
Introduzione:
Dell’Insulina ho già parlato abbondantemente non molto tempo fa attraverso la serie di quattro articoli dedicati a questo peptide nella ricorrenza dei cento anni dalla sua scoperta e prima applicazione in medicina (per approfondire clicca qui, qui, qui e qui). Nella quarta e ultima parte della prima citata serie di articoli, mi concentrai sulle pratiche protocollari dell’Insulina utilizzate in campo culturistico e, tra queste, vi era anche il protocollo di GH/Insulina nelle sue varianti applicative. Vi starete sicuramente chiedendo del perchè io scriva nuovamente di un argomento già trattato, oltretutto di recente. Beh, la risposta è abbastanza semplice: è un argomento che richiede per complessità degli approfondimenti aggiuntivi. Soprattutto alla luce delle “leggende da spogliatoio” e della limitatezza dei divulgatori “naturnazi”.
Come mi è già capitato di precisare, la visione dell’Insulina come ormone puramente anticatabolico è mentalmente riduttivo tanto quanto affermare che, di per se, l’Insulina sia “l’ormone anabolico per eccellenza”. Ed ecco perchè sopra ho specificato come sia necessario chiarire alcuni aspetti in contrapposizione al settarismo “magna e spigni” e “orcojo natty”. Sono volutamente ironico e provocatorio, ovviamente.
Giova anche ripetere che non sono l’unico a ritenere che la narrazione secondo la quale l’Insulina sia prettamente anti-catabolica sia simile alla visione precedente a quella di Bhasin (2001), secondo cui gli AAS non avrebbero effetti anabolici nel muscolo-scheletrico. Attualmente, la comunità (o, per meglio dire, “cupola”) scientifica vuole far credere che l’Insulina esogena non sia particolarmente anabolizzante nel muscolo-scheletrico umano.
Le nozioni e i dati provenienti dai miei appunti sull’Insulina, in parte già riportati negli articoli dedicati prima citati, sono una forte contestazione di questo pregiudizio istituzionale (analogo alla visione pre-Bhasin sugli AAS).
Il punto cieco istituzionale deriva dalla mancanza di studi sull’uso dell’Insulina esogena con:
- dosaggio sovrafisiologico;
- somministrazione sistemica;
- iperamminoacidemia;
- associazione a rhGH sovrafisiologico (e, più spesso, a AAS).
Ed è proprio su questi punti che si diramerà la mia disamina in questo articolo: valutazione teorico/pratica della co-somministrazione di GH e Insulina in condizioni sovra-fisiologiche.
Il motivo? Oltre le affermazioni arbitrarie e riduttive di alcuni divulgatori, vi è anche il risultato emerso dallo studio di Fryburg et al. i quali proposero che il rhGH e l’Insulina esogena lavorassero in modo incrociato, che la crescita fosse una conseguenza del turnover delle proteine muscolari piuttosto che dell’azione ormonale ipo/iperglicemica. La co-somministrazione di livelli endogeni (quindi fisiologici) di Insulina e rhGH sembrò dimostrare semplicemente una attenuazione degli effetti anabolici proteici del GH e che non fosse additiva. Ovviamente questa conclusione era limitata da: 1) il contesto di “picco ormonale” e della sua curva mantenuta in fisiologia e, cosa da non trascurare, 2) è stata trascurata l’importanza della disponibilità intracellulare di AA sulla sintesi proteica.
Ma andiamo per ordine…
Breve ripasso su azione dell’Insulina e meccanismi nell’ipertrofia muscolare:
L’effetto primario dell’Insulina nel muscolo scheletrico è sul gradiente elettrochimico di transmembrana, dove induce un’iperpolarizzazione nelle cellule muscolari attivando direttamente la pompa Na⁺-K⁺-ATPasi (46). [Biolo, G. et al. (1995)]. L’Insulina aumenta anche il numero di pompe Na+/K+-ATPasi nella membrana, determinando uno spostamento intracellulare del potassio e causando ipopotassiemia (basso livello di K/potassio) nello spazio extracellulare del sangue (siero).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
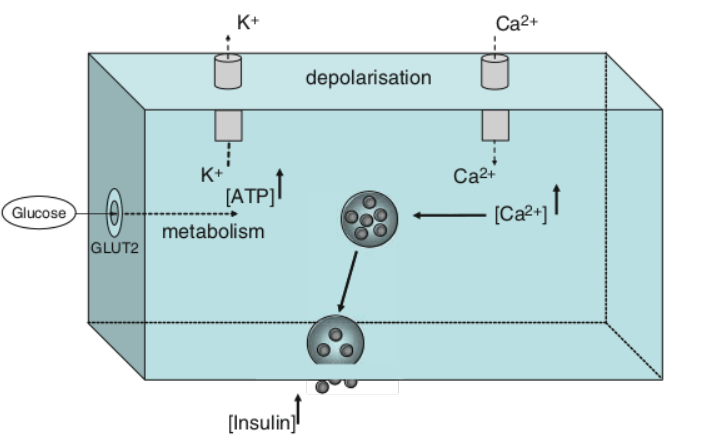
L’Insulina stimola l’espressione genica di MHCα (isoforma lenta) [Toniolo, L. (2005).] nel muscolo scheletrico e dell’albumina nel fegato (Dillman, 1988).
