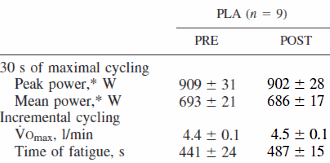Quanto sono realmente efficaci gli AAS nello stimolo della crescita muscolare?
Riportare in sintesi il contenuto di un vecchio studio che prende in esame l’uso di dosi sovra fisiologiche di Testosterone per rispondere a tale quesito potrebbe sembrare al quanto superfluo. Ma, in realtà, per il suo spettro d’analisi, non risulta affatto superfluo o scontato per farsi un idea del potenziale di questi abusati composti.
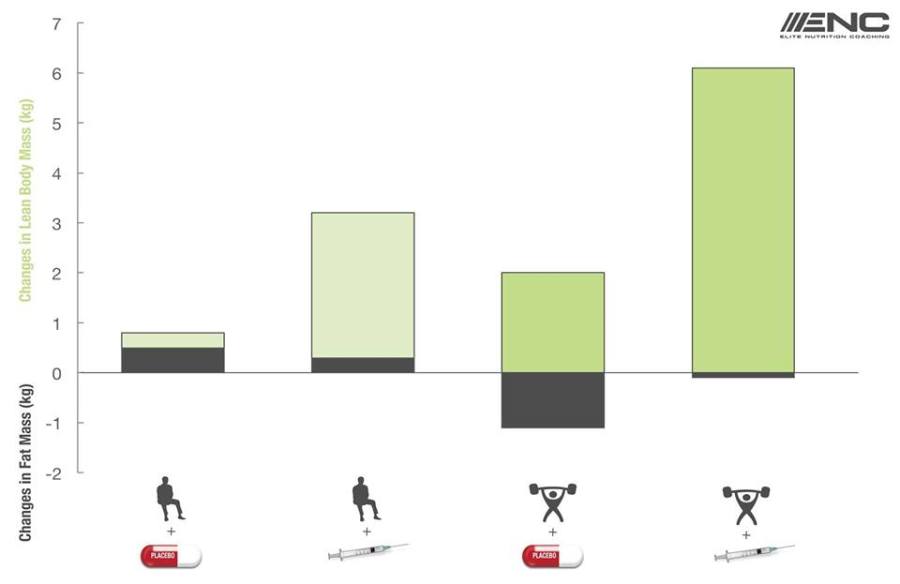
Il seguente grafico riporta i dati conclusivi dello studio pubblicato nel 1996.(1)
Fonte immagine(2).
Lo studio è stato suddiviso in tre periodi distinti:
Un periodo di controllo di 4 settimane;
Un periodo di trattamento di 10 settimane;
Un periodo di recupero di 16 settimane.
Durante il periodo di controllo di quattro settimane, ai soggetti dello studio è stato chiesto di non sollevare pesi o di impegnarsi in un intenso esercizio aerobico.
I soggetti presi in esame (43 uomini allenati per la forza) sono stati suddivisi in quattro gruppi:
No esercizio più placebo
No esercizio più 600mg di Testosterone Enantato a settimana
Esercizio più placebo
Esercizio più 600mg di Testosterone Enantato a settimana
I soggetti del gruppo “Esercizio più placebo” e “Esercizio più 600mg di Testosterone Enantato a settimana” sono stati sottoposti ad un programma di allenamento con i pesi su 3 giorni a settimana per 10 settimane.
Due settimane prima del giorno 1, ai soggetti dello studio (di tutti e 4 i gruppi) è stato chiesto di seguire una dieta giornaliera standard contenente 36kcal per Kg di peso corporeo, con una quota proteica pari a 1,5g per Kg e un assunzione del 100% della dose giornaliera raccomandata di vitamine, minerali e oligoelementi. La conformità con la dieta è stata verificata ogni quattro settimane con registrazioni di tre giorni del consumo di cibo. L’introito alimentare è stato aggiustato ogni due settimane sulla base dei cambiamenti nel peso corporeo.
Dei 43 soggetti reclutati per lo studio, 3 sono usciti durante la fase di trattamento: 1 a causa di problemi di conformità, 1 perché faceva uso di droghe illegali rilevate dallo screening di routine e 1 a causa di un incidente automobilistico. Quaranta uomini hanno completato lo studio: 10 nel gruppo placebo, nessun esercizio; 10 nel gruppo Testosterone, nessun esercizio; 9 nel gruppo placebo più esercizio; e 11 nel gruppo di Testosterone più esercizio.
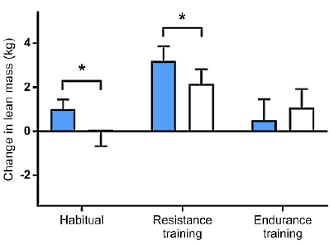
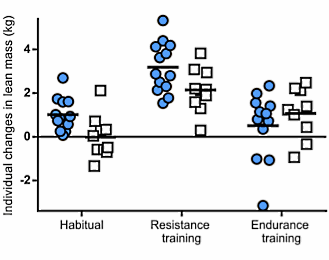
La cosa che può stupire maggiormente vagliando i risultati dello studio è che il gruppo che non si è allenato e ha ricevuto la somministrazione settimanale di Testosterone per dieci settimane ha avuto un guadagno di massa muscolare maggiore rispetto al gruppo che si allenava e veniva trattato con il placebo (3,2 vs 2kg).
I restanti risultati sono decisamente prevedibili, con il gruppo sottoposto ad allenamento e alla somministrazione di Testosterone che presenta i guadagni maggiori in termini di massa muscolare (6,1kg).
La forza muscolare espressa nel beach-press e negli squat non ha mostrato cambiamenti significativi nel corso del periodo di 10 settimane nel gruppo placebo senza esercizio. I soggetti del gruppo Testosterone senza esercizio e quelli del gruppo placebo più esercizio hanno avuto aumenti significativi nel carico massimale negli esercizi di squat, con una media rispettivamente del 19% e del 21%. Allo stesso modo, la forza media nella bench-press è aumentata in questi due gruppi rispettivamente del 10% e dell’11 %. Nel gruppo Testosterone più esercizio, l’aumento della forza muscolare durante l’esercizio di squat (38%) è stato maggiore rispetto a quello di qualsiasi altro gruppo, così come l’aumento della forza nella bench-press (22%).
Nota interessante: Nessuno dei soggetti presi in esame (tutti e quattro i gruppi) ha mostrato cambiamenti nell’umore nel comportamento.
Non fraintendetemi, con questo nessuno vuole affermare che gli atleti che assumono farmaci non si allenino duramente. Loro fanno e come. In realtà, gli atleti supplementati chimicamente tendono ad allenarsi più intensamente in senso assoluto, anche a causa delle maggiori capacità di recupero offerta dall’uso di AAS.
Per fare una sintesi di quanto esposto e rispondere alla domanda iniziale: quanto sono efficaci gli AAS nello stimolo della crescita muscolare? Beh, la loro efficacia è indubbia anche se le variabili sul espressione di tale efficacia sono legate essenzialmente alla base genetica dell’utilizzatore.
In questa terza parte della serie di articoli dedicati all’approfondimento sul Trenbolone descriverò nel dettaglio le proprietà anabolizzanti della molecola. Vale sempre la pena ribadire che tutti gli studi relativi agli argomenti qui trattati provengono da studi svolti sugli animali. Quindi, il mio obiettivo è quello di riportare ciò che è disponibile nella letteratura scientifica discutendo su ciò che è potenzialmente rilevante per l’essere umano e, più nel dettaglio, per il BodyBuilder. Molti meccanismi biologici riportati di seguito sono simili tra i mammiferi e l’uomo, e farò del mio meglio per sottolineare quando questo sussiste o meno.
Nota: se non avete letto la prima e la seconda parte vi invito a farlo prima di procedere con la lettura del presente articolo.
IX. Anabolismo
Prima di entrare nel dettaglio, è importante sottolineare una cosa ovvia ma che ad alcuni sfugge, e cioè che vi sono differenze tra gli esseri umani e gli animali più comunemente usati negli studi che hanno preso in esame il Trenbolone (ad esempio pecore, topi, mucche, ecc.). Mentre ci si addentra nell’analisi della documentazione scientifica disponibile, è utile che teniate a mente queste differenze, in quanto possono sicuramente avere un impatto rilevante per gli esseri umani nella loro differenza di risposta.
Levator ani
La maggior parte dell’apparato muscolo-scheletrico dei roditori possiede una percentuale molto bassa di nuclei AR positivi. Un esempio è dato dalle caratteristiche del extensor digitorum longus, situato vicino alla parte anteriore della zampa posteriore, il quale presenta solo il 7% di mionuclei AR positivi.(1) Questo però non è universalmente vero, poiché il complesso muscolare levator ani / bulbocavernosus (LABC) (situato vicino alla pelvi) presenta il 70-75% di mionuclei AR-positivi e, tale caratteristica, si traduce in una marcata risposta miotropica alla somministrazione di androgeni.(2)(3)(4) Quindi, se si prendono in esame diversi studi svolti sui roditori, nei quali sono stati analizzati i muscoli sopra riportati in modo separato, i risultati saranno nettamente differenti.
In alto un esemplare di Toro, in basso un esemplare di Manzo.
Viceversa, i bovini sono generalmente molto sensibili agli stimoli indotti dagli androgeni a causa delle alte concentrazioni di AR nel loro apparato muscolo-scheletrico e nelle loro cellule satelliti.(5)(6)(7) E’ necessario capire anche che, quando si parla di tori, si parla di bovini di sesso maschile maturi e intatti, mentre quando si parla di manzi, si parla di bovini di sesso maschile i quali hanno subito castrazione prima di raggiungere la maturità sessuale. La stragrande maggioranza degli studi esistenti è stata svolta sui manzo sottoposti ad impianti di Trenbolone, dal momento che tali impianti non presentano effetti rilevanti sui tori. Probabilmente questi animali hanno raggiunto il loro massimo potenziale di crescita muscolare data dai loro livelli elevati di endogeni endogeni, comunque sia sono necessari impianti che combinano TBA ed E2 per produrre la massima crescita ed efficienza alimentare nei bovini castrati.(8) Le giovenche sono giovani bovini di sesso femminile che non hanno mai partorito; sono usate occasionalmente per testare gli impianti.
I tori sintetizzano livelli molto elevati di Testosterone. Oltre ad avere una risposta molto scarsa agli impianti, generalmente hanno anche fibre muscolari più grandi rispetto ai manzi.(9) I tori tendono inoltre ad avere una percentuale più elevata di fibre glicolitiche-ossidanti a contrazione rapida combinate con una percentuale inferiore di fibre glicolitiche a contrazione rapida nei muscoli longissimus dorsi (LD) rispetto ai manzi.(10) È per questi motivi che i tori producono un totale di carne più elevate, anche se generalmente di qualità inferiore. I manzi tendono ad avere una percentuale di grasso più elevata ma, tuttavia, sono compensati da una minore velocità di aumento di peso e una minore efficienza alimentare. Quindi, nel tentativo di ottenere maggiori rese con carni di qualità superiore, i ricercatori hanno iniziato a studiare l’effetto degli impianti per verificare se fosse possibile ottenere il meglio di entrambe le caratteristiche che caratterizzano i tori ed i manzi .
Infine, una breve nota a margine: noi esseri umani siamo molto simili ai bovini per quanto concerne la risposta agli androgeni, in quanto rispondiamo in modo marcato agli stimoli androgenici a causa delle alte percentuali di mionuclei AR-positivi.(11)
Affinità per il Recettore degli Androgeni
È stato dimostrato che il Trenbolone si lega con il AR umano, e con il AR di varie specie animali, con una affinità approssimativa pari a tre volte superiore a quella del Testosterone, o approssimativamente uguale a quella del DHT.(12)(13)(14)(15)(16) Nei AR umani, il metabolita attivo 17β-TbOH ha mostrato un’affinità di legame del 109% rispetto al DHT, mentre il metabolita inattivo 17α-TbOH ha mostrato una affinità di legame del 4,5%.(13) Detto questo, gli studi sul legame recettoriale dovrebbero essere visti come uno strumento per una rapida valutazione iniziale di un ligando, non prendendo in considerazione aspetti come la successiva attivazione della trascrizione genica, ecc. In altre parole, anche se il Trenbolone presenta una affinità di legame al AR tre volte superiore rispetto al Testosterone, questo non significa letteralmente che produrrà un ipertrofia tre volte maggiore.
Oltre a questo, negli studi comparativi è stato dimostrato che il Trenbolone produce una crescita uguale o leggermente superiore nel complesso muscolare del LABC rispetto al Testosterone.(14)(17)(18)(19)(20)(21)(22) Il LABC è un tessuto androgeno reattivo che manca dell’enzima 5α-reduttasi. Mentre il Testosterone esercita effetti potenziati nei tessuti che esprimono l’enzima 5α-reduttasi, il Trenbolone esercita effetti uguali in questi tessuti rispetto a quelli che non causano una anabolico:androgeno ratio favorevole in comparazione al Testosterone.(23) Approfondirò questo argomento quando tratterò dei rischi di sviluppare cancro alla prostata.
X. Ipertrofia
Inizialmente avevo in programma di esporre un’analisi approfondita dei meccanismi alla base dell’ipertrofia, ma trattandosi di un argomento decisamente complesso occorrerebbe un articolo a parte esclusivamente dedicato a questa tematica. Avrò comunque bisogno di esporre alcuni elementi fondamentali dell’ipertrofia, altrimenti molto di quanto tratterò in seguito potrebbe risultare più complicato di quanto dovrebbe. Quindi, non mi dilungherò troppo nella descrizione dei percorsi di segnalazione intracellulare, poiché questo renderebbe il presente articolo inutilmente prolisso. Comunque, se l’argomento dovesse suscitare abbastanza interesse, forse in futuro realizzerò un articolo dedicato esclusivamente all’ipertrofia.
Fondamenti di ipertrofia
Prima di tutto è necessario parlare un po ‘di cos’è l’ipertrofia e di come essa si manifesta nei mammiferi. Questo è un passaggio volto a fornire al lettore nozioni base sull’argomento, affinché i termini utilizzati in seguito vengano meglio compresi.
Il numero di fibre muscolari nei mammiferi è essenzialmente fissato alla nascita, quindi la crescita muscolare postnatale è la risultante dell’ipertrofia delle fibre muscolari esistenti. Questa ipertrofia delle fibre richiede un aumento del numero dei mionuclei presenti all’interno di esse; tuttavia i nuclei presenti nelle fibre muscolari non sono in grado di dividersi, quindi, essi, devono provenire dall’esterno della fibra.(24) La fonte dei nuclei necessari per supportare l’ipertrofia delle fibre è rappresentata da un gruppo di cellule miogeniche mononucleate (cellule satelliti) situate tra la lamina basale e la membrana plasmatica (sarcolemma) delle fibre muscolari.(25) Esiste una forte correlazione tra i tassi di crescita postnatale e i tassi con cui le cellule satellite si accumulano all’interno dei tessuti muscolari. Ciò sembrerebbe avere un senso in quanto saranno disponibili più meccanismi generali per alimentare il processo ipertrofico.(26)
Queste cellule satelliti svolgono un ruolo cruciale nella crescita muscolare postnatale fondendosi con le fibre muscolari esistenti, fornendo i nuclei necessari per la crescita delle fibre postnatali. Negli animali appena nati, si riscontra una percentuale molto più alta di nuclei muscolari legati alle cellule satelliti, ma questa percentuale diminuisce significativamente con l’età.(27) Non solo c’è una riduzione del numero di cellule satelliti, ma quelle cellule ancora presenti si ritirano dallo stato proliferativo del ciclo cellulare e entrano in uno stato di quiescenza, che di conseguenza porta ad un plateau della crescita. Quindi, trovare modi per superare questi limiti fisiologici può ipoteticamente portare a tassi di crescita postnatale superiori.
Per garantire che ci sia un numero adeguato di cellule satelliti utilizzabili, devono prima essere attivate per consentire loro di progredire attraverso il ciclo cellulare e infine di contribuire con il DNA della fibra muscolare esistente. Dopo che queste cellule satelliti dormienti sono state attivate, vi è successivamente la necessità di un rilascio dei fattori di crescita in grado di stimolare la proliferazione e la differenziazione delle cellule satelliti. Sia l’IGF-1 che il Fattore di Crescita dei Fibroblasti 2 (FGF-2) sono esempi di potenti stimolatori della proliferazione delle cellule satelliti.(28)(29) L’IGF-1 è unico in quanto promuove la differenziazione delle cellule muscolari nei muscoli-scheletrici, mentre l’FGF-2 inibisce la differenziazione [30]. Tra poco parlerò più nel dettaglio della relazione tra Trenbolone e IGF-1.
Quindi, facendo un leggero passo indietro, quando si verifica un evento di attivazione dell’ipertrofia (ad esempio esercizio o danno muscolare), questo porta anche alla proliferazione delle cellule satelliti. Questa proliferazione delle cellule satelliti induce queste a fondersi con le fibre muscolari esistenti, fornendo nuovi nuclei per l’ipertrofia e la riparazione e per supportare l’aumentata sintesi proteica. Un modo semplice per comprendere quanto appena detto è il seguente: le cellule satellite possono essere attivate per proliferare (dividere) e donare il loro DNA (nuclei) alle fibre muscolari esistenti (differenziazione).
Questo DNA donato porta le fibre muscolari a generare la fusione di mioblasti (cellule proliferanti) in fibre muscolari multinucleate denominate miotubi. Questi miotubi possono fondersi con le miofibre esistenti, o anche l’uno con l’altro, generando direttamente nuove fibre muscolari. Per il momento questo è l’approfondimento necessario per tale l’argomento.
Effetti di stimolo della crescita
Gli effetti di stimolo della crescita dati dal Trenbolone sono ben noti e sono stati studiati dai ricercatori per decenni. L’obiettivo è sempre stato quello di trovare metodiche per promuovere maggiori rese di carne insieme ad un prodotto finito di qualità superiore. Ci concentreremo principalmente sulle rese di carne per ora, poiché la qualità della carne tende spesso a coincidere con la quantità di grassi intramuscolari ivi contenuti. Questo rientra maggiormente nel campo della lipolisi, che tratteremo nel prossimo articoli.
È stato dimostrato che il Trenbolone aumenta la crescita totale della massa corporea e della massa muscolo-scheletrica in vari studi svolti su animali quando somministrato da solo (3)(14)(17) (19)(31)(32)(33)(34)(35)(36)(37)(38)(39)(40)(41)(42)(43)(44), in combinazione con Estradiolo (45) (46) (47)(48)(49)(50)(51)(52)(53)(54)(55)(56)(57)(58)(59)(60)(61)(62)(63)(64)(65)(66), in combinazione con Testosterone ed Estradiolo (67), nonché in combinazione con Estradiolo e Ormone della Crescita.(68) Questo potenziale ipertrofico è stato universalmente osservato attraverso molteplici metodi di somministrazione in diverse specie animali.
È interessante notare che numerosi studi hanno dimostrato che un impianto contenente la combinazione TBA / E2 è più efficace rispetto ad un impianto contenente soltanto TBA o E2 nello stimolare la crescita dei manzi all’ingrasso.(8)(45)(52)(54)(55)(69)(70)(71)(72)(73)(74) L’ipotesi secondo la quale l’Estradiolo aumenterebbe gli effetti anabolizzanti del Trenbolone è emersa intorno agli anni ’70 (75)(76). Un fatto ancora più interessante è che il trattamento combinato (TBA/E2) aumenta il potenziale di ipertrofia nonostante il fatto che tale metodo si traduce in livelli di Trenbolone serici effettivamente inferiori, di circa la metà (8).
Asse GH/IGF-1
Uno dei motivi che mi spinge a credere nella concretezza di un effetto potenziale nella cosomministrazione di Trenbolone ed Estradiolo, in particolare nei manzi , è il fatto che gli impianti contenenti solo Trenbolone portano alla soppressione dei livelli di Estradiolo endogeno a causa del impatto del Trenbolone sull’asse HPG limitandone la crescita potenziale. L’Estrogeno e, più specificamente, l’attività dell’aromatasi sono un potente stimolatore dell’asse GH/IGF-1. A supporto di questa ipotesi, gli impianti con solo Trenbolone hanno dimostrato di abbassare i livelli serici di GH.(8,68,69,70,71) Al contrario, è stato dimostrato che i manzi impiantati con E2 hanno aumentato le concentrazioni ematiche sia di GH che di IGF-1.(77-78) Gli impianti contenenti la combinazione TBA / E2 probabilmente determinano un aumento dei livelli di GH e possono persino alterare il numero e/o l’affinità dei GHR nei tessuti come quello epatico.(79) Come abbiamo appreso in precedenza, avere livelli adeguati di fattori di crescita per stimolare la proliferazione e la differenziazione delle cellule satelliti è un fattore significativo nel processo ipertrofico.
