Nella 1° parte abbiamo analizzato le caratteristiche e funzioni biochimiche degli Aminoacidi. In questa seconda parte, invece, inizieremo ad analizzare l’aspetto legato all’influenza degli Aminoacidi sulla sintesi proteica nel muscolo scheletrico e sulle potenzialità della loro integrazione in ambito sportivo e, in particolar modo, nel BodyBuilding.
Per tre parti di questa lunga disamina sugli AA, verranno trattati nello specifico cinque integratori di AA largamente utilizzati negli sport di potenza (ma non solo): la L-Glutammina, gli Aminoacidi a Catena Ramificata [BCAA], l’Acido β-idrossi β-metilbutirrico [HMB], la L-Arginina e la L-Citrullina. Al termine verranno trattati gli Aminoacidi Essenziali [EAA].
In questa seconda parte inizieremo con la trattazione della L-Glutammina…
Ma prima un piccolo ripasso…
Aminoacidi non-essenziali, condizionatamente-essenziali e essenziali:
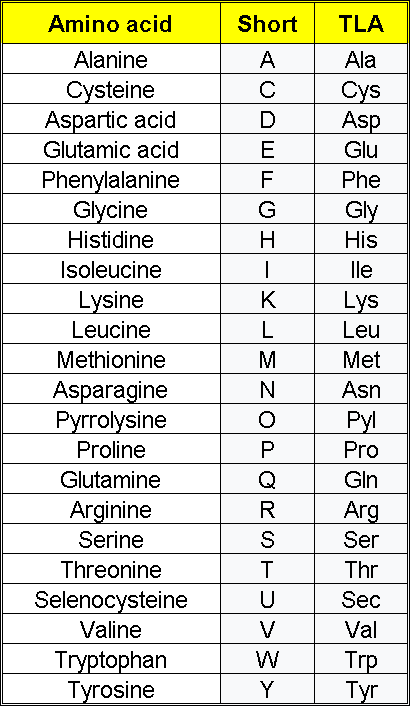
Dei 21 aminoacidi comuni a tutte le forme di vita, sei aminoacidi non sono essenziali per l’uomo, ovvero possono essere sintetizzati in quantità sufficienti dall’organismo. Questi sei sono l’alanina, l’acido aspartico, l’asparagina, l’acido glutammico, la serina[1] e la selenocisteina (considerata il 21° amminoacido). La pirrolisina (considerata il 22° aminoacido),[2] che è proteinogenica solo in alcuni microrganismi, non è utilizzata e quindi non è essenziale per la maggior parte degli organismi, compreso l’uomo.
Altri sei aminoacidi sono considerati condizionatamente essenziali nella dieta umana, il che significa che la loro sintesi può essere limitata in particolari condizioni fisiopatologiche, come la prematurità nel bambino o gli individui in grave difficoltà catabolica.[1] Questi sei aminoacidi sono arginina, cisteina, glicina, glutammina, prolina e tirosina.
I nove aminoacidi essenziali che l’uomo non può sintetizzare ex novo in modo sufficientemente rapido da soddisfarne il fabbisogno e devono quindi provenire dalla dieta sono valina, isoleucina, leucina, metionina, fenilalanina, triptofano, treonina, istidina e lisina.[3][1]
Caratteristiche base della Glutammina:
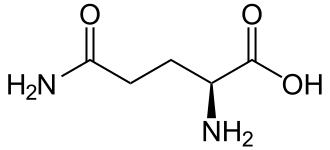
Scheletro molecolare della L-Glutammina.
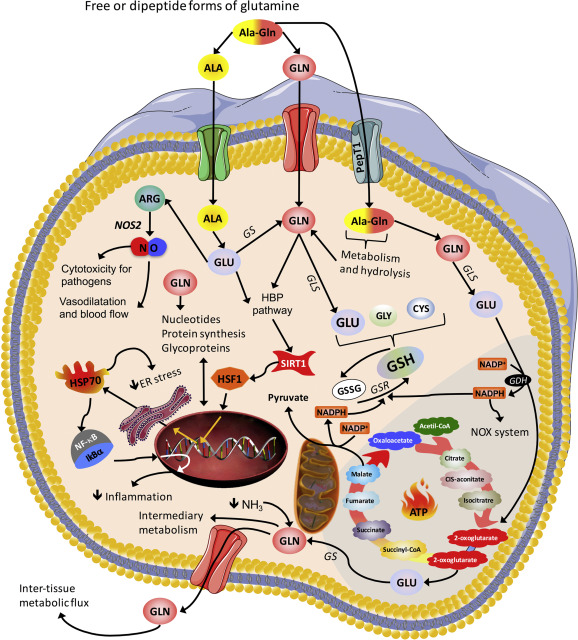
La Glutammina (simbolo Gln o Q)[4] è un α-amminoacido utilizzato nella biosintesi delle proteine. La sua catena laterale è simile a quella dell’acido glutammico, con la differenza che il gruppo carbossilico è sostituito da un’ammide. È classificato come un aminoacido polare a carica neutra. Nell’uomo è un amminoacido condizionatamente essenziale, il che significa che l’organismo è solitamente in grado di sintetizzarne quantità sufficienti, ma in alcuni casi di significativo stress la richiesta di Glutammina da parte dell’organismo aumenta e questo AA deve essere ottenuta dalla dieta.[5][6] È codificata dai codoni CAA e CAG. Prende il nome dall’acido glutammico, che a sua volta prende il nome dalla sua scoperta nelle proteine dei cereali, il glutine.[7]
Nel sangue umano, la Glutammina è l’amminoacido libero più abbondante.[8]
La Glutammina si trova in quantità elevate nella maggior parte delle carni e dei prodotti animali, nonché in qualsiasi prodotto o sottoprodotto lattiero-caseario come le proteine del siero o della caseina.[9] E’ presente anche nei fagioli, nelle barbabietole, nei cavoli, negli spinaci, nelle carote, nel prezzemolo, nei succhi di verdura e anche nel grano, nella papaia, nei cavoletti di Bruxelles, nel sedano e negli alimenti fermentati come il miso. I livelli di Glutammina nei vari alimenti variano da:
– Manzo al 4,7% di proteine[9], mentre la carne in generale oscilla tra il 4,4% e il 4,8%[9]. – Latte scremato all’8,08% di proteine[9], mentre i prodotti lattiero-caseari in generale tendono a fluttuare tra l’8,7% e il 9,2%[9]. – Riso bianco all’11,1% di proteine[9] – Mais con il 16,2% di proteine[9] – Tofu con il 9,1% di proteine[9] – Uova con il 4,3% di proteine[9]
Si osserva inoltre che alcuni di questi livelli di Glutammina possono essere sottovalutati e, di conseguenza, i livelli di glutammato più alti del previsto; ciò è dovuto a uno dei metodi storicamente utilizzati per l’analisi degli aminoacidi, l’idrolisi, che induce la conversione della Glutammina in glutammato[10][11] o acido piroglutammico. Il confronto dei risultati tra i metodi convenzionali e il sequenziamento genico può produrre differenze fino al 4% negli aminoacidi totali (l’influenza sulla Glutammina dipenderebbe dal contenuto di questa negli alimenti).[9]
Biodisponibilità e funzioni di interesse principale per lo sportivo inerenti alla Glutammina:
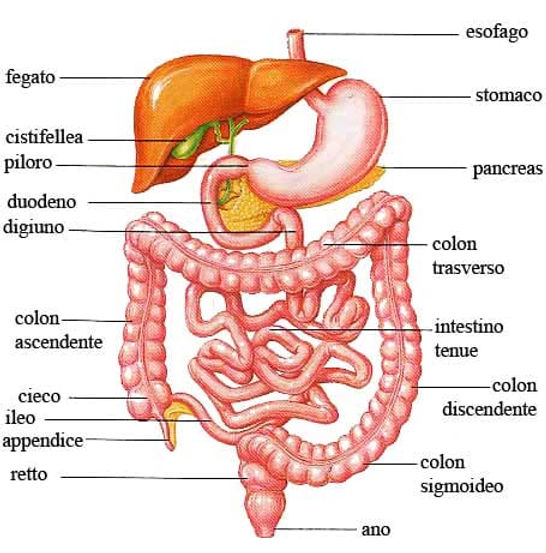
Fino al 13% della Glutammina circolante tende a essere reindirizzata al letto splancnico per essere utilizzata come substrato energetico dal fegato e dagli enterociti intestinali.[12]
La quantità di Glutammina destinata ai tessuti intestinali ed epatici (estrazione splancnica) non differisce tra le fonti legate agli alimenti e i dosaggi degli integratori.[13]
Quando viene somministrata Glutammina per via orale o endovenosa, i tassi di sintesi de novo della Glutammina diminuiscono.[14][15] Ciò può preservare indirettamente gli aminoacidi che potrebbero essere convertiti in Glutammina, come la Leucina che subisce una riduzione dei tassi di ossidazione.[15]
È stato dimostrato che la Glutammina è in grado di “smorzare” i picchi di glucosio nel sangue in risposta ai carboidrati alimentari, attenuando gli aumenti e i valori di Cmax della glicemia e dell’Insulina in risposta all’ingestione di carboidrati con la dieta.[16] E’ stato esaminato se ciò sia dovuto a ritardi non significativi nello svuotamento gastrico, ma non sembra essere così.[16]
La Glutammina è un aminoacido intimamente legato in vitro all’omeostasi muscolare e alla sintesi proteica muscolare, in cui un eccesso provoca anabolismo e previene la disgregazione, mentre un deficit provoca catabolismo.[17][18][19] Questa correlazione è stata osservata in vivo quando la Glutammina viene infusa[18][20] (alcune controprove[21][22]) e sembra essere specifica per la Glutammina.[18][23]
In vitro, la Glutammina è nota per ridurre i tassi di ossidazione della Leucina e aumentare il deposito di Leucina, che aumenta gli effetti della stessa in una cellula muscolare scheletrica.[15]
Gli studi che hanno utilizzato la Glutammina in persone altrimenti sane e che hanno analizzato la sintesi proteica muscolare o l’aumento della massa magra hanno rilevato un fallimento con 900mg/kg di massa magra (il placebo era di 900mg/kg di Maltodestrina) nei giovani abbinati all’allenamento contro-resistenza.[24]
Anche l’aggiunta di Glutammina alla Creatina[25] o di Glutammina extra (300mg/kg di peso corporeo) a un frullato di proteine e carboidrati[26] o di aminoacidi e carboidrati[26] non è riuscita a superare gli integratori ingeriti senza Glutammina, il che suggerisce che non ha alcun ruolo sinergico.
I livelli plasmatici di Glutammina sono aumentati o invariati nelle attività di breve durata e ad alta intensità[27][28] e tendono a rimanere invariati in caso di danno muscolare eccentrico[29], il che suggerisce che un’integrazione extra di Glutammina non apporterà benefici all’esercizio fisico di breve durata o al sollevamento pesi con qualsiasi mezzo che agisca sui livelli sierici di Glutammina (come l’immunosoppressione o il catabolismo). Al contrario, gli eventi di resistenza che superano le 2 ore tendono a mostrare una diminuzione dei livelli di Glutammina nel siero.[30][31] Sia l’integrazione di Glutammina che l’aumento dell’apporto proteico dall’alimentazione (nella dose di 20-30g di proteine di origine animale) possono alleviare questa diminuzione della Glutammina nel siero[32] e potenzialmente possono ridurre i danni alle cellule immunitarie associati all’esercizio cardiovascolare prolungato. [33] Questa diminuzione dei livelli sierici di Glutammina può anche sopprimere il rilascio di interleuchina-6 (IL-6) dal tessuto muscolare e l’integrazione di Glutammina può preservare i livelli di IL-6.[34]
Questi risultati sono coerenti con la teoria secondo cui l’esercizio cardiovascolare prolungato, attraverso la riduzione della Glutammina, può sopprimere la funzione immunologica ostacolando la differenziazione dei leucociti.[30][35]
L’ingestione di 300mg/kg di Glutammina in sollevatori di pesi altrimenti sani non è riuscita a modificare la produzione di potenza più del placebo[36] e dosi più elevate (900mg/kg di massa corporea magra) hanno fallito allo stesso modo altrove in popolazioni attive.[24]
Attenuando o prevenendo in altro modo la deplezione di Glutammina negli esercizi che durano più di un’ora, le prestazioni possono indirettamente aumentare rispetto allo stato di deplezione di Glutammina. Non si tratta tanto di un “miglioramento” delle prestazioni quanto di una loro “conservazione”.[37]
Prednisone
L’ingestione di Glutammina, a 0,5g/kg al giorno, in un piccolo studio su pazienti ipercortisolemici (indotti con Prednisone a una dose tale da indurre la disgregazione delle proteine muscolari) ha mostrato di apportare un minore stato catabolico attraverso la riduzione della conversione degli aminoacidi essenziali in Glutammina e un minore dispendio di Leucina.[38]
Ci sono alcune prove che la Glutammina orale può aumentare il tasso di risintesi del glicogeno se consumata insieme ai carboidrati[39], ma sono necessari ulteriori studi per verificare se questo metodo sia più vantaggioso rispetto alle fonti alimentari di Glutammina o se sia vero con un’assunzione di carboidrati più elevata. La Glutammina stessa, in assenza di carboidrati, può aumentare le scorte di glicogeno muscolare[40].
È stato dimostrato che l’integrazione di Glutammina stimola la sintesi proteica nell’intestino di persone sane con una potenza simile a quella degli aminoacidi misti.[41]
I benefici della Glutammina nelle persone gravemente ferite o malate hanno indotto alcuni ricercatori a proporre che potrebbe essere un integratore utile per gli atleti impegnati nell’esercizio fisico contro-resistenza, e che è anche di natura catabolica.[42] Questi ricercatori hanno testato la loro ipotesi attraverso un RCT in doppio cieco che ha coinvolto 6 uomini allenati contro-resistenza che hanno consumato Glutammina o Glicina (0,3 grammi per chilogrammo di peso corporeo) un’ora prima di una sessione di sollevamento pesi.[43] La Glutammina non ha apportato benefici alle prestazioni. Un altro gruppo di ricercatori ha testato la Glutammina (0,9g/kg) contro il placebo in 31 uomini e donne allenati contro-resistenza durante un programma di allenamento di 6 settimane.[44] Anche una dose giornaliera così elevata di Glutammina non ha influenzato la forza o la massa corporea magra (LBM) più del placebo (la forza e la LBM sono aumentate in entrambi i gruppi). Naturalmente, nessuno dei due studi ha esposto i suoi partecipanti agli alti livelli di stress sperimentati, per esempio, dalle vittime di ustioni. Un RCT che ha coinvolto 18 lottatori maschi universitari ha cercato di risolvere questo problema confrontando il placebo con la Glutammina (0,35g/kg) durante un Cut intensivo di 12 giorni.[45] Entrambi i gruppi hanno perso 2 kg, senza differenze significative tra i gruppi per quanto riguarda i cambiamenti nella LBM o nella massa grassa. Anche una meta-analisi del 2018 di 5 studi non ha rilevato alcun beneficio della Glutammina sulla composizione corporea.[46] Sebbene la Glutammina svolga un ruolo nella sintesi proteica muscolare (è un attivatore indipendente di mTOR[47]), ciò che otteniamo attraverso gli alimenti sembra essere sufficiente; l’integrazione non sembra conferire ulteriori benefici. L’integrazione di Glutammina non ha alcun effetto sulla massa magra o sulla massa grassa, nemmeno durante una dieta ipocalorica “aggressiva”.
L’integrazione di Glutammina non ha quindi alcun effetto sulla composizione corporea; ma potrebbe facilitare il recupero dopo sessioni di allenamento di resistenza?
Per rispondere a questa domanda, i ricercatori hanno somministrato placebo o glutammina (0,3 g/kg) a 15 uomini attivi a livello ricreativo subito dopo un esercizio che danneggiava i muscoli (100 drop jump) e per i quattro giorni successivi. Hanno riferito che, rispetto al placebo, la glutammina ha ridotto significativamente l’indolenzimento muscolare e ha migliorato il recupero della forza.[48]
Tuttavia, uno studio condotto su 17 giovani uomini non allenati ha riportato che l’assunzione di glutammina (0,1 g/kg) tre volte alla settimana per 4 settimane non ha avuto alcun effetto sull’indolenzimento muscolare, sul range di movimento o sull’attività EMG fino a 48 ore dopo un esercizio dannoso per la muscolatura (leg extension eccentrica al 75% dell’1-RM).[49] È importante notare che quest’ultimo studio ha coinvolto uomini non allenati e ha utilizzato una dose minore, un diverso schema di dosaggio e un diverso protocollo di esercizio: tutti fattori che potrebbero spiegare la discrepanza tra i due studi. Una contrazione è isometrica quando la lunghezza del muscolo non cambia e isotonica in caso contrario. Una contrazione isotonica è detta concentrica quando il muscolo si accorcia sotto carico (come quando si solleva un manubrio) ed eccentrica quando si allunga sotto carico (come quando si controlla il manubrio in discesa). Il massimo ad una ripetizione (1-RM) è il peso più pesante che si può sollevare (contrazione concentrica) per un determinato esercizio.
Più di recente, uno studio condotto su 23 uomini allenati alla resistenza ha esaminato gli effetti dell’assunzione di glutammina insieme alla leucina.[50] Gli uomini sono stati randomizzati in tre gruppi e hanno assunto leucina (0,087 g/kg), leucina con glutammina (0,087 g/kg + 0,3 g/kg) o un placebo 30 minuti prima e dopo un esercizio che danneggiava i muscoli (100 drop jump), e di nuovo prima e dopo i test di recupero effettuati 24, 48 e 72 ore dopo. La leucina ha portato a un migliore recupero della forza solo a 72 ore. La leucina con glutammina ha portato a un migliore recupero della forza a 24, 48 e 72 ore. L’indolenzimento muscolare, invece, non differiva tra i gruppi. Si noti che questi tre studi sono stati condotti solo su uomini. Un altro studio ha reclutato 8 uomini e 8 donne, tutti attivi a livello ricreativo, e ha somministrato loro placebo o glutammina (0,3 g/kg) un’ora prima e dopo un esercizio che danneggiava i muscoli (80 contrazioni eccentriche al 125% dell’1-RM), e di nuovo prima dei test di recupero condotti 24, 48 e 72 ore dopo.[51] Il recupero della forza è stato modestamente migliorato negli uomini ma non nelle donne, anche se entrambi i sessi hanno registrato una riduzione significativa dell’indolenzimento muscolare.
Negli uomini attivi a livello ricreativo, l’integrazione di Glutammina dopo l’esercizio sembra migliorare il recupero della forza e potrebbe ridurre l’indolenzimento muscolare. Tuttavia, solo uno studio è durato più di 72 ore, il che impedisce di trarre conclusioni sugli effetti dell’integrazione cronica. Allo stesso modo, solo uno studio ha incluso donne, il che impedisce di trarre conclusioni sugli effetti dell’integrazione nelle donne.
La Glutammina è un importante carburante per le cellule del sistema immunitario.[52] I livelli plasmatici di Glutammina si riducono dopo un esercizio di resistenza prolungato e questa riduzione è correlata a un aumento del rischio di infezioni.[53]
Un primo studio su atleti di resistenza (maratoneti e ultramaratoneti) ha riportato che l’assunzione di 5 grammi di Glutammina subito dopo un evento atletico e 2 ore dopo ha ridotto significativamente l’insorgenza di infezioni nella settimana successiva.[54] In particolare, il 19% del gruppo Glutammina ha riportato malattie, rispetto alla metà del gruppo placebo. Nessun altro studio ha studiato gli effetti della glutammina sulle infezioni come risultato negli atleti, ma altri studi hanno esaminato vari aspetti del sistema immunitario,[46] come la funzione dei globuli bianchi[55][56] e le concentrazioni di IgA salivari,[57] e nessuno ha trovato una relazione tra la diminuzione dei livelli plasmatici di glutammina indotta dall’esercizio fisico e i cambiamenti del sistema immunitario.[58]
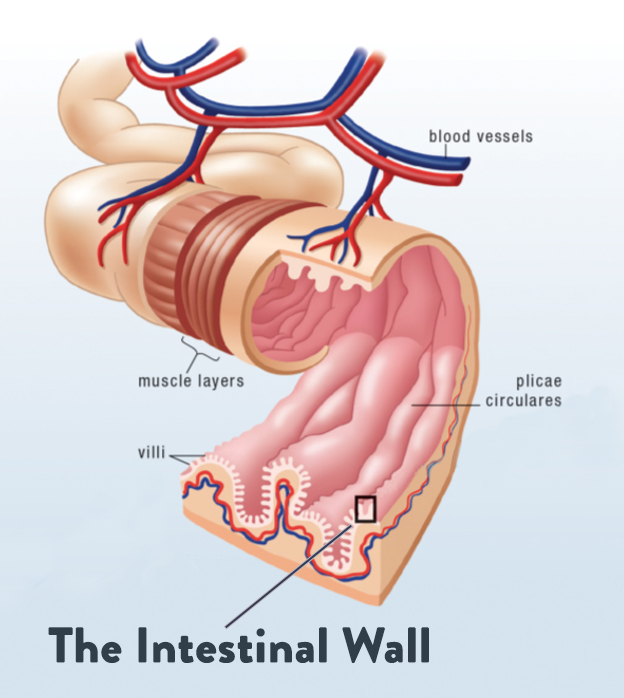
I benefici della Glutammina potrebbero essere mediati dai suoi effetti sulla barriera intestinale, come hanno iniziato a suggerire prove più recenti. È noto che l’esercizio fisico di resistenza prolungato provoca il leaky gut, una condizione in cui lo stress da calore e la riduzione del flusso sanguigno nel tratto gastrointestinale causano danni alle cellule intestinali che allentano le giunzioni strette tra le cellule, consentendo l’assorbimento di elementi che non dovrebbero passare attraverso la barriera intestinale.[59]
In uno studio recente, la Glutammina (0,25, 0,5 e 0,9 g/kg) ha mostrato una riduzione dose-dipendente della permeabilità intestinale indotta dall’esercizio fisico.[60] In uno studio precedente, la riduzione della permeabilità intestinale derivante dall’integrazione di glutammina era correlata a riduzioni dell’endotossina sierica e dei marcatori infiammatori.[61]
La Glutammina è un’importante fonte di energia per le cellule intestinali e per il sistema immunitario. L’integrazione può ridurre le disfunzioni del tratto intestinale indotte dall’esercizio fisico e potrebbe diminuire il rischio di ammalarsi in seguito a un esercizio di resistenza prolungato.
Conclusioni sulla Glutammina:
Che che se ne dica, non è dimostrato che la Glutammina supplementare aiuti a incentivare significativamente la sintesi proteica o a migliorare la composizione corporea.
È dimostrato, invece, che l’integrazione di Glutammina migliora il recupero negli allenamenti di forza/contro-resistenza. Può anche aiutare a mantenere un buon stato di salute intestinale in contesti di stress alimentare e mantenere l’integrità del tratto gastrointestinale durante l’esercizio contro-resistenza prolungato e quindi a ridurre il rischio che si incappi in condizioni croniche/patologiche.
Quindi, se il vostro obiettivo è incentivare la sintesi proteica, non sprecate i vostri soldi con la Glutammina. È più probabile che altri integratori siano utili, in particolare la Creatina, e naturalmente assicuratevi di assumere una quantità sufficiente di proteine.
Ma se volete sperimentare con questo AA allora potete farlo nella maniera più logica basandosi sulle attuali evidenze:
Come booster per la risintesi di Glicogeno [post-workout o durante un refeed] = 8g nel post workout o 10g nei pasti principali del Refeed (30g totali);
Anticatabolico durante la restrizione calorica: >20g divisi durante la giornata.
Dal momento che la Glutammina ha mostrato di poter apportare benefici a livello intestinale, essa sta venendo riproposta sia nel mercato del fitness, sia in quello dell’integrazione su larga scala. Parlando di Bodybuilding non si può non accennare ai problemi intestinali digestivi correlati ai periodi di dieta ipercalorica. In questi casi la Glutammina potrebbe svolgere un ruolo importante a livello di integrazione, anche se non in tutti i soggetti. Un dosaggio di 0.3-0.5g/Kg/die potrebbe apportare dei benefici a livello intestinale.
Nota importante: Il limite di sicurezza osservato per l’integrazione di Glutammina, ovvero la quantità massima che si può assumere con la certezza di non avere effetti collaterali, è stato suggerito in 14g/die in forma di integratore (al di sopra dell’assunzione di cibo). Livelli più alti sono stati testati e ben tollerati, ma non ci sono prove sufficienti per suggerire che dosi più elevate siano completamente prive di danni nel corso di una integrazione cronica, né prove sufficienti per presumere l’esistenza di danni. Prove limitate suggeriscono che 50-60g/die per un periodo di alcune settimane non sono associati a effetti avversi significativi.
A livello acuto, dosi di circa 0,75g/kg di peso corporeo sono state implicate nell’aumento dei livelli di ammoniaca plasmatica al di sopra del limite di sicurezza tollerato. Uno studio condotto su persone anziane (69+/-8,8 anni) con 0,5g/kg di glutammina per via orale non ha mostrato effetti sui livelli di ammoniaca plasmatica, ma è stato associato a un aumento dell’urea e della creatinina sieriche che non è stato ritenuto clinicamente rilevante. È stata osservata una diminuzione transitoria della velocità di filtrazione glomerulare dei reni.
Continua…
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Otten, Jennifer J.; Hellwig, Jennifer Pitzi; Meyers, Linda D., eds. (2006) [1943]. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements (Technical report). doi:10.17226/11537. ISBN978-0-309-15742-1.
Gli aminoacidi sono una classe di molecole biologiche costituenti le unità che formano le proteine, e svolgono numerose funzioni fondamentali per la corretta funzione del corpo umano. Rappresentano un elemento conosciuto a grandi linee da tutti gli assidui frequentatori di sala pesi, ma la maggior parte di loro è all’oscuro delle loro caratteristiche e reali richieste, quando una loro supplementazione risulta funzionale e quando, invece, si traduce in una pratica pressoché sterile. Questo primo articolo è finalizzato ad iniziare una approfondita disamina sugli aminoacidi facendo chiarezza sul significato biochimico e sulla loro più aggiornata applicazione in ambito sportivo soprattutto per quanto concerne gli Aminoacidi Essenziali.
Cosa sono gli Aminoacidi?
Gli aminoacidi sono composti organici che contengono sia gruppi funzionali amminici che carbossilici.[1] Sebbene in natura esistano oltre 500 amminoacidi, i più importanti sono i 22 α-amminoacidi incorporati nelle proteine.[2] Solo questi 22 compaiono nel codice genetico della vita.[3][4]
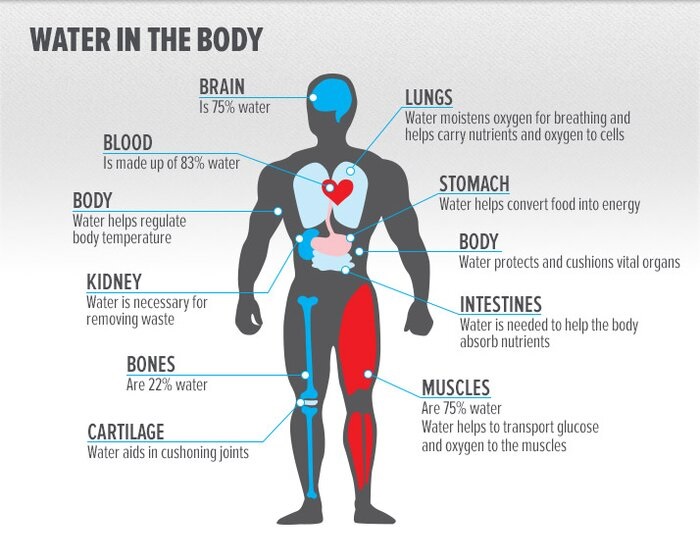
Gli aminoacidi possono essere classificati in base alla posizione dei gruppi funzionali strutturali principali (amminoacidi alfa (α-), beta (β-), gamma (γ-), ecc.); altre categorie riguardano la polarità, la ionizzazione e il tipo di catena laterale (alifatica, aciclica, aromatica, polare, ecc.). Sotto forma di proteine, i residui di aminoacidi costituiscono la seconda componente (l’acqua è la più grande) dei muscoli e degli altri tessuti umani.[5] Oltre al ruolo di residui nelle proteine, gli aminoacidi partecipano a una serie di processi come il trasporto e la biosintesi dei neurotrasmettitori.[6]
Storia:
Formula scheletrica di L-asparagina
I primi amminoacidi furono scoperti all’inizio del 1800.[7][8] Nel 1806, i chimici francesi Louis-Nicolas Vauquelin e Pierre Jean Robiquet isolarono dagli asparagi un composto che fu poi chiamato asparagina, il primo amminoacido ad essere scoperto.[9][10] La cistina fu scoperta nel 1810,[11] anche se il suo monomero, la cisteina, rimase sconosciuto fino al 1884. [La glicina e la leucina furono scoperte nel 1820.[12[13] L’ultimo dei 20 aminoacidi comuni ad essere scoperto fu la treonina nel 1935 da William Cumming Rose, che determinò anche gli aminoacidi essenziali e stabilì il fabbisogno minimo giornaliero di tutti gli aminoacidi per una crescita ottimale.[14][15]
L’unità della categoria chimica fu riconosciuta da Wurtz nel 1865, ma non le diede un nome particolare.[16] Il primo uso del termine “aminoacido” in lingua inglese risale al 1898,[17] mentre il termine tedesco, Aminosäure, era già stato usato in precedenza.[18] Si è scoperto che le proteine producono aminoacidi in seguito a digestione enzimatica o idrolisi acida. Nel 1902, Emil Fischer e Franz Hofmeister proposero indipendentemente che le proteine sono formate da molti amminoacidi, per cui si formano legami tra il gruppo amminico di un amminoacido e il gruppo carbossilico di un altro, dando luogo a una struttura lineare che Fischer definì “peptide”.[19]
Struttura generale
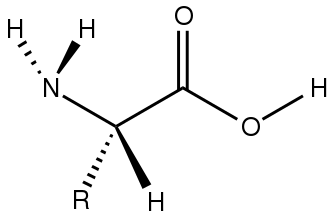
I 2-, alfa- o α-amminoacidi[20] hanno nella maggior parte dei casi la formula generica H2NCHRCOOH, dove R è un sostituente organico noto come “catena laterale”.[21]
Struttura di un tipico L-alfa-amminoacido nella forma “neutra”.
Delle centinaia di amminoacidi descritti, 22 sono proteinogenici (“costruiscono proteine”).[22][23][24] Sono questi 22 composti che si combinano per dare una vasta gamma di peptidi e proteine assemblate dai ribosomi.[25] Gli amminoacidi non proteinogenici o modificati possono derivare da modificazioni post-traslazionali o durante la sintesi di peptidi nonribosomiali.
I 21 α-amminoacidi proteinogenici presenti negli eucarioti, raggruppati in base ai valori pKa delle loro catene laterali e alle cariche trasportate a pH fisiologico (7,4).
L’atomo di carbonio vicino al gruppo carbossilico è chiamato carbonio α. Negli amminoacidi proteinogenici, porta l’ammina e il gruppo R o la catena laterale specifica di ciascun amminoacido. Con quattro sostituenti distinti, il carbonio α è stereogenico in tutti gli α-amminoacidi tranne la glicina. Tutti gli amminoacidi proteogenici chirali hanno la configurazione L. Sono “sinistrorsi”. Si tratta di enantiomeri “sinistrorsi”, che si riferiscono agli stereoisomeri del carbonio alfa.
Alcuni amminoacidi D (“destrorsi”) sono stati trovati in natura, ad esempio negli involucri batterici, come neuromodulatore (D-serina) e in alcuni antibiotici.[26][27] Raramente, i residui di amminoacidi D si trovano nelle proteine e vengono convertiti dall’amminoacido L come modifica post-traduzionale.[28]
Cinque amminoacidi possiedono una carica a pH neutro. Spesso queste catene laterali appaiono sulla superficie delle proteine per consentirne la solubilità in acqua, e le catene laterali con cariche opposte formano importanti contatti elettrostatici chiamati ponti salini che mantengono le strutture all’interno di una singola proteina o tra proteine interfacciate.[29] Molte proteine legano il metallo nelle loro strutture in modo specifico, e queste interazioni sono comunemente mediate da catene laterali cariche come l’aspartato, il glutammato e l’istidina. In determinate condizioni, ogni gruppo che forma ioni può essere carico, formando sali doppi.[30]
Gruppi funzionali presenti nell’istidina (a sinistra), nella lisina (al centro) e nell’arginina (a destra)
I due aminoacidi carichi negativamente a pH neutro sono l’aspartato (Asp, D) e il glutammato (Glu, E). I gruppi carbossilati anionici si comportano come basi di Brønsted nella maggior parte dei casi.[29] Gli enzimi in ambienti a pH molto basso, come la pepsina, proteasi aspartica nello stomaco dei mammiferi, possono avere residui catalitici di aspartato o glutammato che agiscono come acidi di Brønsted.
Formula scheletrica dell’Istidina (forma zwitterionica)
Ci sono tre amminoacidi con catene laterali che sono cationi a pH neutro: l’arginina (Arg, R), la lisina (Lys, K) e l’istidina (His, H). L’arginina ha un gruppo guanidino carico e la lisina un gruppo alchilico amminico carico e sono completamente protonati a pH 7. Il gruppo imidazolico dell’istidina ha un pKa di 6,0 ed è protonato solo per il 10% circa a pH neutro. Poiché l’istidina si trova facilmente nelle sue forme basiche e acide coniugate, partecipa spesso ai trasferimenti catalitici di protoni nelle reazioni enzimatiche.[29]
Formula scheletrica di L-glutammina
Gli aminoacidi polari e privi di carica serina (Ser, S), treonina (Thr, T), asparagina (Asn, N) e glutammina (Gln, Q) formano prontamente legami a idrogeno con l’acqua e con altri aminoacidi.[29] Non si ionizzano in condizioni normali; un’eccezione importante è rappresentata dalla serina catalitica nelle serina-proteasi. Questo è un esempio di grave perturbazione e non è caratteristico dei residui di serina in generale. La treonina ha due centri chirali, non solo il centro chirale L (2S) sul carbonio α condiviso da tutti gli amminoacidi, a parte la glicina achirale, ma anche (3R) sul carbonio β. La specifica stereochimica completa è (2S,3R)-L-treonina.
Formula scheletrica di L-tirosina
Le interazioni tra gli amminoacidi non polari sono la forza motrice principale dei processi di ripiegamento delle proteine nelle loro strutture tridimensionali funzionali.[29] Nessuna delle catene laterali di questi amminoacidi si ionizza facilmente e quindi non hanno pKas, ad eccezione della tirosina (Tyr, Y). L’idrossile della tirosina può deprotonarsi ad alto pH formando un fenolato carico negativamente. Per questo motivo si potrebbe collocare la tirosina nella categoria degli aminoacidi polari e privi di carica, ma la sua bassissima solubilità in acqua corrisponde bene alle caratteristiche degli aminoacidi idrofobici.
Formula scheletrica di L-glicina neutrale
Diverse catene laterali non sono ben descritte dalle categorie cariche, polari e idrofobiche. La glicina (Gly, G) potrebbe essere considerata un amminoacido polare, poiché le sue piccole dimensioni fanno sì che la sua solubilità sia determinata in gran parte dai gruppi amminici e carbossilici. Tuttavia, la mancanza di catene laterali conferisce alla glicina una flessibilità unica tra gli amminoacidi, con ampie ramificazioni nel ripiegamento delle proteine.[29] Anche la cisteina (Cys, C) può formare facilmente legami idrogeno, il che la collocherebbe nella categoria degli amminoacidi polari, anche se spesso si trova nelle strutture proteiche a formare legami covalenti, detti legami disolfuro, con altre cisteine. Questi legami influenzano il ripiegamento e la stabilità delle proteine e sono essenziali nella formazione degli anticorpi. La prolina (Pro, P) ha una catena laterale alchilica e potrebbe essere considerata idrofobica, ma poiché la catena laterale si unisce di nuovo al gruppo alfa-amminico, diventa particolarmente inflessibile quando viene incorporata nelle proteine. Come la glicina, influenza la struttura delle proteine in un modo unico tra gli aminoacidi. La selenocisteina (Sec, U) è un raro amminoacido non codificato direttamente dal DNA, ma incorporato nelle proteine attraverso il ribosoma. La selenocisteina ha un potenziale redox più basso rispetto alla cisteina simile e partecipa a diverse reazioni enzimatiche uniche.[31] La pirrolisina (Pyl, O) è un altro aminoacido non codificato nel DNA, ma sintetizzato nelle proteine dai ribosomi.[34] Si trova in specie arcaiche dove partecipa all’attività catalitica di diverse metiltransferasi.
Gli amminoacidi con struttura NH+3-CXY-CXY-CO-2, come la β-alanina, componente della carnosina e di alcuni altri peptidi, sono β-amminoacidi. Quelli con la struttura NH+3-CXY-CXY-CXY-CO-2 sono γ-amminoacidi, e così via, dove X e Y sono due sostituenti (uno dei quali è normalmente H).[6]
Le forme naturali comuni di amminoacidi hanno una struttura zwitterionica, con gruppi funzionali -NH+3 (-NH+2- nel caso della prolina) e -CO-2 attaccati allo stesso atomo di C; sono quindi α-amminoacidi e sono gli unici che si trovano nelle proteine durante la traduzione nel ribosoma. In soluzione acquosa, a pH prossimo alla neutralità, gli amminoacidi esistono come zwitterioni, cioè come ioni dipolari con entrambi i gruppi NH+3 e CO-2 in stati carichi, per cui la struttura complessiva è NH+3-CHR-CO-2. A pH fisiologico le cosiddette “forme neutre” -Sebbene le due cariche della struttura zwitterionica si sommino a zero, è fuorviante definire “scarica” una specie con carica netta pari a zero.
In condizioni di forte acidità (pH inferiore a 3), il gruppo carbossilato viene protonato e la struttura diventa un acido carbossilico ammonio, NH+3 -CHR-CO2H. Ciò è rilevante per gli enzimi come la pepsina che sono attivi in ambienti acidi come lo stomaco e i lisosomi dei mammiferi, ma non si applica in modo significativo agli enzimi intracellulari. In condizioni altamente basiche (pH superiore a 10, normalmente non riscontrabile in condizioni fisiologiche), il gruppo ammonio viene deprotonato per dare NH2-CHR-CO-2.
Ionizzazione e carattere di Brønsted dell’ammino N-terminale, del carbossilato C-terminale e delle catene laterali dei residui amminoacidici
Sebbene in chimica si utilizzino varie definizioni di acidi e basi, l’unica utile per la chimica in soluzione acquosa è quella di Brønsted:[32][33] un acido è una specie che può donare un protone a un’altra specie, mentre una base è una specie che può accettare un protone. Questo criterio viene utilizzato per etichettare i gruppi nell’illustrazione precedente. Le catene laterali carbossilate dei residui di aspartato e glutammato sono le principali basi di Brønsted nelle proteine. Allo stesso modo, la lisina, la tirosina e la cisteina agiscono tipicamente come acidi Brønsted. L’istidina, in queste condizioni, può agire sia come acido che come base di Brønsted.
Per gli amminoacidi con catene laterali non cariche, lo zwitterione predomina a valori di pH compresi tra i due valori di pKa, ma coesiste in equilibrio con piccole quantità di ioni netti negativi e positivi. Nel punto intermedio tra i due valori di pKa, la traccia di ioni negativi netti e la traccia di ioni positivi netti si bilanciano, in modo che la carica netta media di tutte le forme presenti sia pari a zero.[34] Questo pH è noto come punto isoelettrico pI, per cui pI = 1/2 (pKa1 + pKa2).
Composito di curve di titolazione di venti aminoacidi proteinogenici raggruppati per categoria di catena laterale
Per gli amminoacidi con catene laterali cariche, è coinvolto il pKa della catena laterale. Così per l’aspartato o il glutammato con catene laterali negative, il gruppo amminico terminale è essenzialmente interamente nella forma carica -NH+3, ma questa carica positiva deve essere bilanciata dallo stato con un solo gruppo carbossilato C-terminale carico negativamente. Questo si verifica a metà strada tra i due valori di pKa del carbossilato: pI = 1/2 (pKa1 + pKa(R)), dove pKa(R) è il pKa della catena laterale.[33]
Considerazioni simili valgono per altri amminoacidi con catene laterali ionizzabili, tra cui non solo il glutammato (simile all’aspartato), ma anche la cisteina, l’istidina, la lisina, la tirosina e l’arginina con catene laterali positive.
Gli amminoacidi hanno mobilità nulla nell’elettroforesi al loro punto isoelettrico, anche se questo comportamento è più sfruttato per i peptidi e le proteine che per i singoli amminoacidi. Gli zwitterioni hanno una solubilità minima al loro punto isoelettrico e alcuni amminoacidi (in particolare quelli con catene laterali non polari) possono essere isolati per precipitazione dall’acqua regolando il pH al punto isoelettrico richiesto.
Proprietà fisico-chimiche
I 20 amminoacidi canonici possono essere classificati in base alle loro proprietà. Fattori importanti sono la carica, l’idrofilia o l’idrofobicità, la dimensione e i gruppi funzionali.[27] Queste proprietà influenzano la struttura delle proteine e le interazioni proteina-proteina. Le proteine idrosolubili tendono ad avere i loro residui idrofobici (Leu, Ile, Val, Phe e Trp) sepolti al centro della proteina, mentre le catene laterali idrofile sono esposte al solvente acquoso. (In biochimica, un residuo si riferisce a uno specifico monomero all’interno della catena polimerica di un polisaccaride, di una proteina o di un acido nucleico). Le proteine integrali di membrana tendono ad avere anelli esterni di aminoacidi idrofobici esposti che le ancorano al bilayer lipidico. Alcune proteine di membrana periferiche hanno una zona di aminoacidi idrofobici sulla loro superficie che si attacca alla membrana. In modo simile, le proteine che devono legarsi a molecole cariche positivamente hanno superfici ricche di aminoacidi carichi negativamente, come il glutammato e l’aspartato, mentre le proteine che si legano a molecole cariche negativamente hanno superfici ricche di aminoacidi carichi positivamente, come la lisina e l’arginina. Ad esempio, la lisina e l’arginina sono presenti in grandi quantità nelle regioni a bassa complessità delle proteine che legano gli acidi nucleici.[35] Esistono varie scale di idrofobicità dei residui di amminoacidi.[36]
Diagramma della struttura delle proteine
Alcuni amminoacidi hanno proprietà speciali. La cisteina può formare legami disolfuro covalenti con altri residui di cisteina. La prolina forma un ciclo alla spina dorsale polipeptidica e la glicina è più flessibile di altri aminoacidi.
La glicina e la prolina sono fortemente presenti all’interno delle regioni a bassa complessità delle proteine sia eucariotiche che procariotiche, mentre l’opposto avviene con la cisteina, la fenilalanina, il triptofano, la metionina, la valina, la leucina, l’isoleucina, che sono altamente reattivi, o complessi, o idrofobici.[35][37][38]
Molte proteine subiscono una serie di modifiche post-traslazionali, in base alle quali gruppi chimici aggiuntivi vengono attaccati alle catene laterali dei residui aminoacidici, producendo talvolta lipoproteine (che sono idrofobiche),[39] o glicoproteine (che sono idrofile)[40] che consentono alla proteina di attaccarsi temporaneamente a una membrana. Ad esempio, una proteina di segnalazione può attaccarsi e poi staccarsi dalla membrana cellulare, perché contiene residui di cisteina a cui può essere aggiunto e successivamente rimosso l’acido grasso palmitico.[41]
Sintesi
Nelle piante, l’azoto viene prima assimilato in composti organici sotto forma di glutammato, formato da alfa-chetoglutarato e ammoniaca nel mitocondrio. Per gli altri amminoacidi, le piante utilizzano le transaminasi per spostare il gruppo amminico dal glutammato ad un altro alfa-chetoacido. Ad esempio, l’aspartato aminotransferasi converte il glutammato e l’ossalacetato in alfa-chetoglutarato e aspartato.[42] Anche altri organismi utilizzano le transaminasi per la sintesi degli aminoacidi.
La sintesi degli aminoacidi di Strecker
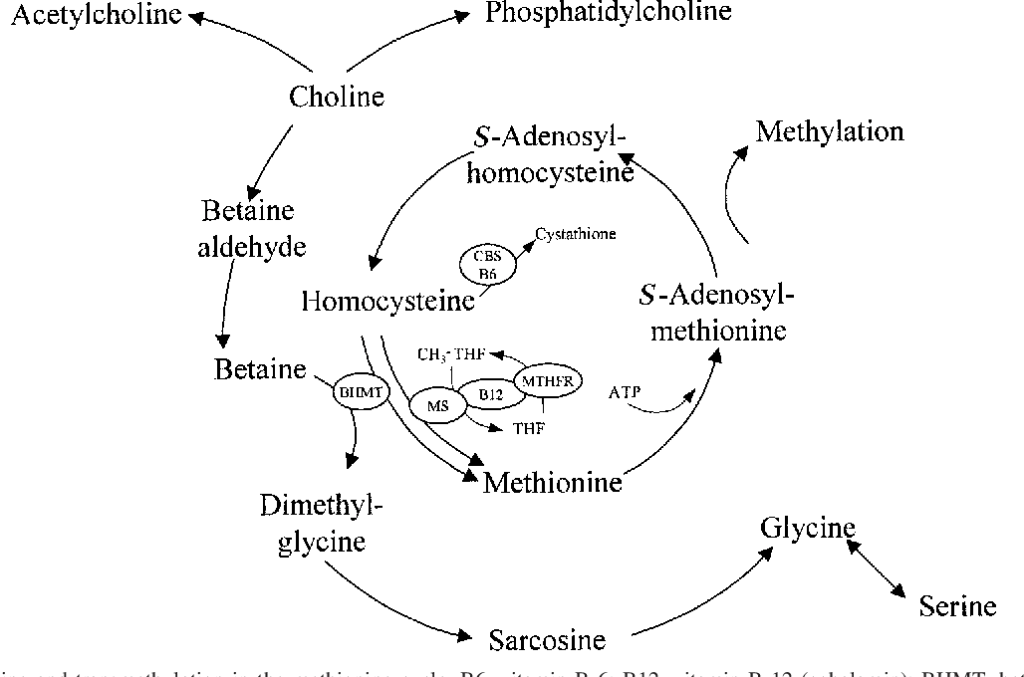
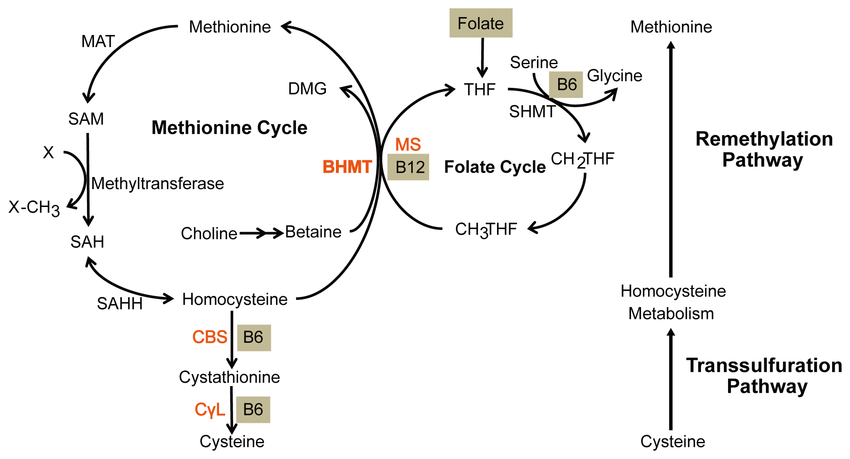
Gli amminoacidi non standard si formano solitamente attraverso modifiche agli amminoacidi standard. Ad esempio, l’omocisteina si forma attraverso la via della transulfurazione o mediante la demetilazione della metionina tramite il metabolita intermedio S-adenosilmetionina,[43] mentre l’idrossiprolina viene prodotta mediante una modifica post-traduzionale della prolina.[44]
Acido 2-amminoisobutirrico
I microrganismi e le piante sintetizzano molti amminoacidi non comuni. Ad esempio, alcuni microbi producono acido 2-amminoisobutirrico e lantionina, che è un derivato con ponti solforati dell’alanina. Entrambi questi amminoacidi si trovano nei lantibiotici peptidici come l’alameticina.[45] Tuttavia, nelle piante, l’acido 1-amminociclopropan-1-carbossilico è un piccolo amminoacido ciclico disostituito che è un intermedio nella produzione dell’ormone vegetale etilene.[46]
La produzione commerciale di amminoacidi si basa solitamente su batteri mutanti che avviano una sovrapproduzione di singoli amminoacidi utilizzando il glucosio come fonte di carbonio. Alcuni amminoacidi sono prodotti mediante conversioni enzimatiche di intermedi sintetici. L’acido 2-amminotiazolin-4-carbossilico è un intermedio in una sintesi industriale della L-cisteina, ad esempio. L’acido aspartico viene prodotto mediante l’aggiunta di ammoniaca al fumarato utilizzando una liasi.[47]
Presenza e funzioni in biochimica
Gli amminoacidi sono i precursori delle proteine[25] e si uniscono tramite reazioni di condensazione per formare catene polimeriche corte chiamate peptidi o catene più lunghe chiamate polipeptidi o proteine. Queste catene sono lineari e non ramificate, con ogni residuo di amminoacido all’interno della catena attaccato a due amminoacidi vicini. In natura, il processo di creazione delle proteine codificate dal materiale genetico DNA/RNA è chiamato traduzione e comporta l’aggiunta graduale di aminoacidi a una catena proteica in crescita da parte di un ribozima chiamato ribosoma.[48] L’ordine di aggiunta degli aminoacidi viene letto attraverso il codice genetico da un modello di mRNA, che è una copia di RNA di uno dei geni dell’organismo.
Struttura Proteina Primaria
Ventidue amminoacidi sono naturalmente incorporati nei polipeptidi e sono chiamati amminoacidi proteinogenici o naturali.[27] Di questi, 20 sono codificati dal codice genetico universale. Gli altri due, la selenocisteina e la pirrolisina, sono incorporati nelle proteine mediante meccanismi sintetici unici. La selenocisteina viene incorporata quando l’mRNA da tradurre include un elemento SECIS, che fa sì che il codone UGA codifichi la selenocisteina invece di un codone di stop.[49] La pirrolisina è utilizzata da alcuni archei metanogeni negli enzimi che usano per produrre metano. È codificata con il codone UAG, che in altri organismi è normalmente un codone di stop.[50] Questo codone UAG è seguito da una sequenza a valle PYLIS.[51]
β-Alanine e il suo isomero α-Alanine
I 20 aminoacidi codificati direttamente dai codoni del codice genetico universale sono chiamati aminoacidi standard o canonici. Una forma modificata di metionina (N-formilmetionina) è spesso incorporata al posto della metionina come aminoacido iniziale delle proteine nei batteri, nei mitocondri e nei cloroplasti. Altri aminoacidi sono chiamati non standard o non canonici. La maggior parte degli amminoacidi non standard sono anche non proteinogenici (cioè non possono essere incorporati nelle proteine durante la traduzione), ma due di essi sono proteinogenici, in quanto possono essere incorporati a livello di traduzione nelle proteine sfruttando informazioni non codificate nel codice genetico universale.
L’aminoacido Selenocisteina
I due aminoacidi non standard proteinogenici sono la selenocisteina (presente in molti non eucarioti e nella maggior parte degli eucarioti, ma non codificata direttamente dal DNA) e la pirrolisina (presente solo in alcuni archei e in almeno un batterio). L’incorporazione di questi aminoacidi non standard è rara. Ad esempio, 25 proteine umane includono la selenocisteina nella loro struttura primaria,[52] e gli enzimi strutturalmente caratterizzati (selenoenzimi) impiegano la selenocisteina come moiety catalitica nei loro siti attivi.[53] La pirrolisina e la selenocisteina sono codificate tramite codoni varianti. Ad esempio, la selenocisteina è codificata dal codone di stop e dall’elemento SECIS.[54][55][56]
La N-formilmetionina (che è spesso l’amminoacido iniziale delle proteine nei batteri, nei mitocondri e nei cloroplasti) è generalmente considerata una forma di metionina piuttosto che un amminoacido proteinogenico separato. Le combinazioni codone-tRNA non presenti in natura possono anche essere utilizzate per “espandere” il codice genetico e formare nuove proteine note come alloproteine che incorporano aminoacidi non proteinogenici.[57][58][59]
Oltre ai 22 aminoacidi proteinogenici, sono noti molti aminoacidi non proteinogenici. Questi non si trovano nelle proteine (ad esempio la carnitina, il GABA, la levotiroxina) o non sono prodotti direttamente e isolatamente dai macchinari cellulari standard. Ad esempio, l’idrossiprolina viene sintetizzata dalla prolina. Un altro esempio è la selenometionina).
Gli aminoacidi non proteici che si trovano nelle proteine si formano tramite modificazioni post-traslazionali. Tali modifiche possono anche determinare la localizzazione della proteina, ad esempio l’aggiunta di lunghi gruppi idrofobici può far sì che una proteina si leghi a una membrana fosfolipidica.[60] Esempi:
la carbossilazione del glutammato permette di legare meglio i cationi di calcio,[61] L’idrossiprolina, generata dall’idrossilazione della prolina, è uno dei principali componenti del collagene del tessuto connettivo[62]. L’ipusina nel fattore di iniziazione della traduzione EIF5A contiene una modifica della lisina.[63] Alcuni aminoacidi non proteici non si trovano nelle proteine. Ne sono un esempio l’acido 2-amminoisobutirrico e il neurotrasmettitore acido gamma-amminobutirrico. Gli aminoacidi non proteinogenici sono spesso presenti come intermedi nelle vie metaboliche degli aminoacidi standard – ad esempio, l’ornitina e la citrullina sono presenti nel ciclo dell’urea, parte del catabolismo degli aminoacidi (vedi sotto).[64] Una rara eccezione alla predominanza degli α-amminoacidi in biologia è rappresentata dal β-amminoacido beta alanina (acido 3-amminopropanoico), utilizzato nelle piante e nei microrganismi nella sintesi dell’acido pantotenico (vitamina B5), un componente del coenzima A.[65]
Gli aminoacidi non sono una componente tipica del cibo: gli animali mangiano proteine. La proteina viene scomposta in aminoacidi durante il processo di digestione. Vengono quindi utilizzati per sintetizzare nuove proteine, altre biomolecole o vengono ossidati in urea e anidride carbonica come fonte di energia.[66] La via dell’ossidazione inizia con la rimozione del gruppo amminico da parte di una transaminasi; il gruppo amminico viene quindi immesso nel ciclo dell’urea. L’altro prodotto della transamidazione è un chetoacido che entra nel ciclo dell’acido citrico.[67] Gli amminoacidi glucogeni possono anche essere convertiti in glucosio, attraverso la gluconeogenesi.[68]
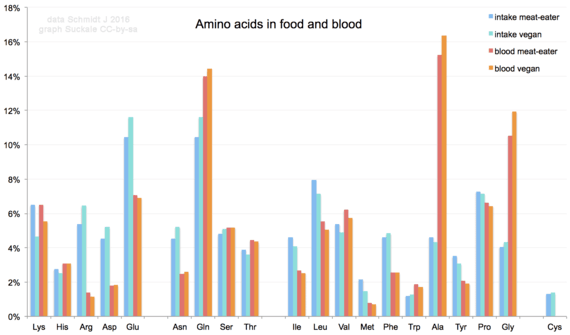
Quota di aminoacidi nelle varie diete umane e miscela risultante di aminoacidi nel siero del sangue umano. Glutammato e glutammina sono i più frequenti negli alimenti (oltre il 10%), mentre alanina, glutammina e glicina sono i più comuni nel sangue.
Dei 20 aminoacidi standard, nove (His, Ile, Leu, Lys, Met, Phe, Thr, Trp e Val) sono chiamati aminoacidi essenziali perché il corpo umano non è in grado di sintetizzarli da altri composti al livello necessario per la normale crescita. quindi devono essere ottenuti dal cibo.[69][70][71]
Inoltre, la cisteina, la tirosina e l’arginina sono considerati aminoacidi semiessenziali e la taurina un acido aminosolfonico semiessenziale nei bambini. Alcuni aminoacidi sono condizionatamente essenziali per determinate età o condizioni mediche. Le vie metaboliche che sintetizzano questi monomeri non sono completamente sviluppate.[72][73]
Molti amminoacidi proteinogenici e non proteogenici hanno funzioni biologiche oltre ad essere precursori di proteine e peptidi. Negli esseri umani, gli amminoacidi hanno anche ruoli importanti in diverse vie biosintetiche. Le difese contro gli erbivori nelle piante a volte impiegano gli aminoacidi.[74] Esempi:
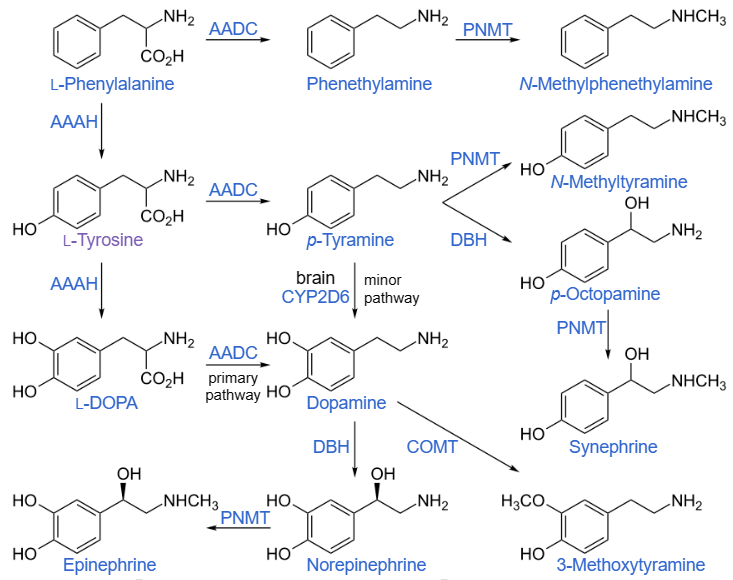
Amminoacidi standard – Il triptofano è un precursore del neurotrasmettitore serotonina.[75] – La tirosina (e il suo precursore fenilalanina) sono precursori dei neurotrasmettitori catecolaminici dopamina, epinefrina e norepinefrina e di varie ammine in traccia. – La fenilalanina è un precursore della fenetilammina e della tirosina nell’uomo. – Nelle piante è un precursore di vari fenilpropanoidi, importanti nel metabolismo vegetale. – La glicina è un precursore delle porfirine come l’eme.[76] – L’arginina è un precursore dell’ossido nitrico.[77] – L’ornitina e la S-adenosilmetionina sono precursori delle poliammine.[78] – Aspartato, glicina e glutammina sono precursori dei nucleotidi.[79] Tuttavia, non tutte le funzioni di altri abbondanti aminoacidi non standard sono note.
Le catecolamine e le ammine in traccia sono sintetizzate dalla fenilalanina e dalla tirosina negli esseri umani.
Ruoli degli amminoacidi non standard – La carnitina è utilizzata nel trasporto dei lipidi. – L’acido gamma-amminobutirrico è un neurotrasmettitore.[80] – Il 5-HTP (5-idrossitriptofano) è utilizzato per il trattamento sperimentale della depressione.[81] – L-DOPA (L-diidrossifenilalanina) per il trattamento del morbo di Parkinson,[82] – L’eflornitina inibisce l’ornitina decarbossilasi e viene utilizzata nel trattamento della malattia del sonno.[83] – La canavanina, un analogo dell’arginina presente in molti legumi, è un antifeedant, che protegge la pianta dai predatori.[84] – La mimosina, presente in alcuni legumi, è un altro possibile antifeedant.[85] Questo composto è un analogo della tirosina e può avvelenare gli animali che pascolano su queste piante.
