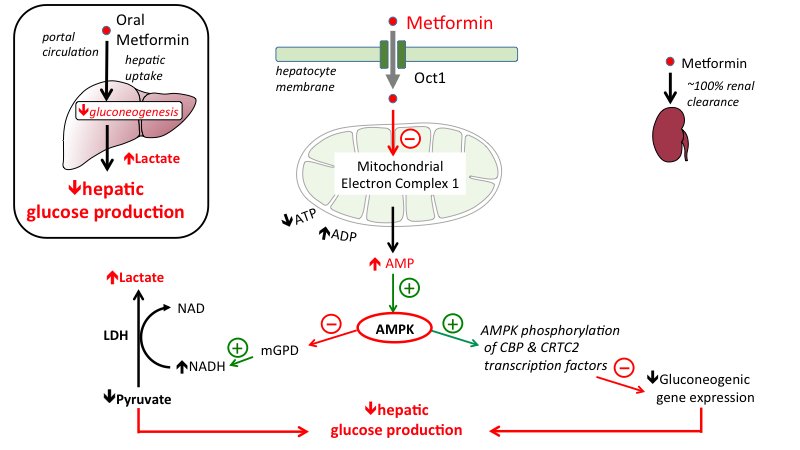
Chi non conosce, nel 2021, dopo quasi un secolo di ricerca, la Metformina ed i suoi effetti sul miglioramento della sensibilità all’Insulina, con conseguente miglioramento del uptake cellulare di glucosio ? E dei vantaggi che esso può apportare ai Bodybuilder in fase di “Refeed”, magari dopo periodi medio-lunghi a bassi CHO e con una capacità di gestirli non proprio ottimale?
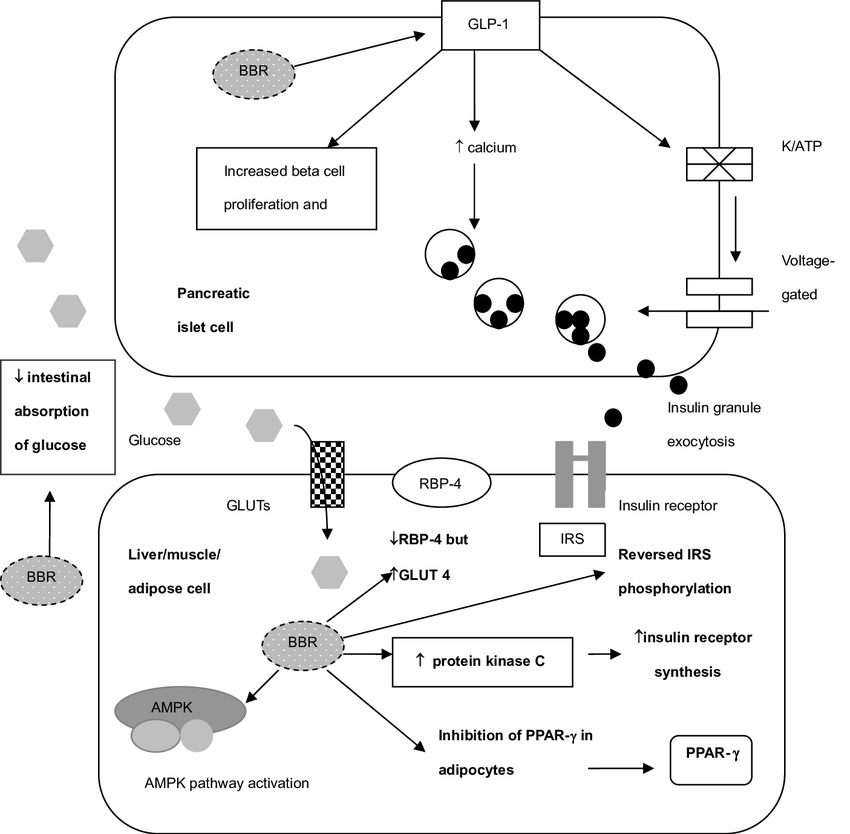
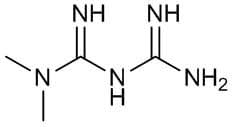
La stessa cosa interessa anche la Berberina, la quale possiede vie farmacodinamiche molto simili alla Metformina. Entrambe le molecole, però, hanno un limite, e questo limite è comune a tutte le Biguanidi oggi in uso clinico o quelle appartenenti ai GDA (come la Berberina): la mancanza di selettività tissutale. Esse, infatti, migliorano sia l’IS del miocita che dell’adipocita, oltre ad attivare l’AMPK con alterazione del mTOR.
Nota: per chi non lo sapesse, le Biguanidi sono una categoria di farmaci ipoglicemizzanti orali di indicazione specifica contro il diabete di tipo II. A differenza di altri farmaci antidiabetici, come ad esempio le sulfaniluree, non determinano un aumento di rilascio di Insulina per cui non causano generalmente ipoglicemia. In questa sede mi riferirò con il termine “Biguanidi” a quelle molecole con tali caratteristiche, sia farmaceutiche (vedi Metformina) che appartenenti al panorama da banco denominato GDA (vedi Berberina).
Ora, potremmo anche dire che in un soggetto con una buona massa contrattile e una massa grassa tendenzialmente bassa questo “difetto” non causa particolari problemi nel complesso della preparazione. Ma c’è da considerare che una selettività miocitaria garantirebbe una ripartizione calorica ottimale in un contesto, per esempio, ipercalorico riducendo gli “approvvigionamenti” degli adipociti e prolungando sensibilmente la soglia temporale durante la quale l’atleta in questione potrebbe crescere in modo qualitativamente soddisfacente. Un pò come quando si ipotizzava sulla applicazione di molecole con teorica attività di riduzione dello stoccaggio degli Acidi Grassi. Discorsi ed effetti diversi, ma il fine è uno: aumentare il tempo di durata della “soglia di crescita qualitativa”.
Per “soglia di crescita qualitativa” intendo la possibilità di proseguire con la programmazione in ipercalorica ottenendo maggiori aumenti ipertrofici del muscolo-scheletrico piuttosto che del tessuto adiposo.
“Ma Gabriel! E l’interferenza con l’mTOR osservata con la Metformina ed altre molecole che stimolano l’attività del AMPK non è forse una limitazione ben più importante???!!!” Calma, piccola zecca interattiva, ne parlerò a tempo debito, come parlerò del fatto che è la dose a determinare se l’alterazione risulterà significativa o meno…. Proseguiamo…
In questo articolo tratterò della nuova molecola sperimentale denominata DS20060511, riporterò quanto è a nostra conoscenza ad oggi e quali sono le sue caratteristiche e possibili applicazioni che, tra l’altro, ho già accennato in questa introduzione…
Il principio della scoperta:
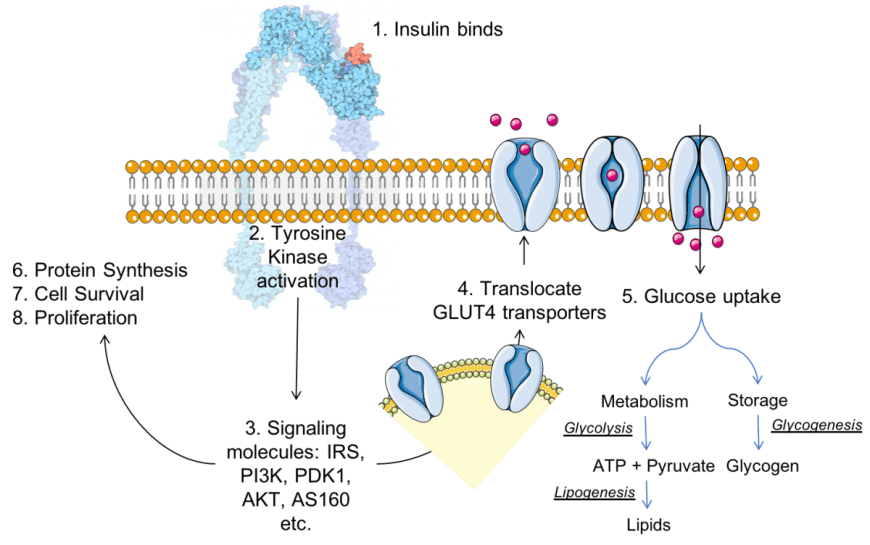
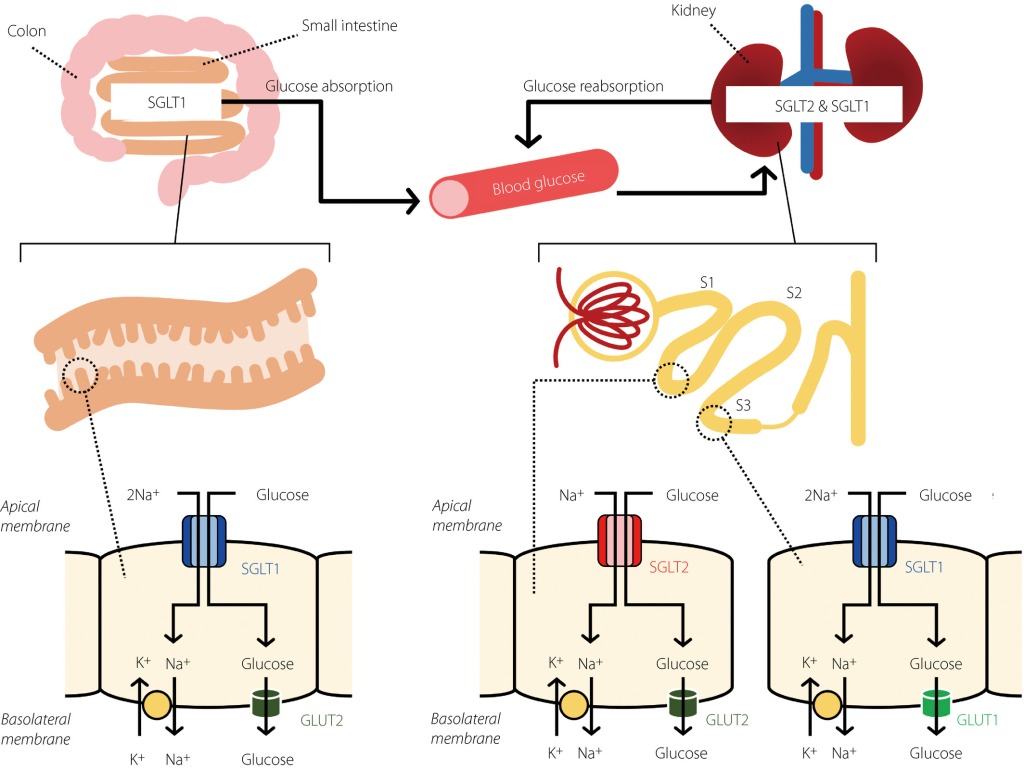
La riduzione dell’assorbimento del glucosio nel muscolo scheletrico è un’importante anomalia fisiopatologica nel diabete di tipo II ed è causata dalla alterazione della funzionalità di traslocazione dei GLUT4 sulla superfice cellulare del miocita nel tessuto muscolo-scheletrico.
Il trasportatore del glucosio di tipo 4 (GLUT-4), noto anche come famiglia di trasportatori di soluti 2, membro 4 del trasportatore di glucosio facilitato, è una proteina codificata, nell’uomo, dal gene SLC2A4. Il GLUT4 è il trasportatore del glucosio regolato dall’insulina, ma non solo, che si trova principalmente nei tessuti adiposo e nel muscolo striato (scheletrico e cardiaco). La prima prova di questa distinta proteina di trasporto del glucosio è stata fornita da David James nel 1988. Il gene che codifica per il GLUT4 è stato clonato e mappato nel 1989.
Struttura molecolare del GLUT-4
Il GLUT4 è il trasportatore che limita la velocità di assorbimento del glucosio e svolge un ruolo cruciale nel mantenimento dell’omeostasi del glucosio [1, 2]. I soggetti con diabete di tipo II mostrano un ridotto assorbimento di glucosio da parte del muscolo scheletrico a causa della ridotta traslocazione di GLUT4 nella superficie delle cellule del muscolo scheletrico[3]. È stato riportato che i topi diabetici con sovraespressione di GLUT4 mostrano livelli di glucosio plasmatico marcatamente ridotti sia a digiuno che in condizioni postprandiali [4,5,6].
Sebbene il GLUT4 sia immagazzinato in vescicole di stoccaggio intracellulari in condizioni basali, l’Insulina, e l’attività di contrazione del muscolo, induce la traslocazione di GLUT4 sulla superficie cellulare, facilitando l’assorbimento del glucosio [7,8]. L’Insulina attiva Akt tramite il substrato del recettore dell’Insulina (IRS)s-fosfoinositide 3-chinasi (PI3K) [9,10] e l’Akt attivato fosforila e di conseguenza inibisce le proteine Akt substrato di 160 kDa (AS160) e membro della famiglia del dominio TBC1 1 (TBC1D1) , entrambi sono proteine attivanti Rab GTPasi (GAP); ciò si traduce nell’attivazione delle proteine Rab e nella traslocazione di GLUT4 sulla superficie della membrana cellulare [11]. È stato riportato che il substrato 1 (Rac1) della tossina botulinica C3 correlato a RAS, un’altra molecola a valle di PI3K, promuove la traslocazione di GLUT4 indipendentemente dalla via Akt-AS160/TBC1D1-Rab. Rac1 stimola la riorganizzazione della polimerizzazione dell’actina corticale, che consente l’inserimento delle vescicole contenenti GLUT4 nella membrana cellulare[12,13]. È noto che l‘Insulina regola la traslocazione di GLUT4 sia attraverso la via di Akt-AS160-Rab che attraverso la via di polimerizzazione di Rac1-actina[14,15]. Nei soggetti con diabete di tipo II, entrambe le vie di segnalazione dell’Insulina sono compromesse nel muscolo scheletrico, con conseguente riduzione dell’assorbimento del glucosio indotto dall’Insulina in questo tessuto.
Schema della traslocazione indotta dall’Insulina del GLUT4 dal citosol alla membrana cellulare. Il legame dell’Insulina ai suoi recettori avvia una cascata di trasduzione del segnale, che si traduce nell’attivazione di Akt. Akt agisce sul GLUT4 contenuto nelle vescicole nel citosol per facilitarne la fusione con la membrana cellulare. Quando più molecole GLUT4 sono presenti nella membrana, più la velocità di assorbimento del glucosio è elevata.
Come già accennato, la contrazione durante l’esercizio è un altro importante potenziatore della traslocazione di GLUT4 nel muscolo scheletrico[16]. All’aumentata richiesta di glucosio durante l’esercizio nel muscolo scheletrico, il GLUT4 si trasloca sulla superficie cellulare per promuovere l’apporto di glucosio al muscolo scheletrico[17,18]. L’esercizio aumenta il rapporto AMP/ATP causato dal consumo di ATP, portando all’attivazione della chinasi attivata dall’AMP (AMPK). Nonostante l’evidenza riportata di una contrazione che induce la fosforilazione di TBC1D1 mediante l’attivazione di AMPK[19] o di un aumento dell’assorbimento del glucosio nel muscolo scheletrico mediante attivazione farmacologica di AMPK da parte di AICAR[20], il significato dell’AMPK nell’assorbimento del glucosio stimolato dall’esercizio in vivo rimane controverso [21,22]. Recentemente, l’induzione da parte di Rac1 della produzione NADPH ossidasi 2-dipendente di specie reattive dell’ossigeno è stata implicata nell’assorbimento del glucosio durante l’esercizio, attraverso la regolazione della traslocazione di GLUT4 [23,24]. La contrazione del muscolo scheletrico non ha indotto la fosforilazione di IRS1 o PI3K[25]. La captazione del glucosio indotta dalla contrazione o la traslocazione di GLUT4 nel muscolo scheletrico non è stata inibita dalla Wortmannina, un inibitore di PI3K [26,27]. Inoltre, la combinazione di Insulina e contrazione del muscolo scheletrico ha causato un ulteriore aumento della traslocazione di GLUT4 e dell’assorbimento di glucosio rispetto alla sola Insulina [27]. Questi dati suggeriscono che la contrazione del muscolo scheletrico stimola la traslocazione di GLUT4 indipendentemente dall’Insulina.
Wortmannina
Nei soggetti con diabete di tipo II, i campioni bioptici del muscolo scheletrico ottenuti durante un clamp insulinico euglicemico hanno mostrato un’alterata segnalazione dell’Insulina, osservata come riduzione della fosforilazione di IRS1 e dell’attività di PI3K, nel muscolo scheletrico[28], mentre non è stato osservato alcun effetto sulla fosforilazione/attività di Akt [29]. Altri studi hanno dimostrato una riduzione della traslocazione di GLUT4 e dell’assorbimento di glucosio in soggetti con diabete di tipo II [23,28]. Inoltre, è stato riportato che la ridotta traslocazione di GLUT4 nei soggetti con diabete di tipo II è stata migliorata dall’esercizio fisico [30,31]. Questi risultati suggeriscono che l’induzione della traslocazione di GLUT4 nel muscolo scheletrico potrebbe essere un potenziale bersaglio terapeutico nei pazienti con diabete di tipo II.
Recentemente, i ricercatori dell’azienda farmaceutica giapponese Daiichi Sankyo hanno dimostrato che il derivato xantenico DS20060511 induce la traslocazione di GLUT4 specifica del muscolo scheletrico, indipendentemente dall’azioni dell’Insulina. Hanno utilizzato miotubi L6 che esprimono GLUT4 marcato con myc (L6-GLUT4myc) per esaminare la libreria di composti chimici in loro possesso e misurare la traslocazione di GLUT4 sulla superficie cellulare mediante dosaggio immunologico anti-myc quantitativo. Gli effetti del composto sull’assorbimento del glucosio e sul metabolismo del glucosio in tutto il corpo sono stati esaminati in una serie di esperimenti in vitro e in vivo. Il meccanismo d’azione del composto è stato esplorato studiando le vie di segnalazione note coinvolte nella traslocazione di GLUT4 indotta dall’Insulina e dall’esercizio fisico. Infine, abbiamo valutato il potenziale terapeutico del composto in un modello murino obeso e insulino-resistente con diabete di tipo II.
Molecola di Xantene, base strutturale dei derivati xantenici.
Nota: I derivati xantenici sono modificazioni molecolari dello Xantene (9H-xantene, 10H-9-ossaantracene), un composto organico con la formula CH2[C6H4]2O. È un solido giallo solubile nei comuni solventi organici. Lo stesso xantene è un composto oscuro, ma molti dei suoi derivati sono coloranti utili.
Il DS20060511, è un induttore specifico per la traslocazione di GLUT4 nelle cellule muscolo-scheletriche:
I ricercatori, come detto pocanzi, hanno esaminato la loro libreria chimica, composta da oltre 100.000 composti, utilizzando miotubi L6-GLUT4myc, per identificare i composti che avrebbero indotto la traslocazione di GLUT4 sulla superficie cellulare. Sono stati identificati due composti completamente diversi ed entrambi hanno superato il test per escludere composti che avrebbero esercitato effetti tossici, come l’inibizione della catena respiratoria. Ulteriori test in vitro hanno rivelato che uno dei due composti ha influenzato la via Akt, così che alla fine hanno selezionato l’altro, un composto xantenico originale, come composto con il potenziale effetto di indurre la traslocazione di GLUT4. L’ottimizzazione della struttura molecolare ha infine prodotto il composto xantenico più potente, DS20060511 (vedi immagine seguente). Il trattamento con DS20060511 ha aumentato la traslocazione di GLUT4 nei miotubi differenziati L6-GLUT4myc in modo concentrazione-dipendente, come nel caso del trattamento con Insulina. Tuttavia, mentre il trattamento con Insulina ha anche aumentato la traslocazione di GLUT4 negli adipociti differenziati 3T3-L1-GLUT4myc, il trattamento con DS20060511 non ha avuto quasi alcun effetto sulla traslocazione di GLUT4 in questi adipociti, suggerendo che l’induzione della traslocazione di GLUT4 da parte di DS20060511 è specifica per le cellule del tessuto muscolo-scheletrico. Coerentemente con questi dati, il trattamento con DS20060511 ha aumentato significativamente l’assorbimento di 2-DG in modo concentrazione-dipendente nei miotubi L6-GLUT4myc, come nel caso del trattamento con Insulina. Ancora una volta, mentre è stato dimostrato che l’Insulina aumenta l’assorbimento di 2-DG negli adipociti differenziati 3T3-L1-GLUT4myc, DS20060511 non ha mostrato tale effetto negli adipociti. Questi dati suggeriscono che il composto xantenico DS20060511 promuove l’assorbimento del glucosio mediante l’attivazione specifica della traslocazione di GLUT4 nelle cellule muscolo-scheletriche.
a Struttura chimica del DS20060511. b, c Induzione concentrazione-dipendente della traslocazione di GLUT4 da parte del DS20060511 e Insulina nei miotubi L6-GLUT4myc (b) e negli adipociti 3T3-L1-GLUT4myc (c). d, e Captazione di 2-DG valutata nei miotubi L6-GLUT4myc (d) e negli adipociti 3T3-L1-GLUT4myc (e). I valori mostrati sono mezzi ± SEM, n = 3. **P < 0,01, ***P < 0,001 rispetto al controllo mediante ANOVA unidirezionale seguito dal test di Dunnett.
Il trattamento con DS20060511, riduzione dei livelli di glucosio ematico e aumento potenziato dell’assorbimento di glucosio per via della traslocazione di GLUT4 nel muscolo scheletrico in vivo:
Per studiare gli effetti del DS20060511 sulla dinamica del glucosio in vivo, il composto è stato somministrato a topi normali. Nei topi che avevano continuato ad accedere al cibo, la sola somministrazione orale di DS20060511 in modo modesto, ma statisticamente significativo, ha ridotto i livelli di glucosio nel sangue, mentre nei topi che avevano negato l’accesso al cibo durante la notte, il composto non ha esercitato alcun effetto sui livelli di glucosio nel sangue. Quando è stato somministrato prima del carico orale di glucosio nel test di tolleranza al glucosio orale (GTT), il DS20060511 ha prodotto una soppressione dose-dipendente dell’aumento dei livelli di glucosio nel sangue dopo un carico orale di glucosio. La secrezione di Insulina durante il GTT orale è stata ridotta in modo piuttosto significativo in tutti i gruppi trattati con DS20060511, suggerendo che il trattamento con DS20060511 riduce i livelli di glucosio nel sangue indipendentemente dalla secrezione di Insulina. Il trattamento con DS20060511 ha prodotto un aumento significativo dell’assorbimento di [3H]-2-DG nei muscoli soleo e gastrocnemio, ma non nel cuore o nel tessuto adiposo bianco (WAT) durante il GTT intraperitoneale. L’analisi Western blot ha rivelato un aumento dei livelli di espressione della proteina GLUT4 nella frazione della membrana plasmatica dei muscoli scheletrici nel gruppo trattato con DS2006511 come osservato in un gruppo trattato con Insulina. Questi dati suggeriscono che il trattamento con DS20060511 riduce i livelli di glucosio nel sangue aumentando l’assorbimento del glucosio nel muscolo scheletrico inducendo la traslocazione di GLUT4 in vivo.
a, b Livelli di glucosio nel sangue dopo il trattamento con DS20060511 (30 mg kg-1) in topi C57BL/6 che avevano ricevuto un accesso continuo al cibo (a) e topi a cui era stato negato l’accesso al cibo durante la notte (b) (n = 8) . I valori mostrati sono mezzi ± SEM. **P < 0.01 vs. 0 min di ANOVA unidirezionale seguito dal test di Dunnett. c Livelli di glicemia e Insulina plasmatica durante GTT orale nei topi C57BL/6 (n = 5–6). I topi hanno ricevuto la somministrazione orale di veicolo o DS20060511 alla dose indicata, 15 min prima della somministrazione di glucosio (1,5 g kg-1). I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01 rispetto al veicolo per ANOVA unidirezionale seguito dal test di Williams. d Captazione di [3H]-2-DG nel muscolo soleo, nel muscolo gastrocnemio (Gastro.), nel cuore e nel tessuto adiposo bianco (WAT) a 60 min durante il GTT intraperitoneale nei topi C57BL/6 (n = 3). I topi hanno ricevuto la somministrazione orale del veicolo o DS20060511 (30 mg kg-1), 15 min prima della somministrazione di glucosio (1 g kg-1 glucosio contenente [3H]-2-DG). I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01 rispetto al veicolo in base al t-test. e Livelli proteici di GLUT4 e Na,K-ATPaseα nella frazione di membrana plasmatica del muscolo tricipite surale asportato dai topi C57BL/6 (n = 2) trattati con DS20060511 (10 mg kg-1), Insulina (5 U kg− 1), o salina come veicolo, attraverso la vena cava inferiore 10 min dopo il trattamento.
Valutazione farmacocinetica del DS20060511 nei topi:
In topi normali sono state esaminate le variazioni della concentrazione plasmatica e della distribuzione del DS20060511 in possibili organi/tessuti bersaglio. I livelli di esposizione sistemica al DS20060511 dopo sua somministrazione orale erano dose dipendenti e le concentrazioni massime a 30 min dopo la somministrazione di 1, 10 e 30 mg kg-1 erano rispettivamente di 0,6, 16,5 e 71,4 μM. La misurazione delle concentrazioni di DS20060511 nei tessuti a 75 min dopo la somministrazione orale (30 mg kg-1) ha rivelato concentrazioni quasi comparabili tra il muscolo scheletrico, il WAT e il cuore. Coerentemente con il suo profilo farmacocinetico stabile, la stabilità metabolica del composto nella frazione microsomiale del fegato era elevata (89% e 79% del composto rimanente dopo 1 h di incubazione con la frazione microsomiale del fegato umano e di topo, rispettivamente).