L’Insulina sembra stimolare in egual misura le fibre di tipo I e II [Albers et al. 2014]
Il muscolo scheletrico è un tessuto eterogeneo composto da diversi tipi di fibre. Gli studi suggeriscono che il metabolismo del glucosio mediato dall’insulina è diverso tra i vari tipi di fibre muscolari. Abbiamo ipotizzato che le differenze siano dovute all’espressione/regolazione specifica di elementi di segnalazione dell’insulina e/o di enzimi metabolici. Sono stati preparati pool di fibre di tipo I e II da biopsie dei muscoli del vasto laterale di soggetti magri, obesi e diabetici di tipo 2 prima e dopo un clamp iperinsulinemico-euglicemico. Le fibre di tipo I rispetto a quelle di tipo II presentano livelli proteici più elevati di recettore dell’insulina, GLUT4, esochinasi II, glicogeno sintasi (GS) e piruvato deidrogenasi-E1α (PDH-E1α) e un contenuto proteico inferiore di Akt2, TBC1 domain family member 4 (TBC1D4) e TBC1D1. Nelle fibre di tipo I rispetto a quelle di tipo II, la risposta di fosforilazione all’insulina era simile (TBC1D4, TBC1D1 e GS) o ridotta (Akt e PDH-E1α). Le risposte di fosforilazione all’Insulina aggiustate per il livello di proteine non erano diverse tra i tipi di fibre. Indipendentemente dal tipo di fibra, la segnalazione dell’insulina era simile (TBC1D1, GS e PDH-E1α) o ridotta (Akt e TBC1D4) nel muscolo di pazienti con diabete di tipo 2 rispetto a soggetti magri e obesi. Concludiamo che le fibre muscolari umane di tipo I rispetto a quelle di tipo II hanno una maggiore capacità di gestione del glucosio ma una sensibilità simile alla fosforegolazione da parte dell’Insulina.[Albers et al. 2014]
Livelli proteici più elevati di IRβ (+16%), esochinasi II impegna il glicogeno nella cellula, GLUT4 trasporta il glucosio nella cellula e complesso II della catena di trasporto degli elettroni ciclo mitocondriale/Kreb sono stati riscontrati nelle fibre di tipo I rispetto a quelle di tipo II.
Akt, mTOR: il contenuto proteico di Akt2 era più basso (-27%) nelle fibre di tipo I rispetto a quelle di tipo II. Gli aumenti medi della fosforilazione di [Akt] sotto stimolazione insulinica (ipertrofia) sono stati rispettivamente di 5,8 e 3,5 volte nelle fibre di tipo I e di 6,1 e 3,7 volte nelle fibre di tipo II. La risposta relativa all’insulina è stata simile tra i tipi di fibre.
Le fibre umane di tipo I hanno una maggiore abbondanza di trasporto (+29% GLUT4), fosforilazione (+470% HKII) e ossidazione (+35% complesso ETC II e +34% complesso piruvato deidrogenasi) del glucosio e di sintesi del glicogeno (+35%) rispetto alle fibre di tipo II.
Le fibre di tipo I possiedono una maggiore capacità di immagazzinamento del glicogeno.
Le differenze apparenti tra i tipi di fibre nella fosforilazione stimolata dall’insulina di Akt, NDRG1 (a valle di mTOR [queste sono le nostre vie dell’ipertrofia])… sono state eliminate quando sono state aggiustate per… l’abbondanza proteica. Questi risultati suggeriscono una sensibilità simile delle fibre muscolari di tipo I e di tipo II alla regolazione da parte dell’insulina delle proteine analizzate. [Albers et al. 2014]
Questi dati [Albers et al. 2014] supportano l’idea che, piuttosto che promuovere in modo preferenziale l’anabolismo proteico muscolare nelle fibre a contrazione lenta, entrambe le fibre muscolari di tipo I e II siano ugualmente sensibili agli effetti anabolici proteici dell’insulina esogena.
L’iperinsulinemia nei soggetti normali diminuisce acutamente le concentrazioni plasmatiche di aminoacidi come conseguenza del deposito netto di proteine. [Biolo, G., & Wolfe, R. R. (1993)].
Una volta secreta dalle beta-cellule pancreatiche, l’insulina circola nel flusso sanguigno con un’emivita di circa 12 minuti. Numerosi tessuti e organi esprimono il recettore dell’insulina e si attivano diverse azioni (Sonksen 2001), alcune delle quali sono di importanza generale e di particolare interesse per gli sport d’élite.
L’effetto centrale dell’insulina, in concerto con altri ormoni come il glucagone o la somatostatina, è il controllo dei livelli di glucosio nel sangue. La secrezione di insulina in risposta a concentrazioni elevate di glucosio (ad esempio, postprandiale) inibisce la produzione epatica di glucosio, abbassando così i livelli di glucosio nel sangue. Il glucagone, invece, composto da 29 residui aminoacidici, è un ormone controregolatore dell’insulina. Aumenta i livelli di glucosio plasmatico in risposta all’ipoglicemia indotta dall’insulina e svolge un ruolo importante nell’omeostasi del glucosio aumentando la gluconeogenesi e diminuendo la glicolisi. Il terzo ormone pancreatico rilevante per il controllo dei livelli di glucosio nel sangue è la somatostatina, un peptide di 14 residui che esercita effetti inibitori sulla secrezione di insulina ma non sulla sua biosintesi. L’intero meccanismo della sua azione non è ancora stato chiarito, ma gli studi suggeriscono un effetto paracrino che inibisce l’esocitosi dell’insulina dalle cellule adiacenti alle cellule D produttrici di somatostatina (Reichlin 1983).