Formazione del legame peptidico
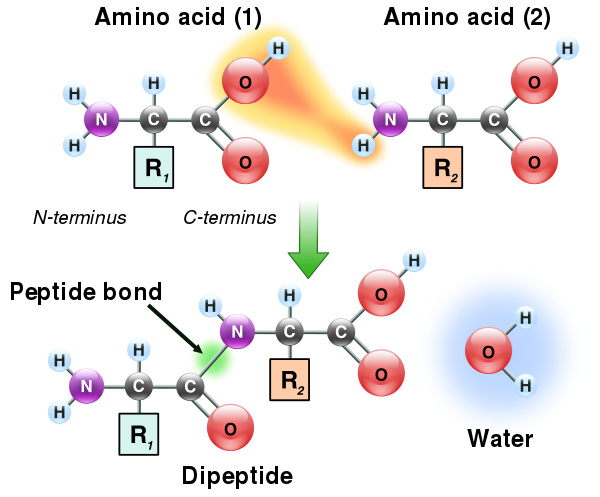
Poiché sia i gruppi amminici che quelli carbossilici degli amminoacidi possono reagire per formare legami ammidici, una molecola di amminoacido può reagire con un’altra e unirsi attraverso un legame ammidico. Questa polimerizzazione degli amminoacidi è ciò che crea le proteine. Questa reazione di condensazione produce il legame peptidico appena formato e una molecola di acqua. Nelle cellule questa reazione non avviene direttamente; invece, l’amminoacido viene prima attivato mediante l’attaccamento a una molecola di RNA di trasferimento attraverso un legame estere. Questo amminoacil-tRNA viene prodotto in una reazione ATP-dipendente effettuata da un’amminoacil tRNA sintetasi.[86] Questo amminoacil-tRNA è quindi un substrato per il ribosoma, che catalizza l’attacco del gruppo amminico della catena proteica allungata sul legame estere.[87] Come risultato di questo meccanismo, tutte le proteine prodotte dai ribosomi vengono sintetizzate a partire dal loro terminale N e spostandosi verso il loro terminale C.
La condensazione di due aminoacidi per formare un dipeptide. I due residui aminoacidici sono legati tramite un legame peptidico
Tuttavia non tutti i legami peptidici si formano in questo modo. In alcuni casi, i peptidi vengono sintetizzati da enzimi specifici. Ad esempio, il tripeptide glutatione è una parte essenziale delle difese delle cellule contro lo stress ossidativo. Questo peptide viene sintetizzato in due fasi da amminoacidi liberi.[88] Nella prima fase, la gamma-glutamilcisteina sintetasi condensa la cisteina e il glutammato attraverso un legame peptidico formato tra il carbossile della catena laterale del glutammato (il carbonio gamma di questa catena laterale) e il gruppo amminico della cisteina. Questo dipeptide viene quindi condensato con la glicina dalla glutatione sintetasi per formare glutatione.[89]
In chimica, i peptidi vengono sintetizzati mediante una varietà di reazioni. Uno dei metodi più utilizzati nella sintesi peptidica in fase solida utilizza i derivati ossimici aromatici degli amminoacidi come unità attivate. Questi vengono aggiunti in sequenza sulla catena peptidica in crescita, che è attaccata a un supporto di resina solida.[90] Le librerie di peptidi vengono utilizzate nella scoperta di farmaci attraverso lo screening ad alto rendimento.[91]
Catabolismo degli aminoacidi proteinogenici. Gli aminoacidi possono essere classificati in base alle proprietà dei loro principali prodotti di degradazione:[139] Glucogenico, con prodotti che hanno la capacità di formare glucosio mediante gluconeogenesi Chetogenico, poiché i prodotti non hanno la capacità di formare glucosio. Questi prodotti possono ancora essere utilizzati per la chetogenesi o la sintesi lipidica. *Amminoacidi catabolizzati sia in prodotti glucogeni che chetogenici.
La combinazione di gruppi funzionali consente agli amminoacidi di essere efficaci ligandi polidentati per chelati metallo-amminoacidi.[92] Le molteplici catene laterali degli amminoacidi possono anche subire reazioni chimiche.
La degradazione di un amminoacido spesso comporta la deaminazione spostando il suo gruppo amminico nell’α-chetoglutarato, formando glutammato. Questo processo coinvolge le transaminasi, spesso le stesse utilizzate nell’amminazione durante la sintesi. In molti vertebrati il gruppo amminico viene poi eliminato attraverso il ciclo dell’urea ed escreto sotto forma di urea. Tuttavia, la degradazione degli aminoacidi può invece produrre acido urico o ammoniaca. Ad esempio, la serina deidratasi converte la serina in piruvato e ammoniaca.[56] Dopo la rimozione di uno o più gruppi amminici, il resto della molecola può talvolta essere utilizzato per sintetizzare nuovi amminoacidi, oppure può essere utilizzato per produrre energia entrando nella glicolisi o nel ciclo dell’acido citrico, come dettagliato nell’immagine a destra.
Valutazione del contenuto di Azoto nella materia organica
Il contenuto totale di azoto della materia organica è formato principalmente dai gruppi amminici delle proteine. L’Azoto Totale Kjeldahl (TKN) è una misura dell’azoto ampiamente utilizzata nell’analisi di acque (reflue), suolo, alimenti, mangimi e materia organica in generale. Come suggerisce il nome, viene applicato il metodo Kjeldahl. Sono disponibili metodi più sensibili.[93][94]
Da sinistra: Digestione Kjeldahl e Distillazione Kjeldahl [Metodo Kjeldahl].
Continua…
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Nelson DL, Cox MM (2005). Principles of Biochemistry (4th ed.). New York: W. H. Freeman. ISBN0-7167-4339-6.
Richard Cammack, ed. (2009). “Newsletter 2009”. Biochemical Nomenclature Committee of IUPAC and NC-IUBMB. Pyrrolysine. Archived from the original on 12 September 2017. Retrieved 16 April 2012.
Wollaston WH (1810). “On cystic oxide, a new species of urinary calculus”. Philosophical Transactions of the Royal Society. 100: 223–230. doi:10.1098/rstl.1810.0015. S2CID110151163.
Braconnot HM (1820). “Sur la conversion des matières animales en nouvelles substances par le moyen de l’acide sulfurique”. Annales de Chimie et de Physique. 2nd Series. 13: 113–125.
Jakubke HD, Sewald N (2008). “Amino acids”. Peptides from A to Z: A Concise Encyclopedia. Germany: Wiley-VCH. p. 20. ISBN9783527621170. Archived from the original on 17 May 2016. Retrieved 5 January 2016 – via Google Books.
Michal G, Schomburg D, eds. (2012). Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology (2nd ed.). Oxford: Wiley-Blackwell. p. 5. ISBN978-0-470-14684-2.
Brønsted, J. N. (1923). “Einige Bemerkungen über den Begriff der Säuren und Basen” [Remarks on the concept of acids and bases]. Recueil des Travaux Chimiques des Pays-Bas. 42 (8): 718–728. doi:10.1002/recl.19230420815.
Urry DW (2004). “The change in Gibbs free energy for hydrophobic association: Derivation and evaluation by means of inverse temperature transitions”. Chemical Physics Letters. 399 (1–3): 177–183. Bibcode:2004CPL…399..177U. doi:10.1016/S0009-2614(04)01565-9.
Marcotte EM, Pellegrini M, Yeates TO, Eisenberg D (October 1999). “A census of protein repeats”. Journal of Molecular Biology. 293 (1): 151–60. doi:10.1006/jmbi.1999.3136. PMID10512723.
Haerty W, Golding GB (October 2010). Bonen L (ed.). “Low-complexity sequences and single amino acid repeats: not just “junk” peptide sequences”. Genome. 53 (10): 753–62. doi:10.1139/G10-063. PMID20962881.
Magee T, Seabra MC (April 2005). “Fatty acylation and prenylation of proteins: what’s hot in fat”. Current Opinion in Cell Biology. 17 (2): 190–196. doi:10.1016/j.ceb.2005.02.003. PMID15780596.
Pilobello KT, Mahal LK (June 2007). “Deciphering the glycocode: the complexity and analytical challenge of glycomics”. Current Opinion in Chemical Biology. 11 (3): 300–305. doi:10.1016/j.cbpa.2007.05.002. PMID17500024.
Smotrys JE, Linder ME (2004). “Palmitoylation of intracellular signaling proteins: regulation and function”. Annual Review of Biochemistry. 73 (1): 559–587.
Kivirikko KI, Pihlajaniemi T (1998). “Collagen Hydroxylases and the Protein Disulfide Isomerase Subunit of Prolyl 4-Hydroxylases”. Advances in Enzymology and Related Areas of Molecular Biology. Advances in Enzymology – and Related Areas of Molecular Biology. Vol. 72. pp. 325–398. doi:10.1002/9780470123188.ch9. ISBN9780470123188. PMID9559057.
Whitmore L, Wallace BA (May 2004). “Analysis of peptaibol sequence composition: implications for in vivo synthesis and channel formation”. European Biophysics Journal. 33 (3): 233–237. doi:10.1007/s00249-003-0348-1. PMID14534753. S2CID24638475.
Krzycki JA (December 2005). “The direct genetic encoding of pyrrolysine”. Current Opinion in Microbiology. 8 (6): 706–712. doi:10.1016/j.mib.2005.10.009. PMID16256420.
Théobald-Dietrich A, Giegé R, Rudinger-Thirion J (2005). “Evidence for the existence in mRNAs of a hairpin element responsible for ribosome dependent pyrrolysine insertion into proteins”. Biochimie. 87 (9–10): 813–817.
Elzanowski A, Ostell J (7 April 2008). “The Genetic Codes”. National Center for Biotechnology Information (NCBI). Archived from the original on 20 August 2016. Retrieved 10 March 2010.
Xie J, Schultz PG (December 2005). “Adding amino acids to the genetic repertoire”. Current Opinion in Chemical Biology. 9 (6): 548–554. doi:10.1016/j.cbpa.2005.10.011. PMID16260173.
Blenis J, Resh MD (December 1993). “Subcellular localization specified by protein acylation and phosphorylation”. Current Opinion in Cell Biology. 5 (6): 984–989. doi:10.1016/0955-0674(93)90081-Z. PMID8129952.
Curis E, Nicolis I, Moinard C, Osowska S, Zerrouk N, Bénazeth S, Cynober L (November 2005). “Almost all about citrulline in mammals”. Amino Acids. 29 (3): 177–205. doi:10.1007/s00726-005-0235-4. PMID16082501. S2CID23877884.
Coxon KM, Chakauya E, Ottenhof HH, Whitney HM, Blundell TL, Abell C, Smith AG (August 2005). “Pantothenate biosynthesis in higher plants”. Biochemical Society Transactions. 33 (Pt 4): 743–746. doi:10.1042/BST0330743. PMID16042590.
Lourenço R, Camilo ME (2002). “Taurine: a conditionally essential amino acid in humans? An overview in health and disease”. Nutricion Hospitalaria. 17 (6): 262–270.
Kostrzewa RM, Nowak P, Kostrzewa JP, Kostrzewa RA, Brus R (March 2005). “Peculiarities of L-DOPA treatment of Parkinson’s disease”. Amino Acids. 28 (2): 157–164. doi:10.1007/s00726-005-0162-4. PMID15750845. S2CID33603501.
Heby O, Persson L, Rentala M (August 2007). “Targeting the polyamine biosynthetic enzymes: a promising approach to therapy of African sleeping sickness, Chagas’ disease, and leishmaniasis”. Amino Acids. 33 (2): 359–366. doi:10.1007/s00726-007-0537-9. PMID17610127. S2CID26273053.
Carpino LA (1992). “1-Hydroxy-7-azabenzotriazole. An efficient peptide coupling additive”. Journal of the American Chemical Society. 115 (10): 4397–4398. doi:10.1021/ja00063a082.
Marasco D, Perretta G, Sabatella M, Ruvo M (October 2008). “Past and future perspectives of synthetic peptide libraries”. Current Protein & Peptide Science. 9 (5): 447–467. doi:10.2174/138920308785915209. PMID18855697.
Konara S, Gagnona K, Clearfield A, Thompson C, Hartle J, Ericson C, Nelson C (2010). “Structural determination and characterization of copper and zinc bis-glycinates with X-ray crystallography and mass spectrometry”. Journal of Coordination Chemistry. 63 (19): 3335–3347.
Martin PD, Malley DF, Manning G, Fuller L (2002). “Determination of soil organic carbon and nitrogen at thefield level using near-infrared spectroscopy”. Canadian Journal of Soil Science. 82 (4): 413–422.
Da quando esistono gli eventi sportivi, il desiderio di ottenere un vantaggio competitivo è sempre stato presente. Con gli enormi incentivi finanziari e le conseguenti pressioni per eccellere associate all’industria sportiva internazionale, i tentativi di ottenere un vantaggio competitivo, in particolare con l’uso di farmaci che migliorano le prestazioni, sono aumentati (Barroso et al., 2008). Nonostante le notizie sull’uso di sostanze per migliorare le prestazioni atletiche risalgano a secoli fa, i test sugli atleti per verificare l’uso di farmaci che migliorano le prestazioni sono iniziati, almeno nel blocco occidentale, solo nel 1968 (Barroso et al., 2008; Botrè, 2008). Da allora, il Comitato Olimpico Internazionale (CIO) e l’Agenzia Mondiale Antidoping (WADA) hanno costantemente aggiornato un elenco di sostanze vietate. I composti e i metodi inclusi nella lista sono quelli che possono essere utilizzati da un atleta per ottenere un vantaggio sleale (WADA, 2009b). Le sostanze presenti nella Lista proibita includono steroidi androgeni anabolizzanti, glucocorticosteroidi, ormoni peptidici e loro modulatori, antagonisti ormonali e loro modulatori, stimolanti, β2-agonisti, narcotici, alcol, β-bloccanti, cannabinoidi, diuretici e agenti mascheranti (WADA, 2009b). L’obiettivo di questo articolo è di rivedere la farmacologia dei diuretici e le applicazioni dei diuretici al doping sportivo, oltre a descrivere in dettaglio le metodologie analitiche attualmente descritte per rilevare e identificare i diuretici nelle urine. Tutte le classi di diuretici (descritte in dettaglio più avanti nel presente articolo) sono vietate nello sport competitivo.
I diuretici sono agenti terapeutici utilizzati per aumentare la velocità del flusso urinario e l’escrezione di sodio al fine di regolare il volume e la composizione dei liquidi corporei o di eliminare i liquidi in eccesso dai tessuti (Jackson, 2006). Sono utilizzati nella terapia clinica per il trattamento di varie malattie e sindromi, tra cui ipertensione, insufficienza cardiaca, cirrosi epatica, insufficienza renale, malattie renali e polmonari, oltre che per una più generale riduzione degli effetti negativi della ritenzione di sali e/o acqua (Jackson, 2006). I diuretici sono stati vietati per la prima volta nello sport (sia in gara che fuori) nel 1988 perché possono essere utilizzati dagli atleti per due motivi principali. In primo luogo, la loro potente capacità di rimuovere l’acqua dal corpo può causare una rapida perdita di peso che può essere necessaria per raggiungere una categoria di peso negli eventi sportivi. In secondo luogo, possono essere utilizzati per mascherare la somministrazione di altri agenti dopanti, riducendo la loro concentrazione nelle urine soprattutto grazie all’aumento del volume di queste ultime. L’effetto di diluizione delle urine dei diuretici permette di classificarli come agenti mascheranti e ne preclude l’uso sia in gara che fuori. Alcuni diuretici provocano un effetto mascherante anche alterando il pH urinario e inibendo l’escrezione passiva di farmaci acidi e basici nelle urine (Ventura e Segura, 1996; Goebel et al., 2004; Trota e Kazlauskas, 2004; Furlanello et al., 2007).
Nel 2008, i diuretici hanno rappresentato il 7,9% di tutti i risultati analitici avversi segnalati dai laboratori WADA, con un numero totale di 436 casi (WADA, 2009a). Tutte le classi di diuretici erano rappresentate nei casi positivi; l’idroclorotiazide è stato il diuretico più comunemente rilevato, con 137 casi. La Tabella 1 riassume le statistiche dei risultati positivi ai diuretici di tutti i laboratori WADA dal 2003 al 2009. In tutti e sei gli anni, tutte le classi di diuretici sono state rappresentate nei risultati positivi (WADA, 2004; 2005; 2006; 2007; 2008a; 2009a;). Nel corso degli anni, il numero totale di casi è aumentato; questa tendenza all’aumento dei risultati positivi può essere dovuta non solo a un aumento dell’abuso, ma anche al miglioramento dei metodi di rilevamento.
Tabella 1.
Sebbene l’applicazione principale dei diuretici sia quella di aumentare l’escrezione renale di sale e acqua, i loro effetti non si limitano al sodio e al cloruro; possono anche influenzare l’assorbimento e l’escrezione renale di altri cationi (K+, H+, Ca2+ e Mg2+), anioni (Cl-, HCO3- e H2PO4-) e acido urico (Jackson, 2006). Questa classe farmacologica di farmaci comprende composti con diverse proprietà farmacologiche e fisico-chimiche. Data la varietà dei composti diuretici, la classificazione di questi farmaci può basarsi su diversi criteri. Le categorie di classificazione più comuni sono: sito d’azione nel nefrone, efficacia relativa, struttura chimica, effetti sull’escrezione di potassio, somiglianza con altri diuretici e meccanismo d’azione (Jackson, 2006). Nella sezione seguente, questo articolo riassumerà brevemente la farmacologia e la tossicologia delle classi di diuretici in base al meccanismo d’azione. La Figura 1 mostra esempi di strutture diuretiche raggruppate per meccanismo d’azione: inibitori dell’anidrasi carbonica (CA), inibitori del simporter Na+/K+/2Cl- (diuretici dell’ansa), inibitori del simporter Na+/Cl- (diuretici tiazidici e simil-tiazidici), diuretici osmotici, inibitori dei canali Na+ dell’epitelio renale (alcuni diuretici risparmiatori di potassio) e antagonisti del recettore mineralcorticoide (MR); si noti la varietà delle strutture molecolari. La Figura 2 illustra in dettaglio il sito e il meccanismo delle classi di diuretici nel nefrone (Figura 2A).
Figura 1 Esempi di strutture diuretiche raggruppate per meccanismo d’azione. (A) inibitori dell’anidrasi carbonica; (B) inibitori del simpatizzante Na+/K+/2Cl- (diuretici dell’ansa); (C) inibitori del simpatizzante Na+/Cl- (diuretici tiazidici e simil-tiazidici); (D) diuretici osmotici; (E) inibitori dei canali Na+ dell’epitelio renale (alcuni diuretici risparmiatori di potassio); (F) antagonisti del recettore mineralcorticoide (MR) (antagonisti dell’aldosterone e alcuni diuretici risparmiatori di potassio).Figura 2 Sito e meccanismo d’azione dei diuretici. (A) Il nefrone con le principali divisioni etichettate. (B) Meccanismo degli inibitori dell’anidrasi carbonica nel tubulo prossimale. (C) Meccanismo degli inibitori del simpatizzante Na+/K+/2Cl- nel tratto ascendente spesso dell’ansa di Henle. (D) Meccanismo degli inibitori del simpatizzante Na+/Cl- nel tubulo distale. (E) Meccanismo degli inibitori dei canali Na+ dell’epitelio renale e degli antagonisti dei recettori mineralcorticoidi nel dotto collettore. Aldo, aldosterone; CA, anidrasi carbonica; MR, recettore dei mineralocorticoidi. Figura modificata da Jackson (2006).
L’identificazione e la quantificazione dei composti proibiti e/o dei loro prodotti metabolici è stato un compito importante nei test antidoping sportivi (Cowan e Kicman, 1997). Storicamente, la rilevazione dei diuretici nei campioni biologici è stata ottenuta utilizzando la cromatografia liquida ad alte prestazioni (HPLC) con rilevazione a raggi ultravioletti (UV-DAD). Tuttavia, la tecnica di rilevamento HPLC-DAD non è specifica per l’identificazione inequivocabile delle sostanze. Pertanto, per la conferma è necessaria la metodologia dello spettro di massa, secondo le normative antidoping internazionali (Trout e Kazlauskas, 2004; Thevis e Schanzer, 2007; WADA, 2009c). La gascromatografia/spettrometria di massa (GC/MS), dopo un’adeguata preparazione e derivatizzazione del campione, è stata, nell’ultimo decennio, la tecnica analitica più utilizzata per la rilevazione dei diuretici. Recentemente, tuttavia, a causa dell’eterogeneità delle strutture chimiche e delle proprietà fisico-chimiche dei diuretici e dell’avvento di una strumentazione più economica, si è diffuso l’uso della cromatografia liquida/MS (LC/MS) (Thevis e Schanzer, 2007). La preparazione del campione prima dell’analisi LC/MS è più semplice rispetto alla GC/MS e non è necessaria alcuna derivatizzazione. Ventura e Segura hanno pubblicato una revisione completa dell’analisi dei diuretici nel 1996 (Ventura e Segura, 1996). Questo articolo si concentrerà principalmente sugli sviluppi e sulle tecniche che sono state sviluppate da allora.
Farmacologia e tossicologia dei diuretici:
Inibitori dell’Anidrasi Carbonica
Gli Inibitori dell’Anidrasi Carbonica (Figura 1A) sono per definizione una classe di sostanze che agiscono come inibitori della CA (carbonato deidratasi, carbonato idrolasi, E.C.4.2.1.1) nelle cellule del tubulo prossimale del nefrone (Figura 2B). La CA è un metalloenzima di zinco espresso nell’uomo come una famiglia di almeno 15 isoenzimi (Tashian, 2000), quattro dei quali (CA II, CA IV, CA XII e CA XIV) sono presenti nel rene (Schwartz, 2002). La CA di tipo II, l’isoenzima più potente, rappresenta il 95% della CA totale nel rene e si trova come proteina solubile nel citoplasma. La CA di tipo IV, un isoenzima legato alla membrana, si trova nelle membrane luminali e basolaterali. Questo enzima svolge un ruolo chiave nel riassorbimento del bicarbonato e nella secrezione acida nel nefrone, catalizzando reversibilmente la reazione di idratazione della CO2 con la produzione di ioni H+ e bicarbonato. Sia la CA II che la CA IV sono inibite dai sulfamidici, in particolare dai sulfamidici aromatici con il gruppo funzionale -SO2NH2 non sostituito. La ridotta capacità di scambiare Na+ con H+ in presenza di questi diuretici determina una debole azione diuretica. Inoltre, il bicarbonato viene trattenuto nel lume con conseguente aumento del pH urinario a circa 8 e successivo sviluppo di un’acidosi metabolica. Anche l’escrezione di fosfato viene aumentata con un meccanismo non del tutto chiarito. L’escrezione di Ca2+ e Mg2+ non viene influenzata.
Secrezione H+ a livello del dotto collettore. Effetto netto: Riassorbimento di NaHCO3 ed H2O.
Acetazolamide
Attualmente sono disponibili tre principali inibitori della CA come diuretici (si veda la Figura 1A per le strutture): l’Acetazolamide (il prototipo della classe, una Sulfonamide senza attività antibatterica), la Diclorfenamide e la Metazolamide. Tutti mostrano una biodisponibilità orale del 100% con un’emivita di 6-14 ore. L’Acetazolamide e la Diclorfenamide sono escrete dai reni come farmaci intatti, mentre la Metazolamide è ampiamente metabolizzata. La principale indicazione terapeutica degli inibitori della CA è il glaucoma ad angolo aperto. L’Acetazolamide è spesso utilizzata per la prevenzione del mal di montagna da alta quota (AMS), un effetto patologico dell’alta quota sull’organismo causato dall’esposizione acuta a una bassa pressione parziale di ossigeno in alta quota che può progredire fino all’edema da alta quota (polmonare e cerebrale). (Coote, 1991; Botrè e Botrè, 1993). L’Acetazolamide aumenta l’escrezione di bicarbonato nelle urine, rendendo il sangue più acido e aumentando la ventilazione, favorendo così l’acclimatazione all’alta quota. L’Acetazolamide è utilizzata anche per il trattamento dell’edema. Gli inibitori della CA possono anche essere utilizzati terapeuticamente per il trattamento della ritenzione di liquidi pre-mestruale.
L’anidrasi carbonica è presente in numerosi tessuti extrarenali, tra cui l’occhio, la mucosa gastrica, il pancreas, il sistema nervoso centrale e gli eritrociti. A causa della diversa localizzazione nell’organismo, gli inibitori della CA sono tipicamente utilizzati per indicazioni non diuretiche, come il glaucoma, per diminuire la velocità di formazione dell’umor acqueo e di conseguenza ridurre la pressione intraoculare. È stato dimostrato che la somministrazione topica di dorzolamide, un inibitore della CA che abolisce l’attività enzimatica nel corpo ciliare, non produce alcun effetto diuretico (Mazzarino et al., 2001). Gli inibitori della CA sono utilizzati anche come farmaci antiepilettici, in parte a causa della produzione di acidosi metabolica.
La maggior parte degli effetti avversi, delle controindicazioni e delle interazioni farmacologiche sono conseguenza dell’alcalinizzazione urinaria o dell’acidosi metabolica. Gli effetti avversi, poco frequenti, sono simili a quelli dei sulfamidici. La deviazione dell’ammoniaca di origine renale dall’urina alla circolazione sistemica, la formazione di calcoli e la colica ureterale che causano la precipitazione di sali di fosfato di calcio nelle urine alcaline, il peggioramento dell’acidosi metabolica o respiratoria e la riduzione del tasso di escrezione urinaria di basi organiche deboli sono altri effetti avversi degli inibitori della CA.
L’efficacia degli inibitori della CA come agenti singoli è bassa e l’utilità a lungo termine degli inibitori della CA è spesso compromessa dallo sviluppo di processi di compensazione come l’acidosi metabolica. Inoltre, l’uso continuo di inibitori della CA può comportare una diminuzione dell’effetto terapeutico desiderato. L’acetazolamide ha rappresentato l’1,4% dei risultati positivi per i diuretici nel 2008 (WADA, 2009a).
Inibitori del co-trasportatore Na+/K+/2Cl- (diuretici dell’ansa):
Gli inibitori del co-trasportatore Na+/K+/2Cl- (Figura 1B) sono una classe di diuretici a breve durata d’azione molto potenti che si legano al sito di legame del Cl- situato nel dominio transmembrana del co-trasportatore Na+/K+/2Cl-, che si trova nell’arto ascendente spesso dell’ansa di Henle (Figura 2C). Il blocco della funzione di questo simpatizzante determina una significativa riduzione della capacità del rene di concentrare l’urina e un conseguente aumento significativo dell’escrezione urinaria di Na+ e Cl-. Si verifica anche un marcato aumento dell’escrezione di Ca2+, Mg2+ e K+. Anche l’escrezione di acido urico aumenta con la somministrazione acuta, mentre la somministrazione cronica ha l’effetto opposto.
Furosemide
Gli inibitori del co-trasportatore Na+/K+/2Cl- sono la Furosemide, la Bumetanide, l’Acido Etacrinico, la Torsemide, l’Assosemide, la Piretanide e la Tripamide (strutture illustrate nella Figura 1B). Oltre il 90% dei farmaci si lega alle proteine plasmatiche. Sono assorbiti rapidamente e ampiamente dal tratto gastrointestinale (65-90%), ma hanno un’emivita molto breve (meno di 1 ora per Bumetanide e Piretanide e un massimo di 3,5 ore per la Torsemide). Questi inibitori del simporto subiscono un parziale metabolismo (epatico per Bumetanide e Torsemide, Glucuronazione renale per gli altri) con escrezione renale come farmaci intatti (Shankar e Brater, 2003).
A causa della loro struttura a base di Sulfonamidi, alcuni diuretici dell’ansa hanno una debole attività inibitoria della CA che aumenta ulteriormente l’effetto diuretico di questi farmaci. Inoltre, hanno effetti vascolari diretti (Dormans et al., 1996) che aumentano acutamente la capacità venosa sistemica e riducono la pressione di riempimento del ventricolo sinistro. Questo effetto, particolarmente evidente per la furosemide, giova ai pazienti con edema polmonare anche prima che si verifichi la diuresi.
Una delle principali indicazioni dei diuretici dell’ansa è il trattamento dell’edema polmonare acuto. Vengono utilizzati anche per il trattamento dell’insufficienza cardiaca congestizia cronica. Ciò comporta una significativa riduzione della mortalità, una diminuzione del rischio di peggioramento dell’insufficienza cardiaca e un miglioramento della capacità di esercizio (Faris et al., 2002). I diuretici dell’ansa sono anche ampiamente utilizzati per il trattamento dell’ipertensione (van der Heijden et al., 1998). Gli inibitori del simpatizzatore Na+/K+/2Cl- sono indicati anche nel trattamento dell’edema e dell’ascite della cirrosi epatica, nel trattamento dell’edema della sindrome nefrosica e per l’iponatriemia a rischio di vita.
Gli effetti avversi sono tutti correlati allo squilibrio di liquidi ed elettroliti. Essi comprendono iponatriemia e/o deplezione del volume del liquido extracellulare (associati a ipotensione, collasso circolatorio ed episodi tromboembolici), alcalosi ipocloremica, ipokaliemia (che induce aritmie cardiache), ipomagnesiemia, iperuricemia (che occasionalmente porta alla gotta) e iperglicemia. Inoltre, aumentano i livelli plasmatici di colesterolo e trigliceridi delle lipoproteine a bassa densità, mentre diminuiscono i livelli plasmatici di colesterolo delle lipoproteine ad alta densità. I diuretici ad ansa possono causare ototossicità, soprattutto l’Acido Etacrinico. Questa classe di diuretici presenta interazioni farmacologiche con diverse sostanze, tra cui Aminoglicosidi, anticoagulanti, glicosidi digitalici, Litio, Propranololo, Sulfoniluree, Cisplatino, Probenecid e Amfotericina B. Il sinergismo dell’attività diuretica dei diuretici dell’ansa e dei diuretici tiazidici associati porta a una diuresi profonda.
Nel 2008, gli inibitori del simpatizzatore Na+/K+/2Cl- hanno rappresentato il 24,6% dei campioni positivi al doping diuretico. La furosemide è stata il secondo diuretico più frequentemente rilevato, con 104 campioni positivi (23,9%) (WADA, 2009a).
Inibitori del co-trasportatore Na+/Cl- (tiazidi e tiazidi-simili):
Gli inibitori del simpatizzatore Na+/Cl- (Figura 1C) hanno un’azione diuretica ottimale nel tubulo convoluto distale iniziale e un effetto diuretico minore nel tubulo prossimale. Inoltre, anche alcuni diuretici tiazidici sono deboli inibitori del CA. Riducono il riassorbimento di Na+ attraverso l’inibizione del co-trasporto Na+/Cl- (Figura 2D). Il legame di Na+ o Cl- al simpatizzatore Na+/Cl- modifica l’inibizione del simpatizzatore indotta dai tiazidici, suggerendo che il sito di legame dei tiazidici è condiviso o alterato sia dal Na+ che dal Cl- (Monroy et al., 2000).
Bendroflumethiazide
Alcuni esempi di farmaci appartenenti a questa classe sono i seguenti (si veda la struttura nella Figura 1C): Bendroflumethiazide, Clorotiazide, Idroclorotiazide, Idroflumetiazide, Meticlorotiazide, Politiazide, Triclormetiazide, clortalidone, Indapamide, Metolazone e Chinetazone. In generale, tutti mostrano una buona biodisponibilità dopo somministrazione orale (100% per la Bendroflumetazide e la Politiazide, almeno il 50% per l’Idroflumetiazide e gli altri). Sono parzialmente metabolizzati da vie sconosciute e sono parzialmente escreti come farmaci intatti dal rene. Il legame con le proteine plasmatiche varia notevolmente tra i vari gruppi. Gli ampi intervalli di emivita variano da 1,5 h per la Clorotiazide a quasi 50 h per il Clortalidone.
Sebbene ci si aspetti che questa classe di diuretici aumenti notevolmente l’escrezione di Na+ e Cl-, questo effetto è moderato poiché circa il 90% del Na+ filtrato viene riassorbito prima di raggiungere il tubulo contorto distale. Come i diuretici dell’ansa, gli inibitori del co-trasportatore Na+/Cl- influenzano l’escrezione di K+ e di acido urico con gli stessi meccanismi; l’escrezione di K+ è marcatamente aumentata dopo la somministrazione e l’escrezione di acido urico è aumentata dopo la somministrazione acuta e diminuisce dopo la somministrazione cronica. Tuttavia, diminuiscono l’escrezione di Ca2+ (Friedman e Bushinsky, 1999).
I diuretici tiazidici sono i più utilizzati. Sono impiegati come terapia di prima linea per l’ipertensione, da soli o in combinazione con altri farmaci antipertensivi (Chobanian et al., 2003). Sono utilizzati anche per il trattamento dell’edema associato a malattie cardiache, epatiche e renali. I diuretici tiazidici sono frequentemente utilizzati per il loro basso costo, l’elevata tolleranza, la buona compliance (somministrazione una volta al giorno), le poche controindicazioni, l’efficacia paragonabile a quella di altre classi di agenti antipertensivi e i comprovati benefici nel ridurre la morbilità e la mortalità cardiovascolare.
Anche in questo caso, come per i diuretici dell’ansa, la maggior parte degli effetti avversi degli inibitori del simporto Na+/Cl- sono dovuti ad anomalie dell’equilibrio dei fluidi e degli elettroliti e comprendono: deplezione del volume extracellulare, ipotensione, ipokaliemia (che compromette l’effetto antipertensivo), iponatremia, ipocloremia, alcalosi metabolica, ipomagnesiemia, ipercalcemia, iperuricemia e iperglicemia (il diabete mellito latente può essere smascherato durante la terapia) (Wilcox et al. , 1999). Tuttavia, a differenza dei diuretici dell’ansa, gli inibitori della simporta Na+/Cl- aumentano i livelli plasmatici di colesterolo delle lipoproteine a bassa densità, colesterolo totale e trigliceridi totali e l’incidenza della disfunzione erettile è maggiore.
Le interazioni farmaco-diuretico tiazidico e tiazidico-simile causano una diminuzione dell’effetto degli anticoagulanti, degli agenti uricosurici, delle sulfoniluree e dell’insulina e aumentano gli effetti dovuti al sinergismo d’azione tra anestetici, diazossido, glicosidi digitalici, litio, vitamina D e diuretici dell’ansa.
Gli inibitori del co-trasportatore Na+/Cl- sono stati la classe di diuretici più abusata nel 2008 secondo le statistiche WADA, con il 38,7% dei campioni positivi. L’idroclorotiazide è stato il diuretico più rilevato, trovato nel 31,4% (137) dei campioni positivi (WADA, 2009a).
Diuretici osmotici:
Isosorbide
I diuretici osmotici sono una classe di composti non metabolizzabili a basso peso molecolare. Solo quattro composti sono inclusi in questa classe di diuretici: Glicerina, Isosorbide, Mannitolo e Urea. Le strutture molecolari sono riportate nella Figura 1D. Questi composti sono relativamente inerti dal punto di vista farmacologico, liberamente filtrabili dal glomerulo e non diffondibili attraverso il nefrone. Vengono somministrati in dosi elevate, non solo per via orale (Glicerina, Isosorbide) ma anche per via endovenosa (Mannitolo, Urea). Tale somministrazione aumenta significativamente l’osmolalità del plasma e del fluido tubulare e, a sua volta, provoca un aumento dell’osmolalità delle urine con conseguente riduzione del riassorbimento di acqua nel nefrone distale/dotti collettori. I diuretici osmotici agiscono sia nel tubulo prossimale che nell’ansa di Henle, con quest’ultima come sito d’azione principale. Questi diuretici agiscono anche attraverso un effetto osmotico nei tubuli e riducendo la tonicità midollare. Le emivite variano da meno di 1 ora nel caso della Glicerina e del Mannitolo a quasi 10 ore per l’Isosorbide.
Estraendo acqua dai compartimenti intracellulari, i diuretici osmotici espandono il volume del fluido extracellulare, riducono la viscosità del sangue e inibiscono il rilascio di renina. Ne consegue un aumento dell’escrezione urinaria di tutti gli elettroliti, Na+, K+, Ca2+, Mg2+, Cl-, HCO3- e PO43-. Il loro uso è limitato a situazioni cliniche ben definite; ad esempio, il mannitolo viene utilizzato per ridurre l’edema cerebrale e la massa cerebrale prima e dopo un intervento di neurochirurgia, nella necrosi tubulare acuta come protettore renale (Levinsky e Bernard, 1988) e per il trattamento della sindrome da disequilibrio dialitico. Poiché i diuretici osmotici estraggono acqua dall’occhio e dal cervello, sono tutti utilizzati per controllare la pressione intraoculare durante gli attacchi acuti di glaucoma e nella chirurgia oculare.
La terapia diuretica osmotica può causare ipernatremia e disidratazione a causa della perdita di acqua in eccesso rispetto alla perdita di elettroliti. Al contrario, il loro uso può portare all’iponatriemia, responsabile dei comuni effetti avversi (cefalea, nausea e vomito). L’iperglicemia può verificarsi come conseguenza del metabolismo della glicerina.
Inibitori dei canali Na+ dell’epitelio renale:
Gli inibitori dei canali Na+ dell’epitelio renale (Figura 1E) agiscono nelle cellule del tubulo distale tardivo e del dotto collettore del nefrone inibendo il riassorbimento di Na+ e la secrezione di K+ e H+ (Figura 2E). Il meccanismo molecolare è il blocco dei canali epiteliali del Na+ nella membrana luminale attraverso la competizione con il Na+ per le aree cariche negativamente all’interno del poro del canale del Na+.
Triamterene
Gli unici due farmaci di questa classe in uso clinico sono il Triamterene e l’Amiloride (strutture illustrate anche nella Figura 1E). Entrambi i farmaci mostrano un modesto effetto diuretico da soli e un piccolo aumento dell’escrezione di Na+ e Cl-. In genere, vengono utilizzati in combinazione con altri diuretici per compensare i loro gravi effetti kaliuretici e preservare i livelli di potassio nei pazienti a rischio di ipokaliemia. Nel trattamento dell’edema o dell’ipertensione, la combinazione di un inibitore dei canali del Na+ con un diuretico tiazidico o dell’ansa potenzia l’effetto diuretico e antipertensivo.
Gli inibitori dei canali del Na+ mostrano una bassa biodisponibilità orale e grandi differenze nell’emivita (più di 20 ore per l’amiloride, meno di 5 ore per il triamterene). La via di eliminazione è prevalentemente renale per l’Amiloride intatta, mentre il Triamterene viene ampiamente metabolizzato nel 4-idrossitriamterene solfato attivo ed escreto nelle urine. Gli effetti avversi più comuni degli inibitori dei canali del Na+ sono nausea, vomito, diarrea, cefalea, crampi alle gambe e vertigini. L’effetto avverso più pericoloso degli inibitori dei canali del Na+ è l’iperkaliemia. Il Triamterene può anche ridurre la tolleranza al glucosio e indurre fotosensibilizzazione.
L’Amiloride e il Triamterene sono stati rilevati nel 3% dei campioni positivi al doping diuretico nel 2008 (WADA, 2009a).
Antagonisti dei recettori dei mineralcorticoidi
Gli antagonisti dei recettori dei mineralcorticoidi (Figura 1F) sono inibitori competitivi dell’Aldosterone che si legano e inibiscono gli MR citosolici presenti nelle cellule epiteliali del tubulo distale tardivo e del dotto collettore del nefrone (Figura 2E).
Il MR è un membro della superfamiglia dei recettori nucleari per gli steroidi. Normalmente, l’Aldosterone entra nella cellula epiteliale e si lega ai MR. Il complesso MR-aldosterone trasloca poi nel nucleo dove si lega a specifiche sequenze di DNA (elementi responsivi all’ormone), regolando così l’espressione di molteplici prodotti genici chiamati proteine indotte dall’aldosterone. A differenza del complesso MR-aldosterone, il complesso MR-antagonista non è in grado di indurre la sintesi di proteine indotte dall’aldosterone.
Spironolattone
I composti appartenenti a questa classe (vedi anche Figura 1F per le strutture molecolari) sono, ad esempio, lo Spironolattone, il Canrenone, il Canrenoato di Potassio e l’Eplerenone. La disponibilità orale dello Spironolattone, la molecola prototipo della classe, è di circa il 65%; è ampiamente metabolizzato, subisce un ricircolo enteroepatico, si lega fortemente alle proteine plasmatiche e ha un’emivita breve (circa 1,6 h) (Beermann, 1984). Il Canrenone è un metabolita attivo dello Spironolattone con un’emivita 10 volte superiore (16,5 h) che prolunga l’effetto del composto madre. Il Canrenoato non è attivo, ma viene convertito in Canrenone nell’organismo. L’Eplerenone ha una buona disponibilità orale ed è ampiamente metabolizzato.
Gli antagonisti dei recettori dei mineralcorticoidi hanno effetti sull’escrezione urinaria simili a quelli degli inibitori dei canali Na+ dell’epitelio renale. L’efficacia clinica degli antagonisti dei MR dipende strettamente dai livelli endogeni di Aldosterone; livelli più elevati provocano effetti maggiori.
Questo gruppo di diuretici è molto utile come alternativa alla terapia sostitutiva del potassio. Di solito vengono impiegati in caso di elevate concentrazioni di potassio. Nel trattamento dell’edema e dell’ipertensione, questi farmaci vengono spesso co-somministrati con i diuretici tiazidici o dell’ansa, oltre che con gli altri diuretici risparmiatori di K+. Lo spironolattone è utile nel trattamento dell’iperaldosteronismo primario e dell’edema refrattario associato all’aldosteronismo secondario (Ouzan et al., 2002). Analogamente agli inibitori dei canali del Na+, l’effetto avverso più comune degli antagonisti del MR è l’iperkaliemia.
A causa della sua struttura molecolare (Figura 1F), lo Spironolattone ha una certa affinità per i recettori del Progesterone e degli Androgeni che causa alcuni effetti collaterali come ginecomastia, impotenza e irregolarità mestruali. Al contrario, grazie al gruppo 9,11-epossido, l’Eplerenone ha un’affinità molto bassa per i Recettori del Progesterone e degli Androgeni (<1% e <0,1%, rispettivamente) rispetto allo Spironolattone. La somministrazione cronica di Spironolattone può indurre tumori maligni; in particolare, è stato osservato il cancro al seno. Per quanto riguarda le interazioni farmaco-farmaco, i salicilati riducono la secrezione tubulare di Canrenone e diminuiscono l’efficacia diuretica dello Spironolattone, mentre quest’ultimo altera la clearance dei glicosidi digitalici.
Canrenone e Spironolattone insieme hanno rappresentato il 4,3% dei campioni positivi al doping diuretico nel 2008 (WADA, 2009a).
Diuretici e Sport:
Osservazioni generali
Come già detto, i diuretici sono comunemente prescritti in medicina clinica per il trattamento dell’ipertensione e di altri disturbi cardiovascolari. Questi composti sono anche frequentemente utilizzati in modo illecito nello sport. I diuretici sono vietati in tutti gli sport perché possono causare una rapida perdita di peso e possono agire come agenti mascheranti (per nascondere gli effetti di altre sostanze proibite) sia in gara che fuori. Tuttavia, il Codice Mondiale Antidoping (WADA, 2009f) consente l’uso terapeutico dei diuretici quando gli atleti e i loro medici richiedono un’esenzione per uso terapeutico (TUE) secondo lo Standard Internazionale per le TUE (WADA, 2009d). La TUE è definita come “l’autorizzazione all’uso, a scopo terapeutico, di sostanze o metodi contenuti nella Lista delle sostanze o dei metodi proibiti, ogni volta che viene approvata da un Comitato per l’esenzione dall’uso terapeutico sulla base di un dossier medico documentato prima dell’uso della sostanza nello sport”. Per i diuretici, l’uso terapeutico principale consentito è quello per l’ipertensione (WADA, 2008b). Va notato che una TUE non è valida se l’urina di un atleta contiene un diuretico in associazione a un livello soglia o sotto-soglia di un’altra sostanza esogena inclusa nella Lista proibita. Grazie alla TUE, alcuni atleti fanno uso di diuretici per scopi medici legittimi; in molti casi, tuttavia, l’uso di diuretici è illecito (Clarkson e Thompson, 1997).
DOPING e Diuretici
Ragionevolmente, l’uso più efficace dei diuretici nel doping sportivo sarebbe prima di un test antidoping. I diuretici aumentano il volume delle urine e diluiscono gli agenti dopanti e i loro metaboliti presenti nelle urine, rendendone più problematica l’individuazione da parte delle analisi antidoping convenzionali. Per questo motivo, i diuretici sono classificati come agenti mascheranti nella Lista proibita della WADA (classe S5: “Diuretici e altri agenti mascheranti”) (WADA, 2009b).
Sebbene vi siano poche prove di un miglioramento delle prestazioni atletiche in seguito alla somministrazione di diuretici, il loro abuso è molto diffuso tra gli atleti che vogliono perdere peso rapidamente. Ad esempio, l’uso di diuretici può consentire a un atleta di ridurre transitoriamente il peso corporeo, il che rappresenta un chiaro vantaggio nella lotta, nel pugilato, nel judo e nel sollevamento pesi, nonché negli sport in generale in cui sono coinvolte categorie di peso e tra gli atleti che desiderano mantenere un peso corporeo basso, come le ginnaste e le ballerine. Gli sciatori e gli alpinisti, tuttavia, fanno un uso legittimo dell’acetazolamide (un inibitore della CA che agisce anche su siti diversi dal rene) per prevenire l’AMS.
Come già detto, i diuretici sono vietati nello sport perché possono essere utilizzati: (i) direttamente, per produrre una rapida perdita di peso che può essere fondamentale per raggiungere una categoria di peso negli eventi sportivi; e/o (ii) indirettamente, per alterare il normale profilo di metabolismo/escrezione di altre sostanze dopanti. In entrambi i casi, discussi più dettagliatamente in seguito, la somministrazione di diuretici può essere acuta o cronica, con dosi somministrate che possono superare notevolmente i livelli terapeutici. In generale, gli atleti possono utilizzare i diuretici in una singola dose alcune ore prima di una gara (ad esempio, lottatori o sportivi a scopo di mascheramento) o abusarne cronicamente per mesi (ad esempio, ginnaste). È importante notare che i diuretici di cui gli atleti abusano maggiormente (furosemide, idroclorotiazide e triamterene) hanno un’emivita breve e sono quindi non rilevabili nelle urine se i campioni non vengono raccolti entro 24-48 ore dall’ultima somministrazione.
Diuretici, esercizio fisico e perdita di peso
Nel tentativo di valutare l’importanza dell’uso di diuretici nella perdita di peso, Caldwell et al. (1984) hanno confrontato il diverso effetto della disidratazione acuta indotta dall’esercizio fisico, dalla sauna e dai diuretici sulla variazione di peso. I risultati hanno mostrato una diminuzione di 2,3 ± 0,8 kg dopo l’esercizio fisico, 3,5 ± 0,8 kg dopo la sauna e 3,1 ± 0,8 kg dopo la somministrazione di furosemide. Inoltre, i bodybuilder abusano di diuretici insieme a steroidi androgeno-anabolizzanti per accentuare la definizione muscolare e il tono corporeo. Nello stesso studio riportato da Caldwell et al. è stato dimostrato che la variazione del volume plasmatico negli atleti è pari a -0,9% dopo l’esercizio fisico, -10,3% dopo la sauna e -14,1% dopo la somministrazione di furosemide (quantità totale di 1,7 mg-kg-1 in due dosi divise, 16 ore prima del test) (Caldwell et al., 1984).
Una freccia indica un effetto moderato; due frecce indicano un effetto profondo. GFR, velocità di filtrazione glomerulare; PRA, attività della renina plasmatica; VO2 max, massima captazione di ossigeno.
I diuretici possono avere diversi effetti fisiologici sulla fisiologia dell’esercizio, tra cui effetti sul metabolismo (termoregolazione, omeostasi del potassio), sul sistema cardiovascolare e sul sistema respiratorio [azioni polmonari, assorbimento di ossigeno (VO2)]. La maggior parte degli effetti è legata alle conseguenze della deplezione di volume e dello squilibrio e della deplezione di elettroliti. L’esercizio fisico può influenzare anche l’azione dei diuretici, con conseguenze sia sulla farmacologia che sulla farmacocinetica. A livello del nefrone, l’esercizio fisico può sia integrare che antagonizzare gli effetti dei diuretici. L’esercizio fisico induce acutamente un bilancio idrico negativo e l’esercizio fisico regolare a lungo termine abbassa la pressione sanguigna, aumentando le proprietà farmacologiche dei diuretici (Zappe et al., 1996). L’esercizio fisico influenza anche le azioni specifiche dei diuretici; può causare uno spostamento acuto del potassio intracellulare nello spazio intravascolare (Young et al., 1992) e potenziare l’effetto kaliuretico dei diuretici. Mentre i diuretici tiazidici sono associati all’insulino-resistenza (Moser, 1998), l’esercizio fisico potenzia l’effetto opposto (Plasqui e Westerterp, 2007). Nella maggior parte dei casi, l’esercizio fisico viene utilizzato come terapia per l’insulino-resistenza perché attiva le cellule β pancreatiche attraverso il sistema neuroadrenergico (Bordenave et al., 2008). Questo riduce i livelli di insulina nel sangue e di conseguenza aumenta il rilascio epatico di glucosio e diminuisce l’utilizzo muscolare dell’insulina (Bonen et al., 2006). Sebbene vi siano poche informazioni su come l’esercizio fisico influisca sulla farmacocinetica dei diuretici, clorotiazide, idroclorotiazide e triamterene hanno un’emivita di eliminazione abbastanza breve (1,5-4 ore) da essere influenzata da 1 ora o più di esercizio fisico prolungato (Somani, 1996), che riduce il flusso sanguigno renale ed epatico. Pertanto, queste sostanze non vengono sempre rilevate nei campioni di urina raccolti dopo una gara o al termine di un’intensa sessione di allenamento. È da notare che sia l’esercizio fisico sia i diuretici possono causare indipendentemente la perdita di liquidi ed elettroliti. La Tabella 2, adattata da Caldwell et al. (1984) e Reents (2000), riassume gli effetti dell’esercizio e dei diuretici sulla fisiologia renale.
È noto che durante l’esercizio fisico la temperatura del muscolo scheletrico supera la temperatura interna entro alcuni minuti, e l’alterazione dei sistemi termoregolatori dell’organismo è uno dei rischi principali dell’abuso di diuretici. La marcata disidratazione conseguente all’assunzione di diuretici esercita un effetto dannoso sui sistemi cardiovascolare e termoregolatorio dell’organismo durante l’esercizio e può portare a esaurimento, battito cardiaco irregolare, infarto e morte. È stato dimostrato che sia l’acetazolamide (Brechue e Stager, 1990), un leggero diuretico, sia la furosemide (Claremont et al., 1976), un potente diuretico, compromettono l’aumento adattativo del flusso sanguigno cutaneo durante l’esercizio.
I diuretici influenzano l’omeostasi del potassio nel muscolo in esercizio; il potassio intracellulare e il potenziale di membrana a riposo della cellula diminuiscono entrambi. Tutti i diuretici, tranne gli agenti risparmiatori di potassio, aumentano la kaliuresi, accelerando la deplezione del potassio intracellulare. L’ipokaliemia che ne consegue può portare a crampi muscolari e ad aritmie cardiache secondarie a spostamenti/perdite di elettroliti. D’altra parte, l’uso eccessivo di diuretici risparmiatori di potassio, come lo spironolattone, il triamterene e l’amiloride, può portare all’iperkaliemia e di conseguenza esporre gli atleti ad aritmie maligne (Appleby et al., 1994). Inoltre, l’interferenza della maggior parte dei diuretici con il metabolismo dell’acido urico può causare un attacco di gotta, che può essere molto doloroso (Koutlianos e Kouidi, 2006).
La disidratazione indotta dai diuretici influenza la frequenza cardiaca da sforzo. In particolare, a bassa intensità di esercizio risulta una frequenza cardiaca più elevata, mentre durante lo sforzo massimale l’effetto è minore o quasi assente (Stager et al., 1990). Ciò è particolarmente vero per l’abuso di acetazolamide (Brechue e Stager, 1990) e, in misura minore, di furosemide (Claremont et al., 1976). Studi condotti sugli inibitori della CA e sui diuretici tiazidici hanno dimostrato che dopo la somministrazione di acetazolamide (Brechue e Stager, 1990) o di una combinazione idroclorotiazide-triamterene (Nadel et al., 1980) il volume plasmatico e il volume dell’ictus sono significativamente diminuiti. La perdita di volume plasmatico e di volume del battito interrompe la termoregolazione attraverso la vasodilatazione periferica (raffreddamento per irraggiamento) e la sudorazione (raffreddamento per evaporazione), compromettendo la risposta vasodilatatoria fisiologica sia acuta che a lungo termine all’esercizio aerobico. Inoltre, gli antagonisti dell’aldosterone, in particolare lo spironolattone, interferiscono con l’aumento della sensibilità dei recettori dell’aldosterone dovuto all’ipervolemia indotta dall’esercizio (una conseguenza del normale adattamento all’esercizio fisico regolare).
Effetti aggiuntivi di classi specifiche di diuretici
Poiché la CA svolge un ruolo chiave nei meccanismi di regolazione acido-base, gli inibitori della CA sono l’unica classe di diuretici che può influenzare la funzione polmonare. È stato dimostrato che l’acetazolamide compromette l’eliminazione di CO2 durante l’esercizio fisico (Scheuermann et al., 1999), ma anche l’efflusso di CO2 dal muscolo inattivo (Kowalchuk et al., 1992). Nell’AMS, l’acetazolamide migliora l’ossigenazione alveolare aumentando le pressioni arteriose di ossigeno e abbassando le pressioni arteriose di anidride carbonica (Bradwell et al., 1986). Gli effetti metabolici cellulari dell’acetazolamide possono prevalere sui suoi effetti polmonari e causare un’inibizione del VO2 durante l’esercizio massimale (Stager et al., 1990; Kowalchuk et al., 1992). La furosemide diminuisce il volume tidalico, la ventilazione minima e il rapporto di scambio respiratorio alla soglia aerobica (Caldwell et al., 1984). Al contrario, i dati clinici indicano che la furosemide inalata riduce la broncocostrizione indotta dall’esercizio fisico nei bambini asmatici (Munyard et al., 1995). Gli effetti dei diuretici sul VO2 sono variabili. La furosemide provoca un effetto dose-dipendente; a basse dosi non ha alcuna influenza sul VO2 (Armstrong et al., 1985; Baum et al., 1986), ma il VO2 diminuisce significativamente a dosi più elevate (Caldwell et al., 1984). L’acetazolamide influisce sul VO2 solo durante l’esercizio massimale (Stager et al., 1990; Kowalchuk et al., 1992), poiché il VO2 non è influenzato in condizioni di normossia (Brechue e Stager, 1990), ma è notevolmente migliorato in condizioni di ipossia (Schoene et al., 1983). Gli effetti dell’acetazolamide sulle prestazioni dipendono dall’altitudine; a livello del mare (Heigenhauser et al., 1980) e in condizioni di normossia (Schoene et al., 1983; Stager et al., 1990) può compromettere le prestazioni aerobiche, ma in condizioni di ipossia diminuisce il tempo di esaurimento durante l’esercizio submassimale (Stager et al., 1990).
Infine, i diuretici tiazidici sono derivati dei sulfamidici e possono causare fotosensibilità se si pratica attività fisica all’aperto nelle ore di mezzogiorno.
Caldwell et al. hanno condotto uno studio sulla riduzione del carico di lavoro ciclistico indotta da diuretici per valutare gli effetti dell’ipoidratazione sulle prestazioni al cicloergometro. In questo studio, il VO2 max (massimo assorbimento di ossigeno) e il carico di lavoro in bicicletta diminuiscono negli atleti dopo l’assunzione di furosemide. Anche dopo la reidratazione, la resistenza muscolare e le prestazioni sono notevolmente compromesse dall’uso di diuretici (Caldwell et al., 1984). Ulteriori studi condotti su corridori di media distanza (Armstrong et al., 1985) e lottatori (Caldwell, 1987) hanno confermato che i diuretici riducono gli effetti sulla prestazione atletica complessiva. Sebbene non siano disponibili dati sufficienti per stabilire l’effetto del trattamento diuretico a lungo termine sulla capacità di esercizio, è stato chiaramente dimostrato che il trattamento diuretico a dose singola e a breve termine influisce negativamente sulla capacità di esercizio massimale e sulla durata dell’esercizio submassimale prolungato (Fagard et al., 1993). Per la moltitudine di ragioni sopra menzionate, gli svantaggi legati alla somministrazione di diuretici superano i potenziali vantaggi della riduzione del peso e della diluizione delle urine; la disidratazione compromette drasticamente la capacità aerobica e la forza muscolare e riduce l’efficienza metabolica. Ciò si traduce in un effetto negativo sulla capacità complessiva di praticare sport ed esercizio fisico e soprattutto sulle prestazioni atletiche (Caldwell et al., 1984; Armstrong et al., 1985). Inoltre, un potenziale effetto dell’abuso di diuretici è la possibile alterazione della dimensione della filtrazione glomerulare, che dipende da una serie di parametri (Edwards et al., 1999), la maggior parte dei quali può essere marcatamente influenzata dal meccanismo d’azione delle diverse classi di diuretici. Infine, va notato che la squalifica dalle competizioni e gli altri effetti dannosi precedentemente menzionati dell’abuso di diuretici compensano qualsiasi beneficio percepito.
Sebbene molti degli studi sopra citati siano stati pubblicati negli anni ’80 e ’90, i diuretici sono ancora ampiamente abusati nello sport (e sono tra gli agenti terapeutici più prescritti). Pochi studi sugli effetti dei diuretici sugli atleti sono stati pubblicati di recente, perché negli ultimi tempi la maggior parte degli studi che valutano gli agenti dopanti e l’esercizio fisico e lo sport si sono concentrati su farmaci e metodi di miglioramento delle prestazioni più recenti. L’uso di diuretici per mascherare altre sostanze proibite rimane comunque un problema serio.
Analisi dei diuretici
Osservazioni generali
Per la rilevazione dei diuretici nelle urine nell’ambito del doping sportivo, la WADA ha fissato un unico livello minimo di prestazione richiesto (MRPL) di 250 ng-mL-1 per i laboratori accreditati (WADA, 2009e). Anche se le potenze relative, il metabolismo e le proprietà di eliminazione variano notevolmente (e determinano livelli urinari diversi) tra le classi di diuretici (Tabella 3), l’MRPL di 250 ng-mL-1 è sufficiente per rilevare l’abuso acuto di diuretici da parte degli atleti. È probabile che dosaggi inferiori di diuretici non siano sufficienti a provocare l’effetto di mascheramento o la drastica e acuta perdita di peso ricercata da chi abusa di diuretici.
*La potenza è relativa ai diuretici della stessa classe. NA, dati non disponibili.
Per l’analisi dei diuretici sono state proposte diverse tecniche analitiche, tra cui principalmente HPLC-UV-DAD, GC/MS, LC/MS e LC/MS-MS, cromatografia elettrocinetica micellare ed elettroforesi capillare. Tuttavia, la soluzione migliore per un metodo di screening completo in grado di rilevare la presenza in un campione biologico di qualsiasi diuretico, soddisfacendo al contempo l’MRPL fissato dalla WADA, è rappresentata dai metodi basati su GC/MS, LC/MS e LC/MS-MS. In genere, l’uso di strumentazioni GC/MS, LC/MS e LC/MS-MS consente di rilevare i composti progenitori dei diuretici e/o i metaboliti più diagnostici e abbondanti. Tuttavia, in alcuni casi, l’analita target può non essere il composto progenitore o i suoi metaboliti, ma uno o più prodotti di degradazione formati dopo l’idrolisi dei diuretici in ambiente acquoso. Questo è il caso dei diuretici tiazidici, tra cui soprattutto l’idroclorotiazide e l’altiazide. Questo fenomeno è più rilevante quando c’è un ritardo tra la raccolta del campione e l’analisi di laboratorio (Thieme et al., 2001; Goebel et al., 2004; Deventer et al., 2009).
Negli anni ’80 e ’90, la GC/MS era la tecnica analitica più comunemente utilizzata dai laboratori antidoping per l’analisi degli xenobiotici nelle urine (Maurer, 1992; Hemmersbach e de la Torre, 1996). Storicamente, anche i diuretici venivano analizzati con la GC/MS [ampiamente rivista da Ventura e Segura, 1996 (Ventura & Segura, 1996)]. La recente evoluzione verso la LC/MS (vedi sotto) è stata guidata da una serie di cause concomitanti che rendono l’approccio basato sulla GC/MS meno preferibile rispetto a quello degli ultimi due decenni: (i) il numero di sostanze target, e in particolare di xenobiotici a basso peso molecolare, da sottoporre a screening nelle analisi antidoping è aumentato drasticamente nel periodo 2002-2007, promuovendo lo sviluppo di tecniche analitiche più “universali” volte a ridurre il rapporto risorse/test; (ii) la necessità di semplificare il pretrattamento dei campioni a causa dell’aumento del numero di procedure analitiche eseguite contemporaneamente nei laboratori antidoping; e (iii) i progressi tecnologici nel campo della strumentazione analitica e, più specificamente, la disponibilità di sistemi LC/MS e LC/MS-MS da banco a un prezzo accessibile. Tutti questi eventi hanno favorito il progressivo passaggio dalla GC/MS alla LC/MS.