TBA/E2 ratio ottimale
Poiché i trattamenti combinati sembrano avere caratteristiche anaboliche migliori, molti studi nel corso degli anni hanno cercato di rispondere alla domanda del rapporto ottimale tra TBA / E2 per ottenere il massimo rendimento di crescita. Ce ne sono stati alcuni che hanno affermato che la risposta a tale quesito era un rapporto di 5: 1 e 8: 1 (52,54), tuttavia i risultati hanno subito delle leggere variazioni nel corso degli anni. Infatti, in uno studio, i tassi medi di crescita giornaliera (ADG) erano abbastanza simili nei manzi impiantati con una combinazione di 120mg TBA+25mg E2 o una combinazione di 120mg TBA+24mg E2 (80).
Un altro studio ha dimostrato che 120mg di TBA+24mg E2 hanno aumentato i tassi medi di crescita del 20-25% e l’efficienza dell’alimentazione del 15-20% (55). Infatti, è stato dimostrato che i trattamenti combinati portano al 50% in più nella proliferazione delle cellule satelliti nel muscolo semimembranoso (tendine del ginocchio) dei manzi. (81) Come detto in precedenza, la proliferazione delle cellule satelliti è un fattore significativo nel processo ipertrofico. Altri studi hanno mostrato un aumento indotto dal Trenbolone sia nell’attivazione che nella proliferazione delle cellule satelliti. (82-83) Sembra che il Trenbolone e il Testosterone aumentino il numero di cellule satelliti per fibra muscolare in misura simile (22), quindi questo non è un effetto dato solo dal Trenbolone. I suoi effetti sulle cellule satelliti possono essere, almeno in parte, mediati attraverso il recettore del IGF-1, come l’inibizione di diversi target a valle del IGF-1 (es. MAPK, MEK / ERK, PI3K / Akt) e la successiva soppressione della proliferazione delle cellule satelliti Trenbolone-indotta osservata nelle colture cellulari.(7)
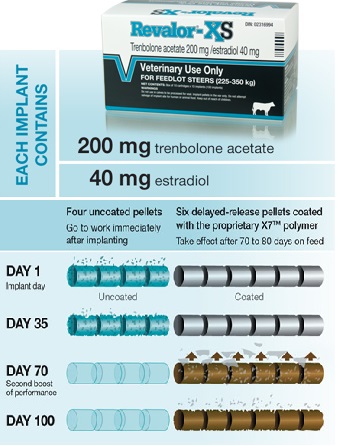
Nel 2007 la FDA ha approvato la commercializzazione del Revalor-XS il quale contiene una combinazione di 200mg di TBA + 40mg di E2, e che è stato progettato per avere un lento rilascio degli ormoni in esso contenuti dovuto all’azione di un rivestimento polimerico presente su sei dei dieci pellet componenti il prodotto. Ciò è utile in quanto i metodi di impianto tradizionali richiedono più impianti con il potenziale di aggiungere stress che potrebbe influire negativamente sul rendimento del bestiame. Gran parte delle variazione dei risultati riscontrate negli studi nel corso degli anni potrebbero benissimo essere correlate a questo fattore. Le studi condotti sul Revalor-XS hanno mostrato che la dose più elevata di TBA / E2 migliora il rendimento dei manzi sono alimentati per periodi più lunghi. (65,140) Nonostante la speculazione sul fatto che più impianti possano causare stress aggiunto ai manzi, il pattern di lento rilascio del Revalor-XS in realtà non fornisce effetti unici sul rendimento delle carni, o della qualità, se confrontato con una strategia di impianto multiplo di pari dosaggio di TBA + E2.
Effetti del IGF-1
Molecola di IGF-1 umano
Gli impianti contenenti TBA / E2 hanno dimostrato di aumentare significativamente i livelli di IGF-1. Questi trattamenti combinati hanno portato a un aumento dei livelli serici di IGF-1 (59,84,85,86), aumenti dell’espressione del mRNA del IGF-1 epatico (56) e aumenti dell’espressione del mRNA del IGF-1 nei tessuti muscolo-scheletrici. (59,61,81)
Anche gli impianti contenenti solo TBA hanno dimostrato di aumentare i livelli di IGF-1 in varie specie, tuttavia, però, il grado di aumento dei livelli di IGF-1 non è stato paragonabile a quello riscontrato con impianti contenenti la combinazione TBA/E2.(87,88,89) In effetti, il Trenbolone non aumenta significativamente l’IGF-1 autocrino o endocrino in maniera maggiore rispetto al Testosterone. Uno studio ha anche dimostrato che il Testosterone aumenta i livelli di IGF-1 autocrino in maniera leggermente più elevata rispetto al TBA.(4) L’evidenza sembra suggerire che qualsiasi effetto sull’IGF-1 possa essere mediato attraverso l’estradiolo e possa anche essere stimolato tramite distinti meccanismi dei recettori degli androgeni e degli estrogeni, che includono il coinvolgimento del recettore accoppiato alla proteina G (GPR30).(90) In particolare, uno studio ha rilevato che l’aumento dell’espressione autocrina del IGF-1 nel muscolo scheletrico richiede la presenza di estrogeni e che gli impianti con solo TBA non hanno determinato aumenti dei livelli di mRNA del IGF-1 muscolare.(91) È certamente ragionevole ipotizzare che possa esserci una soglia da rispettare, nonostante il fatto che essa potrebbe non essere realisticamente rapportabile dagli studi sugli animali e l’uomo.
Studi in vitro svolti utilizzando cellule satelliti bovine (BSC) hanno mostrato una relazione dose-dipendente tra Trenbolone e livelli del mRNA del IGF-1. Nelle cellule trattate con 1nM o 10nM di Trenbolone per 48 ore, i livelli del mRNA del IGF-1 erano 1,7 volte più alti, tuttavia i livelli di mRNA non erano influenzati dai trattamenti con 0,001, 0,01 o 0,1 nM. (92) È anche emerso che gli effetti sono stati almeno parzialmente AR-mediati in quanto il co-trattamento con Flutamide (inibitore AR) ha completamente annullato l’aumento dell’espressione di IGF-1 osservata in queste colture cellulari.(90)
Non ci vuole molto tempo per osservare un aumento dei livelli di IGF-1 dopo l’applicazione di un impianto. In uno studio, agnelli impiantati con Revalor-S (120mg TBA / 24mg E2) hanno mostrato un aumento dei livelli serici di IGF-1 entro il 3° e il 10 ° giorno rispettivamente del 43% e del 62% (56). Questo aumento del IGF-1 è stato mantenuto per tutti i 24 giorni dello studio e i livelli del mRNA del IGF-1 epatico erano risultati del 150% più alti negli agnelli impiantati rispetto a quelli di controllo, suggerendo che il fegato è probabilmente una fonte primaria dalla quale provengono gli aumenti di IGF-1 circolante. I livelli del mRNA del IGF-1 autocrino erano del 68% più alti nei muscoli longissimus degli agnelli impiantati rispetto a quelli osservati negli animali di controllo. Il dosaggio di TBA ed E2, per chilogrammo di peso corporeo, era circa tre volte superiore a quello approvato per l’uso nei manzi. A causa delle differenze tra le specie e il dosaggio, si dovrebbe usare cautela quando si tenta di prendere questi risultati e applicarli ai manzi.
L’uso di questa stessa dose nei manzi ha dimostrato di produrre livelli di IGF-1 serici più elevati rispetto ai bovini non impiantati, entro 6-42 giorni dall’impianto.(93) In sole 48 ore, i manzi impiantati hanno avuto un aumento del 13,4% delle concentrazioni seriche di IGF-1.(84) Nei giorni 21 e 40, i manzi impiantati presentavano livelli di IGF-1 superiori del 16% e del 22% rispetto agli animali di controllo. La cosa interessante è che i livelli di IGF-1 raggiunsero il picco durante questo periodo di tempo e successivamente iniziarono a calare fino al giorno 115 dello studio, dove finirono per assomigliare ai valori del giorno 1. Detto questo, i manzi di controllo avevano ancora livelli IGF-1 più bassi rispetto al primo giorno. Quindi, anche se gli aumenti dei livelli di IGF-1 appaiono transitori, gli impianti sembrano ancora fornire un effetto additivo complessivo, anche con un uso a lungo termine.
Altri studi sul bestiame hanno mostrato campioni muscolari con un più alto livello del mRNA del IGF-1 entro 30-40 giorni dall’applicazione dell’impianto. (56,61) Questi animali impiantati mostravano anche cellule satelliti più proliferanti rispetto ai manzi non impiantati suggerendo che gli aumenti della massa muscolare osservati con la combinazione TBA / E2 possono essere almeno in parte ricondotti all’aumento del IGF-1. Come abbiamo discusso in precedenza, è ben noto che la crescita muscolare postnatale dipende dalla fusione delle cellule satelliti con le fibre esistenti per fornire i mionuclei necessari a sostenere la crescita.(24) Questo aumento è importante anche perché solo un piccolo numero di cellule satelliti sono presenti nei bovini di un anno e molte delle cellule esistenti sono diventate inattive o hanno lasciato il ciclo cellulare. Vale anche la pena ricordare che la sovraespressione del IGF-1 estende la durata della vita replicativa delle cellule satelliti, almeno nelle colture cellulari. (94) Pertanto, sembra ragionevole ipotizzare che l’aumentata espressione del IGF-I muscolare giochi un ruolo nell’aumento indotto dall’AAS nel numero delle cellule satelliti muscolari.
In un altro studio, i livelli del mRNA del IGF-1 allo stato stazionario sono stati mostrati essere del 69% più alti nei manzi impiantati, suggerendo ancora una volta che il fegato può essere un fattore importante nell’aumento dell’IGF-1 circolante negli animali sottoposti ad impiantati.(61) Si prega di notare che c’è stato almeno uno studio, di cui sono a conoscenza, che non ha mostrato differenze nelle concentrazioni di IGF-1 tra manzi impiantati e di controllo.(95) Comunque, questo risultato può benissimo essere considerato come l’eccezione che conferma la regola.
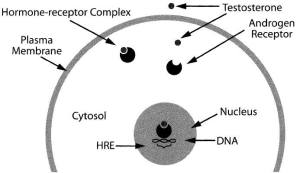
Un elemento di risposta agli androgeni (ARE) è stato identificato nella regione del promotore del gene IGF-1, suggerendo che il complesso recettore-ligando androgeno possa interagire con questo ARE per stimolare la trascrizione del gene IGF-1. Gli androgeni tendono ad agire attraverso meccanismi multipli sui muscoli, e l’estrogeno tende ad agire sull’ipotalamo / pituitaria anteriore per stimolare l’asse GH / IGF-1.(96) La relazione tra l’estrogeno e l’asse GH / IGF-1 ha dimostrato di essere additiva.(97-98)
Elementi sugli Estrogeni
Dato che l’impatto degli estrogeni è stato parzialmente trattato, è corretto concentrarsi sul loro ruolo nel contesto di cui stiamo trattando prima di andare avanti.
Studi in vitro hanno dimostrato che il trattamento di colture di cellule satelliti bovine con E2 per 48 ore porta ad un aumento significativo dell’espressione del mRNA del IGF-1.(92) Ciò è in linea con quello che già sappiamo sull’E2, poiché è stato dimostrato che stimola l’espressione dell’mRNA dell’IGF-1 in un certo numero di tessuti. (99-100) È interessante notare che il co-trattamento con ICI (antagonista del recettore dell’estrogeno) non ha soppresso l’espressione del IGF-1 stimolata con l’E2. Ciò sembra suggerire che il meccanismo attraverso il quale l’E2 stimola l’espressione dell’mRNA del IGF-I nelle BSC può essere diverso dal meccanismo che si attua in altri tessuti che sono stati esaminati fino ad oggi.
Anche se il gene del IGF-1 non contiene un tradizionale elemento di risposta agli estrogeni (ERE) nella sua regione di regolazione, la stimolazione da parte dell’E2 dell’espressione dell’mRNA del IGF-1 può avvenire tramite un percorso che coinvolge il fattore di trascrizione AP-1.(101) Come accennato in precedenza, oltre ai recettori degli estrogeni classici, il recettore GRP30) può svolgere un ruolo nella mediazione delle azioni degli estrogeni.(102,103,104) Questo è rilevante per i nostri interessi in quanto il tessuto muscolare contiene l’mRNA del GPR30 e gli studi immunoistochimici hanno localizzato la proteina del recettore GPR30 all’interno delle cellule muscolo-scheletriche.(105) Inoltre, gli effetti dell’agonista / antagonista GPR indicano fortemente che il recettore GPR30 è coinvolto nell’aumento dello stimolo del E2 sul mRNA del IGF-1 osservato nelle colture di cellule satelliti bovine.(90)
Effetti sulle fibre muscolari
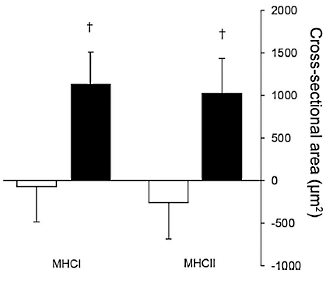
(Fonte immagine “Project Invictus”)
Gli impianti contenenti TBA / E2 aumentano l’area della sezione trasversale delle fibre muscolari (CSA) a causa di un iniziale aumento della trascrizione del DNA seguito da un aumento dei nuclei all’interno delle fibre muscolari che supportano l’ipertrofia.(106) Il TBA (da solo o in combinazione con E2) ha dimostrato di aumentare il CSA delle fibre muscolari di tipo I ma non di quelle di tipo II (9,107). È stato anche dimostrato che l’impianto nei manzi combinato con iperalimentazione aumenta il CSA nei muscoli LM nelle fibre di tipo I e IIA.(110) Ci sono state comunque delle eccezioni, dal momento che è stato dimostrato che uno studio ha aumentato le fibre di tipo IIB senza alcun impatto sulla dimensione o sul numero delle fibre di tipo I.(57) Questi studi, se considerati nel loro complesso, sembrano suggerire che il Trenbolone induca la conversione delle fibre muscolari da glicolitiche a ossidative, il che indica un aumento della capacità ossidativa delle fibre del muscolo-scheletrico.
Tornando alle potenziali differenze osservate tra le diverse specie animali, nonostante l’aumento del peso muscolare e della dimensione delle fibre muscolari, il numero dei mionuclei per fibra non ha subito miglioramenti nei ratti ai quali è stato somministrato Trenbolone o Testosterone.(22) Ciò contraddice i risultati di uno studio precedente il quale, però, ha visto la somministrazione di Testosterone all’inizio della pubertà, una fase caratterizzata da una crescita rapida del muscolo LABC rispetto a quanto osservabile negli esemplari maturi dello studio precedente.(108) È altamente probabile che l’ipertrofia indotta dagli androgeni nei ratti adulti senza stimoli fisici non richieda l’aggiunta mionucleare (109), il che va contro senso rispetto a ciò di cui abbiamo parlato nell’intero articolo. Ma queste sono le differenze da tenere in considerazione quando si osservano studi su animali e si cerca di stabilire modelli che potrebbero potenzialmente essere tradotti negli esseri umani.
Effetti dei Recettori degli Androgeni
Recettore degli Androgeni, modello molecolare.
Ci sono stati più esperimenti in vitro che hanno indicano una sovra regolazione dell’espressione del mRNA del AR (111-112). Tuttavia, sembra che vi sia un limite massimo oltre il quale anche con dosi più elevate non si riesce a modificare i livelli di mRNA in misura relativa a quelli presenti nelle colture di controllo non trattate.(92)
Tuttavia, ciò non è stato universalmente dimostrato negli studi, in quanto alcuni non hanno mostrato effetti mediati dal Trenbolone sull’espressione del mRNA del AR.(4,91) Questa discrepanza può essere dovuta al fatto che l’espressione elevata del AR si verifica in un punto temporale precedente rispetto alla raccolta dei dati effettuata in questi studi, ma ciò è speculativo. L’evidenza in vitro indica anche che il Trenbolone induce la traslocazione del AR umano nel nucleo cellulare in modo dose-dipendente e stimola anche la trascrizione del gene almeno allo stesso grado del DHT.(14)
XI. Atrofia/Anti-Catabolismo
La reputazione del Trenbolone come AAS fortemente anti-catabolico è in realtà ben meritata. Anche in questo caso, è utile trattare brevemente le basi dell’atrofia muscolo-scheletrica prima di proseguire con l’esposizione della letteratura associata al Trenbolone.
Il Proteosoma spezza i substrati ubiquitinilati in corti peptidi, riciclando le molecole di Ubiquitina.
Durante diversi stati catabolici, la via dell’ubiquitina-proteasoma aumenta la disgregazione proteica che porta all’atrofia muscolare. Nello specifico, due ubiquitin ligasi, MuRF1 e MAFbx (anche denominate Atrogin-1) fungono da marker dell’atrofia muscolare in diverse condizioni cataboliche come il digiuno, il cancro, l’insufficienza renale e il diabete.(113,114,115) È stato dimostrato che il Trenbolone riduce significativamente l’espressione del mRNA del MuRF1 e del Atrogin-1 nei tessuti muscolo-scheletrici di un fattore 3 nei ratti castrati. I tassi di Atrogina-1 sono stati soppressi in questi animali ad un livello ancora maggiore rispetto a quanto osservato con la somministrazione di Testosterone.(4)
Glucocorticoidi
Cortisolo, il principale Glucocorticoide sintetizzato dalla Corteccia Surrenale.
I Glucocorticoidi sono ormoni steroidei che aiutano a regolare l’omeostasi metabolica dell’intero organismo. Essi esercitano anche la loro influenza sul muscolo-scheletrico, infatti una elevata esposizione del tessuto muscolare a questi ormoni può potenzialmente portare all’atrofia dei tessuti. I membri principali della famiglia dei Glucocorticoidi sono il Cortisolo, il Corticosterone e il Cortisone. Si legano con il recettore Glucocorticoidi intracellulare (GRα) attivandolo ed esercitando i loro effetti. Vale la pena ricordare che il Cortisolo può legarsi sia al recettore glucocorticoide (GRα) che al recettore mineralcorticoide (MR), tuttavia un approfondimento in tal senso va oltre lo scopo di questa serie di articoli.
È stato dimostrato che il Trenbolone riduce la capacità di legame dei Glucocorticoidi causando una riduzione nel numero di GRα nei tessuti muscolo-scheletrici. (36,116) Studi in vitro hanno dimostrato che il Trenbolone agisce come un antagonista del recettore glucocorticoide [14] nonostante il 17β-TbOH possegga solo un’affinità di legame relativa del 9,4% per il recettore glucocorticoide bovino rispetto al cortisolo [13]. Altri studi hanno dimostrato che il Trenbolone riduce la capacità del Cortisolo di legarsi ai GR nel muscolo-scheletrico e sottoregola l’espressione complessiva dei GR. (117-118) Infatti, il Trenbolone sopprime l’espressione dei GRα del 50% in più rispetto al Testosterone.(4) E le sue azioni anti-glucocorticoidi probabilmente aiutano a creare un’inibizione significativamente più marcata della degradazione delle proteine muscolari (MPB) rispetto al Testosterone, che riduce solo leggermente l’MPB aumentando contemporaneamente l’MPS. (119)
È stato dimostrato che il Trenbolone riduce le concentrazioni circolanti di Corticosterone nei roditori (37,39,116,120) e del Cortisolo nei bovini impiantati.(50) L’evidenza suggerisce che il Trenbolone agisca a livello delle ghiandole surrenali sopprimendo la sintesi del Cortisolo ACTH-stimolata e sopprimendo il rilascio di Cortisolo.(121)
Ora possiamo provare ad estrapolare un po’ di informazioni correlate alle possibilità che si presentano in un contesto dove i livelli di glucocorticoidi sono abbassati. Ad esempio, i glucocorticoidi inibiscono l’assorbimento del glucosio e aiutano a stimolare la disgregazione del glicogeno immagazzinato nel muscolo-scheletrico attenuando la traslocazione dei GLUT4 nelle membrane cellulari indotta dall’insulina.(122) La segnalazione dell’insulina nei tessuti muscolari è essenzialmente soppressa dai glucocorticoidi.(123) Detto questo, è possibile che la somministrazione di Trenbolone possa portare ad un maggiore utilizzo del glucosio? Abbiamo già visto la capacità del Trenbolone di aumentare la sensibilità all’insulina nei ratti, ma cosa succederebbe se lo si co-somministrasse con l’insulina esogena?
I Glucocorticoidi tendono anche ad aumentare i livelli di trigliceridi intramuscolari.(124) È quindi ragionevole ipotizzare che gli effetti cosmetici tradizionalmente attribuiti al Trenbolone possano avere qualcosa a che fare con questo? Se il Trenbolone riduce ì i livelli di trigliceridi intramuscolari, allora questo potrebbe essere un fattore primario dietro all’aspetto muscolare poco “voluminoso” (nonostante la presenza di un carico glicemico pienamente sufficiente) che molti tendono ad avere? Cercherò di tornare su queste domande nelle mie osservazioni conclusive di questa serie di articoli.
Effetti sulla sintesi e degradazione proteica
Una delle caratteristiche più paradossali riscontrate sul Trenbolone è rappresentata da un tasso di sintesi proteica muscolare (MPS) diminuito in seguito alla somministrazione di questo AAS. Ciò è stato dimostrato in studi nei quali si sono osservati gli effetti del trattamento con impianti contenenti TBA o TBA+E2.(17,32,48) Molti si chiedono come sia possibile che una molecola notoriamente molto anabolizzante come il Trenbolone riduca i tassi di MPS. La chiave per comprendere ciò risiede nell’impatto del Trenbolone sul MPB, in quanto il Trenbolone possiede un azione riduttiva sui tassi di MPB maggiore rispetto a quelli sul MPS, il che si traduce in uno stato nettamente anabolico.
