
Estrogeni, questi sconosciuti…
La questione degli Estrogeni nell’ambito delle preparazioni comprendenti farmaci per il miglioramento delle prestazioni, in special modo nel Bodybuilding, è generalmente mal compresa e, di conseguenza, mal gestita. Complice di questa “mala gestione” è l’ignoranza sia specifica, cioè portata da una scarsa disponibilità di testi di facile accesso (e comprensione) che trattano l’argomento in modo esaustivo, sia indotta dal comportamento della maggior parte dei preparatori o degli atleti “solisti” che ignorano totalmente la necessità di un aggiornamento continuo in tutte le componenti della preparazione (allenamento, nutrizione e supplementazione). Con questa pubblicazione è mia intenzione fornire una buona base di conoscenza sugli Estrogeni (loro biosintesi, azioni ecc…) e sulla migliore gestione di questi nel contesto di una preparazione.
Iniziamo per gradi…
Estrogeni e loro caratteristiche/azioni principali (nella donna e nell’uomo)

Gli Estrogeni sono i principali ormoni sessuali femminili. Si tratta di ormoni steroidei responsabili dello sviluppo e della regolazione del sistema riproduttivo femminile e delle caratteristiche sessuali secondarie. Con il termine estrogeno ci si può anche riferire a qualsiasi sostanza, naturale o sintetica, che mima gli effetti dell’ormone naturale.(1) L’estrogeno prevalente in quantità e potenza è l’Estradiolo, anche se diversi metaboliti dell’estradiolo hanno anche una marcata attività ormonale estrogenica. Versioni sintetiche degli estrogeni vengono utilizzate come principio attivo di farmaci contraccettivi orali, nella terapia ormonale sostitutiva per le donne in postmenopausa, nelle donne ipogonadiche e nei transgender, oltre che nel trattamento di alcuni tumori ormone-sensibili come il cancro alla prostata e il cancro al seno. Gli Estrogeni sono una delle tre classi di ormoni sessuali, insieme agli Steroidi Androgeni/Anabolizzanti come il Testosterone e i progestinici come il Progesterone.
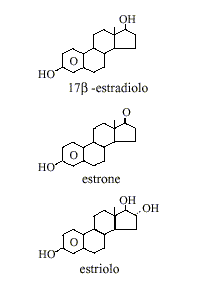
Gli Estrogeni sono sintetizzati in tutti i vertebrati (2) così come in alcuni insetti.(3) Le tre principali forme di estrogeno presenti naturalmente nelle donne sono l’Estrone (E1), l’Estradiolo (E2) e l’Estriolo (E3). Un altro tipo di Estrogeno chiamato Estetrolo (E4) viene prodotto solo durante la gravidanza. Quantitativamente, gli Estrogeni circolano a livelli inferiori rispetto agli Androgeni sia negli uomini che nelle donne.(4) Mentre i livelli di Estrogeni sono significativamente più bassi nei maschi rispetto alle femmine, gli Estrogeni, tuttavia, hanno anche importanti ruoli fisiologici nei soggetti di sesso maschile.(5)
Come tutti gli ormoni steroidei, gli Estrogeni penetrano facilmente attraverso la membrana cellulare. Una volta all’interno della cellula, si legano e attivano i recettori degli estrogeni (ER) che a loro volta modulano l’espressione di molti geni.(6) Inoltre, gli Estrogeni si legano e attivano rapidamente i recettori degli estrogeni di membrana (mER), (7)(8) come i GPER (GPR30).(9)
Come già precedentemente accennato, i tre principali Estrogeni presenti naturalmente nelle donne sono l’Estrone (E1), l’Estradiolo (E2) e l’Estriolo (E3). L’Estradiolo è l’estrogeno predominante negli anni riproduttivi sia in termini assoluti nel siero che in termini di attività estrogenica. Durante la menopausa, l’Estrone è l’Estrogeno circolante predominante mentre durante la gravidanza è l’Estriolo ad essere l’Estrogeno circolante predominante in termini di livelli sierici. Anche se l’Estriolo è il più abbondante dei tre estrogeni è anche il più debole, mentre l’Estradiolo è il più forte con una potenza di circa 80 volte quella dell’Estriolo.(10) Quindi, l’Estradiolo è l’Estrogeno più importante nelle donne non in gravidanza che si trovano tra le fasi del menarca e della menopausa.

Tuttavia, durante la gravidanza questo ruolo passa all’Estriolo e nelle donne in postmenopausa l’Estrone diventa la forma primaria di Estrogeno nel corpo. Come già accennato, un altro tipo di Estrogeno chiamato Estetrolo (E4) viene prodotto solo durante la gravidanza. Oltre che dai follicoli ovarici, tutte le diverse forme di estrogeni sono sintetizzate a partire dagli Androgeni, in particolare dal Testosterone e dal Androstenedione, da parte dell’enzima aromatasi.
Altri substrati utilizzati dal corpo per la sintesi estrogenica, senza il coinvolgimento primario dell’enzima aromatasi, sono il 27-idrossicholesterolo, il Deidroepiandrosterone (DHEA), il 7-oxo-DHEA, il 7α-idrossi-DHEA, il 16α-idrossi-DHEA, il 7β-idrossiepiandrosterone, l’Androstenedione (A4), l’Androstenediolo (A5), il 3α-androstanediolo e il 3β-androstanediolo. Alcuni metaboliti estrogenici, come i catecol-estrogeni 2-idrossiestradiolo, 2-idrossestrone, 4-idrossiestradiolo e 4-idrossestrone, nonché il 16α-idrossiestrone, sono composti estrogenici con vari gradi di attività. L’importanza biologica di questi estrogeni minori non è del tutto chiara. Tutti questi composti possono avere importanti funzioni estrogeniche nel corpo.


Le azioni degli Estrogeni sono mediate dal Recettore Estrogeno (ER), una proteina nucleare dimerica che si lega al DNA e controlla l’espressione genica. Come altri ormoni steroidei, l’Estrogeno entra passivamente nella cellula nella quale si lega e attiva il Recettore dell’Estrogeno. Il complesso ER si lega a specifiche sequenze di DNA chiamate “elemento di risposta ormonale” per attivare la trascrizione di geni bersaglio: in uno studio nel quale si è utilizzate una linea di cellule del cancro al seno estrogeno-dipendente come modello, sono stati identificati 89 di questi geni.(11) Poiché l’Estrogeno entra in tutte le cellule, le sue azioni dipendono dalla presenza del ER nella cellula. L’ER è espresso in tessuti specifici tra cui l’ovaio, l’utero e il seno. Gli effetti metabolici degli Estrogeni nelle donne in postmenopausa sono stati legati al polimorfismo genetico dell’ER. (12)
Mentre gli Estrogeni sono presenti sia negli uomini che nelle donne, sono solitamente presenti a livelli significativamente più alti nelle donne in età riproduttiva. Essi promuovono lo sviluppo delle caratteristiche sessuali secondarie femminili, come i seni, e sono anche coinvolti nell’ispessimento dell’endometrio e altri aspetti della regolazione del ciclo mestruale. Nei maschi, l’Estrogeno regola alcune funzioni del sistema riproduttivo importanti per la maturazione dello sperma (13)(14)(15) e sembra essere necessario per una sana libido.(16) Inoltre, vi sono molti altri cambiamenti strutturali indotti dall’Estrogeno oltre ad altre funzioni.
Gli Estrogeni, nelle femmine, sono prodotti principalmente dalle ovaie e, durante la gravidanza, dalla placenta.(17) L’Ormone Follicolo Stimolante (FSH) stimola la produzione ovarica di Estrogeni da parte delle cellule della granulosa dei follicoli ovarici e dei corpi lutei. Alcuni Estrogeni vengono anche prodotti in quantità minori in altri tessuti come il fegato, le ghiandole surrenali e i seni. Queste fonti secondarie di Estrogeni sono particolarmente importanti nelle donne in postmenopausa. Anche nelle cellule adipose vengono sintetizzati Estrogeni (vedi azione dell’Enzima Aromatasi)(18)

