Dan: Benvenuti a BioGenTech Sport and Health, dove oggi parliamo di quello che succede quando il tuo ciclo decide di regolare i conti con il tuo stomaco.
Denise: Gabriel Bellizzi, noto anche come Ružička, The Biochemist e CEO di BioGenTech, firma l’analisi di oggi: un territorio che copre microbiota intestinale, bruciore di stomaco da AAS e strategie concrete per gestire entrambi. Partiamo da lì.
AAS, intestino e reflusso: cause, rimedi e prevenzione
Dan: Il punto di partenza è questo: gli AAS non agiscono solo sui muscoli. Agiscono anche sull’intestino, sul microbiota e sulla mucosa gastrica, e capire perché è la condizione necessaria per gestire i danni.
Denise: Il post apre con una premessa fondamentale sul ruolo del microbiota intestinale nel metabolismo degli androgeni: “Il microbiota intestinale è costituito da trilioni di batteri, virus e funghi che sono stati descritti nel loro insieme come un organo endocrino virtuale in grado di produrre ed espellere numerose sostanze nel flusso sanguigno del suo ospite, influenzandone così la fisiologia.”
Dan: Quindi non stiamo parlando solo di digestione. Stiamo parlando di un sistema che modifica attivamente i livelli ormonali, e che viene a sua volta modificato dall’eccesso di androgeni esogeni.
Denise: Esattamente. Il post descrive questa rete come “testobolome”: la specifica comunità batterica che metabolizza gli androgeni intestinali. In condizioni di iperandrogenismo da AAS, questa rete va in sovraccarico. Si riduce l’alfa-diversità batterica, si selezionano ceppi patogeni, e si apre la strada a disbiosi, SIBO, permeabilità intestinale aumentata e sindrome dell’intestino irritabile.
Dan: E gli AAS orali peggiorano tutto questo in modo diretto, perché attraversano fisicamente il tratto gastrointestinale prima ancora di arrivare al fegato.
Denise: Il post lo specifica chiaramente: gli AAS orali alchilati in C-17 sono progettati per resistere al primo passaggio epatico, ma questo li rende aggressivi per le mucose. Il danno intestinale si collega poi a quello epatico tramite la colestasi, che riduce la bile disponibile nell’intestino e favorisce la steatorrea e la proliferazione batterica patogena.
Dan: E qui entra il bruciore di stomaco, che non è un fastidio secondario ma un meccanismo preciso.
Denise: Il meccanismo centrale riguarda i recettori H2 dell’istamina sulle cellule parietali gastriche. Gli androgeni disregolano questi recettori, aumentano la secrezione acida, riducono il flusso sanguigno alla mucosa e innalzano le citochine proinfiammatorie come IL-1β e TNF-α. Il post propone anche un’equazione sintetica: bruciore di stomaco uguale androgenicità meno estrogenicità.
Dan: Il che spiega perché Trenbolone e Fluoximesterone sono i peggiori, mentre il Testosterone classico, aromatizzando in estrogeni, attenua parzialmente il problema.
Denise: Per il sollievo immediato, il post indica gli antagonisti del recettore H2, in particolare la Famotidina, come gold standard. La ragione è strutturale: Famotidina e Ranitidina non presentano anelli imidazolici, quindi non si legano al citocromo P-450 e non interferiscono con il metabolismo degli steroidi, a differenza della Cimetidina.
Dan: Insomma, il Tums non basta.
Denise: Il post lo dice esplicitamente: il carbonato di calcio non è raccomandato nemmeno per una settimana continuativa. Per la prevenzione a lungo termine, le strategie includono la somministrazione notturna degli antagonisti H2, una dieta pre-allenamento a basso contenuto proteico e ricca di carboidrati, e l’evitare caffeina, cioccolato e cibi piccanti.
Denise: Sul versante del microbiota, il ripristino richiede un approccio multifasico: cambio molecolare verso androgeni attenuati come il Methenolone, supporto della barriera intestinale con L-Glutammina e colostro, probiotici ad alta sopravvivenza, enzimi digestivi e antiossidanti come glutatione liposomiale e EPA-DHA. Nei casi più gravi, il passaggio in TRT o il recupero dell’asse HPTA rimangono le uniche soluzioni strutturali.
Dan: La salute gastrointestinale non è un optional da gestire a sintomi conclamati, ma una variabile da presidiare dall’inizio del ciclo, prima che i danni si accumulino.
Denise: Il quadro che emerge è coerente: gli AAS modificano l’ambiente intestinale e gastrico attraverso meccanismi precisi, e le soluzioni esistono, ma richiedono continuità, non interventi reattivi.
Dan: La prossima volta che qualcuno dice che il bruciore di stomaco da ciclo è normale, ricordagli che normale non significa inevitabile, e che la Famotidina costa meno di un flacone di Trenbolone.
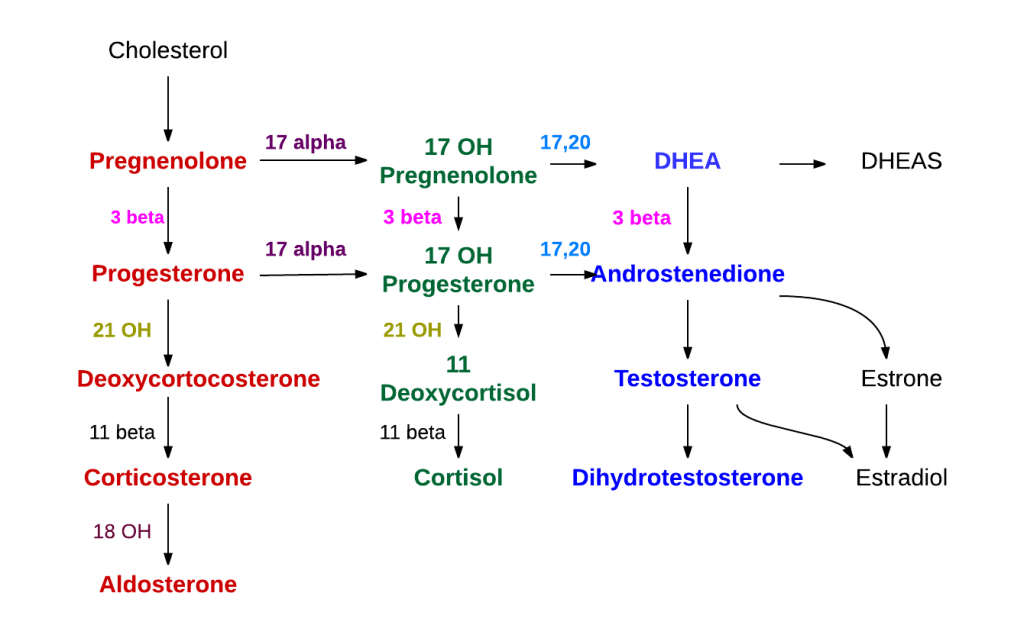
Nei tessuti bersaglio e nel fegato, il Testosterone può essere ulteriormente metabolizzato da una serie di diverse reazioni di fase I (ad esempio, riduzione) e reazioni di fase II (ad esempio, glucuronidazione). Sia il T che il DHT possono essere coniugati, principalmente nel fegato ma anche in altri tessuti, mediante glucuronidazione, che aumenta la solubilità in acqua dei composti. Gli androgeni glucuronidati vengono escreti nelle urine o tramite la bile nell’intestino tenue.
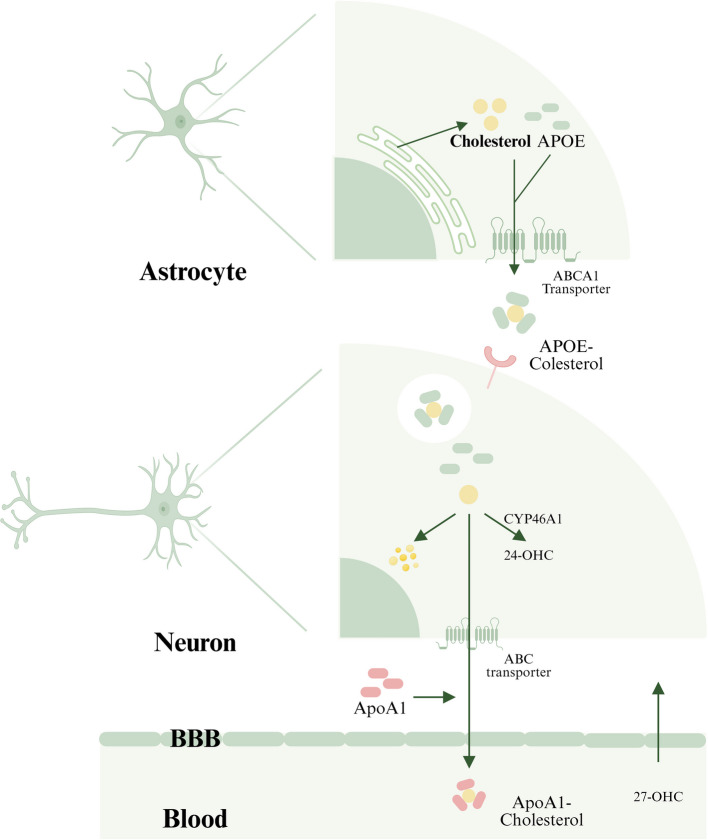
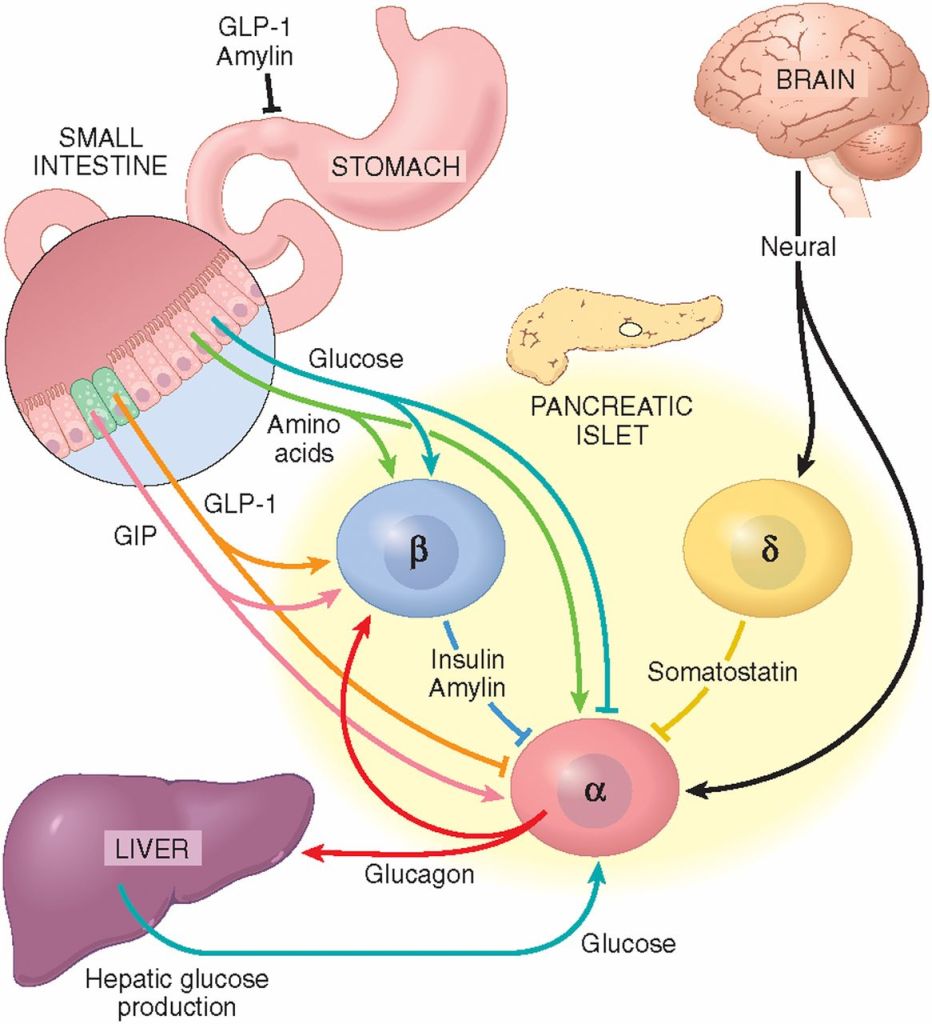
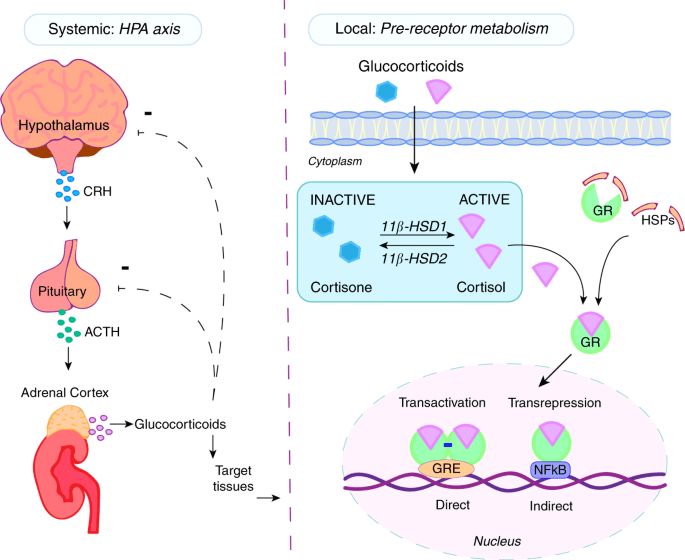
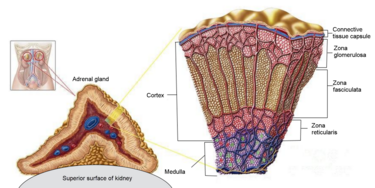
Il microbiota intestinale (GM) è costituito da trilioni di batteri, virus e funghi che sono stati descritti nel loro insieme come un organo endocrino virtuale in grado di produrre ed espellere numerose sostanze nel flusso sanguigno del suo ospite, influenzandone così la fisiologia. Il numero di batteri e la capacità metabolica del GM sono maggiori nel cieco e nel colon rispetto all’intestino tenue. Anche la composizione del GM cambia lungo il tratto gastrointestinale (GI). È stato dimostrato in vitro che alcuni ceppi batterici hanno la capacità di metabolizzare gli androgeni, ad esempio convertendo il T in DHT. La rilevanza fisiologica del GM nel metabolismo degli androgeni e nei livelli di androgeni glucuronidati e liberi in diverse regioni dell’intestino è in gran parte sconosciuta. In un recente studio, è stato osservato un legame tra il GM e le concentrazioni di androgeni nei topi, con topi femmina che avevano ricevuto contenuto intestinale maschile e che mostravano livelli sierici di T aumentati. I livelli locali di androgeni nel contenuto intestinale, tuttavia, non sono stati analizzati nello studio.
E’ stato ipotizzato e osservato che il GM, con bassa abbondanza nell’intestino tenue e alta abbondanza nell’intestino distale, può modulare il metabolismo degli androgeni e quindi influenzare i livelli locali di androgeni in modo sito-specifico lungo il tratto gastrointestinale. Per consentire la quantificazione degli androgeni nel contenuto intestinale, è stato modificato e validato un metodo specifico di gascromatografia-spettrometria di massa tandem (GC-MS/MS) precedentemente sviluppato per le analisi degli steroidi sessuali nel siero. Sono state osservate importanti differenze dipendenti dal GM nei livelli di androgeni glucuronidati e liberi nel contenuto intestinale dell’intestino tenue e delle regioni più distali dell’intestino. Sono stati osservati anche cambiamenti dipendenti dal GM nell’azione degli androgeni nei tessuti extraintestinali e intestinali. Nel complesso, questi risultati identificano il GM come un importante regolatore dell’azione locale degli androgeni nell’intestino e in altri tessuti periferici. Questo delicato equilibrio viene sensibilmente alterato da condizioni di iperandrogenismo sia patologico-organico che indotto dall’uso/abuso di AAS creando alterazioni nella popolazione microbica intestinale con conseguenti effetti connessi alla disbiosi.
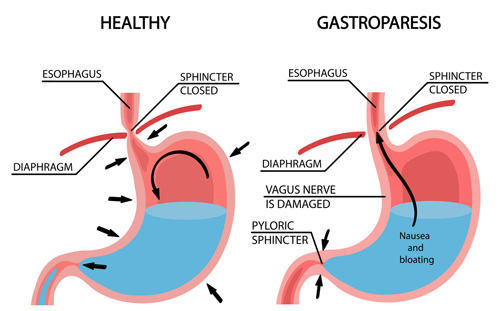
Un altro problema correlato all’uso/abuso di AAS è la comparsa di bruciore di stomaco, inappetenza e reflusso gastroesofageo. Chi conosce bene questo fastidioso effetto collaterale androgeno/antiestrogenico che si manifesta quando si assumono dosi elevate di forti androgeni, soprattutto quelli non aromatizzabili e orali, durante un ciclo di AAS, sa bene che il bruciore di stomaco può causare gravi danni d’organo, scatenando iperacidità. Per chi fa uso di AAS (e sì, la situazione peggiora con l’età), la salute gastrointestinale si preserva al meglio con l’assunzione quotidiana di un antiacido a basso dosaggio e l’uso di AAS “attenuati”… Ovviamente, non con un semplice antiacido come il Tums®, ma un antiacido vero e proprio per una soluzione rapida e semplice, oltre a seguire strategie di prevenzione a lungo termine per proteggere la mucosa gastrica. Rinunciare a un antiacido quando si soffre anche di un lieve bruciore di stomaco danneggia la mucosa gastrica!
Trascurare un adeguato trattamento del bruciore di stomaco provoca un deterioramento della mucosa gastrica. Questo articolo, nel suo insieme, propone di assumere quotidianamente e in modo continuativo gli antiacidi basali appropriati non appena compaiono i primi sintomi in qualsiasi ciclo o fase di trattamento, e non solo in modo reattivo quando i sintomi si riacutizzano.
*Bruciore di stomaco da steroidi anabolizzanti: avverti quella sensazione di bruciore a metà ciclo? Il reflusso gastroesofageo (bruciore di stomaco) colpisce molti utilizzatori di AAS, eppure rimane un problema poco compreso e non adeguatamente trattato. La domanda che molti si pongono è: “Gli steroidi possono causare bruciore di stomaco?”. La risposta è sì. Gli steroidi causano bruciore di stomaco attraverso molteplici meccanismi, tra cui l’aumento della secrezione di acido gastrico. [1] [2] [3] È importante essere a conoscenza del perché gli steroidi anabolizzanti causano bruciore di stomaco, perché un intervento adeguato dipende dall’affrontare la causa principale, non solo dal mascherare i sintomi. Questo articolo fornisce soluzioni basate sull’evidenza scientifica sia per un sollievo immediato che per la prevenzione a lungo termine. In questo modo, i problemi di stomaco non comprometteranno i progressi dell’atleta.
Androgeni/AAS e modulazione/alterazione del microbiota intestinale
Sono state segnalate differenze di genere nel microbiota intestinale anche negli esseri umani. Ad esempio, gli uomini presentano livelli più elevati di Bacteroidetes e Prevotella rispetto alle donne, suggerendo un ruolo per fattori sessuali come la diversa espressione genica dei cromosomi sessuali o le differenze nei livelli di ormoni gonadici nella modulazione del microbiota intestinale. Tuttavia, va notato che studi metagenomici più recenti hanno riportato risultati contrastanti per quanto riguarda le differenze di genere nel microbiota negli esseri umani. Alcuni studi rilevano effetti modesti o nulli del sesso sul microbiota intestinale umano, mentre altri risultati rivelano differenze di genere sostanziali. Altri studi suggeriscono che gli steroidi influenzino le comunità microbiche intestinali. Ad esempio, il microbiota intestinale subisce profondi cambiamenti durante la gravidanza nelle donne. Koren et al. hanno riscontrato un grande cambiamento nel microbiota intestinale dal primo al terzo trimestre, con un aumento della diversità complessiva e una proliferazione di Proteobacteria e Actinobacteria, con conseguenti cambiamenti nel metabolismo. Sebbene il drastico cambiamento degli ormoni steroidei (ad esempio, estrogeni e progestinici) potrebbe benissimo contribuire direttamente a questi cambiamenti nel microbiota intestinale, è stato anche suggerito che i cambiamenti nel sistema immunitario a livello delle superfici mucose potrebbero alterare il microbiota.
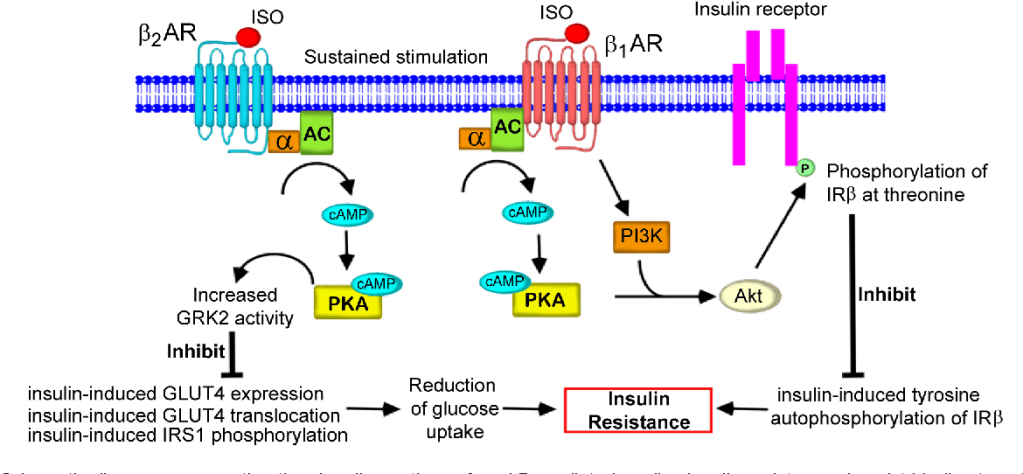
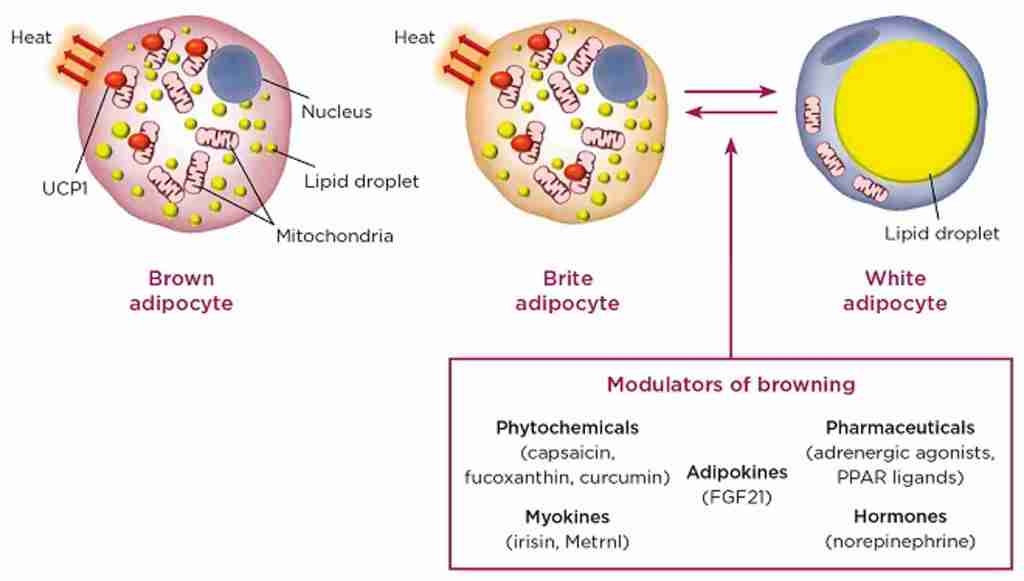
Illustrazione delle vie del recettore H2 dell’istamina sulle cellule parietali gastriche che mostra come gli steroidi anabolizzanti interrompono la secrezione acida e causano bruciore di stomaco. [Fonte immagine: Gear, Growth, and Gains | Enhanced Bodybuilding | Type-IIx | Substack]
Sebbene gli steroidi gonadici possano alterare il microbiota intestinale, sembra che, a sua volta, il microbioma intestinale possa influenzare i livelli ormonali. Nelle donne in postmenopausa, la diversità del microbiota intestinale è stata associata positivamente al rapporto dei metaboliti degli estrogeni nelle urine. Nei topi, il trasferimento del microbiota intestinale di maschi adulti a femmine immature ha causato un aumento dei livelli di Testosterone e cambiamenti metabolomici nelle femmine riceventi. Inoltre, il trasferimento del microbiota intestinale da un modello murino NOD (non obeso diabetico) maschio di diabete di tipo 1 (T1D) a topi NOD T1D femmina ha portato ad un aumento del Testosterone, cambiamenti metabolici e protezione contro il diabete di tipo 1, che di solito mostra una forte predisposizione femminile rispetto a quella maschile. Questi studi evidenziano le connessioni reciproche tra ormoni sessuali e microbiota intestinale. Nel complesso, questi studi forniscono una solida prova che il microbiota intestinale è influenzato dagli ormoni steroidei gonadici e che i livelli di steroidi possono essere alterati dal microbioma intestinale.
Oltre agli ormoni steroidei gonadici che influenzano il microbiota intestinale, vi sono prove che alcune sostanze chimiche interferenti endocrine (EDC), come livelli sovrafisiologici di Testosterone esogeno, AAS sintetici e loro metaboliti, possano alterare la composizione batterica intestinale. Il 17α-ME2 [metabolita aromatico del Methandienone o del Methyltestosterone] o il 7α-ME2 [metabolita aromatico del Trestolone], compresi i SERM, che si legano ai recettori degli estrogeni, possono essere classificati sia come EDC che come composti che altera il metabolismo (MDC) e che possono causare alterazione microbica-intestinale. Gli EDC e gli MDC esercitano i loro effetti sugli organi periferici e sul sistema nervoso centrale (SNC) per tutta la vita, sebbene possano essere particolarmente pericolosi durante il periodo critico, quando possono avere effetti deleteri permanenti che possono causare una varietà di malattie. Presumibilmente, gli EDC possono influenzare il microbiota intestinale in parte alterando l’equilibrio endocrino e forse anche agendo direttamente sul microbiota intestinale.
Sappiamo, quindi, che gli androgeni esercitano effetti importanti anche nel tratto intestinale. In uomini giovani e adulti sono stati rilevati nelle feci livelli elevati di DHT non coniugato, >70 volte superiori a quelli del siero. Questo accade, come abbiamo già accennato, perché il microbiota intestinale [GM] è coinvolto nel metabolismo intestinale e nella deglucuronidazione di DHT e Testosterone, con conseguenti livelli liberi estremamente elevati dell’androgeno più potente, il DHT, nel contenuto del colon di uomini giovani e sani. Abbiamo anche compreso che tale interazione tra GM e androgeni influenza la stessa popolazione batterica portando potenzialmente a SIBO in caso di squilibrio androgeno per eccesso. Ciò potrebbe essere particolarmente vero con molecole con elevata potenzialità androgena.
Considerando poi che un ampio sottoinsieme di neuroni enterici diventa androgeno-responsivo al momento della pubertà, e che la segnalazione degli androgeni a questi neuroni sembra essere necessaria per la normale motilità del colon, la situazione che appare è caratterizzata da un ruolo degli androgeni gonadici nella regolazione neurale della funzione intestinale e ci permette di collegare i livelli alterati di androgeni a un comune disturbo digestivo sia per eccesso che per difetto. Quest’ultimo punto ci collega anche alla possibilità di risposte psicosomatiche date da sovrastimolo della rete di comunicazione nervosa che collega l’intestino e il cervello.
L’uso di steroidi anabolizzanti androgeni (AAS) altera profondamente il microbiota umano, compromettendo la diversità batterica e influenzando l’asse intestino-ormoni. L’introduzione di testosterone sintetico o dei suoi derivati interrompe i normali meccanismi di feedback dell’organismo, modificando non solo l’ambiente intestinale ma anche quello orale. Il microbiota orale è il complesso ecosistema di oltre 700 specie di batteri, funghi e virus che popolano la bocca. In condizioni di equilibrio (eubiosi) protegge denti e gengive. Se alterato si sviluppa una disbiosi che favorisce carie, parodontite e patologie sistemiche.
Punti chiave sui principali meccanismi e gli effetti di queste alterazioni AAS-correlate:
Il microbiota intestinale agisce come un vero e proprio organo endocrino virtuale in grado di metabolizzare gli androgeni. Questa specifica rete di batteri e relativi geni viene definita testobolome.
In condizioni normali, i batteri intestinali regolano i livelli locali di testosterone modificando gli steroidi ed eliminando o facilitando il loro riassorbimento.
L’abuso di AAS immette nell’organismo quantità elevate di ormoni sintetici, sovraccaricando le vie metaboliche batteriche e alterando la composizione del microbiota.
Livelli eccessivi di androgeni riducono la ricchezza e la diversità generale del microbiota (alfa-diversità), una condizione tipicamente associata a stati infiammatori e metabolici disfunzionali.
Gli studi indicano che l’aumento dei livelli di testosterone influisce negativamente su phyla chiave come Actinobacteria, Proteobacteria, Firmicutes e Verrucomicrobia. Al contempo, si osserva la selezione di ceppi batterici specifici in grado di tollerare o metabolizzare l’eccesso di steroidi.
Gli AAS possono alterare la peristalsi intestinale. Questo cambiamento nei tempi di transito, combinato con le diete iperproteiche e/o ipercaloriche spesso seguite da chi assume queste molecole, causa disbiosi, diarrea, stitichezza e un marcato gonfiore addominale.
Gli ormoni steroidei fungono da veri e propri nutrienti per alcuni microrganismi parodontali, alterando l’ambiente subgengivale.
Gli utilizzatori di AAS mostrano una prevalenza significativamente più alta di parodontite severa e infiammazione gengivale. L’assunzione di AAS modifica la flora orale, portando a una selezione sfavorevole di specie fungine (come alcune varietà di Candida) e batteri patogeni.
L’assunzione di AAS orali danneggia direttamente l’intestino e l’intero apparato digerente a causa del loro transito forzato nel sistema gastrointestinale. A differenza delle formulazioni iniettabili, gli AAS orali sono spesso alchilati in C-17 per resistere al primo passaggio epatico, diventando altamente aggressivi per le mucose e per l’equilibrio digestivo.
Questo comporta:
Alterazione del microbiota intestinale: Modificano la flora batterica simbiotica. Questa alterazione compromette le difese immunitarie e causa disbiosi profonda.
Permeabilità intestinale (Leaky Gut): Inducono l’infiammazione delle pareti intestinali. Le giunzioni strette si allentano, permettendo a tossine e frammenti di cibo indigerito di entrare nel flusso sanguigno.
Sindrome del colon irritabile (IBS): Scatenano o peggiorano crampi addominali, meteorismo, flatulenza, diarrea cronica o stitichezza ostinata.
Malassorbimento dei nutrienti: Danneggiano i villi intestinali. Il corpo perde la capacità di assimilare correttamente vitamine, minerali e macro-nutrienti essenziali.
Il danno intestinale è strettamente connesso a quello epatico. Gli steroidi orali sono notoriamente epatotossici. Quando il fegato subisce uno stress da steroidi, la produzione e il flusso della bile si bloccano o rallentano (colestasi).
La carenza di bile nell’intestino impedisce la corretta digestione dei grassi, provocando feci oleose (steatorrea), ulteriore fermentazione intestinale e una proliferazione incontrollata di batteri patogeni nel piccolo intestino (SIBO).
L’abuso di AAS orali si manifesta rapidamente attraverso segnali specifici:
Gonfiore addominale cronico e persistente.
Acidità di stomaco, reflusso gastroesofageo forte e rischio di ulcera peptica.
Nausea frequente e inappetenza, spesso legate, ma non solo, al sovraccarico epatici.
Dolori e fitte nella zona addominale inferiore.
–Quali soluzioni al problema dell’alterazione microbiotica AAS-correlata?
Il ripristino del microbiota alterato dall’uso di AAS richiede un approccio multifasico incentrato sulla riduzione/selezione delle molecole o sulla loro totale sospensione, sul supporto della barriera intestinale e sulla modulazione batterica.
–Cambio molecolare, riduzione del dosaggio o sospensione con recupero endogeno o TRT:
La gestione della causa primaria è il passo fondamentale per consentire al “testobolome” di stabilizzarsi e/o migliorare l’alterazione.
Cambio molecolare: si tratta di cessare l’uso di forti androgeni [vedi Trenbolone e Fluoxymesterone] in favore di AAS “attenuati” [vedi Methenolone] mantenendo una base minima di Testosterone. L’osservazione e l’andamento dei sintomi dovrebbero permettere al consumatore di comprendere se passare alla seconda fase oppure procedere in tal modo
Passaggio in TRT o recupero funzionalità HPTA: nel caso la prima gestione del problema non abbia portato a miglioramenti sensibili, la cosa migliore è quella di entrare in fisiologia o attraverso una TRT o per via di un processo di recupero dell’Asse HPT che può richiedere 3-4 mesi in media.
–Ripristino dell’integrità della mucosa intestinale e della salute gastrointestinale
Gli steroidi orali, come visto in precedenza, possono infiammare la mucosa, portando a iperpermeabilità intestinale (“sindrome dell’intestino permeabile”).
L-Glutammina: Agisce come substrato energetico primario per gli enterociti, aiutando a riparare le giunzioni serrate della barriera intestinale danneggiate dall’infiammazione.
Colostro liquido: Supporta lo strato di muco protettivo e rafforza le difese immunitarie localizzate nel tratto gastrointestinale.
Ottimizzazione delle fibre: Integrare fibre prebiotiche (da verdure, frutta e semi) per nutrire i ceppi batterici benefici e contrastare la perdita di biodiversità.
Enzimi digestivi: L’uso di proteasi, amilasi e lipasi esogene riduce il carico digestivo sullo stomaco e sull’intestino, limitando i fenomeni di fermentazione anomala e lo “steroid bloating”.
Probiotici ad alta sopravvivenza: Ceppi resistenti agli acidi gastrici per reintegrare l’alfa-diversità perduta.
Antiossidanti (Glutatione liposomiale e EPA-DHA): Riducono il carico infiammatorio e lo stress ossidativo a livello epatico e intestinale causato dagli AAS C17-alchilati.
Igiene orale rigorosa: Utilizzo di antisettici orali delicati non alcolici per evitare la selezione di biofilm patogeni.
Risciacquo post-assunzione: Se si utilizzano formulazioni orali o sublinguali, è fondamentale sciacquare accuratamente la bocca per evitare il ristagno locale di residui ormonali.
Perché gli AAS causano bruciore di stomaco?
Gli steroidi anabolizzanti alterano la fisiologia gastrica attraverso percorsi mediati dagli ormoni con metabolismo epatico e interazioni recettoriali. La ricerca mostra che il testosterone e i suoi derivati sintetici inibiscono gli enzimi epatici del citocromo P-450, lo stesso sistema che regola i modulatori della secrezione acida gastrica. Studi su ratti maschi trattati con analoghi del testosterone hanno mostrato importanti alterazioni nell’idrossilazione ormonale e hanno influenzato le funzioni gastriche.
Il meccanismo si basa sui recettori H2 dell’istamina situati sulle cellule parietali gastriche. [1-3] Questi recettori stimolano la produzione di acido attraverso le vie dell’AMP ciclico. Gli androgeni ritardano la guarigione dell’ulcera gastrica in modo dose-dipendente e potenza-dipendente, riducono il flusso sanguigno, aumentano la secrezione di acido gastrico e innalzano le citochine proinfiammatorie (ad esempio, IL-1β e TNF-α). Gli steroidi anabolizzanti causano bruciore di stomaco interrompendo i normali meccanismi di feedback che regolano l’attività del recettore H2. Gli androgeni esogeni creano un ambiente ormonale in cui la secrezione di acido gastrico diventa disregolata, come nelle condizioni osservate negli stati ipersecretori.
La capacità degli steroidi anabolizzanti di provocare bruciore di stomaco dipende dalla loro potenza androgena meno la loro potenza estrogenica e può essere rappresentata dall’equazione:
Bruciore di stomaco = Adrogenicità – Estrogenicità
Equazione 1: Equazione della potenza androgenica e del rischio di bruciore di stomaco.
Utilizzando questo semplice approccio, possiamo rapidamente capire perché potenti androgeni non aromatizzabili come il Trenbolone, il Fluoximesterone (Halotestin®), il Methyldrostanolone (Superdrol) e persino l’Oxymetholone (Anadrol), che ha una androgenicità contenuta, sono diventati famigerati per causare bruciore di stomaco da steroidi, mentre il Testosterone (dal quale la conversione in E2 tende ad attenuare il bruciore di stomaco) e il Metandienone (Dianabol) [aromatizzabile in 17α-ME2, da non confondere con il 7α-ME2, il prodotto aromatico del Trestolone/MENT] sono raramente incolpati di causare questo side.
Una domanda frequente su questo effetto collaterale da AAS è:
D: Gli steroidi anabolizzanti possono scatenare un grave reflusso gastroesofageo?
R: Sì, gli steroidi anabolizzanti possono scatenare una grave malattia da reflusso gastroesofageo (GERD). Gli AAS alterano la fisiologia gastrica compromettendo i normali meccanismi di regolazione dell’acidità, inibendo gli enzimi epatici che controllano la funzione gastrica e stimolando un’eccessiva produzione di acido. Alcuni utilizzatori manifestano i sintomi più gravi di GERD durante i cicli di AAS, necessitando di dosi di farmaci più elevate rispetto a prima.
Il metabolismo epatico fornisce un’altra via. Gli steroidi competono con i composti endogeni per i siti di legame del citocromo P-450, come dimostrato dalla ricerca sull’interazione della cimetidina con i microsomi epatici. [4-1] Questa competizione rallenta il metabolismo degli ormoni regolatori gastrici e ne prolunga gli effetti stimolanti l’acidità. La struttura ad anello imidazolico presente in alcuni steroidi si lega come ligando di tipo II al citocromo P-450 e inibisce la normale funzione enzimatica.
C’è un altro motivo: il metabolismo del testosterone produce metaboliti che stimolano la mucosa gastrica. Studi hanno documentato un aumento dei prodotti idrosolubili e alterazioni nei modelli di idrossilazione degli steroidi, creando composti con proprietà irritanti per lo stomaco.
Soluzioni rapide per il bruciore di stomaco indotto dagli steroidi
Gli antagonisti del recettore H2 apportano il sollievo più diretto per il bruciore di stomaco causato dagli AAS bloccando la via di produzione di acido mediata dall’istamina discussa in precedenza. La Ranitidina (Zantac®) e la Famotidina (Pepcid®) rappresentano opzioni di seconda generazione che affrontano l’ipersecrezione di acido gastrico senza interferire con il metabolismo degli steroidi.
Tabella comparativa delle classi di antiacidi per il sollievo dal bruciore di stomaco indotto da steroidi, con particolare attenzione agli antagonisti del recettore H2 preferiti come Famotidina e Ranitidina.
Attenzione:
Dosaggio clinicamente rilevante: Dosaggio secondo i regimi clinici standard per l’uomo: cimetidina (300 mg), ranitidina (50 mg) e famotidina (10 mg).
Ranitidina: Si prega di notare che in alcuni paesi regolamentati (tra cui Stati Uniti, Regno Unito e Canada), la ranitidina è stata ritirata dal mercato o richiamata a causa di problemi di sicurezza relativi alle impurità di NDMA.
Il carbonato di calcio (ad es. Tums®) non è raccomandato nemmeno quotidianamente per una (1) settimana, ma per le riacutizzazioni gravi, è consigliabile utilizzarlo tamponato da un antagonista del recettore H2 per non più di tre (3) giorni nella stessa (1) settimana.
Un altra domanda frequente
D: Qual è il miglior antiacido da assumere con gli steroidi anabolizzanti?
R: La Famotidina (Pepcid®) è considerata il gold standard per chi assume steroidi perché agisce sul problema dell’istamina alla radice, senza interferire con il metabolismo ormonale. Gli antagonisti dei recettori H2 sopprimono efficacemente la produzione di acido gastrico, preservando al contempo gli effetti desiderati degli steroidi anabolizzanti, a differenza della Cimetidina che può alterare il metabolismo del testosterone, influenzando negativamente il testosterone libero e biodisponibile. Gli antagonisti dei recettori H2 sono altamente efficaci nella guarigione delle ulcere duodenali esistenti (76% di remissione dopo 4 settimane)
La differenza strutturale è importante. Ranitidina e famotidina non presentano anelli imidazolici ed evitano il legame con il citocromo P-450, che causa interazioni farmacologiche. Questi composti non hanno mostrato alcun effetto sui percorsi di idrossilazione ormonale identificabili, a differenza della cimetidina che ha ridotto la formazione di metaboliti polari e aumentato la produzione di 3-androstanediolo.
Le evidenze cliniche ne supportano l’uso: studi controllati condotti in tutto il mondo hanno mostrato tassi di guarigione del 76% a 4 settimane per entrambi i farmaci. La Ranitidina ha dimostrato una particolare efficacia nel sopprimere l’ipersecrezione gastrica resistente ad altri trattamenti.
Strategie di prevenzione a lungo termine
L’infografica mostra l’efficacia dell’assunzione serale di antagonisti dei recettori H2 e di un’alimentazione strategica pre-allenamento nel prevenire le recidive di bruciore di stomaco durante i cicli di steroidi anabolizzanti.[Fonte immagine: Gear, Growth, and Gains | Enhanced Bodybuilding | Type-IIx | Substack]
Assunzione notturna
I protocolli di mantenimento vanno oltre la gestione dei sintomi acuti e affrontano la natura ricorrente e corrosiva del bruciore di stomaco causato dagli steroidi anabolizzanti durante i cicli prolungati. La somministrazione notturna mira a controllare la secrezione acida notturna, quando il pH gastrico si abbassa e i sintomi del reflusso si intensificano.
Un altra domanda frequente
D: Come possono i bodybuilder prevenire il reflusso acido durante l’uso di AAS?
R: Le strategie di prevenzione includono l’assunzione di antagonisti dei recettori H2 prima di coricarsi durante tutto il ciclo, occasionalmente tamponati con carbonato di calcio, il consumo di pasti a basso contenuto proteico e ad alto contenuto di carboidrati prima dell’allenamento, l’assunzione di una quantità adeguata di acqua e l’evitare alimenti scatenanti come cioccolato, cibi piccanti e caffeina.
I pazienti che interrompono l’assunzione di inibitori della pompa protonica (IPP) dopo la risoluzione dei sintomi spesso vanno incontro a una rapida ricaduta. Gli studi hanno documentato che i tassi di guarigione raggiungono il 76% a 4 settimane e l’87% a 6 settimane, tuttavia l’interruzione del trattamento provoca una ricaduta nella maggior parte dei soggetti entro 12 mesi.
Strategie nutrizionali pre-allenamento:
Limita proteine, spezie e caffeina. Preferisci carboidrati pre-allenamento (neutralità a livello dell’indice glicemico e substrato energetico glicolitico per un intenso lavoro muscolare) e liquidi ad alto pH neutro, come un frullato contenente Ciclodestrine altamente ramificate o Maltodestrina in un rapporto di 3:1 o superiore rispetto alle proteine (inclusi gli EAA).
Punti chiave sottoforma di domande:
D1. Cosa aiuta ad alleviare il bruciore di stomaco causato dagli steroidi? Assumere antiacidi con ogni dose di steroidi è molto efficace nel ridurre i sintomi digestivi. Gli antagonisti dei recettori H2 come la famotidina e la ranitidina forniscono un sollievo diretto bloccando la produzione di acido senza interferire con il metabolismo degli steroidi. Questi farmaci agiscono sull’ipersecrezione di acido gastrico e possono ridurre significativamente i sintomi in pochi giorni se usati con costanza.
D2. Quale antiacido è meglio assumere con gli steroidi anabolizzanti? La famotidina (Pepcid) e la ranitidina sono le opzioni preferite da chi assume steroidi perché non interferiscono con il metabolismo ormonale. Questi antagonisti dei recettori H2 sopprimono efficacemente la produzione di acido gastrico preservando gli effetti desiderati degli steroidi anabolizzanti, a differenza della cimetidina che può alterare il metabolismo del testosterone.
D3. Gli steroidi anabolizzanti possono scatenare un grave reflusso acido? Sì, gli steroidi anabolizzanti possono scatenare una grave malattia da reflusso gastroesofageo (GERD). Gli steroidi alterano la fisiologia gastrica compromettendo i normali meccanismi di regolazione dell’acidità, inibendo gli enzimi epatici che controllano la funzione gastrica e stimolando un’eccessiva produzione di acido. Alcuni utilizzatori manifestano i sintomi più gravi di reflusso gastroesofageo durante i cicli di steroidi, richiedendo dosi di farmaci più elevate rispetto a prima.
D4. Come possono i bodybuilder prevenire il reflusso acido durante l’uso di steroidi? Le strategie di prevenzione includono l’assunzione di antagonisti dei recettori H2 prima di coricarsi durante tutto il ciclo, occasionalmente tamponati con carbonato di calcio, pasti a basso contenuto proteico e ad alto contenuto di carboidrati prima dell’allenamento, bere acqua a sufficienza ed evitare cibi che scatenano il reflusso come cioccolato, cibi piccanti e caffeina.
D5. Perché il bruciore di stomaco ritorna dopo aver interrotto l’assunzione di farmaci per il reflusso acido durante un ciclo di steroidi? Il bruciore di stomaco ritorna perché gli antagonisti dei recettori H2 riparano il danno ma non curano la disfunzione di base che gli steroidi creano nella regolazione dell’acidità gastrica. Gli studi mostrano tassi di recidiva del 50-90% entro un anno dall’interruzione del trattamento, motivo per cui è necessaria una terapia di mantenimento continua a basse dosi durante tutto il ciclo di steroidi, piuttosto che un uso intermittente.
Conclusione
Gli steroidi anabolizzanti alterano la normale regolazione dell’acidità gastrica e danneggiano la mucosa dello stomaco, causando bruciore di stomaco e reflusso acido in molti utilizzatori. Questo articolo analizza i meccanismi alla base del bruciore di stomaco indotto dagli steroidi, evidenziando il ruolo degli antagonisti dei recettori H2, come famotidina e ranitidina, come efficaci opzioni di sollievo che non interferiscono con il metabolismo degli steroidi. Sottolinea inoltre l’importanza di una terapia di mantenimento continua a basse dosi durante i cicli di steroidi per prevenire le ricadute e proteggere la salute dell’apparato digerente. Consigli pratici sulla scelta dei farmaci, le strategie di dosaggio e le modifiche dello stile di vita assicurano agli utilizzatori la possibilità di gestire i sintomi senza compromettere l’allenamento o i protocolli ormonali.
Inoltre, abbiamo visto che l’uso/abuso di AAS altera profondamente il microbiota umano, compromettendo la diversità batterica e influenzando l’asse intestino-ormoni. L’assunzione di AAS orali danneggia direttamente l’intestino e l’intero apparato digerente a causa del loro transito forzato nel sistema gastrointestinale.
Attualmente, chi fa uso di steroidi anabolizzanti-androgeni (AAS) a lungo termine ha a disposizione soluzioni basate su evidenze scientifiche per gestire il bruciore di stomaco e le problematiche intestinali. Famotidina e ranitidina offrono un sollievo immediato senza interferire con il metabolismo e agiscono sull’ipersecrezione di acido gastrico, preservando al contempo i protocolli ormonali. La scelta di “androgeni attenuati” e/o l’evitamento di AAS orali può essere una pratica riduttiva delle problematiche intestinali; l’interruzione dell’uso o il passaggio in TRT risultano, a volte, le uniche scelte intelligenti da applicare. È comunque fondamentale comprendere che la terapia di mantenimento “nati-acido” durante i cicli previene le ricadute, non si tratta solo di un trattamento reattivo. Chi avverte bruciore può seguire questi protocolli e aspettarsi una riduzione dei sintomi più importanti entro pochi giorni. Una corretta gestione gastrica garantirà che le complicazioni digestive non compromettano i progressi dell’allenamento o i risultati del ciclo.
Machowska, A., et al. “Gastric Secretion, Proinflammatory Cytokines and Epidermal Growth Factor (EGF) in the Delayed Healing of Lingual and Gastric Ulcerations by Testosterone.” Inflammopharmacology, vol. 16, no. 1, 2008, pp. 40–47, [https://doi.org/10.1007/s10787- 007-1600 – 6](https://doi.org/10.1007/s10787- 007-1600 – 6).↩︎↩︎
Kowalewski, K., et al. “Effect of Sex Hormones on Gastric Secretion and on Gastric Mucosa in Oophorectomized Histamine Stimulated Rats.” Digestion, vol. 3, no. 1, 1970, pp. 13–19. DOI.org (Crossref), https://doi.org/10.1159/000196983.↩︎↩︎↩︎
Galbraith, Richard A., and Peter H. Jellinck. “Differential Effects of Cimetidine, Ranitidine and Famotidine on the Hepatic Metabolism of Estrogen and Testosterone in Male Rats.” Biochemical Pharmacology, vol. 38, no. 12, 1989, pp. 2046–49, [https://doi.org/10.1016/0006- 2952(89)90507 – 8](https://doi.org/10.1016/0006- 2952(89)90507 – 8).↩︎↩︎↩︎↩︎↩︎↩︎↩︎
Adelakun, Sunday Aderemi, et al. “Long-Term Exposure to Cimetidine Induced Gonado-Toxicity in Male Rats: Modulating Role of Ocimum Gratissimum.” Revista Internacional De Andrologı́a, vol. 20, 2022, pp. S2–16, https://doi.org/10.1016/j.androl.2020.10.012.↩︎
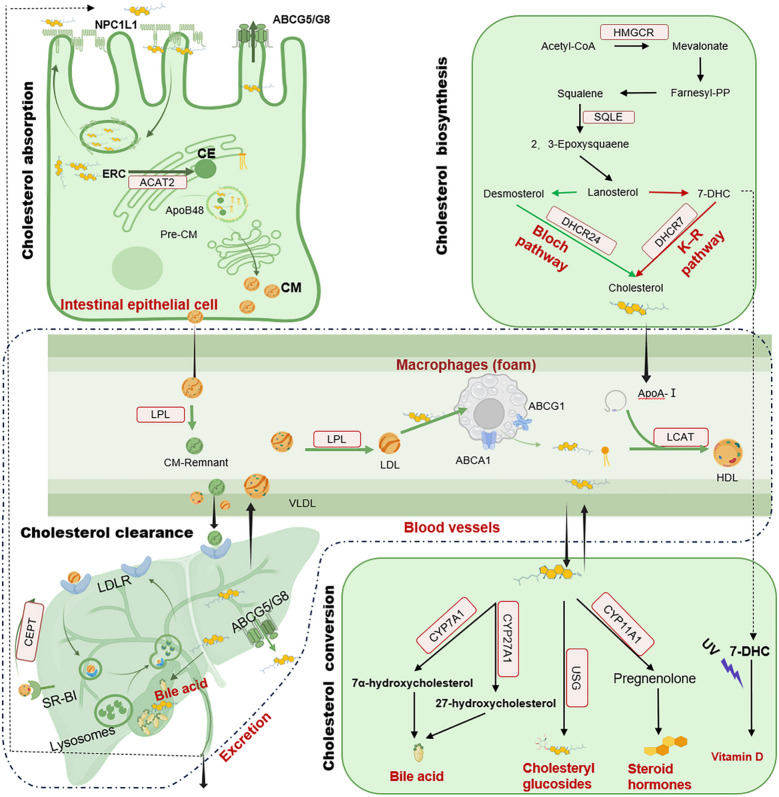
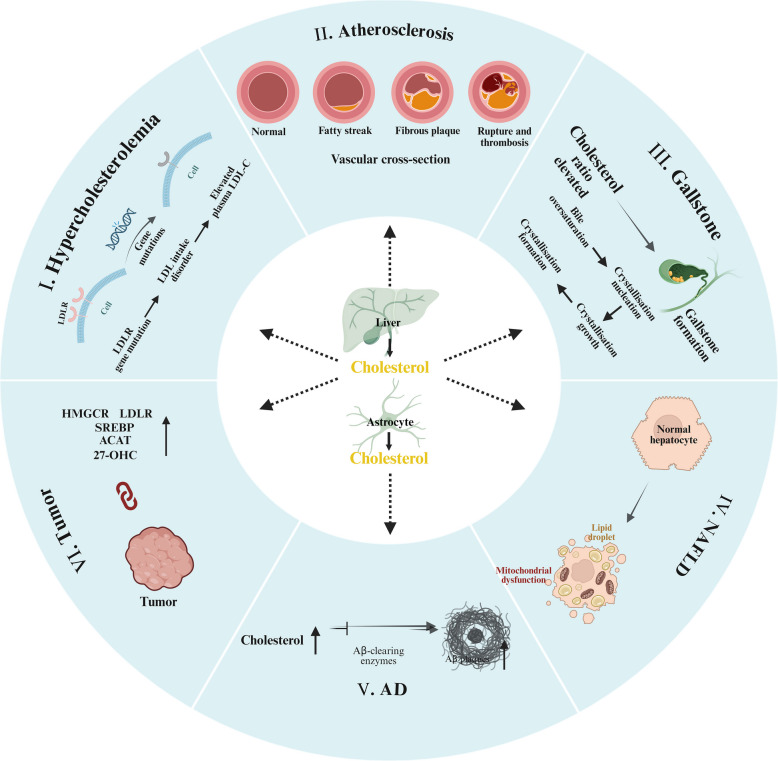
Nella prima parte di questa disamina sul Colesterolo e quanto ne concerne partendo dalla biosintesi per poi passare alle funzioni fisiologiche e le alterazioni con patologie connesse, in questa seconda ed ultima parte vedremo da vicino le terapia ad oggi in uso clinico per trattare l’ipercolesterolemia.
La terapia classica, esemplificata dalle Statine, ha rivoluzionato la prevenzione delle malattie cardiovascolari inibendo l’HMGCR. Nonostante la loro efficacia, l’intolleranza alle Statine e il rischio cardiovascolare residuo persistono, in particolare nei disturbi genetici come l’ipercolesterolemia familiare omozigote (HoFH), spingendo lo sviluppo di terapie di nuova generazione. Una pietra miliare significativa in questa evoluzione è l’avvento degli inibitori di PCSK9. Gli anticorpi monoclonali, come Evolocumab e Alirocumab, raggiungono una sostanziale riduzione del LDL bloccando l’interazione PCSK9-LDLR. Il paradigma è ulteriormente cambiato con Inclisiran, una terapia a base di siRNA che consente la somministrazione semestrale attraverso il targeting degli epatociti mediato da GalNAc. Ciò esemplifica la transizione dal trattamento cronico a strategie di silenziamento genico durature. Inoltre, agenti non statinici come Ezetimibe e sequestranti degli acidi biliari hanno ampliato l’arsenale terapeutico. Essi agiscono sull’assorbimento intestinale del Colesterolo e sul ricircolo degli acidi biliari. Nei casi refrattari come l’ipercolesterolemia familiare omozigote (HoFH), gli inibitori di ANGPTL3 (come Evinacumab) e gli inibitori di MTP (come Lomitapide) aprono nuove prospettive di sopravvivenza per i pazienti che in precedenza non avevano “alcun farmaco disponibile” attraverso vie indipendenti da LDLR.
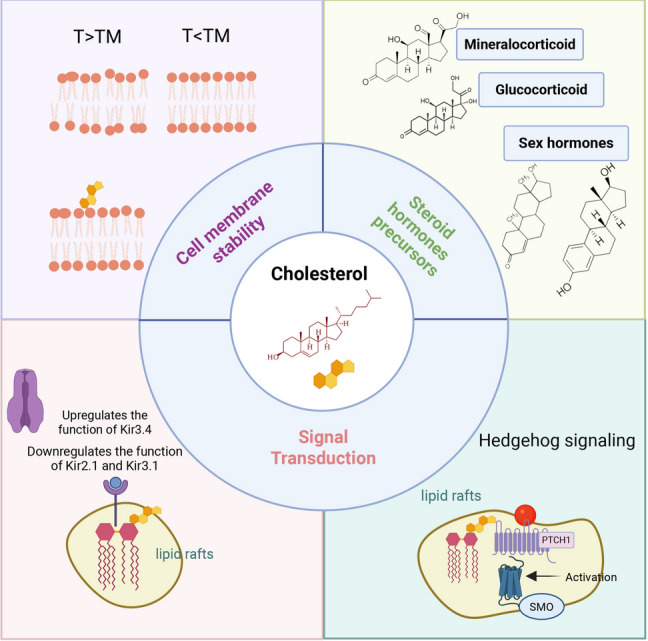
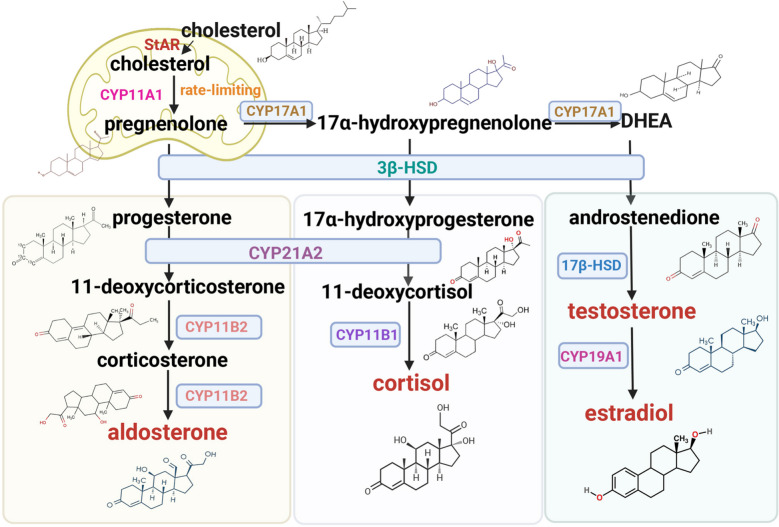
Sebbene questi interventi riducano efficacemente l’LDL circolante, il loro approccio fondamentale, basato sulla soppressione della biodisponibilità del Colesterolo endogeno, merita un esame critico. Dato il ruolo indispensabile del Colesterolo nell’integrità delle membrane, nella sintesi dei neurosteroidi e nella produzione di acidi biliari, le conseguenze a lungo termine della deplezione sistemica di colesterolo rimangono ancora poco chiare. Le evidenze emergenti di effetti avversi associati alle Statine (ad esempio, aumento del rischio di diabete, miopatia e incidenza paradossale di tumori con l’uso prolungato) e il “paradosso della Niacina” (riduzione del colesterolo LDL senza benefici cardiovascolari) sottolineano i potenziali limiti di questa strategia.
Ogni classe farmacologica verrà esaminata criticamente in termini di meccanismi molecolari, farmacocinetica, indicazioni cliniche, profili di sicurezza, potenziale di interazione farmacologica e considerazioni farmacogenetiche rilevanti. Sebbene l’introduzione di nuovi agenti ipolipemizzanti rappresenti un importante progresso nella prevenzione cardiovascolare, la loro ampia diffusione è mitigata da considerazioni di costo-efficacia e allocazione delle risorse sanitarie. Terapie come gli inibitori di PCSK9, Inclisiran ed Evinacumab sono associate a costi di acquisizione sostanzialmente più elevati rispetto alle Statine tradizionali o all’Ezetimibe, sollevando interrogativi sulla loro sostenibilità economica, soprattutto in contesti di prevenzione primaria. Integrando i risultati di studi clinici randomizzati cardine, dati del mondo reale e linee guida in continuo aggiornamento, questa seconda parte offre una valutazione completa e aggiornata dell’armamentario terapeutico per la dislipidemia. L’adozione di questi agenti emergenti preannuncia un passaggio verso strategie di gestione dei lipidi più personalizzate, basate sui meccanismi e adattate al rischio. Man mano che il settore continua a progredire, clinici, ricercatori e responsabili politici avranno bisogno di una comprensione approfondita di queste innovazioni per ottimizzare la prevenzione cardiovascolare e migliorare gli esiti a lungo termine in diverse popolazioni di pazienti.
–Trattamenti farmacologici per la dislipidemia/ipercolesterolemia–
Statine [inibitori dell’HMGCR].
La scoperta delle Statine rappresenta una pietra miliare nel campo della gestione del colesterolo. Nel 1973, lo screening pionieristico di 3.800 ceppi fungini condotto da Akira Endo portò alla scoperta della Mevastatina (Compactina), il primo inibitore dell’HMGCR identificato. Questa scoperta fondamentale aprì la strada al successivo sviluppo della Lovastatina da parte di Merck Pharmaceuticals nel 1987, che divenne la prima Statina approvata clinicamente. Grazie alla continua ottimizzazione molecolare, le successive generazioni di Statine hanno raggiunto una maggiore potenza e specificità, in particolare la Simvastatina (Zocor®), l’Atorvastatina (Lipitor®) e la Rosuvastatina (Crestor®), dimostrando un’efficacia nella riduzione del LDL che varia dal 25 al 60% negli studi clinici. Essendo i farmaci ipocolesterolemizzanti più prescritti al mondo, le Statine rimangono la pietra angolare della terapia per la gestione dell’ipercolesterolemia.
Dal punto di vista meccanicistico, le Statine esercitano i loro effetti inibendo l’attività dell’HMGCR, riducendo così la sintesi del Colesterolo e attivando l’SREBP2 nel reticolo endoplasmatico. Legandosi agli elementi regolatori degli steroli (SRE) nel promotore dell’LDLR, l’SREBP2 potenzia la sintesi dell’LDLR, portando a un maggiore assorbimento di LDL-C circolante. Questo meccanismo di feedback finemente regolato bilancia i livelli di Colesterolo intracellulare riducendo al contempo le concentrazioni di LDL-C circolante. In particolare, le Statine conferiscono una protezione cardiovascolare persistente attraverso effetti pleiotropici, tra cui la stabilizzazione della placca e il miglioramento della funzione endoteliale, che contribuiscono alla riduzione del rischio residuo anche dopo la cessazione del trattamento. Tuttavia, gli effetti avversi dose-dipendenti, come i sintomi muscolari associati alle Statine (SAMS), l’aumento asintomatico delle transaminasi epatiche, la calcificazione della placca e la rara rabdomiolisi, richiedono un attento monitoraggio clinico.
Simvastatina (Zocor®), l’Atorvastatina (Lipitor®) e la Rosuvastatina (Crestor®) sono generalemte somministrate al dosaggio dai 10 ai 40mg/die per la Simvastatina, da 10 a 80mg/die per la Atorvastatina e da 5 a 40mg/die per la Rosuvastatina.
