
Introduzione alla questione “High-Low gainer/responder”:
I fattori alla base dell’eterogeneità dell’ipertrofia muscolare in seguito all’allenamento contro-resistenza (RET) rimangono in gran parte sconosciuti. E la questione ormonale connessa è senza dubbio una parte poco conosciuta e spesso fraintesa.
Tutti i frequentatori di sala pesi, improvvisati o meno, hanno sentito parlare almeno una volta di “High gainers/responders” e di “Low gainers/responders” in riferimento alla possibilità individuale di aumento dell’ipertrofia muscolo-scheletrica. In teoria, possiamo classificare 7 tipi di “gainers/responders” diversi:
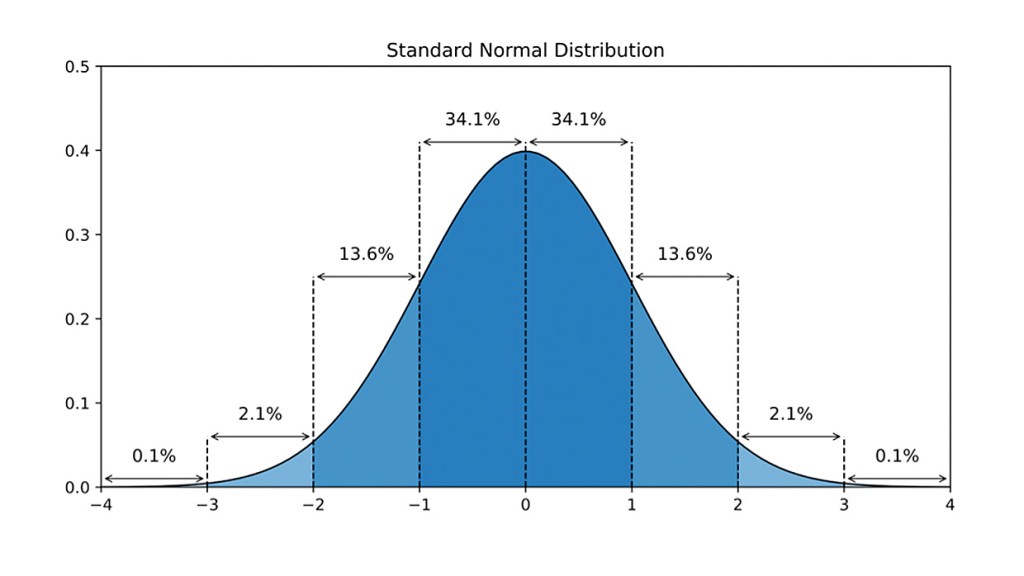
1.Non-Responders.
Coloro che guadagneranno una quantità insignificante di muscoli, anche se stanno facendo tutto correttamente. Queste persone sono estremamente rare e rappresentano circa lo 0,1% della popolazione.
2. Very Low Responders.
Coloro che sono in grado di aggiungere solo una piccola quantità di muscoli nel corso della loro carriera… e a un ritmo faticosamente lento. Anche loro non sono comuni: rappresentano circa il 2,1% della popolazione. È probabile che possano guadagnare solo 2.5-5Kg di muscoli durante la loro carriera di sollevatori (da 1.5 a 3Kg per le donne).
3. Low Responders.
Insieme ai non-responder e ai very low responders, questi soggetti completano la categoria dei veri hardgainer. Questi soggetti hanno guadagni muscolari molto lenti e di solito devono accettare di aggiungere una buona quantità di grasso per aumentare la massa muscolare. Sono i più comuni hardgainer, circa il 13,6% della popolazione. Possono guadagnare 5-10Kg di muscoli nel corso della loro carriera di sollevatori (3-6Kg per le donne).
4. Normal Responders.
È molto probabile che voi facciate parte di questa categoria. Questo gruppo rappresenta quasi il 70% della popolazione. Anche se ci saranno differenze nel potenziale di crescita muscolare all’interno di questo gruppo, tutti possono guadagnare una discreta quantità di muscoli se si allenano, mangiano e riposano correttamente. Gli uomini di questa categoria possono sperare di aumentare la massa muscolare tra i 10 e i 18Kg rispetto al loro peso da adulti senza allenamento. Le donne si avvicinano a 6-9Kg.
5. Easy Gainers.
Nel corso della loro carriera di sollevatori, questo gruppo può guadagnare il 15-20% di muscoli in più – 2.5-4Kg in più rispetto a un normale responder. Inoltre, possono aumentare più velocemente. Il loro potenziale di aumento muscolare potrebbe essere di 15-20Kg (8-11Kg per le donne). Rappresentano circa il 13,6% della popolazione.
6. Very Easy Gainers.
Questi soggetti spesso appaiono muscolosi già prima di iniziare ad allenarsi in sala pesi. E quando iniziano ad allenarsi, rispondono rapidamente e possono guadagnare un altro 10% di muscoli, per un potenziale di crescita muscolare totale di circa 16-24Kg (9-12Kg per le donne).
7. Freaks.
Questi soggetti sono sempre muscolosi e/o forti (e spesso esplosivi) prima ancora di mettere piede in palestra. Sono i “veri naturl” che, prima di iniziare ad assumere PEDs finiscono per assomigliare a chi ne fa già uso. Ma rappresentano lo 0,1% della popolazione, il che significa che la maggior parte degli “influencer” che affermano di avere una buona genetica e non di usare farmaci, stanno mentendo.
Nota: la quantità di potenziale di crescita muscolare può sembrare bassa, ma è chiaro che non sto parlando di peso corporeo. Ogni chilo di aumento muscolare porta normalmente a un aumento di 0,25-0,5 chili di “qualcos’altro” senza aggiungere grasso corporeo. Un aumento muscolare di 13.5Kg porterebbe in realtà a un aumento della massa magra da 16.8Kg a 20.4Kg sulla bilancia.
Ma quali sono le determinanti genetiche che separano una “High gainer/responder” da un “Low gainer/responder”?
In ordine di importanza teorica:
- GENOTIPO ACTN3
Senza entrare troppo nel merito, esistono due genotipi ACTN3 “puri”: ACTN3 RR e ACTN3 XX. Esistono anche tipi misti. Il tipo di ACTN3 determina diversi elementi che svolgono un ruolo importante nel potenziale di crescita muscolare.