Gascromatografia/spettrometria di massa:
La gascromatografia/spettrometria di massa è ancora utilizzata da molti laboratori antidoping e può ancora rappresentare una valida alternativa per l’analisi antidoping dei diuretici. Una procedura analitica generale basata sulla GC/MS è strutturata come una serie di fasi di pretrattamento (come minimo: estrazione dei diuretici dalla matrice biologica e derivatizzazione chimica) da eseguire prima della corsa cromatografica.
Pretrattamento del campione Come è noto, l’analisi GC/MS di campioni biologici per lo screening dei diuretici richiede una serie di procedure prestrumentali volte a rendere il campione adatto all’analisi. Fondamentalmente, le fasi critiche sono rappresentate dall’estrazione dei diuretici dalla matrice biologica e dalla derivatizzazione chimica eseguita per aumentare la volatilità e la stabilità termica dei composti target. Sono stati pubblicati diversi metodi per la rilevazione dei diuretici nelle urine utilizzando procedure di estrazione liquido/liquido (L/L) e fase solida (SPE). La SPE può consentire il recupero dei diuretici con rese più elevate, ma allo stesso tempo l’uso di cartucce monouso aumenta il costo complessivo della procedura di pretrattamento, soprattutto nel caso di supporti più complessi, come i supporti a superficie interna in fase inversa (ISRP-size exclusion).
Le colonne pre-attivate disponibili in commercio sono state testate per la loro efficacia e la scelta migliore dovrebbe dipendere dalle caratteristiche della matrice e dalla composizione prevista del campione [rivista da Ventura e Segura nel 1996 (Ventura e Segura, 1996)].
D’altra parte, l’estrazione L/L richiede generalmente più procedure di estrazione. Quando si desidera rilevare tutti i diuretici (basici, acidi e neutri), la soluzione ottimale è un processo basato su due procedure di estrazione L/L separate (una in mezzo neutro o basico e un’altra in mezzo acido) utilizzando acetato di etile o una miscela di solventi organici. È possibile aggiungere solfato di sodio anidro per favorire l’effetto di salatura. Particolare attenzione deve essere dedicata allo studio dei potenziali processi di degradazione che potrebbero coinvolgere i composti target. È stata dimostrata l’ossidazione dei tiazidi (althiazide, benzthiazide e politiazide) in presenza di acetato di etile, pertanto è necessario valutare preliminarmente l’efficacia e la non reattività di diversi solventi di estrazione.
In alcuni casi, due o più fasi di pretrattamento possono essere combinate, come nel caso della metilazione estrattiva in cui sia l’estrazione che la derivatizzazione sono combinate in un’unica procedura.
Procedure di derivatizzazione
Come già detto, la derivatizzazione è necessaria prima dell’analisi GC/MS, poiché la maggior parte dei diuretici non è sufficientemente volatile, lipofila o termicamente stabile per essere analizzata direttamente con questa tecnica analitica. Le procedure di derivatizzazione più comuni sono la sililazione e la metilazione, ma quest’ultima è solitamente preferita in quanto consente di ottenere rese sufficienti di derivati più stabili per la maggior parte dei diuretici [rivisto da Carreras et al. nel 1994 (Carreras et al., 1994)]. La metilazione può essere eseguita “staticamente” (con una miscela di ioduro di metile e acetone sotto riscaldamento termico) o “dinamicamente” mediante metilazione estrattiva (Lisi et al., 1991; Lisi et al., 1992) o metilazione “in colonna” (flash methylation) (Beyer et al., 2005). Quando la metilazione viene eseguita con un processo autonomo, il tempo può essere drasticamente ridotto dall’irradiazione a microonde, in combinazione o in alternativa all’incubazione termica (Amendola et al., 2003).
Condizioni cromatografiche e spettrometriche La fase stazionaria migliore per l’analisi dei composti diuretici è il fenilmetilsilicone, che consente di separare efficacemente tutti i diuretici in tempi ragionevoli (<15 min). Tempi drasticamente più brevi possono essere ottenuti con sistemi fast-GC, in cui vengono accoppiate con successo colonne di ultima generazione e rivelazione spettrometrica di massa basata su un’elettronica veloce. I sistemi Fast-GC consentono di ridurre di 10 volte la durata complessiva della corsa cromatografica (Morra et al., 2006). La ionizzazione a impatto elettronico e la rivelazione MS sono i metodi più descritti [rivisti in Ventura e Segura, 1996 e da Müller et al. nel 1999 (Ventura e Segura, 1996; Müller et al., 1999)]. Gli spettri di massa dei derivati metilici dei diuretici sono stati descritti da diversi autori e i profili di frammentazione sono stati interpretati anche per confronto con i derivati metilici deuterati (Yoon et al., 1990).
Cromatografia liquida/spettrometria di massa
Quando i diuretici sono stati introdotti nell’elenco delle sostanze proibite dalle autorità sportive internazionali, i primi tentativi di creare un metodo di screening per il loro rilevamento si sono basati sull’HPLC. All’epoca, come rivelatore fu utilizzato un diode array UV che facilitava l’identificazione dei picchi (Ventura e Segura, 1996). Secondo i requisiti del CIO/WADA, le procedure di conferma necessarie per sostenere un caso positivo devono basarsi sulla MS. Per questo motivo, nella maggior parte dei casi, la tecnica scelta è stata un metodo GC/MS dopo metilazione dei composti. Per i motivi illustrati nelle sezioni precedenti, alla fine degli anni ’90, quando sono diventati disponibili strumenti LC/MS più robusti, affidabili ed economici, sono stati introdotti importanti cambiamenti nelle strategie di rilevamento dei diuretici nel campo del doping. I primi tentativi di utilizzare la LC/MS per la rilevazione dei diuretici sono iniziati all’inizio degli anni ’90, utilizzando interfacce termospray o a fascio di particelle (Ventura et al., 1991) nelle analisi di conferma. La mancanza di robustezza delle apparecchiature non consentiva un metodo di screening quotidiano basato su questi strumenti.
Thieme et al. (Thieme et al., 2001) hanno descritto un metodo per l’analisi di 32 diuretici nelle urine umane mediante LC/MS/MS utilizzando una tecnica di ionizzazione electrospray. Questa tecnica ha il vantaggio di poter utilizzare le tradizionali velocità di flusso LC e le colonne LC a fase inversa (colonne di ottadecilsilano-ODS con particelle di 5 o 3 µm) solitamente utilizzate nei metodi LC-UV. Inoltre, è possibile utilizzare contemporaneamente le modalità di ionizzazione positiva e negativa, consentendo la rilevazione di composti acidi e basici inclusi tra i diuretici. L’analisi mediante MS tandem con quadrupoli a triplo stadio è risultata sufficientemente selettiva e sensibile rispetto ai metodi precedenti e ha reso possibile la semplificazione della preparazione dei campioni, in quanto la pulizia degli estratti urinari era meno critica rispetto ai metodi LC-UV progettati in precedenza.
Lo sviluppo di nuovi analizzatori (trappole ioniche) accoppiati alla LC ha creato ulteriori alternative per l’analisi dei diuretici mediante LC/MS (Deventer et al., 2002). Ancora più recentemente, la necessità di strategie più universali per l’analisi degli agenti dopanti ha introdotto l’uso di analizzatori time-of-flight (Georgakopoulos et al., 2007) che possono essere accoppiati alla LC. Per alcuni composti e ai fini dell’identificazione, la ionizzazione mediante ionizzazione chimica a pressione atmosferica (un’altra possibile tecnica di ionizzazione delle interfacce LC/MS) è interessante in quanto produce una frammentazione aggiuntiva (Qin et al., 2003).
La selettività e la sensibilità di queste tecniche hanno permesso di includere nelle stesse procedure di screening anche altre droghe non diuretiche, anch’esse vietate nello sport (Deventer et al., 2005; Mazzarino et al., 2008). Inoltre, sono stati esplorati diversi approcci per la preparazione dei campioni. In passato, le classiche doppie estrazioni con solventi organici a pH acido e basico venivano utilizzate per consentire il recupero di diuretici con proprietà fisico-chimiche diverse.
Le nuove caratteristiche degli strumenti e l’estensione dei metodi di screening ad altri composti ampliano le possibilità di preparazione dei campioni. Specifiche procedure SPE possono essere eseguite in sistemi robotici (Goebel et al., 2004) e alcune procedure analitiche non richiedono alcuna preparazione del campione, ma solo una diluizione del campione di urina e la successiva iniezione diretta nel sistema LC/MS (Politi et al., 2007; Thorngren et al., 2008). I miglioramenti nella velocità di scansione degli spettrometri di massa, così come le colonne LC e le pompe LC più performanti, consentono di aumentare la velocità di analisi (UPLC o fast LC) e di effettuare procedure di screening più eterogenee mediante LC/MS/MS. Attualmente, esistono analisi che includono i diuretici tra le altre sostanze dopanti, in cui più di 100 composti diversi possono essere analizzati in meno di 10 minuti (Thorngren et al., 2008; Ventura et al., 2008).
Sintesi e conclusione:
I membri della classe dei farmaci diuretici variano notevolmente per struttura, proprietà fisico-chimiche, sito e meccanismo d’azione. Negli anni ’90 l’analisi dei diuretici nel doping (mediante LC-UV o GC/MS) rappresentava una sfida per i laboratori antidoping a causa dell’eterogeneità delle sostanze incluse. Dall’avvento di strumenti LC/MS consolidati e affidabili, la loro individuazione nei campioni di urina non è più un problema. Gli obiettivi futuri dell’analisi dei diuretici comprendono lo sviluppo di metodi di rilevamento più efficienti ed economici. Aumentare la sensibilità dei metodi e il numero di composti nello screening, riducendo al contempo i tempi e i costi di analisi per i laboratori, sarebbe un miglioramento auspicabile. Inoltre, lo sviluppo di metodi che combinino il rilevamento dei diuretici con altre sostanze proibite migliorerà la capacità dei laboratori di monitorare gli abusi e il doping nello sport.
In conclusione, l’uso dei diuretici, se specificatamente inteso in ambito Bodybuilding, viste anche le tecniche di ratio Sodio/Sale:Acqua, e l’utilizzo di ACE II inibitori per finalità lipolitiche indirette, nonché un adeguato rapporto tra introito di Sodio e Potassio, risulta molto limitato in senso di vantaggi concreti per l’atleta. L’effetto di aumento dell’Aldosterone androgeno-dipendente è facilmente gestibile con altri interventi fermo restando che la presenza di un ACE II inibitore nella preparazione rappresenta di per se un limite sensibile alla manifestazione tangibile del problema.
Amendola L, Colamonici C, Mazzarino M, Botrè F. Rapid determination of diuretics in human urine by gas chromatography – mass spectrometry following microwave assisted derivatization. Anal Chim Acta. 2003;475:125–136. [Google Scholar]
Appleby M, Fisher M, Martin M. Myocardial infarction, hyperkalaemia and ventricular tachycardia in a young male body-builder. Int J Cardiol. 1994;44:171–174. [PubMed] [Google Scholar]
Armstrong LE, Costill DL, Fink WJ. Influence of diuretic-induced dehydration on competitive running performance. Med Sci Sports Exerc. 1985;17:456–461. [PubMed] [Google Scholar]
Barroso O, Mazzoni I, Rabin O. Hormone abuse in sports: the antidoping perspective. Asian J Androl. 2008;10:391–402. [PubMed] [Google Scholar]
Baum K, Essfeld D, Stegemann J. The influence of furosemide on heart rate and oxygen uptake in exercising man. Eur J Appl Physiol Occup Physiol. 1986;55:619–623. [PubMed] [Google Scholar]
Beermann B. Aspects on pharmacokinetics of some diuretics. Acta Pharmacol Toxicol (Copenh) 1984;54(Suppl. 1):17–29. [PubMed] [Google Scholar]
Beyer J, Bierl A, Peters FT, Maurer HH. Screening procedure for detection of diuretics and uricosurics and/or their metabolites in human urine using gas chromatography-mass spectrometry after extractive methylation. Ther Drug Monit. 2005;27:509–520. [PubMed] [Google Scholar]
Bonen A, Dohm GL, van Loon LJ. Lipid metabolism, exercise and insulin action. Essays Biochem. 2006;42:47–59. [PubMed] [Google Scholar]
Bordenave S, Brandou F, Manetta J, Fedou C, Mercier J, Brun JF. Effects of acute exercise on insulin sensitivity, glucose effectiveness and disposition index in type 2 diabetic patients. Diabetes Metab. 2008;34:250–257. [PubMed] [Google Scholar]
Botrè F. New and old challenges of sports drug testing. J Mass Spectrom. 2008;43:903–907. [PubMed] [Google Scholar]
Botrè F, Botrè C. The carbonic anhydrases: are these enzymes still a good deal for a pharmacologist? Curr Top Med Chem. 1993;1:313–347. [Google Scholar]
Bradwell AR, Dykes PW, Coote JH, Forster PJ, Milles JJ, Chesner I, et al. Effect of acetazolamide on exercise performance and muscle mass at high altitude. Lancet. 1986;1:1001–1005. [PubMed] [Google Scholar]
Brechue WF, Stager JM. Acetazolamide alters temperature regulation during submaximal exercise. J Appl Physiol. 1990;69:1402–1407. [PubMed] [Google Scholar]
Caldwell JE, Ahonen E, Nousiainen U. Differential effects of sauna-, diuretic-, and exercise-induced hypohydration. J Appl Physiol. 1984;57:1018–1023. [PubMed] [Google Scholar]
Carreras D, Imaz C, Navajas R, Garcia MA, Rodriguez C, Rodriguez AF, et al. Comparison of derivatization procedures for the determination of diuretics in urine by gas chromatography-mass spectrometry. J Chromatogr A. 1994;683:195–202. [PubMed] [Google Scholar]
Chobanian AV, Bakris GL, Black HR, Cushman WC, Green LA, Izzo JL, et al. Seventh report of the Joint national committee on prevention, detection, evaluation, and treatment of high blood pressure. Hypertension. 2003;42:1206–1252. [PubMed] [Google Scholar]
Claremont AD, Costill DL, Fink W, Van Handel P. Heat tolerance following diuretic induced dehydration. Med Sci Sports. 1976;8:239–243. [PubMed] [Google Scholar]
Clarkson PM, Thompson HS. Drugs and sport. Research findings and limitations. Sports Med. 1997;24:366–384. [PubMed] [Google Scholar]
Coote JH. Pharmacological control of altitude sickness. Trends Pharmacol Sci. 1991;12:450–455. [PubMed] [Google Scholar]
Cowan DA, Kicman AT. Doping in sport: misuse, analytical tests, and legal aspects. Clin Chem. 1997;43:1110–1113. [PubMed] [Google Scholar]
Deventer K, Delbeke FT, Roels K, Van Eenoo P. Screening for 18 diuretics and probenecid in doping analysis by liquid chromatography-tandem mass spectrometry. Biomed Chromatogr. 2002;16:529–535. [PubMed] [Google Scholar]
Deventer K, Van Eenoo P, Delbeke FT. Simultaneous determination of beta-blocking agents and diuretics in doping analysis by liquid chromatography/mass spectrometry with scan-to-scan polarity switching. Rapid Commun Mass Spectrom. 2005;19:90–98. [PubMed] [Google Scholar]
Deventer K, Pozo OJ, Van Eenoo P, Delbeke FT. Detection of urinary markers for thiazide diuretics after oral administration of hydrochlorothiazide and altizide-relevance to doping control analysis. J Chromatogr A. 2009;1216:2466–2473. [PubMed] [Google Scholar]
Dormans TP, van Meyel JJ, Gerlag PG, Tan Y, Russel FG, Smits P. Diuretic efficacy of high dose furosemide in severe heart failure: bolus injection versus continuous infusion. J Am Coll Cardiol. 1996;28:376–382. [PubMed] [Google Scholar]
Edwards A, Daniels BS, Deen WM. Ultrastructural model for size selectivity in glomerular filtration. Am J Physiol. 1999;276:F892–F902. 6 Pt 2. [PubMed] [Google Scholar]
Fagard R, Staessen J, Thijs L, Amery A. Influence of antihypertensive drugs on exercise capacity. Drugs. 1993;46(Suppl. 2):32–36. [PubMed] [Google Scholar]
Faris R, Flather M, Purcell H, Henein M, Poole-Wilson P, Coats A. Current evidence supporting the role of diuretics in heart failure: a meta analysis of randomised controlled trials. Int J Cardiol. 2002;82:149–158. [PubMed] [Google Scholar]
Friedman PA, Bushinsky DA. Diuretic effects on calcium metabolism. Semin Nephrol. 1999;19:551–556. [PubMed] [Google Scholar]
Furlanello F, Serdoz LV, Cappato R, De Ambroggi L. Illicit drugs and cardiac arrhythmias in athletes. Eur J Cardiovasc Prev Rehabil. 2007;14:487–494. [PubMed] [Google Scholar]
Georgakopoulos CG, Vonaparti A, Stamou M, Kiousi P, Lyris E, Angelis YS, et al. Preventive doping control analysis: liquid and gas chromatography time-of-flight mass spectrometry for detection of designer steroids. Rapid Commun Mass Spectrom. 2007;21:2439–2446. [PubMed] [Google Scholar]
Goebel C, Trout GJ, Kazlauskas R. Rapid screening method for diuretics in doping control using automated solid phase extraction and liquid chromatography electrospray tandem mass spectrometry. Anal Chim Acta. 2004;502:65–74. [Google Scholar]
Heigenhauser GJF, Sutton JR, Jones NL. Ventilation and carbon dioxide output during exercise: effects of glycogen depletion and carbonic anhydrase inhibition. Med Sci Sports Exerc. 1980;12:123. [Google Scholar]
van der Heijden M, Donders SH, Cleophas TJ, Niemeyer MG, van der Meulen J, Bernink PJ, et al. A randomized, placebo-controlled study of loop diuretics in patients with essential hypertension: the bumetanide and furosemide on lipid profile (BUFUL) clinical study report. J Clin Pharmacol. 1998;38:630–635. [PubMed] [Google Scholar]
Hemmersbach P, de la Torre R. Stimulants, narcotics and beta-blockers: 25 years of development in analytical techniques for doping control. J Chromatogr B Biomed Appl. 1996;687:221–238. [PubMed] [Google Scholar]
Jackson EK. Diuretics. In: Brunton L, Lazo J, Parker K, editors. Goodman and Gilman’s The Pharmacological Basis of Therapeutics. 11th edn. New York: McGraw-Hill; 2006. pp. 737–770. [Google Scholar]
Koutlianos N, Kouidi E. Renal disorders and electrolyte metabolism. In: Sarikaya H, Peters C, Schulz T, Schonfelder M, Michna H, editors. Congress Manual: Biomedical Side Effects of Doping. Munich: A Project of the European Union; 2006. pp. 112–118. Available for order from the Institute of Public Health Research, Technische Universität München. [Google Scholar]
Kowalchuk JM, Heigenhauser GJ, Sutton JR, Jones NL. Effect of acetazolamide on gas exchange and acid-base control after maximal exercise. J Appl Physiol. 1992;72:278–287. [PubMed] [Google Scholar]
Levinsky NG, Bernard DB. Mannitol and loop diuretics in acute renal failure. In: Brenner BM, Lazarus JM, editors. Acute Renal Failure. 2nd edn. New York: Churchill-Livingstone; 1988. pp. 841–856. [Google Scholar]
Lisi AM, Trout GJ, Kazlauskas R. Screening for diuretics in human urine by gas chromatography-mass spectrometry with derivatisation by direct extractive alkylation. J Chromatogr. 1991;563:257–270. [PubMed] [Google Scholar]
Lisi AM, Kazlauskas R, Trout GJ. Diuretic screening in human urine by gas chromatography-mass spectrometry: use of a macroreticular acrylic copolymer for the efficient removal of the coextracted phase-transfer reagent after derivatization by direct extractive alkylation. J Chromatogr. 1992;581:57–63. [PubMed] [Google Scholar]
Maurer HH. Systematic toxicological analysis of drugs and their metabolites by gas chromatography-mass spectrometry. J Chromatogr. 1992;580:3–41. [PubMed] [Google Scholar]
Mazzarino M, Botrè F, Rossi F, Paone E, Pecori-Giraldi J. Lack of increased diuresis after topical administration of the carbonic anhydrase inhibitor dorzolamide. In: Schänzer W, Geyer H, Gotzman A, Mareck-Engelke U, editors. Recent Advances in Doping Analysis (9) Köln: Sport & Buch Strauß; 2001. pp. 269–273. [Google Scholar]
Mazzarino M, de la Torre X, Botre F. A screening method for the simultaneous detection of glucocorticoids, diuretics, stimulants, anti-oestrogens, beta-adrenergic drugs and anabolic steroids in human urine by LC-ESI-MS/MS. Anal Bioanal Chem. 2008;392:681–698. [PubMed] [Google Scholar]
Monroy A, Plata C, Hebert SC, Gamba G. Characterization of the thiazide-sensitive Na(+)-Cl(-) cotransporter: a new model for ions and diuretics interaction. Am J Physiol Renal Physiol. 2000;279:F161–F169. [PubMed] [Google Scholar]
Morra V, Davit P, Capra P, Vincenti M, Di Stilo A, Botre F. Fast gas chromatographic/mass spectrometric determination of diuretics and masking agents in human urine: development and validation of a productive screening protocol for antidoping analysis. J Chromatogr A. 2006;1135:219–229. [PubMed] [Google Scholar]
Moser M. Why are physicians not prescribing diuretics more frequently in the management of hypertension? JAMA. 1998;279:1813–1816. [PubMed] [Google Scholar]
Müller RK, Grosse J, Thieme D, Lang R, Teske J, Trauer H. Introduction to the application of capillary gas chromatography of performance-enhancing drugs in doping control. J Chromatogr A. 1999;843:275–285. [PubMed] [Google Scholar]
Munyard P, Chung KF, Bush A. Inhaled frusemide and exercise-induced bronchoconstriction in children with asthma. Thorax. 1995;50:677–679. [PMC free article] [PubMed] [Google Scholar]
Nadel ER, Fortney SM, Wenger CB. Effect of hydration state of circulatory and thermal regulations. J Appl Physiol. 1980;49:715–721. [PubMed] [Google Scholar]
Ouzan J, Perault C, Lincoff AM, Carre E, Mertes M. The role of spironolactone in the treatment of patients with refractory hypertension. Am J Hypertens. 2002;15:333–339. 4 Pt 1. [PubMed] [Google Scholar]
Plasqui G, Westerterp KR. Physical activity and insulin resistance. Curr Nutr Food Sci. 2007;3:157–160. [Google Scholar]
Politi L, Morini L, Polettini A. A direct screening procedure for diuretics in human urine by liquid chromatography-tandem mass spectrometry with information dependent acquisition. Clin Chim Acta. 2007;386:46–52. [PubMed] [Google Scholar]
Qin Y, Wang XB, Wang C, Zhao M, Wu MT, Xu YX, et al. Application of high-performance liquid chromatography-mass spectrometry to detection of diuretics in human urine. J Chromatogr B Analyt Technol Biomed Life Sci. 2003;794:193–203. [PubMed] [Google Scholar]
Reents S. Sport and Exercise Pharmacology. Champaign, IL: Human Kinetics; 2000. [Google Scholar]
Scheuermann BW, Kowalchuk JM, Paterson DH, Cunningham DA. VCO2 and VE kinetics during moderate- and heavy-intensity exercise after acetazolamide administration. J Appl Physiol. 1999;86:1534–1543. [PubMed] [Google Scholar]
Schoene RB, Bates PW, Larson EB, Pierson DJ. Effect of acetazolamide on normoxic and hypoxic exercise in humans at sea level. J Appl Physiol. 1983;55:1772–1776. [PubMed] [Google Scholar]
Schwartz GJ. Physiology and molecular biology of renal carbonic anhydrase. J Nephrol. 2002;15(Suppl. 5):S61–S74. [PubMed] [Google Scholar]
Shankar SS, Brater DC. Loop diuretics: from the Na-K-2Cl transporter to clinical use. Am J Physiol Renal Physiol. 2003;284:F11–F21. [PubMed] [Google Scholar]
Somani SM. Pharmacology in Exercise and Sports. Boca Raton, FL: CRC Press; 1996. [Google Scholar]
Stager JM, Tucker A, Cordain L, Engebretsen BJ, Brechue WF, Matulich CC. Normoxic and acute hypoxic exercise tolerance in man following acetazolamide. Med Sci Sports Exerc. 1990;22:178–184. [PubMed] [Google Scholar]
Tashian RE. Keeping pace with a fast enzyme: steps and missteps. EXS. 2000:569–596. [PubMed] [Google Scholar]
Thevis M, Schanzer W. Mass spectrometry in sports drug testing: Structure characterization and analytical assays. Mass Spectrom Rev. 2007;26:79–107. [PubMed] [Google Scholar]
Thieme D, Grosse J, Lang R, Mueller RK, Wahl A. Screening, confirmation and quantification of diuretics in urine for doping control analysis by high-performance liquid chromatography-atmospheric pressure ionisation tandem mass spectrometry. J Chromatogr B Biomed Sci Appl. 2001;757:49–57. [PubMed] [Google Scholar]
Thorngren JO, Ostervall F, Garle M. A high-throughput multicomponent screening method for diuretics, masking agents, central nervous system (CNS) stimulants and opiates in human urine by UPLC-MS/MS. J Mass Spectrom. 2008;43:980–992. [PubMed] [Google Scholar]
Trout GJ, Kazlauskas R. Sports drug testing – an analyst’s perspective. Chem Soc Rev. 2004;33:1–13. [PubMed] [Google Scholar]
Ventura R, Segura J. Detection of diuretic agents in doping control. J Chromatogr B Biomed Appl. 1996;687:127–144. [PubMed] [Google Scholar]
Ventura R, Fraisse D, Becchi M, Paisse O, Segura J. Approach to the analysis of diuretics and masking agents by high-performance liquid chromatography-mass spectrometry in doping control. J Chromatogr. 1991;562:723–736. [PubMed] [Google Scholar]
Ventura R, Roig M, Montfort N, Saez P, Berges R, Segura J. High-throughput and sensitive screening by ultra-performance liquid chromatography tandem mass spectrometry of diuretics and other doping agents. Eur J Mass Spectrom (Chichester, Eng) 2008;14:191–200. [PubMed] [Google Scholar]
Wilcox CS, Welch WJ, Schreiner GF, Belardinelli L. Natriuretic and diuretic actions of a highly selective adenosine A1 receptor antagonist. J Am Soc Nephrol. 1999;10:714–720. [PubMed] [Google Scholar]
Yoon CN, Lee TH, Park J. Mass spectrometry of methyl and methyl-d3 derivatives of diuretic agents. J Anal Toxicol. 1990;14:96–101. [PubMed] [Google Scholar]
Young DB, Srivastava TN, Fitzovich DE, Kivlighn SD, Hamaguchi M. Potassium and catecholamine concentrations in the immediate post exercise period. Am J Med Sci. 1992;304:150–153. [PubMed] [Google Scholar]
Zappe DH, Helyar RG, Green HJ. The interaction between short-term exercise training and a diuretic-induced hypovolemic stimulus. Eur J Appl Physiol Occup Physiol. 1996;72:335–340. [PubMed] [Google Scholar]
A causa della natura catabolica del Cortisolo e del desiderio viscerale di molti bodybuilder di mantenere uno stato di anabolismo muscolare costante si è speso molto tempo per cercare di contenere il rilascio di Cortisolo, soprattutto quando non necessario. Gli allenamenti sono stati ridotti in volume e intensità nella sciocca speranza di tenere sotto controllo il Cortisolo nei momenti cruciali della sua funzione fisiologica.
Tuttavia, questa visione cortisolocentrica e del suo impatto in acuto è sia riduttiva che controproducente. Essa non tiene conto della differenza tra gli aumenti acuti e cronici del corticosteroide in questione.
Detto ciò, approfondiamo il ruolo dell’ormone Cortisolo nel processo di ipertrofia muscolare.
Caratteristiche principali del Cortisolo:
Il Cortisolo è un ormone steroideo, appartenente alla classe degli ormoni glucocorticoidi. Quando viene utilizzato come farmaco, è noto come Idrocortisone.
Viene sintetizzato in molti animali, principalmente dalla zona fascicolata della corteccia surrenale nella ghiandola surrenale.[1][2] Viene prodotto in altri tessuti in quantità inferiori.[3] Viene rilasciato con un ciclo diurno e il suo rilascio aumenta in risposta allo stress e a una bassa concentrazione di glucosio nel sangue. Funziona per aumentare la glicemia ematica attraverso la gluconeogenesi, per sopprimere il sistema immunitario e per coadiuvare il metabolismo di grassi, proteine e carboidrati.[4] Diminuisce anche la formazione delle ossa.[5] Molte di queste funzioni sono svolte dal Cortisolo che si lega ai recettori dei glucocorticoidi o dei mineralocorticoidi all’interno della cellula, che poi si legano al DNA per influenzare l’espressione genica.[6][7]
Asse Ipotalamo-Ipofisi-Surrene
Grazie alle proprietà immunoregolatrici del Cortisone, i derivati farmaceutici del Cortisolo, come il Prednisone, sono utilizzati per controllare forti reazioni allergiche, artrite e altre condizioni infiammatorie. I pericoli di un aumento cronico del Cortisolo sono evidenti nel modo attento in cui questi farmaci vengono dosati e nella breve durata dei trattamenti che li utilizzano.
Il Cortisolo è sintetizzato a partire dal Colesterolo. Come già accennato, la sua sintesi avviene nella zona fascicolata della corteccia surrenale (il nome Cortisolo deriva da corteccia). Sebbene la corteccia surrenale produca anche Aldosterone (nella zona glomerulosa) e alcuni ormoni sessuali (nella zona reticolare), il Cortisolo è la sua secrezione principale nell’uomo e in molte altre specie. La midollare della ghiandola surrenale si trova sotto la corteccia e secerne principalmente le catecolamine Adrenalina (Epinefrina) e Noradrenalina (Norepinefrina) sotto stimolazione simpatica.
Ormone Adrenocorticotropo (ACTH)
La sintesi di Cortisolo nella ghiandola surrenale è stimolata dal lobo anteriore dell’ipofisi con l’ACTH; la produzione di ACTH è a sua volta stimolata dal CRH, rilasciato dall’ipotalamo. L’ACTH aumenta la concentrazione di Colesterolo nella membrana mitocondriale interna, attraverso la regolazione della proteina regolatrice steroidogenica acuta. Stimola inoltre la principale fase limitante della sintesi del Cortisolo, in cui il Colesterolo viene convertito in Pregnenolone e catalizzato dal citocromo P450SCC (enzima di scissione della catena laterale).[8]
Il Cortisolo viene metabolizzato reversibilmente a Cortisone[9-89] dal sistema dell’11-beta idrossisteroide deidrogenasi (11-beta HSD), che consiste in due enzimi:11-beta HSD1 e 11-beta HSD2. Il metabolismo del Cortisolo a Cortisone comporta l’ossidazione del gruppo ossidrilico in posizione 11-beta.[10]
L’11-beta HSD1 utilizza il cofattore NADPH per convertire il Cortisone biologicamente inerte in Cortisolo biologicamente attivo.
L’11-beta HSD2 utilizza il cofattore NAD+ per convertire il Cortisolo in Cortisone.
Conversione enzimatica da Cortisone a Cortisolo [11-beta HSD1] e da quest’ultimo a Cortisone [11-beta HSD2].
Nel complesso, l’effetto netto è che l’11-beta HSD1 serve ad aumentare le concentrazioni locali di Cortisolo biologicamente attivo in un dato tessuto; l’11-beta HSD2 serve a diminuire le concentrazioni locali di Cortisolo biologicamente attivo. Se è presente l’esoso-6-fosfato deidrogenasi (H6PDH), l’equilibrio può favorire l’attività dell’11-beta HSD1. L’H6PDH rigenera NADPH, aumentando l’attività dell’11-beta HSD1 e diminuendo quella dell’11-beta HSD2.[11]
È stato ipotizzato che un’alterazione dell’11-beta HSD1 svolga un ruolo nella patogenesi dell’obesità, dell’ipertensione e dell’insulino-resistenza, note come sindrome metabolica.[12]
Un’alterazione dell’11-beta HSD2 è stata implicata nell’ipertensione essenziale ed è nota per portare alla sindrome da eccesso apparente di mineralcorticoidi (SAME).
A breve termine, l’aumento del Cortisolo è associato a una diminuzione della sintesi proteica. Il motivo è che una delle azioni del Cortisolo è quella di fornire substrati energetici alternativi all’organismo quando non c’è abbastanza glucosio. Ciò si verifica durante la restrizione calorica o il digiuno, ma anche durante l’esercizio fisico intenso. Il Cortisolo media la degradazione muscolare in modo che gli aminoacidi presenti nel tessuto muscolare possano essere utilizzati per creare glucosio, attraverso la gluconeogenesi.
Il Cortisolo svolge anche un ruolo importante, ma indiretto, nella glicogenolisi epatica e muscolare (la scissione del glicogeno in glucosio-1-fosfato e glucosio) che si verifica in seguito all’azione del Glucagone e dell’Adrenalina. Inoltre, il Cortisolo facilita l’attivazione della glicogeno fosforilasi, necessaria affinché l’Adrenalina abbia effetto sulla glicogenolisi.[13][14]
È paradossale che il Cortisolo promuova non solo la gluconeogenesi nel fegato, ma anche la glicogenesi: è quindi meglio pensare che il Cortisolo stimoli il turnover di glucosio/glicogeno nel fegato. [Questo è in contrasto con l’effetto del cortisolo nel muscolo scheletrico, dove la glicogenolisi è promossa indirettamente attraverso le catecolamine.[15] In questo modo, il Cortisolo e le catecolamine lavorano sinergicamente per promuovere la scissione del glicogeno muscolare in glucosio, che viene poi utilizzato da altri tessuti.
Il Cortisolo aumenta anche i livelli di glucosio nel sangue riducendo l’assorbimento del glucosio nei tessuti muscolari e adiposi, diminuendo la sintesi proteica e aumentando la scomposizione dei trigliceridi di deposito in grassi acidi liberi (lipolisi). Tutte queste modifiche metaboliche hanno l’effetto netto di aumentare i livelli di glucosio nel sangue, che alimentano il cervello e altri tessuti durante la risposta di lotta o fuga [16] … e i workout…
Livelli elevati di Cortisolo, se prolungati, quindi elevati in cronico, possono portare alla proteolisi (disgregazione delle proteine) protratta e al deperimento muscolare.[17] La ragione della proteolisi è quella di fornire ai tessuti interessati una materia prima per la gluconeogenesi; si vedano gli aminoacidi glucogenici.[18] Gli effetti del Cortisolo sul metabolismo lipidico sono più complicati, poiché la lipogenesi è osservata in pazienti con livelli cronici elevati di glucocorticoidi circolanti,[18] mentre un aumento acuto del Cortisolo circolante promuove la lipolisi. La spiegazione abituale di questa apparente discrepanza è anche l’aumento della concentrazione di glucosio nel sangue (per azione del Cortisolo) stimola il rilascio di Insulina. L’Insulina stimola la lipogenesi, quindi questa è una conseguenza indiretta dell’aumento della concentrazione di cortisolo nel sangue, ma si verifica solo su una scala temporale più lunga. Stiamo parlando sempre di condizioni croniche e non in range fisiologici.
Il Cortisolo è un ormone controinsulinare, contribuisce quindi all’iperglicemia stimolando la gluconeogenesi[19] e inibisce l’utilizzo periferico del glucosio (insulino-resistenza)[19] diminuendo la traslocazione dei trasportatori del glucosio (in particolare GLUT4) sulla membrana cellulare. Il Cortisolo aumenta anche la sintesi di glicogeno (glicogenesi) nel fegato, immagazzinando il glucosio in forma facilmente accessibile.[20] L’effetto permissivo del Cortisolo sull’azione dell’Insulina nella glicogenesi epatica è stato osservato in coltura di epatociti in laboratorio, anche se il meccanismo di questo fenomeno è sconosciuto.
Il Cortisolo aumenta gli aminoacidi liberi nel siero inibendo la formazione di Collagene, diminuendo l’assorbimento di aminoacidi da parte del muscolo e inibendo la sintesi proteica.[21] Il Cortisolo (sotto forma di Opticortinolo) può inibire inversamente le cellule precursori delle IgA nell’intestino dei vitelli.[22] Il Cortisolo inibisce anche le IgA nel siero, come le IgM; tuttavia, non è dimostrato che inibisca le IgE.[23]
Il Cortisolo diminuisce la velocità di filtrazione glomerulare e il flusso plasmatico renale dai reni, aumentando così l’escrezione di fosfati e aumentando la ritenzione di Sodio e acqua e l’escrezione di Potassio agendo sui recettori dei mineralocorticoidi. Aumenta inoltre l’assorbimento di Sodio e acqua e l’escrezione di Potassio nell’intestino.[24] Il Cortisolo favorisce l’assorbimento del Sodio attraverso l’intestino tenue dei mammiferi.[25] La deplezione di Sodio, tuttavia, non influisce sui livelli di Cortisolo[26] e quindi questo ormone non può essere utilizzato per regolare il Sodio sierico. Un carico di Sodio aumenta l’intensa escrezione di Potassio da parte del Cortisolo. In questo caso, il Corticosterone è paragonabile al Cortisolo.[27] Affinché il Potassio esca dalla cellula, il Cortisolo sposta un numero uguale di ioni Sodio all’interno della cellula.[28] Ciò dovrebbe facilitare la regolazione del pH (a differenza della normale situazione di carenza di Potassio, in cui due ioni Sodio si spostano all’interno per ogni tre ioni Potassio che si spostano all’esterno, il che si avvicina all’effetto del Desossicorticosterone).
Cortisolo e workout:
Nell’articolo del 1998 “Stress-Related Cortisol Secretion in Men: Relationships with Abdominal Obesity and Endocrine, Metabolic, and Hemodynamic Abnormalities”, i ricercatori del Sahlgrenska University Hospital in Svezia hanno dato diversi contributi preziosi alla nostra comprensione del Cortisolo e delle sue attività differenti in acuto e in cronico. Innanzitutto, le singole letture dei livelli di Cortisolo di un soggetto “non sono altamente informative, perché il Cortisolo viene secreto in modo molto irregolare”.
I livelli di Cortisolo in genere salgono e scendono nel corso della giornata e un livello elevato in un determinato momento non è indicativo di un problema. Al contrario, livelli di Cortisolo variabili, flessibili e reattivi riflettono un sistema endocrino sano. Se il corpo perdesse la capacità di rispondere ai fattori di stress e di regolare in modo appropriato i livelli di Cortisolo, sarebbe un problema.
Un secondo punto che lo studio svedese fornisce riguarda un aspetto che molte persone sbagliano nella loro ricerca di una body fat ridotta, soprattutto addominale. Il Cortisolo viene spesso definito “l’ormone del grasso della pancia”, ma la verità è che il Cortisolo ha il suo maggiore impatto sul grasso viscerale, che è il grasso che circonda gli organi, non il grasso sottocutaneo che copre gli addominali. Se la body fat rende poco visibile il retto addominale, il problema principale non è il Cortisolo.
Nel 2006, Stephen Bird ha pubblicato una serie di articoli che tracciano un buon quadro dei cambiamenti ormonali che si verificano in seguito al sollevamento pesi e di come i diversi interventi nutrizionali influiscano su tali cambiamenti. Nel loro insieme, questi lavori forniscono un quadro della differenza tra i cambiamenti ormonali a breve termine, ad esempio durante o dopo una sessione di allenamento, e quelli a lungo termine.
Nello studio di Bird erano presenti quattro gruppi di soggetti, suddivisi in base a ciò che potevano bere durante gli allenamenti: acqua, aminoacidi essenziali, carboidrati o aminoacidi essenziali più carboidrati. Nell’arco di 12 settimane, tutti i gruppi hanno perso all’incirca la stessa quantità di grasso corporeo, mentre il gruppo che aveva una supplementazione più completa durante l’allenamento (EAA + carboidrati) ha guadagnato più muscoli.
Esaminiamo ora i cambiamenti acuti che hanno accompagnato questa differente risposta. I ricercatori hanno misurato l’aminoacido 3-metil-istidina nelle urine come marcatore della degradazione muscolare. Come mostra il grafico sottostante, il gruppo che ha bevuto solo acqua (il placebo) ha registrato un aumento della disgregazione muscolare 48 ore dopo la sessione di allenamento. I gruppi che hanno assunto aminoacidi essenziali o carboidrati non hanno subito variazioni. Il gruppo che ha assunto la bevanda combinata per l’allenamento ha registrato una diminuzione dei livelli di 3-metil-istidina dopo l’allenamento.
Ciò che è successo, molto banalmente, è che i substrati ingeriti con la bevanda intra-workout hanno tamponato l’uso delle proteine strutturali.
cosa è successo al Cortisolo? Come si può vedere qui sotto, i livelli di Cortisolo 30 minuti dopo l’esercizio fisico sono aumentati di oltre il 50% nel gruppo che ha bevuto acqua, mentre sono rimasti praticamente invariati nel gruppo EAA. Il Cortisolo è diminuito in entrambi i gruppi che hanno assunto Carboidrati come parte dell’alimentazione peri-workout.
Pensate alla gluconeogenesi, il processo che nel fegato crea glucosio da fonti non glucidiche per fornire energia alle cellule del corpo che ne hanno essenziale bisogno. L’organismo non ha bisogno di generare glucosio – un processo ad alto costo metabolico – quando nel flusso ematico c’è glucosio extra dato da una bevanda sportiva. Pertanto, non si è verificato alcun aumento sensibile del Cortisolo in presenza di carboidrati.
Il catabolismo muscolare a breve termine e il picco di Cortisolo in acuto per il gruppo che beveva acqua possono sembrare significativi, ma non bisogna dimenticare che tutti i gruppi hanno guadagnato massa muscolare nel corso dello studio. Il gruppo che ha bevuto solo acqua ha aggiunto quasi due chili di massa muscolare in 12 settimane! Adesso cominciate ad avere chiara la differenza tra effetto in acuto e effetto in cronico?… Il catabolismo è propedeutico all’anabolismo! Eventi in acuto sono largamente compensati dai processi di recupero, in fisiologia.
I picchi di Cortisolo decrescono nel breve termine!
Quindi le persone con la più alta risposta catabolica in acuto hanno comunque guadagnato muscoli? Certo che si! Ed è piuttosto semplice, in realtà: oltre all’attività propedeutica del catabolismo per avviare i processi anabolici, la fisiologia dei soggetti osservati si è adattata allo stimolo dell’allenamento contro-resistenza nel corso del tempo e ha rilasciato sempre meno Cortisolo, anche senza alcun intervento nutrizionale. Nel gruppo che beveva acqua, i livelli di Cortisolo post-esercizio sono diminuiti del 28% nel corso delle 12 settimane dello studio.
Sì, è emerso che i livelli di Cortisolo e la degradazione muscolare acuta non hanno affatto un impatto negativo sull’aumento dei muscoli o sulla perdita di grasso per un periodo di 12 settimane.
I ricercatori della McMaster hanno analizzato la relazione tra i livelli di Cortisolo post-allenamento e i cambiamenti nella forza, nella massa magra e nella sezione trasversale delle fibre muscolari. Hanno scoperto che dopo 12 settimane di allenamento contro-resistenza, alti livelli di Cortisolo post-allenamento erano correlati (anche se debolmente) con l’aumento della massa magra e con le variazioni delle dimensioni delle fibre muscolari di tipo II.
È bene ripeterlo: Le persone con livelli di Cortisolo più elevati in acuto erano quelle che avevano maggiori probabilità di guadagnare più muscoli nel corso dello studio. Tutto il contrario di quello che i limitati detrattori del Cortisolo si sarebbero aspettati!
Conclusioni:
In definitiva, i dati della ricerca sottolineano che l’interruzione di un allenamento contro-resistenza e/o l’abbassamento del intensità e del volume per paura che i livelli di Cortisolo post-allenamento impennassero, è potenzialmente controproducente ai fini ipertrofici. Lo studio della McMaster ha lasciato intendere che potrebbe addirittura esistere una correlazione tra l’innalzamento acuto del Cortisolo e la crescita muscolare a lungo termine. Detto ciò potreste chiedervi: “Se gli innalzamenti acuti del Cortisolo riflettono una buona sessione di allenamento, allora dovrei smettere di usare i protocolli nutrizionali che riducono il Cortisolo?”. Direi di no, non è assolutamente necessaria l’eliminazione del intra-workout. Le proteine e i carboidrati assunti prima, durante e dopo l’allenamento sono comunque importanti per avviare il processo di recupero.
In queste situazioni, il Cortisolo elevato è semplicemente un indicatore di un allenamento produttivo. E, per non dimenticare, nello studio iniziale di 12 settimane il gruppo che ha assunto aminoacidi e carboidrati ha guadagnato più del doppio dei muscoli rispetto a chi ha bevuto solo acqua; per ovvie ragioni di substrati disponibili e migliore prestazione data dal consumo di CHO.
Un ultimo dubbio: se dobbiamo ignorare i livelli di cortisolo post-allenamento, questo significa che dobbiamo dimenticarci del tutto del Cortisolo e ignorare qualsiasi cambiamento a lungo termine nei nostri livelli?
Assolutamente no!
I cambiamenti a lungo termine del Cortisolo e la diminuzione della sua flessibilità circadiana dovrebbero essere monitorati. Gli effetti sistemici di questo ormone catabolico devono essere presi in considerazione quando si guarda al quadro generale dell’allenamento, della alimentazione e dello stile di vita in generale.
Sonno adeguato, calorie e attenzione al recupero sono i tre fattori più importanti su cui abbiamo il controllo quotidiano. Oltre a questi fattori, è stato suggerito l’uso di integratori come il SAMe o l’Ashwagandha per favorire l’adattamento allo stress e prevenire ulteriormente l’aumento cronico del Cortisolo, o, se atleti “enhanced” farmaci come il Trilostano o l’Aminoglutettimide che sono inibitori della biosintesi steroidea.
Indipendentemente dalla forza e dalla forma fisica, gli elevati livelli di Cortisolo indotti dallo stress cronico possono compromettere il benessere psicofisico. Aumentano il rischio di ipertensione e di malattie cardiovascolari e aggravano qualsiasi altro problema di cui si possa soffrire. Ricordate, tuttavia, di mantenere le cose in prospettiva e di guardare al lungo termine.
Jump up to:ab Chyun YS, Kream BE, Raisz LG (February 1984). “Cortisol decreases bone formation by inhibiting periosteal cell proliferation”. Endocrinology.
DeRijk RH, Schaaf M, de Kloet ER (June 2002). “Glucocorticoid receptor variants: clinical implications”. The Journal of Steroid Biochemistry and Molecular Biology.
Margioris AN, Tsatsanis C (2011). “ACTH Action on the Adrenal”. In Chrousos G (ed.). Adrenal physiology and diseases. Endotext.org. Archived from the original on 29 November 2011. Retrieved 5 June 2012.
^ Jump up to:ab Finken MJ, Andrews RC, Andrew R, Walker BR (September 1999). “Cortisol metabolism in healthy young adults: sexual dimorphism in activities of A-ring reductases, but not 11beta-hydroxysteroid dehydrogenases”. The Journal of Clinical Endocrinology and Metabolism. 84 (9): 3316–3321. doi:10.1210/jcem.84.9.6009. PMID10487705.
^ Dammann C, Stapelfeld C, Maser E (April 2019). “Expression and activity of the cortisol-activating enzyme 11β-hydroxysteroid dehydrogenase type 1 is tissue and species-specific”. Chemico-Biological Interactions. 303: 57–61. doi:10.1016/j.cbi.2019.02.018. PMID30796905. S2CID73467693.
^ Atanasov AG, Nashev LG, Schweizer RA, Frick C, Odermatt A (July 2004). “Hexose-6-phosphate dehydrogenase determines the reaction direction of 11beta-hydroxysteroid dehydrogenase type 1 as an oxoreductase”. FEBS Letters. 571 (1–3): 129–133. doi:10.1016/j.febslet.2004.06.065. PMID15280030. S2CID6360244.
Martin PA, Crump MH (2003). “The adrenal gland”. In Dooley MP, Pineda MH (eds.). McDonald’s veterinary endocrinology and reproduction (5th ed.). Ames, Iowa: Iowa State Press. ISBN978-0-8138-1106-2.
^ Coderre L, Srivastava AK, Chiasson JL (June 1991). “Role of glucocorticoid in the regulation of glycogen metabolism in skeletal muscle”. The American Journal of Physiology. 260 (6 Pt 1): E927–32.
Kuo T, McQueen A, Chen TC, Wang JC (2015). “Regulation of Glucose Homeostasis by Glucocorticoids”. In Wang JC, Harris C (eds.). Glucocorticoid Signaling: From Molecules to Mice to Man. Advances in Experimental Medicine and Biology. Vol. 872. Springer. pp. 99–126.
Khani S, Tayek JA (December 2001). “Cortisol increases gluconeogenesis in humans: its role in the metabolic syndrome”. Clin Sci (Lond).
Brown DF, Brown DD (2003). USMLE Step 1 Secrets: Questions You Will Be Asked on USMLE Step 1. Philadelphia: Hanley & Belfus. p. 63.
Baynes J, Dominiczak M (2009). Medical biochemistry. Mosby Elsevier.
Manchester, KL (1964). “Sites of Hormonal Regulation of Protein Metabolism”. In Allison, NH; Munro JB (eds.). Mammalian Protein Metabolism. New York: Academic Press.
Husband AJ, Brandon MR, Lascelles AK (October 1973). “The effect of corticosteroid on absorption and endogenous production of immunoglobulins in calves”. The Australian Journal of Experimental Biology and Medical Science.
^ Posey WC, Nelson HS, Branch B, Pearlman DS (December 1978). “The effects of acute corticosteroid therapy for asthma on serum immunoglobulin levels”. The Journal of Allergy and Clinical Immunology.
^ Sandle GI, Keir MJ, Record CO (1981). “The effect of hydrocortisone on the transport of water, sodium, and glucose in the jejunum. Perfusion studies in normal subjects and patients with coeliac disease”. Scandinavian Journal of Gastroenterology.
^ Mason PA, Fraser R, Morton JJ, Semple PF, Wilson A (August 1977). “The effect of sodium deprivation and of angiotensin II infusion on the peripheral plasma concentrations of 18-hydroxycorticosterone, aldosterone and other corticosteroids in man”. Journal of Steroid Biochemistry.
Muller AF, Oconnor CM (1958). An International Symposium on Aldosterone. Little Brown & Co. p. 58.
Knight RP, Kornfeld DS, Glaser GH, Bondy PK (February 1955). “Effects of intravenous hydrocortisone on electrolytes of serum and urine in man”. The Journal of Clinical Endocrinology and Metabolism.
La Betaina è un integratore che probabilmente la maggior parte dei frequentatori di palestra non ha mai sentito nominare, eppure è un’aggiunta preziosa alla propria gamma di integratori. L’integratore esiste da anni e si trova in diverse fonti alimentari [1], ma solo da meno di un decennio che è tornato all’attenzione grazie a uno studio pubblicato sul Journal of the International Society of Sports Nutrition [2]. L’autore principale, Jason Cholewa, non è nuovo al settore del fitness. Ha scritto per diversi anni per BodyBuilding.com (attivo con il suo nickname online “Big Red”), tra gli altri, e attualmente è assistente alla cattedra di scienze dell’esercizio e studi sportivi presso la Coastal Carolina University. Dirige anche la società di consulenza sul fitness “Big Red Physical Performance” ed è stato lui stesso un bodybuilder attivo per molti anni.
Introduzione alla Betaina:
Ma cos’è esattamente la Betaina? In poche parole, essa svolge due funzioni nel corpo umano. Una è quella di osmolita e l’altra è quella di donatore di metile.
Struttura molecolare della Betaina (Trimetylglicina).
Un osmolita non fa altro che portare con sé l’acqua come risultato dell’osmosi. Quando si aumenta la concentrazione di una sostanza tra due compartimenti (separati da una membrana semipermeabile, come la membrana cellulare), l’acqua si diffonde dal compartimento con la bassa concentrazione della sostanza al compartimento con l’alta concentrazione della sostanza, finché le concentrazioni non si equivalgono. È simile al modo in cui la creatina provoca la ritenzione di liquidi, con la sottile differenza che quasi tutta la creatina presente nell’organismo si trova nel tessuto muscolare, e quindi il liquido si accumula nei muscoli. La betaina si accumula in quasi tutti i tessuti per regolare il volume cellulare ed è uno dei principali osmoliti organici [4]. In pratica, quindi, regola l’idratazione cellulare. Pertanto, può anche assumere un ruolo protettivo nella formazione dell’urea (metabolita del catabolismo degli aminoacidi). L’urea è molto efficace nel denaturare (“dispiegare” la struttura spaziale) delle proteine e quindi influisce sulla loro funzione. Influisce anche sulla miosina, che contrae i muscoli insieme alla proteina actina. L’attività ATPasica della miosina è inibita dall’urea, e l’ATPasica è necessaria per idrolizzare l’ATP in energia: tale energia è necessaria per far “correre” le teste della miosina sull’actina e quindi contrarre il muscolo. La betaina contrasta questo effetto [3]. Inoltre, il rigonfiamento della cellula stimola la sintesi proteica (e la contrazione stimola la proteolisi) [4].
Come si evince dalla formula strutturale riportata in precedenza, la betaina contiene tre gruppi metilici legati all’atomo di azoto. La transmetilazione, il processo biochimico mediante il quale un gruppo metile viene ceduto da una sostanza a un’altra, è molto importante per il funzionamento delle cellule. La betaina è un donatore di metile per l’omocisteina per formare metionina. Questa reazione è catalizzata dalla betaina-omocisteina metiltransferasi (BHMT). Il sottoprodotto di questa reazione è la dimetilglicina, e questa reazione è guidata dall’osmosi, vale a dire che ad alte osmalità, l’espressione di questo enzima è downregolata per mantenere la concentrazione di betaina. Al contrario, a bassa osmalità, l’enzima è più espresso e viene utilizzata più betaina per metilare l’omocisteina.
La metionina che si forma è un aminoacido essenziale (si noti che la metilazione dell’omocisteina è in realtà una rimetilazione). Oltre al suo ruolo di amminoacido che può essere utilizzato nella traduzione dell’mRNA in proteine, è anche il precursore della S-adenosil metionina (SAM). Questa reazione è catalizzata dalla metionina adenosiltransferasi e utilizza metionina e ATP come substrati; oltre alla SAM formata, si formano anche fosfato e difosfato come sottoprodotti.
Il ciclo della Metionina. La Betaina dona un gruppo metile all’Omocisteina per formare Metionina. La Metionina costituisce un substrato per la Metionina adenosiltransferasi per formare SAM. La SAM è un importante donatore di metile in numerose reazioni. Dopo la donazione del suo gruppo metile, l’Omocisteina si forma nuovamente. Figura riprodotta da [1].
Il SAM è una molecola particolare. Alcuni potrebbero riconoscere il nome dalla biosintesi della creatina. L’acido guanidinoacetico, precursore della creatina, riceve un gruppo metilico dal SAM per formare la creatina [5]. Tuttavia, ciò che molti non sanno è che SAM non solo dona un gruppo metile per la biosintesi della creatina, ma lo fa per numerose reazioni. Tra cui la sintesi proteica, la formazione dei fosfolipidi, gli ormoni, le poliammine, la carnitina, l’adrenalina e la metilazione del DNA [1]. Inoltre, inibisce la segnalazione dell’insulina diminuendo la fosforilazione (e quindi l’attivazione) del substrato 1 del recettore insulinico (IRS-1) negli adipociti [8].
Inoltre, le concentrazioni di omocisteina nel sangue sono inversamente correlate alle malattie croniche (comprese quelle cardiovascolari). Questo aspetto esula dallo scopo di questo articolo, ma è discusso in [5].
Ma funziona? Come citato precedentemente, l’integratore è stato recentemente riportato sotto i riflettori da uno studio condotto da Jason Cholewa e dai suoi colleghi. Hanno condotto uno studio in doppio cieco controllato con placebo. Sono stati reclutati 23 uomini esperti nell’allenamento della forza a livello amatoriale (peso medio: 86,8 kg SD: 9,1 kg), di età compresa tra i 18 e i 35 anni. In altre parole: un campione molto rilevante che ho particolarmente apprezzato. Il gruppo di trattamento ha ricevuto 2x 1,25 g di betaina al giorno (n=11). Si sono allenati per sei settimane, quattro giorni alla settimana, suddivisi in microcicli di due settimane. Sono state riscontrate differenze significative nella composizione corporea tra i due gruppi, come mostrato nella tabella seguente:
Gli effetti dell’integrazione di Betaina sulla composizione corporea che sono stati misurati nello studio in questione.
La percentuale di grasso è stata determinata mediante la misurazione delle pliche cutanee. Da questa percentuale e dal peso misurato sono state ricavate la massa magra e la massa grassa.
Inoltre, è stata riscontrata una differenza significativa tra l’area della sezione trasversale tra il periodo precedente e quello successivo al trattamento nel gruppo di integrazione di betaina, che non è stata riscontrata nel gruppo placebo (significatività fissata a P < 0,05). Tuttavia, non è stata riscontrata alcuna differenza in termini di sezione trasversale del femore (P = 0,254).
In termini di forza, non c’è molto di entusiasmante, come si può vedere nella tabella seguente che mostra l’1RM di tre gruppi di atleti.
Effetti sulle misure di forza dell’integrazione di Betaina come misurato nello studio.
Sono state riscontrate alcune differenze significative tra il gruppo Betaina e il gruppo placebo quando si è guardato specificamente ai singoli microcicli, ma ciò è avvenuto anche viceversa.
Degno di nota è soprattutto l’effetto sulla composizione corporea. Non è chiaro in che misura ciò sia dovuto alla funzione osmolitica della Betaina. L’unico studio comparabile in termini di durata non ha riscontrato alcun miglioramento della composizione corporea dopo 12 settimane di integrazione in soggetti obesi [6]; tuttavia, ciò potrebbe essere dovuto al fatto che si trattava di soggetti obesi che mascheravano l’eventuale ritenzione di liquidi dovuta all’integrazione di Betaina. Sono quindi necessarie ulteriori ricerche per chiarire questo aspetto. In ogni caso, l’effetto positivo sul braccio trasversale lascia ben sperare.
Betaina e sistemi energetici:
Struttura molecolare dell’ATP
L’articolo di Cholewa et al. si apre con la proposta che la Betaina possa potenziare la glicolisi aumentando il rapporto NAD+:NADH accettando ioni H+ nucleofili e di conseguenza, incidentalmente, agendo come un leggero tampone del pH. Per coloro che non hanno la memoria fresca sulla glicolisi, essa produce un netto di 2 ATP dalla scissione anaerobica del glucosio ematico (o 3 ATP dal glucosio-6-fosfato dal glicogeno). L’equazione di reazione è la seguente:
Sul lato sinistro dell’equazione di reazione si può notare il NAD+ che è l’agente ossidante della reazione. Inoltre, vengono generati ioni H++. Un’ipotesi di Ghyczy e Boros [5] ci dice che molecole come la Betaina, a causa del loro gruppo azotato trigemetilato (carente di elettroni), accettano coppie di elettroni da altre molecole. Di conseguenza, ipotizzano che reagisca anche con il donatore di elettroni NADH, per cui la Betaina è in grado di diminuire la concentrazione di NADH e aumentare quella di NAD+. La rigenerazione del NAD+ può quindi stimolare la glicolisi. E la reazione con gli ioni H+, per definizione, farà scendere il pH.
Dato che la glicolisi è importante per la generazione di ATP durante l’esercizio fisico intenso, come quello eseguito dagli atleti di forza, questo è un fatto piuttosto interessante. Un ulteriore sostegno a questa ipotesi viene dall’osservazione che l’integrazione di Betaina porta a un maggiore aumento del lattato plasmatico durante lo sprint [6]. Infatti, dopo la formazione di acido piruvico da parte della glicolisi, l’acido piruvico può essere trasportato in due modi:
1) viene trasportato nei mitocondri dove viene convertito in acetil-CoA ed entra nel ciclo dell’acido citrico, oppure
2) si forma lattato. Un aumento del lattato nel plasma può quindi essere indicativo di un aumento della glicolisi (o di un aumento della clearance da parte del muscolo, come sottolineato anche da Cholewa et al.)
In seguito all’effetto della Betaina sulla glicolisi, è stato ipotizzato che essa promuova anche il ciclo dell’acido citrico. Ciò si basa sul fatto che la Betaina protegge la citrato sintasi dalla denaturazione termica [7]. La citrato sintasi è l’enzima che catalizza la prima reazione del ciclo dell’acido citrico, la condensazione dell’acetil-CoA e dell’ossalacetato, formando acido citrico nel processo. La citrato sintasi è molto importante per il flusso del ciclo dell’acido citrico, poiché l’enzima opera lontano dall’equilibrio con un ΔG stimato di -31,5 kJ/mol-1 ed è quindi un passo importante da regolare.