I più comuni effetti collaterali derivanti dall’uso di Statine includono dolori muscolari (mialgie), lievi disturbi gastrointestinali e un potenziale aumento della glicemia. In rari casi, possono provocare tossicità muscolare grave o alterazioni epatiche. Alcuni pazienti riportano stanchezza generale, spesso legata alla riduzione dei livelli di Coenzima Q10 cellulare e ne viene spesso indicata la co-somministrazione.
Niacina
Molecola di Niacina
La Niacina, nota anche come vitamina B3 o acido nicotinico, è un micronutriente essenziale necessario per la sintesi di NAD. Storicamente, la niacina è stata una terapia di prima linea per la dislipidemia. Inibendo la lipolisi del tessuto adiposo, riduce l’apporto di acidi grassi liberi necessari per la sintesi epatica dei trigliceridi, diminuendo così la produzione di VLDL e, secondariamente, l’LDL circolante; allo stesso tempo, rimane l’agente più potente per aumentare il Colesterolo HDL e può ridurre i Trigliceridi del 20-35%. Questi effetti hanno reso la Niacina un valido coadiuvante per l’iperlipidemia familiare e altre forme gravi di ipercolesterolemia. Nell’era delle Statine, tuttavia, ampi studi clinici hanno dimostrato che l’aggiunta di Niacina alla terapia intensiva con Statine, nonostante un ulteriore miglioramento del profilo lipidico, non conferisce un’ulteriore riduzione del rischio cardiovascolare e può persino aumentare la mortalità complessiva, un’osservazione definita “paradosso della Niacina”. Ciò suggerisce che l’influenza della Niacina sugli eventi cardiovascolari coinvolge percorsi che sono indipendenti o non adeguatamente catturati dai suoi tradizionali meccanismi di modulazione del Colesterolo.
Negli adulti, il dosaggio giornaliero della Niacina è dai 500mg a 2g da una a tre volte al giorno.
I più comuni effetti collaterali da assunzione di Niacina includono vampate di calore, arrossamenti, prurito, nausea, alterazioni delle transaminasi, aumento della glicemia e vertigini. Le vampate e arrossamenti (Flushing) sono l’effetto più comune della Niacina. Si manifesta con sensazione di calore, rossore e formicolio a viso e collo, spesso accentuati da alcol, attività fisica o cibi piccanti. Alcuni studi indicano che dosi eccessive (e l’accumulo di specifici metaboliti) sono associati a infiammazione vascolare e aumento del rischio di ictus o infarto.
In Italia non esistono più farmaci registrati come ipolipemizzanti a base di Niacina pura (Acido Nicotinico) per il trattamento del colesterolo e dei Trigliceridi.
I vecchi medicinali specialistici (come Tredaptive o Pelzont) sono stati ritirati dal commercio e la loro autorizzazione è stata revocata a causa di un bilancio rischio/beneficio sfavorevole, legato soprattutto agli effetti collaterali.
In sostituzione della Niacina tradizionale per il controllo dei lipidi nel sangue, l’unico farmaco prescrivibile in Italia è un suo derivato sintetico: Acipimox.
L’Acipimox (noto in commercio come Olbetam) è un farmaco ipolipemizzante derivato dell’Acido Nicotinico. Viene utilizzato per ridurre i livelli ematici di Trigliceridi e VLDL in pazienti con dislipidemie primitive o secondarie che non rispondono adeguatamente a dieta, statine o fibrati.
L’Acipimox inibisce la lipolisi nel tessuto adiposo, riducendo la quantità di acidi grassi liberi che raggiungono il fegato. Di conseguenza, il fegato produce meno lipoproteine a bassissima densità (VLDL), abbassando i livelli di Trigliceridi e, indirettamente, del Colesterolo nel sangue.
Il farmaco è indicato per il trattamento di specifiche alterazioni dei lipidi nel sangue:
Ipertrigliceridemia isolata (iperlipoproteinemia di tipo IV).
Ipercolesterolemia e ipertrigliceridemia miste (iperlipoproteinemia di tipo IIb).
Va sempre associato a modifiche dello stile di vita (dieta ipolipidica ed esercizio fisico).
Essendo una molecola modificata dalla Niacina, è un agonista meno potente sui recettori cutanei che scatenano il flushing. Pur potendo causare arrossamento, questo è nettamente meno frequente, meno severo e più tollerabile rispetto alla Niacina.
Ai dosaggi standard non interferisce con il metabolismo del glucosio né altera l’acido urico. Per questa ragione, l’Acipimox è storicamente preferito nei pazienti dislipidemici che soffrono anche di diabete di tipo 2 o sindrome metabolica.
L’Acipimox mantiene un’ottima efficacia nella riduzione dei trigliceridi (e VLDL), ma ha un effetto decisamente più modesto o limitato sulla riduzione del colesterolo LDL e sull’innalzamento dell’HDL rispetto alla Niacina tradizionale.
Nelle prime fasi del trattamento con Acipimox possono manifestarsi alcuni disturbi, che spesso tendono a regredire con la continuazione della terapia:
Rossore (vasodilatazione cutanea), sensazione di calore e prurito.
Originariamente sviluppato da Schering-Plough Pharmaceuticals, l’ezetimibe rappresenta il primo inibitore dell’assorbimento del colesterolo di questa classe che si rivolge specificamente al recettore NPC1L1. Meccanicisticamente, questo agente terapeutico riduce l’assorbimento intestinale del colesterolo attraverso l’inibizione selettiva del sistema di trasporto del colesterolo mediato dal recettore NPC1L1 nella membrana a orletto a spazzola degli enterociti. Legandosi in modo competitivo al dominio extracellulare di NPC1L1, l’Ezetimibe blocca efficacemente il processo di internalizzazione del colesterolo e dei fitosteroli dalle micelle intestinali, impedendone così la successiva incorporazione nei chilomicroni per il trasporto epatico. Questa azione farmacologica crea un duplice effetto terapeutico: (1) riduzione diretta dell’assorbimento del colesterolo alimentare e (2) miglioramento indiretto della clearance delle LDL circolanti. La diminuzione dell’afflusso di colesterolo epatico innesca una sovraregolazione compensatoria dell’LDLR sugli epatociti, migliorando significativamente la rimozione delle particelle LDL circolanti dal flusso sanguigno. In particolare, l’attività terapeutica dell’ezetimibe è prevalentemente localizzata all’interno del sistema circolatorio enteroepatico, con conseguente minima esposizione sistemica (biodisponibilità < 5%) e, di conseguenza, bassi rischi di interazioni farmacocinetiche tra farmaci. Clinicamente, l’ezetimibe dimostra effetti sinergici se combinato con la terapia con statine, producendo una riduzione incrementale del 15-20% dei livelli di LDL-C rispetto alla monoterapia con statine. Questa strategia di combinazione è stata validata in numerosi studi clinici randomizzati controllati per migliorare significativamente gli esiti cardiovascolari nelle popolazioni ad alto rischio. Il profilo di sicurezza dell’ezetimibe rimane favorevole, con ampi studi clinici che documentano l’assenza di un aumento significativo di gravi effetti avversi rispetto al placebo, rendendolo particolarmente adatto per la gestione lipidica a lungo termine.
La dose abituale di Ezetimibe per adulti in caso di iperlipidemia è di 10mg per via orale una volta al giorno.
L’Ezetimibe è generalmente ben tollerato, con effetti collaterali lievi e transitori:
Disturbi gastrointestinali: dolore addominale, diarrea, flatulenza e senso di gonfiore.
Stanchezza generale: astenia o affaticamento insolito.
Sintomi simil-influenzali: tosse o lievi infezioni alle vie respiratorie.
Più rari:
Miopatia e rabdomiolisi: grave danno o debolezza muscolare con rischio di tossicità renale (le urine possono diventare scure). Il rischio aumenta se combinato con statine o fibrati. Epatite: infiammazione del fegato che si manifesta con ingiallimento degli occhi o della pelle (ittero) e dolore addominale superiore destro. Pancreatite: grave infiammazione del pancreas con forti dolori allo stomaco che si irradiano alla schiena. Reazioni allergiche severe: angioedema (gonfiore di viso, labbra o lingua), orticaria ed eruzioni cutanee estese.
L’aggiunta della Statine può modificare la frequenza o la tipologia di alcuni sintomi:
Cefalea: mal di testa frequente.
Mialgia: dolori o indolenzimenti muscolari.
Disturbi gastrici: nausea, indigestione o bruciore di stomaco.
Alterazioni epatiche: aumento delle transaminasi (enzimi del fegato) nei test del sangue.
Parestesie: sensazioni di formicolio, intorpidimento o pizzicore agli arti.
Acido Bempedoico
Molecola di Acido Bempedoico
L’Acido Bempedoico è un profarmaco orale, da assumere una volta al giorno, che viene attivato selettivamente negli epatociti dalla sintetasi 1 dell’acil-CoA a catena molto lunga (ACSVL1, codificata dal gene SLC27A2). Una volta attivato, l’acido bempedoico viene convertito in bempedoico acido-CoA, che funge da potente e selettivo inibitore dell’ATP-citrato liasi (ACLY), l’enzima che catalizza la fase limitante della biosintesi del colesterolo e degli acidi grassi. Riducendo la biosintesi epatica del colesterolo, l’acido bempedoico innesca la sovraregolazione del LDLR mediata da SREBP2, accelerando la clearance del LDL-C e abbassando i livelli plasmatici di LDL-C. Poiché l’ACSVL1 è assente nel muscolo scheletrico, l’acido bempedoico evita gli effetti collaterali mialgici comuni alle statine, rendendolo una terapia autonoma o aggiuntiva approvata dalla FDA (febbraio 2020) — commercializzata come Nexletol — per i pazienti intolleranti alle statine.
La bioattivazione specifica del Acido Bempedoico nel fegato minimizza l’esposizione sistemica e riduce significativamente il rischio di effetti avversi a carico del muscolo osservati con le statine. Una volta convertito nella sua forma attiva (metabolita attivo), il bempedoil-CoA inibisce l’enzima ACL, riducendo così la produzione epatica di acetil-CoA, un precursore essenziale nella sintesi del colesterolo e degli acidi grassi. La diminuzione del colesterolo intracellulare porta a una sovraregolazione dei LDLR, con conseguente aumento della clearance del colesterolo LDL circolante. La sovraregolazione dei LDLR, a seguito della diminuzione del colesterolo intracellulare, fa parte di un meccanismo di autoregolazione degli epatociti (feedback negativo) con un ruolo nel mantenimento dell’omeostasi lipidica intracellulare, incluso il colesterolo. Pertanto, le cellule epatiche assicurano il loro livello ottimale di colesterolo per le esigenze metaboliche, senza produrre un eccessivo accumulo intracellulare di colesterolo. Più specificamente, la diminuzione del colesterolo intracellulare innesca una cascata di regolazione genica mediata dal fattore SREBP-2 (Sterol Regulatory Element-Binding Protein 2), uno dei tre membri della famiglia SREBP. Si tratta di un fattore di trascrizione ubiquitario, espresso nella maggior parte delle cellule, ma con un’attività particolarmente intensa nel fegato. In condizioni normali, SREBP-2 viene sintetizzato come una proteina inattiva legata alla membrana e attaccata al reticolo endoplasmatico. Nelle cellule carenti di colesterolo/con carenza di colesterolo, SREBP-2 viene trasportato all’apparato di Golgi, dove subisce una scissione proteolitica che rilascia il suo frammento attivo (nSREBP2). Successivamente, nSREBP-2 viene traslocato nel nucleo, dove si lega all’elemento regolatore degli steroli (SRE) nel promotore del gene LDLR, stimolandone la trascrizione. Poiché l’ACL è posizionato a monte dell’HMG-CoA reduttasi nella via del mevalonato, l’acido bempedoico può essere utilizzato in sinergia con le statine o l’ezetimibe. Inoltre, evidenze precliniche suggeriscono che l’acido bempedoico possa attivare la proteina chinasi attivata da AMP (AMPK), contribuendo potenzialmente agli effetti antinfiammatori e metabolici, sebbene la rilevanza clinica di questo meccanismo sia ancora oggetto di studio.
L’acido bempedoico è indicato per pazienti adulti con ipercolesterolemia primaria (sia familiare che non familiare), ASCVD accertata o intolleranza alle statine. Il suo ruolo è sempre più riconosciuto nei pazienti ad alto e altissimo rischio cardiovascolare che non raggiungono i valori target di LDL-C con il solo trattamento con statine, fornendo riduzioni di LDL-C di circa il 17-28% in monoterapia e fino al 48% in combinazione con ezetimibe. Nell’ipercolesterolemia familiare eterozigote (HeFH), in cui i pazienti presentano mutazioni che influenzano la funzione del recettore LDLR, l’acido bempedoico può aumentare la clearance dell’LDL-C mediante la regolazione positiva di questi recettori. Nelle forme poligeniche o non familiari di ipercolesterolemia, rappresenta un valido coadiuvante quando le misure dietetiche e la terapia con statine sono insufficienti.
La dose raccomandata di acido bempedoico è di 180 mg per via orale una volta al giorno, con o senza cibo. È disponibile in compresse per monoterapia e in combinazione a dose fissa con ezetimibe (10 mg). Non è necessario alcun aggiustamento del dosaggio in base all’età, al sesso o in presenza di insufficienza renale o epatica da lieve a moderata.
L’acido bempedoico ha dimostrato un profilo di sicurezza favorevole, con una bassa incidenza di eventi avversi a carico della muscolatura. A differenza delle statine, non aumenta il rischio di mialgia, miopatia o rabdomiolisi, rappresentando quindi un’opzione valida per i pazienti con una storia di sintomi muscolari associati all’uso di statine. Nonostante il suo profilo di sicurezza complessivamente favorevole, l’acido bempedoico è associato ad alcuni effetti avversi che richiedono un’attenta valutazione clinica. In particolare, può indurre iperuricemia a causa dell’inibizione del trasportatore renale di anioni organici 2 (OAT2), che riduce l’escrezione di acido urico e aumenta i livelli sierici di urato. Sono stati inoltre osservati lievi aumenti delle transaminasi epatiche, in particolare dell’alanina aminotransferasi (ALT) e dell’aspartato aminotransferasi (AST), solitamente asintomatici e reversibili dopo l’interruzione del trattamento con acido bempedoico. Altri eventi avversi comunemente riportati, come nasofaringite, infezioni del tratto urinario e mal di schiena, si sono verificati con frequenze simili a quelle del placebo e generalmente non sono considerati clinicamente significativi. Nel complesso, l’acido bempedoico dimostra un profilo di sicurezza favorevole, soprattutto se confrontato con le statine, grazie alla bassa incidenza di tossicità a carico della muscolatura.
L’acido bempedoico presenta un profilo di interazione farmacologica favorevole, in gran parte dovuto al fatto che il suo metabolismo è indipendente dal sistema del citocromo P450. Non è né substrato, né induttore, né inibitore degli enzimi CYP, il che consente una co-somministrazione più sicura con una varietà di farmaci cardiovascolari. Tuttavia, l’acido bempedoico interagisce con le proteine di trasporto renali ed epatiche. È un debole inibitore dei trasportatori di anioni organici OAT2 e OAT3, il che può portare a lievi aumenti dell’acido urico sierico e richiede cautela nei pazienti con gotta o disfunzione renale. L’acido bempedoico inibisce anche i trasportatori di captazione epatica OATP1B1 e OATP1B3, il che può influenzare la farmacocinetica di alcune statine. In particolare, l’esposizione alla simvastatina risulta aumentata e pertanto la sua dose dovrebbe essere limitata a un massimo di 20 mg/giorno. Altre statine (ad esempio, pravastatina, atorvastatina, rosuvastatina) possono richiedere un attento monitoraggio, soprattutto nei pazienti anziani o in quelli che assumono più farmaci.
Inibitori di PCSK9
Gli inibitori di PCSK9 sono una moderna classe di farmaci ipolipemizzanti utilizzati principalmente per la gestione della dislipidemia e delle malattie cardiovascolari aterosclerotiche (ASCVD). Questa classe farmacologica è stata sempre più integrata nella pratica clinica contemporanea, con il suo ruolo formalmente riconosciuto nelle principali linee guida internazionali come pilastro della terapia ipolipemizzante aggiuntiva nel contesto della prevenzione cardiovascolare ad alto rischio. I due principali agenti attualmente approvati per l’uso clinico sono Alirocumab ed Evolocumab. Si tratta di anticorpi monoclonali completamente umani che si legano specificamente al PCSK9 circolante, inibendone l’interazione con LDLR. Questa classe di inibitori è emersa per rispondere a importanti esigenze cliniche. Molti pazienti ad altissimo rischio di eventi cardiovascolari non raggiungono i loro obiettivi di LDL-C nonostante l’uso di statine ad alta intensità ed ezetimibe. Inoltre, alcuni pazienti sono intolleranti alle statine o manifestano effetti avversi che ne impediscono l’uso ottimale. Gli inibitori di PCSK9 offrono una soluzione efficace, sicura e ben tollerata in questi casi, con solide prove derivanti da ampi studi randomizzati a supporto dei loro benefici cardiovascolari.
La proteina PCSK9 è sintetizzata prevalentemente dagli epatociti e secreta nella circolazione sistemica. In condizioni fisiologiche, la PCSK9 si lega al recettore LDLR sulla superficie delle cellule epatiche. Dopo l’internalizzazione del complesso PCSK9-LDLR, la PCSK9 promuove la degradazione lisosomiale del recettore, riducendone così la disponibilità per il riciclo sulla superficie cellulare. Questo meccanismo determina una riduzione del numero di recettori LDLR sulla superficie degli epatociti e, di conseguenza, una minore eliminazione del colesterolo LDL dalla circolazione. Gli anticorpi monoclonali, come alirocumab ed evolocumab, inibiscono questo processo legandosi alla PCSK9 circolante, impedendone l’interazione con i recettori LDLR.
Questa preservazione dei recettori LDLR migliora il riciclo dei recettori e aumenta la clearance epatica del colesterolo LDL. Di conseguenza, il trattamento con questi agenti può ridurre i livelli di colesterolo LDL di circa il 50-70%, a seconda dei valori basali e della terapia ipolipemizzante concomitante. Questo meccanismo rappresenta un cambio di paradigma nella gestione della dislipidemia, in quanto mira a prevenire la degradazione dei recettori LDLR, piuttosto che inibire la sintesi del colesterolo, e ha dimostrato di avere un impatto clinico significativo nella riduzione del rischio cardiovascolare.
Questi farmaci sono raccomandati principalmente per individui ad alto o altissimo rischio cardiovascolare che non riescono a raggiungere i target terapeutici per l’LDL, nonostante l’uso della dose massima tollerata di statine, da sole o in combinazione con Ezetimibe. Un’indicazione chiave per gli inibitori di PCSK9 è la prevenzione secondaria delle malattie cardiovascolari aterosclerotiche in pazienti con anamnesi di infarto miocardico, ictus ischemico o arteriopatia periferica sintomatica. In questa popolazione, il mancato raggiungimento di livelli di LDL inferiori a 55 mg/dL, secondo le linee guida europee, o inferiori a 70 mg/dL, secondo le raccomandazioni americane, indica chiaramente la necessità di iniziare la terapia con inibitori di PCSK9.
Sia alirocumab che evolocumab vengono somministrati tramite iniezione sottocutanea e possono essere auto-iniettati dai pazienti dopo aver ricevuto le opportune istruzioni. Evolocumab offre due regimi di trattamento: 140 mg ogni due settimane o 420 mg una volta al mese. Entrambe le opzioni dimostrano un’equivalenza clinica nella riduzione dei livelli di LDL-C e nella protezione cardiovascolare. Il trattamento con alirocumab viene generalmente iniziato con una dose di 75 mg ogni due settimane, con la possibilità di aumentare a 150 mg ogni due settimane in base ai livelli di LDL-C e agli obiettivi terapeutici. In determinate situazioni cliniche, può essere utilizzata anche una dose mensile di 300 mg.
Gli effetti collaterali sistemici sono rari e possono includere nasofaringite, sintomi simil-influenzali e cefalea. In particolare, si osserva una bassa incidenza di sintomi muscoloscheletrici e il trattamento con inibitori di PCSK9 non è stato associato alla miopatia o alla rabdomiolisi comunemente riscontrate nei pazienti intolleranti alle statine. Inoltre, i dati di follow-up a lungo termine del trattamento con inibitori di PCSK9 non hanno confermato i timori relativi a epatotossicità, insufficienza renale o declino neurocognitivo.
Si raccomanda cautela nella somministrazione di inibitori di PCSK9 in concomitanza con agenti che influenzano la risposta immunitaria, come corticosteroidi o immunosoppressori, a causa del rischio teorico di ridotta efficacia o di una maggiore risposta immunitaria agli anticorpi monoclonali. Sebbene la combinazione di inibitori di PCSK9 con anticoagulanti o antiaggreganti piastrinici non sia strettamente vietata, è necessario procedere con cautela, poiché sono stati segnalati rari eventi emorragici in ampi studi clinici incentrati sugli esiti cardiovascolari, sebbene non sia stata stabilita una chiara relazione causale. Non vi sono prove di interazioni significative tra inibitori di PCSK9 e ipoglicemizzanti orali o farmaci antipertensivi, il che consente la loro co-somministrazione sicura. Tuttavia, in ambito clinico, è fondamentale monitorare i potenziali effetti additivi sulla funzione epatica, in particolare quando vengono utilizzati insieme più farmaci ipolipemizzanti.
Terapie con siRNA
Gli siRNA (small interfering RNA) rappresentano una classe innovativa di terapie di silenziamento genico che modulano l’espressione genica a livello post-trascrizionale attivando la via endogena dell’interferenza dell’RNA (RNAi). Nel contesto della dislipidemia, le terapie basate sugli siRNA si sono rivelate soluzioni promettenti per colpire i geni epatici essenziali coinvolti nel metabolismo lipidico. Queste terapie forniscono effetti ipolipemizzanti di lunga durata con ridotte esigenze di somministrazione. Tra queste, l’inclisiran è attualmente l’agente ipolipemizzante siRNA più avanzato e clinicamente approvato. È specificamente progettato per inibire la sintesi epatica di PCSK9, un regolatore centrale della clearance del colesterolo LDL. A differenza delle terapie ipolipemizzanti convenzionali, come le statine, l’Ezetimibe e gli anticorpi monoclonali che prendono di mira PCSK9, che agiscono a livello proteico o recettoriale, l’Inclisiran agisce a monte degradando l’RNA messaggero di PCSK9. Questo meccanismo blocca la sintesi proteica inibendo la traduzione proteica. Inclisiran esemplifica l’integrazione della farmacologia di precisione con la biotecnologia dell’RNA. Utilizza un sistema di rilascio mirato agli epatociti basato sulla coniugazione con N-acetilgalattosamina (GalNAc), garantendo specificità tissutale, maggiore potenza ed efficacia sostenuta a lungo termine.
L’inclisiran esercita il suo effetto ipolipemizzante sfruttando la via endogena dell’interferenza dell’RNA, riducendo specificamente la sintesi epatica di PCSK9. La molecola è un siRNA a doppio filamento chimicamente stabilizzato, coniugato a N-acetilgalattosamina triantennaria, che facilita l’assorbimento selettivo negli epatociti legandosi al recettore delle asialoglicoproteine (ASGPR), abbondantemente espresso sulla superficie delle cellule epatiche. Dopo l’internalizzazione, l’inclisiran si dissocia e il filamento antisenso viene incorporato nel complesso di silenziamento indotto dall’RNA (RISC). Questo complesso si lega successivamente all’mRNA complementare di PCSK9, causandone la degradazione e bloccandone la traduzione in proteine funzionali. Riducendo i livelli intracellulari di PCSK9, l’inclisiran aumenta indirettamente il numero di LDLR disponibili sulla superficie degli epatociti. Poiché PCSK9 promuove la degradazione lisosomiale dei LDLR, la sua inibizione della sintesi determina un aumento del riciclo del recettore e una maggiore eliminazione del colesterolo LDL circolante. È importante notare che questo meccanismo causa un effetto prolungato sui livelli di colesterolo LDL, con riduzioni che si mantengono per diversi mesi dopo una singola somministrazione. Questo effetto ipolipemizzante di lunga durata è spiegato meccanicisticamente dall’attività intracellulare prolungata del complesso RISC-siRNA, che consente la continua degradazione dell’mRNA di PCSK9 anche molto tempo dopo la somministrazione del farmaco, garantendo così una soppressione duratura della sintesi di PCSK9 e un’attività sostenuta dei LDLR. L’effetto farmacodinamico di Inclisiran è in gran parte indipendente dalle concentrazioni plasmatiche ed è invece sostenuto dalla persistenza intracellulare e dal turnover dell’siRNA legato al RISC negli epatociti. Questi aspetti conferiscono a inclisiran un profilo farmacologico distinto/unico, caratterizzato da un’azione lenta e da un’efficacia di lunga durata, che richiede solo due somministrazioni di mantenimento all’anno, a seguito di uno schema di carico con due dosi iniziali somministrate al giorno 0 e al mese 3.
L’inclisiran è approvato per il trattamento degli adulti con ipercolesterolemia primaria, inclusa l’ipercolesterolemia familiare eterozigote (HeFH) o la dislipidemia mista. È indicato come coadiuvante della dieta. Viene spesso utilizzato in combinazione con le statine o in associazione a statine e altre terapie ipolipemizzanti nei casi in cui gli obiettivi di LDL-C non vengano raggiunti nonostante l’uso delle dosi massime tollerate. L’inclisiran può essere preso in considerazione anche nei pazienti intolleranti alle statine o che manifestano effetti avversi significativi con altri farmaci ipolipemizzanti. Le attuali linee guida raccomandano inclisiran come farmaco di seconda o terza linea nei regimi ipolipemizzanti, in particolare quando le modifiche dello stile di vita, le statine e l’ezetimibe non riescono a raggiungere i livelli target di colesterolo LDL.
L’inclisiran viene somministrato tramite iniezione sottocutanea nell’addome, nella parte superiore del braccio o nella coscia. Il regime posologico raccomandato prevede una dose iniziale al giorno 0, seguita da una seconda dose a 3 mesi e da successive dosi di mantenimento ogni 6 mesi. La dose fissa è di 284 mg per iniezione e non è necessario alcun aggiustamento in base al peso corporeo, all’età, al sesso o a un’insufficienza epatica o renale da lieve a moderata. L’iniezione viene in genere somministrata in un contesto sanitario per garantire la corretta somministrazione e facilitare il monitoraggio del paziente. L’intervallo di dosaggio prolungato è possibile grazie alla persistenza intracellulare dell’inclisiran negli epatociti e all’attività prolungata del RISC, che continua a sopprimere la sintesi di PCSK9 per diversi mesi dopo la somministrazione.
Inclisiran si è dimostrato sicuro e ben tollerato sia negli studi clinici che nella pratica clinica reale. Nella maggior parte dei casi, le reazioni avverse sono limitate al sito di iniezione e consistono in dolore da lieve a moderato, eritema, edema o prurito. Queste manifestazioni sono transitorie, non si accumulano nel tempo e di solito si risolvono senza bisogno di intervento medico. Gli effetti collaterali sistemici sono rari. In particolare, l’inclisiran non è stato associato a tossicità muscolare, epatotossicità o alterazioni significative del controllo glicemico, effetti avversi che si osservano occasionalmente con le statine. L’aumento delle transaminasi epatiche è raro durante la terapia con inclisiran e si verifica con una frequenza simile a quella osservata nei gruppi placebo. L’effetto dell’inclisiran sull’immunogenicità sembra essere minimo, poiché ad oggi non è stato segnalato alcuno sviluppo rilevante di anticorpi neutralizzanti o reazioni di ipersensibilità durante il trattamento. È importante sottolineare che l’inclisiran non attraversa la barriera emato-encefalica e non è stato associato ad effetti neurocognitivi avversi.
L’inclisiran non viene metabolizzato tramite il sistema del citocromo P450 e non coinvolge le comuni vie di trasporto epatiche o renali, rendendo altamente improbabili interazioni farmacologiche clinicamente significative. Questo profilo farmacocinetico favorevole è attribuito al suo rilascio epatico mirato tramite coniugazione con GalNAc e alla sua degradazione da parte di nucleasi endogene in nucleotidi naturali. Pertanto, l’inclisiran non induce né inibisce gli enzimi CYP e non agisce come substrato o inibitore di trasportatori chiave come OATP1B1/1B3, BCRP o P-gp (BCRP—proteina di resistenza al cancro al seno, P-gp—glicoproteina P). L’uso concomitante con altre terapie ipolipemizzanti come le statine o l’ezetimibe non ha dimostrato alcuna interazione clinicamente significativa e la co-somministrazione sembra essere sia sicura che sinergica nella riduzione dei livelli di LDL-C. Tuttavia, come per qualsiasi nuova classe di terapie, la sorveglianza post-marketing è essenziale per rilevare eventuali interazioni farmacologiche rare o ritardate.
Lomitapide
Lomitapide è un inibitore di MTP di prima classe e anche un derivato benzimidazolico a piccola molecola, originariamente sviluppato come farmaco orfano per il trattamento dell’ipercolesterolemia familiare omozigote (HoFH). Questa rara, grave e potenzialmente letale malattia genetica è caratterizzata da livelli marcatamente elevati di colesterolo LDL fin dalla nascita e da un rischio sostanzialmente aumentato di malattie cardiovascolari aterosclerotiche precoci. Negli individui con mutazioni nulle nei geni essenziali per la funzione del recettore LDLR, come i geni LDLR, ApoB, PCSK9 o LDLRAP1, le terapie ipolipemizzanti convenzionali (ad esempio, statine ad alta intensità, ezetimibe, inibitori di PCSK9) sono spesso inadeguate, poiché dipendono dall’attività residua del recettore LDLR. Lomitapide risponde a un’esigenza terapeutica fondamentale nella gestione della dislipidemia e dell’HoFH esercitando il suo effetto ipolipemizzante indipendentemente dalla via del recettore LDLR. Agisce inibendo la MTP, un enzima chiave coinvolto nell’assemblaggio e nella secrezione delle lipoproteine contenenti ApoB sia nel fegato che nell’intestino, in particolare le lipoproteine a bassissima densità (VLDL) e i chilomicroni. Questa inibizione della MTP determina una profonda riduzione della produzione di VLDL, portando in definitiva a una significativa diminuzione dei livelli plasmatici di LDL-C e di altre lipoproteine aterogene. Studi clinici hanno dimostrato che la lomitapide, se utilizzata in combinazione con una dieta a basso contenuto di grassi e trattamenti ipolipemizzanti standard, rappresenta una valida opzione terapeutica per i pazienti con ipercolesterolemia familiare omozigote (HoFH) che non riescono a raggiungere i livelli target di LDL-C con le sole terapie convenzionali o con la sola aferesi delle lipoproteine.
La lomitapide appartiene alla classe di farmaci ipolipemizzanti noti come inibitori della MTP, una proteina chaperon intracellulare localizzata principalmente nel lume del reticolo endoplasmatico all’interno di epatociti ed enterociti. Il suo ruolo fisiologico consiste nel trasferimento di trigliceridi, esteri del colesterolo e fosfolipidi sull’ApoB durante l’assemblaggio delle particelle lipoproteiche.
Nel fegato, la MTP è essenziale per la formazione e la secrezione delle VLDL, il precursore primario delle LDL. Nell’intestino tenue, la MTP facilita l’assemblaggio dei chilomicroni, che trasportano i trigliceridi e il colesterolo assunti con la dieta. Inibendo la MTP, la lomitapide blocca il processo di lipidazione dell’ApoB, impedendo così la formazione e la secrezione intracellulare di lipoproteine contenenti ApoB. Questo blocco porta a una sostanziale riduzione della concentrazione plasmatica di VLDL e, di conseguenza, di LDL-C. È importante notare che questo meccanismo agisce indipendentemente dalla funzione del recettore LDLR, il che differenzia la lomitapide dalle statine e dagli inibitori di PCSK9, che richiedono almeno una certa attività residua di questi recettori per essere efficaci. In questo contesto, la lomitapide è particolarmente vantaggiosa per i pazienti con mutazioni nulle o gravemente difettose del gene LDLR.
Inoltre, riducendo la sintesi di particelle contenenti ApoB, la lomitapide può anche ridurre i livelli circolanti di [Lp(a)] e di lipoproteine postprandiali, entrambe considerate aterogene. L’effetto complessivo è un’ampia attenuazione del carico lipidico, che contribuisce al suo beneficio terapeutico in una popolazione ad alto rischio cardiovascolare.
La lomitapide è approvata come agente complementare alle misure dietetiche e ad altre terapie ipolipemizzanti negli adulti con ipercolesterolemia familiare omozigote (HoFH), con o senza aferesi delle LDL. Negli studi clinici, la lomitapide ha dimostrato una notevole efficacia, riducendo i livelli di LDL-C del 40-60% in modo dose-dipendente. Studi longitudinali e osservazionali condotti nella pratica clinica reale confermano la durabilità della riduzione dell’LDL-C a lungo termine. È importante notare che il trattamento con lomitapide ha permesso a molti pazienti di ridurre o addirittura interrompere le sedute di aferesi ed è stato associato a una minore frequenza di eventi cardiovascolari, nonché alla stabilizzazione dell’aterosclerosi subclinica. Questi benefici terapeutici sono più pronunciati quando la somministrazione del farmaco viene iniziata nelle fasi iniziali della malattia e integrata in una strategia ipolipemizzabile completa.
La dose iniziale raccomandata è di 5 mg una volta al giorno, da assumere a stomaco vuoto almeno due ore dopo il pasto serale per ottimizzare l’assorbimento e ridurre al minimo gli effetti collaterali gastrointestinali. L’aumento della dose deve avvenire gradualmente, in genere ogni 2-4 settimane, ed essere guidato dalla risposta del paziente al colesterolo LDL, dalla tollerabilità e dai livelli degli enzimi epatici. La tipica sequenza di titolazione prevede l’aumento della dose da 5 mg a 10 mg, 20 mg, 40 mg e fino a un massimo di 60 mg al giorno, a seconda della tollerabilità. Una dieta rigorosamente a basso contenuto di grassi è obbligatoria durante tutto il trattamento per ridurre gli effetti avversi gastrointestinali e supportare l’efficacia. Per mitigare il rischio di carenze di vitamine liposolubili e acidi grassi essenziali, è necessaria un’integrazione giornaliera con vitamina E (400 UI) e acidi grassi essenziali. L’educazione del paziente e la consulenza nutrizionale continua sono cruciali per garantire l’aderenza e promuovere la sicurezza a lungo termine.
Il profilo degli eventi avversi del lomitapide è strettamente correlato al suo meccanismo d’azione, che prevede l’inibizione dell’assorbimento lipidico e alterazioni del metabolismo lipidico epatico. Gli effetti collaterali più comunemente osservati sono di natura gastrointestinale e includono diarrea, nausea, dispepsia, flatulenza, gonfiore addominale e vomito. Questi sintomi sono in genere più pronunciati durante la fase di aumento graduale del dosaggio e tendono a correlarsi con la quantità di grassi assunti con la dieta. Pertanto, è fondamentale seguire scrupolosamente una dieta a basso contenuto di grassi per minimizzare l’intolleranza gastrointestinale. Con un’adeguata consulenza dietetica, questi effetti avversi tendono a diminuire nel tempo, migliorando la tollerabilità del trattamento a lungo termine. Sebbene non comune, il lomitapide è stato associato a casi di epatotossicità più grave, tra cui danno epatocellulare che può rendere necessaria l’interruzione temporanea o permanente della terapia.
La lomitapide viene ampiamente metabolizzata dall’enzima citocromo P450 3A4 (CYP3A4), il che spiega il suo considerevole potenziale di interazioni farmacologiche clinicamente rilevanti. La somministrazione concomitante di inibitori del CYP3A4 forti o moderati, come itraconazolo, fluconazolo o claritromicina, è severamente controindicata, poiché questi agenti possono aumentare significativamente le concentrazioni plasmatiche di lomitapide, con conseguente aumento del rischio di epatotossicità e reazioni avverse sistemiche. Un’attenta valutazione della terapia farmacologica e l’evitamento degli inibitori del CYP3A4 sono pertanto componenti essenziali per un uso sicuro della lomitapide.
Inibitori della proteina angiopoietina-simile 3
Gli inibitori di ANGPTL3 rappresentano una nuova classe di farmaci ipolipemizzanti che agiscono sul metabolismo lipidico attraverso un meccanismo distinto dalle terapie tradizionali come le statine o gli inibitori di PCSK9. Le proteine angiopoietina-simili (ANGPTL) comprendono un gruppo di proteine che include i membri 1-8 delle angiopoietine, con differenze nella regolazione tissutale e nell’espressione. Ciascuna di esse include un dominio comune all’estremità ammino-terminale, un dominio simil-fibrinogeno all’estremità C-terminale del carbossile, una regione di legame e un dominio elicoidale. ANGPTL3 è una proteina secreta dal fegato che svolge un ruolo cruciale nella regolazione dei lipidi plasmatici inibendo la lipoproteina lipasi (LPL) e la lipasi endoteliale (EL), enzimi chiave coinvolti rispettivamente nel catabolismo delle lipoproteine ricche di trigliceridi e nel metabolismo delle HDL. Studi genetici hanno dimostrato che gli individui con mutazioni con perdita di funzione nel gene ANGPTL3 presentano livelli significativamente più bassi di LDL-C, trigliceridi e HDL-C, nonché un rischio ridotto di ASCVD, evidenziando ANGPTL3 come un promettente bersaglio terapeutico.
Gli inibitori di ANGPTL3 agiscono neutralizzando l’attività di ANGPTL3, potenziando così l’attività di LPL ed EL. Le ANGPTL costituiscono una famiglia distinta di proteine con molteplici ruoli in processi biologici e patologici, tra cui la regolazione ormonale, il metabolismo del glucosio e la sensibilità all’insulina. LPL agisce come parte della classe delle lipasi presenti sulla superficie luminale dei capillari, che catabolizzano i trigliceridi plasmatici dalle lipoproteine, inclusi i chilomicroni e le VLDL. Bassi livelli di LPL causano grave ipertrigliceridemia e una diminuzione dei livelli di HDL, aumentando così il rischio di cardiopatia ischemica. Le ANGPTL rappresentano un gruppo di proteine in grado di mediare la regolazione post-traduzionale di LPL, inibendola. Pertanto, l’inibizione farmacologica di ANGPTL3 porterà a quanto segue: a. potenziamento dell’attività di LPL ed EL; b. riduzione di VLDL, LDL-C, trigliceridi e particelle contenenti ApoB; c. aumento della clearance delle lipoproteine residue. Questo meccanismo funziona indipendentemente da LDLR, il che rappresenta un vantaggio terapeutico fondamentale. L’anticorpo monoclonale Evinacumab è attualmente la terapia mirata ad ANGPTL3 più avanzata, approvata dalla FDA per l’uso in pazienti con ipercolesterolemia familiare omozigote (HoFH). A differenza delle statine o degli inibitori di PCSK9, come già accennato, l’inibizione di ANGPTL3 riduce i livelli di LDL-C indipendentemente dall’attività di LDLR, risultando particolarmente efficace nei pazienti con perdita completa o quasi completa della funzione di LDLR, come quelli affetti da HoFH.
L’evinacumab è indicato come coadiuvante di altre terapie ipolipemizzanti per i pazienti con ipercolesterolemia familiare omozigote (HoFH) che non riescono a ottenere un’adeguata riduzione del colesterolo LDL con i trattamenti convenzionali. Gli studi clinici hanno dimostrato riduzioni del colesterolo LDL di circa il 47% in questa popolazione, insieme a significative diminuzioni dei trigliceridi e del colesterolo non-HDL. L’esclusivo meccanismo indipendente dal recettore LDLR consente all’evinacumab di essere efficace laddove altri trattamenti hanno un’efficacia limitata. Sebbene l’approvazione attuale sia limitata all’HoFH, studi in corso stanno indagando il suo ruolo in popolazioni dislipidemiche più ampie, tra cui ipertrigliceridemia grave/refrattaria e dislipidemia mista. Più specificamente, i pazienti che non raggiungono il target di colesterolo LDL con statine, ezetimibe o inibitori di PCSK9 possono beneficiare di un’ulteriore riduzione del colesterolo LDL attraverso l’inibizione di ANGPTL3, in quanto possono ridurre il colesterolo LDL, i trigliceridi e il colesterolo non-HDL senza aumentare il rischio di miopatia. Le linee guida ESC/EAS del 2019 per la gestione delle dislipidemie riconoscono la necessità di terapie emergenti nei pazienti con HoFH e alto rischio residuo nonostante una terapia ottimale con statine ed ezetimibe. Gli inibitori di ANGPTL3 sono indicati come un approccio promettente in questo gruppo ad alto rischio. Allo stesso modo, il percorso decisionale di consenso degli esperti ACC del 2022 sul ruolo delle terapie non statiniche nella riduzione del colesterolo LDL identifica l’evinacumab come potenziale coadiuvante nei pazienti con HoFH o in quelli con disturbi lipidici geneticamente confermati che non riescono a raggiungere i livelli target di colesterolo LDL.
La dose raccomandata di evinacumab per i pazienti con ipercolesterolemia familiare omozigote (HoFH) è di 15 mg/kg somministrata tramite infusione endovenosa della durata di 60 minuti una volta ogni quattro settimane. Non è necessario alcun aggiustamento del dosaggio in base all’età, al sesso o alla presenza di insufficienza renale o epatica da lieve a moderata (Appendice A, Tabella A6 e Tabella A7). A causa dei dati limitati nei pazienti con grave insufficienza epatica, si consiglia cautela e monitoraggio clinico. L’aderenza del paziente alla terapia può essere favorita dalla frequenza di somministrazione relativamente bassa, che può rappresentare un vantaggio rispetto ad altri farmaci che richiedono somministrazioni più frequenti. Tuttavia, le difficoltà logistiche associate alla somministrazione endovenosa devono essere considerate nella pianificazione della gestione a lungo termine.
L’Evinacumab non viene metabolizzato dagli enzimi del citocromo P450 e non interferisce con i comuni trasportatori epatici, minimizzando così il rischio di interazioni farmacologiche. Pertanto, l’evinacumab può essere somministrato in sicurezza in concomitanza con statine, ezetimibe, inibitori di PCSK9 e altri farmaci ipolipemizzanti. Tuttavia, i dati sulle interazioni con immunosoppressori, farmaci antidiabetici o altri anticorpi monoclonali sono limitati, e si raccomanda un’attenta valutazione in caso di politerapia complessa.
Gli inibitori di ANGPTL3, come evinacumab, hanno dimostrato un profilo di sicurezza favorevole negli studi clinici. Gli eventi avversi più comunemente riportati includono nasofaringite, sintomi simil-influenzali, vertigini e reazioni correlate all’infusione. Questi eventi sono in genere di gravità da lieve a moderata. Non sono stati segnalati casi significativi di epatotossicità o effetti collaterali a carico della muscolatura, il che distingue gli inibitori di ANGPTL3 dalle statine. Tuttavia, essendo un anticorpo monoclonale che richiede la somministrazione endovenosa, evinacumab necessita di monitoraggio per ipersensibilità e complicanze correlate all’infusione, soprattutto durante le infusioni iniziali.
Conclusioni:
La gestione dell’ipercolesterolemia è entrata in un’era di trasformazione, spinta da una comprensione più approfondita del metabolismo lipidico e dall’avvento di terapie innovative che vanno ben oltre l’ambito delle statine convenzionali. In questa disamina ha sottolineato l’arsenale terapeutico in espansione, che comprende gli inibitori di PCSK9, gli agenti a base di siRNA come l’inclisiran, gli inibitori di ACL come l’acido bempedoico, gli inibitori di MTP e gli inibitori di ANGPTL3, ognuno dei quali offre meccanismi d’azione unici, profili di efficacia distinti e considerazioni di sicurezza specifiche.
Nel loro insieme, queste strategie emergenti non solo forniscono valide alternative per i pazienti con ipercolesterolemia familiare o intolleranza alle statine, ma rappresentano anche strumenti promettenti per mitigare il rischio cardiovascolare residuo persistente in popolazioni complesse ad alto rischio. La loro integrazione in algoritmi di trattamento individualizzati, guidati dalla farmacogenomica, da una stratificazione del rischio più precisa e dalle preferenze del paziente, segna un cambio di paradigma verso una gestione lipidica più precisa e personalizzata.
E’ corretto sottolineare, arrivati a questo punto che, nonostante il ventaglio di applicazioni terapeutiche per il trattamento delle dislipidemie/ipercolesterolemia si sia largamente ampliato, l’uso di farmaci come gli inibitori di PCSK9 ed i siRNA, per fare due esempi di una certo significato, per le pratiche di Harm Reduction negli atleti enhanced risultano costosi ed eccessivi; in definitiva non necessari alla luce della causa radicale della dislipidemia/ipercolesterolemia in tale contesto. Infatti, visti i meccanismi alla base della alterazione del Colesterolo in ambito d’uso di AAS [viste nella I° Parte], le terapie applicabili, se i soggetti non hanno mutazioini geniche che li obbligherebbero a percorrere vie più complesse, sono:
Statine [per es., in specie, Simvastatina, Atorvastatina o Rosuvastatina]; la Monacolina K può essere un ripiego in caso di inaccessibilità alle Statine di sintesi.
In caso di intolleranza alle Statine [manifestazione statistica del 9% nei pazienti trattati] optare potenzialmente per lAcido Bempedoico
In caso di mancato accesso ai suddetti farmaci e alla intolleranza alle Statine, il Bergamotto può essere un buon ripiego; alcuni flavonoidi unici del Bergamotto (come Melitidina e Brutieridina) presentano una struttura molecolare simile all’HMG-CoA. Questo permette loro di legarsi e inibire l’enzima HMG-CoA reduttasi, riducendo la sintesi endogena di Colesterolo nel fegato con un meccanismo analogo a quello dei farmaci Statinici ma senza i tipici sides connessi.
In caso di alimentazione con una significativa presenza di Colesterolo alimentare e/o necessità di un controllo ancillare marcato della dislipidemia, l’aggiunta di Ezetimibe può migliorare ulteriormente il controllo del profilo lipidico ematico; l’uso di 1-2g di Lecitina di Soia possono risultare un discreto sostituto.
L’inserimento di Niacina [500mg-1.5mg/die] per periodi controllati riducono “il paradosso della Niacina” migliorando il tamponamento della riduzione delle HDL-C; qual ora possibile, quest’ultima può essere sostituita con l’Acipimox.
L’aggiunta di 2,5-5gm di EPA e DHA possono aitare a ridurre i livelli di Trigliceridi e migliorare lo stato infiammatorio.
In passato mi sono già cimentato nella trattazione del Colesterolo, soprattutto in ambito Enhanced, esplicando in modo chiaro i meccanismi che portano l’utilizzatore di AAS in una condizione dislipidemica/ipercolesterolemica potenzialmente negativa specie se mantenuta nel tempo. In questo artricolo, invece, è mia intenzione approfondire le caratteristiche di questa molecola tanto vituperata quando mal intesa al fine di far conoscere al lettore non semplicemente le basi di biochimica che reggono la sua omeostasi ma, soprattutto, quali sono ad oggi i trattamenti farmacologici applicati per intervenire sulle alterazione del Colesterolo ematico.
Il Colesterolo, un lipide strutturale e di segnalazione indispensabile, è fondamentale per l’integrità delle membrane cellulari, la steroidogenesi e le vie metaboliche dei morfogeni durante lo sviluppo. La sua omeostasi dipende dal preciso coordinamento di quattro moduli metabolici interdipendenti: biosintesi de novo, assorbimento intestinale, conversione enzimatica ed eliminazione sistemica. In questo articolo tratteremo i meccanismi molecolari che regolano questi processi, dalle vie di sintesi di Bloch/Kandutsch-Russell e dall’assorbimento di colesterolo mediato da NPC1L1 (Niemann-Pick C1-like 1) alla sintesi degli acidi biliari guidata dalla colesterolo 7α-idrossilasi (CYP7A1) e al trasporto inverso dipendente dalle HDL. Vengono inoltre approfonditi i molteplici ruoli del colesterolo nell’assemblaggio dei lipid raft, nella trasduzione del segnale Hedgehog e nella produzione di vitamina D e ormoni.
La disregolazione del flusso di Colesterolo è alla base della patogenesi dell’aterosclerosi, della steatosi epatica associata a disfunzione metabolica (MAFLD), delle malattie neurodegenerative e dell’oncogenesi, con cascate di sintesi, efflusso o esterificazione alterate che fungono da fattori chiave. Le strategie terapeutiche emergenti vanno oltre le statine convenzionali e gli inibitori della proproteina convertasi subtilisina/kexina di tipo 9 (PCSK9) per includere modalità innovative: l’editing genetico in vivo basato su CRISPR (ad esempio, VERVE-101 che ha come bersaglio PCSK9), le terapie con RNA interferente (siRNA) (inclisiran) e gli interventi mirati al microbiota. Approcci pionieristici contro bersagli quali l’angiopoietina-simile 3 (ANGPTL3), la lipoproteina(a) [Lp(a)] e il recettore 1 dell’asialoglicoproteina (ASGR1), insieme ad agenti naturali riproposti (berberina, probiotici), offrono promettenti soluzioni per mitigare il rischio cardiovascolare residuo e promuovere la medicina cardiometabolica di precisione. Integrando le conoscenze sui meccanismi d’azione con i progressi clinici, andrà sottolineata la transizione dalle terapie ad ampio spettro a regimi personalizzati multi-bersaglio, offrendo una tabella di marcia per la mitigazione delle malattie correlate al Colesterolo nell’era della medicina genomica e metabolica.
Questa disamina si dividerà in due parti al fine di dare la possibilità al lettore di assimilare le nozioni e poter essere preparato alle successive presenti della seconda parte.
Regolazione dell’omeostasi del Colesterolo
L’omeostasi del colesterolo è meticolosamente orchestrata attraverso l’integrazione di quattro moduli metabolici chiave: sintesi, assorbimento, conversione ed eliminazione (vedi la figura seguente). Questo ciclo metabolico completo garantisce che il Colesterolo svolga i suoi ruoli fondamentali nella costruzione delle membrane, nella trasduzione del segnale e nell’omeostasi sistemica.
Vie metaboliche intracellulari della biosintesi, dell’assorbimento e della conversione del Colesterolo. La via biosintetica de novo del Colesterolo ha origine dall’acetil-CoA e procede attraverso intermedi chiave come HMG-CoA, Mevalonato, farnesil-PP e squalene, formando infine il Colesterolo tramite il Lanosterolo. Gli enzimi HMGCR e SQLE sono evidenziati come fasi critiche limitanti la velocità. I destini metabolici a valle del Colesterolo comprendono la sua conversione in ormoni steroidei, acidi biliari e vitamina D, nonché l’immagazzinamento in goccioline lipidiche come CE tramite ACAT, o il trasporto dentro e fuori dalle cellule attraverso recettori lipoproteici (LDLR, SR-BI) e trasportatori (ABCA1, ABCG1, ABCG5/8). Viene inoltre illustrato l’assorbimento del Colesterolo dal lume intestinale tramite NPC1L1 e il suo efflusso verso le particelle HDL, riassumendo le principali vie di flusso intracellulare del Colesterolo. Farnesil-PP, farnesil pirofosfato; HMG-CoA, 3-idrossi-3-metilglutaril-CoA; HMGCR, HMG-CoA reduttasi; SQLE, squalene epossidasi; CE, esteri del colesterolo; ACAT, acil-CoA:colesterolo aciltransferasi; SR-BI, recettore scavenger di classe B tipo I; NPC1L1, niemann-pick C1-like 1
Biosintesi del Colesterolo
La sintesi de novo del Colesterolo nei vertebrati avviene prevalentemente nel fegato (80%), con contributi minori dai tessuti extraepatici (20%). Questo processo è metabolicamente impegnativo, consumando 18 molecole di adenosina trifosfato (ATP) e 16 molecole di nicotinammide adenina dinucleotide (NAD) in forma ridotta (NADPH), oltre ad acetil-CoA e ossigeno, per ogni molecola di colesterolo prodotta. Procede attraverso due vie metaboliche conservate nel design: la via canonica di Bloch (> 90% della produzione totale), chiarita dagli studi premiati con il Nobel da Konrad Bloch negli anni ’50, e la via di Kandutsch-Russell (K-R), scoperta negli anni ’60. La via K-R è dominante in condizioni di ipossia o stress da raggi ultravioletti (UV) in tessuti come la pelle e le gonadi. Entrambi i percorsi metabolici condividono le stesse fasi iniziali, a partire dalla condensazione delle molecole di acetil-CoA. La tiolasi combina due molecole di acetil-CoA per formare acetoacetil-CoA, al quale la 3-idrossi-3-metilglutaril-CoA (HMG-CoA) sintasi aggiunge una terza molecola di acetil-CoA, producendo HMG-CoA. L’enzima limitante la velocità di reazione, l’HMG-CoA reduttasi (HMGCR), riduce quindi l’HMG-CoA a Mevalonato (MVA) utilizzando NADPH. L’MVA subisce fosforilazione e isomerizzazioni sequenziali per produrre farnesil pirofosfato (FPP). La squalene sintasi (SQS) dimerizza l’FPP in squalene, che viene successivamente ossidato dal secondo enzima limitante la velocità di reazione, la squalene epossidasi (SQLE), per formare 2,3-ossidosqualene. La lanosterolo sintasi (LSS) ciclizza il 2,3-ossidosqualene in lanosterolo, il primo intermedio steroideo. Oltre il lanosterolo, la via di Bloch e la via di Krebs-Ribbent-Robinson (K-R) divergono. Nella via di Bloch, il lanosterolo subisce demetilazione, desaturazione e riduzione per produrre infine Colesterolo. Nella via di K-R, il lanosterolo viene convertito sequenzialmente in 24,25-diidrolanosterolo e 7-deidrocolesterolo (7-DHC), seguito dalla riduzione del 7-DHC a Colesterolo tramite la 7-DHC reduttasi (DHCR7). Sebbene meno efficiente, la via di K-R evita le fasi dipendenti dall’ossigeno, fornendo un vantaggio fisiologico in ambienti con limitata disponibilità di ossigeno.
Assorbimento del Colesterolo
L’assorbimento del Colesterolo, che avviene principalmente nel duodeno e nel digiuno prossimale, è un processo strettamente regolato, fondamentale per il mantenimento dell’omeostasi lipidica sistemica, garantendo un assorbimento efficiente del Colesterolo alimentare e biliare e prevenendone l’accumulo eccessivo. Il principale mediatore dell’assorbimento intestinale del colesterolo è la proteina di membrana degli enterociti niemann-pick C1-like 1 (NPC1L1), che si sposta tra la superficie cellulare e i compartimenti di riciclo endocitico (ERC) per facilitarne l’assorbimento. NPC1L1 contiene cinque domini transmembrana, tra cui un dominio di rilevamento degli steroli che rileva i livelli di Colesterolo . Un elevato livello di colesterolo luminale induce l’integrazione del colesterolo libero nella membrana degli enterociti, dove NPC1L1 lo lega e ne facilita l’internalizzazione. Questo avviene principalmente tramite endocitosi mediata da clatrina/AP2, trasportando il Colesterolo lungo i filamenti di actina verso gli ERC per l’immagazzinamento. Studi di microscopia crioelettronica rivelano un meccanismo aggiuntivo: il legame del Colesterolo induce un cambiamento conformazionale in NPC1L1, formando un tunnel di trasporto transmembrana che facilita direttamente l’assorbimento indipendentemente dall’endocitosi. In condizioni di basso Colesterolo, NPC1L1 viene riciclato verso la membrana per riprendere l’assorbimento.
Una volta internalizzato, il Colesterolo subisce esterificazione o efflusso per bilanciare i livelli cellulari. Il colesterolo libero si sposta nel reticolo endoplasmatico, dove l’acil-CoA:colesterolo aciltransferasi 2 (ACAT2) lo esterifica con acidi grassi in esteri del Colesterolo. Questi esteri idrofobici vengono impacchettati nei chilomicroni, trasportati all’apparato di Golgi per essere elaborati e infine entrano nella circolazione sistemica attraverso il sistema linfatico (dotto toracico) per essere distribuiti ai tessuti periferici. Nel frattempo, il Colesterolo libero in eccesso viene attivamente pompato di nuovo nel lume intestinale dai trasportatori eterodimerici ATP-binding cassette (ABC) G5/G8 (ABCG5/G8). Questo meccanismo di efflusso limita in modo critico l’assorbimento netto e protegge dal sovraccarico cellulare.
Conversione del Colesterolo
All’interno delle cellule, il Colesterolo funge da precursore versatile per la sintesi di composti biologicamente essenziali, tra cui acidi biliari, glucosidi del Colesterolo, vitamina D e vari ormoni steroidei come Androgeni, Estrogeni, Progesterone, Glucocorticoidi e Mineralcorticoidi.
Sintesi degli Acidi Biliari
Nel fegato, la maggior parte del Colesterolo viene convertita in Acidi Biliari, costituiti principalmente da Acidi Biliari primari come l’Acido Colico (CA) e l’Acido Chenodesossicolico (CDCA), insieme ad acidi biliari secondari come l’Acido Desossicolico (DCA) e tracce di Acido Litocolico (LCA), attraverso due percorsi distinti. Il percorso classico, responsabile di oltre il 90% della produzione di acidi biliari, inizia con la Colesterolo 7α-idrossilasi (CYP7A1) che idrossila il Colesterolo in posizione 7α. Questa fase limitante la velocità produce 7α-idrossicolesterolo, che viene poi convertito in 7α-idrossi-4-colesten-3-one (C4) dalla 3β-idrossi-Δ5-C27-steroide deidrogenasi (3β-HSD). In particolare, il C4 funge da precursore comune sia per il CA che per il CDCA ed è spesso utilizzato come biomarcatore sierico per la velocità di sintesi degli acidi biliari. Al contrario, la via alternativa agisce come un meccanismo compensatorio essenziale durante l’alterazione della via classica o lo stress metabolico, inizia con la sterolo 27-idrossilasi mitocondriale (CYP27A1) che converte il Colesterolo in 27-idrossicolesterolo (27-OHC), seguita dall’ossisterolo 7α-idrossilasi (CYP7B1) che catalizza un’ulteriore idrossilazione per generare acido 3β,7α-diidrossi-5-colestenoico, che subisce ossidazione della catena laterale e accorciamento tramite 3β-HSD tipo 7 (HSD3B7) per produrre CDCA. Sebbene questa via alternativa rappresenti solo circa il 10% della sintesi degli acidi biliari in condizioni normali, diventa di fondamentale importanza negli stati patologici.
Glucosidi del Colesterolo (CG)
I CG sono glicosidi sterolici in cui una porzione di Glucosio è esterificata al gruppo idrossilico del Colesterolo. I CG contribuiscono alla stabilità della membrana alterando l’impacchettamento e la fluidità dei lipidi, in particolare nei raft lipidici. I CG possono anche agire come molecole di segnalazione nelle risposte immunitarie. Ad esempio, sono implicati nell’attivazione dei macrofagi e nella produzione di citochine. Nelle piante, i CG difendono dai patogeni microbici, suggerendo ruoli analoghi nell’immunità innata dei mammiferi. Nei mammiferi, i CG vengono sintetizzati tramite glicosilazione enzimatica del colesterolo principalmente nell’apparato di Golgi o nel reticolo endoplasmatico, dove il colesterolo e l’UDP-glucosio sono accessibili. L’UDP-glucosio:sterolo glucosiltransferasi (USG) catalizza il trasferimento del glucosio dall’UDP-glucosio al gruppo 3β-idrossilico del colesterolo. La produzione di CG è regolata dalla disponibilità di colesterolo e dallo stress cellulare; in condizioni di sovraccarico di colesterolo o stress ossidativo, la sintesi di CG può aumentare per modulare la fluidità della membrana o sequestrare il Colesterolo in eccesso. La disregolazione della sintesi di CG è collegata a disturbi dell’accumulo di lipidi e malattie infiammatorie. Ad esempio, livelli elevati di CG si osservano nella malattia di Niemann-Pick di tipo C, una malattia da accumulo lisosomiale. È interessante notare che alcuni organismi patogeni, come Helicobacter pylori, sono anche in grado di sintetizzare glucosidi del colesterolo, sebbene i loro percorsi biosintetici differiscano fondamentalmente da quelli dei mammiferi.
Sintesi della Vitamina D
La vitamina D, un secosteroide fondamentale per l’omeostasi del Calcio e la regolazione immunitaria, viene sintetizzata a partire dal 7-DHC attraverso un percorso dipendente dai raggi UV e successive modifiche enzimatiche. Nello strato basale e spinoso dell’epidermide, il 7-DHC, un intermedio nella sintesi del Colesterolo, viene convertito in pre-vitamina D3 in seguito all’esposizione alle radiazioni UVB (290-315 nm). Questa reazione non enzimatica avviene spontaneamente. La pre-vitamina D3 subisce un’isomerizzazione dipendente dalla temperatura in colecalciferolo (vitamina D3) nell’arco di circa 48 ore. La vitamina D3 viene idrossilata in due fasi sequenziali per acquisire l’attività biologica. Nel fegato, il citocromo P450 2R1 (CYP2R1) converte la vitamina D3 in 25-idrossivitamina D3 [25(OH)D3], la principale forma circolante. Nel rene, il citocromo P450 27B1 (CYP27B1) idrossila la 25(OH)D3 in 1,25-diidrossivitamina D3 [1,25(OH)2D3], che aumenta l’assorbimento intestinale di Calcio e fosfato e promuove il riassorbimento renale.
Biosintesi steroidea
Il Colesterolo, precursore universale di tutti gli ormoni steroidei, inclusi mineralcorticoidi, glucocorticoidi e ormoni sessuali, subisce una biotrasformazione tessuto-specifica all’interno della corteccia surrenale. Questo sito di sintesi primario è organizzato in tre zone: la zona glomerulare, che produce mineralcorticoidi (ad esempio, Aldosterone) per regolare l’equilibrio elettrolitico e la pressione sanguigna; la zona fascicolata, che sintetizza glucocorticoidi (ad esempio, Cortisolo) che regolano la risposta allo stress e il metabolismo; e la zona reticolare, che genera steroidi sessuali (ad esempio, Androgeni) essenziali per la fisiologia riproduttiva.