Infatti, nonostante la riduzione dei tassi di MPS, è stato dimostrato che il Trenbolone aumenta la ritenzione di azoto di tutto il corpo in varie specie animali. (32,125,126,127) Ancora una volta, questo ha molto a che fare con l’impatto del Trenbolone sui tassi di MPB. È stato dimostrato che causa una riduzione significativa dei tassi di MPB totale e miofibrillare in varie specie animali.(32,34,36,120,128)
Vale la pena notare che studi in vitro hanno effettivamente mostrato aumenti dipendenti dalla concentrazione indotti dal Trenbolone nei tassi di MPS. Possono essere significativi, con un aumento fino a 1,7 volte utilizzando la dose più alta di 10 nM [129]. Quindi, similmente a quello che abbiamo visto prima con l’espressione del IGF-1, potrebbe esserci una soglia oltre la quale il Trenbolone smette di sopprimere la MPS e inizia ad aumentarla. È probabile che questa soglia si estenda oltre i realistici casi d’uso del mondo reale, dato che studi in vivo su vari animali non mostrano questo effetto. A livello cellulare, anche le percentuali di degradazione delle proteine sono state abbassate, con la dose massima di TBA che causa un tasso di degradazione pari al 70% in meno di quello mostrato nelle colture senza TBA. Questo è stato un effetto parzialmente AR-mediato in quanto il Flutamide sopprime la capacità del Trenbolone di stimolare la sintesi proteica e di sopprimere i tassi di degradazione delle proteine. Anche il trattamento delle colture cellulari con JB1 (inibitore del IGF-1) influisce sugli effetti del Trenbolone sulla sintesi / degradazione delle proteine, quindi è altamente probabile che questi effetti richiedano in una certa misura l’attivazione dei recettori degli androgeni e del IGF-1.
È stato anche dimostrato che il Trenbolone sopprime la degradazione degli amminoacidi nel fegato.(37,130) Questo può anche essere un fattore chiave per gli effetti complessivi sul MPB, in quanto il primo passo nella degradazione degli aminoacidi avviene a livello epatico – la rimozione dell’azoto. In effetti, il principale sito di degradazione degli amminoacidi nei mammiferi è il fegato.
Effetti sull’osso
L’ipogonadismo legato all’età è un fattore importante che contribuisce alla perdita di tessuto osseo negli uomini anziani.(131) Come abbiamo discusso in precedenza, il trattamento di fatto per l’ipogonadismo è la terapia sostitutiva del Testosterone (TRT). Il problema è che con le TRT si vengono a creare solo modesti miglioramenti della densità minerale ossea nei soggetti trattati. (132-133) Al contrario, le dosi sovrafisiologiche di Testosterone offrono una protezione completa dalla perdita ossea ma, tuttavia, a tali dosaggi si manifestano molti effetti collaterali indesiderati. (134,135,136) Quindi, sono tornato a trattare un punto già esposto nella prima parte di questa serie di articoli, e cioè cercare di ottenere con l’uso del Trenbolone gli effetti protettivi dati dalla somministrazione di dosi sovra fisiologiche di Testosterone, ma senza gli effetti indesiderati.
Le prime indicazioni in questo senso sono promettenti in quanto gli studi sui roditori dimostrano che il Trenbolone impedisce la perdita di tessuto osseo indotta dal ipogonadismo in ratti castrati a un livello uguale a quello rilevato con dosi sovra fisiologiche di Testosterone, ma senza causare ipertrofia prostatica o aumenti dell’emoglobina che sono frequentemente osservati con trattamenti a base di Testosterone.(3,20)
Corticosterone
Il Trenbolone potenzialmente esercita parte della sua influenza sull’osso attraverso la riduzione del Corticosterone circolante, attraverso la sua attività anti-glucocorticoide.(14,39) E, nonostante l’azione soppressiva del Trenbolone sui livelli estrogenici, questa molecola possiede ancora caratteristiche di conservazione del tessuto osseo simili al Testosterone. Questo è similare a quanto osservato con il DHT, quindi sembra che gli androgeni non aromatizzabili siano in grado di proteggere le ossa direttamente attraverso percorsi AR-mediati.(137-138) Ci sono ancora persone convinte che un lieve grado di aromatizzazione del testosterone in E2 a livello scheletrico sia essenziale per garantire la protezione delle ossa nei maschi.(139) Quindi, prima di poter trarre conclusioni, dovranno essere condotti studi sul Trenbolone a lungo termine.
Anche per questa terza parte siamo arrivati al termine. Nella quarta ed ultima parte di questa serie di articoli tratterò della lipolisi, dei potenziali rischi legati all’uso del Trenbolone ed esporrò le mie osservazioni e raccomandazioni riguardo a quanto trattato.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
Monks DA, O’Bryant EL, Jordan CL. Androgen receptor immunoreactivity in skeletal muscle: enrichment at the neuromuscular junction. J Comp Neurol. 2004 May 17;473(1):59-72.
Monks DA, Kopachik W, Breedlove SM, Jordan CL. Anabolic responsiveness of skeletal muscles correlates with androgen receptor protein but not mRNA. Can J Physiol Pharmacol. 2006 Feb;84(2):273-7
McCoy SC, Yarrow JF, Conover CF, Borsa PA, Tillman MD, Conrad BP, Pingel JE, Wronski TJ, Johnson SE, Kristinsson HG, Ye F, Borst SE. 17β-Hydroxyestra-4,9,11-trien-3-one (Trenbolone) preserves bone mineral density in skeletally mature orchiectomized rats without prostate enlargement. Bone. 2012 Oct;51(4):667-73.
Ye F, McCoy SC, Ross HH, Bernardo JA, Beharry AW, Senf SM, Judge AR, Beck DT, Conover CF, Cannady DF, Smith BK, Yarrow JF, Borst SE. Transcriptional regulation of myotrophic actions by testosterone and trenbolone on androgen-responsive muscle. Steroids. 2014 Sep;87:59-66.
Sauerwein H, Meyer HH. Androgen and estrogen receptors in bovine skeletal muscle: relation to steroid-induced allometric muscle growth. J Anim Sci. 1989 Jan;67(1):206-12.
Brandstetter AM, Pfaffl MW, Hocquette JF, Gerrard DE, Picard B, Geay Y, Sauerwein H. Effects of muscle type, castration, age, and compensatory growth rate on androgen receptor mRNA expression in bovine skeletal muscle. J Anim Sci. 2000 Mar;78(3):629-37.
Kamanga-Sollo E, White ME, Hathaway MR, Chung KY, Johnson BJ, Dayton WR. Roles of IGF-I and the estrogen, androgen and IGF-I receptors in estradiol-17beta- and trenbolone acetate-stimulated proliferation of cultured bovine satellite cells. Domest Anim Endocrinol. 2008 Jul;35(1):88-97.
Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci. 1991 Jun;69(6):2452-62.
Clancy MJ, Lester JM, Roche JF. The effects of anabolic agents and breed on the fibers of the longissimus muscle of male cattle. J Anim Sci. 1986 Jul;63(1):83-91.
Young OA, Bass JJ. Effect of castration on bovine muscle composition. Meat Sci. 1984;11(2):139-56.
Sinha-Hikim I, Taylor WE, Gonzalez-Cadavid NF, Zheng W, Bhasin S. Androgen receptor in human skeletal muscle and cultured muscle satellite cells: up-regulation by androgen treatment. J Clin Endocrinol Metab. 2004 Oct;89(10):5245-55.
Sinnett-Smith PA, Palmer CA, Buttery PJ. Androgen receptors in skeletal muscle cytosol from sheep treated with trenbolone acetate. Horm Metab Res. 1987 Mar;19(3):110-4.
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Wilson VS, Lambright C, Ostby J, Gray LE Jr. In vitro and in vivo effects of 17beta-trenbolone: a feedlot effluent contaminant. Toxicol Sci. 2002 Dec;70(2):202-11.
Ankley GT, Defoe DL, Kahl MD, Jensen KM, Makynen EA, Miracle A, Hartig P, Gray LE, Cardon M, Wilson V. Evaluation of the model anti-androgen flutamide for assessing the mechanistic basis of responses to an androgen in the fathead minnow (Pimephales promelas). Environ Sci Technol. 2004 Dec 1;38(23):6322-7.
Lee HS, Jung DW, Han S, Kang HS, Suh JH, Oh HS, Hwang MS, Moon G, Park Y, Hong JH, Koo YE. Veterinary drug, 17β-trenbolone promotes the proliferation of human prostate cancer cell line through the Akt/AR signaling pathway. Chemosphere. 2018 May;198:364-369.
Vernon BG, Buttery PJ. Protein turnover in rats treated with trienbolone acetate. Br J Nutr. 1976 Nov;36(3):575-9.
Freyberger A, Ellinger-Ziegelbauer H, Krötlinger F. Evaluation of the rodent Hershberger bioassay: testing of coded chemicals and supplementary molecular-biological and biochemical investigations. Toxicology. 2007 Sep 24;239(1-2):77-88.
Hotchkiss AK, Nelson RJ. An environmental androgen, 17beta-trenbolone, affects delayed-type hypersensitivity and reproductive tissues in male mice. J Toxicol Environ Health A. 2007 Jan 15;70(2):138-40.
Yarrow JF, Conover CF, McCoy SC, Lipinska JA, Santillana CA, Hance JM, Cannady DF, VanPelt TD, Sanchez J, Conrad BP, Pingel JE, Wronski TJ, Borst SE. 17β-Hydroxyestra-4,9,11-trien-3-one (trenbolone) exhibits tissue selective anabolic activity: effects on muscle, bone, adiposity, hemoglobin, and prostate. Am J Physiol Endocrinol Metab. 2011 Apr;300(4):E650-60.
Beck DT, Yarrow JF, Beggs LA, Otzel DM, Ye F, Conover CF, Miller JR, Balaez A, Combs SM, Leeper AM, Williams AA, Lachacz SA, Zheng N, Wronski TJ, Borst SE. Influence of aromatase inhibition on the bone-protective effects of testosterone. J Bone Miner Res. 2014 Nov;29(11):2405-13.
Dalbo VJ, Roberts MD, Mobley CB, Ballmann C, Kephart WC, Fox CD, Santucci VA, Conover CF, Beggs LA, Balaez A, Hoerr FJ, Yarrow JF, Borst SE, Beck DT. Testosterone and trenbolone enanthate increase mature myostatin protein expression despite increasing skeletal muscle hypertrophy and satellite cell number in rodent muscle. Andrologia. 2017 Apr;49(3).
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
Campion DR. The muscle satellite cell: a review. Int Rev Cytol. 1984;87:225-51. Review.
Moss FP, Leblond CP. Satellite cells as the source of nuclei in muscles of growing rats. Anat Rec. 1971 Aug;170(4):421-35.
Allen Trenkle, D. L. DeWitt, David G. Topel; Influence of Age, Nutrition and Genotype on Carcass Traits and Cellular Development of the M. Longissimus of Cattle, Journal of Animal Science, Volume 46, Issue 6, 1 June 1978, Pages 1597–1603
Cardasis CA, Cooper GW. A method for the chemical isolation of individual muscle fibers and its application to a study of the effect of denervation on the number of nuclei per muscle fiber. J Exp Zool. 1975 Mar;191(3):333-46.
Johnson SE, Allen RE. The effects of bFGF, IGF-I, and TGF-beta on RMo skeletal muscle cell proliferation and differentiation. Exp Cell Res. 1990 Apr;187(2):250-4.
Allen RE, Rankin LL. Regulation of satellite cells during skeletal muscle growth and development. Proc Soc Exp Biol Med. 1990 Jun;194(2):81-6. Review.
Allen RE, Boxhorn LK. Regulation of skeletal muscle satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J Cell Physiol. 1989 Feb;138(2):311-5.
Best JM. The use of trienbolone acetate implants in heifer beef production at pasture. Vet Rec. 1972 Dec 16;91(25):624-6
Vernon BG, Buttery PJ. The effect of trenbolone acetate with time on the various responses of protein synthesis of the rat. Br J Nutr. 1978 Nov;40(3):563-72.
Galbraith H. Effect of trenbolone acetate on growth, blood metabolites and hormones of cull beef cows. Vet Rec. 1980 Dec 13;107(24):559-60.
Ranaweera KN, Wise DR. The effects of trienbolone acetate on carcass composition, conformation and skeletal growth of turkeys. Br Poult Sci. 1981 Mar;22(2):105-14.
Henricks DM, Edwards RL, Champe KA, Gettys TW, Skelley GC Jr, Gimenez T. Trenbolone, estradiol-17 beta and estrone levels in plasma and tissues and live weight gains of heifers implanted with trenbolone acetate. J Anim Sci. 1982 Nov;55(5):1048-56.
Santidrian S, Thompson JR. Effect of sex, testosterone propionate and trienbolone acetate on the rate of growth and myofibrillar protein degradation in growing young rats. Reproduccion. 1982 Jan-Mar;6(1):33-41.
Thomas KM, Rodway RG. Effects of trenbolone acetate on adrenal function and hepatic enzyme activities in female rats. J Endocrinol. 1983 Jul;98(1):121-7.
Hunter RA, Vercoe JE. Reduction of energy requirements of steers fed on low-quality-roughage diets using trenbolone acetate. Br J Nutr. 1987 Nov;58(3):477-83.
Sillence MN, Rodway RG. Effects of trenbolone acetate and testosterone on growth and on plasma concentrations of corticosterone and ACTH in rats. J Endocrinol. 1990 Sep;126(3):461-6.
Castaldo DJ, Jones JE, Maurice DV. Growth and carcass composition of female turkeys implanted with anabolic agents and fed high-protein and low-protein diets. Arch Tierernahr. 1990 Aug;40(8):703-12.
Apple JK, Dikeman ME, Simms DD, Kuhl G. Effects of synthetic hormone implants, singularly or in combinations, on performance, carcass traits, and longissimus muscle palatability of Holstein steers. J Anim Sci. 1991 Nov;69(11):4437-48.
Freyberger A, Hartmann E, Krötlinger F. Evaluation of the rodent Hershberger bioassay using three reference (anti)androgens. Arh Hig Rada Toksikol. 2005 Jun;56(2):131-9.
Donner DG, Beck BR, Bulmer AC, Lam AK, Du Toit EF. Improvements in body composition, cardiometabolic risk factors and insulin sensitivity with trenbolone in normogonadic rats. Steroids. 2015 Feb;94:60-9.
Galbraith H, Watson HB. Performance, blood and carcase characteristics of finishing steers treated with trenbolone acetate and hexoestrol. Vet Rec. 1978 Jul 8;103(2):28-31.
Galbraith H, Dempster DG. Effect of hexoestrol on the response of finishing steers to treatment with trenbolone acetate. Vet Rec. 1979 Sep 22;105(12):283-4.
Wise DR, Ranaweera KN. The effects of trienbolone acetate and other anabolic agents in growing turkeys. Br Poult Sci. 1981 Mar;22(2):93-104.
Sinnett-Smith PA, Dumelow NW, Buttery PJ. Effects of trenbolone acetate and zeranol on protein metabolism in male castrate and female lambs. Br J Nutr. 1983 Sep;50(2):225-34.
Renaville R, Burny A, Sneyers M, Rochart S, Portetelle D, Théwis A. Effects of an anabolic treatment before puberty with trenbolone acetate-oestradiol or oestradiol alone on growth rate, testicular development and luteinizing hormone and testosterone plasma concentrations. Theriogenology. 1988 Feb;29(2):461-76.
Lee CY, Henricks DM, Skelley GC, Grimes LW. Growth and hormonal response of intact and castrate male cattle to trenbolone acetate and estradiol. J Anim Sci. 1990 Sep;68(9):2682-9.
Perry TC, Fox DG, Beermann DH. Effect of an implant of trenbolone acetate and estradiol on growth, feed efficiency, and carcass composition of Holstein and beef steers. J Anim Sci. 1991 Dec;69(12):4696-702.
Bartle SJ, Preston RL, Brown RE, Grant RJ. Trenbolone acetate/estradiol combinations in feedlot steers: dose-response and implant carrier effects. J Anim Sci. 1992 May;70(5):1326-32.
Cecava MJ, Hancock DL. Effects of anabolic steroids on nitrogen metabolism and growth of steers fed corn silage and corn-based diets supplemented with urea or combinations of soybean meal and feathermeal. J Anim Sci. 1994 Feb;72(2):515-22.
Herschler RC, Olmsted AW, Edwards AJ, Hale RL, Montgomery T, Preston RL, Bartle SJ, Sheldon JJ. Production responses to various doses and ratios of estradiol benzoate and trenbolone acetate implants in steers and heifers. J Anim Sci. 1995 Oct;73(10):2873-81.
Johnson BJ, Anderson PT, Meiske JC, Dayton WR. Effect of a combined trenbolone acetate and estradiol implant on feedlot performance, carcass characteristics, and carcass composition of feedlot steers. J Anim Sci. 1996 Feb;74(2):363-71.
Johnson BJ, White ME, Hathaway MR, Christians CJ, Dayton WR. Effect of a combined trenbolone acetate and estradiol implant on steady-state IGF-I mRNA concentrations in the liver of wethers and the longissimus muscle of steers. J Anim Sci. 1998 Feb;76(2):491-7.
Fritsche S, Solomon MB, Paroczay EW, Rumsey TS. Effects of growth-promoting implants on morphology of Longissimus and Semitendinosus muscles in finishing steers. Meat Sci. 2000 Nov;56(3):229-37.
McClure KE, Solomont MB, Loerch SC. Body weight and tissue gain in lambs fed an all-concentrate diet and implanted with trenbolone acetate or grazed on alfalfa. J Anim Sci. 2000 May;78(5):1117-24.
Dunn JD, Johnson BJ, Kayser JP, Waylan AT, Sissom EK, Drouillard JS. Effects of flax supplementation and a combined trenbolone acetate and estradiol implant on circulating insulin-like growth factor-I and muscle insulin-like growth factor-I messenger RNA levels in beef cattle. J Anim Sci. 2003 Dec;81(12):3028-34.
Scheffler JM, Buskirk DD, Rust SR, Cowley JD, Doumit ME. Effect of repeated administration of combination trenbolone acetate and estradiol implants on growth, carcass traits, and beef quality of long-fed Holstein steers. J Anim Sci. 2003 Oct;81(10):2395-400.
White ME, Johnson BJ, Hathaway MR, Dayton WR. Growth factor messenger RNA levels in muscle and liver of steroid-implanted and nonimplanted steers. J Anim Sci. 2003 Apr;81(4):965-72.
Kreikemeier WM, Mader TL. Effects of growth-promoting agents and season on yearling feedlot heifer performance. J Anim Sci. 2004 Aug;82(8):2481-8.
Lefebvre B, Malouin F, Roy G, Giguère K, Diarra MS. Growth performance and shedding of some pathogenic bacteria in feedlot cattle treated with different growth-promoting agents. J Food Prot. 2006 Jun;69(6):1256-64.
Mader TL, Kreikemeier WM. Effects of growth-promoting agents and season on blood metabolites and body temperature in heifers. J Anim Sci. 2006 Apr;84(4):1030-7.
Parr SL, Chung KY, Hutcheson JP, Nichols WT, Yates DA, Streeter MN, Swingle RS, Galyean ML, Johnson BJ. Dose and release pattern of anabolic implants affects growth of finishing beef steers across days on feed. J Anim Sci. 2011 Mar;89(3):863-73.
Parr SL, Chung KY, Galyean ML, Hutcheson JP, DiLorenzo N, Hales KE, May ML, Quinn MJ, Smith DR, Johnson BJ. Performance of finishing beef steers in response to anabolic implant and zilpaterol hydrochloride supplementation. J Anim Sci. 2011 Feb;89(2):560-70.
Cranwell CD, Unruh JA, Brethour JR, Simms DD, Campbell RE. Influence of steroid implants and concentrate feeding on performance and carcass composition of cull beef cows. J Anim Sci. 1996 Aug;74(8):1770-6.
Hongerholt DD, Crooker BA, Wheaton JE, Carlson KM, Jorgenson DM. Effects of a growth hormone-releasing factor analogue and an estradiol-trenbolone acetate implant on somatotropin, insulin-like growth factor I, and metabolite profiles in growing Hereford steers. J Anim Sci. 1992 May;70(5):1439-48.
Heitzman RJ, Harwood DJ, Kay RM, Little W, Mallinson CB, Reynolds IP. Effects of implanting prepuberal dairy heifers with anabolic steroids on hormonal status, puberty and parturition. J Anim Sci. 1979 Apr;48(4):859-66.
Hancock, D. L., J. F. Wagner, and D. B. Anderson 1991. Effects of estrogens and androgens on animal growth. Pages 255–297 in Growth Regulation in Farm Animals. Advances in Meat Research. Vol. 7. A. M. Pearson and T. R. Dutson ed. Elsevier Applied Science, New York, NY.