Rapporto tra fibre a contrazione rapida e lenta. Un maggior numero di fibre a contrazione rapida significa un maggior potenziale di crescita e di forza.
Livello di attivazione del mTOR. Più si riesce ad attivare l’mTOR dopo l’allenamento e i pasti, più si aumenta la sintesi proteica e più si può crescere.
Riparazione del danno muscolare. Più lenta è la riparazione, meno ci si può allenare proficuamente e più è difficile far crescere nuovo tessuto contrattile.
Il tipo ACTN3 RR presenta un maggior numero di fibre a contrazione rapida, una maggiore attivazione del mTOR e una rapida riparazione del danno muscolare. Tutto ciò favorisce una crescita muscolare più rapida.
All’opposto, ACTN3 XX significa meno fibre a contrazione rapida, minore attivazione del mTOR e riparazione lenta dei danni muscolari. Ma hanno un VO2 max naturale più elevato e sono più resistenti all’affaticamento muscolare.
- Espressione della Miostatina
Sicuramente molti di voi avranno visto le foto degli esemplari di Belgian Blue, una razza di bovini da carne del Belgio la cui caratteristica peculiare sono le accentuate masse muscolari. Non si tratta di un esperimento in cui le mucche vengono sottoposte a dosi massicce di steroidi anabolizzanti, ma semplicemente di una razza di bovini nati senza la capacità di produrre Miostatina.
La Miostatina è una miochina (una proteina rilasciata dai muscoli). Agisce come un fattore limitante nella quantità di muscoli che si possono sviluppare. Più ci si avvicina al proprio potenziale genetico, più la Miostatina limiterà la crescita muscolare.
Alcune persone hanno naturalmente livelli di Miostatina più elevati, quindi il loro tasso di crescita muscolare totale risulta inferiori con un tasso catabolico più accentuato.
Le persone con meno Miostatina possono sviluppare più muscoli e più rapidamente. Sembra anche che siano più a rischio di strappi muscolari.
- Numero, densità e sensibilità recettoriale
Sebbene diversi fattori possano influenzare i livelli di Testosterone, IGF-1 e Ormone della Crescita (alimentazione, stress, sonno, ecc.), alcune persone hanno un numero e potenziale di espressività dei recettori ormone specifici (es. AR) maggiore rispetto alla norma. “Natural” o “Doped”, in entrambi i casi, le persone con questa caratteristica hanno un maggiore potenziale di crescita muscolare grazie a una maggiore sintesi proteica indotta dalla risposta ormone-recettoriale.
Questi articoli saranno però incentrati sull’analisi dei due fattori ormone-genici determinanti l’ipertrofia del muscolo-scheletrico: l’espressività recettoriale e la mutazione del gene della Miostatina.
In questa 1° parte tratterò dell’espressione recettoriale.
Introduzione al “Fattore Recettoriale”:
In un interessante studio del 2018 [1] Sono stati esaminati gli ormoni circolanti, gli ormoni intramuscolari e le variabili correlate agli ormoni intramuscolari in uomini allenati alla resistenza prima e dopo 12 settimane di RET. L’eliminazione a ritroso e la regressione delle componenti principali hanno valutato la significatività statistica degli ormoni anabolici circolanti proposti (ad esempio, Testosterone, Testosterone libero, Deidroepiandrosterone, Diidrotestosterone, Fattore di Crescita Insulino-Simile-1, Fattore di Crescita Insulino-Simile-1 libero, Ormone Luteinizzante e Ormone della Crescita) e i cambiamenti della massa muscolare indotti dalla RET (n = 49). Sono stati utilizzati immunoblots e immunodosaggi per valutare i livelli di Testosterone libero intramuscolare, i livelli di Diidrotestosterone, l’espressione della 5α-reduttasi e il contenuto del Recettore degli Androgeni nei soggetti che hanno risposto in modo più elevato (HIR; n = 10) e più basso (LOR; n = 10) alle 12 settimane di RET. Nessun ormone misurato prima dell’esercizio, dopo l’esercizio, prima dell’intervento o dopo l’intervento è risultato costantemente significativo o selezionato nel modello finale per la variazione di: area trasversale di tipo 1 (CSA), CSA di tipo 2 o massa grassa e ossea (LBM). L’analisi delle componenti principali non ha portato a una grande riduzione delle dimensioni e la regressione delle componenti principali non è stata più efficace delle analisi di regressione non aggiustate. Nessun ormone misurato nel sangue o nel muscolo è risultato diverso tra HIR e LOR. L’enzima steroidogenico 5α-reduttasi è aumentato dopo la RET nell’HIR (P < 0,01) ma non nel LOR (P = 0,32). Il contenuto di recettori per gli androgeni è rimasto invariato con la RET, ma è stato più elevato in ogni momento nell’HIR. A differenza del Testosterone libero intramuscolare, del Diidrotestosterone o della 5α-reduttasi, è stata riscontrata una relazione lineare tra il contenuto dei recettori degli androgeni e la variazione della LBM (P < 0,01), del CSA di tipo 1 (P < 0,05) e del CSA di tipo 2 (P < 0,01) sia prima che dopo l’intervento. Questi risultati indicano che il contenuto intramuscolare di recettori per gli androgeni, ma non gli ormoni circolanti o intramuscolari (o gli enzimi che ne regolano la produzione intramuscolare), influenzano l’ipertrofia del muscolo scheletrico dopo la RET in giovani uomini precedentemente allenati.
Variabili nell’ipertrofia indotta da RET e livelli ormonali:
Esiste una sostanziale variabilità individuale nell’ipertrofia muscolare scheletrica indotta da RET (Hubal et al., 2005; Davidsen et al., 2011). Si ritiene che l’aumento post-esercizio degli ormoni circolanti, presumibilmente anabolici (ad esempio, T, GH e IGF-1), sia causale nel determinare l’ipertrofia muscolare scheletrica indotta da RET (Kraemer et al., 2017; Mangine et al., 2017). Tuttavia, esistono sostanziali prove contrarie di un ruolo causale, o addirittura correlato (cioè che condivide una varianza comune) di tali ormoni sia nell’aumento della sintesi proteica muscolare indotto da RET (West et al., 2009) sia nell’ipertrofia (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016; Mobley et al., 2018).
