Le radici di ginseng contengono diverse sostanze simili agli steroidi e una di esse è il ginsenoside Rg1. In seguito all’assunzione di 2 capsule contenenti 5 mg di Rg1, sono stati osservati miglioramenti nelle prestazioni in soggetti giovani inattivi. Durante l’esercizio intenso l’effetto è risultato maggiore con una azione protettiva a livello muscolare e un tempo di recupero ridotto.(1)
Gli scienziati dell’Università di Taipei (Taiwan) hanno reclutato studenti di sesso maschile, sani ma non allenati. Li hanno sottoposti ad esercizio fisico, somministrando loro una dose di 5mg di ginsenoside Rg1 la sera prima della sessione allenante, e un’altra dose eguale un’ora prima della sessione. In un’altra occasione gli studenti hanno ricevuto un placebo.
I ricercatori stavano studiando il ginsenoside Rg1 da diverso tempo. La loro ricerca è stata finanziata dal governo di Taiwan e dalla NuLiv, con sede negli Stati Uniti [nulivscience.com]. La NuLiv produce estratti erboristici che successivamente vengono commercializzati sotto forma di integratori alimentari.
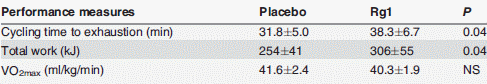
Quando i ricercatori hanno chiesto ai soggetti presi in esame di pedalare il più a lungo possibile ad un’intensità pari all’80% del loro VO2max, hanno osservato che la supplementazione con Rg1 causava un aumento del 20% del tempo di resistenza.
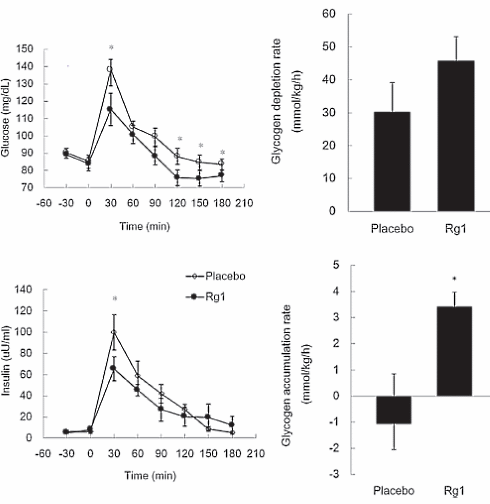
Dopo la sessione allenante, i soggetti dello studio hanno consumato un pasto ricco di carboidrati. Quando i ricercatori hanno analizzato il sangue dei soggetti, hanno notato che lo steroide aveva accelerato l’assorbimento del glucosio [in basso a sinistra]. In basso a destra si può vedere che la supplementazione con ginsenoside Rg1 ha aumentato la quantità di glicogeno nelle cellule muscolari.
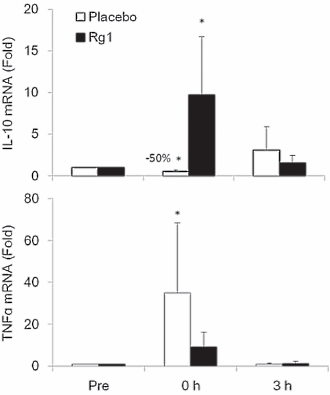
Dopo l’esercizio, la concentrazione cellulare di TNF-alfa dei soggetti trattati con placebo era aumentata. L’Rg1 ha ridotto tale concentrazione. Allo stesso tempo, la sostanza ha indotto le cellule muscolari a produrre più interleuchina-10 (anti-infiammatoria).
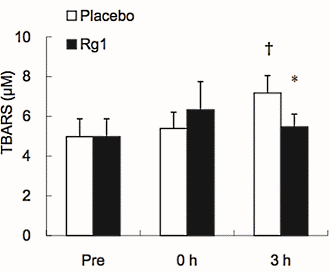
Dopo la sessione allenante, il danno delle cellule muscolari causato dall’attività dei radicali liberi [TBARS] era aumentata nei soggetti trattati con placebo. La supplementazione con ginsenoside Rg1 ha ridotto questo aumento.
I ricercatori affermano che, il risultato principale dello studio è stato quello di dimostrare come la supplementazione con Rg1 pre-workout aumenti significativamente le prestazioni di resistenza durante attività fisica ad alta intensità.
I ricercatori concludono dicendo che, i partecipanti allo studio non allenati sono stati in grado di aumentare la loro capacità di lavoro su un cicloergometro del 20% circa rispetto ai soggetti trattati con placebo. Lo steroide Rg1 può offrire benefici ergogenici per l’uomo.
Un altro studio, svolto dagli stessi ricercatori del precedente, questa volta di recente realizzazione, che ha preso in esame l’azione del Rg1 sugli esseri umani, ha osservato che la combinazione di esercizio fisico intenso e supplementazione con ginseng ha un effetto protettivo a livello muscolare. Lo studio svolto dai biochimici taiwanesi sarà pubblicato sul Journal of Ginseng Research.(2) I ricercatori hanno somministrato ai soggetti dello studio 5mg di ginsenoside Rg1 prima di una intensa sessione su un cicloergometro.
Come per il precedente studio, anche questo è stato in parte finanziato dalla NuLiv Science, [nulivscience.com] e alcuni degli autori sono affiliati alla NuLiv come consulenti scientifici.
Una dei prodotti realizzati dalla NuLiv è l’ActiGin, una combinazione di estratti da Rosa Roxberghii e Panax ginseng. L’ActiGin dovrebbe migliorare le prestazioni sportive, aumentare l’assorbimento di glucosio da parte delle cellule muscolari e accelerare il recupero. Nella pubblicazione della quale stiamo parlando ora si può leggere che questo lavoro è stato finanziato allo scopo di produrre un supplemento ergogenico ActiGin per la Nuliv Science.
Tuttavia, in questo studio i ricercatori non hanno usato l’ActiGin, ma il ginsenoside Rg1. Non mi è stato però possibile constatare il reale contenuto di ginsenoside Rg1 nel ActiGin.
I ricercatori hanno somministrato a 12 studenti maschi non allenati una capsula contenete 5mg di ginsenoside Rg1, un’ora prima che iniziassero la sessione su cicloergometro ad un’intensità pari all’80% del loro VO2max. I soggetti presi in esame dovevano continuare a pedalare fino ad esaurimento. In un’altra occasione, i ricercatori hanno ripetuto l’esperimento, ma somministrando ai soggetti un placebo.
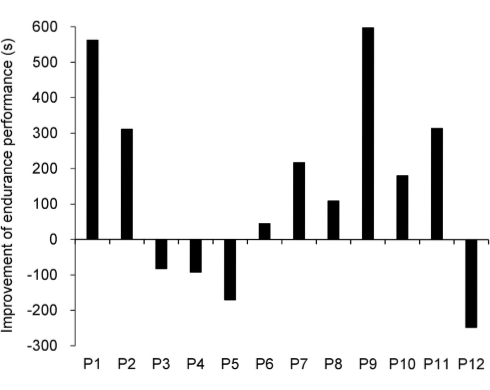
Per la maggior parte dei soggetti presi in esame la supplementazione con Rg1 ha prolungato il loro tempo di resistenza sul cicloergometro. L’evento osservato è paragonabile a quanto riportato per il precedente studio.
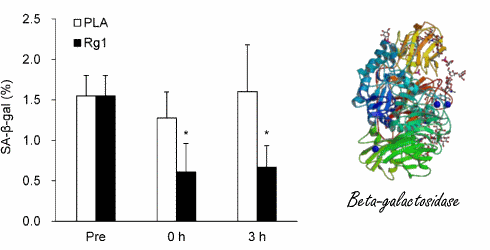
Prima e durante la sessione su cicloergometro, i ricercatori hanno misurato la concentrazione dell’enzima beta-galattosidasi senescente-associato nelle cellule muscolari dei soggetti dello studio. Si tratta di un marker per l’invecchiamento cellulare. Maggiori sono le concentrazioni dell’enzima, più una cellula è senescente.(3)
La combinazione di cicloergometro e supplementazione con ginsenoside Rg1 ha ridotto la concentrazione di beta-galattosidasi.
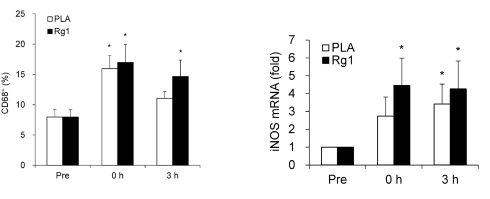
In basso a sinistra si può notare come il ginsenoside Rg1 può portare ad un “ringiovanimento” muscolare. Se i soggetti dello studio avevano assunto l’Rg1, i ricercatori osservavano una maggiore concentrazione di CD68+ nei loro muscoli dopo lo sforzo. Si tratta di cellule le quali ripuliscono le cellule muscolari danneggiate stimolando le cellule staminali a sostituirle.
I ricercatori concludono affermando che, i risultati dello studio suggeriscono che l’attivazione dei macrofagi dopo supplementazione con Rg1 è associata alla clearance delle cellule senescenti osservate nel muscolo scheletrico umano allenato.
Se non avete ancora letto le precedenti parti componenti questa serie di articoli vi invito a farlo prima di procedere con la lettura di questa quarta ed ultima parte: 1° Parte – 2° Parte – 3° Parte.
Farmacocinetica, Farmacodinamica e Feedback Negativi
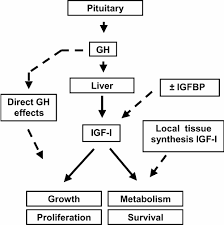
Come già detto, il fegato rappresenta il principale bersaglio del GH, il quale è il principale regolatore della sintesi epatica di IGF-1. Per causare tale effetto, il GH si lega con i GHR localizzati nel dominio extracellulare degli epatociti stimolando successivamente la produzione di IGF-1 endocrino tramite la trascrizione genica, utilizzando la via di segnalazione JAK-STAT. Inoltre, è stato dimostrato che la somministrazione di GH causa una rapida sovraregolazione dell’mRNA del IGF-1 nel fegato.[338]
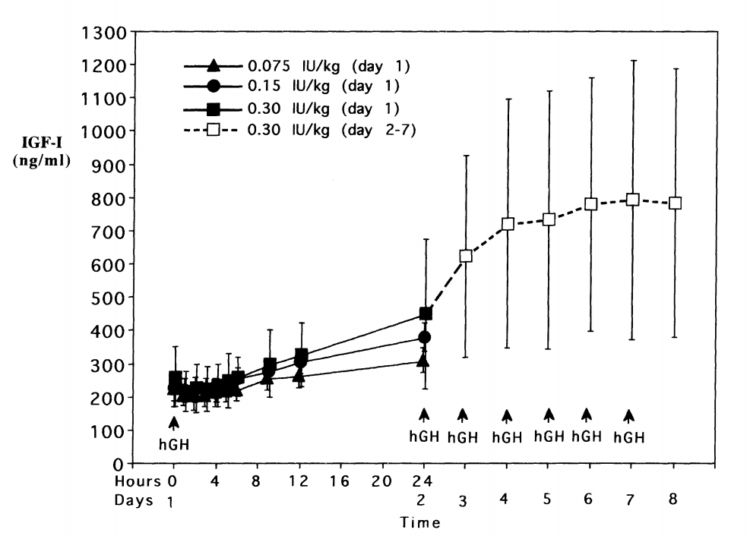
Aumenti dei livelli serici di IGF-1 si verificano molto rapidamente anche in presenza di un grande bolo di rHGH. Incrementi significativi di IGF-1 sono già osservabili dopo 6-12h dall’iniezione.[339] Questi livelli serici di IGF-1 continuano ad aumentare fino a raggiungere il loro punto di saturazione dose-dipendente entro 4-7 giorni, anche quando si utilizzano dosi estremamente elevate che ammontano a 20-30UI al giorno di rHGH.[340] In particolare, il punto di saturazione si è rivelato essere compreso nell’intervallo dei 700-800 ng/mL e sembra suggerire che i livelli endocrini di IGF-1 hanno un tetto massimo negli adulti sani. I meccanismi esatti devono ancora essere chiariti, ma sono probabilmente il risultato dei complessi meccanismi di controllo intrinseci all’Asse GH/IGF-1. Coloro i quali desiderano elevare i livelli endocrini di IGF-1 al fine di ottenerne un vantaggio sull’ipertrofia dovrebbe tenerlo a mente, in quanto vi è un punto in cui l’uso di dosi maggiori di rHGH semplicemente non si traducono in elevati livelli serici di IGF-1. Qui di seguito ho riportato il grafico dello studio di Tanaka il quale mostra la relazione tra l’rhGH e i livelli serici di IGF-:
Ora, vorrei dedicarmi brevemente all’analisi dell’azione del IGF-1autocrino e del perché esso rappresenti un mediatore cruciale del processo ipertrofico, prima di tornare nuovamente a discutere su questioni inerenti alla farmacodinamica e farmacocinetica. La segnalazione recettoriale del IGF-1 è unica nel suo genere, e questo lo si deve al fatto che utilizza due percorsi distinti per stimolare la proliferazione o la differenziazione.[341-343] Questo è un comportamento abbastanza interessante, poiché nessun altro membro della famiglia dei fattori di crescita ha dimostrato di agire in tal modo. Poiché la proliferazione e la differenziazione sono processi opposti, inizialmente era difficile per i ricercatori capire come un singolo fattore di crescita, attraverso un singolo recettore, potesse inviare un segnale che attivasse entrambi.[294] Da quando sono state fatte queste prime scoperte, è stato ulteriormente chiarito che l’IGF-1 non svolge simultaneamente queste azioni. Test su varie linee di coltura cellulare hanno dimostrato che gli effetti proliferativi arrivano prima, durando tra le 24 e le 36 ore. È solo dopo questa fase proliferativa iniziale che si verifica la differenziazione miogenica.[344]
Gli effetti proliferativi mediati dall’IGF-1 sui mioblasti sono noti sin dagli anni ’70, quando vennero osservati per la prima volta nelle cellule epatiche di ratto.[345] Questa stimolazione proliferativa del IGF-1 si traduce in un aumento del numero di cellule, nei livelli di proteine, nella sintesi del DNA, nell’assorbimento di aminoacidico, nell’assorbimento del glucosio e nella soppressione della proteolisi.[346] Nelle colture cellulari umane, l’IGF-1 ha anche dimostrato di aumentare la dimensione dei miotubi indipendentemente dal fatto che i mioblasti proliferino attivamente o che la proliferazione sia cessata. Regola la dimensione dei miotubi attivando la sintesi proteica, inibendo la degradazione proteica e inducendo la fusione delle cellule di riserva.[347-348] La capacità dell’IGF di sopprimere la proteolisi nel muscolo scheletrico, la scomposizione delle proteine in aminoacidi, è stata dimostrata innumerevoli volte nel corso degli anni.[349-352] È stato anche dimostrato che l’IGF-1 induce la proliferazione e la differenziazione delle cellule satelliti in miociti maturi, come determinato da un aumento del numero di miofibre nucleate a livello centrale rispetto a quelle periferiche.[148,353-354]
La capacità dell’IGF-1 autocrino di causare la differenziazione dei mioblasti è stata in realtà una scoperta che potremmo definire quasi “ibrida” dal momento che degli studi svolti negli anni ’60 avevano mostrato che questo effetto si verifica con alti livelli di Insulina.[355] Successivamente è stato dimostrato che gli IGF sono stimolatori molto più potenti nella differenziazione miogenica rispetto all’Insulina e si è concluso che la stessa Insulina agisce realmente come un analogo dell’IGF-1 in questo sistema.[356-357] Gli effetti di differenziazione dati dall’IGF-1 autocrino sono bifasici, con basse concentrazioni che stimolano progressivamente la differenziazione dei mioblasti mentre concentrazioni molto elevate mostrano una cessazione dell’attività di differenziazione. Il limite massimo per la differenziazione sembra attestarsi a circa 100ng/mL per l’IGF-1 e 300ng/mL per IGF-2.[358] Questo effetto non è legato alla proliferazione, poiché non si osservano ulteriori aumenti nel numero complessivo delle cellule.[294] È possibile che le molecole di segnalazione coinvolte nella regolazione negativa del sistema miogenico siano aumentate, ma questa è una affermazione puramente speculativa.[359-360]
La somministrazione di rHGH eleva l’espressione dell’mRNA dell’IGF-1 nel muscolo scheletrico in numerosi modelli cellulari, umani e animali.[127,150,361-364] Ciò avviene abbastanza rapidamente, entro 60 minuti dall’iniezione sottocutanea di rHGH ed i picchi sono segnalati tra le 6 e le 12 ore post iniezione.[363] In questo particolare modello animale citato, il raddoppio della dose di GH non ha portato ad ulteriori aumentati dei livelli di mRNA dell’IGF-1, il che suggerisce che esiste un sistema di regolazione che determina quanto GH sia necessario per stimolare al massimo l’espressione locale dell’IGF-1 nel muscolo scheletrico. In precedenza si è potuto appurare che la differenziazione dei miociti mediata dall’IGF-1 si arresta quando le concentrazioni locali raggiungono circa i 100ng/mL, ma quanto GH è necessario per raggiungere il punto di saturazione dell’espressione dell’mRNA dell’IGF-1?
Gli studi sui miociti umani mostrano che il GH aumenta l’espressione dell’mRNA dell’IGF-1 entro 30-60 minuti con picchi molto più rapidi rispetto a quelli osservati negli studi sugli animali, entro 1-2 ore, usando la via di segnalazione JAK / STAT5b.[365] Questi livelli elevati di mRNA hanno dimostrato di durare fino a 48 ore dopo una singola esposizione al GH. La quantità di GH necessaria per stimolare al massimo l’espressione dell’mRNA dell’IGF-1 è risultata essere una dose compresa tra i 7,5ng/mL e 30ng/ml [366], con una dose media efficace che si attesta a 3ng/ml. Questi numeri sono in linea con gli intervalli di dose fisiologica osservati negli animali, che sono effettivamente compresi tra i 2-100 ng/mL.[367] Inoltre, si collocano esattamente in linea con quanto si osserva endogenamente nell’uomo, con concentrazioni normali di picco comprese tra i 22,4 e 32,4ng/mL.[368-369,436] Ci sono stati casi in cui gli uomini presi in esame hanno mostrato concentrazioni di picco leggermente più alte, ma questi devono essere considerati valori anomali.[370] In ogni caso, ciò che questi dati tendono a suggerire è che il corpo umano è particolarmente adatto a gestire livelli naturali di picco della secrezioni di GH endogeno. Cercare di incidere ulteriormente il sistema elevando i livelli di GH oltre quelli endogeni, unicamente per tentare di potenziare i processi ipertrofici, potrebbe in realtà non tradursi nell’effetto desiderato.
Gli studi che mettono a confronto le infusioni locali con le infusioni sistemiche di GH o IGF-1 sono un po’ più difficili da trovare di quanto si vorrebbe. Le poche sperimentazioni sugli animali che sono riuscito a trovare indicano che l’infusione diretta di GH o IGF-1 nei tessuti bersaglio determina un aumento della massa muscolare. Questo aumento dell’ipertrofia si verifica anche senza che il muscolo bersaglio sia stato sottoposto ad attività motoria.[371-372] Gli studi dimostrano anche che le iniezioni locali di GH portano a livelli sostanzialmente più alti nell’espressione dell’mRNA dell’ IGF-1 locale rispetto alle iniezioni locali di IGF-1, di un fattore di oltre venti.[127] Sono riuscito a trovare uno studio nel quale si confrontavano le risposte dei ratti (attivi e non) all’infusione locale di IGF-1. Il gruppo “IGF-1 plus training” ha mostrato un aumento sia della massa muscolare che della forza locale maggiore rispetto al semplice trattamento in isolamento.[373] Quindi, anche se limitata, la letteratura disponibile è apparentemente in grado di dimostrare che le iniezione locali di GH o IGF-1 hanno effettivamente valore.
Ne ho già parlato diverse volte ma, nel tentativo di imprimere ulteriormente questo concetto, è necessario ricordarsi che i livelli autocrini di IGF-1 sembrano essere molto più importanti dei livelli endocrini di IGF-1 in relazione alla regolazione della massa muscolare. Oltre a questo punto, la sovraespressione dell’IGF-1 autocrino nel muscolo provoca l’ipertrofia delle fibre.[374] La sovraespressione dell’IGF-1 autocrino ha anche mostrato effetti anti-catabolici, con modelli animali tendenti a mostrare una resistenza generale all’atrofia muscolare normalmente osservata con l’invecchiamento.[375] L’IGF-1 localizzato fornisce anche capacità rigenerative indipendenti dall’età nelle cellule muscolari.[376]
Vi sono anche alcune prove convincenti che suggeriscono che l’IGF-1 endocrino agisce direttamente come un regolatore di feedback negativo sulla produzione di IGF-1 autocrino. Questo meccanismo di feedback negativo è dipendente dal pathway PI3K/Akt [377-378]. Inoltre, elevati livelli di IGF-1 endocrino possono anche agire indirettamente per sopprimere la produzione di IGF-1 autocrino. Quindi, in altre parole, non solo l’IGF-1 endocrino ha un impatto diretto minore sulla regolazione della massa muscolare, ma può anche sopprimere l’IGF-1 autocrino che ha impatti maggiori sull’ipertrofia.
Elevati livelli di IGF-1 circolante e, nello specifico, di IGF-1 libero elevati agiscono in modo negativo sul GH determinando un tasso di soppressione della produzione di IGF-1 autocrino a valle.[379] Non è del tutto chiaro, tuttavia, se la regolazione negativa dell’IGF-1 modifichi l’emivita dell’mRNA dell’IGF-1 o influenzi direttamente l’espressione del gene IGF-1. Oltre a questo, è stato anche dimostrato che l’espressione dell’IGF-1 autocrino è sottoregolata nelle cellule muscolari dopo trattamento con IGF-1.[366] È stato anche dimostrato che l’espressione epatica dell’mRNA dell’IGF-1 è sottoregolata dall’esposizione acuta all’IGF-1.[127] Quindi, mantenere livelli endocrini il più possibile soppressi con rispettiva dose di rHGH, elevando contemporaneamente i livelli autocrini, dovrebbe essere un fattore prioritario in un protocollo di GH volto all’ipertrofia.
Il GH è pulsatile per natura sia nell’uomo che nelle specie animali. Quindi, sarebbe logico pensare che molti dei processi intrinseci del corpo saranno tarati in modo tale da rispondere in maniera ottimale all’esposizione al GH in modo simile. In accordo con questa affermazione è stato dimostrato che solo la somministrazione di GH pulsatile, e non l’infusione continua, ha la capacità di stimolare massimamente l’espressione dell’mRNA dell’IGF-1 nel muscolo scheletrico.[366,380-381] È stato anche dimostrato che la somministrazione pulsatile porta ad un aumento del potenziale di crescita postnatale complessivo rispetto all’infusione continua.[89,382] La somministrazione pulsatile può anche portare a livelli endocrini di IGF-1 serici comparabili, o addirittura diminuiti [383], il che è vantaggioso a causa delle potenziali capacità di regolazione negativa che possiede sull’espressione dell’IGF-1 autocrino e che sono state discusse in precedenza. L’evidenza suggerisce anche che il picco stesso, e non necessariamente il numero di picchi, potrebbe essere della massima importanza per i tessuti bersaglio.[384] Per la massima crescita e potenziale ipertrofico, l’evidenza tende a suggerire che creare picchi di GH elevati, e quindi tornare ai livelli basali più volte al giorno, può essere preferibile rispetto a mantenerli elevati per periodi di tempo più lunghi. Questo pratica permette di riprodurre gli schemi secretori in vivo.
I pathways del GH coinvolti nell’anabolismo sono anche suscettibili alla desensibilizzazione, che è parte della fisiologia del GH endogeno.[385] A causa della natura intrinsecamente pulsatile del GH in vivo, l’attività dei recettori e dei pathways sono regolati da un impulso seguito da un periodo di inattività.[386] L’esposizione continua o ripetuta al GH senza un adeguato lasso di tempo refrattario comporterà livelli di attività fortemente soppressi. In effetti, nel corso degli anni numerosi studi hanno dimostrato che tale effetto si verifica. Le cellule ed il tessuto muscolare richiedono un periodo refrattario piuttosto lungo prima che la loro piena risposta al GH venga recuperata. Dopo l’esposizione al GH, le cellule muscolari non sono nemmeno in grado di rispondere alle successive dosi di GH. In realtà, occorrono due ore complete per riprendere parzialmente la reattività nei modelli cellulari, con un totale di 6-8 ore di astinenza dall’uso di GH necessarie per ripristinare la piena sensibilità.[366] Viceversa, quando il GH è micro-dosato in impulsi di dieci minuti, seguiti da intervalli di otto ore, è stato mostrato aumentare progressivamente l’mRNA dell’IGF-1 con ogni impulso successivo.[386]
Questo fenomeno è potenzialmente il risultato di una desensibilizzazione complessiva all’interno della via JAK-STAT5, poiché è stato dimostrato che l’esposizione al GH negli studi sulle cellule epatiche causa resistenza alla successiva attivazione della via STAT5 per 4-8 ore.[387-388] Questo lasso di tempo è sufficiente per sincronizzarsi abbastanza bene con ciò che è stato visto nei modelli di cellule miocitarie citati in precedenza. Nei modelli di cellule epatiche, il GH ha stimolato un significativo aumento dell’espressione del SOCS3, che è un potente inibitore dell’azione del GH.[389]. Poiché il GH non ha avuto alcun effetto sull’espressione del SOCS3 nelle cellule muscolari, questo deve essere un altro meccanismo causante il periodo refrattario. Questo meccanismo può essere dipeso dalla sottoregolazione dei GHR, dall’inibizione mediata da un’altra proteina SOCS, o dall’induzione di una tirosina fosfatasi che semplicemente inattiva la via JAK / STAT.[390] La via JAK-STAT5b, che come ricorderete è intimamente associata al muscolo scheletrico e all’espressione dell’IGF-1, è di natura transitoria – con attivazione massima raggiunta entro 10-30 minuti, seguita da un prolungato periodo di inattivazione.
Una scoperta piuttosto nuova di Xu et al. [391] ha dimostrato che anche distanziare le esposizioni al GH di cinque ore lasciava entrambi i percorsi a valle MEK1/2 e ERK1/2 significativamente soppressi rispetto a tutti i percorsi a monte, a causa di una potenziale disconnessione nella trasduzione del segnale . Ciò è di particolare interesse in quanto questi stessi due percorsi a valle sono stati coinvolti in modo significativo sia nella crescita sia nella proliferazione.[392-393] È stato anche scoperto che l’attivazione indotta da GH di STAT1 e STAT3 è stata desensibilizzata, ma l’esposizione all’Insulina inverte la desensibilizzazione osservata in tutti i percorsi interessati. Anche se non sto per trattare approfonditamente l’Insulina, ci sono un paio di importanti punti da dovere prendere in considerazione. Bisogna comprendere innanzitutto che ci sono molti obiettivi a valle del recettore del GH e molti di questi hanno il potenziale per essere desensibilizzati dopo l’esposizione al GH. Bisogna comprendi anche che l’Insulina possiede l’abilità unica di risensibilizzare molti di questi percorsi. Ciò ha un senso vista la relazione tipo yin-yang tra i due composti. È noto che il GH e l’Insulina possiedono una relazione anabolica sinergica a causa di molti effetti che esercitano l’uno sull’altro. Questo sembra essere soltanto un’anteprima di uno di questi effetti.
Asse GH/IGF-1 – Relazione con altri ormoni
Prima di passare alle note conclusive, vorrei trattare brevemente alcuni altri ormoni connessi a diverso grado con l’Asse GH/IGF-1. Per prima cosa, voglio trattare brevemente l’Asse Tiroideo dal momento che l’inserimento di composti tiroidei insieme al GH è una pratica comune anche durante i protocolli di massa.
Il muscolo scheletrico è il principale bersaglio di segnalazione dell’ormone tiroideo, con trasportatori degli ormoni tiroidei e enzimi di conversione espressi localmente.[394] È ben noto che il GH potenzia la deiodinazione periferica che converte il T4 in T3, riducendo così il T4 e il reverse T3, aumentando contemporaneamente i livelli di T3.[395-398] Ciò che molte persone non riescono a capire è che questo è un effetto transitorio, e studi a lungo termine sembrano indicare che gli effetti mediati dal GH sulla conversione periferica si stabilizzino con il tempo.[399-402]
Via ubiquitina/proteasoma
Invece di proseguire ulteriormente su questo, avendo già trattato la questione nel dettaglio in un mio vecchio articolo, preferirei concentrarmi su alcune pubblicazioni relative alla tiroide che non vengono discusse abbastanza spesso. Gli ormoni tiroidei, per loro natura, sono composti tendenzialmente catabolici in quanto stimolano la disgregazione proteica dell’intero corpo in misura maggiore rispetto alla sintesi proteica.[403] A livello locale, nel muscolo scheletrico stimolano un aumento dell’attività all’interno della via ubiquitina/proteasoma, che è ampiamente coinvolta nella proteolisi.[404-406] Il risultato di questo è un tasso accelerato del turnover proteico e una perdita netta complessiva degli aminoacidi situati all’interno dei muscoli scheletrici.
Inoltre, negli esseri umani, sia gli stati di ipertiroidismo che di ipotiroidismo sono stati associati a livelli di IGF-1 soppressi con una tendenza alla normalizzazione quando viene ristabilita una condizione di eutiroidismo. L’ipertiroidismo è anche associato ad una bassa attività di legame recettoriale del GH, che si ipotizza essere il risultato di una ridotta capacità di elaborazione dei recettori del GH.[407] E’ stato anche ipotizzato che l’ipertiroidismo sia in grado anche di accelerare la clearance del GH urinario.[408] Inoltre, studi su animali hanno dimostrato che gli ormoni tiroidei possono avere importanti effetti soppressivi sulla sintesi di IGF-1 stimolata con il GH.[409] Ovviamente, a causa della complessa relazione che l’Asse Tiroideo ha con l’Asse GH/IGF-1, raggruppando tutte le interazioni che hanno tra loro in pochi paragrafi, trattare l’argomento diventerebbe poco pratico. Tuttavia, quando il corpo della letteratura scientifica viene esaminato nella sua interezza, ci sono molte prove che suggeriscono che la supplementazione con composti tiroidei esogeni potrebbe non essere l’ideale quando l’obiettivo di un individuo è l’ipertrofia, anche se la regolazione del dosaggio dei tiroidei in tale contesto rimane la misura di “sicurezza” più intelligente visto l’impatto negativo sulla funzionalità tiroidea dato dal GH. Comunque, per tutti coloro che sono interessati ad approfondire questo argomento, consiglio di iniziare con la review nella nota seguente.[410]
Miostatina
Mi piacerebbe trattare anche la Miostatin, che rappresenta un argomento molto discusso nei vari forum di BodyBuilding presenti in rete. La sua fama proviene dai risultati ipertrofici espressi dai bovini privi per mutazione del gene della Miostatina, i quali mostrano una massa muscolare significativamente maggiore rispetto ai loro simili non mutati.[411] La Miostatina, un fattore di crescita e differenziazione appartenente alla superfamiglia dei TGF-beta, ha dimostrato di inibire selettivamente la miogenesi, in gran parte tramite il suo effetto soppressivo sulla proliferazione dei mioblasti.[412] È espressa e secreta prevalentemente dal muscolo scheletrico. Come molti sanno, se riesci a sopprimere o inibire la Miostatina, di conseguenza il potenziale ipertrofico aumenta significativamente.
Le mutazioni della Miostatina sono state osservate sia negli animali che nell’uomo. Queste mutazioni del gene della Miostatina portano ad un fenotipo ipertrofico negli animali, come accennato in precedenza.[413-415] L’Asse GH/IGF-1 e la Miostatina sembrano avere una relazione regolativa diretta tra loro, come osservato nei pazienti affetti contemporaneamente da GHD e HIV che mostrano marcati aumenti nell’espressione dell’mRNA della Miostatina.[416] Quindi, è possibile che attraverso una supplementazione di dosi sovrafisiologiche di rHGH si possa indurre una diminuzione dell’mRNA della Miostatina [209,417-419]? Sfortunatamente, nonostante la presenza di alcuni casi studio selezionati, non credo che si abbiano abbastanza dati in questo momento per sapere se ciò possa dare risultati apprezzabili in seguito alla sua applicazione.
Quello che sappiamo è che aumenti dell’espressione dell’mRNA dell’IGF-1 e le concentrazioni circolanti di IGF-1 sono state osservati dopo inibizione della Miostatina.[419-421] Sappiamo anche che l’inibizione della Miostatina tende a causare l’ipertrofia attraverso molte delle stesse modalità osservate con l’IGF-1 autocrino, cioè l’aumento della sintesi proteica e l’attivazione delle cellule satelliti.[422-425] E sappiamo anche che l’ipertrofia indotta dalla sovraespressione dell’IGF-1 o dall’inibizione della Miostatina utilizza la stessa identica via – PI3K/Akt/mTOR.[426-428] Tuttavia, l’IGF-1 non è un requisito per l’ipertrofia indotta dalla Follistatina, tranne nel caso di livelli di Insulina estremamente bassi – come ben sappiamo, la Follistatina è un inibitore della Miostatina [429]. E l’esposizione cronica al GH può in realtà portare ad un’espressione sovrastimolata della Miostatina e del suo recettore.[209]
Quindi quello che possiamo dire, con certezza, è che l’espressione della Miostatina non sarà un fattore diretto o indiretto per quanto riguarda il potenziamento dei processi ipertrofici, né dell’attività contrattile, nei muscoli scheletrici umani.[430] Proprio per questo motivo, non ritengo che sia un fattore sul quale gli atleti debbano eccessivamente concentrarsi, al di fuori di un utile arricchimento delle proprie conoscenze in materia.
Applicazioni pratiche e pensieri conclusivi
Arrivati a questo punto è mia intenzione unire tutto ciò che è stato esposto in questa serie di articoli e esporlo sotto forma di alcuni suggerimenti pratici rivolti a tutti coloro i quali vogliono semplicemente massimizzare la loro capacità ipertrofica.
Ora è chiaro che il GH possiede pochissimi, se non nulli, effetti diretti sull’ipertrofia. Pertanto, qualsiasi protocollo di massa che contempli il suo uso dovrà tenere in considerazione questo punto includendo gli AAS, i quali, per l’appunto, hanno anche una proficua sinergia con il GH. Sia la letteratura scientifica che i dati aneddotici dimostrano chiaramente che l’uso combinato di entrambi i composti ha un massimale ipertrofico significativamente più alto rispetto all’uso singolo. Personalmente, penso che i BodyBuilder dovrebbero sempre optare per l’utilizzo di una base di Testosterone e Boldenone (correttamente rapportati in base a contesto e alle caratteristiche individuali) anche in una fase “Bulk” nella quale viene inserito l’uso del GH. Il Trenbolone può essere considerato come parte “accessoria” di un protocollo di massa, a causa della sua intrinseca difficoltà gestazionale come sostanza anabolizzante. Dovrebbe essere usato con parsimonia e con cautela poiché, insieme ai suoi numerosi punti di forza come composto anabolizzante, presenta alcune limitazioni. Queste limitazioni derivano per lo più, come già accennato, dalla difficile gestione del composto in quanto esso non è facilmente tollerato da una buona parte degli individui. Quindi, se viene usato il Trenbolone, dovrebbe essere inserito calcolando con attenzione il dosaggio e la tolleranza individuale, anche per quanto concerne la tolleranza temporale individuale all’uso di tale composto.
Dopo un periodo d’uso prolungato di dosi sovrafisiologico di AAS (Ciclo+Bridge), è buona cosa procedere con l’interruzione o con una marcata riduzione del numero e degli AAS utilizzati. Detta in parole semplici, questa interruzione può contemplare una completa astinenza dagli AAS, svolgendo un adeguata PCT al fine di ristabilire una omeostasi ormonale fisiologica, o una transizione ad una TRT, metodologia comunemente chiamata “blast and cruise”. La struttura del protocollo di supplementazione farmacologica dovrebbe sempre seguire i principi del dosaggio minimo efficace con aumenti nei dosaggi degli AAS solo nel caso si sia raggiunto il limite di crescita con il precedente dosaggio, assicurandosi che tutte le altre variabili nello stile di vita siano correttamente regolate. L’utilizzo di questo approccio limita il rischio che si sviluppino effetti collaterali indesiderati sul lungo termine.
Quando si decide di usare il GH, la dove ce ne sia la possibilità economica, esso dovrebbe provenire da uno dei prodotti presenti nel mercato farmaceutico. Questi prodotti approvati devono superare anni di studi strettamente controllati per dimostrare la loro sicurezza, purezza ed efficacia su soggetti umani. I progressi tecnologici nel corso degli anni hanno reso molto più facile la produzione di rHGH. Per questo motivo, i produttori ora provengono da tutto il mondo. Spesso questi produttori realizzano ciò che viene definito “GH generico” sui forum, ma tale termine non mi piace molto. Definire qualcosa come “generico” implica che sia una replica perfetta di prodotti legati a specifici marchi farmaceutici approvati dell’agenzia del farmaco che hanno perso il cui brevetto è scaduto, il che non è il caso del GH. Infatti, a causa della natura estremamente complessa del processo di produzione del rHGH, per esempio, la FDA non consente nemmeno l’uso del termine “generico” quando si tratta di rHGH e utilizza invece il termine ” follow-on protein product ” o FOPP.
Spesso questi marchi off-label sono venduti ad un costo molto ridotto, ed è qui che sta il dilemma, in quanto questo può essere molto allettante. Tuttavia, con questo costo ridotto per il consumatore, non ci sarà nemmeno la garanzia del produttore su cosa ci sia realmente nella fiala o persino su come è stato prodotto. Il problema di fondo è che il processo di produzione del rHGH è estremamente complessa, ed è molto facile che nelle fasi di questo processo si commettano errori con conseguenti variazioni nella catena proteica che potenzialmente portano ad effetti indesiderati, o anche a risposte autoimmuni.