Nelle femmine, la sintesi degli Estrogeni inizia nelle cellule della teca follicolare interna nell’ovaio, dalla sintesi del Androstenedione a partire dal Colesterolo. L’Androstenedione è uno steroide con debole attività androgenica il cui scopo è prevalentemente quello di fungere da precursore di più potenti androgeni come il Testosterone così come dell’Estrogeno. Questo steroide attraversa la membrana basale nelle cellule della granulosa circostanti, dove o viene immediatamente convertito in Estrone, o in Testosterone e poi successivamente in Estradiolo. La conversione del Androstenedione in Testosterone è catalizzata dal 17β-idrossisteroide deidrogenasi (17β-HSD), mentre la conversione del Androstenedione e del Testosterone in Estrone ed Estradiolo è rispettivamente catalizzata dal aromatasi, entrambi enzimi che sono espressi nelle cellule della granulosa. Al contrario, le cellule della granulosa non dispongono di 17α-idrossilasi e 17,20-liasi, mentre le cellule della teca follicolare interna esprimono questi enzimi e il 17β-HSD, ma non l’aromatasi. Quindi sia le cellule della granulosa che le cellule della teca follicolare interna sono essenziali per la produzione di Estrogeni nelle ovaie.
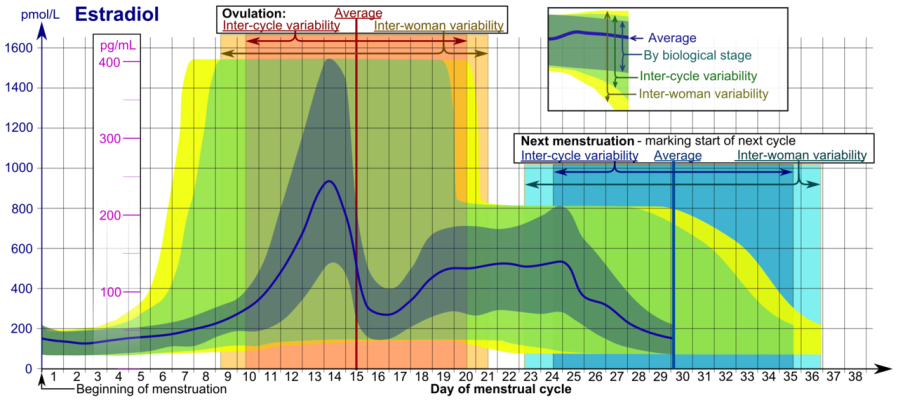
I livelli di Estrogeni variano durante il ciclo mestruale, con livelli più alti vicino alla fine della fase follicolare e poco prima dell’ovulazione.

Gli Estrogeni sono metabolizzati tramite idrossilazione da enzimi citocromo P450 quali CYP1A1 e CYP3A4 e tramite coniugazione del estrogeno sulfotransferasi (sulfato) e del UDP-glucuroniltransferasi (glucuronidazione). Inoltre, l’Estradiolo viene deidrogenato dal 17β-idrossiesteroide deidrogenasi nel molto meno potente Estrone. Queste reazioni si verificano principalmente nel fegato, ma avvengono anche in altri tessuti.
Gli estrogeni sono implicati nel rilascio di GH e IGF-1 sia nell’uomo che nella donna. Le donne hanno mediamente livelli di GH più elevati sebbene sembri che ne siano meno sensibili (vedi percentuale di grasso corporeo femminile).
Quindi, per riepilogare, gli Estrogeni nelle femmine regolano la maturazione sessuale intervenendo nello sviluppo dell’apparato genitale.
La loro massiccia secrezione in epoca puberale induce la chiusura delle cartilagini di coniugazione delle ossa lunghe, terminando di fatto, la fase di accrescimento staturale.
Gli estrogeni stimolano lo sviluppo stromale della mammella e il mantenimento delle caratteristiche femminili secondarie (crescita delle mammelle, distribuzione dei peli, voce, statura, ossatura, distribuzione del grasso).
Permettono la fecondazione e la gravidanza, intervenendo nella regolazione del ciclo mestruale.
Regolano la distribuzione del grasso corporeo, favorendone il deposito nelle anche, nelle natiche, nelle cosce e nell’addome al di sotto dell’ombelico.
Mantengono il trofismo osseo ed hanno quindi azione protettiva nei confronti dell’osteoporosi
Stimolano la sintesi di trigliceridi e l’aumento delle lipoproteine ad alta densità (HDL) proteggendo le pareti vasali dal danno arteriosclerotico.
Stimolano la lipolisi nel tessuto muscolare ed adiposo. Per questo motivo gli estrogeni migliorano la prestazione degli sport di durata risparmiando il glicogeno muscolare a scapito degli acidi grassi
Regolano molte funzioni cerebrali fra cui l’attenzione e la memoria.
Stimolano la sintesi epatica di numerosi enzimi e proteine (SHBG, angiotensinogeno).
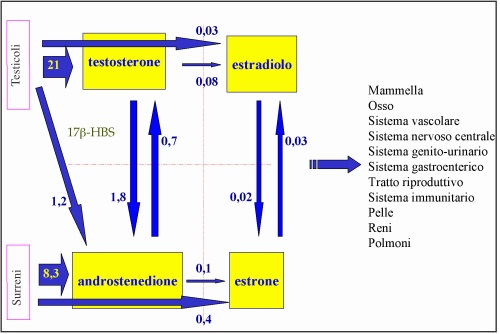
Nell’uomo, invece, gli estrogeni circolanti derivano per la maggior parte dall’aromatizzazione degli Androgeni circolanti (19). Il complesso enzimatico conosciuto come aromatasi è responsabile dell’aromatizzazione dell’anello A degli Androgeni che comporta la trasformazione in sostanze ad attività estrogenica. (20) (21) (22)

La trascrizione del gene per l’aromatasi e l’espressione dell’enzima aromatasi avviene in un ampio numero di tessuti, come i testicoli, il tessuto adiposo, il muscolo, il fegato, il cervello, i follicoli piliferi, i fibroblasti etc. (20). L’azione degli Estrogeni, come già accennato, si esplica attraverso il loro legame a recettori specifici, che appartengono alla ‘superfamiglia’ dei recettori nucleari (23). Finora sono stati caratterizzati due tipi di recettori per gli Estrogeni (ER): l’ER classico, ora denominato ERα (24) (25) (26), ed un altro sottotipo denominato ERβ (27). Entrambi sono recettori intranucleari che, una volta attivati, modulano a livello genomico l’attività di trascrizione genica. Di recente è stata documentata anche una via non-genomica dell’azione degli Estrogeni, che prevede un’interazione plasma-membrana del recettore estrogenico (28) (29) (30).