Tuttavia, gli effetti dell’insulina sull’intero organismo sono molteplici e complessi. L’insulina provoca, ad esempio, la traslocazione del GLUT-4 (il trasportatore di glucosio che si trova prevalentemente nel muscolo scheletrico e nel tessuto adiposo) dalle vescicole intracellulari alla membrana cellulare e, quindi, aumenta la velocità di ingresso del glucosio per una determinata concentrazione nel tessuto bersaglio. Un eccesso di glucosio trasferito nelle cellule stimola successivamente la formazione di glicogeno (Halse et al. 2001; Yeaman et al. 2001), un fatto di notevole interesse negli sport di resistenza, dove la quantità di glicogeno immagazzinata nelle cellule muscolari può influenzare le prestazioni atletiche. Inoltre, il metabolismo proteico (muscolare) è significativamente influenzato dalle proprietà chaloniche (Rooyackers e Nair 1997; Sonksen 2001) e stimolanti dell’insulina (Biolo et al. 1995; Biolo e Wolfe 1993; Tipton e Wolfe 2001; Wolfe 2000, 2005). Grazie all’effetto anticatabolico dell’insulina, la disgregazione proteica è significativamente ridotta e consente la conservazione degli elementi muscolari contrattili. Inoltre, in numerosi studi che hanno dimostrato le proprietà anaboliche dell’insulina, sono stati osservati effetti di stimolazione della sintesi, utilizzando ad esempio analoghi marcati con isotopi stabili. L’anabolismo, tuttavia, è fortemente dipendente dalla disponibilità di aminoacidi (Fujita et al.2006; Garlick e Grant 1988; Zhang et al.1999).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
Giova anche sottolineare i punti chiave dell’effetto dell’Insulina sul metabolismo delle proteine:
- aumenta la velocità di trasporto di alcuni aminoacidi nei tessuti
- aumenta il tasso di sintesi proteica nel muscolo, nel tessuto adiposo, nel fegato e in altri tessuti
- diminuisce il tasso di degradazione delle proteine nel muscolo (e forse anche in altri tessuti)
- diminuisce la velocità di formazione dell’urea. Questi effetti dell’insulina servono a favorire la sintesi di carboidrati, grassi e proteine.[Newsholme, E., & Dimitriadis, G. (2001).]
Un ormone anabolico può indurre l’ipertrofia del muscolo scheletrico attraverso:
- l’aumento della MPS o
- la riduzione della proteolisi.
- Inoltre, il trasporto transmembrana degli AA (AA trafficking) può essere sotto controllo ormonale.
L’Insulina induce l’ipertrofia del muscolo scheletrico attraverso tutti e tre questi aspetti del metabolismo proteico muscolare:
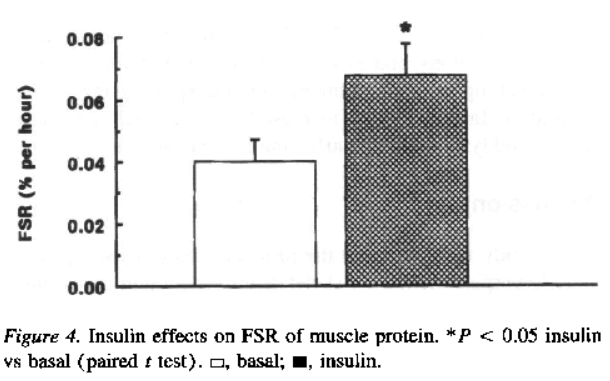
- profondamente (“il tasso di sintesi frazionale delle proteine muscolari era superiore del 65% (P = 0,02) durante l’infusione di insulina”) [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)].
- in misura modesta (nelle proteine non miofibrillari del muscolo scheletrico)
- influenzando l’attività di almeno quattro distinti sistemi di trasporto degli AA (non è un mediatore primario dell’azione dell’insulina sull’ipertrofia del muscolo scheletrico):
- Primariamente: trasporta gli AA con catene laterali polari corte, come l’Alanina e la Glicina; non limita il ritmo della sintesi proteica.
- ASC
- Nᵐ, sostanzialmente
- Xc [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)]
In pazienti gravemente ustionati l’infusione di Insulina (10 – 12 UI/ora, paziente di 70 kg) ↑MPS senza influenzare la MPB.[Hadley JS et al.2002]. Nessun cambiamento nell’assorbimento degli AA nel muscolo scheletrico, suggerendo che gli effetti anabolici muscolari derivino da un più efficiente riutilizzo degli AA generati dalla proteolisi.[Hadley JS et al.2002]
La sintesi proteica del muscolo scheletrico indotta dall’Insulina è modulata dalle variazioni di:
- del flusso sanguigno muscolare e
- disponibilità di AA. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)]. In altre parole, l’insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
- Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in mancanza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)].
Adesso è giunto il momento di aprire una parentesi sul GH…
Effetti dell’Ormone della Crescita e del IGF-1 sulla sintesi proteica:
La maggior parte degli studi svolti sul GH, quando considerati nel loro insieme, suggeriscono che il peptide in questione sia anabolico. Più specificamente, il GH è anabolico perché stimola la sintesi proteica di tutto il corpo con o nessun effetto, o un effetto soppressivo, sui tassi di degradazione proteica. [Møller N et asl. 2007] Tuttavia, quando si approfondisce l’argomento, le cose diventano un po’ meno chiare dal momento che i risultati degli studi tendono ad essere diversi. I diversi risultati sono un riflesso diretto dell’ immensa complessità del GH.