Spesso gli atleti si affidano semplicemente ai test serici per misurare i livelli di GH e/o IGF-1 al fine di concludere che un prodotto contenete GH sia “buono”, ma dobbiamo ricordarci che raggiungere livelli ematici ormonali elevati è la parte relativamente facile. Anche le molecole di GH che sono state alterate o danneggiate durante la produzione possono dare questo esito. Tuttavia, queste stesse molecole di GH danneggiate o mutate possono spesso stimolare risposte autoimmuni. Ciò potrebbe indurre il corpo ad avere una risposta recettoriale degradata, che può anche riflettersi sulla secrezione endogena nel tempo. [431-432] Rimane poi il problema del reale contenuto della vial o fiala, la quale può non presentare nessun principio attivo al suo interno.
Il GH dovrebbe essere usato in modo pulsatile, per mimare le condizioni in vivo. Tra queste iniezioni, deve esserci un periodo di refrattarietà o si deve consumare un pasto che abbia un buon stimolo sull’Insulina. Può anche essere utilizzata l’Insulina esogena al fine di bypassare molte delle limitazioni del periodo refrattario, ma questo va oltre lo scopo di questo articolo. Anche se il cumulo delle dosi giornaliere dovrebbe essere sovrafisiologico, le dosi individuali non hanno bisogno di essere ad alto dosaggio, poiché la massima stimolazione dell’IGF-1 autocrino nel tessuto muscolo scheletrico si verifica ben all’interno delle concentrazioni fisiologiche di GH. Aneddoticamente, sembra anche esserci un limite con il quale l’uso di rHGH diventa additivo in presenza di AAS. Potrebbe essere necessario un po’ di ponderata (e supportata da personale qualificato) auto-sperimentazione per scoprire dove si trova questa singola dose di saturazione, ma la maggior parte dei soggetti troverà questo limite tra le 4 e le 8 UI/die. Oltre questo dosaggio, la maggior parte degli utilizzatori tenderà a scoprire che la giustificazione dei costi e il rapporto rischio /beneficio tendono a diminuire rapidamente.
Non bisognerebbe passare troppo tempo a riflettere sul tempo delle iniezioni di GH, dal momento che gli aumenti dei livelli di IGF-1 autocrino avvengono rapidamente e possono rimanere elevati per giorni. Bisognerebbe concentrati invece sul programma di iniezione che si addice meglio al contesto della giornata, tenendo contemporaneamente presenti le linee guida per il periodo di refrattarietà del GH. Si possono anche prendere in considerazione piccole o grandi iniezioni, poiché alcuni potrebbero trovare più pratiche e funzionali iniezioni con un dosaggio inferiore e somministrate più frequenti mentre altri potrebbero preferire iniezioni con un dosaggio maggiore e somministrazioni meno frequenti. Naturalmente, maggiore è il contenuto dell’iniezione, maggiore è la probabilità che si superi la soglia massima di espressione dell’IGF-1 autocrino.
Massimizzare l’espressione dell’IGF-1 autocrino, mentre contemporaneamente si sopprimono i livelli di IGF-1 endocrino, sarà una priorità. Esistono prove a sostegno dell’ipotesi secondo cui un iniezione locale di GH possa aiutare a raggiungere questo obiettivo, con una conseguente minore possibilità di feedback negativo. Sono stati osservati aumenti significativi della massa muscolare in appena due settimane di iniezioni locali di IGF-1.[441]
La dove cause di forza maggiore non lo impediscano, è consigliabile evitare o comunque regolare attentamente l’uso di tutti quei composti che possono avere interazioni negative con l’uso del GH a fini ipertrofici. Inibitori della Aromatasi, Modulatori Selettivi del Recettore degli Estrogeni e T3 hanno tutti dimostrato di avere un potenziale effetto negativo sul processo globale ipertrofico legato al GH e per tale motivo dovrebbero essere usati con parsimonia, se non omessi del tutto dove possibile (vedi in particolare il T3).
Questo non dovrebbe sorprendere nessuno e non dovrebbe nemmeno essere troppo difficile da tenere a mente: allenarsi duramente, allenarsi in modo intelligente e allenarsi in modo coerente. Sebbene non sia stato affrontato direttamente nell’articolo, bisogna capire che l’allenamento contro resistenza ha impatti unici e additivi sull’ipertrofia. In effetti, alcuni di questi meccanismi non sono nemmeno mediati dall’Asse AR e/o GH/IGF-1. [433] Bisogna anche comprendere che non esiste una “magica” routine allenante universalmente applicabile, la chiave di volta sarà la coerenza nel garantire un carico di lavoro adeguato, con elementi di sovraccarico progressivo nel tempo assicurando un adeguato stimolo meccanico gestendo al meglio le variabili allenanti (intensità, volume, densità e intensità percepita). La logica composizione del piano allenante servirà a garantire lo stimolo ipertrofico di base che sarà coadiuvato dall’azione dei composti utilizzati.
Nonostante la mole e l’importanza delle informazioni presentate in questa serie di articoli, è necessario ricordare che i meccanismi d’azione ormonali sono governati da innumerevoli fattori. Anche esaminando l’intero corpo della letteratura scientifica equivarrebbe a poco più che accumulare una serie di utili nozioni garanti di assicurare una solida base conoscitiva sull’argomento la quale rappresenterà un punto di partenza intelligente per l’applicazione pratica, rimanendo sempre soggetti alle variabili di risposta individuale. Seguendo questa linea, i migliori risultati nella pratica spesso provengono da coloro i quali posseggono una buona conoscenza dei principi scientifici e la capacità innata di saperli applicare non solo su se stessi ma, soprattutto, su terzi. Infatti, molto raramente due persone rispondono in modo identico alla supplementazione di ormoni esogeni (e non solo), quindi, non bisogna assolutamente pensare che basti semplicemente applicare su se stessi o su terzi un protocollo che ha portato benefici realmente apprezzabili su un soggetto per ottenere la medesima risposta.
A tal fine, invito atleti e Preparatori a utilizzare queste pubblicazioni come punto di partenza per essere in grado di gestire l’applicazione pratica in modo più consapevole e produttivo. Inoltre, chi ne fosse in grado, può consultare il vasto numero di riferimenti riportati nel corso di queste pubblicazioni è tentare di ragionare sulle mie conclusioni. Ad ogni citazione presente in questa serie di articoli, assicuratevi che il riferimento elencato supporti effettivamente le affermazioni fatte. Mantenere sempre una mente aperta ma con i giusti “filtri”, e cercare di non credere per partito preso ad una singola opinione, specialmente di fronte alle nuove evidenze scientifiche. Infine, è buona cosa verificare sempre la veridicità di quanto è stato affermato.
Punti conclusivi per un corretto utilizzo della chimica e del GH a fini ipertrofici:
Usare il GH in combinazione con gli AAS
Usare GH e AAS di grado farmaceutico la dove ciò è possibile
Assicurarsi una base di Testosterone correttamente rapportata al Boldenone aggiungendo (in base a maturità e tolleranza) il Trenbolone
Iniettare il GH in modo pulsatile, considerare l’opzione delle iniezioni locali nei gruppi carenti
Mantenere un dosaggio complessivo ottimale di GH il quale si attesta tra le 4 e le 8UI/die
Evitare o regolare attentamente l’uso dei composti che possono interagire negativamente con i processi ipertrofici legati al uso di GH (vedi AI, SERM e T3)
Dopo un periodo di tempo (variabile) nel quale si è stati sottoposti a dosaggi ormonali sovrafisiologici optare per una PCT o per una TRT (a seconda delle proprie necessità e priorità)
Essere a conoscenza dei potenziali effetti collaterali legati all’auso/abuso di GH (nausea, vomito, cefalea, ritenzione idrica e sodica, edemi, parestesie, sindrome del tunnel carpale, rigidità articolare, dolori articolari, artrite, dolori muscolari, ipertensione, insulino-resistenza, diabete di tipo II, acromegalia, dilatazione addominale, ipertrofia cardiaca ecc…)
Svolgere regolarmente esami del sangue; sia durante i periodi di picco nell’uso della farmacologia sia nel periodo successivo (vedi PCT/OCT o TRT)
Gestire al meglio le variabili legate agli stressor ambientali, all’allenamento, all’alimentazione e al sonno.
Gabriel Bellizzi
Riferimenti:
338. Mathews LS, Norstedt G, Palmiter RD. Regulation of insulin-like growth factor I gene expression by growth hormone. Proc Natl Acad Sci U S A. 1986 Dec;83(24):9343-7.
339. Keller A, Wu Z, Kratzsch J, Keller E, Blum WF, Kniess A, Preiss R, Teichert J, Strasburger CJ, Bidlingmaier M. Pharmacokinetics and pharmacodynamics of GH: dependence on route and dosage of administration. Eur J Endocrinol. 2007 Jun;156(6):647-53.
340. Tanaka T, Seino Y, Fujieda K, Igarashi Y, Yokoya S, Tachibana K, Ogawa Y. Pharmacokinetics and metabolic effects of high-dose growth hormone administration in healthy adult men. Endocr J. 1999 Aug;46(4):605-12.
341. Quinn LS, Steinmetz B, Maas A, Ong L, Kaleko M. Type-1 insulin-like growth factor receptor overexpression produces dual effects on myoblast proliferation and differentiation. J Cell Physiol. 1994 Jun;159(3):387-98.
342. Coolican SA, Samuel DS, Ewton DZ, McWade FJ, Florini JR. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J Biol Chem. 1997 Mar 7;272(10):6653-62.
343. Foulstone EJ, Huser C, Crown AL, Holly JM, Stewart CE. Differential signalling mechanisms predisposing primary human skeletal muscle cells to altered proliferation and differentiation: roles of IGF-I and TNFalpha. Exp Cell Res. 2004 Mar 10;294(1):223-35.
344. Ewton DZ, Roof SL, Magri KA, McWade FJ, Florini JR. IGF-II is more active than IGF-I in stimulating L6A1 myogenesis: greater mitogenic actions of IGF-I delay differentiation. J Cell Physiol. 1994 Nov;161(2):277-84.
345. Florini JR, Nicholson ML, Dulak NC. Effects of peptide anabolic hormones on growth of myoblasts in culture. Endocrinology. 1977 Jul;101(1):32-41.
346. Laviola L, Natalicchio A, Giorgino F. The IGF-I signaling pathway. Curr Pharm Des. 2007;13(7):663-9. Review.
347. Jacquemin V, Furling D, Bigot A, Butler-Browne GS, Mouly V. IGF-1 induces human myotube hypertrophy by increasing cell recruitment. Exp Cell Res. 2004 Sep 10;299(1):148-58.
348. Jacquemin V, Butler-Browne GS, Furling D, Mouly V. IL-13 mediates the recruitment of reserve cells for fusion during IGF-1-induced hypertrophy of human myotubes. J Cell Sci. 2007 Feb 15;120(Pt 4):670-81. Epub 2007 Jan 30.
349. Ballard FJ, Francis GL. Effects of anabolic agents on protein breakdown in L6 myoblasts. Biochem J. 1983 Jan 15;210(1):243-9.
350. Ewton DZ, Falen SL, Florini JR. The type II insulin-like growth factor (IGF) receptor has low affinity for IGF-I analogs: pleiotypic actions of IGFs on myoblasts are apparently mediated by the type I receptor. Endocrinology. 1987 Jan;120(1):115-23.
351. Hembree JR, Hathaway MR, Dayton WR. Isolation and culture of fetal porcine myogenic cells and the effect of insulin, IGF-I, and sera on protein turnover in porcine myotube cultures. J Anim Sci. 1991 Aug;69(8):3241-50.
352. Hong D, Forsberg NE. Effects of serum and insulin-like growth factor I on protein degradation and protease gene expression in rat L8 myotubes. J Anim Sci. 1994 Sep;72(9):2279-88.
353. Florini JR, Ewton DZ, Roof SL. Insulin-like growth factor-I stimulates terminal myogenic differentiation by induction of myogenin gene expression. Mol Endocrinol. 1991 May;5(5):718-24.
354. Musarò A, McCullagh KJ, Naya FJ, Olson EN, Rosenthal N. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature. 1999 Aug 5;400(6744):581-5.
355. Haba GDL, Cooper GW, Elting V. HORMONAL REQUIREMENTS FOR MYOGENESIS OF STRIATED MUSCLE IN VITRO: INSULIN AND SOMATOTROPIN. Proceedings of the National Academy of Sciences of the United States of America. 1966;56(6):1719-1723.
356. Florini JR, Ewton DZ. Insulin acts as a somatomedin analog in stimulating myoblast growth in serum-free medium. In Vitro. 1981 Sep;17(9):763-8.
357. Schmid C, Steiner T, Froesch ER. Preferential enhancement of myoblast differentiation by insulin-like growth factors (IGF I and IGF II) in primary cultures of chicken embryonic cells. FEBS Lett. 1983 Sep 5;161(1):117-21.
358. Florini JR, Ewton DZ, Falen SL, Van Wyk JJ. Biphasic concentration dependency of stimulation of myoblast differentiation by somatomedins. Am J Physiol. 1986 May;250(5 Pt 1):C771-8.
359. Quinn LS, Ehsan M, Steinmetz B, Kaleko M. Ligand-dependent inhibition of myoblast differentiation by overexpression of the type-1 insulin-like growth factor receptor. J Cell Physiol. 1993 Sep;156(3):453-61.
360. Olson EN. Signal transduction pathways that regulate skeletal muscle gene expression. Mol Endocrinol. 1993 Nov;7(11):1369-78. Review.
361. Murphy LJ, Bell GI, Friesen HG. Growth hormone stimulates sequential induction of c-myc and insulin-like growth factor I expression in vivo. Endocrinology. 1987 May;120(5):1806-12.
362. Turner JD, Rotwein P, Novakofski J, Bechtel PJ. Induction of mRNA for IGF-I and -II during growth hormone-stimulated muscle hypertrophy. Am J Physiol. 1988 Oct;255(4 Pt 1):E513-7.
363. Isgaard J, Nilsson A, Vikman K, Isaksson OG. Growth hormone regulates the level of insulin-like growth factor-I mRNA in rat skeletal muscle. J Endocrinol. 1989 Jan;120(1):107-12.
364. Bichell DP, Kikuchi K, Rotwein P. Growth hormone rapidly activates insulin-like growth factor I gene transcription in vivo. Mol Endocrinol. 1992 Nov;6(11):1899-908.
365. Sadowski CL, Wheeler TT, Wang LH, Sadowski HB. GH regulation of IGF-I and suppressor of cytokine signaling gene expression in C2C12 skeletal muscle cells. Endocrinology. 2001 Sep;142(9):3890-900.
366. Frost RA, Nystrom GJ, Lang CH. Regulation of IGF-I mRNA and signal transducers and activators of transcription-3 and -5 (Stat-3 and -5) by GH in C2C12 myoblasts. Endocrinology. 2002 Feb;143(2):492-503.
367. MacLeod JN, Pampori NA, Shapiro BH. Sex differences in the ultradian pattern of plasma growth hormone concentrations in mice. J Endocrinol. 1991 Dec;131(3):395-9.
368. Rochiccioli P, Messina A, Tauber MT, Enjaume C. Correlation of the parameters of 24-hour growth hormone secretion with growth velocity in 93 children of varying height. Horm Res. 1989;31(3):115-8.
369. Hansen TK, Gravholt CH, ØRskov H, Rasmussen MH, Christiansen JS, Jørgensen JO. Dose dependency of the pharmacokinetics and acute lipolytic actions of growth hormone. J Clin Endocrinol Metab. 2002 Oct;87(10):4691-8.
370. Baum WF, Klöditz E, Hesse V, Jahreis G, Schneyer U, Giebler H. [Increase in spontaneous growth hormone secretion in asthmatic children–a symptom of atopic disposition?]. Kinderarztl Prax. 1993 Nov;61(9):323-8.
371. Adams GR, McCue SA. Localized infusion of IGF-I results in skeletal muscle hypertrophy in rats. J Appl Physiol (1985). 1998 May;84(5):1716-22.
372. Alzghoul MB, Gerrard D, Watkins BA, Hannon K. Ectopic expression of IGF-I and Shh by skeletal muscle inhibits disuse-mediated skeletal muscle atrophy and bone osteopenia in vivo. FASEB J. 2004 Jan;18(1):221-3. Epub 2003 Nov 3.
373. Lee S, Barton ER, Sweeney HL, Farrar RP. Viral expression of insulin-like growth factor-I enhances muscle hypertrophy in resistance-trained rats. J Appl Physiol (1985). 2004 Mar;96(3):1097-104. Erratum in: J Appl Physiol. 2004 Jun;96(6):2343.
374. Coleman ME, DeMayo F, Yin KC, Lee HM, Geske R, Montgomery C, Schwartz RJ. Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J Biol Chem. 1995 May 19;270(20):12109-16.
375. Barton-Davis ER, Shoturma DI, Musaro A, Rosenthal N, Sweeney HL. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proc Natl Acad Sci U S A. 1998 Dec 22;95(26):15603-7.
376. Musarò A, McCullagh K, Paul A, Houghton L, Dobrowolny G, Molinaro M, Barton ER, Sweeney HL, Rosenthal N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat Genet. 2001 Feb;27(2):195-200
377. Lewis MI, Bulut Y, Biring MS, Da X, Fournier M. (1999) IGF-I administration prevents corticosteroids-induced diaphragm atrophy in emphysema . Am J Respir Crit Care Med 159:A580
378. Fournier M, Huang ZS, Cercek B, Li H, Bykhovskaya I, Lewis MI. (2000) Administration of insulin-like growth factor-1 (IGF-I) and corticosteroids in emphysematous hamsters: influences on diaphragm IGF-I . Am J Respir Crit Care Med 161:A18
379. Shavlakadze T, Grounds M. Of bears, frogs, meat, mice and men: complexity of factors affecting skeletal muscle mass and fat. Bioessays. 2006 Oct;28(10):994-1009. Review.
380. Maiter D, Underwood LE, Maes M, Davenport ML, Ketelslegers JM. Different effects of intermittent and continuous growth hormone (GH) administration on serum somatomedin-C/insulin-like growth factor I and liver GH receptors in hypophysectomized rats. Endocrinology. 1988 Aug;123(2):1053-9.
381. Isgaard J, Carlsson L, Isaksson OG, Jansson JO. Pulsatile intravenous growth hormone (GH) infusion to hypophysectomized rats increases insulin-like growth factor I messenger ribonucleic acid in skeletal tissues more effectively than continuous GH infusion. Endocrinology. 1988 Dec;123(6):2605-10.
382. Clark RG, Jansson JO, Isaksson O, Robinson IC. Intravenous growth hormone: growth responses to patterned infusions in hypophysectomized rats. J Endocrinol. 1985 Jan;104(1):53-61.
383. Bick T, Hochberg Z, Amit T, Isaksson OG, Jansson JO. Roles of pulsatility and continuity of growth hormone (GH) administration in the regulation of hepatic GH-receptors, and circulating GH-binding protein and insulin-like growth factor-I. Endocrinology. 1992 Jul;131(1):423-9.
384. Weltman A, Weltman JY, Schurrer R, Evans WS, Veldhuis JD, Rogol AD. Endurance training amplifies the pulsatile release of growth hormone: effects of training intensity. J Appl Physiol (1985). 1992 Jun;72(6):2188-96.
385. Flores-Morales A, Greenhalgh CJ, Norstedt G, Rico-Bautista E. Negative regulation of growth hormone receptor signaling. Mol Endocrinol. 2006 Feb;20(2):241-53. Epub 2005 Jul 21. Review.
386. Hartman ML, Veldhuis JD, Thorner MO. Normal control of growth hormone secretion. Horm Res. 1993;40(1-3):37-47. Review.
387. Fernández L, Flores-Morales A, Lahuna O, Sliva D, Norstedt G, Haldosén LA, Mode A, Gustafsson JA. Desensitization of the growth hormone-induced Janus kinase 2 (Jak 2)/signal transducer and activator of transcription 5 (Stat5)-signaling pathway requires protein synthesis and phospholipase C. Endocrinology. 1998 Apr;139(4):1815-24.
388. Gebert CA, Park SH, Waxman DJ. Termination of growth hormone pulse-induced STAT5b signaling. Mol Endocrinol. 1999 Jan;13(1):38-56.
389. Ram PA, Waxman DJ. SOCS/CIS protein inhibition of growth hormone-stimulated STAT5 signaling by multiple mechanisms. J Biol Chem. 1999 Dec 10;274(50):35553-61.
390. Ram PA, Waxman DJ. Role of the cytokine-inducible SH2 protein CIS in desensitization of STAT5b signaling by continuous growth hormone. J Biol Chem.2000 Dec 15;275(50):39487-96.
391. Xu J, Keeton AB, Franklin JL, Li X, Venable DY, Frank SJ, Messina JL. Insulin enhances growth hormone induction of the MEK/ERK signaling pathway. J Biol Chem. 2006 Jan 13;281(2):982-92. Epub 2005 Nov 4.
392. Lewis TS, Shapiro PS, Ahn NG. Signal transduction through MAP kinase cascades. Adv Cancer Res. 1998;74:49-139. Review.
394. Mebis L, Paletta D, Debaveye Y, Ellger B, Langouche L, D’Hoore A, Darras VM, Visser TJ, Van den Berghe G. Expression of thyroid hormone transporters during critical illness. Eur J Endocrinol. 2009 Aug;161(2):243-50.
395. Jørgensen JO, Pedersen SA, Laurberg P, Weeke J, Skakkebaek NE, Christiansen JS. Effects of growth hormone therapy on thyroid function of growth hormone-deficient adults with and without concomitant thyroxine-substituted central hypothyroidism. J Clin Endocrinol Metab. 1989 Dec;69(6):1127-32.
396. Jørgensen JO, Pedersen SB, Børglum J, Møller N, Schmitz O, Christiansen JS, Richelsen B. Fuel metabolism, energy expenditure, and thyroid function in growth hormone-treated obese women: a double-blind placebo-controlled study. Metabolism. 1994 Jul;43(7):872-7.
397. Wolthers T, Grøftne T, Møller N, Christiansen JS, Orskov H, Weeke J, Jørgensen JO. Calorigenic effects of growth hormone: the role of thyroid hormones. J Clin Endocrinol Metab. 1996 Apr;81(4):1416-9.
398. Feldt-Rasmussen U. Interactions between growth hormone and the thyroid gland — with special reference to biochemical diagnosis. Curr Med Chem. 2007;14(26):2783-8. Review.
399. Kalina-Faska B, Kalina M, Koehler B. Effects of recombinant growth hormone therapy on thyroid hormone concentrations. Int J Clin Pharmacol Ther. 2004 Jan;42(1):30-4.
400. Hubina E, Mersebach H, Rasmussen AK, Juul A, Sneppen SB, Góth MI, Feldt-Rasmussen U. Effect of growth hormone replacement therapy on pituitary hormone secretion and hormone replacement therapies in GHD adults. Horm Res. 2004;61(5):211-7. Epub 2004 Jan 30.
401. Seminara S, Stagi S, Candura L, Scrivano M, Lenzi L, Nanni L, Pagliai F, Chiarelli F. Changes of thyroid function during long-term hGH therapy in GHD children. A possible relationship with catch-up growth? Horm Metab Res. 2005 Dec;37(12):751-6.
402. Losa M, Scavini M, Gatti E, Rossini A, Madaschi S, Formenti I, Caumo A, Stidley CA, Lanzi R. Long-term effects of growth hormone replacement therapy on thyroid function in adults with growth hormone deficiency. Thyroid. 2008 Dec;18(12):1249-54.
403. Müller MJ, Seitz HJ. Thyroid hormone action on intermediary metabolism. Part III. Protein metabolism in hyper- and hypothyroidism. Klin Wochenschr. 1984 Feb 1;62(3):97-102.
404. Tawa NE Jr, Odessey R, Goldberg AL. Inhibitors of the proteasome reduce the accelerated proteolysis in atrophying rat skeletal muscles. J Clin Invest. 1997 Jul 1;100(1):197-203. PubMed PMID: 9202072
405. Dace A, Zhao L, Park KS, et al. Hormone binding induces rapid proteasome-mediated degradation of thyroid hormone receptors. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(16):8985-8990.
406. Clément K, Viguerie N, Diehn M, Alizadeh A, Barbe P, Thalamas C, Storey JD, Brown PO, Barsh GS, Langin D. In vivo regulation of human skeletal muscle gene expression by thyroid hormone. Genome Res. 2002 Feb;12(2):281-91.
407. Miell JP, Taylor AM, Zini M, Maheshwari HG, Ross RJ, Valcavi R. Effects of hypothyroidism and hyperthyroidism on insulin-like growth factors (IGFs) and growth hormone- and IGF-binding proteins. J Clin Endocrinol Metab. 1993 Apr;76(4):950-5.
408. Murao K, Takahara J, Sato M, Tamaki M, Niimi M, Ishida T. Relationship between thyroid functions and urinary growth hormone secretion in patients with hyper- and hypothyroidism. Endocr J. 1994 Oct;41(5):517-22.
409. Wolf M, Ingbar SH, Moses AC. Thyroid hormone and growth hormone interact to regulate insulin-like growth factor-I messenger ribonucleic acid and circulating levels in the rat. Endocrinology. 1989 Dec;125(6):2905-14.
410. Laron Z. Interactions between the thyroid hormones and the hormones of the growth hormone axis. Pediatr Endocrinol Rev. 2003 Dec;1 Suppl 2:244-9-discussion 250. Review.
411. Fiems LO. Double Muscling in Cattle: Genes, Husbandry, Carcasses and Meat. Animals : an Open Access Journal from MDPI. 2012;2(3):472-506.
412. Langley B, Thomas M, Bishop A, Sharma M, Gilmour S, Kambadur R. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. J Biol Chem. 2002 Dec 20;277(51):49831-40. Epub 2002 Sep 18.
413. McPherron AC, Lee SJ. Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci U S A. 1997 Nov 11;94(23):12457-61.
414. Schuelke M, Wagner KR, Stolz LE, Hübner C, Riebel T, Kömen W, Braun T, Tobin JF, Lee SJ. Myostatin mutation associated with gross muscle hypertrophy in a child. N Engl J Med. 2004 Jun 24;350(26):2682-8.
415. Clop A, Marcq F, Takeda H, Pirottin D, Tordoir X, Bibé B, Bouix J, Caiment F, Elsen JM, Eychenne F, Larzul C, Laville E, Meish F, Milenkovic D, Tobin J, Charlier C, Georges M. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat Genet. 2006 Jul;38(7):813-8.
416. Gonzalez-Cadavid NF, Taylor WE, Yarasheski K, Sinha-Hikim I, Ma K, Ezzat S, Shen R, Lalani R, Asa S, Mamita M, Nair G, Arver S, Bhasin S. Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting. Proc Natl Acad Sci U S A. 1998 Dec 8;95(25):14938-43.
417. Liu W, Thomas SG, Asa SL, Gonzalez-Cadavid N, Bhasin S, Ezzat S. Myostatin is a skeletal muscle target of growth hormone anabolic action. J Clin Endocrinol Metab. 2003 Nov;88(11):5490-6.
418. Oldham JM, Osepchook CC, Jeanplong F, Falconer SJ, Matthews KG, Conaglen JV, Gerrard DF, Smith HK, Wilkins RJ, Bass JJ, McMahon CD. The decrease in mature myostatin protein in male skeletal muscle is developmentally regulated by growth hormone. J Physiol. 2009 Feb 1;587(3):669-77.
419. Williams NG, Interlichia JP, Jackson MF, Hwang D, Cohen P, Rodgers BD. Endocrine actions of myostatin: systemic regulation of the IGF and IGF binding protein axis. Endocrinology. 2011 Jan;152(1):172-80.
420. Winbanks CE, Weeks KL, Thomson RE, Sepulveda PV, Beyer C, Qian H, Chen JL, Allen JM, Lancaster GI, Febbraio MA, Harrison CA, McMullen JR, Chamberlain JS, Gregorevic P. Follistatin-mediated skeletal muscle hypertrophy is regulated by Smad3 and mTOR independently of myostatin. J Cell Biol. 2012 Jun 25;197(7):997-1008.
421. Lach-Trifilieff E, Minetti GC, Sheppard K, Ibebunjo C, Feige JN, Hartmann S, Brachat S, Rivet H, Koelbing C, Morvan F, Hatakeyama S, Glass DJ. An antibody blocking activin type II receptors induces strong skeletal muscle hypertrophy and protects from atrophy. Mol Cell Biol. 2014 Feb;34(4):606-18.
422. Bark TH, McNurlan MA, Lang CH, Garlick PJ. Increased protein synthesis after acute IGF-I or insulin infusion is localized to muscle in mice. Am J Physiol. 1998 Jul;275(1 Pt 1):E118-23.
423. Barton-Davis ER, Shoturma DI, Sweeney HL. Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiol Scand. 1999 Dec;167(4):301-5.
424. Suryawan A, Frank JW, Nguyen HV, Davis TA. Expression of the TGF-beta family of ligands is developmentally regulated in skeletal muscle of neonatal rats. Pediatr Res. 2006 Feb;59(2):175-9.
425. Gilson H, Schakman O, Kalista S, Lause P, Tsuchida K, Thissen JP. Follistatin induces muscle hypertrophy through satellite cell proliferation and inhibition of both myostatin and activin. Am J Physiol Endocrinol Metab. 2009 Jul;297(1):E157-64.
426. Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, Zlotchenko E, Scrimgeour A, Lawrence JC, Glass DJ, Yancopoulos GD. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol. 2001 Nov;3(11):1014-9.
428. Kalista S, Schakman O, Gilson H, Lause P, Demeulder B, Bertrand L, Pende M, Thissen JP. The type 1 insulin-like growth factor receptor (IGF-IR) pathway is mandatory for the follistatin-induced skeletal muscle hypertrophy. Endocrinology. 2012 Jan;153(1):241-53.
429. Barbé C, Kalista S, Loumaye A, Ritvos O, Lause P, Ferracin B, Thissen JP. Role of IGF-I in follistatin-induced skeletal muscle hypertrophy. Am J Physiol Endocrinol Metab. 2015 Sep 15;309(6):E557-67. doi: 10.1152/ajpendo.00098.2015. Epub 2015 Jul 28.
430. Coffey VG, Shield A, Canny BJ, Carey KA, Cameron-Smith D, Hawley JA. Interaction of contractile activity and training history on mRNA abundance in skeletal muscle from trained athletes. Am J Physiol Endocrinol Metab. 2006 May;290(5):E849-55.
431. Moore WV, Leppert P. Role of aggregated human growth hormone (hGH) in development of antibodies to hGH. J Clin Endocrinol Metab. 1980 Oct;51(4):691-7
432. Dannies PS. Protein folding and deficiencies caused by dominant-negative mutants of hormones. Vitam Horm. 2000;58:1-26. Review.
433. DeVol DL, Rotwein P, Sadow JL, Novakofski J, Bechtel PJ. Activation of insulin-like growth factor gene expression during work-induced skeletal muscle growth. Am J Physiol. 1990 Jul;259(1 Pt 1):E89-95.
434. Hermansen K, Bengtsen M, Kjær M, Vestergaard P, Jørgensen JOL. Impact of GH administration on athletic performance in healthy young adults: A systematic review and meta-analysis of placebo-controlled trials. Growth Horm IGF Res. 2017 Jun;34:38-44.
435. de Souza GL, Hallak J. Anabolic steroids and male infertility: a comprehensive review. BJU Int. 2011 Dec;108(11):1860-5.
436. Kraemer WJ, Marchitelli L, Gordon SE, Harman E, Dziados JE, Mello R, Frykman P, McCurry D, Fleck SJ. Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol (1985). 1990 Oct;69(4):1442-50.
437. Pfeffer LA, Brisson BK, Lei H, Barton ER. The insulin-like growth factor (IGF)-I E-peptides modulate cell entry of the mature IGF-I protein. Mol Biol Cell. 2009 Sep;20(17):3810-7.
438. Mills P, Dominique JC, Lafrenière JF, Bouchentouf M, Tremblay JP. A synthetic mechano growth factor E Peptide enhances myogenic precursor cell transplantation success. Am J Transplant. 2007 Oct;7(10):2247-59.
439. Brisson BK, Barton ER. Insulin-like growth factor-I E-peptide activity is dependent on the IGF-I receptor. PLoS One. 2012;7(9):e45588.
440. Brisson BK, Spinazzola J, Park S, Barton ER. Viral expression of insulin-like growth factor I E-peptides increases skeletal muscle mass but at the expense of strength. Am J Physiol Endocrinol Metab. 2014 Apr 15;306(8):E965-74.
441. Goldspink G, Harridge S. Mechanism for adaptation in skeletal muscle In: Komi P, editor. Strength and power in sport: Olympic encyclopedia of sports medicine. Oxford: Blackwell; 2002. p. 231–51.
442. Janssen JA, Hofland LJ, Strasburger CJ, van den Dungen ES, Thevis M. Potency of Full-Length MGF to Induce Maximal Activation of the IGF-I R Is Similar to Recombinant Human IGF-I at High Equimolar Concentrations. PLoS One. 2016 Mar 18;11(3):e0150453.
Nei primi anni del XXI secolo, negli scaffali dei negozi di integratori (fisici o online) emersero alcuni nuovi supplementi e tra questi emerse l’Acido Tetradeciltioacetico [TTA]. In quel periodo questo particolare composto era uno dei principi attivi, insieme a molte altre sostanze, di alcuni controversi supplementi per la perdita di grasso. La maggior parte di questi prodotti è scomparsa dal mercato, ma l’Acido Tetradeciltiocetico è stato recentemente reinserito nel mercato. Ora, spesso in concentrazioni significativamente più elevate rispetto a un decennio fa, il TTA è spesso l’unico ingrediente attivo presente nei supplementi che lo contengono. E alcuni produttori di integratori vendono tale composto sotto forma di polvere. È quindi interessante approfondire le caratteristiche del composto e di cosa è ipoteticamente in grado di fare.
L’Acido Tetradeciltioacetico è un acido grasso sintetico che non può essere convertito in energia.
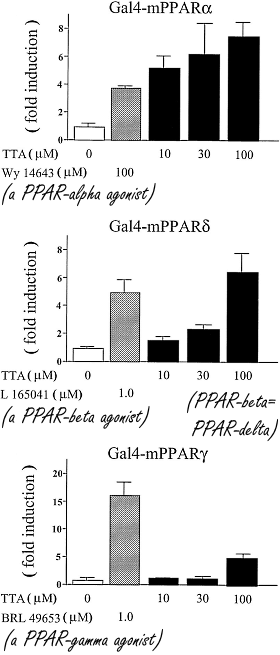
Negli anni ’90 gli scienziati scoprirono che l’Acido Tetradeciltioacetico stimolava il metabolismo dei grassi attivando i PPAR-α. I ricercatori hanno scoperto anche che, attraverso lo stesso meccanismo, la sostanza era in grado di inibire l’infiammazione, la crescita delle cellule tumorali e lo sviluppo/peggioramento dell’insulino-resistenza.
Esistono tre tipi di recettori PPAR: α, γ e δ. Il recettore α è espresso a livello epatico. Quando viene attivato, il fegato utilizza più acidi grassi per produrre energia. Il recettore γ è espresso nelle cellule adipose. Se viene stimolato, la cellula adiposa immagazzina acidi grassi. Il recettore δ è espresso a livello muscolare. Se il recettore δ viene stimolato, i muscoli utilizzano maggiormente gli acidi grassi per produrre energia.
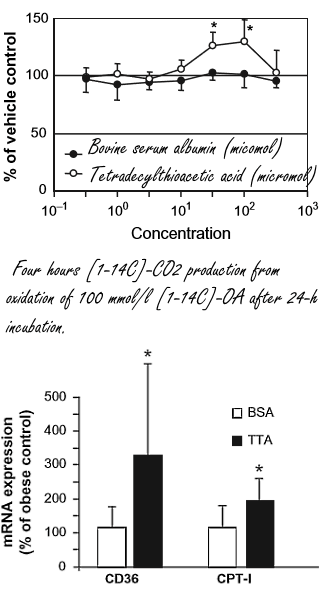
Nel 2002, scienziati norvegesi hanno riferito che gli animali presi in esame, e che avevano ricevuto una dose significativa di Acido Tetradeciltioacetico per via orale, non hanno mostrato un aumento della percentuale di grasso corporeo in concomitanza con un regime alimentare ipercalorico.(1) La suddetta pubblicazione ha anche riportato che l’Acido Tetradeciltioacetico non solo stimolava il PPAR- α, ma anche il PPAR- γ e, in misura minore, il PPAR-δ.
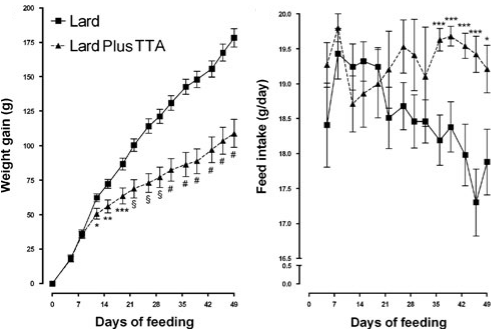
Nel 2009 nutrizionisti affiliati all’Università di Oslo, hanno pubblicato un altro studio svolto su animali. In questo studio, i norvegesi hanno fatto ingrassare i ratti dando loro un mangime che conteneva una elevata percentuale di grasso [Lard] per 7 settimane.
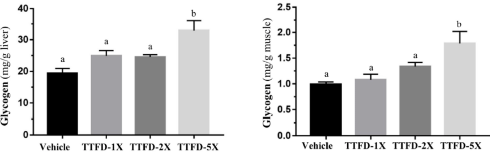
La metà dei ratti presi in esame ha ricevuto anche Acido Tetradeciltioacetico [Lard Plus TTA]. Se i topi fossero stati esseri umani adulti, avrebbero consumato giornalmente circa 1,5g di Acido Tetradeciltioacetico.
L’integrazione con TTA ha significativamente inibito l’aumento del peso corporeo degli animali trattati. Tuttavia, però, la supplementazione ha indotto gli animali a consumare più cibo. Tale comportamento è strano, dal momento che la maggior parte degli agonisti PPAR-α diminuiscono l’appetito interferendo con la biosintesi di alcuni neuropeptidi ipotalamici.
Al termine delle 7 settimane del test, i ratti trattati con Acido Tetradeciltioacetico non erano solo più magri dei ratti non trattati. La loro composizione corporea differiva anche dai ratti del gruppo “Lard”: presentavano una miglior massa muscolare e una ridotta percentuale di grasso.
Di seguito viene mostrato come l’Acido Tetradeciltioacetico stimola il metabolismo nei ratti. Nel fegato, l’attività del gene UCP3, che potenzia il metabolismo, ha subito un incremento di non meno di un fattore di duemila.