È plausibile che, al contrario degli ormoni circolanti a livello sistemico, l’androgenesi locale intramuscolare possa mediare l’ipertrofia muscolare indotta da RET, come è stato proposto per gli uomini anziani (Sato et al., 2014). Inoltre, l’aumento del contenuto di recettori androgeni intramuscolari indotto da RET è stato significativamente correlato all’ipertrofia muscolare indotta da RET (Ahtiainen et al., 2011; Mitchell et al., 2013). Pertanto, è possibile che un aumento degli androgeni intramuscolari e/o dei loro recettori, attraverso un meccanismo autocrino, sia importante nel determinare l’ipertrofia indotta da RET.
Lo scopo dello studio di base trattato in questo articolo [1] è stata quella di determinare se l’eterogeneità dell’ipertrofia del muscolo scheletrico indotta da RET, misurata mediante indici multipli, fosse associata agli ormoni circolanti, agli ormoni intramuscolari, al contenuto di enzimi steroidogenici intramuscolari o al contenuto di recettori per gli androgeni. Sono stati eseguite ulteriori analisi statistiche e intramuscolari sui dati di uno studio precedente condotto su uomini sani e allenati contro-resistenza (n = 49; Morton et al., 2016). Per esplorare ulteriormente la relazione tra ormoni sistemici e ipertrofia, è stato utilizzato l’eliminazione a ritroso e la regressione delle componenti principali sulle concentrazioni di ormoni sistemici sia a riposo che dopo l’esercizio contro-resistenza con gli indici di ipertrofia come variabili di esito separate in tutti i partecipanti. Per valutare l’importanza dell’androgenesi intramuscolare, abbiamo completato un’analisi solo sui rispondenti più alti (HIR – quintile superiore) e più bassi (LOR – quintile inferiore) che comprendeva la valutazione del T intramuscolare, del DHT, dell’espressione della 5α-reduttasi e del contenuto del recettore degli androgeni. Coerentemente con il lavoro precedente (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016), i ricercatori hanno ipotizzato che gli ormoni sistemici circolanti non fossero correlati a nessuna misura di ipertrofia; tuttavia, hanno ipotizzato, visti i risultati precedenti (Ahtiainen et al., 2011; Mitchell et al., 2013), che il contenuto di recettori per gli androgeni fosse associato all’ipertrofia indotta da RET.
Partecipanti e intervento con allenamento contro-resistenza:
Quarantanove giovani uomini allenati alla resistenza (eseguendo RET almeno 2 giorni/settimana [range 3-6 giorni/settimana] per 4 ± 6 anni) si sono offerti volontari per questo studio. In breve, i partecipanti sono stati assegnati in modo casuale a un gruppo ad alte ripetizioni (HR) o a basse ripetizioni (LR). Il gruppo HR ha eseguito tutti gli esercizi con una resistenza relativamente leggera [∼30-50% del loro massimo di ripetizioni (RM)] fino al cedimento volitivo (20-25 ripetizioni) e il gruppo LR ha eseguito tutti gli esercizi con una resistenza relativamente pesante (∼75-90% RM), anch’essi fino al cedimento volitivo (8-12 ripetizioni). Ogni partecipante è stato sottoposto a un intervento RET di 12 settimane in cui ha eseguito RET su tutto il corpo per 4 giorni a settimana e ha ricevuto 30g di proteine isolate del siero di latte due volte al giorno (BioPRO; Davisco Foods International, Le Sueur, MN, Stati Uniti).
Prelievo di sangue e analisi ormonali:
Il giorno del test pre e post intervento è stato eseguito dopo un digiuno notturno alla stessa ora del giorno per ogni partecipante. Ogni partecipante ha eseguito un allenamento acuto contro-resistenza nell’ambito del gruppo designato (HR o LR) e il sangue è stato prelevato da un catetere endovenoso inserito in una vena antecubitale. Due provette vacutainer da 4 ml (Becton, Dickinson and Company, Franklin Lakes, NJ, Stati Uniti) sono state prelevate prima dell’esercizio e dopo 0, 15, 30 e 60 minuti dall’esercizio. Una provetta da 4 mL è stata lasciata coagulare per 30 minuti a temperatura ambiente per isolare successivamente il siero e l’altra è stata eparinizzata per isolare successivamente il plasma. L’analisi del campione di sangue è stata eseguita in cieco per: Cortisolo (nM), LH (IU/L), Lattato (mM), DHEA (ng/mL), T (ng/mL), T libero (fT; ng/dL; cioè, Testosterone non legato alla globulina legante gli ormoni sessuali o all’albumina nel sangue), DHT (ng/mL) e GH (ng/mL) utilizzando test immunometrici a chemiluminescenza in fase solida a due siti (Immulite 2000 Immunoassay System; Siemens Healthineers, Erlangen, Germania) e IGF-1 (μg/dL) e IGF-1 libero (fIGF-1; ng/mL) utilizzando radio-immunoassaggi (Diagnostics Products Corporation, Los Angeles, CA, Stati Uniti). L’AUC a 60 minuti dall’esercizio di resistenza è stata calcolata per ciascun ormone, utilizzando la regola trapezoidale, con punti temporali a 0, 15, 30 e 60 minuti.
Regressioni a scalare:
I dati di HR e LR sono stati eliminati a causa della mancanza di differenze tra i gruppi per quanto riguarda gli ormoni circolanti e gli esiti (Morton et al., 2016). Gli esiti considerati sono stati CSA delle fibre di tipo 1, CSA delle fibre di tipo 2 e massa corporea (LBM) priva di grasso e ossa. Ciascun risultato in ciascun momento della misurazione (ossia, la variazione, i valori assoluti prima e dopo l’intervento) è stato regredito rispetto agli ormoni di ciascun punto temporale: AUC pre-intervento a riposo, AUC post-esercizio pre-intervento, AUC post-intervento a riposo e AUC post-esercizio post-intervento. Per scegliere il modello finale è stata utilizzata l’eliminazione a ritroso, con il criterio di eliminazione Akaike Information Criterion (AIC). I valori di AUC post-esercizio utilizzati nell’analisi non hanno sottratto le concentrazioni a riposo. Tuttavia, abbiamo eseguito l’analisi con le concentrazioni a riposo sottratte dai valori grezzi dell’AUC e non abbiamo riscontrato differenze sostanziali nei risultati.