Formazione del citrato. Condensazione dell’Acetil-CoA con ossalacetato per formare il citrato, reazione catalizzata dalla citrato sintasi.
Infine, oltre all’ipotizzato effetto positivo sui due sistemi energetici sopra citati, la Betaina può avere anche un possibile effetto positivo sul sistema energetico dei fosfati. È stato infatti ipotizzato che l’integrazione di Betaina possa aumentare la biosintesi di Creatina. Questo effetto è stato dimostrato negli animali, ma non nell’uomo [1] (oltre a quanto sottolineato da Cholewa et al., mi sembra che ciò sia dovuto semplicemente a un lasso di tempo troppo breve [10 d]). In effetti, il SAM è il donatore di metile per l’acido guanidinoacetico, il precursore diretto della Creatina.
Betaina e segnalazione dell’Insulina:
Struttura del Recettore dell’Insulina
Oltre agli effetti sui sistemi energetici, può anche avere un effetto benefico sulla composizione corporea, rimetilando l’omocisteina e abbassandone di conseguenza la concentrazione. Oltre al fatto che l’iperomocisteinemia è dannosa per la salute, essa porta a una diminuzione della fosforilazione della Tirosina del recettore dell’Insulina e del substrato del Recettore dell’Insulina 1 (IRS-1), a un aumento della fosforilazione della serina di IRS-1 e a una conseguente minore fosforilazione di Akt [8]. Ciò è stato riscontrato negli adipociti e quindi corrisponde concretamente a un aumento dell’insulino-resistenza. Se lo stesso effetto si verifica nei miociti (cellule muscolari), corrisponde concretamente a una riduzione della sintesi proteica attraverso l’inibizione della via IR/PI3K/Akt/mTOR.
Via PI3K/Akt/mTOR. Ristampato e adattato da [9].
Struttura molecolare del IGF-1
Uno studio clinico ci fornisce un quadro più chiaro al riguardo [10]. La Betaina è stata integrata in individui allenati in un design crossover. Qui, dopo l’allenamento, sono stati misurati alcuni ormoni (GH, IGF-I, Cortisolo, Insulina) e molecole di segnalazione anabolica (Akt, p70S6K, AMPK). È stato riscontrato un aumento significativo dell’IGF-I, un aumento quasi significativo del GH, una diminuzione significativa del Cortisolo e nessun effetto sull’Insulina. Il moderato aumento del GH è positivo, ma in realtà dice ben poco. Anche l’effetto inibitorio sull’aumento del Cortisolo dopo l’esercizio fisico è difficile da mettere in prospettiva. E sebbene l’IGF-I sia strettamente coinvolto nell’ipertrofia muscolare, resta da vedere quanto ci dica in resa l’IGF-I in circolazione. L’IGF-I agisce solo brevemente come ormone autocrino/paracrino in termini di ipertrofia muscolare e, anche se l’IGF-I intramuscolare fosse misurato, sarebbe solo una parte del puzzle. L’attività finale dell’IGF-I è anche fortemente regolata dalle proteine leganti l’IGF. Quindi, idealmente, si vorrebbe misurare anche questo insieme a qualcosa che rifletta l’attività dell’IGF-I. Fortunatamente, in questo studio è stato preso in considerazione l’Akt. L’Akt (noto anche come protein chinasi B) è un importantissimo regolatore a monte di mTORC1 ed è quindi intimamente coinvolto nella regolazione dell’ipertrofia muscolare. L’Akt è un effettore del recettore IGF attraverso l’attivazione di PI3K, che genera PIP3. Il PIP3 causa il reclutamento di Akt, interagendo con il suo dominio di omologia pleckstrin, nel sarcolemma, dove Akt può essere attivato (fosforilato). Pertanto, la misurazione della fosforilazione di Akt riflette in qualche modo l’attività dell’IGF-I. In un certo senso, considerando che ci sono anche altri regolatori a monte.
Struttura molecolare del Akt
Lo studio ha evidenziato che l’integrazione di Betaina aumenta la quantità di Akt nelle cellule muscolari a riposo. Naturalmente, questo non dice ancora nulla, dato che la chinasi deve essere attivata (tramite fosforilazione su Ser473 e Thr308 nel caso dell’isoforma Akt1) prima di agire effettivamente. Anche questo aspetto è stato esaminato. Nel gruppo placebo, la fosforilazione di Akt su Ser473 è diminuita subito dopo l’esercizio, mentre è stata mantenuta un po’ di più nel caso della Betaina. Una bella scoperta. Ancora più interessante, gli autori hanno anche esaminato la fosforilazione di p70S6K, un effettore a valle di Akt che fosforila la proteina ribosomiale S6 (che causa la traduzione dell’mRNA in proteina) e quindi aumenta la sintesi proteica. Non a caso, l’integrazione di Betaina ha portato a livelli più elevati di fosforilazione di p70S6K rispetto al placebo dopo l’esercizio fisico. Infine, la fosforilazione di AMPK è stata ancora misurata, ma è diminuita in entrambi i gruppi (cosa non molto sorprendente se si considera che ai partecipanti è stato somministrato 300ml di Gatorade).
Betaina e osmolarità:
Come scritto in precedenza, la Betaina è un importante osmolita. La concentrazione di Betaina nelle cellule muscolari è regolata dal trasportatore di betaina (BTG-1). Cholewa et al. ipotizzano che l’iperidratazione che ne deriva (a causa dell’aumento della pressione osmotica, vengono prelevati più liquidi nelle cellule) porti alla stimolazione della sintesi proteica e all’inibizione della proteolisi. In effetti, l’aumento del volume cellulare porta a un aumento della sintesi proteica e a una diminuzione della proteolisi negli epatociti. Il volume cellulare è direttamente correlato a questo aspetto [11]. È stato effettivamente riscontrato che la Betaina provoca un aumento del volume cellulare degli epatociti [12], ma le concentrazioni nel fegato sono circa 20 volte superiori a quelle del tessuto muscolare scheletrico [13] (almeno nei ratti), quindi resta da vedere in che misura questo fenomeno sarà significativo nel tessuto muscolare umano.
Conclusioni La letteratura è sufficiente a supportare i risultati mostrati, tra gli altri, dallo studio clinico di Cholewa et al [14]. L’effetto sulla via Akt è particolarmente interessante e suggerisce che la Betaina promuove l’ipertrofia muscolare attraverso un aumento della sintesi proteica e non è puramente il risultato, ad esempio, della ritenzione di liquidi. Ritengo inoltre che sarà interessante vedere in futuri studi clinici in che misura le dosi (più elevate) siano importanti. Probabilmente ci saranno molte ricerche di follow-up su questo tema.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Craig, Stuart AS. “Betaine in human nutrition.” The American journal of clinical nutrition 80.3 (2004): 539-549.
Cholewa, Jason M., et al. “Effects of betaine on body composition, performance, and homocysteine thiolactone.” Journal of the International Society of Sports Nutrition 10.1 (2013): 39.
Ortiz-Costa, Susana, M. M. Sorenson, and Mauro Sola-Penna. “Counteracting effects of urea and methylamines in function and structure of skeletal muscle myosin.” Archives of biochemistry and biophysics 408.2 (2002): 272-278.
Lang, Florian. “Mechanisms and significance of cell volume regulation.” Journal of the American College of Nutrition 26.sup5 (2007): 613S-623S.
Bera, Soumen, et al. “Enzymes of creatine biosynthesis, arginine and methionine metabolism in normal and malignant cells.” FEBS journal 275.23 (2008): 5899-5909.
Schwab, Ursula, et al. “Betaine supplementation decreases plasma homocysteine concentrations but does not affect body weight, body composition, or resting energy expenditure in human subjects.” The American journal of clinical nutrition 76.5 (2002): 961-967.
Lever, Michael, and Sandy Slow. “The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism.” Clinical biochemistry 43.9 (2010): 732-744.
Li, Yin, et al. “Homocysteine upregulates resistin production from adipocytes in vivo and in vitro.” Diabetes 57.4 (2008): 817-827.
Klement, Rainer J., and U. Kammerer. “Is there a role for carbohydrate restriction in the treatment and prevention of cancer.” Nutr Metab (Lond) 8.75 (2011): 75.
Apicella, Jenna M., et al. “Betaine supplementation enhances anabolic endocrine and Akt signaling in response to acute bouts of exercise.” European journal of applied physiology 113.3 (2013): 793-802.
Lang, Florian, et al. Functional significance of cell volume regulatory mechanisms.” Physiological reviews 78.1 (1998): 247-306.
Hoffmann, Lars, et al. “Osmotic regulation of hepatic betaine metabolism.” American Journal of Physiology-Gastrointestinal and Liver Physiology 304.9 (2013): G835-G846.
Slow, Sandy, et al. “Plasma dependent and independent accumulation of betaine in male and female rat tissues.” Physiological Research 58.3 (2009).
Cholewa, Jason M., et al. “Effects of betaine on body composition, performance, and homocysteine thiolactone.” Journal of the International Society of Sports Nutrition 10.1 (2013): 39.
L’uso di AAS può portare a una condizione chiamata eritrocitosi. Si tratta di una condizione in cui il corpo ha troppi eritrociti (globuli rossi) nel sangue. Talvolta si parla anche di policitemia. La percentuale del volume sanguigno occupata dai globuli rossi è chiamata ematocrito. Per gli uomini l’intervallo di riferimento va dal 41 al 51% e per le donne dal 36 al 47%. (Esistono leggere deviazioni da laboratorio a laboratorio in questi intervalli di riferimento).
I globuli rossi sono responsabili del trasporto dei gas. In particolare, trasportano l’ossigeno dai polmoni al resto del corpo. Inoltre, trasportano anche parte dell’anidride carbonica generata dal corpo verso i polmoni. I globuli rossi possono trasportare l’ossigeno grazie a una speciale proteina che contengono in grandi quantità: l’emoglobina. I livelli di emoglobina possono essere misurati anche nel sangue e il loro intervallo di riferimento va da 13,5 a 17 g/dL negli uomini e da 12 g/dL a 15,5 g/dL nelle donne.
I livelli di ematocrito e di emoglobina possono essere facilmente derivati l’uno dall’altro, in quanto i due valori sono in genere strettamente correlati. La concentrazione di emoglobina in g/dL moltiplicata per tre dà il valore di ematocrito in %. Pertanto, una persona con un livello di emoglobina di 15 g/dL ha un valore di ematocrito di circa il 45%.
Quando l’ematocrito diventa troppo basso, l’organismo ha una minore capacità di soddisfare il fabbisogno di ossigeno delle sue cellule. In altre parole: anemia. Questo può far sentire stanchi, deboli e facilmente senza fiato. Questo fenomeno è più evidente durante l’esercizio fisico (faticoso) che richiede una grande quantità di ossigeno.
Quando l’ematocrito diventa troppo alto, il problema è un altro. Più è alto, più il sangue diventa “denso”: aumenta la viscosità del sangue. Questo comporta un aumento del rischio di trombosi (correlata anche ai fattori di coagulazione). Vediamo come gli AAS aumentano l’ematocrito e di quanto. Dopodiché, analizziamo il rischio di trombosi e cosa si può fare al riguardo.
In che modo gli AAS causano l’eritrocitosi?
Struttura della Eritropoietina [EPO]
Due fattori principali sembrano coinvolti nell’eritrocitosi indotta dagli steroidi anabolizzanti. Uno è l’aumento dell’eritropoietina (EPO). Un ormone che talvolta fa notizia, in quanto utilizzato come doping per migliorare le prestazioni negli sport di resistenza come il ciclismo.
L’EPO viene prodotta nei reni in risposta a una ridotta disponibilità di ossigeno. L’EPO indica al midollo osseo, dove vengono prodotti i globuli rossi in un processo chiamato eritropoiesi, di aumentare la produzione di eritrociti. L’aumento dei globuli rossi aumenta la disponibilità di ossigeno e di conseguenza abbassa nuovamente l’EPO. Si crea quindi uno stretto equilibrio tra ematocrito ed EPO.
Struttura del Epcidina
L’altro fattore che sembra essere coinvolto è la soppressione di una proteina chiamata Epcidina. L’Epcidina è un regolatore chiave del metabolismo del ferro, in quanto regola la quantità di ferro assorbita dal tratto gastrointestinale [1]. Il ferro è una parte fondamentale dell’emoglobina: è responsabile del legame con l’ossigeno. La soppressione dell’Epcidina porta a un aumento dell’assorbimento di ferro, mentre un aumento dell’Epcidina ne diminuisce l’assorbimento. Se l’apporto di ferro è inadeguato, si sviluppa l’anemia. Al contrario, il sovraccarico di ferro (emocromatosi) sembra predisporre a un aumento dei livelli di emoglobina/ematocrito. In effetti, nell’emocromatosi (ereditaria) questi livelli possono essere elevati [2, 3].
Mentre la somministrazione di Testosterone per 20 settimane mostra un aumento dose-dipendente (fino a 600mg di Testosterone Enantato alla settimana) dell’emoglobina e dell’ematocrito, soprattutto negli uomini più anziani, non lo fa l’EPO [4]. Al contrario, la somministrazione di Testosterone ha portato alla soppressione dell’Epcidina sierica in uomini giovani e anziani [5]. Il dosaggio del Testosterone (fino a 600mg di Testosterone Enantato a settimana) era altamente correlato con l’ampiezza di questa soppressione. L’aumento dell’ematocrito in seguito alla somministrazione di Testosterone è più pronunciato negli uomini anziani. In particolare, questo studio ha rilevato che i maggiori aumenti dell’ematocrito negli uomini anziani durante la terapia con Testosterone erano correlati a una maggiore soppressione dell’Epcidina. Gli autori hanno inoltre misurato l’Epcidina in più momenti: al basale, alla prima settimana, alla seconda, quarta, ottava e ventesima settimana. In questo modo hanno potuto vedere come si è evoluta nel tempo. Nelle prime due settimane si è verificata una forte diminuzione, dopodiché i livelli di Epcidina sono risaliti verso i livelli di base, anche se ancora più bassi rispetto all’inizio.
Questo è uno dei problemi dello studio che non ha dimostrato alcun cambiamento nell’EPO dopo 20 settimane [4]. Forse l’EPO è aumentata inizialmente, ma è tornata ai valori di base al termine delle 20 settimane. In questo modo, si sarebbe perso un effetto iniziale. Inoltre, i livelli di EPO presentano un’ampia variabilità, il che rende difficile trovare cambiamenti statisticamente significativi, poiché è necessaria una dimensione del campione relativamente ampia.
Uno studio successivo ha misurato l’EPO e l’Epcidina in diversi momenti in uomini anziani con Testosterone basso che hanno ricevuto un placebo o il Testosterone per 6 mesi [6]. I valori sono stati misurati al basale, a 1 mese, 3 mesi, 6 mesi e 9 mesi (quindi 3 mesi dopo la sospensione del placebo o del Testosterone). I risultati hanno dimostrato un aumento iniziale dell’EPO in risposta al gel di Testosterone rispetto al placebo. In seguito, è sceso lentamente, non mostrando più alcuna differenza rispetto al placebo al termine dei 6 mesi:
Variazioni dei livelli di EPO durante la somministrazione di Testosterone o placebo. Il periodo di trattamento è stato di 6 mesi.
Una variazione simile, ma inversa, è stata riscontrata per l’Epcidina:
Ciò ha indotto gli autori a proporre che la somministrazione di Testosterone stabilisca un nuovo set point EPO/emoglobina. Normalmente, quando l’ematocrito aumenta, l’EPO diminuisce in risposta. In questa situazione, tuttavia, l’ematocrito aumenta, molto probabilmente come risultato dell’aumento dell’EPO e della diminuzione delle concentrazioni di Epcidina. Tuttavia, una volta stabilito l’aumento completo dell’ematocrito, i livelli di EPO rimangono simili a quelli precedenti, invece di diminuire. In altre parole, si stabilisce un nuovo setpoint. Ciò è illustrato dagli autori nella figura seguente:
La somministrazione di Testosterone sposta la curva log EPO-ematocrito, mentre il placebo non ha alcun effetto su questa relazione. Lo spostamento verticale indica un aumento dell’EPO per ematocrito come risultato del trattamento con Testosterone.
In sintesi, gli androgeni aumentano l’ematocrito/emoglobina attraverso un aumento iniziale dei livelli di EPO e una contemporanea diminuzione dei livelli di Epcidina, che poi scendono gradualmente ai livelli di base di fronte all’aumento dell’ematocrito/emoglobina: un nuovo set point EPO/emoglobina. I meccanismi d’azione responsabili e il loro contributo relativo a questo fenomeno sono ancora da stabilire.
In che misura gli AAS aumentano l’ematocrito/emoglobina?
In uno studio dose-risposta, con dosaggi fino a 600mg di Testosterone Enantato alla settimana, l’emoglobina ha mostrato un aumento di 1,42g/dL nei giovani uomini dopo 20 settimane [7]. Ciò si traduce in un aumento dell’ematocrito di poco superiore al 4%. L’aumento è maggiore negli uomini più anziani, che mostrano un aumento di 2,94g/dL di emoglobina in risposta allo stesso dosaggio per lo stesso periodo di tempo [8].
E per quanto riguarda i dosaggi più elevati? Fortunatamente, sembra esserci un limite alla misura in cui gli AAS possono aumentare l’ematocrito. Vediamo lo studio HAARLEM. Lo studio HAARLEM, come ormai sappiamo bene, è uno studio prospettico condotto dall’ambulatorio per i consumatori di steroidi anabolizzanti nei Paesi Bassi [9]. In breve, 100 consumatori di steroidi anabolizzanti sono stati seguiti nel tempo mentre si autosomministravano AAS. Il dosaggio medio, basato sulle informazioni riportate sull’etichetta, era di 898mg a settimana, rendendo così il loro ciclo di AAS abbastanza rappresentativo dell’uso comune da parte dei bodybuilder. Le misurazioni sono state effettuate prima, durante, 3 mesi dopo la fine del ciclo e 1 anno dopo l’inizio del ciclo. I ricercatori hanno riscontrato un aumento del 3% dell’ematocrito dei soggetti dello studio al termine del ciclo. Questo dato è in linea con l’aumento del 4% osservato nello studio nei giovani uomini. L’autore principale ha fatto sapere che l’aumento dell’ematocrito sembra stabilizzarsi a un dosaggio di androgeni di circa 500mg a settimana. Infine, vorrei aggiungere che i soggetti dello studio non hanno effettuato donazioni di sangue, quindi questo non è stato un fattore confondente.
Naturalmente, questi risultati presentano alcune variazioni. Alcuni rispondono agli AAS con un aumento dell’ematocrito maggiore di altri. Tuttavia, livelli molto elevati di ematocrito sembrano essere rari, come si può vedere nei grafici a scatola e baffi dei partecipanti allo studio HAARLEM (T0 = subito prima del ciclo di AAS, T1 = alla fine, T2 = 3 mesi dopo la cessazione dell’uso, T3 = 1 anno dopo l’inizio del ciclo):
L’aumento dell’ematocrito è un fattore di rischio per la trombosi.
Come già detto, un ematocrito più alto rende il sangue più viscoso. Di conseguenza, aumenta il rischio di eventi trombotici, sebbene vadano presi in considerazione anche i fattori della coagulazione. Questo fenomeno è più evidente nei pazienti affetti da policitemia vera. Malattia in cui il midollo osseo produce in eccesso globuli rossi e, comunemente, anche globuli bianchi e piastrine (trombociti). La concentrazione mediana di emoglobina riscontrata in un’ampia coorte di pazienti con policitemia vera era del 57% (negli uomini) [10]. In rare occasioni, questi pazienti possono avere valori di ematocrito superiori al 70%. Il valore più alto registrato in questa coorte è stato di ben il 78%. Nel 18% di questi pazienti è stata riscontrata una complicanza trombotica arteriosa prima o al momento della diagnosi, e una trombosi venosa nel 5,4% (L’età media era di 59 anni). Si tratta di un dato piuttosto elevato, poiché il tasso di incidenza della trombosi venosa è di circa 1 per 1.000 anni-persona nel gruppo di età 55-59 [11]. Il rischio di trombosi venosa è semplicemente piuttosto basso all’inizio della vita, ma inizia ad aumentare sostanzialmente dopo i 50-60 anni, con un picco di incidenza di 7,9 per 1.000 anni-persona nella fascia di età 80-84 anni. Il rischio per tutta la vita è dell’8%.
Naturalmente, considerare i pazienti affetti da policitemia vera non è un paragone equo: anche loro, come già detto, producono comunemente globuli bianchi e trombociti.
Diamo un’occhiata allo studio norvegese di Tromsø [12]. Hanno seguito 26.108 soggetti nel tempo (follow-up mediano di 12,5 anni) per mettere in relazione gli eventi tromboembolici venosi con i livelli di ematocrito. Hanno corretto per età, indice di massa corporea e fumo. Ovviamente ci sono altri fattori che avrebbero potuto influenzare i risultati e che non sono stati corretti, ma almeno questo corregge alcuni importanti fattori confondenti. Lo studio ha dimostrato che per ogni aumento del 5% dell’ematocrito c’era un aumento del 33% della probabilità di un evento tromboembolico venoso (negli uomini).
Per mettere in prospettiva questo aumento del rischio: i ricercatori hanno riscontrato circa 1,6 eventi tromboembolici venosi ogni 1.000 anni-persona negli uomini con un ematocrito medio (43-46%). Quindi, se si ha un livello medio di ematocrito, ci sarebbe l’1,6% di probabilità di un evento tromboembolico nei 10 anni successivi. Se il vostro ematocrito fosse del 5% più alto, cioè 48-51%, il rischio aumenterebbe del 33%: 2.1%. Naturalmente questo dato non è lineare per sempre, ma credo che fornisca una buona indicazione dell’aumento del rischio di trombosi venosa dovuto all’aumento dell’ematocrito indotto dagli AAS.
Un altro studio danese ha riscontrato un rischio di tromboembolismo venoso superiore del 26% negli uomini con un ematocrito superiore al 48% rispetto a quelli con un ematocrito compreso tra 41,1 e 45% [13]. Tuttavia, questo dato non era statisticamente significativo. Gli autori riportano anche i rischi di trombosi arteriosa nel cervello e nel cuore. Si è registrato un aumento non statisticamente significativo del 27% nel primo caso e un aumento statisticamente significativo del 46% nel secondo, confrontando gli uomini con un ematocrito superiore al 48% con quelli con un ematocrito compreso tra 41,1 e 45%. I tassi di incidenza della trombosi arteriosa nel cervello e nel cuore sono paragonabili a quelli della tromboembolia venosa.
L’aspetto importante è che, pur essendoci un aumento sostanziale del rischio relativo, il rischio assoluto rimane basso. Questo vale soprattutto per le persone di giovane età (meno di 40 anni) e in assenza di altri fattori di rischio (ad esempio, un precedente evento trombotico o un disturbo della coagulazione come il fattore V Leiden). L’opportunità o meno di intervenire per contrastare l’aumento del rischio di trombosi dovuto all’aumento dell’ematocrito AAS-indotto sembra quindi dipendere in larga misura dalla presenza di altri fattori di rischio. Anche se si potrebbe sostenere che, in un certo senso, l’uso di steroidi anabolizzanti è di per sé un fattore di rischio. Dopo tutto, ha un impatto negativo su più di un semplice ematocrito.
Trattamento:
Forse il trattamento migliore consiste nel ridurre notevolmente il dosaggio (ben al di sotto dei 500mg settimanali) o nell’interrompere del tutto l’uso di AAS [tornando in fisiologia controllata]. In questo modo si abbasserà l’ematocrito, con un effetto completo dopo un paio di mesi, e si annullerà il rischio. Tuttavia, questo non è probabilmente il metodo più gradito per contrastare questo problema.
Una pratica che vedo comunemente eseguita è quella di assumere un basso dosaggio di CardioAspirina (Acido Acetilsalicilico con gastroprotettore). Sebbene non influisca sui livelli di ematocrito, è ampiamente utilizzata per la prevenzione delle malattie cardiovascolari [14]. Più precisamente, è utilizzata nella prevenzione secondaria delle malattie cardiovascolari. Previene la coagulazione del sangue inibendo un enzima chiamato ciclossigenasi (COX) nei trombociti. Se da un lato riduce il rischio di trombosi, dall’altro aumenta il rischio di emorragie. E, naturalmente, non solo il sanguinamento del dito tagliato mentre si taglia il pollo. Ma anche di emorragie interne, come l’ictus emorragico. I benefici devono quindi essere attentamente soppesati rispetto ai rischi del suo utilizzo. Attualmente, le linee guida europee sulla prevenzione delle malattie cardiovascolari nella pratica clinica ne sconsigliano l’uso nella prevenzione primaria (anche se questo potrebbe cambiare per alcune popolazioni, come i diabetici) [15].
Un dosaggio comune a questo scopo è di 81 o 100mg al giorno. È difficile fare una raccomandazione generale per il suo uso, anche se si può fare un’argomentazione per coloro che hanno un rischio maggiore (ma forse è saggio interrompere del tutto l’uso di AAS sovraffisiologici in questo caso).
Come nota finale, nelle settimane successive all’interruzione del farmaco vi è un aumento del rischio di eventi trombotici [16, 17]. Pertanto, anche l’assunzione e la sospensione frequente del farmaco sono sconsigliate.
Prima di un eventuale uso, è caldamente consigliato, oltre il parere medico, un controllo accurato dei fattori che regolano la coagulazione: tempo di tromboplastinaparziale attivata (aPTT), tempo di protrombina (PT) Fibrinogeno, D-Dimero, Antitrombina e Inibitore C1 Esterasi.
Struttura cristallina della Nattochinasi di Bacillus subtilis natto.
Il meccanismo d’azione attualmente ipotizzato è l’inattivazione dell’inibitore 1 dell’attivatore del plasminogeno (PAI-1) che determina un contemporaneo aumento del livello di attivatore del plasminogeno tissutale (tPA). [Questo è stato dimostrato nel siero in seguito all’ingestione orale di Nattochinasi, anche se con una certa variabilità interindividuale (con nattokinase orale a 1.300 mg tre volte al giorno), con un aumento del tPA di breve durata, solo 3 ore dopo la somministrazione orale. L’aumento del tPA, almeno in vitro, è stato osservato con nattokinase trattata termicamente che non ha alcuna attività fibrinolitica intrinseca ed è probabilmente dovuto a peptidi bioattivi.
In vitro, la Nattochinasi mostra proprietà fibrinolitiche dirette che possono degradare la fibrina e ridurre il rischio di complicazioni cardiovascolari. Questo potrebbe però essere un depistaggio meccanicistico, in quanto questa attività diretta si perde con la distruzione dell’enzima (trattamento termico o digestione) e i peptidi della Nattochinasi sembrano aumentare i livelli di attivatore del plasminogeno, che ha anche un effetto fibrinolitico indiretto. La Nattochinasi ha mostrato per la prima volta la sua efficacia in seguito alla somministrazione orale nei cani, dove il trombo è stato indotto sperimentalmente e gli effetti anti-clottici sono stati confermati tramite angiografia (il coagulo è stato eliminato entro 5 ore dall’ingestione orale). Un piccolo studio condotto su 12 persone a cui sono stati somministrati 12 g di nattō per 2 settimane (seguito da un periodo in cui sono stati ingeriti fagioli di soia bolliti per 2 settimane al posto del nattō) ha osservato che il consumo di nattō era associato a una maggiore attività fibrinolitica; ciò è stato successivamente replicato con un supplemento di Nattochinasi a 1.300mg assunto tre volte al giorno.
Un intervento sull’uomo che ha utilizzato un integratore combinato (Nattochinasi e Picnogenolo) in 186 soggetti ad alto rischio di trombosi venosa profonda prima di un volo di lunga durata (7-8 ore) ha osservato che, mentre cinque persone nel gruppo placebo hanno sperimentato una trombosi venosa profonda e due un trombo superficiale (7,6% del gruppo placebo), nel gruppo di intervento non è stato rilevato alcun trombo. Nel gruppo di intervento è stata rilevata una diminuzione dell’edema (15%), mentre nel gruppo placebo è stato rilevato un aumento (12%).
Non ci sono prove sufficienti sull’uomo che utilizzano la Nattochinasi isolatamente e che valutano la formazione di trombi per raccomandarne l’uso come farmaco anti-clottico, anche se sembra esserci qualche promessa. L’assunzione di nattō con la dieta potrebbe conferire un effetto protettivo simile, secondo uno studio preliminare sull’argomento.
Come per la CardioAspirina, la necessità di utilizzo andrebbe valutata per via esami dei fattori della coagulazione.
Struttura molecolare del IP6
Nota: L’Acido Fitico, un estere diidrogenofosfato sestuplo dell’Inositolo (in particolare dell’isomero myo), chiamato anche Esacisfosfato di Inositolo (IP6) o polifosfato di inositolo, sembra avere effetti positivi sul controllo dell’ematocrito a dosaggi aneddotici di 1-2g/die. Ad oggi, non esiste sufficiente letteratura ad avvalorare l’effettiva efficacia terapeutica di questa molecola.
La flebotomia (salasso) è un modo per ridurre efficacemente l’ematocrito. Un modo per farlo è la donazione di sangue a una banca del sangue. Tuttavia, molti Paesi (e giustamente) limitano il numero di volte in cui è possibile farlo ogni anno. Vi sono paesi dove è limitato a cinque volte l’anno. Questo potrebbe non essere sufficiente a mantenere i valori nel range desiderato, dato che uno studio ha rilevato livelli di emoglobina persistentemente elevati in occasione di visite ripetute in un numero elevato di pazienti TRT che hanno donato il sangue [18]. Se la donazione di sangue non è sufficiente, si può sempre consultare un medico generico per eseguire una flebotomia terapeutica a intervalli più frequenti.
Procedura schematica della flebotomia
Tenete presente che ad ogni donazione di sangue si perde ferro. Di conseguenza, si corre il rischio di esaurire le proprie riserve di ferro e, consequenzialmente, anche l’emoglobina rimarrà molto bassa e si diventerà temporaneamente anemici. È possibile contrastare questo fenomeno integrando il ferro, ma questo riduce drasticamente il tempo necessario all’organismo per recuperare i livelli di emoglobina/ematocrito [19]. (nello studio è stato utilizzato un dosaggio di 37,5mg di ferro elementare al giorno). Pertanto, a intervalli più frequenti, è consigliabile un controllo con analisi del sangue.
Ricordiamoci, inoltre, che una flebotomia non monitorata nei tempi di prelievo può portare a rebound dell’ematocrito con peggioramento del quadro clinico ematico.
Conclusioni:
Abbiamo analizzato le cause del eritrocitosi AAS-indotta. Abbiamo visto come può essere gestita e risolta. Sappiamo che l’uso di anticoagulanti ha senso soprattutto se ci sono alterazioni dei fattori di coagulazione, e che il loro uso non è scevro da effetti avversi potenzialmente nefasti.
Non sarebbe nemmeno da dire, ma visto il livello del lettore medio nel capire i concetti è giusto sottolineare che l’uso di farmaci per ridurre l’eritropoiesi e, di conseguenza, l’ematocrito, come l’Hydroxyruea, l’Interferone alfa-2b, il Ruxolitinib e il Busulfano è caldamente sconsigliato per i possibili e gravi effetti collaterali che questi farmaci danno; e ovviamente perchè, come si è visto, non è necessario utilizzarli in caso di eritrocitosi AAS-indotta.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Rauf, Abdur, et al. “Hepcidin, an overview of biochemical and clinical properties.” Steroids 160 (2020): 108661.
Barton, James C., Luigi F. Bertoli, and Barry E. Rothenberg. “Peripheral blood erythrocyte parameters in hemochromatosis: evidence for increased erythrocyte hemoglobin content.” Journal of Laboratory and Clinical Medicine 135.1 (2000): 96-104.
Khan, Adnan Aman, et al. “Polycythemia and Anemia in Hereditary Hemochromatosis.” Cureus 12.4 (2020).
Coviello, Andrea D., et al. “Effects of graded doses of testosterone on erythropoiesis in healthy young and older men.” The Journal of Clinical Endocrinology & Metabolism 93.3 (2008): 914-919.
Bachman, Eric, et al. “Testosterone suppresses hepcidin in men: a potential mechanism for testosterone-induced erythrocytosis.” The Journal of Clinical Endocrinology & Metabolism 95.10 (2010): 4743-4747.
Bachman, Eric, et al. “Testosterone induces erythrocytosis via increased erythropoietin and suppressed hepcidin: evidence for a new erythropoietin/hemoglobin set point.” Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences 69.6 (2014): 725-735.
Bhasin, Shalender, et al. “Testosterone dose-response relationships in healthy young men.” American Journal of Physiology-Endocrinology And Metabolism (2001).
Bhasin, Shalender, et al. “Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle.” The Journal of Clinical Endocrinology & Metabolism 90.2 (2005): 678-688.
Smit, Diederik L., et al. “Baseline characteristics of the HAARLEM study: 100 male amateur athletes using anabolic androgenic steroids.” Scandinavian journal of medicine & science in sports 30.3 (2020): 531-539.
Tefferi, Ayalew, et al. “Survival and prognosis among 1545 patients with contemporary polycythemia vera: an international study.” Leukemia 27.9 (2013): 1874-1881.
Bell, Elizabeth J., et al. “Lifetime risk of venous thromboembolism in two cohort studies.” The American journal of medicine 129.3 (2016): 339-e19.
Brækkan, Sigrid K., et al. “Hematocrit and risk of venous thromboembolism in a general population. The Tromsø study.” haematologica 95.2 (2010): 270.
Warny, Marie, et al. “Arterial and venous thrombosis by high platelet count and high hematocrit: 108 521 individuals from the Copenhagen General Population Study.” Journal of Thrombosis and Haemostasis 17.11 (2019): 1898-1911.
Baigent, Colin, et al. “Aspirin in the primary and secondary prevention of vascular disease: collaborative meta-analysis of individual participant data from randomised trials.” Lancet 373.9678 (2009): 1849-1860.
F. Hobbs, M. Piepoli, A. Hoes, S. Agewall, C. Albus, C. Brotons, A. Catapano, M. Cooney, U. Corra, B. Cosyns, et al. 2016 european guidelines on cardiovascular disease prevention in clinical practice. European Heart Journal, 37(29):2315–2381, 2016.
M. Lordkipanidzé, J. G. Diodati, and C. Pharand. Possibility of a rebound phenomenon following antiplatelet therapy withdrawal: a look at the clinical and pharmacological evidence. Pharmacology & therapeutics, 123(2):178–186, 2009.
L. A. G. Rodríguez, L. C. Soriano, C. Hill, and S. Johansson. Increased risk of stroke after discontinuation of acetylsalicylic acid a uk primary care study. Neurology, pages WNL–0b013e31820d62b5, 2011
B. Chin-Yee, A. Lazo-Langner, T. Butler-Foster, C. Hsia, and I. Chin-Yee. Blood donation and testosterone replacement therapy. Transfusion, 57(3):578–581, 2017
Kiss, Joseph E., et al. “Oral iron supplementation after blood donation: a randomized clinical trial.” Jama 313.6 (2015): 575-583.
Dell’Insulina ho già parlato abbondantemente non molto tempo fa attraverso la serie di quattro articoli dedicati a questo peptide nella ricorrenza dei cento anni dalla sua scoperta e prima applicazione in medicina (per approfondire clicca qui, qui, qui e qui). Nella quarta e ultima parte della prima citata serie di articoli, mi concentrai sulle pratiche protocollari dell’Insulina utilizzate in campo culturistico e, tra queste, vi era anche il protocollo di GH/Insulina nelle sue varianti applicative. Vi starete sicuramente chiedendo del perchè io scriva nuovamente di un argomento già trattato, oltretutto di recente. Beh, la risposta è abbastanza semplice: è un argomento che richiede per complessità degli approfondimenti aggiuntivi. Soprattutto alla luce delle “leggende da spogliatoio” e della limitatezza dei divulgatori “naturnazi”.
Come mi è già capitato di precisare, la visione dell’Insulina come ormone puramente anticatabolico è mentalmente riduttivo tanto quanto affermare che, di per se, l’Insulina sia “l’ormone anabolico per eccellenza”. Ed ecco perchè sopra ho specificato come sia necessario chiarire alcuni aspetti in contrapposizione al settarismo “magna e spigni” e “orcojo natty”. Sono volutamente ironico e provocatorio, ovviamente.
Giova anche ripetere che non sono l’unico a ritenere che la narrazione secondo la quale l’Insulina sia prettamente anti-catabolica sia simile alla visione precedente a quella di Bhasin (2001), secondo cui gli AAS non avrebbero effetti anabolici nel muscolo-scheletrico. Attualmente, la comunità (o, per meglio dire, “cupola”) scientifica vuole far credere che l’Insulina esogena non sia particolarmente anabolizzante nel muscolo-scheletrico umano. Le nozioni e i dati provenienti dai miei appunti sull’Insulina, in parte già riportati negli articoli dedicati prima citati, sono una forte contestazione di questo pregiudizio istituzionale (analogo alla visione pre-Bhasin sugli AAS). Il punto cieco istituzionale deriva dalla mancanza di studi sull’uso dell’Insulina esogena con:
dosaggio sovrafisiologico;
somministrazione sistemica;
iperamminoacidemia;
associazione a rhGH sovrafisiologico (e, più spesso, a AAS).
Ed è proprio su questi punti che si diramerà la mia disamina in questo articolo: valutazione teorico/pratica della co-somministrazione di GH e Insulina in condizioni sovra-fisiologiche.
Il motivo? Oltre le affermazioni arbitrarie e riduttive di alcuni divulgatori, vi è anche il risultato emerso dallo studio di Fryburg et al. i quali proposero che il rhGH e l’Insulina esogena lavorassero in modo incrociato, che la crescita fosse una conseguenza del turnover delle proteine muscolari piuttosto che dell’azione ormonale ipo/iperglicemica. La co-somministrazione di livelli endogeni (quindi fisiologici) di Insulina e rhGH sembrò dimostrare semplicemente una attenuazione degli effetti anabolici proteici del GH e che non fosse additiva. Ovviamente questa conclusione era limitata da: 1) il contesto di “picco ormonale” e della sua curva mantenuta in fisiologia e, cosa da non trascurare, 2) è stata trascurata l’importanza della disponibilità intracellulare di AA sulla sintesi proteica.
Ma andiamo per ordine…
Breve ripasso su azione dell’Insulina e meccanismi nell’ipertrofia muscolare:
L’effetto primario dell’Insulina nel muscolo scheletrico è sul gradiente elettrochimico di transmembrana, dove induce un’iperpolarizzazione nelle cellule muscolari attivando direttamente la pompa Na⁺-K⁺-ATPasi (46). [Biolo, G. et al. (1995)]. L’Insulina aumenta anche il numero di pompe Na+/K+-ATPasi nella membrana, determinando uno spostamento intracellulare del potassio e causando ipopotassiemia (basso livello di K/potassio) nello spazio extracellulare del sangue (siero).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
L’Insulina stimola l’espressione genica di MHCα (isoforma lenta) [Toniolo, L. (2005).] nel muscolo scheletrico e dell’albumina nel fegato (Dillman, 1988).
L’Insulina sembra stimolare in egual misura le fibre di tipo I e II [Albers et al. 2014]
Il muscolo scheletrico è un tessuto eterogeneo composto da diversi tipi di fibre. Gli studi suggeriscono che il metabolismo del glucosio mediato dall’insulina è diverso tra i vari tipi di fibre muscolari. Abbiamo ipotizzato che le differenze siano dovute all’espressione/regolazione specifica di elementi di segnalazione dell’insulina e/o di enzimi metabolici. Sono stati preparati pool di fibre di tipo I e II da biopsie dei muscoli del vasto laterale di soggetti magri, obesi e diabetici di tipo 2 prima e dopo un clamp iperinsulinemico-euglicemico. Le fibre di tipo I rispetto a quelle di tipo II presentano livelli proteici più elevati di recettore dell’insulina, GLUT4, esochinasi II, glicogeno sintasi (GS) e piruvato deidrogenasi-E1α (PDH-E1α) e un contenuto proteico inferiore di Akt2, TBC1 domain family member 4 (TBC1D4) e TBC1D1. Nelle fibre di tipo I rispetto a quelle di tipo II, la risposta di fosforilazione all’insulina era simile (TBC1D4, TBC1D1 e GS) o ridotta (Akt e PDH-E1α). Le risposte di fosforilazione all’Insulina aggiustate per il livello di proteine non erano diverse tra i tipi di fibre. Indipendentemente dal tipo di fibra, la segnalazione dell’insulina era simile (TBC1D1, GS e PDH-E1α) o ridotta (Akt e TBC1D4) nel muscolo di pazienti con diabete di tipo 2 rispetto a soggetti magri e obesi. Concludiamo che le fibre muscolari umane di tipo I rispetto a quelle di tipo II hanno una maggiore capacità di gestione del glucosio ma una sensibilità simile alla fosforegolazione da parte dell’Insulina.[Albers et al. 2014]
Livelli proteici più elevati di IRβ (+16%), esochinasi II impegna il glicogeno nella cellula, GLUT4 trasporta il glucosio nella cellula e complesso II della catena di trasporto degli elettroni ciclo mitocondriale/Kreb sono stati riscontrati nelle fibre di tipo I rispetto a quelle di tipo II.
Akt, mTOR: il contenuto proteico di Akt2 era più basso (-27%) nelle fibre di tipo I rispetto a quelle di tipo II. Gli aumenti medi della fosforilazione di [Akt] sotto stimolazione insulinica (ipertrofia) sono stati rispettivamente di 5,8 e 3,5 volte nelle fibre di tipo I e di 6,1 e 3,7 volte nelle fibre di tipo II. La risposta relativa all’insulina è stata simile tra i tipi di fibre.
Le fibre umane di tipo I hanno una maggiore abbondanza di trasporto (+29% GLUT4), fosforilazione (+470% HKII) e ossidazione (+35% complesso ETC II e +34% complesso piruvato deidrogenasi) del glucosio e di sintesi del glicogeno (+35%) rispetto alle fibre di tipo II.
Le fibre di tipo I possiedono una maggiore capacità di immagazzinamento del glicogeno.
Le differenze apparenti tra i tipi di fibre nella fosforilazione stimolata dall’insulina di Akt, NDRG1 (a valle di mTOR [queste sono le nostre vie dell’ipertrofia])… sono state eliminate quando sono state aggiustate per… l’abbondanza proteica. Questi risultati suggeriscono una sensibilità simile delle fibre muscolari di tipo I e di tipo II alla regolazione da parte dell’insulina delle proteine analizzate. [Albers et al. 2014]
Questi dati [Albers et al. 2014] supportano l’idea che, piuttosto che promuovere in modo preferenziale l’anabolismo proteico muscolare nelle fibre a contrazione lenta, entrambe le fibre muscolari di tipo I e II siano ugualmente sensibili agli effetti anabolici proteici dell’insulina esogena.
L’iperinsulinemia nei soggetti normali diminuisce acutamente le concentrazioni plasmatiche di aminoacidi come conseguenza del deposito netto di proteine. [Biolo, G., & Wolfe, R. R. (1993)].
Una volta secreta dalle beta-cellule pancreatiche, l’insulina circola nel flusso sanguigno con un’emivita di circa 12 minuti. Numerosi tessuti e organi esprimono il recettore dell’insulina e si attivano diverse azioni (Sonksen 2001), alcune delle quali sono di importanza generale e di particolare interesse per gli sport d’élite.
L’effetto centrale dell’insulina, in concerto con altri ormoni come il glucagone o la somatostatina, è il controllo dei livelli di glucosio nel sangue. La secrezione di insulina in risposta a concentrazioni elevate di glucosio (ad esempio, postprandiale) inibisce la produzione epatica di glucosio, abbassando così i livelli di glucosio nel sangue. Il glucagone, invece, composto da 29 residui aminoacidici, è un ormone controregolatore dell’insulina. Aumenta i livelli di glucosio plasmatico in risposta all’ipoglicemia indotta dall’insulina e svolge un ruolo importante nell’omeostasi del glucosio aumentando la gluconeogenesi e diminuendo la glicolisi. Il terzo ormone pancreatico rilevante per il controllo dei livelli di glucosio nel sangue è la somatostatina, un peptide di 14 residui che esercita effetti inibitori sulla secrezione di insulina ma non sulla sua biosintesi. L’intero meccanismo della sua azione non è ancora stato chiarito, ma gli studi suggeriscono un effetto paracrino che inibisce l’esocitosi dell’insulina dalle cellule adiacenti alle cellule D produttrici di somatostatina (Reichlin 1983).
Tuttavia, gli effetti dell’insulina sull’intero organismo sono molteplici e complessi. L’insulina provoca, ad esempio, la traslocazione del GLUT-4 (il trasportatore di glucosio che si trova prevalentemente nel muscolo scheletrico e nel tessuto adiposo) dalle vescicole intracellulari alla membrana cellulare e, quindi, aumenta la velocità di ingresso del glucosio per una determinata concentrazione nel tessuto bersaglio. Un eccesso di glucosio trasferito nelle cellule stimola successivamente la formazione di glicogeno (Halse et al. 2001; Yeaman et al. 2001), un fatto di notevole interesse negli sport di resistenza, dove la quantità di glicogeno immagazzinata nelle cellule muscolari può influenzare le prestazioni atletiche. Inoltre, il metabolismo proteico (muscolare) è significativamente influenzato dalle proprietà chaloniche (Rooyackers e Nair 1997; Sonksen 2001) e stimolanti dell’insulina (Biolo et al. 1995; Biolo e Wolfe 1993; Tipton e Wolfe 2001; Wolfe 2000, 2005). Grazie all’effetto anticatabolico dell’insulina, la disgregazione proteica è significativamente ridotta e consente la conservazione degli elementi muscolari contrattili. Inoltre, in numerosi studi che hanno dimostrato le proprietà anaboliche dell’insulina, sono stati osservati effetti di stimolazione della sintesi, utilizzando ad esempio analoghi marcati con isotopi stabili. L’anabolismo, tuttavia, è fortemente dipendente dalla disponibilità di aminoacidi (Fujita et al.2006; Garlick e Grant 1988; Zhang et al.1999).[Thevis, M., Thomas, A., & Schänzer, W. (2009)]
Giova anche sottolineare i punti chiave dell’effetto dell’Insulina sul metabolismo delle proteine:
aumenta la velocità di trasporto di alcuni aminoacidi nei tessuti
aumenta il tasso di sintesi proteica nel muscolo, nel tessuto adiposo, nel fegato e in altri tessuti
diminuisce il tasso di degradazione delle proteine nel muscolo (e forse anche in altri tessuti)
diminuisce la velocità di formazione dell’urea. Questi effetti dell’insulina servono a favorire la sintesi di carboidrati, grassi e proteine.[Newsholme, E., & Dimitriadis, G. (2001).]
Un ormone anabolico può indurre l’ipertrofia del muscolo scheletrico attraverso:
l’aumento della MPS o
la riduzione della proteolisi.
Inoltre, il trasporto transmembrana degli AA (AA trafficking) può essere sotto controllo ormonale.
L’Insulina induce l’ipertrofia del muscolo scheletrico attraverso tutti e tre questi aspetti del metabolismo proteico muscolare:
profondamente (“il tasso di sintesi frazionale delle proteine muscolari era superiore del 65% (P = 0,02) durante l’infusione di insulina”) [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)].
in misura modesta (nelle proteine non miofibrillari del muscolo scheletrico)
influenzando l’attività di almeno quattro distinti sistemi di trasporto degli AA (non è un mediatore primario dell’azione dell’insulina sull’ipertrofia del muscolo scheletrico):
Primariamente: trasporta gli AA con catene laterali polari corte, come l’Alanina e la Glicina; non limita il ritmo della sintesi proteica.
ASC
Nᵐ, sostanzialmente
Xc [Biolo, G., Declan Fleming, R. Y., & Wolfe, R. R. (1995)]
In pazienti gravemente ustionati l’infusione di Insulina (10 – 12 UI/ora, paziente di 70 kg) ↑MPS senza influenzare la MPB.[Hadley JS et al.2002]. Nessun cambiamento nell’assorbimento degli AA nel muscolo scheletrico, suggerendo che gli effetti anabolici muscolari derivino da un più efficiente riutilizzo degli AA generati dalla proteolisi.[Hadley JS et al.2002]
La sintesi proteica del muscolo scheletrico indotta dall’Insulina è modulata dalle variazioni di:
del flusso sanguigno muscolare e
disponibilità di AA. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)]. In altre parole, l’insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in mancanza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S., Rasmussen, B. B., Cadenas, J. G., Grady, J. J., & Volpi, E. (2006)].
Adesso è giunto il momento di aprire una parentesi sul GH…
Effetti dell’Ormone della Crescita e del IGF-1 sulla sintesi proteica:
Struttura molecolare di hGH
La maggior parte degli studi svolti sul GH, quando considerati nel loro insieme, suggeriscono che il peptide in questione sia anabolico. Più specificamente, il GH è anabolico perché stimola la sintesi proteica di tutto il corpo con o nessun effetto, o un effetto soppressivo, sui tassi di degradazione proteica. [Møller N et asl. 2007] Tuttavia, quando si approfondisce l’argomento, le cose diventano un po’ meno chiare dal momento che i risultati degli studi tendono ad essere diversi. I diversi risultati sono un riflesso diretto dell’ immensa complessità del GH.
Il GH esplica i suoi effetti sulla sintesi proteica legandosi prima con il suo recettore specifico (GHR) e successivamente aumentando la trascrizione del gene muscolare attraverso i percorsi di segnalazione a valle, in definitiva attivando la segnalazione del mTOR. [Hayashi AA et al.2007] Questi effetti si manifestano in acuto, spesso si verificano in pochi minuti e sono di natura simile all’Insulina, usando molte delle stesse vie anaboliche. [Costoya JA et al.1999] La rapida comparsa di questi cambiamenti metabolici legati alle proteine suggerisce che essi siano direttamente causati dal GH e non secondariamente mediati tramite l’IGF-1 [Copeland KC et al.1994]. L’impatto del GH sulla proteolisi, d’altro canto, è molto probabilmente di natura indiretta. A detta di tutti, ciò ha più a che fare con i suoi effetti inibitori sull’Insulina, che è stata vista avere effetti diretti sulla proteolisi.[Umpleby AM et al.]
È stato dimostrato che l’IGF-1 inibisce allo stesso modo la disgregazione proteica a livello sistemico [Fryburg DA et al.1994], il che avrebbe senso a causa della stretta correlazione con il GH. Quando gli amminoacidi e l’Insulina vengono somministrati ai soggetti esaminati, già sottoposti a somministrazione di IGF-1, è stato dimostrato, sia negli uomini che negli animali, che i tassi di sintesi proteica aumentano a livello sistemico [Jacob R et al.1996]. Vale la pena notare che l’IGF-1 è bifasico nel senso che quando è somministrato ad alto dosaggio e, di conseguenza, i livelli serici diventano elevati, il suo comportamento cambia passando da un azione “GH-simile” ad una “insulino-simile”.
Per riassumere, il GH è molto adatto per prevenire la degradazione proteica, e lo fa in una vasta gamma di condizioni alimentari a ristretto apporto calorico. Tuttavia, in presenza di un apporto energetico sufficiente (e di AA), il suo comportamento cambia. L’effetto principale del GH sul metabolismo proteico è volto dapprima a creare un ambiente con una ossidazione amminoacidica ridotta [Buijs MM et al.2002] e successivamente ad aumentare la sintesi proteica sistemica. [Gibney J et al.2005]
L’Ormone della Crescita è noto per aumentare i livelli di IGF-1 circolante così come la sintesi locale di IGF-1, in modo autocrino. Entrambe queste azioni giocano un ruolo fondamentale nella regolazione della massa muscolare e, quindi, in conseguenza di ciò, risulta utile comprendere meglio come la secrezione di GH porta ad un aumento dei livelli endocrino e autocrino di IGF-1.
La stragrande maggioranza dell’Ormone della Crescita negli adulti sani è secreto dalla ghiandola pituitaria e, più specificamente, dalle cellule somatotrope nel lobo anteriore mediate dal fattore di trascrizione Prophet of Pit-1 (PROP1).[Hemchand K et al.2011] Il GH può anche essere sintetizzato localmente in molti tessuti come il cervello, le cellule immunitarie, il tessuto mammario, i denti e la placenta che sono tutti al di fuori della regolazione dell’ipofisi. [Waters MJ et al.1999] Questo supporta l’idea secondo cui il GH abbia ruoli autocrini oltre ai suoi già consolidati ruoli endocrini.
Il GH circolante si lega al GHR, recettore dell’Ormone della Crescita appartenente alla superfamiglia delle proteine transmembrana presenti in tutte le cellule del corpo e che include il recettore della Prolattina e un certo numero di recettori delle citochine [Zou L et al. 1997]. I livelli sulla superficie cellulare, o la densità recettoriale, dei GHR sono il determinante principale dell’affinità di legame del GH con le cellule. La traslocazione del GHR, cioè il recettore che si sposta dal nucleo di una cellula alla sua membrana esterna, è direttamente inibita dall’IGF-1 – che è uno dei molti meccanismi di feedback che esistono tra questi ormoni strettamente correlati. Mediante l’inibizione della traslocazione dei GHR, l’IGF-1 contribuisce direttamente ad abbassare la reattività di queste cellule a uno stimolo esterno di GH. [Leung KC et al.1997]
I GHR esistono sulle membrane cellulari come omodimeri preformati e inattivi. Questo significa che il GHR ha due dimeri identici del recettore della proteina, e questi omodimeri saranno sempre accoppiati al JAK2 quando sono privi di attività enzimatica. Questo accoppiamento al JAK2 provoca un’azione inibitoria complessiva sul recettore. [Sawada T et al.2017] In altre parole, il GHR rimane dormiente finché non viene attivato come parte del processo di legame GH/GHR. Quando una molecola di GH si lega al GHR, si verifica un cambiamento strutturale all’interno del GHR che si traduce in movimento effettivo dei domini intracellulari del recettore separatamente l’uno dall’altro. Questo smorza quell’azione inibitoria data dal JAK2 e consente loro di attivarsi l’un l’altro.[Brown RJ et al.2005]
Successivamente, la molecola di GH si lega sequenzialmente a uno dei due omodimeri di GHR e il completamento di questo processo di legame facilita le interazioni con il secondo omodimero. Dopo questo, i domini intracellulari di questo dimero-GHR appena formato subiscono una rotazione effettiva. La rotazione del nuovo dimero-GHR consente ai domini chinasi del JAK2 di essere in contatto l’uno con l’altro, consentendo loro di transactivare e ciascuno successivamente si lega a una molecola di JAK2. [Lanning NJ et al.2006] Ciascuna molecola di JAK2 eseguirà quindi la fosforilazione incrociata (attivazione) dei residui di Tirosina, e sono proprio questi residui che formano “punti di attracco” per molte delle diverse molecole di segnalazione che costituiscono le vie di segnalazione a valle, e alla fine portano all’espressione genica. [Brooks AJ et al.2010] Uno dei più importanti percorsi a valle di maggior interesse per l’argomento trattato è il percorso JAK-STAT. Questo percorso è di vitale importanza sia per la trascrizione epatica di IGF-1 dal GH che per molti dei processi anabolici mediati dal GH all’interno del tessuto muscolare.
IGF-1:
Struttura molecolare di IGF-1
Gli IGF sono una famiglia di peptidi, in gran parte dipendenti dal GH, che mediano molte delle azioni di stimolo della crescita date dal GH.[Cohen P. et al.2006] Il fegato è il principale responsabile di tutta la produzione endocrina di IGF-1, con circa il 75% della sintesi a carico epatico sotto la regolazione del GH.[Laron Z. et al. 2001] Ciò presuppone che ci sia un apporto macro-calorico sufficiente e livelli elevati di Insulina portale.[D’Ercole AJ et al.1984] La sintesi autocrina di IGF-1 è anche regolata dal GH, in aggiunta ad altri fattori autocrini dipendenti dal tessuto.[Gunawardane K et al.2000]
Alla famiglia degli IGF appartengono oltre dieci proteine strutturalmente simili tra cui IGF-1, IGF-2, Insulina, Relaxina e Pro-Insulina.[Lu C et al.2005] Sono tutti altamente omologhi sia nella struttura che nella funzione e gli effetti metabolici dell’IGF-1 sono stati definiti “insulino-simili” proprio a causa delle somiglianze e dei percorsi che condividono l’uno con l’altro. L’IGF-1 ha un’omologia di sequenza aminoacidica superiore al 50% con l’Insulina e il recettore del IGF-1 ha un’omologia della sequenza aminoacidica del 60% con il recettore dell’Insulina.[Samani AA et al.2006] A causa di queste somiglianze strutturali, i membri della famiglia IGF possono spesso legarsi con i recettori nativi in modo “incrociato”.[Kim JJ et al.2002] Per riassumere brevemente queste relazioni di legame, la molecola di IGF-1 si lega con il recettore del IGF-1 con un elevata affinità, tuttavia sia l’IGF-2 che l’Insulina possono legarsi al recettore del IGF-1, ma con una affinità significativamente inferiori. L’IGF-2 si lega al recettore del IGF-2 con una elevata affinità, e l’IGF-1 si lega a questo recettore con un’affinità inferiore mentre l’Insulina non presenta alcun legame con esso.
La famiglia dei recettori IGF ha densità che variano significativamente in base ai tipi di cellule in cui sono presenti.[Clemmons DR. et al. 2012] Questo è uno dei motivi per cui l’Insulina e l’IGF-1 possono avere diverse azioni metaboliche nonostante siano strutturalmente simili. Cellule come gli epatociti e gli adipociti hanno molti più recettori dell’Insulina rispetto ai recettori del IGF-1. Al contrario, le cellule muscolari lisce vascolari situate nei vasi sanguigni hanno un numero significativamente più elevato di recettori del IGF-1 rispetto ai recettori dell’Insulina.
E’ necessario comprendere che la famiglia dei recettori IGF è anche attivata dalla Tirosina Chinasi che, come ora sappiamo, porta alla fosforilazione dei substrati, all’attivazione delle vie cellulari e infine all’espressione genica e alla sintesi proteica.[Cohen P. et al.2006] L’attivazione del recettore del IGF-1 sembra essere indipendente dall’isoforma da cui è stato prodotto l’IGF-1. Inoltre, si noti che entrambi i tipi di recettori IGF sono stati trovati nelle cellule muscolari umane.[Shimizu M et al.1986]
Nel flusso sanguineo, l’IGF-1 esiste principalmente in forma legata a proteine leganti l’IGF (IGFBP). La superfamiglia IGFBP comprende sei proteine ad alta affinità che vanno dal IGFBP-1 al IGFBP-6, nonché un certo numero di proteine a bassa affinità denominate proteine legate all’IGFBP.[Hwa V et al.1999] Quasi il 95% di tutto l’IGF-1 circolante esiste in forma legata, con circa il 75% legato specificamente con l’IGFBP-3.[Firth SM et al.2002] Una piccola frazione di IGF-1 (normalmente inferiore al 5%) può anche esistere in forma libera, e queste molecole non legate agiscono come regolatore negativo della secrezione di GH. Gli IGFBP possono legarsi con l’IGF-1 e l’IGF-2, ma non con l’Insulina. [Bach LA et al.1993] L’IGF-1 legato esiste più comunemente in un complesso ternario da 150-kDa mentre è nel circolo ematico. Questo complesso ternario è costituito da una molecola di IGF-1, dal IGFBP-3 e dalla subunità labile acida (ALS) – sebbene possa esistere in un complesso binario con altri IGFBP.[Duan C. et al.1996] Questi complessi servono a scopi come l’aumento della biodisponibilità degli IGF circolanti, estendendo la loro emivita serica, trasportando gli IGF alle cellule bersaglio e modulando l’interazione degli IGF con i loro rispettivi recettori di membrana posti sulla superficie delle cellule.[Hwa V et al.1996] Ad esempio, nel plasma, il complesso ternario stabilizza IGF-1, aumentando significativamente la sua emivita da meno di 5 minuti a oltre 16 ore in alcuni casi.[Firth SM et al. 2002]
Struttura molecolare IGFBP-1
Gli IGFB sembrano normalmente inibire l’azione degli IGF, e questo perché competono con i recettori IGF per l’affinità di legame con gli IGF.[Collett-Solberg PF et al.2000] Tuttavia, non è sempre così, poiché gli IGFBP sono anche in grado di potenziare le azioni dell’IGF, potenzialmente facilitando la consegna dell’IGF al recettore.[Wetterau LA et al. 1999] Sebbene esista un’interazione piuttosto complessa, basti ricordare che il ruolo principale degli IGFBP è quello di trasportare gli IGF dal flusso ematico ai tessuti periferici. Una volta che ciò è avvenuto, gli IGFBP vengono rilasciati dai complessi binari e ternari mediante proteolisi o tramite legame alla matrice extracellulare del recettore del IGF-1.[Parker A et al.1999] Una volta rilasciate, le molecole di IGF-1 diventano libere, attive e possono quindi esplicare la loro azione.[Monzavi R et al.2002]
Una volta nei tessuti, gli IGFBP modulano le azioni dell’IGF in quanto hanno una maggiore affinità per il sito recettore rispetto all’IGF stesso [Velloso CP et al.2008], tuttavia essi possono anche esercitare effetti indipendenti dall’IGF.[Jones JI et al.1995] Alcuni degli effetti diretti del IGFBP che sono già stati chiariti includono l’inibizione della crescita, l’induzione diretta dell’apoptosi e la modulazione degli effetti dei fattori di crescita non-IGF.[Cohen P. et al.2006]
Additività Insulina/rhGH sovrafisiologici:
Come ho già riportato in passato, la somministrazione di Insulina esogena capace di creare un livello ematico oltre i 1.200pmol/lt ha effetti significativi e tangibili sulla sintesi proteica muscolare. Ma non si esaurisce tutto a meri e limitati numeri statistici.