L’Estradiolo (E2) nel maschio è sintetizzato dal Testosterone attraverso l’enzima aromatasi o dall’Estrone attraverso la 17β-idrossisteroido deidrogenasi (17β-HSD) (31). L’E2 prodotto giornalmente nel maschio è stimato essere circa 35-45 μg (0.130- 0.165 μmol), dei quali il 15-20% è direttamente prodotto dal testicolo (32) (19), un 60% è prodotto dall’aromatizzazione periferica del Testosterone circolante ed il restante 20% dalla conversione periferica dell’Estrone. L’Estrone (E1) può anche essere direttamente secreto dai surreni (33) o derivare dall’aromatizzazione periferica dell’Androstenedione, quest’ultimo prodotto in parte dal surrene ed in parte derivante dalla conversione periferica del Testosterone. I testicoli comunque contribuiscono maggiormente, rispetto ai surreni, alla totale produzione di Estradiolo circolante e la via enzimatica di sintesi maggiormente coinvolta nella secrezione di Estrogeni nel maschio è quella aromatasica, mentre la via della 17β-HSD ha una minore rilevanza “in vivo”. Solo il 2-3% dell’Estradiolo circolante è libero, la maggior parte dell’ormone è legato all’albumina o alla globulina legante gli ormoni sessuali (SHBG) (34) e la frazione non legata alla SHBG è considerata bioattiva a livello dei tessuti bersaglio.

I progressi negli studi effettuati negli ultimi anni hanno infatti dimostrato in maniera inequivocabile il ruolo fisiologico che gli Estrogeni hanno anche nell’uomo. Gli Estrogeni svolgono un ruolo chiave sul sistema immunitario, a livello del timo, agiscono a livello ipofisario e cardiovascolare, osseo e del metabolismo glucidico e lipidico.

Un eccesso estrogenico nella donna favorisce l’accumulo di tessuto adiposo e la comparsa di ritenzione idrica, oltre ad esporre la donna ad un elevato rischio di sviluppare alcune forme di cancro come quello alla mammella, l’insulinoresistenza, l’infertilità e l’ovaiopolicistico. Nell’uomo l’eccesso estrogenico è legato ad effetti collaterali spiacevoli tra i quali troviamo la ginecomastia, la diminuzione del desiderio sessuale, problemi di erezione (che si verifica anche con un calo estrogenico eccessivo), e diminuzione della fertilità.
Ricordiamoci però che, tanto nella donna come nell’uomo, un livello estremamente basso di Estrogeni comporta diversi problemi tra i quali emergono: elevati livelli sierici di Trigliceridi, LDL e colesterolo totale con ridotti livelli di HDL (35)(36)(37)(38)(39), ridotti livelli di LDL, HDL e colesterolo totale con Trigliceridi normali (40)(41), osteoporosi, dolore osseo, e diabete mellito di tipo II (37).
Si noti che la descrizione di pazienti maschi affetti da deficit congenito di Estrogeni (42) (43) (44) (45) (46) (47) (48) (49) (50) (51) ha fornito un modello in vivo unico che ha permesso di stabilire che la completa ossificazione delle cartilagini epifisarie, anche nel maschio, non può avvenire senza l’azione degli Estrogeni. Inoltre, è stato dimostrato che gli Androgeni da soli non sono sufficienti per garantire una normale mineralizzazione dello scheletro in età adulta, dal momento che un quadro di osteopenia e/o osteoporosi è stato documentato in tutti i pazienti con deficit congenito di Estrogeni (42) (43) (44) (45) (46) (47) (48) (49) (50) (51). Ad oggi è stato chiarito che gli effetti degli steroidi sessuali sul processo di mineralizzazione ossea nel maschio sono solo in parte da ascrivere agli Androgeni di origine testicolare, mentre l’azione degli Estrogeni, che derivano dall’aromatizzazione degli androgeni stessi, sembra giocare un ruolo di maggiore importanza (52) (53). Infatti in letteratura vi sono evidenze riguardo l’associazione tra ipogonadismo e riduzione della massa ossea in entrambi i sessi (54) (55) (56) (57) (58) (59). Inoltre, il concetto generale che gli Androgeni mantengono la massa ossea nell’uomo, così come gli Estrogeni la mantengono nella donna, continua ad essere il punto di vista prevalente tra i medici, sebbene recenti studi suggeriscano il contrario (42) (43) (44) (45) (60) (61). Nel 1997, infatti, è stata descritta l’efficacia della terapia Estrogenica nell’ottenere sia la completa ossificazione delle epifisi che l’aumento di densità minerale ossea (BMD) in un paziente maschio affetto da deficit congenito di aromatasi (44).
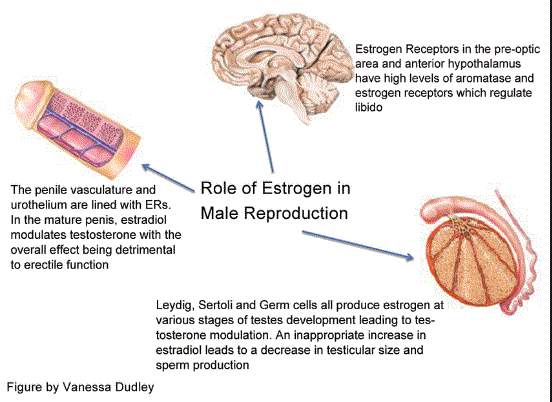
Una funzione interessante dell’Estradiolo nel maschio è rappresentata dal suo impatto sulla funzione sessuale. Come già accennato, l‘Estradiolo negli uomini è essenziale per modulare la libido, la funzione erettile e la spermatogenesi. I recettori degli Estrogeni, così come l’aromatasi, sono abbondanti nel cervello, nel pene e nei testicoli, organi importanti per la funzione sessuale. Nel cervello, la sintesi dell’Estradiolo è elevata nelle aree correlate all’auda sessuale. Inoltre, nel pene, i recettori degli Estrogeni si trovano in tutto il corpus cavernosum con elevata concentrazione intorno ai fasci neurovascolari. Un livello basso di Testosterone e elevato di Estrogeni aumenta l’incidenza della disfunzione erettile indipendentemente l’uno dall’altro. Nei testicoli, la spermatogenesi è modulata a tutti i livelli dagli Estrogeni, a partire dall’asse ipotalamo-ipofisi-gonadi, seguita dalle cellule di Leydig, Sertoli e germinali e terminando con l’epitelio ductale, l’epididimo e lo sperma maturo. La regolazione delle cellule testicolari mediante l’Estradiolo mostra sia un’influenza inibitoria che una stimolatoria, indicando un intricata sinfonia di modulazione dose-dipendente e temporalmente sensibile.(62)
Adesso che abbiamo una buona conoscenza di base sugli Estrogeni e le loro azioni sia nell’uomo che nella donna, possiamo passare a trattare la questione della gestione funzionale degli Estrogeni durante la preparazione.
AAS aromatizzabili e gestione estrogenica