Analisi immunoblot:
Come descritto in precedenza (Aizawa et al., 2010), dopo l’omogenizzazione, la concentrazione proteica del surnatante risultante è stata determinata mediante un saggio proteico di Bradford e le proteine muscolari (sia citoplasmatiche che nucleari, 20μg di proteine) sono state separate su gel di SDS-poliacrilammide al 10% e poi trasferite su membrane di polivinilidene difluoruro (Millipore, Billerica, MA, Stati Uniti). Le membrane sono state bloccate per 1 ora con tampone bloccante (5% latte scremato in soluzione salina tamponata con fosfato e 0,1% Tween 20) e quindi incubate per 12 ore a 4°C con anticorpi primari contro il recettore degli androgeni (#3202, Cell Signaling Technology, Beverly, MA, Stati Uniti) e la 5α-reduttasi (H00006715, Abnova, Taipei, Taiwan) diluiti a 1:1000 in tampone bloccante. Le membrane sono state lavate tre volte con PBST prima di essere incubate per 1 ora con un anticorpo secondario coniugato con perossidasi di rafano e immunoglobulina anti-rabbit (#7074, Cell Signaling Technology, Beverly, MA, Stati Uniti) diluito a 1:3000 nel tampone di blocco. Le membrane sono state poi lavate tre volte con PBST. Le proteine sono state rilevate con un sistema di chemiluminescenza potenziata plus (GE Healthcare Biosciences) e visualizzate su un imager LAS4000 (GE Healthcare Biosciences). L’intensità delle bande è stata quantificata utilizzando ImageJ versione 1.46 (National Institutes of Health, Bethesda, MD, Stati Uniti).
Test immunoenzimatici per gli ormoni intramuscolari:
Il campione di muscolo è stato omogeneizzato con lo stesso metodo dell’analisi immunoblot. I livelli di T e DHT nel muscolo scheletrico sono stati determinati utilizzando un kit per il dosaggio immunoenzimatico, dopo essere stati diluiti 200 volte con ciascun tampone di dosaggio come precedentemente descritto (Horii et al., 2016). Gli anticorpi policlonali immobilizzati sono stati sollevati contro il T (Cayman Chemical, Ann Arbor, MI, Stati Uniti) e il DHT (IBL Hamburg, Germania) prima dell’aggiunta di anticorpi secondari alla perossidasi di rafano. La densità ottica a 450 nm è stata qualificata su un lettore di micropiastre (BioLumin 960; Molecular Dynamics, Tokyo, Giappone) e le analisi sono state eseguite in duplicato. Il valore del coefficiente di variazione era 3,0 e r2 = 0,974 nel presente studio. I ricercatori che hanno eseguito le analisi intramuscolari (KS e SF) non erano ciechi rispetto ai campioni HIR e LOR.
Analisi delle componenti principali e regressione:
I dati sono stati centrati e scalati prima di eseguire l’analisi delle componenti principali (PCA) sugli ormoni di ciascun momento della misurazione (riposo pre-intervento, AUC post-esercizio pre-intervento, riposo post-intervento e AUC post-esercizio post-intervento). Lo scopo della PCA è quello di utilizzare la trasformazione ortogonale per creare un insieme di nuove variabili lineari e non correlate (componenti principali), di cui viene preso un sottoinsieme che rappresenta effettivamente la maggior parte della variabilità osservata nei dati originali. In definitiva, queste componenti principali sono combinazioni lineari delle variabili originali (ad esempio, gli ormoni) che vengono poi utilizzate come covariate nelle analisi di regressione. Presentiamo qui la PCA sotto forma di scree plot. L’eliminazione a ritroso è stata eseguita sulle componenti principali (cioè la regressione delle componenti principali) utilizzando l’AIC come criterio di adattamento del modello. La PCA e la regressione delle componenti principali sono state eseguite in R (R Core Team, 2017).
High- vs. Low-Responders:
Le biopsie del muscolo scheletrico del vasto laterale di ciascun partecipante e la DXA sono state utilizzate per valutare la variazione della CSA delle fibre (sia di tipo 1 che di tipo 2) e della LBM, rispettivamente, come descritto in dettaglio altrove (Morton et al., 2016). La determinazione dell’HIR e del LOR è stata effettuata classificando individualmente (da 1 a 49) la variazione di ciascun risultato per ogni partecipante e quindi calcolando la media della classifica di ciascun partecipante per tutti e tre i risultati (CSA di tipo 1, CSA di tipo 2 e LBM). Con una probabilità di errore di tipo II (alfa) di 0,05, una probabilità di errore di tipo I (beta) di 0,20 e una differenza attesa relativamente moderata nelle variazioni della massa muscolare indotte dalla RET tra HIR e LOR (dimensione dell’effetto, f = 0,60), il calcolo delle dimensioni del campione a priori richiedeva 18 partecipanti (nove in ciascun gruppo). Pertanto, il quintile superiore (n = 10) dei partecipanti classificati è stato classificato come HIR e il quintile inferiore (n = 10) dei partecipanti classificati è stato classificato come LOR. Le analisi statistiche tra HIR e LOR sono state eseguite utilizzando SPSS (versione 22.0, Chicago, IL, Stati Uniti). Il CSA di tipo 1, il CSA di tipo 2, la LBM e tutti i dati relativi agli ormoni intramuscolari sono stati analizzati utilizzando un’analisi della varianza a due fattori (gruppo × tempo) a misure ripetute (ANOVA) con il gruppo (HIR vs. LOR) e il tempo (pre- e post-intervento) come variabili sperimentali. Se indicato, sono stati eseguiti t-test indipendenti a due code per valutare eventuali differenze tra i gruppi in uno specifico punto temporale (ad esempio, la T intramuscolare pre-intervento). Le correlazioni tra i risultati intramuscolari e la variazione di CSA di tipo 1, CSA di tipo 2 e LBM sono state eseguite in SPSS (versione 22.0, Chicago, IL, Stati Uniti). La significatività statistica è stata accettata quando P < 0,05. I dati sono presentati come diagrammi a riquadro e a baffi (comprendenti la mediana [linea], la media [croce], l’intervallo interquartile [riquadro] e i valori minimo e massimo [baffi]) nelle Figure 1 e 3 e media ± SD nel testo e nelle tabelle.