La sintesi proteica nel muscolo-scheletrico indotta dall’Insulina è modulata dalle variazioni di:
1- del flusso sanguigno muscolare e
2- disponibilità di AA. [Fujita, S. et al. (2006).]. In altre parole, l’Insulina deve essere considerata nel contesto di (concentrazione di AA x flusso [cioè, aumento dell’apporto di AA al muscolo]). [Wolfe, R. R. (2000)].
Ciò rappresenta una controargomentazione rispetto al punto di vista di Wolfe, secondo cui l’Insulina deve stimolare la reincorporazione dell’AA dall’MPB all’MPS intracellulare… Wolfe sosteneva che, in assenza di ciò, l’Insulina deve aumentare la disponibilità di AA in altro modo (rispetto alla reincorporazione), ad esempio tramite ingestione o infusione (che causa ipoaminoacidemia). [Wolfe, R. R. (2000)].
In particolare, in tutti gli studi in cui la MPS è stata stimolata dall’Insulina si è registrato anche un aumento dell’apporto di AA al tessuto muscolare (concentrazione di aminoacidi x flusso sanguigno)…
Le differenze nell’apporto di AA erano dovute principalmente alle differenze nelle concentrazioni di AA, che, a loro volta, erano determinate dalla modalità di infusione dell’Insulina (sistemica o locale) e/o dalla concomitante infusione di AA esogeni. Questo perché l’infusione sistemica di Insulina diminuisce le concentrazioni di AA nel sangue, a meno che gli AA non vengano sostituiti dall’infusione esogena.
Al contrario, l’infusione locale di Insulina in una gamba o in un avambraccio consente di esporre il tessuto muscolare a livelli di Insulina relativamente elevati, evitando al contempo una riduzione importante della concentrazione di AA nel sangue. [Fujita, S. et al. (2006)].
l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1, riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1. L’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1. Il discorso precedentemente fatto sulle IGFB deve essere compreso correttamente alla luce di modifiche sostanziali in concentrazione (di IGF-1) e sua frazione legata (minoritaria ma presente).
Quindi, l’Insulina può aumentare la biodisponibilità di IGF-I inibendo la produzione di IGFBP-1 e aumentando l’attività proteolitica di IGFBP-3, che riduce l’affinità di IGF-I per IGFBP-3. [Hadley JS et al. 2002]. Esemplificato in formula:
IGF-I (libero) ↑AA e assorbimento del glucosio e ripartizione delle proteine.[Hadley JS et al. 2002]
L’Insulina dirige la MPS attraverso l’attivazione diretta della via PI3K/Akt/mTORC1 e grazie all’aumento del flusso sanguigno muscolare tramite proprietà vasoattive.[Trommelen et al. 2015]
Variabile della disponibilità intracellulare di AA:
Come già alcuni di voi sapranno, la prima fase della sintesi proteica avviene nel nucleo delle cellule e comporta la trascrizione di un gene specifico in molecole di mRNA. Gli mRNA sono trasportati nel citosol, dove si associano ai ribosomi per la traduzione della sequenza di basi in una sequenza di AA. Il processo di traduzione può essere suddiviso in tre fasi: iniziazione, allungamento e terminazione della catena peptidica. Molti ribosomi possono associarsi a molecole di mRNA, formando poliribosomi. Un poliribosoma può sintetizzare diverse catene peptidiche da un singolo mRNA. Gli AA liberi nel citoplasma non vengono utilizzati direttamente per la sintesi proteica. Il processo di traduzione prevede il legame degli AA a specifiche molecole di RNA di trasferimento (tRNA), formando gli aminoacil-tRNA. Gli aminoacil-tRNA costituiscono pool molto piccoli che si trasformano rapidamente. [Biolo, G., & Wolfe, R. R. (1993)]
È stato dimostrato che l’insulina stimola sia i processi di trascrizione che di traduzione di proteine specifiche (Kimball e Jefferson, 1988). È stato dimostrato che l’espressione genica (riflessa dai livelli di mRNA) di molte proteine è stimolata dall’insulina, tra cui l’albumina nel fegato e l’MHC α nel muscolo scheletrico (Dillman, 1988). Il contenuto di mRNA riflette il potenziale ultimo di sintesi proteica nella cellula (tessuto), ma potrebbe non essere direttamente correlato al tasso di traduzione e quindi di sintesi delle proteine. L’insulina migliora anche il processo di traduzione nel muscolo scheletrico, stimolando l’iniziazione della catena peptidica (Jefferson et al, 1974; Fulks e Goldberg, 1975). Pertanto, da una base molecolare, ci si aspetterebbe che in vivo l’insulina aumenti in generale la capacità di sintesi proteica (contenuto di mRNA) e stimoli la traduzione e la produzione di proteine specifiche, come quelle muscolari. [Biolo, G., & Wolfe, R. R. (1993)]
Esistono due meccanismi attraverso i quali l’aumento dell’AA intracellulare può stimolare la sintesi proteica muscolare. In primo luogo, un effetto di massa dovuto agli AA esogeni forniti; il tRNA necessario per la traduzione degli AA in proteine è disponibile nel muscolo in quantità superiori al pool di AA disponibile. Di conseguenza, l’aumento della disponibilità di AA spingerà la MPS semplicemente caricando più tRNA. [Tipton, K. D., & Wolfe, R. R. (2001).].
È anche possibile che alcuni AA, singoli o gruppi di AA (ad esempio, i BCAA), segnalino l’avvio del processo di traduzione (ribosomiale). Rennie et al. hanno dimostrato che i singoli AA essenziali somministrati in dosi massicce (cioè riempiendo lo spazio AV e lo spazio intracellulare) stimolano la MPS, mentre gli AA non essenziali non hanno questo effetto. Inoltre, abbiamo recentemente dimostrato che non è necessario includere gli AA non essenziali in una soluzione che simula l’anabolismo muscolare in volontari a riposo . [Tipton, K. D., & Wolfe, R. R. (2001).].
Questi studi sono coerenti con l’ipotesi che uno o più degli AA essenziali possano agire come segnale per l’avvio delle MPS. Tuttavia, la stimolazione della sintesi proteica attraverso l’azione di massa non può essere esclusa solo sulla base di queste evidenze. È possibile che siano presenti sufficienti AA non essenziali e che l’aggiunta di grandi quantità di AA essenziali fornisca una carica di tRNA sufficiente a guidare la sintesi proteica. [Tipton, K. D., & Wolfe, R. R. (2001)].
Il punto di vista di Wolfe: L’insulina sembra aumentare l’efficienza della reincorporazione degli AA derivanti dalla disgregazione delle proteine (intracellulare) nelle proteine di nuova sintesi. [Biolo et al. (1995)] [Wolfe, R. R. (2000)].
Disponibilità di AA come segnale per attivare la traduzione
Sistema ubiquitina-proteosoma (sistema MPB predominante nel muscolo a riposo)
I fattori di iniziazione eucariotici (eIF), in particolare eIF4E, sono componenti importanti del controllo dell’iniziazione della traduzione. Quando eIF4E forma un complesso con eIF4G, viene promossa l’iniziazione della traduzione. Il legame di eIF4E con eIF4G è normalmente impedito perché eIF4E è legato a una proteina vincolante, eIF4E-BP1, la cui fosforilazione libera eIF4E. L’apporto di AA favorisce la sua fosforilazione, permettendo così alla sintesi proteica di procedere . In altre parole, la formazione del complesso attivo eIF4E-eIF4G aumenta in risposta alla somministrazione di proteine. [Tipton, K. D., & Wolfe, R. R. (2001)].
La disponibilità di AA come chalonico per la proteolisi muscolare
Anche il pool intracellulare di AA sembra seguire un effetto di massa per evitare la disgregazione delle proteine muscolari.
Un aumento degli AA intracellulari inibisce la MPB solo in determinate condizioni. Ad esempio, l’iperamminoacidemia a riposo ha un effetto minimo o nullo sulla MPB. Tuttavia, quando gli AA vengono somministrati dopo la RT, quando la MPB sarebbe normalmente elevata, non si verifica alcun aumento della MPB. Questa inibizione condizionale della MPB è coerente con l’idea che questa relazione tra disponibilità di AA e riduzione della MPB si verifichi principalmente quando il pool intracellulare di AA disponibili si esaurisce. Ad esempio, se la risposta iniziale all’esercizio fisico è un aumento della MPS, il pool di AA si riduce, portando alla MPB per mantenere il pool intracellulare di AA. [Tipton, K. D., & Wolfe, R. R. (2001)].
In alternativa, poiché esistono diverse vie di MPB, è possibile che in seguito all’esercizio fisico la via lisosomiale diventi predominante nel mediare l’accelerazione della MPB, e che questa via sia reattiva alla disponibilità di AA [e all’insulina esogena!] [Tipton, K. D., & Wolfe, R. R. (2001)].
…L’unico effetto probabile dei carboidrati e dei grassi sul bilancio proteico netto è l’influenza ormonale come substrati [ad esempio, l’influenza dei CHO sulla secrezione di Insulina]. [Tipton, K. D., & Wolfe, R. R. (2001)].
Trasporto transmembrana AA
Alanina +48%, Lisina +75%, Leucina +22%. [Biolo et al. (1995)].
L’alanina è uno dei principali substrati del sistema A, un sistema sodio-dipendente che mantiene ripidi gradienti transmembrana delle concentrazioni di AA ed è regolato dall’insulina [Biolo et al. (1995)] . Inoltre, la sintesi di alanina nella cellula muscolare aumenta a causa dell’incremento insulino-mediato dell’assorbimento di glucosio e della produzione intracellulare di piruvato nel muscolo. [Biolo et al. (1995)].
La lisina è un AA cationico che viene trasportato dal sistema sodio-indipendente y⁺ che è fortemente influenzato dal potenziale elettrochimico della membrana cellulare. Poiché l’insulina induce un’iperpolarizzazione nelle cellule muscolari scheletriche attivando direttamente la pompa Na⁺-K⁺-ATPae, l’accelerazione del trasporto della lisina può essere secondaria all’effetto primario dell’insulina sul gradiente elettrochimico di transmembrana. [Biolo et al. (1995)].
Gli AA a catena ramificata (leucina, isoleucina e valina) e aromatici (fenilalanina e tirosina) vengono trasportati preferibilmente attraverso il sistema L. Questo sistema, indipendente dal sodio, non è in grado di generare elevati gradienti transmembrana per i suoi substrati. È stato dimostrato che le caratteristiche cinetiche del sistema L non sono influenzate dall’insulina. [Biolo et al. (1995)].
L’azione dell’insulina sulla leucina è particolarmente modulata dall’aumento del flusso sanguigno muscolare! La leucina è soggetta a ossidazione e transaminazione che devono essere sottratte dal tasso di scomparsa nel muscolo per ottenere i tassi di sintesi proteica con la tecnica del bilancio arteriovenoso. [Rooyackers et al. (1997).].
In presenza di un adeguato apporto di AA, il sistema degradativo più importante nel muscolo è il sistema dell’ubiquitina indipendente dall’ATP. [Biolo et al. (1995)]. Questo sistema non è sensibile all’insulina [Biolo et al. (1995)]. L’insulina sembra avere un ruolo solo nella regolazione dell’attività dei lisosomi [Biolo et al. (1995)]. I lisosomi (organelli) non sono coinvolti nella degradazione delle proteine miofibrillari in condizioni normali, ma solo in presenza di bassi livelli di insulina o di una ridotta disponibilità di AA. [Biolo et al. (1995)].
La riduzione della degradazione proteica del muscolo scheletrico indotta dall’insulina si verifica in proteine non miofibrillari (soprattutto epatiche). [Rooyackers et al. (1997)].
E’ utile inoltre ricordare che il GH aumenta sia l’assorbimento degli AA dal lume intestinale che a livello cellulare nel muscolo-scheletrico, azione, quest’ultima, che va a sommarsi a quella esercitata da IGF-1 e Insulina.
Conclusioni:
Qual conclusione allora?
Oggettivamente, parlare di protocolli di Insulina/hGH ha senso quando l’atleta in questione è un avanzato, con una carriera “Natty” alle spalle ad OC e una esperienza ben controllata e graduale con i PEDs. Ma andando oltre a ciò che dovrebbe essere scontato, da quanto riportato in questo articolo, si può ben capire che essa rappresenti una pratica con un margine di guadagno in termini ipertrofici decisamente significativo.
I punti chiave degli effetti consequenziali del protocollo Insulina/hGH sono:
Aumento della sensibilità epatica al GH per via dell’attività insulinica di picco;
risposta massiva consequenziale sulla sintesi e rilascio sistemico di IGF-I;
riduzione delle proteine di trasporto IGFBP-1 e 3 insulino-correlato;
aumento della frazione libera e attiva di IGF-I.
I limiti della presente disamina non sono pochi basandosi in gran parte su dati teorici ed estrapolazioni aneddotiche tratte da piccoli casi studio. Ma ciò nonostante è sufficiente a far comprendere, almeno a livello base, come le modifiche dell’omeostasi attraverso l’uso di farmaci possa dare risultanti anche di molto distanti da quelle ottenute in contesti fisiologici.
Inoltre, se volessimo trovare un altro limite al presente lavoro, non sono stati trattati gli effetti sommativi dati dalla co-assunzione di AAS. Ciò è ovviamente legato al fatto di non volere disperdere l’attenzione del lettore dal concetto fondamentale trattato: la validità dei protocolli Insulina/hGH.
Sicuramente, i vantaggi del protocollo Insulina/hGH vanno ben oltre il banale e poco sensato assunto secondo il quale l’Insulina esogena vada semplicemente a compensare in un certo senso la ridotta sensibilità all’Insulina GH correlata.
Nel 1957 sono stati pubblicati i primi risultati del Framingham Heart Study [1]. Si trattava (o dovrei dire si tratta, visto che è ancora in corso) di uno studio epidemiologico che cercava di individuare i fattori di rischio per le malattie cardiovascolari. Lo studio prende il nome dalla città di Framingham, Massachusetts, negli Stati Uniti. Sono stati reclutati 5.209 residenti della città di età compresa tra i 30 e i 62 anni. Diversi dati di questo gruppo di persone (coorte) sono stati raccolti nel tempo per scoprire questi fattori di rischio. Nella loro importante pubblicazione, hanno identificato tre fattori di rischio per le malattie cardiovascolari: ipertensione, obesità e ipercolesterolemia (alti livelli di colesterolo).
Ai fini di questo articolo, mi concentrerò sul Colesterolo. Prima di parlare dell’effetto degli Steroidi Anabolizzanti Androgeni sul colesterolo HDL, fornirò alcune informazioni di base.
Relazione tra malattie cardiovascolari e colesterolo LDL e HDL:
Dopo i risultati iniziali del Framingham Heart Study, il ruolo del colesterolo nello sviluppo del rischio di malattie cardiovascolari è stato ulteriormente chiarito. Un primo passo avanti in quest’area di ricerca è stata la suddivisione del colesterolo in colesterolo a bassa densità (LDL) e colesterolo ad alta densità (HDL) e il loro rispettivo contributo al rischio di malattie cardiovascolari. Queste due frazioni di colesterolo sono note al grande pubblico rispettivamente come colesterolo “cattivo” e “buono”.
Il colesterolo LDL elevato è stato associato a un aumento del rischio di malattie cardiovascolari. Dopo decenni di ricerche, una pletora di prove ha stabilito con certezza che questa associazione è causale [2]. In effetti, la terapia per abbassare le LDL, ad esempio con l’uso di statine, è una pietra miliare del trattamento delle dislipidemie. L’associazione tra colesterolo HDL e rischio di malattie cardiovascolari è opposta a quella del colesterolo LDL: un colesterolo HDL elevato è risultato associato a una diminuzione del rischio di malattie cardiovascolari. Gli studi epidemiologici rilevano una riduzione del rischio cardiovascolare di circa il 2-3% per ogni aumento di 1 mg/dL di colesterolo HDL [3].
A differenza del colesterolo LDL, tuttavia, non sembra esistere un legame causale diretto tra i livelli di colesterolo HDL e il rischio di malattie cardiovascolari [4]. Studi genetici sull’uomo, in cui sono state analizzate alcune mutazioni genetiche che portano a livelli più o meno elevati di colesterolo HDL, non hanno dimostrato chiaramente un’associazione con il rischio di malattie cardiovascolari. Questo sarebbe stato prevedibile se ci fosse stato un legame causale diretto. Lo scollamento tra i livelli di colesterolo HDL e il rischio di malattie cardiovascolari è diventato forse più dolorosamente evidente negli studi clinici sui farmaci. Sono stati sviluppati (o esistevano già) alcuni farmaci che aumentano i livelli di colesterolo HDL in modo significativo, ma non riescono a ridurre la mortalità o l’incidenza di eventi cardiovascolari, come ictus o infarto del miocardio [5]. Ciò include anche l’uso di integratori da banco, come la Niacina [5, 6].
Nota: quando si parla di “colesterolo LDL”, “colesterolo HDL” o “colesterolo VLDL” ci si riferisce alle lipoproteine trasportatrici. La sigla VLDL sta per “very low density lipoproteins”, LDL per “low density lipoproteins” e HDL per “high density lipoproteins”. La densità a cui si fa riferimento è legata al loro contenuto lipidico. In particolare la densità è tanto più bassa quanto maggiori sono i trigliceridi racchiusi all’interno della particella. Ne deriva che le VLDL sono lipoproteine ad alto contenuto in trigliceridi, le LDL sono lipoproteine a basso contenuto in trigliceridi e le HDL sono lipoproteine estremamente povere di trigliceridi. In compenso LDL e HDL sono caratterizzate da un alto contenuto in colesterolo. Ognuna di queste lipoproteine ricopre ruoli diversi. Le VLDL hanno il compito di trasferire trigliceridi dal fegato ai tessuti; in particolare, dopo essere state sintetizzate nel fegato, vengono riversate nel circolo ematico e cedute soprattutto al tessuto muscolare e a quello adiposo. LDL ed HDL trasportano il colesterolo nel circolo sanguigno. Mentre le LDL hanno lo scopo di cederlo ai tessuti, le HDL sono deputate alla rimozione del colesterolo presente in eccesso nel plasma. Le VLDL vengono sintetizzate soprattutto nelle cellule epatiche (epatociti) e trasportano principalmente Trigliceridi di origine endogena.
Le IDL (lipoproteine a densità intermedia) sono il prodotto del catabolismo parziale delle VLDL. Sono più piccole (da 25 a 30 nm), hanno più Colesterolo e meno Trigliceridi. Hanno una densità compresa tra 1,006 e 1,019 g/ml e migrano elettroforeticamente con le ß-globuline. Per ogni molecola tipica di VLDL che viene degradata, viene prodotta una IDL.
Per la stima del VLDL-C è necessario dividere il valore dei Trigliceridi misurato per 5, nel caso in cui siano espressi in mg/dL, o 2.2, nel caso siano espressi in mmol/L. Nella maggior parte dei casi, la formula consente di effettuare una stima accurata del reale valore del VLDL-C.
AAS e riduzione del HDL:
Ora siete un po’ più informati sulla relazione tra malattie cardiovascolari e colesterolo LDL e HDL. Diversi studi interventistici hanno esaminato l’effetto dell’uso di AAS sul colesterolo. Peter Bond ha fatto un piccolo riassunto di questi studi nel suo libro “Book on Steroids” il quale riporto nella tabella sottostante. Sebbene non tutti gli studi abbiano riscontrato una diminuzione statisticamente significativa del colesterolo HDL (↔️), molti lo fanno e nel complesso mostrano inequivocabilmente una diminuzione. Ciò è particolarmente vero per gli AAS orali, che sembrano avere l’effetto maggiore sul colesterolo HDL.
In uno studio, condotto dal gruppo di Bhasin [11], sono stati somministrati dosaggi graduali di Testosterone (25, 50 125, 300 e 600mg di Testosterone Enantato alla settimana). Gli autori hanno quindi potuto valutare se esisteva una relazione dose-risposta tra il dosaggio di Testosterone e il colesterolo HDL, e così è stato. Hanno riscontrato una moderata relazione inversa (r = -0,40) tra i livelli di Testosterone e il colesterolo HDL. Quindi, almeno fino a una dose compresa tra 300 e 600mg settimanali, più alto è il dosaggio, maggiore è la diminuzione del colesterolo HDL.
In un recente studio, 100 consumatori di AAS sono stati seguiti nel tempo durante l’autosomministrazione di questa classe di farmaci. Il dosaggio medio, basato sulle informazioni riportate sull’etichetta, era di 898mg a settimana, rendendo così il loro ciclo di AAS abbastanza rappresentativo dell’uso comune da parte dei bodybuilder. Le misurazioni sono state effettuate prima, durante, 3 mesi dopo la fine del ciclo e 1 anno dopo l’inizio del ciclo. Il colesterolo HDL è diminuito di 0,4 mmol/L (da 1,2 a 0,8) durante l’uso. Si tratta di una diminuzione sostanziale. I valori erano tornati ai valori di base 3 mesi dopo la cessazione dell’uso di AAS.
Meccanismo attraverso il quale gli AAS abbassano l’HDL:
Si ritiene che gli steroidi anabolizzanti riducano il colesterolo HDL aumentando l’attività di un enzima chiamato lipasi epatica [7, 8, 9, 10]. Si tratta di un enzima prodotto principalmente dal fegato. Essendo una lipasi, catalizza le reazioni di idrolisi dei lipidi. In particolare, scinde gli acidi grassi dal triacilglicerolo (Trigliceride) e i fosfolipidi dalle particelle lipoproteiche, come il colesterolo HDL. Idrolizzando il triacilglicerolo e i fosfolipidi dal colesterolo HDL, riduce le dimensioni di queste particelle. Queste particelle più piccole vengono catabolizzate a un ritmo più elevato [12].
Thompson et al. hanno esaminato queste sottofrazioni di colesterolo HDL che differiscono per dimensioni [8]. Hanno misurato i livelli di colesterolo HDL2 e HDL3: le particelle di colesterolo HDL2 sono più grandi e di densità inferiore rispetto a quelle HDL3. Gli uomini partecipanti hanno ricevuto 200mg di Testosterone Enantato alla settimana o 6mg di Stanozololo orale (Winstrol) al giorno per 6 settimane in un design crossover. I risultati sono stati i seguenti:
*Differenza significativa (P < 0,05) rispetto al valore basale.
Come si può notare, la maggiore diminuzione relativa è stata osservata nella frazione HDL2 più grande a seguito del trattamento con Stanozololo. Al contrario, il Testosterone non ha mostrato una diminuzione statisticamente significativa nella frazione HDL2, ma ha fatto altrettanto nella frazione HDL3 più piccola. Non è del tutto chiaro cosa provochi la diminuzione di questa frazione.
Quando dei bodybuilder sono stati randomizzati a ricevere 200mg di Nandrolone Decanoato alla settimana o un placebo [13]. Non sono stati riscontrati cambiamenti statisticamente significativi nel colesterolo totale, nel colesterolo LDL e nel colesterolo HDL. Analogamente, non sono stati riscontrati cambiamenti significativi nelle sottofrazioni di colesterolo HDL2 e HDL3. In particolare, nella stessa pubblicazione, gli autori riferiscono anche di uno studio in cui hanno seguito un gruppo di atleti di forza che si autosomministravano steroidi anabolizzanti. Sono stati utilizzati diversi composti in vari dosaggi, ma vale la pena sottolineare che la maggior parte di essi comprendeva anche uno steroide anabolizzante orale (soprattutto Stanozololo). In questo caso, il colesterolo HDL è sceso in picchiata: da 1,08 mmol/L a 0,43 mmol/L dopo 8 settimane. La sottofrazione di colesterolo HDL2 è scesa da 0,21 a 0,05 e la sottofrazione di colesterolo HDL3 è scesa da 0,87 a 0,40 mmol/L.
Effetto degli AAS sulla funzione del colesterolo HDL:
Dato il legame tra l’effetto di un farmaco sui livelli di colesterolo HDL e il rischio di malattie cardiovascolari, la ricerca ha iniziato a concentrarsi sulla funzione del colesterolo HDL. Il colesterolo HDL è il protagonista di un processo chiamato trasporto inverso del colesterolo. Nell’aterosclerosi, il colesterolo si accumula nelle cellule del sistema immunitario (macrofagi) e nelle cellule muscolari lisce che circondano i vasi sanguigni [14]. Queste cellule, a loro volta, diventano le cosiddette cellule schiumose, che segnano il punto di partenza dell’aterosclerosi. Le particelle di colesterolo HDL sono in grado di raccogliere il colesterolo da queste cellule – efflusso di colesterolo. L’efflusso del colesterolo dalle cellule schiumose nelle particelle di colesterolo HDL è uno dei modi in cui si ritiene che il colesterolo HDL eserciti i suoi effetti protettivi sulle arterie. Il colesterolo HDL raccolto può poi essere riportato al fegato, che lo incorpora nella bile e può quindi essere secreto nelle feci. Allo stesso modo, le particelle di colesterolo HDL possono trasferire parte del loro contenuto alle particelle LDL, che possono finire nuovamente nelle cellule schiumose o essere assorbite dal fegato.
Esistono metodi per misurare la capacità di efflusso del colesterolo HDL e l’idea attuale è che la sua modulazione possa influire sul rischio di malattie cardiovascolari, contrariamente ai livelli di colesterolo HDL in sé. Esistono diversi modi in cui il colesterolo HDL può assorbire il colesterolo dalle cellule schiumose. Uno di questi coinvolge un trasportatore chiamato ATP-binding casette transporter A1 (ABCA1), che si ritiene sia il più importante [15, 16]. Contribuiscono anche altri trasportatori, come ABCG1 e il recettore scavenger B1, oltre alla diffusione semplice.
Diamo uno sguardo agli studi che hanno valutato l’impatto dell’uso di steroidi anabolizzanti sulla capacità di efflusso del colesterolo HDL. In uno studio (non controllato), uomini anziani ipogonadici sono stati randomizzati alla TRT con o senza Dutasteride (un inibitore della 5a-reduttasi) [17]. Dopo 3 mesi, la TRT era riuscita a riportare i livelli di Testosterone di questi uomini all’interno del range di normalità. Il colesterolo HDL e la capacità di efflusso del colesterolo HDL sono rimasti inalterati.
Un altro studio, randomizzato e controllato, ha applicato un approccio leggermente diverso [18]. Uomini sani, di età compresa tra i 19 e i 55 anni, sono stati castrati medicalmente per sopprimere completamente la loro produzione endogena. In seguito, hanno ricevuto un placebo, una TRT a basso dosaggio, una TRT sostitutiva completa o una TRT sostitutiva completa con Letrozolo, un inibitore dell’Aromatasi che inibisce la conversione del Testosterone in Estradiolo. Il colesterolo HDL è aumentato leggermente nel gruppo placebo e in quello a basso dosaggio, mentre è rimasto inalterato nei due gruppi che hanno ricevuto una dose sostitutiva completa. Inoltre, mentre è stata riscontrata una piccola diminuzione della capacità di efflusso di ABCA1 nel gruppo che ha ricevuto anche il Letrozolo, non sono stati osservati cambiamenti negli altri tre gruppi. A causa delle dimensioni ridotte dei gruppi, è possibile che un piccolo effetto non sia stato notato.
E i dosaggi elevati? Sfortunatamente, esiste un solo studio che ha esaminato questo aspetto, ed era di natura trasversale (si tratta di misurazioni effettuate in un solo momento, il che rende impossibile/difficile trarre conclusioni)[19]. I ricercatori hanno confrontato le misurazioni di un gruppo di utilizzatori di AAS con quelle di non utilizzatori e controlli sedentari, che avevano un’età corrispondente. I consumatori di AAS erano forti utilizzatori, avendo fatto uso di AAS in media per circa 8 anni con un dosaggio medio di (apparentemente) 2,5g settimanali. La capacità delle HDL di effluire il colesterolo dai macrofagi è risultata inferiore del 13% nei consumatori di AAS rispetto ai non consumatori con allenamento della forza. Anche in questo caso, a causa della natura trasversale dello studio, è difficile dire se questo sia causale.
Conclusioni:
Gli AAS riducono il colesterolo HDL, in modo dose-dipendente, e questo sembra verificarsi in modo particolarmente marcato con gli AAS orali17α-alchilati. Non è certo come questo si traduca in un rischio di malattia cardiovascolare, in quanto esiste una discrepanza tra la capacità di un farmaco di alterare il colesterolo HDL e il suo effetto su di esso. La correlazione tra i livelli di colesterolo HDL misurati e il rischio di malattie cardiovascolari non è causale. I ricercatori ritengono che la capacità di efflusso del colesterolo HDL possa avere una migliore capacità predittiva, oltre a essere causalmente correlata. Pertanto, i farmaci che influiscono sulla capacità di efflusso potrebbero influenzare il rischio di malattie cardiovascolari. L’effetto degli AAS su questo aspetto non è ancora così chiaro a causa della scarsità di dati, soprattutto per quanto riguarda i dosaggi sovrafisiologici. Alcuni dati suggeriscono che potrebbero avere un impatto negativo sulla capacità di efflusso del colesterolo. Uno studio di coorte longitudinale probabilmente risponderà a questa domanda con maggiore certezza in futuro. Se la capacità di efflusso del colesterolo HDL diminuisce effettivamente in seguito all’uso di AAS, questa diminuzione indotta dagli AAS potrebbe essere dannosa per la salute cardiovascolare.
Certo, vi sono farmaci, non che integratori erboristici da banco, con azione di “tampone” della dislipidemia ematica. Ma ciò non elimina il problema lo rallenta nella sua potenziale comparsa soprattutto agendo sui rapporti tra i marcatori del profilo lipidico ematico. Ciò significa che potenzialmente, e il condizionale è d’obbligo vista la sensibile differenza soggettiva riscontrabile, l’uso di Monacolina-K, Niacina e EPA, nei corretti dosaggi, potrà causare una riduzione del HDL leggermente/moderatamente inferiore rispetto all’utilizzatore meno accorto, con conseguente alterazione delle ratio HDL:LDL, HDL:Trigliceridi e HDL:Colesterolo totale “rallentata” e meno marcata. Di per se questa pratica supplementativa potrebbe portare anche ad una riduzione anche della capacità di efflusso del colesterolo HDL, ma non vi sono, ad oggi, conferme inoppugnabili che ciò avvenga.
Ah, quasi dimenticavo di ricordare ai meno informati che anche i SARM non steroidei (vedi Ostarina, LGD-4033, ecc…) alterano il profilo lipidico ematico a diverso grado. Anche i SERM (Tamoxifene, Clomifene, Raloxifene ecc…) e AI (Letrozolo, Anastrozolo, Exemestane ecc…) hanno un potenziale di alterazione della lipidemia ematica.
Gabriel Bellizzi
Riferimenti:
T. R. Dawber, F. E. Moore, and G. V. Mann. Measuring the risk of coronary heart disease in adult population groups: Ii. coronary heart disease in the framingham study. American Journal of Public Health and the Nations Health, 47(4 Pt 2):4, 1957
Ference, Brian A., et al. “Low-density lipoproteins cause atherosclerotic cardiovascular disease. 1. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel.” European heart journal 38.32 (2017): 2459-2472.
Gordon, David J., et al. “High-density lipoprotein cholesterol and cardiovascular disease. Four prospective American studies.” Circulation 79.1 (1989): 8-15.
Rader, Daniel J., and G. Kees Hovingh. “HDL and cardiovascular disease.” The Lancet 384.9943 (2014): 618-625.
Keene, Daniel, et al. “Effect on cardiovascular risk of high density lipoprotein targeted drug treatments niacin, fibrates, and CETP inhibitors: meta-analysis of randomised controlled trials including 117 411 patients.” Bmj 349 (2014).
Schandelmaier, Stefan, et al. “Niacin for primary and secondary prevention of cardiovascular events.” Cochrane Database of Systematic Reviews 6 (2017).
Friedl, Karl E., et al. “High-density lipoprotein cholesterol is not decreased if an aromatizable androgen is administered.” Metabolism 39.1 (1990): 69-74.
Thompson, Paul D., et al. “Contrasting effects of testosterone and stanozolol on serum lipoprotein levels.” Jama 261.8 (1989): 1165-1168.
Zmuda, Joseph M., et al. “The effect of testosterone aromatization on high-density lipoprotein cholesterol level and postheparin lipolytic activity.” Metabolism 42.4 (1993): 446-450.
Herbst, Karen L., et al. “Testosterone administration to men increases hepatic lipase activity and decreases HDL and LDL size in 3 wk.” American Journal of Physiology-Endocrinology and Metabolism 284.6 (2003): E1112-E1118.
Singh, Atam B., et al. “The effects of varying doses of T on insulin sensitivity, plasma lipids, apolipoproteins, and C-reactive protein in healthy young men.” The Journal of Clinical Endocrinology & Metabolism 87.1 (2002): 136-143.
Jin, Weijun, Dawn Marchadier, and Daniel J. Rader. “Lipases and HDL metabolism.” Trends in Endocrinology & Metabolism 13.4 (2002): 174-178.
Hartgens, F., et al. “Effects of androgenic-anabolic steroids on apolipoproteins and lipoprotein (a).” British journal of sports medicine 38.3 (2004): 253-259.
Ouimet, Mireille, Tessa J. Barrett, and Edward A. Fisher. “HDL and reverse cholesterol transport: Basic mechanisms and their roles in vascular health and disease.” Circulation research 124.10 (2019): 1505-1518.
Du, Xian-Ming, et al. “HDL particle size is a critical determinant of ABCA1-mediated macrophage cellular cholesterol export.” Circulation research 116.7 (2015): 1133-1142.
Adorni, Maria Pia, et al. “The roles of different pathways in the release of cholesterol from macrophages.” Journal of lipid research 48.11 (2007): 2453-2462.
Rubinow, Katya B., et al. “Testosterone replacement in hypogonadal men alters the HDL proteome but not HDL cholesterol efflux capacity.” Journal of lipid research 53.7 (2012): 1376-1383.
Rubinow, Katya B., et al. “Sex steroids mediate discrete effects on HDL cholesterol efflux capacity and particle concentration in healthy men.” Journal of clinical lipidology 12.4 (2018): 1072-1082.
de Souza, Francis Ribeiro, et al. “Diminished cholesterol efflux mediated by HDL and coronary artery disease in young male anabolic androgenic steroid users.” Atherosclerosis 283 (2019): 100-105.
Il BodyBuilding si differenzia dagli sport di prestazione perché il giorno della gara gli atleti vengono giudicati in base all’aspetto piuttosto che alle capacità atletiche. I bodybuilder posano sul palco dove vengono giudicati per la muscolatura, la definizione e la simmetria. Nel corso di una stagione, i bodybuilder attraversano tre fasi diverse: la fase di crescita muscolare (Off-Season), la dieta per la competizione (preparazione alla gara) e la gara stessa. La maggior parte della letteratura riguarda la fase di dieta pre-gara e la peak week.[1]
Tuttavia, la letteratura scientifica sulle raccomandazioni alimentari per i bodybuilder durante la Off-Season è carente. Si tratta di una lacuna importante, poiché la maggior parte della carriera di un bodybuilder si svolge in questa fase, in cui l’obiettivo è aumentare la massa muscolare riducendo al minimo l’aumento eccessivo della massa grassa. I bodybuilder sono noti per avere atteggiamenti rigidi nei confronti della selezione degli alimenti, della frequenza dei pasti, dei tempi di alimentazione e dell’integrazione [2]. Storicamente, le informazioni sull’alimentazione e l’integrazione sono state trasmesse dalle riviste di bodybuilding e dai concorrenti di successo, ma recentemente sono emerse più informazioni attraverso Internet e i forum [3,4]. Di conseguenza, molte delle strategie alimentari utilizzate dai bodybuilder non hanno un solido supporto scientifico e la letteratura scientifica dimostra che alcune di queste strategie, tra cui l’uso massiccio di farmaci, ma anche di integratori più in generale, possono essere ovviamente dannosi per la salute [5,6,7].
Poiché i bodybuilder trascorrono la maggior parte del loro tempo in Off-Season, è evidente la necessità di raccomandazioni nutrizionali e di supplementazione, sia OTC che PEDs, il più possibile “sicure” e basate sull’evidenza per questa popolazione. È stato inoltre dimostrato che alcuni bodybuilder, e non soltanto i concorrenti di alto livello nel bodybuilding “Natural”, potrebbero essere interessati a informazioni basate sull’evidenza [8]. Con il supporto della review realizzata e pubblicata da Juma Iraki et al. che tratta del Off-Season a livello alimentare e integrativo, lo scopo di questo articolo sarà quello di riportare quanto evidenziato dalla letteratura scientifica sugli argomenti relativi all’alimentazione e all’integrazione alimentare e supplementazione PEDs rilevanti per i bodybuilder nella Off-Season e di fornire raccomandazioni pratiche sull’assunzione di energia, macronutrienti, frequenza dei pasti, tempistica dei nutrienti, integratori alimentari e PEDs .
Transizione dalla dieta pre-gara/peak week alla dieta in Off-Season – Reverse Diet Vs. Recovery Diet:
Il primo step che il bodybuilder si trova davanti è la gestione del passaggio da una dieta ipocalorica ad una ipercalorica. Ed è in questo frangente che emergono due strategie simili all’apparenza ma in realtà diverse: la “Recovery Diet” e la “Reverse Diet”.
Ora, molto semplicemente, la “Recovery Diet” consiste in un graduale aumento calorico ma di consistenza tale che l’atleta esca dalla condizione di ipocalorica nel giro di due settimane circa. Con la “Reverse Diet”, invece, abbiamo sempre un graduale aumento calorico ma caratterizzato da una ridotta consistenza dello stesso (si parla di circa 100Kcal/die a settimana). In questo caso specifico, il bodybuilder rimarrebbe in ipocalorica per diverse settimane con possibile emersione di problemi psicofisici legati al protrarsi dello stato stressorio.
Quindi, con il termine “Recovery Diet” ci riferiamo ad uno schema alimentare avente l’obiettivo generale di RECUPERARE da un periodo di dieta cronica sperimentato durante la preparazione alla gara. La “Recovery Diet” incoraggia i bodybuilder a guadagnare il 5-10% del loro peso di gara nelle prime 4-8 settimane successive all’evento. Questo con l’intento di accelerare l’aumento di grasso corporeo e far rientrare il soggetto in un range di grasso corporeo “sano”, fisiologico, il prima possibile. In seguito, si consiglia agli atleti di rallentare il ritmo di aumento del peso e di mantenere un surplus controllato, con un aumento medio dello 0,5-1% del peso corporeo al mese passando pienamente nella Off-Season. Questo fino a quando non raggiungono un punto in cui un ulteriore aumento di peso è considerato improduttivo. Con il termine “Reverse Diet” ci si riferisce ad una strategia la quale può ancora essere attuata con discreti vantaggi per aiutare un agonista a recuperare dopo il contest. Tuttavia, se rispettata e seguita correttamente, piccoli aumenti di cibo di ~100 Kcal/die a settimana potrebbero comunque protrarre il deficit calorico del soggetto, prolungando così il periodo di dieta ipocalorica. Sebbene questa possa essere una strategia utile in alcune circostanze, ad esempio durante l’avvicinamento alla competizione, le modalità di applicazione non permettono un recupero di una bf salubre in tempi ottimali. È risaputo che un bodybuilder in condizioni di picco non è necessariamente al massimo della salute, e questo è in gran parte correlato al livello di grasso corporeo. Accettare un certo aumento di grasso avrà effetti positivi su tutti gli aspetti della Off-Season come le prestazioni in allenamento, i marcatori ormonali, la disponibilità di energia, la qualità del sonno e, inoltre, sarà vantaggioso sulla longevità complessiva dello sport praticato.
In definitiva, se si parte da body fat estremamente basse, tipiche da gara, allora la “Recovery Diet” è la scelta migliore per shiftare dal regime ipocalorico che ha caratterizzato il periodo di preparazione alla gara a quello ipercalorico del Off-Season. Discorso diverso se ci troviamo di fronte ad un soggetto amatoriale, con una body fat del 8-10% arrivato al termine del percorso di “Cut”. In questo caso la “Reverse Diet” è la scelta più funzionale permettendo un controllo migliore degli incrementi calorici evitando che la massa grassa sfori eccessivamente e che il lavoro precedentemente svolto in “Cut” venga facilmente e totalmente compromesso. Anche “ibridazioni” con aumenti settimanali di 45-50g di CHO die possono essere applicati con buoni risultati.
Energia:
Durante la Off-Season, l’obiettivo principale di un bodybuilder è quello di aumentare la massa muscolare riducendo al minimo l’aumento della massa grassa attraverso l’uso di allenamenti contro-resistenza e il mantenimento di un bilancio energetico positivo. Per valutare con precisione il fabbisogno energetico dei bodybuilder durante la bassa stagione, è necessario considerare il volume, la frequenza e l’intensità dell’allenamento. Durante la fase off-season, è stato riportato che i bodybuilder si allenano alla resistenza 5-6 volte a settimana, esercitando ogni gruppo muscolare 1-2 volte a settimana [9]. È stato inoltre riferito che seguono una routine di allenamento ad alto volume con 4-5 esercizi per gruppo muscolare, eseguendo 3-6 serie per esercizio, 7-12 ripetizioni massime (RM) per ogni serie con 1-2 minuti di riposo tra le serie. La durata della sessione di allenamento è stata indicata in ~40-90 minuti. Tuttavia, i piani di allenamento possono variare notevolmente da atleta ad atleta. È necessario valutare anche l’apporto calorico medio dei bodybuilder. Nella fase off-season, l’apporto energetico è di solito sostanzialmente più elevato rispetto alla fase di dieta: tra i bodybuilder maschi è stato riportato un apporto medio di ~3800 kcal/giorno durante la fase off-season e di ~2400 kcal/giorno durante la fase di dieta [2].
Bilancio energetico positivo:
È stato dimostrato che un bilancio energetico positivo ha un importante effetto anabolico, anche in assenza di allenamento contro-resistenza [10]. Tuttavia, la combinazione di un bilancio energetico positivo con l’allenamento contro-resistenza rappresenta il metodo più efficace per garantire che gli effetti anabolici siano diretti all’aumento della massa muscolo-scheletrica [11,12]. L’entità del surplus energetico ideale per guadagnare massa muscolare limitando l’accumulo di tessuto adiposo può variare in base allo stato di allenamento. Nei soggetti non allenati, è stato dimostrato che un surplus energetico sostanziale di circa 2.000 kcal, combinato con l’allenamento contro-resistenza, fornisce un robusto aumento di peso, in cui il contributo della massa magra (LBM) può raggiungere il 100% [12]. Tuttavia, nei soggetti allenati, un surplus energetico sostanziale potrebbe non essere necessario o vantaggioso. Uno studio condotto su atleti d’élite ha esaminato l’effetto delle indicazioni dietetiche sui cambiamenti della composizione corporea tra gli atleti d’élite quando l’allenamento contro-resistenza è stato combinato con diverse entità di surplus energetico. Un gruppo con un peso corporeo medio di 75kg ha consumato energia ad libitum (2964 kcal) per raggiungere un surplus molto ridotto, mentre un secondo gruppo con un peso corporeo medio di 71kg ha ricevuto una consulenza dietetica e ha consumato ~600 kcal in più rispetto al gruppo ad libitum [13].
Entrambi i gruppi hanno seguito lo stesso programma di allenamento contro-resistenza di 4 giorni alla settimana per un periodo di 8-12 settimane. I ricercatori hanno ipotizzato che il gruppo ipercalorico avrebbe avuto un aumento maggiore del peso corporeo e della LBM. Sebbene il gruppo ipercalorico abbia ottenuto un aumento maggiore della LBM rispetto a quelli che mangiavano ad libitum, questo non ha raggiunto la significatività statistica (1,7kg contro 1,2kg, rispettivamente). Inoltre, rispetto al gruppo che mangiava a sazietà, hanno registrato un aumento significativamente maggiore della massa grassa (1,1kg contro 0,2kg, rispettivamente). I ricercatori hanno concluso che un surplus di 200-300 kcal al giorno negli atleti altamente allenati potrebbe essere più appropriato di 500 kcal per minimizzare il rischio di inutili aumenti di grasso corporeo. I soggetti non allenati, più lontani dal loro tetto genetico di massa muscolare, possono essere in grado di aumentare i muscoli a un ritmo più veloce rispetto agli individui allenati.
Il tasso di crescita muscolare può rallentare con l’avanzare dell’età [14]. Pertanto, un maggiore surplus energetico può essere più vantaggioso per i bodybuilder alle prime armi, mentre i bodybuilder avanzati potrebbero trarre maggiore beneficio da diete ipercaloriche conservative per limitare inutili aumenti di grasso corporeo. Studi precedenti hanno raccomandato ai bodybuilder di consumare una dieta leggermente ipercalorica, con un aumento dell’apporto energetico di circa il 15% rispetto al mantenimento nella Off-Season [15]. Tuttavia, ciò non tiene conto della storia di allenamento e del livello di esperienza del singolo bodybuilder. Poiché la capacità di aumentare la massa muscolare è limitata, un surplus aggressivo può portare a un inutile aumento del grasso corporeo, che aumenterebbe la durata o la gravità dei successivi periodi di preparazione alle gare, aumentando di conseguenza la durata o la gravità della scarsa disponibilità energetica. Pertanto, il numero di calorie che un bodybuilder consuma al di sopra del livello di mantenimento può essere stabilito in base al livello di esperienza e poi regolato in base al tasso di aumento di peso e ai cambiamenti nella composizione corporea. Dato che i bodybuilder spesso aumentano rapidamente di peso dopo una gara, potrebbe essere utile avere un obiettivo di aumento di peso per settimana e regolarsi di conseguenza [16,17].
Tuttavia, come detto precedentemente, inizialmente, dopo la gara, potrebbe essere utile un aumento di peso più rapido per aiutare a riportare il concorrente a uno stato di salute sia psicologico che fisiologico, prima che il tasso di aumento di peso venga rallentato per limitare l’accumulo eccessivo di tessuto adiposo. Nella letteratura scientifica si raccomanda di puntare a un aumento di peso di circa 0,25-0,5 kg a settimana per cercare di aumentare la LBM e ridurre al minimo l’aumento della massa grassa [14,18]. Per un bodybuilder avanzato, un potenziale aumento di 2kg di peso corporeo su base mensile potrebbe essere eccessivo e comportare un’inutile accumulazione di grasso corporeo; pertanto, questo tasso dovrebbe essere considerato con cautela. Sulla base delle prove attuali, potrebbe essere opportuno raccomandare ai bodybuilder di consumare una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) nella Off-Season e raccomandare ai bodybuilder avanzati di puntare all’estremità inferiore di questa raccomandazione, o addirittura di essere più conservativi se si verificano aumenti sostanziali della massa grassa. Dato che i bodybuilder consumano in media 45 kcal/kg durante la bassa stagione, il surplus raccomandato equivale a circa 42-48 kcal/kg [2]. Potrebbe essere utile puntare a un aumento di peso di circa 0,25-0,5% del peso corporeo a settimana, regolando al contempo l’apporto energetico in base alle variazioni della composizione corporea. Inoltre, potrebbe essere più appropriato considerare le variazioni di peso medie settimanali basate su pesate giornaliere (o più volte alla settimana) per limitare gli errori delle fluttuazioni giornaliere del peso che possono verificarsi durante la settimana. Una volta determinato il surplus calorico, il passo successivo sarà quello di distribuire le calorie tra proteine, grassi e carboidrati.
Proteine:
Il turnover proteico del muscolo scheletrico è il rapporto tra la sintesi proteica muscolare (MPS) e la degradazione proteica muscolare (MPB). L’ipertrofia del muscolo scheletrico richiede un equilibrio netto in cui la MPS supera la MPB. L’esercizio contro-resistenza fornisce lo stimolo di tensione iniziale che induce l’ipertrofia risultante dall’aumento cumulativo della MPS dopo l’esercizio cronico [19]; tuttavia, l’aumento della massa grassa (FFM) può essere limitato se l’apporto proteico giornaliero è insufficiente [20]. Oltre alla quantità totale consumata al giorno, i ricercatori hanno ipotizzato che la qualità delle proteine possa aumentare il guadagno muscolare indotto dall’allenamento contro-resistenza [21]. Pertanto, entrambi questi argomenti saranno discussi nelle sezioni seguenti.
Introito proteico giornaliero:
Mentre l’attuale RDA per le proteine negli individui sani sedentari è di 0,8 g/kg, in una meta-analisi del 2018 di Morton e colleghi [22] è stato osservato che il doppio di questa quantità massimizza l’ipertrofia indotta dall’allenamento contro-resistenza. Inoltre, gli autori hanno osservato che “potrebbe essere prudente raccomandare ~2,2g di proteine/kg/die per coloro che cercano di massimizzare i guadagni di FFM indotti dall’allenamento contro-resistenza”, poiché 2,2g/kg era l’estremità superiore del limite di confidenza [22] e le differenze individuali impongono che alcuni atleti abbiano un fabbisogno proteico più elevato di altri [23]. Inoltre, la raccomandazione “meglio prevenire che curare” è probabilmente sicura, vista l’assenza di danni apparenti in studi di 1-2 anni tra i sollevatori che consumavano apporti proteici di almeno 2,2 g/kg [24,25]. Infine, la media e il limite superiore di confidenza del 95% per il fabbisogno proteico utilizzando la tecnica di ossidazione degli aminoacidi con indicatore tra i bodybuilder maschi nei giorni di non allenamento sono stati riportati rispettivamente come 1,7 e 2,2g/kg [26], che è simile al fabbisogno tra le donne quando è normalizzato alla FFM [27].
Tuttavia, è stato riportato che i bodybuilder consumano fino a 4,3g/kg di proteine al giorno tra i soggetti di sesso maschile e 2,8g/kg tra quelli di sesso femminile, superando di gran lunga queste raccomandazioni [2]. Le linee guida precedentemente fornite per i bodybuilder nella Off-Season erano di consumare il 25-30% del loro apporto energetico dalle proteine [15]. Potrebbe essere ragionevole opporsi all’indicazione di raccomandazioni basate su percentuali dell’apporto energetico totale, poiché un individuo con un peso non particolarmente elevato ma con un alto fabbisogno energetico potrebbe finire per consumare proteine che superano di gran lunga quelle necessarie e quindi richieste. Inoltre, questo può portare a un’assunzione insufficiente di carboidrati e grassi se l’atleta mira a un apporto calorico specifico. Pertanto, potrebbe essere più appropriato raccomandare un fabbisogno proteico basato sul peso corporeo. Pertanto, i bodybuilder dovrebbero consumare un minimo di 1,6g/kg di proteine nella Off-Season, anche se un obiettivo più vicino a 2,2 g/kg potrebbe garantire una risposta ottimizzata in modo più coerente in una maggiore percentuale di atleti.
E per i “Doped”? Dovremo ormai sapere che la fisiologia di base è la medesima per ogni individuo con le consuete variabili. Detto ciò, l’uso di PEDs va si ad alterare la fisiologia ma in questo specifico ambito, ossia introito proteico per massimizzare lo stimolo ipertrofico, hanno una azione di perfezionamento dell'”economia proteica cellulare”: in parole più semplici, sembra che l’uso di AAS porti ad una migliore resa nell’utilizzo degli amminoacidi scissi e assorbiti dalle proteine alimentari. Di conseguenza, a parità di apporto proteico, la veicolazione degli amminoacidi a scopo plastico è maggiore come minore è l’attività catabolica. Ciò significa che abusare delle proteine, in special modo durante una fase ipercalorica, perchè si è sotto AAS potrebbe risultare più inutile di quanto non lo sia in contesto “Natural”.
Infine, ed è necessario sottolinearlo, tra i bodybuilder che lottano con la fame in Off-Season e che di conseguenza assumono quantità caloriche che portano a un aumento di peso più rapido e all’accumulo di grasso in eccesso, un apporto proteico più elevato può essere utile (se non controindicato per motivi clinici). In uno studio condotto da Antonio e colleghi, i partecipanti ad allenamenti contro-resistenza che consumavano più proteine (4,4g/kg al giorno) e più calorie hanno guadagnato una quantità simile di FFM, ma non hanno guadagnato ulteriore grasso corporeo rispetto al gruppo che consumava meno proteine e meno calorie [28]. Allo stesso modo, in uno studio di follow-up, un gruppo che consumava 3,4g/kg di proteine al giorno ha guadagnato una quantità simile di FFM, ma ha perso una percentuale maggiore di grasso corporeo rispetto a un gruppo a basso contenuto proteico, ancora una volta, nonostante un apporto energetico più elevato [29]. Gli autori di questi studi sulla “vita libera” hanno ipotizzato che i loro risultati fossero dovuti a un aumento della termogenesi indotta dalla dieta attraverso protocolli alimentari ad alto contenuto proteico. Tuttavia, ciò è in contrasto con uno studio di Bray e colleghi del 2012 sul reparto metabolico, più strettamente controllato, in cui il contenuto proteico della dieta influenzava la percentuale di massa corporea acquisita, mentre la massa corporea totale era dettata dal solo contenuto energetico della dieta [30].
Pertanto, mentre la termogenesi indotta dalla dieta potrebbe essere significativamente più elevata con assunzioni di proteine nell’intervallo di 3 g/kg o superiore, la perdita di grasso o la mancanza di aumento di peso osservata da Antonio e colleghi, nonostante un apporto energetico più elevato, potrebbe con più probabilità riflettere l’effetto saziante di assunzioni proteiche molto elevate che diminuiscono l’assunzione calorica effettiva, piuttosto che un aumento della sola termogenesi.
Qualità delle Proteine:
Gli aminoacidi essenziali (EAA) sono gli unici aminoacidi necessari per stimolare il processo di MPS [31]. Sebbene tutti gli aminoacidi forniscano i “mattoni” necessari per la sintesi di nuovi tessuti, l’aminoacido Leucina in particolare sembra essere particolarmente importante come “innesco metabolico” della MPS [32]. È stato suggerito che una concentrazione sufficiente di Leucina è necessaria per raggiungere una “soglia di Leucina” che è richiesta per stimolare al massimo la MPS [33]. In breve, dal punto di vista della costruzione muscolare, le fonti proteiche che innescano una consistente risposta della MPS (quantità sufficiente di Leucina) e forniscono i mattoni essenziali per la costruzione di nuovo tessuto muscolare (contengono l’intero spettro di aminoacidi essenziali in abbondanza) possono essere considerate di “qualità superiore”.
Sebbene l’effetto meccanicistico della Leucina sulle MPS esuli dallo scopo di questo articolo, si invitano i lettori a leggere una rassegna che tratta questo argomento in dettaglio [34]. In generale, su una base di grammo per grammo, le fonti proteiche di origine animale contengono in genere più Leucina ed EAA, anche se ci sono eccezioni degne di nota. Le proteine della soia, uno dei più comuni integratori proteici di origine vegetale, contengono tutti gli EAA, ma in una quantità inferiore per grammo rispetto alle proteine del latte e quindi, in uno studio, hanno prodotto un aumento minore delle MPS rispetto al siero di latte dopo un’ingestione acuta [35]. È interessante notare che in questo stesso studio la soia ha prodotto un aumento maggiore delle MPS rispetto alla caseina, anch’essa una proteina casearia di “alta qualità”, presumibilmente a causa della più lenta velocità di digestione della caseina [35]. Rammentate sempre la differenza tra risposta “acuta” e “cronica”. Per l’appunto, ciò significa che, sebbene il contenuto di Leucina e di EAA di una fonte proteica debba essere preso in considerazione, la risposta acuta alla MPS non è l’unica variabile legata all’ipertrofia a lungo termine. Infatti, una proteina di alta qualità ma “lenta” come la caseina produce inizialmente una risposta MPS di minore ampiezza. Tuttavia, la caseina (e altre proteine a lenta digestione) può produrre un’area MPS sotto la curva simile o maggiore se osservata longitudinalmente rispetto a una fonte proteica “veloce” come il siero di latte, che determina un aumento iniziale maggiore e poi una brusca riduzione [36].
Inoltre, la risposta acuta della MPS a un determinato tipo di proteina non deve essere vista in una prospettiva riduzionista. Nel mondo reale si consumano quotidianamente più porzioni di varie fonti proteiche, rendendo probabilmente superflue alcune di queste distinzioni nel profilo aminoacidico e nella cinetica di digestione. Infatti, in una meta-analisi che ha confrontato i cambiamenti longitudinali della composizione corporea con diversi tipi di integratori proteici, non sono state riscontrate differenze significative tra i partecipanti che consumavano soia rispetto al siero di latte, ad altre proteine del latte o alle proteine isolate del manzo [37].
Come dimostrato in uno studio che ha messo a confronto gruppi che consumavano proteine dopo l’allenamento (in aggiunta a una dieta già composta dal 25% di proteine), sia che venissero forniti 48g di proteine del siero del latte (contenenti 5,5g di Leucina), sia che venissero forniti 48g di proteine del riso (contenenti 3,8g di Leucina), non è stato osservato alcun impatto sui cambiamenti della composizione corporea tra i gruppi dopo otto settimane [38]. Pertanto, se consumate in quantità sufficienti (soprattutto se si considera l’apporto proteico totale giornaliero), la qualità delle proteine di un singolo pasto è meno preoccupante. Tuttavia, se si volesse consumare una dieta dominata da fonti proteiche di origine vegetale, esistono alternative alla soia e al riso. Ad esempio, le proteine isolate del pisello sono ricche di EAA e di Leucina. In uno studio di 12 settimane, un gruppo che consumava 50g di proteine isolate di pisello al giorno ha registrato un aumento maggiore dello spessore muscolare indotto dall’allenamento di resistenza rispetto al placebo, non significativamente diverso da un gruppo che consumava 50g di siero di latte [39].
Pertanto, nel contesto delle indicazioni di questo articolo, la qualità delle proteine può essere un problema solo se si utilizza la fascia bassa delle linee guida sulle proteine (1,6g/kg) o se si consuma una dieta a base prevalentemente vegetale. In entrambi i casi, potrebbe essere utile integrare con fonti proteiche ricche di Leucina e di EAA, a seconda delle preferenze alimentari (ad esempio, proteine del latte o del pisello se si è vegani), per garantire la risposta attesa della MPS all’assunzione di proteine.
Grassi:
Il grasso è un nutriente fondamentale per molte funzioni dell’organismo. Tuttavia, non si sa molto dell’effetto dei grassi alimentari sull’ipertrofia del muscolo scheletrico. È stato riportato che l’assunzione di grassi alimentari tra i bodybuilder varia dall’8 al 33% delle calorie totali [2]. Sebbene i trigliceridi intramuscolari possano fungere da substrato energetico durante l’allenamento di resistenza, non sono un fattore limitante poiché i substrati derivano principalmente da processi anaerobici [40]. Di interesse per il bodybuilder, è dimostrato che negli atleti allenati contro-resistenza [41] e nei giocatori di hockey [42] le diete a basso contenuto di carboidrati (30-45% dell’energia o meno) possono influire sul rapporto Testosterone libero/Cortisolo (fTC), il che potrebbe avere un impatto negativo sul recupero. D’altra parte, la riduzione dei grassi alimentari nelle diete isocaloriche da ~30-40% a ~15-25% ha portato a riduzioni significative ma modeste dei livelli di Testosterone [43,44,45,46].
Tuttavia, non è chiaro se le variazioni di Testosterone all’interno di intervalli normali influenzino in modo significativo l’aumento della massa muscolare [47]. Nonostante la possibilità che i livelli di testosterone possano essere più elevati quando si consuma una percentuale maggiore di energia proveniente dai grassi alimentari, i cambiamenti effettivi nella massa muscolare durante gli studi longitudinali di individui allenati alla resistenza che seguono diete “chetogeniche” ad alto contenuto di grassi sono stati costantemente inferiori rispetto ad approcci moderati o a basso contenuto di grassi con ampi carboidrati [48,49,50,51]. Non è ancora stato chiarito se ciò sia dovuto a cambiamenti nella capacità di esercizio, ad alterazioni del rapporto fTC o a qualche altro meccanismo legato alla componente ad alto contenuto di grassi o a basso contenuto di carboidrati della dieta.