Hayden JM, Bergen WG, Merkel RA. Skeletal muscle protein metabolism and serum growth hormone, insulin, and cortisol concentrations in growing steers implanted with estradiol-17 beta, trenbolone acetate, or estradiol-17 beta plus trenbolone acetate. J Anim Sci. 1992 Jul;70(7):2109-19.
Henricks DM, Brandt RT Jr, Titgemeyer EC, Milton CT. Serum concentrations of trenbolone-17 beta and estradiol-17 beta and performance of heifers treated with trenbolone acetate, melengestrol acetate, or estradiol-17 beta. J Anim Sci. 1997 Oct;75(10):2627-33.
Schneider BA, Tatum JD, Engle TE, Bryant TC. Effects of heifer finishing implants on beef carcass traits and longissimus tenderness. J Anim Sci. 2007 Aug;85(8):2019-30.
Heitzman RJ. The effectiveness of anabolic agents in increasing rate of growth in farm animals; report on experiments in cattle. Environ Qual Saf Suppl. 1976;(5):89-98. Review.
Buttery, P., Vernon, B., & Pearson, J. (1978). Anabolic agents—some thoughts on their mode of action. Proceedings of the Nutrition Society, 37(3), 311-315
Grigsby ME, Trenkle 1986 Plasma growth hormone, insulin, glucocorticoids and thyroid hormones in large, medium and small breeds of steers with and without an estradiol implant Domestic Animal Endocrinology p.261-267
Breier, B. H., P. D. Gluckman, and J. J. Bass. 1988. Influence of nutritional status and oestradiol-17b on plasma growth hormone, insulin-like growth factors-I and -II and the response to exogenous growth hormone in young steers. J. Endocrinol. 118:243
Breier BH, Gluckman PD, Bass JJ. The somatotrophic axis in young steers: influence of nutritional status and oestradiol-17 beta on hepatic high- and low-affinity somatotrophic binding sites. J Endocrinol. 1988 Feb;116(2):169-77.
Chung KY, Baxa TJ, Parr SL, Luqué LD, Johnson BJ. Administration of estradiol, trenbolone acetate, and trenbolone acetate/estradiol implants alters adipogenic and myogenic gene expression in bovine skeletal muscle. J Anim Sci. 2012 May;90(5):1421-7.
Johnson BJ, Halstead N, White ME, Hathaway MR, DiCostanzo A, Dayton WR. Activation state of muscle satellite cells isolated from steers implanted with a combined trenbolone acetate and estradiol implant. J Anim Sci. 1998 Nov;76(11):2779-86.
Thompson SH, Boxhorn LK, Kong WY, Allen RE. Trenbolone alters the responsiveness of skeletal muscle satellite cells to fibroblast growth factor and insulin-like growth factor I. Endocrinology. 1989 May;124(5):2110-7.
Johnson BJ, Chung KY. Alterations in the physiology of growth of cattle with growth-enhancing compounds. Vet Clin North Am Food Anim Pract. 2007 Jul;23(2):321-32, viii. Review.
Johnson BJ, Hathaway MR, Anderson PT, Meiske JC, Dayton WR. Stimulation of circulating insulin-like growth factor I (IGF-I) and insulin-like growth factor binding proteins (IGFBP) due to administration of a combined trenbolone acetate and estradiol implant in feedlot cattle. J Anim Sci. 1996 Feb;74(2):372-9.
Schoonmaker JP, Loerch SC, Fluharty FL, Turner TB, Moeller SJ, Rossi JE, Dayton WR, Hathaway MR, Wulf DM. Effect of an accelerated finishing program on performance, carcass characteristics, and circulating insulin-like growth factor concentration of early-weaned bulls and steers. J Anim Sci. 2002 Apr;80(4):900-10.
Pampusch MS, Johnson BJ, White ME, Hathaway MR, Dunn JD, Waylan AT, Dayton WR. Time course of changes in growth factor mRNA levels in muscle of steroid-implanted and nonimplanted steers. J Anim Sci. 2003 Nov;81(11):2733-40.
Lee CY, Lee HP, Jeong JH, Baik KH, Jin SK, Lee JH, Sohnt SH. Effects of restricted feeding, low-energy diet, and implantation of trenbolone acetate plus estradiol on growth, carcass traits, and circulating concentrations of insulin-like growth factor (IGF)-I and IGF-binding protein-3 in finishing barrows. J Anim Sci. 2002 Jan;80(1):84-93.
Walker DK, Titgemeyer EC, Sissom EK, Brown KR, Higgins JJ, Andrews GA, Johnson BJ. Effects of steroidal implantation and ractopamine-HCl on nitrogen retention, blood metabolites and skeletal muscle gene expression in Holstein steers. J Anim Physiol Anim Nutr (Berl). 2007 Oct;91(9-10):439-47.
Winterholler SJ, Parsons GL, Walker DK, Quinn MJ, Drouillard JS, Johnson BJ. Effect of feedlot management system on response to ractopamine-HCl in yearling steers. J Anim Sci. 2008 Sep;86(9):2401-14.
Kamanga-Sollo E, White ME, Chung KY, Johnson BJ, Dayton WR. Potential role of G-protein-coupled receptor 30 (GPR30) in estradiol-17beta-stimulated IGF-I mRNA expression in bovine satellite cell cultures. Domest Anim Endocrinol. 2008 Oct;35(3):254-62.
Pampusch MS, White ME, Hathaway MR, Baxa TJ, Chung KY, Parr SL, Johnson BJ, Weber WJ, Dayton WR. Effects of implants of trenbolone acetate, estradiol, or both, on muscle insulin-like growth factor-I, insulin-like growth factor-I receptor, estrogen receptor-{alpha}, and androgen receptor messenger ribonucleic acid levels in feedlot steers. J Anim Sci. 2008 Dec;86(12):3418-23.
Kamanga-Sollo E, Pampusch MS, Xi G, White ME, Hathaway MR, Dayton WR. IGF-I mRNA levels in bovine satellite cell cultures: effects of fusion and anabolic steroid treatment. J Cell Physiol. 2004 Nov;201(2):181-9.
Bryant TC, Engle TE, Galyean ML, Wagner JJ, Tatum JD, Anthony RV, Laudert SB. Effects of ractopamine and trenbolone acetate implants with or without estradiol on growth performance, carcass characteristics, adipogenic enzyme activity, and blood metabolites in feedlot steers and heifers. J Anim Sci. 2010 Dec;88(12):4102-19.
Barton-Davis ER, Shoturma DI, Sweeney HL. Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiol Scand. 1999 Dec;167(4):301-5.
Reinhardt CD, Lee TL, Thomson DU, Mamedova LK, Bradford BJ. Restricted nutrient intake does not alter serum-mediated measures of implant response in cell culture. J Anim Sci Biotechnol. 2013 Nov 19;4(1):45.
Wu Y, Zhao W, Zhao J, Pan J, Wu Q, Zhang Y, Bauman WA, Cardozo CP. Identification of androgen response elements in the insulin-like growth factor I upstream promoter. Endocrinology. 2007 Jun;148(6):2984-93.
Preston RL, Bartle SJ, Kasser TR, Day JW, Veenhuizen JJ, Baile CA. Comparative effectiveness of somatotropin and anabolic steroids in feedlot steers. J Anim Sci. 1995 Apr;73(4):1038-47.
Elsasser TH, Rumsey TS, Kahl S, Czerwinski SM, Moseley WM, Ono Y, Solomon MB, Harris F, Fagan JM. Effects of Synovex-S and recombinant bovine growth hormone (Somavubove) on growth responses of steers: III. Muscle growth and protein responses. J Anim Sci. 1998 Sep;76(9):2346-53.
Martin MB, Stoica A. Insulin-like growth factor-I and estrogen interactions in breast cancer. J Nutr. 2002 Dec;132(12):3799S-3801S.
Venken K, Schuit F, Van Lommel L, Tsukamoto K, Kopchick JJ, Coschigano K, Ohlsson C, Movérare S, Boonen S, Bouillon R, Vanderschueren D. Growth without growth hormone receptor: estradiol is a major growth hormone-independent regulator of hepatic IGF-I synthesis. J Bone Miner Res. 2005 Dec;20(12):2138-49.
Umayahara Y, Kawamori R, Watada H, Imano E, Iwama N, Morishima T, Yamasaki Y, Kajimoto Y, Kamada T. Estrogen regulation of the insulin-like growth factor I gene transcription involves an AP-1 enhancer. J Biol Chem. 1994 Jun 10;269(23):16433-42.
Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science. 2005 Mar 11;307(5715):1625-30.
Prossnitz ER, Arterburn JB, Sklar LA. GPR30: A G protein-coupled receptor for estrogen. Mol Cell Endocrinol. 2007 Feb;265-266:138-42.
Filardo E, Quinn J, Pang Y, Graeber C, Shaw S, Dong J, Thomas P. Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology. 2007 Jul;148(7):3236-45.
Prossnitz ER, Arterburn JB, Smith HO, Oprea TI, Sklar LA, Hathaway HJ. Estrogen signaling through the transmembrane G protein-coupled receptor GPR30. Annu Rev Physiol. 2008;70:165-90.
Kellermeier JD, Tittor AW, Brooks JC, Galyean ML, Yates DA, Hutcheson JP, Nichols WT, Streeter MN, Johnson BJ, Miller MF. Effects of zilpaterol hydrochloride with or without an estrogen-trenbolone acetate terminal implant on carcass traits, retail cutout, tenderness, and muscle fiber diameter in finishing steers. J Anim Sci. 2009 Nov;87(11):3702-11.
Gonzalez JM, Carter JN, Johnson DD, Ouellette SE, Johnson SE. Effect of ractopamine-hydrochloride and trenbolone acetate on longissimus muscle fiber area, diameter, and satellite cell numbers in cull beef cows. J Anim Sci. 2007 Aug;85(8):1893-901.
Joubert Y, Tobin C, Lebart MC. Testosterone-induced masculinization of the rat levator ani muscle during puberty. Dev Biol. 1994 Mar;162(1):104-10.
McCarthy JJ, Mula J, Miyazaki M, Erfani R, Garrison K, Farooqui AB, Srikuea R, Lawson BA, Grimes B, Keller C, Van Zant G, Campbell KS, Esser KA, Dupont-Versteegden EE, Peterson CA. Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development. 2011 Sep;138(17):3657-66.
Hughes, N. J., G. T. Schelling, M. J. Garber, J. S. Eastridge, M. B. Solomon, and R. A. Roeder 1998. Skeletal muscle morphology alterations due to Posilac® and Revalor S® treatments alone or in combination in feedlot steers. J. Anim. Sci. 49(Proceedings Western Section):90–93
Sone K, Hinago M, Itamoto M, Katsu Y, Watanabe H, Urushitani H, Tooi O, Guillette LJ Jr, Iguchi T. Effects of an androgenic growth promoter 17beta-trenbolone on masculinization of Mosquitofish (Gambusia affinis affinis). Gen Comp Endocrinol. 2005 Sep 1;143(2):151-60.
Zhao JX, Hu J, Zhu MJ, Du M. Trenbolone enhances myogenic differentiation by enhancing β-catenin signaling in muscle-derived stem cells of cattle. Domest Anim Endocrinol. 2011 May;40(4):222-9.
Bodine SC, Latres E, Baumhueter S, Lai VK, Nunez L, Clarke BA, Poueymirou WT, Panaro FJ, Na E, Dharmarajan K, Pan ZQ, Valenzuela DM, DeChiara TM, Stitt TN, Yancopoulos GD, Glass DJ. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science. 2001 Nov 23;294(5547):1704-8.
Gomes MD, Lecker SH, Jagoe RT, Navon A, Goldberg AL. Atrogin-1, a muscle-specific F-box protein highly expressed during muscle atrophy. Proc Natl Acad Sci U S A. 2001 Dec 4;98(25):14440-5.
Lecker SH, Jagoe RT, Gilbert A, Gomes M, Baracos V, Bailey J, Price SR, Mitch WE, Goldberg AL. Multiple types of skeletal muscle atrophy involve a common program of changes in gene expression. FASEB J. 2004 Jan;18(1):39-51.
Danhaive PA, Rousseau GG. Evidence for sex-dependent anabolic response to androgenic steroids mediated by muscle glucocorticoid receptors in the rat. J Steroid Biochem. 1988 Jun;29(6):575-81.
Sharpe PM, Buttery PJ, Haynes NB. The effect of manipulating growth in sheep by diet or anabolic agents on plasma cortisol and muscle glucocorticoid receptors. Br J Nutr. 1986 Jul;56(1):289-304.
Reiter M, Walf VM, Christians A, Pfaffl MW, Meyer HH. Modification of mRNA expression after treatment with anabolic agents and the usefulness for gene expression-biomarkers. Anal Chim Acta. 2007 Mar 14;586(1-2):73-81.
Sheffield-Moore M. Androgens and the control of skeletal muscle protein synthesis. Ann Med. 2000 Apr;32(3):181-6. Review.
Santidrián S, Thompson JR, Young VR. Effect of trienbolone acetate on the rate of myofibrillar protein breakdown in young adrenalectomized male rate treated with corticosterone. Arch Farmacol Toxicol. 1981 Dec;7(3):333-40.
Isaacson WK, Jones SJ, Krueger RJ. Testosterone, Dihydrotestosterone, trenbolone acetate, and zeranol alter the synthesis of cortisol in bovine adrenocortical cells. J Anim Sci. 1993 Jul;71(7):1771-7.
Morgan SA, Sherlock M, Gathercole LL, Lavery GG, Lenaghan C, Bujalska IJ, Laber D, Yu A, Convey G, Mayers R, Hegyi K, Sethi JK, Stewart PM, Smith DM, Tomlinson JW. 11beta-hydroxysteroid dehydrogenase type 1 regulates glucocorticoid-induced insulin resistance in skeletal muscle. Diabetes. 2009 Nov;58(11):2506-15.
Coderre L, Srivastava AK, Chiasson JL. Role of glucocorticoid in the regulation of glycogen metabolism in skeletal muscle. Am J Physiol. 1991 Jun;260
Gounarides JS, Korach-André M, Killary K, Argentieri G, Turner O, Laurent D. Effect of dexamethasone on glucose tolerance and fat metabolism in a diet-induced obesity mouse model. Endocrinology. 2008 Feb;149(2):758-66.
Chan KH, Heitzman RJ, Kitchenham BA. Digestibility and N-balance studies on growing heifers implanted with trienbolone acetate. Br Vet J. 1975 Mar-Apr;131(2):170-4.
van Weerden EJ, Grandadam JA. The effect of an anabolic agent on N deposition, growth, and slaughter quality in growing castrated male pigs. Environ Qual Saf Suppl. 1976;(5):115-22.
Lobley GE, Connell A, Mollison GS, Brewer A, Harris CI, Buchan V, Galbraith H. The effects of a combined implant of trenbolone acetate and oestradiol-17 beta on protein and energy metabolism in growing beef steers. Br J Nutr. 1985 Nov;54(3):681-94.
Kerth CR, Montgomery JL, Morrow KJ, Galyean ML, Miller MF. Protein turnover and sensory traits of longissimus muscle from implanted and nonimplanted heifers. J Anim Sci. 2003 Jul;81(7):1728-35.
Kamanga-Sollo E, White ME, Hathaway MR, Weber WJ, Dayton WR. Effect of trenbolone acetate on protein synthesis and degradation rates in fused bovine satellite cell cultures. Domest Anim Endocrinol. 2011 Jan;40(1):60-6.
Rodway RG, Galbraith H. Effects of anabolic steroids on hepatic enzymes of amino acid catabolism. Horm Metab Res. 1979 Aug;11(8):489-90.
Compston JE. Sex steroids and bone. Physiol Rev. 2001 Jan;81(1):419-447. Review.
Leifke E, Körner HC, Link TM, Behre HM, Peters PE, Nieschlag E. Effects of testosterone replacement therapy on cortical and trabecular bone mineral density, vertebral body area and paraspinal muscle area in hypogonadal men. Eur J Endocrinol. 1998 Jan;138(1):51-8.
Martin AC. Osteoporosis in men: a review of endogenous sex hormones and testosterone replacement therapy. J Pharm Pract. 2011 Jun;24(3):307-15.
Bhasin S, Tenover JS. Age-associated sarcopenia–issues in the use of testosterone as an anabolic agent in older men. J Clin Endocrinol Metab. 1997 Jun;82(6):1659-60.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7.
Bhasin S, Cunningham GR, Hayes FJ, Matsumoto AM, Snyder PJ, Swerdloff RS, Montori VM; Task Force, Endocrine Society. Testosterone therapy in men with androgen deficiency syndromes: an Endocrine Society clinical practice guideline. J Clin Endocrinol Metab. 2010 Jun;95(6):2536-59.
Kasperk CH, Wakley GK, Hierl T, Ziegler R. Gonadal and adrenal androgens are potent regulators of human bone cell metabolism in vitro. J Bone Miner Res. 1997 Mar;12(3):464-71.
Wiren KM, Zhang X-W, Olson DA, Turner RT, Iwaniec UT. Androgen prevents hypogonadal bone loss via inhibition of resorption mediated by mature osteoblasts/osteocytes. Bone. 2012;51(5):835-846.
Vandenput L, Ohlsson C. Estrogens as regulators of bone health in men. Nat Rev Endocrinol. 2009 Aug;5(8):437-43.
Smith ZK, Thompson AJ, Hutcheson JP, Nichols WT, Johnson BJ. Evaluation of Coated Steroidal Implants Containing Trenbolone Acetate and Estradiol-17β on Live Performance, Carcass Traits, and Sera Metabolites in Finishing Steers. J Anim Sci. 2018 Mar 9.
L’integrazione con Picnogenolo potrebbe aumentare la perdita di grasso. Questo effetto potenziale è suggerito da uno studio svolto su animali da parte di ricercatori cinesi, il quale è stato pubblicato sul Journal of Diabetes Research.(1) Secondo questo studio, il Picnogenolo modifica il funzionamento degli adipociti. Il composto fa si che le cellule adipose rilascino più acidi grassi nel flusso ematico elevando al tempo stesso il dispendio energetico cellulare.
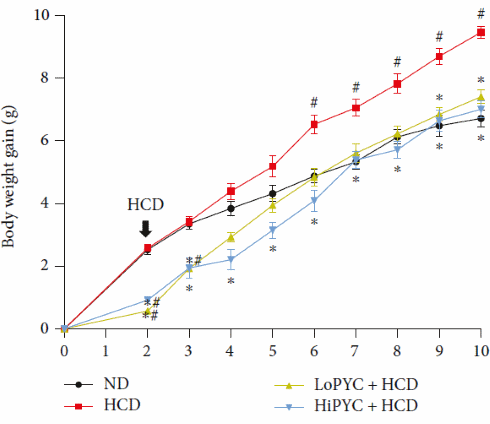
I ricercatori hanno sottoposto dei topi ad un regime alimentare ipercalorico [HCD] per 8 settimane. I topi, ovviamente, hanno subito un aumento della massa grassa. Un gruppo di controllo è stato sottoposto ad un regime alimentare standard [ND] durante lo stesso periodo di tempo. E, come prevedibile, i topi del gruppo di controllo non hanno subito modifiche nella composizione corporea.
Ad un gruppo di topi è stata somministrata una dose giornaliera relativamente bassa (30mg/Kg) di Picnogenolo [LoPYC + HCD], oltre ad essere sottoposti ad una alimentazione ad alto contenuto calorico. La dose utilizzata rapportata ad un essere umano equivarrebbe a circa 300mg di Picnogenolo al giorno.
Ad un altro gruppo di topi è stata somministrata una dose relativamente alta (100mg/Kg)di Picnogenolo [HiPYC + HCD]. La dose utilizzata rapportata ad un essere umano equivarrebbe a circa 1g di Pycnogenolo al giorno.
I ricercatori hanno utilizzato l’estratto di Picnogenolo commercializzato dalla svizzera Horphag.(2) Nonostante ciò, i ricercatori non hanno ricevuta alcun finanziamento dalla Horphag; hanno ottenuto i finanziamenti dal governo cinese.
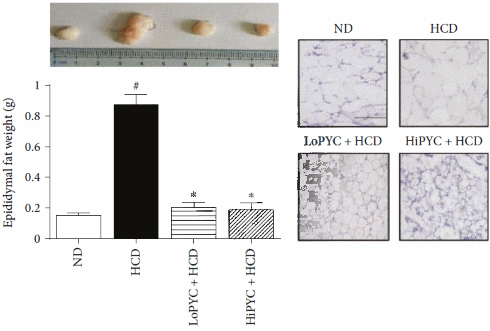
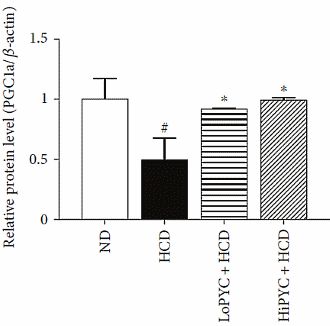
La supplementazione sia a basso [LoPYC + HCD] che ad alto dososaggio di Picnogenolo [HiPYC + HCD] ha mostrato un potenziale di eliminazione dell’effetto ingrassante dato dall’alimentazione ad alto contenuto calorico [HCD], e ha inibito la crescita del tessuto adiposo.