I ricercatori scrivono che i loro dati sono in accordo nel riconoscere l’attivazione del PPAR- α come mediatore principale degli effetti dell’Acido Tetradeciltiocetico sull’espressione del gene epatico. L’effetto dell’Acido Tetradeciltiocetico sull’assunzione di cibo è risultato opposto a quanto riportato per altri ligandi il PPAR- α e può essere correlato all’attivazione concomitante del PPAR- γ, alla riduzione della segnalazione della Leptina o alla riduzione della sensibilità ipotalamica del malonil-CoA.
Per chiarire ulteriormente gli effetti dell’Acido Tetradeciltioacetico sull’assunzione di cibo e sull’efficienza del cibo, continuano i ricercatori, l’uso di gabbie metaboliche potrebbe chiarire laquestione. Inoltre, dovrebbero essere prese in considerazione le indagini sull’eventualità che l’Acido Tetradeciltioacetico si accumuli nell’ipotalamo e promuova […] i livelli di sazietà regolati dai neuropeptidi.
Quando i primi supplementi per la perdita di grasso contenenti Acido Tetradeciltioacetico sono comparsi nel mercato degli integratori nel primo decennio del XXI secolo, i produttori hanno cercato di prevenire l’aumento dell’appetito combinando il TTA con composti aventi azione anoressizzante l’Oleoietanolamide.
Se quei supplementi funzionassero davvero è un altro discorso. C’è da considerare anche che la quantità di Acido Tetradeciltioacetico in essi contenuta era bassa. Tuttavia, con i nuovi prodotti presenti sul mercato, le dosi di TTA sono molto più alte.
Sebbene in numero limitato, esistono degli studi svolti sull’uomo che hanno osservato l’effetto del TTA… e i risultati ottenuti non sono così promettenti come quelli osservati sugli animali.
Nel 2004, i ricercatori del Rikshospitalet di Oslo hanno somministrato a 10 pazienti affetti da HIV 1g di TTA al giorno per 4 settimane.(2) L’integrazione non ha avuto alcun effetto sulle cellule T, ma ha ridotto la concentrazione di TNF-α, LDL e Trigliceridi nel sangue. Ma la perdita di peso? Non si è verificata.
Nel 2008, i ricercatori norvegesi, affiliati all’Università di Bergen, hanno somministrato a 18 volontari 200mg, 600mg o 1g di TTA al giorno durante uno studio di fase 1 per 7 giorni. Non sono emersi effetti collaterali degni di nota.(3) Per quanto possibile dedurre da un test della ristretta durata di 7 giorni, il TTA è risultato essere sicuro.
Nel 2009, i ricercatori norvegesi dell’Università di Bergen hanno pubblicato un piccolo studio svolto su esseri umani nel quale hanno somministrato a 16 uomini con diabete di tipo II 1g di TTA ogni giorno per 4 settimane. Gli uomini seguivano una dieta apposita o usavano farmaci per il trattamento del diabete. Alcuni di loro usavano anche statine.
Ovviamente, i livelli di colesterolo e trigliceridi degli uomini sono migliorati (effetto da attribuirsi maggiormente ai farmaci utilizzati), ma non hanno mostrato alcuna perdita di peso.
Nella stessa pubblicazione, i ricercatori parlano anche di esperimenti con cellule muscolari umane. In concentrazioni di diverse decine di micromoli, il TTA ha aumentato l’ossidazione degli acidi grassi.
Nelle cellule muscolari la TTA ha aumentato la produzione di Carnitina Palmitoiltransferasi-I, un enzima coinvolto nell’ossidazione degli acidi grassi, e di CD36, una proteina che è coinvolta nell’assorbimento cellulare degli acidi grassi.
Tuttavia, questi effetti apparentemente non sono abbastanza incisivi da causare una perdita di peso. Forse sono annullati da un aumento dell’appetito, che è stato dimostrato negli studi svolti su animali precedentemente esposti.
Come accennato al principio di questo articolo, il TTA è un acido grasso sintetico, che non si trova in natura. Necessita di essere consumato nel ordine dei grammi [una capsula generalmente può contenerne 0,5g]. Il TTA non viene utilizzato come fonte energetica dall’organismo. Inoltre, il TTA è una sostanza con attività farmacologica. È a tutti gli effetti un farmaco sperimentale, la cui sicurezza non è stata ancora studiata adeguatamente. E negli studi sull’uomo, non sembra funzionare.
In seguito a quanto esposto, non considero una supplementazione con TTA una buona idea sia per la probabile inefficacia in termini di perdita di grasso e sia per le incognite di un suo uso sul lungo termine.
Se non avete ancora letto la prima e la seconda parte di questa serie di articoli vi invito a farlo prima di procedere con la lettura di questa terza parte: 1° Parte – 2° Parte.
Effetti diretti di GH e IGF-1 sull’ipertrofia muscolare
Andiamo dritti al punto – di per sé, il GH non causa direttamente ipertrofia muscolare. Questa caratteristica è stata ampiamente osservata per decenni e, finora, nessuno studio attendibile è stato in grado di mostrare un chiaro effetto della somministrazione di rHGH a medio-lungo termine sull’ipertrofia – anche in dosi sovrafisiologiche somministrate ad atleti di alto livello sottoposti ad intensi allenamenti contro resistenza.
Il fatto che questa correlazione non sia stata dimostrata non è certamente dovuto alla mancanza di tentativi in tal senso. Diversi gruppi di ricerca nel corso degli anni hanno tentato di identificare un ipotetica ipertrofia GH-mediata in soggetti adulti sani [48,195-199] o in soggetti anziani [188,198,200-203] senza successo, o in modo inconcludente. Inoltre, è stato dimostrato che gli aumenti della secrezione di GH indotta dall’esercizio in acuto non hanno prodotto cambiamenti nella MPS o nell’ipertrofia [204-205]. È interessante notare che, sebbene la quantità di studi presenti in letteratura siano significativamente meno comuni di quelli in cui è stato somministrato GH, anche la somministrazione sistemica di rhIGF-1 non ha prodotto alcun effetto ipertrofico misurabile sia in soggetti giovani [63,206] che negli anziani [207- 208].
Vi sono prove recenti che suggeriscono che l’esposizione cronica al GH aumenta l’espressione delle vie intramuscolari responsabili dell’atrofia.[209] È ovvio che molte delle caratteristiche anaboliche dimostrate dal GH possono essere compensate da questa maggiore espressione del catabolismo, che potrebbe essere il motivo per cui l’esposizione cronica al GH non porta all’ipertrofia. Potrebbe anche essere responsabile del perché l’esposizione cronica al GH può produrre muscoli meno efficienti e più deboli.[210] Molto semplicemente, questo potrebbe essere ancora un altro fattore nella lunga serie di effetti regolatori negativi intrinseci nell’Asse GH/IGF-1, ma ulteriori studi dovranno essere condotti per chiarire maggiormente questa ipotesi.
Per quelli che difficilmente accettano le informazioni riportate in questa serie di articoli, e vogliono addentrarsi loro stessi nella letteratura scientifica sperando di trovare informazioni che confutino quanto da me riportato, voglio esporre un concetto molto importante da me già espresso nella prima parte di questa serie di articoli. La maggior parte degli studi che hanno preso in esame il GH segnalano un aumento della massa magra nei soggetti dei gruppi trattati con tale ormone. Quindi, ad un lettore inesperto, potrebbero risultare infondate le affermazioni sulla mancanza di un effetto ipertrofico diretto del GH. Bisogna ricordare, però, che il GH causa una non indifferente ritenzione idrica, oltre ad aumentare la massa dei tessuti molli della quale parlerò brevemente. Nello specifico, il GH aumenta la ritenzione di sodio e, di conseguenza, dell’acqua extracellulare, in modo dose-dipendente, attraverso i suoi effetti sul sistema renina-angiotensina.[211] Questi incrementi nella ritenzione idrica e sodica sono stati osservati anche con la somministrazione di IGF-1, poiché l’IGF-1 stesso sembra essere un regolatore chiave del tasso di escrezione renale di sodio.[83,212-213] Quindi il punto della questione è che bisogna stare molto attenti nel trarre conclusioni quando non si ha ben chiara la differenza tra aumento della massa magra e crescita effettiva del tessuto muscolare.
Effetti indiretti di GH e IGF-1 sull’ipertrofia muscolare
Nella sezione precedente si è dimostrato che il GH e l’IGF-1, da soli, non hanno alcun impatto diretto sull’ipertrofia. Tuttavia, questo non significa che questi peptidi non abbiano un ruolo nei processi ipertrofici. L’obiettivo principale di questa sezione è quello di spiegare alcuni dei molti meccanismi che interessano in modo indiretto il GH e l’IGF-1 nei processi ipertrofici, molti dei quali saranno fattori importanti per gli atleti interessati a massimizzare il loro potenziale ipertrofico.
L’Ormone della Crescita è un potente stimolatore della sintesi di collagene sia nei tendini che nei muscoli. Questo effetto è probabilmente mediato dalla capacità dell’IGF-1 autocrino di stimolare i fibroblasti per sintetizzarlo.[214-215] In realtà, questo processo avviene senza che il peptide influenzi la sintesi proteica muscolare, nonostante i livelli di IGF-1 circolante e di quello locale siano significativamente più alti. Questo effetto è anche indotto indipendentemente dall’allenamento contro resistenza ed è stato persino osservato in soggetti immobilizzati hai quali è stato somministrato il GH.[216] La componente connettiva del muscolo scheletrico è vitale per la trasmissione della forza, che è prodotta dalle fibre muscolari, ai tendini e alle ossa affinché si verifichi il movimento. In particolare, il collagene è un importante componente nella trasmissione della forza della matrice extracellulare, che viene continuamente sottoposta a carichi intesi durante i movimenti.
A causa dei potenti effetti del GH sui componenti della matrice extracellulare, si può chiaramente iniziare a capire il perché gli aneddoti nel corso degli anni suggerivano che l’aggiunta del GH in una preparazione farmacologica produceva impatti positivi sulla riduzione del dolore tendineo-articolare. D’altra parte però, questo potrebbe anche essere un fattore che contribuisce in primo luogo al motivo per cui vari effetti collaterali sono segnalati dagli utilizzatori di GH come l’edema dei tessuti molli, il dolore articolare e la sindrome del tunnel carpale.[217-219] Ci sono anche molti che credono che il GH possa accelerare i tempi di recupero dagli infortuni, ma questo è un argomento complesso che verrà discusso in futuro.
Gli impatti del GH sulla sintesi di collagene potrebbero anche essere di grande interesse per gli atleti di forza che non sono necessariamente interessati all’ipertrofia, ma il cui obiettivo principale è la creazione di una condizione favorevole allo spostamento del carico massimale. Stimolare la sintesi del collagene potrebbe aiutare a rafforzare l’intero sistema di supporto dei muscoli scheletrici. Ora, vale la pena aggiungere una piccola nota di chiarimento. Nonostante questo suoni positivo in linea di principio, la supplementazione di GH non ha mai portato direttamente a guadagni di forza in nessuno degli studi svolti su soggetti sani, coprendo vari gruppi.[48,50,184,196-198,200-201,203,220-221] Naturalmente, se il GH fosse usato insieme ad un composto con la capacità di aumentare direttamente la forza, non è difficile supporre che in questo caso l’effetto addizionale potrebbe fare del GH un prezioso componente della preparazione.
Decorina
La Decorina è una proteina strutturale, che risiede principalmente nella matrice extracellulare del muscolo scheletrico e il cui ruolo è correlato alla crescita e alla riparazione dei muscoli.[222-223] Qualche decennio fa è stato dimostrato per la prima volta che la somministrazione di GH potrebbe aumentare direttamente l’espressione del gene della Decorina negli animali [224]. Recentemente, è stato dimostrato che questo effetto si manifesta anche in soggetti umani sottoposti ad allenamenti ricreativi.[225] Nello studio più recente, i livelli di Decorina sono fortemente correlati a quelli di PIIINP, che induce la stimolazione del Decorina GH-mediata che può essere coinvolta nel processo di assemblaggio della matrice di collagene osseo. Questo effetto è più pronunciato negli uomini rispetto alle donne e può essere un sottoprodotto dei livelli più alti di IGF-1 osservati negli uomini, sebbene questa affermazione sia speculativa. L’aumento dell’espressione di Decorina non è stato alterato dall’aggiunta di Testosterone, quindi questo effetto è indipendente dagli androgeni. Dopo aver visto gli effetti che il GH ha sulla sintesi di collagene e Decorina, sta diventando piuttosto chiaro che l’asse GH/IGF-1 è molto più importante per rafforzare la matrice extracellulare di supporto piuttosto che contribuire direttamente alla crescita del tessuto muscolare.
Adenosina Trifosfato (ATP)
La somministrazione acuta di GH in soggetti sani ha anche dimostrato di causare una maggiore produzione di ATP mitocondriale e una maggiore attività della citrato sintasi nel muscolo scheletrico, con una maggiore abbondanza di mRNA muscolari codificanti l’IGF-1.[226] Non solo questo potrebbe essere un fattore che contribuisce al motivo per cui il GH potrebbe avere la capacità di promuovere un aumento dei tassi di spesa energetica giornaliera ma potrebbe anche essere coinvolto nello spostamento verso la preferenza dei lipidi come substrato energetico. Anche se, come abbiamo visto in precedenza, il corpo della letteratura non supporta questa pratica, l’aumento della produzione di ATP mitocondriale potrebbe avere un ruolo nella capacità aerobica.[227]
È stato dimostrato che il GH promuove la fusione dei mioblasti con i miotubi in modelli cellulari [84], un effetto completamente indipendente dalla sovraregolazione locale del IGF-1. Per capire perché questo possa essere importante, bisogna approfondire maggiormente la questione dei fattori cellulari coinvolti nell’ipertrofia del muscolo scheletrico. L’ipertrofia dei muscoli scheletrici negli esseri umani si basa sulle cellule satelliti, che sono cellule dormienti situate all’interno delle miofibre, proprio sotto lo strato di lamina basale nella matrice extracellulare.[148] Una volta attivate queste cellule satellite, spesso con l’esercizio fisico o il danno muscolare, proliferano. Dopo la proliferazione, queste cellule satellite migrano verso siti ove si trova il danno differenziandosi e fondendosi con le miofibre esistenti che forniscono nuovi nuclei per l’ipertrofia e la riparazione. [228] Non è ancora del tutto chiaro se il GH abbia un effetto diretto sulla proliferazione e la differenziazione delle cellule satelliti.[229-230] Vale anche la pena di affermare che ciò che si osserva nelle colture cellulari potrebbe non essere del tutto indicativo di ciò che accade in vivo a causa di vari fattori esterni che non possono essere spiegati in condizioni di laboratorio.
Ci sono due fasi distinte in questa fusione dei mioblasti che si verifica.[85] Il primo sarebbe lo stadio iniziale della differenziazione in cui un sottoinsieme di cellule mononucleate si fondono per formare miotubi nascenti (fusione mioblasto/mioblasto). Questo è seguito dal secondo stadio che coinvolge ulteriori cellule disponibili che si fondono con questi miotubi nascenti e dove avviene la crescita muscolare effettiva (fusione mioblasto/miotubo). È all’interno di quest’ultimo stadio in cui il GH esercita i suoi effetti.
Questa è una scoperta piuttosto interessante, ma ancora una volta, numerosi studi sugli esseri umani non sono riusciti a dimostrare un effetto ipertrofico del GH in condizioni reali. Quindi, possiamo probabilmente dedurre da ciò che gli effetti che il GH ha sulla fusione dei nascenti miotubi non si traducono direttamente nell’ipertrofia. Tuttavia, cosa accadrebbe se si aggiungesse un’altra variabile nell’equazione in grado di creare un ambiente in cui esistessero numeri di celle satelliti potenziati, creando più materiale sul quale l’azione del GH possa manifestarsi?[231]
Introduzione degli AAS
A differenza del GH e del IGF, l’uso degli AAS ha mostrato di avere un impatto pronunciato su ipertrofia e forza. Questo è stato ben noto per decenni anche perché sono stati usati e abusati da atleti fin dalla loro creazione negli anni ’30 (con buona pace dei “puritani” della “old school”).[232] La famiglia degli AAS consiste in un gruppo di potenti composti sintetici che sono simili nella struttura chimica al Testosterone e/o al suo derivato 5α-ridotto (DHT). Vari tipi di singoli composti AAS sono stati creati nel corso degli anni usando come base la molecola naturale di Testosterone manipolandola, quindi, attraverso l’aggiunta di un gruppo etile, metile, idrossile o benzile in uno o più siti lungo la sua struttura.[233-234 ] Alcune delle varianti AAS più facilmente riconoscibili includono i composti metilati in C-17, i quali sono notoriamente dotati di un elevata biodisponibilità orale, e le varianti del 19-nortestosterone nelle quali viene rimosso il gruppo 19-metilico dalla molecola di Testosterone nel tentativo di aumentare la sua attività anabolica diminuendo al contempo la sua androgenicità e tendenza all’aromatizzazione. Conosciuti anche come “19-norsteroidi”, questa famiglia include ben note varianti di AAS come il Nandrolone ed il Trenbolone.
Molte di questi composti sono nati come risultato di un desiderio di aumentare le caratteristiche anaboliche del Testosterone a livello muscolare, abbassando allo stesso tempo gli effetti collaterali androgenici intrinsecamente inerenti alla molecola di Testosterone.[235] In generale, i rischi complessivi degli effetti collaterali derivanti dall’uso cronico di AAS sembrano essere relativamente bassi rispetto a molte sostanze socialmente accettate come l’alcol, il tabacco e vari farmaci da prescrizione.[233,236] Ovviamente, l’uso e l’abuso sono termini che si escludono a vicenda e abusare di qualsiasi sostanza tende a creare un ambiente a più alto rischio di effetti collaterali. A meno che non sia diversamente specificato, da questo punto in poi, parlerò specificamente del Testosterone in quanto è l’ormone sessuale endogeno maschile nativo, nonché l’androgeno più approfonditamente studiato in letteratura. Tanto per avere un riferimento, i maschi adulti sani producono una media di 14-77mg/settimana di Testosterone endogeno. [435]
Testosterone
Il Testosterone è un noto regolatore della massa muscolare, e gli aumenti della massa muscolare mediati dal Testosterone sono associati all’ipertrofia delle fibre, così come ad un aumento delle cellule satelliti e del numero mio nucleare.[231,237-240] Il muscolo scheletrico sembra essere uno dei tessuti più reattivi al Testosterone e si stima che i livelli circolanti di Testosterone rappresentino il fattore causale significativo (incidenza del 40-75%) dei guadagni nella massa muscolare osservati in studi di controllo randomizzati. Se ricordate ciò che è stato affermato e dimostrato nella precedente sezione, il GH possiede un’abilità molto particolare in quanto può aumentare la fusione dei mioblasti durante il processo ipertrofico. La massimizzazione di questa capacità si basa sull’avere un numero adeguato di cellule satellitari disponibili.
I risultati mostrano costantemente che i trattamenti con Testosterone determinano aumenti dose-dipendenti della massa e della forza del muscolo scheletrico, indipendentemente dal fatto che i soggetti presi in esame siano maschi più giovani o più anziani.[241-243] Al contrario, negli studi con soggetti sani giovani in cui il Testosterone endogeno è stato soppresso artificialmente, la forza e la composizione corporea hanno entrambi subito peggioramenti significativi.[244] Per ribadire il nostro precedente punto, gli aumenti della massa muscolare mediati dal Testosterone sono basati sull’ipertrofia e non sono il risultato di una transizione della fibra (cambiando le fibre di tipo I in fibre di tipo II o viceversa) o di una sua scissione.[245] A causa dei loro meccanismi anabolici unici e non sovrapposti, la somministrazione di Testosterone e l’allenamento contro resistenza hanno anche mostrato effetti sinergici e additivi l’uno sull’altro per quanto riguarda lo stimolo dell’aumento della massa muscolare.[246]
Recettore degli Androgeni (AR)
Gli Androgeni mediano principalmente i loro effetti attraverso il gene del recettore degli androgeni (AR) che è espresso in mioblasti, miofibre e cellule satelliti.[247-248] I AR sono stati rilevati anche nelle cellule che supportano i muscoli, come i fibroblasti e le cellule endoteliali. La densità dei AR sembra essere specifica per i gruppi muscolari, con l’allenamento contro resistenza e l’uso di AAS che hanno la capacità di influenzare il numero di AR presenti in questi gruppi muscolari. Oltre ai suoi effetti sulla densità dei AR, l’uso di AAS ha anche dimostrato la capacità di influenzare i livelli di attività dei AR sia in modo acuto che sul lungo termine.[249-250] Questi sono fattori piuttosto importanti da considerare quando ci si imbatte in individui che sostengono che il precedente utilizzo di AAS non offre necessariamente un vantaggio competitivo permanente.
A causa della complessità dell’argomento, ci sono state diverse ipotesi riguardo ai meccanismi con cui l’AAS esercita le sue azioni anaboliche sul muscolo scheletrico.[251] È stato dimostrato che i trattamenti con Testosterone aumentano i tassi di sintesi proteica muscolare (MPS) [252], riducono i tassi di degradazione delle proteine [253] facendo persino in modo che il corpo utilizzi più efficientemente gli amminoacidi prontamente disponibili. Quindi, ancora una volta, questo è un sistema abbastanza complesso che può anche essere semplificato ricordando che gli AAS promuovono l’anabolismo muscolare attraverso la loro capacità di incidere positivamente sull’equilibrio degli aminoacidi.
Via Wnt/B-catenina
È generalmente accettato che gli AAS esercitino i loro effetti anabolizzanti attraverso il legame e attivazione del AR che successivamente attiva cascate di segnalazione a valle che coinvolgono la via Wnt/β-catenina.[254-256]. I Wingless-INT (Wnt) sono una famiglia di glicoproteine secrete che regolano la proliferazione e la differenziazione cellulare.[257-258] I modelli cellulari hanno mostrato che il AR forma un complesso con la beta-catenina che si potenzia in presenza di AAS. [259-260] Una volta attivato questo complesso, migra nei nuclei in cui regola l’espressione dei geni target e la differenziazione delle cellule satelliti.[261-262] Questo è anche il pathway degli AAS in gran parte responsabile della miogenesi, la formazione del tessuto muscolare.[263-265]
Vale la pena notare che l’AAS possiede anche caratteristiche non genomiche che possono influenzare rapidamente numerosi processi ormonali e metabolici al di fuori del classico legame con il recettore. In letteratura sono stati riportati casi di maschi adulti con disturbi di insensibilità agli androgeni, causati da mutazioni del AR, che hanno risposto al Testosterone in modo molto simile ai soggetti sani. Questi casi studio rafforzano l’ipotesi che gli effetti anabolici dell’AAS possano essere mediati indipendentemente dal AR.[266] Le azioni non genomiche degli Androgeni possono in realtà essere un argomento piuttosto affascinante, ma che va oltre lo scopo previsto per questo articolo. Per coloro che vogliono approfondire l’argomento, consiglio di iniziare con le seguenti note di riferimento.[267-268]
AAS e potenziale sinergico con l’Asse GH/IGF-1
Ho riportato molte informazioni utili fino a questo punto, ma è qui che le cose cominciano davvero a farsi interessanti. A questo punto una domanda logica sarebbe: esistono studi svolti su soggetti sani nei quali si sono confrontate le differenze tra trattamenti singoli con GH o Androgeni e trattamenti combinati? Fortunatamente per noi, la risposta è “sì” in quanto vi sono stati alcuni studi, principalmente utilizzando soggetti anziani, sia maschi che femmine. I risultati di ognuno di questi studi dimostrano chiaramente che il GH ha un effetto additivo sui benefici consolidati che la terapia con ormoni sessuali fornisce: l’ipertrofia, la lipolisi, la sintesi del collagene, la funzione fisica, la qualità della vita e altri vari indicatori di prestazione.[187-188,269-270] Dato che è abbastanza chiaro che esiste un effetto additivo, vediamo se è possibile approfondire ulteriormente l’argomento al fine di scoprire alcuni dei meccanismi sottostanti che operano nella sinergia tra Androgeni e GH.
GHRH
Deve essere chiaro che il Testosterone, di per sé, ha un effetto additivo sull’intero Asse GH/IGF-1. Questo è stato osservato sia in modelli umani che animali, con la somministrazione di Testosterone che porta ad un aumento dei livelli circolanti di GH e IGF-1.[241,271-276] Viceversa, il deficit di Testosterone è comunemente associato a livelli significativamente ridotti di IGF-1.[277] L’effetto stimolante del Testosterone sull’Asse GH/IGF-1 sembra essere mediato a livello ipotalamico da una promozione della funzionalità del GHRH.[278]
Inoltre, e questo è bene ricordarlo, gli AAS non aromatizzabili sembrano non possedere lo stesso effetto stimolante sull’Asse GH/IGF-1.[279] Gli inibitori dell’aromatasi (AI), progettati per sopprimere il processo di aromatizzazione degli Androgeni, hanno dimostrato di attenuare direttamente la stimolazione del GH seguente la somministrazione di Testosterone. Questi indizi forniscono prove abbastanza convincenti sul fatto che gli estrogeni locali, derivanti dall’aromatizzazione, svolgono un ruolo fondamentale nella regolazione della secrezione di GH nei maschi.[280-281] Poiché l’aromatasi non è espressa nel fegato, gli AI non influenzano l’azione epatica del GH, mentre influenzano la sua azione a livello del sistema centrale.[282-283] Tuttavia i modulatori selettivi del recettore degli estrogeni (SERM) sono ancora più soppressivi in quanto agiscono ad entrambi i livelli a causa del loro meccanismo d’azione.[284-285]
Anche gli Androgeni che aumentano i livelli serici di Estrogeni, come il Nandrolone (Nor-Estrogeni), mostrano un effetto minimo sui livelli sistemici di GH e IGF-1 rispetto al Testosterone.[286] Ciò è legato ipoteticamente sia al fatto che il Nandrolone è poco soggetto all’aromatizzazione e sia al fatto che la forma estrogenica derivata (Nor-Estrogeno) abbia un ridotto potenziale di attività.[287] Ricordiamoci che l’aromatizzazione sembra essere il passo più cruciale nella stimolazione ipotalamica mediata dagli Androgeni. Ora, è ovvio che un utilizzatore di rHGH deve preoccuparsi parzialmente di questo effetto rispetto ad un non utilizzatore, considerando il fatto che, in tal caso, i livelli ormonali sono controllati quasi esclusivamente da mezzi esogeni. Detto questo, è sempre utile analizzare il quadro generale, soprattutto se l’obiettivo è massimizzare l’ipertrofia.
GHR
Un altro potenziale motivo legato agli aumenti dei livelli di GH e IGF osservati durante i trattamenti con Testosterone, è rappresentato dagli effetti diretti di quest’ultimo sui GHR. Sia gli studi sull’uomo che sugli animali hanno fornito prove del fatto che il Testosterone modifica i GHR sia nel fegato che nei tessuti periferici, migliorandone l’espressione.[288-289] Inoltre, soggetti umani ipopituitari e ipogonadici sottoposti a trattamenti con GH hanno mostrato una aumentata risposta sia all’IGF-1 locale che all’espressione del gene del recettore degli androgeni quando sottoposti anche a trattamento con Testosterone.[187,290-291] Inoltre, i maschi ipopituitari sottoposti a trattamenti con Testosterone hanno mostrato effetti notevoli sull’anabolismo proteico in presenza di GH, con il sito primario di interazione ormonale a livello epatico.[292] Quindi anche quando i livelli ormonali sono carenti, c’è ancora un’interazione molto importante tra il Testosterone e l’Asse GH/IGF-1.
Come accennato in precedenza, il GHR è espresso in quasi tutti i principali tipi di tessuto. Vale la pena sottolineare che il GHR è espresso in quantità molto bassa nel muscolo scheletrico – solo circa il 4-33% dei livelli osservati in altri tessuti. D’altra parte, il recettore del IGF-1 è espresso in maniera molto più elevata nei muscoli scheletrici, proprio come nel tessuto epatico.[293-294] Detto questo, l’aumento della sensibilità del GHR in un ambiente con livelli sovra fisiologici di GH andrà certamente a beneficio del BodyBuilder. I BodyBuilder cercano continuamente di utilizzare quantità elevate di rHGH nel tentativo di massimizzare i processi ipertrofici, e se il GH vede potenziata la sua azione e, quindi, il suo stimolo diretto sulla sintesi e rilascio del IGF-1, il quale opera direttamente sul tessuto muscolare, venendosi a creare quindi una vantaggiosa alterazione dell’Asse GH/IGF-1, l’effetto ipertrofico potenziato sarà indubbio.
IGFBP-4
Gli Androgeni hanno mostrato di avere la capacità di aumentare l’espressione del mRNA del IGF-1 locale nel muscolo scheletrico. Possiamo pertanto ipotizzare che gli Androgeni, in particolare a dosi più elevate, creino un ambiente all’interno del muscolo scheletrico estremamente favorevole alla gestione di livelli più alti di IGF-1 seguenti alla somministrazione di dosi sovrafisioogiche di rHGH. Ci sono persino stati studi sull’uomo che hanno mostrato livelli ridotti di IGFBP-4 locale nei campioni di tessuto muscolare, oltre a maggiori livelli di mRNA del IGF-1. Ciò potrebbe indicare che i cambiamenti hanno avuto luogo in quei muscoli per liberare più IGF-1 locale affinché esso possa legarsi ai suoi recettori.[277,295] Non ho molto approfondito la questione delle proteine leganti l’IGF, ma l’IGFBP-4 inibisce l’azione dell’IGF-1 e, quindi, minore è il livello della proteina legante, maggiori saranno i livelli di IGF-1 libero e attivo.[149,296]
È stato anche dimostrato che il Testosterone promuove l’ipertrofia negli stati deficitari di GH/IGF-1. [297-298] Questo è interessante in quanto dimostra che il Testosterone possiede sia vie metaboliche IGF-indipendenti che IGF-indipendenti nel tessuto muscolare.[299] Inoltre, i modelli cellulari hanno dimostrato che il Testosterone può sovraregolare l’espressione delle varie isoforme di IGF nei muscoli scheletrici, anche in assenza di GH/IGF-1.[298] E, anche se questo è stato dimostrato nei fibroblasti, il Testosterone ha mostrato di aumentare l’espressione del IGFBP-3 – un effetto che è stato ulteriormente migliorato dalla somministrazione di IGF-1.[300] È abbastanza chiaro, comunque, che il Testosterone eserciti effetti sinergici e additivi sull’anabolismo mediato da GH/IGF-1.
Fino ad ora mi sono concentrato sul Testosterone, tuttavia, però, sono stati osservati comportamenti leggermente diversi in relazione alle varianti androgene e al loro impatto sull’espressione sistemica e locale del IGF-1. Volevo analizzare un paio di specifici composti che sono spesso presenti nei protocolli farmacologici: sto parlando del Trenbolone e del Nandrolone. Salvo diverse indicazione, i risultati in seguito esposti provengono da studi svolti su animali.
Nandrolone
La somministrazione di Nandrolone ha costantemente dimostrato di non causare cambiamenti nei livelli di IGF-1 endocrino, nonostante produca simultaneamente un’espressione del IGF-1 muscolare significativamente più alta e un aumento della CSA delle fibre muscolari.[286,301-302] Inoltre, i livelli di IGFBP-3 locali sono stati riportati come significativamente più alti mentre i livelli di IGFBP-4 sono stati significativamente soppressi, il che ci rimanda a quanto esposto in precedenza – cioè ad un fattore che determina un aumento dei livelli di IGF-1 locale libero. In tutti gli studi, la somministrazione di Nandrolone ha portato direttamente ad un aumento dell’ipertrofia, nonostante non abbia avuto alcun impatto sui livelli di IGF-1 sistemici. Ciò rafforza ulteriormente l’ipotesi secondo cui l’IGF-1 endocrino non sia un fattore primario nell’ipertrofia dei muscoli scheletrici e quindi i livelli elevati non sono un prerequisito per l’aumento della massa muscolare.[303-305]
Trenbolone
È stato inoltre universalmente dimostrato che il Trenbolone aumenta i tassi di crescita muscolare in tutte le varie specie nelle quali è stato testato. A differenza del Testosterone e del Nandrolone, non si converte in estrogeno ed è stato suggerito fin dagli anni ’70 che l’aggiunta di Estradiolo al Trenbolone sembra migliorare gli effetti anabolici del composto.[306-307] Ci sono stati anche effetti potenziati sull’ipertrofia quando il Trenbolone è stato somministrato insieme ad un fattore di rilascio dell’Ormone della Crescita (GHRF).[308] Come accennato in precedenza, l’aumento di GH/IGF-1 necessita di livelli adeguati di estrogeni (derivanti principalmente dall’aromatizzazione) affinché questi esercitino uno stimolo dell’asse GH/IGF-1. Poiché la somministrazione di Trenbolone diminuisce intrinsecamente i livelli di Estradiolo, mediante inibizione a feedback negativo del Testosterone attraverso l’Asse Ipotalamo-Ipofisi-Gonadi (HPG) [309], la somministrazione di Estradiolo, tecnicamente, dovrebbe migliorare la funzionalità dell’asse GH/IGF-1. Questo dovrebbe quindi favorire ulteriormente la sinergia anabolica che avrebbe con l’androgeno. Questa ipotesi è in linea con ciò che vari studi hanno dimostrato nel corso degli anni.
Nelle colture cellulari, è stato anche dimostrato che gli Estrogeni alterano direttamente i tassi di MPS e MPB del Trenbolone attraverso meccanismi che coinvolgono sia il recettore estrogenico che il recettore del IGF-1.[310-311] In effetti, in linea di massima, i trattamenti con solo Trenbolone non mostrano aumenti significativi dei livelli endocrini o autocrini del IGF-1. Tuttavia, la co-somministrazione con Estradiolo ha tradizionalmente mostrato aumenti nei livelli di IGF-1 autocrini, similmente a quanto osservato con il Testosterone.[312-314] Questa è solo un’ulteriore prova che suggerisce che l’Estrogeno, sia di origine sistemica che derivata dall’aromatasi, è un componente chiave sia della massima stimolazione dell’Asse GH/IGF-1 che delle capacità anaboliche massimali degli Androgeni.
Proprio come il suo “cugino” 19-nor, il Trenbolone ha anche mostrato una maggiore espressione del fattore di crescita nei tessuti muscolo scheletrici, oltre ad una maggiore reattività dei muscoli a tali fattori di crescita.[315] Il Trenbolone ha anche mostrato di operare un aumento dell’attivazione e della proliferazione delle cellule satelliti in varie specie, in misura simile al Testosterone.[316-317] Ora, essendo a conoscenza delle interazioni legate al GH, si può facilmente giungere alla conclusione che entrambi questi effetti sarebbero vantaggiosi in un protocollo che includesse entrambi i composti.
PIIINP
Tornando al Testosterone, sia il GH che questo Androgeno aumentano i marker di sintesi del collagene come il PIIINP. Inoltre, il Testosterone ha anche dimostrato di potenziare le capacità del GH di aumentare la sintesi di collagene sia nei muscoli che nei tendini.[318] A sostegno di ciò, la somministrazione concomitante di GH e Testosterone in soggetti umani allenati (a livello amatoriale) ha causato aumenti significativi sia dei marker del IGF-1 che del collagene.[319] E’ interessante notare come alcuni di questi stessi marker del collagene dei quali si sta discutendo sono indicatori esatti che vengono esaminati come parte dei test antidoping per la rilevazione del GH.[320-321]
In precedenza ho solo brevemente toccato il percorso JAK-STAT ma, arrivati a questo punto, è necessario trattare l’argomento in maniera leggermente più approfondita. La via JAK-STAT è un componente critico del GH e riguarda sia la trascrizione del gene del IGF-1 che la crescita postnatale. Una delle proteine STAT in particolare, la STAT5, sembra essere intimamente coinvolta anche nella regolazione del muscolo scheletrico.[322] Ci sono due sottoproteine nella famiglia STAT5 e sono indicate come STAT5a e STAT5b. Sebbene siano identiche al 96%, è la variante STAT5b che è abbondante nei muscoli e nel tessuto epatico ed è quindi la proteina specifica su cui mi concentrerò da questo punto in avanti.[323-324]
Una revisione completa del percorso di segnalazione renderebbe questo articolo estremamente prolisso, ma ritengo importante che si comprenda che il percorso JAK/STAT5b è stato ripetutamente dimostrato avere una relazione diretta con l’espressione dell’IGF-1 locale nel tessuto muscolare e sul’ipertrofia.[248,325-332] Per questo motivo, se ci fossero modi per migliorare o ottimizzare questo specifico percorso, sembrerebbe che ciò si possa tradurre non solo in un aumento dell’attivazione del gene del IGF-1 [333-335] ma anche in un maggiore potenziale ipertrofico.
Fortunatamente, alcuni nuovi studi svolti su animali hanno già mostrato come i percorsi AR e JAK-STAT siano intimamente correlati.[248,336] Per essere precisi, il percorso STAT5a/b è a monte e l’AR è un bersaglio diretto a valle tramite la regolazione dell’espressione genica del AR. Studi su esseri umani hanno anche dimostrato che questo si verifica anche nell’uomo, con l’attività della STAT5 correlata positivamente all’espressione AR nelle linee cellulari del cancro alla prostata. [337]
Nella prossima parte di questa serie di articoli, parlerò dei modi in cui è possibile tentare di garantire che la via JAK-STAT5b-AR sia massimamente sensibilizzata, assicurando così che il potenziale ipertrofico sia potenziato quando Androgeni e GH vengono co-somministrati. Inoltre tratterò le applicazioni pratiche dell’uso del GH a fini ipertrofici esponendo infine le mie conclusioni in merito.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
Yarasheski KE, Campbell JA, Smith K, Rennie MJ, Holloszy JO, Bier DM. Effect of growth hormone and resistance exercise on muscle growth in young men. Am J Physiol. 1992 Mar;262(3 Pt 1):E261-7.
Yarasheski KE, Zachwieja JJ, Campbell JA, Bier DM. Effect of growth hormone and resistance exercise on muscle growth and strength in older men. Am J Physiol. 1995 Feb;268(2 Pt 1):E268-76.
Fryburg DA. Insulin-like growth factor I exerts growth hormone- and insulin-like actions on human muscle protein metabolism. Am J Physiol. 1994 Aug;267(2 Pt 1):E331-6.