Il GH esplica i suoi effetti sulla sintesi proteica legandosi prima con il suo recettore specifico (GHR) e successivamente aumentando la trascrizione del gene muscolare attraverso i percorsi di segnalazione a valle, in definitiva attivando la segnalazione del mTOR. [Hayashi AA et al.2007] Questi effetti si manifestano in acuto, spesso si verificano in pochi minuti e sono di natura simile all’Insulina, usando molte delle stesse vie anaboliche. [Costoya JA et al.1999] La rapida comparsa di questi cambiamenti metabolici legati alle proteine suggerisce che essi siano direttamente causati dal GH e non secondariamente mediati tramite l’IGF-1 [Copeland KC et al.1994]. L’impatto del GH sulla proteolisi, d’altro canto, è molto probabilmente di natura indiretta. A detta di tutti, ciò ha più a che fare con i suoi effetti inibitori sull’Insulina, che è stata vista avere effetti diretti sulla proteolisi.[Umpleby AM et al.]
È stato dimostrato che l’IGF-1 inibisce allo stesso modo la disgregazione proteica a livello sistemico [Fryburg DA et al.1994], il che avrebbe senso a causa della stretta correlazione con il GH. Quando gli amminoacidi e l’Insulina vengono somministrati ai soggetti esaminati, già sottoposti a somministrazione di IGF-1, è stato dimostrato, sia negli uomini che negli animali, che i tassi di sintesi proteica aumentano a livello sistemico [Jacob R et al.1996]. Vale la pena notare che l’IGF-1 è bifasico nel senso che quando è somministrato ad alto dosaggio e, di conseguenza, i livelli serici diventano elevati, il suo comportamento cambia passando da un azione “GH-simile” ad una “insulino-simile”.
Per riassumere, il GH è molto adatto per prevenire la degradazione proteica, e lo fa in una vasta gamma di condizioni alimentari a ristretto apporto calorico. Tuttavia, in presenza di un apporto energetico sufficiente (e di AA), il suo comportamento cambia. L’effetto principale del GH sul metabolismo proteico è volto dapprima a creare un ambiente con una ossidazione amminoacidica ridotta [Buijs MM et al.2002] e successivamente ad aumentare la sintesi proteica sistemica. [Gibney J et al.2005]
L’Ormone della Crescita è noto per aumentare i livelli di IGF-1 circolante così come la sintesi locale di IGF-1, in modo autocrino. Entrambe queste azioni giocano un ruolo fondamentale nella regolazione della massa muscolare e, quindi, in conseguenza di ciò, risulta utile comprendere meglio come la secrezione di GH porta ad un aumento dei livelli endocrino e autocrino di IGF-1.
La stragrande maggioranza dell’Ormone della Crescita negli adulti sani è secreto dalla ghiandola pituitaria e, più specificamente, dalle cellule somatotrope nel lobo anteriore mediate dal fattore di trascrizione Prophet of Pit-1 (PROP1).[Hemchand K et al.2011] Il GH può anche essere sintetizzato localmente in molti tessuti come il cervello, le cellule immunitarie, il tessuto mammario, i denti e la placenta che sono tutti al di fuori della regolazione dell’ipofisi. [Waters MJ et al.1999] Questo supporta l’idea secondo cui il GH abbia ruoli autocrini oltre ai suoi già consolidati ruoli endocrini.
Il GH circolante si lega al GHR, recettore dell’Ormone della Crescita appartenente alla superfamiglia delle proteine transmembrana presenti in tutte le cellule del corpo e che include il recettore della Prolattina e un certo numero di recettori delle citochine [Zou L et al. 1997]. I livelli sulla superficie cellulare, o la densità recettoriale, dei GHR sono il determinante principale dell’affinità di legame del GH con le cellule. La traslocazione del GHR, cioè il recettore che si sposta dal nucleo di una cellula alla sua membrana esterna, è direttamente inibita dall’IGF-1 – che è uno dei molti meccanismi di feedback che esistono tra questi ormoni strettamente correlati. Mediante l’inibizione della traslocazione dei GHR, l’IGF-1 contribuisce direttamente ad abbassare la reattività di queste cellule a uno stimolo esterno di GH. [Leung KC et al.1997]
I GHR esistono sulle membrane cellulari come omodimeri preformati e inattivi. Questo significa che il GHR ha due dimeri identici del recettore della proteina, e questi omodimeri saranno sempre accoppiati al JAK2 quando sono privi di attività enzimatica. Questo accoppiamento al JAK2 provoca un’azione inibitoria complessiva sul recettore. [Sawada T et al.2017] In altre parole, il GHR rimane dormiente finché non viene attivato come parte del processo di legame GH/GHR. Quando una molecola di GH si lega al GHR, si verifica un cambiamento strutturale all’interno del GHR che si traduce in movimento effettivo dei domini intracellulari del recettore separatamente l’uno dall’altro. Questo smorza quell’azione inibitoria data dal JAK2 e consente loro di attivarsi l’un l’altro.[Brown RJ et al.2005]
Successivamente, la molecola di GH si lega sequenzialmente a uno dei due omodimeri di GHR e il completamento di questo processo di legame facilita le interazioni con il secondo omodimero. Dopo questo, i domini intracellulari di questo dimero-GHR appena formato subiscono una rotazione effettiva. La rotazione del nuovo dimero-GHR consente ai domini chinasi del JAK2 di essere in contatto l’uno con l’altro, consentendo loro di transactivare e ciascuno successivamente si lega a una molecola di JAK2. [Lanning NJ et al.2006] Ciascuna molecola di JAK2 eseguirà quindi la fosforilazione incrociata (attivazione) dei residui di Tirosina, e sono proprio questi residui che formano “punti di attracco” per molte delle diverse molecole di segnalazione che costituiscono le vie di segnalazione a valle, e alla fine portano all’espressione genica. [Brooks AJ et al.2010] Uno dei più importanti percorsi a valle di maggior interesse per l’argomento trattato è il percorso JAK-STAT. Questo percorso è di vitale importanza sia per la trascrizione epatica di IGF-1 dal GH che per molti dei processi anabolici mediati dal GH all’interno del tessuto muscolare.
