Sato K., Gallaugher M.P.B., Oikawa S.Y., McNicholas P.D., Fujita S., Phillips S.M. Muscle Androgen Receptor Content but Not Systemic Hormones Is Associated With Resistance Training-Induced Skeletal Muscle Hypertrophy in Healthy, Young Men. Front. Physiol. 2018;9:9. doi: 10.3389/fphys.2018.01373. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Tinsley G.M., Willoughby D.S. Fat-Free Mass Changes During Ketogenic Diets and the Potential Role of Resistance Training. Int. J. Sport Nutr. Exerc. Metab. 2016;26:78–92. doi: 10.1123/ijsnem.2015-0070. [PubMed] [CrossRef] [Google Scholar]
49. Vargas S., Romance R., Petro J.L., Bonilla D.A., Galancho I., Espinar S., Kreider R.B., Benítez-Porres J. Efficacy of ketogenic diet on body composition during resistance training in trained men: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2018;15:31. doi: 10.1186/s12970-018-0236-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
50. Kephart W.C., Pledge C.D., Roberson P.A., Mumford P.W., Romero M.A., Mobley C.B., Martin J.S., Young K.C., Lowery R.P., Wilson J.M., et al. The Three-Month Effects of a Ketogenic Diet on Body Composition, Blood Parameters, and Performance Metrics in CrossFit Trainees: A Pilot Study. Sports. 2018;6:1. doi: 10.3390/sports6010001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
51. Greene D.A., Varley B.J., Hartwig T.B., Chapman P., Rigney M. A Low-Carbohydrate Ketogenic Diet Reduces Body Mass Without Compromising Performance in Powerlifting and Olympic Weightlifting Athletes. [(accessed on 26 March 2019)];J. Strength Cond. Res. 2018 32:3373–3382. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30335720 [PubMed] [Google Scholar]
52. Bird S. Strength Nutrition: Maximizing Your Anabolic Potential. Strength Cond. J. 2010;32:80–86. doi: 10.1519/SSC.0b013e3181d5284e. [CrossRef] [Google Scholar]
53. American Dietetic Association. Dietitians of Canada. American College of Sports Medicine. Rodriguez N.R., Di Marco N.M., Langley S. American College of Sports Medicine position stand. Nutrition and athletic performance. [(accessed on 26 March 2019)];Med. Sci. Sports Exerc. 2009 41:709–731. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19225360 [PubMed] [Google Scholar]
54. Chung S.T., Chacko S.K., Sunehag A.L., Haymond M.W. Measurements of Gluconeogenesis and Glycogenolysis: A Methodological Review. Diabetes. 2015;64:3996–4010. doi: 10.2337/db15-0640. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Azizi F. Effect of dietary composition on fasting-induced changes in serum thyroid hormones and thyrotropin. Metabolism. 1978;27:935–942. doi: 10.1016/0026-0495(78)90137-3. [PubMed] [CrossRef] [Google Scholar]
56. Mathieson R.A., Walberg J.L., Gwazdauskas F.C., Hinkle D.E., Gregg J.M. The effect of varying carbohydrate content of a very-low-caloric diet on resting metabolic rate and thyroid hormones. Metabolism. 1986;35:394–398. doi: 10.1016/0026-0495(86)90126-5. [PubMed] [CrossRef] [Google Scholar]
57. Leveritt M., Abernethy P.J. Effects of Carbohydrate Restriction on Strength Performance. J. Strength Cond. Res. 1999;13:52–57. [Google Scholar]
58. Jacobs I., Kaiser P., Tesch P. Muscle strength and fatigue after selective glycogen depletion in human skeletal muscle fibers. Graefe’s Arch. Clin. Exp. Ophthalmol. 1981;46:47–53. doi: 10.1007/BF00422176. [PubMed] [CrossRef] [Google Scholar]
59. Ray S., Sale D.G., Lee P., Garner S., MacDougall J.D., McCartney N. Muscle Substrate Utilization and Lactate Production During Weightlifting. Can. J. Appl. Physiol. 1999;24:209–215. [PubMed] [Google Scholar]
60. Tesch P.A., Colliander E.B., Kaiser P. Muscle metabolism during intense, heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1986;55:362–366. doi: 10.1007/BF00422734. [PubMed] [CrossRef] [Google Scholar]
63. Mitchell J.B., DiLauro P.C., Pizza F.X., Cavender D.L. The Effect of Preexercise Carbohydrate Status on Resistance Exercise Performance. Int. J. Sport Nutr. 1997;7:185–196. doi: 10.1123/ijsn.7.3.185. [PubMed] [CrossRef] [Google Scholar]
64. Lima-Silva A.E., Silva-Cavalcante M.D., Oliveira R.S., Kiss M.A., Pires F.O., Bertuzzi R., Bishop D. Effects of a low- or a high-carbohydrate diet on performance, energy system contribution, and metabolic responses during supramaximal exercise. Appl. Physiol. Nutr. Metab. 2013;38:928–934. doi: 10.1139/apnm-2012-0467. [PubMed] [CrossRef] [Google Scholar]
65. Vega F., Jackson R. Dietary habits of bodybuilders and other regular exercisers. Nutr. Res. 1996;16:3–10. doi: 10.1016/0271-5317(95)02054-3. [CrossRef] [Google Scholar]
66. Chappell A.J., Simper T., Barker M.E. Nutritional strategies of high level natural bodybuilders during competition preparation. J. Int. Soc. Sports Nutr. 2018;15:4. doi: 10.1186/s12970-018-0209-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
67. Atherton P.J., Etheridge T., Watt P.W., Wilkinson D., Selby A., Rankin D., Smith K., Rennie M.J. Muscle full effect after oral protein: Time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am. J. Clin. Nutr. 2010;92:1080–1088. doi: 10.3945/ajcn.2010.29819. [PubMed] [CrossRef] [Google Scholar]
68. Res P.T., Groen B., Pennings B., Beelen M., Wallis G.A., Gijsen A.P., Senden J.M., Van Loon L.J. Protein ingestion before sleep improves postexercise overnight recovery. [(accessed on 25 March 2019)];Med. Sci. Sports Exerc. 2012 44:1560–1569. doi: 10.1249/MSS.0b013e31824cc363. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22330017 [PubMed] [CrossRef] [Google Scholar]
69. Moore D.R., Robinson M.J., Fry J.L., Tang J.E., Glover E.I., Wilkinson S.B., Prior T., Tarnopolsky M.A., Phillips S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2009 89:161–168. doi: 10.3945/ajcn.2008.26401. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19056590 [PubMed] [CrossRef] [Google Scholar]
70. Witard O.C., Jackman S.R., Breen L., Smith K., Selby A., Tipton K.D. Muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2014 99:86–95. doi: 10.3945/ajcn.112.055517. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24257722 [PubMed] [CrossRef] [Google Scholar]
71. Macnaughton L.S., Wardle S.L., Witard O.C., McGlory C., Hamilton D.L., Jeromson S., Lawrence C.E., Wallis G.A., Tipton K.D. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol. Rep. 2016;4:e12893. doi: 10.14814/phy2.12893. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
72. Schoenfeld B.J., Aragon A.A., Krieger J.W. The effect of protein timing on muscle strength and hypertrophy: A meta-analysis. J. Int. Soc. Sports Nutr. 2013;10:53. doi: 10.1186/1550-2783-10-53. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Areta J.L., Burke L.M., Ross M.L., Camera D.M., West D.W.D., Broad E.M., Jeacocke N.A., Moore D.R., Stellingwerff T., Phillips S.M., et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J. Physiol. 2013;591:2319–2331. doi: 10.1113/jphysiol.2012.244897. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Hudson J.L., Bergia R.E., Campbell W.W. Effects of protein supplements consumed with meals, versus between meals, on resistance training–induced body composition changes in adults: A systematic review. Nutr. Rev. 2018;76:461–468. doi: 10.1093/nutrit/nuy012. [PubMed] [CrossRef] [Google Scholar]
75. Trommelen J., Kouw I.W.K., Holwerda A.M., Snijders T., Halson S.L., Rollo I., Verdijk L.B., Van Loon L.J.C. Pre-sleep dietary protein-derived amino acids are incorporated in myofibrillar protein during post-exercise overnight recovery. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2018 1:457–467. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28536184 [Google Scholar]
76. Kouw I.W., Holwerda A.M., Trommelen J., Kramer I.F., Bastiaanse J., Halson S.L., Wodzig W.K., Verdijk L.B., Van Loon L.J. Protein Ingestion before Sleep Increases Overnight Muscle Protein Synthesis Rates in Healthy Older Men: A Randomized Controlled Trial. [(accessed on 25 March 2019)];J. Nutr. 2017 147:2252–2261. doi: 10.3945/jn.117.254532. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28855419 [PubMed] [CrossRef] [Google Scholar]
77. Snijders T., Res P.T., Smeets J.S., Van Vliet S., Van Kranenburg J., Maase K., Kies A.K., Verdijk L.B., Van Loon L.J. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. [(accessed on 25 March 2019)];J. Nutr. 2015 145:1178–1184. doi: 10.3945/jn.114.208371. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25926415 [PubMed] [CrossRef] [Google Scholar]
78. Joy J.M., Vogel R.M., Broughton K.S., Kudla U., Kerr N.Y., Davison J.M., Wildman R.E.C., DiMarco N.M. Daytime and nighttime casein supplements similarly increase muscle size and strength in response to resistance training earlier in the day: A preliminary investigation. J. Int. Soc. Sports Nutr. 2018;15:24. doi: 10.1186/s12970-018-0228-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Antonio J., Ellerbroek A., Peacock C., Silver T. Casein Protein Supplementation in Trained Men and Women: Morning versus Evening. Int. J. Exerc. Sci. 2017;10:479–486. [PMC free article] [PubMed] [Google Scholar]
80. Schoenfeld B.J., Aragon A.A. How much protein can the body use in a single meal for muscle-building? Implications for daily protein distribution. J. Int. Soc. Sports Nutr. 2018;15:10. doi: 10.1186/s12970-018-0215-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
81. Pennings B., Groen B.B., Van Dijk J.-W., De Lange A., Kiskini A., Kuklinski M., Senden J.M., Van Loon L.J. Minced beef is more rapidly digested and absorbed than beef steak, resulting in greater postprandial protein retention in older men. Am. J. Clin. Nutr. 2013;98:121–128. doi: 10.3945/ajcn.112.051201. [PubMed] [CrossRef] [Google Scholar]
82. Kim I.Y., Schutzler S., Schrader A., Spencer H.J., Azhar G., Ferrando A.A., Wolfe R.R. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2016 310:73–80. doi: 10.1152/ajpendo.00365.2015. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26530155 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Jentjens R., Jeukendrup A.E. Determinants of Post-Exercise Glycogen Synthesis During Short-Term Recovery. Sports Med. 2003;33:117–144. doi: 10.2165/00007256-200333020-00004. [PubMed] [CrossRef] [Google Scholar]
84. Biolo G., Williams B.D., Fleming R.Y., Wolfe R.R. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes. 1999;48:949–957. doi: 10.2337/diabetes.48.5.949. [PubMed] [CrossRef] [Google Scholar]
85. Greenhaff P.L., Karagounis L.G., Peirce N., Simpson E.J., Hazell M., Layfield R., Wackerhage H., Smith K., Atherton P., Selby A., et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am. J. Physiol. Metab. 2008;295:E595–E604. doi: 10.1152/ajpendo.90411.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
86. Glynn E.L., Fry C.S., Timmerman K.L., Drummond M.J., Volpi E., Rasmussen B.B., Leroy J.L., Gadsden P., De Cossío T.G., Gertler P. Addition of Carbohydrate or Alanine to an Essential Amino Acid Mixture Does Not Enhance Human Skeletal Muscle Protein Anabolism123. J. Nutr. 2013;143:307–314. doi: 10.3945/jn.112.168203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Koopman R., Beelen M., Stellingwerff T., Pennings B., Saris W.H.M., Kies A.K., Kuipers H., Van Loon L.J.C. Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am. J. Physiol. Metab. 2007;293:E833–E842. doi: 10.1152/ajpendo.00135.2007. [PubMed] [CrossRef] [Google Scholar]
88. Aragon A.A., Schoenfeld B.J. Nutrient timing revisited: Is there a post-exercise anabolic window? J. Int. Soc. Sports Nutr. 2013;10:5. doi: 10.1186/1550-2783-10-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
89. Jäger R., Kerksick C.M., Campbell B.I., Cribb P.J., Wells S.D., Skwiat T.M., Purpura M., Ziegenfuss T.N., Ferrando A.A., Arent S.M., et al. International Society of Sports Nutrition position stand: Protein and exercise. [(accessed on 25 March 2019)];J. Int. Soc. Sport. Nutr. 2017 4:20. doi: 10.1186/s12970-017-0177-8. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28642676 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Darrabie M.D., Arciniegas A.J.L., Mishra R., Bowles D.E., Jacobs D.O., Santacruz L. AMPK and substrate availability regulate creatine transport in cultured cardiomyocytes. Am. J. Physiol. Metab. 2011;300:870–876. doi: 10.1152/ajpendo.00554.2010. [PubMed] [CrossRef] [Google Scholar]
91. Purchas R., Busboom J., Wilkinson B. Changes in the forms of iron and in concentrations of taurine, carnosine, coenzyme Q10, and creatine in beef longissimus muscle with cooking and simulated stomach and duodenal digestion. Meat Sci. 2006;74:443–449. doi: 10.1016/j.meatsci.2006.03.015. [PubMed] [CrossRef] [Google Scholar]
92. Branch J.D. Effect of Creatine Supplementation on Body Composition and Performance: A Meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2003;13:198–226. doi: 10.1123/ijsnem.13.2.198. [PubMed] [CrossRef] [Google Scholar]
93. Hultman E., Söderlund K., Timmons J.A., Cederblad G., Greenhaff P.L. Muscle creatine loading in men. [(accessed on 25 March 2019)];J. Appl. Physiol. Soc. 1996 81:232–237. doi: 10.1152/jappl.1996.81.1.232. Available online: https://www.ncbi.nlm.nih.gov/pubmed/8828669 [PubMed] [CrossRef] [Google Scholar]
94. Jagim A.R., Oliver J.M., Sanchez A., Galvan E., Fluckey J., Riechman S., Greenwood M., Kelly K., Meininger C., Rasmussen C., et al. A buffered form of creatine does not promote greater changes in muscle creatine content, body composition, or training adaptations than creatine monohydrate. J. Int. Soc. Sports Nutr. 2012;9:43. doi: 10.1186/1550-2783-9-43. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