Ohlsson C, Mohan S, Sjögren K, Tivesten A, Isgaard J, Isaksson O, Jansson JO, Svensson J. The role of liver-derived insulin-like growth factor-I. Endocr Rev. 2009 Aug;30(5):494-535.
Wakelam MJ. The fusion of myoblasts. Biochem J. 1985 May 15;228(1):1-12. Review.
Crist DM, Peake GT, Egan PA, Waters DL. Body composition response to exogenous GH during training in highly conditioned adults. J Appl Physiol (1985). 1988 Aug;65(2):579-84.
Deyssig R, Frisch H, Blum WF, Waldhör T. Effect of growth hormone treatment on hormonal parameters, body composition and strength in athletes. Acta Endocrinol (Copenh). 1993 Apr;128(4):313-8.
Yarasheski KE, Zachweija JJ, Angelopoulos TJ, Bier DM. Short-term growth hormone treatment does not increase muscle protein synthesis in experienced weight lifters. J Appl Physiol (1985). 1993 Jun;74(6):3073-6.
Lange KH, Andersen JL, Beyer N, Isaksson F, Larsson B, Rasmussen MH, Juul A, Bülow J, Kjaer M. GH administration changes myosin heavy chain isoforms in skeletal muscle but does not augment muscle strength or hypertrophy, either alone or combined with resistance exercise training in healthy elderly men. J Clin Endocrinol Metab. 2002 Feb;87(2):513-23.
Ehrnborg C, Ellegård L, Bosaeus I, Bengtsson BA, Rosén T. Supraphysiological growth hormone: less fat, more extracellular fluid but uncertain effects on muscles in healthy, active young adults. Clin Endocrinol (Oxf). 2005 Apr;62(4):449-57.
Rudman D, Feller AG, Nagraj HS, Gergans GA, Lalitha PY, Goldberg AF, Schlenker RA, Cohn L, Rudman IW, Mattson DE. Effects of human growth hormone in men over 60 years old. N Engl J Med. 1990 Jul 5;323(1):1-6.
Taaffe DR, Pruitt L, Reim J, Hintz RL, Butterfield G, Hoffman AR, Marcus R. Effect of recombinant human growth hormone on the muscle strength response to resistance exercise in elderly men. J Clin Endocrinol Metab. 1994 Nov;79(5):1361-6.
Taaffe DR, Jin IH, Vu TH, Hoffman AR, Marcus R. Lack of effect of recombinant human growth hormone (GH) on muscle morphology and GH-insulin-like growth factor expression in resistance-trained elderly men. J Clin Endocrinol Metab. 1996 Jan;81(1):421-5.
Hennessey JV, Chromiak JA, DellaVentura S, Reinert SE, Puhl J, Kiel DP, Rosen CJ, Vandenburgh H, MacLean DB. Growth hormone administration and exercise effects on muscle fiber type and diameter in moderately frail older people. J Am Geriatr Soc. 2001 Jul;49(7):852-8.
West DW, Kujbida GW, Moore DR, Atherton P, Burd NA, Padzik JP, De Lisio M, Tang JE, Parise G, Rennie MJ, Baker SK, Phillips SM. Resistance exercise-induced increases in putative anabolic hormones do not enhance muscle protein synthesis or intracellular signalling in young men. J Physiol. 2009 Nov 1;587(Pt 21):5239-47.
West DW, Burd NA, Tang JE, Moore DR, Staples AW, Holwerda AM, Baker SK, Phillips SM. Elevations in ostensibly anabolic hormones with resistance exercise enhance neither training-induced muscle hypertrophy nor strength of the elbow flexors. J Appl Physiol (1985). 2010 Jan;108(1):60-7.
Fryburg DA, Jahn LA, Hill SA, Oliveras DM, Barrett EJ. Insulin and insulin-like growth factor-I enhance human skeletal muscle protein anabolism during hyperaminoacidemia by different mechanisms. J Clin Invest. 1995 Oct;96(4):1722-9
Butterfield GE, Thompson J, Rennie MJ, Marcus R, Hintz RL, Hoffman AR. Effect of rhGH and rhIGF-I treatment on protein utilization in elderly women. Am J Physiol. 1997 Jan;272(1 Pt 1):E94-9.
Friedlander AL, Butterfield GE, Moynihan S, Grillo J, Pollack M, Holloway L, Friedman L, Yesavage J, Matthias D, Lee S, Marcus R, Hoffman AR. One year of insulin-like growth factor I treatment does not affect bone density, body composition, or psychological measures in postmenopausal women. J Clin Endocrinol Metab. 2001 Apr;86(4):1496-503.
Consitt LA, Saneda A, Saxena G, List EO, Kopchick JJ. Mice overexpressing growth hormone exhibit increased skeletal muscle myostatin and MuRF1 with attenuation of muscle mass. Skelet Muscle. 2017 Sep 4;7(1):17.
Wolf E, Wanke R, Schenck E, Hermanns W, Brem G. Effects of growth hormone overproduction on grip strength of transgenic mice. Eur J Endocrinol. 1995 Dec;133(6):735-40.
Ho KY, Weissberger AJ. The antinatriuretic action of biosynthetic human growth hormone in man involves activation of the renin-angiotensin system. Metabolism. 1990 Feb;39(2):133-7.
Blazer-Yost BL, Cox M. Insulin-like growth factor 1 stimulates renal epithelial Na+ transport. Am J Physiol. 1988 Sep;255(3 Pt 1):C413-7.
Giordano M, DeFronzo RA. Acute effect of human recombinant insulin-like growth factor I on renal function in humans. Nephron. 1995;71(1):10-5.
Ehrnborg C, Lange KH, Dall R, Christiansen JS, Lundberg PA, Baxter RC, Boroujerdi MA, Bengtsson BA, Healey ML, Pentecost C, Longobardi S, Napoli R, Rosén T; GH-2000 Study Group. The growth hormone/insulin-like growth factor-I axis hormones and bone markers in elite athletes in response to a maximum exercise test. J Clin Endocrinol Metab. 2003 Jan;88(1):394-401.
Doessing S, Heinemeier KM, Holm L, Mackey AL, Schjerling P, Rennie M, Smith K, Reitelseder S, Kappelgaard AM, Rasmussen MH, Flyvbjerg A, Kjaer M. Growth hormone stimulates the collagen synthesis in human tendon and skeletal muscle without affecting myofibrillar protein synthesis. J Physiol. 2010 Jan 15;588(Pt 2):341-51.
Boesen AP, Dideriksen K, Couppé C, Magnusson SP, Schjerling P, Boesen M, Kjaer M, Langberg H. Tendon and skeletal muscle matrix gene expression and functional responses to immobilisation and rehabilitation in young males: effect of growth hormone administration. J Physiol. 2013 Dec 1;591(23):6039-52.
Cohn L, Feller AG, Draper MW, Rudman IW, Rudman D. Carpal tunnel syndrome and gynaecomastia during growth hormone treatment of elderly men with low circulating IGF-I concentrations. Clin Endocrinol (Oxf). 1993 Oct;39(4):417-25.
Sullivan DH, Carter WJ, Warr WR, Williams LH. Side effects resulting from the use of growth hormone and insulin-like growth factor-I as combined therapy to frail elderly patients. J Gerontol A Biol Sci Med Sci. 1998 May;53(3):M183-7.
Dickerman RD, Douglas JA, East JW. Bilateral median neuropathy and growth hormone use: a case report. Arch Phys Med Rehabil. 2000 Dec;81(12):1594-5.
Papadakis MA, Grady D, Black D, Tierney MJ, Gooding GA, Schambelan M, Grunfeld C. Growth hormone replacement in healthy older men improves body composition but not functional ability. Ann Intern Med. 1996 Apr 15;124(8):708-16.
Zachwieja JJ, Yarasheski KE. Does growth hormone therapy in conjunction with resistance exercise increase muscle force production and muscle mass in men and women aged 60 years or older? Phys Ther. 1999 Jan;79(1):76-82. Review.
Kishioka Y, Thomas M, Wakamatsu J, Hattori A, Sharma M, Kambadur R, Nishimura T. Decorin enhances the proliferation and differentiation of myogenic cells through suppressing myostatin activity. J Cell Physiol. 2008 Jun;215(3):856-67.
Kanzleiter T, Rath M, Görgens SW, Jensen J, Tangen DS, Kolnes AJ, Kolnes KJ, Lee S, Eckel J, Schürmann A, Eckardt K. The myokine decorin is regulated by contraction and involved in muscle hypertrophy. Biochem Biophys Res Commun. 2014 Jul 25;450(2):1089-94.
Zhang CZ, Li H, Bartold PM, Young WG, Waters MJ. Effect of growth hormone on the distribution of decorin and biglycan during odontogenesis in the rat incisor. J Dent Res. 1995 Oct;74(10):1636-43.
Bahl N, Stone G, McLean M, Ho KKY, Birzniece V. Decorin, a growth hormone regulated protein in humans. Eur J Endocrinol. 2017 Nov 14. pii: EJE-17-0844.
Short KR, Moller N, Bigelow ML, Coenen-Schimke J, Nair KS. Enhancement of muscle mitochondrial function by growth hormone. J Clin Endocrinol Metab. 2008 Feb;93(2):597-604. Epub 2007 Nov 13.
Lange KH, Isaksson F, Juul A, Rasmussen MH, Bülow J, Kjaer M. Growth hormone enhances effects of endurance training on oxidative muscle metabolism in elderly women. Am J Physiol Endocrinol Metab. 2000 Nov;279(5):E989-96.
Chargé SB, Rudnicki MA. Cellular and molecular regulation of muscle regeneration. Physiol Rev. 2004 Jan;84(1):209-38. Review.
Halevy O, Hodik V, Mett A. The effects of growth hormone on avian skeletal muscle satellite cell proliferation and differentiation. Gen Comp Endocrinol. 1996 Jan;101(1):43-52.
Kim H, Barton E, Muja N, Yakar S, Pennisi P, Leroith D. Intact insulin and insulin-like growth factor-I receptor signaling is required for growth hormone effects on skeletal muscle growth and function in vivo. Endocrinology. 2005 Apr;146(4):1772-9. Epub 2004 Dec 23.
Sinha-Hikim I, Roth SM, Lee MI, Bhasin S. Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men. Am J Physiol Endocrinol Metab. 2003 Jul;285(1):E197-205. Epub 2003 Apr 1.
Ruzicka, L., Wettstein, A. and Kägi, H. (1935), Sexualhormone VIII. Darstellung von Testosteron unter Anwendung gemischter Ester. HCA, 18: 1478–1482.
Haupt HA, Rovere GD. Anabolic steroids: a review of the literature. Am J Sports Med. 1984 Nov-Dec;12(6):469-84. Review.
Shahidi NT. A review of the chemistry, biological action, and clinical applications of anabolic-androgenic steroids. Clin Ther. 2001 Sep;23(9):1355-90. Review.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7
Hartgens F, Kuipers H. Effects of androgenic-anabolic steroids in athletes. Sports Med. 2004;34(8):513-54. Review.
Kadi F, Eriksson A, Holmner S, Thornell LE. Effects of anabolic steroids on the muscle cells of strength-trained athletes. Med Sci Sports Exerc. 1999 Nov;31(11):1528-34.
Herbst KL, Bhasin S. Testosterone action on skeletal muscle. Curr Opin Clin Nutr Metab Care. 2004 May;7(3):271-7. Review.
Eriksson A, Kadi F, Malm C, Thornell LE. Skeletal muscle morphology in power-lifters with and without anabolic steroids. Histochem Cell Biol. 2005 Aug;124(2):167-75.
Kadi F. Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle. A basis for illegal performance enhancement. Br J Pharmacol. 2008 Jun;154(3):522-8. Epub 2008 Apr 14. Review.
Bhasin S, Woodhouse L, Casaburi R, Singh AB, Bhasin D, Berman N, Chen X, Yarasheski KE, Magliano L, Dzekov C, Dzekov J, Bross R, Phillips J, Sinha-Hikim I, Shen R, Storer TW. Testosterone dose-response relationships in healthy young men. Am J Physiol Endocrinol Metab. 2001 Dec;281(6):E1172-81.
Woodhouse LJ, Reisz-Porszasz S, Javanbakht M, Storer TW, Lee M, Zerounian H, Bhasin S. Development of models to predict anabolic response to testosterone administration in healthy young men. Am J Physiol Endocrinol Metab. 2003 May;284(5):E1009-17.
Bhasin S, Woodhouse L, Casaburi R, Singh AB, Mac RP, Lee M, Yarasheski KE, Sinha-Hikim I, Dzekov C, Dzekov J, Magliano L, Storer TW. Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle. J Clin Endocrinol Metab. 2005 Feb;90(2):678-88. Epub 2004 Nov 23.
Kvorning T, Andersen M, Brixen K, Madsen K. Suppression of endogenous testosterone production attenuates the response to strength training: a randomized, placebo-controlled, and blinded intervention study. Am J Physiol Endocrinol Metab. 2006 Dec;291(6):E1325-32. Epub 2006 Jul 25.
Sinha-Hikim I, Artaza J, Woodhouse L, Gonzalez-Cadavid N, Singh AB, Lee MI, Storer TW, Casaburi R, Shen R, Bhasin S. Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. Am J Physiol Endocrinol Metab. 2002 Jul;283(1):E154-64.
Bhasin S, Storer TW, Berman N, Callegari C, Clevenger B, Phillips J, Bunnell TJ, Tricker R, Shirazi A, Casaburi R. The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. N Engl J Med. 1996 Jul 4;335(1):1-7.
Sinha-Hikim I, Taylor WE, Gonzalez-Cadavid NF, Zheng W, Bhasin S. Androgen receptor in human skeletal muscle and cultured muscle satellite cells: up-regulation by androgen treatment. J Clin Endocrinol Metab. 2004 Oct;89(10):5245-55.
Klover P, Chen W, Zhu BM, Hennighausen L. Skeletal muscle growth and fiber composition in mice are regulated through the transcription factors STAT5a/b: linking growth hormone to the androgen receptor. FASEB J. 2009 Sep;23(9):3140-8.
Sheffield-Moore M, Urban RJ, Wolf SE, Jiang J, Catlin DH, Herndon DN, Wolfe RR, Ferrando AA. Short-term oxandrolone administration stimulates net muscle protein synthesis in young men. J Clin Endocrinol Metab. 1999 Aug;84(8):2705-11.
Kadi F, Bonnerud P, Eriksson A, Thornell LE. The expression of androgen receptors in human neck and limb muscles: effects of training and self-administration of androgenic-anabolic steroids. Histochem Cell Biol. 2000 Jan;113(1):25-9.
de Rooy C, Grossmann M, Zajac JD, Cheung AS. Targeting muscle signaling pathways to minimize adverse effects of androgen deprivation. Endocr Relat Cancer. 2016 Jan;23(1):R15-26.
Brodsky IG, Balagopal P, Nair KS. Effects of testosterone replacement on muscle mass and muscle protein synthesis in hypogonadal men–a clinical research center study. J Clin Endocrinol Metab. 1996 Oct;81(10):3469-75.
Ferrando AA, Sheffield-Moore M, Paddon-Jones D, Wolfe RR, Urban RJ. Differential anabolic effects of testosterone and amino acid feeding in older men. J Clin Endocrinol Metab. 2003 Jan;88(1):358-62.
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Singh R, Artaza JN, Taylor WE, Gonzalez-Cadavid NF, Bhasin S. Androgens stimulate myogenic differentiation and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinology. 2003 Nov;144(11):5081-8. Epub 2003 Jul 24.
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
Mulholland DJ, Dedhar S, Coetzee GA, Nelson CC. Interaction of nuclear receptors with the Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know? Endocr Rev. 2005 Dec;26(7):898-915. Epub 2005 Aug 26. Review.
Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006 Nov 3;127(3):469-80. Review.
Singh R, Bhasin S, Braga M, Artaza JN, Pervin S, Taylor WE, Krishnan V, Sinha SK, Rajavashisth TB, Jasuja R. Regulation of myogenic differentiation by androgens: cross talk between androgen receptor/ beta-catenin and follistatin/transforming growth factor-beta signaling pathways. Endocrinology. 2009 Mar;150(3):1259-68.
Zhao JX, Hu J, Zhu MJ, Du M. Trenbolone enhances myogenic differentiation by enhancing β-catenin signaling in muscle-derived stem cells of cattle. Domest Anim Endocrinol. 2011 May;40(4):222-9.
Armstrong DD, Esser KA. Wnt/beta-catenin signaling activates growth-control genes during overload-induced skeletal muscle hypertrophy. Am J Physiol Cell Physiol. 2005 Oct;289(4):C853-9. Epub 2005 May 11.
Takada I, Kouzmenko AP, Kato S. Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat Rev Rheumatol. 2009 Aug;5(8):442-7.
Cossu G, Borello U. Wnt signaling and the activation of myogenesis in mammals. EMBO J. 1999 Dec 15;18(24):6867-72. Review.
Buckingham M. Skeletal muscle formation in vertebrates. Curr Opin Genet Dev. 2001 Aug;11(4):440-8. Review.
Polesskaya A, Seale P, Rudnicki MA. Wnt signaling induces the myogenic specification of resident CD45+ adult stem cells during muscle regeneration. Cell. 2003 Jun 27;113(7):841-52.
Tincello DG, Saunders PT, Hodgins MB, Simpson NB, Edwards CR, Hargreaves TB, Wu FC. Correlation of clinical, endocrine and molecular abnormalities with in vivo responses to high-dose testosterone in patients with partial androgen insensitivity syndrome. Clin Endocrinol (Oxf). 1997 Apr;46(4):497-506.
Foradori CD, Weiser MJ, Handa RJ. Non-genomic Actions of Androgens. Frontiers in neuroendocrinology. 2008;29(2):169-181.
Lucas-Herald AK, Alves-Lopes R, Montezano AC, Ahmed SF, Touyz RM. Genomic and non-genomic effects of androgens in the cardiovascular system: clinical implications. Clin Sci (Lond). 2017 Jul 1;131(13):1405-1418.
Münzer T, Harman SM, Hees P, Shapiro E, Christmas C, Bellantoni MF, Stevens TE, O’Connor KG, Pabst KM, St Clair C, Sorkin JD, Blackman MR. Effects of GH and/or sex steroid administration on abdominal subcutaneous and visceral fat in healthy aged women and men. J Clin Endocrinol Metab. 2001 Aug;86(8):3604-10.
Sattler FR, Castaneda-Sceppa C, Binder EF, Schroeder ET, Wang Y, Bhasin S, Kawakubo M, Stewart Y, Yarasheski KE, Ulloor J, Colletti P, Roubenoff R, Azen SP. Testosterone and growth hormone improve body composition and muscle performance in older men. J Clin Endocrinol Metab. 2009 Jun;94(6):1991-2001.
Illig R, Prader A. Effect of testosterone on growth hormone secretion in patients with anorchia and delayed puberty. J Clin Endocrinol Metab. 1970 May;30(5):615-8.
Pfeilschifter J, Scheidt-Nave C, Leidig-Bruckner G, Woitge HW, Blum WF, Wüster C, Haack D, Ziegler R. Relationship between circulating insulin-like growth factor components and sex hormones in a population-based sample of 50- to 80-year-old men and women. J Clin Endocrinol Metab. 1996 Jul;81(7):2534-40.
Erfurth EM, Hagmar LE, Sääf M, Hall K. Serum levels of insulin-like growth factor I and insulin-like growth factor-binding protein 1 correlate with serum free testosterone and sex hormone binding globulin levels in healthy young and middle-aged men. Clin Endocrinol (Oxf). 1996 Jun;44(6):659-64.
van Kesteren P, Lips P, Deville W, Popp-Snijders C, Asscheman H, Megens J, Gooren L. The effect of one-year cross-sex hormonal treatment on bone metabolism and serum insulin-like growth factor-1 in transsexuals. J Clin Endocrinol Metab. 1996 Jun;81(6):2227-32.
Veldhuis JD, Keenan DM, Mielke K, Miles JM, Bowers CY. Testosterone supplementation in healthy older men drives GH and IGF-I secretion without potentiating peptidyl secretagogue efficacy. Eur J Endocrinol. 2005 Oct;153(4):577-86.
Lewis MI, Fournier M, Storer TW, Bhasin S, Porszasz J, Ren SG, Da X, Casaburi R. Skeletal muscle adaptations to testosterone and resistance training in men with COPD. J Appl Physiol (1985). 2007 Oct;103(4):1299-310.
Mauras N, Hayes V, Welch S, Rini A, Helgeson K, Dokler M, Veldhuis JD, Urban RJ. Testosterone deficiency in young men: marked alterations in whole body protein kinetics, strength, and adiposity. J Clin Endocrinol Metab. 1998 Jun;83(6):1886-92.
Bondanelli M, Ambrosio MR, Margutti A, Franceschetti P, Zatelli MC, degli Uberti EC. Activation of the somatotropic axis by testosterone in adult men: evidence for a role of hypothalamic growth hormone-releasing hormone. Neuroendocrinology. 2003 Jun;77(6):380-7.
Veldhuis JD, Metzger DL, Martha PM Jr, Mauras N, Kerrigan JR, Keenan B, Rogol AD, Pincus SM. Estrogen and testosterone, but not a nonaromatizable androgen, direct network integration of the hypothalamo-somatotrope (growth hormone)-insulin-like growth factor I axis in the human: evidence from pubertal pathophysiology and sex-steroid hormone replacement. J Clin Endocrinol Metab. 1997 Oct;82(10):3414-20.
Weissberger AJ, Ho KK. Activation of the somatotropic axis by testosterone in adult males: evidence for the role of aromatization. J Clin Endocrinol Metab. 1993 Jun;76(6):1407-12.
Veldhuis JD, Mielke KL, Cosma M, Soares-Welch C, Paulo R, Miles JM, Bowers CY. Aromatase and 5alpha-reductase inhibition during an exogenous testosterone clamp unveils selective sex steroid modulation of somatostatin and growth hormone secretagogue actions in healthy older men. J Clin Endocrinol Metab. 2009 Mar;94(3):973-81.
Yamamoto T, Sakai C, Yamaki J, Takamori K, Yoshiji S, Kitawaki J, Fujii M, Yasuda J, Honjo H, Okada H. Estrogen biosynthesis in human liver–a comparison of aromatase activity for C-19 steroids in fetal liver, adult liver and hepatoma tissues of human subjects. Endocrinol Jpn. 1984 Jun;31(3):277-81.
Hata S, Miki Y, Saito R, Ishida K, Watanabe M, Sasano H. Aromatase in human liver and its diseases. Cancer Med. 2013 Jun;2(3):305-15.
Riggs BL, Hartmann LC. Selective estrogen-receptor modulators — mechanisms of action and application to clinical practice. N Engl J Med. 2003 Feb 13;348(7):618-29. Review. Erratum in: N Engl J Med. 2003 Mar 20;348(12):1192.
Löfgren L, Wallberg B, Wilking N, Fornander T, Rutqvist LE, Carlström K, von Schoultz B, von Schoultz E. Tamoxifen and megestrol acetate for postmenopausal breast cancer: diverging effects on liver proteins, androgens, and glucocorticoids. Med Oncol. 2004;21(4):309-18.
Centrella M, McCarthy TL, Chang WZ, Labaree DC, Hochberg RB. Estren (4-estren-3alpha,17beta-diol) is a prohormone that regulates both androgenic and estrogenic transcriptional effects through the androgen receptor. Mol Endocrinol. 2004 May;18(5):1120-30.
Yu YM, Domené HM, Sztein J, Counts DR, Cassorla F. Developmental changes and differential regulation by testosterone and estradiol of growth hormone receptor expression in the rabbit. Eur J Endocrinol. 1996 Nov;135(5):583-90.
Zung A, Phillip M, Chalew SA, Palese T, Kowarski AA, Zadik Z. Testosterone effect on growth and growth mediators of the GH-IGF-I axis in the liver and epiphyseal growth plate of juvenile rats. J Mol Endocrinol. 1999 Oct;23(2):209-21.
Hayes VY, Urban RJ, Jiang J, Marcell TJ, Helgeson K, Mauras N. Recombinant human growth hormone and recombinant human insulin-like growth factor I diminish the catabolic effects of hypogonadism in man: metabolic and molecular effects. J Clin Endocrinol Metab. 2001 May;86(5):2211-9.
Sculthorpe N, Solomon AM, Sinanan AC, Bouloux PM, Grace F, Lewis MP. Androgens affect myogenesis in vitro and increase local IGF-1 expression. Med Sci Sports Exerc. 2012 Apr;44(4):610-5.
Birzniece V, Meinhardt UJ, Umpleby MA, Handelsman DJ, Ho KK. Interaction between testosterone and growth hormone on whole-body protein anabolism occurs in the liver. J Clin Endocrinol Metab. 2011 Apr;96(4):1060-7.
Mertani HC, Delehaye-Zervas MC, Martini JF, Postel-Vinay MC, Morel G. Localization of growth hormone receptor messenger RNA in human tissues. Endocrine. 1995 Feb;3(2):135-42.
Florini JR, Ewton DZ, Coolican SA. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr Rev. 1996 Oct;17(5):481-517. Review.
Urban RJ, Bodenburg YH, Gilkison C, Foxworth J, Coggan AR, Wolfe RR, Ferrando A. Testosterone administration to elderly men increases skeletal muscle strength and protein synthesis. Am J Physiol. 1995 Nov;269(5 Pt 1):E820-6.
Ewton DZ, Coolican SA, Mohan S, Chernausek SD, Florini JR. Modulation of insulin-like growth factor actions in L6A1 myoblasts by insulin-like growth factor binding protein (IGFBP)-4 and IGFBP-5: a dual role for IGFBP-5. J Cell Physiol. 1998 Oct;177(1):47-57.
Venken K, Movérare-Skrtic S, Kopchick JJ, Coschigano KT, Ohlsson C, Boonen S, Bouillon R, Vanderschueren D. Impact of androgens, growth hormone, and IGF-I on bone and muscle in male mice during puberty. J Bone Miner Res. 2007 Jan;22(1):72-82.
Serra C, Bhasin S, Tangherlini F, Barton ER, Ganno M, Zhang A, Shansky J, Vandenburgh HH, Travison TG, Jasuja R, Morris C. The role of GH and IGF-I in mediating anabolic effects of testosterone on androgen-responsive muscle. Endocrinology. 2011 Jan;152(1):193-206.
Spangenburg EE, Le Roith D, Ward CW, Bodine SC. A functional insulin-like growth factor receptor is not necessary for load-induced skeletal muscle hypertrophy. J Physiol. 2008 Jan 1;586(1):283-91.
Yoshizawa A, Clemmons DR. Testosterone and insulin-like growth factor (IGF) I interact in controlling IGF-binding protein production in androgen-responsive foreskin fibroblasts. J Clin Endocrinol Metab. 2000 Apr;85(4):1627-33.
Gayan-Ramirez G, Rollier H, Vanderhoydonc F, Verhoeven G, Gosselink R, Decramer M. Nandrolone decanoate does not enhance training effects but increases IGF-I mRNA in rat diaphragm. J Appl Physiol (1985). 2000 Jan;88(1):26-34.
Lewis MI, Horvitz GD, Clemmons DR, Fournier M. Role of IGF-I and IGF-binding proteins within diaphragm muscle in modulating the effects of nandrolone. Am J Physiol Endocrinol Metab. 2002 Feb;282(2):E483-90
Salmons S. Myotrophic effects of an anabolic steroid in rabbit limb muscles. Muscle Nerve. 1992 Jul;15(7):806-12.
Bisschop A, Gayan-Ramirez G, Rollier H, Dekhuijzen PN, Dom R, de Bock V, Decramer M. Effects of nandrolone decanoate on respiratory and peripheral muscles in male and female rats. J Appl Physiol (1985). 1997 Apr;82(4):1112-8
Lewis MI, Fournier M, Yeh AY, Micevych PE, Sieck GC. Alterations in diaphragm contractility after nandrolone administration: an analysis of potential mechanisms. J Appl Physiol (1985). 1999 Mar;86(3):985-92.
Heitzman RJ. The effectiveness of anabolic agents in increasing rate of growth in farm animals; report on experiments in cattle. Environ Qual Saf Suppl. 1976;(5):89-98. Review.
Buttery, P., Vernon, B., & Pearson, J. (1978). Anabolic agents—some thoughts on their mode of action. Proceedings of the Nutrition Society, 37(3), 311-315.
Hongerholt DD, Crooker BA, Wheaton JE, Carlson KM, Jorgenson DM. Effects of a growth hormone-releasing factor analogue and an estradiol-trenbolone acetate implant on somatotropin, insulin-like growth factor I, and metabolite profiles in growing Hereford steers. J Anim Sci. 1992 May;70(5):1439-48.
Tan RS, Scally MC. Anabolic steroid-induced hypogonadism–towards a unified hypothesis of anabolic steroid action. Med Hypotheses. 2009 Jun;72(6):723-8.
Kamanga-Sollo E, White ME, Hathaway MR, Weber WJ, Dayton WR. Effect of Estradiol-17beta on protein synthesis and degradation rates in fused bovine satellite cell cultures. Domest Anim Endocrinol. 2010 Jul;39(1):54-62.
Kamanga-Sollo E, Thornton KJ, White ME, Dayton WR. Role of G protein-coupled estrogen receptor-1, matrix metalloproteinases 2 and 9, and heparin binding epidermal growth factor-like growth factor in estradiol-17β-stimulated bovine satellite cell proliferation. Domest Anim Endocrinol. 2014 Oct;49:20-6.
Dunn JD, Johnson BJ, Kayser JP, Waylan AT, Sissom EK, Drouillard JS. Effects of flax supplementation and a combined trenbolone acetate and estradiol implant on circulating insulin-like growth factor-I and muscle insulin-like growth factor-I messenger RNA levels in beef cattle. J Anim Sci. 2003 Dec;81(12):3028-34.
Pampusch MS, Johnson BJ, White ME, Hathaway MR, Dunn JD, Waylan AT, Dayton WR. Time course of changes in growth factor mRNA levels in muscle of steroid-implanted and nonimplanted steers. J Anim Sci. 2003 Nov;81(11):2733-40.
Pampusch MS, White ME, Hathaway MR, Baxa TJ, Chung KY, Parr SL, Johnson BJ, Weber WJ, Dayton WR. Effects of implants of trenbolone acetate, estradiol, or both, on muscle insulin-like growth factor-I, insulin-like growth factor-I receptor, estrogen receptor-{alpha}, and androgen receptor messenger ribonucleic acid levels in feedlot steers. J Anim Sci. 2008 Dec;86(12):3418-23.
Thompson SH, Boxhorn LK, Kong WY, Allen RE. Trenbolone alters the responsiveness of skeletal muscle satellite cells to fibroblast growth factor and insulin-like growth factor I. Endocrinology. 1989 May;124(5):2110-7.
Johnson BJ, Halstead N, White ME, Hathaway MR, DiCostanzo A, Dayton WR. Activation state of muscle satellite cells isolated from steers implanted with a combined trenbolone acetate and estradiol implant. J Anim Sci. 1998 Nov;76(11):2779-86.
Dalbo VJ, Roberts MD, Mobley CB, Ballmann C, Kephart WC, Fox CD, Santucci VA, Conover CF, Beggs LA, Balaez A, Hoerr FJ, Yarrow JF, Borst SE, Beck DT. Testosterone and trenbolone enanthate increase mature myostatin protein expression despite increasing skeletal muscle hypertrophy and satellite cell number in rodent muscle. Andrologia. 2017 Apr;49(3).
Bhasin S, He EJ, Kawakubo M, Schroeder ET, Yarasheski K, Opiteck GJ, Reicin A, Chen F, Lam R, Tsou JA, Castaneda-Sceppa C, Binder EF, Azen SP, Sattler FR. N-terminal propeptide of type III procollagen as a biomarker of anabolic response to recombinant human GH and testosterone. J Clin Endocrinol Metab. 2009 Nov;94(11):4224-33.
Nelson AE, Meinhardt U, Hansen JL, Walker IH, Stone G, Howe CJ, Leung KC, Seibel MJ, Baxter RC, Handelsman DJ, Kazlauskas R, Ho KK. Pharmacodynamics of growth hormone abuse biomarkers and the influence of gender and testosterone: a randomized double-blind placebo-controlled study in young recreational athletes. J Clin Endocrinol Metab. 2008 Jun;93(6):2213-22.
Holt RI. Detecting growth hormone misuse in athletes. Indian J Endocrinol Metab. 2013 Oct;17(Suppl 1):S18-22.
Tan SH, Lee A, Pascovici D, Care N, Birzniece V, Ho K, Molloy MP, Khan A. Plasma biomarker proteins for detection of human growth hormone administration in athletes. Sci Rep. 2017 Aug 30;7(1):10039.
Jørgensen JO, Jessen N, Pedersen SB, Vestergaard E, Gormsen L, Lund SA, Billestrup N. GH receptor signaling in skeletal muscle and adipose tissue in human subjects following exposure to an intravenous GH bolus. Am J Physiol Endocrinol Metab. 2006 Nov;291(5):E899-905.
Liu X, Robinson GW, Gouilleux F, Groner B, Hennighausen L. Cloning and expression of Stat5 and an additional homologue (Stat5b) involved in prolactin signal transduction in mouse mammary tissue. Proc Natl Acad Sci U S A. 1995 Sep 12;92(19):8831-5.
Hennighausen L, Robinson GW. Interpretation of cytokine signaling through the transcription factors STAT5A and STAT5B. Genes Dev. 2008 Mar 15;22(6):711-21.
Eshet R, Laron Z, Pertzelan A, Arnon R, Dintzman M. Defect of human growth hormone receptors in the liver of two patients with Laron-type dwarfism. Isr J Med Sci. 1984 Jan;20(1):8-11.
Teglund S, McKay C, Schuetz E, van Deursen JM, Stravopodis D, Wang D, Brown M, Bodner S, Grosveld G, Ihle JN. Stat5a and Stat5b proteins have essential and nonessential, or redundant, roles in cytokine responses. Cell. 1998 May 29;93(5):841-50.
Hwa V, Little B, Adiyaman P, Kofoed EM, Pratt KL, Ocal G, Berberoglu M, Rosenfeld RG. Severe growth hormone insensitivity resulting from total absence of signal transducer and activator of transcription 5b. J Clin Endocrinol Metab. 2005 Jul;90(7):4260-6. Epub 2005 Apr 12.
Rowland JE, Lichanska AM, Kerr LM, White M, d’Aniello EM, Maher SL, Brown R, Teasdale RD, Noakes PG, Waters MJ. In vivo analysis of growth hormone receptor signaling domains and their associated transcripts. Mol Cell Biol. 2005 Jan;25(1):66-77. Erratum in: Mol Cell Biol. 2005 Mar;25(5):2072.
Klover P, Hennighausen L. Postnatal body growth is dependent on the transcription factors signal transducers and activators of transcription 5a/b in muscle: a role for autocrine/paracrine insulin-like growth factor I. Endocrinology. 2007 Apr;148(4):1489-97.
Barclay JL, Kerr LM, Arthur L, Rowland JE, Nelson CN, Ishikawa M, d’Aniello EM, White M, Noakes PG, Waters MJ. In vivo targeting of the growth hormone receptor (GHR) Box1 sequence demonstrates that the GHR does not signal exclusively through JAK2. Mol Endocrinol. 2010 Jan;24(1):204-17.
Hwa V, Nadeau K, Wit JM, Rosenfeld RG. STAT5b deficiency: lessons from STAT5b gene mutations. Best Pract Res Clin Endocrinol Metab. 2011 Feb;25(1):61-75.
Varco-Merth B, Feigerlová E, Shinde U, Rosenfeld RG, Hwa V, Rotwein P. Severe growth deficiency is associated with STAT5b mutations that disrupt protein folding and activity. Mol Endocrinol. 2013 Jan;27(1):150-61.
Davey HW, Xie T, McLachlan MJ, Wilkins RJ, Waxman DJ, Grattan DR. STAT5b is required for GH-induced liver IGF-I gene expression. Endocrinology. 2001 Sep;142(9):3836-41.
Woelfle J, Chia DJ, Rotwein P. Mechanisms of growth hormone (GH) action. Identification of conserved Stat5 binding sites that mediate GH-induced insulin-like growth factor-I gene activation. J Biol Chem. 2003 Dec 19;278(51):51261-6. Epub 2003 Oct 7.
Woelfle J, Billiard J, Rotwein P. Acute control of insulin-like growth factor-I gene transcription by growth hormone through Stat5b. J Biol Chem. 2003 Jun 20;278(25):22696-702. Epub 2003 Apr 7.
MacLean HE, Chiu WS, Notini AJ, Axell AM, Davey RA, McManus JF, Ma C, Plant DR, Lynch GS, Zajac JD. Impaired skeletal muscle development and function in male, but not female, genomic androgen receptor knockout mice. FASEB J. 2008 Aug;22(8):2676-89.
Tan SH, Dagvadorj A, Shen F, Gu L, Liao Z, Abdulghani J, Zhang Y, Gelmann EP, Zellweger T, Culig Z, Visakorpi T, Bubendorf L, Kirken RA, Karras J, Nevalainen MT. Transcription factor Stat5 synergizes with androgen receptor in prostate cancer cells. Cancer Res. 2008 Jan 1;68(1):236-48.
de Souza GL, Hallak J. Anabolic steroids and male infertility: a comprehensive review. BJU Int. 2011 Dec;108(11):1860-5.
Sono ormai alcuni decenni che i fautori de “le calorie non contano” e quelli, conservatori, de “le calorie contano” si scontrano cercando di argomentare (più o meno) scientificamente le loro affermazioni. Tralasciando la questione non poco importante della trattazione superficiale che spesso caratterizza entrambe le fazioni, la schiera dei “negazionisti delle calorie” tocca livelli di scientismo e di contraddizione abbastanza evidenti (mai letto un loro libro?). Essi cercano spiegazioni (all’apparenza) plausibili per avvalorare la loro tesi. Una di esse è perfettamente rappresentata da un recente studio svolto da scienziati dello sport dell’Università di Malaga i quali hanno osservato l’effetto di una dieta chetogenica ipercalorica su atleti di forza.(1)
I ricercatori hanno reclutato 24 uomini in buona salute, che si erano allenati con pesi per almeno 2 anni, e gli hanno sottoposti ad un identico allenamento finalizzato all’ipertrofia per 8 settimane: il piano allenante era suddiviso in 2 giorni di Upper Body e 2 giorni Lower Body, con 72 ore di riposo tra le sessioni per favorire il recupero. I ricercatori hanno diviso i partecipanti allo studio in 3 gruppi e hanno dato ad ogni gruppo una dieta diversa.