Gli ormoni sessuali (androgeni, estrogeni e progestinici) sono regolatori essenziali dello sviluppo riproduttivo, delle caratteristiche sessuali secondarie e del dimorfismo fisiologico. L’attività 17,20-liasi del CYP17A1 converte il 17α-idrossipregnenolone in deidroepiandrosterone (DHEA), precursore universale degli steroidi gonadici. Il metabolismo del DHEA diverge attraverso l’ossidazione mediata da 3β-HSD-1 ad androstenedione o la riduzione dipendente da 17β-HSD-1 a 5-androstenediolo, entrambe convergenti nel testosterone. La successiva elaborazione enzimatica da parte dell’aromatasi (CYP19A1/P450aro) o della 5α-reduttasi produce rispettivamente 17β-estradiolo o 5α-diidrotestosterone (5α-DHT).
Eliminazione del Colesterolo
L’omeostasi del Colesterolo nei mammiferi si basa su meccanismi di eliminazione strettamente regolati per prevenire l’accumulo patologico nei tessuti. Due principali vie complementari mediate dalle lipoproteine, tra cui il trasporto inverso del Colesterolo (RCT) guidato dalle lipoproteine ad alta densità (HDL) e l’assorbimento epatico dipendente dalle lipoproteine a bassa densità (LDL), sono essenziali per il mantenimento dell’omeostasi del Colesterolo. Le HDL rimuovono il Colesterolo in eccesso dai tessuti periferici, mentre le LDL e il loro recettore LDLR assicurano un efficiente assorbimento epatico del colesterolo circolante. Le strategie terapeutiche che prendono di mira queste vie, come gli agenti che potenziano LDLR o HDL, sono promettenti per il trattamento della dislipidemia e dell’aterosclerosi.
Trasporto del Colesterolo mediato dalle HDL
Il trasporto inverso del colesterolo (RCT) è un processo critico attraverso il quale il colesterolo viene trasportato dai tessuti periferici al fegato per essere escreto tramite la bile o le feci. Un componente principale del percorso RCT è l’HDL. Le particelle HDL nascenti hanno una forma discoidale e sono composte da apolipoproteina A1 (ApoA1) e fosfolipidi [24]. Le particelle HDL acquisiscono colesterolo libero dalle cellule schiumose periferiche, come i macrofagi o le cellule muscolari lisce vascolari, tramite i trasportatori ABC A1 (ABCA1) e G1 (ABCG1). ABCA1 interagisce con l’apoA1 povera di lipidi, mentre ABCG1 facilita l’efflusso di colesterolo verso l’HDL matura. La lecitina-colesterolo aciltransferasi (LCAT) esterifica il colesterolo libero all’interno dell’HDL, convertendolo in esteri del colesterolo. Questo nucleo idrofobico trasforma l’HDL in particelle sferiche. L’HDL matura trasporta il colesterolo al fegato attraverso l’assorbimento selettivo mediato dal recettore scavenger di classe B tipo I (SR-BI) o il trasferimento facilitato dalla proteina di trasferimento degli esteri del colesterolo (CETP). L’SR-BI estrae selettivamente gli esteri del colesterolo dall’HDL senza degradare l’intera particella, mentre la CETP trasporta gli esteri del colesterolo dall’HDL alle lipoproteine contenenti apolipoproteina B (ApoB) (ad esempio, LDL, lipoproteine a bassissima densità (VLDL)), che vengono successivamente eliminate tramite LDLR epatico.
Eliminazione del Colesterolo mediata dalle LDL
Le particelle LDL, che trasportano esteri del Colesterolo e apolipoproteina B-100 (ApoB-100), vengono eliminate prevalentemente dal fegato. I recettori LDLR, altamente espressi negli epatociti, legano l’ApoB-100 presente sulle particelle LDL tramite interazioni elettrostatiche. I complessi LDL-LDLR vengono internalizzati in vescicole rivestite di clatrina, che si fondono con i lisosomi. All’interno dei lisosomi, gli esteri del colesterolo vengono idrolizzati a colesterolo libero, mentre i recettori LDLR vengono riciclati sulla superficie cellulare. In alcuni casi, le particelle HDL che esprimono apolipoproteina E (ApoE) possono legarsi ai recettori LDLR o alla proteina 1 correlata al recettore LDL (LRP1), consentendo la loro internalizzazione e degradazione epatica. Questa via metabolica fornisce un percorso ausiliario per l’eliminazione del Colesterolo HDL.
La proproteina convertasi subtilisina/kexina di tipo 9 (PCSK9) è un membro della famiglia delle serina proteasi, nota per il suo ruolo critico nell’attivazione proteolitica, nella modificazione e nella degradazione delle proteine secrete. Gli LDLR epatici fungono da recettori per la PCSK9 circolante. La PCSK9 si lega al dominio EGF-A (epidermal growth factor-like repeat A) degli LDLR tramite il suo dominio catalitico, promuovendo l’endocitosi degli LDLR negli endosomi. Il pH acido all’interno degli endosomi aumenta l’interazione PCSK9-LDLR di 150 volte, impedendo il riciclo degli LDLR. Di conseguenza, il complesso PCSK9-LDLR viene indirizzato alla degradazione lisosomiale, riducendo la densità degli LDLR sulla superficie degli epatociti, diminuendo così la clearance delle particelle di colesterolo LDL (LDL-C) epatiche e aumentando i livelli circolanti di LDL-C. L’inibizione di PCSK9 migliora la clearance del Colesterolo LDL, una strategia terapeutica per l’ipercolesterolemia.
Funzione biologica del Colesterolo
Il Colesterolo è una molecola multifunzionale con un’ampia gamma di ruoli biologici. Le sue funzioni spaziano dal mantenimento dell’integrità e della fluidità delle membrane cellulari al ruolo di precursore di ormoni essenziali e acidi biliari. Inoltre, il Colesterolo svolge un ruolo cruciale nella regolazione del metabolismo lipidico, nella modulazione delle vie di trasduzione del segnale e nell’influenza sulla funzione immunitaria. Comprendere i diversi ruoli del Colesterolo è fondamentale per apprezzarne l’importanza nel mantenimento della salute generale e per sviluppare strategie terapeutiche volte a contrastare i disturbi correlati all’alterazione del Colesterolo.
Ruoli biologici del Colesterolo. Il Colesterolo è una molecola multifunzionale che svolge ruoli essenziali in diversi processi biologici. È un componente chiave della membrana cellulare, rappresentando circa il 20-30% dei lipidi di membrana. Il Colesterolo è fondamentale per il mantenimento dell’integrità e della fluidità della membrana. Un doppio strato fosfolipidico puro passa da una fase gel (rigida) a una fase liquido-cristallina (fluida) a livello della sua membrana transmembrana (TM), e il colesterolo modula dinamicamente questa transizione per ottimizzare le proprietà della membrana. Oltre ai suoi ruoli strutturali, il colesterolo è un regolatore centrale del metabolismo lipidico e un precursore di ormoni essenziali. Modula inoltre le vie di trasduzione del segnale attraverso molteplici meccanismi. I microdomini ricchi di Colesterolo, noti come zattere lipidiche (Lipid rafts), fungono da piattaforme per l’organizzazione e l’attivazione delle molecole di segnalazione. Ad esempio, il Colesterolo interagisce direttamente con la proteina transmembrana PTCH1, che inibisce strutturalmente la SMO. Questa interazione consente un controllo spazio-temporale preciso della via di segnalazione canonica Hedgehog, fondamentale per lo sviluppo, l’immunità e le malattie. Il Colesterolo regola bidirezionalmente la funzione dei canali ionici attraverso meccanismi sia strutturali che allosterici. Nello specifico, aumenta la funzione del canale Kir3.4 e al contempo riduce quella dei canali Kir2.1 e Kir3.1. Queste interazioni evidenziano i diversi ruoli del Colesterolo nella segnalazione cellulare e nell’omeostasi. PTCH1, patched1; SMO, smoothened; TM, temperatura di fusione. Questa figura è stata creata tramite BioRender.
Tra le principali funzioni biologiche del Colesterolo troviamo:
Colesterolo e stabilità della membrana cellulare
Il Colesterolo è un componente chiave delle membrane cellulari eucariotiche, rappresentando circa il 20-30% dei lipidi di membrana. Un doppio strato fosfolipidico puro esiste in una fase gel (rigida) al di sotto della sua temperatura di transizione (TM) e in una fase liquido-cristallina (fluida) al di sopra di Tₘ . Intercalandosi tra le catene fosfolipidiche, il colesterolo può smussare il punto di flesso netto della transizione di fase e contribuire a tamponare il cambiamento di fase, stabilizzando così la fase liquido-cristallina in un intervallo di temperature più ampio. Pertanto, quando l’ambiente è vicino o al di sopra della temperatura di transizione, il Colesterolo limita i movimenti vigorosi, in particolare i movimenti laterali, delle catene fosfolipidiche, prevenendo così un’eccessiva fluidità della membrana. Al contrario, a temperature più basse, l’anello sterolico del colesterolo ostacola la disposizione ordinata delle catene fosfolipidiche, impedendo la formazione di uno stato gel rigido.
L’architettura anfifilica del Colesterolo consente una duplice funzione regolatrice. Da un lato, l’anello steroideo idrofobico si inserisce nelle code idrofobiche dei fosfolipidi, associandosi alle catene di acidi grassi saturi per aumentare la densità della membrana. Riempiendo gli spazi tra le catene fosfolipidiche, il colesterolo riduce i difetti della membrana e ne migliora la resistenza meccanica. Dall’altro lato, la porzione flessibile della catena alchilica del colesterolo può interagire con le code idrofobiche delle catene di acidi grassi insaturi, diminuendo l’impacchettamento disordinato causato dalle loro pieghe e prevenendo un’eccessiva fluidità.
Inoltre, il Colesterolo collabora con gli sfingolipidi per formare i domini lipidici (lipid raft), microdomini spazialmente segregati caratterizzati da un elevato grado di ordine pur mantenendo un certo livello di fluidità, noti come fase liquido-ordinata (Lo). Al contrario, le regioni circostanti, ricche di fosfolipidi insaturi, presentano un’elevata fluidità che facilita la diffusione delle sostanze e la deformazione della membrana, e sono note come fase liquido-disordinata (Ld). Regolando l’assemblaggio e il disassemblaggio dei lipid raft, il Colesterolo influenza indirettamente la fluidità delle regioni non-raft. Le variazioni di fluidità di questi lipid raft possono anche attivare proteine di membrana come le tirosin chinasi recettoriali, influenzando così l’efficienza della trasduzione del segnale.
Modulazione dei canali ionici tramite interazioni con il Colesterolo
Il Colesterolo regola bidirezionalmente la funzione dei canali ionici attraverso meccanismi strutturali e allosterici. Le alterazioni dei livelli di Colesterolo possono influenzare la funzione dei canali ionici. Ad esempio, è noto che i canali del potassio a rettificazione interna (Kir) sono influenzati da tali cambiamenti. Nello specifico, il Colesterolo aumenta la funzione di Kir3.4 mentre riduce le funzioni di Kir2.1 e Kir3.1. Un fattore che contribuisce a queste differenze è l’alterazione della distribuzione del Colesterolo. Rispetto a Kir2.1, il potenziale sito di legame del Colesterolo di Kir3.4 ha subito delle alterazioni. L’accumulo preferenziale di colesterolo nell’elica transmembrana distale è accoppiato allostericamente con la dinamica conformazionale a livello del filtro di selettività. Questo accoppiamento allosterico tra la funzione del canale e il legame dei lipidi è un meccanismo necessario per l’attivazione dei canali Kir mediata da PIP [109].
Il Colesterolo come precursore universale della sintesi degli ormoni steroidei
Come già accennato in precedenza, il Colesterolo funge da precursore fondamentale per tutti gli ormoni steroidei, operando come impalcatura molecolare per questi regolatori critici dei processi di sviluppo, dell’omeostasi metabolica e dell’adattamento allo stress. La steroidogenesi impiega due sistemi enzimatici conservati nei tessuti endocrini: le ossidasi del citocromo P450 (CYP) e le idrossisteroide deidrogenasi (HSD), nonostante i modelli di espressione specifici delle ghiandole. Gli enzimi CYP contengono domini di legame all’eme conservati, mentre le HSD sono prive di eme ma richiedono i cofattori NAD e NADP. La cascata steroidogenica inizia con la fase limitante: la traslocazione del colesterolo alle membrane interne mitocondriali mediata dalla proteina regolatrice acuta steroidogenica (StAR). All’interno dei mitocondri, l’enzima di scissione della catena laterale del citocromo P450 (CYP11A1/P450scc) catalizza la conversione del colesterolo in pregnenolone, il precursore universale degli ormoni steroidei.
Il Colesterolo come precursore della sintesi degli ormoni steroidei. La sintesi degli ormoni steroidei inizia con la traslocazione del Colesterolo nei mitocondri mediata da StAR. All’interno della membrana mitocondriale interna, l’enzima di scissione della catena laterale del citocromo P450 CYP11A1 catalizza la conversione del colesterolo in Pregnenolone, il precursore universale di tutti gli ormoni steroidei. (A sinistra) Biosintesi dell’Aldosterone. Il pregnenolone viene convertito in Progesterone tramite 3β-HSD nel reticolo endoplasmatico liscio. La successiva idrossilazione in posizione C21 mediata da CYP21A2 genera 11-deossicorticosterone, che subisce modifiche sequenziali da parte di CYP11B1 e dell’Aldosterone sintasi CYP11B2 per produrre corticosterone e infine aldosterone. (Al centro) Produzione di cortisolo. La biosintesi del cortisolo inizia con l’idrossilazione in posizione 17α del pregnenolone, mediata dal CYP17A1, a 17α-idrossipregnenolone, seguita dalla catalisi sequenziale attraverso HSD3B2, CYP21A2 e CYP11B1 per generare rispettivamente 17α-Idrossiprogesterone, 11-deossicortisolo e cortisolo. (A destra) Sintesi degli ormoni sessuali. L’attività 17,20-liasi del CYP17A1 converte il 17α-idrossipregnenolone in DHEA, che si differenzia attraverso l’ossidazione mediata da 3β-HSD-1 ad Androstenedione o la riduzione dipendente da 17β-HSD-1 a 5-androstenediolo, entrambe convergenti nel Testosterone. La successiva elaborazione enzimatica da parte dell’Aromatasi (CYP19A1) o della 5α-reduttasi produce rispettivamente 17β-estradiolo o 5α-DHT. StAR, proteina regolatrice acuta della steroidogenesi; 3β-HSD, 3β-idrossisteroide deidrogenasi; DHEA, Deidroepiandrosterone; 5α-DHT, 5α-diidrotestosterone
Colesterolo e mTOR
Una funzione biologica poco conosciuta che interessa il Colesterolo è che esso rappresenta un elemento essenziale per l’attivazione della via metabolica di mTOR. La presenza di Colesterolo libero a livello delle membrane cellulari e il suo corretto trasporto intracellulare sono infatti necessari per permettere l’assemblaggio e l’attivazione del complesso mTORC1, fondamentale per la crescita e la sintesi proteica. L’mTORC1 è un regolatore primario dei processi anabolici. Aumenta i livelli di nSREBP2 fosforilando e impedendo l’ingresso nucleare di Lipin 1. Viceversa, il fattore di trascrizione lipogenico ChREBP (proteina legante l’elemento di risposta ai carboidrati) promuove l’ubiquitinazione e la degradazione proteasomica di nSREBP2 attraverso un meccanismo sconosciuto. Il digiuno attiva la deacetilazione di SREBP2 mediata da SIRT1, arrestando il processo di biosintesi del colesterolo, che consuma energia, in condizioni di carenza di nutrienti. Oltre all’acetilazione, nSREBP2 può subire fosforilazione da parte di ERK e AMPK, portando rispettivamente a un aumento o a una diminuzione dell’attività trascrizionale. Anche la sumoilazione di nSREBP2 riduce la sua attività trascrizionale. AMPK è stata identificata come una chinasi a monte di LXR. L’AMPK attivata (AMPK fosforilata) inibisce la produzione di ligandi LXR endogeni, riducendo così l’espressione di LXR e bloccando la regolazione trascrizionale mediata da LXR.
L’mTOR agisce come un regolatore di molteplici stimoli cellulari. I principali attivatori includono:
1- Nutrienti: Aminoacidi, in particolare la leucina, e il glucosio. 2- Segnali ormonali: Insulina e fattori di crescita (come l’IGF-1). 3- Stimoli meccanici: Sforzi fisici intensi e allenamento contro resistenza (ipertrofia).
Malattie associate al metabolismo del Colesterolo
Come abbiamo visto, il metabolismo del Colesterolo è caratterizzato da un complesso equilibrio dinamico. La disregolazione dell’omeostasi del Colesterolo, caratterizzata da squilibri nella sintesi, nell’assorbimento, nel trasporto o nell’escrezione, rappresenta un fattore patogenetico critico per una serie di malattie croniche e degenerative. Queste condizioni interessano i sistemi cardiovascolare, epatico, neurologico e oncologico, riflettendo l’impatto sistemico del dismetabolismo del Colesterolo.
Malattie associate al metabolismo del Colesterolo. La disregolazione del metabolismo del Colesterolo può portare a diverse patologie. I. Le mutazioni genetiche nel gene LDLR riducono la quantità e la funzionalità dell’LDLR, compromettendo l’assorbimento cellulare di LDL-C e determinando livelli elevati di LDL-C nel plasma, una condizione nota come ipercolesterolemia familiare (FH). II. Il deposito di LDL ossidate sulle pareti arteriose innesca la fagocitosi da parte dei macrofagi, portando alla formazione di cellule schiumose macrofagiche e allo sviluppo di placche aterosclerotiche. III. La sovrasaturazione del Colesterolo nella bile può portare alla sua precipitazione e cristallizzazione nella cistifellea, con conseguente formazione di calcoli biliari. IV. L’aumentata sintesi di colesterolo, associata a una ridotta secrezione di VLDL da parte degli epatociti, porta all’accumulo di lipidi, potenzialmente causando steatosi epatica non alcolica (NAFLD). V. Livelli elevati di Colesterolo sono stati osservati in individui affetti da malattia di Alzheimer. L’eccesso di colesterolo può ostacolare l’attività dell’enzima responsabile della scissione dell’Aβ, esacerbandone l’accumulo intracellulare e peggiorandone la progressione. VI. Le anomalie nel metabolismo del colesterolo sono collegate alla tumorigenesi. Le cellule tumorali spesso mostrano livelli di Colesterolo aumentati, che correlano con la disregolazione dell’HMGCR, la sovraespressione dell’LDLR, l’iperattività dell’ACAT, il metabolismo anomalo del 27-OHC e l’attivazione persistente della via SREBP. FH, ipercolesterolemia familiare; Aβ, beta-amiloide; HMGCR, HMG-CoA reduttasi; ACAT, acil-CoA:Colesterolo aciltransferasi; 27-OHC, 27-idrossicolesterolo; SREBP, proteina legante l’elemento regolatore degli steroli. Questa figura è stata creata tramite BioRender.
Ipercolesterolemia
L’ipercolesterolemia, una dislipidemia clinicamente significativa caratterizzata da livelli cronicamente elevati di Colesterolo totale plasmatico (≥ 5,2 mmol/L [200 mg/dL]) e/o livelli di LDL-C, costituisce un importante fattore di rischio modificabile per le malattie cardiovascolari aterosclerotiche. Classificata secondo le Linee guida cinesi del 2024 e 2026 per la gestione dei lipidi nel sangue e allineata con i criteri dell’American College of Cardiology, questa patologia metabolica si manifesta in due forme eziologiche principali: primaria (ereditaria) e secondaria. L’ipercolesterolemia primaria deriva prevalentemente da difetti genetici, in particolare dall’ipercolesterolemia familiare (FH), una condizione autosomica dominante guidata principalmente da varianti patogene nel gene LDLR. Queste mutazioni compromettono la clearance dell’LDL-C, con conseguente accumulo patologico di LDL-C. L’ipercolesterolemia secondaria deriva da fattori acquisiti, tra cui comorbilità (ad esempio, coronaropatia, diabete mellito), farmaci (ad esempio, diuretici, beta-bloccanti, glucocorticoidi, AAS) e fattori legati allo stile di vita. In particolare, l’obesità amplifica il rischio attraverso la disregolazione del metabolismo lipidico e l’iperlipidemia, mentre l’eccessivo consumo di grassi saturi, la sedentarietà, il consumo cronico di alcol e lo stress psicosociale aggravano ulteriormente l’alterazione dell’omeostasi del Colesterolo.
Data la sua natura multifattoriale, l’ipercolesterolemia può essere inizialmente gestita attraverso interventi sullo stile di vita mirati a fattori estrinseci modificabili. Le attuali strategie di intervento includono: 1) la modifica della composizione e della struttura della dieta, limitando il consumo di cibi ad alto contenuto di grassi e riducendo l’assunzione di Colesterolo esogeno. La ricerca indica che la dieta DASH, specificamente progettata per prevenire e gestire l’ipertensione, è efficace nel ridurre i livelli di LDL-C. Inoltre, i risultati di Luiza et al. suggeriscono che l’adesione alla dieta mediterranea può anche migliorare la dislipidemia negli individui affetti. 2) Impegnarsi in un’attività fisica regolare per migliorare i processi metabolici, migliorando così il metabolismo lipidico anomalo; e 3) Praticare un consumo moderato di alcol per mitigare l’assunzione complessiva di alcol. Quando le modifiche dello stile di vita non producono una risposta sufficiente, diventa imperativo intensificare la farmacoterapia. Gli agenti di prima linea includono statine, inibitori dell’assorbimento del Colesterolo, inibitori di PCSK9, sequestranti degli acidi biliari e fibrati. Queste modalità terapeutiche, con i loro distinti meccanismi e applicazioni cliniche, saranno analizzate in modo esaustivo nelle sezioni successive.
Aterosclerosi
L’aterosclerosi (AS) è una malattia immunoinfiammatoria cronica del sistema arterioso, caratterizzata patologicamente dall’accumulo di placche ricche di lipidi all’interno delle pareti dei vasi, che porta a stenosi luminale e ridotta compliance vascolare. Colpisce principalmente le arterie di grandi e medie dimensioni ed è guidata da una varietà di fattori di rischio, tra cui la dislipidemia, in particolare i livelli elevati di LDL, costituisce il principale determinante patogeno. Le particelle di LDL circolanti migrano attraverso l’endotelio e subiscono una modificazione ossidativa (ad esempio, coniugazione con malondialdeide), formando LDL ossidate pro-infiammatorie (oxLDL). Le oxLDL attivano i recettori scavenger dei macrofagi (ad esempio, CD36, LOX-1), dando inizio a un ciclo autosostenuto di formazione di cellule schiumose. I macrofagi internalizzano le oxLDL ma non possono esportare efficacemente il colesterolo a causa della disfunzione delle HDL. In condizioni di ipercolesterolemia, il sistema RCT, mediato dal legame dell’HDL ai recettori epatici SR-B1, viene sovraccaricato. Ciò promuove lo sviluppo di un nucleo necrotico e di una capsula fibrosa, che sono i tratti distintivi delle placche vulnerabili. Pertanto, il metabolismo del Colesterolo disregolato, in particolare la ridotta clearance lipidica dovuta a livelli elevati di LDL e a una disfunzione dell’HDL, funge da meccanismo centrale nella patogenesi dell’AS, guidando l’intero processo dal danno endoteliale alla formazione della placca. Sebbene l’ipercolesterolemia rimanga il principale fattore di rischio modificabile, altri fattori contribuenti includono ipertensione, diabete mellito, obesità e fattori legati allo stile di vita come il fumo.
Malattia del fegato grasso associata a disfunzione metabolica (MAFLD)
La MAFLD è una condizione epatica cronica caratterizzata dall’accumulo di lipidi nel fegato (steatosi) in assenza di altre cause identificabili, come disturbi genetici o consumo eccessivo di alcol. Lo spettro della malattia spazia dalla semplice steatosi epatica (FL), alla steatoepatite non alcolica (NASH), alla fibrosi e alla cirrosi. La sua progressione è guidata da molteplici fattori interagenti, tra cui predisposizione genetica, influenze ambientali, disbiosi del microbiota intestinale, stress ossidativo, insulino-resistenza, infiammazione e dislipidemia, che possono agire in parallelo o in sequenza nelle diverse fasi della malattia. Il Colesterolo è riconosciuto come il principale agente lipotossico nella patogenesi della MAFLD, e la sua disregolazione gioca un ruolo centrale nella progressione della MAFLD verso la NASH e la fibrosi. Un’eccessiva assunzione e accumulo di Colesterolo promuovono la progressione della malattia e le risposte infiammatorie indotte dal Colesterolo sono state identificate come un fattore determinante in queste condizioni.
La gestione della MAFLD comprende strategie sia non farmacologiche che farmacologiche, adattate ai profili di rischio individuali. Modifiche dietetiche, come la dieta mediterranea [143] e la dieta chetogenica, sono opzioni terapeutiche efficaci. Anche l’esercizio fisico regolare dimostra effetti benefici nel trattamento della MAFLD. L’asse intestino-fegato gioca un ruolo importante, poiché la salute intestinale influenza significativamente la funzione epatica, rendendo il microbiota intestinale un potenziale bersaglio terapeutico. Ad esempio, i probiotici possono modulare la composizione del microbiota intestinale ed esercitare effetti benefici sulla MAFLD, e alcune caratteristiche della flora potrebbero fungere da biomarcatori diagnostici o prognostici [147]. Tuttavia, uno studio del 2021 condotto da Nor et al. ha suggerito che sei mesi di integrazione probiotica non hanno prodotto un miglioramento clinico significativo, indicando che i probiotici potrebbero essere più adatti come terapia aggiuntiva piuttosto che come terapia autonoma. Il primo trattamento farmacologico per la NASH è Rezdiffra, un agonista orale del THR-β che attiva selettivamente il recettore epatico dell’ormone tiroideo β. Modula il metabolismo lipidico, promuove il dispendio energetico e riduce il grasso epatico e l’infiammazione. Altri approcci farmacologici mirano alla patogenesi sottostante, tra cui agenti ipolipemizzanti (ad esempio, statine), agonisti del PPARα (ad esempio, acido obeticolico) e agonisti dell’FXR (ad esempio, cilofexor ed elafibranor).
Calcoli biliari
I calcoli biliari sono depositi cristallini solidi che si formano nella cistifellea o nelle vie biliari e rappresentano una comune patologia gastrointestinale. Si distinguono principalmente in due tipi: calcoli di colesterolo e calcoli pigmentati. La patogenesi della calcolosi biliare è multifattoriale e coinvolge una complessa interazione di predisposizioni genetiche, fattori legati allo stile di vita e altre influenze. Il meccanismo predominante alla base della formazione dei calcoli biliari è la sovrasaturazione di colesterolo nella bile, che favorisce lo sviluppo dei calcoli di colesterolo. In condizioni fisiologiche, il colesterolo è solubilizzato all’interno di micelle e vescicole formate da acidi biliari e lecitina. Tuttavia, quando i livelli di Colesterolo superano la capacità di solubilizzazione di questi composti, si verifica la sovrasaturazione e la conseguente cristallizzazione. In particolare, studi recenti suggeriscono che la sovrasaturazione di colesterolo nei pazienti con calcoli biliari potrebbe essere dovuta principalmente a una carenza di acidi biliari piuttosto che a un’eccessiva produzione di colesterolo.
Le strategie di trattamento per i calcoli biliari includono interventi farmacologici e procedure chirurgiche. L’Acido Ursodesossicolico (UDCA) è un farmaco standard per i calcoli biliari di Colesterolo, poiché promuove la dissoluzione dei calcoli aumentando la secrezione di acidi biliari e sopprimendo la sintesi epatica di Colesterolo, riducendo così la saturazione di Colesterolo nella bile. Anche le Statine sono emerse come potenziale trattamento grazie alla loro capacità di modulare il metabolismo del Colesterolo e diminuire la formazione di calcoli biliari. Secondo Georgescu et al., le statine possono inoltre agire attraverso la modulazione del microbiota intestinale. La medicina tradizionale cinese contribuisce ulteriormente alla gestione dei calcoli biliari. Ad esempio, Huang et al. hanno riportato che i polisaccaridi di Ganoderma lucidum alleviano la formazione di calcoli biliari di Colesterolo attraverso la regolazione dipendente da FXR del metabolismo del Colesterolo e degli acidi biliari. Altre formulazioni a base di erbe, come i granuli Shugan Lidan Xiaoshi (SLXG), hanno anche dimostrato un potenziale terapeutico prendendo di mira geni tra cui HMGCR, SOAT2 e UGT1A1, modulando così l’omeostasi del Colesterolo. Inoltre, prove sempre più numerose indicano il ruolo del microbiota intestinale nella prevenzione e nel trattamento dei calcoli biliari di colesterolo. Ad esempio, Wang et al. hanno dimostrato che l’integrazione di lattobacilli ha ridotto l’incidenza e la gravità dei calcoli biliari indotti da una dieta ricca di grassi, indicando una nuova promettente direzione terapeutica.
Malattie neurodegenerative
Il cervello umano contiene la più alta concentrazione di Colesterolo tra tutti gli organi, rappresentando circa il 25% del Colesterolo totale del corpo. Questo pool strettamente regolato è essenziale per il mantenimento dell’architettura neuronale, della plasticità sinaptica e della neurotrasmissione, con prove emergenti che collegano il metabolismo del Colesterolo disregolato alle malattie neurodegenerative. Separato dalla circolazione sistemica dalla barriera emato-encefalica (BBB), il Colesterolo cerebrale opera come un sistema metabolico autonomo. Gli astrociti sono la fonte primaria di Colesterolo cerebrale, producendolo attraverso la sintesi de novo e consegnandolo ai neuroni tramite il trasporto mediato da ApoE. Il Colesterolo in eccesso subisce tre destini regolatori: (1) accumulo come goccioline lipidiche citoplasmatiche, (2) efflusso mediato dai trasportatori ABC tramite particelle legate ad ApoA1, o (3) conversione enzimatica in ossisteroli (24-idrossicolesterolo [24-OHC] e 27-OHC) per l’eliminazione attraverso la BBB.
La malattia di Alzheimer (AD), una patologia neurodegenerativa prototipica, è caratterizzata dalla deposizione di placche di beta-amiloide (Aβ) e da grovigli neurofibrillari. L’aumento dei livelli di Colesterolo nelle membrane neuronali è correlato alla progressione dell’AD [160, 161]. L’ipercolesterolemia promuove la produzione di Aβ facilitando la processazione della proteina precursore dell’amiloide (APP) all’interno dei lipid raft ricchi di colesterolo tramite le β- e γ-secretasi. L’allele ApoE4 (ε4), il più forte fattore di rischio genetico per l’AD ad esordio tardivo, compromette il trasporto del Colesterolo e la clearance dell’Aβ. In particolare, le dinamiche degli ossisteroli mostrano ruoli opposti: il 27-OHC aggrava la produzione di Aβ e la progressione dell’AD, mentre il 24-OHC promuove l’attività dell’α-secretasi per ridurre il carico di Aβ, esercitando effetti neuroprotettivi [159].
Il limitato successo clinico delle attuali terapie mirate all’Aβ ha spostato l’attenzione verso interventi sul metabolismo del Colesterolo. Le statine, tradizionalmente utilizzate per la dislipidemia, mostrano un potenziale nella gestione dell’AD. Una meta-analisi di 55 studi osservazionali ha indicato che la terapia con statine è associata a una riduzione del 14% del rischio di demenza e a una diminuzione del 18% dell’incidenza dell’AD. Questi benefici possono derivare dall’inibizione della sintesi del Colesterolo e dalla ridotta aggregazione di Aβ, in particolare con statine lipofile che attraversano la barriera emato-encefalica come la Simvastatina. Prove emergenti evidenziano la rilevanza terapeutica del CYP46A1, un enzima neuronale specifico che converte il colesterolo in 24-OHC. La sovraespressione di CYP46A1 migliora la cognizione nei modelli murini e la sua modulazione farmacologica si dimostra promettente: ad esempio, KaiXinSan regola positivamente CYP46A1 per alleviare i deficit cognitivi indotti da 27-OHC, mentre il farmaco anti-HIV efavirenz agisce come modulatore dipendente dalla concentrazione di CYP46A1. Nuovi analoghi di efavirenz progettati specificamente per colpire l’attività di CYP46A1 offrono un ulteriore potenziale per il trattamento dell’AD . Insieme, questi risultati stabiliscono la regolazione del metabolismo del colesterolo come una promettente frontiera terapeutica nella patogenesi dell’AD.
Potenziale impatto del Colesterolo nella patologia della malattia di Alzheimer. Il metabolismo del Colesterolo nel cervello segue una complessa sequenza di eventi. Inizialmente, il colesterolo viene sintetizzato all’interno del reticolo endoplasmatico degli astrociti. Successivamente si associa all’APOE per formare i granuli di APOE-colesterolo. Questi granuli vengono secreti nel fluido extracellulare, un processo facilitato dalle proteine trasportatrici ABC. In seguito, il Colesterolo viene internalizzato dai neuroni che esprimono i recettori LDLR. All’interno delle cellule neuronali, il colesterolo segue tre distinti percorsi metabolici. Una parte del Colesterolo viene convertita in goccioline lipidiche per l’immagazzinamento intracellulare, mentre un’altra frazione viene rilasciata dalla cellula e si combina con l’ApoA1 per l’escrezione diretta. La maggior parte del Colesterolo viene metabolizzata dal CYP46A1, con conseguente produzione di 24-OHC. Questo metabolita può attraversare la barriera emato-encefalica ed entrare nel plasma, dove facilita l’afflusso di 27-OHC nel cervello. ABC, cassetta legante ATP; 24-OHC, 24-idrossicolesterolo; 27-OHC, 27-idrossicolesterolo; CYP46A1, colesterolo 24-idrossilasi. Questa figura è stata creata tramite BioRender.
Tumori
Prove emergenti identificano il metabolismo del Colesterolo disregolato come un segno distintivo metabolico critico del cancro. Rispetto alle cellule normali, le cellule maligne mostrano profonde alterazioni nell’omeostasi del Colesterolo, caratterizzate da quattro modifiche chiave: (1) iperattivazione della biosintesi de novo del Colesterolo, (2) aumento dell’assorbimento mediato da LDLR, (3) alterazione dell’efflusso di colesterolo e (4) accumulo patologico di derivati del Colesterolo. Studi comparativi indicano che i tessuti tumorali presentano livelli di colesterolo significativamente più elevati rispetto ai tessuti normali adiacenti, il che è correlato a una maggiore aggressività e a un maggiore potenziale metastatico. Ciò è supportato da una marcata sovraregolazione dei geni coinvolti nella biosintesi del Colesterolo, come SREBP2, HMGCR, SQS, OSC e SQLE, insieme a un’elevata espressione di LDLR e a una maggiore esterificazione del Colesterolo.
Le attuali strategie antitumorali che prendono di mira il metabolismo del Colesterolo si concentrano principalmente sull’inibizione della biosintesi, del trasporto e dell’esterificazione. Le Statine, inibitori classici dell’HMGCR, mostrano un’attività antitumorale ad ampio spettro nei tumori della prostata, dello stomaco, dell’esofago e della mammella. Agiscono riducendo i livelli di Colesterolo cellulare e sopprimendo la sintesi degli isoprenoidi (ad esempio, geranilgeranil pirofosfato (GGPP) e FPP), interrompendo così la funzione GTPasi nelle cellule tumorali. Tuttavia, l’uso a lungo termine delle Statine (> 4 anni) è stato associato a un aumento del rischio di diversi tumori, tra cui quelli del colon, della vescica e del polmone. Oltre agli effetti ipocolesterolemizzanti, le Statine esercitano un’azione antitumorale attraverso molteplici meccanismi: inducono l’apoptosi tramite l’attivazione di FOXO3a e l’inibizione dell’autofagia nei tumori del cavo orale e del colon, innescano la ferroptosi tramite l’interruzione della via del mevalonato, promuovono la piroptosi e modulano il microambiente tumorale. Clinicamente, le statine mostrano effetti sinergici se combinate con le terapie convenzionali, migliorando gli esiti nei pazienti con carcinoma squamocellulare dell’esofago sottoposti a chemioradioterapia e superando la resistenza ai farmaci nel carcinoma polmonare a piccole cellule recidivante. Per ridurre gli effetti collaterali come la miopatia, sono in fase di sviluppo Statine nanoformulate con un targeting tumorale migliorato.
Il fattore di trascrizione SREBP2 è un regolatore critico della sintesi del Colesterolo. La sua attivazione tramite isomerizzazione mediata dalla prolil isomerasi fosfato-dipendente (PIN1) promuove l’accumulo di Colesterolo oncogenico e il silenziamento di PIN1 riduce i livelli di colesterolo cellulare, mostrando un potenziale terapeutico nel cancro alla vescica. Anche l’inibizione del trasporto del Colesterolo attraverso il blocco del rilascio lisosomiale di Colesterolo o il targeting di NPC1 con agenti come l’itraconazolo sopprime efficacemente la crescita tumorale. Inoltre, l’esterificazione del Colesterolo attraverso ACAT/SOAT1 rappresenta un nodo vulnerabile, con inibitori come l’Avasimibe che esercitano potenti effetti antitumorali in modelli di melanoma. Le evidenze cliniche mostrano un accumulo patologico di esteri del colesterolo nel cancro al pancreas, al colon-retto e alla prostata metastatica, dove l’inibizione di ACAT1 riduce sostanzialmente la progressione e le metastasi.
Sebbene il targeting del metabolismo del Colesterolo abbia un ampio potenziale antitumorale, sempre più evidenze suggeriscono la necessità di approcci terapeutici specifici per tipo di cancro. Le strategie attuali includono l’inibizione della sintesi, l’interruzione del trasporto e il blocco dell’esterificazione, ognuna delle quali richiede un’applicazione personalizzata in base alle dipendenze metaboliche del tumore. Le prospettive future prevedono lo sviluppo di terapie di precisione che combinino il targeting metabolico con trattamenti convenzionali e sistemi di somministrazione avanzati come la nanotecnologia.
AAS e dislipidemia
Gli steroidi anabolizzanti-androgeni (AAS) causano, a diverso grado dipendente dalla natura strutturale di sintesi della molecola, una significativa dislipidemia aterogena alterando il metabolismo del Colesterolo. In genere, riducono sensibilmente l’HDL-C dal 20% al 70% e aumentano l’LDL-C dal 30% al 50%, oltre a causare un aumento dei Trigliceridi ematici. Queste anomalie delle lipoproteine possono aumentare il rischio di cardiopatia coronarica da tre a sei volte.
AAS e riduzione dell’HDL
Diversi studi interventistici hanno esaminato l’effetto dell’uso di AAS sul colesterolo. Peter Bond ha fatto un piccolo riassunto di questi studi nel suo libro “Book on Steroids” il quale riporto nella tabella sottostante. Sebbene non tutti gli studi abbiano riscontrato una diminuzione statisticamente significativa del colesterolo HDL (↔️), molti lo fanno e nel complesso mostrano inequivocabilmente una diminuzione. Ciò è particolarmente vero per gli AAS orali, che sembrano avere l’effetto maggiore sul colesterolo HDL.
Si ritiene che gli AAS riducano l’HDL aumentando l’attività di un enzima chiamato lipasi epatica. Si tratta di un enzima prodotto principalmente dal fegato. Essendo una lipasi, catalizza le reazioni di idrolisi dei lipidi. In particolare, scinde gli acidi grassi dal triacilglicerolo (Trigliceride) e i fosfolipidi dalle particelle lipoproteiche, come l’HDL. Idrolizzando il triacilglicerolo e i fosfolipidi dall’HDL, riduce le dimensioni di queste particelle. Queste particelle più piccole vengono catabolizzate a un ritmo più elevato.
Thompson et al. hanno esaminato queste sottofrazioni di colesterolo HDL che differiscono per dimensioni. Hanno misurato i livelli di colesterolo HDL2 e HDL3: le particelle di colesterolo HDL2 sono più grandi e di densità inferiore rispetto a quelle HDL3. Gli uomini partecipanti hanno ricevuto 200mg di Testosterone Enantato alla settimana o 6mg di Stanozololo orale (Winstrol) al giorno per 6 settimane in un design crossover. I risultati sono stati i seguenti:
*Differenza significativa (P < 0,05) rispetto al valore basale.
Come si può notare, la maggiore diminuzione relativa è stata osservata nella frazione HDL2 più grande a seguito del trattamento con Stanozololo. Al contrario, il Testosterone non ha mostrato una diminuzione statisticamente significativa nella frazione HDL2, ma ha fatto altrettanto nella frazione HDL3 più piccola. Non è del tutto chiaro cosa provochi la diminuzione di questa frazione.
Quando dei bodybuilder sono stati randomizzati a ricevere 200mg di Nandrolone Decanoato alla settimana o un placebo. Non sono stati riscontrati cambiamenti statisticamente significativi nel colesterolo totale, nel colesterolo LDL e nel colesterolo HDL. Analogamente, non sono stati riscontrati cambiamenti significativi nelle sottofrazioni di colesterolo HDL2 e HDL3. In particolare, nella stessa pubblicazione, gli autori riferiscono anche di uno studio in cui hanno seguito un gruppo di atleti di forza che si autosomministravano steroidi anabolizzanti. Sono stati utilizzati diversi composti in vari dosaggi, ma vale la pena sottolineare che la maggior parte di essi comprendeva anche uno steroide anabolizzante orale (soprattutto Stanozololo). In questo caso, il colesterolo HDL è sceso in picchiata: da 1,08 mmol/L a 0,43 mmol/L dopo 8 settimane. La sottofrazione di colesterolo HDL2 è scesa da 0,21 a 0,05 e la sottofrazione di colesterolo HDL3 è scesa da 0,87 a 0,40 mmol/L.
AAS e funzione dell’HDL
Dato il legame tra l’effetto di un farmaco sui livelli di HDL e il rischio di malattie cardiovascolari, la ricerca ha iniziato a concentrarsi sulla funzione del HDL. L’HDL è il protagonista di un processo chiamato trasporto inverso del colesterolo. Nell’aterosclerosi, il colesterolo si accumula nelle cellule del sistema immunitario (macrofagi) e nelle cellule muscolari lisce che circondano i vasi sanguigni. Queste cellule, a loro volta, diventano le cosiddette cellule schiumose, che segnano il punto di partenza dell’aterosclerosi. Le particelle di HDL sono in grado di raccogliere il colesterolo da queste cellule – efflusso di Colesterolo. L’efflusso del colesterolo dalle cellule schiumose nelle particelle di HDL è uno dei modi in cui si ritiene che il HDL eserciti i suoi effetti protettivi sulle arterie. Il HDL raccolto può poi essere riportato al fegato, che lo incorpora nella bile e può quindi essere secreto nelle feci. Allo stesso modo, le particelle di HDL possono trasferire parte del loro contenuto alle particelle LDL, che possono finire nuovamente nelle cellule schiumose o essere assorbite dal fegato.
Esistono metodi per misurare la capacità di efflusso del HDL e l’idea attuale è che la sua modulazione possa influire sul rischio di malattie cardiovascolari, contrariamente ai livelli di HDL in sé. Esistono diversi modi in cui l’HDL può assorbire il Colesterolo dalle cellule schiumose. Uno di questi coinvolge un trasportatore chiamato ATP-binding casette transporter A1 (ABCA1), che si ritiene sia il più importante. Contribuiscono anche altri trasportatori, come ABCG1 e il recettore scavenger B1, oltre alla diffusione semplice.
In uno studio (non controllato), uomini anziani ipogonadici sono stati randomizzati alla TRT con o senza Dutasteride (un inibitore della 5a-reduttasi). Dopo 3 mesi, la TRT era riuscita a riportare i livelli di Testosterone di questi uomini all’interno del range di normalità. L’HDL e la capacità di efflusso del HDL sono rimasti inalterati.
Un altro studio, randomizzato e controllato, ha applicato un approccio leggermente diverso. Uomini sani, di età compresa tra i 19 e i 55 anni, sono stati castrati medicalmente per sopprimere completamente la loro produzione endogena. In seguito, hanno ricevuto un placebo, una TRT a basso dosaggio, una TRT sostitutiva completa o una TRT sostitutiva completa con Letrozolo, un inibitore dell’Aromatasi che inibisce la conversione del Testosterone in Estradiolo. L’HDL è aumentato leggermente nel gruppo placebo e in quello a basso dosaggio, mentre è rimasto inalterato nei due gruppi che hanno ricevuto una dose sostitutiva completa. Inoltre, mentre è stata riscontrata una piccola diminuzione della capacità di efflusso di ABCA1 nel gruppo che ha ricevuto anche il Letrozolo, non sono stati osservati cambiamenti negli altri tre gruppi. A causa delle dimensioni ridotte dei gruppi, è possibile che un piccolo effetto non sia stato notato.
Esiste un solo studio che ha esaminato questo aspetto, ed era di natura trasversale (si tratta di misurazioni effettuate in un solo momento, il che rende impossibile/difficile trarre conclusioni). I ricercatori hanno confrontato le misurazioni di un gruppo di utilizzatori di AAS con quelle di non utilizzatori e controlli sedentari, che avevano un’età corrispondente. I consumatori di AAS erano forti utilizzatori, avendo fatto uso di AAS in media per circa 8 anni con un dosaggio medio di (apparentemente) 2,5g settimanali. La capacità delle HDL di effluire il Colesterolo dai macrofagi è risultata inferiore del 13% nei consumatori di AAS rispetto ai non consumatori con allenamento della forza. Anche in questo caso, a causa della natura trasversale dello studio, è difficile dire se questo sia causale.
Effetti clinici principali:
Riduzione dell’HDL: le riduzioni possono essere estreme, soprattutto con gli AAS non aromatizzabili come lo Stanozololo o l’Oxandrolone.
Aumento dell’LDL: dosi soprafisiologiche di AAS sovrastimolano le lipasi epatiche, alterando la sintesi delle apolipoproteine e accelerando la circolazione delle particelle di LDL.
Alterazione del eflusso del efflusso del HDL: dosi soprafisiologiche di AAS alterano l’efflusso del HDL contribuendo nel lungo termine al deposito di placche aterosclerotiche.
Trigliceridi: livelli elevati sono comunemente osservati anche durante l’uso attivo di AAS.
Insorgenza e reversibilità:
Insorgenza: Le alterazioni del profilo lipidico si manifestano rapidamente, spesso entro le prime 9 settimane di somministrazione di AAS, anche se con l’uso di alcuni AAS orali metilati in C-17 l’insorgenza di una alterazione lipidica può manifestarsi entro 3-4 settimane.
Reversibilità: Gli studi dimostrano che i profili lipidici sono in gran parte reversibili e tendono a normalizzarsi entro 2,5-5 mesi dalla completa interruzione dell’uso di AAS.
Conseguenze a lungo termine:
Sebbene i marcatori lipidici in genere tornino ai valori basali al termine dei cicli, l’uso cronico e ripetuto accelera l’accumulo di placche aterosclerotiche, aumentando il rischio di infarto miocardico precoce e ictus ischemico.
I medici spesso raccomandano pannelli lipidici a digiuno completi, test di funzionalità epatica e monitoraggio cardiovascolare per le persone con una storia di abuso di AAS.
La ricerca scientifica è in continuo sviluppo, e nonostante abbia scritto un corposo articolo sugli incretino-mimetici qualche tempo fa, l’avanzare dei dati disponibili su nuove molecole appartenenti a questa famiglia, ormai largamente in uso in campo medico e off-label, mi spingono a scrivere questo nuovo articolo in tema.
Introduzione:
E’ risaputo che l’intervento attraverso la modifica dello stile di vita rimane la strategia terapeutica di prima linea per il trattamento dell’obesità. In particolare, l’adesione alla dieta Mediterranea — un modello alimentare ricco di frutta, verdura, cereali integrali, olio d’oliva e proteine magre — ha dimostrato miglioramenti significativi nella steatosi epatica e nell’insulino-resistenza tra i pazienti con MAFLD. Inoltre, è stato dimostrato che programmi di esercizio fisico strutturati, che integrano sia l’allenamento aerobico che quello contro-resistenza, riducono il contenuto di grasso nel fegato e migliorano il controllo glicemico. Tuttavia, nonostante i benefici ben documentati dei cambiamenti nello stile di vita, mantenere l’aderenza a lungo termine rimane una sfida per molte persone, che spesso richiede un supporto multidisciplinare e strategie di cambiamento comportamentale a lungo termine. Quando necessario, gli interventi farmacologici sono essenziali per colmare il divario e garantire una prevenzione e una gestione efficaci di queste condizioni correlate.
Numerosi trattamenti farmacologici mirano a interrompere queste connessioni dannose, e molti di essi si concentrano sulle vie ormonali delle incretine e sulla regolazione dell’appetito, come già visto nel precedente articolo. Tre dei più comuni ormoni peptidici incretinici sono il Peptide Glucagone Simile-1 (GLP-1), il Polipeptide Insulinotropico glucosio-dipendente (GIP) e il Glucagone. L’emergere degli agonisti del recettore del GLP-1, come il primo agonista clinicamente approvato, l’Exenatide, nel 2005, ha aperto la strada a un’approfondita esplorazione di queste molecole. In breve tempo sono state introdotte altre molecole, come la Liraglutide, la Dulaglutide e la Semaglutide, che offrono una somministrazione settimanale. Oltre a garantire un controllo più efficace del glucosio e a favorire una significativa perdita di peso, questi farmaci hanno consentito una riduzione dei tassi di gravi effetti avversi cardiovascolari, insufficienza cardiaca, malattie renali e morte cardiovascolare. Questi risultati promettenti hanno portato allo sviluppo dei primi agonisti duali, tra cui spicca la Tirzepatide, un agonista dei recettori GIP e GLP-1. La Tirzepatide ha ottenuto riduzioni significative dei livelli di emoglobina glicata A1c (HbA1c) e del peso corporeo rispetto al placebo. Inoltre, è stato riscontrato che la Tirzepatide ha un effetto più pronunciato rispetto all’agonista del GLP-1 Semaglutide nelle persone con DMT2. L’incidenza di eventi avversi gastrointestinali (GI) è aumentata rispetto al placebo; tuttavia, né la Tirzepatide né il Semaglutide hanno aumentato il rischio di eventi avversi gravi o di ipoglicemia grave.
La Retatrutide, un agonista triplo recettoriale, mira a rivoluzionare la farmacoterapia dell’obesità e del diabete di tipo 2. Questo farmaco innovativo ha dimostrato di migliorare i risultati terapeutici, determinando riduzioni più marcate sia del peso corporeo che dei livelli di HbA1c rispetto alle attuali terapie farmacologiche.
-Recettori Molecolari Bersaglio-
Peptide Glucagone Simile-1 (GLP-1)
Il GLP-1 è un ormone incretinico secreto prevalentemente dal tratto intestinale, soprattutto in risposta ai pasti. Le cellule L dell’intestino tenue sono responsabili della sua produzione a partire da un gene del proglucagone. È stato inoltre dimostrato che la produzione di questa molecola avviene anche nelle cellule α del pancreas e nel sistema nervoso centrale, in aree quali il nucleo del tratto solitario e la microglia. Agisce prevalentemente attivando un recettore che induce la stimolazione del rilascio di insulina, diminuisce la secrezione di glucagone, riduce l’assunzione di cibo e ritarda lo svuotamento gastrico (GE) influenzando la motilità gastrica e intestinale. I recettori del GLP-1 si trovano in una varietà di tessuti quali il pancreas, la mucosa gastrica, i reni, i polmoni, il cuore, la pelle, le cellule immunitarie e il cervello. Si tratta di recettori accoppiati alle proteine G eptelicoidi composti da 463 aminoacidi che, una volta attivati, stimolano la formazione di adenosina monofosfato ciclico (cAMP) seguita dall’attivazione delle vie della proteina chinasi A.
In particolare per quanto riguarda il sistema nervoso centrale, l’espressione dei recettori del GLP-1 in regioni correlate alla gratificazione, quali l’ipotalamo, l’amigdala, il nucleo accumbens, il nucleo paraventricolare, l’area tegmentale ventrale, il locus coeruleus e il tronco encefalico, potrebbe suggerire una possibile associazione tra l’apprendimento della gratificazione e sintomi quali l’anedonia in patologie quali la depressione. Tuttavia, la natura multifattoriale e poligenica di questa diagnosi richiede ulteriori indagini prima di giungere a una conclusione definitiva. È stato inoltre dimostrato che l’attivazione del GLP-1 riduce l’infiammazione diminuendo l’attività dei macrofagi. I suoi effetti sulla sazietà e sull’assunzione di cibo si riscontrano sia in individui sani che in pazienti obesi e diabetici.
Secreto anche dalle cellule K intestinali, il GIP è una proteina secreta prevalentemente in fase postprandiale. È costituito da 42 aminoacidi che si formano in seguito alla proteolisi di un precursore di 153 aminoacidi. I recettori del GIP esistono in due isoforme, costituite rispettivamente da 466 e 493 aminoacidi, che, una volta attivate, mostrano un aumento del calcio intracellulare e dell’acido arachidonico. La loro attivazione è correlata anche all’attivazione dell’adenilato ciclasi. Agendo su questi recettori, espressi in diversi tessuti, tra cui principalmente le cellule β pancreatiche e, in misura minore, il tessuto adiposo e quello nervoso, il GIP stimola la secrezione di glicogeno sia in condizioni di normoglicemia che di iperglicemia. Riduce inoltre l’appetito esercitando un effetto negativo sulla GE e favorendo il rilascio di insulina da parte del pancreas, agendo in sinergia con il GLP-1. Tuttavia, agisce in contrasto con il GLP-1 per quanto riguarda il rilascio di glucagone, potenziandone il rilascio. Inoltre, si è riscontrato che le vie dipendenti dai fattori di crescita, come la MAPK (chinasi 1 e 2 regolate da segnali extracellulari [ERK 1/2]) e la proteina chinasi B, sono influenzate dall’attivazione del GIP. Sono stati descritti anche gli effetti del GIP sul tessuto adiposo, poiché è stato dimostrato che stimola l’adipogenesi, inibisce la lipolisi e stimola la lipogenesi de novo. Inoltre, è stato dimostrato che il GIP ha un’influenza diretta sulle ossa, inducendo l’attività degli osteoblasti.
Glucagone
Infine, il Glucagone, derivato da una molecola precursore denominata pro-glucagone composta da 180 aminoacidi, è un ormone presente principalmente nelle cellule α del pancreas. È stata descritta anche la sua produzione nell’intestino tenue. Dopo la sua scoperta nel 1921, il suo contributo alla fisiopatologia del diabete mellito di tipo 2 ha inaugurato una nuova era nei farmaci antidiabetici e nello studio delle malattie metaboliche. È costituito da 29 aminoacidi e il suo gene è localizzato sul cromosoma 2. I recettori del Glucagone si trovano principalmente negli epatociti del fegato e nei reni. È stata descritta un’espressione minore anche in altri tessuti come il cuore, il pancreas, il tessuto adiposo e il tratto gastrointestinale. I recettori del Glucagone sono recettori accoppiati alle proteine G che, quando stimolati, determinano un aumento dell’AMPc e del Calcio intracellulari che, a loro volta, attivano la via della proteina chinasi A. Il principale fattore che ne determina il rilascio è il basso livello di glucosio nel sangue, e i recettori del Glucagone si trovano in molti tessuti, con gli epatociti come sede più comune. Il Glucagone consente la gluconeogenesi e la glicogenolisi nel fegato, bloccando al contempo la glicogenesi. Questi effetti portano a livelli elevati di glucosio nel sangue. Il Glucagone riduce anche la motilità gastrointestinale e, agendo sul tessuto adiposo, diminuisce la lipogenesi e induce la lipolisi, portando ad un aumento della produzione di acidi grassi non esterificati e corpi chetonici. È stato inoltre osservato che il Glucagone agisce sul tessuto adiposo bruno nei modelli animali, inducendo la termogenesi e il dispendio energetico. Tuttavia, gli studi condotti sull’uomo non hanno dimostrato più di un effetto modesto sul dispendio energetico, senza raggiungere una rilevanza clinica.
Recettore del Glucagone
-Aspetti generali della Retatrutide-
Struttura molecolare — Farmacologia
La Retatrutide è quindi una terapia promettente che agisce su tutti i recettori e attività molecolari correlate sopra menzionate; si tratta di un agonista triplo recettoriale. La Retatrutide è un peptide sintetico che agisce come agonista dei recettori del GLP-1, del GIP e del Glucagone (Figura seguente). La struttura chimica della Retatrutide gli consente di legarsi in modo specifico a questi recettori.
La Retatrutide si dispiega in un’unica struttura elicoidale continua che gli consente di attraversare il dominio transmembranario del recettore con il suo segmento N-terminale. Il segmento C-terminale partecipa alle interazioni con l’elica α N-terminale del dominio extracellulare, con l’estremità extracellulare dell’elica transmembranaria 1 del recettore del GLP-1 e con l’ansa extracellulare 1 del recettore del GIP. Questa molecola è più potente sul recettore GIP umano (EC50: 0,0643 nM) e meno potente sui recettori del GLP-1 (EC50: 0,775 nM) e del Glucagone (EC50: 5,79 nM). Ha un’azione dose-dipendente e provoca una diminuzione dello svuotamento gastrico, mentre, con un’emivita di 6 giorni, può essere somministrato prevalentemente su base settimanale. La Retatrutide viene metabolizzata prevalentemente a livello epatico, ma non interagisce con gli enzimi del citocromo P450.
Struttura molecolare della Retatrutide
I suoi effetti determinano una significativa riduzione del peso e una diminuzione dei livelli di HbA1c. Gli effetti indesiderati più frequenti sono nausea, diarrea, vomito e stitichezza. Tuttavia, questi hanno portato a casi non rari di interruzione della terapia, e la loro intensità era chiaramente correlata a dosaggi più elevati. Studi pratici indicano che i tassi di interruzione dei GLP-1RA possono raggiungere valori compresi tra il 20% e il 50% entro il primo anno e che i pazienti spesso utilizzano dosaggi significativamente inferiori rispetto a quelli testati negli studi clinici. Effetti avversi meno comuni sono stati un aumento temporaneo dei livelli di alanina aminotransferasi (ALT), aumento della frequenza cardiaca e iperestesia cutanea.
-Risultati emersi dagli studi sull’uomo-
Trials di Fase I
Coksun et al. a Singapore hanno condotto uno studio di fase 1 per valutare la sicurezza e il profilo farmacocinetico della Retatrutide. Allo studio hanno partecipato 47 soggetti sani, 45 dei quali hanno ricevuto almeno una dose. La concentrazione massima è stata raggiunta entro 12-72 ore dalla somministrazione. L’emivita media era di circa 6 giorni. Rispetto al placebo, la Retatrutide ha determinato variazioni simili nei livelli basali di glucosio a digiuno. Dopo la somministrazione di dosi da 4,5 e 6 mg, i livelli medi di glucagone a digiuno sono diminuiti a partire da 24 ore dopo la somministrazione fino al giorno 15. Il retatrutide ha determinato una riduzione del peso corporeo medio a tutti i livelli di dosaggio, tranne che alla dose di 0,1 mg. La riduzione di peso ha raggiunto il picco ai livelli di dosaggio compresi tra 3 e 6 mg. Gli effetti avversi più comuni correlati al trattamento dello studio sono stati disturbi gastrointestinali quali vomito, distensione addominale e nausea, tuttavia per lo più di intensità lieve.
A seguito degli studi condotti sugli animali, Urva et al. hanno progettato uno studio di fase 1b a dosi multiple crescenti su pazienti affetti da diabete di tipo 2. A questi pazienti è stato somministrato Retatrutide una volta alla settimana (in gruppi di dosaggio da 0,5, 1,5, 3, 3/6 e 3/6/9/12 mg), placebo o 1,5 mg di Dulaglutide. Retatrutide 0,5, 1,5 e 3 mg sono stati somministrati per 12 settimane. Il gruppo da 3/6 mg ha ricevuto 3 mg di Retatrutide nelle settimane 1-4 e 6 mg nelle settimane 5-12. Infine, il gruppo da 3/6/9/12 mg ha ricevuto 3 mg nelle prime due settimane, 6 mg nelle settimane 3-4, 9 mg nelle settimane 5-8 e 12 mg nelle settimane 9-12. Lo svuotamento gastrico è stato valutato 2 giorni prima del trattamento e 24 ore dopo la somministrazione della dose nei giorni 2, 30 e 79. In sintesi, 72 partecipanti hanno ricevuto almeno una dose del farmaco, mentre 43 hanno completato lo studio. L’età media dei partecipanti era di 58,4 ± 7,4 anni, l’IMC medio era di 32,1 ± 5,1 kg/m², l’HbA1c era dell’8,66 ± 0,92%. Il maggiore effetto sullo svuotamento gastrico è stato riscontrato dopo la prima dose di Retatrutide, mentre era inferiore con le dosi successive, nonostante l’aumento del dosaggio.
Trials di Fase II
In uno studio di fase II in doppio cieco, randomizzato e controllato con placebo, Jastreboff et al. hanno somministrato Retatrutide in dosi fino a 8 mg per un totale di 48 settimane a pazienti con un BMI ≥ 30 kg/m² o con un BMI compreso tra 27 e 30 kg/m² e una patologia concomitante correlata al peso. L’endpoint primario dello studio era la variazione percentuale del peso corporeo dal basale alle 24 settimane di terapia. Gli endpoint secondari erano la variazione percentuale del peso corporeo dal basale alle 48 settimane e la presenza di una riduzione di peso ≥5%, ≥10% e ≥15%. Sono stati arruolati in totale 338 adulti. La variazione percentuale media del peso corporeo a 24 settimane nei gruppi trattati con Retatrutide, calcolata con il metodo dei minimi quadrati, è stata del −7,2% nel gruppo da 1 mg e fino al −17,5% nel gruppo da 12 mg, rispetto al −1,6% nel gruppo placebo. Inoltre, a 48 settimane, le stesse percentuali erano pari a −8,7% nel gruppo da 1 mg, −24,2% nel gruppo da 12 mg e −2,1% nel gruppo placebo. A 48 settimane, si è verificata una riduzione di peso ≥5%, ≥10% e ≥15% nel 100%, 93% e 83% dei soggetti che hanno ricevuto 12 mg di Retatrutide e nel 27%, 9% e 2% di quelli che hanno ricevuto il placebo. Gli effetti avversi più comuni erano anch’essi di natura gastrointestinale e correlati alla dose e sono stati parzialmente mitigati con una dose iniziale più bassa (2 mg vs. 4 mg). Pertanto, negli adulti con obesità, il trattamento con Retatrutide per 48 settimane ha determinato riduzioni sostanziali del peso corporeo.