95. Spillane M., Schoch R., Cooke M., Harvey T., Greenwood M., Kreider R., Willoughby D.S., Cooke M. The effects of creatine ethyl ester supplementation combined with heavy resistance training on body composition, muscle performance, and serum and muscle creatine levels. J. Int. Soc. Sports Nutr. 2009;6:6. doi: 10.1186/1550-2783-6-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Childs E., De Wit H., Wit H. Subjective, behavioral, and physiological effects of acute caffeine in light, nondependent caffeine users. Psychopharmacology. 2006;185:514–523. doi: 10.1007/s00213-006-0341-3. [PubMed] [CrossRef] [Google Scholar]
97. Bellar D., Kamimori G.H., Glickman E.L. The Effects of Low-Dose Caffeine on Perceived Pain During a Grip to Exhaustion Task. J. Strength Cond. Res. 2011;25:1225–1228. doi: 10.1519/JSC.0b013e3181d9901f. [PubMed] [CrossRef] [Google Scholar]
98. Davis J.K., Green J.M. Caffeine and anaerobic performance: Ergogenic value and mechanisms of action. [(accessed on 25 March 2019)];Sport. Med. 2009 39:813–832. doi: 10.2165/11317770-000000000-00000. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19757860 [PubMed] [CrossRef] [Google Scholar]
99. Wickwire P.J., McLester J.R., Gendle S., Hudson G., Pritchett R.C., Laurent C.M., Green J.M. Effects of Caffeine on Repetitions to Failure and Ratings of Perceived Exertion during Resistance Training. Int. J. Sports Physiol. Perform. 2007;2:250–259. [PubMed] [Google Scholar]
100. Duncan M.J., Oxford S.W. The effect of caffeine ingestion on mood state and bench press performance to failure. [(accessed on 25 March 2019)];J. Strength Cond. Res. 2001 25:178–185. doi: 10.1519/JSC.0b013e318201bddb. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22124354 [PubMed] [CrossRef] [Google Scholar]
101. Williams A.D., Cribb P.J., Cooke M.B., Hayes A. The Effect of Ephedra and Caffeine on Maximal Strength and Power in Resistance-Trained Athletes. J. Strength Cond. Res. 2008;22:464–470. doi: 10.1519/JSC.0b013e3181660320. [PubMed] [CrossRef] [Google Scholar]
102. Tarnopolsky M.A., Atkinson S.A., MacDougall J.D., Sale D.G., Sutton J.R. Physiological responses to caffeine during endurance running in habitual caffeine users. Med. Sci. Sports Exerc. 1989;21:418–424. doi: 10.1249/00005768-198908000-00013. [PubMed] [CrossRef] [Google Scholar]
103. Blanchard J., Sawers S.J.A. The absolute bioavailability of caffeine in man. Eur. J. Clin. Pharmacol. 1983;24:93–98. doi: 10.1007/BF00613933. [PubMed] [CrossRef] [Google Scholar]
104. Hobson R.M., Saunders B., Ball G., Harris R.C., Sale C. Effects of β-alanine supplementation on exercise performance: A meta-analysis. Amino Acids. 2012;43:25–37. doi: 10.1007/s00726-011-1200-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
105. Hoffman J., Ratamess N.A., Ross R., Kang J., Magrelli J., Neese K., Faigenbaum A.D., Wise J.A. Beta-alanine and the hormonal response to exercise. [(accessed on 25 March 2019)];Int. J. Sports Med. 2008 29:952–958. doi: 10.1055/s-2008-1038678. Available online: https://www.ncbi.nlm.nih.gov/pubmed/18548362 [PubMed] [CrossRef] [Google Scholar]
106. Hoffman J., Ratamess N., Kang J., Mangine G., Faigenbaum A., Stout J. Effect of creatine and β-alanine supplementation on performance and endocrine responses in strength/power athletes. [(accessed on 25 March 2019)];Int. J. Sport Nutr. Exerc. Metab. 2006 16:430–446. doi: 10.1123/ijsnem.16.4.430. Available online: https://www.ncbi.nlm.nih.gov/pubmed/17136944 [PubMed] [CrossRef] [Google Scholar]
108. Wax B., Kavazis A.N., Weldon K., Sperlak J. Effects of Supplemental Citrulline Malate Ingestion During Repeated Bouts of Lower-Body Exercise in Advanced Weightlifters. J. Strength Cond. Res. 2015;29:786–792. doi: 10.1519/JSC.0000000000000670. [PubMed] [CrossRef] [Google Scholar]
109. Wax B., Kavazis A.N., Luckett W. Effects of Supplemental Citrulline-Malate Ingestion on Blood Lactate, Cardiovascular Dynamics and Resistance Exercise Performance in Trained Males. [(accessed on 25 March 2019)];J. Diet. 2016 13:269–282. doi: 10.3109/19390211.2015.1008615. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25674699 [PubMed] [CrossRef] [Google Scholar]
110. Glenn J.M., Gray M., Wethington L.N., Stone M.S., Stewart R.W., Jr., Moyen N.E. Acute citrulline malate supplementation improves upper- and lower-body submaximal weightlifting exercise performance in resistance-trained females. [(accessed on 25 March 2019)];Eur. J. Nutr. 2017 56:775–784. doi: 10.1007/s00394-015-1124-6. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26658899 [PubMed] [CrossRef] [Google Scholar]
111. Glenn J.M., Gray M., Jensen A., Stone M.S., Vincenzo J.L. Acute citrulline-malate supplementation improves maximal strength and anaerobic power in female, masters athletes tennis players. Eur. J. Sport Sci. 2016;16:1–9. doi: 10.1080/17461391.2016.1158321. [PubMed] [CrossRef] [Google Scholar]
112. Gonzalez A.M., Spitz R.W., Ghigiarelli J.J., Sell K.M., Mangine G.T. Acute Effect of Citrulline Malate Supplementation on Upper-Body Resistance Exercise Performance in Recreationally Resistance-Trained Men. J. Strength Cond. Res. 2018;32:3088–3094. doi: 10.1519/JSC.0000000000002373. [PubMed] [CrossRef] [Google Scholar]
113. Farney T.M., Bliss M.V., Hearon C.M., Salazar D.A. The Effect of Citrulline Malate Supplementation On Muscle Fatigue Among Healthy Participants. J. Strength Cond. Res. 2017:1. doi: 10.1519/JSC.0000000000002356. [PubMed] [CrossRef] [Google Scholar]
114. Trexler E.T., Persky A.M., Ryan E.D., Schwartz T.A., Stoner L., Smith-Ryan A.E. Acute Effects of Citrulline Supplementation on High-Intensity Strength and Power Performance: A Systematic Review and Meta-Analysis. Sports Med. 2019;49:707–718. doi: 10.1007/s40279-019-01091-z. [PubMed] [CrossRef] [Google Scholar]
115. Kleiner S.M., Bazzarre T.L., Litchford M.D. Metabolic profiles, diet, and health practices of championship male and female bodybuilders. J. Am. Diet. Assoc. 1990;90:962–967. [PubMed] [Google Scholar]
116. Kleiner S.M., Bazzarre T.L., Ainsworth B.E. Nutritional Status of Nationally Ranked Elite Bodybuilders. Int. J. Sport Nutr. 1994;4:54–69. doi: 10.1123/ijsn.4.1.54. [PubMed] [CrossRef] [Google Scholar]
117. Sandoval W.M., Heyward V.H. Food Selection Patterns of Bodybuilders. Int. J. Sport Nutr. 1991;1:61–68. doi: 10.1123/ijsn.1.1.61. [PubMed] [CrossRef] [Google Scholar]
118. Ismaeel A., Weems S., Willoughby D.S. A Comparison of the Nutrient Intakes of Macronutrient-Based Dieting and Strict Dieting Bodybuilders. Int. J. Sport Nutr. Exerc. Metab. 2018;28:502–508. doi: 10.1123/ijsnem.2017-0323. [PubMed] [CrossRef] [Google Scholar]
119. Nelson J.R., Raskin S. The eicosapentaenoic acid:arachidonic acid ratio and its clinical utility in cardiovascular disease. Postgrad. Med. 2019;131:268–277. doi: 10.1080/00325481.2019.1607414. [PubMed] [CrossRef] [Google Scholar]
120. Harris W.S. The Omega-6: Omega-3 ratio: A critical appraisal and possible successor. [(accessed on 15 June 2019)];Prostaglandins Leukot Essent Fatty Acids. 2018 132:34–40. doi: 10.1016/j.plefa.2018.03.003. Available online: https://www.ncbi.nlm.nih.gov/m/pubmed/29599053/ [PubMed] [CrossRef] [Google Scholar]
121. Tachtsis B., Camera D., Lacham-Kaplan O. Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration. Nutrients. 2018;10:309. doi: 10.3390/nu10030309. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
122. Di Girolamo F.G., Situlin R., Mazzucco S., Valentini R., Toigo G., Biolo G. Omega-3 fatty acids and protein metabolism: Enhancement of anabolic interventions for sarcopenia. [(accessed on 15 June 2019)];Curr. Opin. Clin. Nutr. Metab Care. 2014 17:145–150. doi: 10.1097/MCO.0000000000000032. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24500439 [PubMed] [CrossRef] [Google Scholar]
123. McGlory C., Wardle S.L., Macnaughton L.S., Witard O.C., Scott F., Dick J., Bell J.G., Phillips S.M., Galloway S.D.R., Hamilton D.L., et al. Fish oil supplementation suppresses resistance exercise and feeding-induced increases in anabolic signaling without affecting myofibrillar protein synthesis in young men. Physiol. Rep. 2016;4:e12715. doi: 10.14814/phy2.12715. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
124. Crestani D.M., Bonin E.F.R., Barbieri R.A., Zagatto A.M., Higino W.P., Milion F. Chronic supplementation of omega-3 can improve body composition and maximal strength, but does not change the resistance to neuromuscular fatigue. [(accessed on 15 June 2019)];Sport Sci. Health. 2017 13:259–265. doi: 10.1007/s11332-016-0322-9. Available online: https://link.springer.com/article/10.1007/s11332-016-0322-9 [CrossRef] [Google Scholar]
125. Lewis E.J.H., Radonic P.W., Wolever T.M.S., Wells G.D. 21 days of mammalian omega-3 fatty acid supplementation improves aspects of neuromuscular function and performance in male athletes compared to olive oil placebo. J. Int. Soc. Sports Nutr. 2015;12:28. doi: 10.1186/s12970-015-0089-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
126. Rossato L.T., Schoenfeld B.J., De Oliveira E.P. Is there sufficient evidence to supplement omega-3 fatty acids to increase muscle mass and strength in young and older adults? Clin. Nutr. 2019 doi: 10.1016/j.clnu.2019.01.001. [PubMed] [CrossRef] [Google Scholar]
127. Mocking R.J.T., Harmsen I., Assies J., Koeter M.W.J., Ruhé H.G., Schene A.H. Meta-analysis and meta-regression of omega-3 polyunsaturated fatty acid supplementation for major depressive disorder. Transl. Psychiatry. 2016;6:e756. doi: 10.1038/tp.2016.29. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
128. Maki K.C., Palacios O.M., Bell M., Toth P.P. Use of supplemental long-chain omega-3 fatty acids and risk for cardiac death: An updated meta-analysis and review of research gaps. J. Clin. Lipidol. 2017;11:1152–1160.e2. doi: 10.1016/j.jacl.2017.07.010. [PubMed] [CrossRef] [Google Scholar]
129. Miller P.E., Van Elswyk M., Alexander D.D. Long-Chain Omega-3 Fatty Acids Eicosapentaenoic Acid and Docosahexaenoic Acid and Blood Pressure: A Meta-Analysis of Randomized Controlled Trials. Am. J. Hypertens. 2014;27:885–896. doi: 10.1093/ajh/hpu024. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
130. Du S., Jin J., Fang W., Su Q. Does Fish Oil Have an Anti-Obesity Effect in Overweight/Obese Adults? A Meta-Analysis of Randomized Controlled Trials. PLoS ONE. 2015;10:e0142652. doi: 10.1371/journal.pone.0142652. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Non è per me raro discutere dell’argomento fertilità negli utilizzatori di AAS o nei soggetti in TRT. Complice una classe medica non sempre aggiornata, molti sono spinti a credere che una condizione di sterilità sia ineluttabile, tanto nei soggetti utilizzatori di dosi sovrafisiologiche di AAS quanto in quelli sottoposti a Terapia Sostitutiva del Testosterone [TRT].
A sottolineare questa eventualità ci ha pensato uno studio pubblicato nel 2019 nel quale veniva riportato che “la terapia con Testosterone è un contraccettivo, anche se di scarsa efficacia. Gli uomini in età riproduttiva con Testosterone basso devono essere informati degli effetti negativi della TRT sulla fertilità. Se la TRT viene prescritta a uomini interessati a preservare la fertilità, è opportuno proporre un’analisi del liquido seminale e l’eventuale crioconservazione dello sperma. Opzioni come il Clomifene Citrato e l’hCG, insieme al rinvio a un urologo della riproduzione, dovrebbero essere prese in considerazione per aumentare naturalmente i livelli di Testosterone negli uomini con testosterone basso che vogliono evitare la TRT.”
Immagine che spiega l’effetto contraccettivo del Testosterone esogeno. In sintesi, agisce attraverso due meccanismi: la diminuzione del Testosterone intratesticolare e l’inibizione della spermatogenesi. La maggior parte del Testosterone intra-testicolare è prodotto dalle cellule di Leydig nel testicolo. In presenza di Testosterone esogeno, esso inibisce la produzione di Ormone di Rilascio delle Gonadotropine (GnRH), che a sua volta inibisce la produzione di Ormone Luteinizzante (LH) e diminuisce la produzione endogena di Testosterone da parte delle cellule di Leydig, diminuendo la concentrazione di Testosterone intra-testicolare. L’inibizione della produzione di GnRH inibisce anche il rilascio dell’Ormone Follicolo-Stimolante (FSH), che compromette la spermatogenesi nelle cellule del Sertoli.
Quindi possiamo chiudere qui e liquidare la questione con un “si, anche in TRT si è destinati ad una condizione di sterilità”? Assolutamente no! Per quanto corretta nei punti espositivi, la conclusione di Amir Shahreza Patel et al. è incompleta. Per quale motivo? Ve lo spiegherò in questo articolo…
Breve panoramica sulla spermatogenesi:
Gli Steroidi Anabolizzanti Androgeni non influiscono solo sulla produzione endogena di Testosterone, ma anche sulla produzione di sperma, un processo chiamato spermatogenesi.
La spermatogenesi è strettamente regolata dalle cellule di Leydig e Sertoli del testicolo. Le cellule di Leydig producono Testosterone in risposta all’attivazione del recettore LHCG (LHCGR). Questo recettore è attivato dal legame con l’Ormone Luteinizzante (LH). Il Testosterone, a sua volta, agisce sulle cellule vicine, comprese le cellule del Sertoli, per controllare la spermatogenesi. L’attivazione del recettore dell’FSH (FSHR) sulle cellule del Sertoli controlla direttamente la spermatogenesi.
La produzione di spermatozoi avviene nei tubuli seminiferi e può essere suddivisa approssimativamente in tre fasi, come illustrato di seguito:
Le diverse fasi della spermatogenesi a partire da uno spermatogonio.