Panoramica del protocollo di allenamento. WK: allenamento (microciclo); UL: Upper-Limb; LL: Lower-Limb; R: Riposo; 30 X: 3s nell’eccentrica e movimento esplosivo durante la fase concentrica.
I soggetti del primo gruppo (gruppo di controllo), ovviamente, hanno continuato a mangiare come era loro abitudine [CG].
I soggetti del secondo gruppo hanno ricevuto un piano alimentare il quale presentava un introito proteico pari a 2g di proteine per chilogrammo di peso corporeo al giorno, e un carico glucidico di massimo 42g di carboidrati al giorno [KD]. L’apporto calorico era alto. Ogni giorno i soggetti presi in esame dovevano ingerire più di 3000Kcal.
In fine, i soggetti del terzo gruppo hanno ricevuto un piano alimentare più o meno “normale” e con un buon apporto proteico [NKD]. Anche i soggetti di questo gruppo dovevano ingerire più di 3000Kcal al giorno.
Al termine del periodo di test, è stato osservato che i soggetti sottoposti ad una dieta a basso contenuto di carboidrati avevano ridotto la loro massa grassa generale, compreso il grasso nella cavità addominale. Quest’ultimo tipo di grasso è noto per essere un fattore predittivo per lo sviluppo di diverse patologie. La massa magra di questi soggetti è rimasta costante.
I ricercatori hanno concluso dicendo che, secondo i loro risultati, i soggetti sottoposti a un allenamento contro-resistenza durante una dieta chetogenica hanno sperimentato una maggiore riduzione della massa grassa e del tessuto adiposo viscerale, rispetto al gruppo che ha seguito un regime alimentare non chetogenico. La maggiore riduzione del tessuto adiposo viscerale può avere una certa rilevanza clinica a causa della sua associazione inversa al rischio cardio-metabolico.
I ricercatori continuano dicendo che, sono necessari ulteriori studi per valutare i vantaggi di questa combinazione (allenamento contro-resistenza e dieta chetogenica) in soggetti con un eccesso di massa grassa, con particolare attenzione alla significativa riduzione riportata a carico del tessuto adiposo viscerale, che potrebbe essere di grande beneficio per questa fascia di popolazione dato anche il fatto che la massa corporea magra è mantenuta. E aggiungono che, in effetti, questa ricerca non ha mostrato cambiamenti significativi sulla massa magra, nonostante la condizione iperenergetica e l’elevato apporto proteico (2,0g/kg/die) assunto dai soggetti sottoposti ad allenamenti contro-resistenza del gruppo “dieta chetogenica”. Quindi, essi concludono, non poco banalmente, che gli approcci dietetici a basso contenuto di carboidrati non sarebbero una strategia ottimale per la costruzione di massa muscolare in uomini allenati nelle condizioni poste da questo studio (protocollo di allenamento contro-resistenza focalizzato sulla tensione meccanica della durata di 8 settimane).
Immagino già la “standing ovation” dei detrattori della termodinamica… mantenete la calma, lo studio qui brevemente esposto ha diverse limitazioni….
Le limitazioni del presente studio vanno dalle limitate misurazioni dei risultati, dal numero ridotto di partecipanti e dal tempo del test (8 settimane); tutti fattori che di per se riducono il valore dello studio. E, cosa assai rilevante, durante lo studio non sono state eseguite valutazioni sulla corretta assunzione calorica dei soggetti presi in esame e non è stata presa in considerazione la soppressione dell’appetito derivante da una dieta ricca di proteine e grassi. Quindi, è possibile, anzi è molto probabile, che si siano verificate variazioni nell’assunzione calorica anche se i partecipanti sono stati istruiti a seguire specifiche raccomandazioni dietetiche. In parole povere, i soggetti sottoposti a regime chetogenico hanno, con tutta probabilità, seguito un regime leggermente ipocalorico in modo indiretto come risposta alle variabili indotte dalla specifica strategia alimentare. Ed ecco spiegato, in modo assai più scientifico, il perché del calo della massa grassa e il mantenimento della massa muscolare. Quest’ultimo elemento trova la sua logica spiegazione nell’introito proteico giornaliero (2g/Kg/die) il quale ha garantito un effetto di “protein sparing”. Il numero limitato delle settimane del test (8 settimane) è stato senza dubbio incisivo anche in questo. E non dimentichiamoci del fatto che, guarda caso, non si è verificato alcun aumento della massa muscolare ne della massa corporea in generale. Evento questo decisamente ai “confini della realtà” se rapportato a soggetti in fisiologia sottoposti ad un regime ipercalorico (di qualsiasi natura esso sia).
Bomba Calorimetrica
Sebbene l’uomo non sia una macchina calorimetrica, che un acido polinsaturo non equivale ad uno saturo, ecc… bisognerebbe tuttavia ricordarsi che facciamo parte di questo universo e come tutta la materia rispondiamo alle Leggi delle Termodinamica e come tutti gli essere viventi a quelle della Bioenergia:
“Queste leggi descrivono come il calore (o energia termica) viene convertito da e per diversi tipi di energia, e l’effetto che questo può avere sulle varie forme di materia.”
Detto in termini semplici, quando si mangia si introduce dell’energia, questa energia non può sparire nel nulla.
Anche se un soggetto possa avere problemi d’assorbimento intestinali, disfunzioni delle proteine mitocondriali UCP2-3, una grave insulino-resistenza, o soffra di ipotiroidismo in ogni caso l’energia che introduce si trasforma sempre seguendo le leggi universali.
Ogni soggetto ha un proprio equilibrio energetico, quanto sia elevato questo equilibrio dipende dallo stato metabolico. Questi due fattori si influenzano a vicenda e sono imprescindibili l’uno dall’alto.
Semplice schema rappresentante le varie componenti di un corretto piano alimentare con il loro grado di importanza.
Per quanto vi possa irritare, alla fine della giostra, una caloria rimane pur sempre una caloria…
Se non avete ancora letto la prima parte di questa serie di articoli vi invito a farlo prima di procedere con la lettura di questa seconda parte: 1° Parte.
Informazioni base sull’IGF-1
IGF-1
Nella prima parte di questa serie di articoli ho esposto la storia e cronologia legate alla scoperta del GH e dll’IGF-1. Tuttavia, vorrei approfondire l’argomento riguardante i membri della famiglia dei fattori di crescita insulino-similimi, esponendo le loro caratteristiche e i loro ruoli nel processo ipertrofico. Gli IGF sono una famiglia di peptidi, in gran parte dipendenti dal GH, che mediano molte delle azioni di stimolo della crescita date dal GH.[121] Il fegato è il principale responsabile di tutta la produzione endocrina di IGF-1, con circa il 75% della sintesi a carico epatico sotto la regolazione del GH.[83,122-123] Ciò presuppone che ci sia un apporto macro-calorico sufficiente e livelli elevati di Insulina portale.[124-125] La sintesi autocrina di IGF-1 è anche regolata dal GH, in aggiunta ad altri fattori autocrini dipendenti dal tessuto.[126-128]
IGF-2
Alla famiglia degli IGF appartengono oltre dieci proteine strutturalmente simili tra cui IGF-1, IGF-2, Insulina, Relaxina e Pro-Insulina.[129] Sono tutti altamente omologhi sia nella struttura che nella funzione e gli effetti metabolici dell’IGF-1 sono stati definiti “insulino-simili” proprio a causa delle somiglianze e dei percorsi che condividono l’uno con l’altro. L’IGF-1 ha un’omologia di sequenza aminoacidica superiore al 50% con l’Insulina e il recettore del IGF-1 ha un’omologia della sequenza aminoacidica del 60% con il recettore dell’Insulina.[121,130-131] Gli evoluzionisti suppongono che la somiglianze nella struttura di questi peptidi sia dovuta al fatto che essi si siano evoluti da una singola molecola precursore trovata in vertebrati di oltre 60 milioni di anni fa.[132] Sia il rilascio di IGF-1 che quello di Insulina sono stimolati dall’assunzione di cibo, rilascio che viene inibito in condizioni di digiuno.[83]
Recettore del IGF-1
A causa di queste somiglianze strutturali, i membri della famiglia IGF possono spesso legarsi con i recettori nativi in modo “incrociato”.[133] Per riassumere brevemente queste relazioni di legame, la molecola di IGF-1 si lega con il recettore del IGF-1 con un elevata affinità, tuttavia sia l’IGF-2 che l’Insulina possono legarsi al recettore del IGF-1, ma con una affinità significativamente inferiori. L’IGF-2 si lega al recettore del IGF-2 con una elevata affinità, e l’IGF-1 si lega a questo recettore con un’affinità inferiore mentre l’Insulina non presenta alcun legame con esso.
Adipocita
La famiglia dei recettori IGF ha densità che variano significativamente in base ai tipi di cellule in cui sono presenti.[132] Questo è uno dei motivi per cui l’Insulina e l’IGF-1 possono avere diverse azioni metaboliche nonostante siano strutturalmente simili. Cellule come gli epatociti e gli adipociti hanno molti più recettori dell’Insulina rispetto ai recettori del IGF-1. Al contrario, le cellule muscolari lisce vascolari situate nei vasi sanguigni hanno un numero significativamente più elevato di recettori del IGF-1 rispetto ai recettori dell’Insulina.
Poiché ho già fatto un’analisi approfondita in precedenza sulle basi chimiche che si verificano durante l’attivazione del GHR, non mi ripeterò su tale questione. Però, è necessario comprendere che la famiglia dei recettori IGF è anche attivata dalla Tirosina Chinasi che, come ora sappiamo, porta alla fosforilazione dei substrati, all’attivazione delle vie cellulari e infine all’espressione genica e alla sintesi proteica.[121] L’attivazione del recettore del IGF-1 sembra essere indipendente dall’isoforma da cui è stato prodotto l’IGF-1. Inoltre, si noti che entrambi i tipi di recettori IGF sono stati trovati nelle cellule muscolari umane.[134]
I livelli serici di IGF-1 sono stabili negli adulti sani e vi sono poche variazioni da un giorno all’altro, o anche da una settimana all’altra. In effetti, osservare i livelli serici di IGF-1 di un individuo può essere un indicatore abbastanza affidabile di eventuali problemi di sensibilità al GH in relazione ad intervalli ben definiti, corretti per età e sesso.[135] Naturalmente, quando si cerca di decidere se esistono reali problemi di sensibilità, è necessario prendere in considerazione aspetti come lo stato nutrizionale generale dell’individuo e la salute epatica.
IGFBP-1
Nel flusso sanguineo, l’IGF-1 esiste principalmente in forma legata a proteine leganti l’IGF (IGFBP). La superfamiglia IGFBP comprende sei proteine ad alta affinità che vanno dal IGFBP-1 al IGFBP-6, nonché un certo numero di proteine a bassa affinità denominate proteine legate all’IGFBP.[136] Quasi il 95% di tutto l’IGF-1 circolante esiste in forma legata, con circa il 75% legato specificamente con l’IGFBP-3.[137] Una piccola frazione di IGF-1 (normalmente inferiore al 5%) può anche esistere in forma libera, e queste molecole non legate agiscono come regolatore negativo della secrezione di GH.[104] Gli IGFBP possono legarsi con l’IGF-1 e l’IGF-2, ma non con l’insulina. [138]
Azioni regolatorie delle proteine leganti l’IGF-1
L’IGF-1 legato esiste più comunemente in un complesso ternario da 150-kDa mentre è nel circolo ematico. Questo complesso ternario è costituito da una molecola di IGF-1, dal IGFBP-3 e dalla subunità labile acida (ALS) – sebbene possa esistere in un complesso binario con altri IGFBP.[139-140] Questi complessi servono a scopi come l’aumento della biodisponibilità degli IGF circolanti, estendendo la loro emivita serica, trasportando gli IGF alle cellule bersaglio e modulando l’interazione degli IGF con i loro rispettivi recettori di membrana posti sulla superficie delle cellule.[141-144] Ad esempio, nel plasma, il complesso ternario stabilizza IGF-1, aumentando significativamente la sua emivita da meno di 5 minuti a oltre 16 ore in alcuni casi.[137]
Gli IGFB sembrano normalmente inibire l’azione degli IGF, e questo perché competono con i recettori IGF per l’affinità di legame con gli IGF.[145] Tuttavia, non è sempre così, poiché gli IGFBP sono anche in grado di potenziare le azioni dell’IGF, potenzialmente facilitando la consegna dell’IGF al recettore.[146] Sebbene esista un’interazione piuttosto complessa, basti ricordare che il ruolo principale degli IGFBP è quello di trasportare gli IGF dal flusso ematico ai tessuti periferici. Una volta che ciò è avvenuto, gli IGFBP vengono rilasciati dai complessi binari e ternari mediante proteolisi o tramite legame alla matrice extracellulare del recettore del IGF-1.[147] Una volta rilasciate, le molecole di IGF-1 diventano libere, attive e possono quindi esplicare la loro azione.[137,143]
Una volta nei tessuti, gli IGFBP modulano le azioni dell’IGF in quanto hanno una maggiore affinità per il sito recettore rispetto all’IGF stesso [148], tuttavia essi possono anche esercitare effetti indipendenti dall’IGF.[149] Alcuni degli effetti diretti del IGFBP che sono già stati chiariti includono l’inibizione della crescita, l’induzione diretta dell’apoptosi e la modulazione degli effetti dei fattori di crescita non-IGF.[121]
È anche noto che lo splicing alternativo del gene IGF-1 produce tre isoforme distinte nell’uomo che hanno sia azioni dirette che indirette che contribuiscono agli effetti di promozione della crescita del IGF-1.[150-151] Sebbene non siano richieste per la secrezione di IGF-1, queste isoforme possono aumentare la reale biodisponibilità del IGF-1 al suo sito recettore.[437] Le tre isoforme sono denominate IGF-1Ea, IGF-1Eb e IGF-1Ec. E’ corretto sottolineare il fatto che roditori e pesci posseggono solo due isoforme, e in questo articolo ci si riferirà solo alle isoforme umane, a meno che non sia chiaramente indicato diversamente, per spera in questo modo di creare meno confusione al lettore.
L’IGF-1Ea è simile all’isoforma IGF principale espressa dagli epatociti e ha l’esone 4 del gene IGF-1 maturo giuntato direttamente all’esone 6.[152] Si pensa che l’IGF-1Eb sia prevalentemente espresso nel fegato, ma il suo ruolo nei muscoli non è ancora del tutto chiaro.[153] Si estende più a valle dell’esone 5, ma solo i primi 17 aminoacidi di questa isoforma sono identici a quelli della variante finale dell’isoforma che tratterò a momenti[154]. Si ritiene che questa isoforma sia unica per i primati, e per l’uomo, in quanto non è stata trovata nei roditori o nei pesci.[155]
MGF
L’IGF-1Ec è anche chiamato Fattore di Crescita Maccanico (MGF) ed è chiamato così perché è espresso in risposta a tensione meccanica e stress.[156-157] Nella prima parte di questa serie di articoli ho riportato in breve come questi siano due dei principali meccanismi alla base del processo ipertrofico all’interno del muscolo scheletrico. Tratterò più nel dettaglio dell’MGF in seguito. Questa isoforma contiene parte dell’esone 5 giuntata all’esone 6 che si traduce in un frame-shift e questo dal mRNA è tradotto in una isoforma con una alternativa sequenza di 25 aminoacidi al C-terminale.[152] Nei roditore l’IGF-1Eb condivide un’alta omologia con l’MGF umano ed entrambi sono spesso usati in modo intercambiabile in letteratura.[158] Riporto ciò solamente perché può diventare forviante quando si esamina la letteratura scientifica trattante questa isoforma, specialmente quando si passa da modelli animali a umani.
L’MGF ha mostrato in modelli di colture cellulari di aumentare la proliferazione e la migrazione dei mioblasti, oltre ad essere coinvolto nell’attivazione delle cellule satelliti. Comunque sia, se ciò si verifichi in vivo è a tutt’oggi una fonte di contesa. Questo comportamento è stato visto anche in presenza dell’inattivazione del IGF-1, il che suggerisce che l’MGF abbia la capacità di operare indipendentemente dall’IGF-1 maturo.[438] Detto questo, tutte le isoforme di IGF-1 richiedono un recettore del IGF-1 funzionale per produrre effettivamente ipertrofia muscolare poiché tale recettore non viene influenzato in assenza di IGF-1 maturo.[439-440] La risposta effettiva al MGF si riscontra in un ambiente con un pool attivo di cellule satelliti, poiché i tessuti muscolari invecchiati sono normalmente in uno stato di dormienza. Infine, sebbene l’MGF iniettabile sia quasi una chimera nel ambiente del BodyBuilding, osservando la totalità del suo effetto sembra avere una minore attività muscolare rispetto all’IGF-1 maturo, quindi il suo valore intrinseco per i Bodybuilder può effettivamente essere stato sovrastimato.[442]
Somatopausa
Lo studio di soggetti anziani offre una prospettiva alquanto singolare dal momento che è noto come la sarcopenia, termine che sta ad indicare una degenerazione del tessuto muscolare, si manifesti con l’avanzare dell’età. È anche noto che i livelli di GH secreto e IGF-1 circolante diminuiscono gradualmente nel corso della vita dopo il picco della pubertà.[159-162] Il declino dei livelli ormonali è piuttosto marcato, con una diminuzione della secrezione di GH del 10-15% ogni decennio dopo i 20 anni.[163] Molti anni fa è stato suggerito che questi cambiamenti senescenti nella composizione corporea e nelle funzioni metaboliche sono direttamente correlati alla diminuzione dei livelli ormonali all’interno dell’Asse GH/IGF-1. La comunità scientifica ha coniato il termine “somatopausa” per descrivere questo fenomeno.[159,164]
In sintesi, i processi che caratterizzano l’”ipotesi della Somatopausa” [128] sono:
I cambiamenti nello stile di vita e le predisposizioni genetiche promuovono l’accumulo di grasso corporeo con l’avanzare dell’età;
Questa maggiore massa grassa aumenta la disponibilità di FFA e quindi induce insulino-resistenza;
Livelli elevati di Insulina sopprimono l’IGFBP-1 determinando un aumento relativo dei livelli di IGF-1 liberi;
Aumenti sistemici degli FFA, dell’Insulina e una soppressione del rilascio ipofisario di GH dato dal aumento del IGF-1, portano ad un ulteriore aumento della massa grassa;
Il GH endogeno viene eliminato più rapidamente nei soggetti con maggiore massa grassa
Come si può vedere, il grasso corporeo aumenta con l’avanzare dell’età, di conseguenza si verifica un peggioramento dell’insulino-resistenza seguita da un aumento dei livelli di Insulina che, attraverso la soppressione del IGFBP-1 e all’aumento del IGF-1 libero, porta alla soppressione della secrezione di GH, che a sua volta peggiora la composizione corporea (vedi aumento della percentuale di grasso). È piuttosto interessante vedere che la frequenza degli impulsi di GH rimane essenzialmente intatta. L’attenuazione legata all’età è in realtà solo una marcata riduzione dell’ampiezza dell’impulso insieme ad un aumento della secrezione di SRIF.[165]
I cambiamenti associati alla somatopausa sono molto simili a quelli osservati negi giovani adulti con deficit dell’Ormone della Crescita (GHD). Sebbene simili, le persone anziane non sono normalmente così gravemente colpite da questa condizione rispetto agli individui con GHD e la somatopause non è considerata uno stato patologico.[160,166-167] Esempi di alcuni dei cambiamenti associati alla somatopausa includono la riduzione della massa muscolare e ossea, riduzione della forza, diminuzione dell’attività fisica e della capacità cardiaca, aumento del grasso corporeo (in particolare nella regione viscerale) e deterioramento cognitivo.
A causa del desiderio di invertire i molti effetti dannosi legati all’invecchiamento, vi è una diffusa ipotesi secondo cui la somministrazione di GH possa essere d’aiuto come parte di un programma di terapia ormonale sostitutiva completa (HRT). Una revisione completa riguardante HRT e anziani va oltre lo scopo di questo articolo, ma quelli che sono interessati possono trovare una discussione recente sull’argomento qui.[168]
L’ormone della crescita migliora le prestazioni atletiche?
Ben Johnson alle Olimpiadi di Seul del 1988.
L’emergere del GH come farmaco per il miglioramento delle prestazioni (PED), al di fuori dei circoli ristretti del BodyBuilding, è in gran parte attribuito al rilascio del famigerato “Underground Steroid Handbook” nei primi anni ’80.[169] Successivamente, il GH ha colpito l’opinione pubblica quando il vincitore della medaglia d’oro delle Olimpiadi del 1988, Ben Johnson, ammise di usarlo insieme agli AAS dopo essere stato privato del suo titolo in seguito alla positività del test anti-doping al quale era stato sottoposto.[170] Durante quel periodo, si credeva comunemente che l’uso del GH avrebbe aumentato la massa muscolare e contemporaneamente migliorato gli aspetti della prestazione atletica.[171] E, come risposta a questa convinzione, nel 1989 il CIO ha vietato l’uso del GH definendolo un PED e inserendolo come parte di una nuova classe di farmaci dopanti denominata “ormoni peptidici e analoghi”. Ha vietato l’uso del GH nonostante in quel momento vi fosse la mancanza di un test legittimo per la rilevazione del rHGH.[172] Nonostante tutto ciò, la domanda rimane ancora: anche con le prove che suggeriscono l’uso decennale del GH nell’atletica competitiva, questo ormone apporta veramente un significativo miglioramento delle prestazioni?
Ci sono state in realtà più revisioni sistematiche che hanno tentato di rispondere a questa domanda, ma sfortunatamente sono state tutt’altro che conclusive per quanto riguarda gli effetti ergogenici del GH.[173-175] E nonostante vari scandali nel corso degli anni, così come la prevalenza d’uso del GH da parte di atleti professionisti, ci sono ancora pochissime prove cliniche che suggeriscono che il GH da solo abbia un impatto significativo sull’aumento delle prestazioni sia negli adulti sani che nei soggetti giovani.[173,176- 179]
Sono stati svolti soltanto pochi studi strettamente controllati che più direttamente hanno tentato di cercare il reale impatto del GH sulle prestazioni fisiche in soggetti sani e allenati. Probabilmente il più interessante di questi ha dimostrato che dosi sovrafisiologiche di GH abbinate agli AAS non hanno apportato miglioramenti significativi del VO2max, nella forza o nella potenza esplosiva misurata attraverso l’altezza del salto.[180] E’ stato notato un leggero miglioramento nella capacità di sprint anaerobico, che era più evidente negli uomini e specialmente in quelli trattati con GH e AAS. Considerando ciò, in un evento sportivo nel quale le frazioni di secondo potrebbero significare la differenza tra vincere e perdere, questo dato ha una certa importanza.
Passando in rassegna il corpo della letteratura scientifica, non si osservano miglioramenti delle prestazioni aerobiche tramite l’uso di GH. Ciò si riscontra sia nel caso in cui il GH viene somministrato a dosi fisiologiche a soggetti sani [181-182] sia nel caso in cui viene somministrato in dosi sovrafisiologiche.[180,183-184] La capacità aerobica non è inoltre influenzata dalla somministrazione acuta di GH prima dell’allenamento.[185] Tutti gli aumenti delle prestazioni aerobiche del GH sembrano essere mediati tramite gli Androgeni, e questo è ulteriormente supportato dai risultati di uno studio che dimostrano come gli ex utilizzatori di AAS esprimano un aumento del VO2max. Sebbene fossero passati alcuni mesi dal loro ultimo utilizzo di AAS, tale lasso di tempo non era sufficiente affinchè l’effetto “di coda” dell’uso di questa categoria di farmaci fosse esaurito.[186]
Soggetti anziani sani a cui erano state somministrate dosi combinate di Testosterone e GH, calibrate affinchè si raggiungesse una soglia ematica nel range dell’età giovanile, hanno sperimentato in qualche misura il miglioramento dell’equilibrio e delle prestazioni fisiche.[187] Questi miglioramenti delle prestazioni sembrano essere più pronunciati negli uomini e, comunque, sono marginali, anche con i trattamenti combinati.[188] Come è stato discusso in precedenza, le dosi soprafisiologiche di GH possono aumentare la capacità anaerobica.[180] Questo è un effetto osservato anche in alcune occasioni in cui i soggetti GHD sono stati trattati con GH, ripristinando i “normali” intervalli dell’ormone.[182,189]
L’Asse GH/IGF-1 può anche svolgere un ruolo nella regolazione del tono vascolare, o nel grado di costrizione rispetto allo stato di massima dilatazione dei vasi sanguigni, regolando in tal modo la resistenza periferico.[190-191] L’IGF-1 è stato identificato come un potente vasodilatatore, un effetto parzialmente mediato dall’aumento dell’emissione di ossido nitrico dall’endotelio, il tessuto che forma uno strato di cellule che rivestono organi come il cuore e i vasi linfatici.[192-194] Questo potenziale di aumento della capacità del flusso sanguigno ha una miriade di benefici ipotetici sugli sforzi atletici.
Per concludere, nonostante la mancanza di prove convincenti in letteratura [434], gli atleti usano spesso dosi e protocolli che non vengono replicati durante gli studi. Detto ciò, si dovrebbe comunque essere cauti prima di liquidare completamente il GH come composto per il miglioramento delle prestazioni basandosi semplicemente su ciò che, al momento, il corpo della letteratura scientifica ci suggerisce. È molto probabile che eserciti un contributo significativo negli atleti di alto livello, e potrebbe farlo attraverso mezzi diretti o indiretti.
Nella terza parte di questa serie di articoli mi occuperò degli effetti diretti e indiretti del GH e del IGF-1 nell’ipertrofia muscolare, dell’uso degli AAS in combinazione con GH e IGF-1 analizzando, infine, il loro potenziale sinergico con l’Asse GH/IGF-1.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
Cohen P. Overview of the IGF-I system. Horm Res. 2006;65 Suppl 1:3-8. Epub 2006 Mar 2. Review.
Yakar S, Liu JL, Stannard B, Butler A, Accili D, Sauer B, LeRoith D. Normal growth and development in the absence of hepatic insulin-like growth factor I. Proc Natl Acad Sci U S A. 1999 Jun 22;96(13):7324-9.
Laron Z. Insulin-like growth factor 1 (IGF-1): a growth hormone. Molecular Pathology. 2001;54(5):311-316.
Wurzburger MI, Prelevic GM, Sönksen PH, Balint-Peric LA, Wheeler M. The effect of recombinant human growth hormone on regulation of growth hormone secretion and blood glucose in insulin-dependent diabetes. J Clin Endocrinol Metab. 1993 Jul;77(1):267-72.
Yakar S, Rosen CJ, Beamer WG, Ackert-Bicknell CL, Wu Y, Liu JL, Ooi GT, Setser J, Frystyk J, Boisclair YR, LeRoith D. Circulating levels of IGF-1 directly regulate bone growth and density. J Clin Invest. 2002 Sep;110(6):771-81.
D’Ercole AJ, Stiles AD, Underwood LE. Tissue concentrations of somatomedin C: further evidence for multiple sites of synthesis and paracrine or autocrine mechanisms of action. Proc Natl Acad Sci U S A. 1984 Feb;81(3):935-9.
Gosteli-Peter MA, Winterhalter KH, Schmid C, Froesch ER, Zapf J. Expression and regulation of insulin-like growth factor-I (IGF-I) and IGF-binding protein messenger ribonucleic acid levels in tissues of hypophysectomized rats infused with IGF-I and growth hormone. Endocrinology. 1994 Dec;135(6):2558-67.
Gunawardane K, Krarup Hansen T, Sandahl Christiansen J, et al. Normal Physiology of Growth Hormone in Adults. [Updated 2015 Nov 12]. In: De Groot LJ, Chrousos G, Dungan K, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-.
Lu C, Lam HN, Menon RK. New members of the insulin family: regulators of metabolism, growth and now … reproduction. Pediatr Res. 2005 May;57(5 Pt 2):70R-73R.
Yakar S, Sun H, Zhao H, Pennisi P, Toyoshima Y, Setser J, Stannard B, Scavo L, Leroith D. Metabolic effects of IGF-I deficiency: lessons from mouse models. Pediatr Endocrinol Rev. 2005 Sep;3(1):11-9. Review.
Samani AA, Yakar S, LeRoith D, Brodt P. The role of the IGF system in cancer growth and metastasis: overview and recent insights. Endocr Rev. 2007 Feb;28(1):20-47. Epub 2006 Aug 24. Review.
Clemmons DR. Metabolic actions of insulin-like growth factor-I in normal physiology and diabetes. Endocrinol Metab Clin North Am. 2012 Jun;41(2):425-43, vii-viii.
Kim JJ, Accili D. Signalling through IGF-I and insulin receptors: where is the specificity? Growth Horm IGF Res. 2002 Apr;12(2):84-90. Review.
Shimizu M, Webster C, Morgan DO, Blau HM, Roth RA. Insulin and insulin-like growth factor receptors and responses in cultured human muscle cells. Am J Physiol. 1986 Nov;251(5 Pt 1):E611-5.
Buckway CK, Guevara-Aguirre J, Pratt KL, Burren CP, Rosenfeld RG. The IGF-I generation test revisited: a marker of GH sensitivity. J Clin Endocrinol Metab. 2001 Nov;86(11):5176-83.
Hwa V, Oh Y, Rosenfeld RG. The insulin-like growth factor-binding protein (IGFBP) superfamily. Endocr Rev. 1999 Dec;20(6):761-87. Review.
Firth SM, Baxter RC. Cellular actions of the insulin-like growth factor binding proteins. Endocr Rev. 2002 Dec;23(6):824-54. Review.
Bach LA, Hsieh S, Sakano K, Fujiwara H, Perdue JF, Rechler MM. Binding of mutants of human insulin-like growth factor II to insulin-like growth factor binding proteins 1-6. J Biol Chem. 1993 May 5;268(13):9246-54.
Boisclair YR, Rhoads RP, Ueki I, Wang J, Ooi GT. The acid-labile subunit (ALS) of the 150 kDa IGF-binding protein complex: an important but forgotten component of the circulating IGF system. J Endocrinol. 2001 Jul;170(1):63-70. Review.
Duan C. Specifying the cellular responses to IGF signals: roles of IGF-binding proteins. J Endocrinol. 2002 Oct;175(1):41-54. Review.
LeRoith D. Insulin-like growth factor receptors and binding proteins. Baillieres Clin Endocrinol Metab. 1996 Jan;10(1):49-73. Review.
Rajaram S, Baylink DJ, Mohan S. Insulin-like growth factor-binding proteins in serum and other biological fluids: regulation and functions. Endocr Rev. 1997 Dec;18(6):801-31. Review.
Monzavi R, Cohen P. IGFs and IGFBPs: role in health and disease. Best Pract Res Clin Endocrinol Metab. 2002 Sep;16(3):433-47. Review.
Giustina A, Mazziotti G, Canalis E. Growth hormone, insulin-like growth factors, and the skeleton. Endocr Rev. 2008 Aug;29(5):535-59. Epub 2008 Apr 24. Review.
Collett-Solberg PF, Cohen P. Genetics, chemistry, and function of the IGF/IGFBP system. Endocrine. 2000 Apr;12(2):121-36. Review.
Wetterau LA, Moore MG, Lee KW, Shim ML, Cohen P. Novel aspects of the insulin-like growth factor binding proteins. Mol Genet Metab. 1999 Oct;68(2):161-81. Review.
Parker A, Rees C, Clarke J, Busby WH Jr, Clemmons DR. Binding of insulin-like growth factor (IGF)-binding protein-5 to smooth-muscle cell extracellular matrix is a major determinant of the cellular response to IGF-I. Mol Biol Cell. 1998 Sep;9(9):2383-92.
Velloso CP. Regulation of muscle mass by growth hormone and IGF-I. Br J Pharmacol. 2008 Jun;154(3):557-68. Review.
Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev. 1995 Feb;16(1):3-34. Review.
Hameed M, Lange KH, Andersen JL, Schjerling P, Kjaer M, Harridge SD, Goldspink G. The effect of recombinant human growth hormone and resistance training on IGF-I mRNA expression in the muscles of elderly men. J Physiol. 2004 Feb 15;555(Pt 1):231-40. Epub 2003 Oct 17.
Pfeffer LA, Brisson BK, Lei H, Barton ER. The insulin-like growth factor (IGF)-I E-peptides modulate cell entry of the mature IGF-I protein. Mol Biol Cell. 2009 Sep;20(17):3810-7.
Zabłocka B, Goldspink PH, Goldspink G, Górecki DC. Mechano-Growth Factor: an important cog or a loose screw in the repair machinery? Front Endocrinol (Lausanne). 2012 Nov 1;3:131.
Rotwein P. Two insulin-like growth factor I messenger RNAs are expressed in human liver. Proc Natl Acad Sci U S A. 1986 Jan;83(1):77-81.
Siegfried JM, Kasprzyk PG, Treston AM, Mulshine JL, Quinn KA, Cuttitta F. A mitogenic peptide amide encoded within the E peptide domain of the insulin-like growth factor IB prohormone. Proc Natl Acad Sci U S A. 1992 Sep 1;89(17):8107-11.
Barton ER, DeMeo J, Lei H. The insulin-like growth factor (IGF)-I E-peptides are required for isoform-specific gene expression and muscle hypertrophy after local IGF-I production. J Appl Physiol (1985). 2010 May;108(5):1069-76.
Yang S, Alnaqeeb M, Simpson H, Goldspink G. Cloning and characterization of an IGF-1 isoform expressed in skeletal muscle subjected to stretch. J Muscle Res Cell Motil. 1996 Aug;17(4):487-95.
McKoy G, Ashley W, Mander J, Yang SY, Williams N, Russell B, Goldspink G. Expression of insulin growth factor-1 splice variants and structural genes in rabbit skeletal muscle induced by stretch and stimulation. J Physiol. 1999 Apr 15;516 (Pt 2):583-92.
Yang SY, Goldspink G. Different roles of the IGF-I Ec peptide (MGF) and mature IGF-I in myoblast proliferation and differentiation. FEBS Lett. 2002 Jul 3;522(1-3):156-60. Erratum in: FEBS Lett. 2006 May 1;580(10):2530.
Rudman D, Kutner MH, Rogers CM, Lubin MF, Fleming GA, Bain RP. Impaired growth hormone secretion in the adult population: relation to age and adiposity. J Clin Invest. 1981 May;67(5):1361-9.
Corpas E, Harman SM, Blackman MR. Human growth hormone and human aging. Endocr Rev. 1993 Feb;14(1):20-39. Review.
Veldhuis JD, Liem AY, South S, Weltman A, Weltman J, Clemmons DA, Abbott R, Mulligan T, Johnson ML, Pincus S, et al. Differential impact of age, sex steroid hormones, and obesity on basal versus pulsatile growth hormone secretion in men as assessed in an ultrasensitive chemiluminescence assay. J Clin Endocrinol Metab. 1995 Nov;80(11):3209-22.
Veldhuis JD, Iranmanesh A, Weltman A. Elements in the pathophysiology of diminished growth hormone (GH) secretion in aging humans. Endocrine. 1997 Aug;7(1):41-8. Review.
Iranmanesh A, Lizarralde G, Veldhuis JD. Age and relative adiposity are specific negative determinants of the frequency and amplitude of growth hormone (GH) secretory bursts and the half-life of endogenous GH in healthy men. J Clin Endocrinol Metab. 1991 Nov;73(5):1081-8
Rudman D. Growth hormone, body composition, and aging. J Am Geriatr Soc. 1985 Nov;33(11):800-7. Review.
Russell-Aulet M, Jaffe CA, Demott-Friberg R, Barkan AL. In vivo semiquantification of hypothalamic growth hormone-releasing hormone (GHRH) output in humans: evidence for relative GHRH deficiency in aging. J Clin Endocrinol Metab. 1999 Oct;84(10):3490-7.
Martin FC, Yeo AL, Sonksen PH. Growth hormone secretion in the elderly: aging and the somatopause. Baillieres Clin Endocrinol Metab. 1997 Jul;11(2):223-50. Review.
Chertman LS, Merriam GR, Kargi AY. Growth Hormone in Aging. [Updated 2015 May 4]. In: De Groot LJ, Chrousos G, Dungan K, et al., editors. Endotext [Internet]. South Dartmouth (MA): MDText.com, Inc.; 2000-.
Sattler FR. Growth hormone in the aging male. Best Pract Res Clin Endocrinol Metab. 2013 Aug;27(4):541-55.
Duchaine D. Underground steroid handbook. 1. California: HLR Technical Books; 1983. p. 84
Erotokritou-Mulligan I, Holt RI, Sönksen PH. Growth hormone doping: a review. Open Access Journal of Sports Medicine. 2011;2:99-111.
Liu H, Bravata DM, Olkin I, Friedlander A, Liu V, Roberts B, Bendavid E, Saynina O, Salpeter SR, Garber AM, Hoffman AR. Systematic review: the effects of growth hormone on athletic performance. Ann Intern Med. 2008 May 20;148(10):747-58. Epub 2008 Mar 17. Review.
Birzniece V, Nelson AE, Ho KK. Growth hormone and physical performance. Trends Endocrinol Metab. 2011 May;22(5):171-8. Mar 17. Review.
Baumann GP. Growth hormone doping in sports: a critical review of use and detection strategies. Endocr Rev. 2012 Apr;33(2):155-86. Epub 2012 Feb 24. Review.
Gibney J, Healy ML, Sönksen PH. The growth hormone/insulin-like growth factor-I axis in exercise and sport. Endocr Rev. 2007 Oct;28(6):603-24. Epub 2007 Sep 4. Review.
Holt RI, Sönksen PH. Growth hormone, IGF-I and insulin and their abuse in sport. Br J Pharmacol. 2008 Jun;154(3):542-56. Epub 2008 Mar 31. Review.
Barroso O, Mazzoni I, Rabin O. Hormone abuse in sports: the antidoping perspective. Asian J Androl. 2008 May;10(3):391-402.
Frystyk J. Exercise and the growth hormone-insulin-like growth factor axis. Med Sci Sports Exerc. 2010 Jan;42(1):58-66.
Meinhardt U, Nelson AE, Hansen JL, Birzniece V, Clifford D, Leung KC, Graham K, Ho KK. The effects of growth hormone on body composition and physical performance in recreational athletes: a randomized trial. Ann Intern Med. 2010 May 4;152(9):568-77.