L’effetto ipoglicemizzante del DS20060511 dipende dal GLUT4:
Per confermare che l’effetto ipoglicemizzante del DS20060511 è mediato dal GLUT4, la molecola è stata somministrata a topi GLUT4KO. L’espressione della proteina GLUT4 non era rilevabile nel muscolo scheletrico, nel cuore e nel WAT dei topi GLUT4KO. Mentre il trattamento con DS20060511 ha causato una significativa diminuzione dei livelli di glucosio nel sangue e di Insulina plasmatica nei topi wild-type (WT) durante GTT orale, questi effetti sono stati completamente aboliti nei topi GLUT4KO. Il trattamento con DS20060511 ha aumentato significativamente l’assorbimento di 2-DG da parte dei muscoli soleo ed estensore lungo delle dita (EDL) dei topi WT, mentre non è stato osservato un tale aumento dell’assorbimento muscolare nei muscoli isolati dei topi GLUT4KO trattati con DS20060511 . Questi dati confermano che l’effetto ipoglicemizzante del DS20060511 è mediato da GLUT4 nel muscolo scheletrico.
a Livelli di glicemia e insulina plasmatica durante GTT orale in topi wild-type (WT, n = 5) e GLUT4 knockout (KO, n = 6). I topi hanno ricevuto la somministrazione orale del veicolo o DS20060511 (30 mg kg-1), 15 min prima della somministrazione di glucosio (1,5 g kg-1). I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01 rispetto al veicolo in base al t-test. b Captazione di [3H]-2-DG stimolata da DS20060511 nei muscoli soleo e EDL isolati asportati da topi WT (n = 6) e KO (n = 6). I valori mostrati sono mezzi ± SEM. **P < 0,01 rispetto al veicolo secondo il t-test.
Il trattamento con DS20060511 induce la traslocazione di GLUT4 senza attivazione delle vie IR-IRS1-PI3K-Akt-AS160 e -PI3K-Rac1:
La traslocazione di GLUT4 indotta dall’Insulina è attivata da (1) la via IR-IRS1-PI3K-Akt-AS16032 e (2) la via IR-IRS1-PI3K-Rac115 nel muscolo scheletrico. L’Insulina lega l’IR, che si traduce nell’attivazione di IRS1, PI3K e Akt. Akt attivato inibisce la proteina di attivazione della Rab GTPasi (GAP) AS160, che si traduce nell’attivazione delle proteine Rab e nella traslocazione di GLUT4 alla membrana plasmatica[33]. D’altra parte, Rac1 è attivato da PI3K e promuove il rimodellamento dell’actina, con conseguente traslocazione di GLUT4[12]. E’ stato esaminato se il trattamento con DS20060511 aumenta la traslocazione di GLUT4 nel muscolo scheletrico attraverso questi percorsi. Sebbene la subunità IRβ e IRS1 siano state fosforilate nei muscoli scheletrici dei topi trattati con Insulina, non è stata osservata tale fosforilazione di queste proteine dopo il trattamento con DS20060511. Allo stesso modo, mentre il trattamento con Insulina ha indotto la fosforilazione di Akt e AS160, il trattamento con DS20060511 non ha avuto tale effetto. Successivamente è stata eseguita la microscopia di immunofluorescenza per indagare se il DS20060511 potesse promuovere la polimerizzazione dell’actina. Sebbene sia stata osservata una forte colorazione di GLUT4 sulla superficie cellulare dopo il trattamento sia con Insulina che con DS20060511, la polimerizzazione dell’actina è stata osservata solo dopo il trattamento con Insulina nei miotubi differenziati L6-GLUT4myc. Inoltre, sebbene la traslocazione di GLUT4 sia stata indotta sia dall’Insulina che dal trattamento con DS20060511, la latrunculina B, un inibitore della polimerizzazione dell’actina, ha soppresso solo la traslocazione di GLUT4 indotta dall’Insulina, ma non quella indotta dal trattamento con DS20060511. Il co-trattamento di DS20060511 e Insulina ha comportato un aumento additivo della traslocazione di GLUT4 nei miotubi L6-GLUT4myc, anche alla concentrazione di Insulina alla quale la traslocazione di GLUT4 da parte della sola Insulina era saturata. Coerentemente con questi dati, anche l’assorbimento di 2-DG indotto dall’Insulina è stato ulteriormente aumentato dal trattamento concomitante con DS20060511 nei muscoli scheletrici isolati. In effetti, i livelli di glucosio nel sangue sono stati ridotti in misura maggiore dopo il trattamento combinato con DS20060511 più Insulina rispetto a quello dopo il solo trattamento con Insulina nei topi trattati con streptozotocina (STZ). Questi dati suggeriscono che l’attivazione né della via IR-IRS1-PI3K-Akt-AS160 né della via IR-IRS1-PI3K-Rac1 è coinvolta nella traslocazione di GLUT 4 indotta dal trattamento con DS20060511.
a, b Fosforilazione di IRβ, IRS1, Akt (Ser473) e AS160 del muscolo tricipite surale asportato da topi C57BL/6 (n = 2) trattati con DS20060511 (10 mg kg-1), Insulina (5 U kg-1 ), o soluzione salina come veicolo, attraverso la vena cava inferiore 10 min dopo il trattamento. c Immunocolorazione in fluorescenza della superficie cellulare GLUT4 e delle fibre intracellulari di actina in miotubi L6-GLUT4myc trattati con 30μM di DS20060511 o 100μnM di Insulina. Le frecce indicano la caratteristica struttura arruffata dell’actina polimerizzata e della superficie associata all’actina GLUT4. d Attività di traslocazione GLUT4 dell’Insulina 30μM DS20060511 o 100μnM in presenza dell’inibitore della polimerizzazione dell’actina, Latrunculin B, alle concentrazioni indicate. I valori mostrati sono mezzi ± SEM, n = 3. e Traslocazione GLUT4 stimolata dall’Insulina concentrazione-dipendente in miotubi L6-GLUT4myc con o senza 30 μM DS20060511 (n = 3). f Captazione di 2-DG stimolata da DS20060511 concentrazione-dipendente con Insulina 100 nM in muscoli isolati da topi C57BL/6 (n = 3). I valori mostrati sono mezzi ± SEM. ***P < 0,001 di ANOVA unidirezionale seguito dal test di Tukey. g Livelli di glucosio nel sangue durante ITT in topi C57BL/6 trattati con STZ (n = 6–7). Il veicolo o la dose indicata di DS20060511 è stata somministrata per via orale contemporaneamente all’iniezione intraperitoneale di Insulina 0,1 U kg-1. I valori mostrati sono mezzi ± SEM. *P < 0.05 vs. veicolo per ANOVA unidirezionale seguito dal test di Dunnett. c Barra della scala in tutti i pannelli, 5 μm. Le macchie non ritagliate per a e b possono essere trovate nella figura seguente.
Il trattamento con DS20060511 aumenta l’ossidazione del glucosio durante l’esercizio fisico:
Poiché l’esercizio fisico, come l’Insulina, è ben noto per migliorare la traslocazione di GLUT4 e aumentare l’assorbimento di glucosio nel muscolo scheletrico[34], i ricercatori hanno successivamente studiato l’effetto del trattamento con DS20060511 sulla capacità di resistenza all’esercizio fisico e l’ossidazione del substrato energetico durante l’esercizio mediante calorimetria. Durante l’esercizio graduale sul tapis roulant, il VO2 è aumentato gradualmente in entrambi i gruppi trattati con il veicolo e DS20060511 e anche la capacità di resistenza all’esercizio era paragonabile tra i due gruppi. Dopo un po’ di tempo dall’inizio della corsa, il gruppo trattato con DS20060511 ha iniziato a mostrare un rapporto di scambio respiratorio (RER) relativamente più elevato rispetto al gruppo trattato con veicolo; inoltre, l’ossidazione stimata del glucosio durante il test era significativamente più alta nei topi trattati con DS20060511 rispetto ai topi trattati con veicolo, mentre l’ossidazione dei grassi era significativamente inferiore. Pertanto, il DS20060511 ha aumentato l’ossidazione del glucosio durante l’esercizio. I livelli di glucosio nel sangue sono diminuiti significativamente dopo l’esercizio nei topi trattati con DS20060511, ma non sono scesi al range di ipoglicemia. I livelli di lattato nel sangue erano comparabili tra i due gruppi.
a–c Rapporto di scambio respiratorio (RER), ossidazione stimata del glucosio e ossidazione dei grassi durante la corsa su tapis roulant graduale nei topi C57BL/6 (n = 7). Il veicolo o DS20060511 (30 mg kg−1) è stato somministrato per via orale 15 min prima di iniziare a correre. Il tapis roulant è partito dalla velocità di 10 m min−1 e aumentato di 2 m min−1 ogni 3 min. I valori mostrati sono mezzi ± SEM. *P < 0.05 vs. veicolo dal t-test.
Mancanza di effetto sulla fosforilazione dell’AMPK con Il trattamento di DS20060511:
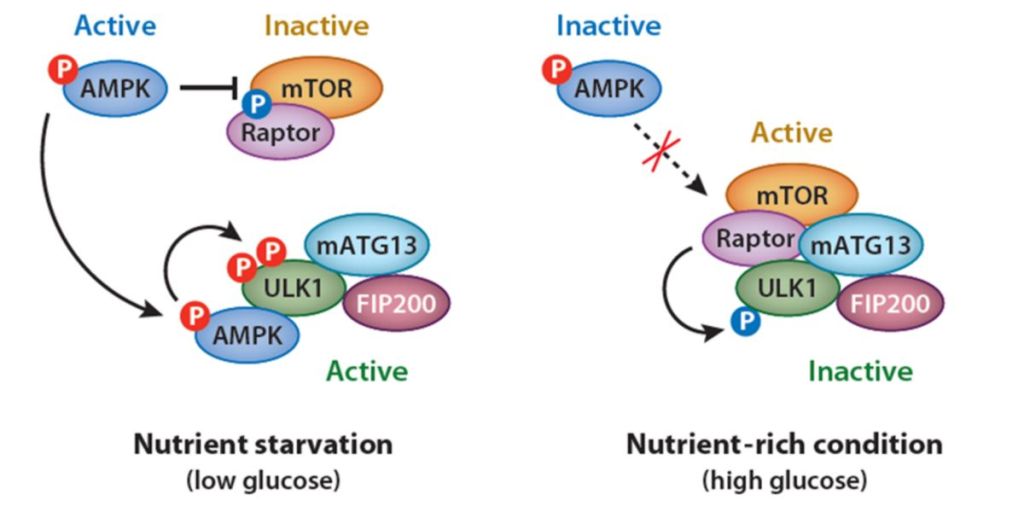
Sulla base della scoperta che il DS20060511 ha aumentato l’utilizzo del glucosio nel muscolo scheletrico durante l’esercizio, i suoi effetti combinati con quelli della contrazione muscolare sono stati ulteriormente valutati utilizzando campioni di muscolo scheletrico isolati. L’assorbimento di 2-DG è stato aumentato in misura maggiore dopo l’elettrostimolazione muscolare combinata con il trattamento DS20060511 rispetto a quello dopo l’elettrostimolazione muscolare senza il trattamento DS20060511. Sebbene recenti scoperte suggeriscano che l’AMPK non svolga alcun ruolo nella traslocazione di GLUT4 e nell’assorbimento di glucosio nel muscolo osservato durante l’esercizio[16,22], l’attivazione di AMPK mediante stimolazione elettrica[21], nonché da AICAR[20], potrebbe aumentare l’assorbimento di glucosio nel muscolo scheletrico isolato. E’ stata esaminata la fosforilazione di AMPK dopo il trattamento con DS20060511 mediante western blotting nel muscolo scheletrico isolato. Sebbene il livello di fosforilazione dell’AMPK sia stato elevato dalla stimolazione muscolare elettrica, non è stato osservato alcun cambiamento di questo tipo dopo il trattamento con DS20060511. Il livello di fosforilazione dell’AMPK nel muscolo scheletrico è rimasto invariato dopo il trattamento con DS20060511 rispetto a quello prima del trattamento in vivo, anche in condizioni di non esercizio. Questi dati suggeriscono che l’aumento dell’assorbimento di glucosio indotto da DS20060511 è indipendente dall’attivazione dell’AMPK.
a Captazione di 2-DG stimolata da DS20060511 dipendente dalla concentrazione con contrazione muscolare (stimolazione elettrica 5 Hz) in muscoli isolati da topi C57BL/6 (n = 3). ***P < 0,001 di ANOVA unidirezionale seguito dal test di Tukey. b La contrazione muscolare (stimolazione elettrica 5 Hz) ha indotto la fosforilazione di AMPK (Thr172) con o senza 10 μM DS20060511 in muscoli isolati da topi C57BL/6. c Livelli di fosforilazione di AMPKα dei muscoli Triceps surae asportati da topi C57BL/6 (n = 2) trattati con DS20060511 (10 mg kg-1) o soluzione salina come veicolo attraverso la vena cava inferiore 10 min dopo il trattamento.
Il trattamento con DS20060511 diminuisce la glicemia in maniera eNOS-indipendente:
È stato dimostrato che il Nitroprussiato di sodio (SNP), un donatore di Ossido Nitrico (NO), aumenta l’assorbimento di glucosio nel muscolo scheletrico e che questo aumento non è inibito dall’inibitore PI3K, Wortmannin[35]. Inoltre, l’assorbimento del glucosio indotto dall’esercizio da parte del muscolo scheletrico non è stato soppresso dall’inibitore di NO NG-monometil-L-arginina (L-NMMA)[35]. Questi dati suggeriscono che il NO induce l’assorbimento del glucosio da parte del muscolo scheletrico attraverso un meccanismo che è distinto sia dall’Insulina che dalle vie di segnalazione dell’esercizio. L’Ossido Nitrico sintasi endoteliale, che è un importante enzima che genera NO, è espresso nel muscolo scheletrico. È stato riportato che l’assorbimento del glucosio è compromesso nei muscoli scheletrici isolati di topi eNOSKO[36]. Per studiare il meccanismo alla base dell’aumento dell’assorbimento di glucosio da parte del muscolo scheletrico indotto da DS20060511, è stato somministrato DS20060511 a topi eNOSKO. Il trattamento con DS20060511 ha ridotto significativamente i livelli di glucosio nel sangue sia nei topi WT che eNOSKO durante GTT orale. Sebbene i livelli di glucosio nel sangue siano stati ridotti dal trattamento con Insulina, i livelli di glucosio nel sangue sono stati ridotti ulteriormente dopo il trattamento con DS20060511, sia nei topi WT che eNOSKO, allo stesso modo. Questi dati suggeriscono che l’effetto ipoglicemizzante di DS20060511 è esercitato in modo eNOS-indipendente.
a, b Livelli di glucosio nel sangue durante GTT orale in topi wild-type (WT, n = 5) ed eNOS-knockout (KO, n = 5–6). I topi hanno ricevuto il veicolo o DS20060511 (10 mg kg-1) per via orale 15 min prima della somministrazione di glucosio (3,0 g kg-1). c, d Livelli di glucosio nel sangue durante ITT nei topi WT (n = 4) e KO (n = 5). Veicolo o DS20060511 (30 mg kg-1) è stato somministrato per via orale contemporaneamente all’iniezione intraperitoneale di insulina 0,5 U kg-1. I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 rispetto al veicolo in base al t-test.
Il trattamento acuto e cronico con DS20060511 migliora l’intolleranza al glucosio nei topi diabetici obesi:
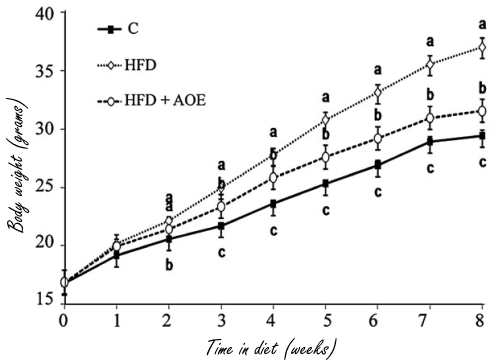
Per indagare se il trattamento con DS20060511 può attenuare l’intolleranza al glucosio nei topi con obesità indotta dalla dieta e resistenza all’Insulina, i ricercatori hanno condotto GTT orale in topi alimentati con dieta ricca di grassi (HFD) dopo il trattamento con DS20060511. Il trattamento con DS20060511 ha ridotto significativamente i livelli di glucosio nel sangue nei topi nutriti con HFD agli stessi livelli di quelli osservati nei topi alimentati con dieta normale durante il GTT orale. I livelli plasmatici di Insulina erano piuttosto diminuiti nei topi nutriti con HFD trattati con DS20060511. La soppressione dell’assorbimento di 2-DG indotto dall’Insulina nel muscolo scheletrico isolato da topi alimentati con HFD è stata completamente ripristinata dal trattamento con DS20060511. Questi dati suggeriscono che il trattamento acuto con DS20060511 migliora l’intolleranza al glucosio nei topi con obesità indotta dalla dieta e resistenza all’Insulina. Successivamente, è stato studiato l’effetto del trattamento cronico con DS20060511 in topi diabetici geneticamente obesi (db/db). I livelli di glucosio nel sangue sono diminuiti significativamente dal primo al 28° giorno di trattamento con DS20060511 nei topi db/db. Coerentemente con questi dati, anche il valore dell’emoglobina glicata (HbA1c) è stato significativamente ridotto dopo il trattamento cronico con DS20060511. Non ci sono state differenze statisticamente significative nel peso corporeo, nell’assunzione di cibo, nel livello di glucosio nel sangue a digiuno o nei livelli di Insulina plasmatica a digiuno tra i topi db/db trattati con DS20060511 e quelli trattati con il veicolo. Non sono stati inoltre rilevati cambiamenti significativi nei pesi dei tessuti di muscolo, cuore, WAT e fegato, o nel contenuto di glicogeno del muscolo, del cuore e del fegato. Questi dati suggeriscono che il trattamento con DS20060511 sia acuto che cronico migliora il diabete ripristinando l’assorbimento alterato del glucosio da parte del muscolo scheletrico.
a Livelli di glucosio nel sangue e di Insulina plasmatica durante un GTT orale in topi alimentati con dieta normale (NC) e ad alto contenuto di grassi (HFD) (n = 5). Veicolo o DS20060511 (30 mg kg-1) è stato somministrato per via orale 15 min prima della somministrazione orale di glucosio (1,5 g kg-1). I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 rispetto al veicolo HFD secondo il t-test. b Effetti dell’Insulina 10 μM DS20060511 e 100 nM sull’assorbimento di 2-DG nei muscoli isolati da topi alimentati con NC (n = 6) e con HFD (n = 5). I valori mostrati sono mezzi ± SEM. **P < 0.01 per ANOVA unidirezionale seguito dal test di Tukey. c, d Cambiamenti nei livelli di glucosio nel sangue e AUC il giorno 1 e il giorno 28 durante la rialimentazione (n = 6) in topi db/db trattati cronicamente con DS20060511 (10 mg kg-1 giorno-1). I valori mostrati sono mezzi ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001 rispetto al veicolo in base al t-test. e Modifica dei livelli di HbA1c nei topi db/db (n = 6) a 4 settimane. I valori mostrati sono mezzi ± SEM. *P < 0.05 vs. veicolo dal t-test.
Discussioni conclusive:
Come abbiamo visto, è stata passata al vaglio la libreria chimica in possesso dei ricercatori i quali hanno utilizzato miotubi L6-GLUT4myc per lo studio di un nuovo farmaco per il trattamento del diabete di tipo II scoprendo il composto xantenico DS20060511. Il DS20060511 ha aumentato la traslocazione di GLUT4 nei miotubi differenziati L6-GLUT4myc, ma non negli adipociti differenziati 3T3-L1-GLUT4myc, suggerendo che agisca principalmente nei muscoli scheletrici. Coerentemente, in vivo, il DS20060511 ha indotto l’assorbimento di 2-DG nei muscoli soleo e gastrocnemio, ma non nel cuore o nel tessuto adiposo. L’Insulina favorisce l’assorbimento del glucosio nel tessuto adiposo e nel muscolo scheletrico, che inevitabilmente, in condizioni metabolicamente alterate e ipercaloriche, porta all’obesità. Tuttavia, il DS20060511 migliora l’assorbimento del glucosio solo nel muscolo scheletrico e riduce la secrezione di Insulina sopprimendo l’aumento dei livelli di glucosio nel sangue dopo il carico di glucosio, sopprimendo così lo sviluppo dell’obesità; quindi, il composto sembra anche offrire una promessa come farmaco per la prevenzione dell’obesità. Il DS20060511 ha ridotto i livelli di glucosio nel sangue nei topi diabetici obesi, senza causare iperfagia, aumento di peso corporeo o ipoglicemia e senza aumentare la secrezione di Insulina. Inoltre, il DS20060511 non sembra abbassare il livello di glucosio nel sangue a digiuno, indicando il rischio relativamente basso di ipoglicemia associato all’uso di questo composto. Queste caratteristiche potrebbero essere preferibili a un farmaco sicuro ed efficace per il trattamento del diabete di tipo II.
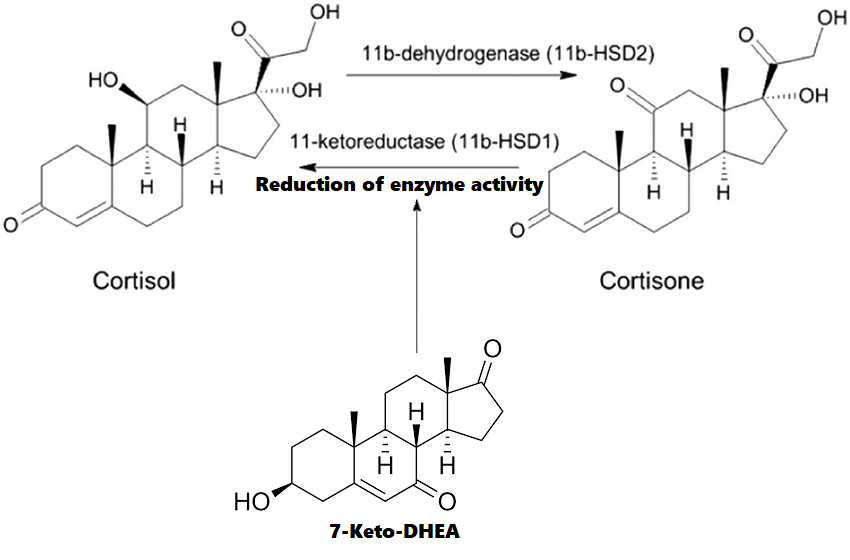
Voglio ricordare che con la sigla “2-DG” ci si riferisce ad un analogo del glucosio tracciabile (vedi immagine a sinistra).
L’effetto ipoglicemizzante del DS20060511 è stato completamente abolito nei topi GLUT4KO, indicando che il DS20060511 aumenta l’assorbimento del glucosio in modo GLUT4-dipendente. È interessante notare che il DS20060511 non è riuscito ad attivare la segnalazione dell’Insulina a monte, inclusa la fosforilazione di AS160 e il rimodellamento dell’actina o il percorso AMPK, che sono anche noti per aumentare la traslocazione di GLUT4 nel muscolo scheletrico. Inoltre, quando somministrato in combinazione con Insulina, il DS20060511 ha ulteriormente migliorato l’assorbimento del glucosio nel muscolo scheletrico sia nei topi normali che in quelli resistenti all’Insulina e ha ulteriormente ridotto i livelli di glucosio nel sangue in un modello murino di diabete di tipo I indotto da STZ. Il DS20060511 ha anche potenziato l’ossidazione del glucosio in tutto il corpo durante l’esercizio fisico, associata a un aumento dell’assorbimento e dell’utilizzo del glucosio nel muscolo scheletrico[16]. Pertanto, il DS20060511 può agire come un agente antidiabetico con un meccanismo d’azione completamente nuovo in pazienti con azioni alterate dell’Insulina nel muscolo scheletrico e in quelli con diabete di tipo I o II che ricevono Insulina e/o terapia fisica.
Alcuni composti sono stati anche segnalati in precedenza per indurre la traslocazione di GLUT4. È stato riportato che nuovi composti della Piridazina inducono fortemente la traslocazione di GLUT4 nei miotubi L6 e mostrano un significativo effetto ipoglicemizzante in un modello murino di diabete grave[37]. È noto che i disaccoppianti protonici, come il 2,4-dinitrofenolo, inducono la traslocazione di GLUT4 in accordo con un rapido calo dei livelli intracellulari di ATP[38]. Tuttavia, a differenza del DS20060511, questi composti promuovono la traslocazione di GLUT4 attraverso la via PI3K o AMPK. È stato riportato che la piccola molecola donatrice di NO NCX 4016 induce la traslocazione di GLUT4 negli adipociti 3T3-L1, ma non nelle cellule del muscolo scheletrico[39]. Questi risultati suggeriscono che un potenziatore della traslocazione di GLUT4 specifico del muscolo scheletrico come il DS20060511 non è mai stato segnalato in precedenza.