Come ormai ben sappiamo, il Testosterone è il substrato primario usato nell’uomo per la sintesi dell’Estradiolo, principale ormone sessuale femminile. Con una leggera alterazione strutturale del Testosterone dovuta all’azione dell’enzima aromatasi, l’Estradiolo viene prodotto nel maschio. L’attività dell’enzima aromatasi nel maschio si verifica in diverse regioni del corpo , compreso il tessuto adiposo, il fegato, le gonadi, il Sistema Nervoso Centrale e il tessuto muscolo scheletrico.(63) (64) (65) (66) (67) In un soggetto sano di sesso maschile , la quantità di Estrogeni prodotta non è generalmente molto significativa ed è basata sulle richieste e caratteristiche fisiologiche divenendo anche un vantaggio, come precedentemente visto, nella qualità e quantità dei valori di colesterolo e nella mineralizzazione ossea. Nei casi in cui le concentrazioni di Estrogeni (in particolare dell’Estradiolo) raggiungono livelli ematici elevati, che ciò avvenga come conseguenza non patologica (vedi obesità e cattiva alimentazione) o patologica (vedi neoplasia al testicolo), o come conseguenza del uso di AAS aromatizzabili e dell’alterazione omeostatica dovuta all’uso di tali composti, il soggetto sperimenta effetti avversi tra i quali vi sono la comparsa di ritenzione idrica, l’accumulo adiposo con modello femminile e la ginecomastia. Per questo motivo, molti atleti si concentrano sulla riduzione dell’incremento e/o dell’attività degli estrogeni nel corpo con l’uso di inibitori dell’aromatasi (es. Anastrozolo, Letrozolo e Exemestane), di Modulatori Selettivi del Recettore degli Estrogeni (SERM) (es. Tamoxifene e Raloxifene) o di inibitori della biosintesi steroidea (es. Aminoglutettimide e Trylostano), in specie quando il ciclo che stanno seguendo è composto da molecole altamente aromatizzabili (es. Testosterone e Methandrostenolone) o quando l’atleta desidera avere un maggior controllo estrogenico in un contesto nel quale si cerca di aumentare la definizione muscolare.

Nota: Con l’uso del Oxymetholone, il quale possiede attività estrogenica intrinseca, la gestione del fattore estrogenico (in particolare con la presenza di substrati soggetti all’aromatizzazione) assume caratteristiche leggermente più “complesse”. L’Oxymetholone è infatti un AAS avente azione altamente estrogenica. Tale azione, per l’appunto, non è dipendente dall’aromatizzazione del composto in estrogeno – essendo un derivato del DHT non può essere aromatizzato. L’uso di inibitori dell’aromatasi come l’Anastrozolo o l’Exemestane non influiscone sulla estrogenicità relativa di questo AAS. Alcuni hanno suggerito che l’alto livello di attività estrogenica del Oxymetholone sia in realtà dovuto alla presunta azione progestinica del composto, simile al Nandrolone. Gli effetti collaterali estrogenici e progestinici possono essere molto simili (anche perché l’azione dei due steroidi è co-attiva), il che ha reso plausibile questa ipotesi. Tuttavia, esiste uno studio nel quale è stata esaminata l’attività progestinica del Oxymetholone. Lo studio in questione stabilì che non vi era alcuna attività progestinica derivante dal composto.(68) Con tali risultati, sembra plausibile che l’Oxymetholone possa attivare il recettore degli estrogeni similmente a, ma più profondamente, dell’androgeno estrogenico Methandriolo. In definitiva, l’attività estrogenica di questo composto può essere gestita solamente con l’uso di SERM come il Tamoxifene o il Raloxifene dal momento che, per i motivi sopra detti, composti anti aromatasi e gli inibitori della biosintesi steroidea non hanno alcuna incidenza sull’attività estrogenica di questo AAS.
Anche alla luce di quanto accennato in precedenza, non dobbiamo tuttavia essere indotti a pensare che gli Estrogeni non diano nessun beneficio prestativo. Livelli di Estradiolo mantenuti nei limiti del range di riferimento offrono diversi vantaggi.
Gli atleti sono da anni a conoscenza del fatto che gli AAS aromatizzabili presentano un vantaggio nell’anabolismo muscolare, ma è solo di recente che siamo arrivati finalmente a capire i meccanismi che stanno alla base di tale effetto. Sembra che i motivi vadano oltre l’aumento di peso e di forza che si attribuiscono alla ritenzione idrica data dall’attività estrogenica, comprendendo che questo ormone (Estradiolo) possiede effettivamente un effetto diretto sui processi anabolici. Ciò si manifesta attraverso l’aumento dell’utilizzo del Glucosio, la secrezione dell’Ormone della Crescita (e di IGF-1) e la proliferazione dei recettori degli Androgeni.
- Estrogeni e utilizzo del Glucosio

Gli Estrogeni possono giocare un ruolo molto importante nella promozione di uno stato anabolico influenzando l’utilizzo del Glucosio nel tessuto muscolare. Ciò avviene attraverso un’alterazione del livello di glucosio 6-fosfato deidrogenasi disponibile, un enzima direttamente legato all’uso del glucosio per la crescita e il recupero del tessuto muscolare.(69) (70) Più specificamente, il G6PD è l’enzima che catalizza la prima reazione della via dei pentoso fosfati (definita anche Shunt dell’Esosomonofosfato [HMP shunt] o PPP da Pentose phosphate pathway), un processo metabolico citoplasmatico, parallelo alla glicolisi, in grado di generare NADPH e zuccheri pentosi (a 5 atomi di carbonio). Durante il periodo di rigenerazione tissutale seguente il danno muscolare, i livelli di G6PD aumentano considerevolmente, il che è ritenuto essere un meccanismo che il corpo attua per migliorare il recupero quando necessario. Sorprendentemente, scopriamo che l’Estrogeno è direttamente legato al livello di G6PD che deve essere messo a disposizione delle cellule in questa fase di recupero. In sintesi, gli Estrogeni svolgono anche una azione metabolica accelerando la sintesi degli acidi nucleici, delle proteine e del glicogeno.
Il legame tra Estrogeno e G6PD è stato stabilito attraverso uno studio nel quale è stato dimostrato che i livelli di questo enzima deidrogenasi aumentano dopo la somministrazione di Testosterone Propionato. Lo studio ha inoltre mostrato che era l’aromatizzazione del Testosterone in Estradiolo ad essere direttamente responsabile di questo aumento e non l’azione Androgena del AAS.(71) In questo studio sono stati presi in esame anche due composti non-aromatizzabili, il Dihydrotestosterone e il Fluoxymesterone, ma l’effetto avuto con l’uso di Testosterone Propionato non è stato replicato. Inoltre, l’effetto positivo del Testosterone Propionato è stato bloccato quando è stato aggiunto l’inibitore dell’aromatasi 4-idrossiandrostenedione (Formestano), mentre la somministrazione di 17-beta Estradiolo da solo ha causato un aumento di G6PD simile a quello osservato con il Testosterone Propionato. Il 17-alfa Estradiolo, isomero ormonalmente inattivo del 17-beta Estradiolo, che non può legarsi al recettore estrogenico, non ha avuto alcun effetto sul G6PD. Ulteriori prove sono state fatte utilizzando il Testosterone Propionato con l’antiandrogeno Flutamide dimostrando che anche quest’ultimo farmaco non ha alterato negativamente le concentrazioni di G6PD, sottolineando l’effetto indipendente dall’azione del Testosterone con il recettore androgeno.
- Estrogeni e GH/IGF-1