Tuttavia, ciò indica che forse si dovrebbe consumare una proporzione più moderata di grassi nella dieta, piuttosto che un apporto basso o alto. In letteratura sono state proposte raccomandazioni del 15-20% e del 20-30% delle calorie provenienti dai grassi alimentari [15,52]. Tuttavia, sono necessarie ulteriori ricerche per stabilire l’effetto e la quantità ottimale di grassi alimentari per favorire l’ipertrofia muscolare.
Sulla base delle evidenze attuali, può essere prudente raccomandare che i grassi alimentari rappresentino il 20-35% delle calorie, in linea con le raccomandazioni dell’American College of Sports Medicine per gli atleti [53], che nella maggior parte dei casi corrispondono a circa 0,5-1,5 g/kg/giorno. Inoltre, va notato che un apporto sufficiente di proteine e carboidrati non deve essere compromesso da un’elevata assunzione di grassi nella dieta.
Anche la qualità dei grassi, come gli essenziali omega 3 e gli omega 6, potrebbe essere importante per i bodybuilder. Se l’apporto di questi acidi grassi è sufficiente, non è necessario integrarli con una dieta di alta qualità contenente buone fonti di acidi grassi. Tuttavia, per alcuni potrebbe essere difficile assumere le quantità ottimali. Per questo motivo, l’argomento verrà trattato in modo più approfondito nella sezione dedicata agli integratori alimentari.
Carboidrati:
A differenza delle proteine e dei grassi, i carboidrati sono considerati non essenziali per la dieta umana perché l’organismo è in grado di produrre il glucosio necessario ai tessuti attraverso la gluconeogenesi [54]. Tuttavia, l’assunzione di carboidrati ha un ruolo importante nella dieta del bodybuilder come regolatore degli ormoni tiroidei e come contributo al fabbisogno di micronutrienti [55,56]. Inoltre, una dieta a basso contenuto di carboidrati potrebbe limitare la rigenerazione dell’adenosina trifosfato (ATP) e limitare la capacità dei muscoli di contrarsi con una forza elevata [57,58]. Durante l’esercizio ad alta intensità, il glicogeno muscolare è il principale contributore di substrato energetico ed è stato dimostrato che la glicolisi fornisce circa l’80% del fabbisogno di ATP di una serie di flessioni del gomito se portata al cedimento muscolare [59]. Nonostante ciò, parte del glicogeno utilizzato durante questo tipo di esercizio può essere risintetizzato dal lattato, il che potrebbe ridurre il fabbisogno di carboidrati. È stato inoltre dimostrato che l’allenamento contro-resistenza riduce il glicogeno muscolare del 24-40% in una singola sessione [59,60].
La quantità esaurita può variare in base alla durata, all’intensità e al lavoro svolto, ma l’allenamento tipico del bodybuilding con ripetizioni più elevate e carichi moderati sembra causare la maggiore riduzione delle scorte di glicogeno muscolare [61]. Inoltre, è stato suggerito che quando le scorte di glicogeno sono troppo basse (~70 mmol/kg), ciò può inibire il rilascio di calcio e accelerare l’insorgenza della fatica muscolare [62]. Un basso livello di glicogeno muscolare riduce significativamente il numero di ripetizioni eseguite quando si eseguono tre serie di Squat all’80% di 1RM [57].
Tuttavia, è stato dimostrato che il consumo di una dieta contenente 7,7 g/kg/die di carboidrati per 48 ore prima di una sessione di allenamento non ha un effetto maggiore sulle prestazioni rispetto a 0,37g/kg/die quando si eseguono 15 serie a 15RM di esercizi per la parte inferiore del corpo [63]. Analogamente, un altro studio ha rilevato che una dieta con il 70% di carboidrati rispetto a una dieta con il 50% di carboidrati non ha un effetto maggiore sulle prestazioni durante l’esercizio sopramassimale; tuttavia, una dieta composta dal 25% di carboidrati ha ridotto significativamente le prestazioni [64].
Inoltre, visti gli effetti negativi a lungo termine sulla massa muscolare osservati di recente in studi su popolazioni allenate alla resistenza che seguono diete chetogeniche [49,51], potrebbe essere prudente per i bodybuilder assicurarsi semplicemente un apporto sufficiente di carboidrati, visti questi risultati disparati. Pertanto, mentre le diete a moderato e alto contenuto di carboidrati sono probabilmente appropriate per il bodybuilding, le diete a bassissimo contenuto di carboidrati possono essere dannose per l’allenamento.
Nei bodybuilder maschi, sono stati riportati apporti medi di carboidrati pari a 5,3g/kg/giorno durante la Off-Season [2]. Tuttavia, non sono state stabilite le quantità ottimali di carboidrati per i bodybuilder. In letteratura sono state proposte raccomandazioni per gli sport di forza, tra cui il bodybuilding, con assunzioni di 4-7g/kg/giorno e 5-6g/kg [15,65]. I carboidrati sembrano essere importanti per il bodybuilder, ma per ottenere benefici possono essere necessarie solo quantità moderate. Pertanto, dopo aver destinato le calorie alle proteine (1,6-2,2g/kg/die) e ai grassi (0,5-1,5g/kg/die), le restanti calorie dovrebbero essere destinate ai carboidrati. Tuttavia, sulla base delle prove attuali, potrebbe essere ragionevole consumare quantità sufficienti di carboidrati nell’intervallo ≥3-5g/kg/giorno, se possibile.
Sono necessarie ulteriori ricerche tra i bodybuilder per stabilire se l’assunzione abituale di carboidrati, superiore o inferiore a quella osservata, possa produrre ulteriori benefici. La Tabella sottostante riassume le raccomandazioni per le calorie e i macronutrienti.
Raccomandazioni dietetiche per i bodybuilder in Off-Season.
Distribuzione e timing dei nutrienti:
Si dice che i bodybuilder consumino in media sei pasti al giorno [66]; tuttavia, non esistono studi che esaminino specificamente quale possa essere la frequenza ottimale dei pasti per questa popolazione [65]. Questa elevata frequenza dei pasti si basa sulla convinzione di un maggiore stato di anabolismo e persino di un migliore utilizzo dei nutrienti durante il giorno, che potrebbe tradursi in un miglioramento della composizione corporea.
Il concetto di temporizzazione dell’assunzione di proteine per massimizzare l’ipertrofia comprende diverse strategie di dosaggio. La prima a comparire in letteratura è stata il consumo di proteine in prossimità dell’allenamento contro-resistenza. I picchi di MPS sono più elevati in questo periodo quando si consumano proteine; pertanto, questa strategia è stata proposta per migliorare l’efficienza della riparazione e del rimodellamento del muscolo scheletrico [31]. Inoltre, a causa dell'”effetto muscolo pieno”, per cui un ulteriore apporto di proteine non aumenta la MPS finché non è trascorso un tempo sufficiente, distribuire uniformemente l’assunzione di proteine tra più pasti è un’altra strategia studiata per massimizzare la MPS totale giornaliera [67]. Infine, il consumo prima di andare a letto di proteine a lenta digestione (come la caseina) per evitare periodi catabolici prolungati durante il sonno è la strategia proposta più di recente per migliorare il bilancio proteico netto giornaliero [68], sebbene si sia dimostrata inutile nel perseguire il fine o, per lo meno, non molto diversa dalla risultante di una assunzione di isolate in un contesto alimentare con parità nel totale proteico giornaliero. Ciascuna di queste tre strategie sarà discussa in seguito.
Dosaggio proteico:
Il periodo post-allenamento consente un picco della MPS più elevato quando si consumano proteine [31] e per raggiungere il picco di MPS può essere necessaria un’adeguata dose di Leucina “soglia” [32]. Diversi studi hanno esaminato il dosaggio proteico necessario per massimizzare la MPS dopo l’allenamento [69,70,71]. In uno studio sono stati consumati 0, 5, 10, 20 o 40g di proteine d’uovo intere dopo l’esercizio contro-resistenza della parte inferiore del corpo, con 20g che stimolavano al massimo la MPS [69]. Risultati simili sono stati riscontrati anche in un altro studio, in cui 20 g di siero di latte sono stati sufficienti a stimolare al massimo i tassi post-assorbitivi di MPS sia a riposo che dopo un lavoro unilaterale delle gambe all’80% del 1RM [70]. Inoltre, 40g di siero di latte non hanno prodotto ulteriori aumenti di MPS in questo studio e hanno portato all’ossidazione amminoacidica e alla produzione di urea.
Tuttavia, uno studio recente ha rilevato che, durante l’esecuzione di esercizi contro-resistenza per tutto il corpo al 75% del 1RM, 40g di siero di latte hanno prodotto una risposta MPS significativamente più elevata rispetto a 20g [71]. Esiste quindi una relazione tra il volume di tessuto muscolare danneggiato e stimolato e l’assunzione adeguata di proteine. È interessante notare che gli autori di una meta-analisi del 2013 hanno osservato che, nonostante gli studi con traccianti a breve termine mostrassero risposte nella MPS maggiori quando le proteine venivano consumate nella “finestra anabolica” post-allenamento, negli studi longitudinali sull’allenamento non è stato riscontrato alcun effetto significativo sull’ipertrofia quando si controllava l’apporto proteico totale giornaliero, indipendentemente dal fatto che le proteine fossero consumate all’interno della “finestra anabolica” o al di fuori di essa [72].
Nutrient Timing:
Analogamente, i ricercatori di uno studio tracciante a breve termine che ha esaminato il dosaggio delle proteine nel corso di 12 ore hanno riportato una maggiore area sotto la curva della MPS quando sono state consumate quattro dosi di proteine del siero di latte da 20g ogni tre ore rispetto a due dosi da 40g a distanza di sei ore e otto dosi da 10g ogni ora e mezza [73]. In teoria, data la soglia oltre la quale le proteine supplementari consumate in una singola seduta non contribuiscono ulteriormente alla MPS [69] e a causa del “periodo refrattario” postprandiale durante il quale la MPS non può essere nuovamente stimolata al massimo [67], si potrebbe concludere che un bodybuilder dovrebbe raggiungere, ma non superare, questa dose soglia ogni poche ore per massimizzare l’ipertrofia a lungo termine. Tuttavia, gli autori di una review sistematica del 2018 sugli integratori proteici, comprendente 34 studi randomizzati e controllati, hanno riportato guadagni di massa magra simili tra i gruppi che utilizzavano un programma di dosaggio con i pasti (che comportava un minor numero di dosi di proteine di entità elevata) e tra i pasti (che comportava un maggior numero di dosi di proteine di entità moderata) [74].
È interessante notare che i dati che esaminano l’alimentazione proteica notturna mostrano uno distacco simile tra gli studi meccanicistici a breve termine e gli interventi di allenamento a lungo termine. Nel 2012 è stata condotta la prima ricerca che esaminava la risposta acuta all’alimentazione notturna con caseina [68]. Gli autori hanno riportato che 40g di caseina consumati prima di andare a letto sono stati digeriti, assorbiti e hanno stimolato la MPS e migliorato l’equilibrio proteico dell’intero corpo durante il periodo notturno in misura maggiore rispetto al placebo. Negli anni successivi sono stati pubblicati altri studi in acuto che hanno confermato [75] e riconfermato questi risultati in una popolazione più anziana [76]. Nel 2015, gli autori del primo studio longitudinale hanno riportato un aumento della forza e dell’ipertrofia in un gruppo a cui era stato somministrato un supplemento proteico notturno rispetto a un gruppo placebo [77].
Tuttavia, la quantità totale di proteine giornaliere non è stata equiparata, in quanto il gruppo con proteine notturne ha consumato 1,9g/kg/giorno, mentre il gruppo placebo ha consumato solo 1,3g/kg. È importante notare che in entrambi gli unici studi longitudinali con corrispondenza proteica che hanno confrontato l’integrazione notturna di caseina con i gruppi che hanno assunto l’integrazione prima, non sono state riportate differenze significative nell’aumento della FFM tra i gruppi [78,79]. Pertanto, la domanda è la stessa per ogni strategia di distribuzione: perché ci sono ripetuti distacchi tra gli studi meccanicistici a breve termine sulle MPS e le ricerche a lungo termine che esaminano l’effettiva ipertrofia? La risposta potrebbe risiedere nei metodi utilizzati negli studi sulla MPS, in quanto i partecipanti sono a digiuno, ricevono solo proteine in polvere in isolamento, spesso viene loro somministrato del siero di latte (che viene digerito molto rapidamente) e vengono osservati per brevi periodi. Questi contesti di laboratorio determinano tempi di digestione e cinetiche degli aminoacidi diversi da quelli che si verificano nel “mondo reale”. In particolare, in queste condizioni di laboratorio i livelli di base degli aminoacidi nel corpo sono più bassi del normale e la digestione e il successivo apporto di aminoacidi al muscolo sono più rapidi.
In condizioni di vita libera, le proteine vengono consumate principalmente da fonti alimentari intere, più volte al giorno e insieme ad altri alimenti, il che ritarda lo svuotamento gastrico. Per questi motivi, gli aminoacidi vengono titolati nel flusso sanguigno in modo più lento e costante; pertanto, in condizioni normali, le scorte sono quasi sempre prontamente disponibili [80]. Pertanto, l’efficacia della “finestra anabolica” e persino delle strategie di distribuzione delle proteine potrebbe non tradursi nella pratica. Inoltre, le limitazioni specifiche del laboratorio si estendono anche agli studi sull’alimentazione notturna. Si consideri, ad esempio, che 26g di proteine provenienti da una bistecca magra determinano un aumento sostenuto della MPS che dura almeno sei ore (l’intero periodo di tempo studiato) [81].
Inoltre, 26g sono solo il ~37% della dose di proteine contenuta in media in una cena americana [82], che richiederebbe più tempo per essere digerita a causa della maggiore porzione di proteine e dell’aggiunta di fibre, lipidi e altri nutrienti che ritarderebbero ulteriormente la digestione [80]. Pertanto, il tipico pasto finale potrebbe già soddisfare lo scopo di un frullato di caseina. Detto questo, nonostante queste discrepanze tra MPS e risultati della composizione corporea, non c’è nulla di male nel tentare queste strategie, soprattutto se attuate in modo pragmatico e senza introdurre ulteriori oneri logistici nel proprio programma quotidiano.
Pertanto, potrebbe essere prudente consigliare ai bodybuilder di suddividere l’assunzione giornaliera di 1,6-2,2 g/kg di proteine in più pasti contenenti ciascuno ~0,40-0,55g/kg [80] e di fare in modo che uno di questi pasti avvenga entro 1-2 ore prima o dopo l’allenamento, mentre un’alimentazione costituita da una fonte proteica e non proteica venga consumata 1-2 ore prima di dormire. Ad esempio, un bodybuilder di 90 kg potrebbe consumare 40-50g di proteine alle 8-9 del mattino per la colazione, allenarsi alle 11, consumare 40-50g di proteine alle 12-13 per il pranzo/post-allenamento, 40-50g di proteine a cena tra le 17-18, e poi un pasto finale di 40-50g di proteine non contenenti fonti proteiche grasse alle 21-10 prima di andare a letto entro le 23.
I carboidrati consumati prima dell’allenamento sono spesso una strategia utilizzata dagli atleti per migliorare le prestazioni negli esercizi ad alta intensità. La completa risintesi del glicogeno può essere raggiunta entro 24 ore da un allenamento che depaupera il glicogeno se si consumano quantità sufficienti di carboidrati [83]. Tuttavia, solo il 24-40% del glicogeno muscolare viene esaurito dopo un allenamento contro-resistenza [59,60]. Pertanto, una quantità di ≥3-5g/kg di carboidrati al giorno sarebbe probabilmente sufficiente per la risintesi del glicogeno. Questo elevato apporto giornaliero di carboidrati probabilmente riduce anche l’impatto della tempistica dei carboidrati pre-allenamento sulle prestazioni dell’esercizio.
Inoltre, per i bodybuilder che non hanno bisogno di enfatizzare il rifornimento di glicogeno, le proteine aumentano la MPS post-allenamento a livelli massimi anche senza l’aggiunta di carboidrati [86,87]. Anche se il consumo di carboidrati nel post-allenamento non è certo dannoso, è improbabile che questo favorisca l’ipertrofia a lungo termine, come discusso in precedenti review [1,88]. Pertanto, è meglio concentrarsi sul consumo di un’adeguata quantità di carboidrati giornalieri e basare la distribuzione dei carboidrati intorno all’allenamento sulle preferenze personali.
Supplementazione OTC:
In un recente sondaggio condotto tra i bodybuilder, è stato riportato che tutti i partecipanti assumevano integratori alimentari [9]. Gli integratori alimentari più comuni erano: integratori di proteine (86%), creatina (68%), aminoacidi a catena ramificata (67%), glutammina (42%), vitamine (40%), olio di pesce (37%) e prodotti contenenti caffeina/efedrina (24%).
Sebbene gli integratori proteici siano molto popolari tra i bodybuilder, vengono utilizzati prevalentemente come gli alimenti interi per raggiungere gli obiettivi proteici. Pertanto, non verranno discussi in dettaglio. I lettori sono invitati a leggere la posizione dell’ISSN su questo argomento [89]. Inoltre, la trattazione di tutti gli integratori comunemente utilizzati dai bodybuilder esula dallo scopo di questo articolo. L’attenzione si concentrerà piuttosto sugli integratori alimentari che potrebbero potenzialmente produrre un effetto ergogenico e sugli integratori che possono garantire un apporto sufficiente di micronutrienti e acidi grassi essenziali.
Creatina Monoidrato:
La Creatin-fosfato si trova in alte concentrazioni nel muscolo scheletrico e cardiaco, dove agisce come fonte di energia [90]. La Creatina può essere ottenuta anche attraverso la dieta nei soggetti che consumano carne; tuttavia, le concentrazioni di Creatina nella carne si riducono con la cottura [91].
Numerosi studi hanno osservato un aumento della massa e della forza muscolare in seguito a fasi di carico di Creatina, in genere di 20g al giorno per circa una settimana, spesso seguite da fasi di mantenimento di 2-3g di Creatina al giorno [92]. Tuttavia, la fase di carico potrebbe non essere necessaria. È stato dimostrato che la saturazione della Creatina muscolare dopo un’integrazione di 3g di Creatina Monoidrato per 28 giorni è simile al consumo di Creatina Monoidrato dopo la tipica fase di carico [93].
La maggior parte degli individui non raggiunge i 3g giornalieri con la dieta e può essere necessaria un’integrazione. Esistono numerose forme di Creatina negli integratori in commercio, tra le quali la Creatina Monoidrato è la più studiata. Le versioni più recenti di Creatina, come la kre-alkalyn [94] e la Creatina etil-estere [95], non si sono dimostrate superiori alla Creatina Monoidrato, nonostante abbiano in genere un prezzo più elevato. Pertanto, si raccomanda il consumo di 3-5g di Creatina Monoidrato al giorno. La tempistica di assunzione della Creatina non sembra avere importanza, poiché la saturazione delle riserve di Creatin-fosfato richiede circa 28 giorni per raggiungere le concentrazioni massime quando si consumano 3g al giorno e non ha un effetto in acuto [93].
Caffeina:
Uno degli integratori alimentari più utilizzati dai bodybuilder sono gli stimolanti, in particolare la Caffeina [9]. Oltre ad aumentare l’eccitazione [96], la Caffeina può ridurre il dolore e lo sforzo percepito durante l’esercizio [97] e migliora la gestione del Calcio, aumentando la potenza [98]. Studi sull’allenamento contro-resistenza hanno rilevato che la Caffeina riduce la fatica e aumenta la forza [99,100]. Tuttavia, non tutti gli studi hanno dimostrato un effetto ergogenico sull’allenamento contro-resistenza [101]. Gli studi che hanno dimostrato un effetto ergogenico hanno utilizzato dosaggi elevati di caffeina (5-6 mg/kg), che sono al limite superiore di quello che è considerato un dosaggio sicuro [99,100]. Tuttavia, può essere consigliabile consumare il dosaggio minimo efficace per individuo, poiché l’assunzione regolare può generare tolleranza [102]. A causa dell’effetto acuto della Caffeina, è consigliabile assumerla circa 1 ora prima dell’esercizio fisico [99]. Tuttavia, l’emivita della Caffeina è di circa 3-9 ore; pertanto, può essere consigliabile consumare la Caffeina all’inizio della giornata per favorire un sonno sano se l’esercizio fisico viene svolto più tardi nel corso della giornata [103]. Sono necessarie ulteriori ricerche per trovare un consenso sull’uso della Caffeina nell’allenamento contro-resistenza, ma sulla base delle prove attuali un dosaggio di 5-6 mg/kg consumato prima dell’esercizio potrebbe produrre un effetto ergogenico sulle prestazioni nell’allenamento contro-resistenza.
Beta-Alanina:
È stato dimostrato che l’ingestione di 4-6 g di beta-alanina aumenta i livelli di carnosina muscolare [104]. La carnosina agisce come tampone del pH nel muscolo scheletrico e può ritardare l’inizio dell’affaticamento muscolare durante l’esercizio ad alta intensità [105]. Una meta-analisi ha concluso che la beta-alanina potrebbe produrre effetti ergogenici durante l’esercizio ad alta intensità della durata di 60-240 secondi [104]. Inoltre, non sono stati riscontrati effetti benefici negli esercizi di durata inferiore a 60 secondi. La maggior parte degli studi inclusi nella meta-analisi riguardava l’esercizio di resistenza.
Tuttavia, è dimostrato che l’integrazione di beta-alanina può migliorare la resistenza muscolare negli atleti allenati alla resistenza [105] e può migliorare la composizione corporea [106]. Sono necessari ulteriori studi per esaminare l’effetto ergogenico della beta-alanina sulla composizione corporea e sulle prestazioni. Tuttavia, dato che i bodybuilder si allenano spesso con più di 10 ripetizioni per serie e spesso includono tecniche di intensità come drop set, pause di riposo, myo reps e altre, la beta-alanina potrebbe apportare un beneficio alla resistenza di queste serie [9].
Pertanto, potrebbe essere ragionevole per un bodybuilder consumare 3-5 g di beta alanina al giorno durante le fasi di allenamento ad alte ripetizioni o nelle fasi di allenamento in cui si incorporano diverse tecniche di intensità che prolungano la durata di un set. Come la creatina monoidrato, la beta-alanina non ha un effetto acuto, in quanto le concentrazioni di carnosina muscolare richiedono circa 4 settimane per raggiungere concentrazioni tali da produrre un effetto ergogenico, a condizione che se ne consumi una quantità sufficiente al giorno [104].
Citrullina Malato:
Recentemente, la Citrullina Malato ha guadagnato popolarità tra i bodybuilder. Il potenziale effetto ergogenico è dovuto all’aumento del flusso ematico al muscolo, alla produzione di ATP e alla potenziale capacità della Citrullina Malato di agire come agente tampone [107]. È stato dimostrato che il consumo di 8g di Citrullina Malato aumenta le ripetizioni fino al cedimento del 50% [107,108,109,110], riduce l’indolenzimento muscolare del 40% [107] e migliora la forza massimale e la potenza anaerobica [111].
Tuttavia, non tutti gli studi hanno osservato effetti ergogenici del consumo di Citrullina Malato. Due studi recenti non hanno mostrato un miglioramento delle prestazioni, un aumento della risposta del gonfiore muscolare dovuto all’allenamento, un’attenuazione della fatica o un aumento dell’attenzione e dell’energia in seguito all’integrazione di Citrullina Malato in uomini allenati contro-resistenza a livello amatoriale [112,113].
Una recente meta-analisi di Trexler et al. ha analizzato 12 studi sullla CM per le prestazioni di forza e potenza [114]. Sebbene abbiano riscontrato solo una piccola dimensione dell’effetto (0,20), hanno concluso che questo potrebbe essere rilevante per gli atleti di alto livello in cui i risultati delle competizioni si decidono su margini ridotti, come i culturisti agonisti di alto livello. Si consiglia di assumere la Citrullina Malato circa 60 minuti prima dell’esercizio fisico per consentire un assorbimento sufficiente.
Sono necessarie ulteriori ricerche per determinare l’efficacia della Citrullina Malato nell’esercizio contro-resistenza. Allo stato attuale, i dati indicano un effetto benefico o neutro sulle prestazioni. Pertanto, sulla base delle prove attuali, 8g al giorno di Citrullina Malato consumati prima dell’esercizio potrebbero avere dei benefici interessanti per i bodybuilder.
Alfa-GPC:
L’Alfa-GPC (alfa-glicerofosfocolina o colina alfoscerato) è un fosfolipide contenente colina. Quando viene ingerita, l’Alfa-GPC viene metabolizzata in colina e glicerolo-1-fosfato. La colina è un precursore dell’acetilcolina, un neurotrasmettitore coinvolto nella memoria, nell’attenzione e nella contrazione dei muscoli scheletrici. Il glicerolo-1-fosfato serve a sostenere le membrane cellulari.[https://pubmed.ncbi.nlm.]
L’Alfa-GPC sembra attraversare facilmente la barriera emato-encefalica e viene assorbito rapidamente. Attualmente è il miglior colinergico per aumentare i livelli plasmatici e cerebrali di colina.[https://pubmed.ncbi.nlm.]
L’integrazione orale di Alfa-GPC è interessante soprattutto per scopi nootropici o di potenziamento cognitivo. Esistono numerosi studi sui roditori che supportano questo effetto, ma non è ancora stato dimostrato negli esseri umani altrimenti sani. Negli anziani affetti da demenza lieve o moderata – che comporta un’alterazione della neurotrasmissione colinergica – l’Alfa-GPC migliora i sintomi cognitivi (ad esempio, disturbi della memoria e dell’attenzione).[https://pubmed.ncbi.nlm] L’Alfa-GPC può anche migliorare l’efficacia degli inibitori dell’acetilcolinesterasi (cioè i farmaci che aumentano la disponibilità di acetilcolina rallentandone la degradazione), utilizzati per il trattamento della malattia di Alzheimer.[https://pubmed.ncbi.nlm.]
Gli atleti sono un’altra popolazione che può trarre beneficio dall’integrazione di Alfa-GPC. Prove preliminari suggeriscono che l’alfa-GPC aumenta la potenza del salto verticale.[https://jissn.biomedcentral.com][https://pubmed.ncbi.nlm.] Inoltre, uno studio pilota ha riportato che l’Alfa-GPC ha aumentato il picco di forza nella panca, ma non la potenza di picco o il tasso di sviluppo della forza.[Ziegenfuss T, Landis J, Hofheins JJ Int Soc Sports Nutr.] Attualmente non è chiaro se l’Alfa-GPC aumenti la forza isometrica, ma i dati empirici e aneddotici sono incoraggianti [https://pubmed.ncbi.nlm.]
L’integrazione di un dosaggio pari a 600mg di Alpha-GPC prima di un test di potenza (spinte su panca) ha riportato un miglioramento della potenza del 14% rispetto al placebo quando assunta 45 minuti prima dell’attività; si trattava di uno studio pilota.[http://www.jissn.com] In media si è notato che il dosaggio di Alfa-GPC efficacie per trarre miglioramenti nella forza è nel range dei 300-600mg 45-30 minuti prima della seduta allenante.
Multi Vitaminico-Multi Minerale:
Storicamente, i bodybuilder hanno utilizzato diete restrittive che eliminano alimenti o interi gruppi di alimenti. Di conseguenza, sono comuni numerose carenze di vitamine e minerali. Nei bodybuilder a dieta sono state osservate carenze di Calcio, vitamina D, Zinco, Ferro e altre ancora [115,116,117]. Tuttavia, la maggior parte della letteratura sulle pratiche alimentari dei bodybuilder risale agli anni ’80 e ’90; pertanto, sono necessari dati più recenti [2].
Più di recente, le pratiche alimentari dei bodybuilder che seguono una dieta tradizionale restrittiva sono state confrontate con quelle degli agonisti che utilizzano un approccio dietetico basato sui macronutrienti, in cui nessun alimento o gruppo alimentare è off limits [118]. Non sorprende che i concorrenti che utilizzano un approccio dietetico più flessibile presentino meno carenze di micronutrienti. In particolare, la vitamina E, la vitamina K e le proteine sono risultate significativamente inferiori nelle donne che utilizzavano approcci dietetici rigidi rispetto a quelle che utilizzavano approcci più flessibili. Nel presente articolo, specie se si parla di Off-Season, si raccomanda di utilizzare un approccio dietetico flessibile, in cui nessun alimento o gruppo viene eliminato dalla dieta.
In questo modo, è meno probabile che si verifichino carenze di micronutrienti, soprattutto se si considera che le atlete in Off-Season hanno a disposizione una maggiore quantità di calorie rispetto a quelle a dieta per un contest, il che dovrebbe consentire loro di incorporare una maggiore varietà di alimenti.
Ciononostante, può essere consigliabile raccomandare un integratore multivitaminico/minerale a basso dosaggio (≤100% RDA) come misura di sicurezza per prevenire eventuali carenze di micronutrienti, sottolineando al contempo il consumo di una buona varietà di alimenti al giorno per soddisfare il fabbisogno di micronutrienti.
Omega 3 (EPA-DHA):
Gli acidi grassi polinsaturi con un doppio legame a tre atomi di distanza dal gruppo metilico terminale sono noti come ω-3 o acidi grassi omega-3 (O3). Un basso apporto di O3 nelle diete occidentali rispetto ad altre fonti di grassi alimentari (come gli acidi grassi omega-6) è associato a un peggioramento della salute multispettrale negli studi epidemiologici [119]. Pertanto, è interessante concentrarsi specificamente sulle modifiche della dieta per fornire acidi eicosapentaenoici e docosaesaenoici (EPA e DHA) – la carenza alimentare più comune nel mondo occidentale; ma vale la pena notare che la misurazione, l’interazione e l’effetto di O3 e acidi grassi omega-6 in relazione alla salute non sono chiari e vanno oltre lo scopo di questo articolo. Per una rassegna si rimanda ad altra pubblicazione [120].
Oltre alla salute, c’è interesse per i potenziali effetti anabolici degli integratori di EPA e DHA [121], che di solito vengono forniti attraverso l’olio di pesce o, in alcuni casi, l’olio di alghe. Tuttavia, ci sono dati contrastanti sulla capacità dell’olio di pesce di aumentare la risposta della sintesi proteica muscolare all’ingestione di proteine. Mentre un articolo di revisione del 2014 ha evidenziato una serie di studi secondo cui l’olio di pesce può aumentare la risposta [122], uno studio recente non ha rilevato alcun effetto sulla risposta della MPS a una sessione di allenamento contro-resistenza e all’ingestione di proteine dopo l’allenamento [123]. Inoltre, i dati sull’ipertrofia longitudinale sono pochi [124] e gli studi sulle prestazioni dell’allenamento contro-resistenza sono contrastanti [125] e in gran parte non applicabili o difficili da valutare a causa dell’uso di partecipanti non allenati o di allenamenti non standardizzati ed ecologicamente non realistici rispetto al bodybuilding.
In una recente review che affronta specificamente la questione se gli integratori di O3 possano o meno aumentare l’ipertrofia [126], gli autori hanno concluso che attualmente non ci sono prove sufficienti per fare tale affermazione. Sebbene siano necessarie ulteriori ricerche prima di poter raccomandare l’integrazione di O3 (o di alterazioni della dieta) a fini di costruzione muscolare, i benefici per la salute dell’integrazione di O3 sono degni di nota. Ad esempio, recenti meta-analisi hanno riportato che l’integrazione di olio di pesce riduce i sintomi della depressione [127], diminuisce il rischio di morte cardiaca [128], riduce la pressione sanguigna [129] e diminuisce la circonferenza vita [130]. Pertanto, gli atleti estetici possono prendere in considerazione l’integrazione giornaliera di olio di pesce (o di alghe) (1.5-2.5g di EPA/DHA) per la salute generale e multi spettro, ma sono necessari studi futuri per formulare raccomandazioni relative alle prestazioni nel bodybuilding.
Acido Arachidonico (AA):
L’Acido Arachidonico (AA) è l’acido grasso omega-6 più rilevante dal punto di vista biologico e, nella membrana lipidica di una cellula, è l’acido grasso che viene confrontato con i due acidi grassi dell’olio di pesce (EPA e DHA) nella costituzione di un rapporto omega-3:6. Dati recenti suggeriscono un’assunzione giornaliera di 50-250mg di Acido Arachidonico[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] con alcune fonti che stimano livelli fino a 500mg al giorno;[https://www.ncbi.nlm.] l’assunzione di Acido Arachidonico sembra essere inferiore nei vegetariani[https://www.ncbi.nlm.].
Si ritiene che l’Acido Arachidonico sia importante per il metabolismo del muscolo scheletrico, poiché si pensa che i fosfolipidi della membrana del sarcoplasma riflettano la dieta,[https://www.ncbi.nlm.][https://www.ncbi.nlm.] l’allenamento stesso sembra alterare il contenuto di fosfolipidi del muscolo (indipendentemente dalla composizione delle fibre muscolari[https://www.ncbi.nlm.] e associato a un rapporto omega 6:3 più basso[https://www.ncbi.nlm.][https://www.ncbi.nlm.]) e gli eicosanoidi dell’Acido Arachidonico interagiscono con la sintesi proteica muscolare attraverso i loro recettori.
L’Acido Arachidonico segnala la sintesi proteica muscolare attraverso una via dipendente dalla COX-2 (che suggerisce il coinvolgimento delle prostaglandine)[https://www.ncbi.nlm.] che è associata ad aumenti sia della prostaglandina E2 (PGE2) che del PGF(2α),[https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.][https://www.ncbi.nlm.] anche se l’incubazione con PGE2 o PGF(2α) isolati non sembra replicare pienamente gli effetti ipertrofici dell’Acido Arachidonico. [https://www.ncbi.nlm.] PGE2 e PGF(2α) sono indotti anche dall’esercizio fisico (nello specifico, dallo stiramento delle cellule muscolari in vitro[https://www.ncbi.nlm.]) ed è stato osservato sia nel siero[https://pubmed.ncbi.nlm.][https://www.ncbi.nlm.] che a livello intramuscolare (quadruplicato, da 0,95+/-0,26ng/mL a 3,97+/-0. La capacità del riflesso da stiramento di aumentare le concentrazioni di PGE2 e PGF(2α)[https://www.ncbi.nlm.] potrebbe essere dovuta semplicemente al fatto che lo stiramento aumenta l’attività delle COX2.[https://www.ncbi.nlm.][https://www.ncbi.nlm.]
Va notato che l’integrazione di 1.500mg di Acido Arachidonico (rispetto a una dieta di controllo contenente 200mg dello stesso) per 49 giorni ha aumentato la secrezione di PGE2 da parte di cellule immunitarie stimolate (del 50-100%) in giovani uomini altrimenti sani,[https://www.ncbi.nlm.] ma la rilevanza di questo studio per il muscolo scheletrico non è nota. Questo studio ha anche osservato che, senza stimolazione, non c’erano differenze significative tra i gruppi.[https://www.ncbi.nlm.] Altrove, è stata osservata una tendenza all’aumento delle concentrazioni sieriche di PGE2 a riposo in uomini allenati a cui sono stati somministrati 1.000mg di Acido Arachidonico per 50 giorni.[https://www.ncbi.nlm.]
L’Acido Arachidonico, attraverso gli eicosanoidi noti come PGF(2α) e PGE2, stimola la sintesi proteica muscolare. Sono prodotti a partire dall’Acido Arachidonico, ma normalmente non formano i rispettivi eicosanoidi per la costruzione del muscolo finché la cellula non viene stimolata da un fattore di stress (come il riflesso di stiramento di una cellula muscolare) che ne induce la produzione.
Il recettore per il PGF(2α) (recettore FP) sembra essere sovraregolato dagli inibitori della COX1 (l’acetaminofene utilizzato in questo studio)[https://www.ncbi.nlm.] e si ritiene che una maggiore segnalazione del PGF(2α) sia alla base del miglioramento della sintesi proteica muscolare osservato nei soggetti anziani con farmaci antinfiammatori. La supplementazione di Acido Arachidonico non sembra influenzare la quantità di recettori FP nei giovani;[https://www.ncbi.nlm.] mentre l’esercizio fisico stesso può aumentare il contenuto di recettori EP3, né gli inibitori della COX1[https://www.ncbi.nlm.] né l’Acido Arachidonico[https://www.ncbi.nlm.] sembrano influenzarlo ulteriormente.
Tuttavia, è stato riscontrato che l’uso di inibitori della COX2 (nei giovani) sopprime l’aumento di PGF(2α) indotto dall’esercizio fisico (Ibuprofene e Acetaminofene)[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e di PGE2,[https://www.ncbi.nlm.] il che si pensa sia dovuto al fatto che la conversione da PGH2 in questi metaboliti dipende dall’attività della COX2.
Poiché la produzione di questi eicosanoidi dipende dall’enzima COX2, si ritiene che l’inibizione di questo enzima riduca gli effetti anabolizzanti dell’esercizio fisico se assunto prima dello stesso.
L’acido arachidonico (così come l’EPA dall’olio di pesce) non ha compromesso l’assorbimento del glucosio nelle cellule muscolari isolate e 10μM di acido grasso sono in grado di attenuare la resistenza all’Insulina indotta dai grassi saturi; [https://pubmed.ncbi.nlm.] un fenomeno osservato con i grassi saturi a 18 o più catene di carbonio[https://www.ncbi.nlm.] che non sembra applicarsi agli acidi grassi polinsaturi di uguale lunghezza di catena[https://www.ncbi.nlm.][https://www.ncbi.nlm.] ed è probabilmente legato all’aumento delle ceramidi intracellulari[https://www.ncbi.nlm.] che compromettono la segnalazione di Akt[https://www.ncbi.nlm.][https://www.ncbi.nlm.] e riducono l’assorbimento di glucosio mediato da GLUT4 con l’Insulina.[https://www.ncbi.nlm.]
L’Acido Arachidonico e i grassi polinsaturi omega-3 sono entrambi associati a una migliore sensibilità all’Insulina delle cellule muscolari, che potrebbe essere secondaria alla riduzione dei livelli di grassi saturi nella membrana lipidica e quindi alla riduzione delle concentrazioni intracellulari di ceramidi. È possibile che ciò non sia correlato agli eicosanoidi o al rapporto omega-3:6.
In 31 uomini allenati, sottoposti a un programma di sollevamento pesi e a una dieta standardizzata (500kcal in eccesso con 2g/kg di proteine) con 1g di Acido Arachidonico al giorno o placebo, l’integrazione per 50 giorni è sembrata aumentare la potenza di picco (7,1%) e la potenza media (3,6%) al test di Wingate, ma non è riuscita a influenzare positivamente la massa muscolare o le misure di potenza del sollevamento pesi (bench press e leg press).[https://www.ncbi.nlm.]
Attualmente non ci sono prove sufficienti per raccomandare una dose ideale di integrazione di Acido Arachidonico, ma aneddoticamente si usa un dosaggio di circa 1.500 mg da assumere 45 minuti prima dell’allenamento per un periodo medio di 8 settimane. Non è certo che si tratti di una dose ottimale o che sia necessaria la tempistica.
Va inoltre notato che per le persone affette da patologie infiammatorie croniche, come l’artrite reumatoide o le malattie infiammatorie intestinali, la dose ideale di Acido Arachidonico può essere in realtà una sua restrizione dietetica. Nei casi di malattie infiammatorie, l’integrazione di Acido Arachidonico è probabilmente controindicata.
Raccomandazioni per gli integratori alimentari e il dosaggio per i bodybuilder in Off-Season:
Creatina Monoidrato= 3-5g/die;
Beta-Alanina= 3-5g/die;
Citrullina Malato= 8g/pre-workout;
Alfa-GPC= 300-600mg/pre-workout;
Caffeina= 5-6mg/Kg/pre-workout (media standard tra 200 e 600mg/die);
Multi Vitaminico – Multi Minerale= ≤100% RDA/die;
Omega 3 (EPA-DHA)= 1.5-2.5g/die;
Acido Arachidonico= 1.5g/pre-workout.
Supplementazione PEDs:
Una cosa occorre premettere prima di procedere con la descrizione delle molecole più utilizzate nel contesto della Off-Season: non esistono PEDs esclusivamente confinabili in uno dei contesti della programmazione di un bodybuilder. Esiste il grado di versatilità il quale sta ad indicare quanto una molecola possa essere gestita con facilità in situazioni preparatorie differenti. Esistono molecole che per caratteristiche possono dare vantaggi maggiori in Off-Season/Bulk per via di alcune loro caratteristiche che in altro contesto, per esempio il pre-contest, risulterebbero più complesse da gestire. Ma questo non significa che tali molecole siano generalemnte da considerarsi “off-limitz” in un altra fase della preparazione annuale.
Premesso ciò, l’attenzione in questo paragrafo si concentrerà sui principali PEDs usati in Off-Season.
Tra tutti gli AAS, il Testosterone è quello che non ha bisogno di particolari presentazioni. Si tratta dell’ormone sessuale maschile per antonomasia. Nell’uomo, il Testosterone svolge un ruolo fondamentale nello sviluppo dei tessuti riproduttivi maschili, come i testicoli e la prostata, oltre a promuovere le caratteristiche sessuali secondarie, come l’aumento della massa muscolare e ossea e la crescita dei peli. Inoltre, in entrambi i sessi, il Testosterone è coinvolto nella salute e nel benessere, compresi gli stati d’animo, il comportamento e la prevenzione dell’osteoporosi in cooperazione con l’Estradiolo. Livelli insufficienti di Testosterone negli uomini possono portare ad anomalie, tra cui la fragilità e la perdita ossea.
In generale, il Testosterone promuove la sintesi proteica e quindi la crescita dei tessuti dotati di recettori per gli androgeni. Il Testosterone può essere descritto come avente effetti virilizzanti e anabolizzanti (anche se queste descrizioni categoriali sono in qualche modo arbitrarie, poiché vi è una grande sovrapposizione reciproca tra di essi).
Gli effetti anabolizzanti comprendono la crescita della massa e della forza muscolare, l’aumento della densità e della resistenza ossea e la stimolazione della crescita lineare e della maturazione ossea.
Gli effetti androgeni comprendono la maturazione degli organi sessuali, in particolare del pene, e la formazione dello scroto nel feto, e dopo la nascita (di solito nella pubertà) l’approfondimento della voce, la crescita dei peli del viso (come la barba) e dei peli ascellari. Molti di questi effetti rientrano nella categoria dei caratteri sessuali secondari maschili.
Al principio degli anni 30 del novecento avvenne la sintesi chimica del Testosterone, quando Butenandt e G. Hanisch pubblicarono un articolo che descriveva “Un metodo per preparare il Testosterone dal colesterolo”. Solo una settimana dopo, il terzo gruppo, Ruzicka e A. Wettstein, annunciò una domanda di brevetto in un documento “Sulla preparazione artificiale dell’ormone testicolare Testosterone (Androsten-3-one-17-ol).” Ruzicka e Butenandt ricevettero il premio Nobel per la chimica nel 1939 per il loro lavoro.
Gli studi clinici sull’uomo, che prevedevano dosi PO (per via orale) di Methyltestosterone o iniezioni di Testosterone Propionato, iniziarono già nel 1937. Il Testosterone Propionato è menzionato in una lettera all’editore della rivista Strength and Health nel 1938; questo è il primo riferimento noto a un AAS in una rivista statunitense di sollevamento pesi o Bodybuilding.
Lo sviluppo delle proprietà di costruzione muscolare del Testosterone proseguì negli anni ’40, in Unione Sovietica e nei paesi del blocco orientale come la Germania dell’Est, dove sono stati utilizzati programmi di AAS per migliorare le prestazioni dei sollevatori di pesi olimpici e di altri dilettanti già prima degli anni ’50. In risposta al successo dei sollevatori di pesi russi, il medico della squadra olimpica statunitense John Ziegler lavorò con un equipe di chimici per sviluppare un AAS con effetti androgeni ridotti. Ma questa è un altra storia.
L’uso del Testosterone nello sport si diffuse tra gli anni ’50 e gli anni ’60. Le forme utilizzate nei primi tempi erano il Testosterone in sospensione e il Testosterone Propionato, che rappresentano con il Methyltestosterone (Testosterone metilato in C-17) le forme più datate dell’ormone in questione (1935).
In ambito culturistico, il Testosterone rappresenta un AAS sufficientemente versatile in maniera dose-dipendente e sensibilità-dipendente dal momento che il dosaggio dovrebbe essere tarato in base alle risposte metaboliche soggettive alle quali è soggetto l’ormone (vedi, ad esempio, aromatizzazione in estrogeni). Questo ultimo punto è di estrema importanza al fine di evitare l’uso/abuso di AI (Inibitori dell’Aromatasi) e/o SERM (Modulatori Selettivi del Recettore degli Estrogeni). Oltre a peggiorare potenzialmente il quadro lipidico, sommandosi all’azione degli AAS utilizzati, essi riducono l’espressione epatica di IGF-1 cosa che può ridurre la risposta anabolizzante del protocollo PEDs. Nei soggetti caratterizzati da una elevata sensibilità all’attività estrogenica, le procedure applicate vedono: 1) l’uso di Raloxifene o Tamoxifene (SERM) a dosi sufficienti a impedire la comparsa o il peggioramento di una ginecomastia in stadio iniziale già presente e non ancora asportata chirurgicamente 2) l’uso di dosi fisiologiche di Testosterone come base onde evitare la comparsa di stati letargici, affaticabilità, disfunzioni sessuali ecc 3) l’uso di un “mix” composto da Testosterone e Boldenone (vedi in seguito) tale da poter usufruire della bassa e diversa sensibilità all’azione dell’Enzima Aromatasi su quest’ultimo riuscendo ad avere un controllo estrogenico teoricamente migliore (Testosterone e Boldenone mostrano qualità anabolizzanti intrinseche simili).
In un contesto Off-Season, quindi, vista l’importanza della presenza di un buon livello di Estradiolo sia sul complesso degli effetti anabolizzanti ricercati sia per la sua attività sessuale e cerebrale, il Testosterone andrebbe inizialmente calibrato sul soggetto e nel caso affiancato da dosi altrettanto ben tarate di SERM la dove ne risultasse un reale bisogno.
L’uso di un estere che garantisca un rilascio graduale della molecola (vedi Enantato o Cypionato) risulta la scelta migliore al fine di creare una soglia ematica stabile e esente da picchi e cali che possono risultare controproducenti a livello psicofisico. Tenere sempre in considerazione l’emivita di una molecola è uno dei punti fondamentali per sfruttarla al meglio. Nel caso degli esteri sopra citati, una divisione del dosaggio settimanale in due somministrazioni uguali distanziate da quattro-cinque giorni l’una dall’altra risulta una pratica ottimale allo scopo di creare una soglia ematica stabile.
I dosaggi comunemente utilizzati, parlando di molecole esterificate, vanno da 200mg ad 1g a settimana. Per quanto riguarda il Testosterone in sospensione, le dosi comunemente utilizzate vanno dai 175mg ai 700mg a settimana.
Il Boldenone [1,4-androstadiene-3-one,17b-ol], commercializzato con il nome di Equipoise, Ganabol, Equigan, Ultragan, e Boldane, è uno steroide anabolizzante-androgeno spesso legato all’estere Undecylenato. Strutturalmente molto simile al Testosterone, il Boldenone differisce da questo per il doppio legame tra C1 e C2.
La Ciba brevettò il Boldenone nel 1949. Successivamente, negli anni ’50 e ’60, sviluppò diversi esteri sperimentali del farmaco. Uno di questi era il Boldenone Undecilenato, che fu introdotto per uso clinico con il marchio Parenabol e fu utilizzato alla fine degli anni ’60 e all’inizio degli anni ’70. Tuttavia, fu sospeso prima della fine degli anni ’70. Ad oggi l’uso del Boldenone è legale in alcuni paesi in campo veterinario.
Se qualcuno volesse usare 500mg di Testosterone, ma non potrebbe usare un tale dosaggio dal momento che presenta particolare difficoltà nella gestione estrogenica in specie senza l’uso di AI come Exemestane o Anastrozolo, una conclusione a cui molti superficialmente sono giunti è che si potrebbe semplicemente usare il Boldenone al dosaggio sopra citato per ridurre della metà l’attività estrogenica, ma comunque supportare un’adeguata produzione di Estradiolo. Ma quando si approfondisci l’ipotesi e la si testa sul campo, è davvero così che stanno le cose? In realtà no, o, comunque, la media delle variabili di risposta spinge a confermare una maggiore validità nel “mixare” Testosterone e Boldenone coprendo la dose base calcolata in precedenza, e con variazione di percentuale T:B ratio da 1:1 a 2:1.
Comunque, oltre a rappresentare genericamente una discreta molecola sia in in preparazione alla gara che in Off-Season, I dosaggi utilizzati si settano nel range tra i 200mg ed i 500mg a settimana, spesso abbinato ad una dose variabile (vedi sopra) di Testosterone.
Il Nandrolone, noto anche come 19-nortestosterone, è uno Steroide Androgeno Anabolizzante (AAS) utilizzato sotto forma di molecola legata a esteri come quello Decanoato (nome commerciale Deca-Durabolin) e il Fenilpropionato (nome commerciale Durabolin). Gli esteri del Nandrolone sono utilizzati nel trattamento di anemie, cachessia (sindrome da deperimento), osteoporosi, cancro al seno e per altre indicazioni mediche.
Il Nandrolone è stato sintetizzato per la prima volta nel 1950. È stato introdotto per la prima volta nel mercato farmaceutico, come Nandrolone Fenilpropionato, nel 1959, e poi come Nandrolone Decanoato nel 1962, seguito da ulteriori esteri.
Il Nandrolone ha una bassa affinità di interazione con l’Enzima Aromatasi convertendo in Estrone, un estrogeno molto meno potente dell’Estradiolo, circa 10 volte meno attivo, e, come tale, è un estrogeno relativamente debole. In una condizione di somministrazione del Nandrolone senza una base di Testosterone, i livelli di Estradiolo calerebbero marcatamente a favore di un aumento del Estrone il quale non potrebbe però sostituire nelle diverse attività tissutali il prima citato E2. Le conseguenze negative si verificherebbero dall’attività sessuale all’attività neurosteroidea.
Infatti, un effetto da non sottovalutare con l’uso di Nandrolone è il suo impatto sul SNC. L’impatto del Nandrolone sul Sistema Nervoso Centrale è stato osservato scientificamente. Nello studio intitolato “The Impact of Nandrolone Decanoate on the Central Nervous System” vengono descritti chiaramente i numerosi effetti psicologici di questa molecola. Essi comprendono e influenzano:
1- Aggressività 2- Ansia, paura e stress 3- Ricompensa e dipendenza 4- Apprendimento, memoria e capacità di lavoro 5- Locomozione e attività fisica 6- Effetti sulla HPAA (Asse Ipotalamo-Pituitaria-Surrene) 7- Effetto sui neurotrasmettitori: Recettore Acido γ-Aminobutirrico Tipo A (GABAA); Recettori 5-idrossitriptamina (5-HT) e 5-HT; Recettori della Dopamina e Recettori Oppioidi.
Questo effetto, unito alla modesta potenzialità anabolizzante se confrontata con altre molecole anche della stessa famiglia, fa pendere l’ago della bilancia verso gli svantaggi d’uso piuttosto che i vantaggi. Sebbene vi sia un rapporto tra Testosterone e Nandrolone finalizzato a ridurre la comparsa di questi effetti avversi (ratio T:N = 2:1) su un buon numero di soggetti risulta dare comunque problemi rilevanti.
Il suo uso principale in Off-Season comprende dosaggi medi tra i 200mg ed i 400mg a settimana, con un adeguato rapporto con il Testosterone. Se utilizzato a fini di recupero articolare viene usato a dosaggi di 100mg a settimana, e con tali dosaggi difficilmente emergono i problemi sopra elencati a patto che ci sia una base di Testosterone.
Il Drostanolone, noto anche come 2α-metil-5α-diidrotestosterone (2α-metil-DHT) o come 2α-metil-5α-androstan-17β-ol-3-one, è uno steroide androstano sintetico e un derivato del DHT. Si tratta nello specifico di DHT con un gruppo metile in posizione C2α. La forma esterificata Drostanolone Propionato è stata usata in passato nel trattamento del cancro al seno nelle donne per via della sua attività antiestrogenica. Questa azione il Drostanolone la esplica sia agendo come antagonista del recettore degli estrogeni e sia come inibitore dell’Enzima Aromatasi. Ed è proprio per questo motivo che una molecola generalmente relegata all’uso in “Cut” o pre-gara trova un suo uso funzionale in Off-Season. La sua attività AI è comunque moderata ma sufficiente in un buon numero di soggetti per evitare l’aggiunta di SERM e/o AI di altro genere. L’attività AI moderata sembra non incidere negativamente in modo sensibile sull’Asse GH/IGF1.
L’effetto miotrofico risulta simile a quello osservato con il Methenolone, in generale moderatamente inferiore al Testosterone. I dosaggi utilizzati in Off-Season per il controllo estrogenico sono nel range dei 200-400mg a settimana (diviso in due iniezioni distanziate da 4-5 giorni) per l’estere Enantato, mentre per il Propionato 150-350mg a settimana (dosi a giorni alterni o giornaliere).
Il Trenbolone, noto anche come 19-nor-δ9,11-testosterone o come estra-4,9,11-trien-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone) sintetizzato per la prima volta nel 1963. Si tratta nello specifico di Nandrolone con due doppi legami aggiuntivi nel nucleo steroideo. Gli esteri del Trenbolone, che hanno un estere in posizione C17β, includono il Trenbolone Acetato, il Trenbolone Enantato, Il Trenbolone Hexahydrobenzylcarbonato e il Trenbolone Undecanoato. Il Trenbolone Acetato (marchi Finajet, Finaplix, e altri) e il Trenbolone Hexahydrobenzylcarbonato (marchi Parabolan, Hexabolan), sono o sono stati commercializzati per uso veterinario e clinico nell’uomo. Il Trenbolone Acetato è utilizzato in medicina veterinaria nel bestiame per aumentare la crescita muscolare e l’appetito degli animali, mentre il Trenbolone Hexahydrobenzylcarbonato è stato utilizzato in passato a livello clinico nell’uomo, ma ora non è più commercializzato.
Si tratta di uno degli AAS più versatili in assoluto, con un ottima resa tanto in preparazione alla gara quanto in Off-Season. L’enorme potenziale anabolizzante del Trenbolone, così come dei suoi analoghi, è stato riportato fin dagli anni ’60. La sua diffusione nel Bodybuilding è iniziata circa negli anni ’80 del secolo scorso. La sua elevata potenzialità miotrofica, lipolitica e di spinta mentale lo resero in poco tempo estremamente popolare tra i culturisti.
In Off-Season viene utilizzato nelle sue forme eseterificate Enantato e Hexahydrobenzylcarbonato a dosaggi nell’ordine dei 100-400mg a settimana (divisa in due somministrazioni distanziate l’una dall’altra da 4-5 giorni), sebbene il trend d’oltre oceano è arrivato a dosaggi decisamente eccessivi e nell’ordine del grammo. Per l’esetere Acetato i dosaggi medi vanno da 150mg a 350mg a settimana con dosaggi a giorni alterni o giornalieri.
E’ necessario ricordare ai lettori che gli effetti collaterali a livello del SNC possono verificarsi in alcuni punti come nel caso del Nandrolone sebbene i vantaggi rendano il Trenbolone più bilanciato tra sides e vantaggi.
Il Trestolone, noto anche come 7α-metil-19-nortestosterone (MENT) o come 7α-metilestr-4-en-17β-ol-3-one, è uno steroide sintetico e un derivato del Nandrolone (19-nortestosterone). È una forma modificata del Nandrolone con un gruppo metile in posizione C7α. Tra gli AAS strettamente correlati vi sono il 7α-metil-19-norandrostenedione (MENT dione, trestione), un pro-ormone androgeno del Trestolone, e il Dimetandrolone (7α, 11β-dimetil-19-nortestosterone), il derivato metilato C11β del Trestolone, nonché il Mibolerone (7α,17α-dimetil-19-nortestosterone) e il Dimetiltrienolone (7α,17α-dimetil-δ9,11-19-nortestosterone). Anche il progestinico Tibolone (7α-metil-17α-etinil-δ5(10)-19-nortestosterone) è strettamente correlato al Trestolone.
Il Trestolone è stato descritto per la prima volta nel 1963. Tuttavia, non è stato successivamente studiato fino al 1990. Lo sviluppo del Trestolone per un potenziale uso nella contraccezione ormonale maschile e nella terapia sostitutiva degli androgeni è stato avviato nel 1993 ed è proseguito in seguito. Non sembra che siano stati condotti ulteriori sviluppi dal 2013. Il Trestolone è stato sviluppato dal Population Council, un’organizzazione non governativa senza scopo di lucro dedicata alla salute riproduttiva.
Come AAS, il Trestolone è un agonista del recettore degli androgeni (AR), analogamente agli androgeni come il Testosterone e il Diidrotestosterone (DHT). Questo AAS presenta spiccate proprietà anticortisolemiche sia attraverso l’inibizione enzimatica sia per attività antagonista recettoriale. Il Trestolone non è un substrato per la 5α-reduttasi e quindi non è potenziato o inattivato nei cosiddetti tessuti “androgeni” come la pelle, i follicoli piliferi e la ghiandola prostatica. Come tale, ha un elevato rapporto tra attività anabolica e androgena, analogamente ad altri derivati del Nandrolone. Il Trestolone è un substrato per l’Aromatasi e quindi produce come metabolita l’estrogeno 7α-metilestradiolo. Tuttavia, il Trestolone ha solo una debole attività estrogenica e una quantità che sembrerebbe essere insufficiente per scopi terapici sostitutivi, come evidenziato dalla diminuzione della densità minerale ossea negli uomini trattati con esso per l’ipogonadismo.
Il potenziale anabolizzante del Trestolone ha mostrato un grado di superiorità miotrofica rispetto al Trenbolone. Le sue caratteristiche ne fanno prediligere l’uso in Off-Season/Bulk. I dosaggi utilizzati con la forma Acetato sono nell’ordine dei 150-350mg a settimana con una cadenza nelle somministrazioni a giorni alterni. Sebbene sia più rara da reperire, la forma Enantato è utilizzato nel range dei 200-400mg a settimana divisi in somministrazioni ogni 4-5 giorni.
L’Oxymetholone, noto anche come 2-idrossimetilene-17α-metil-4,5α-diidrotestosterone (2-idrossimetilene-17α-metil-DHT) o come 2-idrossimetilene-17α-metil-5α-androstan-17β-olo-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del DHT. L’Oxymetholone è stato descritto per la prima volta in un articolo del 1959 da scienziati della Syntex. È stato introdotto per uso medico dalla Syntex e dalla Imperial Chemical Industries nel Regno Unito con il marchio Anapolon nel 1961. L’Oxymetholone è stato introdotto anche con i marchi Adroyd (Parke-Davis) nel 1961 e Anadrol (Syntex) nel 1962. Il farmaco è stato commercializzato negli Stati Uniti nei primi anni ’60.
Come altri AAS, l’Oxymetholone è un agonista del recettore degli androgeni (AR). Non è un substrato per la 5α-reduttasi (dal momento che è già 5α-ridotto) ed è uno substrato scarso per il 3α-idrossisteroide deidrogenasi (3α-HSD), e quindi mostra un alto rapporto di attività anabolizzante rispetto all’effetto androgenico.
Data la sua derivanza dal DHT, l’Oxymetholone non è un substrato per l’Enzima Aromatasi e quindi non può essere aromatizzato in metaboliti estrogenici. Tuttavia, caratteristica unica tra i derivati del DHT, l’Oxymetholone è comunque associato a un’estrogenicità relativamente elevata ed è noto per avere il potenziale di produrre effetti collaterali estrogenici come ginecomastia (anche se non comune) e ritenzione idrica. È stato suggerito che questo può essere una conseguenza del legame diretto a l’attivazione del recettore degli estrogeni da parte dell’Oxymetholone (estrogenicità intrinseca). L’Oxymetholone non possiede alcuna attività progestinica significativa. Per via della caratteristica attività estrogenica intrinseca, con l’uso di Oxymetholone è spesso necessario l’uso di un SERM onde avere un controllo sulla aumentata attività estrogenica.
A causa della sua struttura 17α-alchilata, l’Oxymetholone è epatotossico. L’uso a lungo termine del farmaco può causare una varietà di disturbi gravi, tra cui l’epatite, il cancro al fegato e la cirrosi; pertanto si raccomandano test periodici di funzionalità epatica per coloro che assumono l’Oxymetholone a fini terapeutici. Questa molecola ha ottenuto, infatti, la nomea di essere uno tra gli AAS più epatotossici. Ciò deriva da i dosaggi comunemente, ed erroneamente, utilizzati in contesto culturistico. Si parla di dosaggi che facilmente sforano i 150mg/die.