Il movimento del GLUT4 negli adipociti. Il tessuto adiposo è costituito da adipociti. Negli adipociti, il GLUT4 si trova nella membrana cellulare e nel citosol. La traslocazione di GLUT4 dalle vescicole citosoliche alla membrana cellulare porta ad un elevato assorbimento di glucosio, mentre l’endocitosi riporta il GLUT4 al citosol. (1): Nelle cellule non stimolate, le porzioni di membrana contenenti GLUT4 sono internalizzate in modo endocitosi per generare vescicole contenenti GLUT4. Le vescicole GLUT4 sono internalizzate negli endosomi precoci (o ordinati). Possono entrare nel corpo endoplasmatico di recupero e seguire la via retrograda verso la rete trans-Golgi e il compartimento intermedio del reticolo endoplasmatico-Golgi o altri compartimenti della membrana donatrice. (2): Le vescicole GLUT4 derivate dalle strutture della membrana del donatore sono fissate da un laccio contenente un dominio UBX per la proteina GLUT4 (TUG). (3): Durante la stimolazione del segnale dell’Insulina, le vescicole GLUT4 vengono rilasciate e caricate sul motore dei microtubuli per essere trasferite alla membrana plasmatica. La continua presenza di Insulina porta al movimento diretto di queste vescicole verso la membrana plasmatica. (4): Le vescicole GLUT4 sono legate alla proteina motoria chinesina e ad altre proteine. Quando ciò si verifica, si forma un complesso SNARE ternario stabile. (5): Il complesso SNARE ternario stabile è ancorato alla membrana bersaglio. (6): Le vescicole ancorate si affidano a SNARE per spostarsi e fondersi con la membrana bersaglio. Fonte immagine: Wang T, Wang J, Hu X, Huang XJ, Chen GX. Current understanding of glucose transporter 4 expression and functional mechanisms. World J Biol Chem 2020; 11(3): 76-98 [PMID: 33274014 DOI: 10.4331/wjbc.v11.i3.76]
Perché il DS2006051 agisce selettivamente sul muscolo scheletrico? La molecola bersaglio del DS2006051 può essere espressa selettivamente nel muscolo scheletrico. La quantità di GLUT4 sulla superficie cellulare è determinata dall’equilibrio tra esocitosi dalle vescicole di stoccaggio intracellulare ed endocitosi dalla membrana cellulare. Il DS2006051 può promuovere l’esocitosi o sopprimere l’endocitosi di GLUT4 tramite l’attivazione della molecola bersaglio. Per studiare il bersaglio selettivo di DS20060511 nel muscolo scheletrico e nei miotubi L6, sono stati adottati tre diversi approcci: legame del composto radiomarcato, purificazione di perline immobilizzate con composto e fotoreticolazione UV di un composto al bersaglio. I composti radiomarcati o modificati chimicamente avevano la capacità di reagire con campioni preparati da tessuto muscolare scheletrico o miotubi L6-GLUT4myc, come lisati, microsomi o cellule viventi. Dopo l’arricchimento e la purificazione abbinati per ciascun approccio, i campioni sono stati analizzati mediante LC-MS/MS. Sfortunatamente, i ricercatori non sono riusciti a identificare nessuna molecola bersaglio specifica che si legasse al DS20060511 con un’alta affinità. Sono necessarie ulteriori indagini per identificare il bersaglio molecolare del DS20060511 e anche la via di segnalazione coinvolta, come la produzione di specie reattive dell’ossigeno associate a Rac1 o NADPH ossidasi 2.
In conclusione, è stato identificato un nuovo composto xantenico, il DS20060511, ed è stato dimostrato che il trattamento con DS20060511 induceva la traslocazione di GLUT4 indipendentemente dalla segnalazione canonica dell’Insulina e dall’attività dell’AMPK, per migliorare l’assorbimento del glucosio da parte del muscolo scheletrico. Inoltre, il trattamento con DS20060511 ha anche migliorato l’intolleranza al glucosio nei topi diabetici obesi. Sebbene non siano stati in grado di identificare la specifica molecola bersaglio del DS20060511 sulla cellula muscolare scheletrica, ulteriori studi con il composto aiuterebbero a sviluppare un nuovo farmaco per il diabete di tipo II.
Le caratteristiche del DS20060511 lo rendono una molecola di particolare interesse per i bodybuilder. La sua selettività per il tessuto muscolo scheletrico e la mancata attivazione dell’AMPK offrono due significativi vantaggi che le molecole con attività di miglioramento del insulino-resistenza (Biguanidi et simili) oggi disponibili non danno:
Punto 1: la selettività della molecola per il tessuto muscolo-scheletrico e il miglioramento in tale sede dell’uptake del Glucosio da parte del miocita garantisce una ripartizione calorica a sensibile svantaggio del tessuto adiposo (quindi dell’adipocita) in un contesto ipercalorico, prolungando in modo indeterminato (almeno secondo i dati attuali) il periodo di vantaggio che l’atleta può sperimentare in un regime di questo tipo, prima che il peggioramento dei parametri del IR portino ad un aumento significativo della massa grassa e una riduzione dei guadagni muscolari sia in rapporto alla precedente che in termini assoluti;
Punto 2: la capacità del DS20060511 di bypassare l’attivazione/stimolo del AMPK permette di non sottoregolare/bloccare l’attività del mTOR e della sua attività sull’ipertrofia muscolare. Questo vantaggio è unico nel suo genere dal momento che, per esempio, sia la Metformina che la Berberina, due molecole largamente utilizzate per il miglioramento del IR, interagiscono per via delle PPAR-α nello stimolo dell’attività del AMPK la quale sottoregola/blocca mTOR.
Riguardo all’ultimo punto, c’è da dire che, da quanto osservato empiricamente ed emerso clinicamente, l’interazione negativa di Metformina e Berberina sul mTOR risulta significativa in modo dosaggio-dipendente. Si ipotizza, ma questa è una semplice ipotesi osservazionale, che l’uso di dosaggi non superiori a 500-750mg/die totali di entrambe le molecole non alteri crescita e/o recupero muscolare. Ricordiamoci inoltre che sia la Metformina che la Berberina (compreso anche l’ALA) sembrano avere potenziali inibitori sugli enzimi implicati nella lipogenesi ed esterificazione degli acidi grassi liberi negli adipociti, ma questa è un altra storia.
È interessante notare che alcuni studi dell’ultimo decennio suggeriscono che la Metformina può inibire direttamente l’azione della Leucina sul mTOR. Non solo questo sarebbe, ovviamente, un fattore negativo per la crescita muscolare, ma ipoteticamente l’effetto inibitorio della Metformina sul mTOR potrebbe avere un effetto maggiore in quanto è correlato alla riduzione del rischio di tumori mortali nei diabetici.
E’ a proposito molto interessante quanto postulato dal Dr. Melnik dell’Università di Osnabrück in Germania: “la Metformina può essere un diretto concorrente della Leucina per il legame e l’attività del mTORC1”.
Il medico ha notato nel suo articolo che la dose giornaliera abituale nei diabetici di Metformina (2g) è nell’ordine dei 2g di Leucina derivati dal consumo giornaliero di 100g di carne o formaggio. Poiché le due molecole sono simili per struttura e dimensioni, possono competere per gli stessi siti nell’attivazione del mTOR. Di conseguenza, possiamo affermare, con un buon margine di ragione, che è una questione “dose-risposta dipendente”, come accennato in precedenza, in rapporto all’attività potenziale di alterazione del mTOR sia diretta (legame attivazione leucina-simile) che indiretta (via AMPK).
Per quanto riguarda la questione della potenziale sotto-regolazione sui AR da parte della Metformina, i dati attuali provengono principalmente da studi di linee cellulari in vitro, in donne con PCOS, e da studi sui pazienti con cancro alla Prostata che però non danno comunque dati chiari sul grado di riduzione dei AR a livello del muscolo-scheletrico, di conseguenza si può speculare ancora ampiamente su quali possano essere gli effetti in vivo nell’uomo sulla crescita del tessuto muscolo-scheletrico durante il trattamento con Metformina. Rimango, al momento, dell’idea che sia fondamentalmente una questione di “soglia di efficacia” in rapporto agli “effetti indesiderati”, e la cosa, però, non è così semplice da calibrare come sembra viste le diversità nelle risposte individuali.
Ma, tornando a parlare del DS20060511, potrebbe avere un potenziale anche in un regime ipocalorico? Si, ovviamente, anche se presumibilmente il calo della Leptina sarà più rapido per via della “carestia glucidica adipocitaria indotta”. Sicuramente risulterebbe un vantaggio nei refeed sia pre-contest che quelli di “routine” settimanale. La superiorità rispetto a quanto oggi utilizzato con tali finalità rimane.
Per il momento, non ci resta che attendere nuovi studi sul DS20060511, possibilmente sull’uomo.
Gabriel Bellizzi
Riferimenti:
Kahn, B. B., Rossetti, L., Lodish, H. F. & Charron, M. J. Decreased in vivo glucose uptake but normal expression of GLUT1 and GLUT4 in skeletal muscle of diabetic rats. J. Clin. Invest. 87, 2197–2206 (1991).
Wallberg-Henriksson, H. & Zierath, J. R. GLUT4: a key player regulating glucose homeostasis? Insights from transgenic and knockout mice (review). Mol. Membr. Biol. 18, 205–211 (2001).
Ryder, J. W. et al. Use of a novel impermeable biotinylated photolabeling reagent to assess insulin- and hypoxia-stimulated cell surface GLUT4 content in skeletal muscle from type 2 diabetic patients. Diabetes 49, 647–654 (2000).
Liu, M. L. et al. Transgenic mice expressing the human GLUT4/muscle-fat facilitative glucose transporter protein exhibit efficient glycemic control. Proc. Natl Acad. Sci. USA 90, 11346–11350 (1993).
Gibbs, E. M. et al. Glycemic improvement in diabetic db/db mice by overexpression of the human insulin-regulatable glucose transporter (GLUT4). J. Clin. Invest. 95, 1512–1518 (1995).
Ren, J. M. et al. Overexpression of Glut4 protein in muscle increases basal and insulin-stimulated whole body glucose disposal in conscious mice. J. Clin. Invest. 95, 429–432 (1995).
Huang, S. & Czech, M. P. The GLUT4 glucose transporter. Cell. Metab. 5, 237–252 (2007).
Bryant, N. J., Govers, R. & James, D. E. Regulated transport of the glucose transporter GLUT4. Nat. Rev. Mol. Cell. Biol. 3, 267–277 (2002).
Kubota, T., Kubota, N. & Kadowaki, T. Imbalanced insulin actions in obesity and type 2 diabetes: key mouse models of insulin signaling pathway. Cell. Metab. 25, 797–810 (2017).
Kubota, N. et al. Dynamic functional relay between insulin receptor substrate 1 and 2 in hepatic insulin signaling during fasting and feeding. Cell. Metab. 8, 49–64 (2008).
Bhuin, T. & Roy, J. K. Rab proteins: the key regulators of intracellular vesicle transport. Exp. Cell. Res. 328, 1–19 (2014).
Chiu, T. T., Jensen, T. E., Sylow, L., Richter, E. A. & Klip, A. Rac1 signalling towards GLUT4/glucose uptake in skeletal muscle. Cell. Signal. 23, 1546–1554 (2011).
Khayat, Z. A., Tong, P., Yaworsky, K., Bloch, R. J. & Klip, A. Insulin-induced actin filament remodeling colocalizes actin with phosphatidylinositol 3-kinase and GLUT4 in L6 myotubes. J. Cell. Sci. 113, 279–290 (2000).
Sano, H. et al. Insulin-stimulated phosphorylation of a Rab GTPase-activating protein regulates GLUT4 translocation. J. Biol. Chem. 278, 14599–14602 (2003).
JeBailey, L. et al. Ceramide- and oxidant-induced insulin resistance involve loss of insulin-dependent Rac-activation and actin remodeling in muscle cells. Diabetes 56, 394–403 (2007).
Sylow, L., Kleinert, M., Richter, E. A. & Jensen, T. E. Exercise-stimulated glucose uptake—regulation and implications for glycaemic control. Nat. Rev. Endocrinol. 13, 133–148 (2017).
Hirshman, M. F., Wallberg-Henriksson, H., Wardzala, L. J., Horton, E. D. & Horton, E. S. Acute exercise increases the number of plasma membrane glucose transporters in rat skeletal muscle. FEBS Lett. 238, 235–239 (1988).
Goodyear, L. J., Hirshman, M. F. & Horton, E. S. Exercise-induced translocation of skeletal muscle glucose transporters. Am. J. Physiol. 261, E795–E799 (1991).
Vichaiwong, K. et al. Contraction regulates site-specific phosphorylation of TBC1D1 in skeletal muscle. Biochem. J. 431, 311–320 (2010).
Merrill, G. F., Kurth, E. J., Hardie, D. G. & Winder, W. W. AICA riboside increases AMP-activated protein kinase, fatty acid oxidation, and glucose uptake in rat muscle. Am. J. Physiol. 273, E1107–E1112 (1997).
Sylow, L. et al. Rac1 and AMPK account for the majority of muscle glucose uptake stimulated by ex vivo contraction but not in vivo exercise. Diabetes 66, 1548–1559 (2017).
McConell, G. K. It’s well and truly time to stop stating that AMPK regulates glucose uptake and fat oxidation during exercise. Am. J. Physiol. Endocrinol. Metab. 318, E564–E567 (2020).
Henríquez-Olguin, C. et al. Cytosolic ROS production by NADPH oxidase 2 regulates muscle glucose uptake during exercise. Nat. Commun. 10, 4623 (2019).
Sylow, L. et al. Rac1 governs exercise-stimulated glucose uptake in skeletal muscle through regulation of GLUT4 translocation in mice. J. Physiol. 594, 4997–5008 (2016).
Goodyear, L. J., Giorgino, F., Balon, T. W., Condorelli, G. & Smith, R. J. Effects of contractile activity on tyrosine phosphoproteins and PI 3-kinase activity in rat skeletal muscle. Am. J. Physiol. 268, E987–E995 (1995).
Yeh, J. I., Gulve, E. A., Rameh, L. & Birnbaum, M. J. The effects of wortmannin on rat skeletal muscle. Dissociation of signaling pathways for insulin- and contraction-activated hexose transport. J. Biol. Chem. 270, 2107–2111 (1995).
Lund, S., Holman, G. D., Schmitz, O. & Pedersen, O. Contraction stimulates translocation of glucose transporter GLUT4 in skeletal muscle through a mechanism distinct from that of insulin. Proc. Natl Acad. Sci. USA 92, 5817–5821 (1995).
Krook, A. et al. Characterization of signal transduction and glucose transport in skeletal muscle from type 2 diabetic patients. Diabetes 49, 284–292 (2000).
Kim, Y. B., Nikoulina, S. E., Ciaraldi, T. P., Henry, R. R. & Kahn, B. B. Normal insulin-dependent activation of Akt/protein kinase B, with diminished activation of phosphoinositide 3-kinase, in muscle in type 2 diabetes. J. Clin. Invest. 104, 733–741 (1999).
Kennedy, J. W. et al. Acute exercise induces GLUT4 translocation in skeletal muscle of normal human subjects and subjects with type 2 diabetes. Diabetes 48, 1192–1197 (1999).
Martin, I. K., Katz, A. & Wahren, J. Splanchnic and muscle metabolism during exercise in NIDDM patients. Am. J. Physiol. 269, E583–E590 (1995).
Kramer, H. F. et al. Distinct signals regulate AS160 phosphorylation in response to insulin, AICAR, and contraction in mouse skeletal muscle. Diabetes 55, 2067–2076 (2006).
Jaldin-Fincati, J. R., Pavarotti, M., Frendo-Cumbo, S., Bilan, P. J. & Klip, A. Update on GLUT4 vesicle traffic: a cornerstone of insulin action. Trends Endocrinol. Metab. 28, 597–611 (2017).
Richter, E. A. & Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 93, 993–1017 (2013).
Higaki, Y., Hirshman, M. F., Fujii, N. & Goodyear, L. J. Nitric oxide increases glucose uptake through a mechanism that is distinct from the insulin and contraction pathways in rat skeletal muscle. Diabetes 50, 241–247 (2001).
Duplain, H. et al. Insulin resistance, hyperlipidemia, and hypertension in mice lacking endothelial nitric oxide synthase. Circulation104, 342–345 (2001).
37.Tsuji, T. et al. Discovery of novel pyridazine derivatives as glucose transporter type 4 (GLUT4) translocation activators. Bioorg. Med. Chem. Lett.29, 1785–1790 (2019).
38.Klip, A., Schertzer, J. D., Bilan, P. J., Thong, F. & Antonescu, C. Regulation of glucose transporter 4 traffic by energy deprivation from mitochondrial compromise. Acta Physiol. (Oxf.).196, 27–35 (2009).
Kaddai, V. et al. The nitric oxide-donating derivative of acetylsalicylic acid, NCX 4016, stimulates glucose transport and glucose transporters translocation in 3T3-L1 adipocytes. Am. J. Physiol. Endocrinol. Metab.295, E162–E169 (2008).
Se si domandasse al bodybuilder nella media quale sia l’oggetto di maggiore interesse nella sua programmazione per il “Bulk” quasi sicuramente, dopo aver detto “raggiungere la massima ipertrofia”, direbbe “ridurre al minimo l’aumento della massa grassa”. Ed è più che logico visto che l’atleta con un minimo di senso logico sa che un peggioramento marcato della “bf” in una fase ipercalorica si tradurrà mediamente in un tempo generalmente più lungo in ipocalorica con restrizioni caloriche (o output calorici) più elevate ed un rischio relativamente aumentato di veder persa più massa contrattile o, peggio ancora, il dover calcare con i PEDs per rientrare nei tempi della preparazione al contest (o a qualsiasi evento dove il soggetto vuole essere nella sua “Top Condition”).
Tralasciando la pratica della “Break Diet”, altamente funzionale se gestita correttamente, esiste una strategia alimentare piuttosto datata ma, complici anche “neo-trafilettari” impomatati, riemersa recentemente, ovvero seguire un regime alimentare “High Fat” nel periodo di “Bulk”.
Siamo soliti collegare le diete “Low Carbs” e “High Fat” nel periodo di restrizione calorica, sapendo benissimo che il vantaggio assoluto esiste solo e soltanto con pratiche supplementative mirate (PEDs), parlando sempre di atleti e non di soggetti in sovrappeso o obesi che inizialmente giovano nel seguire una dieta Chetogenica o iperproteica in quanto a compliance insulino-resistenza correlata. Ma in ipercalorica? Possiamo veramente aspettarci una qualità migliore in quanto aumento della massa muscolare ed il peso guadagnato rispetto ad una dieta con prevalenza glucidica?
Con questo articolo cercherò di dare una risposta la più oggettiva possibile avvalendomi della conoscenza scientifica in nostro possesso che riguarda il metabolismo lipidico, l’impatto dei lipidi sulla massa muscolo-scheletrica, sulle prestazioni e i raffronti con il metabolismo glucidico partendo però dalla storia pratica della metodica alimentare qui trattata…
Back to the Future: Mauro Di Pasquale e la sua “Soluzione Anabolica”
Chi è che non ricorda Mauro Di Pasquale e il suo (a detta dell’autore) “Santo Graal” delle diete, “La Dieta Metabolica”? E la sua versione per i Bodybuilder ” La Soluzione Anabolica”? Immagino che molti di voi conosceranno entrambi i libri e i concetti viziati da bias in essi contenuti.
Visto l’argomento partirò proprio da qui…
Per chi non lo sapesse, Mauro G. Di Pasquale è un Powerlifter campione del mondo, autore di articoli sul bodybuilding e opinionista. Di Pasquale è stato assistente professore all’Università di Toronto dal 1988 al 1998. Ha tenuto conferenze e fatto ricerche sulle prestazioni atletiche, sugli integratori alimentari e sull’uso di farmaci nello sport. Ha conseguito una laurea con lode in scienze biologiche, specializzandosi in biochimica molecolare (1968) e ha una laurea in medicina (1971) – entrambe presso l’Università di Toronto. Di Pasquale è certificato come Medical Review Officer (MRO) dal Medical Review Officer Certification Council (MROCC). Era il MRO per la National Association for Stock Car Auto Racing (NASCAR). Dal 1997 al 1999 Di Pasquale si è occupato di redazione, ricerca e sviluppo di prodotti per le Scienze Sperimentali e Applicate (EAS). Come autore di bodybuilding, Di Pasquale ha scritto migliaia di articoli per molte grandi riviste di bodybuilding e fitness come Muscle & Fitness e Iron Man;[1-4] I suoi articoli e libri sono stati tradotti in lingua italiana e pubblicati in Italia da Sandro Ciccarelli per la rivista Olympian’s News.
Di Pasquale divenne anche un oppositore all’uso del doping pubblicizzando i suoi regimi alimentari e integrativi come “sostitutivi salutari” dei farmaci per il miglioramento delle prestazioni.
Da destra: Mauro di Pasquale insieme al bodybuilder professionista Eddie Robinson.
Nonostante la titolatura sopra esposta, Di Pasquale divulgò alcune teorie decisamente opinabili abbracciando la filosofia delle diete “Low Carb” come soluzione universale per perdere grasso e aumentare la massa muscolare. Egli rappresenta una sorta di “paladino” per la fazione dei sostenitori della così detta “ipotesi dell’Insulina”, tra l’altro smentita scientificamente, la quale ipotizza che non sia l’eccesso energetico (calorie) a causare l’aumento di peso/grasso ma i Carboidrati ed il loro stimolo sull’Insulina. Peccato però che la termodinamica ci dimostri il contrario, ovvero che è l’eccesso energetico ha causare una conservazione dello stesso e non il semplice stimolo ormonale (sul quale, tra l’altro, ci sarebbe molto da dire). Oltretutto, alcune fonti proteiche (vedi prodotti lattiero caseari) hanno un impatto insulinico maggiore del pane bianco, sebbene questo venga regolato da una concomitante risposta del Glucagone. Comunque sia, di quest’ultimo punto, il Di Pasquale sembra non curarsene più di tanto, similmente a quanto fa della risposta insulinica data dalla coingestione di grassi e proteine preoccupandosi del carico glucidico da ciclicizzare (cosa, quest’ultima, condivisibile ma da contestualizzare).
Ma arriviamo al dunque sul suo approccio “High Fat” in Bulk…
Nel 1995 pubblica il libro “The Anabolic Diet“, pubblicato poi in Italia con il nome di “La Soluzione Anabolica”.
Il libro in questione espone le vedute dell’autore, accuratamente servite per vendere il prodotto e convincere il lettore, dividendo in sei fasi la preparazione:
Fase di Inizio;
Fase di Massa;
Fase di Forza;
Fase di Definizione;
Fase Pregara;
Fase di Recupero.
Tralasciando le fasi 1, 3,4,5 e 6 che, per argomento trattato nel presente articolo, non ci interessano, concentriamoci sulla “Fase di Massa“.
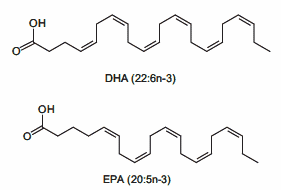
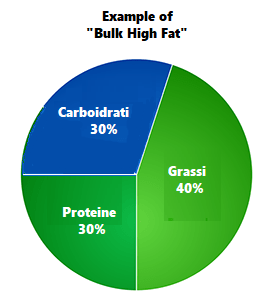
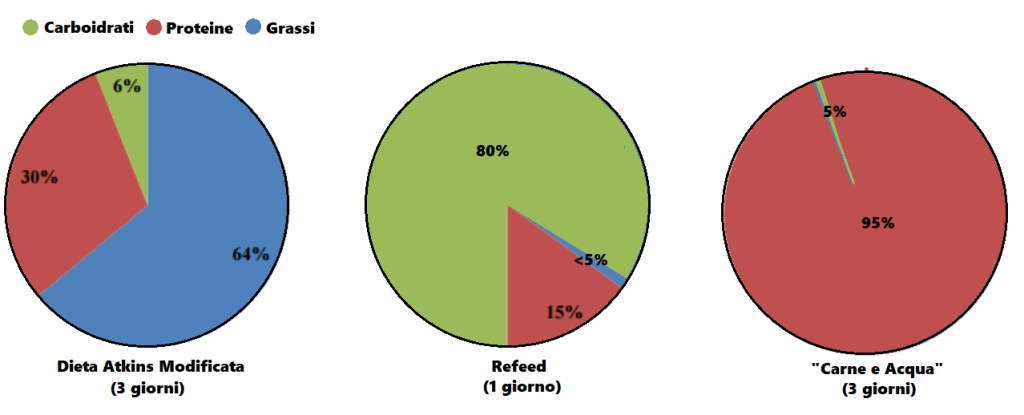
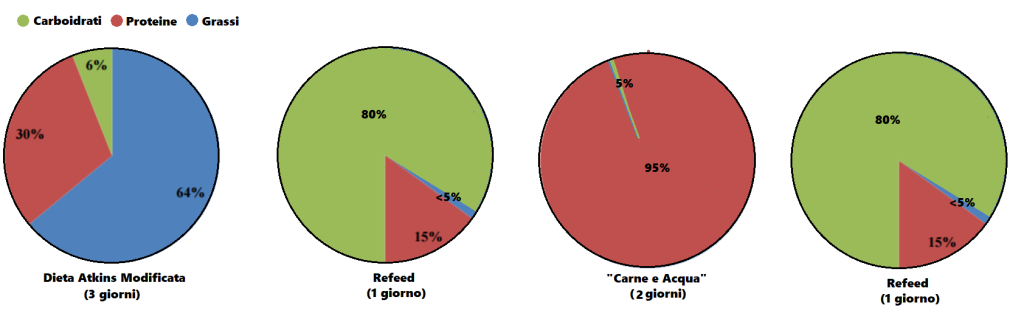
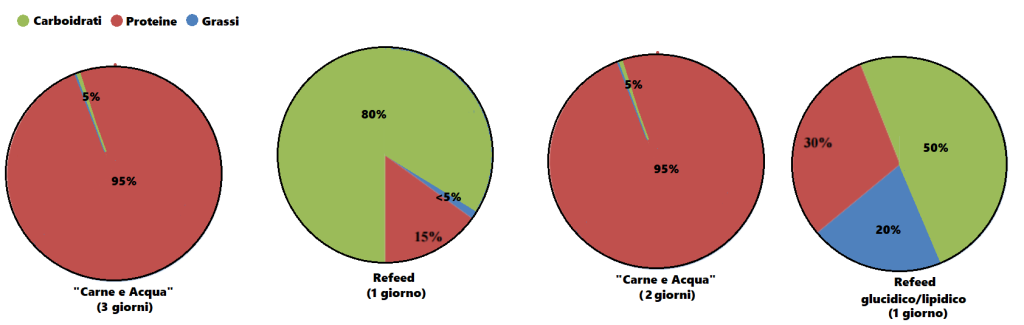
Dopo aver trovato la quota calorica di “mantenimento”, procedendo con i primi 12 giorni sostituendo le kcal tipicamente consumate dai Carboidrati con Grassi e Proteine riducendo significativamente i primi (es. 1g/CHO-Pro = 4Kcal; 1g Fat = 9Kcal; 400g di CHO/120g di Pro/50g di Fat = circa 30g di CHO, 220g/die di Pro e 134g/die di Grassi), si dovrebbe passare alla prima “ricarica di Carboidrati” seguita a sua volta da 5-6 giorni “High Fat” e 1 o 2 giorni “High Carbs” per 3-4 settimane.