L’Estrogeno può anche svolgere un ruolo importante nella produzione dell’Ormone della Crescita e del IGF-1. L’IGF-1 (Fattore di Crescita Insulino-Simile) è un ormone anabolizzante rilasciato nel fegato e in vari tessuti periferici attraverso lo stimolo dell’Ormone della Crescita. L’IGF-1 è il principale responsabile dell’attività anabolica dell’Ormone della Crescita, attraverso una maggiore ritenzione di azoto / sintesi proteica e iperplasia cellulare (proliferazione). Uno dei primi studi da portare all’attenzione riguardante la questione qui trattata ha esaminato gli effetti del SERM Tamoxifene sui livelli di IGF-1, dimostrando che il composto esaminato ha un effetto soppressivo.(72) Un secondo studio, forse più degno di nota, svolto nel 1993, ha esaminato gli effetti della terapia sostitutiva del Testosterone sui soli livelli di GH e IGF-1 e li ha confrontati con gli effetti del Testosterone combinato nuovamente con il Tamoxifene. (73) Quando è stato somministrato Tamoxifene, i livelli di GH e IGF-1 hanno subito un notevole calo, mentre entrambi i valori sono stati elevati con la somministrazione di solo Testosterone Enantato. Un altro studio ha mostrato come la somministrazione settimanale di 300mg di Testosterone Enantato causi un lieve aumento del IGF-1 in uomini normali. Questi 300mg di Testosterone Enantato hanno causato un aumento dei livelli di Estradiolo, cosa normale con tale dose. Questo è stato confrontato con l’effetto portato con lo stesso dosaggio di Nandrolone Decanoato; tuttavia, questo AAS non è riuscito a produrre lo stesso aumento nei livelli di IGF-1. Questo risultato è molto interessante, specialmente quando notiamo che i livelli di Estrogeni sono stati effettivamente ridotti quando questo AAS è stato somministrato (ricordiamoci che il Nandrolone aromatizza il 20% del Testosterone). (74) Un altro studio ha dimostrato che la secrezione di GH e IGF-1 è aumentata con la somministrazione di Testosterone in maschi con pubertà ritardata, mentre il Dihydrotestosterone (non aromatizzabile) sembra sopprimere la secrezione di GH e IGF-1. (75)
- Estrogeni e Recettori degli Androgeni

È stato anche dimostrato che l’Estrogeno può aumentare la concentrazione dei recettori degli androgeni in alcuni tessuti. Ciò è stato dimostrato in studi svolti su ratti che hanno esaminato gli effetti degli Estrogeni sui Recettori degli Androgeni cellulari in animali sottoposti ad orchiectomia (rimozione dei testicoli, spesso effettuata per diminuire la produzione endogena di Androgeni). Secondo lo studio, la somministrazione di Estrogeni ha determinato un aumento del legame recettoriale nel muscolo levator ani del Metribolone pari al 480%. (76) La spiegazione per tale evento suggerita è che l’Estrogeno stimola direttamente la proliferazione dei Recettori Androgeni, o forse ne diminuisce il tasso di danneggiamento. Anche se la crescita del muscolo levator ani è comunemente usata come riferimento per determinare l’attività anabolica dei composti steroidei, esso è un muscolo dell’organo sessuale ed è diverso dal tessuto muscolo scheletrico in quanto possiede una concentrazione molto più alta di Recettori Androgeni. Questo studio, tuttavia, ha esaminato l’effetto sui AR dato dagli Estrogeni nei tessuti musco scheletrici veloci (tibialis anterior e extensor digitorum longus), ma senza notare lo stesso aumento visto nel levator ani. Anche se scoraggiante come notizia, il fatto che l’Estrogeno possa aumentare il legame del Recettore degli Androgeni in qualsiasi tessuto rimane una scoperta estremamente significativa, soprattutto alla luce del fatto che ora sappiamo che gli Androgeni hanno alcuni effetti positivi sulla crescita muscolare mediati al di fuori del tessuto muscolare.
- Estrogeni e fatica/stanchezza

La così detta “Steroid Fatigue” è una frase comunemente utilizzata in riferimento ad un’altra importante funzione dell’Estrogeno sia nel corpo maschile che femminile, vale a dire la sua capacità di promuovere uno stato mentale di vigilanza. Data la larga disponibilità di potenti inibitori dell’aromatasi di terza generazione, i Bodybuilder odierni raggiungono (a volte) un livello di soppressione degli Estrogeni molto più marcata di quanto fosse stato possibile in passato. Spesso associati a questa marcata soppressione si manifestano stati di stanchezza. In tali condizioni, l’atleta, anche se sta seguendo un ciclo correttamente formulato, potrebbe non essere in grado di massimizzare i propri risultati di miglioramento della condizione fisica a causa di un’incapacità di allenarsi con pieno vigore. Questo effetto è talvolta anche soprannominato “letargia steroidea”. La ragione principale per cui ciò accade è legata all’importante azione di supporto all’attività della Serotonina data dall’Estrogeno. La Serotonina è uno dei principali neurotrasmettitori del corpo, di essenziale importanza per un adeguata lucidità mentale e un regolare ciclo sonno / veglia. (77) (78)L’alterazione di questo neurotrasmettitore è associata anche alla sindrome da affaticamento cronico, (79) (80) e ciò ci fa comprendere quanto possa essere incisiva in particolare per la stanchezza. L’abbassamento dei livelli estrogenici nella menopausa è stata associata anche alla stanchezza, (81) così come l’uso clinico di inibitori dell’aromatasi più recenti (e più potenti) come l’Anastrozolo, (82) il Letrozolo, (83) l’Exemestane, (84) e il Fadrozolo (85) in alcuni pazienti. Questo effetto deve essere preso in importante considerazione quando si pianifica un ciclo. Anche se non tutti notano questo problema quando il livello estrogenico è basso, per coloro i quali lo subiscono, l’aggiunta di un po’ di Testosterone o di altro AAS soggetto all’aromatizzazione (oltre che l’abbassamento del composto AI) può aiutare a correggere di molto questo sintomo. È anche da notare che l’uso di AAS non aromatizzabili a volte causa questo effetto, probabilmente dovuto alla soppressione della produzione naturale di Testosterone (abbassando la disponibilità del principale substrato usato dal corpo maschile per sintetizzare Estradiolo).