- Risultati dello studio
Cambiamenti nella massa muscolare con allenamenti contro-resistenza:
Sono stati reclutati 56 partecipanti e 49 hanno completato l’intero intervento (HR: n = 24, LR: n = 25; 23 ± 2 anni, 86 ± 5 kg, 181 ± 6 cm). Due persone hanno abbandonato il gruppo LR a causa di un trasferimento di lavoro e di un infortunio non legato all’intervento, mentre cinque persone hanno abbandonato il gruppo HR a causa di un cambiamento di sede o di un infortunio non legato all’intervento. Dodici settimane di RET hanno portato a un aumento della CSA di tipo 1 (665 ± 149 μm2), della CSA di tipo 2 (978 ± 189 μm2) e della LBM (1,22 ± 1,37 kg, P < 0,01; Figure 1A-C, rispettivamente; Morton et al., 2016). Non sono state riscontrate differenze tra i gruppi di ripetizioni (HR contro LR – vedi Morton et al., 2016) per nessuno dei risultati.
Regressioni a scalare:
Per ciascun risultato (variazione di CSA di tipo 1, CSA di tipo 2 e LBM) nessuna delle AUC post-esercizio (Tabella 1) o delle concentrazioni a riposo (Tabella 2) di qualsiasi ormone misurato prima o dopo l’intervento è risultata costantemente significativa (cioè significativa con più risultati o in più momenti di misurazione) nei modelli finali. Inoltre, i valori dei coefficienti di determinazione (cioè R2) erano bassi (<0,25) per tutti gli esiti in ogni momento della misurazione, indicando che la variazione osservata nella risposta ipertrofica può essere spiegata solo in minima parte da qualsiasi modello adattato. Risultati simili sono stati riscontrati valutando il CSA di tipo 1, il CSA di tipo 2 e la LBM prima e dopo l’intervento rispetto alle concentrazioni ormonali a riposo (Tabella supplementare 1).


Analisi delle componenti principali:
L’analisi delle componenti principali è stata eseguita su predittori centrati e scalari ed è qui presentata (vedi figure) come scree plot per l’AUC post-esercizio pre-intervento, l’AUC post-esercizio post-intervento, le concentrazioni a riposo pre-intervento e le concentrazioni a riposo post-intervento. Come illustrato dagli scree plot a bassa pendenza, nessuna componente principale è risultata particolarmente efficace nello spiegare la varianza nel set di dati originale.

I ricercatori hanno scelto di mantenere il numero di componenti principali che spiegano ≥80% della varianza dei predittori originali, il che ha portato a sette componenti principali incluse in ciascuna delle regressioni a scalare a componenti principali. L’esecuzione della regressione a componenti principali (indipendentemente dal fatto che gli ormoni siano stati valutati a riposo, dopo l’esercizio, prima dell’intervento o dopo l’intervento) ha rivelato che nessuna componente principale è stata costantemente significativa o inclusa in nessuno dei modelli finali e che l’R2 finale non ha mai superato lo 0,25 ed è stato inferiore allo 0,05. Questi risultati indicano che la variazione osservata nella risposta ipertrofica alla RET può essere spiegata solo in minima parte da uno qualsiasi dei modelli adattati.
High- vs. Low-Responders:
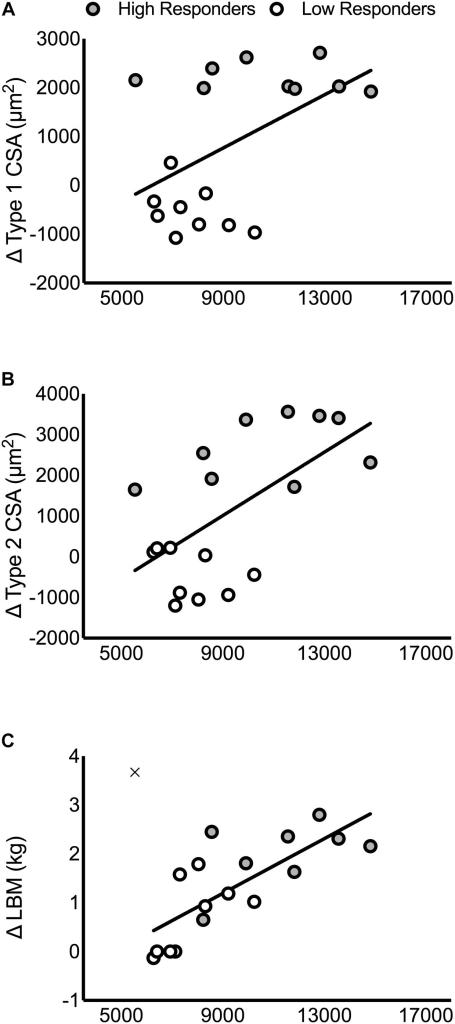
C’è stata una differenza significativa tra HIR e LOR nella variazione della CSA di tipo 1 (HIR: 2106 ± 412, LOR: -520 ± 450 μm2), della CSA di tipo 2 (HIR: 2642 ± 756, LOR: -373 ± 593 μm2) e della LBM (HIR: 2,1 ± 0,8, LOR: 0,6 ± 0,8 kg, P ≤ 0,001; Figure 1D-F). Non c’erano differenze nel numero di partecipanti di ciascun gruppo di allenamento (HIR: quattro e sei e LOR: sei e quattro da HR e LR, rispettivamente).
Non vi è stata alcuna differenza nella concentrazione ormonale a riposo tra HIR e LOR, ad eccezione della concentrazione a riposo post-intervento di LH (HIR: 3,67 ± 0,63; LOR 4,59 ± 1,15 UI/L, P < 0,01) e lattato (HIR: 0,52 ± 0,05; LOR: 0,55 ± 0,07 mM, P = 0,02), che erano maggiori nel LOR. Non c’è stata differenza nell’AUC post-esercizio per nessun ormone tra HIR e LOR, ad eccezione del cortisolo pre-intervento post-esercizio, che era più alto nell’HIR (HIR: 576 ± 100; LOR: 508 ± 199 nM; P < 0,001).