A questo punto l’autore consiglia di permettere al peso un aumento del 15% oltre il peso ideale (es. Peso ideale = 98Kg, peso da raggiungere = 113Kg [+15%]) premurandosi di avvisare il lettore che non deve commettere l’errore di mangiare eccessivamente e vanificare il potenziale della sua strategia di farvi aumentare di massa magra con poco incremento della massa grassa…
Quindi, il bodybuilder dovrebbe seguire lo schema “High Fat” nei giorni feriali ed effettuare le “ricariche” glucidiche nel fine settimana, con un monte calorico indicativo pari a 55Kcal per Kg di peso corporeo desiderato (es. 90Kg = 4950Kcal/die). E le differenze di risposta delle vie metaboliche? Capacità di gestione delle Kcal connesse alle richieste prestative e al livello dell’atleta?… Punti non pervenuti… Per lo meno, il Di Pasquale dice che se il soggetto presenta difficoltà nell’ingestione di grossi quantitativi calorici giornalieri può far diventare il totale calorico da raggiungere un obbiettivo settimanale, abbandonando saggiamente il concetto forviante di “evento in acuto”.
Nelle indicazioni viene riportata anche la soglia del grasso corporeo da non superare. La quota riportata è del 10%… Ora, fatta eccezione per soggetti con un set point adipocitario sensibilmente basso, arrivare a pesare il 15% in più del peso ideale mantenendo una “bf” del 10% è cosa non proprio semplice, leggermente di più se parliamo di atleti “Natural”. Molto più logico sarebbe stato indicare una soglia tra il 12 ed il 15%. Ma l’autore scrive che la Fase di Massa deve essere interrotta quando si raggiunge il peso desiderato o il 10% di “bf”… Se siete agonisti dovete interromperla se arrivate al limite delle 12 settimane dalla gara.
Per quanto concerne l’obbiettivo di incremento settimanale di peso, viene indicato come “ideale” 1Kg a settimana.
Ma quanti glucidi dovrebbero essere consumati durante il/i giorno/i di “ricarica”? Di Pasquale afferma che si possono raggiungere “quantità enormi” di CHO e Kcal nel week and, anche fino a 12.000Kcal tanto il sabato quanto la domenica. Inoltre sia la quota di CHO giornaliera che il numero di giorni con rialzo glucidico possono essere aumentati in base alle risposte. E già questo punto denota una mancanza di funzionalità di base del suo piano alimentare “High Fat” per una “massa pulita” che però va oltre il concetto di “pulito” arrivando al “nodo metabolico”: il vantaggio indiscutibile del substrato glucidico per sostenere l’attività fisica in uno sport contro-resistenza che sfrutta prevalentemente un metabolismo anaerobico glicolitico.
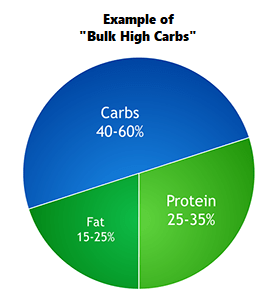
Percentuali di ripartizione dei macronutrienti durante i giorni “High Fat” e le “ricariche di Carboidrati”. (fonte immagine “THE ANABOLIC DIET”, pag. 30 Capitolo 3).
Lascerò da parte la sfilza di integratori marca “MetabolicDiet”, che per lo più altro non sono che Creatina e Proteine in polvere, e le riduttive, ridicole e confutate chiacchiere sulla manipolazione ormonale “simil-doped”, riflettendo direttamente su quanto detto fino a questo punto.
Ora, abbiamo una dieta per il “Bulk” a tutti gli effetti “High Fat” per 3-4/5 della settimana con delle “ricariche glucidiche” di base di 1 o 2 giorni ma incrementabili a bisogno. Ricordo che questa dieta ha la bellezza di 26 anni, ed il sottoscritto, ai tempi in cui ero fortemente tentato a prestar orecchio più alla “carne ed al sangue” piuttosto che alla logica e al buon senso, la conosce e osserva i suoi effetti sugli atleti che l’hanno voluta sperimentare da circa 15 anni. Beh, posso assicurarvi che i risultati con questo tipo di alimentazione in “Bulk” erano apprezzabili (questo non vuol dire che erano spettacolari o che riflettevano le promesse dell’autore) principalmente in due contesti:
Alteti “doped” con un set point adipocitario basso e una media di 3 copiose “ricariche” a settimana;
Atleti “natural” con un set point adipocitario basso e una media di 3 copiose “ricariche” a settimana e un minimo di CHO giornalieri pari a 2-3g/Kg.;
Atleti di sesso femminile con variabili sul numero di “ricariche” e tempi di svolgimento delle medesime.
Ed i casi osservati con risultanti apprezzabili, comunque, sono pochi rispetto ad una maggioranza di individui non significativamente avvantaggiati dal protocollo.
Come potete vedere, il concetto di base della dieta crolla inesorabilmente sotto il peso di dati empirici raccolti direttamente ed indirettamente nel corso degli anni.
“Mah Gabriel! Esistono altre metodologie “High Fat” per una “massa pulita” e funzionano!” Alt, non ti agitare piccolo “troll”, perchè con il termine “High Fat” mi stai dicendo “tutto e niente”, un pò come succede quando si parla di “dieta iperproteica”.
Si possono considerare “High Fat” tutte le diete che in ipercalorica superano il 20-25% delle calorie giornaliere dai grassi! E’ ovvio che possono esistere degli abissi programmatici tra uno schema alla Di Pasquale e un altro che presenta linee di percentuali macro-caloriche nei range sopra indicati.
Prima di proseguire con esempi di schemi alimentari “High Fat” ben più logici rispetto a quanto presentato ne “La Soluzione Anabolica”, e considerare le argomentazioni a favore di questo tipo di dieta ipercalorica, vediamo come i diversi acidi grassi influenzano la massa muscolare. Passiamo a un pò di teoria per tornare successivamente alla pratica con maggiori conoscenze…
La modulazione nella massa e funzione del muscolo-scheletrico dei lipidi
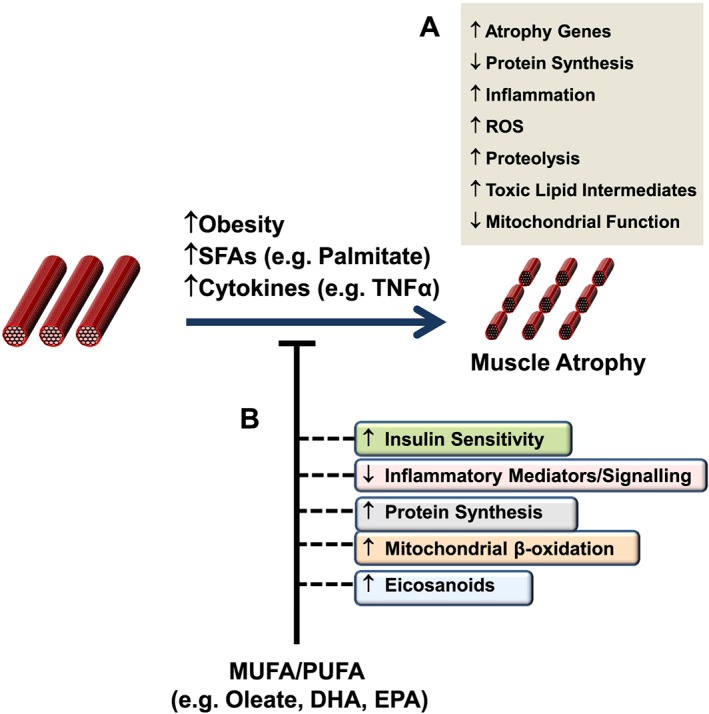
È importante iniziare il discorso sottolineando che vi sono prove crescenti che supportano un ruolo degli acidi grassi e dei loro intermedi lipidici derivati nella regolazione della massa e della funzione del muscolo scheletrico. E’ mia intenzione quindi discutere le prove relative a quei percorsi che sono coinvolti nella riduzione, aumento e/o conservazione della massa muscolare scheletrica da parte dei lipidi.
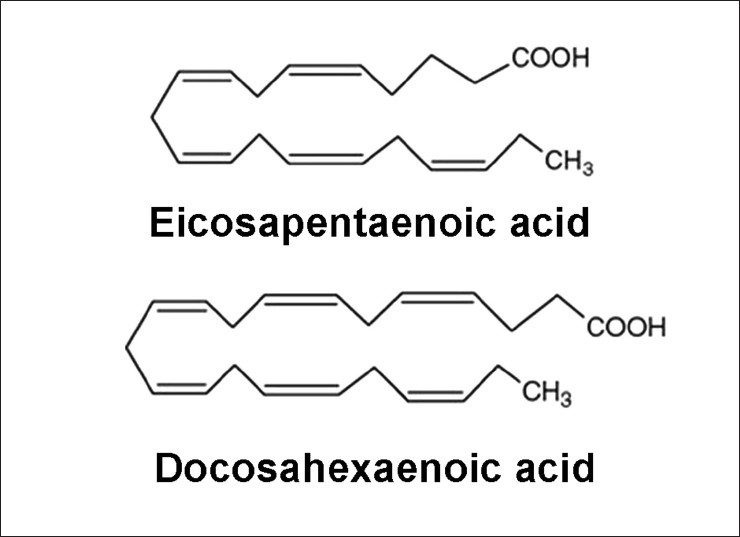
L’evidenza di diversi studi suggerisce che gli acidi grassi saturi e insaturi possono agire per regolare in modo differenziale la massa e la funzione del muscolo scheletrico. Ad esempio, è stato dimostrato che l’esposizione dei miotubi C2C12 al palmitato (C16:0), l’acido grasso saturo circolante più abbondante, riduce il diametro del miotubo e sopprime la segnalazione dell’Insulina. [2] In accordo con questo, è stato riportato che la fornitura di palmitato nelle cellule muscolari induce l’espressione di geni pro-atrofici come atrogin-1/MAFbx, in concomitanza con una maggiore localizzazione nucleare del suo regolatore trascrizionale FoxO3. [3] Al contrario, l’applicazione di acido docosaesaenoico (DHA), un acido grasso polinsaturo omega-3 (PUFA), non ha alterato la morfologia dei miotubi quando applicato da solo ed è stato dimostrato che contromodula l’atrofia indotta dal palmitato nei miotubi C2C12. [2] Coerentemente con ciò, uno studio separato ha riportato il miglioramento della degradazione proteica indotta dal palmitato nei miotubi C2C12 dopo il co-trattamento con DHA. [3] In particolare, ciò ha coinciso con la capacità del DHA di mitigare la localizzazione nucleare potenziata di FoxO3 e l’espressione genica di atrogin-1/MAFbx in risposta alla fornitura di palmitato. [3]
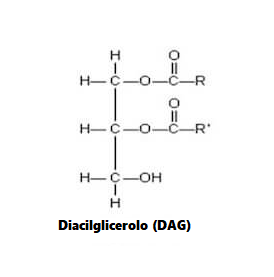
In accordo con questi risultati nelle cellule muscolari di coltura, diversi studi in vivo hanno anche riportato la capacità degli acidi grassi insaturi di trasmettere risposte benefiche che agiscono per prevenire l’atrofia muscolare. Ad esempio, l’alimentazione di topi portatori dell’adenocarcinoma del colon-26, un modello animale di cachessia tumorale, con una dieta integrata con acido linoleico coniugato, ha dimostrato di preservare la massa muscolare del gastrocnemio. [4] In particolare, questo effetto protettivo ha coinciso con una riduzione dell’espressione del recettore TNF-α del muscolo scheletrico, suggerendo che i PUFA possono agire per prevenire l’atrofia muscolare, almeno in parte, riducendo le azioni cataboliche della citochina TNF-α. [4 , 5] In uno studio separato, l’integrazione alimentare con acido eicosapentaenoico (EPA; C20:5 (n-3)) ha attenuato la degradazione proteica nel muscolo gastrocnemio di topi portatori del tumore MAC16 che induce la cachessia. [6] Il trattamento con EPA è stato riportato anche per evitare riduzioni dell’artrite indotta e aumento del peso del muscolo gastrocnemio nei ratti dopo somministrazione di adiuvante di Freund, concomitante con la normalizzazione dell’espressione genica di atrogin-1 / MAFbx e MuRF1. [7] Inoltre, i criceti distrofici alimentati con una dieta arricchita in acido α-linolenico PUFA (ALA) (C18:3(n-6)) hanno mostrato miglioramenti nella morfologia e nella funzione muscolare, compreso l’ingrossamento delle miofibre . [8] In accordo con questi risultati, è stato anche dimostrato che i PUFA omega-3 e omega-6 aumentano la fosforilazione di p70S6K1 a Thr389, indicativo della sua maggiore attività, durante la differenziazione miogenica dei miociti L6. [9] Insieme, questi studi supportano l’idea che gli acidi grassi insaturi possono fornire protezione contro l’atrofia muscolare in risposta a varie condizioni patologiche e potenzialmente migliorare le condizione trofiche in soggetti sani. Inoltre, questi risultati evidenziano le risposte distinte che gli acidi grassi saturi e insaturi inducono rispettivamente per promuovere o contrastare l’atrofia muscolare e la degradazione proteica.
Un certo numero di differenti vie di segnalazione e/o intermedi sono stati implicati come potenziali mediatori della atrofia muscolare, che a loro volta possono essere regolati in risposta alla assunzione di acidi grassi (vedi Figura seguente). Ad esempio, è noto che il palmitato agisce come un potente repressore della segnalazione diretta di PKB/Akt nel muscolo scheletrico, almeno in parte attraverso la sua capacità di indurre l’accumulo di intermedi lipidici tossici come la Ceramide. [10 , 11] Infatti, tali sfingolipidi possono agire stimolando le isoforme della proteina fosfatasi 2A (PP2A) o della proteina chinasi C (PKC) atipica (PKCζ) per inibire PKB/Akt. [12] In accordo con ciò, è stato riportato che l’atrofia del miotubo C2C12 indotta da TNF-α coincide con livelli elevati di Ceramide intracellulare, [13] mentre è stato dimostrato che il blocco della sintesi di Ceramide attenua l’atrofia muscolare indotta dal TNF-α nei miotubi L6, oltre a proteggere i topi contro l’atrofia del muscolo scheletrico tumore indotto ( via impianto di carcinoma C26) in vivo. [13] In particolare, queste risposte benefiche hanno contribuito a una maggiore sintesi proteica e a una diminuzione della proteolisi, in concomitanza con una ridotta espressione del gene atrogin-1/MAFbx tramite la funzione Foxo3 soppressa, nonché una maggiore abbondanza di mediatori chiave della sintesi proteica tra cui S6K1 e PKB/Akt. [13] Inoltre, è stato riportato che la fornitura esogena di Ceramide nelle cellule muscolari L6 riduce i livelli proteici del fattore di trascrizione miogenico miogenina attraverso l’ inibizione della fosfolipasi D, mentre l’inibizione della sintesi di Ceramide migliora l’espressione della miogenina e accelera la formazione di miotubi. [14]Uno studio di Turpin e colleghi ha anche dimostrato un aumento del contenuto di Ceramide muscolare dopo l’infusione acuta (5 h) di intralipid®, che ha coinciso con l’attivazione della segnalazione pro-apoptotica, come dimostrato dall’aumento dell’attività della caspasi-3 nel muscolo gastrocnemio. [15] Tuttavia, il ruolo della Ceramide nel promuovere questo aumento dell’apoptosi muscolare guidato dai lipidi non è stato studiato, ad esempio mediante la co-somministrazione di inibitori della sintesi della Ceramide. In alternativa, livelli elevati di Ceramide associati all’iperlipidemia possono anche agire per sopprimere la sintesi proteica inducendo l’espressione e/o l’attività di repressori chiave della segnalazione mTORC1-S6K come Regulated in Development e DNA Damage 1 (REDD1). [16 , 17]In particolare, va anche evidenziato che il ganglioside GM3 (trisialotetrahexosylganglioside), un glicosfingolipide contenente acido sialico derivato dalla Ceramide, è stato anche implicato come regolatore negativo della crescita e/o differenziazione del muscolo scheletrico, in concomitanza con la sua capacità segnalata di modificare l’azione dell’insulina alterando la funzione del recettore specifico (Recettore dell’Insulina). [18 , 19 , 20 , 21] Inoltre, è stato dimostrato che un altro lipide derivato dalla Ceramide, ceramide-1-fosfato, stimola la proliferazione dei mioblasti C2C12 attraverso un meccanismo che coinvolge l’attivazione di Akt, mTOR e ERK1/2. [22] In effetti, un ulteriore lavoro che utilizza topi carenti di GM3 sintasi, l’enzima responsabile della sintesi di GM3, potrebbe far luce sul ruolo di questo ganglioside nel controllo della massa muscolare scheletrica, ad esempio in risposta all’obesità e/o all’invecchiamento.
Riassunto delle vie che mediano l’atrofia muscolare da acidi grassi saturi. L’esposizione delle cellule muscolari ad acidi grassi saturi come il palmitato (C16:0) provoca l’accumulo intracellulare di intermedi lipidici tossici come Ceramide e Diacilglicerolo. (A) L’aumento dei livelli di Ceramide può portare all’inibizione della proteina chinasi B/Akt attraverso l’attivazione di isoforme atipiche della proteina chinasi C (ξ/λ) e/o della proteina fosfatasi 2A. Inoltre, la Ceramide agisce come precursore per la sintesi del glicosfingolipide GM3 che ha dimostrato di compromettere la funzione del recettore dell’Insulina. Inoltre, la Ceramide può anche agire per modulare l’assorbimento dei nutrienti, ad esempio reprimendo l’espressione del trasportatore amminoacidico neutro SNAT2, riducendo così l’apporto cellulare di amminoacidi. (B) È stato dimostrato che la stimolazione indotta da diacilglicerolo della protein chinasi Cθ promuove la fosforilazione della serina dell’IRS-1, con conseguente sua funzione compromessa. La risultante inibizione della protein chinasi B/Akt a sua volta può portare alla repressione della sintesi proteica attraverso la soppressione del segnale meccanicistico bersaglio della rapamicina (mTOR)/p70-S6 chinasi 1-dipendente (C), l’attivazione del forkhead box O (FoxO ) fattori di trascrizione e induzione dei loro geni atrofici bersaglio (D) e/o l’attivazione della proteolisi dipendente dalla caspasi (E). Inoltre, la stimolazione della segnalazione proinfiammatoria da parte degli acidi grassi saturi a catena lunga può portare alla sovraregolazione dei geni atrofici dipendente dal fattore nucleare kappa B (F).
Oltre agli sfingolipidi, i diacilgliceroli (DAG) sono una classe alternativa di lipidi che possono essere generati in risposta alla ingestione di acidi grassi. In particolare, l’aumento dei livelli di DAG è stato associato allo sviluppo dell’insulino-resistenza, fattore di per se limitante sul corretto ripartizionamento calorico a favore del miocita e, quindi, sulla sintesi proteica . [23] sono stati rilevati Inoltre, un aumento dei livelli di DAG muscolare dopo infusione di lipidi nei topi, con concomitante aumento della attività nel muscolo gastrocnemio della caspasi-3 .[15] Sebbene si sappia poco sul ruolo dei DAG nella regolazione della massa muscolare scheletrica, è stato riportato che l’attivazione meccanica ex-vivo della DAG chinasiζ (DGKζ), un enzima che catalizza la conversione di DAG in acido fosfatidico (PA), favorisce un aumento Segnalazione dipendente da mTOR e ipertrofia associata nel muscolo estensore lungo delle dita (EDL) di topo isolato, in concomitanza con la capacità riportata di PA di legare e attivare direttamente mTOR. [24 , 25] In accordo con ciò, è stato anche dimostrato che la sovraespressione cardiaca specifica di DGKζ migliora l’atrofia miocardica nei topi diabetici indotti da streptozotocina. [26] Pertanto, questi risultati suggeriscono che l’attivazione e/o la sovraespressione di DGKζ può fornire un mezzo per stimolare i tassi di sintesi proteica e le risposte ipertrofiche, e quindi migliorare le perdite di massa muscolare, sia riducendo i livelli cellulari di DAG e/o aumentando l’attivazione della segnalazione mTOR indotta dal PA. È importante sottolineare che il lavoro futuro potrebbe comportare lo studio dei potenziali effetti benefici della sovraespressione di DGKζ nel muscolo come mezzo per contrastare l’atrofia muscolare indotta dall’età e/o dalla dieta. Inoltre, modelli animali che mostrano livelli elevati di DAG nel muscolo scheletrico, compresi i topi che sono carenti di lipasi ormone-sensibile (HSL), [27] possono anche essere utili per chiarire il ruolo di DAG nell’atrofia del muscolo scheletrico.
Un’altra considerazione importante riguarda la possibilità che specie di DAG distinte possano avere un impatto diverso sulle vie coinvolte nella regolazione della massa muscolare, ad esempio come determinato dalla composizione dei gruppi acilici grassi che si esterificano a livello sn-1,2, sn‐ 1,3, o le posizioni sn-2,3 della base di glicerolo del DAG. [28 , 29] Infatti, è stato dimostrato che il trattamento dei miotubi L6 di ratto con palmitato porta ad aumenti significativi dei livelli cellulari di alcune specie di DAG, nonché del contenuto totale di DAG cellulare. [30] Inoltre, è stato dimostrato che il co-trattamento con l’acido grasso monoinsaturo (MUFA) palmitoleato (C16:1) sopprime selettivamente gli aumenti indotti dal palmitato nei livelli di specie DAG contenenti porzioni di acidi grassi saturi C18:0 e C20:0, in coincidenza con l’azione antinfiammatoria del MUFA. [30] Sebbene non determinati in questo studio, stereoisomeri distinti di DAG possono anche regolare in modo differenziale la segnalazione anabolica/catabolica muscolare. Per supportare questa nozione, è stato riportato che gli stereoisomeri sn-1,2 DAG (rispetto agli isomeri sn-1,3) sono più potenti nell’attivare le vie di segnalazione legate all’insulino-resistenza, inclusa l’attivazione della PKC. [31] Insieme, questi studi forniscono prove emergenti che alcune molecole/isomeri DAG possono svolgere un ruolo più importante nello sviluppo dell’atrofia muscolare, ad esempio promuovendo la resistenza all’insulina e/o aumentando la spinta proinfiammatoria. Tuttavia, sarà necessario un ulteriore lavoro per determinare quali di queste molecole di DAG, se presenti, sono responsabili delle azioni di atrofia muscolare. Nel tentativo di affrontare questo problema, studi futuri potrebbero comportare il trattamento di cellule muscolari in coltura con diverse molecole/stereoisomeri DAG al fine di determinare i loro effetti sulla miogenesi e/o sull’atrofia muscolare. In alternativa, un ulteriore lavoro può anche incorporare un’analisi lipidomica dettagliata di varie specie di DAG intramuscolari in tessuti isolati da modelli animali di atrofia muscolare, oltre a monitorare potenziali cambiamenti nella loro abbondanza a seguito di interventi noti per aumentare la massa muscolare (ad es. apporto dietetico di PUFA o aumento dell’attività fisica). Infatti, se tali studi dovessero rivelare un ruolo chiave per l’accumulo di DAG nello sviluppo dell’atrofia del muscolo scheletrico, il lavoro successivo potrebbe implicare la determinazione dell’origine di tali specie di DAG, ad esempio inibendo l’attività degli enzimi implicati nella formazione del triacilglicerolo dal DAG ( sintesi di TAG) (es. glicerolo fosfato transferasi (GPAT), acilglicerolfosfato aciltransferasi (AGPAT) e lipina), o alterando l’attività di enzimi implicati nell’idrolisi di TAG e/o DAG (es. lipasi dei trigliceridi adiposa (ATGL) o HSL). A questo fine, i lavori di Badin e collaboratori hanno riportato un’elevata abbondanza di proteine ATGL nel muscolo scheletrico di individui diabetici di tipo 2 rispetto a soggetti di controllo magri, nonché una ridotta espressione di HSL muscolare in individui obesi.[32] Inoltre, gli autori dello stesso studio hanno ulteriormente dimostrato che la sovraespressione di ATGL o l’inibizione dell’attività dell’HSL nei miotubi primari umani determinava l’accumulo di DAG cellulare e una compromissione associata nella segnalazione dell’Insulina. Tuttavia, in questo studio non è stato determinato se questi cambiamenti nei livelli di DAG siano collegati all’atrofia muscolare.