- IGF-1:

Gli IGF sono una famiglia di peptidi, in gran parte dipendenti dal GH, che mediano molte delle azioni di stimolo della crescita date dal GH.[Cohen P. et al.2006] Il fegato è il principale responsabile di tutta la produzione endocrina di IGF-1, con circa il 75% della sintesi a carico epatico sotto la regolazione del GH.[Laron Z. et al. 2001] Ciò presuppone che ci sia un apporto macro-calorico sufficiente e livelli elevati di Insulina portale.[D’Ercole AJ et al.1984] La sintesi autocrina di IGF-1 è anche regolata dal GH, in aggiunta ad altri fattori autocrini dipendenti dal tessuto.[Gunawardane K et al.2000]
Alla famiglia degli IGF appartengono oltre dieci proteine strutturalmente simili tra cui IGF-1, IGF-2, Insulina, Relaxina e Pro-Insulina.[Lu C et al.2005] Sono tutti altamente omologhi sia nella struttura che nella funzione e gli effetti metabolici dell’IGF-1 sono stati definiti “insulino-simili” proprio a causa delle somiglianze e dei percorsi che condividono l’uno con l’altro. L’IGF-1 ha un’omologia di sequenza aminoacidica superiore al 50% con l’Insulina e il recettore del IGF-1 ha un’omologia della sequenza aminoacidica del 60% con il recettore dell’Insulina.[Samani AA et al.2006] A causa di queste somiglianze strutturali, i membri della famiglia IGF possono spesso legarsi con i recettori nativi in modo “incrociato”.[Kim JJ et al.2002] Per riassumere brevemente queste relazioni di legame, la molecola di IGF-1 si lega con il recettore del IGF-1 con un elevata affinità, tuttavia sia l’IGF-2 che l’Insulina possono legarsi al recettore del IGF-1, ma con una affinità significativamente inferiori. L’IGF-2 si lega al recettore del IGF-2 con una elevata affinità, e l’IGF-1 si lega a questo recettore con un’affinità inferiore mentre l’Insulina non presenta alcun legame con esso.
La famiglia dei recettori IGF ha densità che variano significativamente in base ai tipi di cellule in cui sono presenti.[Clemmons DR. et al. 2012] Questo è uno dei motivi per cui l’Insulina e l’IGF-1 possono avere diverse azioni metaboliche nonostante siano strutturalmente simili. Cellule come gli epatociti e gli adipociti hanno molti più recettori dell’Insulina rispetto ai recettori del IGF-1. Al contrario, le cellule muscolari lisce vascolari situate nei vasi sanguigni hanno un numero significativamente più elevato di recettori del IGF-1 rispetto ai recettori dell’Insulina.