Tutte queste fasi si svolgono nei tubuli seminiferi. Durante la prima fase, gli spermatogoni migrano tra le cellule del Sertoli verso il lume dei tubuli. Mentre migrano lungo le cellule del Sertoli, questi spermatogoni si dividono lentamente e si differenziano in cellule spermatiche mature. In primo luogo, subiscono la mitosi, ossia la divisione in due cellule figlie identiche. Alcune di queste cellule figlie subiranno ulteriori modifiche e ingrandimenti, un processo noto come spermatocitogenesi, che darà origine agli spermatociti primari. Queste cellule, a loro volta, subiranno la meiosi. In questo caso, si verificano due divisioni cellulari consecutive, che danno origine a un totale di quattro cellule figlie. Ognuna di queste cellule avrà la metà del numero di cromosomi della cellula madre. Dopo la prima divisione cellulare chiamiamo queste cellule spermatociti secondari, mentre dopo la seconda divisione meiotica le chiamiamo spermatidi. Infine, gli spermatidi si differenziano in spermatozoi (spermatozoi maturi) durante la spermiogenesi.
L’intero processo di spermatogenesi richiede circa 74 giorni per essere completato [1]. Dopodiché, ci vorranno altri 1-21 giorni prima che gli spermatozoi finiscano nell’eiaculato [2]. Di conseguenza, quando la spermatogenesi si interrompe e riprende, ci vorrà un po’ di tempo prima che ciò si rifletta in un’analisi del liquido seminale.
La spermatogenesi dipende in larga misura dalla concentrazione di Testosterone intratesticolare (ITT). Poiché l’LH stimola i testicoli a produrre Testosterone e quindi è responsabile della concentrazione di ITT, l’LH è importante per la spermatogenesi. Normalmente, la concentrazione di ITT è circa 100 volte superiore a quella del sangue [3]. La somministrazione settimanale di 200mg di Testosterone Enantato la riduce notevolmente, fino a circa il 2% dei livelli basali. Sebbene non sia mai stato studiato nell’uomo, il limite inferiore della concentrazione di ITT necessaria per una spermatogenesi quantitativamente normale nei ratti è circa il 20% del livello basale [4]. Una volta scesi al di sotto di questo valore, esiste una chiara relazione tra il calo della concentrazione di ITT e la conta spermatica.
Breve parentesi su Estrogeni e fertilità:
Il Recettore α degli Estrogeni (ERα) è essenziale per la fertilità maschile. La sua attività è responsabile del mantenimento della citoarchitettura epiteliale nei dotti efferenti e del riassorbimento del liquido per la concentrazione degli spermatozoi nella testa dell’epididimo. Queste e altre scoperte hanno contribuito a stabilire il ruolo bisessuale degli estrogeni nell’importanza riproduttiva. È stato dimostrato che gli Estrogeni regolano l’espressione dello scambiatore Na+/H+-3 (NHE3) e la velocità di trasporto del 22Na+, sensibile a un inibitore di NHE3. Pertanto, nel maschio, gli estrogeni regolano uno dei più importanti trasportatori epiteliali di ioni e mantengono la differenziazione morfologica epiteliale nei dotti efferenti del maschio, indipendentemente dalla regolazione del trasporto di Na+.[https://www.pnas.org/]
17 β-estradiolo (E2) legato a ERα (giallo) e ERβ (blu). Solo due residui, cioè L384/M336 e M421/I373 (Erα/ERβ), differiscono nelle tasche di legame di ERα e ERβ. Non sorprende che l’E2 si leghi ai sottotipi in modo leggermente diverso.
Così come una concentrazione ottimale di E2 porta ad un miglioramento dei quadri della fertilità, livelli elevati di Estradiolo sono correlati all’infertilità maschile. Le cause dell’iperestrogenismo includono malattie della corteccia surrenale, del testicolo o uso di farmaci che influenzano l’asse ipotalamo-ipofisi-gonadi.[https://www.nature.com/]
Da notare che i dati raccolti hanno sollevato la possibilità di puntare sul ERα nello sviluppo di un contraccettivo per l’uomo.
AAS è soppressione della spermatogenesi:
E’ un dato di fatto che l’uso di AAS sopprime la produzione endogena di Testosterone. Lo fa attraverso un feedback negativo a livello dell’ipotalamo e dell’ipofisi. In breve, l’ipotalamo secerne un ormone chiamato Ormone di Rilascio delle Gonadotropine (GnRH) che viene rilasciato nel sistema portale ipofisario. Attraverso questo sistema, può raggiungere l’ipofisi anteriore. Qui, si legherà al suo recettore cognitivo che porterà alla secrezione di gonadotropine da parte dell’ipofisi anteriore. Queste gonadotropine, l’Ormone Luteinizzante (LH) e l’Ormone Follicolo-Stimolante (FSH), raggiungono la circolazione sistemica che le trasporta all’organo bersaglio: i testicoli. Il legame dell’LH al suo recettore specifico porta alla produzione di Testosterone. Il legame dell’FSH con il suo recettore specifico svolge un ruolo importante nella spermatogenesi. E, come descritto in precedenza, anche il Testosterone prodotto è fondamentale nella spermatogenesi.
Gli AAS inibiscono la secrezione di GnRH da parte dell’ipotalamo e la secrezione di gonadotropine da parte dell’ipofisi. Di conseguenza, sia la produzione di Testosterone che quella di spermatozoi vengono soppresse. Questo può portare a una condizione chiamata azoospermia, in cui non si trovano spermatozoi in un campione di sperma. Oppure può portare all’oligozoospermia, in cui la concentrazione di spermatozoi è molto bassa (inferiore a 15 milioni per mL o 39 milioni per eiaculato).[5]
Ad esempio, in uno studio, il 65% degli uomini è diventato azoospermico entro 6 mesi dalla somministrazione di Testosterone Enantato a 200mg settimanali [6]. Poiché l’LH e l’FSH non sono stati completamente soppressi (rispettivamente -66,7 e -62,5%), si può ipotizzare che un numero maggiore di uomini sarebbe diventato azoospermico con un dosaggio più elevato e più soppressivo. In effetti, in combinazione con un progestinico (che porterebbe a una più forte soppressione di LH e FSH), si registrano generalmente tassi di azoospermia di quasi il 90% [7]. Tuttavia, uno studio prospettico osservazionale (lo studio HAARLEM) che ha seguito 100 consumatori di AAS prima, durante e in due momenti successivi al ciclo di AAS, ha visto risultati simili a quelli dello studio in cui il 65% degli uomini è diventato azoospermico [8]. I dati relativi all’analisi dello sperma erano disponibili per 91 utilizzatori al termine del ciclo. Nonostante la soppressione praticamente totale di LH e FSH in quasi tutti gli utilizzatori, la concentrazione di spermatozoi era inferiore a 15 milioni per mL nel 68% degli utilizzatori (la conta totale degli spermatozoi era inferiore a 40 milioni nel 77%). Una differenza fondamentale in questo caso potrebbe essere il tempo di soppressione, in quanto l’altro studio ha mostrato il tasso cumulativo di azoospermia fino a 6 mesi, mentre gli utilizzatori di AAS si sono sottoposti a somministrazioni per periodi di tempo variabili, con una durata mediana di 13 settimane. Inoltre, alcuni dei consumatori di AAS hanno utilizzato l’hCG durante il ciclo, che potrebbe aver stimolato in qualche misura la spermatogenesi (tornerò su questo punto più avanti). Anche se gli autori scrivono: “(…) l’uso di hCG non ha avuto effetti rilevabili sulle dimensioni dei testicoli o sulla spermatogenesi”. Questo potrebbe essere attribuito a un sottodosaggio di hCG, a un uso non corretto o forse, in qualche misura, alla mancanza di potenza statistica. Infine, alti dosaggi di AAS – in modo del tutto casuale – potrebbero stimolare la spermatogenesi sostituendo una parte dell’attività androgena endogena mancante, come spiegato nella sezione precedente.
In ogni caso, è chiaro che l’uso di AAS di per se compromette in modo significativo la spermatogenesi.
Uso di AAS e atrofia testicolare: I testicoli comprendono il compartimento interstiziale, che ospita le cellule di Leydig, e il compartimento dei tubuli seminiferi, che ospita la spermatogenesi. Quest’ultimo è responsabile della maggior parte del volume del testicolo, con valori che in letteratura variano dal 60 al 90% [9, 10]. Gran parte di questo volume è costituito da cellule spermatiche in via di sviluppo. Di conseguenza, quando la spermatogenesi è compromessa, i testicoli diminuiscono di dimensioni. Ad esempio, lo studio citato in precedenza, in cui il 65% degli uomini è diventato azoospermico entro 6 mesi dalla somministrazione di Testosterone, ha visto una diminuzione del volume testicolare del 16,5% [6]. Uno studio in cui il Testosterone è stato combinato con un dosaggio molto basso di un progestinico orale (Levonorgestrel) per ottenere una soppressione più forte ha registrato una riduzione maggiore del volume testicolare, pari a circa il 30% [11]. Lo studio HAARLEM, citato in precedenza, ha registrato una riduzione del 24%. È interessante notare che i consumatori di AAS hanno visto il loro volume testicolare tornare a quello che era 3 mesi dopo la cessazione dell’uso (c’era solo una differenza del -4% a quel punto).
La terapia con gonadotropine (hCG e hMG/FSH) può preservare la spermatogenesi: L’effetto dell’hCG e dell’FSH o dell’hMG sulla spermatogenesi è forse dimostrato in modo più elegante da una serie di esperimenti di Matsumoto et al. [12]. In primo luogo, soggetti maschi sani hanno ricevuto 5000 UI di hCG due volte alla settimana per 7 mesi. Questo stimola fortemente la produzione di Testosterone da solo e di conseguenza l’FSH viene completamente soppresso. Ciononostante, è stata mantenuta una certa produzione di spermatozoi, la cui concentrazione è stata ridotta da 88 milioni/mL a 22 milioni/mL dopo 4 mesi. Dopo questi 7 mesi, il Testosterone Enantato (200mg settimanali) è stato aggiunto all’hCG per altri 6 mesi in questi uomini. Le concentrazioni di sperma sono rimaste praticamente inalterate (26 milioni/mL negli ultimi 3 mesi).
Dopo questo periodo, 4 soggetti hanno continuato l’hCG per altri 3 mesi senza Testosterone. Successivamente, in due dei soggetti è stato aggiunto l’FSH (100 UI al giorno) e negli altri due l’hMG (75 UI al giorno). L’aggiunta di FSH o hMG ha portato a un forte aumento della produzione di spermatozoi, raggiungendo una media di 103 milioni/mL negli ultimi 2 mesi:
Allo stesso modo, l’FSH da solo può preservare una parte della spermatogenesi durante la soppressione della terapia con testosterone, come illustrato nella figura seguente [13]:
Ciò che si può dedurre da questi risultati è che sia l’FSH che l’hCG possono preservare una certa spermatogenesi durante la soppressione delle gonadotropine da parte del Testosterone, ma che entrambi sono necessari per una spermatogenesi quantitativamente normale. Va notato, tuttavia, che ci sono state marcate differenze interindividuali. Nel precedente studio con hCG, un uomo è diventato azoospermico durante il trattamento con hCG.
Un piccolo studio retrospettivo suggerisce che l’hCG da solo, al dosaggio di 500 UI a giorni alterni, può preservare completamente la spermatogenesi in associazione alla Terapia Sostitutiva del Testosterone [14]. Forse in questi uomini c’era una secrezione residua di FSH sufficiente a consentire la piena conservazione della spermatogenesi. Inoltre, la natura retrospettiva dello studio potrebbe aver portato a una distorsione dei risultati.
Differenze tra hCG, LH e FSH.
Questo mi porta a un altro aspetto che vorrei discutere: il dosaggio. Uno studio ha rilevato che iniettando hCG al dosaggio di 250 UI a giorni alterni si ottiene una concentrazione di Testosterone intratesticolare praticamente uguale a quella del basale [15]. Dato il ruolo centrale del Testosterone intratesticolare nella spermatogenesi, si potrebbe sostenere che questo basso dosaggio dovrebbe essere sufficiente per preservare la spermatogenesi durante l’uso di AAS. Tuttavia, questo aspetto non è stato studiato direttamente in uno studio controllato.
L’hMG (chiamata anche Menotropina o Gonadotropina Umana della Menopausa – human Menopausal Gonadotropin), commercializzato in Italia sotto il nome di MENOGON ®, è un principio attivo per il trattamento dei disordini della fertilità. Si compone di gonadotropine che vengono estratte dalle urine di donne in post-menopausa, gonadotropine che sono solitamente l’Ormone Luteinizzante (LH) e l’Ormone Follicolo-Stimolante (FSH). Spesso, contiene anche Gonadotropina Corionica umana (hCG).
Un dosaggio più elevato, ma comunque inferiore a quello utilizzato negli studi di Matsumoto, ha dimostrato la conservazione di una certa spermatogenesi in pazienti con ipogonadismo secondario con hCG dosato a 500-2500 UI due volte alla settimana [16]. I dosaggi sono stati titolati in base ai livelli di Testosterone raggiunti. Per ottenere una spermatogenesi quantitativamente normale era necessaria l’aggiunta di FSH (3x 150 UI hMG settimanali). Anche in questo caso, però, si trattava di uno studio retrospettivo.
Infine, sono state sollevate alcune perplessità sull’effetto dell’hCG sulla morfologia degli spermatozoi. Uno studio finlandese suggerisce che l’uso concomitante di hCG e AAS ad alti dosaggi può avere un impatto negativo sulla morfologia dello sperma [17]. Lo studio ha seguito 18 atleti di forza amatoriali, 16 dei quali hanno utilizzato l’hCG insieme ad alti dosaggi di AAS. I campioni di sperma sono stati prelevati alla fine del ciclo di AAS, circa 1,5 mesi dopo il ciclo e circa 6 mesi dopo il ciclo. Naturalmente, la produzione di sperma era compromessa, con una conta media di 33 milioni di spermatozoi/mL alla fine del ciclo di AAS. Un soggetto è diventato azoospermico (e lo è rimasto per tutto il successivo periodo di sospensione). Ciò sembra dimostrare che l’uso di hCG può preservare una certa spermatogenesi durante l’uso di AAS. La morfologia dello sperma, tuttavia, era solo del 15% rispetto a una media del 40% di una coorte finlandese di donatori di banche del seme. Inoltre, hanno trovato una correlazione tra la dose totale di hCG utilizzata e gli spermatozoi morfologicamente anormali.
Quando hanno stratificato gli utilizzatori in due gruppi: un gruppo ad alta dose di hCG (>12.000 UI totali) e un gruppo a bassa dose (<12.000 UI totali), hanno notato che c’era una differenza significativa nella morfologia dello sperma tra i due. In media, il 22% era normale nel gruppo ad alto dosaggio e il 72% nel gruppo a basso dosaggio alla fine del ciclo di AAS. Ma come? Se la media del gruppo è del 15%, come può essere più alta sia nel gruppo ad alta dose che in quello a bassa dose? C’è qualcosa di sbagliato nei dati. Questo è un problema dello studio in questione. Da notare che, poiché il gruppo ad alta dose aveva una concentrazione di spermatozoi quasi cinque volte superiore, la quantità assoluta di spermatozoi morfologicamente normali era maggiore nel gruppo ad alta dose.