Analogamente, Rosenstock et al. hanno condotto uno studio randomizzato, in doppio cieco, di fase 2, in cui erano ammessi all’arruolamento adulti di età compresa tra i 18 e i 75 anni affetti da diabete di tipo 2, con HbA1c compresa tra il 7,0% e il 10,5% e BMI compreso tra 25 e 50 kg/m². I partecipanti sono stati trattati con dieta ed esercizio fisico da soli o con una dose stabile di metformina per almeno 3 mesi prima dello screening. Sono stati inoltre assegnati in modo casuale a ricevere iniezioni settimanali di placebo, 1,5 mg di Dulaglutide o dosi di mantenimento di Retatrutide pari a 0,5 mg, 4 mg (dose iniziale 2 mg), 4 mg (senza aumento), 8 mg (dose iniziale 2 mg), 8 mg (dose iniziale 4 mg) o 12 mg (dose iniziale 2 mg). L’endpoint primario era la variazione dell’HbA1c dal basale a 24 settimane, mentre gli endpoint secondari includevano la variazione dell’HbA1c e del peso corporeo a 36 settimane. A 24 settimane, le variazioni medie dei minimi quadrati rispetto al basale dell’HbA1c con retatrutide erano pari a –0,43% per il gruppo da 0,5 mg, –1,39% per il gruppo con aumento graduale a 4 mg, –1,30% per il gruppo da 4 mg, –1,99% per il gruppo con aumento graduale lento a 8 mg, –1,88% per il gruppo con aumento rapido della dose a 8 mg e –2,02% per il gruppo con aumento della dose a 12 mg, contro –0,01% per il gruppo placebo e –1,41% per il gruppo trattato con Dulaglutide 1,5 mg. Le riduzioni dell’HbA1c con Retatrutide sono risultate significativamente maggiori (p < 0,0001) rispetto al placebo in tutti i gruppi tranne quello da 0,5 mg e maggiori rispetto al Dulaglutide da 1,5 mg nel gruppo con aumento graduale a 8 mg (p = 0,0019) e nel gruppo con aumento graduale a 12 mg (p = 0,0002). I risultati sono stati confermati a 36 settimane. Il peso corporeo è diminuito in modo dose-dipendente con Retatrutide a 36 settimane fino al 16,94% per il gruppo con aumento progressivo della dose a 12 mg, rispetto al 3,00% con il placebo e al 2,02% con 1,5 mg di Dulaglutide. È risultato evidente che Retatrutide ha mostrato miglioramenti clinicamente significativi nel controllo glicemico e riduzioni impressionanti del peso corporeo nei pazienti con diabete di tipo 2, pur essendo relativamente sicuro.
Un altro studio di fase II condotto da Sanyal et al. ha cercato di dimostrare gli effetti del Retatrutide sulla steatosi epatica. 98 pazienti con un contenuto di grasso epatico ≥10% (come definito dalla frazione di grasso in base alla densità protonica mediante risonanza magnetica) sono stati assegnati in modo casuale a un trattamento di 48 settimane con Retatrutide per via sottocutanea una volta alla settimana (a dosi di 1, 4, 8 o 12 mg) o a un trattamento con placebo. In totale, 76 partecipanti sono riusciti a completare questo sottostudio. La variazione relativa media rispetto al basale del grasso epatico a 24 settimane è stata di −42,9% (1 mg), −57,0% (4 mg), −81,4% (8 mg), −82,4% (12 mg) e +0,3% (placebo). A 24 settimane, il grasso epatico normale (<5%) è stato raggiunto dal 27% (1 mg), dal 52% (4 mg), dal 79% (8 mg), dall’86% (12 mg) e dallo 0% (placebo) dei partecipanti. A 48 settimane, la variazione relativa media rispetto al basale del grasso epatico era pari a −51,3% (1 mg), −59,0% (4 mg), −81,7% (8 mg), −86,0% (12 mg) e −4,6% (placebo). Inoltre, un contenuto totale di grasso epatico < 5% a 48 settimane è stato raggiunto con dosi da 8 mg (nell’89% dei partecipanti) e da 12 mg (nel 93% dei partecipanti). La riduzione del grasso epatico è stata associata a perdita di peso, riduzione del grasso addominale e miglioramento della sensibilità all’insulina e del metabolismo dei lipidi.
Trials di Fase III
Risultati dello studio TRIUMPH-1 [2026], uno studio clinico di fase 3 volto a valutare l’efficacia e la sicurezza della Retatrutide in adulti affetti da obesità o sovrappeso, con almeno una comorbilità correlata al peso e senza diabete. A 80 settimane, tutte le dosi della Retatrutide (4mg, 9mg e 12mg) hanno raggiunto gli endpoint primari e secondari chiave per l’obesità, determinando una perdita di peso clinicamente significativa.
I soggetti con un BMI basale ≥35 che hanno partecipato a un’estensione dello studio hanno continuato a perdere peso, raggiungendo una perdita media di 85,0 libbre (30,3%) a 104 settimane. Alla dose di 4mg, raggiunta con un solo aumento, i partecipanti hanno perso in media 47,2 libbre (19,0%) a 80 settimane, con un tasso di interruzione osservato inferiore a causa di eventi avversi rispetto al placebo.
Per quanto riguarda l’endpoint primario, i partecipanti trattati con Retatrutide 9mg e 12mg hanno perso in media rispettivamente 64,4 libbre (25,9%) e 70,3 libbre (28,3%). Coloro che hanno assunto la dose da 4mg di Retatrutide, con un solo aumento del dosaggio, hanno perso in media 47,2 libbre (19,0%). In particolare, il 65,3% dei partecipanti trattati con Retatrutide 12mg ha raggiunto un BMI <30, scendendo al di sotto della soglia di obesità a 80 settimane, compreso il 37,5% di coloro che hanno iniziato con obesità di classe 3 (BMI ≥40). 1 In un’estensione in cieco prestabilita per i soggetti con un BMI ≥35, i partecipanti che hanno continuato con Retatrutide 12mg fino a 104 settimane hanno perso in media 85,0 libbre (30,3%). Inoltre, la Retatrutide ha mostrato miglioramenti significativi rispetto al basale in alcuni fattori di rischio cardiovascolare, tra cui la circonferenza della vita, il colesterolo non-HDL, i Trigliceridi, la pressione sistolica e la proteina C-reattiva ad alta sensibilità (hsCRP).
Per quanto riguarda l’estimando del regime terapeutico, ogni livello di dosaggio di Retatrutide ha portato a miglioramenti negli endpoint primari e secondari chiave, nonché nell’estensione prestabilita, tra cui:
Variazione percentuale del peso corporeo a 104 settimane: -25,7% (-30,6 kg; -67,5 libbre; da 4 mg a MTD); -28,7% (-35,6 kg; -78,4 libbre; da 9 mg a MTD); -29,9% (-38,1 kg; -83,9 libbre; da 12 mg a MTD) e -18,9% (-22,3 kg; -49,1 libbre; da placebo a MTD)
I tipi di eventi avversi osservati erano generalmente in linea con quelli riscontrati negli studi clinici condotti su altre terapie a base di incretine. Gli eventi avversi più comuni tra i partecipanti trattati con retatrutide (4 mg, 9 mg, 12 mg, rispetto al placebo, rispettivamente) sono stati nausea (28,6%, 38,4% e 42,4% contro il 14,8%), diarrea (25,2%, 34,1% e 32,0% contro il 13,5%), stitichezza (23,8%, 25,9% e 26,1% rispetto al 10,9%), vomito (10,6%, 22,8% e 25,3% rispetto al 4,8%) e infezione del tratto respiratorio superiore (14,2%, 12,2% e 13,1% rispetto all’11,6%) . Si sono verificati casi di disestesia nel 5,1%, 12,3% e 12,5% dei pazienti trattati con retatrutide 4 mg, 9 mg e 12 mg, rispettivamente, rispetto allo 0,9% con il placebo, mentre l’incidenza di infezioni del tratto urinario si è verificata nel 7,5%, 8,8% e 8,4% dei pazienti trattati con retatrutide 4 mg, 9 mg e 12 mg, rispettivamente, rispetto al 5,3% con il placebo. Gli episodi di disestesia e le infezioni del tratto urinario sono stati generalmente di entità da lieve a moderata; la maggior parte si è risolta durante il trattamento e la maggior parte dei partecipanti ha continuato ad assumere retatrutide. I tassi di interruzione del trattamento a causa di eventi avversi sono stati del 4,1%, 6,9% e 11,3% rispettivamente con retatrutide 4 mg, 9 mg e 12 mg, rispetto al 4,9% con il placebo.
Principali risultati della fase III:
Perdita di peso (TRIUMPH-1): negli adulti affetti da obesità o sovrappeso, la dose settimanale da 12 mg ha determinato una notevole perdita di peso media del 28,3% a 80 settimane. Uno studio di estensione di due anni (104 settimane) condotto sul sottogruppo con il peso maggiore (BMI ≥ 35) ha mostrato una notevole riduzione media del peso corporeo del 30,3% (con una perdita media di 85 libbre).
Diabete di tipo 2 (TRANSCEND-T2D-1): Negli adulti con diabete di tipo 2, la dose da 12 mg ha prodotto una perdita di peso media del 15,3% a 40 settimane. In particolare, fino al 40% dei pazienti ha raggiunto livelli normali di HbA1c (<5,7%) e quasi il 90% ha raggiunto valori <7,0%.
Osteoartrite (TRIUMPH-4): I pazienti affetti da obesità e osteoartrite del ginocchio hanno registrato una perdita di peso del 28,7% insieme a una significativa riduzione del dolore al ginocchio a 68 settimane.
Conclusioni:
È fondamentale comprendere che la perdita di peso non è sempre benefica per la salute; in determinate circostanze, può portare a una riduzione clinicamente significativa della massa muscolare e ossea, anziché alla riduzione desiderata del grasso corporeo. Questo fenomeno solleva importanti interrogativi sulla qualità e sulla composizione della perdita di peso, poiché il mantenimento della massa magra è essenziale per preservare la forza complessiva, la mobilità e la salute metabolica. Pertanto, è necessario progettare studi che non si concentrino solo sulla percentuale, ma che indaghino anche sui cambiamenti della composizione corporea che accompagnano la perdita di peso.
Va tuttavia sottolineato che, sebbene la Retatrutide appaia promettente come terapia dimagrante per gli adulti, il suo impiego sperimentale non è attualmente approvato per bambini e adolescenti. La sicurezza e l’efficacia della Retatrutide non sono state stabilite nella popolazione pediatrica. Gli studi clinici attualmente in corso si sono concentrati esclusivamente sugli adulti e mancano dati relativi al suo impiego nei bambini e negli adolescenti. Pertanto, la Retatrutide non è indicato per i soggetti di età inferiore ai 18 anni. Per i bambini e gli adolescenti, la prevenzione e la gestione dell’obesità dovrebbero dare priorità agli interventi non farmacologici. Le modifiche dello stile di vita, tra cui una dieta equilibrata e un’attività fisica regolare, sono la pietra angolare della prevenzione dell’obesità pediatrica. La U.S. Preventive Services Task Force raccomanda ai medici di effettuare screening per l’obesità nei bambini e negli adolescenti di età pari o superiore a 6 anni e di offrire loro o indirizzarli verso interventi comportamentali intensivi e completi per promuovere miglioramenti nel peso corporeo.
Tuttavia, non si può negare che gli studi di fase III sulla Retatrutide hanno fino a questo momento dimostrato una riduzione del peso e miglioramenti cardiometabolici senza precedenti.
In conclusione, il Retatrutide sembra preannunciare un significativo progresso nel campo della farmacoterapia per l’obesità e il controllo del peso. Grazie al suo eccellente profilo di sicurezza e alle notevoli percentuali di perdita di peso, suscita indubbiamente grande entusiasmo nella comunità medica.
La presentazione della domanda di autorizzazione per la commercializzazione della Retatrutide è prevista intorno al 2027.
Nella prima parte di questa disamina sono state passate in rassegna diverse informazioni necessarie per comprendere cosa sono le Incretine e quali sono le loro principali azioni. Si è poi passati a descrivere la classe di farmaci degli Incretino-Mimetici discorrendo sulla loro sintesi, tipologia molecolare [in ordine cronologico in base alla data di commercializzazione] e caratteristiche di azione sia per quanto concerne le attività positive che quelle negative. In conclusione, si è accennato alla consistente diffusione dell’uso “cosmetico” di tali farmaci.
In questa seconda ed ultima parte vedremo come questa relativamente nuova classe di farmaci abbia trovato un certo spazio di diffusione nel Fitness e (sebbene in minor parte) nel BodyBuilding. Verranno discussi i punti di attrattiva e le limitazioni (e rischi) legati al loro uso.
Le “attrattive” degli Incretino-Mimetici:
L’obesità è una grave epidemia che affligge la società del così detto “occidente americanizzato”. I farmaci iniettabili sottocutanei inizialmente concepiti per la gestione del diabete di tipo II, come la Semaglutide e altri agonisti del recettore del GLP-1, stanno rapidamente guadagnando popolarità per i loro effetti sulla perdita di peso. Questi farmaci (Ozempic, Wegovy, Saxenda e Mounjaro) sono onnipresenti sui social media e sono promossi da celebrità di tutte le fasce demografiche. “Viso da Ozempic” e ‘sedere da Ozempic’ sono ormai concetti mainstream che evidenziano i possibili cambiamenti morfologici che si verificano con questi farmaci. Con la diffusione non controllata da personale qualificato dell’uso di questi farmaci, è aumentato anche l’elenco dei potenziali effetti avversi.
Nella prima parte di questa disamina si è constatato che gli Incretino-Mimetici hanno potenzialità che si esprimono su modifiche della composizione corporea e che riguardano principalmente:
Rallentamento dello svuotamento gastrico con riduzione del senso di fame derivante dal aumento del senso di sazietà;
Gli effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare riducendo il senso di fame e il consumo di cibo;
Indirettamente, per via della stimolazione insulinica, aumento del senso di sazietà dato dal picco insulinico a livello ipotalamico;
Miglioramento del ripartizionamento Kcal attraverso il miglioramento della sensibilità all’insulina.
L'”attrattiva” che ha spinto (e che spinge) diverse persone di diverse classi sociali e di ambo i sessi a prendere in considerazione e concretizzare l’uso di questi farmaci è fondamentalmente ridotta alla riduzione della fame/appetito. Ma questo riguarda la persona “nella media”, la ragazza/donna alla ricerca di rapide soluzioni per l’imminente prova costume o per il servizio fotografico, sfilata ecc…
Nel Bodybuilding questa classe di farmaci ebbe una serie di attrattive comprendenti lo sfruttamento di tutti i punti sopra elencati. E, di conseguenza, si ipotizzarono fasi della preparazione nelle quali applicare tale categoria farmaceutica. Limitandone l’uso in “Cut” per ragioni legate al rischio (seppur limitato in monoterapia) di incorrere in eventi ipoglicemici, l'”attrattiva d’uso” riguardava il potenziale in fasi di “Recomp”; quindi non propriamente “ipocalorico” o, al massimo, leggermente ipocaloriche e non ipoglucidiche.
Ricomposizione corporea, Ripartizionamento calorico e Insulino-Sensibilità:
Ma chiariamo alcuni termini per schiarirci le idee…
Ricomposizione corporea: si intende il raggiungimento di un obiettivo o un risultato desiderato di un regime alimentare e di allenamento (e di farmaci), come il “Cut” o il “Bulk”.
“Cut”: si intende, propriamente, una diminuzione della massa grassa (FM) e il mantenimento della massa muscolare (LBM).
“Bulk”: si intende un aumento della LBM con una concomitante attenuazione dell’aumento della FM.
“Recomp” [comunemente intesa]: è definita come un aumento della LBM e la diminuzione della FM.
La suddivisione terminologica corretta è un concetto funzionale all’obiettivo di tutti gli interventi dietetici razionali.
Il rapporto p (rapporto di ripartizione) descrive le proteine depositate nei tessuti della LBM in relazione all’apporto energetico e, viceversa, le proteine perse dai tessuti della LBM in relazione al deficit energetico.
Il p-ratio comprende i fattori di:
stato ormonale (cioè i livelli assoluti di ormoni chiave noti);
sensibilità all’insulina;
sensibilità alla Leptina.
Esiste un’interazione tra il punto 1 e 3.
La prima chiave è…
Sensibilità all’Insulina: quando si è a dieta (cioè in uno stato di deficit energetico), la resistenza all’Insulina fisiologica è una condizione favorevole all’uso del grasso di deposito limitando l’uso del glucosio da parte del muscolo come substrato energetico, risparmiando il glucosio per il cervello e l’utilizzo degli Acidi Grassi intramuscolare. In condizioni di uscita da un regime ipocalorico, in uno stato di migliorata insulino-sensibilità, l’aumento calorico di una fase di “Bulk” vede, almeno inizialmente, un ripartizionamento delle Kcal al miocita a discapito del adipocita; tale condizione di inverte col protrarsi del regime ipercalorico.
I fattori che influenzano la sensibilità all’insulina includono [1]:
Livelli di grasso corporeo; B.F. % (predittore primario): ↑B.F. ⇒ ↑Acidi Grassi come substrato energetico (risparmiando glucosio e proteine [che possono essere utilizzate dal fegato nella gluconeogenesi]) e detta la segnalazione delle adipochine (cioè gli ormoni secernenti gli adipociti [leptina, TNF-α, IL-…, adiponectina, etc.]) ⇒ ↓ sensibilità all’insulina;
Contrazione muscolare (cioè attività, come ad esempio locomozione, allenamento contro-resistenza) ⇒ ↑ assorbimento di glucosio nella cellula muscolare; traslocazione di GLUT-4 ⇒ ↑ sensibilità all’Insulina;
Dieta: elevata quantità di carboidrati (in ipercalorica), grassi saturi e poche fibre ⇒ ↓ sensibilità all’Insulina;
Stoccaggio del Glicogeno o Supercompensazione [successivo ad una deplezione] ⇒ ↑assorbimento di glucosio e glicogenesi ⇒↑sensibilità Insulinica;
Deplezione di glicogeno (ad esempio, nel periodo successivo a un allenamento intenso, prima di un’alimentazione particolarmente ricca di carboidrati) ⇒ sottoregolazione (deplezione pressoché totale) della disponibilità di glucosio e promozione dell’ossidazione di Acidi Grassi dopo il depauperamento delle scorte di glicogeno muscolare (in media < 700 g negli adulti) ⇒ ↑ Acidi Grassi liberi nel sangue (circolanti) ⇒ ↓ sensibilità all’Insulina;
Fattori genetici in parte modificabili dai farmaci, ad esempio, nei casi di ipogonadismo, l’applicazione TRT inverte chiaramente l’insulino-resistenza nei casi in cui l’eziologia dell’insulino-resistenza è riconducibile alla carenza di Testosterone.
La seconda chiave è…
Struttura molecolare della Leptina
La Leptina: come sappiamo bene, la Leptina è un ormone, più precisamente è una adipochina, secreta principalmente dagli adipociti, che si correla con la %B.F.;↑%B.F. ⇒ ↑Leptina. (i depositi viscerali e quelli sottocutanei hanno rapporti diversi con la Leptina). A una data percentuale di B.F., le donne producono ~2 – 3 volte più Leptina rispetto agli uomini. Le concentrazioni di Leptina cambiano con la restrizione energetica e la sovralimentazione. La Leptina è un segnalatore primario di regolazione dell’accumulo di energia che riflette:
la percentuale di B.F.;
l’assunzione di energia.
Esempio 1: all’inizio di una dieta ipocalorica, la Leptina può diminuire del 50% entro 1 settimana (o meno) – anche se ovviamente il soggetto a dieta non ha perso il 50% di B.F. – quindi, in un primo momento, i cambiamenti della concentrazione di Leptina non sono correlati alla B.F. (piuttosto segnalano l’assunzione di energia).
Dopo il calo iniziale, si assiste a un declino più graduale della Leptina in relazione alla perdita di %B.F. .
Esempio 2: in caso di sovralimentazione, la Leptina subisce un incremento in modo altrettanto rapido (cioè senza relazione con la %B.F., ma in relazione all’assunzione di energia).
A breve termine, la secrezione di Leptina è determinata principalmente dalla disponibilità di glucosio, per cui la riduzione della disponibilità di glucosio nella cellula adiposa (dieta ipocalorica) ⇒ ↓Leptina e viceversa.
Gli effetti specifici dell’ormone Leptina includono effetti sul pancreas e sul fegato, nel muscolo scheletrico ↑FA e ↓AA e l’uso del glucosio come substrato energetico (aumentando la perdita di grasso, promuovendo il risparmio di proteine)… [1]
Modello classico Leptina-Melanocortina
In sostanza, il partizionamento (p-ratio) è un concetto che associa la Leptina e la sensibilità all’Insulina come fattori principali che determinano il modo in cui le variazioni dell’apporto calorico e del contenuto di macronutrienti influiscono sul metabolismo (influenzando profondamente la composizione corporea) e sullo stato ormonale. Possiamo modificarlo e migliorarlo, tenendo conto dei tessuti bersaglio e del nostro obiettivo (ad esempio, se Bulk o Cut).
Inoltre, non bisogna confondere il potenziamento dell’insulino-resistenza fisiologica sulla perdita di grasso con l’erronea valutazione che l’insulino-resistenza sia salutare. L’insulino-resistenza, soprattutto in una persona sedentaria, è associata alla sindrome metabolica, al diabete di tipo II, per non parlare del grasso viscerale, ecc.
L’insulino-resistenza è uno stato in cui i tessuti dell’organismo (ad esempio, fegato, pancreas, muscolo scheletrico) presentano una scarsa recettività con l’Insulina continuando, se si parla in particolare del fegato, a produrre glucosio in quantità inappropriate. Questo stato di iperglicemia è un effetto piuttosto che la causa dell’insulino-resistenza, anche se i livelli tossici di glucosio degradano la reattività delle isole pancreatiche all’Insulina rappresentando così una delle vie/meccanismi dell’insulino-resistenza, peggiorando la stessa condizione.
Ma tutto questo cosa centra con gli Incretino-Mimetici? Se non ci siete ancora arrivati, calma e capirete …
Agonisti del recettore GLP-1 e GIP:
La perdita di grasso si verifica con gli agonisti del GLP-1 e della GIP (Incretino-mimetici) – come la Semaglutide e la Tirzepatide – che sono veri e propri agenti sensibilizzanti dell’Insulina. Tuttavia, non è la sensibilità all’Insulina di per sé che è responsabile della perdita di grasso con questa classe di farmaci – ma piuttosto, come abbiamo già visto, la perdita di grasso avviene grazie agli altri effetti di questi farmaci, come l’alterazione potenziale delle preferenze alimentari, il ritardo dello svuotamento gastrico, il senso di sazietà, che promuovono il controllo dell’appetito e riducono l’assunzione di energia.
Sappiamo che gli agonisti del GLP-1 e del GIP migliorano direttamente la sensibilità all’insulina modulando la secrezione di Insulina – accoppiandola alla presenza di elevate concentrazioni di glucosio. Questa secrezione di Insulina si attenua quando le concentrazioni di glucosio nel sangue diminuiscono e si avvicinano all’euglicemia. Inoltre, anche se indirettamente, riducendo l’assunzione di cibo, questi farmaci determinano una riduzione della %B.F.. La riduzione della percentuale di B.F. dovuta alla riduzione dell’assunzione di cibo riduce le riserve di massa grassa (e quindi gli FFA circolanti), migliorando ulteriormente la sensibilità all’Insulina.
La stragrande maggioranza degli agenti per la perdita di grasso, in quanto agenti lipolitici, favoriscono l’insulino-resistenza. Ad esempio i β-agonisti, non selettivi come l’Efedrina o selettivi come il Clenbuterolo, oppure lo stimolante da banco per eccellenza la caffeina, agendo in modo analogo o aumentano l’azione delle catecolamine (epinefrina e noradrenalina, o adrenalina e noradrelanina) possono portare ad un peggioramento di questa condizione.
Diagramma schematico che rappresenta la via di segnalazione dell’insulino-resistenza cardiaca mediata dai β-AR. Tale meccanismo interessa (tra gli altri tessuti) anche il muscolo-scheletrico.
Quando tessuti come il fegato e le cellule adipose vedono ridotta l’interazione con l’Insulina, il glucosio non viene ottimamente assorbito dalle cellule. Con un marcato calo del glucosio, il fegato inizia a metabolizzare gli acidi grassi liberi (FFA), aumentando così i livelli di chetoni nel sangue e impedendo che vengano riesterificati nelle cellule adipose (in ipocalorica). Nel fegato e nelle cellule adipose, senza che l’Insulina interagisca ottimamente con questi tessuti, si verifica una soppressione della sintesi/lipogenesi dei grassi (negli adipociti) e della sintesi di lipoproteine a bassissima densità (VLDL) (nel fegato).
Incretino-mimetici, miglioramento della sensibilità all’Insulina e preservazione della massa muscolo-scheletrica:
La classificazione degli incretino-mimetici come agenti di ripartizione calorica:
Nel BodyBuilding, il fascino verso questa classe di farmaci si concentra anche sul funzionamento degli agonisti del GLP-1 e della GIP sull’insulino-resistenza, poiché quest’ultima durante la restrizione calorica nel muscolo scheletrico (>60% del peso corporeo, più nei bodybuilder) è un’immagine non proprio esaltante, con le riserve di glicogeno che vengono prima catabolizzate abbastanza rapidamente; poi i trigliceridi intramuscolari (che rappresentano solo l’1% del peso del muscolo idratato, fino al 2% del volume, dato che il grasso è meno denso del muscolo scheletrico, e ~1/3 dell’energia muscolare, dato che il grasso è energeticamente denso) e infine, se necessario, l’organismo utilizzerà gli AA (catabolizzando le proteine muscolari; proteolisi) per ottenere l’energia necessaria a svolgere le attività giornaliere. Questi agenti, quindi, nella misura in cui sono sensibilizzanti per l’Insulina, dovrebbero servire a promuovere il mantenimento della LBM durante il “Cut”.
In effetti, come si evince dalla seguente immagine tratta da un articolo di ricerca di Volpe et. al del 2022 [2], la Semaglutide preserva in modo abbastanza efficace la LBM e riduce in modo preferenziale la FM, con riduzioni solo clinicamente insignificanti dell’indice di massa magra (FFMI, kg/m²) e dell’indice della muscolatura scheletrica durante il periodo iniziale di adattamento, che poi si attenua:
In un certo senso, quindi, migliorando il grado di sensibilità all’Insulina dell’equazione della p-ratio, gli incretino-mimetici possono essere classificati come agenti di ripartizione calorica, a grandi linee come il Clenbuterolo, ma invece di promuovere l’insulino-resistenza come i β-agonisti, la migliorano.
* Ovviamente, creare un ambiente significativamente insulino-sensibile in un contesto ipocalorico (soprattutto se ipoglucidico) può mettere l’utilizzatore a maggior rischio (sebbene limitato) di ipoglicemia o calo glicemico borderline con effetti simili allo stato di ipoglicemia (tremore, sudorazione copiosa ecc…).
Per coloro che hanno familiarità con questi concetti, derivanti dalle discussioni sul bodybuilding, può risultare molto confuso il fatto che l’iperglicemia (elevata quantità di glucosio nel sangue) è solo uno dei fattori associati all’insulino-resistenza, ma in realtà non è sinonimo di insulino-resistenza (iperglicemia ≠ insulino-resistenza). Sì, ridurre la glicemia a livelli normali è molto importante per migliorare la sensibilità all’insulina durante l’uso dell’ormone della crescita esogeno (rhGH), perché il glucosio è tossico per le cellule β pancreatiche. Questa glucotossicità a livello delle cellule pancreatiche si traduce in una diminuzione della risposta secretoria dell’insulina all’iperglicemia, alimentando così il fuoco dell’iperglicemia e della glucotossicità, contribuendo all’insulino-resistenza – ma non costituendone l’unica eziologia.
La sensibilità all’Insulina è multifattoriale e comprende componenti sistemiche (ad esempio, QUICKI) e periferiche (ad esempio, GLUT-4) ed è regolata a livello centrale da GLP-1 e GIP. L’iperglicemia è solo uno dei fattori (l’altro è l’Insulina) che funge da proxy dell’insulino-resistenza sistemica. Vi sono altri aspetti, tra cui la tolleranza ai carboidrati [vedi anche capacità di tolleranza del metabolismo glucidico], ecc.
In tema di insulino-resistenza, è utile ricordare che l’Insulina endogena viene secreta in modo pulsatile per regolare il metabolismo glucidico e lipidico, la crescita cellulare ecc…, a differenza del Testosterone che viene secreto in modo più stabile (rilascio graduale nel sangue, ma soggetto a variazioni diurne, ad esempio una maggiore secrezione al mattino rispetto a mezzogiorno/sera). Gli aumenti cronici di Insulina, ad esempio quelli relativi al profilo di rilascio di una bassa dose giornaliera di Insulina Glargine (Lantus), presentano un’area sotto la curva (AUC) relativamente ampia a causa del profilo di rilascio (concentrazioni elevate per lunghi periodi di tempo) rispetto ai normali profili di rilascio dell’Insulina endogena (paragonabili alla farmacocinetica dell’Insulina regolare, ad esempio Actrapid, Novolin o HumuLin -R). L’elevata AUC di Lantus e/o le dosi di Insulina regolare esogena moderatamente elevate e frequenti sono descritte come iperinsulinemia cronica.
Questa resistenza non avviene per feedback negativo a livello delle cellule β.
Al contrario, l’iperinsulinemia cronica che causa l’insulino-resistenza è multifattoriale e comprende:
L’aumento dell’HOMA-IR e la diminuzione del QUICKI (misure biochimiche dell’insulino-resistenza e della sensibilità all’Insulina, rispettivamente).
L’alterata trasduzione del segnale insulinico dovuta alla disfunzione del recettore dell’Insulina (IR) e alla diminuzione dell’autofosforilazione dell’IR, che blocca la traslocazione del GLUT-4 sulla superficie cellulare nelle cellule muscolari e adipose, con conseguente aumento del glucosio nel sangue [3]:
Segnalazione dell’Insulina durante l’insulino-resistenza. Durante l’insulino-resistenza, la segnalazione attraverso le chinasi AKT è parzialmente compromessa. Non tutte le vie AKT-dipendenti sono interessate, così come altre vie di segnalazione, indicando che l’insulino-resistenza è selettiva. Pertanto, l’iperinsulinemia, in presenza di insulino-resistenza, promuove le attività anaboliche delle cellule attraverso la via MEK-ERK e attraverso mTORC1. Sebbene la via PIK/AKT sia compromessa durante l’insulino-resistenza e fornisca solo una traslocazione insufficiente di GLUT4 per l’assorbimento del glucosio e un’attivazione carente di eNOS, sembra esserci un’attivazione normale di mTORC1. Oltre alle conseguenze anaboliche della segnalazione attraverso la via MEK/ERK descritte nella figura, si osserva un’aumentata espressione di ET-1 e PAt-1 (non mostrato), nonché l’inibizione dell’autofagia e del fattore nucleare Nrf2, che compromettono rispettivamente il turnover dei costituenti cellulari e i meccanismi di difesa delle cellule dallo stress radicale. L’iperinsulinemia riduce l’assorbimento del glucosio non solo attraverso lo smorzamento della via PIK/AKT (resistenza all’Insulina), ma anche attraverso altre vie ancora sconosciute.
3. Aumento dei livelli e dell’attività di sn-1,2-diacilglicerolo (DAG) grazie alla sintesi de novo.
Le limitazioni degli Incretino-Mimetici dietro all’iniziale entusiasmo:
Se ci dovessimo basare su quanto esposto fino a questo punto, saremo tutti d’accordo nell’ammantare della nomea di “farmaci prodigiosi per la ricomposizione corporea” tanto la Semaglutide quanto il Tirzepatide e tutti gli altri membri di questa classe. Ma, dal momento che, la conoscenza per essere utile deve essere sufficientemente approfondita, occorre indagare meglio sulle caratteristiche di questi farmaci.
Sappiamo che la Semaglutide è effettivamente associata alla perdita di peso con una differenza media dell’11,85% rispetto al placebo emersa dalle ultime review. Il consolidamento degli studi ha mostrato che nausea, vomito, costipazione e diarrea sono gli eventi avversi più comuni. Nonostante questi effetti sembrano essere per lo più di gravità da lieve a moderata, la loro risoluzione totale era spesso connessa al termine del trattamento.
Dal momento che gli Incretino-mimetici causano una riduzione del senso di fame, in modo anche significativo (dose dipendente), rallentano lo svuotamento gastrico portando anche ad eventi diarroici preceduti da mal assorbimento dei nutrienti, senza sottovalutare il rischio di paresi gastrica, la facilità di perdere massa muscolare in un contesto ipocalorico è molto alta, specie se non supportata da agenti anabolizzanti; l’inserimento di questi ultimi, però, non “tampona” la possibile condizione di mal assorbimento e/o scarsa assunzione proteica/alimentare.
Inoltre, la paresi gastrica è una condizione che un bodybuilder sano di mente cerca di evitare a tutti i costi combattendoci già se sussiste abuso di Insulina e/o hGH.
Trattando la limitazione data da una potenziale eccessiva inappetenza, è giusto specificare un impatto singolare che le Incretine (e gli Incretino-Mimetici) hanno sulle preferenze alimentari. Sappiamo, infatti, che gli alimenti maggiormente palatabili sono tipicamente ricchi di grassi e/o zuccheri e tendono a essere preferiti a quelli a basso contenuto di grassi/zuccheri. L’entità di questa preferenza, tuttavia, può essere influenzata da peptidi intestinali quali la Grelina e il GLP-1. La Grelina e il GLP-1 sono influenzati in modo differenziato dal consumo di alimenti palatabili. Tralasciando la Grelina, le concentrazioni di GLP-1 a digiuno predicono negativamente l’assunzione di alimenti ricchi di zuccheri semplici in un paradigma di distributori automatici, che gli autori dello studio hanno interpretato come prova del fatto che il GLP-1 svolge un ruolo nelle vie di ricompensa che regolano l’assunzione di zuccheri semplici. Diversi studi riportano anche un’alterazione delle preferenze alimentari dopo l’intervento di bypass gastrico, con un allontanamento dalla preferenza per gli zuccheri/grassi elevati. L’assunzione di alimenti appetibili, in particolare di soluzioni zuccherate, è aumentata dalla Grelina, mentre il GLP-1 riduce preferenzialmente l’assunzione di alimenti ad alto contenuto di grassi e zuccheri, almeno dopo una somministrazione acuta. Inoltre, i lavori condotti sull’uomo rivelano che la preferenza per i grassi e gli zuccheri può essere alterata dalla chirurgia bariatrica e contribuire alla perdita di peso, ma non è ancora stato stabilito se questi effetti siano legati a un’alterazione del segnale della Grelina o del GLP-1. Infine, i livelli circolanti di Grelina e GLP-1 possono essere indicativi del consumo di cibo appetibile nell’uomo.[https://www.frontiersin.org/]
Schema che descrive i principali effetti elettrofisiologici dell’attivazione del GLP-1R nelle aree cerebrali coinvolte nella modulazione del comportamento alimentare. (A) Il GLP-1 (compresi i suoi agonisti) si lega al GLP-1R postsinaptico per depolarizzare il potenziale di membrana e/o aumentare la frequenza di sparo nella maggior parte delle regioni cerebrali, ma iperpolarizza il potenziale di membrana in alcune aree cerebrali. Diversi meccanismi ionici, tra cui il canale cationico non selettivo, il canale K+ e il canale TRPC5, possono essere coinvolti nell’attivazione della depolarizzazione o iperpolarizzazione indotta dal GLP-1R. (B) Oltre ai recettori postsinaptici, il GLP-1 agisce sui GLP-1R presinaptici per modulare la neurotrasmissione glutammatergica e GABAergica. ARC, nucleo arcuato; BNST, nucleo del letto della stria terminale; Glu, glutammato; CRH, ormone di rilascio della corticotropina; HC, ippocampo; LH, ipotalamo laterale; NAc, nucleo accumbens; NPY/AgRP, Neuropeptide Y/Peptide legato al gene Agouti; OB, bulbo olfattivo; PBN, nucleo parabrachiale; POMC, proopiomelanocortina; PVN, nucleo paraventricolare; PVT, nucleo talamico paraventricolare; VTA, area tegmentale ventrale.
Di conseguenza, nonostante gli indizi di cui sopra, l’uso di Incretino-Mimetici potrebbe ridurre marcatamente il consumo di cibo indipendentemente dalla fonte anche se, come abbiamo visto, l’effetto anoressizzante del GLP-1 sembra essere a maggior carico della componente alimentare glucidico-lipidica.
Tornando invece sulla questione legata al catabolismo muscolo-scheletrico e l’uso concomitante di agenti anabolizzanti, per ovviare a questo problema nei pazienti trattati con Semaglutide, è stato avviato uno studio clinico di fase 2b, multicentrico, in doppio cieco, controllato con placebo, randomizzato, per la determinazione della dose e per valutare la sicurezza e l’efficacia di Ostarina 3mg, Ostarina 6mg o placebo come trattamento per preservare la massa muscolare e aumentare la perdita di grasso in circa 150 pazienti con obesità sarcopenica o pazienti anziani in sovrappeso (>60 anni di età) trattati con Semaglutide (Wegovy®). L’endpoint primario è la massa corporea magra totale e gli endpoint secondari chiave sono la massa grassa corporea totale e la funzione fisica misurata dal test di salita delle scale a 16 settimane.
Dopo aver completato la parte di determinazione della dose di efficacia dello studio clinico di Fase 2b, si prevede che i partecipanti continueranno in cieco in uno studio clinico di estensione di Fase 2b in cui tutti i pazienti smetteranno di ricevere un GLP-1 RA, ma continueranno ad assumere un placebo, Ostarina 3mg o Ostarina 6mg per ulteriori 12 settimane. Lo studio clinico di estensione di fase 2b valuterà se l’Ostarina è in grado di mantenere la massa muscolare e prevenire l’aumento di grasso e peso che si verifica dopo l’interruzione di un GLP-1 RA.
Lo studio clinico è condotto in 14 centri clinici negli Stati Uniti. È stato raggiunto l’arruolamento completo dei circa 150 pazienti nello studio di fase 2b QUALITY. L’azienda prevede ora che l’ultimo paziente a completare lo studio di fase 2b QUALITY sarà nel dicembre 2024, con i risultati clinici di prima linea per lo studio clinico di fase 2b QUALITY attesi nel gennaio 2025. Inoltre, i risultati principali per lo studio clinico di estensione di Fase 2b in cieco separato possono ora essere attesi nel secondo trimestre solare del 2025.
Ovviamente, queste limitazioni, incisive nel Culturismo, interessano tutti gli Incretino-Mimetici, compresa la Tirzepatide la quale sembrerebbe avere un maggior margine di efficacia e “sicurezza” per quanto riguarda il rischio (seppur limitato in monoterapia) di eventi ipoglicemici.
Il motivo di questa riduzione di rischio ipoglicemico è dovuta alla doppia affinità recettoriale della Tirzepatide la quale, come abbiamo visto, possiede una attività agonista per i recettori del GLP-1 e del GIP. Ed è proprio il legame e l’attivazione di quest’ultimo recettore (GIP) da parte della Tirzepatide a permettere ciò. Infatti, l’attivazione del recettore GIP stimola la secrezione di Glucagone in modo glucosio-dipendente nelle persone sane, con un’attività maggiore in presenza di glicemie più basse. Ciò significa che, raggiunta la soglia euglicemica i livelli di glucosio nel sangue verranno mantenuti più facilmente all’interno di questa soglia per via dell’attività del Glucagone secreto dalle cellule α delle isole di Langerhans.[https://pubmed.ncbi.nlm.nih.gov/]
Sebbene i trials clinici suggeriscano che la Tirzepatide riduca la glucagonemia, un recente studio dimostra che la Tirzepatide è un potente stimolatore della secrezione di Glucagone nelle condizioni sopradette.
Quindi la Tirzepatide è superiore alla Semaglutide?
Alcuni studi recenti hanno messo a confronto la Semaglutide e la Tirzepatide per la perdita di peso. Studi di ricerca del 2021, del 2023 e del 2024 suggeriscono che la Tirzepatide può determinare una maggiore perdita di peso rispetto alla Semaglutide.
Variazione percentuale media del peso corporeo a 3, 6 e 12 mesi di trattamento per la popolazione complessiva, i soggetti con diabete di tipo II (TD2) e quelli senza TD2. Le barre rappresentano le variazioni medie del peso corporeo dal basale al punto di riferimento tra la popolazione di pazienti ancora in trattamento, abbinata in base al punteggio di propensione. Barre scure (Semaglutide); Barre chiare (Tirzepatide).
Ma questi studi presentano alcuni importanti limiti.
Le dosi di Semaglutide e Tirzepatide somministrate ai partecipanti non erano, per ovvie ragioni, uguali. La Semaglutide e Tirzepatide funzionano in modo leggermente diverso, come ormai sappiamo, e quindi i ricercatori hanno scelto livelli di dose comparabili. Tuttavia, la dose di Tirzepatide era più alta, il che potrebbe aver, anche di poco, influenzato i risultati.
Tabella comparativa tra il dosaggio della Semaglutide [Wegovy] e quella della Tirzepatide [Zepbound] usato negli studi.
Sappiamo altresì che la Tirzepatide ha una maggiore affinità con i recettori GIP rispetto ai recettori GLP-1. Di conseguenza, la ratio della dose di Tirzepatide con quella di Semaglutide risulta maggiormente a carico della prima.
La Tirzepatide è attualmente approvata dalla FDA per l’uso in persone in sovrappeso o con obesità, indipendentemente dal fatto che soffrano o meno di diabete di tipo II. Tuttavia, alcuni studi suggeriscono che la Tirzepatide sia un farmaco che non ha bisogno di essere somministrato in caso di mancanza della condizione diabetica.
Negli studi dove la Tirzepatide è stata somministrata a soggetti obesi, sono comunque stati osservati miglioramenti in tutte le misure cardiometaboliche. Gli eventi avversi più comuni con la Tirzepatide sono i medesimi riscontrati con la Semaglutide o altri membri della stessa famiglia. Essi sono stati di tipo gastrointestinale e la maggior parte di questi sono stati di gravità lieve o moderata e si sono verificati principalmente durante l’aumento della dose. Gli eventi avversi hanno causato l’interruzione del trattamento nel 4,3%, 7,1%, 6,2% e 2,6% dei partecipanti che hanno ricevuto dosi di Tirzepatide da 5, 10 e 15mg e placebo, rispettivamente.
In uno studio di 72 settimane su partecipanti con obesità, 5mg, 10mg o 15mg di Tirzepatide una volta alla settimana hanno fornito riduzioni sostanziali e durature del peso corporeo.
Effetto della Tirzepatide somministrata una volta alla settimana, rispetto al placebo, sul peso corporeo. Le medie dei minimi quadrati sono presentate, se non diversamente specificato. Il pannello A mostra la variazione percentuale del peso corporeo dal basale alla settimana 72, derivata da un modello di analisi della covarianza per la stima del regime di trattamento (TRE). Il pannello B mostra la variazione percentuale del peso corporeo in base alle settimane dalla randomizzazione, derivata da un modello misto per misure ripetute (MMRM) per la stima dell’efficacia; sono riportate anche le stime alla settimana 72 per la stima del regime di trattamento. I riquadri C e D mostrano le percentuali di partecipanti che hanno avuto riduzioni di peso di almeno il 5%, 10%, 15%, 20% e 25% dal basale alla settimana 72. Per il riquadro C, la percentuale è stata calcolata in base al numero di settimane di randomizzazione. Per il pannello C, la percentuale è stata calcolata con l’uso delle regole di Rubin, combinando le percentuali dei partecipanti che hanno raggiunto l’obiettivo nei set di dati imputati. I valori mancanti alla settimana 72 sono stati imputati con MMRM se la mancanza era dovuta esclusivamente a Covid-19 e con imputazione multipla se la mancanza non era dovuta a Covid-19. Per il pannello D, la percentuale di partecipanti che hanno raggiunto gli obiettivi di riduzione del peso è stata ottenuta dividendo il numero di partecipanti che hanno raggiunto i rispettivi obiettivi alla settimana 72 per il numero di partecipanti con un valore al basale e almeno un valore post-base non mancante. I valori mancanti alla settimana 72 sono stati imputati dall’analisi MMRM. Le barre 𝙸 indicano gli intervalli di confidenza al 95%.
Nella pratica su bodybuilder, si sono osservati i minori sides comparati a ottimi risultati su insulino-resistenza e riduzione contenuta della fame con dosaggi settimanali di 2.5mg.
Alla luce di ciò, e in contesto aspecifico, la Tirzepatide mostra un moderato vantaggio gestionale rispetto alla Semaglutide.
Si è parlato di un ipotetico “rebound” di Grelina con incremento della fame e del consumo calorico (con aumento di peso) dopo la cessazione d’uso di Semaglutide o Tirzepatide. Al momento non esistono dati certi che ci indichino un reale collegamento equazionale tra cessazione d’uso di GLP-1 agonisti > picco in cronico della Grelina > iperfagia > aumento ponderale di peso. Sappiamo, però, che le variazioni di Grelina e GLP-1 a 6 mesi dalla cessazione di una dieta ipocalorica non hanno predetto il recupero del peso da 6 a 18 mesi. Ciò significa che, in un soggetto sano, potrebbe si esserci una maggiore attività della Grelina nelle prime settimane dopo cessazione d’uso di un incretino-mimetico (calo soglia ematica del farmaco e stabilizzazione dei livelli endogeni di GLP-1), ma l’aumento del peso successivo potrebbe risultare con maggiore probabilità dalla modifica omeostatica multi-fattoriale la quale, per trovare un nuovo equilibrio, richiede per lo meno un arco di tempo direttamente proporzionale al tempo di trattamento. Si consideri, inoltre, che un anno dopo la sospensione della Semaglutide sottocutanea a 2,4mg una volta alla settimana e dell’intervento sullo stile di vita, i soggetti possono mostrare una riacquisizione di due terzi della perdita di peso precedente, con cambiamenti simili nelle variabili cardiometaboliche. Qualcosa di un possibile rebound grelinico…
Concentrazione totale di GLP-1 durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli mattutini (P = 0,10) e notturni (P = 0,12) tendevano a essere più bassi e i livelli pomeridiani erano significativamente più bassi (P = 0,016) durante il sonno breve rispetto al sonno abituale nelle donne, mentre negli uomini le concentrazioni pomeridiane di GLP-1 tendevano a essere più alte dopo il sonno breve rispetto al sonno abituale (P = 0,10). I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.Concentrazione di Grelina totale durante un giorno di frequenti prelievi di sangue dopo 3 notti di sonno abituale (9 ore a letto, riquadri neri) o breve (4 ore a letto, riquadri bianchi) in uomini (pannello A) e donne (pannello B). Il tempo è presentato come minuti dal campione a digiuno. Il campione a digiuno è stato prelevato alle 08:00. I pasti e gli spuntini sono stati serviti dopo i prelievi di 0, 240 e 480 minuti e a 660 minuti. L’ora di andare a letto era a 840 minuti (sonno abituale) e a 1.020 minuti (sonno breve) rispetto al prelievo a digiuno (equivalente alle 22:00 e alle 01:00 per il sonno abituale e breve, rispettivamente). I livelli di Grelina mattutina più elevati sono stati osservati dopo il sonno breve rispetto al sonno abituale negli uomini; nelle donne non sono state osservate differenze tra i periodi di sonno. I dati sono medie non aggiustate e SEM, n = 14 uomini o 13 donne.
Conclusioni:
Arrivati alla conclusione di questa disamina abbiamo tutti gli elementi per valutare l’eventuale senso di utilizzo degli Incretino-Mimetici in contesti al di fuori del trattamento del diabete di tipo II o di soggetti obesi.
Le limitazioni date dagli effetti collaterali più comuni, se contestualizzate in una preparazione di un bodybuilder, possono causare non poche problematiche specie nelle vicinanze di una gara; vedi, ad esempio, estrema riduzione dell’assunzione calorica e proteica, mal assorbimento e gonfiore addominale o paresi gastrica.
Sebbene la possibilità di eventi ipoglicemici sia contenuta, e ancor più rara con la Tirzepatide in monoterapia, il rischio, in concomitanza dell’effetto sulla insulino-sensibilità degli AAS o di altre molecole co-somministrate, di questo sides può aumentare in modo sensibile durante una dieta ipocalorica.
In tal sede non ho preso in considerazione i due più preoccupanti, e più rari, effetti collaterali legati all’uso di incretino-mimetici: Tumore Midollare della Tiroide [MTC] e Pancreatite. Quest’ultima può manifestarsi anche con l’uso di AAS, seppur raramente, specie in caso d’uso di molecole aromatizzabili; la presenza di un incretino-mimetico in tali circostanze potrebbe avere un incidenza maggiore nello sviluppo e manifestazione della Pancreatite.[https://jmedicalcasereports.biomedcentral.com/]
In definitiva, i vantaggi potenziali di una una insulino-sensibiltà maggiore iatrogeno-dipendente (visti in precedenza) con l’uso di Incretino-Mimetici è, con i dovuti distinguo complessivi, ottenibile con l’uso di Metformina la quale presenta un margine di sicurezza decisamente più ampio.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Semaglutide and Tirzepatide are More Than Just Weight Loss Drugs [di Type-IIx]
[1] McDonald, L. The Ultimate Diet 2.0: Advanced Cyclical Dieting for Achieving Super Leanness. (2003). Lyle McDonald Publishing.
[2] Volpe S, Lisco G, Racaniello D, Fanelli M, Colaianni V, Vozza A, Triggiani V, Sabbà C, Tortorella C, De Pergola G, Piazzolla G. Once-Weekly Semaglutide Induces an Early Improvement in Body Composition in Patients with Type 2 Diabetes: A 26-Week Prospective Real-Life Study. Nutrients. 2022 Jun 10;14(12):2414. doi: 10.3390/nu14122414.
[3] Kolb H, Kempf K, Röhling M, Martin S. Insulin: too much of a good thing is bad. BMC Med. 2020;18(1):224. Published 2020 Aug 21. doi:10.1186/s12916-020-01688-6
Gli incretino-mimetici sono una recente classe di farmaci antidiabete che prevede la modulazione del sistema delle Incretine. Si legano e attivano i recettori del peptide glucagone-simile-1 (GLP-1) sulle beta-cellule pancreatiche, dando inizio alla secrezione e alla sintesi di Insulina. Gli incretino-mimetici di ultima generazione hanno attività sia sul recettore del GLP-1 sia su quello del Polipeptide Inibitore Gastrico (GIP), un ormone inibitore della famiglia delle secretine (Incretine) con ruolo principale, essendo un’Incretina, di stimolare la secrezione di Insulina.
Poiché questi composti non hanno attività insulinotropica a concentrazioni di glucosio inferiori, il rischio di ipoglicemia – una nota carenza degli attuali trattamenti antidiabete – è basso. Inoltre, è stato dimostrato che gli incretino-mimetici sono associati a effetti benefici sui fattori di rischio cardiovascolare, come la perdita di peso, la riduzione della pressione sanguigna e le modifiche del profilo lipidico. Ciò nonostante, gli effetti avversi che possono causare non sono trascurabili e la loro diffusione per uso “Off-Label” ne ha mostrato le limitazioni sia in soggetti obesi che in utilizzatori per finalità estetiche.
La disamina che seguirà sarà divisa in due parti preposte a spiegare, a partire dalla loro genesi, il funzionamento delle incretine per poi passare in rassegna le caratteristiche di quelle che sono le forme farmaceutiche di incretino-mimetici utilizzate in medicina e in ambito “Off-Label/ricreativo”.
In questa prima parte vedremo nel dettaglio le incretine endogene e le molecole incretino-mimentiche presenti nel mercato…
Ormoni Incretinici [Incretine]:
Joel Habener
Negli anni ’70, Jens Juul Holst e Joel Habener iniziarono la ricerca sull’ormone GLP-1, inizialmente in relazione alla malattia dell’ulcera duodenale.[1] Esaminarono gli ormoni secreti durante l’alimentazione e li testarono su pancreas di maiale, portando alla scoperta della notevole potenza del GLP-1 nel 1988. Il loro lavoro, che in seguito ha contribuito in modo significativo al trattamento del diabete e dell’obesità, è valso a loro e a Daniel Drucker il premio della Fondazione Warren Alpert del 2021.[1] La ricerca è proseguita e nel 1993 Michael Nauck è riuscito a infondere il GLP-1 in persone affette da diabete di tipo II, stimolando l’Insulina e inibendo il Glucagone e portando la glicemia a livelli normali. Tuttavia, il trattamento dei pazienti diabetici con gli ormoni GLP-1 ha provocato notevoli effetti collaterali, inducendo i ricercatori finanziati da Novo Nordisk a cercare di sviluppare un composto adatto all’uso terapeutico.[1]
Nel 1998 un team di ricercatori di Novo Nordisk, guidato dalla scienziata Lotte Bjerre Knudsen, ha sviluppato la Liraglutide, un agonista del recettore del peptide glucagone-simile-1 con potenziale di utilizzo per il trattamento del diabete di tipo II.[2]
Gli ormoni incretinici GLP-1 e il GIP sono ormoni peptidici intestinali rilasciati in risposta all’ingestione di cibo.(3) L’effetto più importante del GLP-1 e del GIP è la loro capacità di potenziare la secrezione di Insulina indotta dal Glucosio da parte del pancreas – il cosiddetto effetto incretinico. Nei soggetti sani l’effetto incretinico rappresenta fino al 70% dell’Insulina secreta in risposta all’ingestione di Glucosio.(4) Il GLP-1 è un polipeptide di 30 aminoacidi sintetizzato dal proglucagone nelle cellule endocrine L distribuite principalmente nella mucosa della parte distale dell’intestino tenue e del colon. Il GIP è un polipeptide di 42 aminoacidi rilasciato dalle cellule endocrine K presenti nella mucosa del duodeno e della parte superiore del digiuno.(5) Mentre il GLP-1 viene rapidamente degradato (dall’enzima ubiquitario dipeptidil peptidasi-4 (DPP-4)) in circolo con un’emivita apparente di 1 – 1,5 minuti,(6) il GIP viene degradato più lentamente, con un’emivita per l’ormone intatto di 7 minuti.(7) L’ormone aumenta la secrezione di Insulina all’inizio del pasto, ma non ha alcuna attività insulinotropica a concentrazioni di glucosio inferiori (meno di 4mM), non favorendo così l’ipoglicemia. Il GLP-1 aumenta anche la biosintesi dell’Insulina e l’espressione genica della stessa. Inoltre, esercita azioni trofiche e protettive sulle beta-cellule(8) e inibisce fortemente la secrezione pancreatica di Glucagone in modo glucosio-dipendente.(9) Al contrario, è stato dimostrato che la GIP stimola la secrezione di Glucagone. Gli ormoni esercitano il loro effetto insulinotropico attraverso recettori accoppiati a proteine G sulle beta-cellule pancreatiche.(10) Oltre agli effetti sul pancreas endocrino, entrambi gli ormoni hanno diverse altre funzioni. I recettori del GLP-1 si trovano in varie regioni del cervello (11) e, una volta attivati, promuovono il senso di sazietà che, in combinazione con l’inibizione della motilità gastrointestinale indotta dal GLP-1 (mediata dal nervo vago (12) ), riduce l’assunzione di cibo e, consequenzialmente, il peso corporeo. I recettori del GLP-1 si trovano anche nel cuore (13) e la maggior parte dei dati suggerisce che il GLP-1 esercita effetti protettivi sul miocardio. È stato inoltre riscontrato che il GLP-1 riduce l’aumento postprandiale dei trigliceridi e abbassa la concentrazione di acidi grassi liberi nell’uomo.(14) Infine, studi su animali e sull’uomo indicano che il GLP-1 ha proprietà natriuretiche e diuretiche attraverso la modulazione dello scambio renale Na+/H+ (15) – un meccanismo che potrebbe servire a ridurre la pressione sanguigna. La GIP non sembra avere effetti fisiologici sul tratto gastrointestinale, sull’appetito o sull’assunzione di cibo, ma può avere un ruolo nel metabolismo dei lipidi (16) e delle ossa.(17)
Azioni del GLP-1 nei tessuti periferici. Il GLP-1 agisce direttamente sul pancreas endocrino, sul cuore, sullo stomaco e sul cervello, mentre le azioni su fegato e muscoli sono indirette.
Nei pazienti con diabete di tipo II l’effetto incretinico è gravemente ridotto.(18, 19) Questo tratto fisiopatologico gioca probabilmente un ruolo centrale nell’incapacità di questi pazienti di secernere una quantità di Insulina sufficiente a prevenire l’iperglicemia in seguito all’assunzione di glucosio per via orale.(20-30) L’attenuata secrezione postprandiale (21) e la diminuita potenza insulinotropica del GLP-1 (22), in combinazione con l’abolizione dell’effetto insulinotropico del GIP (23) , sembrano essere responsabili del ridotto effetto incretinico nei pazienti con diabete di tipo II. Poiché l’effetto insulinotropico del solo GLP-1 (e non del GIP) è conservato nei pazienti con diabete di tipo II, sono state sviluppate modalità di trattamento antidiabete basate sull’effetto di questo peptide. È interessante notare che l’infusione endovenosa (iv) di GLP-1 nativo è in grado di normalizzare la glicemia nei pazienti con diabete di tipo 2, (24) ma a causa della breve emivita del GLP-1, la somministrazione terapeutica del GLP-1 nativo non è praticabile. Pertanto, per sfruttare le azioni benefiche del GLP-1 nel diabete di tipo II, sono stati sviluppati agonisti stabili a lunga durata d’azione del recettore del GLP-1 (incretino-mimetici). Nella sezione seguente verranno descritti gli incretino-mimetici disponibili, le loro caratteristiche e applicazioni.
Ruoli contrastanti di GLP-1 e GIP sull’omeostasi del glucosio. Il GLP-1 secreto nella vena porta attiva un sensore del glucosio portale che segnala, tramite afferenze vagali, il sistema nervoso centrale e, a sua volta, le efferenze vagali aumentano la secrezione di Insulina. Sia il GLP-1 che il GIP attivano anche direttamente la secrezione di Insulina attraverso il legame con i loro recettori distinti sulle cellule β dell’isolotto.
Farmaci Incretino-mimetici:
Exenatide[approvato nel 2005/2012]
L’Exenatide, il primo di questa classe di farmaci, è stato introdotto sul mercato negli Stati Uniti nel 2005 e in Europa nel 2007 con il nome commerciale Byetta® (Amylin Pharmaceuticals/Eli Lilly).
L’Exenatide è un peptide di 39 aminoacidi; è una versione sintetica dell’exendin-4, un peptide presente nel veleno del “mostro di Gila”.
L’ Exenatide è stata isolata per la prima volta nel 1992 presso il Veterans Administration Medical Center di New York City.[25]
L’Exenatide si lega al recettore umano intatto del Peptide Glucagone-Simile-1 (GLP-1R) in modo simile al GLP-1; l’Exenatide ha un’omologia aminoacidica del 50% con il GLP-1 e ha un’emivita più lunga in vivo.[26]
Si ritiene che l’Exenatide faciliti il controllo del glucosio in almeno cinque modi:
L’Exenatide aumenta la risposta del Pancreas[27] (cioè aumenta la secrezione di Insulina) in risposta al consumo dei pasti; il risultato è il rilascio di una quantità di Insulina maggiore e più appropriata che aiuta a ridurre l’aumento della glicemia dovuto al consumo di cibo. Una volta che i livelli di glucosio nel sangue si avvicinano ai valori normali, la risposta del Pancreas alla produzione di Insulina si riduce; altri farmaci (come la rInsulina ) sono efficaci nell’abbassare la glicemia, ma possono “superare” il loro obiettivo e causare un abbassamento eccessivo della glicemia, provocando la pericolosa condizione di ipoglicemia.
L’Exenatide sopprime anche il rilascio di Glucagone da parte del Pancreas in risposta al pasto, impedendo al fegato di produrre eccessivamente glucosio quando non è necessario e prevenendo così l’iperglicemia (livelli elevati di glucosio nel sangue).
L’Exenatide contribuisce a rallentare lo svuotamento gastrico e quindi a ridurre la velocità di comparsa nel sangue del glucosio derivato dai pasti.
L’Exenatide ha un effetto sottile ma prolungato di riduzione dell’appetito e di promozione della sazietà attraverso i recettori ipotalamici (recettori diversi da quelli dell’Amilina). La maggior parte delle persone che utilizzano l’Exenatide perdono lentamente peso e, in genere, la perdita di peso maggiore è ottenuta dalle persone più in sovrappeso all’inizio della terapia con l’Exenatide. Gli studi clinici hanno dimostrato che l’effetto di riduzione del peso continua allo stesso ritmo per 2,25 anni di uso continuato. Se suddivisi in quartili di perdita di peso, il 25% più alto registra una sostanziale perdita di peso, mentre il 25% più basso non registra alcuna perdita o un lieve aumento di peso.
L’Exenatide riduce il contenuto di grasso nel fegato. L’accumulo di grasso nel fegato o la malattia del fegato grasso non alcolico (NAFLD) è fortemente correlata a diversi disturbi metabolici, in particolare a un basso livello di HDL e a Trigliceridi elevati, presenti nei pazienti con diabete di tipo II. È emerso che l’Exenatide ha ridotto il grasso epatico nei topi[28], nei ratti[29] e nell’uomo.[30]
I principali effetti collaterali del Exenatide (che condivide con tutte le molecole apopartenenti alla sua “famiglia”) sono nausea e vomito da lievi a moderati e transitori. L’incidenza dell’ipoglicemia associata al trattamento è bassa (31) – apparentemente dovuta agli effetti insulinotropi e glucagonostatici del GLP-1, dipendenti dal glucosio. Tuttavia, in combinazione con altri ipoglicemizzanti l’incidenza aumenta e dipende dalla dose di questi. Nella maggior parte degli studi con Exenatide gli episodi ipoglicemici minori sono definiti come glucosio plasmatico <3,3mM; negli studi LEAD sono definiti come glucosio plasmatico <3,1 mM. Negli studi che utilizzano Exenatide in combinazione con ipoglicemizzanti il rischio di episodi ipoglicemici minori è riportato tra il 15% e il 36%.(32) Nello studio Exenatide/Insulina Glargine l’1,5% dei pazienti ha sperimentato un’ipoglicemia grave.(33) Non c’è stata differenza tra i gruppi e l’incidenza è stata simile nei due gruppi.