E’ necessario comprendere che la famiglia dei recettori IGF è anche attivata dalla Tirosina Chinasi che, come ora sappiamo, porta alla fosforilazione dei substrati, all’attivazione delle vie cellulari e infine all’espressione genica e alla sintesi proteica.[Cohen P. et al.2006] L’attivazione del recettore del IGF-1 sembra essere indipendente dall’isoforma da cui è stato prodotto l’IGF-1. Inoltre, si noti che entrambi i tipi di recettori IGF sono stati trovati nelle cellule muscolari umane.[Shimizu M et al.1986]
Nel flusso sanguineo, l’IGF-1 esiste principalmente in forma legata a proteine leganti l’IGF (IGFBP). La superfamiglia IGFBP comprende sei proteine ad alta affinità che vanno dal IGFBP-1 al IGFBP-6, nonché un certo numero di proteine a bassa affinità denominate proteine legate all’IGFBP.[Hwa V et al.1999] Quasi il 95% di tutto l’IGF-1 circolante esiste in forma legata, con circa il 75% legato specificamente con l’IGFBP-3.[Firth SM et al.2002] Una piccola frazione di IGF-1 (normalmente inferiore al 5%) può anche esistere in forma libera, e queste molecole non legate agiscono come regolatore negativo della secrezione di GH. Gli IGFBP possono legarsi con l’IGF-1 e l’IGF-2, ma non con l’Insulina. [Bach LA et al.1993] L’IGF-1 legato esiste più comunemente in un complesso ternario da 150-kDa mentre è nel circolo ematico. Questo complesso ternario è costituito da una molecola di IGF-1, dal IGFBP-3 e dalla subunità labile acida (ALS) – sebbene possa esistere in un complesso binario con altri IGFBP.[Duan C. et al.1996] Questi complessi servono a scopi come l’aumento della biodisponibilità degli IGF circolanti, estendendo la loro emivita serica, trasportando gli IGF alle cellule bersaglio e modulando l’interazione degli IGF con i loro rispettivi recettori di membrana posti sulla superficie delle cellule.[Hwa V et al.1996] Ad esempio, nel plasma, il complesso ternario stabilizza IGF-1, aumentando significativamente la sua emivita da meno di 5 minuti a oltre 16 ore in alcuni casi.[Firth SM et al. 2002]

Gli IGFB sembrano normalmente inibire l’azione degli IGF, e questo perché competono con i recettori IGF per l’affinità di legame con gli IGF.[Collett-Solberg PF et al.2000] Tuttavia, non è sempre così, poiché gli IGFBP sono anche in grado di potenziare le azioni dell’IGF, potenzialmente facilitando la consegna dell’IGF al recettore.[Wetterau LA et al. 1999] Sebbene esista un’interazione piuttosto complessa, basti ricordare che il ruolo principale degli IGFBP è quello di trasportare gli IGF dal flusso ematico ai tessuti periferici. Una volta che ciò è avvenuto, gli IGFBP vengono rilasciati dai complessi binari e ternari mediante proteolisi o tramite legame alla matrice extracellulare del recettore del IGF-1.[Parker A et al.1999] Una volta rilasciate, le molecole di IGF-1 diventano libere, attive e possono quindi esplicare la loro azione.[Monzavi R et al.2002]
Una volta nei tessuti, gli IGFBP modulano le azioni dell’IGF in quanto hanno una maggiore affinità per il sito recettore rispetto all’IGF stesso [Velloso CP et al.2008], tuttavia essi possono anche esercitare effetti indipendenti dall’IGF.[Jones JI et al.1995] Alcuni degli effetti diretti del IGFBP che sono già stati chiariti includono l’inibizione della crescita, l’induzione diretta dell’apoptosi e la modulazione degli effetti dei fattori di crescita non-IGF.[Cohen P. et al.2006]
Additività Insulina/rhGH sovrafisiologici:
Come ho già riportato in passato, la somministrazione di Insulina esogena capace di creare un livello ematico oltre i 1.200pmol/lt ha effetti significativi e tangibili sulla sintesi proteica muscolare. Ma non si esaurisce tutto a meri e limitati numeri statistici.
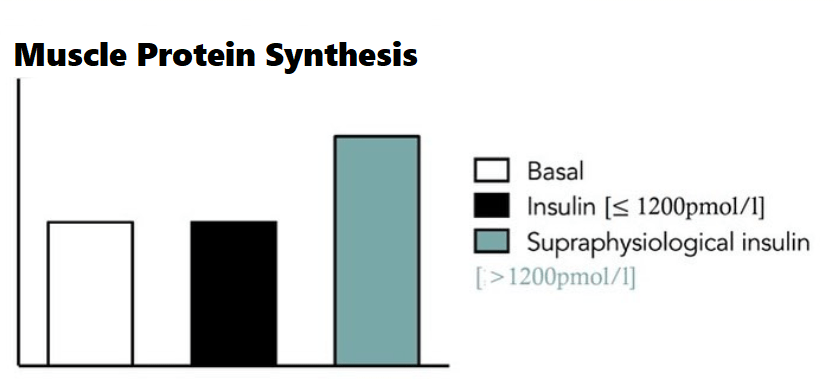
La sintesi proteica nel muscolo-scheletrico indotta dall’Insulina è modulata dalle variazioni di:
1- del flusso sanguigno muscolare e
2- disponibilità di AA. [Fujita, S. et al. (2006).]. In altre parole, l’Insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
- Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in assenza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S. et al. (2006)].
l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1, riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1. L’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1. Il discorso precedentemente fatto sulle IGFB deve essere compreso correttamente alla luce di modifiche sostanziali in concentrazione (di IGF-1) e sua frazione legata (minoritaria ma presente).