Wallace JD, Cuneo RC, Baxter R, Orskov H, Keay N, Pentecost C, Dall R, Rosén T, Jørgensen JO, Cittadini A, Longobardi S, Sacca L, Christiansen JS, Bengtsson BA, Sönksen PH. Responses of the growth hormone (GH) and insulin-like growth factor axis to exercise, GH administration, and GH withdrawal in trained adult males: a potential test for GH abuse in sport. J Clin Endocrinol Metab. 1999 Oct;84(10):3591-601.
Chikani V, Ho KK. Action of GH on skeletal muscle function: molecular and metabolic mechanisms. J Mol Endocrinol. 2013 Dec 19;52(1):R107-23.
Lange KH, Larsson B, Flyvbjerg A, Dall R, Bennekou M, Rasmussen MH, Ørskov H, Kjaer M. Acute growth hormone administration causes exaggerated increases in plasma lactate and glycerol during moderate to high intensity bicycling in trained young men. J Clin Endocrinol Metab. 2002 Nov;87(11):4966-75.
Berggren A, Ehrnborg C, Rosén T, Ellegård L, Bengtsson BA, Caidahl K. Short-term administration of supraphysiological recombinant human growth hormone (GH) does not increase maximum endurance exercise capacity in healthy, active young men and women with normal GH-insulin-like growth factor I axes. J Clin Endocrinol Metab. 2005 Jun;90(6):3268-73. Epub 2005 Mar 22.
Irving BA, Patrie JT, Anderson SM, Watson-Winfield DD, Frick KI, Evans WS, Veldhuis JD, Weltman A. The effects of time following acute growth hormone administration on metabolic and power output measures during acute exercise. J Clin Endocrinol Metab. 2004 Sep;89(9):4298-305. Epub 2004 Aug 24.
Graham MR, Baker JS, Evans P, Kicman A, Cowan D, Hullin D, Davies B. Evidence for a decrease in cardiovascular risk factors following recombinant growth hormone administration in abstinent anabolic-androgenic steroid users. Growth Horm IGF Res. 2007 Jun;17(3):201-9. Epub 2007 Feb 26.
Brill KT, Weltman AL, Gentili A, Patrie JT, Fryburg DA, Hanks JB, Urban RJ, Veldhuis JD. Single and combined effects of growth hormone and testosterone administration on measures of body composition, physical performance, mood, sexual function, bone turnover, and muscle gene expression in healthy older men. J Clin Endocrinol Metab. 2002 Dec;87(12):5649-57.
Blackman MR, Sorkin JD, Münzer T, Bellantoni MF, Busby-Whitehead J, Stevens TE, Jayme J, O’Connor KG, Christmas C, Tobin JD, Stewart KJ, Cottrell E, St Clair C, Pabst KM, Harman SM. Growth hormone and sex steroid administration in healthy aged women and men: a randomized controlled trial. JAMA. 2002 Nov 13;288(18):2282-92.
Chikani V, Cuneo RC, Hickman I, Ho KK. Growth hormone (GH) enhances anaerobic capacity: impact on physical function and quality of life in adults with GH deficiency. Clin Endocrinol (Oxf). 2016 Oct;85(4):660-8.
Saccà L, Cittadini A, Fazio S. Growth hormone and the heart. Endocr Rev. 1994 Oct;15(5):555-73. Review.
Svensson J, Tivesten A, Isgaard J. Growth hormone and the cardiovascular function. Minerva Endocrinol. 2005 Mar;30(1):1-13. Review.
Copeland KC, Nair KS. Recombinant human insulin-like growth factor-I increases forearm blood flow. J Clin Endocrinol Metab. 1994 Jul;79(1):230-2.
Pete G, Hu Y, Walsh M, Sowers J, Dunbar JC. Insulin-like growth factor-I decreases mean blood pressure and selectively increases regional blood flow in normal rats. Proc Soc Exp Biol Med. 1996 Nov;213(2):187-92.
Walsh MF, Barazi M, Pete G, Muniyappa R, Dunbar JC, Sowers JR. Insulin-like growth factor I diminishes in vivo and in vitro vascular contractility: role of vascular nitric oxide. Endocrinology. 1996 May;137(5):1798-803.
Hermansen K, Bengtsen M, Kjær M, Vestergaard P, Jørgensen JOL. Impact of GH administration on athletic performance in healthy young adults: A systematic review and meta-analysis of placebo-controlled trials. Growth Horm IGF Res. 2017 Jun;34:38-44.
Mi sono sempre discostato dalle mode e dalle “etichette commerciali” che sorgono e tramontano nel mondo del Fitness, in generale, e nel BodyBuilding, in particolare. Dalla “carbofobia” alla presunta superiorità universale delle tecniche HIT. L’inserimento di sedute di allenamento cardiovascolare nel BodyBuilding è diventato, da diverso tempo, l’ennesimo argomento da utilizzare a “fini commerciali” in senso denigratorio. In passato ho già trattato l’argomento esponendo con logica e basi concrete come tale pratica, se ben calibrata, può portare a dei vantaggi anche in una disciplina come il Culturismo. Una recente review, svolta da scienziati dello sport australiani, ha evidenziato come l’inserimento di sedute di Cardio HIT in concomitanza con un allenamento contro-resistenza non comporta perdita di massa muscolare (dipendente questo fattore dal carico allenante e dall’introito macro-calorico). I ricercatori riportano al massimo una leggera diminuzione della forza nelle gambe, problema che gli stessi considerano ovviabile.(1)
I ricercatori, affiliati all’Università di Sydney, hanno tracciato 13 studi presenti nella letteratura scientifica, in cui i loro colleghi hanno confrontato l’effetto della combinazione dell’allenamento contro-resistenza con l’HIIT(High Intensity Interval Training). I dettagli relativi alle pianificazioni utilizzate sono disponibili nella tabella riportata qui di seguito.
I ricercatori hanno aggregato i risultati dei precedenti studi analizzandoli nuovamente.
In seguito all’analisi dei risultati derivanti dalla ricerca disponibile, si è evidenziato che l’aggiunta del HIIT all’allenamento contro resistenza non ha effetti negativi sull’aumento della massa muscolare, né sull’aumento della forza della parte superiore del corpo. Tuttavia, si verifica un effetto negativo sull’aumento della forza nella parte inferiore del corpo.
Tale effetto negativo è comunque contenuto. Secondo la meta-analisi, la forza massima espressa dalla parte inferiore del corpo nei soggetti che avevano svolto solo l’allenamento contro-resistenza era aumentata del 23,9% contro un aumento del 19,4% riscontrato nei soggetti che avevano svolto sedute HIIT insieme all’allenamento contro-resistenza.
Tuttavia, quando i ricercatori hanno esaminato attentamente i dati in loro possesso, hanno “scoperto” il modo attraverso il quale gli atleti possono ridurre il piccolo effetto negativo dato dall’HIIT sull’aumento della forza.
In sintesi i ricercatori hanno riportato che, i risultati di questa review mostrano come l’HIIT può essere affiancato ad un allenamento contro resistenza senza che questo influenzi negativamente sui cambiamenti nella massa muscolare e che qualsiasi attenuazione della forza muscolare della parte inferiore può essere migliorata inserendo l’HIIT a debita distanza dalla sessione di allenamento contro-resistenza. Quest’ultima è una pratica che da i medesimi riscontri positivi anche con l’applicazione del Cardio LISS.
È importante tenere in considerazione che l’allenamento contro-resistenza abbinato all’HIIT e il solo allenamento contro resistenza hanno dato come risultato un miglioramento della forza dinamica in tutti gli studi vagliati e che la differenza nell’espressione della forza della parte inferiore del corpo tra i due gruppi era minima.
Inoltre, poiché è stato dimostrato che l’allenamento HIIT migliora il VO2max, il tempo di sprint e la massima velocità aerobica, la leggera riduzione [dell’aumento] della forza corporea espressa dalla parte inferiore del corpo può essere un piccolo prezzo da pagare per ottenere miglioramenti su altri aspetti chiave della prestazione sportiva in particolare se vincolata da limiti di tempo.
Quello che verrà riportato in questo e negli altri articoli della serie che seguiranno, è il frutto di un lavoro di ricerca ed estrapolazione di informazioni che trovano la loro origine nel ottimo lavoro di Chest Rockwell sull’uso del Ormone della Crescita a fini ipertrofici intitolato “The Most Effective Growth Hormone Protocol for Hypertrophy”.
Nota introduttiva
Se volete un metodo altamente efficace per spingere un Bodybuilder a guardarvi prima con confusione e, in seguito, con totale irritazione, basta che usiate qualsiasi variazione della seguente affermazione:
L’Ormone della Crescita determina un aumento significativo della massa corporea magra, è altamente anabolico, ma non causa la crescita del tessuto muscolare…
Nel corso di questi articoli, spiegherò come questa affermazione sia precisa al 100% nonostante sembri un po’ assurdo ad una prima lettura. Per impostare alcuni parametri in anticipo, si parlerà di AAS e della loro relazione sinergica con il GH, ma ci saranno solo alcune menzioni sull’Insulina. Questa serie di articoli rappresenta già di suo un’impresa enorme. Penso che per impostare in modo corretto l’argomento, il tema GH/Insulina meriti un articolo a parte (cosa parzialmente già fatta) a causa delle immense complessità che esistono nella loro correlazione. Non ho nemmeno intenzione di toccare l’argomento dei secretagoghi del GH, dei peptidi sperimentali o di altri analoghi in modo che ci si possa concentrare principalmente sui fondamentali della questione GH ed Ipertrofia. Se non diversamente specificato nell’articolo, si parlerà esplicitamente dell’Ormone della Crescita di origine endogena o ricombinante (esogeno).
E, infine, questi articoli sono rivolti esclusivamente agli uomini. Mi dispiace per le atlete, ma i maschi sono molto più sensibili agli effetti anabolici della supplementazione con GH rispetto alle donne, data la spiccata natura sessualmente dimorfica del GH.(1) Quindi, a meno che non dichiari specificatamente il contrario, le informazioni che seguiranno saranno orientate verso l’uomo. In futuro è probabile che realizzerò un articolo in merito dedicato alle donne.
Introduzione all’Anabolismo e alla Crescita Muscolare
La prima cosa da fare è conoscere concisamente le corrette definizioni di termini quali anabolismo, ipertrofia e iperplasia. Li vedo spesso usati quasi in modo intercambiabile, ma ci sono alcune differenze chiave. Ad esempio, solo perché qualcosa è anabolico non significa automaticamente che causerà la crescita del tessuto muscolare. Viceversa solo perché qualcosa è catabolico non significa automaticamente che non possa contribuire alla crescita del tessuto muscolare in modo positivo.
Schema dei processi anabolici
L’anabolismo può essere definito come qualsiasi stato in cui la ritenzione d’azoto nella massa magra è positiva, sia attraverso la stimolazione della sintesi proteica che attraverso la soppressione dei tassi di proteolisi.(2) Il primo chiarimento da fare in riferimento alla mia precedente affermazione è che la misurazione della massa corporea magra non include soltanto la massa muscolare ma, per esempio, anche gli organi, le ossa e l’acqua ivi contenuta (3), fattore quest’ultimo soggetto all’azione dell’Ormone della Crescita. Quindi, leggere uno studio nel quale si afferma che la massa corporea magra è aumentata in seguito al trattamento con GH, non vuol dire che ci si riferisca ad un aumento della massa muscolare.
L’apparato muscolare è un tessuto plastico molto complesso e in grado di adattarsi alle mutevoli esigenze funzionali alle quali viene sottoposto. Quando parliamo di aumento della massa muscolare, stiamo parlando principalmente di un evento che si verifica attraverso due meccanismi primari – ipertrofia e iperplasia.
L’ipertrofia è il processo attraverso il quale l’aumento della massa muscolare si verifica tramite l’aumento delle dimensioni dell’area della sezione trasversa di una fibra muscolare esistente (CSA). Il processo ipertrofico è mediato da molti fattori come con l’ipertrofia indotta dall’esercizio, di particolare interesse per i Bodybuilder, essendo essa mediata da una combinazione di tensione meccanica, danno muscolare e stress metabolico.(4)
L’iperplasia, ivece, è il processo mediante il quale l’aumento della massa muscolare viene raggiunto attraverso un incremento del numero effettivo di fibre muscolari. È generalmente accettato che, negli esseri umani, il numero di fibre all’interno del muscolo sia geneticamente predeterminato e fissato durante il periodo perinatale.(5) Esistono alcuni studi svolti su animali i quali hanno dimostrato che l’iperplasia può verificarsi (6-7), spesso in condizioni di test unici, ma il tentativo di dedurre da ciò che essa si possa verificare negli esseri umani (8-9) è altamente speculativo nella migliore delle ipotesi. Anche se l’iperplasia si verifica nell’apparato muscolare umano, è molto probabile che sia solo un fattore minore che determina la massa complessiva. Tuttavia, a causa di una cattiva comprensione (se così la vogliamo chiamare) degli studi prima citati spesso vengono formulate affermazioni definitive secondo cui il GH causa iperplasia. Vale quindi la pena di ribadire che questi tipi di affermazioni dovrebbero essere considerate nient’altro che speculative.
La scoperta dell'”Ormone della Crescita”
Ormone della Crescita
Sono trascorsi oltre 100 anni da quando Harvey Cushing suppose per la prima volta l’esistenza di un “Ormone della Crescita” (10), e questo ormone venne successivamente isolato, identificato ed estratto dall’ipofisi umana negli anni ’40.(11) Nel decennio successivo, si fece largo un’ipotesi cardine secondo cui la “crescita” non era direttamente attribuibile al GH, ma piuttosto ad un gruppo di fattori serici regolati dall’espressione del GH.(12) Questi fattori serici sono stati in seguito chiamati fattori di solfatazione, termine volto ad indicare sostanze controllate dal GH che stimolavano l’assorbimento di solfato nella cartilagine e nei tessuti. Questa ipotesi ha cercato di aiutare i ricercatori a conciliare meglio il modo in cui la crescita somatica veniva regolata da una sostanza secreta dall’Ipofisi, mentre allo stesso tempo rinforzava il fatto che questa sostanza ipofisaria non agiva direttamente sui suoi tessuti bersaglio per promuovere la crescita.(13-14)
Nei decenni successivi, molti esperimenti hanno dimostrato l’ampia varietà di funzioni possedute da questa famiglia di fattori di solfatazione e il termine “somatomedina” è stato proposto per includere tutti i membri di questa famiglia.(15) È interessante notare che l’ipotesi originale che descrive la regolazione di questi fattori di solfatazione mediati dal GH è ancora oggi generalmente indicata come “l’ipotesi della somatomedina”.(16) L’ipotesi originale affermava che la crescita somatica è causata dall’azione del GH a livello epatico, dove stimola la sintesi di IGF-1 che viene successivamente rilasciato agendo sui tessuti bersaglio con modello endocrino. Questa ipotesi è rimasta il modello accettato per decenni fino a quando non sono stati identificati i ruoli autocrini del IGF (17-18) insieme agli effetti diretti che il GH ha nella crescita ossea.(19-20)Per riconciliare queste nuove scoperte, l’ipotesi originaria è stata leggermente modificata prendendo il nome di “Teoria del Doppio Effetto”, che ha definito sia un ruolo autocrino che endocrino del IGF-1.(21)
Tornando indietro di qualche tempo, verso la fine degli anni ’70, l’identità chimica di questi fattori di solfatazione era più intimamente riconosciuta come la manifestazione di due peptidi con un’elevatissima somiglianza con l’Insulina e quindi ribattezzati “fattori di crescita isulino-simili (IGF)”.(22-24) Fu all’incirca nello stesso periodo nel quale vennero identificate anche le proteine leganti l’IGF (IGFBP), e di conseguenza la conoscenza della biologia dell’IGF-1 decollò ad un tasso esponenziale.(25)
Nel frattempo, gli estratti ipofisari umani diventarono disponibili tramite l’estrazione dai cadaveri e gli esperimenti iniziali svolti su animali ed esseri umani mostrarono quanto complesse fossero realmente le azioni del GH umano. (26-33) La pratica dell’uso di questi estratti ipofisari venne interrotta dai medici in seguito alla comparsa della malattia di Creutzfeldt-Jakob (CJD) in pazienti a cui era stato precedentemente somministrato il GH contenuto nel Ipofisi estratta dai cadaveri.(34) La CJD è una malattia cerebrale particolarmente sgradevole e universalmente fatale, con la stragrande maggioranza dei soggetti affetti che muoiono poco dopo la sua diagnosi.
Fortunatamente, nel 1985, nello stesso periodo in cui questi casi di CJD cominciarono a fare notizia, la FDA approvò il primo GH ricombinante sintetico (rHGH) per l’utilizzo su soggetti con deficit dell’Ormone della Crescita. Questa versione di rHGH è stata prodotta dalla Genentech e denominata Protropin.(35-36) Vale la pena notare che, analizzando la composizione dei vecchi farmaci contenenti GH si può osservare che erano costituiti da una catena di 192 amminoacidi (met-hGH) rispetto al rHGH che consiste in una catena di 191 amminoacidi.(37) In ogni caso, con l’rHGH, ora di facile disponibilità, è stata inaugurata un’era completamente nuova, con condizioni cliniche molto più sicure, per il numero in continua crescita di pazienti trattati.
Effetti dell’Ormone della Crescita sulla sintesi proteica
All’inizio dell’articolo, ho definito a grandi linee cos’è l’anabolismo ed ho anche affermato che l’Ormone della Crescita possiede una natura anabolica. Adesso, descriverò ciò che rende anabolizzante l’Ormone della Crescita e approfondirò parte della letteratura scientifica esistente sull’argomento.
Nel corso degli anni, il GH è stato ampiamente studiato in diversi modi. La maggior parte degli studi svolti, quando considerati nel loro insieme, suggeriscono che il GH sia anabolico. Più specificamente, il GH è anabolico perché stimola la sintesi proteica di tutto il corpo con o nessun effetto, o un effetto soppressivo, sui tassi di degradazione proteica. [38] Tuttavia, quando si approfondisce l’argomento, le cose diventano un po’ meno chiare dal momento che i risultati degli studi tendono ad essere diversi. I diversi risultati sono un riflesso diretto dell’ immensa complessità del GH.
Il GH esplica i suoi effetti sulla sintesi proteica legandosi prima con il suo recettore specifico (GHR) e successivamente aumentando la trascrizione del gene muscolare attraverso i percorsi di segnalazione a valle, in definitiva attivando la segnalazione del mTOR. [39-40] Questi effetti si manifestano in acuto, spesso si verificano in pochi minuti e sono di natura simile all’Insulina, usando molte delle stesse vie anaboliche. [41-44] La rapida comparsa di questi cambiamenti metabolici legati alle proteine suggerisce che essi siano direttamente causati dal GH e non secondariamente mediati tramite l’IGF-1 [45]. L’impatto del GH sulla proteolisi, d’altro canto, è molto probabilmente di natura indiretta. A detta di tutti, ciò ha più a che fare con i suoi effetti inibitori sull’Insulina, che è stata vista avere effetti diretti sulla proteolisi.[46]
Ora, poiché sarete interessati principalmente alla crescita muscolare, riporterò come il GH influenza specificamente i tassi di sintesi delle proteine muscolari (MPS). Vi sono numerosi studi in letteratura nei quali il GH è stato somministrato a soggetti adulti sani senza aver mostrato alcun impatto sui tassi di MPS. [45,47-52] Ciò che risulta affascinante di questi risultati è che un paio di studi includevano anche una allenamento di resistenza, sebbene, nella fattispecie, non sono stati comunque riscontrati aumenti nei tassi di MPS locali. Al contrario, ci sono una serie di studi che hanno riportato di aumenti nei tassi di MPS senza cambiamenti significativi nei tassi di sintesi proteica dell’intero corpo. [53-55]
Ci sono molte ragioni per cui questi risultati appaiono non del tutto coerenti all’interno del corpo della letteratura scientifica. Uno dei motivi principali è rappresentato dal modo in cui gli studi sono stati svolti per quanto riguarda il tipo di somministrazione del GH (ad esempio, le concentrazione di dosaggio, se somministrato localmente o in modo sistemico, nonché se l’ormone è stato somministrato in modo pulsatile o costante). Alcune delle altre ragioni includono come è stata misurata la sintesi proteica, se i soggetti esaminati erano a digiuno o nutriti, che tipo di muscolo scheletrico è stato esaminato e, anche, per quanto tempo è durato lo studio. Come menzionato in precedenza, molti degli effetti che il GH ha sul metabolismo proteico avvengono in acuto.
Sebbene questa serie di articoli è principalmente incentrata sull’ipertrofia, ritengo comunque che valga la pena di discutere le differenze che il GH ha sul metabolismo delle proteine in condizioni di digiuno e nutrizione. Ciò aiuterà ad avere un quadro generale più chiaro di come il comportamento di questo ormone sia spesso il risultato diretto dell’ambiente nel quale è introdotto. Come ho spiegato dettagliatamente nell’articolo dedicato a GH e lipolisi, la secrezione di GH subisce un aumento durante periodi prolungati di digiuno. Questo è un meccanismo di sopravvivenza, con l’obiettivo principale di conservare preziosi pool amminoacidici immagazzinati prevenendo la degradazione delle proteine. [56] Questo stesso comportamento di risparmio di proteine può essere visto, in misura minore, in soggetti trattati con GH e sottoposti a severe restrizioni dietetiche [57], soggetti obesi sottoposti a vari tipi di dieta ipocalorica [58-61] e soggetti privati di proteine dietetiche [ 62].
IGF-1
È stato dimostrato che l’IGF-1 inibisce allo stesso modo la disgregazione proteica a livello sistemico [63], il che avrebbe senso a causa della stretta co-relazione con il GH. Quando gli amminoacidi e l’Insulina vengono somministrati ai soggetti esaminati, già sottoposti a somministrazione di IGF-1, è stato dimostrato, sia negli uomini che negli animali, che i tassi di sintesi proteica aumentano a livello sistemico [64-65]. Vale la pena notare che l’IGF-1 è bifasico nel senso che quando è somministrato ad alto dosaggio e, di conseguenza, i livelli serici diventano elevati, il suo comportamento cambia passando da un azione “GH-simile” ad una “insulino-simile”. Tratterò nel dettaglio l’argomento in seguito.
Per riassumere, il GH è molto adatto per prevenire la degradazione proteica, e lo fa in una vasta gamma di condizioni alimentari a ristretto apporto calorico. Tuttavia, in presenza di un apporto energetico sufficiente, il suo comportamento cambia. L’effetto principale del GH sul metabolismo proteico è volto dapprima a creare un ambiente con una ossidazione amminoacidica ridotta [47,66] e successivamente ad aumentare la sintesi proteica sistemica. [67]
Il ruolo di GH e IGF-1 nella crescita postnatale
È noto che il GH regola la crescita postnatale e che questi effetti di promozione della crescita sono principalmente mediati dall’IGF-1. [68-69] Tuttavia, giusto per ribadirlo, è necessario chiarire che questi effetti di induzione della crescita non sono esclusivi dell’ipertrofia muscolare. La crescita lineare di un organismo include cambiamenti nell’apparato scheletrico, negli organi e nei muscoli.
Per riportare esempi di un certo rilievo sull’importanza che GH e IGF-1 hanno sulla crescita postnatale, basta osservare gli individui con mutazioni o disturbi correlati all’Asse GH/IGF. Una revisione dettagliata su questo tema va oltre lo scopo di questo articolo ma, in generale, e penso sia abbastanza risaputo, i disturbi che sopprimono/inibiscono l’Asse GH/IGF in soggetti in giovane età si caratterizzano in una bassa statura di questi mentre i disturbi che stimolano l’Asse GH/IGF si caratterizzano nel gigantismo. [70-71]
La stragrande maggioranza degli effetti del GH sulla promozione della crescita sono mediati tramite l’IGF-1, tuttavia ci sono una serie di processi che sono GH-mediati o IGF-indipendenti. L’esempio più eloquente di questo è rappresentato nei modelli animali in cui i topi “knockout” mutati e deficitari di GH e IGF-1 hanno mostrato una maggiore gravità nel ritardo della crescita rispetto ai “knockout” GH-deficienti o IGF-deficienti [72]. Ma osserviamo nel dettaglio alcuni degli effetti più specifici che sono stati identificati nei modelli umani.
Il primo è legato alla steatosi epatica, nota anche come “malattia del fegato grasso” [73]. È stato dimostrato sia negli esseri umani con la sindrome di Laron che negli animali con soppressione del GHR che questa condizione può ancora verificarsi in presenza di livelli di IGF-1 soppressi[74]. La sindrome di Laron fornisce alcune intuizioni piuttosto uniche sul sistema GH/IGF-1 a causa del fatto che è causata da una mutazione nel GHR che si traduce in livelli significativamente bassi di IGF-1 endocrino, dal momento che al GH viene impedito di legarsi al suo sito recettore e di stimolare efficacemente la produzione di IGF-1.
Un’altra azione specifica del GH è legata alla sua capacità di migliorare lo sviluppo del follicolo preantrale ovarico. In effetti, il GH è stato anche recentemente oggetto di studi per i suoi potenziali miglioramenti sulla fertilità femminile. I risultati indicano che alcuni gruppi che utilizzano la fecondazione in vitro (FIV) sembrano beneficiare della somministrazione di GH. [75-76] Anche i modelli animali con deficit di GHR hanno mostrato un numero inferiore di follicoli primari preantrali e antrali rispetto ai loro simili di controllo.
Una delle prime e più interessanti scoperte riguardanti l’azione del GH , è stata il ruolo specifico del GH nel promuovere la crescita ossea longitudinale attraverso i suoi effetti sulla generazione dei condrociti (cellule cartilaginee) nella regione delle placche di crescita epifisaria. [19-20,77-80]. Il GH ha in realtà due ruoli nella promozione della crescita ossea longitudinale; sia attraverso i suddetti effetti sulla generazione dei condrociti nella piastra di crescita che attraverso un ruolo mediato dall’IGF-1 nella promozione dell’ipertrofia dei condrociti.[81] Vale la pena notare che i livelli endocrini di IGF-1 completamente intatti non sono nemmeno una necessità quando si tratta di crescita ossea postnatale. Infatti, fino a quando sono presenti almeno il 10-20% dei livelli di IGF-1 endocrino circolante, la combinazione di IGF-1 e GH autocrini può comunque garantire la normale crescita ossea postnatale. [82] Ciò è probabilmente dovuto ai ruoli sovrapposti di IGF-1 autocrino ed endocrino in relazione alla crescita ossea longitudinale. [83]
Il GH promuove un aumento dei tassi di fusione delle cellule muscolari tardive che possono avere la capacità di aumentare la dimensione delle fibre muscolari in un modo completamente indipendente dalla sovraregolazione dell’IGF-1.[84] Utilizzando una nuova tecnica di sperimentazione su cellule animali, i ricercatori sono stati in grado di dimostrare che il GH promuove la fusione dei mioblasti con i miotubi nascenti senza un aumento dell’espressione effettiva dell’mRNA del IGF-1. I miotubi nascenti sono presenti negli stadi successivi della fusione delle cellule muscolari [85] e il GH ha dimostrato di aumentare il numero di nuclei per miotubo.
La relazione tra secrezione di GH e IGF-1
L’Ormone della Crescita è noto per aumentare i livelli di IGF-1 circolante così come la sintesi locale di IGF-1, in modo autocrino. Entrambe queste azioni giocano un ruolo fondamentale nella regolazione della massa muscolare e, quindi, in conseguenza di ciò, risulta utile comprendere meglio come la secrezione di GH porta ad un aumento dei livelli endocrino e autocrino di IGF-1.
La stragrande maggioranza dell’Ormone della Crescita negli adulti sani è secreto dalla ghiandola pituitaria e, più specificamente, dalle cellule somatotrope nel lobo anteriore mediate dal fattore di trascrizione Prophet of Pit-1 (PROP1).[86-87] Il GH può anche essere sintetizzato localmente in molti tessuti come il cervello, le cellule immunitarie, il tessuto mammario, i denti e la placenta che sono tutti al di fuori della regolazione dell’ipofisi. [88] Questo supporta l’idea secondo cui il GH abbia ruoli autocrini oltre ai suoi già consolidati ruoli endocrini.
Il GH è secreto in modo pulsatile in tutte le specie [89-90] e questo modello secretorio gioca un ruolo fisiologico importante in tutte le sue caratteristiche, da quelle dimorfiche sessuali all’RNA del IGF-1, di cui scopriremo di più in seguito. I giovani maschi adulti sani secernono tra 0,4-0,5mg di GH ogni 24 ore e molte di queste secrezioni si verificano come “impulsi all’interno di impulsi”. [91] Normalmente, ci sono circa 10-12 impulsi secretorie ogni giorno con gli uomini che presentano un modello di impulso significativamente più regolare rispetto alle donne. Nei maschi, il GH viene secreto in modo episodico, con la ben nota grande ondata serale che si verifica in prossimità dell’inizio del sonno ad onde lente. I maschi hanno anche secrezioni più contenute che si verificano poche ore dopo aver consumato i pasti.[92-94] Le femmine hanno livelli ematici inter-secretori più elevati, in particolare nella fase follicolare del ciclo mestruale, con impulsi di GH più frequenti durante il giorno e un impulso notturno significativamente più basso rispetto ai maschi [95-96]. Non è del tutto chiaro perché esista questo schema secretivo sessualmente dimorfico.
GHRH
La secrezione di GH è regolata in un modo molto complesso che coinvolge la partecipazione di diversi neurotrasmettitori, così come di feedback sia ormonali che metabolici. È regolato principalmente in modo positivo dal GHRH (Ormone di Rilascio della Somatotropina o Somatorelina) [97], e in modo negativo dalla SRIF (Somatostatina). Entrambi questi peptidi sono prodotti all’interno dell’ipotalamo. Infatti, oltre al suo ruolo di base nella produzione ormonale, l’ipotalamo monitora costantemente il rapporto GHRH/SRIF e di conseguenza opera un controllo sulla secrezione ipofisaria. [98]
Oltre alla sua funzione primaria di stimolazione della secrezione di GH, il GHRH svolge anche un ruolo essenziale nella proliferazione e nello sviluppo delle suddette cellule somatotrope. Infatti, in un sistema con GHRH alterato o assente, è stata osservata ipoplasia anteriore dell’ipofisi che è probabilmente il risultato di un alterato sviluppo somatotropo.[99-100] Gli esseri umani con il GHRH soppresso per azione di un antagonista hanno mostrato un rilascio pulsatile di GH gravemente compromesso e una risposta soppressa del GH al GHRH [101], quindi è chiaramente una componente vitale all’interno dell’Asse GH/IGF-1.
SRIF
La SRIF, il principale regolatore negativo della secrezione di GH, sopprime il TSH. Inoltre, in misura minore, sopprime sia la Prolattina che l’Ormone Adrenocorticotropo [97]. Tutti questi ormoni sono noti per avere strette relazioni con l’Asse GH/IGF-1. La SRIF ha un’emivita breve di circa 2-3 minuti nel siero e viene quindi rapidamente inattivata dalle peptidasi tissutali. Durante la sua vita attiva, sopprime non solo il rilascio spontaneo di GH, ma anche la risposta del GH a tutti gli stimoli esterni incluso il GHRH, l’ipoglicemia, l’Arginina e l’esercizio fisico solo per nominarne alcuni. I suoi effetti soppressivi apparentemente sono limitati all’ampiezza della liberazione basale e pulsatile di GH, poiché non è stato dimostrato che alteri la frequenza degli impulsi di GH.[102]
Il GH circolante è in gran parte legato alle proteine di trasporto chiamate GHBP o proteine leganti l’ormone della crescita. Queste proteine carrier sono essenzialmente una forma solubile e troncata del dominio extracellulare del GHR – GHR circolanti mobili che non si trovano all’interno delle membrane cellulari.[103] Il GH nel flusso ematico può anche esistere in forma libera o non legata e il rapporto tra frazione legata e non legata dipende dal modello pulsatile della sua secrezione. [104] I complessi di GH circolante nell’uomo possono essere costituiti da una delle due distinte forme di GH (22-kDa e 20-kDa), con circa il 90% rappresentato dalla forma molecolare da 22-kDa nonostante le prime stime abbiano ridotto tale numero.[105-106] Le moderne metodologie utilizzate nei test antidoping per la rilevazione indiretta del GH possono effettivamente sfruttare i rapporti in tempo reale delle molecole di GH circolanti all’interno del sistema di un atleta per determinare se questo ha usato rHGH nelle ultime 24-36 ore prima del test. La molecola dell’Ormone della Crescita, nella sua forma corretta da 22-kDa, è raffigurata di seguito. [107]:
Molecola dell’Ormone della Crescita nella sua forma corretta da 22kDa
In definitiva, questo GH circolante si lega al GHR, recettore dell’Ormone della Crescita appartenente alla superfamiglia delle proteine transmembrana presenti in tutte le cellule del corpo e che include il recettore della Prolattina e un certo numero di recettori delle citochine [108-110]. I livelli sulla superficie cellulare, o la densità recettoriale, dei GHR sono il determinante principale dell’affinità di legame del GH con le cellule. La traslocazione del GHR, cioè il recettore che si sposta dal nucleo di una cellula alla sua membrana esterna, è direttamente inibita dall’IGF-1 – che è uno dei molti meccanismi di feedback che esistono tra questi ormoni strettamente correlati. Mediante l’inibizione della traslocazione dei GHR, l’IGF-1 contribuisce direttamente ad abbassare la reattività di queste cellule a uno stimolo esterno di GH. [111]
Una analisi profonda dell’attivazione della Tirosina Chinasi, delle vie di segnalazione a valle e dell’espressione genica va oltre lo scopo di questo articolo. Quindi, mi limiterò a trattarlo superficialmente. E’ necessario comunque toccare alcuni dei punti più importanti della segnalazione intracellulare per comprendere veramente le basi del processo ipertrofico e del perché esiste sia sinergie composte che potenziali strategie ottimizzate per massimizzare il processo.
Come già accennato, i GHR esistono sulle membrane cellulari come omodimeri preformati e inattivi. Questo significa che il GHR ha due dimeri identici del recettore della proteina, e questi omodimeri saranno sempre accoppiati al JAK2 quando sono privi di attività enzimatica. Questo accoppiamento al JAK2 provoca un’azione inibitoria complessiva sul recettore. [112-113] In altre parole, il GHR rimane dormiente finché non viene attivato come parte del processo di legame GH/GHR. Quando una molecola di GH si lega al GHR, si verifica un cambiamento strutturale all’interno del GHR che si traduce in movimento effettivo dei domini intracellulari del recettore separatamente l’uno dall’altro. Questo smorza quell’azione inibitoria data dal JAK2 e consente loro di attivarsi l’un l’altro.[114-116]
Successivamente, la molecola di GH si lega sequenzialmente a uno dei due omodimeri di GHR e il completamento di questo processo di legame facilita le interazioni con il secondo omodimero. Dopo questo, i domini intracellulari di questo dimero-GHR appena formato subiscono una rotazione effettiva. La rotazione del nuovo dimero-GHR consente ai domini chinasi del JAK2 di essere in contatto l’uno con l’altro, consentendo loro di transactivare e ciascuno successivamente si lega a una molecola di JAK2. [117-118] Ciascuna molecola di JAK2 eseguirà quindi la fosforilazione incrociata (attivazione) dei residui di Tirosina, e sono proprio questi residui che formano “punti di attracco” per molte delle diverse molecole di segnalazione che costituiscono le vie di segnalazione a valle, e alla fine portano all’espressione genica. [114,118 -120] Uno dei più importanti percorsi a valle di maggior interesse per l’argomento specifico che qui vado trattando è il percorso JAK-STAT. Questo percorso è di vitale importanza sia per la trascrizione epatica di IGF-1 dal GH che per molti dei processi anabolici mediati dal GH all’interno del tessuto muscolare.
Nella seconda parte di questa serie di articoli mi occuperò più nel dettaglio del IGF-1, della “somatopausa” e della reale efficacia dell’uso del GH nello sport.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
The Most Effective Growth Hormone Protocol for Hypertrophy – by Chest Rockwell
Dall R, Longobardi S, Ehrnborg C, Keay N, Rosén T, Jørgensen JO, Cuneo RC,Boroujerdi MA, Cittadini A, Napoli R, Christiansen JS, Bengtsson BA, Sacca L,Baxter RC, Basset EE, Sönksen PH. The effect of four weeks of supraphysiological growth hormone administration on the insulin-like growth factor axis in women and men. GH-2000 Study Group. J Clin Endocrinol Metab. 2000 Nov;85(11):4193-200.
Kuhn CM. Anabolic steroids. Recent Prog Horm Res. 2002;57:411-34. Review.
Ribeiro SML, Kehayias JJ. Sarcopenia and the Analysis of Body Composition. Advances in Nutrition. 2014;5(3):260-267.
Schoenfeld BJ. The mechanisms of muscle hypertrophy and their application to resistance training. J Strength Cond Res. 2010 Oct;24 (10):2857-72.
Stickland NC. Muscle development in the human fetus as exemplified by m.sartorius: a quantitative study. J Anat. 1981 Jun;132(Pt 4):557-79.
Antonio J, Gonyea WJ. Skeletal muscle fiber hyperplasia. Med Sci Sports Exerc. 1993 Dec;25(12):1333-45. Review.
Fernández AM, Dupont J, Farrar RP, Lee S, Stannard B, Le Roith D.Muscle-specific inactivation of the IGF-I receptor induces compensatory hyperplasia in skeletal muscle. J Clin Invest. 2002 Feb;109(3):347-55.
Kadi F, Thornell LE. Training affects myosin heavy chain phenotype in the trapezius muscle of women. Histochem Cell Biol. 1999 Jul;112(1):73-8.
D’Antona G, Lanfranconi F, Pellegrino MA, Brocca L, Adami R, Rossi R, Moro G, Miotti D, Canepari M, Bottinelli R.Skeletal muscle hypertrophy and structure and function of skeletal muscle fibres in male body builders. J Physiol. 2006 Feb 1;570(Pt 3):611-27.
Cohen-Gadol AA, Liu JK, Laws ER Jr. Cushing’s first case of transsphenoidal surgery: the launch of the pituitary surgery era. J Neurosurg. 2005 Sep;103(3):570-4.
Li CH, Evans HM. THE ISOLATION OF PITUITARY GROWTH HORMONE. Science. 1944 Mar 3;99(2566):183-4.