Si potrebbe obiettare che potrebbe essere l’assenza di FSH, piuttosto che l’hCG di per sé, ad avere un impatto sulla morfologia. Infatti, è stato riscontrato che dosi elevate di hCG migliorano la motilità degli spermatozoi e la morfologia normale in uomini subfertili con livelli normali di FSH [17]. Inoltre, si potrebbe sostenere che l’AAS stesso potrebbe avere un effetto negativo diretto sulla morfologia degli spermatozoi a dosi elevate [18]. Questo potrebbe non manifestarsi se vengono prodotte solo piccole quantità di spermatozoi, come nel caso del gruppo a basso dosaggio. Anche Matsumoto et al. hanno dimostrato che l’hCG (3x 5000 UI settimanali) non ha alcun effetto sulla morfologia degli spermatozoi in associazione al testosterone in un piccolo studio [19]. Infine, anche l’abuso di altre sostanze non dichiarate potrebbe aver avuto un impatto.
Conclusioni:
Ricapitolando, la spermatogenesi è strettamente regolata da LH e FSH. Quando si somministrano AAS, la secrezione di questi due ormoni viene fortemente ridotta. Di conseguenza, anche la spermatogenesi viene fortemente ridotta. Nella maggior parte degli uomini questo porta all’azoospermia. È stato riscontrato che l’uso di hCG mantiene una certa spermatogenesi, anche se a un livello inferiore al normale. L’aggiunta di FSH (direttamente o come parte di hMG) è necessaria per preservare completamente la spermatogenesi. Il dosaggio necessario per mantenere in modo ottimale la spermatogenesi con il solo hCG durante un ciclo AAS non è del tutto chiaro. Dato l’importante ruolo del Testosterone intratesticolare nel mantenimento della spermatogenesi, si potrebbe sostenere che un dosaggio che sostenga questo aspetto sostenga in modo ottimale anche la spermatogenesi. Si potrebbe quindi arrivare a un dosaggio di circa 250 UI a giorni alterni. Tuttavia, gli studi clinici (controllati) che hanno valutato direttamente l’impatto sulla spermatogenesi con la soppressione delle gonadotropine hanno tutti utilizzato dosaggi nettamente superiori. I dati di studi retrospettivi suggeriscono che potrebbero essere sufficienti da 500 a 2500 UI due volte alla settimana. L’ideale sarebbe testare il proprio sperma per capire quale sia il dosaggio più adatto. Si tenga presente che l’intero processo di spermatogenesi e la successiva comparsa di spermatozoi nell’eiaculato possono richiedere fino a circa 3 mesi. I cambiamenti nella terapia potrebbero quindi richiedere almeno 3 mesi prima che i loro effetti si riflettano nell’analisi dello sperma.
Dopo le informazioni fin qui riportate, non ci si stupisce del fatto che nello studio citato nell’introduzione la TRT fosse stata classificata come un “contraccettivo di bassa efficacia”. Se infatti togliamo dall’equazione la somministrazione esogena di hCG e/o FSH [o in alternativa hMG], la condizione di azoospermia è praticamente una certezza. Da considerarsi anche i dosaggi di questi ancillari della TRT. Dosaggi che devono tenere conto della risposta terapeutica soggettiva. Ciò significa che i dosaggi standard per l’hCG, per esempio, rappresentano per la maggior parte dei soggetti solo un punto di partenza che dovrà essere riconsiderato alla luce di esami specifici [vedi spermiogramma].
Un protocollo di fertilità nel quale mi sono imbattuto spesso parlando con atleti o preparatori d’oltre oceano è costituito da una hCG, hMG e Clomifene Citrato. L'”invenzione” di questo protocollo si attribuisce a Dave Palumbo. Non propriamente un luminare dell’endocrinologia ma sicuramente un vagliatore di tester non da poco.
Il protocollo è il seguente:
hCG – 2000 UI a giorni alterni hMG – 75 UI a giorni alterni Clomifene Citrato – 50mg al giorno
Clomifene Citrato
In alternativa all’uso di Clomifene Citrato si opta per Enclomifene Citrato, l’isomero trans del Clomifene Citrato. Ma di lui parlerò in un articolo apposito. Rimane comunque il dubbio di una loro sufficiente efficacia additiva.
Encolimfene Citrato
Ho avuto l’opportunità di raccogliere molte testimonianze di utilizzatori. Alcuni di loro hanno trascorso più di un decennio tra cicli, bridge e fasi in TRT. Anche nei casi più estremi, quando è arrivato il momento di avere un figlio, una parte consistente di quelli che hanno seguito questo semplice protocollo sono riusciti a ingravidare la propria moglie/fidanzata:
La sospensione temporanea della TRT per seguire un protocollo di ristabilizzazione dell’Asse HPT e aumentare il numero di spermatozoi con il protocollo di fertilità, non rappresentava una costante ma una eventualità che poteva interessare alcuni individui .
Ovviamente, quanto detto non rappresenta assolutamente una prescrizione medica o un consiglio terapeutico! Si tratta, come sempre, di pura divulgazione scientifica volta alla formazione di una cultura di base utile alla tutela della propria e altrui salute.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
“Book on Steroids” di Peter Bond [capitolo 6 – Side effects and managing them – sezione 6.12. – Low/undetectable sperm count (oligo-/azoospermia)].
Amann, Rupert P. “The cycle of the seminiferous epithelium in humans: a need to revisit?.” Journal of andrology 29.5 (2008): 469-487.
Rowley, Mavis J., Florence Teshima, and Carl G. Heller. “Duration of transit of spermatozoa through the human male ductular system.” Fertility and sterility 21.5 (1970): 390-396.
McLachlan, Robert I., et al. “Effects of testosterone plus medroxyprogesterone acetate on semen quality, reproductive hormones, and germ cell populations in normal young men.” The Journal of Clinical Endocrinology & Metabolism 87.2 (2002): 546-556.
Zirkin, Barry R., et al. “Maintenance of advanced spermatogenic cells in the adult rat testis: quantitative relationship to testosterone concentration within the testis.” Endocrinology 124.6 (1989): 3043-3049.
T. G. Cooper, E. Noonan, S. Von Eckardstein, J. Auger, H. Baker, H. M. Behre, T. B. Haugen, T. Kruger, C. Wang, M. T. Mbizvo, et al. World health organization reference values for human semen characteristics. Human reproduction update, 16(3):231–245, 2010.
W. H. O. T. F. on Methods for the Regulation of Male Fertility. Contraceptive efficacy of testosterone-induced azoospermia in normal men. The Lancet, 336(8721):955–959, 1990.
Page, Stephanie T., John K. Amory, and William J. Bremner. “Advances in male contraception.” Endocrine reviews 29.4 (2008): 465-493.
Smit, D. L., et al. “Disruption and recovery of testicular function during and after androgen abuse: the HAARLEM study.” Human Reproduction 36.4 (2021): 880-890.
S. Melmed. Williams textbook of endocrinology. 13th edition. Elsevier Health Sciences, 2016.
M. Simoni and I. T. Huhtaniemi. Endocrinology of the Testis and Male Reproduction. Springer, 2017.
Anawalt, Bradley D., et al. “Intramuscular testosterone enanthate plus very low dosage oral levonorgestrel suppresses spermatogenesis without causing weight gain in normal young men: a randomized clinical trial.” Journal of andrology 26.3 (2005): 405-413.
Matsumoto, Alvin M., Anthony E. Karpas, and William J. Bremner. “Chronic human chorionic gonadotropin administration in normal men: evidence that follicle-stimulating hormone is necessary for the maintenance of quantitatively normal spermatogenesis in man.” The Journal of Clinical Endocrinology & Metabolism 62.6 (1986): 1184-1192.
Matsumoto, Alvin M., et al. “Reinitiation of sperm production in gonadotropin-suppressed normal men by administration of follicle-stimulating hormone.” The Journal of clinical investigation 72.3 (1983): 1005-1015.
Hsieh, Tung-Chin, et al. “Concomitant intramuscular human chorionic gonadotropin preserves spermatogenesis in men undergoing testosterone replacement therapy.” The Journal of urology 189.2 (2013): 647-650.
Coviello, Andrea D., et al. “Low-dose human chorionic gonadotropin maintains intratesticular testosterone in normal men with testosterone-induced gonadotropin suppression.” The Journal of Clinical Endocrinology & Metabolism 90.5 (2005): 2595-2602.
Depenbusch, Marion, et al. “Maintenance of spermatogenesis in hypogonadotropic hypogonadal men with human chorionic gonadotropin alone.” European journal of endocrinology 147.5 (2002): 617-624.
Homonnai, Z. T., M. Peled, and G. F. Paz. “Changes in semen quality and fertility in response to endocrine treatment of subfertile men.” Gynecologic and obstetric investigation 9.5 (1978): 244-255.
Torres-Calleja, J., et al. “Effect of androgenic anabolic steroids on sperm quality and serum hormone levels in adult male bodybuilders.” Life sciences 68.15 (2001): 1769-1774.
Matsumoto, Alvin M., et al. “Human chorionic gonadotropin and testicular function: stimulation of testosterone, testosterone precursors, and sperm production despite high estradiol levels.” The Journal of Clinical Endocrinology & Metabolism 56.4 (1983): 720-728.
Introduzione alla questione “High-Low gainer/responder”:
I fattori alla base dell’eterogeneità dell’ipertrofia muscolare in seguito all’allenamento contro-resistenza (RET) rimangono in gran parte sconosciuti. E la questione ormonale connessa è senza dubbio una parte poco conosciuta e spesso fraintesa.
Tutti i frequentatori di sala pesi, improvvisati o meno, hanno sentito parlare almeno una volta di “High gainers/responders” e di “Low gainers/responders” in riferimento alla possibilità individuale di aumento dell’ipertrofia muscolo-scheletrica. In teoria, possiamo classificare 7 tipi di “gainers/responders” diversi:
Se si osserva il modello di distribuzione normale (curva gaussiana), si possono creare sette diversi livelli di “guadagnatori”.
1.Non-Responders.
Coloro che guadagneranno una quantità insignificante di muscoli, anche se stanno facendo tutto correttamente. Queste persone sono estremamente rare e rappresentano circa lo 0,1% della popolazione.
2. Very Low Responders. Coloro che sono in grado di aggiungere solo una piccola quantità di muscoli nel corso della loro carriera… e a un ritmo faticosamente lento. Anche loro non sono comuni: rappresentano circa il 2,1% della popolazione. È probabile che possano guadagnare solo 2.5-5Kg di muscoli durante la loro carriera di sollevatori (da 1.5 a 3Kg per le donne).
3. Low Responders. Insieme ai non-responder e ai very low responders, questi soggetti completano la categoria dei veri hardgainer. Questi soggetti hanno guadagni muscolari molto lenti e di solito devono accettare di aggiungere una buona quantità di grasso per aumentare la massa muscolare. Sono i più comuni hardgainer, circa il 13,6% della popolazione. Possono guadagnare 5-10Kg di muscoli nel corso della loro carriera di sollevatori (3-6Kg per le donne).
4.Normal Responders. È molto probabile che voi facciate parte di questa categoria. Questo gruppo rappresenta quasi il 70% della popolazione. Anche se ci saranno differenze nel potenziale di crescita muscolare all’interno di questo gruppo, tutti possono guadagnare una discreta quantità di muscoli se si allenano, mangiano e riposano correttamente. Gli uomini di questa categoria possono sperare di aumentare la massa muscolare tra i 10 e i 18Kg rispetto al loro peso da adulti senza allenamento. Le donne si avvicinano a 6-9Kg.
5. Easy Gainers. Nel corso della loro carriera di sollevatori, questo gruppo può guadagnare il 15-20% di muscoli in più – 2.5-4Kg in più rispetto a un normale responder. Inoltre, possono aumentare più velocemente. Il loro potenziale di aumento muscolare potrebbe essere di 15-20Kg (8-11Kg per le donne). Rappresentano circa il 13,6% della popolazione.
6. Very Easy Gainers. Questi soggetti spesso appaiono muscolosi già prima di iniziare ad allenarsi in sala pesi. E quando iniziano ad allenarsi, rispondono rapidamente e possono guadagnare un altro 10% di muscoli, per un potenziale di crescita muscolare totale di circa 16-24Kg (9-12Kg per le donne).
7. Freaks. Questi soggetti sono sempre muscolosi e/o forti (e spesso esplosivi) prima ancora di mettere piede in palestra. Sono i “veri naturl” che, prima di iniziare ad assumere PEDs finiscono per assomigliare a chi ne fa già uso. Ma rappresentano lo 0,1% della popolazione, il che significa che la maggior parte degli “influencer” che affermano di avere una buona genetica e non di usare farmaci, stanno mentendo.
Nota:la quantità di potenziale di crescita muscolare può sembrare bassa, ma è chiaro che non sto parlando di peso corporeo. Ogni chilo di aumento muscolare porta normalmente a un aumento di 0,25-0,5 chili di “qualcos’altro” senza aggiungere grasso corporeo. Un aumento muscolare di 13.5Kg porterebbe in realtà a un aumento della massa magra da 16.8Kg a 20.4Kg sulla bilancia.
Ma quali sono le determinanti genetiche che separano una “High gainer/responder” da un “Low gainer/responder”?
In ordine di importanza teorica:
GENOTIPO ACTN3 Senza entrare troppo nel merito, esistono due genotipi ACTN3 “puri”: ACTN3 RR e ACTN3 XX. Esistono anche tipi misti. Il tipo di ACTN3 determina diversi elementi che svolgono un ruolo importante nel potenziale di crescita muscolare.
Rapporto tra fibre a contrazione rapida e lenta. Un maggior numero di fibre a contrazione rapida significa un maggior potenziale di crescita e di forza. Livello di attivazione del mTOR. Più si riesce ad attivare l’mTOR dopo l’allenamento e i pasti, più si aumenta la sintesi proteica e più si può crescere. Riparazione del danno muscolare. Più lenta è la riparazione, meno ci si può allenare proficuamente e più è difficile far crescere nuovo tessuto contrattile. Il tipo ACTN3 RR presenta un maggior numero di fibre a contrazione rapida, una maggiore attivazione del mTOR e una rapida riparazione del danno muscolare. Tutto ciò favorisce una crescita muscolare più rapida.
All’opposto, ACTN3 XX significa meno fibre a contrazione rapida, minore attivazione del mTOR e riparazione lenta dei danni muscolari. Ma hanno un VO2 max naturale più elevato e sono più resistenti all’affaticamento muscolare.