Circa il 40% dei pazienti trattati con Exenatide in studi a lungo termine, controllati con placebo, ha sviluppato anticorpi contro il peptide durante le prime 30 settimane di trattamento.(32) L’esatto impatto degli anticorpi a lungo termine deve essere stabilito. L’Exenatide non è raccomandata durante la gravidanza o l’allattamento a causa della mancanza di dati sufficienti. L’Exenatide non deve essere utilizzata in pazienti con insufficienza renale, poiché viene eliminata principalmente nei reni attraverso la filtrazione glomerulare (31) e sono stati segnalati casi di insufficienza renale acuta. Nessun dato indica l’inibizione o l’induzione degli enzimi di metabolizzazione dei farmaci del citocromo P450. Dal 2005 la Food and Drug Administration (FDA) statunitense ha ricevuto 170 segnalazioni di pancreatite in pazienti trattati con Exenatide e ha ricevuto segnalazioni di pancreatite acuta, alcune delle quali erano casi gravi di pancreatite emorragica o necrotizzante in pazienti che assumevano Exenatide. Negli studi LEAD sono state osservate in totale (al 2010) 9 segnalazioni di pancreatite, nei pazienti trattati con Exanatide. E’ tutt’ora poco chiaro il nesso causale tra pancreatite ed Exenatide. Tuttavia, si raccomanda di interrompere il trattamento con incretino-mimetici in caso di sospetta pancreatite.(34) È stato suggerito che i risultati sul cancro nei roditori siano causati da un meccanismo non genotossico e specifico mediato dal recettore GLP-1 a cui i roditori sono particolarmente sensibili. La rilevanza per l’uomo è probabilmente insignificante dal punto di vista clinico, ma sono necessari ulteriori studi per chiarire i potenziali meccanismi alla base dello sviluppo del tumore delle cellule C nei pazienti trattati con analoghi del GLP-1 e le loro possibili implicazioni cliniche.(35)
Nel marzo 2013, la FDA ha pubblicato una Drug Safety Communication in cui annunciava l’avvio di indagini sui mimetici dell’incretina a causa dei risultati ottenuti da ricercatori accademici.[36] Poche settimane dopo, l’Agenzia europea per i medicinali ha avviato un’indagine simile sugli agonisti del GLP-1 e sugli inibitori della DPP-4.[37]
53 cause consolidate contro i produttori di “prodotti GLP-1/DPP-4” sono state archiviate nel 2015.[38]
Nel 2016 è stato pubblicato un lavoro che dimostra che è in grado di invertire l’alterata segnalazione del calcio nelle cellule epatiche steatotiche, che a sua volta potrebbe essere associata a un corretto controllo del glucosio.[39]
È in fase di valutazione per il trattamento del morbo di Parkinson.[40] Uno studio clinico di fase 3, iniziato nel gennaio 2020, ha avuto la sua data di completamento il 30 giugno 2024 (NCT04232969).[41]
L’Exenatide si presenta come soluzione parenterale destinata all’iniezione. Può essere somministrata nel sottocute dell’addome, del braccio o della coscia. Per migliorare la tollerabilità gastrointestinale, la terapia con Exenatide deve essere iniziata a 5mcg per dose somministrata due volte al giorno per almeno un mese. La dose di Exenatide può poi essere aumentata a 10mcg due volte al giorno.(31) La soddisfazione dei pazienti che hanno utilizzato Exenatide è stata valutata in un paio di studi. L’aggiunta di Exenatide a Metformina e altri ipoglicemizzanti ha determinato un miglioramento significativo della soddisfazione per il trattamento e della qualità di vita correlata alla salute dei pazienti dal basale a 26 settimane.(42) Il miglioramento è stato simile per i pazienti trattati con Insulina Glargine.
Liraglutide [approvato nel 2010]
La Liraglutide è un analogo acilato del GLP-1 umano e presenta il 97% di omologia di sequenza con il GLP-1 nativo. L’analogo è prodotto con la tecnologia del DNA ricombinante nel lievito.(43) Ha un effetto sul recettore del GLP-1 simile a quello descritto per l’Exenatide. Un elevato grado di legame con le proteine plasmatiche causa una minore suscettibilità al metabolismo da parte della DPP-4 e l’emivita dopo la somministrazione di Liraglutide è di circa 13 ore.(44) Questo profilo d’azione prolungato rende Liraglutide adatto alla somministrazione una volta al giorno. Non ci sono differenze clinicamente significative nella farmacocinetica di Liraglutide tra soggetti di sesso maschile e femminile, soggetti di razza diversa o soggetti anziani e giovani.(45)
La Liraglutide, venduta tra l’altro con i marchi Victoza e Saxenda, è un farmaco antidiabetico utilizzato per il trattamento del diabete di tipo II e dell’obesità cronica.[46][47] Si tratta di una terapia di seconda linea per il diabete dopo la terapia di prima linea con la Metformina.[46][48] Non sono chiari i suoi effetti sugli esiti di salute a lungo termine, come le malattie cardiache e l’aspettativa di vita.[46][49] Viene somministrata mediante iniezione sotto cutanea.[46]
La Liraglutide è stata approvata per uso medico nell’Unione Europea nel 2009 e negli Stati Uniti nel 2010.[50][51] Nel 2021 è stato il 166° farmaco più comunemente prescritto negli Stati Uniti, con oltre 3 milioni di prescrizioni.[52][53]
L’azione prolungata della Liraglutide si ottiene attaccando una molecola di acido grasso in una posizione della molecola GLP-1-(7-37), consentendole di auto-associarsi e di legarsi all’albumina nel tessuto sottocutaneo e nel flusso sanguigno. Il GLP-1 attivo viene quindi rilasciato dall’albumina a un ritmo lento e costante. Il legame con l’albumina determina inoltre una degradazione più lenta e un’eliminazione renale ridotta rispetto a quella del GLP-1-(7-37).[54]
A) La struttura molecolare del GLP-1 umano. B) La struttura molecolare di Exenatide (il colore grigio indica le differenze di struttura rispetto al GLP-1 umano). C) La struttura molecolare della Liraglutide (il colore grigio indica le differenze di struttura rispetto al GLP-1 umano).
Come abbiamo visto, La Liraglutide è un agonista acilato del recettore del GLP-1, derivato dal GLP-1-(7-37) umano, una forma meno comune di GLP-1 endogeno.
Riduce l’iperglicemia correlata ai pasti (per 24 ore dopo la somministrazione) aumentando la secrezione di Insulina (solo) quando richiesto dall’aumento dei livelli di glucosio, ritardando lo svuotamento gastrico e sopprimendo la secrezione prandiale di Glucagone.[54][55]
Quindi, la Liraglutide provoca il rilascio di Insulina nelle cellule beta pancreatiche in presenza di una glicemia elevata. Questa secrezione di Insulina si attenua quando le concentrazioni di glucosio diminuiscono e si avvicinano all’euglicemia (livello normale di glucosio nel sangue). Diminuisce inoltre la secrezione di Glucagone in modo glucosio-dipendente e ritarda lo svuotamento gastrico. A differenza del GLP-1 endogeno, la Liraglutide è stabile contro la degradazione metabolica da parte delle peptidasi, con un’emivita plasmatica di 13 ore.[56][54]
Nei pazienti ad alto rischio cardiovascolare, è stato dimostrato che la Liraglutide riduce il rischio di morte per cause cardiovascolari, infarto miocardico non fatale o ictus non fatale. Le linee guida dell’ADA considerano attualmente Liraglutide una terapia farmacologica di prima linea per il diabete di tipo II (di solito insieme alla Metformina), in particolare per i pazienti con malattie cardiovascolari aterosclerotiche o obesità.[57] Una revisione Cochrane del 2011 ha dimostrato una riduzione dell’HbA1c dello 0,24% in più con Liraglutide. Del 24% in più con Liraglutide a 1,8 mg rispetto a Insulina Glargine, 0,33% in più rispetto a Exenatide 10mcg due volte al giorno, Sitagliptin e Rosiglitazone. In uno studio randomizzato e controllato (RCT) che ha confrontato Liraglutide, Insulina Glargine, Glimepiride e Sitagliptin (tutti aggiunti alla Metformina) con un follow-up di cinque anni, Insulina Glargine e Liraglutide sono risultate modestamente più efficaci nel raggiungimento e nel mantenimento dell’HbA1c target,[58] senza alcuna differenza negli esiti delle malattie microvascolari e cardiovascolari.[59]
La Liraglutide può anche essere utilizzata insieme alla dieta e all’esercizio fisico per la gestione cronica del peso negli adulti.[46] La Liraglutide ha portato a una perdita di peso maggiore rispetto ad alcuni precedenti analoghi del peptide glucagone-simile,[60] ma è meno efficace della dose standard di Semaglutide per la perdita di peso.[61][62]
In un recente studio pubblicato nel settembre 2024, Liraglutide ha aiutato i bambini di età compresa tra i 6 e i 12 anni a ridurre l’indice di massa corporea del 7,4% in uno studio di 56 settimane.[63] Se da un lato lo studio ha mostrato i potenziali benefici del farmaco, dall’altro solleva preoccupazioni riguardo all’uso di farmaci contro l’obesità in bambini così piccoli.[64] Novo Nordisk, l’azienda innovatrice che commercializza Liraglutide, ha chiesto alle autorità di regolamentazione statunitensi ed europee di estendere l’approvazione di Saxenda anche a questa fascia d’età più giovane, dato che attualmente è approvato solo per adolescenti e adulti.[65]
aUso di 1.8mg di Liraglutide.
Come per l’Exenatide, la Liraglutide ha un effetto significativo sul peso corporeo, come dimostrano i dati relativi a Liraglutide somministrata a 1,8mg/die. Liraglutide ha ridotto il peso corporeo medio o è stato neutro rispetto al placebo o ai comparatori attivi, in monoterapia (66) e in combinazione con uno (67) o due (68) agenti antidiabete orali. Lo studio LEAD 662 ha esaminato il profilo lipidico con Exenatide e Liraglutide. Sono state osservate riduzioni significative maggiori dei trigliceridi (-0,4 vs -0,2 mM) e degli acidi grassi liberi (-0,17 vs -0,10 mM) nel gruppo Liraglutide. Entrambi i composti hanno causato una riduzione significativa della pressione arteriosa (pressione sistolica -2,2 mmHg e pressione diastolica -1,5 mmHg) senza differenze significative tra i due composti.
Tra gli effetti collaterali si annoverano ipoglicemia, nausea, vertigini, dolore addominale e dolore nel sito di iniezione.[46] Gli effetti collaterali gastrointestinali tendono a essere più forti all’inizio del periodo di trattamento e si attenuano con il tempo.[60] Altri effetti collaterali gravi possono includere angioedema, pancreatite, malattie della cistifellea e problemi renali. L’uso in gravidanza e durante l’allattamento non è sicuro.[46] Una black box warning avverte che nei ratti trattati con Liraglutide sono stati osservati tumori midollari della tiroide, ma è “Sconosciuto se Liraglutide causi tumori delle cellule C della tiroide, incluso il carcinoma midollare della tiroide (MTC), nell’uomo, poiché la rilevanza per l’uomo di tali tumori nei roditori non è stata determinata.”[46]
A proposito del MTC, a esposizioni otto volte superiori a quelle utilizzate nell’uomo, la Liraglutide ha causato un aumento statisticamente significativo dei tumori alla tiroide nei ratti. La rilevanza clinica di questi risultati è sconosciuta.[69] Negli studi clinici, il tasso di tumori alla tiroide nei pazienti trattati con Liraglutide è stato di 1,3 per 1000 anni-paziente (4 persone) rispetto a 1,0 per 1000 pazienti (1 persona) nei gruppi di confronto. L’unica persona nel gruppo di confronto e quattro delle cinque persone nel gruppo Liraglutide avevano marcatori sierici (calcitonina elevata) suggestivi di una malattia preesistente al basale.[69]
L’FDA ha dichiarato che la calcitonina sierica, un biomarcatore del carcinoma midollare della tiroide, era leggermente aumentata nei pazienti con Liraglutide, ma ancora nei limiti della norma, e che era necessario un monitoraggio continuo per 15 anni in un registro dei tumori.[70]
Un altro effetto collaterale preoccupante è rappresentato dalla possibilità (sebbene rara) di sviluppare pancreatite.
Nel 2013, un gruppo della Johns Hopkins ha riportato un’associazione con apparenza statisticamente significativa tra l’ospedalizzazione per pancreatite acuta e un precedente trattamento con derivati del GLP-1 (come la precedentemente vista Exenatide) e inibitori della DPP-4 (come il Sitagliptin).[71] In risposta, la FDA degli Stati Uniti e l’Agenzia Europea per i Medicinali hanno condotto una revisione di tutti i dati disponibili in merito alla possibile connessione tra i mimetici dell’Incretina e la pancreatite o il cancro al pancreas. In una lettera congiunta del 2014 al New England Journal of Medicine, le agenzie hanno concluso che “Un’analisi congiunta dei dati di 14.611 pazienti con diabete di tipo II provenienti da 25 studi clinici nel database di sitagliptin non ha fornito alcuna prova convincente di un aumento del rischio di pancreatite o di cancro al pancreas” e “Entrambe le agenzie concordano sul fatto che le affermazioni relative a un’associazione causale tra i farmaci a base di Incretine e la pancreatite o il cancro al pancreas, espresse di recente nella letteratura scientifica e nei media, non sono coerenti con i dati attuali”. L’FDA e l’EMA non hanno ancora raggiunto una conclusione definitiva su tale relazione causale. Sebbene la totalità dei dati esaminati fornisca rassicurazioni, la pancreatite continuerà a essere considerata un rischio associato a questi farmaci finché non saranno disponibili ulteriori dati; entrambe le agenzie continuano a indagare su questo segnale di sicurezza”[72].
Albiglutide [approvato nel 2014]
L’Albiglutide (nome commerciale Eperzan in Europa e Tanzeum negli Stati Uniti) è un farmaco agonista del GLP-1 commercializzato da GlaxoSmithKline (GSK) per il trattamento del diabete di tipo II.
L’Albiglutide è un peptide composto da 645 aminoacidi proteinogenici con 17 ponti disolfuro. Gli aminoacidi 1-30 e 31-60 costituiscono due copie di GLP-1 umano modificato, in cui l’alanina in posizione 2 è stata scambiata con una glicina per migliorare la resistenza alla DPP-4.[73] La sequenza rimanente è costituita da albumina umana.
Viene bioingegnerizzata nel lievito Saccharomyces cerevisiae utilizzando la tecnologia del DNA ricombinante.[74]
Fasi applicative della tecnologia del DNA ricombinante.
Il farmaco è stato brevettato dalla Human Genome Sciences e sviluppato in collaborazione con GSK.[75]
La GSK ha presentato domanda di approvazione alla FDA statunitense il 14 gennaio 2013 e all’Agenzia europea per i medicinali (EMA) il 7 marzo 2013. Nel marzo 2014, GSK ha ricevuto dalla Commissione Europea l’approvazione per la commercializzazione di Albiglutide con il nome di Eperzan.[76] Nell’aprile 2014, la FDA statunitense ha approvato Albiglutide con il nome di Tanzeum.[77]
Nell’agosto 2017, GSK annunciò l’intenzione di ritirare il farmaco dal mercato mondiale entro luglio 2018 per motivi economici.[78]
L’Albiglutide, come gli altri incretino-mimetici, è stato utilizzato per il trattamento del diabete di tipo II negli adulti. Può essere utilizzato da solo (se la terapia con Metformina è inefficace o non tollerata) o in combinazione con altri farmaci antidiabetici, comprese le forme di Insulina.[74]
Secondo un’analisi del 2015, l’Albiglutide è meno efficace di altri agonisti del GLP-1 per la riduzione dell’HbA1c e la perdita di peso. Sembra inoltre avere meno effetti collaterali rispetto alla maggior parte degli altri farmaci di questa classe, ad eccezione delle reazioni nel sito di iniezione che sono più comuni con Albiglutide rispetto, ad esempio, a Liraglutide.[79]
Dopo l’iniezione sottocutanea, l’Albiglutide raggiunge le massime concentrazioni ematiche dopo tre-cinque giorni. Le concentrazioni allo stato stazionario vengono raggiunte dopo tre-cinque settimane. Essendo resistente alla dipeptidil peptidasi-4 (DPP-4),[73] l’enzima che scompone il GLP-1, l’Albiglutide ha un’emivita biologica di cinque (da quattro a sette) giorni, notevolmente più lunga rispetto agli analoghi del GLP-1 più vecchi, l’Exenatide e la Liraglutide. [80][81] Ciò consente una somministrazione una volta alla settimana,[74] a differenza della Liraglutide ma come la forma a rilascio prolungato dell’Exenatide.
L’Albiglutide agisce come agonista del recettore GLP-1, il che lo rende un tipo di incretino-mimetico. Questo provoca un aumento della secrezione di insulina, soprattutto in presenza di glucosio elevato nel sangue, e rallenta anche lo svuotamento gastrico.[74]
La differenza nel meccanismo d’azione del Albiglutide con gli altri agonisti del recettore del GLP-1, dipende dalla sua struttura la quale rende difficile l’attraversamento della barriera emato-encefalica. Ciò significa che non influisce sul sistema nervoso centrale come altri agonisti del recettore del GLP-1 e potrebbe essere responsabile della limitata perdita di peso osservata con questo farmaco.[81]
Dulaglutide[approvato nel 2014]
La Dulaglutide, venduta tra l’altro con il nome commerciale Trulicity,[8] è un farmaco utilizzato per il trattamento del diabete di tipo II in combinazione con la dieta e l’esercizio fisico.[83][84] È inoltre approvato negli Stati Uniti per la riduzione degli eventi cardiovascolari avversi maggiori negli adulti con diabete di tipo II che presentano una malattia cardiovascolare conclamata o molteplici fattori di rischio cardiovascolare.[85]
Come per gli altri incretino-mimetici visti in precedenza, la Dulaglutide si lega ai recettori del GLP-1, rallentando lo svuotamento gastrico e aumentando la secrezione di Insulina da parte delle cellule β pancreatiche. Contemporaneamente, il peptide riduce l’elevata secrezione di Glucagone inibendo le cellule α del pancreas, poiché è noto che il Glucagone è elevato in modo inappropriato nei pazienti diabetici.
Più precisamente, la Dulaglutide è un agonista del recettore del GLP-1 costituito da GLP-1(7-37) legato covalentemente a un frammento Fc di IgG4 umana.
La sicurezza e l’efficacia della Dulaglutide sono state valutate in sei studi clinici in cui 3.342 soggetti con diabete di tipo II hanno ricevuto Dulaglutide. I soggetti che hanno ricevuto Dulaglutide hanno registrato un miglioramento del controllo glicemico, osservato con riduzioni del livello di HbA1c.[86]
La Food and Drug Administration (FDA) statunitense ha approvato la Dulaglutide con una strategia di valutazione e mitigazione dei rischi (REMS),[86] e ha concesso l’approvazione di Trulicity a Eli Lilly and Company.[86] La REMS consiste in una serie di misure che Eli Lilly adotterà per sensibilizzare i medici sul rischio di pancreatite e sul potenziale rischio di carcinoma midollare della tiroide associato al farmaco.[87]
Nel 2020, la FDA ha approvato due dosi più elevate del farmaco, 3,0mg e 4,5mg, sulla base dei risultati dello studio AWARD-11 che hanno dimostrato una migliore riduzione del glucosio e benefici sul peso.[88]
Il peptide è indicato per gli adulti con diabete di tipo II come aggiunta alla dieta e all’esercizio fisico per migliorare il controllo glicemico. La Dulaglutide non è indicato nel trattamento di soggetti con diabete di tipo I o di pazienti con chetoacidosi diabetica perché questi problemi sono dovuti all’incapacità delle isole pancreatiche di produrre Insulina e una delle azioni della Dulaglutide è quella di stimolare le isole funzionanti a produrre più Insulina. La Dulaglutide può essere utilizzata da solo o in combinazione con altri farmaci per il diabete di tipo II, in particolare Metformina, Sulfoniluree, Tiazolidinedioni e Insulina da assumere contemporaneamente ai pasti.[89]
Il programma di sperimentazione clinica di fase 3 del farmaco ha dimostrato riduzioni dell’emoglobina A1c di circa l’1% con le dosi di 0,75mg e 1,5mg del farmaco, insieme a una perdita di peso media di circa 5Kg. Le dosi più elevate da 3,0mg e 4,5mg, approvate nel 2020, hanno dimostrato riduzioni dell’emoglobina A1c più vicine all’1,5% e una perdita di peso leggermente superiore.[90]
DPP-4
Una meta-analisi del 2017 non ha supportato l’ipotesi che il trattamento con agonisti del GLP-1 o inibitori della DPP-4 aumenti la mortalità per tutte le cause nei diabetici di tipo II.[91]
La Dulaglutide viene assorbita lentamente dopo l’iniezione sottocutanea. In uno studio farmacocinetico condotto su 20 adulti sani, la Cmax si è verificata entro 24-48 ore dalla somministrazione. La biodisponibilità assoluta media di Dulaglutide dopo iniezioni sottocutanee di dosi singole da 0,75mg e 1,5mg è stata rispettivamente del 65% e del 47%. L’emivita media della Dulaglutide somministrato a varie dosi è stata di circa 3,75 giorni (89,9 ore). Questa emivita prolungata consente la somministrazione una volta alla settimana. Le informazioni di prescrizione indicano un’emivita di circa 5 giorni.
Gli effetti collaterali più comuni includono disturbi gastrointestinali, come dispepsia, inappetenza, nausea, vomito, dolore addominale, diarrea.[92] Alcuni pazienti possono manifestare reazioni avverse gravi: pancreatite acuta (i sintomi includono dolore addominale persistente e grave, che talvolta si irradia alla schiena ed è accompagnato da vomito), ipoglicemia, insufficienza renale (che talvolta può richiedere l’emodialisi). Il rischio di ipoglicemia aumenta se il farmaco è usato in combinazione con Sulfoniluree o Insulina.[93][94] Esiste anche un rischio potenziale di carcinoma midollare della tiroide associato all’uso del farmaco.[87]
Lixisenatide [approvato nel 2016]
La Lixisenatide (nome commerciale Lyxumia nell’Unione Europea e Adlyxin negli Stati Uniti e prodotto da Sanofi) è un agonista del recettore GLP-1 iniettabile una volta al giorno per il trattamento del diabete di tipo II.
È stato sintetizzato dalla danese Zealand Pharma A/S;[95] nel 2003 Zealand lo ha concesso in licenza a Sanofi, che ha sviluppato il farmaco.[96] La Lixisenatide è stata approvata dalla Commissione europea nel febbraio 2013.
La Lixisenatide è un peptide composto da 44 aminoacidi, con un gruppo amidico sul suo terminale C.[97]
E’ stata descritta come “des-38-prolina-exendin-4 (Heloderma suspectum)-(1-39)-peptidilpenta-L-lisil-L-lisinamide”, ovvero è derivata dai primi 39 aminoacidi della sequenza del peptide exendin-4, isolato dal veleno del “mostro di Gila”, omettendo la Prolina in posizione 38 e aggiungendo sei residui di Lisina. La sua sequenza completa è:
La Lixisenatide, appartenendo alla classe dei farmaci agonisti del GLP-1, come per i precedentemente trattati composti agisce rallentando lo svuotamento gastrico e aumentando la secrezione di Insulina da parte delle cellule β pancreatiche.
I risultati di una ricerca condotta da McClean PL et al. hanno dimostrato che la Liraglutide e la Lixisenatide sono promettenti come potenziali trattamenti farmacologici della malattia di Alzheimer AD. La Lixisenatide è risultata ugualmente efficace a una dose inferiore rispetto alla Liraglutide in alcuni dei parametri misurati dopo dieci settimane di iniezioni intraperitoneali giornaliere di Liraglutide (2,5 o 25 nmol/kg) o Lixisenatide (1 o 10 nmol/kg) o soluzione fisiologica in topi APP/PS1 a un’età in cui le placche amiloidi si erano già formate. Analizzando la plasticità sinaptica nell’ippocampo, l’LTP è stato fortemente aumentato nei topi APP/PS1 da entrambi i farmaci, con maggiore efficacia con la Lixisenatide. La riduzione del numero di sinapsi osservata nei topi APP/PS1 è stata evitata dai due farmaci. Il carico di placche amiloidi e il carico di placche Congo rosso positivo a nucleo denso nella corteccia sono stati ridotti da entrambi i farmaci a tutte le dosi. Anche la risposta infiammatoria cronica (attivazione microgliale) è stata ridotta da tutti i trattamenti.[98]
Cai HY et al. hanno dimostrato in uno studio che la lixisenatide è in grado di ridurre le placche amiloidi, i grovigli neurofibrillari e la neuroinfiammazione negli ippocampi di topi femmina APP/PS1/tau di 12 mesi; l’attivazione della via di segnalazione PKA-CREB e l’inibizione della p38-MAPK potrebbero essere i meccanismi importanti nella funzione neuroprotettiva della lixisenatide. Pertanto, la lixisenatide potrebbe avere il potenziale per essere sviluppata come nuova terapia per l’AD. [99] Liu Wet al hanno trovato risultati interessanti confrontando exendin-4 (10 nmol/kg), liraglutide (25 nmol/kg) e lixisenatide (10 nmol/kg): è emerso che exendin-4 non ha mostrato effetti protettivi alla dose scelta, mentre sia liraglutide che lixisenatide hanno mostrato effetti nel prevenire la compromissione motoria indotta da MPTP (Rotarod, locomozione in campo aperto, test di catalessi), la riduzione dei livelli di tirosina idrossilasi (TH) (sintesi di dopamina) nella substantia nigra e nei gangli della base, una riduzione della molecola di segnalazione pro-apoptotica BAX e un aumento della molecola di segnalazione anti-apoptotica B-cell lymphoma-2. I risultati precedenti dimostrano che sia la liraglutide che la lixisenatide sono superiori all’exendin-4 ed entrambi i farmaci sono promettenti come nuovo trattamento della malattia di Parkinson.[100]
Un altro studio condotto da Kerry Hunter et al. ha analizzato gli agonisti del recettore GLP-1 liraglutide e lixisenatide. Sono state valutate le cinetiche di attraversamento della barriera ematoencefalica (BBB), l’attivazione del GLP-1R attraverso la misurazione dei livelli di cAMP e gli effetti fisiologici nel cervello sulla proliferazione delle cellule staminali neuronali e sulla neurogenesi. Entrambi i farmaci sono stati in grado di attraversare la BBB. La lixisenatide ha attraversato la BBB a tutte le dosi testate (2,5, 25 o 250 nmol/kg ip.) quando misurate 30 minuti dopo l’iniezione e a 2,5-25 nmol/kg ip. 3 ore dopo l’iniezione. La lixisenatide ha anche aumentato la neurogenesi nel cervello. La liraglutide ha attraversato la BBB a 25 e 250 nmol/kg ip. ma nessun aumento è stato rilevato a 2,5 nmol/kg ip. 30 minuti dopo l’iniezione, e a 250 nmol/kg ip. a 3 ore dopo l’iniezione. Liraglutide e lixisenatide hanno aumentato i livelli di cAMP nel cervello, con lixisenatide più efficace. I risultati precedenti suggeriscono che questi nuovi analoghi dell’incretina attraversano la BBB mostrando attività fisiologica e neurogenesi nel cervello, il che li rende buoni candidati per essere utilizzati come trattamento delle malattie neurodegenerative.[101]
Anche la Lixisenatide è utilizzata come coadiuvante della dieta e dell’esercizio fisico per il trattamento del diabete di tipo II.[97] Nell’Unione Europea il suo uso è limitato all’integrazione della terapia Insulinica.[102][103] Al 2017 non è chiaro se influisca sul rischio di morte di una persona.[104]
Viene fornito in un autoiniettore contenente quattordici dosi e viene iniettato per via sottocutanea.[97]
La Lixisenatide non deve essere utilizzata da persone che hanno problemi di svuotamento gastrico.[97] La Lixisenatide ritarda lo svuotamento gastrico, il che può modificare la velocità con cui altri farmaci assunti oralmente esplicano la loro efficacia.[97]
Dopo la somministrazione sottocutanea nell’uomo, la Lixisenatide mostra una farmacocinetica lineare e un’emivita di eliminazione dipendente dall’assorbimento di 2-3 ore.
La dose iniziale di Lixisenatide è di 10mcg una volta al giorno, per 14 giorni. La dose di mantenimento è successivamente di 20mcg una volta al giorno nell’ora che precede il primo pasto della giornata o il pasto serale.
In circa lo 0,1% dei casi le persone hanno avuto reazioni anafilattiche alla lixisenatide e in circa lo 0,2% dei casi il farmaco ha causato pancreatite.[97] L’uso con insulina o sulfonilurea può causare ipoglicemia.[97] In alcuni casi, persone senza malattie renali hanno avuto lesioni renali acute e in alcune persone con malattie renali esistenti la condizione è peggiorata. Poiché la Lixisenatide è un peptide, le persone possono sviluppare una risposta immunitaria nei suoi confronti che finirà per rendere il farmaco inefficace; le persone che hanno sviluppato anticorpi contro la Lixisenatide tendono ad avere una maggiore infiammazione nel sito di iniezione.[97]
Almeno il 5% delle persone ha avuto nausea, vomito, diarrea, mal di testa o vertigini dopo l’assunzione di Lixisenatide.[97]
Semaglutide [approvata nel 2017]
La Semaglutide è chimicamente simile al GLP-1 umano.[105-41] Mancano i primi sei aminoacidi del GLP-1.[105] Le sostituzioni sono effettuate nelle posizioni 8 e 34 del GLP-1 (posizioni 2 e 28 della Semaglutide), dove l’Alanina e la Lisina sono sostituite rispettivamente dall’acido 2-aminoisobutirrico e dall’Arginina. La sostituzione dell’Alanina impedisce la degradazione chimica da parte della dipeptidil peptidasi-4.[106] La Lisina in posizione 26 del GLP-1 (posizione 20 del Semaglutide) ha una lunga catena attaccata, che termina con una catena di 17 atomi di carbonio e un gruppo carbossilico.[106] Ciò aumenta il legame del farmaco con le proteine trasportatrici nel sangue (albumina), consentendo una più lunga presenza nella circolazione sanguigna.[106]
L’emivita del Semaglutide nel sangue è di circa sette giorni (165-184 ore).
Come per gli altri incretino-mimetici, la Semaglutide è un agonista del recettore del GLP -1.[107][108][109] Il farmaco riduce i livelli di glucosio nel sangue. Sembra inoltre che aumenti la crescita delle cellule β pancreatiche, responsabili della produzione e del rilascio di Insulina.[110][111] Inoltre, inibisce la produzione di Glucagone, l’ormone che aumenta la glicogenolisi (rilascio dei carboidrati immagazzinati dal fegato) e la Gluconeogenesi (sintesi di nuovo glucosio). Riduce l’assunzione di cibo abbassando l’appetito e rallentando la digestione nello stomaco e suo svuotamento,[112] contribuendo a ridurre il peso corporeo.[113][114]
Effetti di svuotamento gastrico degli agonisti del recettore del glucagone peptide-1 ad azione breve rispetto a quelli ad azione prolungata (GLP-1RA). (A) I GLP-1RA a breve durata d’azione sopprimono lo svuotamento gastrico, prolungando la presenza di cibo nello stomaco e nella parte superiore dell’intestino tenue; il ridotto flusso transpilorico provoca un ritardo nell’assorbimento intestinale del glucosio e una diminuzione della secrezione insulinica postprandiale. I GLP-1RA a breve durata d’azione possono anche sopprimere direttamente la secrezione di glucagone. (B) I GLP-1RA a lunga durata d’azione non influenzano significativamente la motilità gastrica, a causa della tachifilassi. Invece, i GLP-1RA ad azione prolungata esercitano maggiormente il loro effetto attraverso il pancreas, aumentando la secrezione di insulina e inibendo la secrezione di glucagone attraverso il rilascio paracrino di somatostatina. Agendo sul sistema nervoso centrale, sia i GLP-1RA a più breve (A) che a più lunga durata d’azione (B) aumentano la sazietà e possono anche indurre la nausea. Adattato da Meier. Adattato su autorizzazione di Macmillan Publishers Ltd: Nature Reviews Endocrinology 2012;8(12):728-42, copyright 2012.
Nel giugno 2008 è stato avviato uno studio clinico di fase II al fine di esaminare la Semaglutide come terapia per il diabete da somministrare una volta alla settimana, come alternativa ad azione prolungata alla Liraglutide .[115][116] Gli studi clinici sono iniziati nel gennaio 2016 e si sono conclusi nel maggio 2017.[117][118]
Nel giugno 2021, una versione iniettabile a dosaggio più elevato, venduta con il marchio Wegovy, è stata approvata dalla Food and Drug Administration (FDA) statunitense come farmaco anti-obesità per la gestione del peso a lungo termine negli adulti.[119-15] Nel novembre 2021, il Comitato per i Medicinali per Uso Umano (CHMP) dell’Agenzia Europea per i Medicinali (EMA) ha raccomandato di concedere a Novo Nordisk A/S l’autorizzazione all’immissione in commercio di Wegovy[120]. Nel gennaio 2022, Wegovy è stato approvato per uso medico nell’Unione Europea.[121]
Nel gennaio 2023, l’etichetta di Rybelsus è stata aggiornata per indicare che può essere utilizzato come trattamento di prima linea per gli adulti con diabete di tipo 2.[122]
Nel marzo 2021, in uno studio di fase III randomizzato, in doppio cieco, 1.961 adulti con un indice di massa corporea pari o superiore a 30 sono stati assegnati, in un rapporto 2:1, a un trattamento con Semaglutide sottocutaneo una volta alla settimana o placebo, più un intervento sullo stile di vita. Gli studi si sono svolti in 129 siti in 16 Paesi di Asia, Europa, Nord America e Sud America. La variazione percentuale media del peso corporeo alla settimana 68 è stata di -14,9% nel gruppo Semaglutide contro -2,4% con placebo, per una differenza di trattamento stimata di -12,4 punti percentuali (95% CI, da -13,4 a -11,5).[123][124][125][126]
Una revisione dei trattamenti anti-obesità del 2022 ha rilevato che il Semaglutide e la Tirzepatide (che ha un meccanismo d’azione sovrapponibile) erano più promettenti dei precedenti farmaci anti-obesità, anche se meno efficaci della chirurgia bariatrica.[127]
Nel marzo 2023, un funzionario di Novo Nordisk ha dichiarato che i pazienti che utilizzano la Semaglutide per perdere peso possono riacquistare il peso originario entro 5 anni dall’interruzione del trattamento.[128]
Nel marzo 2024, l’FDA ha esteso l’indicazione di Semaglutide (Wegovy) per ridurre il rischio di morte cardiovascolare, infarto e ictus in adulti con malattie cardiovascolari e obesità o sovrappeso. L’efficacia e la sicurezza di questa nuova indicazione sono state studiate in uno studio multinazionale, multicentrico, in doppio cieco, controllato con placebo, che ha assegnato in modo casuale oltre 17.600 partecipanti a ricevere Semaglutide (Wegovy) o placebo.[129] I partecipanti di entrambi i gruppi hanno ricevuto anche un trattamento medico standard (ad es, Semaglutide (Wegovy) ha ridotto significativamente il rischio di eventi cardiovascolari avversi maggiori (morte cardiovascolare, infarto e ictus), che si sono verificati nel 6,5% dei partecipanti che hanno ricevuto Semaglutide (Wegovy) rispetto all’8% dei partecipanti che hanno ricevuto placebo.[129]
Una meta-analisi del 2014 ha rilevato che la Semaglutide può essere efficace nell’abbassare gli enzimi epatici (transaminite) e nel migliorare alcune caratteristiche radiologicamente osservate della malattia epatica steatotica associata a disfunzione metabolica.[130]
Nel luglio 2023, l’Agenzia islandese per i medicinali ha segnalato due casi di pensieri suicidi e un caso di autolesionismo tra i consumatori del farmaco, inducendo a valutare la sicurezza di Ozempic,[131] Wegovy, Saxenda e altri farmaci simili.[132] Nel gennaio 2024, una revisione preliminare condotta dalla FDA ha confermato che non sono state trovate prove che suggeriscano che il farmaco causi pensieri o azioni suicide.[133][134]
La Semaglutide ha dimostrato di poter ridurre l’interesse per il consumo di alcol tra gli utilizzatori. Gli scienziati ipotizzano che il Semaglutide possa influenzare le regioni cerebrali coinvolte nella dipendenza e nella regolazione dell’appetito, sebbene i meccanismi esatti siano ancora in fase di studio. La ricerca sugli animali ha indicato che farmaci simili alla Semaglutide possono ridurre l’assunzione di alcolici.[135]
La Semaglutide e farmaci simili, come la Dulaglutide e la Liraglutide, sono stati utilizzati per trattare il disturbo da alimentazione incontrollata (BED), in quanto possono minimizzare i pensieri ossessivi sul cibo e gli impulsi ad abbuffarsi.[136][137] Alcuni utilizzatori di questi farmaci hanno riferito di aver ridotto in modo significativo quello che è colloquialmente noto come “food noise” (pensieri costanti e inarrestabili di mangiare nonostante non si abbia fisicamente fame), che può essere un fattore di BED.[138][139]
Attualmente, la Semaglutide indicata come coadiuvante della dieta e dell’esercizio fisico per migliorare il controllo glicemico negli adulti con diabete di tipo II.[140][141]
La formulazione a dosi più elevate di Semaglutide è indicata come coadiuvante della dieta e dell’esercizio fisico per la gestione del peso a lungo termine negli adulti con obesità (indice di massa corporea (IMC) iniziale ≥ 30 kg/m2) o in sovrappeso (IMC iniziale ≥ 27 kg/m2) e con almeno una comorbidità correlata al peso.[142]
Nel marzo 2024, la Food and Drug Administration (FDA) statunitense ha ampliato l’indicazione di Semaglutide (Wegovy), in combinazione con una dieta a ridotto contenuto calorico e un aumento dell’attività fisica, per ridurre il rischio di morte cardiovascolare, infarto e ictus in adulti obesi o in sovrappeso con malattie cardiovascolari.[143]
La dose iniziale è di 0,25mg di Semaglutide una volta alla settimana. Dopo 4 settimane, la dose deve essere aumentata a 0,5 mg una volta alla settimana. Dopo almeno 4 settimane con una dose da 0,5 mg una volta alla settimana, la dose può essere aumentata a 1 mg una volta alla settimana per migliorare ulteriormente il controllo glicemico. Dopo almeno 4 settimane con una dose da 1 mg una volta alla settimana, la dose può essere aumentata a 2 mg una volta alla settimana per migliorare ulteriormente il controllo glicemico.
Semaglutide 0,25mg non è una dose di mantenimento. Non sono raccomandate dosi superiori a 2 mg alla settimana.
Quando Ozempic viene aggiunto alla terapia in atto a base di Metformina e/o Tiazolidinedione o dell’ inibitore del cotrasportatore sodio-glucosio (SGLT2), la dose di Metformina e/o Tiazolidinedione o dell’inibitore SGLT2 può essere mantenuta senza variazioni.
Quando Ozempic viene aggiunto alla terapia in atto con Sulfanilurea o con un’insulina, è necessario considerare una riduzione della dose di Sulfanilurea o di insulina per ridurre il rischio di ipoglicemia (vedere paragrafi 4.4 e 4.8).
Non è necessario automonitorare la glicemia per aggiustare la dose di Ozempic. L’auto-monitoraggio della glicemia è necessario per correggere la dose di Sulfanilurea e insulina, in particolare quando si inizia Ozempic e si riduce l’insulina. Si raccomanda un approccio graduale alla riduzione dell’insulina.
Similmente agli altri incretino-mimetici, possibili effetti avversi con l’uso di questo peptide includono nausea, diarrea, vomito, costipazione, dolore addominale, cefalea, affaticamento, indigestione/bruciore di stomaco, vertigini, distensione addominale, eruttazioni, ipoglicemia (basso livello di glucosio nel sangue) nelle persone con diabete di tipo II (ma non limitato ad esse), flatulenza, gastroenterite e malattia da reflusso gastroesofageo (GERD). In passato è stato sospettato di causare pancreatite e può causare gastroparesi e ostruzione intestinale.[144]Tra le persone a cui è stato prescritto un agonista del recettore del GLP-1, lo 0,1% ha ricevuto una diagnosi di gastroparesi. L’1% ha ricevuto una diagnosi di gastroparesi almeno sei mesi dopo, il che equivale a un aumento del 52% del rischio di diagnosi di gastroparesi durante l’assunzione di un farmaco di questa classe.[145] Una meta-analisi del 2019 non ha indicato un rischio significativamente elevato di pancreatite acuta.[146]Secondo il sistema di segnalazione degli eventi avversi dell’FDA (FAERS), più di 150 pazienti che assumevano Ozempic hanno riportato ileo o ostruzioni intestinali dopo l’assunzione del farmaco.[147]
Confronto visivo tra stomaco sano e stomaco con gastroparesi.
L’etichetta dell’FDA statunitense per il Semaglutide contiene un boxed warning per i tumori della tiroide a cellule C nei roditori.[148] Non è noto se il Semaglutide causi tumori della tiroide a cellule C, incluso il carcinoma midollare della tiroide, nell’uomo.[149]
Tirzepatide[approvato nel 2022]
La Tirzepatide è un farmaco antidiabetico utilizzato per il trattamento del diabete di tipo II [150][151][152][153] e per la perdita di peso.[154][155] La Tirzepatide viene somministrata tramite iniezioni sottocutanee.[150][151] Viene venduta con i marchi Mounjaro per il trattamento del diabete,[150] e Zepbound per la perdita di peso.[154] La Tirzepatide è un agonista del recettore del GIP e del GLP-1.[154]
La sintesi della Tirzepatide è stata divulgata per la prima volta nei brevetti depositati da Eli Lilly and Company.[156] Questa utilizza la sintesi standard di peptidi in fase solida, con un gruppo protettivo allilossicarbonilico sulla Lisina in posizione 20 della catena lineare degli amminoacidi, consentendo una serie finale di trasformazioni chimiche in cui l’ammina della catena laterale di tale Lisina viene derivatizzata con il frammento contenente lipidi.
Per questo composto sono stati riportati processi di produzione su larga scala.[157]
La Tirzepatide è un analogo dell’ormone GIP umano con una porzione diacidica grassa C20, utilizzata per ottimizzare l’assorbimento e il metabolismo del composto.[158] La sezione diacidica grassa (acido eicosanedioico) è legata tramite un acido glutammico e due unità di acido (2-(2-aminoetossi)etossico)acetico alla catena laterale del residuo di Lisina. Questa disposizione consente un’emivita molto più lunga, prolungando il tempo tra una dose e l’altra, grazie alla sua elevata affinità con l’albumina.[159]
Quindi, la Tirzepatide è un polipeptide lineare di 39 aminoacidi che è stato modificato chimicamente mediante lipidazione per migliorarne l’assorbimento nelle cellule e la stabilità al metabolismo.[158] Ha completato la sperimentazione di fase III a livello globale nel 2021.[160][161]
La Tirzepatide ha un’affinità maggiore per i recettori GIP rispetto ai recettori GLP-1 e questo comportamento da doppio agonista ha dimostrato di produrre una maggiore riduzione dell’iperglicemia rispetto a un agonista selettivo dei recettori GLP-1.[162] Studi di segnalazione hanno riportato che la Tirzepatide imita le azioni del GIP naturale sul recettore GIP. [Studi di segnalazione hanno riportato che la Tirzepatide imita le azioni del GIP naturale sul recettore del GIP.[163] Tuttavia, sul recettore del GLP-1, la Tirzepatide mostra una predilezione per la generazione di cAMP (un messaggero associato alla regolazione del metabolismo del glicogeno, degli zuccheri e dei lipidi), piuttosto che per il reclutamento della β-arrestina. Questa combinazione di preferenza verso il recettore GIP e di proprietà di segnalazione distinte del GLP-1 suggerisce che questo agonismo distorto aumenta la secrezione di Insulina.[163] È stato riportato che la Tirzepatide aumenta i livelli di adiponectina, un’adipochina coinvolta nella regolazione del metabolismo del glucosio e dei lipidi, con un aumento massimo del 26% rispetto al basale dopo 26 settimane, al dosaggio di 10mg.[162]
GIP e GLP-1: somiglianze e differenze. La GIP è secreta dalle cellule K dell’intestino tenue prossimale (duodeno e digiuno), mentre il GLP-1 è secreto dalle cellule L dell’intestino tenue e crasso (ileo distale e colon), in seguito all’introduzione di carboidrati, trigliceridi, proteine o aminoacidi. Un’importante eccezione è rappresentata dalla glutammina, che è uno stimolatore specifico del GLP-1. GIP e GLP-1 determinano la secrezione di Insulina in modo dipendente dal glucosio. Le azioni del GLP-1 e del GIP sulla secrezione di Glucagone sono diverse: il GLP-1 sopprime il Glucagone durante l’iperglicemia, ma non in presenza di una normale concentrazione di glucosio plasmatico a digiuno, mentre il GIP può stimolare la secrezione di Glucagone a digiuno, durante l’ipoglicemia e l’iperglicemia. Per quanto riguarda il tessuto adiposo, il GLP-1 stimola la lipolisi, mentre il GIP determina un accumulo di grasso corporeo.
Negli studi preliminari finanziati dall’industria che hanno confrontato la Tirzepatide con la Semaglutide, la Tirzepatide ha mostrato un miglioramento minore delle riduzioni (2,01%-2,30% a seconda del dosaggio) nei test dell’emoglobina glicata rispetto alla Semaglutide (1,86%). [164] Una dose di 10mg si è dimostrata efficace anche nel ridurre l’insulino-resistenza, con una riduzione di circa l’8% rispetto al basale, misurata utilizzando l’HOMA2-IR (calcolato con l’Insulina a digiuno).[162] I livelli a digiuno delle proteine che legano l’IGF, come IGFBP1 e IGFBP2, sono aumentati in seguito al trattamento con Tirzepatide, aumentando la sensibilità all’Insulina.[162]
IGFBP1
Una meta-analisi del 2021 ha mostrato che, nell’arco di un anno di utilizzo clinico, la Tirzepatide è risultata superiore a Dulaglutide, Semaglutide, Degludec e Insulina glargine per quanto riguarda l’efficacia glicemica e la riduzione dell’obesità.[165]
In uno studio di fase III, in doppio cieco, randomizzato e controllato, sostenuto da Eli Lilly, adulti non diabetici con un indice di massa corporea pari o superiore a 30, o pari o superiore a 27 e almeno una complicazione correlata al peso, escluso il diabete, sono stati randomizzati a ricevere Tirzepatide sottocutanea una volta alla settimana (5mg, 10mg o 15mg) o placebo. La variazione percentuale media del peso alla settimana 72 è stata di -15,0% (intervallo di confidenza [IC] al 95%, da -15,9 a -14,2) con dosi settimanali di Tirzepatide di 5mg, -19,5% (IC al 95%, da -20,4 a -18,5) con dosi di 10mg e -20,9% (IC al 95%, da -21,8 a -19,9) con dosi di 15mg. La variazione di peso nel gruppo placebo è stata del -3,1% (95% CI, da -4,3 a -1,9).[166][167][168]
La Tirzepatide è stata approvata per uso medico nell’Unione Europea nel settembre 2022.[169][170]
La dose iniziale di Tirzepatide è 2,5mg una volta a settimana. Dopo 4 settimane, la dose deve essere aumentata a 5mg una volta a settimana. Se necessario, è possibile aumentare la dose con incrementi di 2,5mg dopo un minimo di 4 settimane con la dose in uso.
Le dosi di mantenimento raccomandate sono 5mg, 10mg e 15mg.
La dose massima è 15mg una volta a settimana.
Quando Tirzepatide viene aggiunto alla terapia esistente con Metformina e/o inibitore del co- trasportatore di sodio-glucosio 2 (SGLT2i), può essere mantenuta la dose in uso di Metformina e/o SGLT2i.
Quando Tirzepatide viene aggiunto alla terapia esistente con una sulfonilurea e/o Insulina, si può considerare una riduzione della dose di sulfonilurea o Insulina per ridurre il rischio di ipoglicemia. L’automonitoraggio della glicemia è necessario per aggiustare la dose di sulfonilurea e Insulina. Si raccomanda un approccio graduale per la riduzione dell’Insulina.
Gli studi preclinici, di fase I e clinici di fase II hanno indicato che la Tirzepatide presenta effetti avversi simili a quelli di altri agonisti del recettore GLP-1, come visto in precedenza. Questi effetti si verificano in gran parte a livello del tratto gastrointestinale.[171] I più frequentemente osservati sono nausea, diarrea e vomito, la cui incidenza è aumentata con l’entità del dosaggio (cioè la probabilità è maggiore quanto più alta è la dose). Anche il numero di pazienti che hanno interrotto l’assunzione di Tirzepatide è aumentato con l’aumentare del dosaggio: i pazienti che assumevano 15mg avevano un tasso di interruzione del 25% rispetto al 5,1% dei pazienti che assumevano 5mg e all’11,1% di quelli che assumevano Dulaglutide.[172] In misura leggermente minore, i pazienti hanno anche riferito una riduzione dell’appetito.[171] Altri effetti collaterali segnalati sono stati dispepsia, costipazione, dolore addominale, vertigini e ipoglicemia.[173][174]
Uso off-label e “ricreativo”:
Oltre ai loro usi medici, gli agonisti del GLP-1 hanno visto una massiva diffusione in ambito della perdita di peso a fini “estetici” nel Fitness e in parte nel BodyBuilding, resa popolare da influencer e celebrità.[175] I venditori del mercato nero offrono online prodotti non autorizzati che si spacciano per agonisti del GLP-1. Questa pratica è illegale sia negli Stati Uniti che in Europa, ma alcuni acquirenti si rivolgono a rivenditori non autorizzati perché non hanno la possibilità di farsi prescrivere legalmente il farmaco.[176][177][178][179][180] Gli acquirenti, ovviamente, corrono rischi dovuti a farmaci contraffatti o di qualità inferiore venduti da soggetti non autorizzati.[181]
L’uso, le modalità di applicazione e le limitazioni degli incretino-mimetici in campo “cosmetico” saranno riportate nella seconda parte…
Holst JJ. The physiology of glucagon-like peptide 1. Physiol Rev. 2007;87:1409–1439. [PubMed] [Google Scholar]
Nauck MA, Homberger E, Siegel EG, et al. Incretin effects of increasing glucose loads in man calculated from venous insulin and C-peptide responses. J Clin Endocrinol Metab. 1986;63:492–498. [PubMed] [Google Scholar]
Brown JC, Mutt V, Pederson RA. Further purification of a polypeptide demonstrating enterogastrone activity. J Physiol. 1970;209:57–64. [PMC free article] [PubMed] [Google Scholar]
Deacon CF, Johnsen AH, Holst JJ. Degradation of glucagon-like peptide-1 by human plasma in vitro yields an N-terminally truncated peptide that is a major endogenous metabolite in vivo. J Clin Endocrinol Metab. 1995;80:952–957. [PubMed] [Google Scholar]
Vilsboll T, Agerso H, Lauritsen T, et al. The elimination rates of intact GIP as well as its primary metabolite, GIP 3–42, are similar in type 2 diabetic patients and healthy subjects. Regul Pept. 2006;137:168–172. [PubMed] [Google Scholar]
Drucker DJ. The biology of incretin hormones. Cell Metab. 2006;3:153–165. [PubMed] [Google Scholar]
Orskov C, Holst JJ, Poulsen SS, Kirkegaard P. Pancreatic and intestinal processing of proglucagon in man. Diabetologia. 1987;30:874–881. [PubMed] [Google Scholar]
Orci L, Bordi C, Unger RH, Perrelet A. Glucagon- and glicentin-producing cells. In: Lefebvre PJ, editor. Glucagon. Berlin: Springer Verlag; 1983. pp. 57–79. [Google Scholar]
Goke R, Larsen PJ, Mikkelsen JD, Sheikh SP. Distribution of GLP-1 binding sites in the rat brain: evidence that exendin-4 is a ligand of brain GLP-1 binding sites. Eur J Neurosci. 1995;7:2294–2300. [PubMed] [Google Scholar]
Flint A, Raben A, Astrup A, Holst JJ. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J Clin Invest. 1998;101:515–520. [PMC free article] [PubMed] [Google Scholar]
Gros R, You X, Baggio LL, et al. Cardiac function in mice lacking the glucagon- like peptide-1 receptor. Endocrinology. 2003;144:2242–2252. [PubMed] [Google Scholar]
Meier JJ, Gethmann A, Gotze O, et al. Glucagon-like peptide 1 abolishes the postprandial rise in triglyceride concentrations and lowers levels of non-esterified fatty acids in humans. Diabetologia. 2006;49:452–458. [PubMed] [Google Scholar]
Carraro-Lacroix LR, Malnic G, Girardi AC. Regulation of Na+/H+ exchanger NHE3 by glucagon-like peptide 1 receptor agonist exendin-4 in renal proximal tubule cells. Am J Physiol Renal Physiol. 2009;297:F1647–F1655. [PubMed] [Google Scholar]
Gault VA, O’Harte FP, Flatt PR. Glucose-dependent insulinotropic polypeptide (GIP): anti-diabetic and anti-obesity potential? Neuropeptides. 2003;37:253–263. [PubMed] [Google Scholar]
Tsukiyama K, Yamada Y, Yamada C, et al. Gastric inhibitory polypeptide as an endogenous factor promoting new bone formation after food ingestion. Mol Endocrinol. 2006;20:1644–1651. [PubMed] [Google Scholar]
Knop FK, Vilsboll T, Hojberg PV, et al. Reduced incretin effect in type 2 diabetes: cause or consequence of the diabetic state? Diabetes. 2007;56:1951–1959. [PubMed] [Google Scholar]
Nauck M, Stockmann F, Ebert R, Creutzfeldt W. Reduced incretin effect in type 2 (non-insulin-dependent) diabetes. Diabetologia. 1986;29:46–52. [PubMed] [Google Scholar]
Bagger JO, Knop FK, Lund A, Vestergaard H, Holst JJ, Vilsboll T. Impaired Regulation of the Incretin Effect in Patients with Type 2 Diabetes Mellitus. Diabetes. 2010;58 (Suppl 1):A369. [PubMed] [Google Scholar]
Vilsboll T, Krarup T, Deacon CF, Madsbad S, Holst JJ. Reduced postprandial concentrations of intact biologically active glucagon-like peptide 1 in type 2 diabetic patients. Diabetes. 2001;50:609–613. [PubMed] [Google Scholar]
Kjems LL, Holst JJ, Volund A, Madsbad S. The influence of GLP-1 on glucose-stimulated insulin secretion: effects on beta-cell sensitivity in type 2 and nondiabetic subjects. Diabetes. 2003;52:380–386. [PubMed] [Google Scholar]
Nauck MA, Heimesaat MM, Orskov C, Holst JJ, Ebert R, Creutzfeldt W. Preserved incretin activity of glucagon-like peptide 1 [7–36 amide] but not of synthetic human gastric inhibitory polypeptide in patients with type-2 diabetes mellitus. J Clin Invest. 1993;91:301–307. [PMC free article] [PubMed] [Google Scholar]
Nauck MA, Kleine N, Orskov C, Holst JJ, Willms B, Creutzfeldt W. Normalization of fasting hyperglycaemia by exogenous glucagon-like peptide 1 (7–36 amide) in type 2 (non-insulin-dependent) diabetic patients. Diabetologia. 1993;36:741–744. [PubMed] [Google Scholar]
European medicines Agency. European Public Assessment Report Byetta. 2006. EMEA/H/C/698. [Google Scholar]
Amori RE, Lau J, Pittas AG. Efficacy and safety of incretin therapy in type 2 diabetes: systematic review and meta-analysis. JAMA. 2007;298:194–206. [PubMed] [Google Scholar]
Heine RJ, Van Gaal LF, Johns D, Mihm MJ, Widel MH, Brodows RG. Exenatide versus insulin glargine in patients with suboptimally controlled type 2 diabetes: a randomized trial. Ann Intern Med. 2005;143:559–569. [PubMed] [Google Scholar]
Cvetkovic RS, Plosker GL. Exenatide: a review of its use in patients with type 2 diabetes mellitus (as an adjunct to metformin and/or a sulfonylurea) Drugs. 2007;67:935–954. [PubMed] [Google Scholar]
Knudsen LB, Nielsen PF, Huusfeldt PO, et al. Potent derivatives of glucagon-like peptide-1 with pharmacokinetic properties suitable for once daily administration. J Med Chem. 2000;43:1664–1669. [PubMed] [Google Scholar]
Elbrond B, Jakobsen G, Larsen S, et al. Pharmacokinetics, pharmacodynamics, safety, and tolerability of a single-dose of NN2211, a long-acting glucagon-like peptide 1 derivative, in healthy male subjects. Diabetes Care. 2002;25:1398–1404. [PubMed] [Google Scholar]
Damholt B, Golor G, Wierich W, Pedersen P, Ekblom M, Zdravkovic M. An open-label, parallel group study investigating the effects of age and gender on the pharmacokinetics of the once-daily glucagon-like peptide-1 analogue liraglutide. J Clin Pharmacol. 2006;46:635–641. [PubMed] [Google Scholar]
O’Neil PM, Birkenfeld AL, McGowan B, Mosenzon O, Pedersen SD, Wharton S, et al. (August 2018). “Efficacy and safety of semaglutide compared with liraglutide and placebo for weight loss in patients with obesity: a randomised, double-blind, placebo and active controlled, dose-ranging, phase 2 trial”. Lancet. 392 (10148): 637–649. doi:10.1016/S0140-6736(18)31773-2. PMID30122305. S2CID52041320.
Fox CK, Barrientos-Pérez M, Bomberg EM, Dcruz J, Gies I, Harder-Lauridsen NM, et al. (September 2024). “Liraglutide for Children 6 to <12 Years of Age with Obesity – A Randomized Trial”. The New England Journal of Medicine. doi:10.1056/NEJMoa2407379. PMID39258838.
“News details”. Novo Nordisk. Retrieved 12 September 2024.
Garber A, Henry R, Ratner R, et al. Liraglutide versus glimepiride monotherapy for type 2 diabetes (LEAD-3 Mono): a randomised, 52-week, phase III, double-blind, parallel-treatment trial. Lancet. 2009;373:473–481. [PubMed] [Google Scholar]
Marre M, Shaw J, Brandle M, et al. Liraglutide, a once-daily human GLP-1 analogue, added to a sulphonylurea over 26 weeks produces greater improvements in glycaemic and weight control compared with adding rosiglitazone or placebo in subjects with Type 2 diabetes (LEAD-1 SU) Diabet Med. 2009;26:268–278. [PMC free article] [PubMed] [Google Scholar]
Buse JB, Rosenstock J, Sesti G, et al. Liraglutide once a day versus exenatide twice a day for type 2 diabetes: a 26-week randomised, parallel-group, multinational, open-label trial (LEAD-6) Lancet. 2009;374:39–47. [PubMed] [Google Scholar]
Parks M, Rosebraugh C (March 2010). “Weighing risks and benefits of liraglutide–the FDA’s review of a new antidiabetic therapy”. The New England Journal of Medicine. 362 (9): 774–7. doi:10.1056/NEJMp1001578. PMID20164475.
Singh S, Chang HY, Richards TM, Weiner JP, Clark JM, Segal JB (April 2013). “Glucagonlike peptide 1-based therapies and risk of hospitalization for acute pancreatitis in type 2 diabetes mellitus: a population-based matched case-control study”. JAMA Internal Medicine. 173 (7): 534–539. doi:10.1001/jamainternmed.2013.2720. PMID23440284. S2CID425632.
Tibble CA, Cavaiola TS, Henry RR (May 2013). “Longer acting GLP-1 receptor agonists and the potential for improved cardiovascular outcomes: a review of current literature”. Expert Review of Endocrinology & Metabolism. 8 (3): 247–259. doi:10.1586/eem.13.20. PMID30780817. S2CID73313508.
Monami M, Dicembrini I, Nardini C, Fiordelli I, Mannucci E (February 2014). “Glucagon-like peptide-1 receptor agonists and pancreatitis: a meta-analysis of randomized clinical trials”. Diabetes Research and Clinical Practice. 103 (2): 269–275. doi:10.1016/j.diabres.2014.01.010. PMID24485345. S2CID33922845.
Christensen M, Knop FK, Holst JJ, Vilsboll T (August 2009). “Lixisenatide, a novel GLP-1 receptor agonist for the treatment of type 2 diabetes mellitus”. IDrugs. 12 (8): 503–13. PMID19629885.
McClean PL, Hölscher C (November 2014). “Lixisenatide, a drug developed to treat type 2 diabetes, shows neuroprotective effects in a mouse model of Alzheimer’s disease”. Neuropharmacology. 86: 241–58. doi:10.1016/j.neuropharm.2014.07.015. PMID25107586. S2CID24550291.
Cai HY, Yang JT, Wang ZJ, Zhang J, Yang W, Wu MN, Qi JS (January 2018). “Lixisenatide reduces amyloid plaques, neurofibrillary tangles and neuroinflammation in an APP/PS1/tau mouse model of Alzheimer’s disease”. Biochemical and Biophysical Research Communications. 495 (1): 1034–1040. doi:10.1016/j.bbrc.2017.11.114. PMID29175324.
Clinical trial number NCT00696657 for “A Randomised Controlled Clinical Trial in Type 2 Diabetes Comparing Semaglutide to Placebo and Liraglutide” at ClinicalTrials.gov
“Wegovy : Pending EC decision”. European Medicines Agency. 11 November 2021. Archived from the original on 13 November 2021. Retrieved 13 November 2021. Text was copied from this source which is copyright European Medicines Agency. Reproduction is authorized provided the source is acknowledged.
^ Abd El Aziz M, Cahyadi O, Meier JJ, Schmidt WE, Nauck MA (April 2020). “Incretin-based glucose-lowering medications and the risk of acute pancreatitis and malignancies: a meta-analysis based on cardiovascular outcomes trials”. Diabetes, Obesity & Metabolism. 22 (4): 699–704. doi:10.1111/dom.13924. PMID31750601.
US patent 9474780, Bokvist BK, Coskun T, Cummins RC, Alsina-Fernandez J, “GIP and GLP-1 co-agonist compounds”, issued 2016-10-25, assigned to Eli Lilly and Co
Ahangarpour M, Kavianinia I, Harris PW, Brimble MA (January 2021). “Photo-induced radical thiol-ene chemistry: a versatile toolbox for peptide-based drug design”. Chemical Society Reviews. 50 (2). Royal Society of Chemistry: 898–944. doi:10.1039/d0cs00354a. PMID33404559. S2CID230783854.
“Mounjaro EPAR”. European Medicines Agency (EMA). 18 July 2022. Archived from the original on 12 December 2022. Retrieved 2 January 2023. Text was copied from this source which is copyright European Medicines Agency. Reproduction is authorized provided the source is acknowledged.