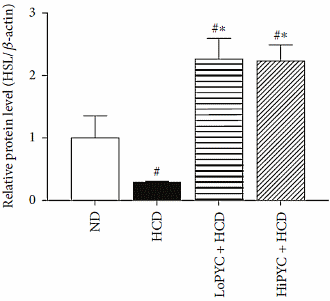
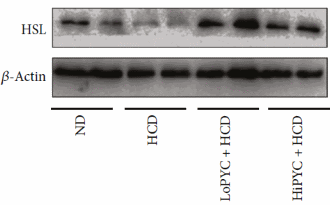
Negli adipociti del tessuto adiposo bianco, il Picnogenolo ha inibito il gene PLN1, che è coinvolto nella conservazione del grasso. Contemporaneamente, il Picnogenolo ha mostrato di attivare i PPAR-α e γ (implicati nella regolazione del deposito degli acidi grassi e del metabolismo del glucosio). Più sorprendentemente, il Picnogenolo ha aumentato la concentrazione dell’Ormone Lipasi Sensibile [HSL] nel tessuto adiposo bianco. Questo enzima è il principale responsabile della mobilizzazione dei Trigliceridi dal tessuto adiposo, e fa sì che gli adipociti rilascino gli acidi grassi liberi nel flusso ematico (anche se esistono altre lipasi coinvolte in questo processo). Il corpo può usare questi acidi grassi liberi come substrato energetico.
Il Picnogenolo ha anche agito mantenendo la concentrazione di PGC-1-α nelle cellule adipose. Il PGC-1-α stimola la biogenesi mitocondriale nelle cellule (aumento del numero dei mitocondri). Maggiore è il numero di mitocondri nelle cellule, più nutrienti possono essere convertiti in energia.
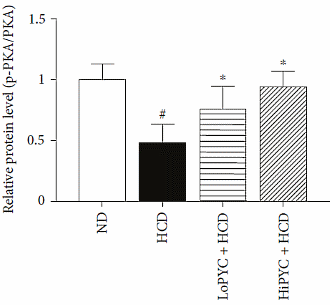
Tutti questi processi sono probabilmente dipendenti dall’azione attivante del Picnogenolo nei confronti della proteina chinasi cAMP–dipendente [PKA].
I ricercatori affermano che, i loro dati dimostrano che il Picnogenolo può ridurre efficacemente il peso corporeo e i depositi adiposi. Questa scoperta suggerisce che la somministrazione di Picnogenolo può essere una nuova strategia per prevenire e trattare l’obesità e le malattie metaboliche associate.
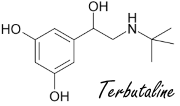
Il meccanismo scoperto suggerisce che l’effetto dimagrante del Picnogenolo aumenta in combinazione con i beta-agonisti. Il PKA aumenta l’attività beta-adrenergica. Il fatto che i fenoli [le sostanze attive contenute nel Picnogenolo sono fenoli] rafforzino l’azione dei beta-agonisti è stato dimostrato in precedenti studi.(3)
Nella prima parte di questa serie di articoli, ho esposto diverse informazioni riguardanti principalmente le caratteristiche metaboliche del Trenbolone nei mammiferi. Se non avete già letto la prima parte, vi esorto a farlo prima di proseguire con la lettura della seconda parte, poiché alcuni degli argomenti trattati in precedenza verranno approfonditi nel presente articolo.
VI. Effetti sull’Asse HPG
Nei vertebrati, l’Asse Ipotalamo-Ipofisi-Gonadi (HPG), conosciuto nell’uomo anche come HPTA, controlla i processi riproduttivi attraverso una varietà di ormoni che agiscono sui tessuti bersaglio direttamente o indirettamente. Ad alti livelli, nei maschi, l’Ormone Rilasciante la Gonadotropina (GnRH) secreto dall’ipotalamo stimola l’ipofisi a rilasciare l’Ormone Luteinizzante (LH) e l’Ormone Follicolo-Stimolante (FSH). Questi, a loro volta, stimolano il rilascio di ormoni sessuali dai testicoli (l’FSH incrementa la risposta all’LH attraverso l’up- regulation dei recettori nelle cellule di Leydig.).(1)(2) La forte connessione tra i diversi componenti del sistema che caratterizza l’Asse HPG si traduce nel fatto che nessuno di questi funzioni in maniera isolata. Entrando in circolo, sia gli ormoni androgenici che quelli estrogenici sono in grado di attraversare la barriera emato-encefalica e di esercitare un feedback negativo sull’ipofisi e sull’ipotalamo, sottoregolando quindi il rilascio del GnRH e, di conseguenza, causando una sottoregolazione/soppressione del funzionamento dell’intero asse.(3)
La somministrazione di Trenbolone è associata a numerosi tipi di alterazioni dell’Asse HPG, il che è in linea con ciò che è stato osservato con vari altri trattamenti androgeni nel corso degli anni. (4) Alcune delle alterazioni indotte dal Trenbolone osservate negli anni includono livelli ridotti di LH serico (5)(6)(7)(8)(9)(10), ridotti livelli di FSH serico (11), ridotti livelli di Testosterone serico (5)(6) (7)(8)(9)(10)(12)(13)(14)(15)(16), ridotti livelli di DHT serico (11), ridotti livelli di estradiolo serico (13)(15), atrofia testicolare (7)(17)(18) e un principio di ritardato della pubertà (19). Questi effetti si verificano abbastanza rapidamente poiché, in uno studio svolto su ratti, sono stati registrati entro dieci giorni dalla somministrazione di Trenbolone Enantato tassi di soppressione del Testosterone serico pari all’80% e tassi di soppressione del DHT serico pari al 70% rispetto agli animali di controllo. (20) Vale la pena notare che essendo il Trenbolone Enantato (TBE) una variante a lunga durata d’azione, con l’uso del Trenbolone Acetato (TBA) gli effetti registrati si sarebbero potenzialmente verificati in tempi più rapidi.
Non è del tutto chiaro quali siano i meccanismi attraverso i quali si esplicano gli effetti di soppressione dati dal Trenbolone sull’Asse HPG, tuttavia ci sono state certamente alcune prove nel corso degli anni che forniscono indizi in merito. Un’ipotesi diffusa prevede l’inibizione diretta del feedback ipotalamico, come evidenziato dalla ridotta trascrizione di GnRH osservata nel cervello dei modelli animali (pesci). Questo può essere additivo ai suoi effetti diretti sulla biosintesi degli steroidi testicolari, come sostenuto dalla sottoregolazione dell’espressione del CYP17 testicolare.(21) Il CYP17 è un enzima molto importante nella biosintesi degli steroidi e catalizza sequenzialmente due reazioni chiave nella produzione di steroidi sessuali nei maschi.
È anche interessante notare che qualunque sia il meccanismo, non sembra essere dipendente dal Recettore degli Androgeni (AR).(22)(23) Sostenendo ulteriormente questa linea di pensiero, nelle colture di tessuto ovarico di pesce, androgeni non aromatizzabili come il Trenbolone hanno mostrato effetti inibitori diretti e non genomici, anti-androgeno-insensibili, sulla sintesi di estrogeni.(24) È altamente probabile che i meccanismi di feedback alla base dell’alterazione dell’Asse HPG da parte del Trenbolone siano del tutto simili ad altri androgeni, con conseguente inibizione dei livelli di GnRH e, in definitiva, la sottoregolazione/inibizione della produzione di FSH e LH.(25)
Un altro potenziale gene candidato coinvolto nelle diminuite concentrazioni di steroidi sessuali, osservate in esperimenti su pesci i quali erano stati esposti al forte androgeno 17-trenbolone, è l’idrossisteroide (17β) deidrogenasi 12a (hsd17b12a). L’Hsd17b12a catalizza la conversione del Androstenedione in Testosterone che, a sua volta, viene convertito in 17β-estradiolo dall’enzima aromatasi. Pertanto, la sottoregolazione del hsd17b12a, come si può osservare nei pesci esposti al Trenbolone, ha come prevedibile risposta un declino sia nei livelli di Testosterone che di Estradiolo.(14)
VII. Effetti sui Pathways Anabolici
Come già detto nella prima parte, il Trenbolone è considerabile quale SARM con il suo indice terapeutico pari a circa 3,4. Vorrei utilizzare questa sezione per discutere parte delle caratteristiche che fanno del Trenbolone un composto SARM-simile.
5α-Reduttasi
Nonostante abbia una somiglianza strutturale con il Testosterone, il Trenbolone non è soggetto alla 5α-riduzione a causa della presenza di una struttura a 3-ossotrieni che impedisce la riduzione dell’anello A.(26) Questo è il percorso enzimatico utilizzato per la conversione del Testosterone nel suo metabolita fortemente androgeno Dihydrotestosterone (DHT). Poiché il Trenbolone non è un substrato soggetto all’azione del enzima 5α-reduttasi, ha dimostrato di stimolare effetti androgenici meno pronunciati rispetto al Testosterone nei tessuti sensibili agli androgeni che esprimono l’enzima 5α-reduttasi, compresi gli organi sessuali accessori e la prostata.(27)(28)(29)(30)(31)(32). Per rendere meglio l’idea di ciò che si è appena affermato, il Testosterone ha una potenza circa tre volte superiore nei tessuti androgenici che esprimono l’enzima 5α-reduttasi nonostante abbia un’affinità di legame ai AR significativamente più bassa rispetto al Trenbolone.(33) Discuteremo di come questa caratteristica influenzi il potenziale ipertrofico in questi tessuti in seguito.
Come presumibile, un motivo particolare per cui il Trenbolone sta iniziando a prendere piede nella comunità scientifica è dovuto al suo potenziale di ridurre i rischi associati al cancro alla prostata nei pazienti trattati per l’ipogonadismo. L’attuale strategia per il trattamento degli individui ipogonadici consiste nella somministrazione di Testosterone al fine di ripristinare i livelli fisiologici dell’ormone. Tuttavia, nei maschi adulti, la crescita benigna e maligna del tessuto prostatico ghiandolare è ampiamente regolata dagli ormoni sessuali. Inoltre, è stato dimostrato che anche aumenti moderati del Testosterone circolante si traducono direttamente in pronunciati effetti iperplastici nei tessuti prostatici, mediati dalla sua 5α riduzione in DHT.(34)(35) Più avanti, indagheremo più a fondo nella letteratura disponibile per constatare se il potenziale attribuito al Trenbolone di abbassare il rischio di cancro alla prostata sia una realtà concreta.
Enzima Aromatasi
Enzima Aromatasi
Stranamente, uno dei quesiti sul Trenbolone maggiormente posti è se questo AAS abbia o meno effetti sui livelli degli estrogeni serici, e se possa aromatizzare o meno come il Testosterone. E’ un fatto pienamente riconosciuto nella comunità scientifica che il Trenbolone, ed altri composti C19 nor-steroidi, non sia soggetto all’azione dell’enzima aromatasi.(36)(37) Detto questo, è necessario che voi capiate che la precedente affermazione non sta a significare che i composti C19 nor-steroidi non possano essere convertiti in estrogeni o non possano indurre effetti estrogenici. (38)(39)
Il Trenbolone è in gran parte ritenuto non avere alcuna attività estrogenica (40)(41) e ci sono stati numerosi studi sugli animali che hanno dimostrato che il suo uso causa una riduzione delle concentrazioni ematiche di Estradiolo. (13)(14)(15)(42)(43)(44)Tenete presente che ci sono stati alcuni studi che non hanno mostrato questo effetto soppressivo sui livelli estrogenici (10)(45)(46) ma, nel complesso, il corpo della letteratura nel suo complesso supporta l’ipotesi che il Trenbolone possieda effetti anti-estrogenici.
Sulla base di ciò che ora sappiamo sull’Asse HPG, avrebbe sicuramente senso pensare che gli effetti anti-estrogenici causati dalla somministrazione di Trenbolone siano probabilmente legati al suo feedback negativo sull’Asse. Questo feedback negativo, causando l’inibizione della produzione endogena di Testosterone, porta ad una riduzione dei substrati soggetti all’azione dell’enzima aromatasi, il quale è necessario per la biosintesi degli estrogeni endogeni nei maschi. Questo impatto sull’Asse HPG causerebbe un tasso di inibizione estrogenica più marcato rispetto a qualsiasi potenziale effetto diretto del Trenbolone sui recettori degli estrogeni e / o l’enzima aromatasi.(5)(6)(8)(21)(47) L’impatto di questo AAS sui livelli estrogenici e la loro attività potrebbe contare anche un meccanismo secondario legato alla capacità del Trenbolone di sottoregolare l’espressione di entrambi i recettori estrogenici α e β (REα/Reβ).(48)
Vi sono state altre scoperte interessanti riguardo ai meccanismi alla base della relazione del Trenbolone con gli estrogeni, così come le risposte compensatorie associate ai livelli dell’ormone soppresso. È stato dimostrato che il Trenbolone riduce le concentrazioni tissutali e l’espressione genica della VTG (Vitellogenina), una proteina positivamente associata all’esposizione a composti estrogenici. (13)(14)(21)(41)(47)(49)(50)(51)(52)(53)(54) E’ stata anche dimostrata una sottoregolazione a livello cerebrale del CYP19B (aromatasi B) e una sovra regolazione gonadica del CYP19A (aromatasi A) in pesci femmina, anche se la cosa non è risultata interessante nei maschi. (14)(54)
Flutamide
L’impatto del Trenbolone sugli estrogeni non sembra essere AR-dipendente, poiché gli studi hanno dimostrato che la co-somministrazione con un antagonista dei AR (Flutamide) ha portato alla stessa attività anti-estrogenica nei pesci.(13) È interessante notare che c’è stato un altro studio sui pesci il quale ha riportato che il Trenbolone possiede una bassa affinità per il recettore degli estrogeni ma può potenzialmente attivarlo.(44) Che questa caratteristica sia specifica o meno per la specie presa in esame è questione di dibattito, poiché in altri studi che ho esaminato non ho riscontrato che tale effetto si verifichi. Tuttavia, esperimenti di coltura cellulare e analisi biologiche dimostrano che il Trenbolone ed i suoi metaboliti hanno un’affinità di legame molto bassa con i recettori degli estrogeni, circa il 20% dell’affinità dell’Estradiolo.(40)
Quindi, il Trenbolone può aromatizzare? Date le informazioni sopra esposte (soppressione dei livelli di E2) e la presenza di un doppio legame inserito in C9– C10 che esclude teoricamente tale possibilità, al momento non ho trovato nulla che suggerisca in via definitiva che ciò possa verificarsi. Esiste però un’ipotesi esposta da Holland et al. (55) che trovo abbastanza interessante da includere nella sua interezza:
“In precedenza abbiamo riportato che il Trenbolone Enantato riduce la massa grassa viscerale in animali giovani e anziani ORX, indicando che la perdita di grasso si verifica in risposta alla somministrazione di androgeni, anche in assenza di un substrato androgenico soggetto all’azione dell’enzima aromatasi. Tuttavia, il nostro precedente lavoro non ha tenuto conto della possibilità che l’Androstenedione (derivato dal Deidroepiandrosterone) possa essere aromatizzato in Estrone e, successivamente, convertito in E2 per azione del 17β-idrossisteroide deidrogenasi nei tessuti, come il grasso, esprimendo gli enzimi richiesti.”
Progesterone/SHBG
Dimero SHBG umano.
È stato dimostrato che il Trenbolone possiede un’alta affinità per il recettore progestinico dei bovini e si presume che abbia un’affinità simile allo stesso Progesterone verso tale recettore.(56) L’analisi in vitro ha rivelato che la relativa affinità di legame con il recettore del progesterone bovino, rispetto al progesterone stesso, era del 137,4% per 17β-TbOH e del 2,1% per 17α-TbOH (57). Infine, la relativa affinità di legame tra Trenbolone e SHBG umano, rispetto al DHT, è del 29,4% per il 17β-TbOH e del 94,8% per il 17α-TbOH.
VIII. Effetti sui marker della salute metabolica
Uno dei motivi principali per cui i detrattori del Trenbolone dispensano ammonimenti contro il suo uso è legato alla severità con cui il composto incide apparentemente sui marker della salute. Speravo di avere qualche riferimento effettivo su esami ematici da aggiungere a questo articolo, ma sfortunatamente i miei sforzi di crowdsourcing non hanno avuto successo poiché non molte persone usano il Trenbolone da solo. Quindi quello che farò in questa sezione è proseguire con l’esposizione della letteratura scientifica disponibile (studi su animali) che espone l’impatto del Trenbolone su diversi marker della salute.
Asse Ipotalamo-Ipofisi-Tiroide (Asse Tiroideo)
Il rapporto tra il Trenbolone e l’Asse Tiroideo è decisamente dibattuto nel mondo del BodyBuilding. Sebbene gli effetti osservati siano stati un po’ incoerenti, sembra esserci un pattern che suggerisce che il Trenbolone abbia un effetto soppressivo complessivo sull’Asse Tiroideo. In uno studio, il Trenbolone ha ridotto il T4 nelle giovenche, il Trenbolone e l’Estradiolo hanno ridotto il T4 nei manzi, mentre non è stato osservato alcun impatto sul uptake del T3.(58) In un altro studio, il Trenbolone e l’Estradiolo hanno effettivamente aumentato il T3, mentre il Trenbolone da solo ha ridotto sia il T3 che il T4.(59) Bisogna ricordare, però, che l’Estradiolo stimola l’asse GH / IGF, il che aumenta acutamente la conversione del T4 in T3. Ciò può aiutare a spiegare il perché la co-somministrazione con estradiolo può portare a livelli più elevati di T3 mentre la somministrazione di solo Trenbolone ha come conseguenza l’effetto opposto, poiché i livelli di Estradiolo sono marcatamente soppressi. Anche negli studi in cui i livelli tiroidei non sono risultati significativamente differenti, il Trenbolone ha ridotto i tassi metabolici a digiuno, portando ad un minor fabbisogno calorico per creare un surplus energetico.(60)
Pertanto, può essere ragionevole ipotizzare che l’aumentata efficienza alimentare vista in numerosi studi nel corso degli anni potrebbe essere correlata ad una alterazione metabolica trenbolone-mediata. Naturalmente, va notato che i bovini più magri tendono a crescere più velocemente e ad utilizzare i mangimi in modo più efficiente, quindi potrebbe anche essere solo un sottoprodotto di questo.(61) Prima di concludere questa serie di articoli, parlerò un po’ più approfonditamente della gestione pratica di questo possibile impatto dato dal Trenbolone.
Colesterolo
In generale, esiste una forte correlazione tra la perdita di grasso e i cambiamenti favorevoli nei livelli serici dei lipidi, in particolare negli uomini. (62)(63) Pertanto, poiché il Trenbolone ha dimostrato costantemente di migliorare la composizione corporea, è ragionevole ipotizzare che possa avere un impatto favorevole sui marker lipidici. Quindi, vediamo quali prove esistono a riguardo analizzando le informazioni provenienti dagli studi svolti sugli animali.
Studi sui ratti hanno dimostrato che sia il Testosterone che il Trenbolone possiedono capacità protettive simili contro il colesterolo elevato nonostante la capacità del Trenbolone di provocare la perdita del grasso viscerale sia maggiore. Ciò suggerisce che i livelli serici di colesterolo possono essere regolati principalmente dalla composizione corporea complessiva, indipendentemente dai cambiamenti nelle riserve adipose viscerali. In uno studio, il colesterolo totale serico, l’HDL e l’LDL erano tutti significativamente più bassi nei ratti trattati con Trenbolone rispetto ai ratti di controllo (- 62%, – 57% e – 78% rispettivamente). Il sottoprodotto di questo era che i ratti trattati avevano un rapporto HDL:LDL maggiore. Anche i Trigliceridi serici erano diminuiti di un significativo 51% rispetto ai ratti di controllo.(15) In un altro studio, sia il Testosterone che il Trenbolone hanno ridotto il colesterolo circolante nei ratti alimentati con una dieta ricca di grassi e zuccheri, ma solo il Trenbolone ha ridotto i livelli di Trigliceridi circolanti.(64) Si consiglia vivamente di tenere d’occhio i livelli di colesterolo quando si usano dosi sovrafisiologiche di androgeni, in quanto tendono ad avere la capacità di aumentare lo stimolo delle catecolamine sull’ormone lipasi sensibile (HSL) nel tessuto epatico e cardiaco.(65)(66) Questa aumentata attività della HSL tende a determinare un aumento del tasso di mobilitazione dei trigliceridi e un aumento della degradazione del HDL. Avere livelli di HDL cronicamente bassi rappresenta un fattore di rischio cardiovascolare indipendente, quindi, un uso prolungato di AAS che portano ad una soppressione marcata del HDL (vedi, per esempio, composti metilati in C-17) dovrebbe essere attentamente monitorato.(67)
Marker epatici
Enzimi epatocellulari (ALT e AST) sono immagazzinati all’interno delle cellule epatiche e gli enzimi epatici colestatici (es. ALP e GGT) sono associati a dotti biliari esterni alle cellule epatiche.