*Nota: gli AAS metilati in C-17, soggetti ad aromatizzazione (vedi Methyltestosterone e Mathandrostenolone), convertono nel potente Methylestradiolo, o 17α-methylestradiolo (17α-ME). A causa della presenza del gruppo 17α-metile, il Methylestradiolo non può essere disattivato per ossidazione del gruppo 17β-idrossile, con conseguente migliorata stabilità metabolica e potenza di legame recettoriale (e conseguente attività estrogenica) rispetto all’Estradiolo. (86)(87)(88)
- Quindi, come gestire il fattore estrogenico durante la preparazione?

Arrivati a questo punto, quale significato ha quanto è stato trattato per un Bodybuilder che cerca di ottenere una forma ottimale? Fondamentalmente penso che il punto chiave sia quello di mantenere un approccio prudente all’uso di farmaci per il controllo estrogenico, un approccio che deve essere dettato dalle reali esigenze verificate attraverso appositi esami ematici. Ovviamente, è chiaro che l’approccio all’uso dei composti per il controllo estrogenico debba variare a seconda dell’obbiettivo (vale a dire se l’obbiettivo primario è l’aumento della massa muscolare o se è l’aumento della definizione muscolare). Ripeto, è ovvio che l’utilizzo di composti volti al controllo estrogenico è una chiara necessità qualora si manifestassero effetti collaterali estrogenici, come lo è, nel caso, il calo dei composti soggetti all’azione dell’enzima aromatasi, ma gli esami ematici sono sempre di primaria importanza ai fini di una gestione ottimale del problema. La ginecomastia è certamente un problema indesiderato per l’utilizzatore di AAS, come lo è la ritenzione idrica ed un eccessivo accumulo di massa grassa. Ma se tali problemi non emergono, o se dagli esami ematici non emerge la reale necessità, l’aggiunta di composti per il controllo estrogenico (che siano AI, SERM o inibitori della biosintesi steroidea) può inficiare i risultati ottenibili in un contesto “Bulk”.
Quindi, concludendo, in un contesto “Bulk”, al fine di mantenere i benefici estrogenici senza incappare negli effetti collaterali estrogeno-dipendenti, la soglia ematica di Estradiolo dovrebbe mantenersi all’interno del range medio di riferimento; ciò significa che, in teoria, non dovrebbe ne scendere al di sotto dei 30pg/pg ne salire al di sopra dei 60 pg/ml. In un contesto “Cut” o “Pre-Gara”, ovviamente, le cose cambiano. In tali contesti, considerando gli effetti estrogenici anche sullo spessore della pelle, la cosa migliore sarebbe optare per il mantenimento di un livello di estrogeni basso-normale (20-30pg/ml) per tutta la fase “Cut” della preparazione per poi, se si è in preparazione alla gara, calare i livelli a 10pg o meno nell’ultimo paio di settimane prima del contest.
Di questi temi ho già parlato in modo approfondito in due articoli (“Crescita muscolare e connessione con gli estrogeni” e “Ormone della crescita, estrogeni, e ispessimento della pelle“).
Per le atlete il discorso è simile anche se diverso sotto alcuni aspetti. In ambito femminile il controllo dei livelli e dell’attività estrogenica è di importanza anche maggiore per il raggiungimento di una forma da gara ottimale (vedere ad esempio “Nolvadex (Tamoxifene Citrato)”e “RALOXIFENE E COMPOSIZIONE CORPOREA NELLE DONNE”).
In definitiva, come logica conclusione, il metodo d’approccio per la gestione estrogenica deve basarsi sulle reali necessità, su dati oggettivi, e non su un fallace e molto poco scientifico “passa parola” dettato dall'”esperienza” di qual si voglia agonista o ex tale…
Gabriel Bellizzi
Riferimenti:
1- “estrogen (CHEBI:50114)”. ChEBI. Retrieved 15 July 2015.
2- Ryan KJ (August 1982). “Biochemistry of aromatase: significance to female reproductive physiology”. Cancer Res. 42 (8 Suppl): 3342s–3344s. PMID 7083198.
3- Mechoulam R, Brueggemeier RW, Denlinger DL (September 2005). “Estrogens in insects” (PDF). Cellular and Molecular Life Sciences. 40 (9): 942–944. doi:10.1007/BF01946450.
4- Burger HG (2002). “Androgen production in women”. Fertility and Sterility. 77 Suppl 4: S3–5. doi:10.1016/S0015-0282(02)02985-0. PMID 12007895.
5- Lombardi G, Zarrilli S, Colao A, Paesano L, Di Somma C, Rossi F, De Rosa M (2001). “Estrogens and health in males”. Molecular and Cellular Endocrinology. 178 (1–2): 51–5. doi:10.1016/S0303-7207(01)00420-8. PMID 11403894.
6- Whitehead SA, Nussey S (2001). Endocrinology: an integrated approach. Oxford: BIOS: Taylor & Francis. ISBN 1-85996-252-1.
7- Soltysik K, Czekaj P (April 2013). “Membrane estrogen receptors — is it an alternative way of estrogen action?”. J. Physiol. Pharmacol. 64 (2): 129–42. PMID 23756388.
8- Micevych PE, Kelly MJ (2012). “Membrane estrogen receptor regulation of hypothalamic function”. Neuroendocrinology. 96 (2): 103–10. doi:10.1159/000338400. PMC 3496782 ![]() . PMID 22538318.
. PMID 22538318.
9- Prossnitz ER, Arterburn JB, Sklar LA (2007). “GPR30: A G protein-coupled receptor for estrogen”. Mol. Cell. Endocrinol. 265–266: 138–42. doi:10.1016/j.mce.2006.12.010. PMC 1847610 ![]() . PMID 17222505.
. PMID 17222505.
10- Files JA, Ko MG, Pruthi S (2011). “Bioidentical hormone therapy”. Mayo Clin. Proc. 86 (7): 673–80, quiz 680. doi:10.4065/mcp.2010.0714. PMC 3127562 ![]() . PMID 21531972.
. PMID 21531972.
11- Lin CY, Ström A, Vega VB, Kong SL, Yeo AL, Thomsen JS, Chan WC, Doray B, Bangarusamy DK, Ramasamy A, Vergara LA, Tang S, Chong A, Bajic VB, Miller LD, Gustafsson JA, Liu ET (2004). “Discovery of estrogen receptor alpha target genes and response elements in breast tumor cells”. Genome Biol. 5 (9): R66. doi:10.1186/gb-2004-5-9-r66. PMC 522873 ![]() . PMID 15345050.
. PMID 15345050.
12- Darabi M, Ani M, Panjehpour M, Rabbani M, Movahedian A, Zarean E (2011). “Effect of estrogen receptor β A1730G polymorphism on ABCA1 gene expression response to postmenopausal hormone replacement therapy”. Genet Test Mol Biomarkers. 15 (1–2): 11–5. doi:10.1089/gtmb.2010.0106. PMID 21117950.
13- Raloff J (December 6, 1997). “Science News Online (12/6/97): Estrogen’s Emerging Manly Alter Ego”. Science News. Retrieved 2008-03-04.