Espressione della Miostatina
Sicuramente molti di voi avranno visto le foto degli esemplari di Belgian Blue, una razza di bovini da carne del Belgio la cui caratteristica peculiare sono le accentuate masse muscolari. Non si tratta di un esperimento in cui le mucche vengono sottoposte a dosi massicce di steroidi anabolizzanti, ma semplicemente di una razza di bovini nati senza la capacità di produrre Miostatina.
La Miostatina è una miochina (una proteina rilasciata dai muscoli). Agisce come un fattore limitante nella quantità di muscoli che si possono sviluppare. Più ci si avvicina al proprio potenziale genetico, più la Miostatina limiterà la crescita muscolare.
Alcune persone hanno naturalmente livelli di Miostatina più elevati, quindi il loro tasso di crescita muscolare totale risulta inferiori con un tasso catabolico più accentuato.
Le persone con meno Miostatina possono sviluppare più muscoli e più rapidamente. Sembra anche che siano più a rischio di strappi muscolari.
Numero, densità e sensibilità recettoriale Sebbene diversi fattori possano influenzare i livelli di Testosterone, IGF-1 e Ormone della Crescita (alimentazione, stress, sonno, ecc.), alcune persone hanno un numero e potenziale di espressività dei recettori ormone specifici (es. AR) maggiore rispetto alla norma. “Natural” o “Doped”, in entrambi i casi, le persone con questa caratteristica hanno un maggiore potenziale di crescita muscolare grazie a una maggiore sintesi proteica indotta dalla risposta ormone-recettoriale.
Questi articoli saranno però incentrati sull’analisi dei due fattori ormone-genici determinanti l’ipertrofia del muscolo-scheletrico: l’espressività recettoriale e la mutazione del gene della Miostatina.
In questa 1° parte tratterò dell’espressione recettoriale.
Introduzione al “Fattore Recettoriale”:
In un interessante studio del 2018 [1] Sono stati esaminati gli ormoni circolanti, gli ormoni intramuscolari e le variabili correlate agli ormoni intramuscolari in uomini allenati alla resistenza prima e dopo 12 settimane di RET. L’eliminazione a ritroso e la regressione delle componenti principali hanno valutato la significatività statistica degli ormoni anabolici circolanti proposti (ad esempio, Testosterone, Testosterone libero, Deidroepiandrosterone, Diidrotestosterone, Fattore di Crescita Insulino-Simile-1, Fattore di Crescita Insulino-Simile-1 libero, Ormone Luteinizzante e Ormone della Crescita) e i cambiamenti della massa muscolare indotti dalla RET (n = 49). Sono stati utilizzati immunoblots e immunodosaggi per valutare i livelli di Testosterone libero intramuscolare, i livelli di Diidrotestosterone, l’espressione della 5α-reduttasi e il contenuto del Recettore degli Androgeni nei soggetti che hanno risposto in modo più elevato (HIR; n = 10) e più basso (LOR; n = 10) alle 12 settimane di RET. Nessun ormone misurato prima dell’esercizio, dopo l’esercizio, prima dell’intervento o dopo l’intervento è risultato costantemente significativo o selezionato nel modello finale per la variazione di: area trasversale di tipo 1 (CSA), CSA di tipo 2 o massa grassa e ossea (LBM). L’analisi delle componenti principali non ha portato a una grande riduzione delle dimensioni e la regressione delle componenti principali non è stata più efficace delle analisi di regressione non aggiustate. Nessun ormone misurato nel sangue o nel muscolo è risultato diverso tra HIR e LOR. L’enzima steroidogenico 5α-reduttasi è aumentato dopo la RET nell’HIR (P < 0,01) ma non nel LOR (P = 0,32). Il contenuto di recettori per gli androgeni è rimasto invariato con la RET, ma è stato più elevato in ogni momento nell’HIR. A differenza del Testosterone libero intramuscolare, del Diidrotestosterone o della 5α-reduttasi, è stata riscontrata una relazione lineare tra il contenuto dei recettori degli androgeni e la variazione della LBM (P < 0,01), del CSA di tipo 1 (P < 0,05) e del CSA di tipo 2 (P < 0,01) sia prima che dopo l’intervento. Questi risultati indicano che il contenuto intramuscolare di recettori per gli androgeni, ma non gli ormoni circolanti o intramuscolari (o gli enzimi che ne regolano la produzione intramuscolare), influenzano l’ipertrofia del muscolo scheletrico dopo la RET in giovani uomini precedentemente allenati.
Variabili nell’ipertrofia indotta da RET e livelli ormonali:
Esiste una sostanziale variabilità individuale nell’ipertrofia muscolare scheletrica indotta da RET (Hubal et al., 2005; Davidsen et al., 2011). Si ritiene che l’aumento post-esercizio degli ormoni circolanti, presumibilmente anabolici (ad esempio, T, GH e IGF-1), sia causale nel determinare l’ipertrofia muscolare scheletrica indotta da RET (Kraemer et al., 2017; Mangine et al., 2017). Tuttavia, esistono sostanziali prove contrarie di un ruolo causale, o addirittura correlato (cioè che condivide una varianza comune) di tali ormoni sia nell’aumento della sintesi proteica muscolare indotto da RET (West et al., 2009) sia nell’ipertrofia (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016; Mobley et al., 2018).
È plausibile che, al contrario degli ormoni circolanti a livello sistemico, l’androgenesi locale intramuscolare possa mediare l’ipertrofia muscolare indotta da RET, come è stato proposto per gli uomini anziani (Sato et al., 2014). Inoltre, l’aumento del contenuto di recettori androgeni intramuscolari indotto da RET è stato significativamente correlato all’ipertrofia muscolare indotta da RET (Ahtiainen et al., 2011; Mitchell et al., 2013). Pertanto, è possibile che un aumento degli androgeni intramuscolari e/o dei loro recettori, attraverso un meccanismo autocrino, sia importante nel determinare l’ipertrofia indotta da RET.
Lo scopo dello studio di base trattato in questo articolo [1] è stata quella di determinare se l’eterogeneità dell’ipertrofia del muscolo scheletrico indotta da RET, misurata mediante indici multipli, fosse associata agli ormoni circolanti, agli ormoni intramuscolari, al contenuto di enzimi steroidogenici intramuscolari o al contenuto di recettori per gli androgeni. Sono stati eseguite ulteriori analisi statistiche e intramuscolari sui dati di uno studio precedente condotto su uomini sani e allenati contro-resistenza (n = 49; Morton et al., 2016). Per esplorare ulteriormente la relazione tra ormoni sistemici e ipertrofia, è stato utilizzato l’eliminazione a ritroso e la regressione delle componenti principali sulle concentrazioni di ormoni sistemici sia a riposo che dopo l’esercizio contro-resistenza con gli indici di ipertrofia come variabili di esito separate in tutti i partecipanti. Per valutare l’importanza dell’androgenesi intramuscolare, abbiamo completato un’analisi solo sui rispondenti più alti (HIR – quintile superiore) e più bassi (LOR – quintile inferiore) che comprendeva la valutazione del T intramuscolare, del DHT, dell’espressione della 5α-reduttasi e del contenuto del recettore degli androgeni. Coerentemente con il lavoro precedente (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016), i ricercatori hanno ipotizzato che gli ormoni sistemici circolanti non fossero correlati a nessuna misura di ipertrofia; tuttavia, hanno ipotizzato, visti i risultati precedenti (Ahtiainen et al., 2011; Mitchell et al., 2013), che il contenuto di recettori per gli androgeni fosse associato all’ipertrofia indotta da RET.
Partecipanti e intervento con allenamento contro-resistenza:
Quarantanove giovani uomini allenati alla resistenza (eseguendo RET almeno 2 giorni/settimana [range 3-6 giorni/settimana] per 4 ± 6 anni) si sono offerti volontari per questo studio. In breve, i partecipanti sono stati assegnati in modo casuale a un gruppo ad alte ripetizioni (HR) o a basse ripetizioni (LR). Il gruppo HR ha eseguito tutti gli esercizi con una resistenza relativamente leggera [∼30-50% del loro massimo di ripetizioni (RM)] fino al cedimento volitivo (20-25 ripetizioni) e il gruppo LR ha eseguito tutti gli esercizi con una resistenza relativamente pesante (∼75-90% RM), anch’essi fino al cedimento volitivo (8-12 ripetizioni). Ogni partecipante è stato sottoposto a un intervento RET di 12 settimane in cui ha eseguito RET su tutto il corpo per 4 giorni a settimana e ha ricevuto 30g di proteine isolate del siero di latte due volte al giorno (BioPRO; Davisco Foods International, Le Sueur, MN, Stati Uniti).
Prelievo di sangue e analisi ormonali:
Il giorno del test pre e post intervento è stato eseguito dopo un digiuno notturno alla stessa ora del giorno per ogni partecipante. Ogni partecipante ha eseguito un allenamento acuto contro-resistenza nell’ambito del gruppo designato (HR o LR) e il sangue è stato prelevato da un catetere endovenoso inserito in una vena antecubitale. Due provette vacutainer da 4 ml (Becton, Dickinson and Company, Franklin Lakes, NJ, Stati Uniti) sono state prelevate prima dell’esercizio e dopo 0, 15, 30 e 60 minuti dall’esercizio. Una provetta da 4 mL è stata lasciata coagulare per 30 minuti a temperatura ambiente per isolare successivamente il siero e l’altra è stata eparinizzata per isolare successivamente il plasma. L’analisi del campione di sangue è stata eseguita in cieco per: Cortisolo (nM), LH (IU/L), Lattato (mM), DHEA (ng/mL), T (ng/mL), T libero (fT; ng/dL; cioè, Testosterone non legato alla globulina legante gli ormoni sessuali o all’albumina nel sangue), DHT (ng/mL) e GH (ng/mL) utilizzando test immunometrici a chemiluminescenza in fase solida a due siti (Immulite 2000 Immunoassay System; Siemens Healthineers, Erlangen, Germania) e IGF-1 (μg/dL) e IGF-1 libero (fIGF-1; ng/mL) utilizzando radio-immunoassaggi (Diagnostics Products Corporation, Los Angeles, CA, Stati Uniti). L’AUC a 60 minuti dall’esercizio di resistenza è stata calcolata per ciascun ormone, utilizzando la regola trapezoidale, con punti temporali a 0, 15, 30 e 60 minuti.
Regressioni a scalare:
I dati di HR e LR sono stati eliminati a causa della mancanza di differenze tra i gruppi per quanto riguarda gli ormoni circolanti e gli esiti (Morton et al., 2016). Gli esiti considerati sono stati CSA delle fibre di tipo 1, CSA delle fibre di tipo 2 e massa corporea (LBM) priva di grasso e ossa. Ciascun risultato in ciascun momento della misurazione (ossia, la variazione, i valori assoluti prima e dopo l’intervento) è stato regredito rispetto agli ormoni di ciascun punto temporale: AUC pre-intervento a riposo, AUC post-esercizio pre-intervento, AUC post-intervento a riposo e AUC post-esercizio post-intervento. Per scegliere il modello finale è stata utilizzata l’eliminazione a ritroso, con il criterio di eliminazione Akaike Information Criterion (AIC). I valori di AUC post-esercizio utilizzati nell’analisi non hanno sottratto le concentrazioni a riposo. Tuttavia, abbiamo eseguito l’analisi con le concentrazioni a riposo sottratte dai valori grezzi dell’AUC e non abbiamo riscontrato differenze sostanziali nei risultati.
Analisi immunoblot:
Come descritto in precedenza (Aizawa et al., 2010), dopo l’omogenizzazione, la concentrazione proteica del surnatante risultante è stata determinata mediante un saggio proteico di Bradford e le proteine muscolari (sia citoplasmatiche che nucleari, 20μg di proteine) sono state separate su gel di SDS-poliacrilammide al 10% e poi trasferite su membrane di polivinilidene difluoruro (Millipore, Billerica, MA, Stati Uniti). Le membrane sono state bloccate per 1 ora con tampone bloccante (5% latte scremato in soluzione salina tamponata con fosfato e 0,1% Tween 20) e quindi incubate per 12 ore a 4°C con anticorpi primari contro il recettore degli androgeni (#3202, Cell Signaling Technology, Beverly, MA, Stati Uniti) e la 5α-reduttasi (H00006715, Abnova, Taipei, Taiwan) diluiti a 1:1000 in tampone bloccante. Le membrane sono state lavate tre volte con PBST prima di essere incubate per 1 ora con un anticorpo secondario coniugato con perossidasi di rafano e immunoglobulina anti-rabbit (#7074, Cell Signaling Technology, Beverly, MA, Stati Uniti) diluito a 1:3000 nel tampone di blocco. Le membrane sono state poi lavate tre volte con PBST. Le proteine sono state rilevate con un sistema di chemiluminescenza potenziata plus (GE Healthcare Biosciences) e visualizzate su un imager LAS4000 (GE Healthcare Biosciences). L’intensità delle bande è stata quantificata utilizzando ImageJ versione 1.46 (National Institutes of Health, Bethesda, MD, Stati Uniti).
Test immunoenzimatici per gli ormoni intramuscolari:
Il campione di muscolo è stato omogeneizzato con lo stesso metodo dell’analisi immunoblot. I livelli di T e DHT nel muscolo scheletrico sono stati determinati utilizzando un kit per il dosaggio immunoenzimatico, dopo essere stati diluiti 200 volte con ciascun tampone di dosaggio come precedentemente descritto (Horii et al., 2016). Gli anticorpi policlonali immobilizzati sono stati sollevati contro il T (Cayman Chemical, Ann Arbor, MI, Stati Uniti) e il DHT (IBL Hamburg, Germania) prima dell’aggiunta di anticorpi secondari alla perossidasi di rafano. La densità ottica a 450 nm è stata qualificata su un lettore di micropiastre (BioLumin 960; Molecular Dynamics, Tokyo, Giappone) e le analisi sono state eseguite in duplicato. Il valore del coefficiente di variazione era 3,0 e r2 = 0,974 nel presente studio. I ricercatori che hanno eseguito le analisi intramuscolari (KS e SF) non erano ciechi rispetto ai campioni HIR e LOR.
Analisi delle componenti principali e regressione:
I dati sono stati centrati e scalati prima di eseguire l’analisi delle componenti principali (PCA) sugli ormoni di ciascun momento della misurazione (riposo pre-intervento, AUC post-esercizio pre-intervento, riposo post-intervento e AUC post-esercizio post-intervento). Lo scopo della PCA è quello di utilizzare la trasformazione ortogonale per creare un insieme di nuove variabili lineari e non correlate (componenti principali), di cui viene preso un sottoinsieme che rappresenta effettivamente la maggior parte della variabilità osservata nei dati originali. In definitiva, queste componenti principali sono combinazioni lineari delle variabili originali (ad esempio, gli ormoni) che vengono poi utilizzate come covariate nelle analisi di regressione. Presentiamo qui la PCA sotto forma di scree plot. L’eliminazione a ritroso è stata eseguita sulle componenti principali (cioè la regressione delle componenti principali) utilizzando l’AIC come criterio di adattamento del modello. La PCA e la regressione delle componenti principali sono state eseguite in R (R Core Team, 2017).