Il concetto di “ricomposizione corporea” nel Fitness e nel BodyBuilding è senza dubbio considerabile come il “fattore dominante” ricercato dal momento che si tratta, molto semplicemente, del miglioramento quantitativo e qualitativo della massa contrattile (muscolo-scheletrico) a discapito della massa grassa e della ritenzione idrica extacellulare. Che si parli di “Natursl” o “Enhanced”, oltre alle variabili alimentari e allenanti vi sono quelle supplementative rappresentate, dipendentemente dalla “filosofia” scelta, da supplementi OTC e da farmaci utilizzati in ambito off-label.
Caffeina e p-Sinefrina rappresentano i lipolitici/termogenici OTC più utilizzati con un discreto margine di efficacia. Nel contesto “Enhanced”, invece, le classi di farmaci utilizzate al fine di accentuare la riduzione (direttamente o indirettamente) della massa grassa sono diverse e comprendono comunemente:
i β2-agonisti (non selettivi e selettivi) come Efedrina, Clenbuterolo e Salbutamolo;
i β3-agonisti selettivi come il Mirabegron;
gli agenti anoressizzanti con azione sui neurotrasmettitori come la Sibutramina, la Lorcaserina, l’Amfepramone e il Benfluorex;
gli anoressizzanti analoghi incretinici come la Semaglutide, il Liraglutide e il Tirzepatide;
i tiroidei Tiroxina (T4), Triiodotironina (T3) e Diiodotironina (T2);
i disaccoppianti della fosforilazione ossidativa come il 2,4-dinitrofenolo (DNP);
stimolanti il Il Peptide Natriuretico Atriale (ANP) – vedi, ad esempio, i β-bloccanti – ;
α2-antagonisti come la Yohimbina e l’α-yohimbina [Rauwolscine];
trattamenti mesoterapici a base di Fosfatidilcolina e/o Acidi Biliari.
A questo elenco, però, andrebbe aggiunta una classe di farmaci che molto poco intuitivamente ci fa pensare alla riduzione della massa grassa. Tale classe di farmaci è rappresentata dagli ACE II inibitori.
Per iniziare a comprendere del perchè questi farmaci possono rappresentare una componente funzionale nel miglioramento della composizione corporea, bisogna parlare di “Stubborn Fat” [“Grasso Testardo”]. Perchè è proprio in questa specifica e caratteristica area del tessuto adiposo che l’ACE II inibitore può contribuire alla riduzione della massa grassa.
Al fine di avere una visione di insieme più completa, è necessario trattare in modo adeguato tutte le componenti dell'”equazione”…
Tessuto adiposo e sue caratteristiche:
Il tessuto adiposo (noto anche come grasso corporeo o semplicemente grasso) è un tessuto connettivo lasso composto principalmente da adipociti.[1][2] Contiene anche la frazione vascolare stromale (SVF) di cellule tra cui preadipociti, fibroblasti, cellule endoteliali vascolari e una varietà di cellule immunitarie come i macrofagi del tessuto adiposo. Il suo ruolo non è semplicemente e solo quello di immagazzinare energia sotto forma di lipidi, ma anche di ammortizzare e isolare il corpo e rappresenta un vero e proprio organo endocrino.
Leptina
Infatti, il tessuto adiposo veniva considerato inerte dal punto di vista ormonale, ma negli ultimi anni è stato riconosciuto come un importante organo endocrino,[3] in quanto produce ormoni come Leptina, Estrogeni, Resistina e Citochine (in particolare il TNFα). Nell’obesità, il tessuto adiposo è coinvolto nel rilascio cronico di marcatori pro-infiammatori noti come adipochine, che sono responsabili dello sviluppo della sindrome metabolica, una costellazione di malattie tra cui il diabete di tipo II, le malattie cardiovascolari e l’aterosclerosi.[2][4]
Preadipociti umani sottocutanei.
Il tessuto adiposo deriva dai preadipociti e la sua formazione sembra essere controllata in parte dal gene dell’adipe. Sappiamo ormai bene che vi sono due principali tipi di tessuto adiposo, il tessuto adiposo bianco (WAT), che immagazzina energia, e il tessuto adiposo bruno (BAT), che genera calore corporeo. Il tessuto adiposo, più precisamente il tessuto adiposo bruno, è stato identificato per la prima volta dal naturalista svizzero Conrad Gessner nel 1551.[5]
Grasso Viscerale e Sottocutaneo:
Grasso Viscerale: Il grasso viscerale o addominale[6] (noto anche come grasso d’organo o grasso intra-addominale) si trova all’interno della cavità addominale, stipato tra gli organi (stomaco, fegato, intestino, reni, ecc.). Il grasso viscerale è diverso dal grasso sottocutaneo e dal grasso intramuscolare presente nei muscoli scheletrici. Il grasso nella parte inferiore del corpo, come nelle cosce e nei glutei, è sottocutaneo e non è un tessuto omogeneo, mentre il grasso nell’addome è per lo più viscerale e semi-fluido.[7] Il grasso viscerale è composto da diversi depositi adiposi, tra cui il tessuto adiposo mesenterico, il tessuto adiposo bianco epididimale (EWAT) e i depositi perirenali. Il grasso viscerale viene spesso espresso in termini di area in cm2 (VFA, visceral fat area).[8]
Da sinistra: Normale funzione dell’insulina nell’adipocita e Resistenza all’Insulina nell’adipocita.
Un eccesso di grasso viscerale è noto come obesità addominale, o “grasso della pancia”, in cui l’addome sporge eccessivamente. Nuovi sviluppi, come il Body Volume Index (BVI), sono specificamente progettati per misurare il volume addominale e il grasso addominale. L’eccesso di grasso viscerale è anche legato al diabete di tipo II,[9] all’insulino-resistenza,[10] alle malattie infiammatorie,[11] e ad altre patologie correlate all’obesità.[12] Allo stesso modo, è stato dimostrato che l’accumulo di grasso del collo (o tessuto adiposo cervicale) è associato alla mortalità.[13] Diversi studi hanno suggerito che il grasso viscerale può essere previsto da semplici misure antropometriche,[14] e predice la mortalità in modo più accurato dell’indice di massa corporea o della circonferenza vita.[15]
Gli uomini hanno maggiori probabilità di accumulare grasso nell’addome a causa delle differenze tra gli ormoni sessuali. L’estrogeno causa l’accumulo di grasso nei glutei, nelle cosce e nei fianchi delle donne.[16][17] Quando le donne raggiungono la menopausa e gli estrogeni prodotti dalle ovaie diminuiscono, il grasso migra dai glutei, dai fianchi e dalle cosce alla vita;[18] in seguito il grasso viene accumulato nell’addome.[7]
Il grasso viscerale può essere causato da un eccesso di livelli di cortisolo.[19] Almeno 10 ore MET a settimana di esercizio aerobico portano a una riduzione del grasso viscerale in chi non ha disturbi legati al metabolismo.[20] Anche l’allenamento contro-resistenza e la restrizione calorica riducono il grasso viscerale, anche se il loro effetto può non essere cumulativo.[21] Sia l’esercizio che la dieta ipocalorica causano la perdita di grasso viscerale, ma l’esercizio ha un effetto maggiore sul grasso viscerale rispetto al grasso totale. [22] L’esercizio fisico ad alta intensità è un modo per ridurre efficacemente il grasso addominale totale.[23][24] Una dieta ipocalorica combinata con l’esercizio fisico riduce il grasso corporeo totale e il rapporto tra tessuto adiposo viscerale e tessuto adiposo sottocutaneo, suggerendo una mobilitazione preferenziale del grasso viscerale rispetto al grasso sottocutaneo.[25] Il grasso addominale è fortemente soggetto alle variabili dell’Insulino-resistenza/sensibilità.
Grasso Sottocutaneo: La maggior parte del grasso non viscerale rimanente si trova appena sotto la pelle, in una regione chiamata ipoderma.[26] Questo grasso sottocutaneo non è correlato a molte delle classiche patologie legate all’obesità, come le malattie cardiache, il cancro e l’ictus, e alcune prove suggeriscono addirittura che potrebbe essere protettivo.[27] Il modello tipicamente femminile (o ginecoide) di distribuzione del grasso corporeo intorno ai fianchi, alle cosce e ai glutei è costituito da grasso sottocutaneo, e quindi rappresenta un rischio minore per la salute rispetto al grasso viscerale.[28][29]
Come tutti gli altri organi adiposi, il grasso sottocutaneo è parte attiva del sistema endocrino e secerne gli ormoni Leptina e Resistina.[26]
La relazione tra lo strato adiposo sottocutaneo e il grasso corporeo totale di una persona viene spesso modellata utilizzando equazioni di regressione. La più popolare di queste equazioni è stata creata da Durnin e Wormersley, che hanno testato in modo rigoroso molti tipi di dermoprotezione e, di conseguenza, hanno creato due formule per calcolare la densità corporea di uomini e donne. Queste equazioni presentano una correlazione inversa tra le pieghe cutanee e la densità corporea: all’aumentare della somma delle pieghe cutanee, la densità corporea diminuisce.[30]
Fattori come il sesso, l’età, le dimensioni della popolazione o altre variabili possono rendere le equazioni non valide e inutilizzabili e, a partire dal 2012, le equazioni di Durnin e Wormersley rimangono solo stime del reale livello di grassezza di una persona. Nuove formule sono ancora in fase di creazione.[30]
Gli adipociti del grasso sottocutaneo sono il target degli sforzi di manipolazione dietetica, allenante e supplementativa per ridurre al massimo la percentuale di grasso corporeo. Vi sono comunque aree di distribuzione del grasso sottocutaneo con tassi di mobilitazione lipidica differenti tra gli individui. Ed è proprio in riferimento alle aree di più difficile mobilitazione che ci si riferisce con il termina “grasso ostinato” .
Fisiologia del tessuto adiposo:
Gli acidi grassi liberi (FFA) vengono rilasciati dalla lipoproteina lipasi (LPL) ed entrano nell’adipocita, dove vengono riassemblati in trigliceridi mediante esterificazione con il glicerolo.[2] Il tessuto adiposo umano contiene circa l’87% di lipidi.[31]
Esiste un flusso costante di FFA che entrano ed escono dal tessuto adiposo.[2] La direzione netta di questo flusso è controllata dall’insulina e dalla leptina: se l’insulina è elevata, c’è un flusso netto di FFA verso l’interno e solo quando l’insulina è bassa gli FFA possono lasciare il tessuto adiposo. La secrezione di Insulina è stimolata dall’aumento della glicemia, dagli AA insulinogenici e in piccola parte dai grassi.[32]
β2-AR
Nell’uomo, la lipolisi (idrolisi dei trigliceridi in acidi grassi liberi) è controllata attraverso il settaggio equilibrato dei recettori β-adrenergici lipolitici e dell’antilipolisi mediata dai recettori α2A-adrenergici.
L’equilibrio tra β2 e α2A-AR determina le caratteristiche peculiari dell’adipocita in termini di lisi dei trigliceridi di deposito (perdita di grasso). Infatti, se l’equilibrio tende a perdersi in favore dei α2A-AR a discapito dei β2-AR ci troviamo di fronte al già prima citato “grasso testardo”.
Distribuzione degli Adrenocettori negli adipociti bianchi, bruni e beige
Gli adipociti bianchi sono il tipo di adipocita predominante nell’organismo e sono localizzati in depositi WAT distinti, caratterizzati da grasso intra-addominale (grasso viscerale che circonda gli organi interni, ovvero grasso mesenterico, perirenale e gonadico) o sottocutaneo (come il grasso inguinale). Gli adipociti bianchi immagazzinano energia (glucosio e acidi grassi) sotto forma di trigliceridi all’interno di un’unica goccia lipidica e il WAT agisce anche come organo endocrino per il rilascio di adipochine come la leptina e l’adiponectina che regolano l’omeostasi energetica dell’intero corpo (Galic, Oakhill, & Steinberg, 2010).
Differenze nella visualizzazione, nella funzione e nell’espressione dei geni firma negli adipociti bianchi, bruni e beige e l’attuale comprensione dell’espressione e della funzione degli adrenocettori (AR) nei roditori e nell’uomo. La mancata menzione di un sottotipo di adrenocettore indica che non esistono prove attuali dell’espressione/funzione della proteina recettoriale. In alcuni casi, l’evidenza funzionale si basa sull’uso di agonisti non selettivi (✦), tra cui l’Isoprenalina (Bartesaghi et al., 2015) e l’Efedrina (Carey et al., 2013) o di antagonisti (✧), tra cui la Fentolamina (Stich et al., 1999) o una combinazione di Propranololo e SR59230A per inibire tutte le risposte mediate dai β-adrenocettori (Imai et al., 2006). *L’assorbimento di 2-[18F]fluoro-2-deossiglucosio è stato misurato in risposta all’agonista selettivo dei β3-adrenocettori mirabegron nel tessuto adiposo bruno umano (Cypess et al., 2015). BA: adipocita bruno; UCP1: proteina di disaccoppiamento 1; WA: adipocita bianco
Nei roditori, tutti e tre i sottotipi di β-adrenocettori sono espressi in una serie di depositi sottocutanei e viscerali (Collins et al., 1994; Collins, Daniel, & Rohlfs, 1999; Germack, Starzec, Vassy, & Perret, 1997; Granneman, 1992; Hollenga & Zaagsma, 1989; Komai et al, 2016; Llado et al., 2002; Susulic et al., 1995), con il β3-adrenocettore che è il principale recettore responsabile della lipolisi mediata dal β-adrenocettore negli adipociti bianchi maturi. L’espressione del β-adrenocettore è influenzata anche dallo stato di differenziazione dell’adipocita bianco. L’agonista generale dei β-adrenocettori, l’Isoprenalina, ma non l’agonista altamente selettivo dei β3-adrenocettori, il CL316243, aumenta la proliferazione dei preadipociti, suggerendo un ruolo mediato dai β1-adrenocettori, mentre sia i β1-adrenocettori che i β3-adrenocettori mediano la lipolisi negli adipociti maturi (Germack et al., 1997; Klaus, Seivert, & Boeuf, 2001; Louis, Jackman, Nero, Iakovidis, & Louis, 2000; Susulic et al., 1995). È stato escluso un ruolo del β2-adrenocettore utilizzando antagonisti e agonisti selettivi del recettore.
Questi studi dimostrano collettivamente che i β-adrenocettori sono essenziali per la funzione del WAT, ma che esistono meccanismi di compensazione quando manca il β3-adrenocettore. Non ci sono prove convincenti di un contributo funzionale da parte degli α1- o α2-adrenocettori negli adipociti bianchi autentici dei roditori (Merlin, Sato, Nowell, et al., 2018). Le conoscenze sulla regolazione dell’adiponectina da parte degli adrenocettori sono meno numerose. L’adiponectina, una seconda adipochina secreta dagli adipociti bianchi e bruni, regola l’assorbimento del glucosio, la lipogenesi, la lipolisi e l’ossidazione degli acidi grassi in diversi tessuti, compreso il WAT, in modo autocrino.
Negli esseri umani, l’α1A-adrenocettore mostra una forte espressione in tutti i campioni adulti nativi, ma un’espressione trascurabile negli adipociti coltivati. Al contrario, l’mRNA per l’α1B-adrenocettore è osservato nei tessuti nativi ma anche negli adipociti differenziati di tutti i depositi, mentre l’espressione dell’α1D-adrenocettore è estremamente bassa sia nei tessuti che nelle colture primarie. L’α2A-adrenocettore mostra una forte espressione nei depositi di WAT adulto, un’espressione molto più bassa nel BAT e un’espressione bassa ma significativa nelle colture di adipociti umani maturi. L’espressione dell’α2B-adrenocettore è massima nel BAT fetale, mentre quella dell’α2C-adrenocettore è elevata nel WAT adulto e nel BAT fetale. Livelli significativi di mRNA di α2C-adrenocettori sono osservati anche negli adipociti bruni interscapolari fetali in coltura. Come accennato in precedenza, esiste un’ampia letteratura sul ruolo dei β-adrenocettori nel tessuto adiposo animale; è quindi interessante che tutti e tre i recettori siano espressi nei depositi adiposi umani nativi. Gli mRNA dei β1- e β2-adrenocettori sono presenti in tutti i depositi del BAT e del WAT, mentre l’mRNA del β3-adrenocettore è espresso principalmente nel BAT sopraclaveare adulto. Come altri marcatori termogenici, il numero di β3-adrenocettori è aumentato nel BAT sovraclaveare di un soggetto esposto al freddo (Chondronikola et al., 2016).
Rapporti precedenti hanno utilizzato la RT-PCR per dimostrare l’espressione dei β3-adrenocettori nel WAT, sebbene i segnali fossero costantemente più elevati nel BAT infantile o nel BAT perirenale (Krief et al., 1993; Lonnqvist et al., 1993; Tavernier et al., 1996). Il riscontro costante di una bassissima espressione di β3-adrenocettori nel WAT, sia da RT-PCR che da RNA-Seq, suggerisce che potrebbero esistere sottopopolazioni minori di cellule positive ai β3-adrenocettori nei depositi di WAT umano.
Le colture di adipociti derivate dalla SVF di depositi adiposi umani mostrano un’espressione trascurabile dei β3-adrenocettori, anche dopo il differenziamento in presenza di cocktail altamente adipogenici (Ding et al., 2018; Shinoda et al., 2015). L’mRNA del β1-adrenocettore è trascurabile anche negli adipociti umani primari, mentre il β2-adrenocettore è espresso nelle colture differenziate con valori medi di frammenti per kilobase per milione di reads di 1,8 (adipociti bruni sopraclavicolari) e 2,2 (adipociti bianchi sottocutanei). La mancanza di espressione dei β3-adrenocettori si verifica parallelamente a bassi livelli di mRNA per PPARGC1A, CPT1B e UCP1, tutti elementi centrali per il controllo cellulare della termogenesi. Ciò suggerisce che la differenziazione di adipociti bruni o beige termogenici è difficile da ottenere sperimentalmente negli adipociti umani primari derivati dalla SVF. Shinoda et al. (2015) hanno osservato che la differenziazione di colture clonali di adipociti bruni sopraclavicolari in presenza di 1 μM di Rosiglitazone e/o il trattamento degli adipociti maturi con 10 μM di Forskolina per 4 ore era sufficiente a indurre livelli di espressione di UCP1 simili a quelli osservati nelle biopsie scBAT native, come osservato nelle colture di adipociti bruni e beige di topo (Merlin, Sato, Chia, et al., 2018). Questo tipo di induzione potrebbe essere necessaria per promuovere l’espressione dei β3-adrenocettori, di PPARGC1A e di CPT1B.
L’espressione a basso livello dei sottotipi di β-adrenocettori è stata rilevata mediante qPCR nelle cellule staminali umane multipotenti di derivazione adiposa, con un rapporto di 3:12:1 per i β1:β2:β3-adrenocettori (Mattsson et al., 2011), ma solo gli agonisti dei β1- e β3-adrenocettori aumentano i livelli di mRNA e di proteina di UCP1 in queste cellule (Mattsson et al., 2011). Le cellule differenziate SGBS e PAZ6 sono state analizzate mediante RNA-Seq (Guennoun et al., 2015). L’espressione del β3-adrenocettore non è rilevabile nelle cellule SGBS, ma è significativa nelle cellule PAZ6 differenziate (2,5 RPKM (reads per kilobase per million mapped reads); Guennoun et al., 2015). È quindi evidente che i livelli di espressione degli adrenocettori e dei marcatori termogenici devono essere considerati in diversi sistemi modello quando si studiano potenziali agenti di “inbrunenti”.
Il WAT umano e gli adipociti bianchi dei roditori differiscono significativamente nell’espressione degli α2-adrenocettori, con un’alta espressione degli α2-adrenocettori nel WAT umano (Galitzky, Larrouy, Berlan, & Lafontan, 1990; Mauriege et al, 1991; Mauriege, Marette, et al, 1995; Mauriege, Prud’homme, Lemieux, Tremblay, & Despres, 1995), ma bassa espressione negli adipociti bianchi dei roditori (Merlin, Sato, Nowell, et al. , 2018; Valet et al., 2000). Ormai sappiamo che l’attivazione di α2-adrenocettori accoppiati a Gαi/o negli adipociti bianchi umani inibisce gli aumenti della lipolisi stimolati dalle catecolamine, contrastando così la lipolisi mediata dai β-adrenocettori (Stich et al, 1999), e gli adipociti bianchi degli esseri umani obesi presentano livelli aumentati di α2-adrenocettori, aumento di α2: β-adrenocettori e un aumento delle risposte mediate dagli α2-adrenocettori (Galitzky et al, 1990; Mauriege et al. , 1991; Mauriege, Marette, et al., 1995; Mauriege, Prud’homme, et al., 1995). Quando l’α2-adrenocettore umano è sovraespresso nel tessuto adiposo di topi KO con β3-adrenocettore, la lipolisi mediata dalla catecolamina negli adipociti bianchi è attenuata e i topi sviluppano una maggiore obesità con una dieta ad alto contenuto di grassi (Valet et al., 2000). Nonostante l’espressione significativa degli α1A- e α1B-adrenocettori nel tessuto adiposo umano nativo, non vi sono prove funzionali convincenti di un’attività diretta delle catecolamine.
“Stubborn Fat”
I due tipi di adrenocettori sopra citati, non controllano solo il metabolismo delle cellule grasse, ma anche il flusso sanguigno in entrata e in uscita da queste ultime. Di conseguenza, i β2-AR aumentano la lipolisi e il flusso sanguigno del tessuto adiposo mentre i α2A-AR inibiscono la lipolisi e il flusso sanguigno del tessuto adiposo.
Quindi, le diverse aree del grasso corporeo hanno una diversa distribuzione degli adrenorecettori β2 e α2A e questo influisce profondamente sulla capacità o meno di mobilitare e trasportare il grasso al di fuori di esse.
L’esempio più estremo è quello del grasso corporeo inferiore (fianchi e cosce), in cui è stato riscontrato un numero di recettori α2A circa 9 volte maggiore rispetto ai recettori β2. Alcune ricerche suggeriscono che il grasso addominale degli uomini ha una maggiore densità di recettori α2A (rispetto, ad esempio, al grasso viscerale), anche se non è così accentuato come per il grasso corporeo inferiore. Sebbene non sia stato studiato, è probabile che anche il grasso della parte inferiore della schiena sia relativamente resistente agli stimoli lipolitici, a causa di un numero maggiore di recettori α2A.
I dismorfismi sessuali sulla ripartizione calorica sembrano mostrare che nelle donne, dopo un pasto, può verificarsi una distribuzione calorica preferenziale nel grasso dell’area inferiore del corpo, oltre ad una ridistribuzione del grasso dalla parte superiore a quella inferiore del corpo.
Non è raro, infatti, che le donne lamentino una perdita sensibile nella parte superiore del corpo con una concomitante ed apparente peggioramento dei depositi adiposi nella parte inferiore. Una donna potrebbe mobilitare bene il grasso della parte superiore del corpo, ma immagazzinare parte di quel grasso nei depositi della parte inferiore del corpo. La parte superiore del corpo diventa più magra, quella inferiore più grassa. Questa possibilità può interessare a diverso grado anche gli uomini.
Come accennato in precedenza, oltre alle differenze nella reattività agli stimoli lipolitici, i depositi di “grasso testardo” hanno un flusso sanguigno significativamente più scarso rispetto ad altri depositi.
Alcuni studi hanno dimostrato che il flusso sanguigno nella parte inferiore del corpo può essere inferiore del 67% rispetto ad altri depositi. Il grasso viscerale ha un flusso sanguigno estremamente buono e viene mobilitato molto rapidamente.
La scarsa circolazione sanguigna ha due conseguenze importanti. In primo luogo, significa che gli ormoni trasportati dal sangue non possono raggiungere a concentrazioni ottimali le cellule adipose. In secondo luogo, un flusso sanguigno insufficiente rende più difficile far uscire il grasso mobilitato dalla cellula grassa per ossidarlo altrove.
Il motivo per cui il flusso sanguigno è così scarso non è ben definito. In parte potrebbe trattarsi semplicemente di un minor numero di vasi sanguigni, visto che gli studi di imaging ne mostrano pochi in quell’area. Inoltre, sembra che i vasi sanguigni della parte inferiore del corpo abbiano più recettori α2A che β2; ciò ha la stessa conseguenza della lipolisi. Più recettori α2A significano più vasocostrizione e meno vasodilatazione, il che si traduce in un minor flusso sanguigno.
Un fattore da tenere in considerazione è che, l’Estradiolo aumenta direttamente il numero di recettori α2A-adrenergici antilipolitici negli adipociti sottocutanei. L’aumento del numero di recettori α2A-adrenergici causa una risposta lipolitica attenuata delle Catecolamine o delle ammine simpaticomimentiche negli adipociti sottocutanei; al contrario, non è stato osservato alcun effetto degli estrogeni sull’espressione dell’mRNA dei recettori α2A-adrenergici negli adipociti del deposito di grasso intra-addominale.
Questi risultati mostrano che una cattiva gestione degli estrogeni abbassa la risposta lipolitica nel deposito di grasso sottocutaneo aumentando il numero di recettori α2A-adrenergici antilipolitici, mentre gli estrogeni non sembrano influenzare la lipolisi negli adipociti del deposito di grasso intra-addominale. Si è scoperto che questo effetto degli estrogeni è causato dal sottotipo α del recettore degli estrogeni (ERα).
Questi risultati dimostrano che una sovraespressione estrogenica attenua la risposta lipolitica attraverso la sovra-regolazione del numero di recettori α2A-adrenergici antilipolitici solo nel sottocutaneo e non nei depositi di grasso viscerale. Ciò rappresenta una spiegazione del modo in cui gli estrogeni mantengono la tipica distribuzione del grasso femminile nel sottocute, poiché gli estrogeni sembrano inibire la lipolisi solo nei depositi sottocutanei, spostando così l’assimilazione del grasso dai depositi intra-addominali a quelli sottocutanei peggiorando la situazione dei depositi di “grasso testardo” pre-esistenti e “generandone” di nuovi.
Antagonisti degli α2-AR:
Fentolamina; un α2 bloccante
Gli α2 bloccanti sono un sottoinsieme della classe dei farmaci α-bloccanti e sono antagonisti del recettore adrenergico α2. Sono utilizzati principalmente nella ricerca, avendo trovato un’applicazione clinica limitata nella medicina umana. Gli α2 bloccanti aumentano il rilascio di Noradrenalina e bloccano, per l’appunto, l’attività recettoriale degli α2-AR.
La Yohimbina, storicamente utilizzata come afrodisiaco, è talvolta impiegata in medicina veterinaria (anche se ora è stata ampiamente sostituita dall’atipamezolo) per invertire gli effetti degli α2-AR, come la Medetomidina, utilizzati come sedativi durante gli interventi chirurgici.[33]
Gli antidepressivi tetraciclici Mianserina e Mirtazapina sono α2-bloccanti , anche se la loro efficacia come antidepressivi può derivare dalla loro attività su altri siti recettoriali.
Meccanicamente, i α2-bloccanti aumentano i neurotrasmettitori adrenergici, dopaminergici e serotoninergici e inducono la secrezione di Insulina, riducendo i livelli di zucchero nel sangue.
La sospensione repentina degli α2-bloccanti può essere difficile o pericolosa, poiché la sottoregolazione globale dei neurotrasmettitori può causare sintomi di depressione e altri problemi neurologici, e l’aumento dei livelli di zucchero nel sangue insieme alla diminuzione della sensibilità all’insulina può causare in alcuni casi stati diabetici. Inoltre, può verificarsi una riduzione della microcircolazione insieme alla supersensibilità all’adrenalina in organi come il fegato.
Yohimbina e α-yohinbina
Yohimbina
Non vi è dubbio che la Yohimbina rappresenti l’α2-antagonista più usato per ridurre il grasso corporeo e, nello specifico, le zone del “grasso testardo”.
Se assunta alla dose raccomandata (≤0,2mg per kg di peso corporeo), la Yohimbina può causare nausea, dolore addominale, vertigini, nervosismo e ansia.[34]
Dosi più elevate di Yohimbina possono essere pericolose; un rapporto del 2005 ha rilevato che la Yohimbina ha il più alto tasso di effetti tossici di qualsiasi prodotto botanico.[35] Casi di ingestione di Yohimbina in eccesso hanno suggerito che l’ansia, l’ipertensione (pressione alta), la tachicardia (frequenza cardiaca elevata), le aritmie e l’agitazione sono tra gli effetti collaterali più gravi di questo composto.[35]
La Yohimbina è un α2-antagonista adrenergico selettivo. In altre parole, ha come bersaglio e inattiva una classe di recettori del sistema nervoso che risponde al neurotrasmettitore Noradrenalina.[36] L’antagonismo dei recettori α2 aumenta il rilascio di Noradrenalina da parte del sistema nervoso simpatico, causando gli effetti stimolanti e “iperadrenergici” della Yohimbina.
La Yohimbina inibisce anche l’attività dei recettori α2 sulle cellule adipose, dove la Noradrenalina agisce normalmente per sopprimere il rilascio di grasso. L’inibizione dell’effetto antilipolitico della Noradrenalina consente una maggiore lipolisi (e conseguente ossidazione lipidica).[37]
Dosi giornaliere totali di 0,2mg/kg di peso corporeo sono state utilizzate con successo per aumentare la mobilitazione lipidica dai depositi di “grasso testardo” e la successiva ossidazione dei grassi senza implicazioni significative sui parametri cardiovascolari come la frequenza cardiaca e la pressione sanguigna. Ciò si traduce in un dosaggio giornaliero totale di:
14 mg per una persona di 68 kg
18 mg per una persona di 91 kg
22 mg per una persona di 113 kg.
Queste dosi totali giornaliere si riferiscono all’uso di Yohimbina come unico agente con azione riduttiva sulla attività dei recettori α2. Tali dosaggi vengono spesso suddivise e assunte in due o quattro dosi nel corso della giornata. Ad esempio, una persona di 68 kg potrebbe assumere 7mg due volte al giorno (lontano dai pasti) per raggiungere una dose totale di 14mg.
Nota: non tutti i soggetti sono in grado di tollerare la “dose piena” ricavata dalla sopra citata formula. In quel caso, l’utilizzatore mantiene la tose tollerabile raggiunta.
Rauwlscina
Se si considera lo stesso recettore α2, la Yohimbina sembra avere una selettività per la subunità α2C piuttosto che per la A o la B; la selettività è compresa tra 4 e 15 volte,[38] mentre la Rauwolscina [α-yohimbina] sembra non essere selettiva tra queste tre subunità.[39][38] La Rauwlscina sembra essere efficace a livello del recettore quanto la Yohimbina ma con una emivita di circa 5h contro i 30 minuti della prima emivita della Yohimbina.[40]
Il fatto che la Yohimbina è selettiva per la subunità α2C più che per altre subunità, compresa l’importante A, se parliamo di α2-AR adipocitari, la sua efficacia risulta moderatamente ridotta per la riduzione del “grasso testardo”, sebbene la subunità α2C sia ad un certo grado espressa anche nel WAT; o per lo meno lo è se utilizzata come unico agente interferente l’attività adipocitaria dei α2-AR.
Introduzione agli ACE II inibitori:
Captopril
Leonard T. Skeggs e i suoi colleghi (tra cui Norman Shumway) scoprirono l’ACE [Inibitori dell’enzima di conversione dell’angiotensina] nel plasma nel 1956.[41] Le scoperte avvenute nel corso di un annosa ricerca hanno portato allo sviluppo del Captopril, il primo ACE-inibitore attivo per via orale, nel 1975.[42]
Bradichinina
Gli ACE inibitori inibiscono l’attività dell’enzima di conversione dell’angiotensina, un componente importante del sistema renina-angiotensina che converte l’angiotensina I in angiotensina II e idrolizza la bradichinina.[43] Pertanto, gli ACE inibitori diminuiscono la formazione di angiotensina II, un vasocostrittore, e aumentano il livello di bradichinina, un vasodilatatore peptidico.[43] Questa combinazione è sinergica nell’abbassare la pressione sanguigna.
Gli ACE-inibitori riducono l’attività del sistema Renina-Angiotensina-Aldosterone (RAAS) come evento eziologico (causale) primario nello sviluppo dell’ipertensione nelle persone con diabete mellito, come parte della sindrome da insulino-resistenza o come manifestazione di una malattia renale.[44][45]
Il sistema renina-angiotensina-aldosterone è un importante meccanismo di regolazione della pressione sanguigna. I marcatori di squilibrio elettrolitico e idrico nell’organismo, come l’ipotensione, la bassa concentrazione di sodio nel tubulo distale, la diminuzione del volume sanguigno e l’elevato tono simpatico, innescano il rilascio dell’enzima renina dalle cellule dell’apparato juxtaglomerulare del rene.
Renina
La renina attiva un proormone circolante derivato dal fegato, l’angiotensinogeno, mediante scissione proteolitica di tutti i suoi residui aminoacidici, tranne i primi dieci, noti come angiotensina I. L’ACE (enzima di conversione dell’angiotensina) rimuove quindi altri due residui, convertendo l’angiotensina I in angiotensina II. L’ACE si trova nella circolazione polmonare e nell’endotelio di molti vasi sanguigni.[46] Il sistema aumenta la pressione sanguigna aumentando la quantità di sale e acqua trattenuta dal corpo, sebbene l’angiotensina II sia anche un potente vasocostrittore.[47]
Struttura dell’Angiotensina I e II
Gli ACE-inibitori sono stati inizialmente approvati per il trattamento dell’ipertensione e possono essere utilizzati da soli o in combinazione con altri farmaci antipertensivi. In seguito, si sono rivelati utili per altre malattie cardiovascolari e renali[48], tra cui:
Complicanze renali del diabete mellito (nefropatia diabetica), grazie alla riduzione della pressione arteriosa e alla prevenzione del danno da iperfiltrazione glomerulare[51].
Angiotesina II e tessuto adiposo:
Noradrenalina
L’angiotensina II determina, tra le atre cose, un aumento del rilascio di catecolamine (Noradrenalina), della sensibilità alle catecolamine e della loro attività.[52]
L’angiotensina II può essere prodotta dal tessuto adiposo umano; a questo proposito, l’angiotensinogeno e gli enzimi coinvolti nella sua conversione in Ang II, nonché le vie RAS (renina, enzima di conversione dell’angiotensina: ACE) e non RAS (catepsina D, catepsina G) sono espressi nel tessuto adiposo umano. Inoltre, anche i recettori dell’Ang II sono espressi nel tessuto adiposo, il che suggerisce un ruolo locale di questo ormone nella regolazione dell’adipogenesi, del metabolismo lipidico e nella patogenesi dell’obesità28,48. L’influenza dell’Ang II sugli adipociti è mediata dall’attivazione dei recettori АТ1 e АТ2, coinvolgendo diversi sistemi di trasduzione del segnale, tra cui le risposte Са 2+, la proliferazione e la differenziazione cellulare, l’accumulo di trigliceridi, l’espressione dei geni delle adipochine e la secrezione di queste ultime [53]. L’angiotensina II ha anche un effetto anti-adipogenico, riducendo la differenziazione delle cellule pre-adipose umane [54]. Pertanto, questo ormone potrebbe rappresentare un fattore protettivo contro l’espansione incontrollata del tessuto adiposo [55].Questo effetto anti-adipogenico dell’Ang II è stato osservato anche nel grasso omentale di esseri umani affetti da obesità, con la partecipazione della via della chinasi regolata dal segnale extracellulare/1,2 (ERK/1,2) e la fosforilazione del recettore gamma attivato dal proliferatore del perossisoma (pPARG) [56]. Durante questo processo, l’origine dell’Ang II può essere sia da RAS che da vie non RAS; queste ultime potrebbero essere più importanti in questo processo [57]. Tuttavia, oltre a questo effetto, l’Ang II può aumentare il contenuto di trigliceridi e l’attività di due enzimi lipogenici (FAS: sintasi degli acidi grassi e GPDH: glicerolo-3-fosfato deidrogenasi) in colture primarie di cellule adipose umane, suggerendo un controllo dell’adiposità attraverso la regolazione della sintesi e dell’immagazzinamento dei lipidi negli adipociti [58]. L’Ang II regola anche il flusso sanguigno regionale verso il tessuto adiposo e le dimensioni e il numero delle cellule grasse [59]. Queste scoperte sono state confermate dal blocco sperimentale dell’Ang II, che influenza direttamente il peso corporeo e l’adiposità [60].
Effetti adipogenici e anti-adipogenici del sistema renina-angiotensina (RAS). La produzione locale di angiotensina II (Ang II) nel tessuto adiposo è coinvolta nella regolazione dell’adipogenesi e del metabolismo lipidico. L’Ang II ha un effetto anti-adipogenico riducendo la differenziazione adipogenica delle cellule pre-adipose umane con la partecipazione di ERK e pPARG. L’Ang II può anche aumentare il contenuto di trigliceridi negli adipociti attivando due enzimi lipogenici, FAS e GPDH. Questo effetto anti-adipogenico dell’Ang II può essere regolato. L’Ang II può essere catabolizzato dall’ACE2 adiposo per formare l’Ang 1-7 che interagisce con i recettori dell’Ang 1-7 (Mas) sugli adipociti, attivando la PI3K/Akt e l’inibizione delle vie MAPK chinasi/ ERK e inducendo un effetto inibitorio nell’Ang II/AT1 anti-adipogenico, promuovendo l’adipogenesi. AT1: Recettore-1 dell’angiotensina II; AT2: Recettore-2 dell’angiotensina II; RAS: Renin Angiotensin System; Cathep D, G: Cathepsin D, Cathepsin G; ACE1: angiotensin-converting enzyme-1; ACE2: angiotensin-converting enzyme-2; Ang 1-7: Angiotensina 1-7; ERK: extracellular signal-regulated kinase; pPARG: phosphorylated peroxisome proliferator-activated receptor gamma; FAS: fatty acid synthase; GPDH: glicerolo-3-fosfato deidrogenasi; MAPK chinasi/ERK: mitogen-activated protein kinases/extracellular signal-regulated kinases; PI3K/Akt: fosfatidilinositolo 3-chinasi/proteina chinasi B.
È stata documentata anche la regolazione autocrina dell’Ang II durante l’adipogenesi. L’angiotensina II può essere catabolizzata nei tessuti adiposi dall’enzima adiposo di conversione dell’angiotensina 2 (ACE2) per formare l’Ang 1-7. La regolazione autocrina del sistema angiotensinico locale implica la coespressione dei recettori dell’Ang II (AT1 e AT2) e dei recettori dell’Ang 1-7 (Mas) sugli adipociti. L’attivazione del recettore Mas da parte dell’Ang 1-7 ha un effetto contrario all’effetto anti-adipogenico dell’Ang II, inducendo l’adipogenesi attraverso l’attivazione delle vie PI3K/Akt e l’inibizione delle vie MAPK chinasi/ERK [61] . In questo contesto, la regolazione autocrina dell’asse Ang II/AT1-ACE2-Ang 1-7/Mas durante l’adipogenesi è in grado di produrre ormoni e citochine che promuovono l’infiammazione, l’accumulo di lipidi, l’IR e le componenti del RAS, che si attivano in presenza di obesità come meccanismi chiave correlati all’obesità dell’ipertensione e di altre componenti della sindrome cardiometabolica [62].
Angiotesina II e α2A-AR
Una caratteristica di particolare interesse in riferimento all’Angiotesina II è il fatto che sia un polipeptide necessario per l’espressione di alcuni recettori α2 (ma non di tutti). Ciò significa che senza l’Angiotensina II i recettori α2 non possono essere sviluppati in alcune cellule. Di conseguenza, se sottoregoliamo l’Angiotensina II, prodotta naturalmente dall’organismo, il normale rinnovamento dei recettori α2 non avverrà. Bisogna capire che in ogni cellula c’è un costante rinnovamento recettoriale. Bloccando la formazione di un tipo specifico di recettore in una cellula (ad esempio i recettori α2), dopo un po’ di tempo non ci saranno più recettori α2 in questa cellula. I vecchi recettori saranno completamente degradati e avremo impedito alla nuova generazione di recettori di sostituire quelli vecchi.
Attività dell’Angiotesina II a livello dei α2A-AR e del Recettore dell’Angiotesina II dell’adipocita del WAT
Quindi, sotto-regolazione marcata dei α2 recettori . Il problema principale è se questa azione dell’Angiotensina II avviene nelle cellule adipose. L’Angiotensina II agisce solo sui recettori α2 che rispondono a due condizioni:
Sembra avere il massimo effetto sui recettori α2 del sottotipo “A”. Ciò è positivo, poiché sono proprio questi recettori a trovarsi nelle cellule adipose. Quindi, la prima condizione è soddisfatta.
L’Angiotensione II agisce solo sulle cellule ricche di recettori α2 e di recettori dell’Angiotensina II. Sappiamo già che le cellule adipose sono molto ricche di recettori α2. Da tempo i ricercatori sanno anche che le cellule adipose sono ricche di recettori dell’Angiotensina II.
Il punto chiave da ricordare è che nelle cellule grasse l’Angiotensina II è necessaria perché i recettori α2 si rinnovino normalmente. Se impediamo in qualche modo la formazione di Angiotensina II, causeremo grossi problemi nel rinnovo dei recettori α2A nelle cellule adipose.
Quindi, tutto ciò che occorre fare è alterare la produzione di Angiotensina II attraverso l’uso principale di ACE II inibitori. Nel giro di poche settimane il numero di recettori α2 diminuirà sensibilmente.
Quinapril
L’uso di ACE II inibitori ha quindi il potenziale di attenuare la sensibilità agli α2-adrenocettori negli adipociti umani. L’effetto del Quinapril, un ACE II inibitore lipofilo, è stato maggiore di quello dell’Enalapril [www.ncbi.nlm.nih.gov/pmc/articles], un ACE II inibitore idrofilo. Gli ACE II inibitori lipofili possono avere un effetto vasodilatatore più potente rispetto agli ACE II inibitori idrofili. La concentrazione di Angiotensina II nei tessuti piuttosto che nel plasma può contribuire alla sensibilità e il numero degli α2-adrenocettori.
Enalapril
È stato riportato che l’ACE inibitore lipofilo, Quinapril, riduce la concentrazione tissutale di Angiotensina II in misura maggiore rispetto all’ACE inibitore idrofilo, Enalapril, da 5 a 24 ore dopo una singola somministrazione orale nei ratti. I tempi di raggiungimento della concentrazione plasmatica massima del Quinapril e del suo metabolita attivo sono stati di 2-3 ore [63, 64]. Pertanto, per esaminare più chiaramente la cosa, ciascun farmaco è stato somministrato 22 e 3 ore prima dell’esame. Entrambi gli ACE inibitori hanno soppresso le attività plasmatiche dell’ACE per oltre il 90%. Questo risultato conferma i precedenti risultati ottenuti in soggetti giapponesi [65]. Sebbene la soppressione dell’attività dell’enzima convertitore dell’Angiotensina nel plasma e la pressione arteriosa sistemica non differissero tra i due farmaci, l’attenuazione della sensibilità degli α-adrenocettori alla Fenilefrina era maggiore nei soggetti trattati con Quinapril rispetto a quelli trattati con Enalapril. Le osservazioni e i rapporti precedenti [66] suggeriscono che la concentrazione di Angiotensina II nei tessuti piuttosto che quella nel plasma può contribuire alla sensibilità dei recettori α-adrenergici nei vasi ed in altri tessuti come quello adiposo. Inoltre, l’ACE inibitore lipofilo può essere più potente dell’ACE inibitore idrofilo. Infatti, il Quinapril ha attenuato la risposta vasopressore della Fenilefrina più dell’Enalapril e l’intervallo di confidenza del 95% per le differenze di ED50 tra Enalapril e Qinapril è stato di 31,1-397,5. Sebbene l’entità dell’attenuazione della sensibilità dei recettori α-adrenergici indotta dalla soppressione dell’ACE tissutale con Quinapril fosse varia, ciò è coerente con un altro esperimento in vitro [67].
Quando osservata, la concentrazione di Noradrenalina nel siero durante il riposo a letto non è cambiata prima e dopo la somministrazione del farmaco ACE inibitore. Rapporti precedenti hanno dimostrato che gli ACE inibitori attenuano il deflusso del nervo simpatico negli animali e nell’uomo [68, 69]. Negli studi in cui non è stato applicato alcun carico al sistema nervoso simpatico, non è stato possibile rilevare alcun cambiamento nel flusso simpatico indotto dagli ACE inibitori.
Applicazione degli ACE II inibitori nel trattamento del “grasso testardo”:
La genesi dell’uso degli ACEI come PEDs
Daniel (“Dan”) Duchaine
Nonostante il potenziale maggiore nella sotto-regolazione degli α2A-AR attribuita agli ACE inibitori con caratteristiche prettamente lipolifiche, la molecola appartenente a questa classe di farmaci maggiormente utilizzata per tale scopo e da più tempo è il Captopril. Questo storico ACE II inibitore mostra però caratteristiche idrofile. Certo, la sua maggiore diffusione è legata senza dubbio agli anni dalla sintesi e immissione nel circuito farmaceutico della molecola, ma anche, e soprattutto, al suo lancio come PEDs da parte, tra i primi, di Dan Duchaine (1952-2000).
Le proprietà potenziali sulla composizione corporea del Captopril vennero individuare per la prima volta in alcune atlete interessate ad assumere un farmaco che le desse un miglioramento della composizione corporea ma senza virilizzazione. Così quella divenne l’occasione giusta per testare il Captopril. La dieta delle atlete non venne cambiata. Le atlete hanno continuato per un paio di mesi ad assumere il Captopril come unico farmaco. Avevano migliorato leggermente il trofismo, ma non molto. Ciò che però colpì i “pionieri della preparazione” fu il fatto che avevano perso grasso in aree in cui prima non erano riuscite a perderlo in modo significativo.
Approfondendo le caratteristiche della molecola attraverso la consultazione di testi accademici reperiti alla biblioteca medica, scoprirono che la relazione tra il Captropril e i recettori α2.
Con il procedere del tempo e le sperimentazione dose-tempo nell’applicazione del Captopril (ma non solo), si è notato che il farmaco poteva rendere possibile la riduzione totale della dose di Yohimbina migliorando notevolmente la compliance dell’utilizzatore.
Sappiamo, infatti, che la Yohimbina presenta una selettività maggiore per i recettori α2C piuttosto che ai sottogruppi “A” e “B”. Questa caratteristica risulta limitativa nell’azione ricercata nella Yohimbina come α2-antagonista adipocitario. L’inserimento del Captopril [o di altro ACE II inibitore] permette di 1) ridurre sensibilmente il numero di α2A-AR nell’adipocita e 2) di permettere, a dosaggio di 1/2 fino a 1/3, un legame antagonista da parte della Yohimbina nei confronti degli α2-AR rimasti. L’uso della α-yohimbina, non presentando tale affinità selettiva, migliora sensibilmente questo effetto sinergico.
Situazione adipocitaria in fisiologia con attività catecolaminergica a livello degli adrenocettori nel adipocita; Impatto sulla attività adrenorecettoriale con somministrazione di Yohimbina; Impatto sulla densità/numero adrenorecettoriale con somministrazione di un ACE II inibitore [Captopril]; Impatto additivo sulla densità, numero, funzionalità e attivazione adrenorecettoriale con somministrazione di Yohimbina e un ACE II inibitore [Captopril].
Le limitazioni degli ACE II inibitori
Il Captopril [e in generale gli ACE II inibitori] non è un farmaco che manifesta rapidamente i suoi effetti dal punto di vista estetico. Bisogna ricordare che la regolazione degli α2-AR richiede almeno due mesi prima di diventare significativa.
È necessario seguire una dieta ipocalorica per vedere ottimi risultati in termini di perdita di grasso ostinato. Abbiamo detto, infatti, che i recettori α2-AR impediscono la normale perdita di grasso la dose si presentano in maggiori concentrazioni. Questo non significa, però, che si perderà automaticamente grasso di deposito solo perché si è ridotto il numero di recettori α2. Significa solo che la perdita di grasso ostinato indotta dalla dieta ipocalorica sarà più “facile”. Avrà un effetto permissivo sulla perdita di grasso ostinato, consentendo di ridurre i depositi adiposi con un rapporto di α2-AR più elevato.
L’ultima limitazione è che esiste ancora una linea di difesa per le cellule adipose e la conservazione delle riserve lipiche. Eliminando parzialmente la linea di difesa rappresentata dagli α2-AR, se ne attiva una nuova costituita da recettori antilipolitici chiamati peptide YY, anch’essi localizzati sulle cellule adipose. Ciò significa che la riduzione del livello dei recettori α2-AR permetterà di perdere più grasso ostinato di quanto sarebbe stato normalmente possibile, ma le limitazioni genetiche saranno sempre presenti.
Ma l’uso di Captopril [o altro ACE II inibitore] può permettere di fare un grande passo avanti nella giusta direzione se l’obbiettivo è una marcata riduzione della body fat, soprattutto le aree ostinate.
Esempi applicativi degli ACE II inibitori per il trattamento del “Stubborn Fat”
Nell’approccio protocollare di base, e se prendiamo come esempio di ACE II inibitore il Captopril:
Captopril = 50mg/die [da raggiungere con gradualità e aumenti giornalieri di 6,25mg];
Yohimbina = 5-10mg/die [dose da raggiungere con aumenti giornalieri (pari a 2,5mg) e test della sensibilità ];
α-yohimbina = 3/5mg die [dose da raggiungere con aumenti giornalieri (pari a 0,5mg) e test della sensibilità ].
Nell’approccio protocollare intermedio:
Captorpil = 50-75mg/die [da raggiungere con gradualità e aumenti giornalieri di 6,25mg];
Yohimbina = 10mg/die [dose da raggiungere con aumenti giornalieri (pari a 2,5mg) e test della sensibilità ];
α-yohimbina = 5-6mg die [dose da raggiungere con aumenti giornalieri (pari a 0,5mg) e test della sensibilità ];
T3 = 25mcg/die [dose da raggiungere con aumenti giornalieri (pari a 12,5mcg) e controllo ematico del FT3].
Nell’approccio protocollare avanzato:
Captorpil = 100mg/die [da raggiungere con gradualità e aumenti giornalieri di 6,25mg];
Yohimbina = 0.2mg/Kg/die [dose da raggiungere con aumenti giornalieri (pari a 2,5mg) e test della sensibilità ];
α-yohimbina = 0.1mg/Kg/die [dose da raggiungere con aumenti giornalieri (pari a 0,5mg) e test della sensibilità ];
T3 = 50mcg/die [dose da raggiungere con aumenti giornalieri (pari a 12,5mcg) e controllo ematico del FT3];
Salbutamolo = 8-12mg/die [dose da raggiungere con aumenti ogni 1-2 giorni (pari a 2mg)];
Alternativa: Clenbuterolo = 1mcg/Kg/die (range 40-80mcg) [dose da raggiungere con aumenti ogni 2 giorni (pari a 10-20mcg) e test della sensibilità/tolleranza];
Nedbivololo = 5mg/die [dose di partenza 2,5mg/die e valutazione della tolleranza].
*Nota bene: Nessuno dei protocolli sopra esposti rappresenta un indicazione d’uso o una prescrizione medica di applicazione. Tali informazioni SONO AD ESCLUSIVO SCOPO ESEMPLIFICATIVO!
Effetti collaterali degli ACE II inibitori
pressione bassa;
tosse. Un altro possibile effetto avverso specifico degli ACE-inibitori, ma non di altri bloccanti del RAAS, è l’aumento del livello di bradichinina. La tosse secca persistente è un effetto avverso relativamente comune che si ritiene sia associato all’aumento dei livelli di bradichinina prodotto dagli ACE inibitori, anche se il ruolo della bradichinina nella produzione di questi sintomi è stato contestato. Tuttavia, molti casi di tosse in persone che assumono ACE inibitori potrebbero non essere dovuti al farmaco stesso. Alcuni (0,7%) sviluppano angioedema a causa dell’aumento dei livelli di bradichinina. Può esistere una predisposizione genetica. ;
iperkaliemia. Il potassio elevato nel sangue è un’altra possibile complicazione del trattamento con un ACE-inibitore, dovuta al suo effetto sull’aldosterone. La soppressione dell’angiotensina II porta a una diminuzione dei livelli di aldosterone. Poiché l’aldosterone è responsabile dell’aumento dell’escrezione di potassio, gli ACE-inibitori possono causare una ritenzione di potassio. Alcune persone, tuttavia, possono continuare a perdere potassio durante l’assunzione di un ACE-inibitore. È necessario un attento monitoraggio dei livelli di potassio nei soggetti in trattamento con ACE-inibitori che sono a rischio di iperkaliemia.;
cefalea;
vertigini;
affaticamento;
nausea e compromissione renale. I soggetti che iniziano la terapia con un ACE-inibitore presentano di solito una modesta riduzione della velocità di filtrazione glomerulare (eGFR). Tuttavia, la riduzione può essere significativa in condizioni di preesistente ridotta perfusione renale, come stenosi dell’arteria renale, insufficienza cardiaca, malattia renale policistica o deplezione di volume. Una moderata riduzione della funzione renale, non superiore al 30% di aumento della creatinina sierica, che si stabilizza dopo una settimana di trattamento. La riduzione del eGFR è un problema soprattutto se il paziente assume contemporaneamente un FANS e un diuretico. Quando i tre farmaci vengono assunti insieme, il rischio di sviluppare un’insufficienza renale aumenta notevolmente.
Una rara reazione allergica grave può colpire la parete intestinale e causare secondariamente dolore addominale.
Ma gli ARB/Sartani possono essere un sostituto agli ACE II inibitori per lo scopo qui discusso?
Telmisartan
Sono circa vent’anni che si è scoperto che il Telmisartan, un Bloccante del Recettore dell’Angiotensina II (ARB) approvato per il trattamento dell’ipertensione, è anche un agonista parziale di PPARγ.[70-71] Mentre gli agonisti completi di PPARγ, come il Rosiglitazone e il Pioglitazone, promuovono l’aumento di peso alterando la distribuzione del grasso e la differenziazione degli adipociti, gli agonisti parziali (agonisti/antagonisti misti) di PPARγ possono avere la capacità di ritardare l’aumento di peso promuovendo al contempo la differenziazione degli adipociti.[72] Ad esempio, è stato scoperto che il Telmisartan può promuovere la differenziazione degli adipociti ma anche attenuare l’aumento di peso, migliorando al contempo il metabolismo del glucosio e dei lipidi nei ratti alimentati con una dieta ad alto contenuto di grassi e carboidrati.[70] Sharma et al[73] hanno riportato che il blocco del recettore dell’angiotensina II di tipo 1, di per sé, può promuovere la differenziazione degli adipociti e hanno proposto che questo possa contribuire agli effetti antidiabetici degli antagonisti del recettore dell’angiotensina II. Non è noto se molecole bifunzionali come il Telmisartan, che attivano PPARγ e bloccano il recettore dell’angiotensina II, esercitino effetti diversi sulle dimensioni degli adipociti e sui determinanti primari del peso corporeo rispetto ai normali bloccanti del recettore dell’angiotensina, come il Valsartan, che non hanno la capacità di attivare PPARγ.
Valsartan
Negli studi si è scoperto che il Telmisartan, ma non il Valsartan, aumenta l’espressione dei geni di un fattore di trascrizione nucleare (TFAM) che regola la funzione mitocondriale e di una proteina mitocondriale (MTCO1) coinvolta nella fosforilazione ossidativa. Rispetto agli agonisti totali convenzionali di PPARγ, come i Tiazolidinedioni, gli agonisti parziali del PPARγ, come il Telmisartan, possono avere la capacità di reclutare in modo preferenziale alcuni coattivatori trascrizionali che sono particolarmente importanti nella regolazione dei geni che controllano la funzione mitocondriale e il metabolismo energetico.[74-75] Ad esempio, gli agonisti parziali sembrano reclutare preferenzialmente il coattivatore 1-α di PPARγ, un coattivatore trascrizionale noto per stimolare l’espressione di TFAM, che, a sua volta, può aumentare l’espressione dei geni mitocondriali (ad esempio, MTCO1) e, in ultima analisi, la biogenesi mitocondriale.[75-76] Sebbene i precisi meccanismi cellulari e molecolari che mediano i robusti effetti del Telmisartan sul peso corporeo, sul dispendio energetico e sul metabolismo dei grassi rimangano da chiarire, gli studi sul reclutamento del coattivatore PPARγ e sull’espressione dei geni target, nonché sul numero, la struttura e la funzione dei mitocondri, potrebbero rappresentare aree di indagine potenzialmente fruttuose in futuro.
Ciò che si è anche notato con gli ARB, ma soprattutto con il Telmisartan, è che ha una azione sulla distribuzione del grasso più che sulla sua riduzione sistemica. Infatti, il Telmisartan ha mostrato di indurre la riduzione del grasso viscerale ma senza cambiamenti statistici sui deposito sottocutanei. Le più recenti review che hanno esaminato l’effetto del Telmisartan sulla condizione metabolica e composizione corporea dei pazienti trattati, hanno evidenziato che i risultati suggeriscono che questo sartano influisce sulla distribuzione del grasso, inducendo una riduzione del grasso viscerale, e quindi potrebbe essere utile nei pazienti ipertesi con obesità/sovrappeso, sindrome metabolica o intolleranza al glucosio.
Anche i dati aneddotici di un certo valore e design suggeriscono uno “spostamento” nell’equilibrio di mobilitazione delle riserve di grasso verso la perdita dei depositi viscerali invece di quelli sottocutanei. Ed è per tale motivo che diversi preparatori ne evitino l’uso sotto gara.
Questo “effetto shift” sul bilancio della mobilitazione delle riserve di grasso dal grasso sottocutaneo ad una prevalenza del viscerale si manifesta in modo significativo nel range di dosaggio di 80-160mg/die.
PPARγ
L’attività come agonista parziale del PPARγ è il motivo principale per il quale in Telmisartan agisce sul metabolismo lipidico adipocitario. Si è affermato che coloro i quali vogliono bypassare il problema dello shift della mobilitazione adiposa possono farlo assumendo l’Oleuropeina. Ora, non vi è nulla di certo e poco che superi la sottile linea tra ipotesi e dato realmente misurato, ma alcuni, soprattutto coloro i quali mal tollerano gli aumenti di bradichinina dati dagli ACE II inibitori, inseriscono questo supplemento erboristico nel tentativo di risolvere la sopra citata limitazione.
Peccato, però, che grazie a questa attività di agonista parziale del PPARγ, il Telmisartan può ridurre lo stoccaggio dei trigliceridi negli adipociti durante una dieta ipercalorica. In topi trattati per 28 giorni con ARB e ACE I, si è osservato un inferiore accumulo adiposo, minor peso corporeo, miglior controllo sull’assunzione di cibo rispetto ai topi non trattati con una dieta ad alto contenuto lipidico.
Nonostante, in teoria, l’effetto sul “grasso testardo” possa essere trattato anche attraverso il blocca del recettore dell’Angiotesina II, i dati a nostra disposizione ci mostrano una superiorità di azione e versatilità legata agli ACE II inibitori. L’uso di ARB, in particolar modo del Telmisartan, potrebbe avere un applicazione logica (se non si parla di soggetti obesi o in sovrappeso) nel gestione del grasso corporeo durante le fasi di ipercalorica, ad un dosaggio ipotetico di 40-80mg/die, al fine di ridurre l’accumulo adiposo e migliorare la qualità complessiva del peso raggiunto in Bulk.
Effetti collaterali degli ARB:
tachicardia e bradicardia (battito cardiaco accelerato o lento);
ipotensione (pressione sanguigna bassa);
edema (gonfiore di braccia, gambe, labbra, lingua o gola, quest’ultimo con conseguenti problemi di respirazione);
potenziale manifestazione di reazioni allergiche;
infezioni del tratto respiratorio superiore;
diarrea;
mal di schiena;
problemi renali;
iperkalemia.
Conclusioni:
Abbiamo visto come gli adrenocettori svolgono un ruolo importante nella biologia e nella fisiologia del tessuto adiposo, che comprende la regolazione della sintesi e dell’immagazzinamento dei trigliceridi (lipogenesi), la degradazione dei trigliceridi immagazzinati (lipolisi), la termogenesi (produzione di calore), il metabolismo del glucosio e la secrezione di ormoni derivati dagli adipociti che possono controllare l’omeostasi energetica dell’intero corpo. Questi processi sono regolati dal sistema nervoso simpatico attraverso l’azione di diversi sottotipi di adrenocettori espressi nei depositi di tessuto adiposo. In questa disamina, abbiamo evidenziato il ruolo dei sottotipi di adrenocettori negli adipociti bianchi, bruni e beige, e nel tessuto adiposo “testardo” ed abbiamo approfondito il ruolo potenziale degli ACE II inibitori nella modulazione sottoregolativa dell’attività degli α2-AR e l’impatto che questo può avere sul miglioramento della composizione corporea. Sono stati anche descritti gli effetti riscontrabili, nel medesimo contesto e fine, dei Sartani con le differenze tra l’applicabilità di questi confronto a quella degli ACE II inibitori.
Mentre il potenziale degli ACE II inibitori di migliorare la perdita di massa grassa in specie a carico dei depositi con una ratio sfavorevole tra α2:β2-AR, permettendo un importante sgravio sui dosaggi di α2-antagonisti, risulta un dato importante per la pianificazioni della preparazione alla gara, il potenziale effetto di riduzione del accumulo lipidico per attività di agonista parziale del PPARγ dato dal Telmisartan amplifica le applicazioni potenziali dei Sartani per il miglioramento della qualità del peso guadagnato in fase Bulk.
Ricordo, in fine, che tutto ciò che è stato detto è informazioni prettamente scientifica e non rappresenta in nessun modo un incitamento all’uso di farmaci fuori dalle linee di prescrizioni.
Kern PA, Ranganathan S, Li C, Wood L, Ranganathan G (May 2001). “Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance”. American Journal of Physiology. Endocrinology and Metabolism. 280 (5): E745–E751. doi:10.1152/ajpendo.2001.280.5.e745. PMID11287357. S2CID24306481
Marette A (December 2003). “Molecular mechanisms of inflammation in obesity-linked insulin resistance”. International Journal of Obesity and Related Metabolic Disorders. 27 (Suppl 3): S46–S48. doi:10.1038/sj.ijo.0802500. PMID14704744. S2CID30693649.
“Reduce Abdominal Fat”. Archived from the original on 2011-09-28. Retrieved 2009-04-10. Estrogen causes fat to be stored around the pelvic region, hips, butt and thighs (pelvic region)
Researchers think that the lack of estrogen at menopause plays a role in driving our fat northward. See: Andrews M (2006-12-01). “A Matter of Fat”. Yahoo Health. Women’s Health. Archived from the original on 2007-03-15.