SALMON WD Jr, DAUGHADAY WH. A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro. J Lab Clin Med. 1957 Jun;49(6):825-36.
Daughaday WH, Reeder C. Synchronous activation of DNA synthesis in hypophysectomized rat cartilage by growth hormone. J Lab Clin Med. 1966 Sep;68(3):357-68.
Garland JT, Lottes ME, Kozak S, Daughaday WH. Stimulation of DNA synthesis in isolated chondrocytes by sulfation factor. Endocrinology. 1972 Apr;90(4):1086-90.
Daughaday WH, Hall K, Raben MS, Salmon WD Jr, van den Brande JL, van Wyk JJ.Somatomedin: proposed designation for sulphation factor. Nature. 1972 Jan 14;235(5333):107.
Le Roith D, Bondy C, Yakar S, Liu JL, Butler A. The somatomedin hypothesis: 2001. Endocr Rev. 2001 Feb;22(1):53-74. Review.
D’Ercole AJ, Applewhite GT, Underwood LE. Evidence that somatomedin is synthesized by multiple tissues in the fetus. Dev Biol. 1980 Mar 15;75(2):315-28.
Han VK, Lund PK, Lee DC, D’Ercole AJ. Expression of somatomedin/insulin-like growth factor messenger ribonucleic acids in the human fetus: identification, characterization, and tissue distribution. J Clin Endocrinol Metab. 1988 Feb;66(2):422-9.
Isaksson OG, Jansson JO, Gause IA. Growth hormone stimulates longitudinal bone growth directly. Science. 1982 Jun 11;216 (4551):1237-9.
Isaksson OG, Lindahl A, Nilsson A, Isgaard J. Mechanism of the stimulatory effect of growth hormone on longitudinal bone growth. Endocr Rev. 1987 Nov;8(4):426-38. Review.
Green H, Morikawa M, Nixon T. A dual effector theory of growth-hormone action. Differentiation. 1985;29(3):195-8. Review.
Rinderknecht E, Humbel RE. The amino acid sequence of human insulin-like growth factor I and its structural homology with proinsulin. J Biol Chem. 1978 Apr 25;253(8):2769-76.
Klapper DG, Svoboda ME, Van Wyk JJ. Sequence analysis of somatomedin-C: confirmation of identity with insulin-like growth factor I. Endocrinology. 1983 Jun;112(6):2215-7.
Daughaday WH, Rotwein P. Insulin-like growth factors I and II. Peptide, messenger ribonucleic acid and gene structures, serum, and tissue concentrations. Endocr Rev. 1989 Feb;10(1):68-91. Review.
Hintz RL, Liu F. Demonstration of specific plasma protein binding sites for somatomedin. J Clin Endocrinol Metab. 1977 Nov;45(5):988-95.
BECK JC, McGARRY EE, DYRENFURTH I, VENNING EH. The metabolic effects of human and monkey growth hormone in man. Ann Intern Med. 1958 Nov;49(5):1090-105.
IKKOS D, LUFT R, GEMZELL CA. The effect of human growth hormone in man. Lancet. 1958 Apr 5;1(7023):720-1.
RABEN MS, HOLLENBERG CH. Effect of growth hormone on plasma fatty acids. J Clin Invest. 1959 Mar;38(3):484-8. PubMed PMID: 13641397
RABEN MS. Growth hormone. 1. Physiologic aspects. N Engl J Med. 1962 Jan 4;266:31-5.
RABEN MS. Growth hormone. 2. Clinical use of human growth hormone. N Engl J Med. 1962 Jan 11;266:82-6 concl.
ZIERLER KL, RABINOWITZ D. ROLES OF INSULIN AND GROWTH HORMONE, BASED ON STUDIES OF FOREARM METABOLISM IN MAN. Medicine (Baltimore). 1963 Nov;42:385-402.
RABINOWITZ D, KLASSEN GA, ZIERLER KL. EFFECT OF HUMAN GROWTH HORMONE ON MUSCLE AND ADIPOSE TISSUE METABOLISM IN THE FOREARM OF MAN. J Clin Invest. 1965 Jan;44:51-61.
Fineberg SE, Merimee TJ. Acute metabolic effects of human growth hormone. Diabetes. 1974 Jun;23(6):499-504.
Appleby BS, Lu M, Bizzi A, et al. Iatrogenic Creutzfeldt-Jakob Disease from Commercial Cadaveric Human Growth Hormone. Emerging Infectious Diseases. 2013;19(4):682-684. doi:10.3201/eid1904.121504.
Flodh H. Human growth hormone produced with recombinant DNA technology: development and production. Acta Paediatr Scand Suppl. 1986;325:1-9.
Crist DM, Peake GT, Egan PA, Waters DL. Body composition response to exogenous GH during training in highly conditioned adults. J Appl Physiol (1985). 1988 Aug;65(2):579-84.
Møller N, Copeland KC, Nair KS. Growth hormone effects on protein metabolism. Endocrinol Metab Clin North Am. 2007 Mar;36(1):89-100. Review.
Argetsinger LS, Carter-Su C. Mechanism of signaling by growth hormone receptor. Physiol Rev. 1996 Oct;76(4):1089-107. Review.
Hayashi AA, Proud CG. The rapid activation of protein synthesis by growth hormone requires signaling through mTOR. Am J Physiol Endocrinol Metab. 2007 Jun;292(6):E1647-55.
Kostyo JL. Rapid effects of growth hormone on amino acid transport and protein synthesis. Ann N Y Acad Sci. 1968 Feb 5;148(2):389-407.
Cameron CM, Kostyo JL, Adamafio NA, Brostedt P, Roos P, Skottner A, Forsman A, Fryklund L, Skoog B. The acute effects of growth hormone on amino acid transport and protein synthesis are due to its insulin-like action. Endocrinology. 1988 Feb;122(2):471-4.
Vanderkuur JA, Butch ER, Waters SB, Pessin JE, Guan KL, Carter-Su C. Signaling molecules involved in coupling growth hormone receptor to mitogen-activated protein kinase activation. Endocrinology. 1997 Oct;138(10):4301-7.
Costoya JA, Finidori J, Moutoussamy S, Seãris R, Devesa J, Arce VM. Activation of growth hormone receptor delivers an antiapoptotic signal: evidence for a role of Akt in this pathway. Endocrinology. 1999 Dec;140(12):5937-43.
Copeland KC, Nair KS. Acute growth hormone effects on amino acid and lipid metabolism. J Clin Endocrinol Metab. 1994 May;78(5):1040-7.
Umpleby AM, Boroujerdi MA, Brown PM, Carson ER, Sönksen PH. The effect of metabolic control on leucine metabolism in type 1 (insulin-dependent) diabetic patients. Diabetologia. 1986 Mar;29(3):131-41.
Horber FF, Haymond MW. Human growth hormone prevents the protein catabolic side effects of prednisone in humans. J Clin Invest. 1990 Jul;86(1):265-72.
Yarasheski KE, Campbell JA, Smith K, Rennie MJ, Holloszy JO, Bier DM. Effect of growth hormone and resistance exercise on muscle growth in young men. Am J Physiol. 1992 Mar;262(3 Pt 1):E261-7.
Zachwieja JJ, Bier DM, Yarasheski KE. Growth hormone administration in older adults: effects on albumin synthesis. Am J Physiol. 1994 Jun;266(6 Pt 1):E840-4.
Yarasheski KE, Zachwieja JJ, Campbell JA, Bier DM. Effect of growth hormone and resistance exercise on muscle growth and strength in older men. Am J Physiol. 1995 Feb;268(2 Pt 1):E268-76.
Healy ML, Gibney J, Russell-Jones DL, Pentecost C, Croos P, Sönksen PH, Umpleby AM. High dose growth hormone exerts an anabolic effect at rest and during exercise in endurance-trained athletes. J Clin Endocrinol Metab. 2003 Nov;88(11):5221-6.
Giannoulis MG, Jackson N, Shojaee-Moradie F, Nair KS, Sonksen PH, Martin FC, Umpleby AM. The effects of growth hormone and/or testosterone on whole body protein kinetics and skeletal muscle gene expression in healthy elderly men: a randomized controlled trial. J Clin Endocrinol Metab. 2008 Aug;93(8):3066-74.
Fryburg DA, Gelfand RA, Barrett EJ. Growth hormone acutely stimulates forearm muscle protein synthesis in normal humans. Am J Physiol. 1991 Mar;260(3 Pt 1):E499-504.
Fryburg DA, Louard RJ, Gerow KE, Gelfand RA, Barrett EJ. Growth hormone stimulates skeletal muscle protein synthesis and antagonizes insulin’s antiproteolytic action in humans. Diabetes. 1992 Apr;41(4):424-9.
Fryburg DA, Barrett EJ. Growth hormone acutely stimulates skeletal muscle but not whole-body protein synthesis in humans. Metabolism. 1993 Sep;42(9):1223-7.
Nørrelund H, Nair KS, Jørgensen JO, Christiansen JS, Møller N. The protein-retaining effects of growth hormone during fasting involve inhibition of muscle-protein breakdown. Diabetes. 2001 Jan;50(1):96-104.
Manson JM, Wilmore DW. Positive nitrogen balance with human growth hormone and hypocaloric intravenous feeding. Surgery. 1986 Aug;100(2):188-97.
Clemmons DR, Snyder DK, Williams R, Underwood LE. Growth hormone administration conserves lean body mass during dietary restriction in obese subjects. J Clin Endocrinol Metab. 1987 May;64(5):878-83.
Snyder DK, Clemmons DR, Underwood LE. Treatment of obese, diet-restricted subjects with growth hormone for 11 weeks: effects on anabolism, lipolysis, and body composition. J Clin Endocrinol Metab. 1988 Jul;67(1):54-61.
Tagliaferri M, Scacchi M, Pincelli AI, Berselli ME, Silvestri P, Montesano A, Ortolani S, Dubini A, Cavagnini F. Metabolic effects of biosynthetic growth hormone treatment in severely energy-restricted obese women. Int J Obes Relat Metab Disord. 1998 Sep;22(9):836-41.
Nørrelund H, Børglum J, Jørgensen JO, Richelsen B, Møller N, Nair KS, Christiansen JS. Effects of growth hormone administration on protein dynamics andsubstrate metabolism during 4 weeks of dietary restriction in obese women. Clin Endocrinol (Oxf). 2000 Mar;52(3):305-12.
Lundeberg S, Belfrage M, Wernerman J, von der Decken A, Thunell S, Vinnars E. Growth hormone improves muscle protein metabolism and whole body nitrogen economy in man during a hyponitrogenous diet. Metabolism. 1991 Mar;40(3):315-22
Fryburg DA. Insulin-like growth factor I exerts growth hormone- and insulin-like actions on human muscle protein metabolism. Am J Physiol. 1994 Aug;267(2 Pt 1):E331-6.
Russell-Jones DL, Umpleby AM, Hennessy TR, Bowes SB, Shojaee-Moradie F, Hopkins KD, Jackson NC, Kelly JM, Jones RH, Sönksen PH. Use of a leucine clamp to demonstrate that IGF-I actively stimulates protein synthesis in normal humans. Am J Physiol. 1994 Oct;267(4 Pt 1):E591-8.
Jacob R, Hu X, Niederstock D, Hasan S, McNulty PH, Sherwin RS, Young LH. IGF-I stimulation of muscle protein synthesis in the awake rat: permissive role of insulin and amino acids. Am J Physiol. 1996 Jan;270(1 Pt 1):E60-6.
Buijs MM, Romijn JA, Burggraaf J, De Kam ML, Cohen AF, Frölich M, Stellaard F, Meinders AE, Pijl H. Growth hormone blunts protein oxidation and promotes protein turnover to a similar extent in abdominally obese and normal-weight women. J Clin Endocrinol Metab. 2002 Dec;87(12):5668-74.
Gibney J, Wolthers T, Johannsson G, Umpleby AM, Ho KK. Growth hormone and testosterone interact positively to enhance protein and energy metabolism in hypopituitary men. Am J Physiol Endocrinol Metab. 2005 Aug;289(2):E266-71.
Le Roith D, Scavo L, Butler A. What is the role of circulating IGF-I? Trends Endocrinol Metab. 2001 Mar;12(2):48-52. Review.
Mauras N, Haymond MW. Are the metabolic effects of GH and IGF-I separable? Growth Horm IGF Res. 2005 Feb;15(1):19-27. Review.
Laron Z. Laron syndrome (primary growth hormone resistance or insensitivity): the personal experience 1958-2003. J Clin Endocrinol Metab. 2004 Mar;89(3):1031-44.
Muhammad A, van der Lely AJ, Neggers SJ. Review of current and emerging treatment options in acromegaly. Neth J Med. 2015 Oct;73(8):362-7. Review.
Lupu F, Terwilliger JD, Lee K, Segre GV, Efstratiadis A. Roles of growth hormone and insulin-like growth factor 1 in mouse postnatal growth. Dev Biol. 2001 Jan 1;229(1):141-62.
Waters MJ. The growth hormone receptor. Growth Horm IGF Res. 2016 Jun;28:6-10.
Laron Z, Ginsberg S, Webb M. Nonalcoholic fatty liver in patients with Laron syndrome and GH gene deletion – preliminary report. Growth Horm IGF Res. 2008 Oct;18(5):434-8.
Sharara FI. Value of growth hormone in ovulation induction? Fertil Steril. 1996 Jun;65(6):1259-61.
Kolibianakis EM, Venetis CA, Diedrich K, Tarlatzis BC, Griesinger G. Addition of growth hormone to gonadotrophins in ovarian stimulation of poor responders treated by in-vitro fertilization: a systematic review and meta-analysis. Hum Reprod Update. 2009 Nov-Dec;15(6):613-22.
Russell SM, Spencer EM. Local injections of human or rat growth hormone or of purified human somatomedin-C stimulate unilateral tibial epiphyseal growth in hypophysectomized rats. Endocrinology. 1985 Jun;116(6):2563-7.
Schlechter NL, Russell SM, Greenberg S, Spencer EM, Nicoll CS. A direct growth effect of growth hormone in rat hindlimb shown by arterial infusion. Am J Physiol. 1986 Mar;250(3 Pt 1):E231-5.
Isaksson OG, Ohlsson C, Nilsson A, Isgaard J, Lindahl A. Regulation of cartilage growth by growth hormone and insulin-like growth factor I. Pediatr Nephrol. 1991 Jul;5(4):451-3. Review.
Wang J, Zhou J, Cheng CM, Kopchick JJ, Bondy CA. Evidence supporting dual, IGF-I-independent and IGF-I-dependent, roles for GH in promoting longitudinal bone growth. J Endocrinol. 2004 Feb;180(2):247-55.
Yakar S, Rosen CJ, Bouxsein ML, Sun H, Mejia W, Kawashima Y, Wu Y, Emerton K, Williams V, Jepsen K, Schaffler MB, Majeska RJ, Gavrilova O, Gutierrez M, Hwang D, Pennisi P, Frystyk J, Boisclair Y, Pintar J, Jasper H, Domene H, Cohen P, Clemmons D, LeRoith D. Serum complexes of insulin-like growth factor-1 modulate skeletal integrity and carbohydrate metabolism. FASEB J. 2009 Mar;23(3):709-19.
Ohlsson C, Mohan S, Sjögren K, Tivesten A, Isgaard J, Isaksson O, Jansson JO, Svensson J. The role of liver-derived insulin-like growth factor-I. Endocr Rev. 2009 Aug;30(5):494-535.
Sotiropoulos A, Ohanna M, Kedzia C, Menon RK, Kopchick JJ, Kelly PA, Pende M. Growth hormone promotes skeletal muscle cell fusion independent of insulin-like growth factor 1 up-regulation. Proc Natl Acad Sci U S A. 2006 May 9;103(19):7315-20.
Wakelam MJ. The fusion of myoblasts. Biochem J. 1985 May 15;228(1):1-12. Review.
Pfäffle RW, Blankenstein O, Wüller S, Kentrup H. Combined pituitary hormone deficiency: role of Pit-1 and Prop-1. Acta Paediatr Suppl. 1999 Dec;88(433):33-41. Review.
Hemchand K, Anuradha K, Neeti S, Vaman K, Roland P, Werner B, Sharmila B. Entire prophet of Pit-1 (PROP-1) gene deletion in an Indian girl with combined pituitary hormone deficiencies. J Pediatr Endocrinol Metab. 2011;24(7-8):579-80.
Waters MJ, Shang CA, Behncken SN, Tam SP, Li H, Shen B, Lobie PE. Growth hormone as a cytokine. Clin Exp Pharmacol Physiol. 1999 Oct;26(10):760-4. Review.
Jansson JO, Edén S, Isaksson O. Sexual dimorphism in the control of growth hormone secretion. Endocr Rev. 1985 Spring;6(2):128-50. Review.
Giustina A, Veldhuis JD. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocr Rev. 1998 Dec;19(6):717-97. Review.
Hartman ML, Faria AC, Vance ML, Johnson ML, Thorner MO, Veldhuis JD. Temporal structure of in vivo growth hormone secretory events in humans. Am J Physiol. 1991 Jan;260(1 Pt 1):E101-10.
Takahashi Y, Kipnis DM, Daughaday WH. Growth hormone secretion during sleep. J Clin Invest. 1968 Sep;47(9):2079-90.
Ho KY, Veldhuis JD, Johnson ML, Furlanetto R, Evans WS, Alberti KG, Thorner MO. Fasting enhances growth hormone secretion and amplifies the complex rhythms of growth hormone secretion in man. J Clin Invest. 1988 Apr;81(4):968-75.
Jaffe CA, Ocampo-Lim B, Guo W, Krueger K, Sugahara I, DeMott-Friberg R, Bermann M, Barkan AL. Regulatory mechanisms of growth hormone secretion are sexually dimorphic. J Clin Invest. 1998 Jul 1;102(1):153-64.
Jessup SK, Dimaraki EV, Symons KV, Barkan AL. Sexual dimorphism of growth hormone (GH) regulation in humans: endogenous GH-releasing hormone maintains basal GH in women but not in men. J Clin Endocrinol Metab. 2003 Oct;88(10):4776-80.
Goldenberg N, Barkan A. Factors regulating growth hormone secretion in humans. Endocrinol Metab Clin North Am. 2007 Mar;36(1):37-55. Review.
Müller EE, Locatelli V, Cocchi D. Neuroendocrine control of growth hormone secretion. Physiol Rev. 1999 Apr;79(2):511-607. Review.
Murray RA, Maheshwari HG, Russell EJ, Baumann G. Pituitary hypoplasia in patients with a mutation in the growth hormone-releasing hormone receptor gene. AJNR Am J Neuroradiol. 2000 Apr;21(4):685-9.
Murray PG, Higham CE, Clayton PE. 60 YEARS OF NEUROENDOCRINOLOGY: The hypothalamo-GH axis: the past 60 years. J Endocrinol. 2015 Aug;226(2):T123-40.
Russell-Aulet M, Dimaraki EV, Jaffe CA, DeMott-Friberg R, Barkan AL. Aging-related growth hormone (GH) decrease is a selective hypothalamic GH-releasing hormone pulse amplitude mediated phenomenon. J Gerontol A Biol Sci Med Sci. 2001 Feb;56(2):M124-9
Dimaraki EV, Jaffe CA, Demott-Friberg R, Russell-Aulet M, Bowers CY, Marbach P, Barkan AL. Generation of growth hormone pulsatility in women: evidence against somatostatin withdrawal as pulse initiator. Am J Physiol Endocrinol Metab. 2001 Mar;280(3):E489-95.
Baumann G. Growth hormone heterogeneity: genes, isohormones, variants, and binding proteins. Endocr Rev. 1991 Nov;12(4):424-49. Review.
Vijayakumar A, Novosyadlyy R, Wu Y, Yakar S, LeRoith D. Biological effects of growth hormone on carbohydrate and lipid metabolism. Growth Horm IGF Res. 2010 Feb;20(1):1-7. doi: 10.1016/j.ghir.2009.09.002. Epub 2009 Oct 1. Review.
Herrington J, Carter-Su C. Signaling pathways activated by the growth hormone receptor. Trends Endocrinol Metab. 2001 Aug;12(6):252-7. Review.
Baumann G. Growth hormone binding protein. The soluble growth hormone receptor. Minerva Endocrinol. 2002 Dec;27(4):265-76. Review.
Bairoch A, Apweiler R. The SWISS-PROT protein sequence data bank and its new supplement TREMBL. Nucleic Acids Res. 1996 Jan 1;24(1):21-5.
Kelly PA, Djiane J, Postel-Vinay MC, Edery M. The prolactin/growth hormone receptor family. Endocr Rev. 1991 Aug;12(3):235-51. Review.
Vikman K, Carlsson B, Billig H, Edén S. Expression and regulation of growth hormone (GH) receptor messenger ribonucleic acid (mRNA) in rat adipose tissue, adipocytes, and adipocyte precursor cells: GH regulation of GH receptor mRNA. Endocrinology. 1991 Sep;129(3):1155-61.
Zou L, Menon RK, Sperling MA. Induction of mRNAs for the growth hormone receptor gene during mouse 3T3-L1 preadipocyte differentiation. Metabolism. 1997 Jan;46(1):114-8.
Leung KC, Waters MJ, Markus I, Baumbach WR, Ho KK. Insulin and insulin-like growth factor-I acutely inhibit surface translocation of growth hormone receptors in osteoblasts: a novel mechanism of growth hormone receptor regulation. Proc Natl Acad Sci U S A. 1997 Oct 14;94(21):11381-6.
Birzniece V, Sata A, Ho KK. Growth hormone receptor modulators. Rev Endocr Metab Disord. 2009 Jun;10(2):145-56.
Sawada T, Arai D, Jing X, Miyajima M, Frank SJ, Sakaguchi K. Molecular interactions of EphA4, growth hormone receptor, Janus kinase 2, and signal transducer and activator of transcription 5B. PLoS One. 2017 Jul 7;12(7):e0180785.
Brooks AJ, Dai W, O’Mara ML, Abankwa D, Chhabra Y, Pelekanos RA, Gardon O, Tunny KA, Blucher KM, Morton CJ, Parker MW, Sierecki E, Gambin Y, Gomez GA, Alexandrov K, Wilson IA, Doxastakis M, Mark AE, Waters MJ. Mechanism of activation of protein kinase JAK2 by the growth hormone receptor. Science. 2014 May 16;344(6185):1249783.
Liu Y, Berry PA, Zhang Y, Jiang J, Lobie PE, Paulmurugan R, Langenheim JF, Chen WY, Zinn KR, Frank SJ. Dynamic analysis of GH receptor conformational changes by split luciferase complementation. Mol Endocrinol. 2014 Nov;28(11):1807-19.
Waters MJ, Brooks AJ. JAK2 activation by growth hormone and other cytokines. Biochem J. 2015 Feb 15;466(1):1-11.
Brown RJ, Adams JJ, Pelekanos RA, Wan Y, McKinstry WJ, Palethorpe K, Seeber RM, Monks TA, Eidne KA, Parker MW, Waters MJ. Model for growth hormone receptor activation based on subunit rotation within a receptor dimer. Nat Struct Mol Biol. 2005 Sep;12(9):814-21. Epub 2005 Aug 21.
Lanning NJ, Carter-Su C. Recent advances in growth hormone signaling. Rev Endocr Metab Disord. 2006 Dec;7(4):225-35. Review.
Carter-Su C, Schwartz J, Smit LS. Molecular mechanism of growth hormone action. Annu Rev Physiol. 1996;58:187-207. Review.
Brooks AJ, Waters MJ. The growth hormone receptor: mechanism of activation and clinical implications. Nat Rev Endocrinol. 2010 Sep;6(9):515-25.
La Fursultiamina è un analogo sintetico della vitamina B1. Se viene somministrata ai topi, questi, come risposta, manifestano un aumento esponenziale della resistenza e della forza. Il tutto senza uno stimolo indotto dall’attività fisica. Ciò è stato riportato in uno studio pubblicato su Nutrients.(1)
Il nome ufficiale della Fursultiamina è Tiamina Tetraidrofurfuril Disolfuro [TTFD]. Come accennato, è un analogo sintetico della Tiamina o Vitamina B1 il quale, grazie alla sua struttura, presenta un assorbimento cellulare migliore rispetto alla normale Tiamina. È facilmente assorbita dal tratto intestinale e trasformata in Pirofosfato di Tiamina (TPP), il quale, al pari della classica Tiamina, svolge un ruolo importante come cofattore per gli enzimi delle vie metaboliche coinvolte nella produzione di Adenosina Trifosfato (ATP). Chimicamente, la Fursulftiamina è un derivato disolfuro della Tiamina ed è simile nella struttura all’Allitiamina.(2)
In molti paesi asiatici, la Fursultiamina è presente sul mercato come farmaco da banco per il trattamento della stanchezza, mentre in occidente è generalmente possibile trovare la Fursultiamina all’interno delle formulazioni di integratori per la perdita di peso e, a volte, nei pre-workout. Di solito la sua presenza all’interno dei supplementi è indicata in modo generale, in quantità sconosciute e, con tutta probabilità, molto piccole.
Gli autori dell’esperimento qui trattato nello specifico, hanno somministrato ai topi una dose orale giornaliera di Fursultiamina pari a 0 [Vehicle], 50mg [TTFD-1X], 100mg [TTFD-2X] e 500mg [TTFD-5X] per kg per 6 settimane. Il dosaggio utilizzato rapportato ad un uomo adulto equivarrebbe, in media, a 0 [Vehicle], 800mg [TTFD-1X], 1600mg [TTFD-2X] e 4000mg [TTFD-5X] di Fursultiamina al giorno.
Dopo il periodo di supplementazione, i ricercatori hanno fatto correre i topi il più a lungo possibile. In basso a sinistra si può osservare che gli animali trattati con il dosaggio più alto di Fursultiamina hanno mostrato una maggiore resistenza all’esercizio.
In alto a destra si può notare che la supplementazione di Fursultiamina ha portato ad una maggiore forza esprimibile da parte degli animali. La forza generata dalla spinta dei loro arti era maggiore nei topi trattati con Fursultiamina.
La supplementazione con Fursultiamina ha aumentato la quantità di glucosio presente nelle cellule. Dopo che i topi avevano corso, l’analisi ematica ha evidenziato una minore presenza di proteine degradate [BUN] e di Creatina Chinasi [CK] nei topi trattati con Fursultiamina. La Fursultiamina sembra avere un effetto protettivo a livello muscolare diminuendo la degradazione proteica indotta dall’esercizio fisico.
La Tiamina, nei dosaggi raccomandati dalle linee guida. è ampiamente accettata come integratore per mantenere le funzioni fisiologiche ottimali. I ricercatori hanno riportato che, per l’applicazioni pratica della supplementazione di Fursultiamina negli atleti è consigliabile un’assunzione giornaliera pari a 40mg/kg – dose equivalentemente convertita dal dosaggio di 500 mg/kg utilizzato nello studio, in base alla superficie corporea tra i topi e gli esseri umani secondo la formula US Food and Drug Administration, per il miglioramento della regolazione energetica e delle prestazioni in una strategia nutrizionale combinata, comprendente un carico glucidico adeguato alla domanda energetica consequenziale all’esercizio prolungato.
Per un atleta di 80Kg, la dose consigliata dai ricercatori sarebbe pari a 3.200mg di Fursultiamina al giorno. C’è il probabile rischio che tale dosaggio sia, in effetti, eccessivamente alto. In base ad un promettente studio svolto su esseri umani (3) il dosaggio efficace, e ragionevolmente “sicuro”, di Fursultiamina risulta essere di 10mg/Kg (800mg/die per un atleta di 80Kg).
La questione degli Estrogeni nell’ambito delle preparazioni comprendenti farmaci per il miglioramento delle prestazioni, in special modo nel Bodybuilding, è generalmente mal compresa e, di conseguenza, mal gestita. Complice di questa “mala gestione” è l’ignoranza sia specifica, cioè portata da una scarsa disponibilità di testi di facile accesso (e comprensione) che trattano l’argomento in modo esaustivo, sia indotta dal comportamento della maggior parte dei preparatori o degli atleti “solisti” che ignorano totalmente la necessità di un aggiornamento continuo in tutte le componenti della preparazione (allenamento, nutrizione e supplementazione). Con questa pubblicazione è mia intenzione fornire una buona base di conoscenza sugli Estrogeni (loro biosintesi, azioni ecc…) e sulla migliore gestione di questi nel contesto di una preparazione.
Iniziamo per gradi…
Estrogeni e loro caratteristiche/azioni principali (nella donna e nell’uomo)
Estradiolo
Gli Estrogeni sono i principali ormoni sessuali femminili. Si tratta di ormoni steroidei responsabili dello sviluppo e della regolazione del sistema riproduttivo femminile e delle caratteristiche sessuali secondarie. Con il termine estrogeno ci si può anche riferire a qualsiasi sostanza, naturale o sintetica, che mima gli effetti dell’ormone naturale.(1) L’estrogeno prevalente in quantità e potenza è l’Estradiolo, anche se diversi metaboliti dell’estradiolo hanno anche una marcata attività ormonale estrogenica. Versioni sintetiche degli estrogeni vengono utilizzate come principio attivo di farmaci contraccettivi orali, nella terapia ormonale sostitutiva per le donne in postmenopausa, nelle donne ipogonadiche e nei transgender, oltre che nel trattamento di alcuni tumori ormone-sensibili come il cancro alla prostata e il cancro al seno. Gli Estrogeni sono una delle tre classi di ormoni sessuali, insieme agli Steroidi Androgeni/Anabolizzanti come il Testosterone e i progestinici come il Progesterone.
Gli Estrogeni sono sintetizzati in tutti i vertebrati (2) così come in alcuni insetti.(3) Le tre principali forme di estrogeno presenti naturalmente nelle donne sono l’Estrone (E1), l’Estradiolo (E2) e l’Estriolo (E3). Un altro tipo di Estrogeno chiamato Estetrolo (E4) viene prodotto solo durante la gravidanza. Quantitativamente, gli Estrogeni circolano a livelli inferiori rispetto agli Androgeni sia negli uomini che nelle donne.(4) Mentre i livelli di Estrogeni sono significativamente più bassi nei maschi rispetto alle femmine, gli Estrogeni, tuttavia, hanno anche importanti ruoli fisiologici nei soggetti di sesso maschile.(5)
Come tutti gli ormoni steroidei, gli Estrogeni penetrano facilmente attraverso la membrana cellulare. Una volta all’interno della cellula, si legano e attivano i recettori degli estrogeni (ER) che a loro volta modulano l’espressione di molti geni.(6) Inoltre, gli Estrogeni si legano e attivano rapidamente i recettori degli estrogeni di membrana (mER), (7)(8) come i GPER (GPR30).(9)
Come già precedentemente accennato, i tre principali Estrogeni presenti naturalmente nelle donne sono l’Estrone (E1), l’Estradiolo (E2) e l’Estriolo (E3). L’Estradiolo è l’estrogeno predominante negli anni riproduttivi sia in termini assoluti nel siero che in termini di attività estrogenica. Durante la menopausa, l’Estrone è l’Estrogeno circolante predominante mentre durante la gravidanza è l’Estriolo ad essere l’Estrogeno circolante predominante in termini di livelli sierici. Anche se l’Estriolo è il più abbondante dei tre estrogeni è anche il più debole, mentre l’Estradiolo è il più forte con una potenza di circa 80 volte quella dell’Estriolo.(10) Quindi, l’Estradiolo è l’Estrogeno più importante nelle donne non in gravidanza che si trovano tra le fasi del menarca e della menopausa.
Estetrolo
Tuttavia, durante la gravidanza questo ruolo passa all’Estriolo e nelle donne in postmenopausa l’Estrone diventa la forma primaria di Estrogeno nel corpo. Come già accennato, un altro tipo di Estrogeno chiamato Estetrolo (E4) viene prodotto solo durante la gravidanza. Oltre che dai follicoli ovarici, tutte le diverse forme di estrogeni sono sintetizzate a partire dagli Androgeni, in particolare dal Testosterone e dal Androstenedione, da parte dell’enzima aromatasi.
Altri substrati utilizzati dal corpo per la sintesi estrogenica, senza il coinvolgimento primario dell’enzima aromatasi, sono il 27-idrossicholesterolo, il Deidroepiandrosterone (DHEA), il 7-oxo-DHEA, il 7α-idrossi-DHEA, il 16α-idrossi-DHEA, il 7β-idrossiepiandrosterone, l’Androstenedione (A4), l’Androstenediolo (A5), il 3α-androstanediolo e il 3β-androstanediolo. Alcuni metaboliti estrogenici, come i catecol-estrogeni 2-idrossiestradiolo, 2-idrossestrone, 4-idrossiestradiolo e 4-idrossestrone, nonché il 16α-idrossiestrone, sono composti estrogenici con vari gradi di attività. L’importanza biologica di questi estrogeni minori non è del tutto chiara. Tutti questi composti possono avere importanti funzioni estrogeniche nel corpo.
Steroidogenesi; gli Estrogeni sono rappresentati in basso a destra.
Recettore degli Estrogeni alfa (ERα)
Le azioni degli Estrogeni sono mediate dal Recettore Estrogeno (ER), una proteina nucleare dimerica che si lega al DNA e controlla l’espressione genica. Come altri ormoni steroidei, l’Estrogeno entra passivamente nella cellula nella quale si lega e attiva il Recettore dell’Estrogeno. Il complesso ER si lega a specifiche sequenze di DNA chiamate “elemento di risposta ormonale” per attivare la trascrizione di geni bersaglio: in uno studio nel quale si è utilizzate una linea di cellule del cancro al seno estrogeno-dipendente come modello, sono stati identificati 89 di questi geni.(11) Poiché l’Estrogeno entra in tutte le cellule, le sue azioni dipendono dalla presenza del ER nella cellula. L’ER è espresso in tessuti specifici tra cui l’ovaio, l’utero e il seno. Gli effetti metabolici degli Estrogeni nelle donne in postmenopausa sono stati legati al polimorfismo genetico dell’ER. (12)
Mentre gli Estrogeni sono presenti sia negli uomini che nelle donne, sono solitamente presenti a livelli significativamente più alti nelle donne in età riproduttiva. Essi promuovono lo sviluppo delle caratteristiche sessuali secondarie femminili, come i seni, e sono anche coinvolti nell’ispessimento dell’endometrio e altri aspetti della regolazione del ciclo mestruale. Nei maschi, l’Estrogeno regola alcune funzioni del sistema riproduttivo importanti per la maturazione dello sperma (13)(14)(15) e sembra essere necessario per una sana libido.(16) Inoltre, vi sono molti altri cambiamenti strutturali indotti dall’Estrogeno oltre ad altre funzioni.
Gli Estrogeni, nelle femmine, sono prodotti principalmente dalle ovaie e, durante la gravidanza, dalla placenta.(17) L’Ormone Follicolo Stimolante (FSH) stimola la produzione ovarica di Estrogeni da parte delle cellule della granulosa dei follicoli ovarici e dei corpi lutei. Alcuni Estrogeni vengono anche prodotti in quantità minori in altri tessuti come il fegato, le ghiandole surrenali e i seni. Queste fonti secondarie di Estrogeni sono particolarmente importanti nelle donne in postmenopausa. Anche nelle cellule adipose vengono sintetizzati Estrogeni (vedi azione dell’Enzima Aromatasi)(18)
Nelle femmine, la sintesi degli Estrogeni inizia nelle cellule della teca follicolare interna nell’ovaio, dalla sintesi del Androstenedione a partire dal Colesterolo. L’Androstenedione è uno steroide con debole attività androgenica il cui scopo è prevalentemente quello di fungere da precursore di più potenti androgeni come il Testosterone così come dell’Estrogeno. Questo steroide attraversa la membrana basale nelle cellule della granulosa circostanti, dove o viene immediatamente convertito in Estrone, o in Testosterone e poi successivamente in Estradiolo. La conversione del Androstenedione in Testosterone è catalizzata dal 17β-idrossisteroide deidrogenasi (17β-HSD), mentre la conversione del Androstenedione e del Testosterone in Estrone ed Estradiolo è rispettivamente catalizzata dal aromatasi, entrambi enzimi che sono espressi nelle cellule della granulosa. Al contrario, le cellule della granulosa non dispongono di 17α-idrossilasi e 17,20-liasi, mentre le cellule della teca follicolare interna esprimono questi enzimi e il 17β-HSD, ma non l’aromatasi. Quindi sia le cellule della granulosa che le cellule della teca follicolare interna sono essenziali per la produzione di Estrogeni nelle ovaie.
I livelli di Estrogeni variano durante il ciclo mestruale, con livelli più alti vicino alla fine della fase follicolare e poco prima dell’ovulazione.
Intervalli di riferimento nelle concentrazioni di Estradiolo, il principale Estrogeno circolante, durante il ciclo mestruale. CYP1A1
Gli Estrogeni sono metabolizzati tramite idrossilazione da enzimi citocromo P450 quali CYP1A1 e CYP3A4 e tramite coniugazione del estrogeno sulfotransferasi (sulfato) e del UDP-glucuroniltransferasi (glucuronidazione). Inoltre, l’Estradiolo viene deidrogenato dal 17β-idrossiesteroide deidrogenasi nel molto meno potente Estrone. Queste reazioni si verificano principalmente nel fegato, ma avvengono anche in altri tessuti.
Gli estrogeni sono implicati nel rilascio di GH e IGF-1 sia nell’uomo che nella donna. Le donne hanno mediamente livelli di GH più elevati sebbene sembri che ne siano meno sensibili (vedi percentuale di grasso corporeo femminile).
Quindi, per riepilogare, gli Estrogeni nelle femmine regolano la maturazione sessuale intervenendo nello sviluppo dell’apparato genitale.
La loro massiccia secrezione in epoca puberale induce la chiusura delle cartilagini di coniugazione delle ossa lunghe, terminando di fatto, la fase di accrescimento staturale.
Gli estrogeni stimolano lo sviluppo stromale della mammella e il mantenimento delle caratteristiche femminili secondarie (crescita delle mammelle, distribuzione dei peli, voce, statura, ossatura, distribuzione del grasso).
Permettono la fecondazione e la gravidanza, intervenendo nella regolazione del ciclo mestruale.
Regolano la distribuzione del grasso corporeo, favorendone il deposito nelle anche, nelle natiche, nelle cosce e nell’addome al di sotto dell’ombelico.