High- vs. Low-Responders:
Le biopsie del muscolo scheletrico del vasto laterale di ciascun partecipante e la DXA sono state utilizzate per valutare la variazione della CSA delle fibre (sia di tipo 1 che di tipo 2) e della LBM, rispettivamente, come descritto in dettaglio altrove (Morton et al., 2016). La determinazione dell’HIR e del LOR è stata effettuata classificando individualmente (da 1 a 49) la variazione di ciascun risultato per ogni partecipante e quindi calcolando la media della classifica di ciascun partecipante per tutti e tre i risultati (CSA di tipo 1, CSA di tipo 2 e LBM). Con una probabilità di errore di tipo II (alfa) di 0,05, una probabilità di errore di tipo I (beta) di 0,20 e una differenza attesa relativamente moderata nelle variazioni della massa muscolare indotte dalla RET tra HIR e LOR (dimensione dell’effetto, f = 0,60), il calcolo delle dimensioni del campione a priori richiedeva 18 partecipanti (nove in ciascun gruppo). Pertanto, il quintile superiore (n = 10) dei partecipanti classificati è stato classificato come HIR e il quintile inferiore (n = 10) dei partecipanti classificati è stato classificato come LOR. Le analisi statistiche tra HIR e LOR sono state eseguite utilizzando SPSS (versione 22.0, Chicago, IL, Stati Uniti). Il CSA di tipo 1, il CSA di tipo 2, la LBM e tutti i dati relativi agli ormoni intramuscolari sono stati analizzati utilizzando un’analisi della varianza a due fattori (gruppo × tempo) a misure ripetute (ANOVA) con il gruppo (HIR vs. LOR) e il tempo (pre- e post-intervento) come variabili sperimentali. Se indicato, sono stati eseguiti t-test indipendenti a due code per valutare eventuali differenze tra i gruppi in uno specifico punto temporale (ad esempio, la T intramuscolare pre-intervento). Le correlazioni tra i risultati intramuscolari e la variazione di CSA di tipo 1, CSA di tipo 2 e LBM sono state eseguite in SPSS (versione 22.0, Chicago, IL, Stati Uniti). La significatività statistica è stata accettata quando P < 0,05. I dati sono presentati come diagrammi a riquadro e a baffi (comprendenti la mediana [linea], la media [croce], l’intervallo interquartile [riquadro] e i valori minimo e massimo [baffi]) nelle Figure 1 e 3 e media ± SD nel testo e nelle tabelle.
Variazione della massa muscolare in tutti i partecipanti (in alto) e in HIR e LOR (in basso). Pannelli superiori: La variazione di (A) CSA di tipo 1, (B) CSA di tipo 2 e (C) LBM di tutti i 49 partecipanti. Pannelli inferiori: La variazione di (D) CSA di tipo 1, (E) CSA di tipo 2 e (F) LBM classificata in HIR e LOR. I valori sono presentati come mediana (linee) con intervallo interquartile (riquadri), range (minimo e massimo) e media (croce). ∗Differenza significativa tra soggetti ad alta e bassa risposta (P < 0,01). Pannelli A-C adattati da Morton et al. (2016).
Risultati dello studio
Cambiamenti nella massa muscolare con allenamenti contro-resistenza:
Sono stati reclutati 56 partecipanti e 49 hanno completato l’intero intervento (HR: n = 24, LR: n = 25; 23 ± 2 anni, 86 ± 5 kg, 181 ± 6 cm). Due persone hanno abbandonato il gruppo LR a causa di un trasferimento di lavoro e di un infortunio non legato all’intervento, mentre cinque persone hanno abbandonato il gruppo HR a causa di un cambiamento di sede o di un infortunio non legato all’intervento. Dodici settimane di RET hanno portato a un aumento della CSA di tipo 1 (665 ± 149 μm2), della CSA di tipo 2 (978 ± 189 μm2) e della LBM (1,22 ± 1,37 kg, P < 0,01; Figure 1A-C, rispettivamente; Morton et al., 2016). Non sono state riscontrate differenze tra i gruppi di ripetizioni (HR contro LR – vedi Morton et al., 2016) per nessuno dei risultati.
Regressioni a scalare:
Per ciascun risultato (variazione di CSA di tipo 1, CSA di tipo 2 e LBM) nessuna delle AUC post-esercizio (Tabella 1) o delle concentrazioni a riposo (Tabella 2) di qualsiasi ormone misurato prima o dopo l’intervento è risultata costantemente significativa (cioè significativa con più risultati o in più momenti di misurazione) nei modelli finali. Inoltre, i valori dei coefficienti di determinazione (cioè R2) erano bassi (<0,25) per tutti gli esiti in ogni momento della misurazione, indicando che la variazione osservata nella risposta ipertrofica può essere spiegata solo in minima parte da qualsiasi modello adattato. Risultati simili sono stati riscontrati valutando il CSA di tipo 1, il CSA di tipo 2 e la LBM prima e dopo l’intervento rispetto alle concentrazioni ormonali a riposo (Tabella supplementare 1).
Regressione ad eliminazione all’indietro finale tra l’AUC dell’ormone sistemico post-esercizio e la variazione di CSA di tipo 1, CSA di tipo 2 e LBM.Risultati finali della regressione ad eliminazione all’indietro tra gli ormoni a riposo e la variazione di CSA di tipo 1, CSA di tipo 2 e LBM.
Analisi delle componenti principali:
L’analisi delle componenti principali è stata eseguita su predittori centrati e scalari ed è qui presentata (vedi figure) come scree plot per l’AUC post-esercizio pre-intervento, l’AUC post-esercizio post-intervento, le concentrazioni a riposo pre-intervento e le concentrazioni a riposo post-intervento. Come illustrato dagli scree plot a bassa pendenza, nessuna componente principale è risultata particolarmente efficace nello spiegare la varianza nel set di dati originale.
I diagrammi di scree illustrano la proporzione di varianza degli ormoni spiegata da ciascuna componente principale. I pannelli includono le componenti principali derivate dagli ormoni sistemici misurati: (A) pre-intervento post-esercizio, (B) post-intervento post-esercizio, (C) pre-intervento a riposo e (D) post-intervento a riposo. La linea orizzontale tratteggiata indica il punto di cut-off in cui le componenti principali di cui sopra hanno spiegato ≥80% di varianza tra l’insieme dei dati originali degli ormoni.
I ricercatori hanno scelto di mantenere il numero di componenti principali che spiegano ≥80% della varianza dei predittori originali, il che ha portato a sette componenti principali incluse in ciascuna delle regressioni a scalare a componenti principali. L’esecuzione della regressione a componenti principali (indipendentemente dal fatto che gli ormoni siano stati valutati a riposo, dopo l’esercizio, prima dell’intervento o dopo l’intervento) ha rivelato che nessuna componente principale è stata costantemente significativa o inclusa in nessuno dei modelli finali e che l’R2 finale non ha mai superato lo 0,25 ed è stato inferiore allo 0,05. Questi risultati indicano che la variazione osservata nella risposta ipertrofica alla RET può essere spiegata solo in minima parte da uno qualsiasi dei modelli adattati.
High- vs. Low-Responders:
C’è stata una differenza significativa tra HIR e LOR nella variazione della CSA di tipo 1 (HIR: 2106 ± 412, LOR: -520 ± 450 μm2), della CSA di tipo 2 (HIR: 2642 ± 756, LOR: -373 ± 593 μm2) e della LBM (HIR: 2,1 ± 0,8, LOR: 0,6 ± 0,8 kg, P ≤ 0,001; Figure 1D-F). Non c’erano differenze nel numero di partecipanti di ciascun gruppo di allenamento (HIR: quattro e sei e LOR: sei e quattro da HR e LR, rispettivamente).
Non vi è stata alcuna differenza nella concentrazione ormonale a riposo tra HIR e LOR, ad eccezione della concentrazione a riposo post-intervento di LH (HIR: 3,67 ± 0,63; LOR 4,59 ± 1,15 UI/L, P < 0,01) e lattato (HIR: 0,52 ± 0,05; LOR: 0,55 ± 0,07 mM, P = 0,02), che erano maggiori nel LOR. Non c’è stata differenza nell’AUC post-esercizio per nessun ormone tra HIR e LOR, ad eccezione del cortisolo pre-intervento post-esercizio, che era più alto nell’HIR (HIR: 576 ± 100; LOR: 508 ± 199 nM; P < 0,001).
Ormoni intramuscolari:
Non sono state riscontrate differenze nei valori pre-intervento, post-intervento o nella variazione di T o DHT intramuscolare tra HIR e LOR (Figure 3A,B, rispettivamente). Il cambiamento nell’espressione della 5α-reduttasi è stato significativo nell’HIR (pre: 1457 ± 450, post: 1957 ± 543 AU, P < 0,01) ma non nel LOR (pre: 1748 ± 559, post: 1994 ± 840 AU, P = 0,32; Figura 3C). Il contenuto di recettori per gli androgeni intramuscolari prima dell’intervento (HIR: 10827 ± 2789, LOR: 7759 ± 1323 AU, P < 0,01) e dopo l’intervento (HIR: 11406 ± 2789, LOR: 7801 ± 1189 AU, P = 0,01; Figura 3D) era significativamente maggiore in HIR rispetto a LOR. Non c’è stato alcun cambiamento nel contenuto dei recettori degli androgeni intramuscolari prima e dopo l’intervento (Δ319 ± 1314 AU, P = 0,75) e c’è stata una relazione lineare tra il contenuto dei recettori degli androgeni dei partecipanti prima e dopo l’intervento (r = 0,92). Non sono state riscontrate correlazioni significative tra il T, il DHT o la 5α-reduttasi intramuscolare prima dell’intervento, dopo l’intervento e la variazione della CSA di tipo 1, della CSA di tipo 2 o della LBM (P > 0,05; Tabella supplementare 5). Al contrario, il contenuto di recettori per gli androgeni prima dell’intervento, dopo l’intervento e la media tra il contenuto di recettori per gli androgeni prima e dopo l’intervento sono stati significativamente correlati con la variazione della LBM (pre: r = 0,76, P < 0,01; post: r = 0,75, P < 0,01; media: r = 0. 77, P < 0,01), CSA di tipo 1 (pre: r = 0,51, P = 0,03; post: r = 0,49, P = 0,04; media: r = 0,51, P = 0,03) e CSA di tipo 2 (pre: r = 0,61, P < 0,01; post: r = 0,65, P < 0,01; media: r = 0,64, P < 0,01; Tabella supplementare 5 e Figura 4). I dati di un partecipante sono stati rimossi dalle analisi di regressione che includevano la variazione della LBM perché identificati come outlier statistici attraverso il metodo di regressione robusta e rimozione degli outlier con un coefficiente dell’1% (Motulsky e Brown, 2006). La posizione di questo partecipante è stata indicata nella Figura seguente a scopo illustrativo.
Correlazioni tra il contenuto di recettori androgeni intramuscolari prima dell’intervento e le variazioni della massa muscolare. Le correlazioni sono presentate nei pannelli per: (A) CSA di tipo 1 (r = 0,51, P = 0,03), (B) CSA di tipo 2 (r = 0,61, P < 0,01) e (C) LBM (r = 0,76, P < 0,01). In (C), l’outlier che è stato rimosso dall’analisi correlazionale tra il contenuto di recettori per gli androgeni prima dell’intervento e la LBM è incluso nella figura come una “×”.
Punto della situazione:
Il risultato principale del presente studio, coerente con il lavoro precedentemente svolto dai ricercatori, è che nessun ormone sistemico condivide una varianza significativa con i cambiamenti indotti da RET nella CSA delle fibre muscolari scheletriche o nella massa muscolare scheletrica negli uomini allenati contro-resistenza. Sono stati estesi questi risultati alle concentrazioni ormonali locali misurate nel muscolo, che non hanno mostrato un’associazione significativa con alcun indice di ipertrofia. E’ stato riscontrato che gli HIR presentavano un aumento del contenuto di 5α-reduttasi dopo 12 settimane di RET e un contenuto di recettori degli androgeni significativamente più alto, che non cambiava con la RET, rispetto ai LOR sia prima che dopo la RET. La conclusione di ciò è che né la disponibilità sistemica né quella locale muscolare di ormoni influenzano l’ipertrofia muscolo-scheletrica indotta dalla RET in giovani uomini sani. Coerentemente con i lavori precedenti, i ricercatori propongono invece che l’entità dell’ipertrofia muscolare scheletrica indotta da RET sia modulata in parte dal contenuto intramuscolare di recettori per gli androgeni e probabilmente da altre variabili intramuscolari.
Ormoni circolanti e allenamento contro-resistenza:
Recenti pubblicazioni (Kraemer et al., 2017; Mangine et al., 2017) e linee guida (Ratamess et al., 2009) sostengono che gli ormoni circolanti sono meccanicamente e direttamente correlati e predittivi dei cambiamenti della massa muscolare scheletrica indotti dal RET, nonostante l’esistenza di prove che dimostrano il contrario (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016; Mobley et al., 2018). In uno studio precedente a quello attualmente trattato, i ricercatori hanno eseguito 120 correlazioni, ciascuna su 49 partecipanti, tra 10 diversi ormoni e varie misure di variazione della massa e della forza muscolare. Hanno riscontrato che solo l’aumento del Cortisolo dopo l’esercizio era correlato con le variazioni del CSA di tipo 2 (pre-intervento: r = -0,34, P = 0,02; post-intervento: r = -0,31, P = 0,04) (Morton et al., 2016). Altri hanno trovato correlazioni significative tra l’aumento post-esercizio del GH circolante (McCall et al., 1999) e del T (Ahtiainen et al., 2003; Brook et al., 2016) con le variazioni della massa muscolare, ma queste correlazioni sono state eseguite su campioni composti da meno di 11 partecipanti, che potrebbero dare origine a correlazioni spurie. Qui sono state eseguite altre 48 regressioni graduali su 49 partecipanti, 10 ormoni e tre risultati distinti legati all’ipertrofia, tra cui la dimensione delle fibre muscolari. E’ stato riscontrato che nessun ormone era costantemente significativo, né alcun modello finale aveva un elevato coefficiente di determinazione, cioè tutti i valori di R2 erano inferiori a 0,25. Inoltre, la PCA non era efficace nel determinare le correlazioni con l’ipertrofia. Inoltre, la PCA non è stata efficace nel ridurre la varianza totale dei dati ormonali originali e non c’è stato alcun modello di regressione con le componenti principali utilizzate come covariate che spiegasse una proporzione significativa della variabilità in qualsiasi risultato. Esistono oggi prove sostanziali che suggeriscono che gli ormoni sistemici circolanti misurati a riposo (McCall et al., 1999; Morton et al., 2016; Mobley et al., 2018) e/o dopo l’esercizio (Ahtiainen et al., 2003; West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016) non condividono alcuna varianza comune e non sono quindi né correlati né predittivi dei cambiamenti della massa muscolare indotti dal RET in giovani partecipanti sani.