Ci sono alcuni marker epatici comunemente utilizzati per valutare la funzionalità e la salute del fegato, così come il danno a quest’organo. L’albumina è un marker indicativo della funzione epatica generale mentre AST, ALT, ALP sono tutti marker generali del danno epatico.
Recenti studi svolti sui roditori hanno dimostrato che il Trenbolone non sembra indurre danni significativi al tessuto epatico. In uno studio, campioni di tessuto epatico di ratti trattati con Trenbolone hanno mostrato una morfologia simile a quella dei ratti di controllo. AST, ALT, ALP e albumina erano tutti a livello simile tra i ratti trattati con Trenbolone ed i ratti del gruppo di controllo.(15) In un altro studio, sono stati osservati valori simili negli enzimi epatici con ratti alimentati con una dieta ricca di grassi e zuccheri in tutti i gruppi di trattamento, compresi i gruppi trattati con Testosterone e Trenbolone.(64) Questo steroide possiede comunque un forte livello di resistenza alla disattivazione epatica, e una significativa tossicità epatica è stata osservato nei body builder che abusano del Trenbolone.(68) Sebbene non sia una costante, l’epatotossicità non può essere completamente esclusa, in particolare con alte dosi.
Insulina
L’Insulina è un altro ormone che tende ad avere una correlazione diretta con il grasso corporeo, e in particolare con i livelli di grasso viscerale. L’accumulo di grasso viscerale e l’aumento dei livelli di trigliceridi circolanti sono entrambi associati con il peggioramento dell’insulino-resistenza. (69) Al contrario, la restrizione calorica e la consequenziale perdita di peso nei soggetti non diabetici visceralmente obesi ha indotto miglioramenti significativi nella sensibilità all’insulina.(70) Oltre all’obesità, esistono anche prove convincenti che dimostrano come bassi livelli di androgeni promuovano il peggioramento dell’insulino-rsistenza. (71) È stato ipotizzato che il trattamento con Trenbolone in modelli animali possa promuovere effetti di miglioramento della sensibilità all’insulina attraverso meccanismi simili a quelli ottenuti dalla restrizione calorica nei soggetti umani di sesso maschile.
Uno studio ha dimostrato che l’Insulina serica è significativamente più bassa nei ratti trattati con Trenbolone (riduzione del 38%) rispetto ai ratti di controllo, e che ciò si è tradotto in un valore HOMA-IR (indice utilizzato per valutare l’insulino resistenza) significativamente più basso.(15) I ratti che sono stati sottoposti a regimi alimentari ad alto contenuto di grassi e zuccheri presentavano livelli di Insulina serica significativamente elevati e che sono stati ripristinati solo parzialmente con la somministrazione di Testosterone, mentre con la somministrazione di Trenbolone i livelli insulinici si sono ridotti significativamente.(64) In effetti, il gruppo trattato con Trenbolone era l’unico a mostrare valori di HOMA-IR ridotti, indicando un aumento della funzionalità delle cellule beta e una riduzione della resistenza all’insulina. Quindi, anche se limitata, l’evidenza suggerisce che il Trenbolone abbia effetti sulla sensibilità all’Insulina superiore al Testosterone.
L’adiponectina è un’adipochina sensibilizzante dell’insulina da 30 kDa che viene principalmente secreta dal tessuto adiposo viscerale.(72)(73) In generale, i livelli serici di adiponectina sono inversamente proporzionali alla massa grassa (74). Il Testosterone ed il Trenbolone tendono a ridurre i livelli totali di adiponectina in misura simile nei ratti (75).
Eritropoiesi
L’eritropoiesi è il processo di formazione dei globuli rossi (RBC) attraverso una serie di elementi cellulari immaturi (serie eritroblastica). Uno degli effetti collaterali più comunemente riportati durante le TRT è l’aumento dei livelli di ematocrito ed emoglobina. Nello specifico, la deprivazione androgenica riduce sia l’ematocrito che l’emoglobina mentre la somministrazione di Testosterone determina un aumento dose-dipendente di entrambi.(76)(77)
Eritropoietina (EPO)
I meccanismi mediante i quali gli androgeni aumentano la produzione di RBC possono essere direttamente correlati alla stimolazione della secrezione di eritropoietina renale o persino del midollo osseo.(78) E sulla base delle prove esistenti, sembrerebbe che gli androgeni elevino direttamente l’eritropoiesi attraverso meccanismi AR-mediati.(11) Non sembra che l’aromatizzazione del Testosterone sia necessaria per l’eritropoiesi poiché la somministrazione di DHT ne causa un analogo aumento nei soggetti di sesso maschile.(79) Inoltre, è stato dimostrato che ciò si verifica anche in soggetti di sesso maschile con carenza dell’enzima aromatasi.(80) Allo stesso modo, per l’eritropoiesi non sembra essere necessaria una riduzione della 5α riduzione del Testosterone poiché la co-somministrazione di Testosterone e Finasteride (inibitore dell’enzima 5α-riduttasi) ha portato ad un aumento sia dell’ematocrito che dell’emoglobina nella stessa misura data dalla sola somministrazione di Testosterone nonostante una riduzione nei livelli di DHT del 65% nel gruppo Finasteride .(81)
Se il Trenbolone non causasse l’innalzamento dell’ematocrito e dell’emoglobina osservati con le tradizionali TRT, allora questo potrebbe essere un altro potenziale motivo che renderebbe questa molecola un candidato interessante per le HTR.
Prove preliminari indicano che il Trenbolone aumenta l’emoglobina nei roditori maschi in modo dose-dipendente, e in misura leggermente superiore rispetto a dosi sovrafisiologiche di Testosterone (8-10%), nonostante il DHT sia soppresso di oltre il 70% dopo la somministrazione.(20) In un altro studio, a dosi somministrate che erano sette volte superiori a quelle del Testosterone, i ratti trattati con Trenbolone avevano livelli di emoglobina quasi identici, sebbene entrambi fossero significativamente elevati rispetto al gruppo di controllo.(82)
Penso che le informazioni esposte fino a questo punto siano più che sufficienti per concludere questa seconda parte. Nella terza parte, inizieremo ad approfondire il potenziale ipertrofico del Trenbolone.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
Simoni M, Weinbauer GF, Gromoll J, Nieschlag E. Role of FSH in male gonadal function. Ann Endocrinol (Paris). 1999 Jul;60(2):102-6. Review.
Liu PY, Iranmanesh A, Nehra AX, Keenan DM, Veldhuis JD. Mechanisms of hypoandrogenemia in healthy aging men. Endocrinol Metab Clin North Am. 2005 Dec;34(4):935-55, ix. Review.
Kumar TR, Low MJ. Hormonal regulation of human follicle-stimulating hormone-beta subunit gene expression: GnRH stimulation and GnRH-independent androgen inhibition. Neuroendocrinology. 1995 Jun;61(6):628-37.
Tan RS, Scally MC. Anabolic steroid-induced hypogonadism–towards a unified hypothesis of anabolic steroid action. Med Hypotheses. 2009 Jun;72(6):723-8.
Fabry J, Renaville R, Halleux V, Burny A. Plasma testosterone and LH responses to LHRH in double-muscled bulls treated with trenbolone acetate and zeranol. J Anim Sci. 1983 Nov;57(5):1138-45.
Gettys TW, D’Occhio MJ, Henricks DM, Schanbacher BD. Suppression of LH secretion by oestradiol, dihydrotestosterone and trenbolone acetate in the acutely castrated bull. J Endocrinol. 1984 Jan;100(1):107-12.
Silcox RW, Keeton JT, Johnson BH. Effects of zeranol and trenbolone acetate on testis function, live weight gain and carcass traits of bulls. J Anim Sci. 1986 Aug;63(2):358-68.
Renaville R, Burny A, Sneyers M, Rochart S, Portetelle D, Théwis A. Effects of an anabolic treatment before puberty with trenbolone acetate-oestradiol or oestradiol alone on growth rate, testicular development and luteinizing hormone and testosterone plasma concentrations. Theriogenology. 1988 Feb;29(2):461-76.
Lee CY, Henricks DM, Skelley GC, Grimes LW. Growth and hormonal response of intact and castrate male cattle to trenbolone acetate and estradiol. J Anim Sci. 1990 Sep;68(9):2682-9.
Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci. 1991 Jun;69(6):2452-62.
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
Galbraith, H. (1982). Growth, hormonal and metabolic response of post-pubertal entire male cattle to trenbolone acetate and hexoestrol. Animal Science, 35(2), 269-276.
Ankley GT, Defoe DL, Kahl MD, Jensen KM, Makynen EA, Miracle A, Hartig P, Gray LE, Cardon M, Wilson V. Evaluation of the model anti-androgen flutamide for assessing the mechanistic basis of responses to an androgen in the fathead minnow (Pimephales promelas). Environ Sci Technol. 2004 Dec 1;38(23):6322-7.
Dorts J, Richter CA, Wright-Osment MK, Ellersieck MR, Carter BJ, Tillitt DE. The genomic transcriptional response of female fathead minnows (Pimephales promelas) to an acute exposure to the androgen, 17beta-trenbolone. Aquat Toxicol. 2009 Jan 18;91(1):44-53.
Donner DG, Beck BR, Bulmer AC, Lam AK, Du Toit EF. Improvements in body composition, cardiometabolic risk factors and insulin sensitivity with trenbolone in normogonadic rats. Steroids. 2015 Feb;94:60-9.
Ma F, Liu D. 17β-trenbolone, an anabolic-androgenic steroid as well as an environmental hormone, contributes to neurodegeneration. Toxicol Appl Pharmacol. 2015 Jan 1;282(1):68-76.
O’Lamhna M, Roche JF. Effect of repeated implantation with anabolic agents on growth rate, carcase weight, testicular size and behaviour of bulls. Vet Rec. 1983 Dec 3;113(23):531-4.
López-Bote C, Sancho G, Martínez M, Ventanas J, Gázquez A, Roncero V. Trenbolone acetate induced changes in the genital tract of male pigs. Zentralbl Veterinarmed B. 1994 Mar;41(1):42-8.
Moran C, Prendiville DJ, Quirke JF, Roche JF. Effects of oestradiol, zeranol or trenbolone acetate implants on puberty, reproduction and fertility in heifers. J Reprod Fertil. 1990 Jul;89(2):527-36.
Yarrow JF, Conover CF, McCoy SC, Lipinska JA, Santillana CA, Hance JM, Cannady DF, VanPelt TD, Sanchez J, Conrad BP, Pingel JE, Wronski TJ, Borst SE. 17β-Hydroxyestra-4,9,11-trien-3-one (trenbolone) exhibits tissue selective anabolic activity: effects on muscle, bone, adiposity, hemoglobin, and prostate. Am J Physiol Endocrinol Metab. 2011 Apr;300(4):E650-60.
Zhang X, Hecker M, Park JW, Tompsett AR, Jones PD, Newsted J, Au DW, Kong R, Wu RS, Giesy JP. Time-dependent transcriptional profiles of genes of the hypothalamic-pituitary-gonadal axis in medaka (Oryzias latipes) exposed to fadrozole and 17beta-trenbolone. Environ Toxicol Chem. 2008 Dec;27(12):2504-11.
Heinlein CA, Chang C. The roles of androgen receptors and androgen-binding proteins in nongenomic androgen actions. Mol Endocrinol. 2002 Oct;16(10):2181-7. Review.
Martinović D, Blake LS, Durhan EJ, Greene KJ, Kahl MD, Jensen KM, Makynen EA, Villeneuve DL, Ankley GT. Reproductive toxicity of vinclozolin in the fathead minnow: confirming an anti-androgenic mode of action. Environ Toxicol Chem. 2008 Feb;27(2):478-88.
Braun AM, Thomas P. Androgens inhibit estradiol-17beta synthesis in Atlantic croaker (Micropogonias undulatus) ovaries by a nongenomic mechanism initiated at the cell surface. Biol Reprod. 2003 Nov;69(5):1642-50.
MacIndoe JH, Perry PJ, Yates WR, Holman TL, Ellingrod VL, Scott SD. Testosterone suppression of the HPT axis. J Investig Med. 1997 Oct;45(8):441-7.
Pottier J, Cousty C, Heitzman RJ, Reynolds IP. Differences in the biotransformation of a 17 beta-hydroxylated steroid, trenbolone acetate, in rat and cow. Xenobiotica. 1981 Jul;11(7):489-500.
Wilson VS, Lambright C, Ostby J, Gray LE Jr. In vitro and in vivo effects of 17beta-trenbolone: a feedlot effluent contaminant. Toxicol Sci. 2002 Dec;70(2):202-11.
Ashby J, Lefevre PA, Tinwell H, Odum J, Owens W. Testosterone-stimulated weanlings as an alternative to castrated male rats in the Hershberger anti-androgen assay. Regul Toxicol Pharmacol. 2004 Apr;39(2):229-38.
Freyberger A, Hartmann E, Krötlinger F. Evaluation of the rodent Hershberger bioassay using three reference (anti)androgens. Arh Hig Rada Toksikol. 2005 Jun;56(2):131-9.
Freyberger A, Ellinger-Ziegelbauer H, Krötlinger F. Evaluation of the rodent Hershberger bioassay: testing of coded chemicals and supplementary molecular-biological and biochemical investigations. Toxicology. 2007 Sep 24;239(1-2):77-88.
Owens W, Gray LE, Zeiger E, Walker M, Yamasaki K, Ashby J, Jacob E. The OECD program to validate the rat Hershberger bioassay to screen compounds for in vivo androgen and antiandrogen responses: phase 2 dose-response studies. Environ Health Perspect. 2007 May;115(5):671-8.
Moon HJ, Kang TS, Kim TS, Kang IH, Ki HY, Kim SH, Han SY. OECD validation of phase 3 Hershberger assay in Korea using surgically castrated male rats with coded chemicals. J Appl Toxicol. 2009 May;29(4):350-5.
Wilson JD. The role of 5alpha-reduction in steroid hormone physiology. Reprod Fertil Dev. 2001;13(7-8):673-8. Review.
Borst SE, Lee JH, Conover CF. Inhibition of 5alpha-reductase blocks prostate effects of testosterone without blocking anabolic effects. Am J Physiol Endocrinol Metab. 2005 Jan;288(1):E222-7.
Vargas RA, Oliveira LP, Frankenfeld S, Souza DB, Costa WS, Favorito LA, Sampaio FJ. The prostate after administration of anabolic androgenic steroids: a morphometrical study in rats. Int Braz J Urol. 2013 Sep-Oct;39(5):675-82.
Donaldson IA, Hart IC, Heitzman RJ. Growth hormone, insulin, prolactin and total thyroxine in the plasma of sheep implanted with the anabolic steroid trenbolone acetate alone or with oestradiol. Res Vet Sci. 1981 Jan;30(1):7-13.
Quinn MJ Jr, Lavoie ET, Ottinger MA. Reproductive toxicity of trenbolone acetate in embryonically exposed Japanese quail. Chemosphere. 2007 Jan;66(7):1191-6.
Kuhl H, Wiegratz I. Can 19-nortestosterone derivatives be aromatized in the liver of adult humans? Are there clinical implications? Climacteric. 2007 Aug;10(4):344-53. Review.
Attardi BJ, Pham TC, Radler LC, Burgenson J, Hild SA, Reel JR. Dimethandrolone (7alpha,11beta-dimethyl-19-nortestosterone) and 11beta-methyl-19-nortestosterone are not converted to aromatic A-ring products in the presence of recombinant human aromatase. J Steroid Biochem Mol Biol. 2008 Jun;110(3-5):214-22.
Le Guevel R, Pakdel F. Assessment of oestrogenic potency of chemicals used as growth promoter by in-vitro methods. Hum Reprod. 2001 May;16(5):1030-6.
Hemmer MJ, Cripe GM, Hemmer BL, Goodman LR, Salinas KA, Fournie JW, Walker CC. Comparison of estrogen-responsive plasma protein biomarkers and reproductive endpoints in sheepshead minnows exposed to 17beta-trenbolone. Aquat Toxicol. 2008 Jun 23;88(2):128-36.
Neumann F. Pharmacological and endocrinological studies on anabolic agents. Environ Qual Saf Suppl. 1976;(5):253-64. Review.
Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci. 1991 Jun;69(6):2452-62.
Ankley GT, Jensen KM, Makynen EA, Kahl MD, Korte JJ, Hornung MW, Henry TR, Denny JS, Leino RL, Wilson VS, Cardon MC, Hartig PC, Gray LE. Effects of the androgenic growth promoter 17-beta-trenbolone on fecundity and reproductive endocrinology of the fathead minnow. Environ Toxicol Chem. 2003 Jun;22(6):1350-60.
Henricks DM, Edwards RL, Champe KA, Gettys TW, Skelley GC Jr, Gimenez T. Trenbolone, estradiol-17 beta and estrone levels in plasma and tissues and live weight gains of heifers implanted with trenbolone acetate. J Anim Sci. 1982 Nov;55(5):1048-56.
Henricks DM, Brandt RT Jr, Titgemeyer EC, Milton CT. Serum concentrations of trenbolone-17 beta and estradiol-17 beta and performance of heifers treated with trenbolone acetate, melengestrol acetate, or estradiol-17 beta. J Anim Sci. 1997 Oct;75(10):2627-33.
Zhang X, Hecker M, Park JW, Tompsett AR, Jones PD, Newsted J, Au DW, Kong R, Wu RS, Giesy JP. Time-dependent transcriptional profiles of genes of the hypothalamic-pituitary-gonadal axis in medaka (Oryzias latipes) exposed to fadrozole and 17beta-trenbolone. Environ Toxicol Chem. 2008 Dec;27(12):2504-11.
Reiter M, Walf VM, Christians A, Pfaffl MW, Meyer HH. Modification of mRNA expression after treatment with anabolic agents and the usefulness for gene expression-biomarkers. Anal Chim Acta. 2007 Mar 14;586(1-2):73-81.
Seki M, Fujishima S, Nozaka T, Maeda M, Kobayashi K. Comparison of response to 17 beta-estradiol and 17 beta-trenbolone among three small fish species. Environ Toxicol Chem. 2006 Oct;25(10):2742-52.
Jensen KM, Ankley GT. Evaluation of a commercial kit for measuring vitellogenin in the fathead minnow (Pimephales promelas). Ecotoxicol Environ Saf. 2006 Jun;64(2):101-5. Epub 2006 Apr 17.
Holbech H, Kinnberg K, Petersen GI, Jackson P, Hylland K, Norrgren L, Bjerregaard P. Detection of endocrine disrupters: evaluation of a Fish Sexual Development Test (FSDT). Comp Biochem Physiol C Toxicol Pharmacol. 2006 Sep;144(1):57-66.
Orn S, Yamani S, Norrgren L. Comparison of vitellogenin induction, sex ratio, and gonad morphology between zebrafish and Japanese medaka after exposure to 17alpha-ethinylestradiol and 17beta-trenbolone. Arch Environ Contam Toxicol. 2006 Aug;51(2):237-43.
Park JW, Tompsett A, Zhang X, Newsted JL, Jones PD, Au D, Kong R, Wu RS, Giesy JP, Hecker M. Fluorescence in situ hybridization techniques (FISH) to detect changes in CYP19a gene expression of Japanese medaka (Oryzias latipes). Toxicol Appl Pharmacol. 2008 Oct 15;232(2):226-35.
Zhang X, Hecker M, Park JW, Tompsett AR, Newsted J, Nakayama K, Jones PD, Au D, Kong R, Wu RS, Giesy JP. Real-time PCR array to study effects of chemicals on the Hypothalamic-Pituitary-Gonadal axis of the Japanese medaka. Aquat Toxicol. 2008 Jul 7;88(3):173-82.
Holland AM, Roberts MD, Mumford PW, Mobley CB, Kephart WC, Conover CF, Beggs LA, Balaez A, Otzel DM, Yarrow JF, Borst SE, Beck DT. Testosterone inhibits expression of lipogenic genes in visceral fat by an estrogen-dependent mechanism. J Appl Physiol (1985). 2016 Sep 1;121(3):792-805.
H. D. MEYER and M. RAPP Reversible binding of the anabolic steroid trenbolone to steroid receptors Acta Endocrinol 110 S129-S130
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Heitzman RJ, Donaldson IA, Hart IC. Effect of anabolic steroids on plasma thyroid hormones in steers and heifers. Br Vet J. 1980 Mar-Apr;136(2):168-74.
Mader TL, Kreikemeier WM. Effects of growth-promoting agents and season on blood metabolites and body temperature in heifers. J Anim Sci. 2006 Apr;84(4):1030-7.
Hunter RA, Vercoe JE. Reduction of energy requirements of steers fed on low-quality-roughage diets using trenbolone acetate. Br J Nutr. 1987 Nov;58(3):477-83.
Field RA. Effect of castration on meat quality and quantity. J Anim Sci. 1971 May;32(5):849-58.
Leenen R, van der Kooy K, Droop A, Seidell JC, Deurenberg P, Weststrate JA, Hautvast JG. Visceral fat loss measured by magnetic resonance imaging in relation to changes in serum lipid levels of obese men and women. Arterioscler Thromb. 1993 Apr;13(4):487-94.
Leenen R, van der Kooy K, Meyboom S, Seidell JC, Deurenberg P, Weststrate JA. Relative effects of weight loss and dietary fat modification on serum lipid levels in the dietary treatment of obesity. J Lipid Res. 1993 Dec;34(12):2183-91.