14- Hess RA, Bunick D, Lee KH, Bahr J, Taylor JA, Korach KS, Lubahn DB (1997). “A role for estrogens in the male reproductive system”. Nature. 390 (6659): 447–8. doi:10.1038/37352. PMID 9393999.
15- “Science Blog – Estrogen Linked To Sperm Count, Male Fertility”. Science Blog. Retrieved 2008-03-04.
16- Hill RA, Pompolo S, Jones ME, Simpson ER, Boon WC (2004). “Estrogen deficiency leads to apoptosis in dopaminergic neurons in the medial preoptic area and arcuate nucleus of male mice”. Mol. Cell. Neurosci. 27 (4): 466–76. doi:10.1016/j.mcn.2004.04.012. PMID 15555924.
17- Marieb, Elaine (2013). Anatomy & physiology. Benjamin-Cummings. p. 903. ISBN 9780321887603.
18- Nelson LR, Bulun SE (September 2001). “Estrogen production and action”. J. Am. Acad. Dermatol. 45 (3 Suppl): S116–24. doi:10.1067/mjd.2001.117432. PMID 11511861.
19- MacDonald PC, Madden JD, Brenner PF, Wilson JD, Siiteri PK: Origin of estrogen in normal men and in women with testicular feminization. J Clin Endocrinol Metab 49:905-916, 1979
20- Simpson ER, Mahendroo MS, Means GD, Kilgore MW, Hinshelwood MM, Graham-Lorence S, Amarneh B, Ito Y, Fisher CR, Michael MD, et al.: Aromatase cytochrome P450, the enzyme responsible for estrogen biosyn- thesis. Endocr Rev 15:342-355, 1994
21- Simpson ER, Zhao Y, Agarwal VR, Michael MD, Bulun SE, Hinshelwood MM, Graham-Lorence S, Sun T, Fisher CR, Qin K, Mendelson CR: Aromatase expression in health and disease. Recent Prog Horm Res 52:185-213; discus- sion 213-184, 1997
22- Simpson ER: Aromatase: biologic relevance of tissue-specific expression. Semin Reprod Med 22:11-23, 2004
23- Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, Evans RM: The nuclear recep- tor superfamily: the second decade. Cell 83:835-839, 1995
24- Green S, Walter P, Kumar V, Krust A, Bornert JM, Argos P, Chambon P: Human oestrogen receptor cDNA: sequence, expression and homology to v-erb-A. Nature 320:134-139, 1986
25- Greene GL, Gilna P, Waterfield M, Baker A, Hort Y, Shine J: Sequence and expression of human estrogen receptor complementary DNA. Science 231:1150-1154, 1986
26- Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson JA: Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology 138:863-870, 1997
27- Enmark E, Gustafsson JA: Oestrogen receptors – an overview. J Intern Med 246:133-138, 1999
28- Levin ER: Cellular Functions of the Plasma Membrane Estrogen Receptor. Trends Endocrinol Metab 10:374-377, 1999
29- Pedram A, Razandi M, Levin ER: Nature of functional estrogen receptors at the plasma membrane. Mol Endocrinol 20:1996-2009, 2006
30- Filardo E, Quinn J, Pang Y, Graeber C, Shaw S, Dong J, Thomas P: Activation of the novel estrogen receptor G protein-coupled receptor 30 (GPR30) at the plasma membrane. Endocrinology 148:3236-3245, 2007
31- de Ronde W, Pols HA, van Leeuwen JP, de Jong FH: The importance of oestrogens in males. Clin Endocrinol (Oxf) 58:529-542, 2003
32- Baird DT, Horton R, Longcope C, Tait JF: Steroid dynamics under steady- state conditions. Recent Prog Horm Res 25:611-664, 1969
33- Baird DT, Uno A, Melby JC: Adrenal secretion of androgens and oestrogens. J Endocrinol 45:135-136, 1969
34- Dunn JF, Nisula BC, Rodbard D: Transport of steroid hormones: bind- ing of 21 endogenous steroids to both testosterone-binding globulin and corticosteroid-binding globulin in human plasma. J Clin Endocrinol Metab 53:58-68, 1981
35- Carr BR: Disorders of the ovaries and female reproductive tract. Williams Textbook of Endocrinology. 11th ed. W.B. Saunders Company ed. Polonsky KS, Melmed S, Kronenberg HM, Larsen PR, Eds. Philadelphia, 2008
36- Weinbauer GF, Nieschlag E: The role of testosterone in spermatogene- sis. Springer-Verlag ed. Nieschlag E, Behre H, Eds. Berlin Heidelberg, Testosterone action deficiency substitution, 1990, p. 23-50
37- Zondek B: Mass excretion of oestrogenic hormone in the urine of the stallion. Nature 33:209-210, 1934
38- Heard RD, Jellinck PH, O’Donnell VJ: Biogenesis of the estrogens: the con- version of testosterone-4-C14 to estrone in the pregnant mare. Endocrinology 57:200-204, 1955.
39- West CD, Damast BL, Sarro SD, Pearson OH: Conversion of testosterone to estrogens in castrated, adrenalectomized human females. J Biol Chem 218:409-418, 1956.
40- Davis JW, Gut M, Lemon HM, Wotiz HH: Studies in steroid metabolism. V. The conversion of testosterone-4-C14 to estrogens by human ovarian tissue. J Biol Chem 222:487-495, 1956
41- Baggett B, Dorfman RI, Engel LL, Savard K: The conversion of testoster- one-3-C14 to C14-estradiol-17beta by human ovarian tissue. J Biol Chem 221:931-941, 1956 8. Baggett B, Engel LL, Balderas L, Lanman G: Conversion of C14-testosterone to C14-estrogenic steroids by endocrine tissues. Endocrinology 64:600-608, 1959
42- Smith EP, Boyd J, Frank GR, Takahashi H, Cohen RM, Specker B, Williams TC, Lubahn DB, Korach KS: Estrogen resistance caused by a mutation in the estrogen-receptor gene in a man. N Engl J Med 331:1056-1061, 1994
43- Morishima A, Grumbach MM, Simpson ER, Fisher C, Qin K: Aromatase deficiency in male and female siblings caused by a novel mutation and the physiological role of estrogens. J Clin Endocrinol Metab 80:3689-3698, 1995
44- Carani C, Qin K, Simoni M, Faustini-Fustini M, Serpente S, Boyd J, Korach KS, Simpson ER: Effect of testosterone and estradiol in a man with aromata- se deficiency. N Engl J Med 337:91-95, 1997
45- Bilezikian JP, Morishima A, Bell J, Grumbach MM: Increased bone mass as a result of estrogen therapy in a man with aromatase deficiency. N Engl J Med 339:599-603, 1998
46- Herrmann BL, Saller B, Janssen OE, Gocke P, Bockisch A, Sperling H, Mann K, Broecker M: Impact of estrogen replacement therapy in a male with con- genital aromatase deficiency caused by a novel mutation in the CYP19 gene. J Clin Endocrinol Metab 87:5476-5484, 2002