Ormoni intramuscolari:
Non sono state riscontrate differenze nei valori pre-intervento, post-intervento o nella variazione di T o DHT intramuscolare tra HIR e LOR (Figure 3A,B, rispettivamente). Il cambiamento nell’espressione della 5α-reduttasi è stato significativo nell’HIR (pre: 1457 ± 450, post: 1957 ± 543 AU, P < 0,01) ma non nel LOR (pre: 1748 ± 559, post: 1994 ± 840 AU, P = 0,32; Figura 3C). Il contenuto di recettori per gli androgeni intramuscolari prima dell’intervento (HIR: 10827 ± 2789, LOR: 7759 ± 1323 AU, P < 0,01) e dopo l’intervento (HIR: 11406 ± 2789, LOR: 7801 ± 1189 AU, P = 0,01; Figura 3D) era significativamente maggiore in HIR rispetto a LOR. Non c’è stato alcun cambiamento nel contenuto dei recettori degli androgeni intramuscolari prima e dopo l’intervento (Δ319 ± 1314 AU, P = 0,75) e c’è stata una relazione lineare tra il contenuto dei recettori degli androgeni dei partecipanti prima e dopo l’intervento (r = 0,92). Non sono state riscontrate correlazioni significative tra il T, il DHT o la 5α-reduttasi intramuscolare prima dell’intervento, dopo l’intervento e la variazione della CSA di tipo 1, della CSA di tipo 2 o della LBM (P > 0,05; Tabella supplementare 5). Al contrario, il contenuto di recettori per gli androgeni prima dell’intervento, dopo l’intervento e la media tra il contenuto di recettori per gli androgeni prima e dopo l’intervento sono stati significativamente correlati con la variazione della LBM (pre: r = 0,76, P < 0,01; post: r = 0,75, P < 0,01; media: r = 0. 77, P < 0,01), CSA di tipo 1 (pre: r = 0,51, P = 0,03; post: r = 0,49, P = 0,04; media: r = 0,51, P = 0,03) e CSA di tipo 2 (pre: r = 0,61, P < 0,01; post: r = 0,65, P < 0,01; media: r = 0,64, P < 0,01; Tabella supplementare 5 e Figura 4). I dati di un partecipante sono stati rimossi dalle analisi di regressione che includevano la variazione della LBM perché identificati come outlier statistici attraverso il metodo di regressione robusta e rimozione degli outlier con un coefficiente dell’1% (Motulsky e Brown, 2006). La posizione di questo partecipante è stata indicata nella Figura seguente a scopo illustrativo.
Punto della situazione:
Il risultato principale del presente studio, coerente con il lavoro precedentemente svolto dai ricercatori, è che nessun ormone sistemico condivide una varianza significativa con i cambiamenti indotti da RET nella CSA delle fibre muscolari scheletriche o nella massa muscolare scheletrica negli uomini allenati contro-resistenza. Sono stati estesi questi risultati alle concentrazioni ormonali locali misurate nel muscolo, che non hanno mostrato un’associazione significativa con alcun indice di ipertrofia. E’ stato riscontrato che gli HIR presentavano un aumento del contenuto di 5α-reduttasi dopo 12 settimane di RET e un contenuto di recettori degli androgeni significativamente più alto, che non cambiava con la RET, rispetto ai LOR sia prima che dopo la RET. La conclusione di ciò è che né la disponibilità sistemica né quella locale muscolare di ormoni influenzano l’ipertrofia muscolo-scheletrica indotta dalla RET in giovani uomini sani. Coerentemente con i lavori precedenti, i ricercatori propongono invece che l’entità dell’ipertrofia muscolare scheletrica indotta da RET sia modulata in parte dal contenuto intramuscolare di recettori per gli androgeni e probabilmente da altre variabili intramuscolari.
Ormoni circolanti e allenamento contro-resistenza:
Recenti pubblicazioni (Kraemer et al., 2017; Mangine et al., 2017) e linee guida (Ratamess et al., 2009) sostengono che gli ormoni circolanti sono meccanicamente e direttamente correlati e predittivi dei cambiamenti della massa muscolare scheletrica indotti dal RET, nonostante l’esistenza di prove che dimostrano il contrario (West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016; Mobley et al., 2018). In uno studio precedente a quello attualmente trattato, i ricercatori hanno eseguito 120 correlazioni, ciascuna su 49 partecipanti, tra 10 diversi ormoni e varie misure di variazione della massa e della forza muscolare. Hanno riscontrato che solo l’aumento del Cortisolo dopo l’esercizio era correlato con le variazioni del CSA di tipo 2 (pre-intervento: r = -0,34, P = 0,02; post-intervento: r = -0,31, P = 0,04) (Morton et al., 2016). Altri hanno trovato correlazioni significative tra l’aumento post-esercizio del GH circolante (McCall et al., 1999) e del T (Ahtiainen et al., 2003; Brook et al., 2016) con le variazioni della massa muscolare, ma queste correlazioni sono state eseguite su campioni composti da meno di 11 partecipanti, che potrebbero dare origine a correlazioni spurie. Qui sono state eseguite altre 48 regressioni graduali su 49 partecipanti, 10 ormoni e tre risultati distinti legati all’ipertrofia, tra cui la dimensione delle fibre muscolari. E’ stato riscontrato che nessun ormone era costantemente significativo, né alcun modello finale aveva un elevato coefficiente di determinazione, cioè tutti i valori di R2 erano inferiori a 0,25. Inoltre, la PCA non era efficace nel determinare le correlazioni con l’ipertrofia. Inoltre, la PCA non è stata efficace nel ridurre la varianza totale dei dati ormonali originali e non c’è stato alcun modello di regressione con le componenti principali utilizzate come covariate che spiegasse una proporzione significativa della variabilità in qualsiasi risultato. Esistono oggi prove sostanziali che suggeriscono che gli ormoni sistemici circolanti misurati a riposo (McCall et al., 1999; Morton et al., 2016; Mobley et al., 2018) e/o dopo l’esercizio (Ahtiainen et al., 2003; West et al., 2010; West e Phillips, 2012; Mitchell et al., 2013; Morton et al., 2016) non condividono alcuna varianza comune e non sono quindi né correlati né predittivi dei cambiamenti della massa muscolare indotti dal RET in giovani partecipanti sani.