Osservazioni e esaminazione di diversi referti di esami ematici hanno evidenziato una soglia di “vantaggio/svantaggio” a favore del primo con un dosaggio calcolato con la formula 1mg/Kg. Genericamente, però, il dosaggio standard e conservativo si attesta nel range dei 50-100mg/die per non più di 28 giorni consecutivi, al fine di ridurre l’impatto negativo sul fegato e lipidemia.
Il Methandrostenolone, noto anche come 17α-metil-δ1-testosterone o come 17α-metilandrost-1,4-dien-17β-ol-3-one, è uno steroide androstanico sintetico e un derivato 17α-alchilato del Testosterone. È una modifica del Testosterone con un gruppo metile in posizione C17α e un doppio legame aggiuntivo tra le posizioni C1 e C2. Il farmaco è anche il derivato 17α-metilato del Boldenone (δ1-testosterone) e l’analogo δ1 del Methyltestosterone (17α-metiltestosterone).
Il Methandrostenolone è stato descritto per la prima volta nel 1955. È stato sintetizzato dai ricercatori dei laboratori CIBA di Basilea, in Svizzera. La CIBA depositò un brevetto statunitense nel 1957 e iniziò a commercializzare il farmaco sotto il nome di Dianabol nel 1958 negli Stati Uniti. Inizialmente veniva prescritto alle vittime di ustioni e agli anziani. Tra i primi utilizzatori vi furono i giocatori dell’Oklahoma University e l’allenatore dei San Diego Chargers Sid Gillman, che somministrò il Dianabol alla sua squadra a partire dal 1963.
Anche se il primo a somministrare il Methandrostenolone agli atleti fu il Dr. John Ziegler, personaggio che ebbe non poca importanza nella storia dell’uso degli AAS negli Stati Uniti. Ziegler contribuì a facilitare l’adozione degli AAS in generale, e del Dianabol in particolare, da parte degli atleti americani. Ziegler fu la prima persona a somministrare il Dianabol agli atleti competitivi poco dopo la sua introduzione da parte della CIBA nel 1958. Ebbe accesso al laboratorio CIBA a Summit (New Jersey) nel corso degli anni 50’ e somministrava già ai pesisti il Testosterone Propionato per “scopi di ricerca”. Da li il passo fu breve per la diffusione a macchia d’olio di questo AAS tra i culturisti.
Data la sua principale modifica strutturale, ossia la metilazione in C-17, il Methandrostenolone mostra un aumentata stabilità del legame recettoriale aumentando così l’affinità sia al AR sia, successivamente all’aromatizzazione nel suo metabolita 17-Methylestradiolo, per i recettori estrogenici rendendo il composto molto più estrogenico del Testosterone. Tale caratteristiche migliora però il potenziale proliferativo dei AR e l’influenza positiva sulla sintesi di IGF-1. Da non dimenticare è il suo significativo impatto anticortisolemico.
Trattandosi di una molecola con una discreta tendenza all’aromatizzazione, il suo uso tipico la vede inserita nelle fasi Off-Season. Il calcolo del dosaggio, per via dati aneddotici e osservativi raccolti, lo si ottiene con la formula 5mg/12Kg di peso corporeo. Trattandosi di un composto orale metilato in C-17 se ne scoraggia l’utilizzo oltre i 28 giorni consecutivi onde ridurre l’impatto negativo su fegato e lipidemia. Data la sua emivita di circa 4h, il dosaggio giornaliero dovrebbe essere diviso in più assunzioni distribuite durante l’arco della giornata.
L’Ormone della Crescita (GH) o Somatotropina, noto anche come Ormone della Crescita Umano (hGH o HGH), è un ormone peptidico che stimola la crescita, la riproduzione e la rigenerazione cellulare nell’uomo e in altri animali. È quindi importante per lo sviluppo umano. Il GH stimola anche la produzione di IGF-1 e aumenta la concentrazione di glucosio e acidi grassi liberi nel sangue. È un tipo di mitogeno specifico solo per i recettori di alcuni tipi di cellule. Il GH è un polipeptide a catena singola di 191 aminoacidi che viene sintetizzato, immagazzinato e secreto dalle cellule somatotrope nelle ali laterali dell’ipofisi anteriore.
Una forma ricombinante di hGH, chiamata Somatropina, viene utilizzata come farmaco da prescrizione per il trattamento dei disturbi della crescita nei bambini e della carenza di Ormone della Crescita negli adulti. Molte delle funzioni dell’hGH rimangono sconosciute.
Nel suo ruolo di agente anabolizzante, l’hGH è stato utilizzato dagli sportivi agonisti almeno dal 1982, quando la sola forma disponibile era quella derivata dall’Ipofisi dei cadaveri, ed è stato vietato dal CIO e dall’NCAA. L’analisi tradizionale delle urine non è in grado di rilevare il doping con HGH, pertanto il divieto è stato applicato solo all’inizio degli anni 2000, quando sono stati sviluppati test del sangue in grado di distinguere tra hGH naturale e artificiale.
In ambiente bodybuilding, l’hGH viene utilizzato in Off-Season (dai soggetti meglio informati) a dosaggi nel range delle 4-8UI al giorno o 8-16UI a giorni alterni. La somministrazione in concomitanza con l’uso di Insulina ha mostrato effetti sinergici molto evidenti che trovano la loro origine nel miglioramento della sintesi di IGF-1 e della sua frazione libera quindi attiva. Ricordo inoltre che l’uso di hGH può causare una sottoregolazione della funzionalità tiroidea per via del feedback negativo causato da un aumento della conversione del T4 in T3 per azione del GH. L’uso di T4, nel periodo d’uso in Off-Season, è in alcuni casi una necessità.
Il Fattore di Crescita Insulino-Simile 1 (IGF-1), chiamato anche Somatomedina C, è un ormone dalla struttura molecolare simile a quella dell’insulina che svolge un ruolo importante nella crescita infantile e ha effetti anabolici negli adulti. L’IGF-1 è costituito da 70 aminoacidi in una singola catena con tre ponti disolfuro intramolecolari.
L’IGF-1 è prodotto principalmente dal fegato. La produzione è stimolata dall’Ormone della Crescita (GH). La maggior parte dell’IGF-1 è legata a una delle 6 proteine di legame (IGF-BP). L’IGFBP-1 è regolato dall’Insulina. L’IGF-1 viene prodotto durante tutta la vita; i tassi più alti di produzione di IGF-1 si verificano durante la crescita puberale. I livelli più bassi si verificano nell’infanzia e nella vecchiaia.
L’IGF-1 lega e attiva il proprio recettore, l’IGF-1R, attraverso l’espressione sulla superficie cellulare delle tirosin-chinasi recettoriali (RTK) e segnala ulteriormente attraverso molteplici cascate di trasduzione intracellulare. L’IGF-1R è l’induttore che svolge un ruolo critico nella modulazione degli effetti metabolici dell’IGF-1 per la senescenza e la sopravvivenza cellulare. L’IGF-1 è responsabile di stimolare la crescita di tutti i tipi di cellule e di provocare effetti metabolici significativi. Un importante effetto metabolico dell’IGF-1 è la sua capacità di segnalare alle cellule che sono disponibili nutrienti sufficienti per l’ipertrofia e la divisione cellulare. Questi segnali consentono inoltre all’IGF-1 di inibire l’apoptosi cellulare e di aumentare la produzione di proteine cellulari. I recettori dell’IGF-1 sono ubiquitari, il che consente che i cambiamenti metabolici causati dall’IGF-1 si verifichino in tutti i tipi di cellule. Gli effetti metabolici dell’IGF-1 sono di vasta portata e possono coordinare il metabolismo delle proteine, dei carboidrati e dei grassi in una varietà di tipi di cellule diverse. La regolazione degli effetti metabolici dell’IGF-1 sui tessuti bersaglio è coordinata anche con altri ormoni, come l’Ormone della Crescita e l’Insulina.
L’IGF-1 da DNA ricombinante è disponibile principalmente in due diversi formati/varianti, lr3 e DES. È importante ricordare che, a prescindere dalla variante, tutti funzionano a livello sistemico nell’organismo e che, nonostante la somministrazione dell’ormone per via intramuscolare direttamente in un muscolo specifico, non genererà una crescita localizzata misurabile.
Ovviamente tralascerò di descrivere l’IGF-1 bioidentico commercializzato come Mecasermina dal momento che la sua farmacocinetica è identica a quella del IGF-1 endogeno. Dirò soltanto che mediamente viene utilizzato in dosi giornaliere nel range tra 60-1.000mcg post-workout. L’emivita di questa forma di IGF-1 è di circa 5.8h.
IGF-1 LR3: Questa forma è la variante di IGF-1 più comune e molto popolare sul mercato e utilizzata da bodybuilder e atleti di altre discipline. Contiene IGF-1 bioidentico costituito dalla catena originale di 70 aminoacidi, ma con 13 aminoacidi in più all’estremità N, per un totale di 83 aminoacidi. Possiede anche una seconda modifica, in cui un’Arginina si trova in 3a posizione invece dell’Acido Glutammico originale. Il risultato di queste modifiche è che l’IGF-1 continua a svolgere la sua attività originaria sul recettore dell’IGF-1 nei tessuti corporei e ha un’affinità di legame molto bassa per le proteine leganti l’IGF menzionate in precedenza. Inoltre, presenta una vita attiva significativamente più lunga, di circa 20-30 ore, rispetto a quella dell’IGF-1 di 12-15 ore. L’insieme di questi fattori ha dimostrato che l’LR3 ha un’efficacia circa tre volte superiore a quella dell’IGF-1.
I dosaggi medi utilizzati per questa forma sono nel range dei 40-80mcg/die. A causa della sua lunga vita attiva nell’organismo, la variante LR3 non dovrebbe essere somministrata più di una volta al giorno per il semplice fatto che non risulta necessario. Nei giorni di allenamento, il dosaggio di IGF-1 è solitamente somministrato subito dopo l’allenamento. La scelta è a discrezione dell’utilizzatore, in quanto può essere benissimo somministrato sia prima che dopo (solo prima dell’allenamento o solo dopo l’allenamento). E’ possibile comunque dividere il dosaggio giornaliero in due somministrazioni nell’arco della giornata, il dosaggio giornaliero completo può essere diviso quindi a metà tra i due (ad esempio, 20mcg prima dell’allenamento e 20mcg dopo l’allenamento, per un totale di 40mcg al giorno). Nei giorni di non allenamento, può essere somministrato in qualsiasi momento della giornata.
IGF-1 DES: Conosciuto anche come DES(1-3)IGF-1, questa è la forma di IGF-1 comunemente conosciuta come ad azione molto rapida e di solito è la meno preferita tra le due. Le sue modifiche rispetto alla molecola originale di IGF-1 sono tali da farle mancare i primi 3 aminoacidi all’N terminale, il che conferisce all’IGF-1 DES un totale di 67 aminoacidi nella sua catena rispetto ai 70 originali. Questa modifica garantisce all’IGF-1 DES una ridotta affinità di legame per le proteine leganti l’IGF menzionate in precedenza, oltre a una maggiore forza di legame e potenziale miotrofico, circa dieci volte superiore a quella dell’IGF-1 originale e cinque volte superiore a quella dell’IGF-1 LR3. A differenza dell’IGF-1 LR3, l’IGF-1 DES ha un’emivita molto più breve, di circa 20-30 minuti. Grazie alla sua attività più rapida e alla maggiore forza/potenza, la variante DES dell’IGF-1 è comunemente ritenuta in grado di ottenere una crescita muscolare localizzata nel sito in cui viene iniettata. Sebbene ciò sia in parte vero, gli studi hanno dimostrato che, come l’IGF-1 in generale, agisce a livello sistemico una volta raggiunti i capillari e il flusso sanguigno. Quindi l’effetto localizzato è minimo e non significativamente differente dall’effetto sistemico.
Il dosaggio della variante DES è leggermente più variabile rispetto a quello del LR3. Per l’IGF-1 DES, il dosaggio varia da 50 a 150 mcg al giorno. A causa della sua emivita molto più breve rispetto alla variante LR3, è possibile utilizzare dosaggi più elevati con una ipotetica riduzione del rischio di effetti a lungo termine sull’organismo, anche se è necessario usare comunque cautela. Può essere utilizzato nello stesso modo dell’IGF-1 LR3 post-workout, ed è infatti comunemente usato in questo modo a causa della sua breve emivita.
Entrambe le forme di IGF-1 possono essere somministrate per via intramuscolare o sottocutanea. L’uso di una delle due forme non deve superare la durata di 30 giorni prima di una pausa di almeno 2 settimane, anche se fare pause più lunghe di 2 settimane tra un ciclo di IGF-1 e l’altro è l’opzione migliore. Questo non solo per ridurre il rischio di effetti sulla salute a lungo termine, ma anche per garantire che i recettori dell’IGF-1 tornino ad un grado di sensibilità ottimale e, quindi, a “rispondere” correttamente dopo un ciclo.
L’insulina è un ormone peptidico prodotto dalle cellule beta delle isole pancreatiche. Regola il metabolismo dei carboidrati, dei grassi e delle proteine promuovendo l’assorbimento del glucosio dal sangue nelle cellule del fegato, dei grassi e dei muscoli scheletrici. In questi tessuti il glucosio assorbito viene convertito in glicogeno attraverso la glicogenesi o in grassi (trigliceridi) attraverso la lipogenesi o, nel caso del fegato, in entrambi. La produzione e la secrezione di glucosio da parte del fegato sono fortemente inibite da alte concentrazioni di Insulina nel sangue. L’Insulina circolante influisce anche sulla sintesi di proteine in un’ampia varietà di tessuti. È quindi un ormone anabolico e anticatabolico, che promuove la conversione di piccole molecole nel sangue in grandi molecole all’interno delle cellule. Bassi livelli di Insulina nel sangue hanno l’effetto opposto, favorendo un diffuso catabolismo, soprattutto del grasso corporeo di riserva.
La maggior parte dei bodybuilder utilizza una sola forma di Insulina (ad azione rapida o ultra-rapida), anche se alcuni utilizzano anche un’Insulina a lunga durata d’azione o in monoterapia insulinica o in conbinazione con le forme ad azione rapida o ultra-rapida.
L’Humalog® (Insulina Lispro) è senza dubbio la forma di Insulina più diffusa tra i bodybuilder insieme all’Humulin-R. L’Humalog è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”. Proprio al fine di potenziare la “finestra anabolica”, l’Humalog viene usata in concomitanza del GH il quale viene somministrato in una tempistica tale che i due picchi di rilascio (curva ematica massima) si “incrocino” andando a creare un affetto additivo di potenziamento della sintesi epatica di IGF-1 e della sua attività per via della riduzione dei trasportatori IGFBP.
Tuttavia, l’uso di una base insulinica composta da Insuline Glargine (Lantus), con una vita attiva di 24-26.5h, la quale sembra avere effetti di maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi, viene da alcuni inserita nei protocolli Off-Season.
I dosaggi di Insulina non andrebbero calcolati in modo distaccato dal piano alimentare e dal suo contenuto glucidico. Se il margine di “sicurezza” indica un assunzione di 10-15g di Carboidrati per UI di Insulina, questi non dovrebbero essere addizionati al piano alimentare già tarata in surplus calorico. Il calcolo delle unità dovrebbe essere tarato sul quantitativo glucidico della dieta e sul rapporto con il peso corporeo dell’atleta. Facciamo un semplice esempio: Soggetto di 90Kg = formula 1UI ogni 10Kg di peso = 9UI massime somministrabili per pasto e in base alla vita attiva della forma utilizzata = assicurarsi che il pasto appena successivo alla somministrazione dell’Insulina a questo dosaggio sia pari o superiore ai 90g di Carboidrati. Il monitoraggio della glicemia attraverso un glucometro è ovviamente d’obbligo in un protocollo di Insulina.
Nota: tali informazioni esposte non rappresentano in nessun modo un parere medico ne tanto meno una prescrizione e/o incentivo all’uso di sostanze dopanti e illegali. Le descrizioni presentate per i PEDs solitamente più utilizzati in Off-Season sono sintetiche sia per motivi di “Off Topic” sia per ragioni legate alla loro descrizione approfondita in altri articoli presenti nel database di questo sito. In queste pubblicazioni potrete trovare informazioni inerenti anche agli affetti collaterali connessi ad un uso/abuso “off-label” dei diversi PEDs.
Conclusioni:
Per concludere e fare una sintesi delle nozioni esposte in questo articolo, dobbiamo ricordarci che i bodybuilder in Off-Season dovrebbero concentrarsi sul consumo di una dieta leggermente ipercalorica (~10-20% sopra le calorie di mantenimento) con l’obiettivo di guadagnare ~0,25-0,5% del peso corporeo a settimana per un “Natural”, mentre nel caso di un “Doped” la soglia può spostarsi tra l’1-2% con variabili connesse a risposte genetiche differenziali e anzianità nella carriera culturistica (principiante, intermedio e avanzato). In ogni caso, in una fetta maggioritaria di praticanti, ai bodybuilder avanzati si consiglia di essere più prudenti con il surplus calorico e il tasso di aumento di peso settimanale. L’assunzione di proteine nella dieta è raccomandata a 1,6-2,2 g/kg/giorno, con particolare attenzione a una quantità sufficiente di proteine a ogni pasto (0,40-0,55 g/kg/pasto) e a una distribuzione uniforme nell’arco della giornata (3-6 pasti). Per i “Doped”, in alcuni casi, l’introito proteico può essere portato, con minimi vantaggi in contesto ipercalorico, a 2,5g/Kg con le medesime linee guida di suddivisione per numero di pasti. I grassi alimentari devono essere consumati a livelli moderati, né troppo bassi né troppo alti (0,5-1,5 g/kg/die), per evitare un rapporto fTC sfavorevole e per prevenire riduzioni dei livelli di testosterone. Nei “Doped” l’obbiettivo con i lipidi è principalmente quello di assumerne una dose necessaria, e altamente qualitativa, al fine di assimilare vitamine liposolubili, per substrato strutturale, per sintesi di eicosanoidi (vedi assunzione EPA, DHA e AA), protezioni epidermide e capelli; di conseguenza attenersi ad un dosaggio medio pari a 35-50g/die. Dopo che le calorie sono state distribuite tra Proteine e Grassi, le restanti calorie dovrebbero provenire dai Carboidrati, assicurandosi di consumarne una quantità sufficiente (≥3-5 g/kg/giorno). Si possono ottenere benefici maggiori consumando proteine (0,40-0,55 g/kg/pasto) in prossimità delle sessioni di allenamento (1-2 ore prima dell’esercizio ed entro 1-2 ore dopo l’esercizio). È opportuno prendere in considerazione la Creatina Monoidrato (3-5 g/giorno) e la Caffeina (5-6 mg/kg), in quanto possono produrre effetti ergogenici per i bodybuilder. Inoltre, Beta-Alanina (3-5 g/die) e Citrullina Malato (8 g/die) sono integratori alimentari che possono essere presi in considerazione in quanto potenzialmente utili per i bodybuilder, a seconda dei regimi di allenamento individuali. I bodybuilder che non sono in grado di assumere un apporto sufficiente di micronutrienti e acidi grassi essenziali nella loro dieta dovrebbero prendere in considerazione l’integrazione di questi nutrienti per evitare carenze. Il limite principale di questo articolo è la mancanza di studi su larga scala e a lungo termine sui bodybuilder durante la Off-Season. Sono necessarie ulteriori ricerche su questa popolazione per ottimizzare la nutrizione e le raccomandazioni sugli integratori alimentari.
4. Philen R.M., Ortiz D.I., Auerbach S.B., Falk H. Survey of Advertising for Nutritional Supplements in Health and Bodybuilding Magazines. JAMA. 1992;268:1008. doi: 10.1001/jama.1992.03490080082029. [PubMed] [CrossRef] [Google Scholar]
5. Giampreti A., Lonati D., Locatelli C., Rocchi L., Campailla M.T. Acute neurotoxicity after yohimbine ingestion by a bodybuilder. [(accessed on 25 March 2019)];Clin. Toxicol. 2009 47:827–829. doi: 10.1080/15563650903081601. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19640235 [PubMed] [CrossRef] [Google Scholar]
7. Della Guardia L., Cavallaro M., Cena H. The risks of self-made diets: The case of an amateur bodybuilder. J. Int. Soc. Sports Nutr. 2015;12:5. doi: 10.1186/s12970-015-0077-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Hackett D.A., Johnson N.A., Chow C.-M. Training Practices and Ergogenic Aids Used by Male Bodybuilders. J. Strength Cond. Res. 2013;27:1609–1617. doi: 10.1519/JSC.0b013e318271272a. [PubMed] [CrossRef] [Google Scholar]
10. Forbes G.B., Brown M.R., Welle S.L., Lipinski B.A. Deliberate overfeeding in women and men: Energy cost and composition of the weight gain. Br. J. Nutr. 1986;56:1–9. doi: 10.1079/BJN19860080. [PubMed] [CrossRef] [Google Scholar]
11. Kreider R.B., Klesges R., Harmon K., Ramsey L., Bullen D., Wood L., Almada A., Grindstaff P., Li Y. Effects of Ingesting Supplements Designed to Promote Lean Tissue Accretion on Body Composition during Resistance Training. Int. J. Sport Nutr. 1996;6:234–246. doi: 10.1123/ijsn.6.3.234. [PubMed] [CrossRef] [Google Scholar]
12. Rozenek R., Ward P., Long S., Garhammer J. Effects of high-calorie supplements on body composition and muscular strength following resistance training. J. Sports Med. Phys. Fit. 2002;42:340–347. [PubMed] [Google Scholar]
13. Garthe I., Raastad T., Refsnes P.E., Sundgot-Borgen J. Effect of nutritional intervention on body composition and performance in elite athletes. Eur. J. Sport Sci. 2013;13:295–303. doi: 10.1080/17461391.2011.643923. [PubMed] [CrossRef] [Google Scholar]
14. American College og Sports Medicine American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. [(accessed on 25 March 2019)];Med. Sci. Sport. Exerc. 2009 41:687–708. doi: 10.1249/MSS.0b013e3181915670. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19204579 [PubMed] [CrossRef] [Google Scholar]
15. Lambert C.P., Frank L.L., Evans W.J., Lambert D.C.P. Macronutrient Considerations for the Sport of Bodybuilding. Sports Med. 2004;34:317–327. doi: 10.2165/00007256-200434050-00004. [PubMed] [CrossRef] [Google Scholar]
16. Walberg-Rankin J., Edmonds C.E., Gwazdauskas F.C. Diet and Weight Changes of Female Bodybuilders Before and After Competition. Int. J. Sport Nutr. 1993;3:87–102. doi: 10.1123/ijsn.3.1.87. [PubMed] [CrossRef] [Google Scholar]
17. Lamar-Hildebrand N., Saldanha L., Endres J. Dietary and exercise practices of college-aged female bodybuilders. J. Am. Diet. Assoc. 1989;89:1308–1310. [PubMed] [Google Scholar]
18. Houston M.E. Gaining Weight: The Scientific Basis of Increasing Skeletal Muscle Mass. Can. J. Appl. Physiol. 1999;24:305–316. doi: 10.1139/h99-024. [PubMed] [CrossRef] [Google Scholar]
19. Phillips S.M. A Brief Review of Critical Processes in Exercise-Induced Muscular Hypertrophy. Sports Med. 2014;44:71–77. doi: 10.1007/s40279-014-0152-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Campbell B.I., Aguilar D., Conlin L., Vargas A., Schoenfeld B.J., Corson A., Gai C., Best S., Galvan E., Couvillion K. Effects of High Versus Low Protein Intake on Body Composition and Maximal Strength in Aspiring Female Physique Athletes Engaging in an 8-Week Resistance Training Program. Int. J. Sport Nutr. Exerc. Metab. 2018;28:580–585. doi: 10.1123/ijsnem.2017-0389. [PubMed] [CrossRef] [Google Scholar]
22. Morton R.W., Murphy K.T., McKellar S.E., Schoenfeld B.J., Henselmans M., Helms E., Aragon A.A., Devries M.C., Banfield L., Krieger J.W., et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. [(accessed on 25 March 2019)];Br. J. Sports Med. 2018 52:376–384. doi: 10.1136/bjsports-2017-097608. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28698222 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Houltham S.D., Rowlands D.S. A snapshot of nitrogen balance in endurance-trained women. Appl. Physiol. Nutr. Metab. 2014;39:219–225. doi: 10.1139/apnm-2013-0182. [PubMed] [CrossRef] [Google Scholar]
25. Antonio J., Ellerbroek A., Silver T., Vargas L., Peacock C. The effects of a high protein diet on indices of health and body composition—A crossover trial in resistance-trained men. J. Int. Soc. Sports Nutr. 2016;13:8. doi: 10.1186/s12970-016-0114-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Bandegan A., Courtney-Martin G., Rafii M., Pencharz P.B., Lemon P.W. Indicator Amino Acid–Derived Estimate of Dietary Protein Requirement for Male Bodybuilders on a Nontraining Day Is Several-Fold Greater than the Current Recommended Dietary Allowance. J. Nutr. 2017;147:850–857. doi: 10.3945/jn.116.236331. [PubMed] [CrossRef] [Google Scholar]
27. Malowany J.M., West D.W.D., Williamson E., Volterman K.A., Sawan S.A., Mazzulla M., Moore D.R. Protein to Maximize Whole-Body Anabolism in Resistance-trained Females after Exercise. Med. Sci. Sports Exerc. 2019;51:798–804. doi: 10.1249/MSS.0000000000001832. [PubMed] [CrossRef] [Google Scholar]
28. Antonio J., Peacock C.A., Ellerbroek A., Fromhoff B., Silver T. The effects of consuming a high protein diet (4.4 g/kg/d) on body composition in resistance-trained individuals. J. Int. Soc. Sports Nutr. 2014;11:19. doi: 10.1186/1550-2783-11-19. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
29. Antonio J., Ellerbroek A., Silver T., Orris S., Scheiner M., Gonzalez A., Peacock C.A. A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women—A follow-up investigation. J. Int. Soc. Sports Nutr. 2015;12:39. doi: 10.1186/s12970-015-0100-0. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Bray G.A., Smith S.R., de Jonge L., Xie H., Rood J., Martin C.K., Most M., Brock C., Mancuso S., Redman L.M. Effect of dietary protein content on weight gain, energy expenditure, and body composition during overeating: A randomized controlled trial. [(accessed on 25 March 2019)];JAMA. 2012 307:47–55. doi: 10.1001/jama.2011.1918. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22215165 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Tipton K.D., Ferrando A.A., Phillips S.M., Doyle D., Wolfe R.R. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am. J. Physiol. Metab. 1999;276:628–634. doi: 10.1152/ajpendo.1999.276.4.E628. [PubMed] [CrossRef] [Google Scholar]
32. Rieu I., Balage M., Sornet C., Giraudet C., Pujos E., Grizard J., Mosoni L., Dardevet D. Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia. J. Physiol. 2006;575:305–315. doi: 10.1113/jphysiol.2006.110742. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Burd N.A., Tang J.E., Moore D.R., Phillips S.M. Exercise training and protein metabolism: Influences of contraction, protein intake, and sex-based differences. [(accessed on 25 March 2019)];J. Appl. Physiol. 2008 106:1692–1701. doi: 10.1152/japplphysiol.91351.2008. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19036897 [PubMed] [CrossRef] [Google Scholar]
34. Drummond M.J., Dreyer H.C., Fry C.S., Glynn E.L., Rasmussen B.B. Nutritional and contractile regulation of human skeletal muscle protein synthesis and mTORC1 signaling. J. Appl. Physiol. 2009;106:1374–1384. doi: 10.1152/japplphysiol.91397.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Tang J.E., Moore D.R., Kujbida G.W., Tarnopolsky M.A., Phillips S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009;107:987–992. doi: 10.1152/japplphysiol.00076.2009. [PubMed] [CrossRef] [Google Scholar]
36. Kanda A., Nakayama K., Sanbongi C., Nagata M., Ikegami S., Itoh H. Effects of Whey, Caseinate, or Milk Protein Ingestion on Muscle Protein Synthesis after Exercise. Nutrients. 2016;8:339. doi: 10.3390/nu8060339. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Messina M., Lynch H., Dickinson J.M., Reed K.E. No Difference Between the Effects of Supplementing With Soy Protein Versus Animal Protein on Gains in Muscle Mass and Strength in Response to Resistance Exercise. Int. J. Sport Nutr. Exerc. Metab. 2018;28:674–685. doi: 10.1123/ijsnem.2018-0071. [PubMed] [CrossRef] [Google Scholar]
38. Joy J.M., Lowery R.P., Wilson J.M., Purpura M., De Souza E.O., Mc Wilson S., Kalman D.S., Dudeck J.E., Jäger R. The effects of 8 weeks of whey or rice protein supplementation on body composition and exercise performance. Nutr. J. 2013;12:86. doi: 10.1186/1475-2891-12-86. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
39. Babault N., Paizis C., Deley G., Guérin-Deremaux L., Saniez M.-H., Lefranc-Millot C., Allaert F.A. Pea proteins oral supplementation promotes muscle thickness gains during resistance training: A double-blind, randomized, Placebo-controlled clinical trial vs. Whey protein. J. Int. Soc. Sports Nutr. 2015;12:1692. doi: 10.1186/s12970-014-0064-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Tesch P.A. Glycogen and triglyceride utilization in relation to muscle metabolic characteristics in men performing heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1990;61:5–10. [PubMed] [Google Scholar]
41. Lane A.R., Duke J.W., Hackney A.C. Influence of dietary carbohydrate intake on the free testosterone: Cortisol ratio responses to short-term intensive exercise training. [(accessed on 25 March 2019)];Eur. J. Appl. Physiol. 2010 108:1125–1131. doi: 10.1007/s00421-009-1220-5. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20091182 [PubMed] [CrossRef] [Google Scholar]
42. Tegelman R., Aberg T., Pousette A., Carlström K. Effects of a diet regimen on pituitary and steroid hormones in male ice hockey players. [(accessed on 25 March 2019)];Int. J. Sports Med. 1992 13:420–430. doi: 10.1055/s-2007-1021292. Available online: https://www.ncbi.nlm.nih.gov/pubmed/1387870 [PubMed] [CrossRef] [Google Scholar]
43. Dorgan J.F., Judd J.T., Longcope C., Brown C., Schatzkin A., Clevidence B.A., Campbell W.S., Nair P.P., Franz C., Kahle L., et al. Effects of dietary fat and fiber on plasma and urine androgens and estrogens in men: A controlled feeding study. Am. J. Clin. Nutr. 1996;64:850–855. doi: 10.1093/ajcn/64.6.850. [PubMed] [CrossRef] [Google Scholar]
44. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Decrease of serum total and free testosterone during a low-fat high-fibre diet. J. Steroid Biochem. 1983;18:369–370. doi: 10.1016/0022-4731(83)90117-6. [PubMed] [CrossRef] [Google Scholar]
45. Hämäläinen E., Adlercreutz H., Puska P., Pietinen P. Diet and serum sex hormones in healthy men. J. Steroid Biochem. 1984;20:459–464. doi: 10.1016/0022-4731(84)90254-1. [PubMed] [CrossRef] [Google Scholar]
46. Wang C., Catlin D.H., Starcevic B., Heber D., Ambler C., Berman N., Lucas G., Leung A., Schramm K., Lee P.W.N., et al. Low-Fat High-Fiber Diet Decreased Serum and Urine Androgens in Men. J. Clin. Endocrinol. Metab. 2005;90:3550–3559. doi: 10.1210/jc.2004-1530. [PubMed] [CrossRef] [Google Scholar]
47. Morton R.W., Sato K., Gallaugher M.P.B., Oikawa S.Y., McNicholas P.D., Fujita S., Phillips S.M. Muscle Androgen Receptor Content but Not Systemic Hormones Is Associated With Resistance Training-Induced Skeletal Muscle Hypertrophy in Healthy, Young Men. Front. Physiol. 2018;9:9. doi: 10.3389/fphys.2018.01373. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Tinsley G.M., Willoughby D.S. Fat-Free Mass Changes During Ketogenic Diets and the Potential Role of Resistance Training. Int. J. Sport Nutr. Exerc. Metab. 2016;26:78–92. doi: 10.1123/ijsnem.2015-0070. [PubMed] [CrossRef] [Google Scholar]
49. Vargas S., Romance R., Petro J.L., Bonilla D.A., Galancho I., Espinar S., Kreider R.B., Benítez-Porres J. Efficacy of ketogenic diet on body composition during resistance training in trained men: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2018;15:31. doi: 10.1186/s12970-018-0236-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
50. Kephart W.C., Pledge C.D., Roberson P.A., Mumford P.W., Romero M.A., Mobley C.B., Martin J.S., Young K.C., Lowery R.P., Wilson J.M., et al. The Three-Month Effects of a Ketogenic Diet on Body Composition, Blood Parameters, and Performance Metrics in CrossFit Trainees: A Pilot Study. Sports. 2018;6:1. doi: 10.3390/sports6010001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
51. Greene D.A., Varley B.J., Hartwig T.B., Chapman P., Rigney M. A Low-Carbohydrate Ketogenic Diet Reduces Body Mass Without Compromising Performance in Powerlifting and Olympic Weightlifting Athletes. [(accessed on 26 March 2019)];J. Strength Cond. Res. 2018 32:3373–3382. Available online: https://www.ncbi.nlm.nih.gov/pubmed/30335720 [PubMed] [Google Scholar]
52. Bird S. Strength Nutrition: Maximizing Your Anabolic Potential. Strength Cond. J. 2010;32:80–86. doi: 10.1519/SSC.0b013e3181d5284e. [CrossRef] [Google Scholar]
53. American Dietetic Association. Dietitians of Canada. American College of Sports Medicine. Rodriguez N.R., Di Marco N.M., Langley S. American College of Sports Medicine position stand. Nutrition and athletic performance. [(accessed on 26 March 2019)];Med. Sci. Sports Exerc. 2009 41:709–731. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19225360 [PubMed] [Google Scholar]
54. Chung S.T., Chacko S.K., Sunehag A.L., Haymond M.W. Measurements of Gluconeogenesis and Glycogenolysis: A Methodological Review. Diabetes. 2015;64:3996–4010. doi: 10.2337/db15-0640. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Azizi F. Effect of dietary composition on fasting-induced changes in serum thyroid hormones and thyrotropin. Metabolism. 1978;27:935–942. doi: 10.1016/0026-0495(78)90137-3. [PubMed] [CrossRef] [Google Scholar]
56. Mathieson R.A., Walberg J.L., Gwazdauskas F.C., Hinkle D.E., Gregg J.M. The effect of varying carbohydrate content of a very-low-caloric diet on resting metabolic rate and thyroid hormones. Metabolism. 1986;35:394–398. doi: 10.1016/0026-0495(86)90126-5. [PubMed] [CrossRef] [Google Scholar]
57. Leveritt M., Abernethy P.J. Effects of Carbohydrate Restriction on Strength Performance. J. Strength Cond. Res. 1999;13:52–57. [Google Scholar]
58. Jacobs I., Kaiser P., Tesch P. Muscle strength and fatigue after selective glycogen depletion in human skeletal muscle fibers. Graefe’s Arch. Clin. Exp. Ophthalmol. 1981;46:47–53. doi: 10.1007/BF00422176. [PubMed] [CrossRef] [Google Scholar]
59. Ray S., Sale D.G., Lee P., Garner S., MacDougall J.D., McCartney N. Muscle Substrate Utilization and Lactate Production During Weightlifting. Can. J. Appl. Physiol. 1999;24:209–215. [PubMed] [Google Scholar]
60. Tesch P.A., Colliander E.B., Kaiser P. Muscle metabolism during intense, heavy-resistance exercise. Graefe’s Arch. Clin. Exp. Ophthalmol. 1986;55:362–366. doi: 10.1007/BF00422734. [PubMed] [CrossRef] [Google Scholar]
63. Mitchell J.B., DiLauro P.C., Pizza F.X., Cavender D.L. The Effect of Preexercise Carbohydrate Status on Resistance Exercise Performance. Int. J. Sport Nutr. 1997;7:185–196. doi: 10.1123/ijsn.7.3.185. [PubMed] [CrossRef] [Google Scholar]
64. Lima-Silva A.E., Silva-Cavalcante M.D., Oliveira R.S., Kiss M.A., Pires F.O., Bertuzzi R., Bishop D. Effects of a low- or a high-carbohydrate diet on performance, energy system contribution, and metabolic responses during supramaximal exercise. Appl. Physiol. Nutr. Metab. 2013;38:928–934. doi: 10.1139/apnm-2012-0467. [PubMed] [CrossRef] [Google Scholar]
65. Vega F., Jackson R. Dietary habits of bodybuilders and other regular exercisers. Nutr. Res. 1996;16:3–10. doi: 10.1016/0271-5317(95)02054-3. [CrossRef] [Google Scholar]
66. Chappell A.J., Simper T., Barker M.E. Nutritional strategies of high level natural bodybuilders during competition preparation. J. Int. Soc. Sports Nutr. 2018;15:4. doi: 10.1186/s12970-018-0209-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
67. Atherton P.J., Etheridge T., Watt P.W., Wilkinson D., Selby A., Rankin D., Smith K., Rennie M.J. Muscle full effect after oral protein: Time-dependent concordance and discordance between human muscle protein synthesis and mTORC1 signaling. Am. J. Clin. Nutr. 2010;92:1080–1088. doi: 10.3945/ajcn.2010.29819. [PubMed] [CrossRef] [Google Scholar]
68. Res P.T., Groen B., Pennings B., Beelen M., Wallis G.A., Gijsen A.P., Senden J.M., Van Loon L.J. Protein ingestion before sleep improves postexercise overnight recovery. [(accessed on 25 March 2019)];Med. Sci. Sports Exerc. 2012 44:1560–1569. doi: 10.1249/MSS.0b013e31824cc363. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22330017 [PubMed] [CrossRef] [Google Scholar]
69. Moore D.R., Robinson M.J., Fry J.L., Tang J.E., Glover E.I., Wilkinson S.B., Prior T., Tarnopolsky M.A., Phillips S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2009 89:161–168. doi: 10.3945/ajcn.2008.26401. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19056590 [PubMed] [CrossRef] [Google Scholar]
70. Witard O.C., Jackman S.R., Breen L., Smith K., Selby A., Tipton K.D. Muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after. [(accessed on 25 March 2019)];Am. J. Clin. Nutr. 2014 99:86–95. doi: 10.3945/ajcn.112.055517. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24257722 [PubMed] [CrossRef] [Google Scholar]
71. Macnaughton L.S., Wardle S.L., Witard O.C., McGlory C., Hamilton D.L., Jeromson S., Lawrence C.E., Wallis G.A., Tipton K.D. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol. Rep. 2016;4:e12893. doi: 10.14814/phy2.12893. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
72. Schoenfeld B.J., Aragon A.A., Krieger J.W. The effect of protein timing on muscle strength and hypertrophy: A meta-analysis. J. Int. Soc. Sports Nutr. 2013;10:53. doi: 10.1186/1550-2783-10-53. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
73. Areta J.L., Burke L.M., Ross M.L., Camera D.M., West D.W.D., Broad E.M., Jeacocke N.A., Moore D.R., Stellingwerff T., Phillips S.M., et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J. Physiol. 2013;591:2319–2331. doi: 10.1113/jphysiol.2012.244897. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Hudson J.L., Bergia R.E., Campbell W.W. Effects of protein supplements consumed with meals, versus between meals, on resistance training–induced body composition changes in adults: A systematic review. Nutr. Rev. 2018;76:461–468. doi: 10.1093/nutrit/nuy012. [PubMed] [CrossRef] [Google Scholar]
75. Trommelen J., Kouw I.W.K., Holwerda A.M., Snijders T., Halson S.L., Rollo I., Verdijk L.B., Van Loon L.J.C. Pre-sleep dietary protein-derived amino acids are incorporated in myofibrillar protein during post-exercise overnight recovery. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2018 1:457–467. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28536184 [Google Scholar]
76. Kouw I.W., Holwerda A.M., Trommelen J., Kramer I.F., Bastiaanse J., Halson S.L., Wodzig W.K., Verdijk L.B., Van Loon L.J. Protein Ingestion before Sleep Increases Overnight Muscle Protein Synthesis Rates in Healthy Older Men: A Randomized Controlled Trial. [(accessed on 25 March 2019)];J. Nutr. 2017 147:2252–2261. doi: 10.3945/jn.117.254532. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28855419 [PubMed] [CrossRef] [Google Scholar]
77. Snijders T., Res P.T., Smeets J.S., Van Vliet S., Van Kranenburg J., Maase K., Kies A.K., Verdijk L.B., Van Loon L.J. Protein ingestion before sleep increases muscle mass and strength gains during prolonged resistance-type exercise training in healthy young men. [(accessed on 25 March 2019)];J. Nutr. 2015 145:1178–1184. doi: 10.3945/jn.114.208371. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25926415 [PubMed] [CrossRef] [Google Scholar]
78. Joy J.M., Vogel R.M., Broughton K.S., Kudla U., Kerr N.Y., Davison J.M., Wildman R.E.C., DiMarco N.M. Daytime and nighttime casein supplements similarly increase muscle size and strength in response to resistance training earlier in the day: A preliminary investigation. J. Int. Soc. Sports Nutr. 2018;15:24. doi: 10.1186/s12970-018-0228-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Antonio J., Ellerbroek A., Peacock C., Silver T. Casein Protein Supplementation in Trained Men and Women: Morning versus Evening. Int. J. Exerc. Sci. 2017;10:479–486. [PMC free article] [PubMed] [Google Scholar]
80. Schoenfeld B.J., Aragon A.A. How much protein can the body use in a single meal for muscle-building? Implications for daily protein distribution. J. Int. Soc. Sports Nutr. 2018;15:10. doi: 10.1186/s12970-018-0215-1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
81. Pennings B., Groen B.B., Van Dijk J.-W., De Lange A., Kiskini A., Kuklinski M., Senden J.M., Van Loon L.J. Minced beef is more rapidly digested and absorbed than beef steak, resulting in greater postprandial protein retention in older men. Am. J. Clin. Nutr. 2013;98:121–128. doi: 10.3945/ajcn.112.051201. [PubMed] [CrossRef] [Google Scholar]
82. Kim I.Y., Schutzler S., Schrader A., Spencer H.J., Azhar G., Ferrando A.A., Wolfe R.R. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. [(accessed on 25 March 2019)];Am. J. Physiol. Metab. 2016 310:73–80. doi: 10.1152/ajpendo.00365.2015. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26530155 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
83. Jentjens R., Jeukendrup A.E. Determinants of Post-Exercise Glycogen Synthesis During Short-Term Recovery. Sports Med. 2003;33:117–144. doi: 10.2165/00007256-200333020-00004. [PubMed] [CrossRef] [Google Scholar]
84. Biolo G., Williams B.D., Fleming R.Y., Wolfe R.R. Insulin action on muscle protein kinetics and amino acid transport during recovery after resistance exercise. Diabetes. 1999;48:949–957. doi: 10.2337/diabetes.48.5.949. [PubMed] [CrossRef] [Google Scholar]
85. Greenhaff P.L., Karagounis L.G., Peirce N., Simpson E.J., Hazell M., Layfield R., Wackerhage H., Smith K., Atherton P., Selby A., et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am. J. Physiol. Metab. 2008;295:E595–E604. doi: 10.1152/ajpendo.90411.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
86. Glynn E.L., Fry C.S., Timmerman K.L., Drummond M.J., Volpi E., Rasmussen B.B., Leroy J.L., Gadsden P., De Cossío T.G., Gertler P. Addition of Carbohydrate or Alanine to an Essential Amino Acid Mixture Does Not Enhance Human Skeletal Muscle Protein Anabolism123. J. Nutr. 2013;143:307–314. doi: 10.3945/jn.112.168203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Koopman R., Beelen M., Stellingwerff T., Pennings B., Saris W.H.M., Kies A.K., Kuipers H., Van Loon L.J.C. Coingestion of carbohydrate with protein does not further augment postexercise muscle protein synthesis. Am. J. Physiol. Metab. 2007;293:E833–E842. doi: 10.1152/ajpendo.00135.2007. [PubMed] [CrossRef] [Google Scholar]
88. Aragon A.A., Schoenfeld B.J. Nutrient timing revisited: Is there a post-exercise anabolic window? J. Int. Soc. Sports Nutr. 2013;10:5. doi: 10.1186/1550-2783-10-5. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
89. Jäger R., Kerksick C.M., Campbell B.I., Cribb P.J., Wells S.D., Skwiat T.M., Purpura M., Ziegenfuss T.N., Ferrando A.A., Arent S.M., et al. International Society of Sports Nutrition position stand: Protein and exercise. [(accessed on 25 March 2019)];J. Int. Soc. Sport. Nutr. 2017 4:20. doi: 10.1186/s12970-017-0177-8. Available online: https://www.ncbi.nlm.nih.gov/pubmed/28642676 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Darrabie M.D., Arciniegas A.J.L., Mishra R., Bowles D.E., Jacobs D.O., Santacruz L. AMPK and substrate availability regulate creatine transport in cultured cardiomyocytes. Am. J. Physiol. Metab. 2011;300:870–876. doi: 10.1152/ajpendo.00554.2010. [PubMed] [CrossRef] [Google Scholar]
91. Purchas R., Busboom J., Wilkinson B. Changes in the forms of iron and in concentrations of taurine, carnosine, coenzyme Q10, and creatine in beef longissimus muscle with cooking and simulated stomach and duodenal digestion. Meat Sci. 2006;74:443–449. doi: 10.1016/j.meatsci.2006.03.015. [PubMed] [CrossRef] [Google Scholar]
92. Branch J.D. Effect of Creatine Supplementation on Body Composition and Performance: A Meta-analysis. Int. J. Sport Nutr. Exerc. Metab. 2003;13:198–226. doi: 10.1123/ijsnem.13.2.198. [PubMed] [CrossRef] [Google Scholar]
93. Hultman E., Söderlund K., Timmons J.A., Cederblad G., Greenhaff P.L. Muscle creatine loading in men. [(accessed on 25 March 2019)];J. Appl. Physiol. Soc. 1996 81:232–237. doi: 10.1152/jappl.1996.81.1.232. Available online: https://www.ncbi.nlm.nih.gov/pubmed/8828669 [PubMed] [CrossRef] [Google Scholar]
94. Jagim A.R., Oliver J.M., Sanchez A., Galvan E., Fluckey J., Riechman S., Greenwood M., Kelly K., Meininger C., Rasmussen C., et al. A buffered form of creatine does not promote greater changes in muscle creatine content, body composition, or training adaptations than creatine monohydrate. J. Int. Soc. Sports Nutr. 2012;9:43. doi: 10.1186/1550-2783-9-43. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
95. Spillane M., Schoch R., Cooke M., Harvey T., Greenwood M., Kreider R., Willoughby D.S., Cooke M. The effects of creatine ethyl ester supplementation combined with heavy resistance training on body composition, muscle performance, and serum and muscle creatine levels. J. Int. Soc. Sports Nutr. 2009;6:6. doi: 10.1186/1550-2783-6-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Childs E., De Wit H., Wit H. Subjective, behavioral, and physiological effects of acute caffeine in light, nondependent caffeine users. Psychopharmacology. 2006;185:514–523. doi: 10.1007/s00213-006-0341-3. [PubMed] [CrossRef] [Google Scholar]
97. Bellar D., Kamimori G.H., Glickman E.L. The Effects of Low-Dose Caffeine on Perceived Pain During a Grip to Exhaustion Task. J. Strength Cond. Res. 2011;25:1225–1228. doi: 10.1519/JSC.0b013e3181d9901f. [PubMed] [CrossRef] [Google Scholar]
98. Davis J.K., Green J.M. Caffeine and anaerobic performance: Ergogenic value and mechanisms of action. [(accessed on 25 March 2019)];Sport. Med. 2009 39:813–832. doi: 10.2165/11317770-000000000-00000. Available online: https://www.ncbi.nlm.nih.gov/pubmed/19757860 [PubMed] [CrossRef] [Google Scholar]
99. Wickwire P.J., McLester J.R., Gendle S., Hudson G., Pritchett R.C., Laurent C.M., Green J.M. Effects of Caffeine on Repetitions to Failure and Ratings of Perceived Exertion during Resistance Training. Int. J. Sports Physiol. Perform. 2007;2:250–259. [PubMed] [Google Scholar]
100. Duncan M.J., Oxford S.W. The effect of caffeine ingestion on mood state and bench press performance to failure. [(accessed on 25 March 2019)];J. Strength Cond. Res. 2001 25:178–185. doi: 10.1519/JSC.0b013e318201bddb. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22124354 [PubMed] [CrossRef] [Google Scholar]
101. Williams A.D., Cribb P.J., Cooke M.B., Hayes A. The Effect of Ephedra and Caffeine on Maximal Strength and Power in Resistance-Trained Athletes. J. Strength Cond. Res. 2008;22:464–470. doi: 10.1519/JSC.0b013e3181660320. [PubMed] [CrossRef] [Google Scholar]
102. Tarnopolsky M.A., Atkinson S.A., MacDougall J.D., Sale D.G., Sutton J.R. Physiological responses to caffeine during endurance running in habitual caffeine users. Med. Sci. Sports Exerc. 1989;21:418–424. doi: 10.1249/00005768-198908000-00013. [PubMed] [CrossRef] [Google Scholar]
103. Blanchard J., Sawers S.J.A. The absolute bioavailability of caffeine in man. Eur. J. Clin. Pharmacol. 1983;24:93–98. doi: 10.1007/BF00613933. [PubMed] [CrossRef] [Google Scholar]
104. Hobson R.M., Saunders B., Ball G., Harris R.C., Sale C. Effects of β-alanine supplementation on exercise performance: A meta-analysis. Amino Acids. 2012;43:25–37. doi: 10.1007/s00726-011-1200-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
105. Hoffman J., Ratamess N.A., Ross R., Kang J., Magrelli J., Neese K., Faigenbaum A.D., Wise J.A. Beta-alanine and the hormonal response to exercise. [(accessed on 25 March 2019)];Int. J. Sports Med. 2008 29:952–958. doi: 10.1055/s-2008-1038678. Available online: https://www.ncbi.nlm.nih.gov/pubmed/18548362 [PubMed] [CrossRef] [Google Scholar]
106. Hoffman J., Ratamess N., Kang J., Mangine G., Faigenbaum A., Stout J. Effect of creatine and β-alanine supplementation on performance and endocrine responses in strength/power athletes. [(accessed on 25 March 2019)];Int. J. Sport Nutr. Exerc. Metab. 2006 16:430–446. doi: 10.1123/ijsnem.16.4.430. Available online: https://www.ncbi.nlm.nih.gov/pubmed/17136944 [PubMed] [CrossRef] [Google Scholar]
108. Wax B., Kavazis A.N., Weldon K., Sperlak J. Effects of Supplemental Citrulline Malate Ingestion During Repeated Bouts of Lower-Body Exercise in Advanced Weightlifters. J. Strength Cond. Res. 2015;29:786–792. doi: 10.1519/JSC.0000000000000670. [PubMed] [CrossRef] [Google Scholar]
109. Wax B., Kavazis A.N., Luckett W. Effects of Supplemental Citrulline-Malate Ingestion on Blood Lactate, Cardiovascular Dynamics and Resistance Exercise Performance in Trained Males. [(accessed on 25 March 2019)];J. Diet. 2016 13:269–282. doi: 10.3109/19390211.2015.1008615. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25674699 [PubMed] [CrossRef] [Google Scholar]
110. Glenn J.M., Gray M., Wethington L.N., Stone M.S., Stewart R.W., Jr., Moyen N.E. Acute citrulline malate supplementation improves upper- and lower-body submaximal weightlifting exercise performance in resistance-trained females. [(accessed on 25 March 2019)];Eur. J. Nutr. 2017 56:775–784. doi: 10.1007/s00394-015-1124-6. Available online: https://www.ncbi.nlm.nih.gov/pubmed/26658899 [PubMed] [CrossRef] [Google Scholar]
111. Glenn J.M., Gray M., Jensen A., Stone M.S., Vincenzo J.L. Acute citrulline-malate supplementation improves maximal strength and anaerobic power in female, masters athletes tennis players. Eur. J. Sport Sci. 2016;16:1–9. doi: 10.1080/17461391.2016.1158321. [PubMed] [CrossRef] [Google Scholar]
112. Gonzalez A.M., Spitz R.W., Ghigiarelli J.J., Sell K.M., Mangine G.T. Acute Effect of Citrulline Malate Supplementation on Upper-Body Resistance Exercise Performance in Recreationally Resistance-Trained Men. J. Strength Cond. Res. 2018;32:3088–3094. doi: 10.1519/JSC.0000000000002373. [PubMed] [CrossRef] [Google Scholar]
113. Farney T.M., Bliss M.V., Hearon C.M., Salazar D.A. The Effect of Citrulline Malate Supplementation On Muscle Fatigue Among Healthy Participants. J. Strength Cond. Res. 2017:1. doi: 10.1519/JSC.0000000000002356. [PubMed] [CrossRef] [Google Scholar]
114. Trexler E.T., Persky A.M., Ryan E.D., Schwartz T.A., Stoner L., Smith-Ryan A.E. Acute Effects of Citrulline Supplementation on High-Intensity Strength and Power Performance: A Systematic Review and Meta-Analysis. Sports Med. 2019;49:707–718. doi: 10.1007/s40279-019-01091-z. [PubMed] [CrossRef] [Google Scholar]
115. Kleiner S.M., Bazzarre T.L., Litchford M.D. Metabolic profiles, diet, and health practices of championship male and female bodybuilders. J. Am. Diet. Assoc. 1990;90:962–967. [PubMed] [Google Scholar]
116. Kleiner S.M., Bazzarre T.L., Ainsworth B.E. Nutritional Status of Nationally Ranked Elite Bodybuilders. Int. J. Sport Nutr. 1994;4:54–69. doi: 10.1123/ijsn.4.1.54. [PubMed] [CrossRef] [Google Scholar]
117. Sandoval W.M., Heyward V.H. Food Selection Patterns of Bodybuilders. Int. J. Sport Nutr. 1991;1:61–68. doi: 10.1123/ijsn.1.1.61. [PubMed] [CrossRef] [Google Scholar]
118. Ismaeel A., Weems S., Willoughby D.S. A Comparison of the Nutrient Intakes of Macronutrient-Based Dieting and Strict Dieting Bodybuilders. Int. J. Sport Nutr. Exerc. Metab. 2018;28:502–508. doi: 10.1123/ijsnem.2017-0323. [PubMed] [CrossRef] [Google Scholar]
119. Nelson J.R., Raskin S. The eicosapentaenoic acid:arachidonic acid ratio and its clinical utility in cardiovascular disease. Postgrad. Med. 2019;131:268–277. doi: 10.1080/00325481.2019.1607414. [PubMed] [CrossRef] [Google Scholar]
120. Harris W.S. The Omega-6: Omega-3 ratio: A critical appraisal and possible successor. [(accessed on 15 June 2019)];Prostaglandins Leukot Essent Fatty Acids. 2018 132:34–40. doi: 10.1016/j.plefa.2018.03.003. Available online: https://www.ncbi.nlm.nih.gov/m/pubmed/29599053/ [PubMed] [CrossRef] [Google Scholar]
121. Tachtsis B., Camera D., Lacham-Kaplan O. Potential Roles of n-3 PUFAs during Skeletal Muscle Growth and Regeneration. Nutrients. 2018;10:309. doi: 10.3390/nu10030309. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
122. Di Girolamo F.G., Situlin R., Mazzucco S., Valentini R., Toigo G., Biolo G. Omega-3 fatty acids and protein metabolism: Enhancement of anabolic interventions for sarcopenia. [(accessed on 15 June 2019)];Curr. Opin. Clin. Nutr. Metab Care. 2014 17:145–150. doi: 10.1097/MCO.0000000000000032. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24500439 [PubMed] [CrossRef] [Google Scholar]
123. McGlory C., Wardle S.L., Macnaughton L.S., Witard O.C., Scott F., Dick J., Bell J.G., Phillips S.M., Galloway S.D.R., Hamilton D.L., et al. Fish oil supplementation suppresses resistance exercise and feeding-induced increases in anabolic signaling without affecting myofibrillar protein synthesis in young men. Physiol. Rep. 2016;4:e12715. doi: 10.14814/phy2.12715. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
124. Crestani D.M., Bonin E.F.R., Barbieri R.A., Zagatto A.M., Higino W.P., Milion F. Chronic supplementation of omega-3 can improve body composition and maximal strength, but does not change the resistance to neuromuscular fatigue. [(accessed on 15 June 2019)];Sport Sci. Health. 2017 13:259–265. doi: 10.1007/s11332-016-0322-9. Available online: https://link.springer.com/article/10.1007/s11332-016-0322-9 [CrossRef] [Google Scholar]
125. Lewis E.J.H., Radonic P.W., Wolever T.M.S., Wells G.D. 21 days of mammalian omega-3 fatty acid supplementation improves aspects of neuromuscular function and performance in male athletes compared to olive oil placebo. J. Int. Soc. Sports Nutr. 2015;12:28. doi: 10.1186/s12970-015-0089-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
126. Rossato L.T., Schoenfeld B.J., De Oliveira E.P. Is there sufficient evidence to supplement omega-3 fatty acids to increase muscle mass and strength in young and older adults? Clin. Nutr. 2019 doi: 10.1016/j.clnu.2019.01.001. [PubMed] [CrossRef] [Google Scholar]
127. Mocking R.J.T., Harmsen I., Assies J., Koeter M.W.J., Ruhé H.G., Schene A.H. Meta-analysis and meta-regression of omega-3 polyunsaturated fatty acid supplementation for major depressive disorder. Transl. Psychiatry. 2016;6:e756. doi: 10.1038/tp.2016.29. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
128. Maki K.C., Palacios O.M., Bell M., Toth P.P. Use of supplemental long-chain omega-3 fatty acids and risk for cardiac death: An updated meta-analysis and review of research gaps. J. Clin. Lipidol. 2017;11:1152–1160.e2. doi: 10.1016/j.jacl.2017.07.010. [PubMed] [CrossRef] [Google Scholar]
129. Miller P.E., Van Elswyk M., Alexander D.D. Long-Chain Omega-3 Fatty Acids Eicosapentaenoic Acid and Docosahexaenoic Acid and Blood Pressure: A Meta-Analysis of Randomized Controlled Trials. Am. J. Hypertens. 2014;27:885–896. doi: 10.1093/ajh/hpu024. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
130. Du S., Jin J., Fang W., Su Q. Does Fish Oil Have an Anti-Obesity Effect in Overweight/Obese Adults? A Meta-Analysis of Randomized Controlled Trials. PLoS ONE. 2015;10:e0142652. doi: 10.1371/journal.pone.0142652. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Per accedere alle precedenti parti, dalla prima alla terza, clicca qui, qui e qui.
Disclaimer: quelle che seguono sono informazioni provenienti da casi studio [tranne dove diversamente specificato] e testimonianze di Bodybuilder sull’uso dell’Insulina, alcune delle quali sono palesemente sbagliate. Non prendete assolutamente queste opinioni come consigli.
Insulina – dall’uso clinico al Bodybuinding:
Come abbiamo visto nella prima parte, inizialmente l’Insulina farmaceutica era di origine animale. In questo caso, l’Insulina viene estratta dal pancreas di suino o di mucca (o di entrambi) e preparata per uso medico. Queste preparazioni sono ulteriormente suddivise nelle categorie “standard” e “purificate”, a seconda del livello di purezza e del contenuto non insulinico della soluzione. Con questi preparati c’è sempre la possibilità che contaminanti pancreatici siano presenti nel farmaco.
Nel 1977, Herbert Boyer e i suoi collaboratori Keiichi Itakura e Arthur Riggs al City of Hope National Medical Center descrisse la prima sintesi ed espressione di un gene codificante per un peptide. Nell’agosto del 1978, Boyer produsse Insulina sintetica utilizzando i suoi nuovi batteri transgenici geneticamente modificati, seguita nel 1979 dall’Ormone della Crescita. La Tecnologia del DNA ricombinante faceva il suo debutto e cambiava la storia dell’Insulina per uso medico.
L’Insulina umana biosintetica (insulina umana rDNA), attualmente e maggiormente utilizzata per uso clinico, è prodotta con la tecnologia del DNA ricombinante. L’Insulina umana biosintetica ha una maggiore purezza rispetto all’Insulina animale estrattiva e riduce la formazione di anticorpi. I ricercatori sono riusciti a introdurre il gene dell’Insulina umana nelle piante come un altro metodo per produrre Insulina (“biopharming”) nel cartamo. Si prevede che questa tecnica ridurrà i costi di produzione.