Quindi, l’Insulina può aumentare la biodisponibilità di IGF-I inibendo la produzione di IGFBP-1 e aumentando l’attività proteolitica di IGFBP-3, che riduce l’affinità di IGF-I per IGFBP-3. [Hadley JS et al. 2002]. Esemplificato in formula:
- IGF-I (libero) ↑AA e assorbimento del glucosio e ripartizione delle proteine.[Hadley JS et al. 2002]
L’Insulina dirige la MPS attraverso l’attivazione diretta della via PI3K/Akt/mTORC1 e grazie all’aumento del flusso sanguigno muscolare tramite proprietà vasoattive.[Trommelen et al. 2015]
Variabile della disponibilità intracellulare di AA:
Come già alcuni di voi sapranno, la prima fase della sintesi proteica avviene nel nucleo delle cellule e comporta la trascrizione di un gene specifico in molecole di mRNA. Gli mRNA sono trasportati nel citosol, dove si associano ai ribosomi per la traduzione della sequenza di basi in una sequenza di AA. Il processo di traduzione può essere suddiviso in tre fasi: iniziazione, allungamento e terminazione della catena peptidica. Molti ribosomi possono associarsi a molecole di mRNA, formando poliribosomi. Un poliribosoma può sintetizzare diverse catene peptidiche da un singolo mRNA. Gli AA liberi nel citoplasma non vengono utilizzati direttamente per la sintesi proteica. Il processo di traduzione prevede il legame degli AA a specifiche molecole di RNA di trasferimento (tRNA), formando gli aminoacil-tRNA. Gli aminoacil-tRNA costituiscono pool molto piccoli che si trasformano rapidamente. [Biolo, G., & Wolfe, R. R. (1993)]
È stato dimostrato che l’insulina stimola sia i processi di trascrizione che di traduzione di proteine specifiche (Kimball e Jefferson, 1988). È stato dimostrato che l’espressione genica (riflessa dai livelli di mRNA) di molte proteine è stimolata dall’insulina, tra cui l’albumina nel fegato e l’MHC α nel muscolo scheletrico (Dillman, 1988). Il contenuto di mRNA riflette il potenziale ultimo di sintesi proteica nella cellula (tessuto), ma potrebbe non essere direttamente correlato al tasso di traduzione e quindi di sintesi delle proteine. L’insulina migliora anche il processo di traduzione nel muscolo scheletrico, stimolando l’iniziazione della catena peptidica (Jefferson et al, 1974; Fulks e Goldberg, 1975). Pertanto, da una base molecolare, ci si aspetterebbe che in vivo l’insulina aumenti in generale la capacità di sintesi proteica (contenuto di mRNA) e stimoli la traduzione e la produzione di proteine specifiche, come quelle muscolari. [Biolo, G., & Wolfe, R. R. (1993)]
Esistono due meccanismi attraverso i quali l’aumento dell’AA intracellulare può stimolare la sintesi proteica muscolare. In primo luogo, un effetto di massa dovuto agli AA esogeni forniti; il tRNA necessario per la traduzione degli AA in proteine è disponibile nel muscolo in quantità superiori al pool di AA disponibile. Di conseguenza, l’aumento della disponibilità di AA spingerà la MPS semplicemente caricando più tRNA. [Tipton, K. D., & Wolfe, R. R. (2001).].
È anche possibile che alcuni AA, singoli o gruppi di AA (ad esempio, i BCAA), segnalino l’avvio del processo di traduzione (ribosomiale). Rennie et al. hanno dimostrato che i singoli AA essenziali somministrati in dosi massicce (cioè riempiendo lo spazio AV e lo spazio intracellulare) stimolano la MPS, mentre gli AA non essenziali non hanno questo effetto. Inoltre, abbiamo recentemente dimostrato che non è necessario includere gli AA non essenziali in una soluzione che simula l’anabolismo muscolare in volontari a riposo . [Tipton, K. D., & Wolfe, R. R. (2001).].
Questi studi sono coerenti con l’ipotesi che uno o più degli AA essenziali possano agire come segnale per l’avvio delle MPS. Tuttavia, la stimolazione della sintesi proteica attraverso l’azione di massa non può essere esclusa solo sulla base di queste evidenze. È possibile che siano presenti sufficienti AA non essenziali e che l’aggiunta di grandi quantità di AA essenziali fornisca una carica di tRNA sufficiente a guidare la sintesi proteica. [Tipton, K. D., & Wolfe, R. R. (2001)].
Il punto di vista di Wolfe: L’insulina sembra aumentare l’efficienza della reincorporazione degli AA derivanti dalla disgregazione delle proteine (intracellulare) nelle proteine di nuova sintesi. [Biolo et al. (1995)] [Wolfe, R. R. (2000)].
- Disponibilità di AA come segnale per attivare la traduzione
Sistema ubiquitina-proteosoma (sistema MPB predominante nel muscolo a riposo)
I fattori di iniziazione eucariotici (eIF), in particolare eIF4E, sono componenti importanti del controllo dell’iniziazione della traduzione. Quando eIF4E forma un complesso con eIF4G, viene promossa l’iniziazione della traduzione. Il legame di eIF4E con eIF4G è normalmente impedito perché eIF4E è legato a una proteina vincolante, eIF4E-BP1, la cui fosforilazione libera eIF4E. L’apporto di AA favorisce la sua fosforilazione, permettendo così alla sintesi proteica di procedere . In altre parole, la formazione del complesso attivo eIF4E-eIF4G aumenta in risposta alla somministrazione di proteine. [Tipton, K. D., & Wolfe, R. R. (2001)].
- La disponibilità di AA come chalonico per la proteolisi muscolare
Anche il pool intracellulare di AA sembra seguire un effetto di massa per evitare la disgregazione delle proteine muscolari.
Un aumento degli AA intracellulari inibisce la MPB solo in determinate condizioni. Ad esempio, l’iperamminoacidemia a riposo ha un effetto minimo o nullo sulla MPB. Tuttavia, quando gli AA vengono somministrati dopo la RT, quando la MPB sarebbe normalmente elevata, non si verifica alcun aumento della MPB. Questa inibizione condizionale della MPB è coerente con l’idea che questa relazione tra disponibilità di AA e riduzione della MPB si verifichi principalmente quando il pool intracellulare di AA disponibili si esaurisce. Ad esempio, se la risposta iniziale all’esercizio fisico è un aumento della MPS, il pool di AA si riduce, portando alla MPB per mantenere il pool intracellulare di AA. [Tipton, K. D., & Wolfe, R. R. (2001)].