Manolopoulos KN, Karpe F, Frayn KN (June 2010). “Gluteofemoral body fat as a determinant of metabolic health”. International Journal of Obesity. 34 (6): 949–959. doi:10.1038/ijo.2009.286. PMID20065965. S2CID21052919.
Brodie D, Moscrip V, Hutcheon R (March 1998). “Body composition measurement: a review of hydrodensitometry, anthropometry, and impedance methods”. Nutrition. 14 (3): 296–310. doi:10.1016/S0899-9007(97)00474-7. PMID9583375.
Dhaliwal SS, Welborn TA (May 2009). “Central obesity and multivariable cardiovascular risk as assessed by the Framingham prediction scores”. The American Journal of Cardiology. 103 (10): 1403–1407. doi:10.1016/j.amjcard.2008.12.048. PMID19427436.
Atzmon G, Yang XM, Muzumdar R, Ma XH, Gabriely I, Barzilai N (November 2002). “Differential gene expression between visceral and subcutaneous fat depots”. Hormone and Metabolic Research. 34 (11–12): 622–628. doi:10.1055/s-2002-38250. PMID12660871. S2CID33960130.
Lemke, KA (June 2004). “Perioperative use of selective alpha-2 agonists and antagonists in small animals”. The Canadian Veterinary Journal. 45 (6): 475–80. PMC 548630. PMID 15283516.
Hedner T, Edgar B, Edvinsson L, Hedner J, Persson B, Pettersson AYohimbine pharmacokinetics and interaction with the sympathetic nervous system in normal volunteersEur J Clin Pharmacol.(1992)
Grossman E, Rosenthal T, Peleg E, Holmes C, Goldstein DSOral yohimbine increases blood pressure and sympathetic nervous outflow in hypertensive patientsJ Cardiovasc Pharmacol.(1993 Jul)
Berlan M, Galitzky J, Riviere D, Foureau M, Tran MA, Flores R, Louvet JP, Houin G, Lafontan MPlasma catecholamine levels and lipid mobilization induced by yohimbine in obese and non-obese womenInt J Obes.(1991 May)
Kaplan’s Essentials of Cardiac Anesthesia. Elsevier. 2018. doi:10.1016/c2012-0-06151-0. ISBN978-0-323-49798-5. Mechanisms of Action:ACE inhibitors act by inhibiting one of several proteases responsible for cleaving the decapeptide Ang I to form the octapeptide Ang II. Because ACE is also the enzyme that degrades bradykinin, ACE inhibitors increase circulating and tissue levels of bradykinin (Fig. 8.4).
Jandeleit-Dahm K, Cooper ME (Sep 2006). “Hypertension and diabetes: role of the renin–angiotensin system”. Endocrinol Metab Clin North Am. 35 (3): 469–90, vii. doi:10.1016/j.ecl.2006.06.007. PMID16959581.
Human Physiology, Silverthorn (Pearson Benjamin Cummings 2004)[page needed]
Weir M (1999). “The renin-angiotensin-aldosterone system: a specific target for hypertension management”. American Journal of Hypertension. 12 (4). Oxford University Press (OUP): 205–213. doi:10.1016/s0895-7061(99)00103-x. ISSN0895-7061. PMID10619573.
“Congestive Heart Failure”. The Lecturio Medical Concept Library. 7 August 2020. Retrieved 27 August 2021.
Kester M, Karpa KD, Vrana KE (2012). “Cardiovascular System”. Elsevier’s Integrated Review Pharmacology. Elsevier. pp. 125–151. doi:10.1016/b978-0-323-07445-2.00008-2. ISBN978-0-323-07445-2. ACE inhibitors also slow progression of kidney disease in patients with diabetic nephropathies. Renal benefits are probably a result of improved renal hemodynamics from decreased glomerular arteriolar resistance.
Long AN, Dagogo-Jack S. Comorbidities of diabetes and hypertension: mechanisms and approach to target organ protection. J Clin Hypertens (Greenwich) 2011; 13:244-251. http://doi:10.1111/j.1751-7176.2011.00434.x.
50. Palominos MM, Dünner DH, Wabitsch M, Rojas CV. 2015. Angiotensin II directly impairs adipogenic differentiation of human preadipose cells. Mol Cell Biochem 2015; 408: 115-122. http://doi:10.1007/s11010-015-2487-y.
51. Schling MM, Dünner NH, Wabitsch M, Rojas CV. Angiotensin II directly impairs adipogenic differentiation of human preadipose cells. Mol Cell Biochem 2015; 408:115-122. http://doi:10.1007/s11010-015-2487-y.
52. Brücher R, Cifuentes M, Acuña MJ, Albala C, Rojas CV. Larger anti-adipogenic effect of angiotensin II on omental preadipose cells of obese humans. Obesity 2007; 15:1643-1646. http://doi:10.1038/oby.2007.196.
53. Fuentes P, Acuña MJ, Cifuentes M, Rojas CV. The anti-adipogenic effect of angiotensin II on human preadipose cells involves ERK1,2 activation and PPARG phosphorylation. J Endocrinol 2010; 206:75-83. http:// doi:10.1677/JOE-10-0049.
Jones BH, Standridge MK, Moustaid N. Angiotensin II increases lipogenesis in 3T3-L1 and human adipose cells. Endocrinology 1997; 138:1512-1519. http://doi:10.1210/endo.138.4.5038.
Townsend RR. The effects of angiotensin-II on lipolysis in humans. Metabolism 2001; 50:468-472. http://doi:10.1053/meta.2001.21021.
Weisinger RS, Begg DP, Jois M. Antagonists of the renin-angiotensin system and the prevention of obesity. Curr Opin Investig Drugs 2009; 10: 1069-1077.
Than A, Leow MK, Chen P. Control of adipogenesis by the autocrine interplays between angiotensin 1-7/Mas receptor and angiotensin II/AT1 receptor signaling pathways. J Biol Chem 2013; 288:15520-15531. http://doi:10.1074/jbc.M113.459792.
Sharma AM, Engeli S. The role of renin-angiotensin system blockade in the management of hypertension associated with the cardiometabolic syndrome. J Cardiometab Syndr 2006; 1:29-35. http://doi:10.1111/j.0197-3118.2006.05422.x.
Vertes V, Haynie R. Comparative pharmacokinetics of captopril, enalapril, and quinapril. Am J Cardiol. 1992;69:8C–16C. [PubMed] [Google Scholar]
Nakajima T, Yamada T, Setoguchi M. Prolonged inhibition of local angiotensin-converting enzyme after single or repeated treatment with quinapril in spontaneously hypertensive rats. J Cardiovasc Pharmacol. 1992;19:102–107. [PubMed] [Google Scholar]
Fukiyama F, Azuma J, Yoshida H, et al. A pharmacokinetic study of quinapril in normal Japanese men. J Clin Ther Med. 1993;9(Suppl 7):3–16. [Google Scholar]
Weishaar RE, Panek RL, Major TC, Simmerman J, Rapundalo ST, Taylor J, DG DG. Evidence for a functional tissue renin-angiotensin system in the rat mesentric vasculature and its involvement in regulation blood pressure. J Pharmacol Exp Ther. 1991;256:568–574. [PubMed] [Google Scholar]
Major TC, Overhiser RW, Taylor DG, Panek RL. Effect of quinapril, a new angiotensin-converting enzyme inhibitor, on vasoconstrictor activity in the isolated, perfused mesenteric vasculature of hypertensive rats. J Pharmacol Exp Ther. 1993;265:187–193. [PubMed] [Google Scholar]
Willenbrock R, Ozcelik C, Osterziel KJ, Dietz R. Angiotensin-converting enzyme inhibition, autonomic activity, and hemodynamics in patients with heart failure who perform isometric exercise. Am Heart J. 1996;131:999–1006. [PubMed] [Google Scholar]
Saitoh M, Miyakoda H, Kitamura H, Kinugawa T, Kotake H, Mashiba H. Effects of an angiotensin-converting enzyme inhibitor, alacepril, on cardiovascular and sympathetic nervous responses to mental stress in patients with essential hypertension. Intern Med. 1993;32:691–694. [PubMed] [Google Scholar]
Schling P, Mallow H, Trindl A, Loffler G. Evidence for a Local Renin Angiotensin System in Primary Cultured Human Preadipocytes. Int J Obes Relat Metab Disord (1999) 23(4):336–41. doi: 10.1038/sj.ijo.080082 PubMed Abstract | CrossRef Full Text | Google Scholar
Weisinger RS, Begg DP, Chen N, Jois M, Mathai ML, Sinclair AJ. The Problem of Obesity: Is There a Role for Antagonists of the Renin-Angiotensin System? Asia Pac J Clin Nutr (2007) 16 S1:359–67.Google Scholar
Takahashi N, Li F, Hua K, Deng J, Wang CH, Bowers RR, et al. Increased Energy Expenditure, Dietary Fat Wasting, and Resistance to Diet-Induced Obesity in Mice Lacking Renin. Cell Metab (2007) 6(6):506–12. doi: 10.1016/j.cmet.2007.10.01 PubMed Abstract | CrossRef Full Text | Google Scholar
Massiera F, Seydoux J, Geloen A, Quignard-Boulange A, Turban S, Saint-Marc P, et al. Angiotensinogen-Deficient Mice Exhibit Impairment of Diet-Induced Weight Gain With Alteration in Adipose Tissue Development and Increased Locomotor Activity. Endocrinology (2001) 142(12):5220–5. doi: 10.1210/endo.142.12.8556 PubMed Abstract | CrossRef Full Text | Google Scholar
Jayasooriya AP, Begg DP, Chen N, Mathai ML, Sinclair AJ, Wilkinson-Berka J, et al. Omega-3 Polyunsaturated Fatty Acid Supplementation Reduces Hypertension in TGR(mRen-2)27 Rats. Prostaglandins Leukot Essent Fatty Acids (2008) 78(1):67–72. doi: 10.1016/j.plefa.2007.11.001 PubMed Abstract | CrossRef Full Text | Google Scholar
De Blasi A, Cortellaro M, Costantini C. Enalapril in Essential Hypertension: A Comparative Study With Propranolol. Enalapril in Hypertension Study Group (Uk). Br J Clin Pharmacol (1984) 18(1):51–6. doi: 10.1111/j.1365-2125.1984.tb05021. PubMed Abstract | CrossRef Full Text | Google Scholar
Zorad S, Dou JT, Benicky J, Hutanu D, Tybitanclova K, Zhou J, et al. Long-Term Angiotensin II AT1 Receptor Inhibition Produces Adipose Tissue Hypotrophy Accompanied by Increased Expression of Adiponectin and Ppargamma. Eur J Pharmacol (2006) 552(1-3):112–22. doi: 10.1016/j.ejphar.2006.08.062 PubMed Abstract | CrossRef Full Text | Google Scholar
I capisaldi della gestione del peso sono gli interventi sullo stile di vita con il ruolo aggiuntivo di supporto dei farmaci anti-obesità e delle procedure bariatriche. Attualmente, i farmaci disponibili approvati per il trattamento dell’obesità agiscono sul bilancio energetico riducendo l’assunzione di cibo e il comportamento di ricompensa alimentare nel sistema nervoso centrale (ad esempio, sopprimendo l’appetito) o riducendo l’assorbimento dei grassi nell’intestino.[1,2,3] Finora non sono disponibili farmaci con un effetto diretto sull’aumento del dispendio energetico attraverso un’influenza sul tessuto adiposo [4,6]. Nell’uomo esistono due tipi di tessuto adiposo con funzioni fisiologiche distinte: il tessuto adiposo bianco (WAT), specializzato nell’immagazzinamento di trigliceridi in eccesso quando l’assunzione di energia supera il dispendio energetico, e il tessuto adiposo bruno (BAT) – con i relativi adipociti “beige”/”bruni” (derivati dal WAT) – che svolge un ruolo centrale nel metabolizzare il glucosio, gli acidi grassi e altre sostanze chimiche per produrre calore attraverso l’attivazione della proteina di disaccoppiamento 1 (UCP1) specifica del tessuto termogenico [4,7]. Alcuni dati suggeriscono che il BAT possa essere funzionale nell’uomo adulto [1]. I recettori β3-adrenergici (AR) sono espressi non solo nella vescica urinaria, ma anche sulla superficie degli adipociti bruni e bianchi [1]. I tessuti adiposi bruni e “beige”, contenenti cellule grasse termogeniche, possono essere attivati da agonisti dei recettori β3-adrenergici (β3-AR) [8]. È stato riportato che il Mirabegron, un agonista β3-AR umano selettivo, può stimolare il BAT e il processo di imbrunimento degli adipociti derivati dal WAT [9,10]. Il fatto che l’attivazione del BAT e degli adipociti “beige” possa aumentare il dispendio energetico rende i tessuti adiposi bruni e “beige” nuovi e promettenti bersagli per il trattamento dell’obesità [4,11].
Il ruolo del tessuto adiposo nella termogenesi e nei processi metabolici associati all’obesità:
Nell’uomo esistono due tipi principali di tessuto adiposo, che svolgono funzioni diverse: il tessuto adiposo bianco e il tessuto adiposo bruno. Oltre al WAT e al BAT, sono state distinte anche cellule adipose “brune”, definite cellule adipose “beige”. Esse derivano dal WAT, ma la loro funzione metabolica è simile a quella del BAT [4,7]. Il WAT è responsabile dell’immagazzinamento di energia sotto forma di trigliceridi, del rilascio di lipidi e della funzione di ghiandola endocrina, secernendo adipochine, come l’adiponectina e la leptina, per promuovere l’omeostasi metabolica [9,12]. Nell’obesità, gli adipociti bianchi si ipertrofizzano, seguiti da fibrosi, necrosi degli adipociti e infiltrazione di cellule immunitarie, che portano a infiammazione locale e sistemica, insulino-resistenza e disfunzione metabolica [9].
Cellule del BAT (colorate di marrone con anticorpi contro la proteina specifica del grasso bruno Ucp1) annidate tra le cellule del WAT grasso bianco.
Il BAT è stato descritto per la prima volta nel 1981 in finlandesi che lavoravano all’aperto e che erano stati esposti a basse temperature ambientali [6]. Il BAT metabolicamente attivo è stato identificato negli adulti mediante imaging PET/CT focalizzato principalmente sulla fossa sopraclavicolare, sull’area succlavia e sull’ascella, seguito dalle aree mediastiniche, paraspinali, perinefriche e sopradrenali [10,12]. Sebbene il BAT sia presente nell’uomo, la sua prevalenza diminuisce con l’età e nelle persone in sovrappeso o obese rispetto ai soggetti magri [6,9,13,14]. Gli anelli mancanti nel trattamento dell’obesità sono i farmaci che possono aumentare la quantità o l’attività del BAT. È stato riportato che il volume del BAT può essere aumentato dopo la chirurgia bariatrica [12]. Il BAT è il principale organo termogenico dei mammiferi, con lo scopo di aumentare il dispendio energetico in risposta al freddo o ad altre stimolazioni nervose simpatiche, rilasciando noradrenalina dai terminali nervosi per attivare i recettori β3-adrenergici attraverso il processo definito termogenesi senza brividi [2,10,11,12,13]. La capacità termogenica del BAT è stata stimata in circa 500 W/kg [6]. Gli adipociti del BAT sono arricchiti di mitocondri (i loro livelli sono più alti di quelli del WAT), nei quali la proteina di disaccoppiamento 1 (UCP1) è altamente espressa. La UCP1 dissipa l’energia in eccesso sotto forma di calore in un processo noto come termogenesi [2,15]. L’attivazione adrenergica della lipolisi stimola l’attività termogenica della UCP1 [2,10]. L’attivazione dell’UCP1 sulla membrana mitocondriale interna disaccoppia la respirazione mitocondriale, separando il trasporto di elettroni dalla produzione di ATP per ossidare il substrato e generare calore [4,8,16]. Gli acidi grassi a catena lunga, generati dai pool lipidici intracellulari, sono trasportati ai mitocondri attraverso la carnitina palmitoiltransferasi 1 (CPT1) e utilizzati come fonte di carburante dagli adipociti bruni per produrre calore. Inoltre, è stato proposto che gli acidi grassi liberi agiscano come attivatori allosterici di UCP1. Oltre agli acidi grassi, anche il glucosio circolante può essere utilizzato dal BAT attivo per alimentare la termogenesi [2,10,17]. In sintesi, il BAT consuma glucosio e lipidi per generare calore attraverso la respirazione disaccoppiata mediata da UCP1, con conseguente miglioramento dell’omeostasi glucidica e lipidica [9,13,18].
Struttura della proteina disaccoppiante umana UCP1
Gli adipociti termogenici umani possono originare da due lignaggi distinti, non solo da adipociti bruni costitutivi ma anche da cellule “beige” reclutabili, definite adipociti “bruni” o “bruno-simili” [5]. Gli adipociti “beige” sono localizzati prevalentemente nei depositi di WAT [16]. Le cellule adipose del WAT possono essere convertite in adipociti “beige” termogenici in un processo chiamato “browning” o “beiging” [12]. Da un lato, è stato dimostrato un sostanziale “beiging” del WAT sottocutaneo umano in alcuni disturbi, come la cachessia da cancro, le ustioni e le condizioni con alti livelli di catecolamine, ad esempio il feocromocitoma [8,13,14]. I pazienti con tumori che secernono catecolamine hanno anche più tessuto adiposo bruno rispetto alla maggior parte delle persone [19]. D’altra parte, gli adipociti “bruni” possono essere attivati con l’induzione dell’espressione di UCP1 da parte di stimoli ambientali, come l’esposizione al freddo e agli agonisti β-adrenergici, mediata dalla via di segnalazione p38-MAPK [9,12,14,16]. La risposta “beiging” dei soggetti obesi al freddo è simile a quella dei soggetti magri [14]. Sebbene queste cellule differiscano dagli adipociti bruni convenzionali – in quanto si sviluppano da una cellula precursore di adipociti bianchi e non da una cellula precursore di adipociti bruni, simile agli adipociti bruni classici nel BAT – gli adipociti “beige” possiedono goccioline lipidiche multiloculari, un gran numero di mitocondri e marcatori unici di espressione genica del grasso bruno, come UCP1, aumentando la capacità del tessuto di ossidazione del carburante e il dispendio energetico [4,16,18]. Inoltre, è stato dimostrato che il “beiging” è associato a una riduzione della fibrosi del tessuto adiposo e della disfunzione adiposa. Questi risultati suggeriscono che l’induzione del tessuto adiposo “beige” può migliorare l’omeostasi metabolica aumentando la capacità del WAT sottocutaneo di funzionare come serbatoio metabolico per il glucosio e i lipidi o riducendo la disfunzione del WAT che si verifica con l’obesità [13]. Così, oltre alla termogenesi e al dispendio energetico, i tessuti adiposi bruni e “beige” sono associati a un miglioramento dell’omeostasi del glucosio e dei lipidi, nonché a una maggiore sensibilità all’insulina nell’uomo e nel topo [14].
Distribuzione del BAT nei neonati, nelle donne e negli uomini. Il BAT è immagazzinato in un deposito interscapolare separato nei neonati che perdono il loro tessuto adiposo bruno con l’avanzare dell’età. Negli esseri umani adulti, la maggior parte degli adipociti bruni si trova nei depositi di BAT sopraclavicolari nella regione del collo. Quantità minori di BAT si trovano nell’aorta, nelle vertebre, nelle aree ascellari e renali. C’è una distribuzione simile del tessuto adiposo bruno sia nelle donne che negli uomini. Tuttavia, le donne hanno una maggiore quantità di massa e attività di BAT.
Considerando il fatto che nelle persone obese adulte c’è meno BAT rispetto ai soggetti magri, il WAT in eccesso, che può essere stimolante e in fase di “beigezzazione/imbrunimento”, può svolgere un ruolo aggiuntivo rispetto al BAT nei processi metabolici [13]. Pertanto, il tessuto adiposo bruno e il tessuto adiposo “beige” sono stati riconosciuti come regolatori critici del metabolismo e del dispendio energetico dell’intero corpo e sono considerati bersagli promettenti per la terapia anti-obesità [2,12,15].
L’Irisina è un ormone sintetizzato in grande quantità dal tessuto muscolare umano durante le attività sportive. La molecola è in grado di operare il meccanismo molecolare detto “browning” [“imbrunimento”], ovvero di conversione del WAT in BAT.
Il grasso bruno, il grasso “beige” e i β3-adrenocettori nel contesto dell’obesità:
Struttura del recettore β3-adrenergico
La famiglia dei recettori β-adrenergici (AR) umani è composta dai recettori β1, β2 e β3, in cui il β1-AR è altamente espresso in tutto il sistema cardiovascolare, il β2-AR si trova nelle vie aeree polmonari, in tutta la vascolarizzazione e nel muscolo scheletrico e l’espressione del β3-AR è limitata soprattutto alla vescica urinaria e alla cistifellea, oltre che al BAT e al WAT [4,10]. Il β3-AR umano, identificato nel 1989, è un recettore a 7 membrane, con una coda N-terminale extracellulare e una coda C-terminale intracellulare, composta da 408 aminoacidi. Si accoppia principalmente a Gs per attivare l’adenilato ciclasi, con conseguente aumento dei livelli intracellulari di cAMP, sebbene sia stato riportato un accoppiamento promiscuo con altri effettori, come Gi [4,16].
La β3-AR svolge un ruolo critico nel tessuto adiposo, nella regolazione della termogenesi, della glicolisi e della lipolisi [16]. Studi sugli animali hanno dimostrato che la stimolazione cronica del BAT porta a un miglioramento della tolleranza al glucosio e della sensibilità all’insulina e a una riduzione dell’obesità, oltre che al rilascio di adipochine che regolano beneficamente il metabolismo [1,8,10,12,20]. Inoltre, l’attivazione β3-AR-mediata del WAT può aumentare la secrezione insulinica delle cellule β pancreatiche [5]. È stato anche riportato che una parte significativa della termogenesi non da brivido ha luogo nel tessuto adiposo bruno ed è mediata principalmente dal β3-adrenocettore [19]. Nei topi alimentati con dieta a base di chow e ad alto contenuto di grassi, il trapianto di BAT ha ridotto il peso corporeo, aumentato il metabolismo del glucosio e la sensibilità all’insulina e incrementato l’assorbimento di glucosio nel BAT e nel WAT [15].
Oltre alla funzione metabolica, la β3-AR svolge un ruolo nel cervello, essendo coinvolta nei processi di memoria, apprendimento e regolazione dell’appetito, nel tratto gastrointestinale, dove partecipa alla regolazione della motilità, e nel sistema genitourinario, dove svolge un ruolo nella regolazione della funzione vescicale [16].
Durante la termogenesi, i β3-adrenocettori aumentano il dispendio energetico, che può portare alla perdita di grasso, in risposta alla stimolazione simpatica [19]. È dimostrato che la stimolazione cronica dell’attività nervosa simpatica e dei β3-AR può attivare il BAT [6]. È stato dimostrato che l’esposizione al freddo stimola il sistema nervoso simpatico a rilasciare noradrenalina dalle terminazioni nervose simpatiche per attivare i β-AR sulle membrane delle cellule del BAT, promuovendo la termogenesi. In questo modo, il BAT umano è in grado di avviare la termogenesi attraverso il consumo di acidi grassi e glucosio e, successivamente, di generare calore [1,3,8]. Inoltre, l’attivazione dei β3-ARs da parte dell’esposizione al freddo o di agenti farmacologici induce un programma di “beiging” nel WAT [18]. Un modo per aumentare la quantità effettiva di tessuto adiposo bruno può essere quello di somministrare l’agonista β3-adrenoccettore in modo cronico [19]. Una singola dose di agonista dei β3-adrenocettori può almeno raddoppiare il dispendio energetico in un modello murino a circa 21 °C [19].
Struttura del gene ADRB3
Il ruolo dei β3-AR nel metabolismo energetico umano è supportato da studi clinici che riportano associazioni tra polimorfismi specifici nel gene umano ADRB3 (il gene che codifica i β3-AR) e tassi più elevati di obesità, insulino-resistenza e diabete [10]. Inoltre, le mutazioni nel gene ADRB3 sono state correlate all’insulino-resistenza, all’aumento del rischio di obesità e diabete e alla malattia del fegato grasso non alcolico negli individui obesi [10]. I dati indicano che il silenziamento di ADRB3 negli adipociti umani “marroni”/”beige” altera il macchinario termogenico cellulare e causa una riduzione dei livelli di espressione dei geni associati al metabolismo degli acidi grassi, alla massa mitocondriale e alla termogenesi, senza compromettere il fenotipo “marrone”/”beige” [10].
Attività agonista dei β3-AR del Mirabegron:
Struttura molecolare del Mirabegron
Mirabegron è una nuova generazione di agonisti dei β3-adrenocettori con una buona biodisponibilità [21]. Gli effetti dell’agonista selettivo dei β3-AR mirabegron sul rilassamento della vescica sono stati scoperti nel 2007. Per la prima volta, la selettività β3 del mirabegron (YM-178) nel contesto della funzione vescicale è stata descritta da Takasu et al. [22]. YM-178 ha aumentato l’accumulo di AMP ciclico in cellule ovariche di criceto cinese che esprimono il β3-adrenocettore umano. Mirabegron ha dimostrato valori di EC50 nanomolari contro il β3-AR umano in saggi biochimici, con una potente selettività rispetto ai β1- e β2-AR [22]. Studi in vivo hanno dimostrato che la somministrazione di mirabegron ha ridotto la pressione intravescicale e le contrazioni spontanee della vescica in modo dose-dipendente [23]. Mirabegron è stato approvato dalla Food and Drug Administration (FDA) statunitense nel 2012 come nuovo tipo di trattamento farmacologico per la vescica iperattiva (OAB) [6,21,24]. Cinquanta milligrammi di mirabegron è la dose raccomandata a tutti i pazienti con OAB [24]. Il farmaco è generalmente ben tollerato e gli effetti collaterali più comuni includono ipertensione, rinofaringite e infezione del tratto urinario [6].
La selettività β3 di Mirabegron è stata confermata in molti studi con l’uso di linee cellulari che esprimono il β3-adrenocettore sia animale che umano [22,23,25]. Mirabegron ha mostrato una selettività per il β3-AR umano superiore di oltre 400 volte rispetto al β1-AR o al β2-AR umano [26]. Ad esempio, Brucker et al. [27] hanno utilizzato cellule di ovaio di criceto cinese (CHO)-K1, cellule di rene embrionale umano 293 esprimenti stabilmente recettori β1-, β2- o β3-adrenergici umani e recettori α1D- e α2B-adrenergici umani per valutare la selettività di mirabegron. A una concentrazione di 10 μM, l’attività β3-adrenergica rispetto all’isoproterenolo (agonista β-adrenergico completo) era dell’88% per mirabegron. A sua volta, l’attività β1- e β2-adrenergica di mirabegron era rispettivamente del 3% e del 15% [27]. In questo studio mirabegron non ha soddisfatto il criterio di significatività per l’inibizione dei recettori α1D- o α2B-adrenergici [27]. Tuttavia, alcuni studi hanno indicato che mirabegron potrebbe svolgere un ruolo come antagonista degli α1-adrenergici [28,29]. Alexandre et al. [28] hanno ipotizzato che mirabegron rilassasse la muscolatura liscia uretrale nei topi attraverso un duplice meccanismo che coinvolge l’attivazione dei β3-adrenocettori e il blocco degli α1-adrenocettori. In un altro studio, mirabegron ha indotto una vasorilassazione endotelio-indipendente nelle arterie del tessuto adiposo viscerale attraverso l’antagonismo degli α1-adrenocettori. Questa azione ha suggerito che mirabegron potrebbe migliorare efficacemente la perfusione del tessuto adiposo viscerale, favorendo così un sano rimodellamento del tessuto adiposo e prevenendo alcune delle conseguenze cardiometaboliche indesiderate dell’obesità e dell’invecchiamento [29]. Resta ancora difficile stabilire in che misura l’antagonismo degli α1-adrenocettori possa contribuire agli effetti clinici di mirabegron [28,29].
I cambiamenti metabolici benefici causati dal trattamento cronico con mirabegron potrebbero derivare dalla stimolazione della β3-AR nel BAT e nel WAT umani [5,12,18]. È stato suggerito che mirabegron potrebbe migliorare le malattie metaboliche legate all’obesità aumentando la termogenesi del BAT, la lipolisi del WAT e la stimolazione del processo di “brunimento” degli adipociti derivati dal WAT [4,5,9,10]. Il trattamento acuto con mirabegron ha aumentato il dispendio energetico [10,15]. Dopo il silenziamento dell’espressione dei β3-AR, il mirabegron non è stato in grado di stimolare la lipolisi e la termogenesi del BAT [10]. Molti studi hanno dimostrato che il trattamento con mirabegron ha aumentato l’assorbimento del glucosio negli adipociti bruni e “beige”, ha migliorato l’omeostasi del glucosio e ha aumentato la sensibilità all’insulina e la funzione delle cellule β [1,9]. Inoltre, è stato dimostrato che il trattamento cronico con agonisti β3-AR nell’uomo può rilasciare adipochine benefiche [1]. Il modo in cui mirabegron migliora il metabolismo del glucosio non è stato finora chiarito [5]. Tuttavia, sono stati ipotizzati alcuni meccanismi. In primo luogo, mirabegron stimola la secrezione di adiponectina, nota adipochina derivata dal WAT e associata a una maggiore sensibilità all’insulina nel muscolo scheletrico e nel fegato. In secondo luogo, mirabegron aumenta la concentrazione di polipeptide inibitore gastrico (GIP), l’incretina collegata alla secrezione di insulina. Infine, il meccanismo di mirabegron potrebbe coinvolgere le stesse cellule β [5].
L’agonista dei recettori β3-adrenergici è un ottimo candidato per il trattamento dell’obesità, poiché l’isoforma β3 è espressa esclusivamente negli adipociti e l’azione su altri tipi di cellule, come i cardiomiociti e le cellule muscolari lisce, attraverso le altre isoforme β – β1 e β2 – è minima e dose-dipendente [11]. Pertanto, come agonista β3-AR, Mirabegron attiverebbe la termogenesi nel tessuto adiposo, stimolando l’ossidazione dei lipidi e il consumo di glucosio per produrre calore, senza causare gravi effetti collaterali cardiovascolari [13].
Mirabegron come agente antiobesità negli studi sperimentali:
Adipocita del BAT
Il trattamento dei roditori con agonisti β3-AR ha attivato il BAT, con conseguente aumento del dispendio energetico, perdita di peso e miglioramento del metabolismo del glucosio e dei lipidi. Inoltre, ha ripristinato l’equilibrio NO/redox, migliorato la funzione endoteliale e, quindi, esercitato effetti protettivi vascolari [4,6,13,17]. L’aumento dell’attività del BAT ha impedito lo sviluppo e la gravità dell’obesità e del diabete di tipo 2, mentre i topi privi di BAT erano inclini all’obesità [16]. È stato riportato che una riduzione della massa del BAT nei topi indotta da un transgene produce obesità e che questi topi presentano un’ulteriore maggiore suscettibilità all’obesità a causa di diete obesitogene [8,30,31].
Adipocita “Beige”
Come si è detto, il Mirabegron può essere efficace come attivatore del BAT, stimolatore delle cellule “beige” e controllore dell’omeostasi metabolica. L’influenza benefica di mirabegron sul metabolismo è stata confermata da studi in vitro e in vivo [2,4,15,18]. Nello studio condotto da Dehvari et al. [15], sono stati riportati gli effetti di mirabegron negli adipociti bruni, bianchi e “beige” in vitro e i suoi effetti sull’utilizzo del glucosio e sulla termogenesi in vivo. È stato dimostrato che mirabegron aumenta l’assorbimento di glucosio e la glicolisi negli adipociti bruni di topo in vitro e promuove l’assorbimento di glucosio nel BAT in vivo. Il mirabegron ha aumentato i livelli di cAMP e l’mRNA di UCP1, con conseguente aumento del consumo di ossigeno mediato da UCP1, nonché l’assorbimento di glucosio e la glicolisi cellulare negli adipociti bruni e “beige” (tale azione è mancata nelle colture cellulari primarie di adipociti bruni provenienti da topi knockout per il β3-adrenocettore), mentre questi effetti erano assenti o ridotti negli adipociti bianchi. In vivo, mirabegron ha aumentato il consumo di ossigeno nell’intero corpo e l’assorbimento di glucosio nel tessuto adiposo bruno e bianco inguinale e ha migliorato la tolleranza al glucosio. Nei topi knockout per il β3-adrenorecettore, mirabegron non è riuscito a indurre l’assorbimento di glucosio nel tessuto adiposo, né ad aumentare il consumo di ossigeno corporeo, il che dimostra che la segnalazione del β3-adrenorecettore è una via principale delle azioni metaboliche di mirabegron [15]. Analogamente a Dehvari et al. [15], Hao et al. [4] hanno studiato gli effetti anti-obesità di mirabegron utilizzando modelli in vitro e in vivo. In entrambe le linee cellulari – preadipociti bruni di topo e preadipociti bianchi 3T3-L1 – mirabegron ha stimolato l’espressione di UCP1. I topi trattati con mirabegron, alimentati con una dieta ad alto contenuto di grassi, presentavano una riduzione del peso corporeo e dell’adiposità, nonché un miglioramento della tolleranza al glucosio e della sensibilità all’insulina. Le goccioline lipidiche nel BAT dei topi trattati con mirabegron erano meno numerose e di dimensioni inferiori rispetto ai controlli. La colorazione H&E e l’immunoistochimica hanno indicato che mirabegron ha aumentato l’abbondanza di cellule “beige” nel WAT [4]. Si è concluso che mirabegron ha aumentato l’espressione di UCP1 e ha promosso la “brunitura” del WAT, che è stata accompagnata da un miglioramento della tolleranza al glucosio, della sensibilità all’insulina e della prevenzione dell’obesità indotta da una dieta ad alto contenuto di grassi [4]. In un altro studio su animali, Valgas da Silva et al. [18] hanno riferito che un trattamento di 2 settimane con mirabegron ha ridotto l’infiammazione, migliorato il metabolismo, impedito l’accumulo di grasso ectopico nel BAT e nel fegato e diminuito l’insulino-resistenza nei topi obesi (riduzione dell’indice HOMA e dei livelli di insulina). Mirabegron ha aumentato l’espressione di UCP1 nel BAT e il dispendio energetico, oltre a ridurre l’adiposità nei topi obesi. Inoltre, mirabegron ha ridotto i livelli circolanti di acidi grassi liberi, glicerolo e TNF-α. È noto che l’aumento dei livelli di FFA circolanti causa insulino-resistenza negli organi bersaglio dell’insulina ed è emerso come uno dei principali collegamenti tra l’obesità e lo sviluppo della sindrome metabolica. È noto anche che il TNF-α ha un effetto lipolitico, che determina un aumento dei livelli di FFA e glicerolo in circolo, contribuendo all’insulino-resistenza. Tuttavia, a differenza dello studio condotto da Dehvari et al. non sono stati riscontrati cambiamenti nel WAT inguinale: il mirabegron non ha indotto il “beiging” del WAT inguinale dei topi obesi. Inoltre, l’obesità indotta dalla dieta ha aumentato significativamente i depositi lipidici nel fegato e nel BAT, ma mirabegron ha parzialmente invertito questi cambiamenti, il che potrebbe indicare un ruolo protettivo di mirabegron nello sviluppo della steatosi epatica e dell’insulino-resistenza [18]. La conferma che mirabegron può essere utile come agente anti-obesità è stata trovata anche nello studio di Hao et al. [4]. È stato dimostrato che mirabegron provoca un aumento di 14 volte dell’espressione genica di UCP1 e può determinare una perdita di peso del 12% e una riduzione dell’adiposità nei topi obesi rispetto all’attività fisica.
Struttura molecolare della Metformina
La terapia combinata, composta da Mirabegron e Metformina, è stata verificata nel modello murino di prevenzione e nel modello murino di trattamento dell’obesità [2]. La metformina, un derivato della biguanide, è uno dei farmaci più comunemente utilizzati per il trattamento del diabete di tipo 2. Inibisce il complesso mitocondriale I, vitale per il trattamento dell’obesità. Inibisce il complesso mitocondriale I, vitale per il trasporto di elettroni, che porta all’attivazione dell’AMPK (proteina chinasi attivata dall’adenosina 5′-monofosfato). Di conseguenza, la produzione di ATP (adenosina trifosfato) diminuisce e la concentrazione intracellulare di ADP (adenosina difosfato) aumenta. Di conseguenza, i livelli cellulari di AMP (adenosina monofosfato) aumentano, attivando infine l’AMPK. L’AMPK è un regolatore chiave di numerose vie metaboliche, tra cui il metabolismo del glucosio e dei lipidi e l’omeostasi energetica. La metformina svolge anche un ruolo importante inibendo la segnalazione dei recettori dell’insulina e dell’IGF, con conseguenti cambiamenti nell’omeostasi metabolica [32]. Zhao et al. [2] hanno indicato che questa terapia complessa potrebbe essere un approccio promettente per la prevenzione e il trattamento dell’obesità, agendo contemporaneamente sull’assunzione e sul dispendio energetico, senza effetti collaterali sulla funzione cardiovascolare. Nel modello di prevenzione, metformina e mirabegron hanno provocato un’ulteriore riduzione del 12% e del 14% dell’aumento di peso corporeo indotto da una dieta ad alto contenuto di grassi, rispetto a metformina o mirabegron da soli, rispettivamente. Nel modello di trattamento, metformina e mirabegron hanno promosso in modo additivo una perdita di peso corporeo del 17% nei topi obesi indotti dalla dieta, superiore del 13% e del 6% rispetto a metformina e mirabegron da soli, rispettivamente. La terapia combinata ha avuto un effetto additivo sulla perdita di peso nei topi, associato a una significativa perdita di grasso, soprattutto nel WAT sottocutaneo [2]. I ricercatori hanno suggerito che l’effetto additivo di metformina e mirabegron sull’aumento del dispendio energetico abbia contribuito in modo determinante alla riduzione del peso corporeo e della massa grassa nei topi [2]. La terapia con metformina e mirabegron ha avuto un effetto additivo sulla termogenesi del BAT e sulla doratura del WAT sottocutaneo. La terapia combinata ha aumentato significativamente l’espressione di UCP1 nel BAT e nel WAT sottocutaneo [2]. Inoltre, metformina e mirabegron hanno migliorato la tolleranza al glucosio e la sensibilità all’insulina, e l’effetto era indipendente dall’assunzione di cibo. Tuttavia, la co-somministrazione di metformina e mirabegron non ha migliorato l’omeostasi del glucosio nei topi in misura maggiore rispetto alla metformina o al mirabegron da soli [2].
Un diagramma che riassume gli effetti combinati di Metformina (Met)/Mirabegron (Mir) sull’obesità nei modelli di prevenzione e trattamento. (A) Nel modello di prevenzione in cui una dieta ricca di grassi (HFD) e farmaci venivano somministrati simultaneamente, il trattamento con Met/Mir ha ridotto l’aumento di peso in modo additivo. Ciò è dovuto principalmente a un miglioramento della spesa energetica (EE) che era accompagnato da un’espressione sovraregolata di marcatori critici nella lipolisi, nell’ossidazione degli acidi grassi e nella termogenesi nel tessuto adiposo bruno (BAT). (B) Nel modello di trattamento, è stato prima stabilito un fenotipo di obesità indotta dalla dieta (DIO), seguito da 5 settimane di trattamenti terapeutici con Met e/o Mir. Il trattamento con Met/Mir ha causato una marcata perdita di peso, derivante dall’aumento di EE
E’ interessante notare che la p-Sinefrina, una agonista selettivo dei β3-Adrenocettoiri di origine naturale, ha mostrato in studi su animali effetti positivi sull’imbrunimento del WAT, sopprimendo così l’obesità e la steatosi epatica.
Mirabegron come farmaco antiobesità: i dati degli studi sull’uomo:
Oltre agli studi sperimentali, esistono numerosi studi clinici in cui è stata dimostrata l’influenza di mirabegron sull’attività del BAT e sulla massa corporea. Gli autori hanno riferito che mirabegron ha portato a un aumento dell’attività del BAT e del dispendio energetico a riposo [1,3,5,10,17,21]. Prove preliminari suggeriscono che gli effetti del mirabegron sul metabolismo del glucosio, sul colesterolo HDL e sugli acidi biliari assomigliano a quelli ottenuti con un lieve esercizio fisico [1,5]. Nel primo gruppo di studi sono state testate soprattutto dosi elevate di mirabegron (100 mg, 150 mg o 200 mg) [1,3,5,17,21].
Cypess et al. [1] hanno usato, per la prima volta, Mirabegron per studiare il BAT umano e hanno confrontato la sua azione in un grado che corrispondeva alle risposte all’esposizione al freddo. La somministrazione di 200mg al giorno di Mirabegron orale per 12 settimane a 12 uomini sani è stata associata a una maggiore attività del BAT (misurata tramite tomografia a emissione di positroni 18F-fluorodesossiglucosio combinata con tomografia computerizzata) e all’aumento del tasso metabolico a riposo di 203 ± 40 kcal/die, rispetto agli individui che hanno ricevuto il placebo. È stato ipotizzato che la perdita di peso calcolata, associata al dispendio energetico, dovrebbe raggiungere i 5 kg nel primo anno e i 10 kg entro la fine dei 3 anni [1]. In questo studio, il dosaggio di 200 mg di mirabegron, una dose molto più alta di quelle attualmente approvate per ridurre i sintomi della vescica iperattiva, è stato generalmente ben tollerato, anche dopo 12 settimane di somministrazione orale giornaliera [1]. L’effetto collaterale più comune era la tachicardia [1].
L’alta dose di Mirabegron [100mg al giorno] è stata testata da O’Mara et al. durante un programma di terapia di 4 settimane su 14 donne sane di varie etnie [5]. Nell’endpoint primario, i ricercatori hanno riferito che la terapia cronica con Mirabegron ha aumentato il volume del BAT e l’attività metabolica, misurati tramite PET/CT con 18F-fluorodesossiglucosio [5]. Inoltre, le donne che avevano avuto principalmente meno BAT hanno finalmente raggiunto un aumento maggiore del volume e dell’attività del BAT dopo il trattamento [5]. Gli endpoint secondari hanno rivelato che la spesa energetica a riposo dell’intero corpo era più alta dopo il trattamento con mirabegron; tuttavia, non sono state riscontrate modifiche nel peso corporeo o nella composizione. Questi risultati dovrebbero essere associati a un intervallo di BMI ristretto e alla partecipazione di donne non obese. Inoltre, è stato riscontrato che la terapia con Mirabegron aumenta i biomarcatori delle lipoproteine come HDL e apolipoproteina A1, apolipoproteina E e peptide inibitorio gastrico (GIP), nonché i livelli di adiponectina, adipochina antidiabetica e antinfiammatoria. Dopo il trattamento con mirabegron, è stata osservata una riduzione del rapporto ApoB100/ApoA1, un biomarcatore del rischio cardiovascolare. Infine, dopo il trattamento cronico con mirabegron, un test di tolleranza al glucosio per via endovenosa ha rivelato una maggiore sensibilità all’insulina, efficacia del glucosio e secrezione di insulina [5]. Tuttavia, il cambiamento nella valutazione del modello omeostatico della resistenza all’insulina (HOMA-IR), una misura della resistenza all’insulina, non è stato significativo dopo il trattamento cronico con mirabegron. Gli autori hanno suggerito che la ragione principale dovrebbe essere il livello HOMA-IR quasi normale all’inizio dello studio [5]. Come è una preoccupazione comune nel trattamento cronico con agonisti adrenergici, 100mg di Mirabegron hanno portato a una variazione diurna della frequenza cardiaca tale che Mirabegron l’ha aumentata di più durante la notte rispetto a quando i soggetti erano svegli e in movimento. D’altra parte, il trattamento con Mirabegron non ha avuto alcun effetto sulla tolleranza all’esercizio [5].
Loh et al. [21] hanno riportato l’efficacia di varie dosi singole di mirabegron (50, 100, 150 e 200 mg) in un gruppo di 17 individui sani (11 uomini, 6 donne) che hanno assunto il farmaco in quattro giorni separati, con 3-14 giorni di wash-out tra ogni dose. Hanno riferito che la spesa energetica (misurata tramite calorimetria indiretta) è aumentata significativamente dopo le dosi da 100 mg e 200 mg e ha mostrato una tendenza all’aumento dopo le dosi da 150 mg, ma non era significativamente diversa dal basale in risposta a 50 mg di mirabegron. La temperatura cutanea sopraclaveare (come indicatore surrogato dell’attività BAT), è aumentata dopo le dosi di mirabegron da 50 mg, 100 mg e 150 mg, ma non era significativamente diversa dal basale in risposta a 200 mg. Considerando gli effetti collaterali, il cambiamento nella pressione sanguigna sistolica è stato significativo dopo le dosi da 150 mg e 200 mg rispetto alla dose da 50 mg e alla dose da 100 mg. Tuttavia, non c’era alcuna differenza nella pressione sanguigna diastolica tra le dosi da 50 mg, 100 mg, 150 mg e 200 mg. Il cambiamento nella frequenza cardiaca è stato maggiore dopo 200 mg rispetto alle dosi rimanenti. Hanno concluso che una dose da 100 mg di mirabegron può essere efficace per aumentare il dispendio energetico e la temperatura cutanea sopraclaveare in modo specifico per il recettore β3-adrenergico, senza gli aumenti significativi della pressione sanguigna o della frequenza cardiaca osservati a dosi più elevate [21].
Baskin et al. [17] hanno studiato le implicazioni cliniche del mirabegron in 12 uomini sani e magri a cui è stata somministrata la dose approvata di 50 mg e una dose elevata di 200 mg. Si è verificato un aumento più che proporzionale alla dose nell’attività metabolica del BAT (misurata tramite PET/CT). Rispetto al placebo, 50 mg di mirabegron hanno aumentato l’attività del BAT nella maggior parte dei soggetti. Tuttavia, l’attivazione del BAT con 50 mg è stata significativamente inferiore rispetto a quella con 200 mg. Solo la dose da 200 mg ha aumentato la spesa energetica a riposo (5,8%). La stimolazione cardiovascolare è stata coerente con studi precedenti, poiché 200 mg di mirabegron hanno aumentato sia la frequenza cardiaca che la pressione sanguigna.
Uno studio randomizzato, in doppio cieco, cross-over costituito da tre interventi (esposizione al freddo a breve termine (~2 h), mirabegron (dose singola da 200 mg) e placebo) in un gruppo di 10 uomini magri olandesi sud asiatici e 10 uomini magri europei, condotto da Nahon et al. [3], ha rivelato che l’esposizione al freddo e il mirabegron hanno indotto effetti metabolici benefici, tra cui un aumento della spesa energetica a riposo (misurata mediante calorimetria indiretta), livelli di acidi grassi liberi nel siero e ossidazione dei lipidi. Il mirabegron ha aumentato la frequenza cardiaca sia nei sud asiatici (+10 battiti/min) che nei caucasici bianchi (+7 battiti/min), mentre la pressione sanguigna sistolica e diastolica non sono cambiate in modo significativo [3]. È stato osservato che una singola dose di mirabegron ha aumentato i livelli di insulina nel siero senza influenzare i livelli di glucosio. Il mirabegron può stimolare il rilascio di insulina direttamente agendo sul β3-AR del pancreas o indirettamente attraverso un aumento degli FFA che possono stimolare il pancreas a rilasciare insulina [3].
L’azione dose-dipendente del mirabegron sul tessuto adiposo, inclusa l’influenza sull’attività BAT e sul dispendio energetico, può essere analoga all’effetto del mirabegron sulla vescica urinaria. L’attivazione dei recettori β3-adrenergici con mirabegron ha determinato risposte dei recettori β3-adrenergici dipendenti dalla concentrazione [27]. Per quanto riguarda la funzione della vescica, negli studi in vivo, la somministrazione di mirabegron ha ridotto la pressione intravescicolare e le contrazioni vescicali spontanee in modo dose-dipendente [23].
È stato riportato che dosi elevate di mirabegron (in particolare 200 mg al giorno), molto più elevate di quelle approvate dalla FDA per l’iperattività della vescica (50 mg al giorno), possono essere associate a effetti collaterali cardiovascolari come mal di testa, tachicardia e pressione sanguigna elevata (per lo più solo pressione sanguigna sistolica) [1,3,5,17,21]. L’aumento della pressione sanguigna sistolica può raggiungere ~10 mm Hg alla dose di 200 mg al giorno [21]. Questo è il risultato della perdita di selettività per il β3-adrenocettore a questa dose, tale che mirabegron attiva indirettamente i β1-adrenocettori che sono ampiamente espressi in vari organi, in particolare il sistema cardiovascolare. Questo meccanismo coinvolge l’assorbimento del trasportatore di noradrenalina del mirabegron nei terminali nervosi simpatici cardiaci, causando successivamente un rilascio di noradrenalina, che attiva i β1-adrenocettori [21]. Tuttavia, il trattamento con mirabegron non ha avuto effetti sulla tolleranza all’esercizio [5]. L’attivazione dei β1-adrenocettori può essere attenuata dalla co-somministrazione di propranololo o bisoprololo [16]. D’altro canto, gli studi clinici hanno rivelato che dosi di mirabegron fino a 100 mg al giorno per almeno 12 mesi hanno mostrato un buon profilo di sicurezza e non hanno determinato un aumento dell’incidenza di tachicardia, pressione sanguigna, alterazioni dell’ECG o eventi cardiovascolari [21]. Dosi terapeutiche inferiori (50 mg) nei pazienti con OAB hanno determinato piccole variazioni della frequenza cardiaca (1 battito al minuto) e della pressione sanguigna (1 mm Hg o meno). Considerando gli effetti collaterali cardiovascolari, il mirabegron non è raccomandato nei pazienti con grave ipertensione incontrollata (pressione sanguigna sistolica ≥ 180 mm Hg e/o pressione sanguigna diastolica ≥ 110 mm Hg) [16].
Nel secondo gruppo di studi, condotto da Finlin et al. [9,13,14], è stata testata una bassa dose di mirabegron, una che è stata approvata per il trattamento dell’OAB. In un gruppo di 13 pazienti obesi di mezza età, 50 mg di mirabegron al giorno durante una terapia di 12 settimane hanno indotto il “beiging” del tessuto adiposo bianco sottocutaneo, nonché un miglioramento della funzione delle cellule β. Mirabegron ha aumentato l’espressione proteica dei marcatori adiposi “beige” UCP1 (2,4 volte), della proteina transmembrana 26 (TMEM26) (4,2 volte) e dell’effettore A simile al DFFA che induce la morte cellulare (CIDEA) (2,4 volte) [13]. Il “beiging” del tessuto adiposo bianco sottocutaneo da parte di mirabegron può ridurre la disfunzione del tessuto adiposo, il che può migliorare la capacità ossidativa muscolare e può migliorare la funzione delle cellule β [13]. Prendendo in considerazione l’omeostasi del glucosio, il trattamento con mirabegron ha migliorato la tolleranza orale al glucosio, portando a convertire il prediabete in una normale concentrazione di glucosio, ha ridotto i livelli di emoglobina A1c e ha migliorato la sensibilità all’insulina e la funzione delle cellule β, senza influenzare la glicemia a digiuno o i livelli di insulina a digiuno e HOMA-IR. Tuttavia, i risultati delle pinze euglicemiche, che sono il gold standard per misurare la sensibilità all’insulina, hanno rivelato che il trattamento con mirabegron ha aumentato in modo coerente e significativo la velocità di infusione del glucosio di circa il 12% [13]. I livelli di lipidi plasmatici sono cambiati in modo significativo, ma, dopo il trattamento con mirabegron, è stata riscontrata una tendenza verso una riduzione del colesterolo totale [13]. Sfortunatamente, una terapia di 12 settimane non ha determinato un aumento significativo della quantità di BAT e del dispendio energetico a riposo, della perdita di peso o dei cambiamenti nella composizione corporea in tali pazienti [13].
L’effetto benefico del mirabegron, simile all’effetto dell’esposizione al freddo, sull’induzione del tessuto adiposo “beige” nel tessuto adiposo sottocutaneo umano è stato riportato anche in un altro studio condotto da Finlin et al. [14]. Hanno esposto al freddo i partecipanti alla ricerca magri e obesi o li hanno trattati con mirabegron. Il trattamento cronico con mirabegron (10 settimane; 50 mg/giorno) ha indotto UCP1 (3 volte) e TMEM26 (8,7 volte) nei soggetti obesi. Inoltre, l’espressione di UCP1 e dei marcatori degli adipociti “beige” è aumentata più che dopo 10 giorni di ripetuta esposizione al freddo [14].
Nello studio successivo, composto da 12 partecipanti obesi insulino-resistenti, Finlin et al. [9] hanno valutato la capacità del trattamento con pioglitazone (30 mg/giorno) o del trattamento con mirabegron (50 mg/giorno) in monoterapia, così come una combinazione di trattamento con pioglitazone (30 mg/giorno) e mirabegron (50 mg/giorno), di aumentare il grasso “beige” o migliorare ulteriormente il metabolismo del glucosio durante 12 settimane di terapia. Il pioglitazone è un attivatore PPARγ che può stimolare il BAT o “grasso beige”. Il trattamento con pioglitazone o la combinazione di pioglitazone e mirabegron hanno aumentato l’espressione del marcatore proteico del tessuto adiposo “beige” e migliorato la sensibilità all’insulina (misurata tramite clamp euglicemico, più efficace nella terapia combinata) e l’omeostasi del glucosio (inclusi test di tolleranza al glucosio migliorati, più efficaci nella terapia combinata), ma nessuno dei due trattamenti ha indotto il BAT o influenzato la spesa energetica nei soggetti obesi. Inoltre, non si è verificato alcun cambiamento significativo nel peso corporeo dopo il trattamento. Nonostante il fatto che mirabegron e pioglitazone somministrati separatamente abbiano indotto il “beiging” del tessuto adiposo, l’aggiunta di pioglitazone a mirabegron non ha migliorato il “beiging”, poiché il trattamento combinato ha prodotto un “beiging” inferiore rispetto a entrambi i farmaci somministrati singolarmente [9].
Sebbene i risultati preliminari degli studi sugli animali abbiano mostrato i benefici della co-somministrazione di mirabegron e metformina nella prevenzione e nel trattamento dell’obesità [2], a nostra conoscenza, l’influenza di tale terapia combinata non è stata verificata in relazione all’attività BAT, al dispendio energetico e alla perdita di peso negli esseri umani. È noto solo che non ci sono interazioni clinicamente significative tra metformina e mirabegron. Nello studio con 32 soggetti maschi sani (BMI: 18–30 kg/m2), mirabegron (160 mg somministrati una volta al giorno) non ha mostrato alcun effetto sulla farmacocinetica di metformina (500 mg somministrati due volte al giorno). La co-somministrazione di mirabegron con metformina ha determinato piccole modifiche nell’esposizione a mirabegron (AUC e Cmax diminuite del 21%). Le modifiche farmacocinetiche osservate non sono state considerate clinicamente rilevanti. Pertanto, non è necessario alcun aggiustamento del dosaggio di mirabegron quando viene co-somministrato con metformina [33].
Sebbene i dati confermino che una bassa dose di mirabegron può indurre il “beiging” del WAT sottocutaneo, è stato riportato che 50 mg di mirabegron durante il trattamento a breve termine (circa 12 settimane di terapia) non hanno alcun effetto sulla quantità di BAT, sul dispendio energetico a riposo e sulla perdita di peso. Pertanto, sono necessari studi clinici di lunga durata, con partecipanti obesi con una dose inferiore di mirabegron, per valutare se il “beiging” del tessuto adiposo si tradurrebbe in un miglioramento del dispendio energetico a riposo e in una significativa perdita di peso.
Conclusioni:
Il BAT metabolicamente attivo è stato correlato positivamente al miglioramento dell’energia, del glucosio e del metabolismo dell’intero corpo [34]. L’attivazione del BAT e l’induzione del processo di “browning” nel WAT sembrano essere un’interessante strategia terapeutica per aumentare la spesa energetica e migliorare il metabolismo. Il mirabegron, come agonista del recettore β3-adrenergico, si è rivelato efficace come attivatore del BAT, stimolatore delle cellule “beige” e regolatore dell’omeostasi metabolica sia negli studi sugli animali che negli esseri umani. Sebbene negli studi sugli animali la somministrazione di mirabegron abbia portato a un miglioramento dell’obesità, non è stata ancora dimostrata una significativa perdita di peso nei pazienti obesi dopo dosi elevate o basse del farmaco. Ciò può essere spiegato dalla durata troppo breve degli studi e dal numero esiguo di partecipanti agli studi. Inoltre, negli esseri umani, il trattamento più efficace per la stimolazione del BAT e del WAT è stato quello con dosi elevate di mirabegron; tuttavia, gli effetti collaterali cardiovascolari possono limitare l’uso di dosi superiori a quelle approvate dalla FDA per il trattamento della vescica iperattiva. Da un lato, considerando l’uso di dosi elevate di mirabegron, deve essere valutata la sicurezza a lungo termine in relazione al sistema cardiovascolare. In caso di attivazione aggravata dei recettori β1 miocardici, la somministrazione concomitante di 100-200 mg di mirabegron con un bloccante β1-AR può essere una strategia terapeutica utile per evitare effetti collaterali cardiovascolari. D’altro canto, dovrebbe essere valutato se dosi più piccole di mirabegron, ad esempio quelle approvate per la vescica iperattiva (50 mg al giorno), assunte per un periodo di tempo più lungo, saranno sufficienti a stimolare la crescita del BAT, l’imbrunimento del WAT e la termogenesi che può portare alla perdita di peso. Negli studi clinici riguardanti l’efficacia e la sicurezza del mirabegron nei pazienti con vescica iperattiva, l’influenza del mirabegron sul peso corporeo non è stata verificata. A nostra conoscenza, l’efficacia del mirabegron in relazione ai disturbi metabolici, inclusa l’obesità, nei soggetti trattati per vescica iperattiva, non è stata finora valutata.
Si potrebbe quindi ipotizzare che, il potenziale ruolo del Mirabegron nel trattamento o nella prevenzione dell’obesità dipenderebbe dai risultati della sua efficacia determinati da studi clinici a lungo termine. In caso di mancanza o insoddisfacente effetto dimagrante (rispetto ai farmaci attualmente disponibili approvati per il trattamento dell’obesità), il mirabegron potrebbe essere utilizzato per migliorare il profilo metabolico nei pazienti obesi. Se l’effetto dimagrante del mirabegron venisse confermato, il farmaco diventerebbe un’opzione alternativa agli attuali agenti anti-obesità, specialmente nei pazienti con controindicazioni o intolleranza ad altri farmaci. Inoltre, un aspetto interessante da valutare negli studi clinici sarebbe se la co-somministrazione di mirabegron e altri farmaci, come metformina, pioglitazone o altri farmaci anti-obesità attualmente utilizzati, potrebbe essere una strategia più efficace rispetto alla somministrazione di tali farmaci da soli per migliorare i profili metabolici o per trattare l’obesità. I benefici della co-somministrazione di mirabegron e metformina nella prevenzione e nel trattamento dell’obesità, dimostrati in studi sugli animali, devono essere confermati in ulteriori studi clinici. Sebbene i risultati preliminari della co-somministrazione di mirabegron e pioglitazone in partecipanti obesi non abbiano indicato alcuna influenza di tale terapia sul peso corporeo, devono essere eseguiti ulteriori studi per confermare questi risultati. Pertanto, l’introduzione di agonisti del recettore β3-adrenergico nel trattamento dell’obesità in futuro richiederà studi a lungo termine con un numero maggiore di soggetti per valutarne l’efficacia, la tollerabilità e la sicurezza.
I tessuti adiposi bruni e “beige” rimangono un bersaglio attraente per combattere le malattie metaboliche. Sono necessari ulteriori studi per confermare se la combinazione di agenti attivatori di BAT e “beige”, esercizi fisici e una dieta ipocalorica sana sarebbe una strategia di successo per ottenere la perdita di peso nei pazienti con obesità.