Oltre a modulare la segnalazione diretta di PKB/Akt e/o mTORC1, gli acidi grassi e/oi loro lipidi derivati possono ulteriormente contribuire alla perdita muscolare modulando il trasporto dei nutrienti (aminoacidi) e/o la segnalazione associata. Ad esempio, lavori svolti da diversi gruppi di ricerca hanno dimostrato la capacità della Ceramide di sottoregolare l’espressione e/o l’attività dei principali trasportatori di nutrienti, incluso il trasportatore di amminoacidi neutri SNAT2 (SLC38A2). [33 , 34] In tal modo, gli acidi grassi che agiscono attraverso tali intermedi lipidici possono agire per compromettere l’assorbimento degli aminoacidi, contribuendo così a una perdita di massa muscolare. È interessante notare che in uno studio separato, è stato dimostrato che l’incubazione di miotubi L6 di ratto con acido linoleico (C18:2) limita la sovraregolazione adattativa dell’espressione e dell’attività di SNAT2 in risposta alla carenza di aminoacidi. [35] In particolare, questa riduzione indotta da acidi grassi nell’attività di trasporto del Sistema A è stata mediata da una maggiore ubiquitinazione e degradazione proteasomica della proteina SNAT2. [35] Al contrario, in uno studio separato di Li e collaboratori, è stato riportato che l’espressione dell’mRNA dei transcettori di aminoacidi LAT1 (un trasportatore di aminoacidi di tipo L) e SNAT2 è sovra-regolata nel longissimus dorsi di suini alimentati con diete n. -6 e n-3 PUFA.[36] Quindi, è possibile che gli acidi grassi e/o i loro lipidi derivati possano funzionare per modulare le strategie adattative che vengono utilizzate da tessuti come il muscolo scheletrico, al fine di massimizzare o minimizzare l’assorbimento di nutrienti durante condizioni di digiuno o privazione di nutrienti e, presumibilmente, tale meccanismo può subire alterazioni durante stati di sovra-alimentazione.
È importante sottolineare che la letteratura attuale descrive prove che suggeriscono che gli acidi grassi insaturi possono agire per contrastare i mediatori pro-atrofici, compresi quelli attivati in seguito all’esposizione agli acidi grassi saturi. Ad esempio, è stato riportato che MUFA e PUFA prevengono le riduzioni indotte dal palmitato della sensibilità all’Insulina e veicolano effetti antinfiammatori nelle cellule del muscolo scheletrico. [30 , 37, 38] In effetti, la regolazione trascrizionale dipendente da NF-kB è stata implicata nel promuovere l’atrofia muscolare da disuso nel muscolo soleo di ratto aumentando l’attivazione mediata da FoxO del promotore MuRF1. [39] Inoltre, uno studio recente ha dimostrato che la diminuzione del segnale anabolico nel muscolo scheletrico di topi anziani coincideva con l’accumulo di Ceramide intramuscolare e DAG, nonché con una maggiore abbondanza di mRNA di TNF-α. [40] È interessante notare che la somministrazione di olio di pesce ai suinetti in fase di svezzamento, che ha portato all’arricchimento di EPA, DHA e contenuto totale di PUFA omega-3 all’interno del muscolo gastrocnemio, ha coinciso con una riduzione dei livelli di TNF-α muscolare e una ridotta espressione del recettore Toll-like 4 (TLR4), un recettore bersaglio per gli acidi grassi saturi che stimola la segnalazione proinfiammatoria in risposta alla sua attivazione. [41] In particolare, è stato riportato che la stimolazione del TLR4 da parte del suo ligando lipopolisaccaride induce il catabolismo muscolare nei miotubi C2C12 attraverso l’attivazione delle vie ubiquitina-proteasoma e autofagia-lisosoma. [42] Inoltre, è stato dimostrato che il trattamento con DHA in cellule muscolari umane co-coltivate con macrofagi attenua il contenuto proteico indotto dai macrofagi di Fn14, un modulatore positivo dell’espressione di MuRF-1. [43 , 44] Pertanto, sulla base di questi risultati, è concepibile che le azioni antinfiammatorie riportate degli acidi grassi insaturi nelle cellule muscolari scheletriche possano contribuire, almeno in parte, alla loro capacità di preservare la massa e/o la funzione muscolare.
In particolare, queste azioni protettive possono essere collegate a miglioramenti della funzione mitocondriale, la cui compromissione è stata suggerita dal tipo di dieta e/o all’atrofia muscolare indotta dall’età. [45 , 46] Ad esempio, un recente studio di Roseno e colleghi ha riportato che una dieta ricca di grassi a breve termine (3 settimane) ha aumentato l’atrofia muscolare da denervazione nei topi inducendo la degradazione proteica nel soleo ricco di mitocondri, ma non nel muscolo EDL glicolitico. In particolare, la denervazione di 14 giorni ha indotto una perdita del contenuto proteico mitocondriale nel soleo ma non nell’EDL, indipendentemente dalla dieta. Pertanto, questi risultati suggeriscono che la perdita di mitocondri indotta dalla denervazione e la compromissione della funzione mitocondriale indotta da una dieta ricca di grassi possono combinarsi per promuovere l’atrofia del muscolo scheletrico. Al contrario, uno studio indipendente di Tardif e collaboratori ha dimostrato che i ratti anziani alimentati con una dieta arricchita di oleati mostrano notevoli miglioramenti nella sensibilità all’insulina e un aumento della sintesi proteica muscolare, in concomitanza con una maggiore espressione di geni implicati nella stimolazione della ossidazione mitocondriale, compreso il recettore attivato dal proliferatore dei perossisomi PPARα e PPARβ, nonché CPT-1β. [47 , 48] Inoltre, è stato dimostrato che i miotubi C2C12 trattati con PUFA acido linolenico e ALA mostrano una maggiore attivazione del AMPK, un altro regolatore positivo chiave della -ossidazione mitocondriale. [49] Inoltre, è stato riportato che il DHA inibisce la degradazione proteica nei miotubi C2C12 attraverso una via PPARγ-dipendente. [50] Infatti, la capacità ossidativa mitocondriale potenziata e/o preservata, come precedentemente riportato in risposta alla sola o co-fornitura di acidi grassi insaturi, può anche aiutare a prevenire l’accumulo intramuscolare di intermedi lipotossici come la Ceramide che sono stati implicati nella promozione dell’atrofia muscolare. [51 , 52] Inoltre, è possibile che l’integrazione di PUFA possa agire per alterare le proprietà contrattili e metaboliche muscolari, ad esempio promuovendo un passaggio da fibre glicolitiche veloci a fibre lente (ossidative), cosa non propriamente positiva per un atleta di potenza. A sostegno di questa idea, lavori precedenti hanno dimostrato che l’alimentazione di ratti Wistar con una dieta arricchita in PUFA n-3 porta alla sovraregolazione delle proteine implicate nell’attivazione del metabolismo ossidativo (ad es. nel muscolo EDL, un tessuto muscolare dominante di tipo veloce). [53] È interessante notare che questo cambiamento metabolico mediato da PUFA ha coinciso anche con livelli ridotti di proteine dell’isoforma di tipo veloce MyHC-2b (catena pesante della miosina 2b) nel muscolo EDL. Pertanto, è concepibile che uno spostamento mediato dai PUFA verso un tipo di fibra muscolare a lenta ossidazione possa contribuire, almeno in parte, a guadagni benefici nella massa muscolare e/o nella funzione metabolica.
In alternativa, la regolazione della massa muscolare da parte dei lipidi può anche comportare la modulazione dell’autofagia, un meccanismo omeostatico che facilita la degradazione e il riciclaggio di proteine e organelli attraverso il macchinario lisosomiale. [54] In particolare, è stato riportato che un aumento della degradazione autofagica coincide con l’atrofia muscolare in varie condizioni e/o patologie tra cui cancro, [55] denervazione, [56] e invecchiamento. [55 , 57] Inoltre, è stato dimostrato che un’alimentazione ricca di grassi a breve termine (3 settimane) aumenta l’abbondanza di marcatori autofagosomiali nel soleo denervato dei topi. In accordo con ciò, Yuzefovych e collaboratori hanno dimostrato un aumento dell’autofagia nei miotubi L6 a seguito della somministrazione di palmitato. Pertanto, sebbene non sia ancora stato stabilito un collegamento diretto in vivo , è ipotizzabile che il turnover proteico alterato tramite l’ autofagia possa, almeno in parte, mediare le alterazioni indotte dai lipidi nella massa muscolare.
Va inoltre evidenziato che alcuni acidi grassi insaturi possono alterare il tasso di proliferazione delle cellule satelliti che funzionano come cellule progenitrici miogeniche necessarie per la crescita e la rigenerazione muscolare. Ad esempio, è stato dimostrato che DHA ed EPA inibiscono la proliferazione dei mioblasti C2C12 e delle cellule satelliti isolate dal muscolo di tacchino. [58 , 59] In particolare, queste azioni di soppressione della crescita sono state collegate a livelli ridotti di ciclina E e CDK2, proteine che svolgono un ruolo critico nella progressione del ciclo cellulare, nonché all’attivazione soppressa di ERK1/2, una proteina chinasi attivata da mitogeni implicata nel promuovere la crescita e la divisione cellulare. [59 , 60] Al contrario, è stato dimostrato che l’alimentazione di criceti distrofici carenti di δ-sarcoglicano con una dieta arricchita in ALA (un PUFA omega-3) aumenta la proliferazione e la differenziazione delle cellule satellite nel muscolo EDL, in concomitanza con un miglioramento dell’istologia muscolare. In particolare, queste risposte benefiche hanno coinciso con la capacità dell’ALA di aumentare la proporzione di miofibre α-MHC positive nel muscolo scheletrico dei criceti distrofici, insieme a una riduzione dell’espressione di β-MHC, contribuendo così alla conservazione di un α/β più fisiologico. Rapporto MHC. Inoltre, nello stesso studio, è stato anche dimostrato che l’integrazione di ALA alimentare previene l’accumulo aberrante citoplasmatico di proteine di membrana chiave nei muscoli adduttori dei criceti distrofici, inclusa la caveolina-3, una proteina coinvolta nella regolazione dell’adesione cellulare e della riparazione della membrana, nonché essendo implicato nel controllo della differenziazione muscolare e della segnalazione indotta dall’insulina. [61 , 62] Infatti, dato che le aberrazioni nella funzione e/o localizzazione della caveolina-3 sono state associate a vari fenotipi di malattie del muscolo scheletrico, [63 , 64 , 65 , 66 , 67] è plausibile che gli acidi grassi e/o i loro derivati lipidici possano influenzare la proliferazione delle cellule satellite e/o la differenziazione muscolare, almeno in parte, alterando la funzione e/o la localizzazione subcellulare delle isoforme della caveolina, nonché di altri componenti chiave della membrana strutturale.
Gli effetti degli acidi grassi sulla massa muscolare e sulla differenziazione possono essere mediati anche da una serie di metaboliti lipidici derivati. Ad esempio, è stato dimostrato che la capacità dell’acido arachidonico PUFA (C20; 4n-6) di aumentare le dimensioni, il contenuto mionucleare e il contenuto proteico dei miotubi C2C12 è mediata dall’attività della cicloossigenasi-2 (COX-2), implicando dipendenza da sintesi delle prostaglandine a valle. [68] In accordo con ciò, è stato riportato che la crescita indotta dall’acido arachidonico dei miociti C2C12 coincide con l’aumento della secrezione degli eicosanoidi PGF2α e PGE. [68]È interessante notare che diversi studi hanno anche documentato il ruolo positivo che le prostaglandine svolgono nel promuovere eventi precoci sulla superficie cellulare, inclusa l’adesione cellula-cellula, che successivamente mediano la fusione dei mioblasti nei miotubi. [69 , 70] Infatti, importanti studi di follow-up possono comportare la determinazione dell’esatta identità dei bersagli molecolari attraverso i quali le prostaglandine mediano le loro azioni, ad esempio agendo sui recettori prostanoidi legati alla proteina G (es. EP1). [69 , 71] Al contrario, è stato recentemente riportato che un altro metabolita lipidico derivato dall’acido arachidonico noto come 2-arachidonoilglicerolo (2-AG), un ligando lipidico endogeno chiave del sistema endocannabinoide, inibisce la differenziazione delle cellule satellite primarie umane e dei mioblasti murini C2C12 prendendo di mira il recettore-G ‐ 1 dei cannabinoidi accoppiati a proteine e sua successiva inibizione dei canali Kv7.4. [72] Pertanto, non si può escludere il possibile coinvolgimento di tali intermedi lipidici nella regolazione del catabolismo muscolare in risposta a determinate condizioni patologiche o alimentari.
L’aumento dell’adiposità osservato nell’invecchiamento umano è stato collegato a risposte sintetiche proteiche muscolari alterate negli individui anziani. [73] Pertanto, è ipotizzabile che i cambiamenti nei livelli e/o nella composizione dei lipidi possano contribuire all’atrofia muscolare in queste condizioni ma non limitatamente ad esse. A sostegno di questa idea, ci sono prove che suggeriscono che la modifica della composizione alimentare può avere un impatto sulla massa muscolare e/o sulla sua funzione negli esseri umani. Ad esempio, uno studio di McGlory e collaboratori ha riportato che il consumo di olio di pesce ha aumentato il contenuto di PUFA omega-3 del muscolo ( Vastus lateralis) in individui maschi sani, che ha coinciso con un’elevata espressione di proteine di segnalazione anabolizzanti incluso mTOR. [74] Inoltre, è stato riportato che l’inclusione di MUFA e PUFA nella dieta riduce l’espressione di geni lipogenici nel muscolo scheletrico di soggetti insulino-resistenti, in concomitanza con ridotti tassi di sintesi frazionaria di DAG intramuscolare e triacilgliceroli. [75] In particolare, queste risposte benefiche possono essere collegate a miglioramenti nella sensibilità all’insulina veicolati dall’integrazione alimentare di PUFA omega-3 negli esseri umani. [76 , 77 , 78]
E’ stato riportato che l’integrazione alimentare con acidi grassi insaturi può anche agire per migliorare le prestazioni fisiche e/o aumentare gli effetti metabolici benefici associati all’esercizio, in particolare in individui sedentari o non allenati . [79] A sostegno di questa idea, è stato dimostrato che un basso apporto alimentare di olio di tonno promuove la resistenza all’affaticamento muscolare nei ratti, in concomitanza con un aumento selettivo del contenuto di fosfolipidi della membrana e DHA all’interno del muscolo gastrocnemio. [80 , 81] Pertanto, un approccio più integrato che coinvolga modifiche al consumo di grassi nella dieta e un aumento dell’esercizio fisico può fornire una strategia più efficace per alleviare gli effetti deleteri associati all’atrofia muscolare. Ma stiamo comunque parlando di effetti apprezzabili su soggetti sedentari e non allenati, non su culturisti intermedi o avanzati.
Tirando le somme di quanto fino qui esposto, c’è un certo interesse giustificato sul ruolo importante che gli acidi grassi e/oi loro derivati lipidici possono svolgere nella modulazione della massa muscolare e della sua funzione. Nel complesso, le prove presentate indicano che gli acidi grassi saturi agiscono per trasmettere effetti dannosi sulla funzione muscolare, ad esempio alterando o riducendo la sintesi proteica e accentuando il catabolismo. Al contrario, è stato dimostrato che diversi acidi grassi insaturi contrastano molte delle azioni pro-cataboliche associate alla assunzione di acidi grassi saturi. Tuttavia, sarà necessari ulteriori studi per delineare le vie e i processi alla base dell’atrofia muscolare indotta da acidi grassi, nonché quelli che mediano miglioramenti nella funzione muscolare in risposta alla fornitura di acidi grassi insaturi (cioè aumento della sintesi proteica, ridotta atrofia, miglioramento della funzione metabolica). A tal fine, strategie volte ad alterare il contenuto e/o la composizione lipidica intramuscolare in quelle condizioni che possono favorire l’atrofia muscolare (es. aumento dell’obesità, invecchiamento, inattività fisica), ad esempio sopprimendo l’accumulo di mediatori lipidici come le Ceramidi (es. de novosintesi di ceramide) o prostaglandine (ad es. inibendo l’attività della COX-2), possono fornire informazioni utili sul ruolo che diverse classi di lipidi svolgono nella modulazione della massa e della funzione muscolare. È importante sottolineare che tale lavoro potrebbe implicare l’uso di modelli animali pertinenti o soggetti umani che richiederebbero di prendere in considerazione fattori come il background genetico, la composizione della dieta e l’apporto calorico. Inoltre, questi studi possono anche comportare la determinazione di potenziali alterazioni indotte dai lipidi dell’architettura muscolare e della composizione del tipo di fibra che possono influenzare la forza muscolare, nonché il monitoraggio dei cambiamenti nella segnalazione intramuscolare e nei metaboliti all’interno di specifici tipi di fibre muscolari. Insieme a questo, ulteriori lavori che esplorano il ruolo degli acidi grassi e degli intermedi lipidici nella regolazione della proliferazione, sarebbe necessario eseguire la differenziazione e/o la funzione delle cellule satellite derivate da muscoli umani e dei miotubi primari al fine di effettuare confronti appropriati con i dati ottenuti in altri modelli sperimentali (ad es. miotubi C2C12), che hanno dimostrato di mostrare differenze funzionali (ad es. grado di maturazione).[82] Nel complesso , i dati ottenuti da tali studi possono portare allo sviluppo di nuove strategie terapeutiche per contrastare l’atrofia muscolare e/o migliorare la capacità rigenerativa a seguito di lesioni o malattie.
Potenziali meccanismi attraverso i quali gli acidi grassi insaturi possono contrastare l’obesità e/o l’atrofia del muscolo scheletrico indotta da acidi grassi saturi. Livelli elevati di acidi grassi saturi e/o citochine proinfiammatorie, ad esempio durante l’obesità e ipoteticamente anche nel sovrappeso marcato, possono favorire lo sviluppo dell’atrofia del muscolo scheletrico attraverso varie vie e processi come indicato (A). È importante sottolineare che gli acidi grassi monoinsaturi e polinsaturi hanno dimostrato di contrastare queste azioni pro-atrofiche, che possono essere mediate dalla loro capacità di aumentare la sensibilità all’insulina e la produzione di eicosanoidi protettivi, oltre a migliorare la capacità ossidativa mitocondriale e la sintesi proteica, mentre riducono contemporaneamente la spinta proinfiammatoria (B).
Torniamo alla pratica: tesi, antitesi e sintesi del “Bulk High Fat”.
Quindi, cosa concludere da tutto ciò che ho esposto proveniente dalla letteratura scientifica se parliamo di Bodybuilding e alimentazione “High Fat” in ipercalorica? Quanto segue:
La cosa certa, e risaputa in tutti i contesti dietetici, è quella di preoccuparsi di ridurre l’assunzione di acidi grassi saturi a circa e non oltre il 10% del totale calorico giornaliero (soprattutto l’AcidoPalmitico, Miristico e Laurico);
Concentrare l’assunzione di acidi grassi utilizzando fonti principalmente di Omega-3 (con particolare attenzione ad assumere EPA e DHA in un range tra 1 e 5g/die)
Assumere una buona dose di GLA (Omega-6) la quale può portare ad una migliore risposta nella biosintesi di prostaglandine anti-infiammatorie;
Possibilità di assumere Acido Arachidonico (Omega-6) in un range di dosaggio tra i 100mg ed 1.5g/die (quest’ultimo dosaggio va calibrato con un apporto di EPA/DHA pari ad un minimo di 2,5g/die) che può portare ad un miglioramento della biosintesi di prostaglandine anti-infiammatorie e con attività anabolizzante (vedi, per esempio, la PGF2-α responsabile con la PGE2 della regolazione della proteolisi e della sintesi proteica; così come della proliferazione, differenziazione e fusione delle celle satellite);
Uno schema di ripartizionamento dei macronutrienti potrebbe essere il seguente: 30-40% di Grassi, 30-20% di Carboidrati e 40-30% di Proteine.
Metabolismo e vie di segnalazione dell’Acido Arachidonico. Sono riassunte 3 delle 4 principali vie metaboliche dell’Acido Arachidonico: il ciclo di Terra, la conversione in prostaglandine che richiede l’attività della cicloossigenasi e la conversione in leucotrieni e lipossine tramite l’attività della 5-lipossigenasi. Non è mostrato il metabolismo del citocromo P-450 dell’Acido Arachidonico. I processi enzimatici sono mostrati in rosso, i substrati ei prodotti sono mostrati in nero e i recettori di superficie della proteina G delle cellule sono mostrati in verde. Gli effetti noti dell’attivazione del recettore sull’adenosina monofosfato ciclico (cAMP) a valle o sui livelli di calcio intracellulare sono mostrati in basso. COX-1, cicloossigenasi-1; COX-2, cicloossigenasi-2; 5-LO, 5-lipossigenasi; FLAP, proteina attivante la 5-lipossigenasi; PGH2, PGD2, PGE2, PGF2a e PGI2, sono prostaglandine H2, D2, E2, F2a e I2, rispettivamente; TXA2: trombossano A2; acido 5-HpETE, 5S-idroperossi-6E,8Z,11Z,14Z-eicosatetraenoico; acido 5-HETE, 5S-idrossi6E,8Z,11Z,14Z-eicosatetraenoico; LTA4, LTB4, LTC4, LTD4 e LTE4 sono i leucotrieni A4, B4, C4, D4 ed E4, rispettivamente; LXA4 e LXB4, lipossine A4 e B4; DP1, EP1-EP4, FP e IP, recettori delle prostaglandine; TP, recettore del trombossano A2; BLT1 e BLT2, recettore dei leucotrieni B4; CysLT1 e CysLT2: recettori dei cisteinil leucotrieni; ALXR: recettore delle lipossine.
Partendo da questi punti si hanno delle “linee guida” finalizzate alla qualità degli acidi grassi e alla quantità media percentuale.
Ora, mettiamo il caso che un soggetto per caratteristiche genetiche non riesca a gestire grosse quantità di glucidi e abbia un buon metabolismo lipidico sul quale poter puntare per eventuali surplus calorici.
Il soggetto in questione si trova bene con un quantitativo di CHO pari a 3g/Kg, e pesando 80Kg la suo quota glucidica giornaliera ammonta a 240g. L’assunzione proteica giornaliera è pari a 2,5g/Kg, per un totale di 200g.
Il suo TDEE di mantenimento è di circa 2.500Kcal. Punta ad un surplus calorico giornaliero di 500Kcal (obbiettivo 3000Kcal/die). Dal momento che sa già quanti CHO e proteine dovrà assumere e a quante calorie ammontano (1760Kcal totali da CHO e Pro) il restante lo andrà ad assumere dai Grassi (1240Kcal = circa 138g di Fat).
Totale macronutrienti del soggetto:
Carboidrati: 240g
Proteine: 200g
Grassi: 138g (di cui il 10% di Saturi ed il restante 90% di insaturi, polinsaturi e monoinsaturi; presenza di 5g/die di EPA e DHA, 300mg/die di GLA e 1.5g/die di Acido Arachidonico per 8-12 settimane).
Il soggetto in questione seguirà questo schema per circa 4 settimana senza effettuare alcuna giornata con un aumento dei CHO e una riduzione della componente proteica e lipidica. In base alle risposte ottenute rimodulerà a bisogno il suo schema alimentare.
Ecco, questo è un banalissimo esempio di come una dieta “Bulk High Fat” possa essere per lo meno impostata in modo logico, anche per quanto riguarda il tipo ipotetico di soggetto menzionato. Nonostante ciò, e vi parlo da persona che ha analizzato i risultati di svariati schemi similari a questo, un vantaggio vero e proprio sulla qualità dell’aumento di peso non è praticamente mai emerso, o per lo meno mai in modo sufficientemente apprezzabile.
A questo punto ci sarebbe anche da riportare quanto affermato in uno studio dal Dr. Jose Antonio, PhD, ricercatore della ISSN, che ha affermato come non vi siano aumenti maggiori di grasso corporeo tra una dieta ipercalorica “High Fat” e una “High Carbs” dopo circa 14 giorni.[https://italia-podcast.it/] Questa affermazione va comunque presa come una ipotesi nata da osservazione di alcuni studi. Sebbene tra la teoria e la pratica spesso c’è differenza, quando si parla di surplus calorico e aumento del grasso corporeo sappiamo tutti quanto gli acidi grassi alimentari vengano facilmente stoccati negli adipociti sotto forma di trigliceridi di deposito quando in surplus calorico e ingeriti in quantità significative con carboidrati. Questi ultimi, o meglio il glucosio, in eccesso, in un soggetto in salute vengono in gran parte dissipati in calore e solo un 5-10% è coinvolto nei processi di De Novo Lipogenesi (DNL).