Uno studio (Mangine et al., 2017) ha utilizzato un modello di equazione strutturale ai minimi quadrati parziali (PLS-SEM) e ha riportato che un modello con punteggi ormonali compositi (T, GH, IGF-1, insulina e cortisolo) e una misura composita di ipertrofia (CSA e spessore muscolare del vasto laterale e del retto femorale) ha prodotto un coefficiente di determinazione significativo (R2= 0,73). L’interpretazione di questo risultato è che il punteggio ormonale composito era correlato a un punteggio composito di ipertrofia. L’aspetto preoccupante di questa interpretazione è che il modello senza T (il miglior predittore ormonale del modello) aveva ancora un coefficiente di determinazione sostanziale (R2 = 0,43) con il punteggio composito di ipertrofia ed era statisticamente significativo. In effetti, la rimozione individuale degli altri ormoni (GH, IGF-1, insulina e cortisolo) ha mostrato un effetto trascurabile sulla varianza condivisa del modello, eppure il modello senza il suo “migliore” ormone predittivo, il T, ha rappresentato quasi il 60% della varianza osservata con tale ormone presente nel modello. Mentre gli autori sostengono che le interazioni inspiegabili tra gli ormoni siano la ragione della varianza del modello senza T, è stato suggerito che è più probabile che i pesi PLS capitalizzino il caso per esagerare le correlazioni (Goodhue et al., 2012). Sebbene riteniamo che il PLS-SEM sia utile per l’esame di grandi insiemi di dati, vi sono limitazioni sostanziali all’interpretazione quando si utilizzano campioni di piccole dimensioni (n = 26) (Goodhue et al., 2012). La definizione di PLS come metodo SEM appropriato è stata messa in discussione anche per quanto riguarda la stima e l’inferenza (Rönkkö e Evermann, 2013) e il coefficiente di determinazione (ad esempio, R2) è un parametro inadeguato per valutare l’adattamento del modello PLS-SEM, poiché stimatori incoerenti possono produrre modelli con R2 elevato. Di conseguenza, non tutti i modelli ben adattati sono predittivi (Henseler et al., 2014) e non tutti i modelli predittivi sono ben adattati (McIntosh et al., 2014).
High- vs. Low-Responders e allenamento contro-resistenza:
Per indagare sui potenziali determinanti dell’eterogeneità dell’ipertrofia muscolare scheletrica indotta da RET (Hubal et al., 2005; Davidsen et al., 2011; Morton et al., 2016), i ricercatori hanno suddiviso 49 partecipanti in HIR (n = 10) e LOR (n = 10) in base alla variazione di tre indicatori della massa muscolare scheletrica (CSA di tipo 1, CSA di tipo 2 e LBM). Nonostante le grandi differenze tra i gruppi in ogni risultato, non ci sono state differenze significative in nessun ormone circolante prima o dopo l’esercizio fisico, misurato sia prima che dopo l’intervento. Considerando che gli ormoni steroidei sono liposolubili (ad esempio, si diffondono attraverso il sarcolemma in base al loro gradiente di concentrazione), non sorprende che anche il T e il DHT intramuscolari misurati prima e dopo l’intervento non fossero diversi tra HIR e LOR. La mancanza di differenze negli ormoni circolanti e intramuscolari tra HIR e LOR dimostra che né l’apporto di ormoni al muscolo né il trasferimento di ormoni steroidei all’interno del muscolo sono fasi limitanti in individui giovani e sani.
Il contenuto di recettori per gli androgeni era significativamente più alto sia prima che dopo l’intervento nell’HIR rispetto al LOR ed era correlato alle variazioni della massa muscolare. Sebbene un altro gruppo non abbia riscontrato alcuna differenza nel contenuto di recettori degli androgeni tra HIR e LOR (Mobley et al., 2018), è importante riconoscere le differenze nel disegno dello studio (ad esempio, partecipanti non allenati rispetto a quelli allenati) e nelle misure di esito (ad esempio, analisi dei cluster basata sullo spessore muscolare rispetto a un punteggio aggregato di DXA e CSA delle fibre) tra loro e il lavoro dei ricercatori, rispettivamente. La funzione del recettore degli androgeni è quella di traslocare nel nucleo e modificare l’espressione dei geni bersaglio [rivisto altrove (Beato e Klug, 2000)], molti dei quali sono bersagli noti coinvolti nella crescita e nello sviluppo del muscolo scheletrico (Wyce et al., 2010). Infatti, quando i recettori degli androgeni vengono eliminati nei topi maschi, si verifica una significativa riduzione della massa e della forza muscolare (MacLean et al., 2008). È importante notare che la maggior parte degli ormoni steroidei ha un’elevata affinità con i propri recettori steroidei. Ad esempio, la costante di dissociazione del recettore degli androgeni nei confronti del T e del DHT è di soli ∼0,2-0,5 nM (Wilson e French, 1976). Nel presente studio, a riposo, la molarità del T sierico (HIR: 28 ± 7; LOR: 31 ± 7 nM), della fT sierica (HIR: 0,5 ± 0,01; LOR: 0,5 ± 0,01 nM) e del DHT sierico (HIR e LOR: 0,7 ± 0,2 nM) superavano tutti 0,2-0,5 nM. Dato che non c’era alcuna differenza negli ormoni circolanti o intramuscolari tra HIR e LOR, insieme all’elevata affinità di legame tra androgeno e recettore degli androgeni, sembra probabile che sia a riposo che dopo l’esercizio i recettori androgeni esistenti siano stati saturati nel muscolo scheletrico. Si ipotizza che, sebbene l’apporto di androgeni possa essere un passo limitante per l’ipertrofia muscolare indotta da RET negli uomini ipogonadici (Bhasin et al., 1997; Kvorning et al., 2013), il contenuto di recettori per gli androgeni sia la variabile più importante nell’accrescimento di proteine del muscolo scheletrico mediato dagli androgeni indotti da RET negli uomini sani (Diver et al., 2003).
Limitazioni:
I ricercatori hanno eseguito 120 correlazioni in uno studio precedente (Morton et al., 2016) e 48 regressioni graduali in questo caso (24 sui dati originali e 24 sulle componenti principali). L’applicazione di analisi multiple sugli stessi dati è stata un’operazione di data mining intenzionale per dimostrare la mancanza di capacità degli ormoni circolanti e intramuscolari a riposo o dopo l’esercizio fisico di prevedere le variazioni della massa muscolare scheletrica al basale o indotte dalla RET. Avrebbero potuto eseguire ulteriori statistiche per tenere conto dei test multipli, ma questo non sarebbe stato informativo perché nessuno dei loro modelli spiegava molta varianza (come valutato dai valori di R2, che non superavano lo 0,25). Riconoscono inoltre che, pur avendo incluso un campione di grandi dimensioni (n = 49) per l’analisi degli ormoni sistemici, essi si sono limitati a un campione relativamente più piccolo (n = 20) per il confronto tra HIR e LOR. Ammettono pienamente che, nel caso della correlazione con il recettore degli androgeni, quella che presentano è una stima gonfiata a causa della scelta di misurare solo i soggetti con risposta più alta e più bassa al loro protocollo di allenamento. Hanno condotto la loro analisi in questo modo per illustrare la differenza nell’ipertrofia muscolare indotta da RET e per indagare l’influenza delle variabili ormonali circolanti e intramuscolari su due gruppi distinti. Sebbene fossero limitati dalla quantità di tessuto raccolto, è giusto criticare il fatto che la loro analisi correlazionale sarebbe stata più eloquente se avessero incluso tutti i partecipanti e se avessero eseguito analisi aggiuntive [ad esempio, frazioni nucleari e citoplasmatiche del contenuto di recettori degli androgeni e espressioni geniche multiple (Cheung et al., 2017)]. Per questo motivo, il lavoro futuro potrà concentrarsi sulla biologia specifica che regola la regolazione e la funzione del recettore degli androgeni. Altri hanno ipotizzato che l’analisi con spettrometria di massa (rispetto ai test immunologici) sia necessaria per rilevare piccole concentrazioni intramuscolari di ormoni steroidei (Handelsman e Wartofsky, 2013); tuttavia, l’intento dei ricercatori era quello di analizzare i loro campioni utilizzando metodi simili a quelli che altri hanno utilizzato nella scienza dell’esercizio fisico, che possono essere diversi da quelli dell’endocrinologia clinica. Riconoscono che l’uso della DXA per misurare i cambiamenti nella LBM non è il gold standard, motivo per cui hanno scelto di includere anche i cambiamenti nella CSA delle fibre di tipo 1 e 2 per determinare i loro HIR e LOR (Buckinx et al., 2018). Per quanto riguarda la loro interpretazione, è ingenuo suggerire che la segnalazione degli androgeni sia esclusivamente operativa attraverso la loro tendenza a legarsi a un recettore androgenico [rivisto altrove (Herbst e Bhasin, 2004; Dubois et al., 2012)]. Sebbene la regolazione trascrizionale (ad esempio, la segnalazione dei recettori degli androgeni) sia qui evidenziata come un potente modulatore dei cambiamenti nella massa muscolare indotti da RET, è anche chiaro che la regolazione post-trascrizionale è almeno altrettanto importante per la sintesi proteica (Schwanhausser et al, 2011), come è stato evidenziato da recenti risultati (Figueiredo et al., 2015; Robinson et al., 2017; Mobley et al., 2018) e review (Chaillou et al., 2014; McGlory et al., 2017). Infine, sebbene vi sia un’influenza genetica alla base dell’ipertrofia muscolare scheletrica indotta da RET, vi sono ancora molte considerazioni ambientali, ad esempio il consumo di proteine alimentari adeguate (Morton et al., 2017), un apporto calorico e stimolo allenante adeguato che modulano l’ipertrofia muscolare indotta da RET.
Riflessioni conclusive sul presente studio:
Ricapitolando, i ricercatori hanno eseguito l’eliminazione a ritroso e la regressione delle componenti principali su una coorte relativamente ampia (n = 49) di uomini allenati contro-resistenza, concludendo che l’AUC post-esercizio (cioè l’esposizione ormonale netta transitoria acuta) e le concentrazioni ormonali a riposo misurate nel sangue non condividono una varianza comune con le variazioni della massa muscolare indotte dalla RET. In altre parole, le concentrazioni ormonali sistemiche non sono correlate o in qualche modo predittive delle variazioni della massa muscolare indotte da RET. L’analisi dei sottoinsiemi dei soggetti con risposta più alta e più bassa ha rivelato che il contenuto di recettori per gli androgeni, e non i livelli di androgeni intramuscolari, non cambia con il RET nei partecipanti allenati, ma è significativamente più alto negli HIR rispetto ai LOR. Questo studio, insieme ad altri (Bamman et al., 2007; Petrella et al., 2008; Davidsen et al., 2011; Eynon et al., 2013), fornisce la prova che l’aumento relativo della massa muscolare scheletrica in seguito alla RET è sostenuto da fattori locali intramuscolari e non da concentrazioni ormonali sistemiche.
Questo è quanto suggerito dall’osservazione di soggetti in stato fisiologico. Individui trattati con dosi esogene sovrafisiologiche di AAS sarebbero teoricamente soggetti alle medesime limitazioni presenti nel confronto tra HIR e LOR dello studio. Questa limitazione sembra essere data dall’espressione dei AR (Recettori degli Androgeni) nel muscolo scheletrico. Sebbene dosi sovrafisiologiche di AAS causino un aumento del numero dei AR presenti nel muscolo scheletrico, tale espressione è comunque soggetta ad una regolazione genica con variabili soggettive di potenziale. Tali variabili sono teoricamente evincibili dall’osservazione degli atleti allenati contro-resistenza, specie Bodybuilder, e della loro differenza di potenziale indipendente nella sua massima espressione. Tale potenziale è diverso tra HIR e LOR sia in fisiologia che in condizione di trattamento farmacologico, indipendentemente dalla dose di AAS utilizzata.
La capacità di riacquisire la condizione della massa muscolare precedente a un periodo di deallenamento o inattività fisica è noto come “memoria muscolare”. Quindi, se un soggetto ha avuto una condizione muscolare ottimale (vedi muscoli più ipertrofici) in passato, ciò lo aiuterà a riportarli nuovamente nelle precedenti condizioni una volta ripreso un regolare stimolo allenante. Il concetto di memoria muscolare si basa in buona parte su qualcosa chiamato permanenza mio-nucleare. Il ‘mio’ in ‘mionucleare’ si riferisce al ‘muscolo’ e il ‘nucleare’ si riferisce alla parola ‘nucleo’: un organello della cellula. Prima di esplorare ulteriormente il concetto di memoria muscolare, e come gli AAS si leghino a questo, cerchiamo prima di rispolverare un po’ di concetti utili sui nuclei muscolari o mionuclei.
Informazioni di base sui nuclei muscolari/mionuclei:
Le cellule muscolo-scheletriche sono le singole cellule contrattili all’interno di un muscolo e sono spesso definite fibre muscolari.[1] Un singolo muscolo come il bicipite in un giovane individuo di sesso maschile adulto contiene circa 253.000 fibre muscolari.[2]
Sezione 3D di una fibra del muscolo-scheletrico
Le fibre muscolo-scheletriche sono le uniche cellule muscolari multinucleate con i nuclei spesso indicati come mionuclei . Ciò si verifica durante la miogenesi con la fusione di mioblasti, ciascuno dei quali contribuisce a un nucleo.[3] La fusione dipende da proteine muscolo-specifiche note come fusogeni chiamate myomaker e myomerger .[4]
Molti nuclei sono necessari alla cellula muscolo-scheletrica per le grandi quantità di proteine ed enzimi necessari per essere prodotti per il normale funzionamento della cellula. Una singola fibra muscolare può contenere da centinaia a migliaia di nuclei.[5] Una fibra muscolare ad esempio nel bicipite umano con una lunghezza di 10cm può avere fino a 3000 nuclei.[5] A differenza di una cellula non muscolare in cui il nucleo è posizionato centralmente, il mionucleo è allungato e si trova vicino al sarcolemma . I mionuclei sono disposti in modo abbastanza uniforme lungo la fibra con ciascun nucleo che ha il proprio dominio mionucleare dove è responsabile del supporto del volume del citoplasma in quella particolare sezione della miofibra.[4,5]
Un gruppo di cellule staminali muscolari conosciute come cellule miosatelliti, anche cellule satelliti che si trovano tra la membrana basale e il sarcolemma delle fibre muscolari, sono normalmente quiescenti ma possono essere attivate dall’esercizio o anche condizioni patologiche per fornire mionuclei aggiuntivi per la crescita o la riparazione muscolare.[6]
Detto più semplicemente, i muscoli sono costituiti da un insieme di fibre muscolari. Ogni fibra muscolare, o cellula muscolare, contiene più nuclei, l’organello di una cellula che contiene il DNA ed è il luogo dove avviene il processo di trascrizione dei geni. La maggior parte degli altri tipi di cellule umane contiene solo un nucleo, o in alcuni casi addirittura nessun nucleo (globuli rossi/Eritrociti). Per dare un’idea di quanti nuclei si stia parlando: le fibre muscolari di ratto contengono da 44 a 116 nuclei per millimetro di lunghezza della fibra, con le fibre muscolari di tipo 1 che contengono più nuclei per millimetro delle fibre muscolari di tipo 2.[7] Il numero sembra più basso negli esseri umani, come riportato da un ricercatore il quale segnala la presenza di circa 30 nuclei per millimetro di lunghezza della fibra nel muscolo del bicipite brachiale.[8] Come tali, le fibre muscolari possono contenere migliaia di mionuclei, dato che possono estendersi per diversi centimetri di lunghezza.