Uno studio (Mangine et al., 2017) ha utilizzato un modello di equazione strutturale ai minimi quadrati parziali (PLS-SEM) e ha riportato che un modello con punteggi ormonali compositi (T, GH, IGF-1, insulina e cortisolo) e una misura composita di ipertrofia (CSA e spessore muscolare del vasto laterale e del retto femorale) ha prodotto un coefficiente di determinazione significativo (R2= 0,73). L’interpretazione di questo risultato è che il punteggio ormonale composito era correlato a un punteggio composito di ipertrofia. L’aspetto preoccupante di questa interpretazione è che il modello senza T (il miglior predittore ormonale del modello) aveva ancora un coefficiente di determinazione sostanziale (R2 = 0,43) con il punteggio composito di ipertrofia ed era statisticamente significativo. In effetti, la rimozione individuale degli altri ormoni (GH, IGF-1, insulina e cortisolo) ha mostrato un effetto trascurabile sulla varianza condivisa del modello, eppure il modello senza il suo “migliore” ormone predittivo, il T, ha rappresentato quasi il 60% della varianza osservata con tale ormone presente nel modello. Mentre gli autori sostengono che le interazioni inspiegabili tra gli ormoni siano la ragione della varianza del modello senza T, è stato suggerito che è più probabile che i pesi PLS capitalizzino il caso per esagerare le correlazioni (Goodhue et al., 2012). Sebbene riteniamo che il PLS-SEM sia utile per l’esame di grandi insiemi di dati, vi sono limitazioni sostanziali all’interpretazione quando si utilizzano campioni di piccole dimensioni (n = 26) (Goodhue et al., 2012). La definizione di PLS come metodo SEM appropriato è stata messa in discussione anche per quanto riguarda la stima e l’inferenza (Rönkkö e Evermann, 2013) e il coefficiente di determinazione (ad esempio, R2) è un parametro inadeguato per valutare l’adattamento del modello PLS-SEM, poiché stimatori incoerenti possono produrre modelli con R2 elevato. Di conseguenza, non tutti i modelli ben adattati sono predittivi (Henseler et al., 2014) e non tutti i modelli predittivi sono ben adattati (McIntosh et al., 2014).
High- vs. Low-Responders e allenamento contro-resistenza:
Per indagare sui potenziali determinanti dell’eterogeneità dell’ipertrofia muscolare scheletrica indotta da RET (Hubal et al., 2005; Davidsen et al., 2011; Morton et al., 2016), i ricercatori hanno suddiviso 49 partecipanti in HIR (n = 10) e LOR (n = 10) in base alla variazione di tre indicatori della massa muscolare scheletrica (CSA di tipo 1, CSA di tipo 2 e LBM). Nonostante le grandi differenze tra i gruppi in ogni risultato, non ci sono state differenze significative in nessun ormone circolante prima o dopo l’esercizio fisico, misurato sia prima che dopo l’intervento. Considerando che gli ormoni steroidei sono liposolubili (ad esempio, si diffondono attraverso il sarcolemma in base al loro gradiente di concentrazione), non sorprende che anche il T e il DHT intramuscolari misurati prima e dopo l’intervento non fossero diversi tra HIR e LOR. La mancanza di differenze negli ormoni circolanti e intramuscolari tra HIR e LOR dimostra che né l’apporto di ormoni al muscolo né il trasferimento di ormoni steroidei all’interno del muscolo sono fasi limitanti in individui giovani e sani.
Il contenuto di recettori per gli androgeni era significativamente più alto sia prima che dopo l’intervento nell’HIR rispetto al LOR ed era correlato alle variazioni della massa muscolare. Sebbene un altro gruppo non abbia riscontrato alcuna differenza nel contenuto di recettori degli androgeni tra HIR e LOR (Mobley et al., 2018), è importante riconoscere le differenze nel disegno dello studio (ad esempio, partecipanti non allenati rispetto a quelli allenati) e nelle misure di esito (ad esempio, analisi dei cluster basata sullo spessore muscolare rispetto a un punteggio aggregato di DXA e CSA delle fibre) tra loro e il lavoro dei ricercatori, rispettivamente. La funzione del recettore degli androgeni è quella di traslocare nel nucleo e modificare l’espressione dei geni bersaglio [rivisto altrove (Beato e Klug, 2000)], molti dei quali sono bersagli noti coinvolti nella crescita e nello sviluppo del muscolo scheletrico (Wyce et al., 2010). Infatti, quando i recettori degli androgeni vengono eliminati nei topi maschi, si verifica una significativa riduzione della massa e della forza muscolare (MacLean et al., 2008). È importante notare che la maggior parte degli ormoni steroidei ha un’elevata affinità con i propri recettori steroidei. Ad esempio, la costante di dissociazione del recettore degli androgeni nei confronti del T e del DHT è di soli ∼0,2-0,5 nM (Wilson e French, 1976). Nel presente studio, a riposo, la molarità del T sierico (HIR: 28 ± 7; LOR: 31 ± 7 nM), della fT sierica (HIR: 0,5 ± 0,01; LOR: 0,5 ± 0,01 nM) e del DHT sierico (HIR e LOR: 0,7 ± 0,2 nM) superavano tutti 0,2-0,5 nM. Dato che non c’era alcuna differenza negli ormoni circolanti o intramuscolari tra HIR e LOR, insieme all’elevata affinità di legame tra androgeno e recettore degli androgeni, sembra probabile che sia a riposo che dopo l’esercizio i recettori androgeni esistenti siano stati saturati nel muscolo scheletrico. Si ipotizza che, sebbene l’apporto di androgeni possa essere un passo limitante per l’ipertrofia muscolare indotta da RET negli uomini ipogonadici (Bhasin et al., 1997; Kvorning et al., 2013), il contenuto di recettori per gli androgeni sia la variabile più importante nell’accrescimento di proteine del muscolo scheletrico mediato dagli androgeni indotti da RET negli uomini sani (Diver et al., 2003).