Mantengono il trofismo osseo ed hanno quindi azione protettiva nei confronti dell’osteoporosi
Stimolano la sintesi di trigliceridi e l’aumento delle lipoproteine ad alta densità (HDL) proteggendo le pareti vasali dal danno arteriosclerotico.
Stimolano la lipolisi nel tessuto muscolare ed adiposo. Per questo motivo gli estrogeni migliorano la prestazione degli sport di durata risparmiando il glicogeno muscolare a scapito degli acidi grassi
Regolano molte funzioni cerebrali fra cui l’attenzione e la memoria.
Stimolano la sintesi epatica di numerosi enzimi e proteine (SHBG, angiotensinogeno).
Enzima Aromatasi
Nell’uomo, invece, gli estrogeni circolanti derivano per la maggior parte dall’aromatizzazione degli Androgeni circolanti (19). Il complesso enzimatico conosciuto come aromatasi è responsabile dell’aromatizzazione dell’anello A degli Androgeni che comporta la trasformazione in sostanze ad attività estrogenica. (20) (21) (22)
Recettore degli Estrogeni beta (ERβ)
La trascrizione del gene per l’aromatasi e l’espressione dell’enzima aromatasi avviene in un ampio numero di tessuti, come i testicoli, il tessuto adiposo, il muscolo, il fegato, il cervello, i follicoli piliferi, i fibroblasti etc. (20). L’azione degli Estrogeni, come già accennato, si esplica attraverso il loro legame a recettori specifici, che appartengono alla ‘superfamiglia’ dei recettori nucleari (23). Finora sono stati caratterizzati due tipi di recettori per gli Estrogeni (ER): l’ER classico, ora denominato ERα (24) (25) (26), ed un altro sottotipo denominato ERβ (27). Entrambi sono recettori intranucleari che, una volta attivati, modulano a livello genomico l’attività di trascrizione genica. Di recente è stata documentata anche una via non-genomica dell’azione degli Estrogeni, che prevede un’interazione plasma-membrana del recettore estrogenico (28) (29) (30).
Conversione del Testosterone in Estradiolo
L’Estradiolo (E2) nel maschio è sintetizzato dal Testosterone attraverso l’enzima aromatasi o dall’Estrone attraverso la 17β-idrossisteroido deidrogenasi (17β-HSD) (31). L’E2 prodotto giornalmente nel maschio è stimato essere circa 35-45 μg (0.130- 0.165 μmol), dei quali il 15-20% è direttamente prodotto dal testicolo (32) (19), un 60% è prodotto dall’aromatizzazione periferica del Testosterone circolante ed il restante 20% dalla conversione periferica dell’Estrone. L’Estrone (E1) può anche essere direttamente secreto dai surreni (33) o derivare dall’aromatizzazione periferica dell’Androstenedione, quest’ultimo prodotto in parte dal surrene ed in parte derivante dalla conversione periferica del Testosterone. I testicoli comunque contribuiscono maggiormente, rispetto ai surreni, alla totale produzione di Estradiolo circolante e la via enzimatica di sintesi maggiormente coinvolta nella secrezione di Estrogeni nel maschio è quella aromatasica, mentre la via della 17β-HSD ha una minore rilevanza “in vivo”. Solo il 2-3% dell’Estradiolo circolante è libero, la maggior parte dell’ormone è legato all’albumina o alla globulina legante gli ormoni sessuali (SHBG) (34) e la frazione non legata alla SHBG è considerata bioattiva a livello dei tessuti bersaglio.
SHBG
I progressi negli studi effettuati negli ultimi anni hanno infatti dimostrato in maniera inequivocabile il ruolo fisiologico che gli Estrogeni hanno anche nell’uomo. Gli Estrogeni svolgono un ruolo chiave sul sistema immunitario, a livello del timo, agiscono a livello ipofisario e cardiovascolare, osseo e del metabolismo glucidico e lipidico.
Un eccesso estrogenico nella donna favorisce l’accumulo di tessuto adiposo e la comparsa di ritenzione idrica, oltre ad esporre la donna ad un elevato rischio di sviluppare alcune forme di cancro come quello alla mammella, l’insulinoresistenza, l’infertilità e l’ovaiopolicistico. Nell’uomo l’eccesso estrogenico è legato ad effetti collaterali spiacevoli tra i quali troviamo la ginecomastia, la diminuzione del desiderio sessuale, problemi di erezione (che si verifica anche con un calo estrogenico eccessivo), e diminuzione della fertilità.
Ricordiamoci però che, tanto nella donna come nell’uomo, un livello estremamente basso di Estrogeni comporta diversi problemi tra i quali emergono: elevati livelli sierici di Trigliceridi, LDL e colesterolo totale con ridotti livelli di HDL (35)(36)(37)(38)(39), ridotti livelli di LDL, HDL e colesterolo totale con Trigliceridi normali (40)(41), osteoporosi, dolore osseo, e diabete mellito di tipo II (37).
Si noti che la descrizione di pazienti maschi affetti da deficit congenito di Estrogeni (42) (43) (44) (45) (46) (47) (48) (49) (50) (51) ha fornito un modello in vivo unico che ha permesso di stabilire che la completa ossificazione delle cartilagini epifisarie, anche nel maschio, non può avvenire senza l’azione degli Estrogeni. Inoltre, è stato dimostrato che gli Androgeni da soli non sono sufficienti per garantire una normale mineralizzazione dello scheletro in età adulta, dal momento che un quadro di osteopenia e/o osteoporosi è stato documentato in tutti i pazienti con deficit congenito di Estrogeni (42) (43) (44) (45) (46) (47) (48) (49) (50) (51). Ad oggi è stato chiarito che gli effetti degli steroidi sessuali sul processo di mineralizzazione ossea nel maschio sono solo in parte da ascrivere agli Androgeni di origine testicolare, mentre l’azione degli Estrogeni, che derivano dall’aromatizzazione degli androgeni stessi, sembra giocare un ruolo di maggiore importanza (52) (53). Infatti in letteratura vi sono evidenze riguardo l’associazione tra ipogonadismo e riduzione della massa ossea in entrambi i sessi (54) (55) (56) (57) (58) (59). Inoltre, il concetto generale che gli Androgeni mantengono la massa ossea nell’uomo, così come gli Estrogeni la mantengono nella donna, continua ad essere il punto di vista prevalente tra i medici, sebbene recenti studi suggeriscano il contrario (42) (43) (44) (45) (60) (61). Nel 1997, infatti, è stata descritta l’efficacia della terapia Estrogenica nell’ottenere sia la completa ossificazione delle epifisi che l’aumento di densità minerale ossea (BMD) in un paziente maschio affetto da deficit congenito di aromatasi (44).
Fonti degli estrogeni circolanti nel maschio ed organi e tessuti bersaglio dell’azione degli estrogeni. La figura indica la produzione in µmol/L.(modificata da Baird et al., 1969a e Saez et al., 1972)
Una funzione interessante dell’Estradiolo nel maschio è rappresentata dal suo impatto sulla funzione sessuale. Come già accennato, l‘Estradiolo negli uomini è essenziale per modulare la libido, la funzione erettile e la spermatogenesi. I recettori degli Estrogeni, così come l’aromatasi, sono abbondanti nel cervello, nel pene e nei testicoli, organi importanti per la funzione sessuale. Nel cervello, la sintesi dell’Estradiolo è elevata nelle aree correlate all’auda sessuale. Inoltre, nel pene, i recettori degli Estrogeni si trovano in tutto il corpus cavernosum con elevata concentrazione intorno ai fasci neurovascolari. Un livello basso di Testosterone e elevato di Estrogeni aumenta l’incidenza della disfunzione erettile indipendentemente l’uno dall’altro. Nei testicoli, la spermatogenesi è modulata a tutti i livelli dagli Estrogeni, a partire dall’asse ipotalamo-ipofisi-gonadi, seguita dalle cellule di Leydig, Sertoli e germinali e terminando con l’epitelio ductale, l’epididimo e lo sperma maturo. La regolazione delle cellule testicolari mediante l’Estradiolo mostra sia un’influenza inibitoria che una stimolatoria, indicando un intricata sinfonia di modulazione dose-dipendente e temporalmente sensibile.(62)
Adesso che abbiamo una buona conoscenza di base sugli Estrogeni e le loro azioni sia nell’uomo che nella donna, possiamo passare a trattare la questione della gestione funzionale degli Estrogeni durante la preparazione.
AAS aromatizzabili e gestione estrogenica
Come ormai ben sappiamo, il Testosterone è il substrato primario usato nell’uomo per la sintesi dell’Estradiolo, principale ormone sessuale femminile. Con una leggera alterazione strutturale del Testosterone dovuta all’azione dell’enzima aromatasi, l’Estradiolo viene prodotto nel maschio. L’attività dell’enzima aromatasi nel maschio si verifica in diverse regioni del corpo , compreso il tessuto adiposo, il fegato, le gonadi, il Sistema Nervoso Centrale e il tessuto muscolo scheletrico.(63) (64) (65) (66) (67) In un soggetto sano di sesso maschile , la quantità di Estrogeni prodotta non è generalmente molto significativa ed è basata sulle richieste e caratteristiche fisiologiche divenendo anche un vantaggio, come precedentemente visto, nella qualità e quantità dei valori di colesterolo e nella mineralizzazione ossea. Nei casi in cui le concentrazioni di Estrogeni (in particolare dell’Estradiolo) raggiungono livelli ematici elevati, che ciò avvenga come conseguenza non patologica (vedi obesità e cattiva alimentazione) o patologica (vedi neoplasia al testicolo), o come conseguenza del uso di AAS aromatizzabili e dell’alterazione omeostatica dovuta all’uso di tali composti, il soggetto sperimenta effetti avversi tra i quali vi sono la comparsa di ritenzione idrica, l’accumulo adiposo con modello femminile e la ginecomastia. Per questo motivo, molti atleti si concentrano sulla riduzione dell’incremento e/o dell’attività degli estrogeni nel corpo con l’uso di inibitori dell’aromatasi (es. Anastrozolo, Letrozolo e Exemestane), di Modulatori Selettivi del Recettore degli Estrogeni (SERM) (es. Tamoxifene e Raloxifene) o di inibitori della biosintesi steroidea (es. Aminoglutettimide e Trylostano), in specie quando il ciclo che stanno seguendo è composto da molecole altamente aromatizzabili (es. Testosterone e Methandrostenolone) o quando l’atleta desidera avere un maggior controllo estrogenico in un contesto nel quale si cerca di aumentare la definizione muscolare.
Oxymetholone
Nota:Con l’uso del Oxymetholone, il quale possiede attività estrogenica intrinseca, la gestione del fattore estrogenico (in particolare con la presenza di substrati soggetti all’aromatizzazione) assume caratteristiche leggermente più “complesse”. L’Oxymetholone è infatti un AAS avente azione altamente estrogenica. Tale azione, per l’appunto, non è dipendente dall’aromatizzazione del composto in estrogeno – essendo un derivato del DHT non può essere aromatizzato. L’uso di inibitori dell’aromatasi come l’Anastrozolo o l’Exemestane non influiscone sulla estrogenicità relativa di questo AAS. Alcuni hanno suggerito che l’alto livello di attività estrogenica del Oxymetholone sia in realtà dovuto alla presunta azione progestinica del composto, simile al Nandrolone. Gli effetti collaterali estrogenici e progestinici possono essere molto simili (anche perché l’azione dei due steroidi è co-attiva), il che ha reso plausibile questa ipotesi. Tuttavia, esiste uno studio nel quale è stata esaminata l’attività progestinica del Oxymetholone. Lo studio in questione stabilì che non vi era alcuna attività progestinica derivante dal composto.(68) Con tali risultati, sembra plausibile che l’Oxymetholone possa attivare il recettore degli estrogeni similmente a, ma più profondamente, dell’androgeno estrogenico Methandriolo. In definitiva, l’attività estrogenica di questo composto può essere gestita solamente con l’uso di SERM come il Tamoxifene o il Raloxifene dal momento che, per i motivi sopra detti, composti anti aromatasi e gli inibitori della biosintesi steroidea non hanno alcuna incidenza sull’attività estrogenica di questo AAS.
Anche alla luce di quanto accennato in precedenza, non dobbiamo tuttavia essere indotti a pensare che gli Estrogeni non diano nessun beneficio prestativo. Livelli di Estradiolo mantenuti nei limiti del range di riferimento offrono diversi vantaggi.
Gli atleti sono da anni a conoscenza del fatto che gli AAS aromatizzabili presentano un vantaggio nell’anabolismo muscolare, ma è solo di recente che siamo arrivati finalmente a capire i meccanismi che stanno alla base di tale effetto. Sembra che i motivi vadano oltre l’aumento di peso e di forza che si attribuiscono alla ritenzione idrica data dall’attività estrogenica, comprendendo che questo ormone (Estradiolo) possiede effettivamente un effetto diretto sui processi anabolici. Ciò si manifesta attraverso l’aumento dell’utilizzo del Glucosio, la secrezione dell’Ormone della Crescita (e di IGF-1) e la proliferazione dei recettori degli Androgeni.
Estrogeni e utilizzo del Glucosio
G6PD
Gli Estrogeni possono giocare un ruolo molto importante nella promozione di uno stato anabolico influenzando l’utilizzo del Glucosio nel tessuto muscolare. Ciò avviene attraverso un’alterazione del livello di glucosio 6-fosfato deidrogenasi disponibile, un enzima direttamente legato all’uso del glucosio per la crescita e il recupero del tessuto muscolare.(69) (70) Più specificamente, il G6PD è l’enzima che catalizza la prima reazione della via dei pentoso fosfati (definita anche Shunt dell’Esosomonofosfato [HMP shunt] o PPP da Pentose phosphate pathway), un processo metabolico citoplasmatico, parallelo alla glicolisi, in grado di generare NADPH e zuccheri pentosi (a 5 atomi di carbonio). Durante il periodo di rigenerazione tissutale seguente il danno muscolare, i livelli di G6PD aumentano considerevolmente, il che è ritenuto essere un meccanismo che il corpo attua per migliorare il recupero quando necessario. Sorprendentemente, scopriamo che l’Estrogeno è direttamente legato al livello di G6PD che deve essere messo a disposizione delle cellule in questa fase di recupero. In sintesi, gli Estrogeni svolgono anche una azione metabolica accelerando la sintesi degli acidi nucleici, delle proteine e del glicogeno.
Il legame tra Estrogeno e G6PD è stato stabilito attraverso uno studio nel quale è stato dimostrato che i livelli di questo enzima deidrogenasi aumentano dopo la somministrazione di Testosterone Propionato. Lo studio ha inoltre mostrato che era l’aromatizzazione del Testosterone in Estradiolo ad essere direttamente responsabile di questo aumento e non l’azione Androgena del AAS.(71) In questo studio sono stati presi in esame anche due composti non-aromatizzabili, il Dihydrotestosterone e il Fluoxymesterone, ma l’effetto avuto con l’uso di Testosterone Propionato non è stato replicato. Inoltre, l’effetto positivo del Testosterone Propionato è stato bloccato quando è stato aggiunto l’inibitore dell’aromatasi 4-idrossiandrostenedione (Formestano), mentre la somministrazione di 17-beta Estradiolo da solo ha causato un aumento di G6PD simile a quello osservato con il Testosterone Propionato. Il 17-alfa Estradiolo, isomero ormonalmente inattivo del 17-beta Estradiolo, che non può legarsi al recettore estrogenico, non ha avuto alcun effetto sul G6PD. Ulteriori prove sono state fatte utilizzando il Testosterone Propionato con l’antiandrogeno Flutamide dimostrando che anche quest’ultimo farmaco non ha alterato negativamente le concentrazioni di G6PD, sottolineando l’effetto indipendente dall’azione del Testosterone con il recettore androgeno.
Estrogeni e GH/IGF-1
L’Estrogeno può anche svolgere un ruolo importante nella produzione dell’Ormone della Crescita e del IGF-1. L’IGF-1 (Fattore di Crescita Insulino-Simile) è un ormone anabolizzante rilasciato nel fegato e in vari tessuti periferici attraverso lo stimolo dell’Ormone della Crescita. L’IGF-1 è il principale responsabile dell’attività anabolica dell’Ormone della Crescita, attraverso una maggiore ritenzione di azoto / sintesi proteica e iperplasia cellulare (proliferazione). Uno dei primi studi da portare all’attenzione riguardante la questione qui trattata ha esaminato gli effetti del SERM Tamoxifene sui livelli di IGF-1, dimostrando che il composto esaminato ha un effetto soppressivo.(72) Un secondo studio, forse più degno di nota, svolto nel 1993, ha esaminato gli effetti della terapia sostitutiva del Testosterone sui soli livelli di GH e IGF-1 e li ha confrontati con gli effetti del Testosterone combinato nuovamente con il Tamoxifene. (73) Quando è stato somministrato Tamoxifene, i livelli di GH e IGF-1 hanno subito un notevole calo, mentre entrambi i valori sono stati elevati con la somministrazione di solo Testosterone Enantato. Un altro studio ha mostrato come la somministrazione settimanale di 300mg di Testosterone Enantato causi un lieve aumento del IGF-1 in uomini normali. Questi 300mg di Testosterone Enantato hanno causato un aumento dei livelli di Estradiolo, cosa normale con tale dose. Questo è stato confrontato con l’effetto portato con lo stesso dosaggio di Nandrolone Decanoato; tuttavia, questo AAS non è riuscito a produrre lo stesso aumento nei livelli di IGF-1. Questo risultato è molto interessante, specialmente quando notiamo che i livelli di Estrogeni sono stati effettivamente ridotti quando questo AAS è stato somministrato (ricordiamoci che il Nandrolone aromatizza il 20% del Testosterone). (74) Un altro studio ha dimostrato che la secrezione di GH e IGF-1 è aumentata con la somministrazione di Testosterone in maschi con pubertà ritardata, mentre il Dihydrotestosterone (non aromatizzabile) sembra sopprimere la secrezione di GH e IGF-1. (75)
Estrogeni e Recettori degli Androgeni
Recettore degli Androgeni
È stato anche dimostrato che l’Estrogeno può aumentare la concentrazione dei recettori degli androgeni in alcuni tessuti. Ciò è stato dimostrato in studi svolti su ratti che hanno esaminato gli effetti degli Estrogeni sui Recettori degli Androgeni cellulari in animali sottoposti ad orchiectomia (rimozione dei testicoli, spesso effettuata per diminuire la produzione endogena di Androgeni). Secondo lo studio, la somministrazione di Estrogeni ha determinato un aumento del legame recettoriale nel muscolo levator ani del Metribolone pari al 480%. (76) La spiegazione per tale evento suggerita è che l’Estrogeno stimola direttamente la proliferazione dei Recettori Androgeni, o forse ne diminuisce il tasso di danneggiamento. Anche se la crescita del muscolo levator ani è comunemente usata come riferimento per determinare l’attività anabolica dei composti steroidei, esso è un muscolo dell’organo sessuale ed è diverso dal tessuto muscolo scheletrico in quanto possiede una concentrazione molto più alta di Recettori Androgeni. Questo studio, tuttavia, ha esaminato l’effetto sui AR dato dagli Estrogeni nei tessuti musco scheletrici veloci (tibialis anterior e extensor digitorum longus), ma senza notare lo stesso aumento visto nel levator ani. Anche se scoraggiante come notizia, il fatto che l’Estrogeno possa aumentare il legame del Recettore degli Androgeni in qualsiasi tessuto rimane una scoperta estremamente significativa, soprattutto alla luce del fatto che ora sappiamo che gli Androgeni hanno alcuni effetti positivi sulla crescita muscolare mediati al di fuori del tessuto muscolare.
Estrogeni e fatica/stanchezza
La così detta “Steroid Fatigue” è una frase comunemente utilizzata in riferimento ad un’altra importante funzione dell’Estrogeno sia nel corpo maschile che femminile, vale a dire la sua capacità di promuovere uno stato mentale di vigilanza. Data la larga disponibilità di potenti inibitori dell’aromatasi di terza generazione, i Bodybuilder odierni raggiungono (a volte) un livello di soppressione degli Estrogeni molto più marcata di quanto fosse stato possibile in passato. Spesso associati a questa marcata soppressione si manifestano stati di stanchezza. In tali condizioni, l’atleta, anche se sta seguendo un ciclo correttamente formulato, potrebbe non essere in grado di massimizzare i propri risultati di miglioramento della condizione fisica a causa di un’incapacità di allenarsi con pieno vigore. Questo effetto è talvolta anche soprannominato “letargia steroidea”. La ragione principale per cui ciò accade è legata all’importante azione di supporto all’attività della Serotonina data dall’Estrogeno. La Serotonina è uno dei principali neurotrasmettitori del corpo, di essenziale importanza per un adeguata lucidità mentale e un regolare ciclo sonno / veglia. (77) (78)L’alterazione di questo neurotrasmettitore è associata anche alla sindrome da affaticamento cronico, (79) (80) e ciò ci fa comprendere quanto possa essere incisiva in particolare per la stanchezza. L’abbassamento dei livelli estrogenici nella menopausa è stata associata anche alla stanchezza, (81) così come l’uso clinico di inibitori dell’aromatasi più recenti (e più potenti) come l’Anastrozolo, (82) il Letrozolo, (83) l’Exemestane, (84) e il Fadrozolo (85) in alcuni pazienti. Questo effetto deve essere preso in importante considerazione quando si pianifica un ciclo. Anche se non tutti notano questo problema quando il livello estrogenico è basso, per coloro i quali lo subiscono, l’aggiunta di un po’ di Testosterone o di altro AAS soggetto all’aromatizzazione (oltre che l’abbassamento del composto AI) può aiutare a correggere di molto questo sintomo. È anche da notare che l’uso di AAS non aromatizzabili a volte causa questo effetto, probabilmente dovuto alla soppressione della produzione naturale di Testosterone (abbassando la disponibilità del principale substrato usato dal corpo maschile per sintetizzare Estradiolo).
Methylestradiolo
*Nota:gli AAS metilati in C-17, soggetti ad aromatizzazione (vedi Methyltestosterone e Mathandrostenolone), convertono nel potente Methylestradiolo, o 17α-methylestradiolo (17α-ME). A causa della presenza del gruppo 17α-metile, il Methylestradiolo non può essere disattivato per ossidazione del gruppo 17β-idrossile, con conseguente migliorata stabilità metabolica e potenza di legame recettoriale (e conseguente attività estrogenica) rispetto all’Estradiolo. (86)(87)(88)
Quindi, come gestire il fattore estrogenico durante la preparazione?
Dave Bautista mostra un classico esempio di cattiva gestione estrogenica.
Arrivati a questo punto, quale significato ha quanto è stato trattato per un Bodybuilder che cerca di ottenere una forma ottimale? Fondamentalmente penso che il punto chiave sia quello di mantenere un approccio prudente all’uso di farmaci per il controllo estrogenico, un approccio che deve essere dettato dalle reali esigenze verificate attraverso appositi esami ematici. Ovviamente, è chiaro che l’approccio all’uso dei composti per il controllo estrogenico debba variare a seconda dell’obbiettivo (vale a dire se l’obbiettivo primario è l’aumento della massa muscolare o se è l’aumento della definizione muscolare). Ripeto, è ovvio che l’utilizzo di composti volti al controllo estrogenico è una chiara necessità qualora si manifestassero effetti collaterali estrogenici, come lo è, nel caso, il calo dei composti soggetti all’azione dell’enzima aromatasi, ma gli esami ematici sono sempre di primaria importanza ai fini di una gestione ottimale del problema. La ginecomastia è certamente un problema indesiderato per l’utilizzatore di AAS, come lo è la ritenzione idrica ed un eccessivo accumulo di massa grassa. Ma se tali problemi non emergono, o se dagli esami ematici non emerge la reale necessità, l’aggiunta di composti per il controllo estrogenico (che siano AI, SERM o inibitori della biosintesi steroidea) può inficiare i risultati ottenibili in un contesto “Bulk”.
Quindi, concludendo, in un contesto “Bulk”, al fine di mantenere i benefici estrogenici senza incappare negli effetti collaterali estrogeno-dipendenti, la soglia ematica di Estradiolo dovrebbe mantenersi all’interno del range medio di riferimento; ciò significa che, in teoria, non dovrebbe ne scendere al di sotto dei 30pg/pg ne salire al di sopra dei 60 pg/ml. In un contesto “Cut” o “Pre-Gara”, ovviamente, le cose cambiano. In tali contesti, considerando gli effetti estrogenici anche sullo spessore della pelle, la cosa migliore sarebbe optare per il mantenimento di un livello di estrogeni basso-normale (20-30pg/ml) per tutta la fase “Cut” della preparazione per poi, se si è in preparazione alla gara, calare i livelli a 10pg o meno nell’ultimo paio di settimane prima del contest.
Per le atlete il discorso è simile anche se diverso sotto alcuni aspetti. In ambito femminile il controllo dei livelli e dell’attività estrogenica è di importanza anche maggiore per il raggiungimento di una forma da gara ottimale (vedere ad esempio “Nolvadex (Tamoxifene Citrato)”e “RALOXIFENE E COMPOSIZIONE CORPOREA NELLE DONNE”).
In definitiva, come logica conclusione, il metodo d’approccio per la gestione estrogenica deve basarsi sulle reali necessità, su dati oggettivi, e non su un fallace e molto poco scientifico “passa parola” dettato dall'”esperienza” di qual si voglia agonista o ex tale…
2- Ryan KJ (August 1982). “Biochemistry of aromatase: significance to female reproductive physiology”. Cancer Res. 42 (8 Suppl): 3342s–3344s. PMID7083198.
5- Lombardi G, Zarrilli S, Colao A, Paesano L, Di Somma C, Rossi F, De Rosa M (2001). “Estrogens and health in males”. Molecular and Cellular Endocrinology. 178 (1–2): 51–5. doi:10.1016/S0303-7207(01)00420-8. PMID11403894.
7- Soltysik K, Czekaj P (April 2013). “Membrane estrogen receptors — is it an alternative way of estrogen action?”. J. Physiol. Pharmacol. 64 (2): 129–42. PMID23756388.
12- Darabi M, Ani M, Panjehpour M, Rabbani M, Movahedian A, Zarean E (2011). “Effect of estrogen receptor β A1730G polymorphism on ABCA1 gene expression response to postmenopausal hormone replacement therapy”. Genet Test Mol Biomarkers. 15 (1–2): 11–5. doi:10.1089/gtmb.2010.0106. PMID21117950.
14- Hess RA, Bunick D, Lee KH, Bahr J, Taylor JA, Korach KS, Lubahn DB (1997). “A role for estrogens in the male reproductive system”. Nature. 390 (6659): 447–8. doi:10.1038/37352. PMID9393999.
16- Hill RA, Pompolo S, Jones ME, Simpson ER, Boon WC (2004). “Estrogen deficiency leads to apoptosis in dopaminergic neurons in the medial preoptic area and arcuate nucleus of male mice”. Mol. Cell. Neurosci. 27 (4): 466–76. doi:10.1016/j.mcn.2004.04.012. PMID15555924.
18- Nelson LR, Bulun SE (September 2001). “Estrogen production and action”. J. Am. Acad. Dermatol. 45 (3 Suppl): S116–24. doi:10.1067/mjd.2001.117432. PMID11511861.
19- MacDonald PC, Madden JD, Brenner PF, Wilson JD, Siiteri PK: Origin of estrogen in normal men and in women with testicular feminization. J Clin Endocrinol Metab 49:905-916, 1979
20- Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham-Lorence S, Amarneh B, Ito Y, Fisher CR, Michael MD, et al.: Aromatase cytochrome P450, the enzyme responsible for estrogen biosyn- thesis. Endocr Rev 15:342-355, 1994
21- Simpson ER, Zhao Y, Agarwal VR, Michael MD, Bulun SE, Hinshelwood MM, Graham-Lorence S, Sun T, Fisher CR, Qin K, Mendelson CR: Aromatase expression in health and disease. Recent Prog Horm Res 52:185-213; discus- sion 213-184, 1997
22- Simpson ER: Aromatase: biologic relevance of tissue-specific expression. Semin Reprod Med 22:11-23, 2004
23- Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, Evans RM: The nuclear recep- tor superfamily: the second decade. Cell 83:835-839, 1995
24- Green S, Walter P, Kumar V, Krust A, Bornert JM, Argos P, Chambon P: Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature 320:134-139, 1986
25- Greene GL, Gilna P, Waterfield M, Baker A, Hort Y, Shine J: Sequence and expression of human estrogen receptor complementary DNA. Science 231:1150-1154, 1986
26- Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson JA: Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 138:863-870, 1997
27- Enmark E, Gustafsson JA: Oestrogen receptors – an overview. J Intern Med 246:133-138, 1999
28- Levin ER: Cellular Functions of the Plasma Membrane Estrogen Receptor. Trends Endocrinol Metab 10:374-377, 1999
29- Pedram A, Razandi M, Levin ER: Nature of functional estrogen receptors at the plasma membrane. Mol Endocrinol 20:1996-2009, 2006
30- Filardo E, Quinn J, Pang Y, Graeber C, Shaw S, Dong J, Thomas P: Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology 148:3236-3245, 2007
31- de Ronde W, Pols HA, van Leeuwen JP, de Jong FH: The importance of oestrogens in males. Clin Endocrinol (Oxf) 58:529-542, 2003
32- Baird DT, Horton R, Longcope C, Tait JF: Steroid dynamics under steady- state conditions. Recent Prog Horm Res 25:611-664, 1969
33- Baird DT, Uno A, Melby JC: Adrenal secretion of androgens and oestrogens. J Endocrinol 45:135-136, 1969
34- Dunn JF, Nisula BC, Rodbard D: Transport of steroid hormones: bind- ing of 21 endogenous steroids to both testosterone-binding globulin and corticosteroid-binding globulin in human plasma. J Clin Endocrinol Metab 53:58-68, 1981
35- Carr BR: Disorders of the ovaries and female reproductive tract. Williams Textbook of Endocrinology. 11th ed. W.B. Saunders Company ed. Polonsky KS, Melmed S, Kronenberg HM, Larsen PR, Eds. Philadelphia, 2008
36- Weinbauer GF, Nieschlag E: The role of testosterone in spermatogene- sis. Springer-Verlag ed. Nieschlag E, Behre H, Eds. Berlin Heidelberg, Testosterone action deficiency substitution, 1990, p. 23-50
37- Zondek B: Mass excretion of oestrogenic hormone in the urine of the stallion. Nature 33:209-210, 1934
38- Heard RD, Jellinck PH, O’Donnell VJ: Biogenesis of the estrogens: the con- version of testosterone-4-C14 to estrone in the pregnant mare. Endocrinology 57:200-204, 1955.
39- West CD, Damast BL, Sarro SD, Pearson OH: Conversion of testosterone to estrogens in castrated, adrenalectomized human females. J Biol Chem 218:409-418, 1956.
40- Davis JW, Gut M, Lemon HM, Wotiz HH: Studies in steroid metabolism. V. The conversion of testosterone-4-C14 to estrogens by human ovarian tissue. J Biol Chem 222:487-495, 1956
41- Baggett B, Dorfman RI, Engel LL, Savard K: The conversion of testoster- one-3-C14 to C14-estradiol-17beta by human ovarian tissue. J Biol Chem 221:931-941, 1956 8. Baggett B, Engel LL, Balderas L, Lanman G: Conversion of C14-testosterone to C14-estrogenic steroids by endocrine tissues. Endocrinology 64:600-608, 1959
42- Smith EP, Boyd J, Frank GR, Takahashi H, Cohen RM, Specker B, Williams TC, Lubahn DB, Korach KS: Estrogen resistance caused by a mutation in the estrogen-receptor gene in a man. N Engl J Med 331:1056-1061, 1994
43- Morishima A, Grumbach MM, Simpson ER, Fisher C, Qin K: Aromatase deficiency in male and female siblings caused by a novel mutation and the physiological role of estrogens. J Clin Endocrinol Metab 80:3689-3698, 1995
44- Carani C, Qin K, Simoni M, Faustini-Fustini M, Serpente S, Boyd J, Korach KS, Simpson ER: Effect of testosterone and estradiol in a man with aromata- se deficiency. N Engl J Med 337:91-95, 1997
45- Bilezikian JP, Morishima A, Bell J, Grumbach MM: Increased bone mass as a result of estrogen therapy in a man with aromatase deficiency. N Engl J Med 339:599-603, 1998
46- Herrmann BL, Saller B, Janssen OE, Gocke P, Bockisch A, Sperling H, Mann K, Broecker M: Impact of estrogen replacement therapy in a male with con- genital aromatase deficiency caused by a novel mutation in the CYP19 gene. J Clin Endocrinol Metab 87:5476-5484, 2002
47- Pura M, Mittre H, Carreau S, Kottler ML: Clinical findings in an adult man with a novel mutation in the aromatase gene. In Program of the 85th Annual Meeting of the Endocrine Society, Abstract Book Philadelphia, PA, 2003
48- Maffei L, Murata Y, Rochira V, Tubert G, Aranda C, Vazquez M, Clyne CD, Davis S, Simpson ER, Carani C: Dysmetabolic syndrome in a man with a novel mutation of the aromatase gene: effects of testosterone, alendronate, and estradiol treatment. J Clin Endocrinol Metab 89:61-70, 2004
49- Maffei L, Rochira V, Zirilli L, Antunez P, Aranda C, Fabre B, Simone ML, Pignatti E, Simpson ER, Houssami S, Clyne CD, Carani C: A novel com- pound heterozygous mutation of the aromatase gene in an adult man: reinforced evidence on the relationship between congenital oestrogen deficiency, adiposity and the metabolic syndrome. Clin Endocrinol (Oxf) 67:218-224, 2007
50- Luberto A, Rochira V, Zirilli L, Pignatti E, Simpson ER, Maffei L, Carani C: A novel compound heterozygous mutation of the aromatase gene in an adult man: a reinforced estrogen deficincy, adiposity and the metabolic syndrome. J Endocrinol Invest 30 Suppl 4:50, 2007
51- Bouillon R, Bex M, Vanderschueren D, Boonen S: Estrogens are essen- tial for male pubertal periosteal bone expansion. J Clin Endocrinol Metab 89:6025-6029, 2004
52- Vanderschueren D, Boonen S, Bouillon R: Action of androgens versus estrogens in male skeletal homeostasis. Bone 23:391-394, 1998
53- Vanderschueren D, Venken K, Ophoff J, Bouillon R, Boonen S: Clinical Review: Sex steroids and the periosteum–reconsidering the roles of androgens and estrogens in periosteal expansion. J Clin Endocrinol Metab 91:378-382, 2006
54- Horsman A, Gallagher JC, Simpson M, Nordin BE: Prospective trial of oestrogen and calcium in postmenopausal women. Br Med J 2:789-792, 1977
55- Lindsay R, Hart DM, Forrest C, Baird C: Prevention of spinal osteoporosis in oophorectomised women. Lancet 2:1151-1154, 1980
56- Christiansen C, Christensen MS, Transbol I: Bone mass in postmenopausal women after withdrawal of oestrogen/gestagen replacement therapy. Lancet 1:459-461, 1981
57- Finkelstein JS, Neer RM, Biller BM, Crawford JD, Klibanski A: Osteopenia in men with a history of delayed puberty. N Engl J Med 326:600-604, 1992
58- Speroff L, Rowan J, Symons J, Genant H, Wilborn W: The comparative effect on bone density, endometrium, and lipids of continuous hormones as replacement therapy (CHART study). A randomized controlled trial. Jama 276:1397-1403, 1996
59- Wang C, Eyre DR, Clark R, Kleinberg D, Newman C, Iranmanesh A, Veldhuis J, Dudley RE, Berman N, Davidson T, Barstow TJ, Sinow R, Alexander G, Swerdloff RS: Sublingual testosterone replacement improves muscle mass and strength, decreases bone resorption, and increases bone formation markers in hypogonadal men–a clinical research center study. J Clin Endocrinol Metab 81:3654-3662, 1996
60- Faustini-Fustini M, Rochira V, Carani C: Oestrogen deficiency in men: where are we today? Eur J Endocrinol 140:111-129, 1999
61- Vanderschueren D, Venken K, Ophoff J, Bouillon R, Boonen S: Clinical Review: Sex steroids and the periosteum–reconsidering the roles of androgens and estrogens in periosteal expansion. J Clin Endocrinol Metab 91:378-382, 2006
69- Pentose Cycle Activity in Muscle from Fetal, Neonatal and Infant Rhesus Monkeys. Arch Biochem Biophys 117:275-81 1966
70- The pentose phosphate pathway in regenerating skeletal muscle. Biochem J 170: 17 1978
71- Aromatization of androgens to estrogens mediates increased activity of glucose 6-phosphate dehydrogenase in rat levator ani muscle. Endocrinol 106(2):440-43 1980
72- Influence of tamoxifen, aminoglutethimide and goserelin on human plasma IGF-1 levels in breast cancer patients. J steroid Biochem Mol Bio 41:541- 3,1992
73- Activation of the somatotropic axis by testosterone in adult males: Evidence for the role of aromatization. J Clin. Endocrinol Metab 76:1407-12 1993
74- Testosterone administration increases insulin-like growth factor-I levels in normal men. J Clin Endocrinol Metab 77(3):776-9 1993
80- Association between serotonin transporter gene polymorphism and chronic fatigue syndrome. Narita M et al. Biochem Biophys Res Commun 2003 Nov 14;311(2)264-6
81- Premenstrual Syndrome. Dickerson LM et al. Am Fam Physician 2003 Apr 15;67(8):1743-52
82- Phase II trial of anastrozole in women with asymptomatic mullerian cancer. Gynecol Oncol. 2003 Dec;91(3):596-602.
83- Letrozole. A review of its use in postmenopausal women with advanced breast cancer. Drugs. 1998 Dec;56(6):1125-40. Review.
84- Exemestane: a review of its clinical efficacy and safety. Breast. 2001 Jun;10(3):198-208.
85- A study of fadrozole, a new aromatase inhibitor, in postmenopausal women with advanced metastatic breast cancer. J Clin Oncol. 1992 Jan;10(1):111-6.
87- Feenstra A, Vaalburg W, Nolten GM, Reiffers S, Talma AG, Wiegman T, van der Molen HD, Woldring MG (1983). “Estrogen receptor binding radiopharmaceuticals: II. Tissue distribution of 17 alpha-methylestradiol in normal and tumor-bearing rats”. J. Nucl. Med. 24 (6): 522–8. PMID6406650.