E per l’uso off-label nella ricomposizione corporea? I test in tal senso sono ancora scarsi e dal design spesso pessimo. Vi sono stati riscontri positivi, almeno preliminarmente parlando, con protocolli di 8-12 settimane a dosaggi di 75-100mg/die. Il dosaggio era stato settato partendo da 25mg/die per poi mantenere e osservare le risposte al dosaggio per qualche giorno [pressione, battito cardiaco ecc…]. L’uso del Mirabegron in ambito sportivo è tanto pionieristico come lo è quello dei tireomimetici. Ci vorranno ancora diversi studi per poter essere maggiormente certi di concreti vantaggi applicativi di questa molecola come PEDs.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Elia, E.F.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell. Metab.2015, 21, 33–38. [Google Scholar] [CrossRef]
Zhao, X.Y.; Liu, Y.; Zhang, X.; Zhao, B.C.; Burley, G.; Yang, Z.C.; Luo, Y.; Li, A.Q.; Zhang, R.X.; Liu, Z.Y.; et al. The combined effect of metformin and mirabegron on diet-induced obesity. Med. Comm.2023, 4, e207. [Google Scholar] [CrossRef]
Nahon, K.J.; Janssen, L.G.M.; Sardjoe Mishre, A.S.D.; Bilsen, M.P.; van der Eijk, J.A.; Botani, K.; Overduin, L.A.; Ruiz, J.R.; Burakiewicz, J.; Dzyubachyk, O.; et al. The effect of mirabegron on energy expenditure and brown adipose tissue in healthy lean South Asian and Europid men. Diabetes Obes. Metab.2020, 22, 2032–2044. [Google Scholar] [CrossRef]
Hao, L.; Scott, S.; Abbasi, M.; Zu, Y.; Khan, M.S.H.; Yang, Y.; Wu, D.; Zhao, L.; Wang, S. Beneficial Metabolic Effects of Mirabegron In Vitro and in High-Fat Diet-Induced Obese Mice. J. Pharmacol. Exp. Ther.2019, 369, 419–427. [Google Scholar] [CrossRef]
Waki, H.; Yamauchi, T. Body-weight-independent glucose-lowering effect of the b3-adrenergic receptor agonist mirabegron in humans. J. Diabetes Investig.2021, 12, 689–690. [Google Scholar] [CrossRef]
Harb, E.; Kheder, O.; Poopalasingam, G.; Rashid, R.; Srinivasan, A.; Izzi-Engbeaya, C. Brown adipose tissue and regulation of human body weight. Diabetes Metab. Ress. Rev.2023, 39, e3594. [Google Scholar] [CrossRef]
Finlin, B.S.; Memetimin, H.; Zhu, B.; Confides, A.L.; Vekaria, H.J.; El Khouli, R.H.; Johnson, Z.R.; Westgate, P.M.; Chen, J.; Morris, A.J.; et al. The β3-adrenergic receptor agonist mirabegron improves glucose homeostatsis in obese humans. J. Clin. Investig.2020, 130, 2319–2331. [Google Scholar] [CrossRef]
Finlin, B.S.; Memetimin, H.; Confides, A.L.; Kasza, I.; Zhu, B.; Vekaria, H.J.; Harfmann, B.; Jones, K.A.; Johnson, Z.R.; Westgate, P.M.; et al. Human adipose beiging in response to cold and mirabegron. JCI Insight2018, 3, e121510. [Google Scholar] [CrossRef]
Dehvari, N.; Sato, M.; Bokhari, M.H.; Kalinovich, A.; Ham, S.; Gao, J.; Nguyen, H.T.M.; Whiting, L.; Mukaida, S.; Merlin, J.; et al. The metabolic effects of mirabegron are mediated primarily by β3-adrenoceptors. Pharmacol. Res. Perspect.2020, 8, e00643. [Google Scholar] [CrossRef]
Dehvari, N.; da Silva Junior, E.D.; Bengtsson, T.; Hutchinson, D.S. Mirabegron: Potential off target effects and uses beyond the bladder. Br. J. Pharmacol.2018, 175, 4072–4082. [Google Scholar] [CrossRef]
Baskin, A.S.; Linderman, J.D.; Brycha, R.J.; McGehee, S.; Anflick-Chames, E.; Cero, C.; Johnson, J.W.; O’Mara, A.E.; Fletcher, L.A.; Leitner, B.P.; et al. Regulation of Human Adipose Tissue Activation, Gallbladder Size, and Bile Acid Metabolism by a b3-Adrenergic Receptor Agonis. Diabetes2018, 67, 2113–2125. [Google Scholar] [CrossRef]
Pereas Valgas da Silva, C.; Calmasini, F.; Alexandre, E.C.; Raposo, H.F.; Delbin, M.A.; Monica, F.Z.; Zanesco, A. The effects of mirabegron on obesity-induced inflammation and insulin resistance are associated with brown adipose tissue activation but not beiging in the subcutaneous white adipose tissue. Clin. Exp. Pharmacol. Physiol.2021, 48, 1477–1487. [Google Scholar] [CrossRef]
Arch, J.R.S. Challenges in β3-adrenoceptor agonist drug development. Ther. Adv. Endocrinol. Metab.2011, 2, 59–64. [Google Scholar] [CrossRef]
Liu, X.; Pérusse, F.; Bukowiecki, L.J. Mechanisms of the antidiabetic effects of the beta 3-adrenergic agonist CL-316243 in obese Zucker-ZDF rats. Am. J. Physiol.1998, 274, R1212–R1219. [Google Scholar]
Takasu, T.; Ukai, M.; Sato, S.; Matsui, T.; Nagase, I.; Maruyama, T.; Sasamata, M.; Miyata, K.; Uchida, H.; Yamaguchi, O. Effect of (R)-2-(2-aminothiazol-4-yl)-4′-{2-[(2-hydroxy-2-phenylethyl)amino]ethyl} acetanilide (YM178), a novel selective beta3-adrenoceptor agonist, on bladder function. J. Pharmacol. Exp. Ther.2007, 321, 642–647. [Google Scholar] [CrossRef]
Igawa, Y.; Michel, M.C. Pharmacological profile of β3-adrenoceptor agonists in clinical development for the treatment of overactive bladder syndrome. Naunyn. Schmiedebergs. Arch. Pharmacol.2013, 386, 177–183. [Google Scholar] [CrossRef] [PubMed]
Krhut, J.; Martan, A.; Zachoval, R.; Hanus, T.; Svabik, K.; Zvara, P. Impact of body mass index on treatment efficacy of mirabegron for overactive bladder in females. Eur. J. Obstet. Gynecol. Reprod. Biol.2016, 196, 64–68. [Google Scholar] [CrossRef] [PubMed]
Hatanaka, T.; Ukai, M.; Watanabe, M.; Someya, A.; Ohtake, A.; Suzuki, M.; Ueshima, K.; Sato, S.; Sasamata, M. In vitro and in vivo pharmacological profile of the selective β3-adrenoceptor agonist mirabegron in rats. Naunyn. Schmiedebergs. Arch. Pharmacol.2013, 386, 247–253. [Google Scholar] [CrossRef]
Nagiri, C.; Kobayashi, K.; Tomita, A.; Kato, M.; Kobayashi, K.; Yamashita, K.; Nishizawa, T.; Inoue, A.; Shihoya, W.; Nureki, O. Cryo-EM structure of the β3-adrenergic receptor reveals the molecular basis of subtype selectivity. Mol. Cell.2021, 81, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
Brucker, B.M.; King, J.; Mudd, P.N., Jr.; McHale, K. Selectivity and Maximum Response of Vibegron and Mirabegron for β3-Adrenergic Receptors. Curr. Ther. Res. Clin. Exp.2022, 96, 100674. [Google Scholar] [CrossRef]
Alexandre, E.C.; Kiguti, L.R.; Calmasini, F.B.; Silva, F.H.; da Silva, K.P.; Ferreira, R.; Ribeiro, C.A.; Mónica, F.Z.; Pupo, A.S.; Antunes, E. Mirabegron relaxes urethral smooth muscle by a dual mechanism involving β3 -adrenoceptor activation and α1 -adrenoceptor blockade. Br. J. Pharmacol.2016, 173, 415–428. [Google Scholar] [CrossRef]
De Stefano, A.; Schinzari, F.; Di Daniele, N.; Sica, G.; Gentileschi, P.; Vizioli, G.; Cardillo, C.; Tesauro, M. Mirabegron relaxes arteries from human visceral adipose tissue through antagonism of α1-adrenergic receptors. Vascul. Pharmacol.2022, 146, 107094. [Google Scholar] [CrossRef]
Lowell, B.B.; S-Susulic, V.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature1993, 366, 740–742. [Google Scholar] [CrossRef]
Hamann, A.; Flier, J.S.; Lowell, B.B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology1996, 137, 21–29. [Google Scholar] [CrossRef] [PubMed]
Lv, Z.; Guo, Y. Metformin and its benefits for various diseases. Front. Endocrinol.2020, 11, 191. [Google Scholar] [CrossRef] [PubMed]
Groen-Wijnberg, M.; van Dijk, J.; Krauwinkel, W.; Kerbusch, V.; Meijer, J.; Tretter, R.; Zhang, W.; van Gelderen, M. Pharmacokinetic Interactions Between Mirabegron and Metformin, Warfarin, Digoxin or Combined Oral Contraceptives. Eur. J. Drug Metab. Pharmacokinet.2017, 42, 417–429. [Google Scholar] [CrossRef] [PubMed]
Bel, J.C.; Tai, T.C.; Khaper, N.; Lees, S.J. Mirabegron: The most promising adipose tissue beiging agent. Physiol. Rep.2021, 9, e14779. [Google Scholar] [CrossRef]
I Glucocorticoidi (o, meno comunemente, glucocorticosteroidi) sono una classe di corticosteroidi, ovvero una classe di ormoni steroidei. I Glucocorticoidi sono corticosteroidi che si legano al recettore dei glucocorticoidi[1], presente in quasi tutte le cellule animali vertebrate. Il nome “Glucocorticoide” è un portmanteau (glucosio + corteccia + steroide) ed è composto dal suo ruolo nella regolazione del metabolismo del glucosio, dalla sintesi nella corteccia surrenale e dalla sua struttura steroidea.
I Glucocorticoidi fanno parte del meccanismo di feedback del sistema immunitario, che riduce alcuni aspetti della funzione immunitaria, come l’infiammazione. Sono quindi utilizzati in medicina per trattare le malattie causate da un sistema immunitario iperattivo, come allergie, asma, malattie autoimmuni e sepsi. I Glucocorticoidi hanno molti effetti diversi, come la pleiotropia, tra cui effetti collaterali potenzialmente dannosi.[2] Inoltre, interferiscono con alcuni meccanismi anomali delle cellule tumorali, per cui vengono utilizzati in dosi elevate per il trattamento del cancro. Ciò include effetti inibitori sulla proliferazione dei linfociti, come nel trattamento di linfomi e leucemie, e l’attenuazione degli effetti collaterali dei farmaci antitumorali.
I glucocorticoidi agiscono sulle cellule legandosi al Recettore dei Glucocorticoidi (GR). Il complesso recettore glucocorticoide-glucocorticoide attivato regola l’espressione di proteine antinfiammatorie nel nucleo (processo noto come transattivazione) e reprime l’espressione di proteine pro-infiammatorie nel citosol impedendo la traslocazione di altri fattori di trascrizione dal citosol al nucleo (transrepressione).[2]
Il Recettore dei Glucocorticoidi (GR o GCR), noto anche come NR3C1 (sottofamiglia 3 del recettore nucleare, gruppo C, membro 1), è il recettore a cui si legano il Cortisolo e altri Glucocorticoidi.
I glucocorticoidi si distinguono dai mineralocorticoidi e dagli steroidi sessuali per i loro specifici recettori, cellule bersaglio ed effetti. In termini tecnici, il termine “corticosteroide” si riferisce sia ai glucocorticoidi che ai mineralocorticoidi (poiché entrambi sono mimici degli ormoni prodotti dalla corteccia surrenale), ma è spesso usato come sinonimo di “glucocorticoide”. I glucocorticoidi sono prodotti principalmente nella zona fascicolata della corteccia surrenale, mentre i mineralocorticoidi sono sintetizzati nella zona glomerulosa.
Il Cortisolo (o idrocortisone) è il più importante glucocorticoide umano. È essenziale per la vita e regola o supporta una serie di importanti funzioni cardiovascolari, metaboliche, immunologiche e omeostatiche. Gli aumenti delle concentrazioni di glucocorticoidi sono parte integrante della risposta allo stress e sono i biomarcatori più comunemente utilizzati per misurare lo stress.[3] I glucocorticoidi hanno anche numerose funzioni non correlate allo stress e le concentrazioni di glucocorticoidi possono aumentare in risposta al piacere o all’eccitazione.[4] Sono disponibili diversi glucocorticoidi sintetici, ampiamente utilizzati nella pratica medica generale e in numerose specialità, come terapia sostitutiva in caso di carenza di glucocorticoidi o per sopprimere il sistema immunitario dell’organismo.
Si sospetta che gli atleti utilizzino i Glucocorticoidi per migliorare le prestazioni atletiche fin dagli anni ’60 (5). I glucocorticoidi possono migliorare le prestazioni fisiche attraverso diverse vie, tra cui una maggiore disponibilità di substrati metabolici attraverso un aumento della lipolisi (6), della proteolisi (7) e della disponibilità di glucosio (5). I Glucocorticoidi possono anche avere effetti immunosoppressivi e antinfiammatori che possono impedire al sistema immunitario di reagire in modo eccessivo a seguito di danni muscolari indotti dall’esercizio fisico (8), e il Cortisolo sembra preparare l’organismo per l’esercizio successivo (9). Inoltre, i glucocorticoidi possono stimolare i recettori cerebrali dei glucocorticoidi, determinando una riduzione del dolore muscolare durante l’esercizio, un aumento della soglia della fatica e risposte edoniche positive, che possono tradursi in un aumento delle prestazioni fisiche (10). I glucocorticoidi hanno un elevato potenziale di effetti avversi, tra cui il deperimento muscolare, e questi effetti dipendono da molteplici fattori come il tipo di glucocorticoide, la durata del trattamento, la dose e la via di somministrazione (11). Pertanto, gli atleti che assumono glucocorticoidi per migliorare le prestazioni probabilmente preferiscono periodi di somministrazione più brevi.
L’Associazione Mondiale Antidoping (WADA) aggiorna annualmente la sua lista proibita, un elenco di sostanze e metodi vietati negli sport d’élite. La lista proibita, in vigore dal 1° gennaio 2022, vieta tutti i glucocorticoidi nelle competizioni quando sono somministrati per via orale, rettale o iniettabile, poiché queste forme di somministrazione sono considerate ad effetto sistemico. Altre vie di somministrazione, come l’applicazione topica per inalazione e le iniezioni locali, sono approvate nelle competizioni, poiché si ritiene che abbiano un minore potenziale di miglioramento delle prestazioni. L’uso dei glucocorticoidi al di fuori delle competizioni è approvato (12). Per aiutare i medici a curare gli atleti e a rispettare le norme antidoping, dal 2022 la WADA raccomanda specifici periodi di wash out (tempo dall’ultima dose al giorno prima della competizione) per i diversi tipi di glucocorticoidi e le diverse vie di somministrazione. Dal 2022 la WADA ha anche introdotto livelli di segnalazione urinaria specifici per i diversi tipi di glucocorticoidi, in quanto i diversi glucocorticoidi hanno un’ampia variazione nel tempo di eliminazione (13).
Dagli anni 90 si è studiato l’effetto dei glucocorticoidi sulla frequenza cardiaca e sul consumo di ossigeno durante intervalli di corsa ad alta intensità. Da allora diversi studi RCT hanno analizzato l’effetto dei glucocorticoidi sulla prestazione fisica submassimale e massimale. Poiché le modalità di somministrazione, il tipo di glucocorticoide, la durata del trattamento, la dose, la popolazione e il protocollo di esercizio possono variare tra gli studi, essi giungono a conclusioni diverse per quanto riguarda l’effetto dei glucocorticoidi sulla prestazione fisica, tuttavia sono soprattutto gli studi che indagano l’effetto di alte dosi orali somministrate nel tempo sulla resistenza in bicicletta fino all’esaurimento a mostrare effetti. L’effetto dei glucocorticoidi sulla prestazione fisica e sul metabolismo dei soggetti sani è stato esaminato in precedenza (5, 10), ma senza una ricerca sistematica della letteratura e senza combinare i risultati degli studi inclusi nell’analisi statistica. Pertanto, in tale sede, cercherò anche di fare chiarezza sull’effetto dei glucocorticoidi sulla prestazione massimale o submassimale in soggetti sani.
Steroidogenesi dei Glucocorticoidi:
La steroidogenesi è il processo biologico attraverso il quale gli steroidi vengono generati dal colesterolo e trasformati in altri steroidi.[14] Le vie della steroidogenesi differiscono tra le specie. Le principali classi di ormoni steroidei, con i loro membri e le loro funzioni principali, sono i progestinici, i corticosteroidi (corticoidi), gli androgeni e gli estrogeni.[15][16] La steroidogenesi umana dei corticosteroidi avviene nella Corteccia Surrenale.
Steroidogenesi che mostra i Glucocorticoidi nell’ellisse verde a destra con l’esempio primario del Cortisolo. Non è un gruppo strettamente delimitato, ma un continuum di strutture con effetto glucocorticoide crescente.
La Corteccia Surrenale è la regione più esterna e anche la parte più grande della ghiandola surrenale. È divisa in tre zone distinte: zona glomerulosa, zona fascicolata e zona reticolare. Ogni zona è responsabile della produzione di ormoni specifici. È anche un sito secondario di sintesi degli androgeni.[17]
Corteccia Surrenale
Zona Glomerulosa
Colorazione H&E della Corteccia Surrenale. La Zona Glomerulare è lo strato più esterno, sotto la capsula renale (vicino all’indicatore)
Lo strato più esterno, la zona glomerulosa, è il sito principale per la produzione di aldosterone, un mineralcorticoide. La sintesi e la secrezione di aldosterone sono regolate principalmente dal sistema renina-angiotensina-aldosterone. Le cellule della zona glomerulosa esprimono un enzima specifico, l’aldosterone sintasi (noto anche come CYP11B2).[18][19] L’aldosterone è ampiamente responsabile della regolazione a lungo termine della pressione sanguigna. [Gli effetti dell’aldosterone si manifestano nel tubulo contorto distale e nel dotto collettore del rene, dove provoca un aumento del riassorbimento del sodio e una maggiore escrezione di potassio (da parte delle cellule principali) e di ioni idrogeno (da parte delle cellule intercalate del dotto collettore).[20] La ritenzione di sodio è anche una risposta del colon distale e delle ghiandole sudoripare alla stimolazione dei recettori dell’aldosterone. Sebbene la produzione sostenuta di aldosterone richieda un ingresso persistente di calcio attraverso canali del Ca2+ attivati a basso voltaggio, le cellule isolate della zona glomerulosa sono considerate non eccitabili, con tensioni di membrana registrate troppo iperpolarizzate per consentire l’ingresso di canali del Ca2+.[21]
La secrezione di aldosterone è stimolata anche dall’ormone adrenocorticotropo (ACTH).[22]
Le cellule della zona glomerulosa non esprimono l’11β-idrossilasi e la 17α-idrossilasi. Per questo motivo la zona glomerulosa non può sintetizzare cortisolo, corticosterone o ormoni sessuali (androgeni). [23] L’espressione di proteine specifiche per i neuroni nelle cellule della zona glomerulosa dei tessuti adrenocorticali umani è stata prevista e riportata da diversi autori [24][25][26] ed è stato suggerito che l’espressione di proteine come la molecola di adesione delle cellule neuronali (NCAM) nelle cellule della zona glomerulosa rifletta la caratteristica rigenerativa di queste cellule, che perderebbero l’immunoreattività della NCAM dopo essersi spostate nella zona fascicolata. [24][27] Tuttavia, insieme ad altri dati sulle proprietà neuroendocrine delle cellule della zona glomerulosa, l’espressione di NCAM potrebbe riflettere una differenziazione neuroendocrina di queste cellule.[24]
Zona Fasciculata
Zona Fasciculata
Situate tra la glomerulosa e la reticolare, le cellule della zona fascicolata sintetizzano e secernono glucocorticoidi (come l’11-deossicorticosterone, il corticosterone e il cortisolo), oltre a piccole quantità di androgeni ed estrogeni surrenalici.[28] La zona fascicolata ha una maggiore attività di 3β-idrossisteroide deidrogenasi rispetto alla zona reticolare. Pertanto, la zona fascicolata produce più 11-deossicorticosterone, corticosterone e cortisolo.[23] Il principale ormone che stimola la secrezione di cortisolo nell’uomo è l’ACTH, rilasciato dall’ipofisi anteriore.[22] È stato dimostrato che la capacità steroidogenica della zona fascicolata aumenta durante la malattia nei neonati.[22]
Zona Reticolare
Zona Reticolare
La zona reticolare, lo strato corticale più interno, produce gli androgeni surrenalici, oltre a piccole quantità di estrogeni e alcuni glucocorticoidi.[28] La zona reticolare possiede una quantità maggiore di cofattori necessari per l’attività della 17,20-liasi della 17α-idrossilasi rispetto alla zona fascicolata. Pertanto, la zona reticolare produce più androgeni,[23] soprattutto deidroepiandrosterone (DHEA), DHEA solfato (DHEA-S) e androstenedione (il precursore del Testosterone e DHT) nell’uomo. La secrezione di DHEAS è stimolata anche dall’ACTH.[22]
Come abbiamo visto, I glucocorticoidi sono prodotti principalmente nella Zona Fascicolata.[23]
Il precursore degli steroidi sintetizzati nella corteccia surrenale è il colesterolo che viene immagazzinato nelle vescicole. Il colesterolo può essere sintetizzato de novo nella corteccia surrenale. Tuttavia, la fonte principale di colesterolo sembra essere il colesterolo assunto dalle lipoproteine circolanti. [29]
I passaggi fino a questo punto avvengono in molti tessuti produttori di steroidi. Le fasi successive per generare aldosterone e cortisolo, tuttavia, avvengono principalmente nella corteccia surrenale:
Progesterone → (idrossilazione a C21) → 11-Deossicorticosterone → (due ulteriori idrossilazioni a C11 e C18) → Aldosterone
Progesterone → (idrossilazione a C17) → 17-alfa-idrossiprogesterone → (idrossilazione a C21) → 11-Deossicortisolo → (idrossilazione a C11) → Cortisolo
Fasi della sintesi dell’ormone steroideo surrenale
Effetto sistemico dei Glucocorticoidi:
Gli effetti dei glucocorticoidi possono essere ampiamente classificati in due categorie principali:
immunologici
metabolici.
Inoltre, i glucocorticoidi svolgono ruoli importanti nello sviluppo fetale e nell’omeostasi dei fluidi corporei.
Immunità
Come già accennato, i glucocorticoidi funzionano anche attraverso l’interazione con il recettore dei glucocorticoidi:
Aumentano l’espressione di proteine antinfiammatorie.
Riducono l’espressione di proteine pro-infiammatorie.
Micrografia elettronica a scansione di un globulo rosso (sinistra), una piastrina (centro) e un linfocita T (destra); colorato
È stato dimostrato che i glucocorticoidi svolgono un ruolo nello sviluppo e nell’omeostasi dei linfociti T. Questo è stato dimostrato in transgenici. Ciò è stato dimostrato in topi transgenici con una maggiore o minore sensibilità della linea delle cellule T ai glucocorticoidi.[30]
Metabolismo
Nello stato di digiuno, il cortisolo stimola diversi processi che servono collettivamente ad aumentare e mantenere le normali concentrazioni di glucosio nel sangue.
Effetti metabolici:
Stimolazione della gluconeogenesi, in particolare nel fegato: Questa via porta alla sintesi del glucosio a partire da substrati non esosi, come gli aminoacidi e il glicerolo proveniente dalla scissione dei trigliceridi, ed è particolarmente importante nei carnivori e in alcuni erbivori. L’aumento dell’espressione degli enzimi coinvolti nella gluconeogenesi è probabilmente la funzione metabolica più nota dei glucocorticoidi.
Mobilitazione di aminoacidi dai tessuti extraepatici: Questi servono come substrati per la gluconeogenesi.
Inibizione della captazione del glucosio nel tessuto muscolare e adiposo: Un meccanismo per conservare il glucosio
Stimolazione della demolizione dei grassi nel tessuto adiposo: Gli acidi grassi rilasciati dalla lipolisi vengono utilizzati per la produzione di energia in tessuti come il muscolo e il glicerolo rilasciato fornisce un altro substrato per la gluconeogenesi.
L’aumento della ritenzione di sodio e dell’escrezione di potassio porta a ipernatremia e ipokaliemia[31].
Aumento della concentrazione di emoglobina, probabilmente dovuto all’ostacolo dell’ingestione di globuli rossi da parte di macrofagi o altri fagociti[32].
Aumento dell’acido urico urinario[33]
Aumento del calcio urinario e ipocalcemia[34]
Alcalosi[35]
Leucocitosi[36]
Il Cortisolo innesca una cascata di eventi che influenzano l’omeostasi del glucosio. Il fegato, i muscoli scheletrici, i tessuti adiposi bianchi e il pancreas svolgono un ruolo chiave nell’assicurare un apporto continuo di energia utilizzabile per la risposta di lotta/fuga.
Livelli eccessivi di glucocorticoidi derivanti dalla somministrazione di farmaci o dall’iperadrenocorticismo hanno effetti su molti sistemi. Alcuni esempi includono l’inibizione della formazione ossea, la soppressione dell’assorbimento del calcio (entrambi possono portare all’osteoporosi), il ritardo nella guarigione delle ferite, la debolezza muscolare e l’aumento del rischio di infezioni. Queste osservazioni suggeriscono una moltitudine di ruoli fisiologici meno drammatici per i glucocorticoidi.[30]
Eccitazione e sfera cognitiva
Principali scissure e lobi del cervello visti lateralmente (il lobo frontale è mostrato in blu).
I glucocorticoidi agiscono sull’ippocampo, sull’amigdala e sui lobi frontali. Insieme all’adrenalina, favoriscono la formazione di ricordi flashbulb di eventi associati a forti emozioni, sia positive che negative.[36] Ciò è stato confermato da studi in cui il blocco dell’attività dei glucocorticoidi o della noradrenalina ha compromesso il richiamo di informazioni emotivamente rilevanti. Ulteriori fonti hanno dimostrato che i soggetti il cui apprendimento della paura è stato accompagnato da alti livelli di cortisolo hanno avuto un migliore consolidamento di questa memoria (questo effetto è stato più importante negli uomini). L’effetto che i glucocorticoidi hanno sulla memoria può essere dovuto a un danno specifico all’area CA1 della formazione dell’ippocampo.
In diversi studi sugli animali, lo stress prolungato (che causa aumenti prolungati dei livelli di glucocorticoidi) ha mostrato la distruzione dei neuroni nell’area dell’ippocampo del cervello, che è stata collegata a prestazioni di memoria inferiori.[32][37][33]
Una rappresentazione grafica della curva di Yerkes-Dodson
È stato inoltre dimostrato che i glucocorticoidi hanno un impatto significativo sulla vigilanza (disturbo da deficit di attenzione) e sulla cognizione (memoria). Questo sembra seguire la curva di Yerkes-Dodson, in quanto gli studi hanno dimostrato che i livelli circolanti di glucocorticoidi rispetto alle prestazioni della memoria seguono un andamento a U rovesciata, proprio come la curva di Yerkes-Dodson. Ad esempio, il potenziamento a lungo termine (LTP, il processo di formazione dei ricordi a lungo termine) è ottimale quando i livelli di glucocorticoidi sono leggermente elevati, mentre si osserva una significativa riduzione dell’LTP dopo la surrenalectomia (stato di basso livello di glucocorticoidi) o dopo la somministrazione di glucocorticoidi esogeni (stato di alto livello di glucocorticoidi). Livelli elevati di glucocorticoidi migliorano la memoria per gli eventi emotivamente eccitanti, ma portano più spesso a una scarsa memoria per il materiale non correlato alla fonte di stress/eccitazione emotiva.[38] In contrasto con gli effetti di potenziamento dose-dipendenti dei glucocorticoidi sul consolidamento della memoria, è stato dimostrato che questi ormoni dello stress inibiscono il recupero di informazioni già memorizzate. [È stato dimostrato che l’esposizione a lungo termine a farmaci glucocorticoidi, come quelli contro l’asma e gli antinfiammatori, crea deficit di memoria e attenzione sia durante che, in misura minore, dopo il trattamento,[39][40] una condizione nota come “demenza da steroidi”.[41]
Omeostasi dei fluidi corporei
I glucocorticoidi potrebbero agire a livello centrale e periferico per contribuire alla normalizzazione del volume dei liquidi extracellulari regolando l’azione dell’organismo nei confronti del peptide natriuretico atriale (ANP). A livello centrale, i glucocorticoidi potrebbero inibire l’assunzione di acqua indotta dalla disidratazione;[42] a livello periferico, i glucocorticoidi potrebbero indurre una potente diuresi.[43]
Metabolismo dei Glucocorticoidi. La secrezione di Glucocorticoidi da parte della ghiandola surrenale è regolata dall’asse HPA tramite secrezione di ACTH. Il cortisolo plasmatico principale (F) è legato alle proteine con una frazione libera del 4-5%. Il Cortisone plasmatico (E) è nella forma libera non legata. L’equilibrio di cortisolo e cortisone tra plasma e tessuti è illustrato con le frecce bidirezionali tratteggiate. È anche raffigurato il metabolismo dei GC tessuto-specifici. I GC sono metabolizzati principalmente nel fegato e i metaboliti sono escreti nelle urine. Sono mostrati solo i tessuti rilevanti per la sindrome metabolica. THE, tetraidrocortisone; THF, tetraidrocortisolo.
Meccanismi d’azione dei Glucocorticoidi:
Transattivazione
I glucocorticoidi si legano al recettore citosolico dei glucocorticoidi, un tipo di recettore nucleare che viene attivato dal legame con il ligando. Dopo che un ormone si lega al recettore corrispondente, il complesso appena formato si trasloca nel nucleo della cellula, dove si lega agli elementi di risposta ai glucocorticoidi nella regione promotrice dei geni bersaglio, determinando la regolazione dell’espressione genica. Questo processo viene comunemente definito attivazione trascrizionale o transattivazione.[44][45]
Le proteine codificate da questi geni regolati hanno un’ampia gamma di effetti, tra cui, ad esempio:[45]
L’Annessina [Lipocortina I]
Antinfiammatori – lipocortina I, proteina legante p11/calpactina, inibitore secretorio della proteasi leucocitaria 1 (SLPI) e fosfatasi della proteina chinasi attivata dal mitogeno (MAPK fosfatasi).
Aumento della gluconeogenesi – glucosio 6-fosfatasi e tirosina aminotransferasi
Transrepressione
Il meccanismo opposto è chiamato repressione trascrizionale o transrepressione. Secondo la concezione classica di questo meccanismo, il recettore dei glucocorticoidi attivato si lega al DNA nello stesso sito in cui si legherebbe un altro fattore di trascrizione, impedendo la trascrizione di geni che vengono trascritti tramite l’attività di quel fattore.[44][45] Sebbene ciò avvenga, i risultati non sono coerenti per tutti i tipi di cellule e per tutte le condizioni; non esiste un meccanismo generale e generalmente accettato per la transrepressione.[45]
Meccanismo d’azione di NF-κB.
Si stanno scoprendo nuovi meccanismi in cui la trascrizione viene repressa, ma il recettore dei glucocorticoidi attivato non interagisce con il DNA, bensì direttamente con un altro fattore di trascrizione, interferendo con esso, o con altre proteine che interferiscono con la funzione di altri fattori di trascrizione. Quest’ultimo meccanismo sembra essere il modo più probabile in cui il recettore glucocorticoide attivato interferisce con NF-κB, ossia reclutando istone deacetilasi, che deacetilano il DNA nella regione del promotore portando alla chiusura della struttura cromatinica in cui NF-κB deve legarsi.[44][45]
Attività non-genomica
Il recettore glucocorticoide attivato ha effetti che, come è stato dimostrato sperimentalmente, sono indipendenti da qualsiasi effetto sulla trascrizione e possono essere dovuti solo al legame diretto del recettore glucocorticoide attivato con altre proteine o con l’mRNA.[44][45]
Effetti genomici e non genomici dei Glucocorticoidi. Trans-attivazione: l’effetto genomico del GC dopo il legame del GR al suo elemento di risposta positiva causa una maggiore trascrizione di proteine antinfiammatorie, ad esempio, lipocortina-1, IL-10, IL-12, MAPK fosfatasi I e IκB. Trans-repressione: l’interazione molecola-molecola tra GR attivato e fattori di trascrizione pro-infiammatori, ad esempio, AP-1 o NF-κB causa una riduzione della trascrizione di mediatori pro-infiammatori, ad esempio, Il-2, IL-3, IL-4, IL-5, IL-6, IL-13, IL-15, TNF-α e VCAM-a.
Ad esempio, la chinasi Src, che si lega al recettore glucocorticoide inattivo, viene rilasciata quando un glucocorticoide si lega al recettore glucocorticoide e fosforila una proteina che a sua volta sposta una proteina adattatrice da un recettore importante nell’infiammazione, il fattore di crescita epidermico, riducendone l’attività, che a sua volta si traduce in una riduzione della creazione di acido arachidonico, una molecola proinfiammatoria chiave. Questo è uno dei meccanismi con cui i glucocorticoidi hanno un effetto antinfiammatorio.[44]
Farmacologia dei Glucocorticoidi:
Fludrocortisone Acetato
Per uso terapeutico sono stati creati diversi Glucocorticoidi sintetici, alcuni molto più potenti del Cortisolo. Si differenziano sia per la farmacocinetica (fattore di assorbimento, emivita, volume di distribuzione, clearance) che per la farmacodinamica (ad esempio la capacità di attività mineralcorticoide: ritenzione di sodio (Na+) e acqua; fisiologia renale). Poiché permeano facilmente l’intestino, vengono somministrati principalmente per os (per bocca), ma anche con altri metodi, ad esempio per via topica sulla pelle. Oltre il 90% di essi lega diverse proteine plasmatiche, anche se con una diversa specificità di legame. I glucocorticoidi endogeni e alcuni corticoidi sintetici hanno un’elevata affinità con la proteina Transcortina (detta anche globulina legante i corticosteroidi), mentre tutti legano l’albumina. Nel fegato, vengono rapidamente metabolizzati mediante coniugazione con un solfato o un acido glucuronico e vengono secreti nelle urine.
La potenza dei Glucocorticoidi, la durata dell’effetto e la sovrapposizione della potenza dei mineralocorticoidi variano. Il cortisolo è lo standard di confronto per la potenza dei glucocorticoidi. Idrocortisone è il nome utilizzato per le preparazioni farmaceutiche di cortisolo.
I dati riportati di seguito si riferiscono alla somministrazione orale. La potenza orale può essere inferiore a quella parenterale perché quantità significative (fino al 50% in alcuni casi) possono non raggiungere la circolazione. Il Fludrocortisone Acetato e il desossicorticosterone acetato sono, per definizione, mineralocorticoidi piuttosto che glucocorticoidi, ma hanno una potenza glucocorticoide minore e sono inclusi in questa tabella per fornire una prospettiva sulla potenza dei mineralocorticoidi.
Usi terapeutici:
I Glucocorticoidi possono essere utilizzati a basse dosi nell’insufficienza surrenalica. A dosi molto più elevate, i glucocorticoidi per via orale o inalatoria sono utilizzati per sopprimere vari disturbi allergici, infiammatori e autoimmuni. I glucocorticoidi per via inalatoria sono il trattamento di seconda linea per l’asma. Sono anche somministrati come immunosoppressori post-trapianto per prevenire il rigetto acuto del trapianto e la malattia del trapianto contro l’ospite. Tuttavia, non prevengono un’infezione e inibiscono anche i successivi processi riparativi. Nuove evidenze hanno dimostrato che i glucocorticoidi potrebbero essere utilizzati nel trattamento dell’insufficienza cardiaca per aumentare la responsività renale ai diuretici e ai peptidi natriuretici. I glucocorticoidi sono storicamente utilizzati per alleviare il dolore nelle condizioni infiammatorie.[46][47][48] Tuttavia, i corticosteroidi mostrano un’efficacia limitata nell’alleviare il dolore e potenziali eventi avversi per il loro uso nelle tendinopatie.[49]
Terapia Sostitutiva
Cortisolo
Qualsiasi glucocorticoide può essere somministrato in una dose che fornisce all’incirca gli stessi effetti glucocorticoidi della normale produzione di cortisolo; si parla di dosaggio fisiologico, sostitutivo o di mantenimento. Si tratta di circa 6-12mg/m2/die di Idrocortisone (m2 si riferisce all’area di superficie corporea (BSA), ed è una misura delle dimensioni del corpo; la BSA di un uomo medio è di 1,9 m2).
Gli usi clinici dei glucocorticoidi comprendono quindi:
Terapia Immunosoppressiva: I glucocorticoidi causano immunosoppressione e la componente terapeutica di questo effetto è principalmente la diminuzione della funzione e del numero di linfociti, compresi i linfociti B e i linfociti T.
Terapia Anti-Infiammatoria: I glucocorticoidi sono potenti antinfiammatori, indipendentemente dalla causa dell’infiammazione; il loro meccanismo antinfiammatorio primario è la sintesi della lipocortina-1 (annexin-1). La lipocortina-1 sopprime la fosfolipasi A2, bloccando così la produzione di eicosanoidi, e inibisce vari eventi infiammatori dei leucociti (adesione epiteliale, emigrazione, chemiotassi, fagocitosi, esplosione respiratoria, ecc.) In altre parole, i glucocorticoidi non solo sopprimono la risposta immunitaria, ma inibiscono anche i due principali prodotti dell’infiammazione, le prostaglandine e i leucotrieni. Inibiscono la sintesi delle prostaglandine a livello della fosfolipasi A2 e a livello della cicloossigenasi/PGE isomerasi (COX-1 e COX-2),[50] quest’ultimo effetto è molto simile a quello dei FANS, potenziando così l’effetto antinfiammatorio. Inoltre, i glucocorticoidi sopprimono anche l’espressione della ciclossigenasi.[51]
Trattamento del Iperaldosteronismo: I glucocorticoidi possono essere utilizzati nella gestione dell’iperaldosteronismo familiare di tipo 1. Non sono efficaci, tuttavia, per l’uso nella condizione di tipo 2.
Trattamento insufficienza cardiaca: I glucocorticoidi possono essere utilizzati nel trattamento dell’insufficienza cardiaca scompensata per potenziare la reattività renale ai diuretici, in particolare nei pazienti con insufficienza cardiaca con resistenza diuretica refrattaria a dosi elevate di diuretici dell’ansa.[52][53][54][55][56][57][58]
Meccanismi di resistenza ai corticosteroidi
La resistenza agli usi terapeutici dei glucocorticoidi può verificarsi in un certo numero di pazienti e presentare delle difficoltà; ad esempio, il 25% dei casi di asma grave può non rispondere agli steroidi. Questo può essere il risultato di una predisposizione genetica, dell’esposizione continua alla causa dell’infiammazione (come gli allergeni), di fenomeni immunologici che bypassano i glucocorticoidi, di disturbi farmacocinetici (assorbimento incompleto o escrezione o metabolismo accelerati) e di infezioni respiratorie virali e/o batteriche.[59][60]
Glucocorticoidi e Sport:
Come abbiamo visto, i Glucocorticoidi sono una delle classi di farmaci più ampiamente utilizzate ed efficaci nella popolazione generale e sono disponibili in una varietà di formulazioni farmaceutiche (ad esempio, iniezioni, compresse, creme, colliri, gocce auricolari, inalatori e spray nasali). Somministrati sia per i loro effetti sistemici che locali, i Glucocorticoidi sono utilizzati a livello globale in una vasta gamma di specialità cliniche, principalmente per le loro proprietà antinfiammatorie e immunosoppressive. In alcuni contesti, l’uso medico dei Glucocorticoidi orali sembra essere aumentato negli ultimi anni poiché questi sono un’alternativa accessibile e conveniente ai farmaci mirati ma più costosi. La prevalenza dell’uso sistemico prevalentemente per uso a breve termine varia tra l’1% e il 3%, sebbene abbia raggiunto il 17,1% in un recente studio sugli adulti in Francia.(https://bjsm.bmj.com) Nelle popolazioni di atleti, vi è una maggiore prevalenza di lesioni muscoloscheletriche e asma, e pertanto un frequente uso legittimo di Glucocorticoidi terapeutici non sarebbe sorprendente. Tuttavia, vi è una scarsità di stime di prevalenza nelle popolazioni di atleti. Un’analisi di TUE abbreviate in cui il CIO è stato informato dell’uso di Glucocorticoidi da parte degli atleti prima dei Giochi olimpici negli anni ’90 e nei primi anni 2000 suggerisce che almeno il 5% al 12% degli atleti d’élite competitivi è stato trattato con Glucocorticoidi tramite tutte le vie, prevalentemente inalatoria. In un recente sondaggio internazionale non pubblicato di medici che lavorano con atleti d’élite, oltre l’85% ha riferito di aver somministrato almeno occasionalmente Glucocorticoidi iniettabili come parte della loro normale pratica (comunicazione personale, Dr David Hughes, Australian Institute of Sport).
I Glucocorticoidi, somministrati tramite determinate vie, sono stati proibiti per la prima volta nello sport dal CIO nel 1985 e sono stati proibiti dalla WADA sin dalla sua Lista iniziale, pubblicata nel 2004. Le sostanze o i metodi sono considerati per l’inclusione nella Lista se soddisfano due dei tre criteri seguenti come stabilito dal Codice mondiale antidoping:
potenziale di migliorare o migliorano le prestazioni sportive;
rappresentano un rischio effettivo o potenziale per la salute dell’atleta;
violano lo spirito dello sport. I Glucocorticoidi sono proibiti in competizione quando somministrati tramite vie “sistemiche” (orali, rettali, intramuscolari o endovenose).[https://www.wada-ama.org/] La somministrazione tramite tutte le altre vie (incluse le iniezioni intra-articolari e altre periarticolari) è considerata somministrazione locale e non è proibita in competizione. La somministrazione di Glucocorticoidi tramite qualsiasi via non è proibita fuori competizione (OOC).
Indipendentemente dalla sostanza specifica del Glucocorticoidi e dalle sue singole caratteristiche farmacologiche, un presunto riscontro analitico avverso (AAF) viene segnalato dai laboratori accreditati WADA quando i livelli urinari dei campioni in gara superano un livello di segnalazione di 30ng/mL. La farmacocinetica dei Glucocorticoidi è complessa e influenzata dalla formulazione, dal tipo di esterificazione e sale, dalla via di somministrazione, dal sito e dal metodo di somministrazione. Di conseguenza, mentre il limite di segnalazione del laboratorio può dimostrare la presenza di un Glucocorticoidi , non può necessariamente indicare se la somministrazione è avvenuta in gara o OOC o se è probabile che abbia un effetto farmacologico o ergogenico. Qualsiasi medico o atleta non sarà sicuro di quando interrompere l’uso di GC sistemici prima del periodo in gara per evitare di superare il limite di segnalazione. Per complicare ulteriormente il quadro farmacocinetico, le iniezioni intra-articolari possono dare origine a livelli sistemici e i medici possono inavvertitamente caratterizzare erroneamente il sito di iniezione in assenza di guida radiologica o ecografica. La definizione di limiti di segnalazione specifici per sostanza è un’area di discussione e ricerca attiva tra gli esperti nominati dalla WADA e va oltre lo scopo del presente documento.
Glucocorticoidi sistemici e performance
Alcuni atleti hanno indubbiamente tentato di sfruttare i presunti effetti di miglioramento delle prestazioni dei Glucocorticoidi sistemici che ritengono benefici nella loro particolare disciplina sportiva. Tuttavia, i meccanismi complessi e pleiotropici dell’azione dei Glucocorticoidi suggeriscono che questi farmaci sono uno strumento poco maneggevole per l’atleta che cerca di ottenere un vantaggio nelle prestazioni e sono considerati una componente meno popolare dei regimi di doping rispetto al passato.[ https://cyclingtips.com/] Alcuni pazienti e atleti hanno riferito di aver sperimentato euforia dopo la somministrazione sistemica.[ http://www.cyclingnews.com] Tuttavia, le prove scientifiche a supporto dell’euforia misurabile nelle popolazioni cliniche sono ambigue e l’interpretazione dei dati è complicata dall’associazione del dolore cronico confondente.[https://bjsm.bmj.com/]
Sembrerà starno, ma non vi è alcuna prova incontrovertibile di effetti di miglioramento delle prestazioni derivanti dall’uso a breve termine di Glucocorticoidi sistemici.[https://bjsm.bmj.com/] Esistono studi randomizzati in doppio cieco cross-over che suggeriscono che gli atleti possono sfruttare cicli di Glucococrticoidi orali ad alto dosaggio della durata di una settimana per migliorare le loro prestazioni di esercizio di intensità submassimale per brevi periodi di tempo.[https://bjsm.bmj.com/] Questi dosaggi sarebbero facilmente rilevati durante i test antidoping, se assunti in gara. Il meccanismo preciso di questo effetto non è chiaro, ma si suggerisce che derivi da una combinazione di effetti sul metabolismo energetico, sui muscoli, sull’infiammazione e sul sistema nervoso. Questo effetto del farmaco è stato dimostrato in uno studio su atleti maschi il cui allenamento era strettamente periodizzato insieme all’uso di Glucocorticoidi orali.[https://bjsm.bmj.com/] Sfruttare questo tipo di regime di miglioramento delle prestazioni evitando efficacemente l’insufficienza surrenalica e il rilevamento tramite controlli antidoping standard in gara richiederebbe una meticolosa supervisione medica. Potrebbe anche richiedere una manipolazione farmacologica più complessa ed esotica dell’asse ipotalamo-ipofisi-surrene rispetto a quella offerta dai Glucocorticoidi prescritti.[https://bjsm.bmj.com/]
Atleti e dottori hanno descritto metodi inappropriati con cui l’uso sistemico di Glucocorticoidi, un’alimentazione limitata e un allenamento a bassa intensità potrebbero essere combinati OOC per perdere peso e preservare la massa muscolare.[ https://www.nytimes.com/]Tuttavia, date le funzioni cataboliche proteiche ampiamente riconosciute dei Glucocorticoidi,[https://bjsm.bmj.com/] questo meccanismo di doping rimane speculativo e controverso. Inoltre, l’efficacia potrebbe dipendere dall’uso di Glucocorticoidi come parte di un cocktail complesso che include altri ormoni proibiti ma scarsamente rilevati come l’insulina.[https://bjsm.bmj.com/]
Schema esemplificativo sull’uso di Glucocorticoidi sistemici e perdita di peso/ricomposizione corporea:la base teorica sulla quale si sostiene la suddetta pratica farmacologica è il ciclo di feedback negativo dell’Asse HPA per via d’uso di Glucocorticoidi esogeni. Tale pratica dovrebbe portare a 1) soppressione del rilascio di —> CRH>ACTH>Cortisone<>Cortisolo con consequenziale prevenzione di 2) aumento della fame/appetito con il procedere del regime ipocalorico 3) prevenzione del aumento del catabolismo muscolare 4) prevenzione di un aumento della ritenzione idrica e 5) prevenzione dell’alterazione del metabolismo lipidico correlato ad un incremento significativo del Cortisolo. Da notare che il momento della somministrazione del Glucocorticoide esogeno può influenzare il grado di soppressione surrenalica. Per esempio, il Prednisone in una dose di 5mg somministrato la sera prima di coricarsi e 2,5mg al mattino produrrà una soppressione dell’Asse HPA più marcata rispetto a 2,5mg la sera e 5mg al mattino. 5mg è un dosaggio basso e generalmente non è promotore di insonnia sebbene in soggetti sensibili può manifestarsi. Il dosaggio comunemente utilizzato varia da 15 a 25mg/die diviso in due dosi dopo i pasti. Tale pratica comunemente è parte di protocolli PEDs più complessi e contenenti uno o più agenti anabolizzanti.
Recenti resoconti sulla presunta potenza dei Glucocorticoidi sistemici provengono da atleti che hanno anche confessato l’uso concomitante di altri metodi e sostanze per migliorare le prestazioni, tra cui agenti anabolizzanti come il testosterone.[https://bjsm.bmj.com/] Tali regimi di Glucocorticoidi potrebbero avere rilevanza solo in un piccolo sottoinsieme di discipline sportive, come nelle ripide tappe di montagna dei Grandi Giri del ciclismo, dove gli atleti potrebbero essere disposti ad accettare compromessi nei loro regimi di allenamento o potenza assoluta in uscita nel perseguimento di un rapporto potenza/peso superiore. L’uso di OOC richiederebbe comunque una continuazione prolungata dell’uso di Glucocorticoidi nel periodo di gara per evitare l’insufficienza surrenalica dovuta a meccanismi di feedback. L’uso prolungato di Glucocorticoidi comporta rischi medici ben noti, alcuni dei quali potrebbero ridurre in modo permanente le prestazioni atletiche.[https://bjsm.bmj.com/]
Rischi per la salute, eventi avversi ed effetti negativi sulle prestazioni
Il trattamento con Glucocorticoidi per molte condizioni ha una lunga storia e un profilo di sicurezza ragionevole. Dosi elevate o uso cronico di Glucocorticoidi sistemici presentano un certo rischio per la salute dell’atleta. Un esame attento, una diagnosi e una deliberazione da parte del medico sono fondamentali e i benefici del trattamento devono essere soppesati rispetto ai potenziali rischi ed effetti avversi. L’uso potenziale per migliorare le prestazioni, descritto sopra e ritenuto limitato a contesti sportivi specifici con uso di GC ad alto dosaggio, è anche potenzialmente associato a rischi significativi per la salute di un atleta.
Gli eventi avversi con associazioni causali ben consolidate all’uso clinicamente appropriato di GC toccano praticamente ogni sistema umano, vanno da esiti negativi sulla salute acuti a cronici e includono insufficienza surrenalica, immunodeficienza, osteoporosi, atrofia muscolare, cedimento di tendini/fasce, necrosi avascolare della testa femorale, vari squilibri elettrolitici, nutrizionali e metabolici, glaucoma e cataratta. Forse perché i GC sono farmaci così comuni e clinicamente versatili, alcuni medici potrebbero sopravvalutare il loro valore terapeutico e sottostimare la gravità degli eventi avversi associati.[https://bjsm.bmj.com/] Anche una singola iniezione intra-articolare potrebbe causare un’insufficienza surrenalica clinicamente significativa che porta a malessere, squilibrio elettrolitico e immunosoppressione per diverse settimane.[https://bjsm.bmj.com/]
È importante sottolineare che l’eziologia di questi sintomi potrebbe non essere riconosciuta dall’atleta e dal personale medico, in particolare in un contesto sportivo in cui gli atleti si allenano ad alta intensità e i sintomi possono mascherarsi da affaticamento correlato al sovrallenamento. Inoltre, un atleta che subisce un trauma o un infortunio grave potrebbe essere a maggior rischio di crisi surrenalica a causa della soppressione ipotalamo-ipofisi-surrene dovuta al precedente utilizzo di GC. Ciò potrebbe essere particolarmente problematico se l’atleta non rivela questo precedente utilizzo.
Sia l’efficacia che il potenziale danno delle iniezioni intra-articolari sono ampiamente dibattuti. Le prove di un recente studio prospettico controllato con placebo su pazienti con osteoartrite hanno suggerito che frequenti iniezioni di triamcinolone al ginocchio, somministrate secondo un programma prestabilito, non sono riuscite a gestire efficacemente il dolore a lungo termine e hanno portato a una riduzione statisticamente significativa dello spessore della cartilagine.[https://bjsm.bmj.com/]Tuttavia, le raccomandazioni della società medica, così come una meta-analisi completa, supportano l’efficacia e la sicurezza dello stesso intervento,[https://bjsm.bmj.com/l] suggerendo fortemente che un uso giudizioso di iniezioni intra-articolari in pazienti e circostanze appropriate può produrre risultati positivi. Vi è una mancanza di prove pubblicate sulla sicurezza o il danno dell’uso di GC intra-articolari nelle popolazioni di atleti e sono urgentemente necessarie ulteriori ricerche a causa dell’uso onnipresente di GC intra-articolari.
Politiche per garantire l’uso appropriato dei GC
Nonostante le preoccupazioni di un possibile abuso per un vantaggio competitivo o potenziali effetti dannosi sulla salute degli atleti, i GC sono ampiamente utilizzati nello sport per legittime ragioni terapeutiche. Considerando che l’elenco è armonizzato in tutti gli sport, dal tiro con l’arco al wakeboard, il doping con i GC non è un problema laddove i presunti benefici dell’uso di GC ad alto dosaggio (potenza prolungata a intensità di esercizio submassimali o gestione aggressiva del peso catabolico) difficilmente miglioreranno le prestazioni. Pertanto, un AAF per i GC non sarebbe probabilmente associato a nessun intento di doping. L’uso di GC sistemici in molti sport deve essere considerato sotto una luce diversa rispetto agli sport ad alto rischio come il ciclismo, dove l’abuso è ben documentato e le prove scientifiche forniscono un certo supporto.
Consapevoli delle sfide specifiche poste dall’uso di GC nello sport, le organizzazioni sportive e antidoping hanno introdotto politiche innovative e stanno rafforzando le normative esistenti per affrontare l’uso terapeutico ragionevole dei GC.
Conclusioni su Glucocorticoidi e prestazioni sportive:
Da recenti review, sappiamo che l’uso dei Glucocorticoidi sistemici può migliorare la prestazione fisica massima rispetto al placebo (SDM 0,300, 95% CI 0,080-0,520). In una recente review [https://www.frontiersin.org/], l’SDM per i 13 confronti inclusi non era eterogeneo (I2 = 35%, p = 0,099). L’analisi di sensibilità escludendo i due studi con alto rischio di bias ha mostrato un effetto simile (SDM 0,349, 95% CI 0,071-0,626). Con la meta-regressione si è scoperto che la durata del trattamento, la via di somministrazione e il tipo di esercizio non hanno influenzato (p > 0,124) l’SDM. Nell’analisi stratificata il trattamento prolungato e l’ingestione orale hanno migliorato la prestazione fisica (p = 0,003). Il trattamento acuto e l’inalazione non hanno avuto alcun effetto sulla prestazione fisica (p > 0,564), l’analisi di sensibilità con studi ad alto rischio di bias rimossi o solo un trattamento per gruppo di controllo, ha mostrato un effetto simile all’analisi completa con SDM 0,334, 95% CI 0,075-0,592 e SDM 0,296 0,059-0,532, rispettivamente. L’analisi di sensibilità escludendo i sei confronti con meno di 10 coppie di dati non ha indicato alcun effetto dei Glucocorticoidi sulla prestazione fisica (p = 0,070). I Glucocorticoidi hanno migliorato la prestazione aerobica (SDM 0,348, 95% CI 0,129-0,567). Tre confronti hanno testato l’effetto dei Glucocorticoidi sulla prestazione anaerobica massima e la meta-analisi dei confronti non ha mostrato alcun effetto (p = 0,573) sulla prestazione fisica. L’effetto è rimasto non statisticamente significativo dopo aver incluso i due studi che misuravano la prestazione anaerobica all’interno di un test di prestazione aerobica (p = 0,491) e quando tutti i risultati della prestazione anaerobica negli studi inclusi (anche più risultati dello stesso studio) sono stati meta-analizzati (p = 0,177). Non è stato trovato alcun effetto dei Glucocorticoidi sulla spesa energetica durante la prestazione submassimale (SDM 0,-332, 95% CI −0,785 a 0,121).
In una review narrativa del 2016, Collomp et al. hanno mirato a riassumere le attuali conoscenze sugli effetti ergogenici dei glucocorticoidi negli esseri umani. Hanno riferito che un effetto ergogenico (sull’esercizio di endurance) dei Glucocorticoidi sistemici a breve termine è stato chiaramente dimostrato e che gli effetti a breve termine (4,5 giorni) e a lungo termine (4 settimane) dell’assunzione di Glucocorticoidi non hanno avuto alcun effetto sul VO2-max o sulla potenza massima in uscita durante protocolli di esercizio graduati. Hanno anche il test sul campo come categoria di test delle prestazioni, ma fanno riferimento solo allo studio di Casuso et al. che riporta un miglioramento delle prestazioni nella corsa a navetta, ma nessun cambiamento nelle prestazioni nello sprint. I risultati successivi supportano e rafforzano la conclusione di Collomp et al. ( per quanto riguarda l’esercizio di endurance poiché scopriamo anche che il trattamento prolungato con glucocorticoidi migliora le prestazioni aerobiche. Tuttavia, a differenza di Collomp et al. nelle ultime review sono stati inclusi sia test di esercizio graduati che test sul campo (della durata di 1 minuto o più) nella definizione di prestazione aerobica e quindi, più studi che hanno aggiunto potenza statistica alla analisi. Non è stato trovato alcun effetto dei Glucocorticoidi sulla prestazione anaerobica quando è stato analizzato secondo il protocollo di studio, tuttavia sono stati inclusi solo tre studi che testavano la prestazione anaerobica. Per aumentare la potenza statistica, è stato anche incluso la prestazione anaerobica all’interno di test aerobici e più test anaerobici dallo stesso studio, ma ancora non era evidente alcun effetto dei glucocorticoidi. Questo approccio può diminuire la validità dell’analisi poiché la prestazione anaerobica all’interno di test aerobici può testare altre abilità rispetto ai test anaerobici e la meta-analisi di più risultati dagli stessi soggetti e l’intervento non è raccomandato dal Cochrane Handbook. Collomp et al. concludono che non è chiaro se i Glucocorticoidi migliorino la prestazione durante l’esercizio breve-intenso. Questa conclusione è ancora valida poiché solo pochi studi hanno indagato l’effetto dei glucocorticoidi sull’esercizio anaerobico/breve intenso. Tuttavia, quando meta-analizziamo tutte le prove disponibili, sembra che i Glucocorticoidi non migliorino le prestazioni anaerobiche. I Glucocorticoidi non hanno influenzato le prestazioni submassimali aumentando l’energia totale spesa e/o il VO2max a un carico fisso, ma questa conclusione dovrebbe anche essere interferita con cationi poiché l’analisi include solo 35 soggetti, il che fornisce una potenza statistica limitata.
L’uso di Glucocorticoidi nello sport è una questione altamente complessa a causa del loro uso diffuso in medicina, delle numerose formulazioni e vie di somministrazione con farmacocinetica variabile, effetti negativi sulla salute e potenziali associazioni di doping.
In definitiva, da quanto recentemente emerso attraverso il riassunto delle migliori prove scientifiche disponibili, i Glucocorticoidi migliorano le prestazioni aerobiche e massime, ma non influenzano le prestazioni anaerobiche nei soggetti sani.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Pelt AC (2011). Glucocorticoids: effects, action mechanisms, and therapeutic uses. Hauppauge, N.Y.: Nova Science. ISBN978-1617287589.[page needed]
Ralph CR, Tilbrook AJ (February 2016). “INVITED REVIEW: The usefulness of measuring glucocorticoids for assessing animal welfare”. Journal of Animal Science. 94 (2): 457–470. doi:10.2527/jas.2015-9645. PMID27065116.
Collomp K, Arlettaz A, Buisson C, Lecoq AM, Mongongu C. Glucocorticoid administration in athletes: performance, metabolism and detection. Steroids. (2016) 115:193–202. doi: 10.1016/j.steroids.2016.09.008 PubMed Abstract | CrossRef Full Text | Google Scholar
Fain JN, Cheema P, Tichansky DS, Madan AK. Stimulation of human omental adipose tissue lipolysis by growth hormone plus dexamethasone. Mol Cell Endocrinol. (2008) 295(1):101–5. doi: 10.1016/j.mce.2008.05.014PubMed Abstract | CrossRef Full Text | Google Scholar
Thomasson R, Rieth N, Jollin L, Amiot V, Lasne F, Collomp K. Short-term glucocorticoid intake and metabolic responses during long-lasting exercise. Horm Metab Res. (2011) 43(3):216–22. doi: 10.1055/s-0030-1269919 PubMed Abstract | CrossRef Full Text | Google Scholar
Duclos M, Guinot M, Le Bouc Y, Cortisol GH. Odd and controversial ideas. Appl Physiol Nutr Metab. (2007) 32(5):895–903. doi: 10.1139/h07-064PubMed Abstract | CrossRef Full Text | Google Scholar
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. (2000) 21(1):55–89. doi: 10.1210/edrv.21.1.0389PubMed Abstract | CrossRef Full Text | Google Scholar
Duclos M. Evidence on ergogenic action of glucocorticoids as a doping agent risk. Phys Sportsmed. (2010) 38(3):121–7. doi: 10.3810/psm.2010.10.1817PubMed Abstract | CrossRef Full Text | Google Scholar
Strehl C, Bijlsma JWJ, de Wit M, Boers M, Caeyers N, Cutolo M, et al. Defining conditions where long-term glucocorticoid treatment has an acceptably low level of harm to facilitate implementation of existing recommendations: viewpoints from an EULAR task force. Ann Rheum Dis. (2016) 75(6):952–7. doi: 10.1136/annrheumdis-2015-20891PubMed Abstract | CrossRef Full Text | Google Scholar
Ventura R, Daley-Yates P, Mazzoni I, Collomp K, Saugy M, Buttgereit F, et al. A novel approach to improve detection of glucocorticoid doping in sport with new guidance for physicians prescribing for athletes. Br J Sports Med. (2021) 55(11):631–42. doi: 10.1136/bjsports-2020-103512
Zhou M, Gomez-Sanchez CE (July 1993). “Cloning and expression of a rat cytochrome P-450 11 beta-hydroxylase/aldosterone synthase (CYP11B2) cDNA variant”. Biochem. Biophys. Res. Commun. 194 (1): 112–7. doi:10.1006/bbrc.1993.1792. PMID8333830.
Lefebvre H, Cartier D, Duparc C, et al. (March 2002). “Characterization of serotonin(4) receptors in adrenocortical aldosterone-producing adenomas: in vivo and in vitro studies”. J. Clin. Endocrinol. Metab. 87 (3): 1211–6. doi:10.1210/jcem.87.3.8327. PMID11889190.
Lupien SJ, McEwen BS, Gunnar MR, Heim C (Jun 2009). “Effects of stress throughout the lifespan on the brain, behaviour and cognition”. Nature Reviews. Neuroscience. 10 (6): 434–445. doi:10.1038/nrn2639. PMID19401723. S2CID205504945.
Belanoff JK, Gross K, Yager A, Schatzberg AF (2001). “Corticosteroids and cognition”. Journal of Psychiatric Research. 35 (3): 127–145. doi:10.1016/S0022-3956(01)00018-8. PMID11461709.
Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE (Dec 2007). “The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition”. Brain and Cognition. 65 (3): 209–237. doi:10.1016/j.bandc.2007.02.007. PMID17466428. S2CID5778988.
Wolkowitz OM, Lupien SJ, Bigler ED (Jun 2007). “The ‘steroid dementia syndrome’: a possible model of human glucocorticoid neurotoxicity”. Neurocase. 13 (3): 189–200. doi:10.1080/13554790701475468. PMID17786779. S2CID39340010.
Liu C, Chen Y, Kang Y, Ni Z, Xiu H, Guan J, et al. (Oct 2011). “Glucocorticoids improve renal responsiveness to atrial natriuretic peptide by up-regulating natriuretic peptide receptor-A expression in the renal inner medullary collecting duct in decompensated heart failure”. The Journal of Pharmacology and Experimental Therapeutics. 339 (1): 203–209. doi:10.1124/jpet.111.184796. PMID21737535. S2CID1892149.
Newton R, Holden NS (Oct 2007). “Separating transrepression and transactivation: a distressing divorce for the glucocorticoid receptor?”. Molecular Pharmacology. 72 (4): 799–809. doi:10.1124/mol.107.038794. PMID17622575. S2CID52803631.
Rado JP, Blumenfeld G, Hammer S (Nov 1959). “The effect of prednisone and 6-methylprednisolone on mercurial diuresis in patients with refractory cardiac edema”. The American Journal of the Medical Sciences. 238 (5): 542–551. doi:10.1097/00000441-195911000-00003. PMID14435747. S2CID38687480.
Riemer AD (Apr 1958). “Application of the newer corticosteroids to augment diuresis in congestive heart failure”. The American Journal of Cardiology. 1 (4): 488–496. doi:10.1016/0002-9149(58)90120-6. PMID13520608.
Newman DA (Feb 1959). “Reversal of intractable cardiac edema with prednisone”. New York State Journal of Medicine. 59 (4): 625–633. PMID13632954.
Massari F, Mastropasqua F, Iacoviello M, Nuzzolese V, Torres D, Parrinello G (Mar 2012). “The glucocorticoid in acute decompensated heart failure: Dr Jekyll or Mr Hyde?”. The American Journal of Emergency Medicine. 30 (3): 517.e5–10. doi:10.1016/j.ajem.2011.01.023. PMID21406321.