Limitazioni:
I ricercatori hanno eseguito 120 correlazioni in uno studio precedente (Morton et al., 2016) e 48 regressioni graduali in questo caso (24 sui dati originali e 24 sulle componenti principali). L’applicazione di analisi multiple sugli stessi dati è stata un’operazione di data mining intenzionale per dimostrare la mancanza di capacità degli ormoni circolanti e intramuscolari a riposo o dopo l’esercizio fisico di prevedere le variazioni della massa muscolare scheletrica al basale o indotte dalla RET. Avrebbero potuto eseguire ulteriori statistiche per tenere conto dei test multipli, ma questo non sarebbe stato informativo perché nessuno dei loro modelli spiegava molta varianza (come valutato dai valori di R2, che non superavano lo 0,25). Riconoscono inoltre che, pur avendo incluso un campione di grandi dimensioni (n = 49) per l’analisi degli ormoni sistemici, essi si sono limitati a un campione relativamente più piccolo (n = 20) per il confronto tra HIR e LOR. Ammettono pienamente che, nel caso della correlazione con il recettore degli androgeni, quella che presentano è una stima gonfiata a causa della scelta di misurare solo i soggetti con risposta più alta e più bassa al loro protocollo di allenamento. Hanno condotto la loro analisi in questo modo per illustrare la differenza nell’ipertrofia muscolare indotta da RET e per indagare l’influenza delle variabili ormonali circolanti e intramuscolari su due gruppi distinti. Sebbene fossero limitati dalla quantità di tessuto raccolto, è giusto criticare il fatto che la loro analisi correlazionale sarebbe stata più eloquente se avessero incluso tutti i partecipanti e se avessero eseguito analisi aggiuntive [ad esempio, frazioni nucleari e citoplasmatiche del contenuto di recettori degli androgeni e espressioni geniche multiple (Cheung et al., 2017)]. Per questo motivo, il lavoro futuro potrà concentrarsi sulla biologia specifica che regola la regolazione e la funzione del recettore degli androgeni. Altri hanno ipotizzato che l’analisi con spettrometria di massa (rispetto ai test immunologici) sia necessaria per rilevare piccole concentrazioni intramuscolari di ormoni steroidei (Handelsman e Wartofsky, 2013); tuttavia, l’intento dei ricercatori era quello di analizzare i loro campioni utilizzando metodi simili a quelli che altri hanno utilizzato nella scienza dell’esercizio fisico, che possono essere diversi da quelli dell’endocrinologia clinica. Riconoscono che l’uso della DXA per misurare i cambiamenti nella LBM non è il gold standard, motivo per cui hanno scelto di includere anche i cambiamenti nella CSA delle fibre di tipo 1 e 2 per determinare i loro HIR e LOR (Buckinx et al., 2018). Per quanto riguarda la loro interpretazione, è ingenuo suggerire che la segnalazione degli androgeni sia esclusivamente operativa attraverso la loro tendenza a legarsi a un recettore androgenico [rivisto altrove (Herbst e Bhasin, 2004; Dubois et al., 2012)]. Sebbene la regolazione trascrizionale (ad esempio, la segnalazione dei recettori degli androgeni) sia qui evidenziata come un potente modulatore dei cambiamenti nella massa muscolare indotti da RET, è anche chiaro che la regolazione post-trascrizionale è almeno altrettanto importante per la sintesi proteica (Schwanhausser et al, 2011), come è stato evidenziato da recenti risultati (Figueiredo et al., 2015; Robinson et al., 2017; Mobley et al., 2018) e review (Chaillou et al., 2014; McGlory et al., 2017). Infine, sebbene vi sia un’influenza genetica alla base dell’ipertrofia muscolare scheletrica indotta da RET, vi sono ancora molte considerazioni ambientali, ad esempio il consumo di proteine alimentari adeguate (Morton et al., 2017), un apporto calorico e stimolo allenante adeguato che modulano l’ipertrofia muscolare indotta da RET.
Riflessioni conclusive sul presente studio:
Ricapitolando, i ricercatori hanno eseguito l’eliminazione a ritroso e la regressione delle componenti principali su una coorte relativamente ampia (n = 49) di uomini allenati contro-resistenza, concludendo che l’AUC post-esercizio (cioè l’esposizione ormonale netta transitoria acuta) e le concentrazioni ormonali a riposo misurate nel sangue non condividono una varianza comune con le variazioni della massa muscolare indotte dalla RET. In altre parole, le concentrazioni ormonali sistemiche non sono correlate o in qualche modo predittive delle variazioni della massa muscolare indotte da RET. L’analisi dei sottoinsiemi dei soggetti con risposta più alta e più bassa ha rivelato che il contenuto di recettori per gli androgeni, e non i livelli di androgeni intramuscolari, non cambia con il RET nei partecipanti allenati, ma è significativamente più alto negli HIR rispetto ai LOR. Questo studio, insieme ad altri (Bamman et al., 2007; Petrella et al., 2008; Davidsen et al., 2011; Eynon et al., 2013), fornisce la prova che l’aumento relativo della massa muscolare scheletrica in seguito alla RET è sostenuto da fattori locali intramuscolari e non da concentrazioni ormonali sistemiche.
Questo è quanto suggerito dall’osservazione di soggetti in stato fisiologico. Individui trattati con dosi esogene sovrafisiologiche di AAS sarebbero teoricamente soggetti alle medesime limitazioni presenti nel confronto tra HIR e LOR dello studio. Questa limitazione sembra essere data dall’espressione dei AR (Recettori degli Androgeni) nel muscolo scheletrico. Sebbene dosi sovrafisiologiche di AAS causino un aumento del numero dei AR presenti nel muscolo scheletrico, tale espressione è comunque soggetta ad una regolazione genica con variabili soggettive di potenziale. Tali variabili sono teoricamente evincibili dall’osservazione degli atleti allenati contro-resistenza, specie Bodybuilder, e della loro differenza di potenziale indipendente nella sua massima espressione. Tale potenziale è diverso tra HIR e LOR sia in fisiologia che in condizione di trattamento farmacologico, indipendentemente dalla dose di AAS utilizzata.
Continua…
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
1- https://www.frontiersin.org/articles/10.3389/fphys.2018.01373/full