47- Pura M, Mittre H, Carreau S, Kottler ML: Clinical findings in an adult man with a novel mutation in the aromatase gene. In Program of the 85th Annual Meeting of the Endocrine Society, Abstract Book Philadelphia, PA, 2003
48- Maffei L, Murata Y, Rochira V, Tubert G, Aranda C, Vazquez M, Clyne CD, Davis S, Simpson ER, Carani C: Dysmetabolic syndrome in a man with a novel mutation of the aromatase gene: effects of testosterone, alendronate, and estradiol treatment. J Clin Endocrinol Metab 89:61-70, 2004
49- Maffei L, Rochira V, Zirilli L, Antunez P, Aranda C, Fabre B, Simone ML, Pignatti E, Simpson ER, Houssami S, Clyne CD, Carani C: A novel com- pound heterozygous mutation of the aromatase gene in an adult man: reinforced evidence on the relationship between congenital oestrogen deficiency, adiposity and the metabolic syndrome. Clin Endocrinol (Oxf) 67:218-224, 2007
50- Luberto A, Rochira V, Zirilli L, Pignatti E, Simpson ER, Maffei L, Carani C: A novel compound heterozygous mutation of the aromatase gene in an adult man: a reinforced estrogen deficincy, adiposity and the metabolic syndrome. J Endocrinol Invest 30 Suppl 4:50, 2007
51- Bouillon R, Bex M, Vanderschueren D, Boonen S: Estrogens are essen- tial for male pubertal periosteal bone expansion. J Clin Endocrinol Metab 89:6025-6029, 2004
52- Vanderschueren D, Boonen S, Bouillon R: Action of androgens versus estrogens in male skeletal homeostasis. Bone 23:391-394, 1998
53- Vanderschueren D, Venken K, Ophoff J, Bouillon R, Boonen S: Clinical Review: Sex steroids and the periosteum–reconsidering the roles of androgens and estrogens in periosteal expansion. J Clin Endocrinol Metab 91:378-382, 2006
54- Horsman A, Gallagher JC, Simpson M, Nordin BE: Prospective trial of oestrogen and calcium in postmenopausal women. Br Med J 2:789-792, 1977
55- Lindsay R, Hart DM, Forrest C, Baird C: Prevention of spinal osteoporosis in oophorectomised women. Lancet 2:1151-1154, 1980
56- Christiansen C, Christensen MS, Transbol I: Bone mass in postmenopausal women after withdrawal of oestrogen/gestagen replacement therapy. Lancet 1:459-461, 1981
57- Finkelstein JS, Neer RM, Biller BM, Crawford JD, Klibanski A: Osteopenia in men with a history of delayed puberty. N Engl J Med 326:600-604, 1992
58- Speroff L, Rowan J, Symons J, Genant H, Wilborn W: The comparative effect on bone density, endometrium, and lipids of continuous hormones as replacement therapy (CHART study). A randomized controlled trial. Jama 276:1397-1403, 1996
59- Wang C, Eyre DR, Clark R, Kleinberg D, Newman C, Iranmanesh A, Veldhuis J, Dudley RE, Berman N, Davidson T, Barstow TJ, Sinow R, Alexander G, Swerdloff RS: Sublingual testosterone replacement improves muscle mass and strength, decreases bone resorption, and increases bone formation markers in hypogonadal men–a clinical research center study. J Clin Endocrinol Metab 81:3654-3662, 1996
60- Faustini-Fustini M, Rochira V, Carani C: Oestrogen deficiency in men: where are we today? Eur J Endocrinol 140:111-129, 1999
61- Vanderschueren D, Venken K, Ophoff J, Bouillon R, Boonen S: Clinical Review: Sex steroids and the periosteum–reconsidering the roles of androgens and estrogens in periosteal expansion. J Clin Endocrinol Metab 91:378-382, 2006
62-https://www.researchgate.net/publication/290473706_The_Role_of_Estradiol_in_Male_Reproductive_Function
64- The aromatization of androstenedione by human adipose and liver tissue. J Steroid Biochem. 1980 Dec;13(12):1427-31.
65- Aromatase expression in the human male. Brodie A, Inkster S, Yue W. Mol Cell Endocrinol 2001 Jun 10;178(1-2):23-8
66- A review of brain aromatase cytochrome P450. Lephart ED. Brain Res Brain Res Rev 1996 Jun;22(1):1-26
69- Pentose Cycle Activity in Muscle from Fetal, Neonatal and Infant Rhesus Monkeys. Arch Biochem Biophys 117:275-81 1966
70- The pentose phosphate pathway in regenerating skeletal muscle. Biochem J 170: 17 1978
71- Aromatization of androgens to estrogens mediates increased activity of glucose 6-phosphate dehydrogenase in rat levator ani muscle. Endocrinol 106(2):440-43 1980
72- Influence of tamoxifen, aminoglutethimide and goserelin on human plasma IGF-1 levels in breast cancer patients. J steroid Biochem Mol Bio 41:541- 3,1992
73- Activation of the somatotropic axis by testosterone in adult males: Evidence for the role of aromatization. J Clin. Endocrinol Metab 76:1407-12 1993
74- Testosterone administration increases insulin-like growth factor-I levels in normal men. J Clin Endocrinol Metab 77(3):776-9 1993
76- Modulation of the cytosolic androgen receptor in striated muscle by sex steroids. Endocrinology. 1984 Sep;115(3):862-6.
78- Serotonin and the sleep/wake cycle: special emphasis on miscodialysis studies. Chiara M Portas et al. Progress in Neurology 60(200) 13-35.
80- Association between serotonin transporter gene polymorphism and chronic fatigue syndrome. Narita M et al. Biochem Biophys Res Commun 2003 Nov 14;311(2)264-6
81- Premenstrual Syndrome. Dickerson LM et al. Am Fam Physician 2003 Apr 15;67(8):1743-52
82- Phase II trial of anastrozole in women with asymptomatic mullerian cancer. Gynecol Oncol. 2003 Dec;91(3):596-602.
83- Letrozole. A review of its use in postmenopausal women with advanced breast cancer. Drugs. 1998 Dec;56(6):1125-40. Review.
84- Exemestane: a review of its clinical efficacy and safety. Breast. 2001 Jun;10(3):198-208.
85- A study of fadrozole, a new aromatase inhibitor, in postmenopausal women with advanced metastatic breast cancer. J Clin Oncol. 1992 Jan;10(1):111-6.
86- Detlef Thieme; Peter Hemmersbach (18 December 2009). Doping in Sports. Springer Science & Business Media. pp. 470–. ISBN 978-3-540-79088-4.
87- Feenstra A, Vaalburg W, Nolten GM, Reiffers S, Talma AG, Wiegman T, van der Molen HD, Woldring MG (1983). “Estrogen receptor binding radiopharmaceuticals: II. Tissue distribution of 17 alpha-methylestradiol in normal and tumor-bearing rats”. J. Nucl. Med. 24 (6): 522–8. PMID 6406650.
88- William Llewellyn (2011). Anabolics. Molecular Nutrition Llc. pp. 533–. ISBN 978-0-9828280-1-4.