Alcuni sostenitori delle diete “High Fat” in “Bulk” affermano che tali strategie alimentari portino ad un miglioramento della “durezza muscolare”, anche grazie ad un basso stimolo insulinico e consequenziale riduzione della ritenzione idrica, e della pienezza dei ventri data dai Trigliceridi intra-muscolari. Peccato che, Insulina o meno, gli acidi grassi in eccesso vengono captati e depositati negli adipociti per intervento, tra l’altro, della Proteina Stimolante l’Acilazione (ASP), la quale ha azione insulino-indipendente. I Trigliceridi intra-muscolari (IMTG) danno si un effetto di pienezza muscolare dal momento che legano in piccola parte acqua, ma parliamo di un effetto additivo al ben maggiore volume dato dal glicogeno muscolare e apprezzabile a “bf” basse, da pre-contest per intenderci.
Come molti di voi già sapranno, la sintesi proteica aumenta principalmente:
In seguito ad adattamenti in risposta a sedute di allenamento contro-resistenza correttamente svolte generando tensione meccanica, stress metabolico e danno muscolare;
Mangiando sufficienti calorie;
Assumendo un quantitativo adeguato di proteine (media 1,5-2,5g/Kg).
Raggiungere il primo punto, però, diventa molto più difficile se si consuma una quota glucidica bassa. Le diete ricche di grassi possono fornire molte calorie e proteine, ma risultano per la stragrande maggioranza dei soggetti disfunzionali alle prestazioni in sala pesi. Ricordo, per l’ennesima volta, che un bodybuilder è un atleta che sfrutta primariamente il metabolismo glucidico al fine della massima prestazione essendo praticante di uno sport anaerobico alattacido e lattacido.
Durante l’esercizio intenso, i muscoli si basano principalmente sul metabolismo glucidico. E come certamente saprete, il glicogeno è un polimero di glucosio immagazzinato nel muscolo-scheletrico e nel fegato, ed è scomposto nelle sue unità di glucosio e utilizzato come substrato energetico durante l’esercizio nel distretto muscolare di deposito oppure venendo rilasciato nel flusso ematico dal deposito epatico.
Le richieste di carboidrati nella dieta variano da soggetto a soggetto, ma un culturista che si allena seriamente richiede in media una consistente quantità di glucidi per mantenere le sue riserve di glicogeno e sostenere una buona prestazione, motivo per cui le diete ricche di grassi e povere di carboidrati riducono significativamente i livelli di questo polimero. Ed i problemi annessi emergono soprattutto quando ci si allena con pesi consistenti.
Fare solo 6-9 serie può ridurre i livelli di glicogeno muscolare di circa il 40% [83] e, se ci si allena con livelli già bassi, è ragionevole presumere che si farà fatica a sollevare più peso.
Ma cosa mostrano gli studi sugli atleti? La maggior parte degli studi dimostrano che le diete ricche di grassi e povere di carboidrati riducono le prestazioni atletiche [84, 85, 86] mentre altri affermano che non vi è alcuna differenza [87]e persino alcuni che mostrano un aumento delle prestazioni.[88] Tuttavia, ci sono altri motivi per pensare che le diete ad alto contenuto di carboidrati potrebbero essere superiori per l’aumento della massa muscolare.
La ricerca mostra che la disponibilità di glicogeno influenza direttamente la sintesi proteica ei tassi di degradazione.[89] In poche parole: le diete ricche di grassi e povere di carboidrati comportano livelli inferiori di sintesi proteica rispetto a quelle ad alto contenuto di carboidrati.
Un altro modo in cui i carboidrati influenzano favorevolmente l’equilibrio proteico muscolare ha a che fare con la produzione di Insulina.
L’insulina è un ormone peptidico rilasciato dal pancreas che agevola l’assorbimento dei nutrienti dal sangue alle cellule. Possiede, in fisiologia, proprietà prettamente anticataboliche, il che significa che quando i livelli di Insulina sono elevati, viene soppressa la lisi delle proteine muscolari.[90]
Ora, poiché la produzione di Insulina è stimolata ingerendo cibo, e mangiando carboidrati in particolare, ma anche proteine con stimoli differenti dati dalla composizione amminoacidica, non sorprende che le persone che seguono una dieta ricca di carboidrati abbiano generalmente livelli ottimali di Insulina rispetto alle persone che seguono una dieta povera di carboidrati e ricca di grassi.
La formazione del complesso Insulina (di colore rosso) – Recettore dell’Insulina (di colore blu) determina una cascata di segnali per l’esplicazione della funzione dell’ormone. Ispirato da discussioni e cifre in Siddle (2011). L’immagine del complesso Recettore Insulina-Insulina è adattata da Goodsell (2015).
I carboidrati sono principalmente energetici e se non si è molto attivi, non si necessita di loro quantità elevate. Tuttavia, livelli ottimali di insulina (e di sensibilità insulinica) sono altamente desiderabili se si sta cercando di costruire massa muscolare, semplicemente perché si viene a creare un ambiente più anabolico in cui i muscoli possono vedere agevolata la loro crescita. E questa non è solo teoria.
La ricerca condotta dagli scienziati della Ball State University ha scoperto che bassi livelli di glicogeno muscolare (che è inevitabile con una dieta povera di carboidrati e ricca di grassi) compromettono la segnalazione cellulare post-allenamento relativa alla crescita muscolare.[91]
Un altro studio condotto da ricercatori dell’Università della Carolina del Nord ha scoperto che, se combinata con l’esercizio quotidiano, una dieta povera di carboidrati e ricca di grassi aumenta i livelli di Cortisolo a riposo e diminuisce i livelli di Testosterone libero.[92] Sicuramente una situazione compromettente soprattutto per un “Natural”. Quando si tratta di costruire massa muscolare, ciò che si vuole sono bassi livelli di Cortisolo a riposo e alti livelli di Testosterone libero, l’esatto opposto di ciò che porta una dieta a basso contenuto di carboidrati.
Quanto detto aiuta a spiegare i risultati di altri studi sulla questione dei carboidrati e della composizione corporea e delle prestazioni.
Ad esempio, uno studio condotto da ricercatori dell’Università del Rhode Island ha esaminato come l’assunzione di carboidrati a basso e ad alto contenuto di carboidrati influenzasse il danno muscolare indotto dall’esercizio, il recupero della forza e il metabolismo proteico di tutto il corpo dopo un intenso allenamento. Quello che hanno scoperto è che i soggetti con una dieta a basso contenuto di carboidrati hanno perso più forza, si sono ripresi più lentamente e hanno mostrato livelli più bassi di sintesi proteica. Vale anche la pena notare che il gruppo “a basso contenuto di carboidrati” (e quindi ad alto contenuto di grassi) non era poi così “Low Carbs”. Stavano ingerendo circa 220g di carboidrati al giorno contro i 350g assunti dal gruppo con dieta ad alto contenuto di carboidrati.[93] E questi effetti diventano ancora più pronunciati man mano che l’assunzione di carboidrati diminuisce.
Ancora un altro studio degno di nota è stato condotto dai ricercatori della McMaster University, che ha confrontato la dieta ad alto e basso contenuto di carboidrati con soggetti che eseguono allenamenti quotidiani per le gambe. I soggetti che seguono una dieta a basso contenuto di carboidrati (30% delle calorie giornaliere) hanno mostrato tassi più elevati di degradazione proteica e tassi più bassi di sintesi proteica rispetto ai soggetti a dieta ricca di carboidrati (60% delle calorie giornaliere), con conseguente minore crescita muscolare complessiva.[94]
Conclusionie ultime indicazioni:
La linea di fondo è questa: Alcuni studi dimostrano che le diete ricche di grassi possono funzionare per la costruzione muscolare, ma un approccio più ideale e redditizio sembra rimanere quello di seguire una dieta ricca di carboidrati secondo capacità individuali.
I dati empirici ci suggeriscono mediamente la stessa medesima cosa, senza però cadere nell’errore di valutare le innumerevoli variabili soggettive in un unico macro-gruppo di risposta positiva ai regimi prevalentemente glucidici.
Questo significa, in poche parole, che la soggettività genetica non va mai sottovalutata o peggio rinnegata per via di futili credenze pseudoscientifiche o convinzioni personali.
Ciò non toglie, però, che i regimi ipercalorici “High Fat” non portano a sostanziali benefici se non ad un ristretto numero di individui caratterizzati da ipotetiche alterazioni metaboliche rispetto alla media della popolazione. Ma anche in questi casi il “tetto favorevole” del “High Fat” si attesta mediamente al 40% delle calorie totali giornaliere con un totale glucidico del 30-25%.
Ciò che dobbiamo ricordare è che i test alimentari soggettivi con modulazioni macro-caloriche sono utili al fine di attestare una quota singolarmente funzionale per ogni macronutriente. Solo in questo modo, e con una valutazione oggettiva, possiamo capire fin dove possiamo spingerci in quanto a quantità di macronutrienti al fine di ottenerne il massimo dei vantaggi prestativi ed estetici. Questo può significare differenze non da poco in un piano ipercalorico tra Grassi e Carboidrati.
Altri tre esempi di variazione nella percentuale dei macronutrienti in funzione delle risposte soggettive.
Personalmente, ritengo che il range della quota lipidica dovrebbe coprire un arco di grammatura da 0,6 a 2g/Kg. Sempre su base soggettiva e rapporto funzionale tra carboidrati e grassi.
In regimi ipercalorici, con l’obbiettivo di non perdere la flessibilità metabolica, si possono utilizzare, oltre alla “C:G ratio” di Ludovico Lemme, uno schema d’esempio di rapporto come segue:
CHO 4-5g/Kg= Fat 0,6g/Kg
CHO 5-6g/Kg= Fat 0,8g/Kg
CHO 6-7g/Kg= Fat 1g/Kg
CHO 7-8g/Kg= Fat 1,5/Kg
CHO 8-10g/Kg= Fat 2g/Kg
E se volessi tarare il quantitativo lipidico per un regime “Bulk High Fat”? Le linee indicative sarebbero:
CHO 3-4g/Kg= Fat 1-1,2g/Kg
CHO 2.5-3g/Kg= Fat 1,5-1,6g/Kg
CHO 2-2.5g/Kg= Fat 1,8-2g/Kg
CHO 0,5-2g/Kg= Fat 2-2,5g/Kg
Queste sono semplici indicazioni generali estrapolate da dati raccolti empiricamente, nulla di scolpito nella roccia o inconfutabilmente dimostrato.
Prima di chiudere con il presente articolo, è logico che vi dica di fare attenzioni alle fonti lipidiche che assumete o che andrete ad assumere (cosa già detta in precedenza). Nessuno vi sta dicendo di evitare la carne rossa (tagli magri) ma di concentrarvi soprattutto su fonti lipidiche quali tuorlo d’uovo, Olio Extravergine di Oliva, EPA, DHA e MCT (controllandone la composizione ed evitando l’olio di Cocco ricco di Acido Laurico). Fonti proteiche quali salmone selvatico, sgombro, pesce spada e carni da allevamenti “Grass Feed” sono pienamente consigliate, se alla propria portata economica.
Gabriel Bellizzi
Riferimenti:
Dr. Mauro Di Pasquale, bodybuilding.com, accessed February 21, 2007. His articles and books have been translated into Italian language and published in Italy by Sandro Ciccarelli Olympian’s News magazine.
Woodworth‐Hobbs ME, Hudson MB, Rahnert JA, Zheng B, Franch HA, Price SR. Docosahexaenoic acid prevents palmitate‐induced activation of proteolytic systems in C2C12 myotubes. J Nutr Biochem 2014;25:868–74.
Graves E, Hitt A, Pariza MW, Cook ME, McCarthy DO. Conjugated linoleic acid preserves gastrocnemius muscle mass in mice bearing the colon‐26 adenocarcinoma. Res Nurs Health 2005;28:48–55.
Langen RC, Schols AM, Kelders MC, van der Velden JL, Wouters EF, Janssen‐Heininger YM. Muscle wasting and impaired muscle regeneration in a murine model of chronic pulmonary inflammation. Am J Respir Cell Mol Biol 2006;35:689–96.
Whitehouse AS, Smith HJ, Drake JL, Tisdale MJ. Mechanism of attenuation of skeletal muscle protein catabolism in cancer cachexia by eicosapentaenoic acid. Cancer Res 2001;61:3604–9.
Castillero E, Martin AI, Lopez‐Menduina M, Villanua MA, Lopez‐Calderon A. Eicosapentaenoic acid attenuates arthritis‐induced muscle wasting acting on atrogin‐1 and on myogenic regulatory factors. Am J Physiol Regul Integr Comp Physiol 2009;297:R1322–31.
Fiaccavento R, Carotenuto F, Vecchini A, Binaglia L, Forte G, Capucci E, et al. An omega‐3 fatty acid‐enriched diet prevents skeletal muscle lesions in a hamster model of dystrophy. Am J Pathol 2010;177:2176–84.
Briolay A, Jaafar R, Nemoz G, Bessueille L. Myogenic differentiation and lipid‐raft composition of L6 skeletal muscle cells are modulated by PUFAs. Biochim Biophys Acta 2013;1828:602–13.
Watson ML, Coghlan M, Hundal HS. Modulating serine palmitoyl transferase (SPT) expression and activity unveils a crucial role in lipid‐induced insulin resistance in rat skeletal muscle cells. Biochem J 2009;417:791–801.
. Yuzefovych L, Wilson G, Rachek L. Different effects of oleate vs. palmitate on mitochondrial function, apoptosis, and insulin signaling in L6 skeletal muscle cells: role of oxidative stress. Am J Physiol Endocrinol Metab 2010;299:E1096–105.
Mahfouz R, Khoury R, Blachnio‐Zabielska A, Turban S, Loiseau N, Lipina C, et al. Characterising the inhibitory actions of ceramide upon insulin signaling in different skeletal muscle cell models: a mechanistic insight. PLoS One 2014;9:e101865.
De Larichaudy J, Zufferli A, Serra F, Isidori AM, Naro F, Dessalle K, et al. TNF‐alpha‐ and tumor‐induced skeletal muscle atrophy involves sphingolipid metabolism. Skelet Muscle 2012;2:2.
Mebarek S, Komati H, Naro F, Zeiller C, Alvisi M, Lagarde M, et al. Inhibition of de novo ceramide synthesis upregulates phospholipase D and enhances myogenic differentiation. J Cell Sci 2007;120:407–16.
Turpin SM, Ryall JG, Southgate R, Darby I, Hevener AL, Febbraio MA, et al. Examination of ‘lipotoxicity’ in skeletal muscle of high‐fat fed and ob/ob mice. J Physiol 2009;587:1593–605.
Williamson DL, Dungan CM, Jadhav KS. High lipid concentrations regulate skeletal muscle REDD1. FASEB J 2013;27:Supplement 942.1.
Gordon BS, Williamson DL, Lang CH, Jefferson LS, Kimball SR. Nutrient‐induced stimulation of protein synthesis in mouse skeletal muscle is limited by the mTORC1 repressor REDD1. J Nutr 2015;145:708–13.
Leskawa KC, Erwin RE, Buse PE, Hogan EL. Glycosphingolipid biosynthesis during myogenesis of rat L6 cells in vitro. Mol Cell Biochem 1988;83:47–54.
Anastasia L, Papini N, Colazzo F, Palazzolo G, Tringali C, Dileo L, et al. NEU3 sialidase strictly modulates GM3 levels in skeletal myoblasts C2C12 thus favoring their differentiation and protecting them from apoptosis. J Biol Chem 2008;283:36265–71.
Papini N, Anastasia L, Tringali C, Dileo L, Carubelli I, Sampaolesi M, et al. MmNEU3 sialidase over‐expression in C2C12 myoblasts delays differentiation and induces hypertrophic myotube formation. J Cell Biochem 2012;113:2967–78.
Lipina C, Hundal HS. Ganglioside GM3 as a gatekeeper of obesity‐associated insulin resistance: evidence and mechanisms. FEBS Lett 2015;589:3221–7.
Gangoiti P, Bernacchioni C, Donati C, Cencetti F, Ouro A, Gomez‐Munoz A, et al. Ceramide 1‐phosphate stimulates proliferation of C2C12 myoblasts. Biochimie 2012;94:597–607.
Szendroedi J, Yoshimura T, Phielix E, Koliaki C, Marcucci M, Zhang D, et al. Role of diacylglycerol activation of PKCtheta in lipid‐induced muscle insulin resistance in humans. Proc Natl Acad Sci U S A 2014;111:9597–602.
You JS, Lincoln HC, Kim CR, Frey JW, Goodman CA, Zhong XP, et al. The role of diacylglycerol kinase zeta and phosphatidic acid in the mechanical activation of mammalian target of rapamycin (mTOR) signaling and skeletal muscle hypertrophy. J Biol Chem 2014;289:1551–63.
Shad BJ, Smeuninx B, Atherton PJ, Breen L. The mechanistic and ergogenic effects of phosphatidic acid in skeletal muscle. Appl Physiol Nutr Metab 2015;40:1233–41.
Bilim O, Takeishi Y, Kitahara T, Arimoto T, Niizeki T, Sasaki T, et al. Diacylglycerol kinase zeta inhibits myocardial atrophy and restores cardiac dysfunction in streptozotocin‐induced diabetes mellitus. Cardiovasc Diabetol 2008;7:2.
Haemmerle G, Zimmermann R, Hayn M, Theussl C, Waeg G, Wagner E, et al. Hormone‐sensitive lipase deficiency in mice causes diglyceride accumulation in adipose tissue, muscle, and testis. J Biol Chem 2002;277:4806–15.
Marignani PA, Epand RM, Sebaldt RJ. Acyl chain dependence of diacylglycerol activation of protein kinase C activity in vitro. Biochem Biophys Res Commun 1996;225:469–73.
Eichmann TO, Kumari M, Haas JT, Farese RV Jr, Zimmermann R, Lass A, et al. Studies on the substrate and stereo/regioselectivity of adipose triglyceride lipase, hormone‐sensitive lipase, and diacylglycerol‐O‐acyltransferases. J Biol Chem 2012;287:41446–57.
Macrae K, Stretton C, Lipina C, Blachnio‐Zabielska A, Baranowski M, Gorski J, et al. Defining the role of DAG, mitochondrial function, and lipid deposition in palmitate‐induced proinflammatory signaling and its counter‐modulation by palmitoleate. J Lipid Res 2013;54:2366–78.
Rando RR, Young N. The stereospecific activation of protein kinase C. Biochem Biophys Res Commun 1984;122:818–23.
Badin PM, Louche K, Mairal A, Liebisch G, Schmitz G, Rustan AC, et al. Altered skeletal muscle lipase expression and activity contribute to insulin resistance in humans. Diabetes 2011;60:1734–42.
Hyde R, Hajduch E, Powell DJ, Taylor PM, Hundal HS. Ceramide down‐regulates System A amino acid transport and protein synthesis in rat skeletal muscle cells. FASEB J 2005;19:461–3.
Guenther GG, Peralta ER, Rosales KR, Wong SY, Siskind LJ, Edinger AL. Ceramide starves cells to death by downregulating nutrient transporter proteins. Proc Natl Acad Sci U S A 2008;105:17402–7.
Nardi F, Hoffmann TM, Stretton C, Cwiklinski E, Taylor PM, Hundal HS. Proteasomal modulation of cellular SNAT2 (SLC38A2) abundance and function by unsaturated fatty acid availability. J Biol Chem 2015;290:8173–84.
Li F, Duan Y, Li Y, Tang Y, Geng M, Oladele OA, et al. Effects of dietary n‐6:n‐3 PUFA ratio on fatty acid composition, free amino acid profile and gene expression of transporters in finishing pigs. Br J Nutr 2015;113:739–48.
Coll T, Eyre E, Rodriguez‐Calvo R, Palomer X, Sanchez RM, Merlos M, et al. Oleate reverses palmitate‐induced insulin resistance and inflammation in skeletal muscle cells. J Biol Chem 2008;283:11107–16.
Salvado L, Coll T, Gomez‐Foix AM, Salmeron E, Barroso E, Palomer X, et al. Oleate prevents saturated‐fatty‐acid‐induced ER stress, inflammation and insulin resistance in skeletal muscle cells through an AMPK‐dependent mechanism. Diabetologia 2013;56:1372–82.
Wu CL, Cornwell EW, Jackman RW, Kandarian SC. NF‐kappaB but not FoxO sites in the MuRF1 promoter are required for transcriptional activation in disuse muscle atrophy. Am J Physiol Cell Physiol 2014;306:C762–7.
Rivas DA, McDonald DJ, Rice NP, Haran PH, Dolnikowski GG, Fielding RA. Diminished anabolic signaling response to insulin induced by intramuscular lipid accumulation is associated with inflammation in aging but not obesity. Am J Physiol Regul Integr Comp Physiol 2016;310:R561–9.
Liu Y, Chen F, Odle J, Lin X, Zhu H, Shi H, et al. Fish oil increases muscle protein mass and modulates Akt/FOXO, TLR4, and NOD signaling in weanling piglets after lipopolysaccharide challenge. J Nutr 2013;143:1331–9.
Doyle A, Zhang G, Abdel Fattah EA, Eissa NT, Li YP. Toll‐like receptor 4 mediates lipopolysaccharide‐induced muscle catabolism via coordinate activation of ubiquitin–proteasome and autophagy–lysosome pathways. FASEB J 2011;25:99–110.
Mittal A, Bhatnagar S, Kumar A, Lach‐Trifilieff E, Wauters S, Li H, et al. The TWEAK‐Fn14 system is a critical regulator of denervation‐induced skeletal muscle atrophy in mice. J Cell Biol 2010;188:833–49.
Finlin BS, Varma V, Nolen GT, Dube J, Starnes CP, Rasouli N, et al. DHA reduces the atrophy‐associated Fn14 protein in differentiated myotubes during coculture with macrophages. J Nutr Biochem 2012;23:885–91.
Jang YC, Lustgarten MS, Liu Y, Muller FL, Bhattacharya A, Liang H, et al. Increased superoxide in vivo accelerates age‐associated muscle atrophy through mitochondrial dysfunction and neuromuscular junction degeneration. FASEB J 2010;24:1376–90.
Gouspillou G, Sgarioto N, Kapchinsky S, Purves‐Smith F, Norris B, Pion CH, et al. Increased sensitivity to mitochondrial permeability transition and myonuclear translocation of endonuclease G in atrophied muscle of physically active older humans. FASEB J 2014;28:1621–33.
Muoio DM, Way JM, Tanner CJ, Winegar DA, Kliewer SA, Houmard JA, et al. Peroxisome proliferator‐activated receptor‐alpha regulates fatty acid utilization in primary human skeletal muscle cells. Diabetes 2002;51:901–9.
Tardif N, Salles J, Landrier JF, Mothe‐Satney I, Guillet C, Boue‐Vaysse C, et al. Oleate‐enriched diet improves insulin sensitivity and restores muscle protein synthesis in old rats. Clin Nutr 2011;30:799–806.
Park SY, Kim MH, Ahn JH, Lee SJ, Lee JH, Eum WS, et al. The stimulatory effect of essential fatty acids on glucose uptake involves both Akt and AMPK activation in C2C12 skeletal muscle cells. Korean J Physiol Pharmacol: Off J Korean Phys Soc Korean Soc Pharm 2014;18:255–61.
Wang Y, Lin QW, Zheng PP, Zhang JS, Huang FR. DHA inhibits protein degradation more efficiently than EPA by regulating the PPARgamma/NFkappaB pathway in C2C12 myotubes. BioMed Res Int 2013;2013:318981.
Lanza IR, Blachnio‐Zabielska A, Johnson ML, Schimke JM, Jakaitis DR, Lebrasseur NK, et al. Influence of fish oil on skeletal muscle mitochondrial energetics and lipid metabolites during high‐fat diet. Am J Physiol Endocrinol Metab 2013;304:E1391–403.
Lim JH, Gerhart‐Hines Z, Dominy JE, Lee Y, Kim S, Tabata M, et al. Oleic acid stimulates complete oxidation of fatty acids through protein kinase A‐dependent activation of SIRT1‐PGC1alpha complex. Nutr Metab 2013;288:7117–26.
Mizunoya W, Iwamoto Y, Shirouchi B, Sato M, Komiya Y, Razin FR, et al. Dietary fat influences the expression of contractile and metabolic genes in rat skeletal muscle. PLoS One 2013;8:e80152.
Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues. Cell 2011;147:728–41.
Penna F, Costamagna D, Pin F, Camperi A, Fanzani A, Chiarpotto EM, et al. Autophagic degradation contributes to muscle wasting in cancer cachexia. Am J Pathol 2013;182:1367–78.
O’Leary MF, Vainshtein A, Carter HN, Zhang Y, Hood DA. Denervation‐induced mitochondrial dysfunction and autophagy in skeletal muscle of apoptosis‐deficient animals. Am J Physiol Cell Physiol 2012;303:C447–54.
Wenz T, Rossi SG, Rotundo RL, Spiegelman BM, Moraes CT. Increased muscle PGC‐1alpha expression protects from sarcopenia and metabolic disease during aging. Proc Natl Acad Sci U S A 2009;106:20405–10.
McFarland DC, Velleman SG, Pesall JE, Coy CS. Effect of lipids on avian satellite cell proliferation, differentiation and heparan sulfate proteoglycan expression. Comp Biochem Physiol A Mol Integr Physiol 2011;159:188–95.
Peng Y, Zheng Y, Zhang Y, Zhao J, Chang F, Lu T, et al. Different effects of omega‐3 fatty acids on the cell cycle in C2C12 myoblast proliferation. Mol Cell Biochem 2012;367:165–73.
Schevzov G, Kee AJ, Wang B, Sequeira VB, Hook J, Coombes JD, et al. Regulation of cell proliferation by ERK and signal‐dependent nuclear translocation of ERK is dependent on Tm5NM1‐containing actin filaments. Mol Biol Cell 2015;26:2475–90, doi:10.1091/mbc.E14-10-1453.