Sono disponibili diversi analoghi dell’Insulina umana. Questi analoghi dell’Insulina sono strettamente correlati alla struttura dell’Insulina umana e sono stati sviluppati per aspetti specifici del controllo glicemico in termini di azione rapida (insuline prandiali) e azione prolungata (insuline basali). Il primo analogo biosintetico dell’insulina è stato sviluppato per l’uso clinico al momento del pasto (insulina prandiale), Humalog (Insulina lispro), è assorbita più rapidamente dopo l’iniezione sottocutanea rispetto all’Insulina normale, con un effetto a 15 minuti dopo l’iniezione. Altri analoghi ad azione rapida sono NovoRapid e Apidra, con profili simili. Tutti vengono assorbiti rapidamente grazie a sequenze aminoacidiche che riducono la formazione di dimeri ed esameri (le insuline monomeriche vengono assorbite più rapidamente). Le insuline ad azione rapida non richiedono l’intervallo iniezione-pasto precedentemente raccomandato per l’Insulina umana e le insuline animali. L’altro tipo è l’Insulina a lunga durata d’azione; la prima di queste è stata Lantus (Insulina glargine). Queste hanno un effetto costante per un periodo prolungato, da 18 a 24 ore. Allo stesso modo, un altro analogo dell’Insulina a lunga durata d’azione (Levemir) si basa su un approccio di acilazione degli acidi grassi. A questo analogo è legata una molecola di acido miristico, che associa la molecola di Insulina all’abbondante albumina sierica, prolungando così l’effetto e riducendo il rischio di ipoglicemia. Entrambi gli analoghi ad azione prolungata devono essere assunti una sola volta al giorno e sono utilizzati nei diabetici di tipo 1 come Insulina basale. È disponibile anche una combinazione di un’Insulina ad azione rapida e di un’Insulina protratta, che consente ai pazienti di ottenere un profilo insulinico simile a quello del rilascio di Insulina da parte dell’organismo. L’Insulina viene utilizzata anche in molte linee cellulari, come CHO-s, HEK 293 o Sf9, per la produzione di anticorpi monoclonali, vaccini virali e prodotti per la terapia genica.
L’Insulina viene solitamente somministrata sotto forma di iniezioni sottocutanee tramite siringhe monouso con aghi, tramite un microinfusore di Insulina o tramite penne da insulina a uso ripetuto con aghi monouso. Sul mercato statunitense è disponibile anche l’Insulina per inalazione.
A differenza di molti farmaci, l’Insulina non può essere assunta per bocca perché, come quasi tutte le altre proteine introdotte nel tratto gastrointestinale, si riduce in frammenti amminoacidici, perdendo tutto il suo potenziale di attività. Sono state condotte alcune ricerche su come proteggere l’Insulina dal tratto digestivo, in modo da poterla somministrare per via orale o sublinguale.
Nel 2021, l’Organizzazione Mondiale della Sanità ha aggiunto l’Insulina al suo modello di elenco di farmaci essenziali.
Complice la descrizione iniziale allettante riportata in letteratura riguardante l’azione dell’Insulina sul metabolismo glucidico e proteico, dove tale peptide veniva descritto come l'”ormone più anabolico”, unita alla maggiore disponibilità di approvvigionamento del farmaco data dalla Tecnologia del DNA ricombinante, nel mondo della cultura fisica di alto livello non mancarono i primi pionieri del suo utilizzo “”off-label”.
Si può stimare che nel giro di 40 anni, vale a dire dagli anni 80 ad oggi, l’uso dell’Insulina nel Bodybuilding abbia subito sia un abuso pratico che teorico. Quello che spero di fare con questa mia piccola opera divulgativa è proprio quello di cambiare questa situazione. Come uomo di scienza con anni di ricerca alle spalle sono qualificato per giudicare le conoscenze dei Bodybuilder. Molti di loro conoscono molto meno di me il funzionamento e l’uso dell’Insulina. Quello che porto sul tavolo sono anni di ricerca nella comunità del bodybuilding e una corposo serie di prospettive diverse di culturisti sull’uso dell’Insulina, raccolte da 20 interviste con utilizzatori di Insulina con un’esperienza che va da mesi a decenni. Tranquilli però, fortunatamente non sono un “classico camicie bianco”, ma uno che analizza con attenzione è sa ammettere quando gli atleti hanno ragione e la ricerca scientifica si sbaglia. È ora quindi di trasformare la “broscience” dell’Insulina in scienza vera e propria e di correggere alcuni miti potenzialmente pericolosi.
Sto scrivendo questo articolo non solo per dimostrare le diverse, e persino contraddittorie, opinioni che i bodybuilder hanno sull’insulina, ma anche per ispirarvi a riflettere lucidamente.
I culturisti, quelli con doti intellettive un minimo sopra la media dei loro colleghi, sanno cose che i medici non potrebbero mai sapere perché hanno un obiettivo diverso e priorità diverse. Ma una cosa è certa, i bodybuilder accorti vogliono praticare il bodybuilding in modo scientificamente informato.
Non saranno presenti i nomi dei culturisti intervistati, dei quali sono stati esaminati i video e di cui sono state raccolte le affermazioni nei forum, per un principio etico che prevede di non rivelare l’identità delle persone che hanno contribuito alla presente ricerca, per evitare qualsiasi rischio di danno alla loro persona e immagine. Pertanto, tutti i nomi sono pseudonimi. È sufficiente dire che sono stati intervistati alcuni individui riconosciuti come esperti mondiali nell’uso dell’Insulina per il bodybuilding. sono stati inclusi anche i bodybuilder medi.
L’Insulina vista dai BodyBuilder:
I bodybuilder hanno tra loro visioni piuttosto diverse riguardo all’Insulina e ai suoi effetti. Per esempio, essi non sono d’accordo all’unanimità su quanto sia anabolizzante l’Insulina. Una minoranza afferma che l’Insulina esogena non è direttamente anabolizzante. Alcuni suggeriscono che l’Insulina induce l’anabolismo solo aumentando l’appetito. Ma, come sappiamo bene, di per se, quest’ultima affermazione ha ben poco senso.
Una piccolissima minoranza di bodybuilder sostiene che l’impatto dell’Insulina esogena sia principalmente, o puramente, cosmetico, in quanto l’Insulina fa apparire il muscolo più “pieno” (piuttosto che aumentare effettivamente le dimensioni del muscolo) aumentando la ritenzione idrica intracellulare.
Al contrario, molti bodybuilder sostengono che l’Insulina sia l’ormone più anabolico.
Alcuni partecipanti hanno descritto l’uso dell’Insulina come il risultato di un aumento muscolare di 3-6kg a settimana rispetto a quello che si potrebbe ottenere con il solo uso di Steroidi Androgeni Anabolizzanti (AAS) o con l’uso combinato di AAS e Ormone della Crescita (GH). Tuttavia, queste affermazioni sono state contestate da alcuni. Vi sono bodybuilder che ritengono che i benefici dell’Insulina non siano sufficienti e non hanno intenzione di utilizzare nuovamente questo peptide. Uno di questi non raccomanda più l’uso dell’Insulina ai suoi clienti.
Mentre alcuni bodybuilder sostengono che l’Insulina da sola sia anabolizzante, altri suggeriscono che essa sia significativamente anabolizzante solo in sinergia con AAS e hGH.
La maggior parte dei bodybuilder concorda sul funzionamento dell’Insulina, anche se le loro spiegazioni sono più o meno scientifiche.
In genere i bodybuilder descrivono l’Insulina come una “navetta” o un “mezzo” che trasporta i nutrienti nelle cellule muscolari. L’Insulina viene descritta come una “chiave” che apre le porte delle cellule o come un “autobus” che trasporta i nutrienti. Alcuni bodybuilder citano la letteratura scientifica quando descrivono l’azione dell’Insulina:
Sembra che l’Insulina abbia almeno un effetto permissivo sulla sintesi proteica, tanto che i suoi livelli basali sono necessari per la normale sintesi proteica miofibrillare (MPS), ma l’aumento dell’Insulina dopo un pasto potrebbe non aumentare la MPS (Greenhaff et al., 2008). Tuttavia, l’Insulina promuove l’anabolismo muscolare (bilancio proteico proattivo) attraverso il suo effetto inibitorio sulla degradazione delle proteine muscolari (MPB) (Deutz e Wolf 2013). Inoltre, l’Insulina può aumentare l’MPS attraverso un maggiore assorbimento di aminoacidi (essenziali) nel muscolo scheletrico, provocato da un aumento del flusso sanguigno associato alla vasodilatazione (Biolo et al., 1995; Fujita et al., 2006; Timmerman et al., 2010). [Chad via e-mail].
Molti concordano sul fatto che:
l’Insulina è molto efficace nel trasportare i nutrienti nelle cellule muscolari, ma anche nelle cellule adipose.
l’Insulina è anabolizzante grazie al suo ruolo nella ripartizione dei nutrienti, in quanto lavora di concerto con GH e IGF-I.
l’Insulina promuove l’anabolismo muscolare grazie al suo effetto inibitorio sulla degradazione delle proteine muscolari.
l’Insulina (così come gli AAS e il GH) promuove la sintesi proteica muscolare solo in presenza di un adeguato apporto di aminoacidi.
l’Insulina svolge un ruolo nel controllo fisiologico della riproduzione, agendo sulla secrezione dell’Ormone di Rilascio delle Gonadotropine (GnRH)/luteinizzante (LH).
L’Insulina è anche descritta da alcuni bodybuilder come anti-catabolica.
Un bodybuilder di alto livello ha dichiarato che l’Insulina dovrebbe essere usata solo se un individuo è carente di Insulina:
A volte è una buona idea prendere l’Insulina solo per aiutare il pancreas a fare il suo lavoro. … se il vostro corpo producesse abbastanza da solo, perché avreste bisogno di assumere Insulina esogena? In altre parole, non apporta alcun beneficio. È utile solo se non si produce abbastanza Insulina. Quindi bisogna innanzitutto stabilire se non si produce abbastanza Insulina. Procuratevi un glucometro e controllate la glicemia. … Quindi l’Insulina dovrebbe essere usata solo se si ha una carenza di insulina, perché si sta usando molto GH o perché si sta mangiando una quantità esorbitante di carboidrati.
Definirei questa ipotesi come “patologica indotta/deficitaria”. Ricordatevi sempre che l’omeostasi organica è regolata da feedback. Ciò significa che l’uso di Insulina esogena causerà una sottoregolazione/inibizione della biosintesi endogena di Insiluna. Di conseguenza, parlare di “pancreas ipoattivi” o “supporto pancreatico” non è in definitiva corretto. In tal caso si parla di una vera e propria terapia ormonale sostitutiva dell’Insulina.
Un bodybuilder ha anche affermato che gli effetti anabolici dell’Insulina sono dovuti alla sua azione osmotica e alla sua capacità di aprire “tutti i recettori del corpo”. Descrizione alquanto particolare ma che può rendere una certa idea di uno degli effetti dell’Insulina.
Mentre alcuni bodybuilder mettono in guardia dall’uso dell’Insulina perché può causare ipoglicemia con conseguente coma ipoglicemico potenzialmente letale, la maggior parte ritiene che i rischi dell’Insulina siano stati sopravvalutati e alcuni suggeriscono che la morte dovuta all’uso di Insulina è estremamente improbabile. Molti pensano che “bisogna essere una testa di cazzo per uccidersi con l’Insulina”.
Sebbene la morte di diversi culturisti di alto livello nel corso della mia ricerca sia stata inizialmente suggerita da membri della comunità dei culturisti come correlata all’Insulina, l’Insulina non è stata implicata nelle cause ufficiali dei loro decessi. Non mi sono imbattuto in un caso confermato di morte o di danni significativi causati dall’uso di Insulina per il bodybuilding, anche se alcuni bodybuilder hanno dichiarato di conoscere qualcuno che è morto a causa dell’uso di Insulina (e uno di loro ha avuto un grave incidente d’auto a causa di una ipoglicemia avuta in autostrada, ma fortunatamente nessuno è rimasto ferito). Nella letteratura medica sono riportati due casi di bodybuilder in coma ipoglicemico (Heidet et al., 2019; Petrovic et al., 2015). Non credo che la mancanza di decessi confermati sia dovuta al fatto che l’Insulina non sia pericolosa, ma più probabilmente perché non è comunemente testata o difficile da rilevare.
Un bodybuilder ha suggerito che le morti premature dovute all’uso di Insulina potrebbero non essere dovute solo all’ipoglicemia, ma ha suggerito che l’aumento dei livelli di Insulina nel corso della vita accorcia la durata della stessa e che quindi i bodybuilder si mettono a rischio in questo senso.
Alcuni bodybuilder sostengono che l’Insulina sia uno dei farmaci più sicuri del loro arsenale, in particolare rispetto al DNP e al Trenbolone. Alcuni suggeriscono addirittura che l’Insulina sia più sicura di qualsiasi AAS.
Mentre tutti i bodybuilder hanno descritto almeno lievi sintomi di ipoglicemia in alcuni momenti del loro utilizzo di Insulina, la maggior parte degli episodi di ipoglicemia si sono verificati durante le prime fasi di utilizzo, quando stavano elaborando il dosaggio dell’Insulina, o sono stati attribuiti alla loro stupidità (ad esempio, dimenticando di mangiare). Tutti i bodybuilder hanno dichiarato che l’ipoglicemia era facilmente gestibile consumando zuccheri.
Molti suggeriscono che per essere competitivi come bodybuilder professionisti è necessario utilizzare l’Insulina. Tuttavia, altri suggeriscono che non è necessario.
L’Insulina fa ormai parte del bodybuilding, ne è parte integrante. È come i denti sbiancati. Tutti sbiancano i denti, tutti hanno denti bianchi e splendenti. Se c’è uno che non li ha, gli si chiede: “Cosa c’è che non va in te?”. … Se non fai l’Insulina, cosa che alcuni professionisti non fanno, alcuni non ne hanno bisogno, allora il tuo aspetto è un po’ diverso da quello degli altri bodybuilder e potresti distinguerti in modo negativo.
Molti attribuiscono il significativo aumento della muscolatura dei mostri di massa all’Insulina e/o all’Ormone della Crescita. Alcuni suggeriscono che l’Insulina abbia rovinato il bodybuilding, poiché l’attenzione si è spostata dall’estetica alle dimensioni a scapito dell’estetica. Altri criticano ulteriormente il look dell’Insulina:
Prima dell’arrivo dell’Insulina, tornando ai primi tempi di Bertil Fox, Tom Platts, Arnold, Sergio, i loro muscoli avevano questo aspetto duro e granitico, sembravano scolpiti nella pietra. Ora ci sono questi ragazzi, certo grandi e stravaganti come i Ramy e tutti questi ragazzi, sono grandi ma non hanno quell’aspetto duro e denso.
Ma altri suggeriscono che Dorian Yates è stato il primo a portare sul palcoscenico del Olympia un fisico potenziato dall’Insulina e viene spesso descritto come se avesse un aspetto granitico.
Alcuni suggeriscono che l’Insulina (e/o l’Ormone della Crescita) provochi la “bolla intestinale” o il “palumboismo” [vedi anche “GH Gut”], e una fonte ha affermato che l’Insulina ha causato l’organomegalia. Per questi motivi alcuni affermano che l’Insulina ha rovinato l’estetica del bodybuilding.
Il dosaggio dell’insulina è molto vario tra i bodybuilder. Ho parlato con culturisti che usano un massimo di 4UI al giorno, e altri che hanno usato un massimo di 360UI al giorno! Anche se lo considero ben poco credibile. Tuttavia, in linea con le precedenti ricerche accademiche che riportavano dosaggi compresi tra 10 e 20 unità al giorno (Dawson e Harrison 1997; Evans 1997; Hildebrandt et al., 2007) e con i sondaggi condotti all’interno della comunità, ho scoperto che la maggior parte dei bodybuilder utilizza dosi che si collocano all’estremità inferiore dello spettro. Tuttavia, una buona parte dei bodybuilder tende a usare più di quanto riportato in precedenza nella letteratura accademica e nella comunità, con una dose giornaliera mediana di 40 unità e una dose mediana di 0,39 unità per chilogrammo di peso corporeo.
La maggior parte dei bodybuilder utilizza una sola forma di insulina (ad azione rapida o ultra-rapida), anche se un quarto degli intervistati (n=20) utilizza anche un’Insulina a lunga durata d’azione.
Ma vediamo nel dettaglio i tipi di Insulina utilizzati:
Humalog® (Insulina Lispro): Humalog® è un analogo a breve durata d’azione dell’Insulina umana, in particolare l’analogo Lys(B28) Pro(B29) dell’Insulina che si crea quando gli aminoacidi in posizione 28 e 29 sono invertiti. È considerata equipotente all’Insulina solubile normale su base unitaria, ma con un’attività più rapida. L’inizio dell’azione del farmaco in seguito alla somministrazione sottocutanea è di circa 10-15 minuti e il suo picco d’effetto viene raggiunto in 30-90 minuti. La durata d’azione totale è compresa tra 3-5 ore. L’Insulina lispro viene solitamente utilizzata come supplemento a un prodotto a base di Insulina a più lunga durata d’azione, fornendo un farmaco ad azione rapida che può essere assunto prima o subito dopo i pasti per imitare la secrezione insulinica naturale dell’organismo. Molti atleti ritengono che la sua breve finestra d’effetto la renda un farmaco insulinico ideale per scopi dopanti, in quanto la maggior parte dell’azione può essere concentrata nel periodo successivo all’allenamento sfruttando l’assimilazione dei nutrienti durante la così detta “finestra anabolica”.
Novolog® (Insulina Aspart):Novolog è un analogo a breve durata d’azione dell’Insulina umana, creato quando l’aminoacido prolina in posizione B28 viene sostituito con l’acido aspartico. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 15 minuti e l’effetto di picco si raggiunge in 1-3 ore. La durata d’azione totale è compresa tra le 3 e le 5 ore. L’Insulina Aspart viene solitamente utilizzata come supporto a un prodotto contenente insulina a più lunga durata d’azione, fornendo un farmaco a rapida azione che può essere assunto prima o subito dopo i pasti per imitare la risposta insulinica dell’organismo. Molti atleti ritengono che la sua breve finestra di effetto la renda ideale per scopi dopanti, tanto quanto la Lispro, in quanto la maggior parte della sua azione si può concentra nel periodo successivo all’allenamento durante la “finestra anabolica”.
Humulin®-R “Regular” (insulina Inj): Identica all’Insulina umana. Venduta in alcuni mercati anche come Humulin-S® (Solubile), questo prodotto è costituito da cristalli di zinco-insulina disciolti in un liquido chiaro. Non viene aggiunto nulla per rallentare il rilascio di questo prodotto, per cui viene genericamente indicato come insulina umana solubile. Questo farmaco agisce rapidamente e ha una breve durata d’azione. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di 20-30 minuti, e il suo picco d’effetto si raggiunge in 1-3 ore. Ha una durata d’azione totale tra le 5 e le 8 ore. Insieme a Humalog, queste due forme di Insulina sono le scelte più popolari tra gli atleti e i culturisti per scopi dopanti.
Humulin®-N, NPH (Insulina Isofana): Una sospensione cristallina di Insulina con protamina e zinco per ritardarne il rilascio e prolungarne l’azione. L’Insulina Isofana è considerata un’Insulina di lunghezza intermedia. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore e il picco d’effetto si raggiunge in 4-10 ore. La durata totale dell’attività è superiore a 14 ore. Questo tipo di Insulina non è comunemente usata come agente dopante.
Humulin®-L, Lente (sospensione media di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-L è considerata un’insulina di lunghezza d’azione intermedia. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 1-3 ore e l’effetto di picco viene raggiunto in 6-14 ore. Ha una durata totale di attività superiore alle 20 ore. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin®-U, Ultralente (sospensione prolungata di Zinco): Una sospensione cristallina di Insulina con zinco per ritardarne il rilascio e prolungarne l’azione. Humulin-U è considerata un’Insulina a lunga durata d’azione. L’inizio dell’azione del farmaco dopo somministrazione sottocutanea è di circa 6 ore, e l’effetto di picco viene raggiunto in 14-18 ore. La durata totale dell’attività è di 18-24 ore. Questo tipo di insulina non è comunemente usato per scopi dopanti.
Lantus (Insulina Glargine): Analogo a lunga durata d’azione dell’Insulina umana. L’Insulina Glargine viene creata quando l’aminoacido asparagina in posizione A21 viene sostituito con la glicina e vengono aggiunte due arginine al C-terminale della catena B dell’Insulina. L’inizio dell’azione del farmaco dopo la somministrazione sottocutanea è di circa 1-2 ore, e il farmaco è considerato privo di un picco significativo (è stato progettato per un modello di rilascio molto stabile per tutta la durata dell’attività). L’Insulina Glargine ha una durata d’azione compresa tra 20-24 ore nell’organismo dopo l’iniezione sottocutanea. Questo tipo di Insulina non è comunemente usato per scopi dopanti.
Humulin® (Miscele) : Sono miscele di Insulina solubile normale per un’azione rapida, e di un’Insulina a lunga durata d’azione o ad azione intermetizzata per un effetto prolungato. Queste miscele sono etichettate con la percentuale di miscela, di solito 10/90, 20/80, 30/70, 40/60 e 50/50. Sono anche disponibili le miscele che utilizzano Humalog come Insulina ad azione rapida.
Indipendentemente dal tipo, tutte le insuline forniscono gli stessi effetti di base, e la durata di azione è la differenziazione primaria. Il trasporto dei nutrienti, l’aumento della sintesi proteica, la diminuzione del catabolismo proteico, l’aumento del IGF-1, l’aumento della biodisponibilità del IGF-1, e una maggiore vasodilatazioni sono i vantaggi più noti.
Non è da molto tempo che un tipo di Insulina precedentemente marginale a fini dopanti è diventata di moda tra alcuni culturisti. Sto parlando della Lantus (Insulina Glargine), appunto.
Di tutte le diverse insuline disponibili, la Lantus è probabilmente quella meno utilizzata anche perchè e paradossalmente la più complessa da gestire. Il suo scarso utilizzo nel bodybuilding ha portato, come ovvia conseguenza, ad una scarsità delle informazioni disponibili su di essa.
A differenza delle insuline a breve durata d’azione, che forniscono i benefici di cui sopra per poche ore al giorno, la Lantus continuerà il trasporto dei nutrienti, l’aumento della sintesi proteica, il miglioramento della vasodilatazioni, ecc, per tutto il giorno, anche mentre dormiamo, ed è questo ultimo punto a renderla di non facile gestione. Ma uno degli svantaggi principali della Lantus risiede nella sua possibilità di essere utilizzata solo per brevi periodi di tempo, in quanto l’esposizione continua a livelli elevati di Insulina esogena, e lo sappiamo bene, porterà ad una riduzione della sensibilità all’insulina, la successiva sotto-regolazione dei trasportatori GLUT-4, cose che si vorrebbero evitare sia da un punto di vista della saluta che della crescita muscolare. Così, mentre la Lantus può essere superiore per lo stimolo della crescita muscolare nel breve termine, troviamo che le cose cominciano a pareggiarsi nel lungo periodo, e protocolli come quelli pre-allenamento più comunemente impiegati possono invece essere utilizzati a tempo indeterminato, senza danneggiare eccessivamente la sensibilità all’Insulina a qualsiasi grado significativo. Questo rende la Lantus ideale per dei “blitz”, in cui l’atleta vuole mettere su muscoli il più rapidamente possibile, ma non è adatta per un uso prolungato.
Differenza nella curva di rilascio tra Humulin N e Lantus in pazienti con Diabete di Tipo I.
Una caratteristica decisamente interessante della Lantus è il suo effetto sul IGF-1 ed i suoi recettori. In diversi studi universitari, la Lantus ha dimostrato una maggiore affinità di legame per il recettore del IGF-1 rispetto all’Insulina umana regolare o uno dei qualsiasi altri analoghi. È interessante notare che Levemir, l’unico altro analogo dell’Insulina ad azione prolungata sul mercato, mostra una ridotta affinità di legame ai recettori del IGF-1 umani. Questo mette la Lantus e la Levemir alle estremità opposte dello spettro in termini di affinità di legame. Mentre un aumento di IGF-1 vincolante è visto generalmente come una cosa positiva per la crescita muscolare, è stato il punto focale del dibattito in corso nella comunità medica per parecchi anni, per il fatto che alcuni studi hanno mostrato un aumento del rischio di cancro quando si usa la Lantus. Da allora, altri studi hanno confutato questa nozione, ma il dibattito continua, con la comunità medica riluttante a prendere una posizione in un modo o nell’altro.
Via di segnalazione del Recettore dell’Insulina (IR) e del Recettore del Fattore di Crescita Insulino-Simile 1 (IGF1R). L’Insulina e l’IGF1 si legano ai loro recettori, inducendo un cambiamento conformazionale e l’autofosforilazione della subunità beta di IR e IGF1R. Successivamente, le proteine substrato del recettore dell’Insulina (IRS) o Shc vengono reclutate e fosforilate. Shc attiva la via della mitogen-activated protein kinase-extracellular signal regulated kinase (MAPK-ERK) e le proteine IRS inducono prevalentemente l’attivazione della via della fosfoinositide 3-chinasi (PI3K)-AKT. In questo caso, l’attivazione di PI3K causa la conversione del fosfatidilinositolo 4,5-bisfosfato (PIP 2 ) in fosfatidilinositolo (3,4,5)-trifosfato (PIP 3 ) e l’attivazione e la fosforilazione di AKT da parte della proteina chinasi 1 dipendente dal fosfoinositide. La regolazione dipendente da AKT della forkhead box O (FoxO), del mammalian target of rapamycin complex 1 (mTORC1) e della glicogeno sintasi chinasi 3b (GSK3b) regola la crescita degli assoni, la trascrizione genica, la sintesi proteica e la plasticità neuronale. MEK, MAPK/ERK chinasi; PDK1, proteina chinasi 1 fosfoinositide-dipendente; SOS, son-of-sevenless. [Adattato da Servier Medical Art di Servier, con licenza Creative CommonsAttribuzione 3.0 Unported].
In definitiva, la Lantus viene solitamente usata come “base” di un protocollo di Insulina affiancata dall’uso del Humalog nei protocolli di Insulina e GH pre o post workout.
UI:CHO ratio e timing di somministrazione
Sappiamo tutti che il rapporto più frequentemente citato tra carboidrati e insulina (UI:CHO ratio) è di 10-15g di carboidrati per 1UI di Insulina. Ma non tutti i bodybuilder si attengono a questo rapporto. Diversi culturisti non hanno stabilito un rapporto fisso tra carboidrati e Insulina, mentre altri hanno utilizzato una gamma di rapporti (5-20g per unità di Insulina) con una media di 9-10g di carboidrati per ogni UI di Insulina.
La maggior parte dei bodybuilder ha assunto l’Insulina durante i pasti, ma considerando che i bodybuilder mangiano spesso, questo non ci dice molto. C’è stato un grande dibattito sul momento più efficace per l’uso dell’Insulina: alcuni suggeriscono che il momento più efficace sia il pre-workout, mentre altri affermano che si tratta di un uso irresponsabile, in quanto è difficile determinare quanti carboidrati verranno ossidati durante l’allenamento e quindi l’uso dell’Insulina nel pre-allenamento potrebbe essere pericoloso. Alcuni bodybuilder usano l’Insulina prima e dopo l’allenamento, altri solo dopo.
Alcuni bodybuilder sostengono che l’uso di Insulina a scopo ricreativo espone i bodybuilder al rischio di sviluppare il diabete. Altri bodybuilder sostengono che l’uso dell’Insulina riduce il rischio di diabete in quanto diminuisce l’impatto delle diete per il bodybuilding. Lasciatemi dire che l’ultima affermazione non ha alcun senso. Il corpo mantiene una condizione di omeostasi attraverso elaborati feedback di controllo. La somministrazione di Insulina esogena causerà un feedback negativo a livello della secrezione endogena pancreatica, e l’Insulina esogena somministrata avrà il medesimo effetto a livello centrale e periferico in un regime alimentare ipercalorico (vedi “dieta per il bodybuilding”) dell’Insulina endogena! E le affermazioni secondo le quali l’uso dell’Insulina esogena sortirebbe un effetto di protezione all’affaticamento pancreatico beh, è un affermazione che non ha basi di riscontro.
La follia del protocollo “No Fat Gain Insulin Program”:
Diversi anni fa riportai un protocollo d’uso dell’Insulina denominato “No Fat Gain Insulin Program”. Questo protocollo “alternativo” fu ideato da Mike Zumpano e Oliver Starr i quali si chiesero se ci poteva essere una strategia che permettesse ai bodybuilder di non aumentare eccessivamente di bf durante l’uso di Insulina. In realtà, la motivazione di base per la quale molti culturisti diventano più grassi che grossi quando nelle loro preparazioni inseriscono l’Insulina è fondamentalmente la “la paura” dell’ipoglicemia. La maggior parte del guadagno di grasso è causato dal consumo eccessivo di carboidrati durante l’uso di Insulina. Una regola di “sicurezza” diffusa con l’uso di Insulina dice che bisogna consumare un minimo di 10-15g di carboidrati per ogni UI di Insulina utilizzata (distribuiti nell’arco di tempo d’azione dell’Insulina utilizzata). Un altro errore commesso da molti bodybuilder e che porta ad un eccesso calorico addizionale è che essi non calcolano le UI in base ai CHO della dieta ma calcolano le UI in base al peso e di conseguenza aggiungono i carboidrati di “sicurezza” a quelli già presenti nel loro programma alimentare. Comunque sia, un Bodybuilder che utilizza 8UI di Insulina 2 volte al giorno, in aggiunta al suo normale apporto di carboidrati, proteine e grassi andrà (con il metodo standard) a consumere una quota addizionale di carboidrati pari a 160g. Difficilmente ci si aspetta che 160g in più di carboidrati, o 640Kcal in più al giorno facciano una differenza significativa su un soggetto che magari mangia 5000 o più calorie al giorno.
Anche se la quantità di carboidrati supplementari (10gXUI) comunemente applicata non sembra terribilmente eccessiva, alcuni “pionieri” dei PEDs alla fine degli anni ‘90 erano certi che fosse il motivo principale per cui gli utilizzatori di Insulina guadagnavano quantità sproporzionate di grasso.
Il plasma umano contiene solo circa 5g di carboidrati in uno specifico momento. I diabetici che hanno preso troppa Insulina di solito possono riportare i loro livelli glicemici nel sangue nel range di normalità consumando cinque grammi (solo 20 calorie!) di Destrosio.
Comunque sia, il metodo “alternativo” lo trovarono e fu ribattezzato, come precedentemente accennato, protocollo “No Fat Gain”. Il trucco, se così possiamo definirlo, sarebbe quello di assumere l’Insulina, ma seguendo un dieta Low-Carb. Proprio così, Low. Con un contenuto glucidico di circa 50g al giorno. Oliver Starr, con rudimentali conoscenze in biochimica e fisiologia umana, si chiese se ci fosse qualche altro modo per mantenere la glicemia nel sangue moderata con un alto grado di stabilità. La sua (riduttiva) risposta è stata la gluconeogenesi. Se si guarda su un grafico dei processi biochimici, si può chiaramente vedere che, quando le riserve di glicogeno epatico e muscolare sono esaurite, ma prima che il soggetto vada in chetosi, il corpo comincia a convertire aminoacidi in glucosio per mantenere stabili i livelli di glucosio nel sangue. Questo processo è noto come gluconeogenesi. Come risaputo, seguire una dieta molto povera di carboidrati provoca un esaurimento delle riserve di glicogeno epatico e muscolare. Questo provoca un sovra-regolazione degli enzimi necessari per la conversione rapida ed efficace degli aminoacidi in glucosio. La parola gluconeogenesi significa letteralmente “la nascita di nuovo glucosio.”
La seconda metà dell’ipotesi applicata, ovviamente, è il contenuto proteico della dieta. Se non si mangiano molti carboidrati, l’unico modo con cui il corpo può produrre glucosio è principalmente quello di convertire gli aminoacidi in glucosio. Questo accade in una certa misura ogni volta che si mangiano proteine, tuttavia, quando si mangia una grande quantità di proteine, si viene a creare ancora più glucosio. E’ il livello di glucosio creato dall’eccesso di proteine che dovrebbe impedire il verificarsi di uno stato ipoglicemico in un contesto nel quale si utilizza Insulina esogena con una dieta a basso contenuto di carboidrati.
Per questo protocollo è necessaria l’assunzione di proteine in polvere, perché non c’è modo di riuscire a essere in grado di mangiare la quantità di proteine che si richiedono da cibi interi. Per sostenere il livello di gluconeogenesi supposto per coprire le necessità durante l’utilizzo di Insulina si è proposto che la migliore strategia è quella in cui si consumano 600g di proteine da una combinazione di proteine del siero di latte e caseina, più un pasto solido che contiene da 50 a 100g di proteine, più alcune verdure fibrose a foglia verde. Il resto delle calorie devono provenire da fonti di grassi con scarsissimo o nullo contenuto di carboidrati.
Si può suddividere l’assunzione consumando una bevanda proteica ogni 30 minuti o un’ora, mescolando in un contenitore 3 litri con 100g di proteine e mantenendo una lista di quante volte si svuota ogni giorno.
Bisogna ricordare, però, che in questo protocollo, l’unica cosa che dovrebbe salvaguardare l’atleta (letteralmente) è l’assunzione di proteine. Se si utilizza l’Insulina con tali modalità e non si mantiene un adeguato apporto di proteine, le conseguenze saranno più che spiacevoli, saranno gravissime.
L’approccio teorico all’uso di Insulina in questo protocollo “No Fat Gains” è:
Giorni da 1 a 3: la deplezione di carboidrati. È necessario diminuire i carboidrati al di sotto dei 100g al giorno. Si suggerisce di arrivare a 50g di carboidrati il giorno 3. L’assunzione proteica deve aumentare a 450g al giorno.
Giorni da 4 a 30: le proteine devono essere pari o superiore a 600g al giorno. I carboidrati devono essere mantenuti tra i 100 e i 50g al giorno (50g è meglio) e si dovrebbero utilizzare i grassi affinché si compensi l’equilibrio delle proprie esigenze caloriche giornaliere. Come già detto, si raccomanda l’uso di proteine in polvere di composizione mista (siero di latte e caseina), anche se è possibile utilizzare alcuni cibi interi, se lo si desidera. (Basta tenere a mente che 600g grammi di proteine corrispondono all’incirca a più di 2.5Kg di petto di pollo o di tacchino al giorno)
Partire da una piccola dose di Insulina (4 UI) per poi aumentarla gradualmente (fino anche a 12UI x 3 volte al giorno).
Monitorare regolarmente la glicemia durante il giorno e in specie nel periodo di massima azione dell’Insulina. Se si inizia a perdere la capacità di rimanere svegli, prendere una zolletta di zucchero.
Si, questo protocollo non è soltanto folle ma, cosa fondamentale, è basato su una conoscenza superficiale e con enormi lacune della fisiologia umana. Perchè? Perchè l’Insulina è un regolatore in negativo della gluconeogenesi!
Il ruolo dell’Insulina nella regolazione della produzione epatica di glucosio è ampiamente accettato. Negli individui sani, l’iperinsulinemia fisiologica sopprime la gluconeogenesi del 20%, mentre la glicogenolisi è completamente soppressa. L’iperglicemia da sola sopprime la glicogenolisi epatica con effetti minimi sull’immagazzinamento del glicogeno. Solo la combinazione di iperglicemia e iperinsulinemia ha un effetto significativo sulla sintesi epatica di glicogeno. Pertanto, l’Insulina svolge un ruolo cruciale nel metabolismo epatico del glucosio.
Effetti dell’Insulina sul metabolismo del glucosio e dei lipidi.
Il meccanismo dominante della regolazione insulino-mediata della gluconeogenesi epatica non è però chiaro. L’Insulina esercita un controllo diretto della gluconeogenesi agendo sul fegato, ma influisce anche indirettamente sulla gluconeogenesi agendo su altri tessuti. L’effetto diretto dell’Insulina è stato dimostrato nei cani a digiuno, dove l’Insulina plasmatica portale ha soppresso la produzione epatica di glucosio, anche in assenza di variazioni del glucagone o dei precursori gluconeogenici. Tuttavia, nei modelli murini, l’Insulina è risultata avere effetti più potenti sulla produzione epatica di glucosio in vivo piuttosto che in vitro. Inoltre, è stato dimostrato che gli effetti indiretti dell’Insulina sui tessuti extraepatici sono sufficienti a mantenere il normale metabolismo del glucosio, suggerendo un ruolo importante per la regolazione indiretta della gluconeogenesi da parte dell’Insulina.
Regolazione dell’espressione genica della gluconeogenesi da parte del segnale insulinico epatico. L’azione dell’Insulina regola l’attività dei fattori di trascrizione che controllano l’espressione dei geni gluconeogenici. La fosforilazione mediata da AKT porta all’esportazione nucleare di FOXO1. La fosforilazione inibitoria di CBP/p300 blocca la formazione del complesso di trascrizione di CREB. La modifica di PGC-1α mediante acetilazione mediata da GCN5 o fosforilazione mediata da AKT/CLK2 riduce l’attività trascrizionale di PGC-1α.
Un sistema di segnalazione insulinica intatto è fondamentale per mantenere i livelli di glucosio nel sangue all’interno di un range glicemico normale e ristretto durante i periodi di digiuno o di eccesso di disponibilità di nutrienti, e questo si ottiene in particolare attraverso la regolazione del flusso metabolico attraverso la via gluconeogenica epatica. Un importante nodo di controllo coinvolge la regolazione trascrizionale dell’espressione dei geni chiave della gluconeogenesi epatica Pck1 e G6pc, che avviene principalmente attraverso il fattore di trascrizione FOXO1 e il recettore nucleare HNF4α e il loro coattivatore trascrizionale PGC-1α. La comprensione di queste vie di regolazione è di estrema importanza per comprendere come un massivo consumo proteico in regime low carb non possa assolutamente garantire una stabilità glicemica in presenza di trattamento con Insulina esogena!
Ma si ingrassa di meno con questo protocollo? Direi di no o, comunque, la differenza è irrisoria… Vorrei ricordare in tal sede che, per quanto dispendioso in termini energetici, in un contesto di eccesso calorico (essenziale in regimi “Bulk”) dove si consumano più proteine del necessario, l’organismo utilizza gli AA che le compongono o come come fonte energetica di scarsa resa [3,3Kcal per 1lt di Ossigeno] o li converte in acidi grassi! Certo, per necessità di deficienza nutrizionale la gluconeogenesi degli AA garantisce una stabilità glicemica, ma in fisiologia e non quando quest’ultima viene marcatamente alterata dall’uso di Insulina esogena!
Per concludere questo paragrafo, è corretto sottolineare che la gluconeogenesi si verifica dopo circa 8 ore di digiuno o scarso apporto glucidico, quando le scorte di glicogeno del fegato iniziano a esaurirsi ed è necessaria una fonte alternativa di glucosio. Inoltre è un processo biochimico piuttosto lento e assolutamente non garante della ben che minima sicurezza in un regime d’uso di Insulina in contesto low carb.
L’uso dell’Insulina pre-workout:
I dibattiti sulla reale efficacia dell’Insulina come agente anabolizzante ha spinto atleti e preparatori a sperimentare protocolli diversi. L’unico di questi che si è avvicinato maggiormente ad una logica d’insieme è ““The Ultimate Insulin Protocol” di Mike Arnold.
Ciò che sta alla base di questo protocollo non è altro che una versione “arricchita” di ciò che sta alla base dell’integrazione “intra-workout”.
Spesso si ragiona sul fatto che durante l’allenamento ci si trova in uno stato fondamentalmente catabolico e dopo in uno stato fondamentalmente anabolico. In realtà però, la sintesi proteica ed i meccanismi di anabolismo e catabolismo sono sempre attivi, con diverse prevalenze, quindi i processi di riparazione tissutale iniziano già nel momento in cui il muscolo viene danneggiato e durante il danneggiamento. Questo è ancor più vero nel momento in cui il workout sarà incentrato su più gruppi muscolari. La presenza di una concentrazione di Insulina esogena in circolo addizionata all’introduzione di macronutrienti ben calibrati (vedi integrazione intra-workout) renderebbe maggiormente incisivo il processo.
Il pump e la pienezza muscolare che si può raggiungere seguendo questo protocollo sono, a detta dei “tester”, impressionanti. Il programma trova la sua “magia” nella sua tempistica e nella sinergia degli ingredienti utilizzati.
Qui di seguito è riportato il protocollo nella sua interezza:
60 minuti pre-workout: *** optional (Uno qualsiasi dei supplementi NO stimolanti sul mercato. Gaspari Nutrition “Vasotropin” è un ottimo prodotto).
45 minuti pre-workout: 15UI di Humulin R.
20 minuti pre-workout: 50g di carboidrati ad alto peso molecolare (ex: Vitargo, Karbolyn, etc). 20g di proteine idrolizzate (es: Hydrowhey, Carnivore). 20g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizata. 2g grammi di Beta-Alanina. 10g di Glutammina. 3g di Taurina. 500mg di Potassio. 1g di Vitamina C.
75 minuti dopo il 1° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 10g di Glicerolo Monostearato. 3g di Leucina. 5g di Creatina Monoidrata Micronizzata. 2g Beta-Alanina. 10g di Glutammina. 3g di Taurina.
75 minuti dopo il 2° shake: 50g di carboidrati ad alto peso molecolare. 20g di protein idrolizzate (es: Hydrowhey, Carnivore). 3g di Leucina. 5g di Glutammina.
Proteine totali: 60g (esclusi gli amminoacidi in forma libera aggiunti)
Carboidrati totali: 150g (escluse le trace di carboidrati contenute nelle polveri proteiche).
Prima di tutto, nel formulare il rapporto macros/Insulina sopra esposto, Arnold ha aumentato la quantità di carboidrati-proteine al di sopra di ciò che è tipicamente necessario per una UI di Insulina, al fine di tenere conto degli utilizzatori che dimostrano un grado superiore alla media di sensibilità all’Insulina. La maggior parte degli utilizzatori di Insulina, richiedono circa 8g di carboidrati-proteine per UI di Insulina, al fine di chiudere in pareggio e mantenere la normale soglia di glucosio nel sangue. Questo protocollo utilizza un rapporto 14:1 (macros/Insulina), che dovrebbe permettere a praticamente chiunque di utilizzare questo programma mantenendo il glucosio nel sangue all’interno di un range di normalità.
Va notato che questo programma è stato progettato per essere seguito “come è scritto”, soprattutto per quanto riguarda i tempi di assunzione dei nutrienti e le loro quantità. Per gli utilizzatori di Insulina già “navigati” che sanno quali rapporti sono ideali per loro, essi hanno la libertà di ridurre la quantità di macros consumati per UI, se necessario, come determinato dalla valutazione della loro risposta metabolica. Per gli utilizzatori inesperti, la componente nutrizionale del programma dovrebbe essere rispettata come scritta per almeno 2 settimane, a questo punto l’utilizzatore può quindi iniziare a personalizzare il suo rapporto macros/Insulina, se necessario.
La base di questo programma poggia sul tipo specifico di macros utilizzati. Senza di loro, ogni altro componente/aspetto del programma è influenzato negativamente e in alcuni casi rende il tutto sensibilmente limitato negli effetti. Carboidrati ad alto peso molecolare, come il Vitargo o le Ciclodestrine Altamente Ramificate, hanno dimostrato di essere superiori ad altre forme di carboidrati in diversi modi, per esempio un rapido e costante rilascio di glucosio nel sangue e una bassissima osmolaritá in soluzione.
Passando alla componente proteica; le proteine idrolizzate sono molto più velocemente assorbite rispetto ad altri tipi di proteine e sono l’unica proteina che può essere consumata insieme ai carboidrati ad alto peso molecolare senza compromettere il loro assorbimento.
In questo protocollo vi è anche la possibilità di un aggiunta di uno stimolatore del NO a scelta. L’aggiunta di questi stimolatori (vedi, ad esempio, la Citrullina Malato), pur non “necessario”, aumenterà ulteriormente la circolazione e il trasporto dei nutrienti ai muscoli che lavorano, così come contribuiscono ad aumentare il pumping sperimentato durante e dopo l’allenamento. Il Glicerolo monostearato è incluso anche tra gli ingredienti per il suo ruolo di volumizzante muscolare. Questo composto viene spesso utilizzato appena prima della gara, al fine di contribuire al raggiungimento di uno aspetto pieno e asciutto durante l’esibizione. Sono presenti anche dei volumizatori tradizionali, come la Glutammina, la Taurina, la Creatina, e il Potassio.
Al fine di promuovere un maggiore recupero e una maggiore risposta per una crescita muscolare, la temporizzazione dell’assunzione degli shake è stata messa a punto per mantenere un flusso costante dei nutrienti per tutta la vita attiva dell’Insulina. L’Humulin R è stata appositamente scelta per questo scopo, dal momento che con la sua emivita permetterà all’utilizzatore di sfruttare le “finestre” sia intra che post-allenamento. L’Humulin R offre anche un picco di Insulina meno pronunciato, che risulta più facile da gestire per una buona parte degli utilizzatori rispetto ad una versione di Insulina ad azione più rapida, come ad esempio l’Humalog che, però, viene in alcuni casi sostituita alla scelta classica.
Per via del tempo di esposizione all’Insulina limitato con la pratica di questo protocollo, la sensibilità all’Insulina è solo moderatamente influenzata in modo diretto quando si utilizza il programma per circa 5-6 volte alla settimana. Per gli individui che scelgono di utilizzare il presente protocollo per 3-4 volte alla settimana, le alterazioni dirette sulla sensibilità all’Insulina non risulta essere un problema. Per coloro che eseguono il protocollo per 5-6 volte a settimana, possono intervenire in due modi per assicurare una pienamente ottimale sensibilità all’insulina:
L’utilizzatore può fare 2 settimane “off” ogni 4 settimane “on”.
L’individuo può aggiungere Metformina nel suo programma di 3-4 volte a settimana ad un dosaggio di 750mg-1g/die.
Per gli utilizzatori che si apprestano all’uso di questo protocollo per la prima volta, mentre il rapporto macros / Insulina di cui sopra è sufficiente, l’autore consiglia sempre di iniziare con un dosaggio ridotto e aumentarlo poco a poco fino al raggiungimento del pieno dosaggio. Per uno novizio, un dosaggio di 6-8UI è ideale. Questo può essere seguito da una seconda iniezione da 8-10UI, per poi passare ad una terza iniezione di 10-12UI prima di arrivare al dosaggio massimo di 15UI.
Il protocollo in questione non contempla il solo uso di “shake e insulina” ma anche di GH (nel pre-workout a distanza dall’Insulina) e del IGF-1lr3 (nel post workout).
Il protocollo inizia con un’assunzione di GH, circa 30 minuti prima del workout per far si che i livelli plasmatici siano ragionevolmente alti, prima di aggiungere la dose di Insulina. L’idea alla base di questo, è assicurarsi che il GH passi attraverso il fegato mentre si ha una notevole quantità di Insulina in circolo. Questo è il modo in cui produciamo grandi picchi di IGF-1. Dopo l’allenamento, si somministra l’IGF-1LR3.
Il protocollo esemplificativo è il seguente:
30 minuti pre-workout: 6-10UI di GH subq
15 minuti pre-workout: 6-16UI di Novalog subq
10 minuti pre-workout: assumere lo shake #1
Dopo ogni set eseguito: sorseggiare lo shake #2, e terminarlo entro la fine dell’allenamento.
Andare a casa
Somministrare la dose di 100mcg di IGF-1lr3 (per i suoi effetti sulla sensibilità all’Insulina
Assumere lo shake #3
Formulazione degli shake:
Shake 1: 10-20g di EAA (Amino Acidi Essenziali) o PeptoPro, 40-60g Vitargo, 5g Creatina Monoidrata Micronizata, 200mg di Caffeina (migliora la resintesi di glicogeno); anche in questo caso la dose di carboidrati per ogni UI si aggira in media sui 7-10g.
Shake 2: 10-20g EAA o PeptoPro, 50-100g Vitargo, 5g di Carnitina Monoidrata Micronizata.
Shake 3: 2 tazze di albume pastorizzato, 1 tazza di avena istantanea, 1 banana o 1 tazza di mirtilli, Splenda o Stevia.
La somministrazione di Insulina pre-workout trova la sua motivazione d’essere nel fatto che durante l’allenamento con i pesi si viene a creare uno stress meccanico che a sua volta crea una sovra regolazione dello stimolo anabolico. Più comunemente, questo stato iper-anabolizzante viene indicato come “finestra anabolica”; un determinato periodo di tempo che dura dall’inizio dell’allenamento fino a poche ore dopo. Un modo in cui il corpo reagisce all’allenamento con i pesi è attraverso un aumento della sensibilità all’Insulina. Questo accade quando i recettori per l’Insulina, che risiedono sulla superficie della cellula, rispondono al segnale dell’Insulina in modo più efficiente, cosa che ci permette un migliore assorbimento dei nutrienti all’interno della cellula. Oltre a ciò, avviene un aumento della proliferazione dei GLUT-4 sulla superficie cellulare in maniera insulino-indipendente, caratteristica che richiede una minore quantità di Insulina per avere i massimi effetti nell’uptake cellulare.
Infatti, l’allenamento promuove anche il recupero e la crescita a livello intracellulare aumentando la sintesi proteica, la glicogeno sintasi, dei GLUT4 e l’espressione del trasportatore degli aminoacidi, e diminuendo i livelli di Miostatina. In combinazione con un aumento della sensibilità all’Insulina, queste cose non solo si traducono in una crescita accelerata, ma forniscono un effetto di ri-partizionamento dei nutrienti, in cui il cibo che assumiamo e i macronutrienti ivi contenuti vengono indirizzati maggiormente verso il miocita (cellule muscolari), piuttosto che immagazzinati come grasso.
L’Insulina è il complemento perfetto per questa “finestra anabolica” durante l’allenamento, in quanto non solo i nutrienti vengono trasportati alle cellule muscolari, permettendo così al corpo di approfittare di questo stato anabolizzante intensificato, ma l’Insulina agisce anche per molti dei processi di costruzione muscolare, fornendo uno stimolo ipertrofico raddoppiato nel momento in cui il corpo è più sensibile alla risposta del segnale dell’Insulina.
Come anti-catabolico, e lo abbiamo anche visto nella review presente nella terza parte di questa serie di articoli, l’Insulina è decisamente efficace, riducendo sensibilmente la degradazione del tessuto muscolare che si somma attivamente con la ripartizione delle proteine muscolari, e il tempo di recupero risulta ridotto. Questo ha un duplice effetto:
alterazione in positivo del rapporto catabolismo:anabolismo e maggiore supercompensazione (crescita muscolare);
il recupero più rapido si traduce in un aumento della possibile frequenza di allenamento, permettendo ai muscoli di essere stimolati più volte entro un determinato periodo di tempo e, in definitiva, di crescere più rapidamente;
infine, le concentrazioni di Insulina sovrafisiologiche aumentano in modo significativo il volume ematico all’interno nel tessuto muscolare, aggiungendo un pump muscolare generale che di per se stimola la crescita.
Tuttavia, affinché il corpo possa mettere a frutto tutto questo, sono necessari lo stimolo allenante adeguato e i nutrienti giusti che devono essere presenti al momento giusto. A questo punto entra in gioco la nutrizione intra-allenamento presentata in precedenza.
Nonostante questo protocollo abbia rappresentato la migliore applicazione dell’uso di Insulina nel Bodybuilding, esso presenta delle lacune oltre che dei problemi tecnici che riducono la pienezza del potenziale e il margine di “sicurezza”.
Alcuni atleti hanno sperimentato una maggiore tendenza all’ipoglicemia durante i loro workout con protocolli di Insulina pre workout. Questo imprevisto è limitante oltre che decisamente pericoloso, specie durante un workout. E’ ovvio che il controllo della glicemia e il rispetto dei punti base del protocollo riducono le possibilità del verificarsi di casi ipoglicemici. Ma il rischio è comunque maggiore rispetto ad una condizione di trattamento base (assunzione classica ai pasti).
Inoltre, tiene poco in considerazione le reale farmacocinetica dei PEDs utilizzati riducendo le piene ed ottimali interazioni ed effetti additivi.
GH/Insulin Protocol – pre e post-workout:
In risposta alle lacune del protocollo di Arnold che abbiamo appena visto, ho realizzato una versione perfezionata denominata “GH/Insulin Protocol”.
Tale perfezionamento prende in considerazione in modo preciso sia la farmacocinetica che la farmacodinamica incrociata dei componenti Insulina e GH.
Ora, sappiamo che la curva di rilascio del hGH somministrato per via sottocutanea raggiunge un picco iniziale dopo 30 minuti post iniezione per poi attestarsi a 2-3h e subire un calo significativo dopo 4h dalla somministrazione. Lo strascico di IGF-1 perdura per circa 24h sopra il basale.
Sappiamo inoltre che l’Insulina aumenta la sensibilità epatica del GH con risposta massiva nella sintesi e rilascio di IGF-1riduzione del IGFBP-1 e IGFBP-2 con conseguente aumento della frazione libera e bioattiva di IGF-1;3) l’aumento della sensibilità del GH a livello epatico porta anche ad una riduzione della IGF-1/IGFBP-3 ratio con ulteriore incremento della frazione libera e bioattiva di IGF-1.
Interazioni biologiche a livello ipofisario ed epatico tra Insulina, Ormone della Crescita (GH), Fattore di Crescita Insulino-Simile-I (IGF-I) e Proteine Leganti il Fattore di Crescita Insulino-Simile (IGFBPs). Le frecce aperte indicano la stimolazione e le linee nere sottili l’inibizione. Livelli elevati di Insulina (a destra) possono aumentare indirettamente la biodisponibilità di IGF-I (cerchi pieni) sopprimendo la produzione di IGFBP-1 e, in misura minore, di IGFBP-2 (simboli ombreggiati). A sua volta, l’aumento della biodisponibilità di IGF-I può aumentare l’effetto di feedback negativo sul GH (quadrato aperto), portando a una riduzione della secrezione di GH e a una minore produzione epatica di IGF-I e IGFBP-3. Tuttavia, livelli elevati di Insulina possono anche aumentare il numero e l’attività dei recettori epatici del GH (barra aperta), riflessi da un aumento dei livelli di Proteina Legante l’Ormone della Crescita (GHBP) circolante. Questo effetto può portare a un aumento della produzione epatica di IGF-I e IGFBP-3 regolata dal GH, con un aumento maggiore dei livelli di IGF-I circolante rispetto a quelli di IGFBP-3. Pertanto, insieme a fattori genetici, ormonali e ambientali, l’entità relativa di questi due effetti opposti dell’Insulina sulla produzione di IGF-I potrebbe determinare i livelli di IGF-I circolante. Nel tempo, un’eccessiva biodisponibilità di IGF-I potrebbe aumentare il rischio di cancro del colon-retto, favorendo la sopravvivenza di cellule trasformate e mutate che normalmente andrebbero incontro ad apoptosi.
Quindi, dal momento che l’obbiettivo è quello di creare un ambiente non fisiologicamente riproducibile senza alterazioni iatrogene al fine di ottenere la massima risposta anabolica dal protocollo, il primo punto di congiunzione, o meglio la chiave di volta del protocollo, deve essere l’incrocio del picco di hGH con quello dell’Insulina. Per fare ciò le modalità di somministrazione dovrebbero contemplare primariamente l’iniezione di hGH pre-workout e quella di Insulina (preferibilmente Humalog per via del picco raggiunto entro 15 minuti dalla somministrazione) nel post-workout. Un vantaggio aggiuntivo di questa modifica è la riduzione del rischio ipoglicemico durante la sessione di allenamento.
E no, il vantaggio dello shake intra-workout non viene perso dal momento che la ripartizione calorica è di per se ottimale per via di meccanismi insulino-indipendenti dati dall’attività contro-resistenza (vedi aumento dei GLUT-4 sulla superficie cellulare in seguito all’attività muscolare). Si veda anche l’assorbimento dilazionato dei composti facenti parte della soluzione ingerita e costituente lo shake intra-workout.
I punti chiave del protocollo sono i seguenti:
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8, in questo ultimo caso divise in una dose da 4UI pre workout e due da 2UI appena sveglio e prima di dormire, a secondo della modalità d’uso del hGH; se somministrato giornalmente o a giorni alterni. Vedi a tal proposito l’articolo dedicato alla somministrazione di hGH a giorni alterni];
1h pre-workout possibilità di assumere tra i 25 ed i 50mg di Sildenafil seguiti a 30 minuti dall’inizio del workout dall’assunzione di 8g di Citrullina Malato;
consumare uno shake intra-workout contenente di base 0,5-1g di carboidrati ad alto peso molecolare, 0,25g di proteine idrolizzate e 5g di Creatina Monoidrato;
somministrazione dell’Insulina (Humalog) post-workout [le UI vanno calibrate in base ai CHO consumati nel pasto post-workout; in linea di massima 1UI ogni 10Kg di peso]. L’Humalog può essere sostituita con l’Humulin-R se per la risposta del soggetto in quanto a tolleranza è migliore.
Schema esemplificativo dell’azione incrociata tra Insulina e GH a livello epatico e sue consequenziali principali aree di influenza ricercate.
Esiste una variante del suddetto protocollo la quale contempla l’uso di due tipi di Insulina: una base (Lantus) e una post-workout (Humalog):
calcolare la dose totale di Insulina giornaliera con la formula 1UI ogni 10Kg di peso;
dividere il totale della dose a metà tra Insulina Glargine (Lantus) e Insulina lispro (Humalog) o Humulin-R;
somministrazione di Insulina glargina (Lantus) dividendo la dose giornaliera in 2: la prima, pari al 65% della dose totale, al mattino in concomitanza con il primo pasto e la seconda, pari al 35% della dose totale, 12h dopo la prima somministrazione;
somministrazione di hGH pre-workout [UI utilizzate dalle 4 alle 8 a secondo della modalità d’uso del hGH];
somministrare la dose di Insulina lispro o Humulin-R nel post-workout.
L’uso di una base di Insulina rappresentata dalla Insulina glargina ha il potenziale di aumentare ulteriormente l’espressione del IGF-1 e, di conseguenza, la risultante anabolizzante del protocollo. Ma, ovviamente, i rischi di ipoglicemia con questa modifica protocollare sono maggiori.
Grafico esemplificativo delle curve ematiche di Insulina, hGH e IGF-1 nel protocollo con Insulina Glargine di base.
Il monitoraggio regolare della glicemia e il rispetto dei punti base del protocollo evitano con un buon margine il verificarsi di casi ipoglicemici. Quindi, il glucometro, che sia classico o con sensore, è essenziale in questi casi.
Repetita iuvant: QUESTO PROTOCOLLO COME I PRECEDENTEMENTE PRESENTATI, E L’USO DI INSULINA, SONO APPANNAGGIO DI ATLETI AVANZATI E MONITORATI DA PERSONALE QUALIFICATO! NULLA DI CIO’ CHE E’ STATO DESCRITTO DEVE ESSERE PRESO COME UNA PRESCRIZIONE MEDICA O UN CONSIGLIO!
Conclusioni “dopo un lungo viaggio”:
Siamo ora giunti alla conclusione di questa serie di articoli dedicati all’Insulina e al suo centesimo anniversario.
Durante questo lungo percorso abbiamo imparato a conoscere meglio questo affascinante e mal compreso peptide. Abbiamo visto come è stato scoperto, abbiamo compreso l’enorme passo avanti nella medicina e nella tutela della vita umana che la sua scoperta ha rappresentato, abbiamo compreso come essa sia fisiologicamente regolata e quali sono le sue reali azioni a livello sistemico ed abbiamo imparato a separare i luoghi comuni che vi aleggiano intorno dai fatti.
Con la terza parte molti sono rimasti delusi nel constatare che in fisiologia l’Insulina abbia un effetto prettamente anticatabolico, effetto preminente che mantiene anche se somministrata esogenamente entro i range fisiologici [<1.200ng/dl]. Altresì gli umori sono migliorati quando siamo venuti a conoscenza del fatto che non solo il dosaggio fa la differenza tra preminenza anticatabolica e anabolica ma anche la sensibilità.
Con la recente constatazione che il risultato dell’equazione “Insulina/anabolismo” cambi drasticamente in positivo se vi si aggiunge la variabile del hGH, abbiamo imparato che l’unico uso minimamente sensato dell’Insulina per il miglioramento della massa muscolo-scheletrica è in associazione con il peptide ipofisario con le ultime due modalità esposte.
Ci tengo però a precisare che l’uso dell’Insulina “off-label” dovrebbe rappresentare la componente più marginale nella carriera di un culturista. Vale a dire che se ne può benissimo fare a meno, in specie quando si è semplici amatori o agonisti di piccoli o medi circuiti competitivi. Non complicatevi la vita.
Fate tesoro delle nozioni che vi ho esposto affinché il confine della conoscenza si espanda e prevalga su quello dell’ignoranza.