In alternativa, poiché esistono diverse vie di MPB, è possibile che in seguito all’esercizio fisico la via lisosomiale diventi predominante nel mediare l’accelerazione della MPB, e che questa via sia reattiva alla disponibilità di AA [e all’insulina esogena!] [Tipton, K. D., & Wolfe, R. R. (2001)].
…L’unico effetto probabile dei carboidrati e dei grassi sul bilancio proteico netto è l’influenza ormonale come substrati [ad esempio, l’influenza dei CHO sulla secrezione di Insulina]. [Tipton, K. D., & Wolfe, R. R. (2001)].
- Trasporto transmembrana AA
Alanina +48%, Lisina +75%, Leucina +22%. [Biolo et al. (1995)].
L’alanina è uno dei principali substrati del sistema A, un sistema sodio-dipendente che mantiene ripidi gradienti transmembrana delle concentrazioni di AA ed è regolato dall’insulina [Biolo et al. (1995)] . Inoltre, la sintesi di alanina nella cellula muscolare aumenta a causa dell’incremento insulino-mediato dell’assorbimento di glucosio e della produzione intracellulare di piruvato nel muscolo. [Biolo et al. (1995)].
La lisina è un AA cationico che viene trasportato dal sistema sodio-indipendente y⁺ che è fortemente influenzato dal potenziale elettrochimico della membrana cellulare. Poiché l’insulina induce un’iperpolarizzazione nelle cellule muscolari scheletriche attivando direttamente la pompa Na⁺-K⁺-ATPae, l’accelerazione del trasporto della lisina può essere secondaria all’effetto primario dell’insulina sul gradiente elettrochimico di transmembrana. [Biolo et al. (1995)].
Gli AA a catena ramificata (leucina, isoleucina e valina) e aromatici (fenilalanina e tirosina) vengono trasportati preferibilmente attraverso il sistema L. Questo sistema, indipendente dal sodio, non è in grado di generare elevati gradienti transmembrana per i suoi substrati. È stato dimostrato che le caratteristiche cinetiche del sistema L non sono influenzate dall’insulina. [Biolo et al. (1995)].
L’azione dell’insulina sulla leucina è particolarmente modulata dall’aumento del flusso sanguigno muscolare! La leucina è soggetta a ossidazione e transaminazione che devono essere sottratte dal tasso di scomparsa nel muscolo per ottenere i tassi di sintesi proteica con la tecnica del bilancio arteriovenoso. [Rooyackers et al. (1997).].
In presenza di un adeguato apporto di AA, il sistema degradativo più importante nel muscolo è il sistema dell’ubiquitina indipendente dall’ATP. [Biolo et al. (1995)]. Questo sistema non è sensibile all’insulina [Biolo et al. (1995)]. L’insulina sembra avere un ruolo solo nella regolazione dell’attività dei lisosomi [Biolo et al. (1995)]. I lisosomi (organelli) non sono coinvolti nella degradazione delle proteine miofibrillari in condizioni normali, ma solo in presenza di bassi livelli di insulina o di una ridotta disponibilità di AA. [Biolo et al. (1995)].
La riduzione della degradazione proteica del muscolo scheletrico indotta dall’insulina si verifica in proteine non miofibrillari (soprattutto epatiche). [Rooyackers et al. (1997)].
E’ utile inoltre ricordare che il GH aumenta sia l’assorbimento degli AA dal lume intestinale che a livello cellulare nel muscolo-scheletrico, azione, quest’ultima, che va a sommarsi a quella esercitata da IGF-1 e Insulina.
Conclusioni:
Qual conclusione allora?
Oggettivamente, parlare di protocolli di Insulina/hGH ha senso quando l’atleta in questione è un avanzato, con una carriera “Natty” alle spalle ad OC e una esperienza ben controllata e graduale con i PEDs. Ma andando oltre a ciò che dovrebbe essere scontato, da quanto riportato in questo articolo, si può ben capire che essa rappresenti una pratica con un margine di guadagno in termini ipertrofici decisamente significativo.
I punti chiave degli effetti consequenziali del protocollo Insulina/hGH sono:
- Aumento della sensibilità epatica al GH per via dell’attività insulinica di picco;
- risposta massiva consequenziale sulla sintesi e rilascio sistemico di IGF-I;
- riduzione delle proteine di trasporto IGFBP-1 e 3 insulino-correlato;
- aumento della frazione libera e attiva di IGF-I.
I limiti della presente disamina non sono pochi basandosi in gran parte su dati teorici ed estrapolazioni aneddotiche tratte da piccoli casi studio. Ma ciò nonostante è sufficiente a far comprendere, almeno a livello base, come le modifiche dell’omeostasi attraverso l’uso di farmaci possa dare risultanti anche di molto distanti da quelle ottenute in contesti fisiologici.
Inoltre, se volessimo trovare un altro limite al presente lavoro, non sono stati trattati gli effetti sommativi dati dalla co-assunzione di AAS. Ciò è ovviamente legato al fatto di non volere disperdere l’attenzione del lettore dal concetto fondamentale trattato: la validità dei protocolli Insulina/hGH.
Sicuramente, i vantaggi del protocollo Insulina/hGH vanno ben oltre il banale e poco sensato assunto secondo il quale l’Insulina esogena vada semplicemente a compensare in un certo senso la ridotta sensibilità all’Insulina GH correlata.
Gabriel Bellizzi [CEO BioGenTech]