Donner DG, Elliott GE, Beck BR, Bulmer AC, Lam AK, Headrick JP, Du Toit EF. Trenbolone Improves Cardiometabolic Risk Factors and Myocardial Tolerance to Ischemia-Reperfusion in Male Rats With Testosterone-Deficient Metabolic Syndrome. Endocrinology. 2016 Jan;157(1):368-81.
Rebuffé-Scrive M, Mårin P, Björntorp P. Effect of testosterone on abdominal adipose tissue in men. Int J Obes. 1991 Nov;15(11):791-5.
Langfort J, Jagsz S, Dobrzyn P, Brzezinska Z, Klapcinska B, Galbo H, Gorski J. Testosterone affects hormone-sensitive lipase (HSL) activity and lipid metabolism in the left ventricle. Biochem Biophys Res Commun. 2010 Sep 3;399(4):670-6.
Goldbourt U, Yaari S, Medalie JH. Isolated low HDL cholesterol as a risk factor for coronary heart disease mortality. A 21-year follow-up of 8000 men. Arterioscler Thromb Vasc Biol. 1997 Jan;17(1):107-13.
Cholestasis induced by Parabolan successfully treated with the molecular adsorbent recirculating system. Anand JS et al. ASAIO 2006. JanFeb;52(1):117-8.
Katsuki A, Sumida Y, Urakawa H, Gabazza EC, Murashima S, Maruyama N, Morioka K, Nakatani K, Yano Y, Adachi Y. Increased visceral fat and serum levels of triglyceride are associated with insulin resistance in Japanese metabolically obese, normal weight subjects with normal glucose tolerance. Diabetes Care. 2003 Aug;26(8):2341-4.
Bruun JM, Verdich C, Toubro S, Astrup A, Richelsen B. Association between measures of insulin sensitivity and circulating levels of interleukin-8, interleukin-6 and tumor necrosis factor-alpha. Effect of weight loss in obese men. Eur J Endocrinol. 2003 May;148(5):535-42.
Kapoor D, Malkin CJ, Channer KS, Jones TH. Androgens, insulin resistance and vascular disease in men. Clin Endocrinol (Oxf). 2005 Sep;63(3):239-50. Review.
Motoshima H, Wu X, Sinha MK, Hardy VE, Rosato EL, Barbot DJ, Rosato FE, Goldstein BJ. Differential regulation of adiponectin secretion from cultured human omental and subcutaneous adipocytes: effects of insulin and rosiglitazone. J Clin Endocrinol Metab. 2002 Dec;87(12):5662-7.
Swarbrick MM, Havel PJ. Physiological, pharmacological, and nutritional regulation of circulating adiponectin concentrations in humans. Metab Syndr Relat Disord. 2008 Jun;6(2):87-102.
Gavrila A, Chan JL, Yiannakouris N, Kontogianni M, Miller LC, Orlova C, Mantzoros CS. Serum adiponectin levels are inversely associated with overall and central fat distribution but are not directly regulated by acute fasting or leptin administration in humans: cross-sectional and interventional studies. J Clin Endocrinol Metab. 2003 Oct;88(10):4823-31.
Yarrow JF, Beggs LA, Conover CF, McCoy SC, Beck DT, Borst SE. Influence of Androgens on Circulating Adiponectin in Male and Female Rodents. Lobaccaro J-MA, ed. PLoS ONE. 2012;7(10):e47315.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7.
Coviello AD, Kaplan B, Lakshman KM, Chen T, Singh AB, Bhasin S. Effects of graded doses of testosterone on erythropoiesis in healthy young and older men. J Clin Endocrinol Metab. 2008 Mar;93(3):914-9.
Shahani S, Braga-Basaria M, Maggio M, Basaria S. Androgens and erythropoiesis: past and present. J Endocrinol Invest. 2009 Sep;32(8):704-16.
Sakhri S, Gooren LJ. Safety aspects of androgen treatment with 5alpha-dihydrotestosterone. Andrologia. 2007 Dec;39(6):216-22. Review.
Rochira V, Zirilli L, Madeo B, Maffei L, Carani C. Testosterone action on erythropoiesis does not require its aromatization to estrogen: Insights from the testosterone and estrogen treatment of two aromatase-deficient men. J Steroid Biochem Mol Biol. 2009 Feb;113(3-5):189-94.
Amory JK, Watts NB, Easley KA, Sutton PR, Anawalt BD, Matsumoto AM, Bremner WJ, Tenover JL. Exogenous testosterone or testosterone with finasteride increases bone mineral density in older men with low serum testosterone. J Clin Endocrinol Metab. 2004 Feb;89(2):503-10.
McCoy SC, Yarrow JF, Conover CF, Borsa PA, Tillman MD, Conrad BP, Pingel JE, Wronski TJ, Johnson SE, Kristinsson HG, Ye F, Borst SE. 17β-Hydroxyestra-4,9,11-trien-3-one (Trenbolone) preserves bone mineral density in skeletally mature orchiectomized rats without prostate enlargement. Bone. 2012 Oct;51(4):667-73.
Se si assume una dose di Terbutalina poco più alta della normale dose terapeutica utilizzata dai soggetti asmatici, è potenzialmente possibile riscontrare un aumento della massa muscolare pari ad 1KG in un mese. Questo, ovviamente, può accadere se ci si allena contro resistenza in modo serio, ma anche quando si svolge sufficiente attività fisica. Gli scienziati dello sport dell’Università di Copenaghen sono arrivati a questa conclusione in seguito ad uno studio svolto su esseri uomani, pubblicato sullo Scandinavian Journal of Medicine & Science in Sports. (1)
La Terbutalina, come il Clenbuterolo e il Salbutamolo, è un beta-2 agonista. I beta-2 agonisti, come suggerisce il nome, si legano selettivamente al recettore beta-2 adrenergico. Quando questo recettore viene attivato, la muscolatura liscia si rilassa e le vie respiratorie si dilatano.
Molti beta-2 agonisti presentano anche un effetto anti-catabolico, il quale (teoricamente) permette agli utilizzatori di aumentare o conservare la massa muscolare (effetto solitamente limitato dalla dose necessaria richiesta e da un aumento del Cortisolo). Come ben sappiamo, gli atleti supplementari farmacologicamente spesso usano i beta-2 agonisti per agevolare la riduzione del grasso corporeo.
Se gli atleti hanno l’asma, è consentito loro usare la Terbutalina. Da quanto si sa, le dosi terapeutiche di Terbutalina non hanno alcun effetto sulla massa muscolare. Ma i ricercatori danesi si sono chiesti cosa succederebbe se gli atleti utilizzassero una dose più elevata di Terbutalina di quanto sia normalmente necessario per il trattamento dell’asma.
I ricercatori, che tra l’altro erano pagati dal governo danese e dalla WADA, hanno reclutato per l’esperimento 66 uomini sani e attivi tra i 18 ed i 36 anni. I soggetti si allenavano per 2-5 ore a settimana. Alcuni giocavano a calcio, altri praticavano jogging o andavano in bici tutti i giorni da e verso il lavoro. I ricercatori hanno diviso i soggetti in 3 gruppi.
Ai soggetti del primo gruppo non sono state applicate modifiche alla loro routine di attività fisica [Habitual].
I soggetti del secondo gruppo sono stati sottoposti ad allenamenti di resistenza. Per 4 settimane, 3 volte a settimana, i soggetti eseguivano un allenamento a intervalli, che consisteva in 3 sessioni da 10 minuti di Cyclette con un’intensità dell’85% del VO2max. Ogni sessione si concludeva con uno sprint finale di 30 secondi. Dopo l’allenamento, i soggetti consumavano uno shake contenente 30g di Whey.
I soggetti del terzo gruppo sono stati sottoposti ad allenamenti contro resistenza 3 volte a settimana. Ogni seduta allenante consisteva il una “Full Body”, in cui i soggetti allenavano i loro principali gruppi muscolari con esercizi di base come leg-press, bench-press, extensions, military-press, lunges, lat-pulldowns, leg-curl e low-row. I soggetti hanno eseguito serie da 12 ripetizioni riposandosi per 2 minuti tra le serie. Dopo l’allenamento, i soggetti consumavano uno shake contenente 30g di Whey.
In ogni gruppo, metà dei soggetti presi in esame assumeva la Terbutalina. Il farmaco è stato somministrato per via inalatoria. I soggetti, per l’esattezza, hanno usato il Bricanyl Turbohaler della AstraZeneca. Ogni giorno i soggetti trattati inalavano 8 erogazioni da 0,5mg di Terbutalina (4mg totali al giorno).
Nei soggetti del primo gruppo e nei soggetti che si allenavano contro resistenza, la somministrazione di Terbutalina a causato un aumento della massa corporea magra di poco più di un chilo. Questo non si è verificato nei soggetti del gruppo sottoposto ad allenamenti di resistenza.
La Terbutalina, hai dosaggi utilizzati nello studio, non ha avuto effetti sulla massa grassa.
I ricercatori scrivono che, il presente studio mostra come l’inalazione quotidiana di un beta2-agonista comunemente somministrato aumenta la massa magra in individui che non sono di per sé allenanti, ma che piuttosto mantengono un basso livello di attività fisica, e che sperimentano un aumentano additivo della massa magra se combinata con esercizi contro resistenza.
I ricercatori proseguono scrivendo che, si tratta di una problematica per l’antidoping e stabilire una soglia delle concentrazioni di Terbutalina nelle urine dovrebbe essere una priorità per la WADA, al fine di evitare un eccessivo uso improprio al fine di un aumento dell’ipertrofia muscolare da parte degli atleti che hanno accesso alla Terbutalina.
Concludendo, i ricercatori affermano che, i dati del presente studio dovrebbero suggerire cautela ai medici che trattano gli atleti d’élite come inalazioni di Terbutalina in dosi di 4 mg/die le quali, come riportato sopra, possono potenziare gli adattamenti dell’allenamento contro resistenza.
Nel 2015, gli stessi ricercatori danesi hanno pubblicato uno studio nel quale i volontari hanno assunto un dosaggio consistente di Terbutalina in compresse. Gli effetti di ricomposizione corporea sono stati decisamente migliori rispetto a quelli legati al precedente studio qui esposto. (2)
I ricercatori hanno reclutato per lo studio 18 maschi attivi di vent’anni. I soggetti praticavano ciclismo, corsa o fitness e si allenavano per 4-8 ore a settimana. Durante l’esperimento i soggetti reclutati hanno continuato a svolgere le loro attività motorie abituali.
I ricercatori hanno diviso i soggetti in 2 gruppi. Per 4 settimane un gruppo ha ricevuto un placebo e l’altro la Terbutalina.
La dose terapeutica massima di Terbutalina per gli adulti è di 15mg/die, suddivisa in tre assunzioni. I ricercatori hanno somministrato ai soggetti del test una dose pari a 2-3 volte il dosaggio terapeutico massimo: per ogni 30 kg di peso corporeo i soggetti assumevano 5mg di Terbutalina Solfato ogni giorno, due volte al giorno. I ricercatori hanno utilizzato il Bricanyl Retard della AstraZeneca.
La forza isometrica [Massima contrazione volontaria] che i soggetti potevano esprimere durante l’esecuzione alla leg extension è rimasta costante nel gruppo placebo, mentre ha subito un aumento di 97 Newton nel gruppo Terbutalina.
Durante uno sprint su Cyclette di 30 secondi, i soggetti del gruppo Terbutalina [TER] hanno potuto sviluppare una potenza maggiore dopo 4 settimane. Ciò significa che sono diventati più veloci. Questo non è accaduto nel gruppo placebo [PLA].
La massa corporea magra nel gruppo trattato con Terbutalina era aumentata di 2Kg rispetto al gruppo placebo. Allo stesso tempo, la massa grassa nel gruppo trattato con Terbutalina era diminuita di 1kg rispetto al gruppo placebo.
Nelle cellule muscolari, prelevate dalle gambe dei soggetti dello studio, i ricercatori hanno scoperto un aumento delle proteine muscolari della catena pesante della miosina I e della catena pesante della miosina II nel gruppo trattato con Terbutalina.
Nelle cellule muscolari, la Terbutalina non ha avuto alcun effetto sul espressione della Miostatina, famoso peptide deputato alla regolazione dell’ipertrofia muscolare. Tuttavia, il farmaco ha aumentato la produzione di Follistatina. E, come ormai ben sappiamo, la Follistatina esercita un azione inibitoria nei confronti della Miostatina.
Ovviamente, la dose di Terbutalina utilizzata non è priva di effetti collaterali. Nella prima settimana dello studio, alcuni soggetti hanno sperimentato tremori, palpitazioni e irrequietezza. Dopo la prima settimana, gli effetti collaterali sono scomparsi.
I ricercatori si sono premurati di concludere dicendo che, dato l’impatto sul miglioramento delle prestazioni in seguito ad uso acuto e cronico di beta-2-adrenergici sulla massa muscolare, la forza e la potenza durante l’esercizio massimale, sembra logico che l’uso sistemico di beta-2-agonisti debba rimanere nell’elenco delle sostanze proibite negli sport competitivi.
Alcuni di voi potrebbero essersi già imbattuti nell’articolo di Chest Rockwell intitolato “The Science of Trenbolone” il quale si trova da qualche tempo su alcuni siti dedicati al BodyBuilding. L’intento dell’autore è stato quello di creare un articolo approfondito sul Trenbolone. Tale articolo è stato progettato per essere una guida di riferimento rapida volta a rispondere a molte domande riguardanti questo AAS. La mole di informazioni presenti in esso ha reso questo articolo un punto di riferimento per comprendere le caratteristiche e le potenziali applicazioni del Trenbolone. Il mio intento è quello di realizzare una “guida” sul Trenbolone, sul modello di quella realizzata da Chest Rockwell, in grado di fornire una serie di importanti informazioni riguardanti questo AAS e utilizzabili da atleti e preparatori.
II. Introduzione
Il Trenbolone è senza dubbio l’AAS con una reputazione quasi mitica all’interno del mondo del BodyBuilding. Poiché i dati sugli esseri umani sono molto limitati, spesso dobbiamo fare affidamento sugli aneddoti nel tentativo di formulare ipotesi su di esso. Come si può leggere in quasi tutti i forum dedicati al BodyBuilding, le esperienze con il Trenbolone variano ampiamente – con alcuni utenti che adorano assolutamente il composto mentre altri ne consigliano estrema cautela nell’uso o dicono agli altri utenti di evitarlo a tutti i costi. Nonostante questa grande divergenza di opinioni, non si può contestare la sua popolarità poiché numerosi studi hanno dimostrato negli anni che tale molecola è uno dei composti anabolizzanti più frequentemente usati, con una percentuale compresa tra il 20 e il 25% dei culturisti supplementari chimicamente che hanno riferito di averlo usato nell’arco degli ultimi dodici mesi. (1)(2)(3)
Il mio obiettivo con questo articolo sarà quello di utilizzare quante più informazioni disponibili per cercare di formulare alcune solide conclusioni riguardo al funzionamento e al potenziale utilizzo del composto. Allo stesso tempo spero di contribuire a dissipare alcuni miti che vengono ancora propagandati troppo spesso.
Come precedentemente accennato, esistono solo un paio di studi sull’uomo di cui sono a conoscenza, quindi la maggior parte del materiale citato proverrà da studi svolti su animali o in vitro. La domanda che dovrebbe essere posta è la seguente: possiamo prendere questi dati e applicarli concretamente sui BodyBuilder? Personalmente, ritengo che ci sono dei dati molto concreti e universalmente applicabili sugli esseri umani e poi ce ne sono altri che potrebbero richiedere un disclaimer. Cercherò di fare del mio meglio per indicarli nel corso di questo articolo.
III. Nozioni di base sul Trenbolone
Trenbolone
Il Trenbolone è considerabile quale Modulatore Selettivo del Recettore degli Androgeni (SARM), non progettato per l’uso umano (4), anche se venne commercializzato come farmaco da prescrizione per uso umano dalla Negma Laboratoires in Francia sotto il nome di Parabolan (Trenbolone Hexahydrobenzylcarbonato). Nonostante la sua primaria designazione, questo AAS continua ad essere pesantemente utilizzato dai Bodybuilder per promuovere la crescita muscolare, la riduzione del grasso e, di conseguenza, migliorare la composizione corporea.(5)(6) Quando sentiamo parlare di SARM colleghiamo tale termine ad una classe di agenti anabolizzanti non steroidei (vedi Ostarina, Andarina, LGD4033, ecc…). Però, tale definizione “particolaristica” non è del tutto corretta. Infatti, tutti gli AAS aventi un indice terapeutico (dato dalla Anabolico:Androgeno ratio) superiore a “1” sono considerabili quali SARM (l’indice terapeutico del Trenbolone è di circa 3,4). Si sa perfettamente che i SARM sono analoghi modificati degli ormoni sessuali maschili che normalmente esibiscono attività anabolica favorevole mentre, contemporaneamente, presentano una attività androgenica da moderata a minima in vivo rispetto agli androgeni endogeni.(7)(8)(9) Sono composti in fase di sviluppo da parte di molte aziende farmaceutiche nel tentativo di creare mezzi alternativi per il trattamento di condizioni come l’ipogonadismo e gli stati di deperimento muscolare e osseo. In sostanza, l’obiettivo è quello di ricreare gli aspetti positivi dati da dosi sovrafisiologiche di testosterone eliminando al contempo il rischio di eventi avversi che tendono a verificarsi quando si utilizzano tali dosaggi.(10) Questo obbiettivo lo si ritrova, appunto, concretizzato in tutti gli AAS che presentano modifiche dello scheletro carbossilico tali da permettere tale effetto.
La maggior parte dei SARM steroidei hanno come base di partenza una molecola di Testosterone. La struttura chimica della molecola di Testosterone viene quindi tradizionalmente modificata in uno dei seguenti tre modi (11)(12):
Esterificazione del gruppo 17β-idrossile che aumenta l’idrofobicità o la probabilità che la molecola possa essere respinta da una massa d’acqua;
Alchilazione in posizione 7α la quale riduce l’affinità di legame con l’enzima 5α-reduttasi;
Modifica strategica dei legami di carbonio in C1, C2, C9, C11 o C19 per ottenere una vasta gamma di effetti terapeutici.
Il Trenbolone è un C19-norsteroide (19-nor), derivato dal Nandrolone (Nortestosterone). La rimozione del gruppo metilico nella posizione C19 dello scheletro carbossilico steroideo riduce significativamente la suscettibilità dei 19-norsteroidi all’azione dell’enzima aromatasi e a quella dell’enzima 5α-reduttasi.(4) Entreremo più nel dettaglio dei meccanismi sopracitati in seguito ma, per ora, è necessario che si comprenda semplicemente come le sottili modifiche allo scheletro carbossilico della molecola di Testosterone possano tradursi direttamente in cambiamenti significativi del comportamento della nuova molecola derivata. Alcuni di questi cambiamenti possono includere l’affinità di legame del composto per il recettore degli androgeni e la sua affinità di legame con numerosi enzimi in grado di convertirlo in altri steroidi.(13)
Il Trenbolone differisce dal suo precursore (Nandrolone) per:
1- Il doppio legame inserito in C9– C10, che inibisce totalmente l’aromatizzazione e aumenta la resistenza al passaggio epatico;
2- l’insaturazione in C11-C12 che aumenta l’affinità per il recettore androgeno, rendendo il Trenbolone uno degli anabolizzante con la più forte affinità AR. (14)
Il valore Anabolico/Androgeno del Trenbolone rispetto al Testosterone (100/100) è pari a 625:185.
Quindi, il Trenbolone ha proprietà SARM-simili in quanto presenta una affinità significativamente minore ai percorsi metabolici ai quali è soggetto il Testosterone. Ma di questo ne parleremo in seguito.
IV. Storia del Trenbolone
Metribolone
L’enorme potenziale anabolizzante del Trenbolone, così come dei suoi analoghi, è stato riportato fin dagli anni ’60. Come molti di voi già sapranno, venne sintetizzata anche una versione orale denominata Metribolone (conosciuta anche come Methyltrienolone o Methyltrenbolone), tuttavia non è mai stata commercializzata come agente anabolizzante a causa della sua tossicità epatica estrema – causando colestasi intraepatica a quantità somministrate per via orale pari a 1 mg/giorno.(15)
A parte la parentesi francese del Parabolan (Trenbolone Hexahydrobenzylcarbonato), commercializzato dalla Negma fino al 1997, il Trenbolone non è mai stato approvato per l’uso umano e il suo utilizzo è stato (ed è) principalmente quello di agente per la promozione della crescita nel bestiame.(16)(17) Per tale scopo viene usato sia singolarmente che in combinazione con Estradiolo (E2).(18) L’uso di impianti per il bestiame contenenti la combinazione di Trenbolone ed Estradiolo è stata approvata dalla FDA nel 1992 (19), e ora circa il 90% dei bovini da carne negli Stati Uniti viene trattato con un mix di estrogeni, androgeni e/o progestinici volto a promuoverne la crescita.(20) Gli impianti per il bestiame sono un grande business con fino a 20 milioni di bovini all’anno impiantati con Trenbolone e un reddito annuo probabilmente superiore a un miliardo di dollari.(21)
Trenbolone Acetato
Nonostante l’approvazione da parte della FDA, sussistono ancora problemi di sicurezza poiché il Trenbolone Acetato (TBA), estere utilizzato per trattare il bestiame, ei suoi metaboliti sono stati identificati come potenziali sostanze chimiche dannose per il sistema endocrino (EDC). Gli EDC sono molecole esogene che possono imitare o inibire l’azione dei recettori ormonali come i recettori degli estrogeni, degli androgeni e degli ormoni tiroidei. Questi EDC possono anche alterare la sintesi, l’azione, il metabolismo e la secrezione di ormoni endogeni la quale può portare a problemi gravi come obesità, diabete e persino il cancro. (22)(23)
Meccanismi d’azione degli EDC. A: Interazione diretta del EDC con un recettore nucleare dell’ormone che porta alla stimolazione (agonismo) o inibizione (antagonismo) della sua attività trascrizionale. B: Stimolazione o inibizione della biosintesi degli ormoni endogeni. C: Stimolazione o inibizione della degradazione degli ormoni endogeni. D: Stimolazione o inibizione della proteina legante gli ormoni endogeni che porta ad un aumento o diminuzione della disponibilità degli ormoni circolanti.