Poiché i nuclei cellulari delle fibre muscolari non sono in grado di dividersi (cioè sono differenziati terminalmente), le fibre muscolari dipendono dalle cellule satelliti circostanti per l’aggiunta di nuovi nuclei. Essenzialmente, le cellule satelliti sono cellule staminali delle fibre muscolari che si trovano schiacciate tra il sarcolemma (la membrana cellulare di una fibra muscolare) e la lamina basale (uno strato di matrice extracellulare che è avvolto intorno al sarcolemma). Sono stati scoperti e descritti per la prima volta da Alexander Mauro nella letteratura scientifica nel 1961.[9] Usando un microscopio elettronico, egli vide delle cellule “incastrate” tra il sarcolemma delle fibre muscolari di rana e la lamina basale. Le descrisse aventi una scarsità di citoplasma, con il nucleo che costituisce quasi l’intero volume della cellula satellite. Ha continuato a speculare sull’origine e sul ruolo delle cellule satelliti, toccando brevemente l’idea che potrebbero essere coinvolte nella risposta al trauma inflitto a una fibra muscolare. Cosa che, in effetti, sono.[10]
L’ipotesi del dominio mionucleare e la permanenza mionucleare
La scoperta delle cellule satelliti e il loro ruolo nella rigenerazione muscolare fanno sorgere la domanda sulla misura in cui le cellule satelliti sono coinvolte nell’ipertrofia. Un’ipotesi chiamata “ipotesi del dominio mionucleare” si è agganciata a questo quesito. Essa postula che un mionucleo controlla una quantità limitata di citoplasma, e quindi, affinché la crescita muscolare abbia luogo, i mionuclei devono essere aggiunti alla fibra muscolare per sostenerla. Tre osservazioni chiave hanno sostenuto questa ipotesi, vale a dire:
L’esposizione alle radiazioni γ rende le cellule satellite incapaci di dividersi e inibisce fortemente l’ipertrofia da sovraccarico nei modelli animali, mantenendo intatto il metabolismo cellulare o la sintesi proteica [11].
I prodotti (organelli, membrane e proteine strutturali) derivati da un nucleo rimangono localizzati nelle sue vicinanze [12].
Il rapporto citoplasma/mionucleo rimane abbastanza costante [13].
Questo implicherebbe un aumento del numero di mionuclei con la crescita di una fibra muscolare (ipertrofia), mentre diminuirebbe con una perdita di dimensioni della stessa (atrofia). Tuttavia, vari studi su animali suggeriscono che i mionuclei non si perdono durante l’atrofia.[14] Così è nato il paradigma della permanenza mionucleare: una volta che i mionuclei sono guadagnati con l’ipertrofia, non vengono persi di nuovo con il deallenamento. Questo potrebbe potenzialmente permettere alle fibre muscolari di ricrescere in modo più efficiente durante il successivo riallenamento e quindi servire come un meccanismo di “memoria muscolare”.
Il concetto di memoria muscolare basato sulla permanenza mionucleare illustrato da Bruusgaard et al.
AAS e permanenza mionucleare:
E gli AAS? Ciò che è chiaro è che l’uso di AAS aumenta il numero di mionuclei. Dosaggi crescenti di Testosterone Enantato portano ad un aumento del numero di mionuclei per mm di fibra muscolare.[15] Questo effetto non è poi così sorprendente: si osserva semplicemente questo effetto con praticamente tutte le modalità di induzione ipertrofica.
Ma che dire della loro permanenza? Questi mionuclei permangono una volta che la massa muscolare diminuisce di nuovo? In un esperimento su animali, da me già riportato anni fa, topi femmina sono stati trattati con Testosterone Propionato per 2 settimane, che ha portato a un aumento del 66% del numero di mionuclei e un aumento del 77% della fibra muscolare CSA [16]. La massa muscolare è tornata alla normalità dopo la successiva interruzione della somministrazione di Testosterone, ma il numero di mionuclei è rimasto elevato per almeno 3 mesi. 3 mesi potrebbe non sembrare molto, ma sulla scala temporale di un topo lo sono: i topi che hanno usato per lo studio vivono per circa 2 anni. Comunque, dopo questi 3 mesi, quando i topi sono stati sottoposti a sovraccarico per induzione ipertrofica, la CSA delle fibre muscolari è aumentata del 30% dopo 6 giorni, mentre quella dei topi di controllo non è aumentata significativamente. Dopo questo, la massa muscolare è aumentata in parallelo tra entrambi i gruppi, ma la CSA era ancora più alta del 20% nel gruppo che era stato precedentemente trattato con Testosterone dopo 14 giorni. Anche se questo non prova un nesso causale tra il numero più alto di mionuclei e l’ipertrofia, è comunque un’osservazione interessante.
Si noti come il gruppo che è stato trattato con Testosterone per 2 settimane, circa 3 mesi prima ha mostrato un forte aumento della massa muscolare rapidamente ottenuto in risposta al sovraccarico.
E negli esseri umani? Due studi hanno valutato questo e sono stati portati all’attenzione da Alexander Kolliari-Turner, uno studente con dottorato di ricerca presso la School of Sport and Health Sciences of the University of Brighton nel Regno Unito. Una è una tesi di master e l’altra è una tesi di dottorato.
Nella tesi di dottorato di Anders Eriksson [17], sono stati reclutati quattro gruppi di soggetti. Un gruppo di soggetti sedentari che fungeva da controllo (gruppo C), un gruppo di PowerLifter natural (gruppo P), un gruppo di powerlifter che usano AAS (gruppo PAS), e un gruppo di PowerLifter che hanno precedentemente usato AAS (gruppo PREV). I mionuclei per fibra muscolare sono stati determinati nei muscoli vasto laterale e trapezio. Il gruppo PREV aveva interrotto l’uso di AAS da almeno un anno (con una media di 8 anni). Infatti, l’area delle fibre muscolari misurata nel gruppo PREV era paragonabile a quella del gruppo P, e notevolmente più piccola di quella del gruppo PAS.
La distribuzione del dominio nucleare (nr. di nuclei per fibra diviso per l’area della fibra) per gruppo si trova nell’immagine qui sotto. Se ci fosse una permanenza dei mioonuclei, ci si aspetterebbe un dominio nucleare più piccolo, cioè più nuclei rispetto all’area delle fibre, nel gruppo PREV rispetto agli altri gruppi.
Chiaramente questo non è il caso del vasto laterale, ma è il caso del trapezio. È difficile dire cosa causa questa apparente discrepanza tra i due muscoli. O qualche proprietà che differisce tra i due muscoli, o il suo modo di utilizzo dopo la cessazione dell’uso di AAS, forse ha portato a apparente permanenza mionucleare nel muscolo trapezio.
Va notato, tuttavia, che questo era uno studio trasversale con un piccolo numero di soggetti (32 in totale). L’ideale sarebbe avere uno studio prospettico che valuti questo, anche se ciò è estremamente difficile su lunghi periodi di tempo, in quanto potrebbe richiedere almeno un anno o più prima che i cambiamenti diventino evidenti. In alternativa, anche uno studio trasversale con un gruppo di soggetti più grande sarebbe piuttosto interessante. Indipendentemente da ciò, questo presta una certa credibilità alla permanenza dei mionuclei negli esseri umani come risultato dell’uso di steroidi anabolizzanti in muscoli selezionati.
In una tesi di laurea di Lindholm et al. sono stati reclutati tre gruppi di soggetti: attuali consumatori di AAS (gruppo CAS), ex consumatori di AAS (gruppo FAS) e controllo allenati alla resistenza (gruppo CON) [18]. Gli ex consumatori di AAS avevano smesso di usarli per una media di 6,5 anni. In questo studio, sono state prese solo biopsie del muscolo vasto laterale. In particolare, non c’erano differenze significative nella CSA delle fibre muscolari tra i tre gruppi. Questo è senza dubbio il risultato delle dimensioni relativamente piccole del gruppo (34 soggetti in totale; un errore di tipo 2).
Una piccola, ma significativa, differenza nel dominio mio-nucleare è stata trovata tra le fibre muscolari di tipo 2 del gruppo FAS rispetto al gruppo CON, come si può vedere nella figura sottostante:
Questo suggerisce una permanenza mionucleare? Forse. La differenza era piccola e può essere facilmente spiegata anche dalla natura trasversale dello studio (e non c’era alcuna differenza rispetto agli attuali utilizzatori di AAS).
Le prove finora sono scarse. In ogni caso, quando si guarda alla permanenza mionucleare in generale, l’evidenza generale indica che questa regge a breve termine, ma mancano prove per il lungo termine [19]. Inoltre, non è chiaro se la permanenza mionucleare possa aiutare o meno il ritorno alla condizione muscolo-scheletrica precedente. E visti i dati di cui sopra, il dibattito sul fatto che l’uso di AAS porti o meno alla manifestazione della memoria muscolare come risultato della permanenza mionucleare, è tutt’altro che risolto.
Conclusione:
Come osservazione conclusiva: c’è anche un concetto di memoria muscolare basato su qualcosa di diverso dalla permanenza mionucleare, vale a dire, la memoria epigenetica.[20] In breve, questa si riferisce a modifiche apportate al DNA senza influenzare la sua sequenza nucleotidica, quindi senza cambiare il codice genetico. Ciò comporta l’aggiunta (o la rimozione) di gruppi metilici ai nucleotidi di Citosina e Adenina o modifiche degli istoni (ad esempio, metilazione o acetilazione di residui di aminoacidi delle proteine istoniche). Il risultato di ciò è che influisce sull’espressione genica. Questo potrebbe forse essere trattato in un futuro articolo, dato che più ricerche vengono gradualmente pubblicate su questa nuova ed interessante strada ipotetica.
A proposito di “memoria epigenetica”: questa figura illustra un modello di sviluppo della persistenza batterica basato sulla presenza di un potenziale effetto di “memoria” epigenetica che include l’eredità stabile di certi modelli di metilazione del DNA. Lo stato di metilazione del DNA cellulare potrebbe portare alla conservazione di alcuni profili di espressione genica che favoriscono la dormienza, conservati in alcune cellule dopo il risveglio dalla dormienza. Cinetica di uccisione bifasica adattata da. (A) Popolazione originale di cellule metabolicamente attive che potrebbero contenere un’intrinseca eterogeneità fenotipica. (B) Quando incontra lo stress, la maggior parte delle cellule metabolicamente attive muore, mentre una piccola frazione di cellule entra nello stato di persistenza. La popolazione di persister può essere in qualche modo eterogenea, cioè formata da diversi percorsi (stocastico contro specifico). (C) Dopo gli stimoli nutrizionali/la rimozione dello stress, alcuni persister si risvegliano. Qui, la maggior parte dei persister inizia rapidamente la crescita e si divide in cellule regolari e metabolicamente attive. Tuttavia, alcune cellule potrebbero sperimentare un effetto di “memoria” epigenetica. Qui, lo stato di metilazione del DNA di alcuni siti che si trovano a monte di regioni codificanti regolate per esprimere tratti che favoriscono la dormienza potrebbe essere mantenuto dopo la replicazione del DNA. (D) A livello di popolazione totale, la popolazione finale dopo il risveglio potrebbe essere ugualmente suscettibile allo stress come la popolazione originale in (A). Tuttavia, a livello di singola cellula, alcune cellule potrebbero contenere un effetto di “memoria” legato alla dormienza, basato sull’eredità di alcuni tratti epigenetici dipendenti dalla metilazione del DNA. (E) L’esistenza di un effetto di “memoria” epigenetica potrebbe potenzialmente aumentare la frequenza dei persister nel tempo durante ripetuti cicli di stress.
Klein, CS; Marsh, GD; Petrella, RJ; Rice, CL (July 2003). “Muscle fiber number in the biceps brachii muscle of young and old men”. Muscle & Nerve. 28 (1): 62–8.
Tseng, Brian S., Christine E. Kasper, and V. Reggie Edgerton. “Cytoplasm-to-myonucleus ratios and succinate dehydrogenase activities in adult rat slow and fast muscle fibers.” Cell and tissue research 275.1 (1994): 39-49.
Schmalbruch H. Skeletal Muscle. Berlin: Springer-Verlag; 1985.
Mauro, Alexander. “Satellite cell of skeletal muscle fibers.” The Journal of Cell Biology 9.2 (1961): 493-495.
Forcina, Laura, et al. “An overview about the biology of skeletal muscle satellite cells.” Current genomics 20.1 (2019): 24-37.
Rosenblatt, J. David, David Yong, and David J. Parry. “Satellite cell activity is required for hypertrophy of overloaded adult rat muscle.” Muscle & nerve 17.6 (1994): 608-613.
Pavlath, Grace K., et al. “Localization of muscle gene products in nuclear domains.” Nature 337.6207 (1989): 570-573.
Allen, David L., Roland R. Roy, and V. Reggie Edgerton. “Myonuclear domains in muscle adaptation and disease.” Muscle & nerve 22.10 (1999): 1350-1360.
Gundersen, Kristian, and Jo C. Bruusgaard. “Nuclear domains during muscle atrophy: nuclei lost or paradigm lost?.” The Journal of physiology 586.11 (2008): 2675-2681.
Sinha-Hikim, Indrani, et al. “Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men.” American Journal of Physiology-Endocrinology and Metabolism 285.1 (2003): E197-E205.
Egner, Ingrid M., et al. “A cellular memory mechanism aids overload hypertrophy in muscle long after an episodic exposure to anabolic steroids.” The Journal of physiology 591.24 (2013): 6221-6230.
Eriksson, Anders. Strength training and anabolic steroids: a comparative study of the trapezius, a shoulder muscle and the vastus lateralis, a thigh muscle, of strength trained athletes. PhD Diss. 2006.
Lindholm, Jesper Bøgh, et al. Effects of Long-Term Supplementation of Androgen Anabolic Steroids on Human Skeletal Muscle – Evidence for Muscle Memory? Master’s Thesis, 2019.
Snijders, Tim, et al. “The concept of skeletal muscle memory: Evidence from animal and human studies.” Acta Physiologica 229.3 (2020): e13465.
Seaborne, Robert A., et al. “Human skeletal muscle possesses an epigenetic memory of hypertrophy.” Scientific reports 8.1 (2018): 1-17.