Parton RG, Way M, Zorzi N, Stang E. Caveolin‐3 associates with developing T‐tubules during muscle differentiation. J Cell Biol 1997;136:137–54.
Oshikawa J, Otsu K, Toya Y, Tsunematsu T, Hankins R, Kawabe J, et al. Insulin resistance in skeletal muscles of caveolin‐3‐null mice. Proc Natl Acad Sci U S A 2004;101:12670–5.
Galbiati F, Volonte D, Minetti C, Chu JB, Lisanti MP. Phenotypic behavior of caveolin‐3 mutations that cause autosomal dominant limb girdle muscular dystrophy (LGMD‐1C). Retention of LGMD‐1C caveolin‐3 mutants within the golgi complex. J Biol Chem 1999;274:25632–41.
Minetti C, Bado M, Broda P, Sotgia F, Bruno C, Galbiati F, et al. Impairment of caveolae formation and T‐system disorganization in human muscular dystrophy with caveolin‐3 deficiency. Am J Pathol 2002;160:265–70.
Park DS, Woodman SE, Schubert W, Cohen AW, Frank PG, Chandra M, et al. Caveolin‐1/3 double‐knockout mice are viable, but lack both muscle and non‐muscle caveolae, and develop a severe cardiomyopathic phenotype. Am J Pathol 2002;160:2207–17.
Capozza F, Combs TP, Cohen AW, Cho YR, Park SY, Schubert W, et al. Caveolin‐3 knockout mice show increased adiposity and whole body insulin resistance, with ligand‐induced insulin receptor instability in skeletal muscle. Am J Physiol Cell Physiol 2005;288:C1317–31.
Gazzerro E, Sotgia F, Bruno C, Lisanti MP, Minetti C. Caveolinopathies: from the biology of caveolin‐3 to human diseases. Eur J Hum Genet: EJHG 2010;18:137–45.
Markworth JF, Cameron‐Smith D. Arachidonic acid supplementation enhances in vitro skeletal muscle cell growth via a COX‐2‐dependent pathway. Am J Physiol Cell Physiol 2013;304:C56–67.
Hausman RE, Velleman SG. Prostaglandin E1 receptors on chick embryo myoblasts. Biochem Biophys Res Commun 1981;103:213–8.
Santini MT, Indovina PL, Hausman RE. Prostaglandin dependence of membrane order changes during myogenesis in vitro. Biochim Biophys Acta 1988;938:489–92.
Hausman RE, elGendy H, Craft F. Requirement for G protein activity at a specific time during embryonic chick myogenesis. Cell Differ Dev: The Official J Int Soc Dev Biol 1990;29:13–20.
Iannotti FA, Silvestri C, Mazzarella E, Martella A, Calvigioni D, Piscitelli F, et al. The endocannabinoid 2‐AG controls skeletal muscle cell differentiation via CB1 receptor‐dependent inhibition of Kv7 channels. Proc Natl Acad Sci U S A 2014;111:E2472–81.
Murton AJ, Marimuthu K, Mallinson JE, Selby AL, Smith K, Rennie MJ, et al. Obesity appears to be associated with altered muscle protein synthetic and breakdown responses to increased nutrient delivery in older men, but not reduced muscle mass or contractile function. Diabetes 2015;64:3160–71.
McGlory C, Galloway SD, Hamilton DL, McClintock C, Breen L, Dick JR, et al. Temporal changes in human skeletal muscle and blood lipid composition with fish oil supplementation. Prostaglandins Leukot Essent Fatty Acids 2014;90:199–206.
Jans A, van Hees AM, Gjelstad IM, Sparks LM, Tierney AC, Riserus U, et al. Impact of dietary fat quantity and quality on skeletal muscle fatty acid metabolism in subjects with the metabolic syndrome. Metabolism 2012;61:1554–65.
Rasic‐Milutinovic Z, Perunicic G, Pljesa S, Gluvic Z, Sobajic S, Djuric I, et al. Effects of N‐3 PUFAs supplementation on insulin resistance and inflammatory biomarkers in hemodialysis patients. Ren Fail 2007;29:321–9.
Dangardt F, Chen Y, Gronowitz E, Dahlgren J, Friberg P, Strandvik B. High physiological omega‐3 fatty acid supplementation affects muscle fatty acid composition and glucose and insulin homeostasis in obese adolescents. J Nutr Metab 2012;2012:395757.
Jimenez‐Gomez Y, Cruz‐Teno C, Rangel‐Zuniga OA, Peinado JR, Perez‐Martinez P, Delgado‐Lista J, et al. Effect of dietary fat modification on subcutaneous white adipose tissue insulin sensitivity in patients with metabolic syndrome. Mol Nutr Food Res 2014;58:2177–88.
Kawabata F, Neya M, Hamazaki K, Watanabe Y, Kobayashi S, Tsuji T. Supplementation with eicosapentaenoic acid‐rich fish oil improves exercise economy and reduces perceived exertion during submaximal steady‐state exercise in normal healthy untrained men. Biosci Biotechnol Biochem 2014;78:2081–8.
Henry R, Peoples GE, McLennan PL. Muscle fatigue resistance in the rat hindlimb in vivo from low dietary intakes of tuna fish oil that selectively increase phospholipid n‐3 docosahexaenoic acid according to muscle fibre type. Br J Nutr 2015;114:873–84.
Peoples GE, McLennan PL. Long‐chain n‐3 DHA reduces the extent of skeletal muscle fatigue in the rat in vivo hindlimb model. Br J Nutr 2014;111:996–1003.
Langelaan ML, Boonen KJ, Rosaria‐Chak KY, van der Schaft DW, Post MJ, Baaijens FP. Advanced maturation by electrical stimulation: differences in response between C2C12 and primary muscle progenitor cells. J Tissue Eng Regen Med 2011;5:529–39.
Il più delle volte risulta inutile ribadire banalità largamente conosciute, ma in questo caso era d’obbligo sottolinearne il fatto dal momento che tornerà utile anche al fine di comprendere come la maggior parte dei (presunti) preparatori agisca quando si parla di PEDs.
Il GH, in modo particolare, è avvolto da dicerie di ogni genere. Dalle convinzioni sul momento in cui “inizia a funzionare” e la leggenda (fuori da ogni base fisiologica e attività espressiva iper-indotta iatrogenamente) del “assottiglia la pelle”. Ma la cosa di cui vorrei parlare riguarda i dosaggi. Nell’agonismo se ne sentono e vedono di tutti i tipi, dosaggi mostruosi da 10-20UI/die portatori di neuropatie croniche e probabili stati diabetici subclinici o patologici. La cosa triste è che tali dosaggi vengono indicati come “funzionali” solo perchè danno l’effetto ricercato che spesso e volentieri si confonde con quello indotto dallo svariato numero di farmaci cosomministrati e dalla additività tra questi. In poche parole: l’effetto lo avrebbero raggiunto comunque e nello stesso arco temporale utilizzando 1/10 della dose somministrata.
Si, mi dispiace infrangere le vostre fantasie ma il GH ha un limite nello stimolo massimo della lipolisi molto sottile…
GH e suo limite di dosaggio per il massimo stimolo lipolitico:
In effetti è stato calcolato un tetto massimo alla lipolisi indotta da una singola somministrazione di GH, ed è molto più bassa di quanto la maggior parte delle persone pensi.
Come riportato nello studio “Pharmacokinetics and acute lipolytic actions of growth hormone. Impact of age, body composition, binding proteins, and other hormonesvedere” nel quale si è valutata la farmacocinetica e l’azione lipolitica acuta del GH, si verificano significativi effetti dose-risposta confrontando l’area incrementale sotto la curva sia degli acidi grassi liberi che del 3-idrossi-butirrato nel sangue dopo la somministrazione di 0, 1 , e 3mcg/kg di GH, mentre non si osservano differenze tra le risposte dopo 3 e 6mcg/kg di GH.[1]
Questi dosaggi sono stati somministrati per via endovenosa.
1mg di Somatropina corrisponde a 3UI (Unità Internazionali) della medesima, come si può anche leggere nelle informazioni riferite al Norditropin presenti di seguito.[2]
Quindi, questo significa che per un soggetto di 100kg, la lipolisi è stimolata al massimo con circa 300mcg di Somatropina somministrata endovena, che equivale a 0,9UI di GH di grado farmaceutico IV (endovena).
Occorre quindi calcolare la dose sottocutanea corrispondente in base ai dati di confronto data dalla biodisponibilità e bioattività che abbiamo sugli esseri umani a cui è stato somministrato hGH.
Ovviamente i dosaggi somministrati endovena non sono rappresentativi di quali sarebbero i dosaggi ideali tramite il metodo di somministrazione molto più realistico e tollerabile quale è quello sottocutaneo. In uno studio, la disponibilità media stimata di hGH iniettato per via sottocutanea ha dimostrato di essere del 63% di quella di hGH somministrato per via endovenosa. dopo aver corretto le differenze nella dose di GH. [3] Un altro studio ha mostrato che la disponibilità di hGH iniettato per via sottocutanea è circa il 70% di quella di hGH somministrato per via endovenosa.[4]
Quindi, per un uomo di 100kg, i benefici della perdita di grasso sarebbero massimizzati per ogni somministrazione iniettando circa 1,35UI di GH (arrotondabili a massimo 2UI). Esiste infatti un periodo di refrattarietà durante il quale le cellule non danno risposta ad altro impulso lipolitico dato dal GH esogeno.
Conclusioni pratiche:
Di conseguenza, la risposta massima dello stimolo della lipolisi per somministrazione si aggira intorno a circa 1.5-2UI (3-4UI/die totali) in un individuo di 100Kg. Questo rispecchia in parte quanto osservato aneddoticamente ed annotato empiricamente con la somministrazione giornaliera di GH ad un dosaggio di 4UI/die divise in due somministrazioni uguali durante periodi di “Cut” in molti atleti di diverse categorie. Ciò vuole anche dire, però, che la maggior parte dei soggetti potrebbero avere risposte ottimali con la somministrazione di appena 2UI/die di GH. Parlo, ovviamente, di GH di grado farmaceutico, cioè equivalente al mcg al contenuto riportato in etichetta, non così abbondante nel mercato nero.
Le risposte temporali alla riduzione del grasso corporeo per azione del GH non sono dovute al dosaggio ma alla percentuale di grasso di partenza e alla distanza tra questo ed il punto percentuale prefissato da raggiungere oltre che dalla additività con altre molecole aventi effetti lipolitici e/o termogenici: più è breve e meno tempo di somministrazione sarà necessario. Ovviamente, i tempi di risposta sono maggiori rispetto ad atleti non trattati, mi pare ovvio e scontato. E, lo ripeto, vanno anche considerate nella somma dell’effetto le altre molecole co-somministrate e aventi attività lipolitica (diretta o indiretta) e/o termogenica.
Ma questo vale anche per le donne? Di questo me ne occuperò in un altro articolo ma vi anticipo già che in termini assoluti il dosaggio non ha variabili significative ma circostanze gestionali diverse.
Nel giugno 2021 è stato pubblicato un eccellente lavoro di Guillermo Escalante, Scott W. Stevenson, Christopher Barakat, Alan A. Aragon e Brad J. Schoenfeld che analizza le comuni pratiche applicate dai bodybuilder nella settimana precedente la gara (Peak Week) esponendone la logica applicativa in base alle evidenze scientifiche in nostro possesso.[Peak week recommendations for bodybuilders: an evidence based approach | BMC Sports Science, Medicine and Rehabilitation | Full Text (biomedcentral.com)] Ho deciso quindi di scrivere un articolo dettagliato sulla settimana pre-contest, utilizzando come base lo studio sopra citato e aggiungendo la mia ricerca personale, al fine di spiegarne le modalità di gestione migliori che, senz’altro, potranno tornare utili tanto agli atleti quanto ai preparatori.
Punto di partenza:
Il Bodybuilding è uno sforzo competitivo per il miglioramento della composizione corporea la quale verrà giudicata con parametri che comprendono la combinazione delle dimensioni muscolari, la simmetria, la “condizione” (bassi livelli di grasso corporeo) e presentazione sul palco. Per avere successo, i concorrenti devono presentare la loro forma fisica migliore durante il giorno (o i giorni) della competizione. I bodybuilder impiegano tipicamente periodi di 8-22 o più settimane di preparazione in cui la dieta e i programmi di esercizio vengono modificati dalla off season nel tentativo di perdere la maggior percentuale di grasso corporeo e guadagnare o mantenere la massa muscolare scheletrica [1,2,3,4,5,6 ,7,8,9,10]. Negli ultimi giorni di preparazione, i concorrenti implementano interventi per “tirare” il loro corpo nel tentativo di massimizzare l’estetica nel giorno della gara [11,12,13,14]. Gli interventi spesso utilizzati includono l’alterazione dei loro regimi di esercizio e l’assunzione di macronutrienti, acqua ed elettroliti con gli obiettivi di:
1-massimizzare il contenuto di glicogeno muscolare come mezzo per migliorare la “pienezza” muscolare (cioè il volume);
2-ridurre al minimo l’acqua sottocutanea (nel tentativo di sembrare “asciutti” anziché “acquosi”, migliorando così l’estetica muscolatura) e
3-ridurre al minimo il gonfiore addominale per mantenere un girovita più piccolo e ottimizzare le proporzioni fisiche e l’estetica generale [11, 12, 14,15 ,16,17].
Sebbene i concorrenti possano utilizzare metodi naturali per raggiungere questi obiettivi, sappiamo benissimo che vi è un ampio uso segnalata di auto-prescrizione di farmaci per il miglioramento delle prestazioni/estetica [8, 18, 19, 20, 21].
Uno studio osservazionale ha raccolto informazioni sulle strategie nutrizionali della Peak Week e dei giorni di gara tra 81 bodybuilder natural (maschi= 59, femmine = 22) tramite un questionario di 34 elementi; l’indagine ha elencato le strategie di picco comunemente utilizzate e ha fornito spazio aggiuntivo per informazioni qualitative [11]. La stragrande maggioranza dei partecipanti (93,8%) ha riferito di aver utilizzato una strategia di picco la settimana prima della competizione (denominata appunto “Peak Week”), con la manipolazione di Carboidrati (CHO), Acqua e/o Sodio segnalata più comunemente [ 11]. L’obiettivo primario dichiarato della manipolazione dei CHO era massimizzare le concentrazioni di glicogeno muscolare utilizzando principi simili al carico dei CHO classico [11]. Inoltre, i concorrenti hanno manipolato l’assunzione di Acqua e/o Sodio nel tentativo di indurre un effetto diuretico/poliuria per eliminare l’acqua superflua.[11]
In un altro studio, i ricercatori hanno condotto interviste approfondite per identificare e descrivere diverse strategie dietetiche utilizzate da sette culturisti maschi natural durante la off season, la stagione, la Peak Week e il post-season [14]. Durante la Peak Week, sei partecipanti hanno riferito di aver utilizzato un regime di carico di carboidrati modificato per tentare di aumentare il contenuto di glicogeno. Inoltre, tutti i partecipanti hanno riferito di aver manipolato l’assunzione di acqua mentre tre hanno manipolato contemporaneamente l’assunzione di sodio nel tentativo di ridurre l’acqua corporea nella speranza di creare un aspetto più “asciutto”.[14]
Sebbene esistano molti protocolli delle Peak Week al fine di tentare il miglioramento dell’estetica, mancano ricerche sull’efficacia e la sicurezza dei metodi comunemente usati dai bodybuilder. Dal momento che lo studio in questione non tratta i bodybuilder supplementati farmacologicamente, aggiungerò delle note esplicative sui metodi aggiuntivi utilizzati da questi atleti.
Lo scopo di questo articolo è:
1-rivedere la letteratura attuale sui protocolli di picco più comunemente impiegati dai bodybuilder;
2-fornire raccomandazioni basate sull’evidenza per le strategie di picco pre-gara per concorrenti e preparatori.
Manipolazione dei Carboidrati:
La manipolazione dell’assunzione di Carboidrati è una popolare strategia di picco pre-gara diffusa tra i bodybuilder [11, 12, 14]. La strategia, generalmente adottata durante la settimana che precede la competizione, prevede la limitazione sostanziale dell’assunzione di Carboidrati per diversi giorni (spesso indicata come fase di esaurimento o scarica) seguita da un breve periodo di consumo elevato di Carboidrati, con l’obiettivo di ottenere una supercompensazione dei livelli di glicogeno quando i carboidrati sono “stoccati” [22]. I livelli di glicogeno muscolare a riposo con una dieta mista (normale) sono ~ 130mmol/kg di muscolo (peso umido) in individui allenati (un po’ più alti dei soggetti sedentari) [23], o circa 23g di glicogeno (unità di glucosio) per chilogrammo di tessuto muscolare. Il glicogeno muscolare è organizzato nella cellula in frazioni subcellulari [24] e immagazzinato come un complesso di glicogeno-glicogenina (“granulo”) [25] che crea un effetto osmotico il quale attira acqua nella cellula mentre il glicogeno viene immagazzinato [26, 27], aumentando così il volume delle cellule muscolari. Le prime ricerche hanno suggerito che ogni grammo di glicogeno muscolare immagazzinato è accompagnato da circa 3-4g di acqua intracellulare [28]. Questo è superiore al valore comunemente indicato di 2,7g di acqua per grammo di glicogeno, a volte arrotondato a 3g di acqua per grammo di glicogeno, derivato da studi sul fegato di ratto [29, 30]. Tuttavia, i livelli di glicogeno muscolare risultanti dopo il carico di glicogeno sono altamente variabili [31], forse a causa della complessità sottostante all’accumulo di glicogeno intramuscolare [25]. Allo stesso modo, mentre è chiaro che il carico di glicogeno può aumentare il contenuto di acqua intracellulare [31], lo spessore muscolare [15] e le stime della massa corporea magra (LBM) [32], l’entità relativa dell’idratazione intracellulare in grammi di acqua per grammo di glicogeno può variare così tanto da non essere statisticamente correlato con il contenuto di glicogeno.[30]
Una panoramica semplificata del metabolismo del glicogeno a riposo e durante l’esercizio. Il sarcolemma separa l’interno della cellula muscolare dal liquido interstiziale che circonda la cellula. A riposo (lato sinistro), il consumo di carboidrati stimola il rilascio di insulina dal pancreas. Le molecole di insulina si legano ai recettori dell’insulina incorporati nel sarcolemma. Quel legame innesca una cascata di risposte intracellulari che provocano il movimento dei trasportatori del glucosio GLUT4 dall’interno della cellula muscolare nel sarcolemma, consentendo al glucosio di spostarsi nella cellula. Una volta all’interno della cellula muscolare, le molecole di glucosio sono pronte per essere stoccate sotto forma di glicogeno. La glicogenina è un enzima che forma il centro delle particelle di glicogeno, consentendo la formazione iniziale di filamenti di glicogeno. Durante l’esercizio (lato destro), i trasportatori GLUT4 si spostano nel sarcolemma senza l’assistenza dell’Insulina, favorendo l’assorbimento del glucosio nella cellula. Contemporaneamente, la degradazione del glicogeno aumenta in risposta ai cambiamenti nella concentrazione dei metaboliti all’interno della cellula. Le molecole di glucosio dal sangue e quelle rilasciate dal glicogeno vengono ossidate per produrre le molecole di adenosina trifosfato (ATP) necessarie per sostenere la contrazione muscolare.
Sebbene la ricerca controllata sull’argomento sia limitata a ciò che è ottimale per i bodybuilder, le prove attuali sembrano indicare un potenziale beneficio della manipolazione dei carboidrati come strategia di picco. Una serie di casi esaminati da Bamman et al., i quali hanno esaminato sei bodybuilder maschi, ha fornito il supporto iniziale di un effetto benefico [1]. Secondo quanto riferito, i bodybuilder si sono impegnati in un protocollo di carico di carboidrati tre giorni prima della competizione (assunzione media di ~ 290g/giorno). Le misurazioni degli ultrasuoni effettuate 24-48 ore in questo periodo di carico di carboidrati hanno mostrato un aumento del 4,9% dello spessore del muscolo bicipite brachiale rispetto alle misurazioni ottenute sei settimane prima. Sebbene questi risultati sembrino suggerire che il protocollo di carico di carboidrati sia stato efficace nel migliorare in modo acuto la dimensione muscolare, va notato che il lungo intervallo tra le sessioni di test rende impossibile trarre conclusioni sulla causalità a questo proposito. Inoltre, gli autori dello studio non hanno valutato l’assunzione di carboidrati durante la fase di esaurimento dei carboidrati, offuscando ulteriormente gli effetti diretti del protocollo di carico. Pertanto, sebbene i risultati siano intriganti, il livello di prove a sostegno può essere considerato basso.
Un recente studio quasi sperimentale di de Moraes et al. [15] getta una luce più obiettiva sull’argomento. Ventiquattro bodybuilder dilettanti di alto livello sono stati divisi in base al fatto se avessero o meno manipolato i carboidrati come strategia di picco; il gruppo che ha manipolato i carboidrati ha impiegato una fase di esaurimento di tre giorni (che porta immediatamente al giorno del peso) seguita da una fase di carico di 24 ore (che porta al giorno della gara). Lo spessore muscolare è stato misurato sia al momento del peso che il giorno della gara. Inoltre, le foto dei concorrenti scattate in questi momenti sono state mostrate a un gruppo di giudici federati di bodybuilding, che hanno valutato soggettivamente il loro fisico; da notare, i giudici erano ciechi alle pratiche nutrizionali dei concorrenti. I risultati hanno mostrato un aumento del 3 % della dimensione muscolare della parte superiore delle braccia per coloro che hanno manipolato l’assunzione di carboidrati prima della competizione rispetto a nessun cambiamento in coloro che non lo hanno fatto. Inoltre, solo il gruppo che ha manipolato l’assunzione di carboidrati ha mostrato miglioramenti nelle misure estetiche soggettive, come determinato dall’ispezione visiva delle foto. Una potenziale limitazione dello studio è che i soggetti non sono stati sottoposti a test anti-doping prima della competizione; pertanto, non è noto se l’uso di steroidi anabolizzanti e/o altre sostanze sintetiche (ad es. synthol) possa aver influenzato i risultati. Gli studi futuri dovrebbero accertare tramite autovalutazione, poligrafo e/o analisi del sangue lo stato di libero/migliorato dei soggetti ed escludere o confrontare i risultati in base all’uso di steroidi da parte del soggetto nonché all’uso di altri farmaci che possono influenzare il bilancio idrico.
Recentemente, Schoenfeld ed Escalante hanno condotto un caso di studio in cui hanno seguito un bodybuilder natural di alto livello nel corso della sua preparazione al contest [33]. A partire dalla settimana prima della data della competizione, il concorrente ha ridotto notevolmente l’assunzione di carboidrati a < 50g/giorno per 3 giorni (domenica, lunedì, martedì) e poi ha eseguito una ricarica dei carboidrati a una quantità > 450g/giorno nei successivi 2 giorni (Mercoledì e giovedì). Simile alla ricerca precedente, la valutazione ecografica ha mostrato che la strategia di picco aumentava notevolmente lo spessore muscolare. In questo particolare caso di studio, gli aumenti sono stati del 5% negli arti superiori e del ~ 2 % negli arti inferiori; a causa delle limitate prove disponibili, è difficile fornire un motivo razionale per cui c’era una differenza tra i gruppi muscolari. Dati i risultati soggettivi riportati da de Moraes et al. [15], si può dedurre che questi risultati erano probabilmente significativi dal punto di vista della concorrenza.
Quando si considera la totalità della ricerca attuale, l’evidenza suggerisce che la manipolazione dei carboidrati è una valida strategia di picco per aumentare il volume della massa muscolare il giorno della gara; tuttavia, l’evidenza dovrebbe essere considerata preliminare data la relativa scarsità di studi pubblicati sull’argomento. Inoltre, la strategia può portare a un aumento dei sintomi gastrointestinali come dolore addominale, bruciore di stomaco, stitichezza e diarrea [15], che a loro volta possono influenzare negativamente la capacità di eseguire in modo ottimale la preparazione al giorno della gara e il contest stesso. Pertanto, i concorrenti dovrebbero sperimentare la strategia con almeno 2-4 settimane di anticipo per determinarne gli effetti a livello individuale e apportare le modifiche necessarie secondo necessità.
Manipolazione di acqua e sodio:
Acqua e sodio sono frequentemente manipolati dai bodybuilder, indipendentemente o contemporaneamente, impiegando una varietà di strategie che comportano il “carico” e la limitazione di entrambi [11], con l’obiettivo di ridurre al minimo l’acqua sottocutanea per massimizzare la definizione del muscolo scheletrico sottostante [8, 11, 12 , 14, 19, 20]. È noto che diversi bodybuilder si auto-prescrivono diuretici farmaceutici per facilitare il processo [8, 19,20,21, 34, 35]. I bodybuilder possono anche impiegare queste strategie per scendere a classi di peso inferiori, il che può fornire un vantaggio competitivo se il concorrente è in grado di recuperare parte del peso sotto forma di volume intramiocellulare (“riempimento” tramite glicogeno e/o stoccaggio di trigliceridi intramiocellulari) prima della competizione. Sebbene l’acqua e il sodio siano due componenti dietetici separati, è fondamentale comprendere che la manipolazione di una variabile influenza l’altra; quindi, esamineremo insieme queste due variabili.