A causa della potenziale gravità degli EDC, negli ultimi due decenni si è registrata una maggiore attenzione internazionale sull’esposizione ambientale e sugli effetti degli EDC nell’uomo e nella fauna selvatica.(24)(25) Come precedentemente accennato, il TBA ei suoi metaboliti sono stati identificati come EDC attraverso numerosi studi, possono essere diffusi in ambienti agricoli e sono associati a tossicità riproduttiva. (26)(27)(28) Il problema è che è necessaria soltanto un’esposizione a concentrazioni molto basse per causare potenziali problemi, come è stato dimostrato negli animali come i pesci, i quali hanno mostrato disordini nel comportamento sessuale e una diminuzione della fertilità. (29)
Sarà anche importante essere in grado di distinguere i vari tipi di impianti contenenti TBA, dal momento che molti degli studi che esamineremo in seguito utilizzeranno tipi diversi sugli animali presi in esame. Quello che segue è un elenco dei tipi di impianti comuni usati negli Stati Uniti, con specificazione delle loro concentrazioni ormonali:
Revalor-XS (200mg TBA / 40mg E2)
Revalor-200 (200mg TBA / 20mg E2)
Revalor-H (140mg TBA / 14mg E2)
Revalor-S (120mg TBA / 24mg E2)
Revalor-IS (80mg TBA / 16mg E2)
Revalor-IH (80mg TBA / 8mg E2)
Revalor-G (40mg TBA / 8mg E2)
Synovex PLUS (200mg TBA / 28mg E2)
Synovex-C (100mg Progesterone / 10mg E2)
Synovex-ONE Grass (150mg TBA / 15mg E2)
Synovex-S (200mg Progesterone / 20mg E2)
Synovex-H (200mg Testosterone / 20mg E2)
V. Metabolismo e fisiologia
Precedentemente ho fatto breve menzione ai metaboliti del TBA, quindi, ora, per completezza è giusto entrare nei dettagli. E’ corretto rammentare che la maggior parte delle informazioni riguardanti il metabolismo del Trenbolone in vivo provengono da osservazioni su bestiame e roditori (30)(31)(32). È fondamentale anche comprendere che vi sono differenze marcate nella quantità di vari metaboliti osservati nei modelli di ratti e mucche, i due mammiferi più intensamente studiati. (33) Torneremo a trattare questo argomento dopo aver esaminato alcuni dei principi fondamentali.
Trendione
Il nome chimico del TBA è 17ß-idrossi-estra-4,9,11-trien-3-one-17-acetato, talvolta abbreviato in 17β-TBOH-acetato. Dopo un’iniezione intramuscolare, viene rapidamente idrolizzato nel metabolita biologicamente attivo noto come 17β-idrossi-estra-4,9,11-trien-3-one o 17β-TBOH.(34) Da lì viene ulteriormente convertita in metaboliti inclusi i glucuronidi (per esempio Trendione / TBO) e altri cinque metaboliti idrossilati polari.(35) La serie di questi processi metabolici può essere riassunta come segue:
Il 17β-TBOH ha una maggiore affinità per gli AR rispetto a qualsiasi altro dei suoi metaboliti primari suggerendo che la biotrasformazione del Trenbolone riduce l’attività biologica dello steroide.(26)(27)(35) Per mettere questo in prospettiva, in uno studio l’elevata affinità del 17β-TBOH nei confronti del recettore degli androgeni umani e del recettore del progesterone bovino è stata ridotta dopo che la molecola è stata metabolizzata in 17α-TBOH e TBO a meno di 1/24 della dose originale del composto.(36) Questo comportamento è in netto contrasto con quello osservato nel Testosterone la cui conversione in DHT e in estrogeni porta a composti più potenti in relazione all’affinità del legame con il recettore bersaglio.(37)(38) Tuttavia, il comportamento del TBA è simile per natura ad altri composti 19-norsteroidei (come il Nandrolone), la cui affinità AR diminuisce una volta 5α-ridotta.(39)
Come accennato in precedenza, vi è una certa variazione nel metabolismo del 17-TBOH tra i mammiferi poiché i metaboliti primari sono il 17ß-idrossi-estra-4,9,11-trien-3-one e l’Estra-4,9,11-triene- 3,17-dione con, a loro volta, metaboliti 16α e 16ß-idrossilati nel ratto. Nella mucca questi metaboliti sono trascurabili e il 17α-TBOH è il prodotto principale insieme a piccole quantità di 16α e 16ß-idrossi-17α-TBOH (30)(31). Segue un grafico dettagliato che confronta le differenze tra gli animali:
Confronto dei metaboliti biliari del 17beta-trenbolone acetato nel ratto e nella mucca.
Fortunatamente per noi, esiste uno studio su esseri umani che ci può aiutare a chiarire come l’uomo metabolizzi il Trenbolone – almeno dopo che questo è stato ingerito per via orale. (35) Lo studio è stato progettato per indagare sugli effetti dell’ingestione di cibo contaminato e quindi il team di ricerca ha iniettato il 17β-TBOH in un pezzo di hamburger fritto da 5g, con una dose di 0,04 mg/kg di peso corporeo. Dopo un singolo consumo, il 63% della dose somministrata è stata escreta tramite l’urina entro 72 ore; a 24 ore il 50% della dose somministrata è stata osservata nei campioni di urina.
I risultati hanno anche rivelato che, negli esseri umani, il 17β-TBOH ingerito è principalmente escreto intatto come 17β-TBOH, come Epitrenbolone (17α-TBOH) o come Trendione (TBO) – con la maggior parte in forma 17α-TBOH. In questo senso, la biotrasformazione del 17ß-TBOH negli esseri umani assomiglia più da vicino a quella delle mucche che a quella dei roditori. Inoltre, nell’urina umana sono stati rilevati numerosi metaboliti polari di 17β-TBOH, sebbene in concentrazione molto inferiore rispetto a quei metaboliti precedentemente menzionati. (40)
Il 17β-TBOH presenta una bassa biodisponibilità orale dal momento che non presenta una metilazione in posizione 17α. I risultati di due saggi di Hershberger dimostrano che il Trenbolone è circa 80-100 volte meno efficace assunto per via orale rispetto a somministrazione tramite iniezione (comportamento legato appunto alla biodisponibilità).(26) Nonostante questo, il TBA e, ovviamente, 17β-TBOH hanno ancora dimostrato di alterare il sistema riproduttivo di umani, maiali, topi, ratti e altre specie di mammiferi a livelli di dosaggio relativamente bassi quando somministrati per via orale.
Nella prossima parte di questa serie di articoli, esploreremo l’impatto del Trenbolone su vari percorsi anabolici e marker della salute metabolica. Vedremo anche come il Trenbolone influenzi la produzione endogena di ormoni e inizieremo ad analizzare in maniera approfondita i suoi effetti su anabolismo e ipertrofia.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
The Science of Trenbolone – By Chest Rockwell
Perry PJ, Lund BC, Deninger MJ, Kutscher EC, Schneider J. Anabolic steroid use in weightlifters and bodybuilders: an internet survey of drug utilization. Clin J Sport Med. 2005 Sep;15(5):326-30.
Parkinson AB, Evans NA. Anabolic androgenic steroids: a survey of 500 users. Med Sci Sports Exerc. 2006 Apr;38(4):644-51.
Ip EJ, Barnett MJ, Tenerowicz MJ, Perry PJ. The Anabolic 500 survey: characteristics of male users versus nonusers of anabolic-androgenic steroids for strength training. Pharmacotherapy. 2011 Aug;31(8):757-66.
Donner DG, Beck BR, Bulmer AC, Lam AK, Du Toit EF. Improvements in body composition, cardiometabolic risk factors and insulin sensitivity with trenbolone in normogonadic rats. Steroids. 2015 Feb;94:60-9.
Daniels JM, van Westerloo DJ, de Hon OM, Frissen PH. [Rhabdomyolysis in a bodybuilder using steroids]. Ned Tijdschr Geneeskd. 2006 May 13;150(19):1077-80. Dutch.
Geraci MJ, Cole M, Davis P. New onset diabetes associated with bovine growth hormone and testosterone abuse in a young body builder. Hum Exp Toxicol. 2011 Dec;30(12):2007-12.
Omwancha J, Brown TR. Selective androgen receptor modulators: in pursuit of tissue-selective androgens. Curr Opin Investig Drugs. 2006 Oct;7(10):873-81. Review.
Gao W, Dalton JT. Expanding the therapeutic use of androgens via selective androgen receptor modulators (SARMs). Drug Discov Today. 2007 Mar;12(5-6):241-8. Review.
Thevis M, Schänzer W. Synthetic anabolic agents: steroids and nonsteroidal selective androgen receptor modulators. Handb Exp Pharmacol. 2010;(195):99-126.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7.
Kicman AT. Pharmacology of anabolic steroids. British Journal of Pharmacology. 2008;154(3):502-521.
Haendler B, Cleve A. Recent developments in antiandrogens and selective androgen receptor modulators. Mol Cell Endocrinol. 2012 Apr 16;352(1-2):79-91.
Fragkaki AG, Angelis YS, Koupparis M, Tsantili-Kakoulidou A, Kokotos G, Georgakopoulos C. Structural characteristics of anabolic androgenic steroids contributing to binding to the androgen receptor and to their anabolic and androgenic activities. Applied modifications in the steroidal structure. Steroids. 2009 Feb;74(2):172-97.
Krüskemper HL, Noell G. Liver toxicity of a new anabolic agent: methyltrienolone (17-alpha-methyl-4,9,11-estratriene-17 beta-ol-3-one). Steroids. 1966 Jul;8(1):13-24.
Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci. 1991 Jun;69(6):2452-62.
Chung KY, Johnson BJ. Application of cellular mechanisms to growth and development of food producing animals. J Anim Sci. 2008 Apr;86(14 Suppl):E226-35.Epub 2007 Oct 26. Review.
Metzler M, Pfeiffer E. Genotoxic potential of xenobiotic growth promoters and their metabolites. APMIS. 2001 Feb;109(2):89-95. Review.
Chung KY, Baxa TJ, Parr SL, Luqué LD, Johnson BJ. Administration of estradiol, trenbolone acetate, and trenbolone acetate/estradiol implants alters adipogenic and myogenic gene expression in bovine skeletal muscle. J Anim Sci. 2012 May;90(5):1421-7.
Balter M. Scientific cross-claims fly in continuing beef war. Science. 1999 May 28;284(5419):1453, 1455.
Lawrence JD, Ibarburu MA. Proceedings of the NCCC-134 Conference on Applied Commodity Price Analysis, Forecasting, and Market Risk Management; 16 and 17 April 2007; Chicago.
Diamanti-Kandarakis E, Bourguignon JP, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009 Jun;30(4):293-342. Review.
Rachoń D. Endocrine disrupting chemicals (EDCs) and female cancer: Informing the patients. Rev Endocr Metab Disord. 2015 Dec;16(4):359-64.
Birnbaum LS. State of the science of endocrine disruptors. Environ Health Perspect. 2013 Apr;121(4):A107.
Baynes RE, Dedonder K, Kissell L, Mzyk D, Marmulak T, Smith G, Tell L, Gehring R, Davis J, Riviere JE. Health concerns and management of select veterinary drug residues. Food Chem Toxicol. 2016 Feb;88:112-22.
Wilson VS, Lambright C, Ostby J, Gray LE Jr. In vitro and in vivo effects of 17beta-trenbolone: a feedlot effluent contaminant. Toxicol Sci. 2002 Dec;70(2):202-11.
Durhan EJ, Lambright CS, Makynen EA, Lazorchak J, Hartig PC, Wilson VS, Gray LE, Ankley GT. Identification of metabolites of trenbolone acetate in androgenic runoff from a beef feedlot. Environ Health Perspect. 2006 Apr;114 Suppl 1:65-8.
Gall HE, Sassman SA, Lee LS, Jafvert CT. Hormone discharges from a midwest tile-drained agroecosystem receiving animal wastes. Environ Sci Technol. 2011 Oct 15;45(20):8755-64.
Jensen KM, Makynen EA, Kahl MD, Ankley GT. Effects of the feedlot contaminant 17alpha-trenbolone on reproductive endocrinology of the fathead minnow. Environ Sci Technol. 2006 May 1;40(9):3112-7.
Pottier J, Busigny M, Grandadam JA. Plasma kinetics, excretion in mild and tissue levels in the cow following implantation of trenbolone acetate. J Anim Sci. 1975 Sep;41(3):962-8.
Pottier J, Cousty C, Heitzman RJ, Reynolds IP. Differences in the biotransformation of a 17 beta-hydroxylated steroid, trenbolone acetate, in rat and cow. Xenobiotica. 1981 Jul;11(7):489-500.
Evrard P, Maghuin-Rogister G, Rico AG. Fate and residues of trenbolone acetate in edible tissues from sheep amd calves implanted with tritium-labeled trenbolone acetate. J Anim Sci. 1989 Jun;67(6):1489-96
Metzler M. Metabolism of some anabolic agents: toxicological and analytical aspects. J Chromatogr. 1989 Apr 7;489(1):11-21.
Dorts J, Richter CA, Wright-Osment MK, Ellersieck MR, Carter BJ, Tillitt DE. The genomic transcriptional response of female fathead minnows (Pimephales promelas) to an acute exposure to the androgen, 17beta-trenbolone. Aquat Toxicol.2009 Jan 18;91(1):44-53.
Spranger B, Metzler M. Disposition of 17 beta-trenbolone in humans. J Chromatogr. 1991 Apr 5;564(2):485-92.
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Wilson JD. The role of 5alpha-reduction in steroid hormone physiology. Reprod Fertil Dev. 2001;13(7-8):673-8. Review.
Santen RJ, Brodie H, Simpson ER, Siiteri PK, Brodie A. History of aromatase: saga of an important biological mediator and therapeutic target. Endocr Rev. 2009 Jun;30(4):343-75.
Sundaram K, Kumar N, Monder C, Bardin CW. Different patterns of metabolism determine the relative anabolic activity of 19-norandrogens. J Steroid Biochem Mol Biol. 1995 Jun;53(1-6):253-7.
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
L’Acido Alfa-Lipoico può dare sollievo ai soggetti che soffrono della sindrome del tunnel carpale. Ricercatori messicani hanno osservato che una dose pari a 600mg di ALA si presta bene a tal fine.(1) Il loro studio è interessante anche per gli atleti supplementari chimicamente. Infatti, circa un terzo degli utilizzatori dell’ormone della crescita sviluppa la sindrome del tunnel carpale.
Gli atleti supplementati farmacologicamente e che usano alte dosi di GH hanno buone possibilità di sviluppare la sindrome del tunnel carpale. Questo accade perché il GH promuove, direttamente ed indirettamente, nella mano e nel polso, la crescita del tessuto osseo, dei muscoli, dei legamenti e del tessuto connettivo. Nel polso, come ben sappiamo, passa il nervo mediano attraverso il tunnel carpale (o canale carpale).
Quando il tunnel carpale si restringe e il nervo mediano viene compresso, si sviluppa formicolio, intorpidimento e spesso dolore. E questa condizione viene chiama sindrome del tunnel carpale.
Quando tale sindrome è causato dall’uso di GH esogeno, il rimedio è semplice: cessare la somministrazione del composto. Secondo quanto riportato nella letteratura medica (2), la sindrome del tunnel carpale causato dall’uso di GH esogeno tende a scomparire nel giro di poche settimane dopo la cessata somministrazione. Ma se la sindrome del tunnel carpale è causata da un’altro fattore- e ce ne sono molti – allora l’unica soluzione risulta essere l’intervento chirurgico.
Nel 2014, ricercatori italiani hanno pubblicato uno studio nel quale si riportava che un integrazione giornaliera composta da 300mg di Acido Alfa-Lipoico, 500mg di Curcumina Fitosoma e vitamine del gruppo B ha alleviato i sintomi della sindrome del tunnel carpale.(3) Gli autori dello studio hanno usavano l’Axin C, un prodotto della Farmacondo.
Anche i ricercatori messicani, affiliati all’Università di Guadalajara, hanno cercato di alleviare i sintomi legati alla sindrome del tunnel carpale con una supplementazione OTC, ma optando per un approccio più semplice. Hanno somministrato a 10 pazienti 600mg di Acido Alfa-Lipoico ogni giorno per 3 mesi. Hanno usato la versione più economica, vale a dire la miscela racemica composta dal 50% di enantiomeri R e dal 50% di enantiomeri S.
Altri nove pazienti hanno ricevuto un placebo.
Dopo un mese di integrazione, i pazienti hanno subito l’intervento chirurgico. Successivamente, hanno continuato a utilizzare la supplementazione di ALA per altri 2 mesi.
Poco prima dell’inizio della supplementazione [BASAL], dopo un mese di supplementazione [PRESURGERY] e un mese dopo la cessazione della supplementazione [FINAL], i medici hanno fatto compilare ai pazienti il Boston Questionnaire. Si tratta di un questionario con il quale i medici determinano la gravità della sindrome del tunnel carpale.
Nel mese precedente l’operazione, l’Acido Alfa-Lipoico ha ridotto la gravità dei sintomi. Tre mesi dopo l’intervento, il gruppo trattato con Acido Alfa-Lipoico presentava meno disturbi rispetto al gruppo placebo. Questo suggerisce che la supplementazione con ALA ha accelerato la guarigione del nervo mediano danneggiato.
Il gruppo trattato con Acido Alfa-Lipoico ha mostrato miglioramenti nei parametri sensoriali e motori, un’osservazione che potrebbe essere spiegata dai meccanismi multimodali dell’Acido Alfa-Lipoico legati alla sua azione antiossidante, agendo come “spazzino” dei radicali liberi e come rigeneratore delle vitamine C ed E, le quali contribuiscono alla riduzione dello stress ossidativo nel nervo periferico.
I risultati ottenuti suggeriscono anche che l’Acido Alfa-Lipoico può accelerare il recupero della funzione del nervo mediano, poiché il punteggio del Boston Questionnaire è migliorato più repentinamente nel gruppo trattato con Acido Alfa-Lipoico rispetto al gruppo placebo.
In conclusione, i ricercatori affermano di aver scoperto che l’Acido Alfa-Lipoico ha un effetto neuroprotettivo, somministrato per 1 mese prima della decompressione chirurgica e per 3 mesi dopo l’intervento.
Gli AAS possono inibire gli effetti estrogenici di sostegno sulla crescita e il mantenimento dello sviluppo dei tessuti mammari e possono causare, consequenzialmente, una riduzione visibile delle dimensioni del seno (atrofia mammaria). E’ giusto sottolineare che tale condizione non è ovviamente paragonabile alla riduzione nelle dimensioni del seno sperimentata dalle atlete “Natural”, soprattutto a causa della diminuzione della massa grassa. È stato dimostrato che l’uso di androgeni da parte di soggetti di sesso femminile causa una riduzione delle dimensioni del tessuto ghiandolare mammario e un aumento del tessuto connettivo fibroso.(1) Questi cambiamenti fisiologici sono simili a quelli osservati dopo la menopausa, quando i livelli estrogenici, in specie quelli dell’Estradiolo, sono marcatamente ridotti. La riduzione delle dimensioni del seno prodotta dall’uso di AAS può persistere in modo significativo anche dopo la loro cessata assunzione, poiché può esserci un sostanziale rimodellamento tissutale locale causato, appunto, da un eccessiva attività androgenica. Anche se il problema è tutto sommato facilmente ovviabile con un intervento di mastoplastica additiva, le utilizzatrici, o aspiranti tali, sono avvertite delle possibili e sostanziali modifiche nella fisiologia del seno conseguenti all’uso/abuso di AAS.
Gabriel Bellizzi
Riferimenti:
ANABOLICS 11th Edition [William Llewellyn]
Effects of long-term androgen administration on breast tissue of female-to-male transsexuals. M Slagter, L Gooren et al. J Histochem Cytochem. 54(8): 905-910, 2006.