In un’indagine precedentemente citata sulle strategie delle Peak Week e dei giorni di gara utilizzate dai bodybuilder natural, la manipolazione dell’acqua è stata la seconda strategia più popolare implementata (dietro la manipolazione dei carboidrati) [11]. I ricercatori hanno riferito che i concorrenti hanno implementato il carico dell’acqua (65,4%), la restrizione dell’acqua (32,1%) o entrambi (25%) per ottenere un aspetto “asciutto”. La quantità di acqua consumata durante la fase di carico variava da 4 a 12L al giorno ed era tipicamente seguita da restrizioni idriche di 10-24 ore prima della competizione. Oltre alla manipolazione dell’acqua, i ricercatori hanno anche riferito che i concorrenti utilizzavano la restrizione di sodio (13,6 %), il carico di sodio (18,5 %), o entrambi (6,2 %) senza un ordine temporale coerente per il regime di carico/restrizione del sodio (un errore limitante); tuttavia, la manipolazione del sodio veniva generalmente praticata tre o quattro giorni prima della competizione. È stato segnalato anche l’uso del tè al dente di leone per le sue presunte proprietà diuretiche.
Nello studio precedentemente discusso di Mitchell et al. [14], i ricercatori hanno riferito che il 100% dei partecipanti (n = 7) ha utilizzato la pratica del carico e del taglio dell’acqua durante la Peak Week. Questa strategia prevedeva di bere >10L di acqua al giorno all’inizio della settimana e quindi di ridurre l’assunzione ogni giorno successivo prima della competizione. La teoria alla base di questa pratica era quella di consumare quantità superflue di acqua per aumentare naturalmente l’escrezione di liquidi nel tentativo di espellere preferenzialmente l’acqua sottocutanea; tuttavia, i partecipanti hanno riferito che i risultati di questa strategia erano in gran parte non significativi [14]. Dei sette partecipanti che hanno manipolato l’acqua durante la Peak Week, tre (42,8%) hanno anche manipolato il sodio per aiutare a rimuovere l’acqua sottocutanea [14]. Hanno riferito di aumentare notevolmente l’assunzione di sodio per i primi tre giorni della Peak Week, seguita da una completa restrizione dell’assunzione di sale per i tre giorni prima della competizione; tuttavia, i risultati sono stati incoerenti e i partecipanti hanno dichiarato che non avrebbero manipolato il sodio in futuro [14]. Si noti che la decisione unanime dei partecipanti di abbandonare queste strategie di manipolazione dell’acqua e del sodio suggerisce che probabilmente non le avevano né eseguite né perfezionate in precedenza (ad esempio, come prova o durante la Peak Week per un’altra competizione).
Altre ricerche supportano i risultati degli studi di cui sopra. Probert et al. ha condotto un sondaggio su 382 bodybuilder competitivi insieme a interviste personali di 30 dei partecipanti e ha riferito che i bodybuilder si sono spesso impegnati in pratiche di deplezione del sodio e disidratazione nei giorni precedenti la competizione [12]. Sebbene i partecipanti abbiano riconosciuto i rischi di queste strategie, le hanno minimizzate come pratiche temporanee ma necessarie [12]. In effetti, i casi clinici documentano condizioni potenzialmente pericolose per la vita dovute a pratiche estreme di manipolazione dell’acqua e del sodio [19, 20]. In un caso, un bodybuilder maschio di 35 anni si è presentato al pronto soccorso dopo essersi sentito debole, stordito e aver avvertito crampi muscolari dolorosi mentre posava durante una gara di bodybuilding; i test hanno rivelato onde T di picco sull’elettrocardiogramma (ECG), iperkaliemia (alti livelli di potassio), iponatriemia (bassi livelli di sodio nel sangue), intossicazione da acqua e rabdomiolisi [20]. Il bodybuilder ha riferito di aver bevuto 12 litri di acqua al giorno per sette giorni prima della competizione insieme a 100 mg al giorno di Spironolattone (un diuretico da prescrizione risparmiatore di potassio) e scarico del sale per due giorni prima della competizione; è stato curato, stabilizzato e dimesso con successo [20]. In un altro caso, un bodybuilder professionista di 26 anni è stato trasportato al pronto soccorso il giorno dopo una gara a causa di palpitazioni cardiache e incapacità di stare in piedi a causa della difficoltà nel muovere le estremità [19]. Ha riferito l’assunzione orale di 2 × 80mg di Furosemide (un diuretico da prescrizione) 48 e 24 ore prima della competizione con l’obiettivo di migliorare la definizione muscolare; ha perso 5-6 kg di peso corporeo a causa della nicturia [19]. I test hanno rivelato ipokaliemia grave (bassi livelli di potassio; al contrario dell’iperkaliemia nel caso di studio discusso in precedenza probabilmente dovuto all’uso di un diuretico dell’ansa rispetto a un diuretico risparmiatore di potassio), iperglicemia (livelli elevati di glucosio nel sangue), iperlattatemia (alti livelli di lattato nel sangue) e tachicardia sinusale con onde U pronunciate all’ECG compatibili con ipokaliemia [19]. Sebbene l’ipokaliemia sia una condizione potenzialmente pericolosa per la vita, il bodybuilder è stato trattato con successo e dimesso la mattina successiva [19].
Nonostante le varie strategie riportate dai bodybuilder per manipolare l’acqua e il sodio allo scopo di sembrare “pieni e asciutti”, le prove attuali non indicano che queste pratiche siano specificamente efficaci e/o sicure. Inoltre, sebbene diverse strategie di manipolazione dell’acqua e del sodio siano state pubblicate da un certo numero di preparatori di bodybuilding che hanno lavorato con bodybuilder di grande successo [16, 17, 36], né l’efficacia né la sicurezza di queste diverse metodologie sono state valutate scientificamente. Quindi, i principi fisiologici della regolazione dei fluidi corporei devono essere considerati quando si tenta di formulare strategie per promuovere un aspetto “pieno e asciutto”, e queste strategie possono essere discordanti con quelle attualmente utilizzate dai bodybuilder e/o suggerite dai loro preparatori.
Il contenuto di acqua corporea totale (TBW) rappresenta circa il 60 % del peso corporeo medio di una persona ed è costituito da acqua intracellulare (ICW) (~ 67 %) e acqua extracellulare (ECW) (~ 33 %). L’ECW è ulteriormente compartimentato nel fluido interstiziale che circonda le cellule (~ 25 %) e il plasma sanguigno (~ 8 %) [37, 38]. Quindi, dal punto di vista di un bodybuilder, ridurre al minimo il fluido interstiziale extracellulare che circonda i miociti, in particolare l’acqua sottocutanea, preservando o aumentando l’ICW intramiocellulare rappresenta lo scenario ideale per un aspetto “pieno e asciutto”, cioè, per cui l’aspetto della muscolarità è massimizzato . Sebbene questo concetto possa sembrare un compito semplice da realizzare manipolando solo l’acqua e il sodio, potrebbero essere necessarie altre strategie incentrate sull’ottimizzazione del volume intramiocellulare (cioè quelle mirate al glicogeno intramiocellulare, ai trigliceridi e al contenuto di potassio) insieme alla manipolazione dell’acqua. e sodio per migliorare l’aspetto della muscolosità.
Compartimenti dei fluidi corporei. Nell’uomo adulto “medio”, i domini del fluido intracellulare (ICF) e del fluido extracellulare (ECF) sono costituiti da circa il 57 e il 43% dell’acqua corporea totale (TBW). Il compartimento ECF è ulteriormente suddiviso in liquido interstiziale (ISF)/linfa, plasma, tessuto osseo e connettivo, tessuto adiposo e acqua transcellulare. Il muscolo scheletrico predomina l’ICF. Le percentuali sono percento di TBW. GR, globuli rossi.
Durante la normale omeostasi fluido-elettrolitica, il compartimento extracellulare contiene la maggior parte del sodio (Na+), cloruro (Cl-) e bicarbonato (HCO3-), mentre il compartimento intracellulare contiene la maggior parte dell’acqua, potassio (K+) e fosfato ( PO43−) [39]. Sebbene entrambi i compartimenti contengano tutti i suddetti composti, la quantità di ciascuno varia tra i compartimenti in modo tale che la concentrazione totale di soluti (osmolarità) sia la stessa [39]. I meccanismi omeostatici controllano l’equilibrio idrico ed elettrolitico per garantire che la TBW e l’osmolarità corporea totale (TBO) rimangano equilibrate e l’acqua si ridistribuisca tra i compartimenti intracellulari ed extracellulari in modo tale che l’osmolarità dei fluidi corporei si avvicini alla TBO [37]. Infatti, Costill et al. hanno studiato le perdite muscolari di acqua ed elettroliti mentre i partecipanti pedalavano in una camera ambientale calda per perdere il 2,2 (% (fase 1), il 4,1 % (fase 2) e il 5,8 % (fase 3) del loro peso corporeo in un periodo stimato di 5,5 ore [40] . Quando i partecipanti hanno perso il 2,2% del loro peso corporeo entro la prima ~ 1,5 h nella fase 1, il 30% dell’acqua persa era ICW mentre il 70% era ECW [40]. Tuttavia, il rapporto tra ICW ed ECW perso è diventato 52 % ICW/48 % ECW allo stadio 2 (~ 3.5 h mark) e 50 % ICW/50 % ECW allo stadio 3 (~ 5.5 h mark) [40]. Gli autori hanno affermato che la grande perdita di ICW nel muscolo allo stadio 1 può essere spiegata dalla significativa perdita di contenuto di glicogeno muscolare (che contiene acqua) dalla pre-disidratazione a 115 mmol/kg fino a 76 mmol/kg; tuttavia, i livelli di contenuto di glicogeno muscolare sono scesi a una velocità molto inferiore a 73 mmol/kg allo stadio 2 e 61 mmol/kg allo stadio 3 quando il rapporto o ICW:ECW si è stabilizzato [40]. Pertanto, il rapporto tra la perdita ECW e ICW sembra rimanere vicino a 1:1 poiché i livelli di glicogeno si stabilizzano nel tempo e vengono raggiunti livelli più elevati di disidratazione. Pertanto, sembra che la ritenzione del glicogeno muscolare, evitando l’esercizio che si basa fortemente sull’uso del glicogeno, possa essere importante se i metodi di perdita di acqua devono effettuare una perdita favorevole di ECW rispetto a ICW (ECW > ICW) in modo tale che la dimensione muscolare venga mantenuta mentre l’ECW interstiziale viene preferibilmente perso, migliorando l’aspetto della “definizione” muscolare. Allo stesso modo, l’immagazzinamento e la ritenzione del glicogeno muscolare dipendono fortemente dalla disponibilità di potassio (un catione intracellulare primario – vedi sopra) [41,42,43,44,45,46], quindi sembra che garantire un’adeguata assunzione di potassio durante le procedure di carico di carboidrati e disidratazione sia fondamentale per ottimizzare l’aspetto scenico.
È importante sottolineare che se le alterazioni dell’osmolarità plasmatica (attraverso i cambiamenti nell’acqua corporea totale e degli elettroliti) raggiungono una soglia fisiologica, allora una complessa rete neuroendocrina in tutto il corpo, nel cervello, vasi sanguigni, reni e ghiandole endocrine, risponderà per stabilizzarlo [47] . L’osmolarità plasmatica è influenzata dalle variazioni (aumento o diminuzione) della concentrazione di soluti (cioè sodio) nel sangue nonché dalle variazioni del volume del fluido; il volume del fluido è influenzato dall’acqua corporea totale (TBW) [48]. L’osmolarità plasmatica può aumentare per un’eccessiva perdita di acqua o per un aumento significativo dell’assunzione di sodio; al contrario, l’osmolarità plasmatica può diminuire con un consumo insufficiente di elettroliti o un’eccessiva assunzione di acqua [49]. L’osmolarità plasmatica e la pressione sanguigna sono regolate in modo tale che l’aumento dell’osmolarità plasmatica si traduca in una diminuzione della pressione sanguigna e viceversa [49]. Inoltre, le variazioni della pressione sanguigna mediate dallo spostamento dell’osmolarità plasmatica sono contrastate dai barocettori arteriosi e renali [50].
Durante la disidratazione, come potrebbe essere impiegato durante la Peak Week, l’osmolarità plasmatica aumenta, la pressione sanguigna diminuisce ed i barocettori renali nell’apparato iuxtaglomerulare (JGA) rilasciano l’ormone Renina; a sua volta, questo attiva il sistema Renina-Angiotensina-Aldosterone (RAAS) [51]. Quando viene attivato il RAAS, viene avviato il processo di mantenimento dell’omeostasi dei fluidi, elettroliti e pressione sanguigna [51] e alla fine rilascia l’ormone Aldosterone dalle ghiandole surrenali per perfezionare ulteriormente l’omeostasi [52, 53]. I barocettori nell’aorta e nelle arterie carotidi rilevano anche una diminuzione della pressione sanguigna e segnalano il rilascio dell’Ormone Antidiuretico (ADH, noto anche come Vasopressina) dalla ghiandola pituitaria per conservare l’acqua, aumentare il volume del sangue e aumentare la pressione sanguigna [48]. Al contrario, se la pressione sanguigna aumenta a causa dell’aumento del volume sanguigno arterioso, gli atri cardiaci percepiscono un allungamento e rilasciano l’ormone Fattore Natriuretico Atriale (ANF) per aumentare l’escrezione di sodio, inibire la vasocostrizione renale, attenuare la secrezione di Renina e infine diminuire il volume sanguigno e la pressione sanguigna [54].
Collettivamente, se l’acqua e il sodio non vengono manipolati e programmati con cura, questi meccanismi fisiologici che lavorano per mantenere il corpo in omeostasi potrebbero non produrre l’effetto desiderato di ridurre selettivamente il fluido nello spazio extracellulare/sottocutaneo. Sebbene questi meccanismi siano in atto per mantenere il corpo in equilibrio, non tutti gli ormoni rilasciati hanno un effetto immediato sul corpo quando l’osmolarità plasmatica è alterata. Ad esempio, uno studio ha mostrato un effetto ritardato dell’ADH quando i ricercatori hanno esaminato gli effetti del carico d’acqua sulla perdita di peso acuta negli atleti di sport da combattimento confrontando una strategia di carico d’acqua per tre giorni in cui il gruppo sperimentale ha consumato 100ml/kg/giorno di acqua rispetto ad un gruppo di controllo che ha consumato 40ml/kg/giorno di acqua [55]. Durante il successivo giorno di disidratazione con entrambi i gruppi che consumavano 15ml/kg/giorno di acqua, i livelli di ADH nel gruppo di carico idrico sono aumentati da ~ 2,3pmol/L a ~ 3,8pmol/L alla 13a ora e ~ 5pmol/L a la 24a ora di restrizione dei liquidi, momento in cui le perdite di massa corporea hanno superato quelle del gruppo di controllo dello 0,6 % (~ 2,5 vs. 3,1 % rispetto al basale) [55]. Pertanto, nonostante l’aumento della produzione totale di liquidi da 3 giorni di carico idrico combinato con un giorno di drastica restrizione dei liquidi, i livelli di ADH stavano ancora salendo oltre le 24 ore di disidratazione [55]. In un altro studio, i ricercatori hanno ridotto l’assunzione di sodio a livelli estremamente bassi (10meq/giorno) per ~ 6 giorni in 16 uomini sani e hanno misurato i livelli di RAAS, Aldosterone plasmatico, sodio urinario e sodio sierico a 24 ore, 48 ore e ~ 6 giorni dopo l’intervento [53]. Sebbene i livelli sierici di sodio siano rimasti abbastanza coerenti tra 137,6 e 139meq/l per il periodo di ~ 6 giorni, i ricercatori hanno riferito che l’attivazione del RAAS era evidente entro 24 ore e diminuiva la produzione di sodio nelle urine da 217meq/24 ore fino a 105meq/24 ore [53]. Inoltre, ci sono volute 48 ore per osservare un forte aumento dei livelli di Aldosterone plasmatico per ridurre ulteriormente la produzione di sodio nelle urine a 59meq/24 ore e altri ~ 4 giorni affinché la produzione di sodio nelle urine si stabilizzasse a 9,9meq/24 ore [53]. Quindi, c’è un ritardo temporale nello stabilire l’omeostasi di fluidi ed elettroliti durante il quale la manipolazione di acqua e sodio può essere implementata per indurre la diuresi prima che i meccanismi omeostatici protettivi si manifestino completamente per arrestare la perdita di acqua.
Mentre i bodybuilder manipolano spesso l’acqua e/o il sodio alterandone l’assunzione [8, 11, 12, 14, 19, 20], può essere presa in considerazione anche un’altra strategia praticabile per aumentare la diuresi. La letteratura sull’atrofia da disuso e gli adattamenti cardiovascolari all’assenza di gravità durante il volo spaziale [56] rivela una strategia precedentemente descritta [36] che i culturisti possono impiegare per promuovere la diuresi durante le ~ 24 ore prima della competizione. Riposare e/o dormire con una posizione di “inclinazione a testa in giù” (HDT) (tipicamente da − 4 a -6˚ per cui l’intera superficie durante il sonno è inclinata verso il basso [57, 58] simula l’aumento del ritorno venoso cardiaco (e la perdita di pressione ortostatica) che si verifica durante la microgravità. Ciò si traduce in diuresi e risposte cardiovascolari simili a quelle osservate acutamente durante il volo spaziale [57,59], mediate in parte da un aumento del Peptide Natriuretico Atriale (rilasciato dal cuore) e da una riduzione della Renina plasmatica [60,61] Mauran et al., ad esempio, hanno dimostrato che queste risposte ormonali e la diuresi e la natriuresi associate ritornano ai valori di base entro 24 ore [62], provocando una perdita di peso corporeo di circa 1,0-1,3 kg senza variazioni della frequenza cardiaca a riposo o del sangue [58, 60, 61] Brevi periodi di HDT più grave fino a -30 % evocano aumenti graduali della pressione venosa centrale oltre quelli di -6 % HDT [63], sebbene le risposte diuretiche all’angolo HDT siano inferiori a −T6 % non sembra siano stati studiati. Brevi (≤ 2 h) periodi di HDT fino a -40˚ sembrano ben tollerati [64, 65], ma una HDT prolungata ad angoli -12 % aumenta significativamente la pressione intracranica e intraoculare [66]. Inoltre, chi soffre di reflusso gastrico dovrebbe essere consapevole che l’HDT potrebbe in teoria peggiorare la sintomatologia, dato che sollevare la testa sopra il livello del letto (l’opposto dell’HDT) è un rimedio efficace [67,68,69,70]. Questo probabilmente non è un problema per coloro che normalmente non soffrono di reflusso gastrico [71]. Pertanto, i bodybuilder potrebbero plausibilmente impiegare l’HDT durante il riposo e il sonno durante le 12-24 ore prima della competizione per incoraggiare ulteriormente la diuresi se necessario.
Un’altra considerazione quando si manipola l’assunzione di acqua e sodio è il ruolo importante che svolgono nell’assorbimento dei carboidrati. I cotrasportatori sodio-glucosio dipendenti (SGLT) sono proteine presenti nell’intestino tenue che consentono il trasporto del glucosio attraverso la membrana cellulare; una forte evidenza suggerisce che la consegna del trasporto di carboidrati è limitata dalla capacità di trasporto SGLT1 [72,73,74,75]. Poiché il carico di carboidrati sembra avere potenziali benefici per i bodybuilder di apparire “pieni”, è importante la disponibilità di sodio per il co-trasporto del glucosio attraverso le membrane cellulari. È interessante notare che lo studio di de Moraes et al. hanno riferito che il carico di carboidrati ha indotto vari sintomi gastrointestinali nei bodybuilder agonisti [15]. Sebbene l’assunzione di sodio non sia stata riportata in questo studio, alcuni dei sintomi potrebbero essere stati dovuti alla mancanza di sodio nella dieta poiché i bodybuilder hanno riferito di ridurre al minimo l’assunzione di sodio mentre si avvicinano al giorno della gara [11, 14, 20]. Inoltre, poiché ogni grammo di glicogeno attira ~ 3–4g di acqua nel muscolo [31] e questo è un processo dipendente dal potassio (vedi sopra), una mancanza di acqua e di potassio può anche ridurre l’efficacia del raggiungimento di un aspetto “pieno”.
Gestione del glucosio tramite cotrasportatore sodio-glucosio dipendenti (SGLT)1 e SGLT2. Nell’intestino tenue, il glucosio alimentare viene assorbito principalmente da SGLT1 sulla membrana del bordo a spazzola. SGLT1 ha un’elevata affinità (costante di Michaelis-Menten [Km] = 0,4 mmol/L) per il glucosio e trasporta sodio e glucosio con una stechiometria 2:1. Nel rene, il glucosio filtrato dal glomerulo renale viene riassorbito da SGLT2 e SGLT1 espressi rispettivamente nella membrana luminale dei segmenti (S)1 e S2 e nel segmento S3 dei tubuli prossimali. L’affinità di SGLT2 per il glucosio è inferiore (Km = 2 mmol/L) e il trasporto di sodio e glucosio da parte di SGLT2 avviene con una stechiometria 1:1. GLUT, trasportatore del glucosio.
Contrariamente al tipico obiettivo di ridurre l’acqua corporea (extracellulare, sottocutanea), il disturbo psicologico/stress emotivo può causare ritenzione di liquidi corporei [76] attraverso l’azione delle catecolamine (in particolare della Dopamina) [77,78,79] e degli ormoni surrenalici includendo sia il Cortisolo [80] che l’Aldosterone [81]. La ritenzione idrica durante condizioni sperimentali di stress che richiedono competizione è soggetta a variabilità interindividuale, forse dovuta in parte a differenze genetiche [82]. In casi estremi, situazioni emotivamente stressanti possono evocare polidipsia e alterare l’omeostasi dei fluidi in modo tale che aumenti fino a 9 kg (~ 20 libbre) di massa corporea possono accumularsi in appena 48 ore [78, 79]. Pertanto, c’è supporto per l’osservazione empirica comune che lo stress psicologico possa contrastare i tentativi del bodybuilder agonista di ridurre l’acqua corporea, specialmente nei casi estremi di ansia pre-gara. Gli autori raccomandano di eseguire una prova pratica della strategia della Peak Week ~ 2–4 settimane prima della competizione effettiva, in parte per ridurre l’ansia e assicurare al concorrente che la strategia della Peak Week è sia gestibile che efficace. Sebbene ciò vada oltre lo scopo di questo articolo, la gestione dello stress è riconosciuta come un aspetto importante della psicologia dello sport [83, 84] ed è molto probabile che sia importante per i concorrenti che trovano gli ultimi giorni prima della competizione così stressanti da influenzare negativamente il loro aspetto sul palco.
Sulla base di questi principi dell’equilibrio idrico-elettrolitico e delle attuali prove disponibili, sembra che la manipolazione dell’acqua e del sodio debba essere attentamente considerata, pianificata e praticata insieme alla manipolazione dei carboidrati se devono essere utilizzate. Sebbene sembrino esserci alcuni potenziali benefici nell’implementazione di queste strategie per migliorare la forma fisica il giorno della competizione, possono verificarsi effetti potenzialmente dannosi se queste variabili vengono calcolate in modo errato e/o con un cronometraggio errato che può causare ai bodybuilder di perdere il loro picco e/o incorrere in problemi di salute; quindi, lasciare queste variabili a se stesse potrebbe essere un’opzione migliore per alcuni concorrenti. Poiché è stato riferito che i bodybuilder considerano la manipolazione del sodio e dell’acqua come pratiche temporanee ma necessarie minimizzando i potenziali rischi coinvolti, è necessario prestare attenzione poiché sono state segnalate misure estreme che hanno portato a condizioni potenzialmente letali [12, 19, 20]. Le sezioni sulle applicazioni pratiche di questo articolo delineeranno ulteriormente come queste variabili possono essere manipolate in modo sicuro sulla base delle prove attualmente disponibili.
Grassi alimentari:
Oltre al glicogeno, le cellule muscolari immagazzinano anche energia sotto forma di Trigliceridi Intramuscolari (IMT). Infatti, nelle cellule muscolari viene immagazzinata quasi la stessa quantità di energia immagazzinata nell’IMT rispetto al glicogeno [85]. Tuttavia, i depositi di IMT variano considerevolmente negli esseri umani, in parte in funzione dello stato di allenamento, del tipo di fibra muscolare, della sensibilità all’Insulina, del sesso e della dieta [85]. L’IMT possono ammontare a ~ 1 % del peso muscolare [86, 87], ma poiché il grasso è meno denso del muscolo scheletrico [88], il volume di IMT in una cellula muscolare completamente “caricata di grasso” potrebbe superare il 2% del volume muscolare [89, 90]. Nei ratti (17), un singolo esercizio può ridurre il contenuto di IMT muscolare del 30% e tre giorni di una dieta ricca di grassi possono aumentare la conservazione dell’IMT di circa il 60% rispetto al basale [91]. Negli esseri umani, il reintegro alimentare di IMT può essere più lento quando anche il ripristino del glicogeno è una priorità [89, 92, 93, 94]. Tuttavia, le riserve di IMT sono aumentate dall’assunzione di grassi nella dieta [91, 95] e ridotte durante l’esercizio di contro-resistenza [96] e di endurance [85].
Sebbene il carico dei grassi sia una strategia nota nell’ambiente del Bodybuilding da molti anni [97, 98], a mia conoscenza la strategia non è stata studiata direttamente nel contesto della Peak Week del bodybuilding (p. es., in combinazione con altre strategie dietetiche come la supercompensazione del glicogeno) . Nello studio sui roditori menzionato sopra [91], tre giorni di dieta ricca di grassi seguiti da tre giorni di dieta ricca di carboidrati (CHO) hanno determinato una supercompensazione sia dei IMT che del glicogeno; tuttavia, e c’era da aspettarselo, 6 giorni di soli CHO elevati hanno prodotto l’effetto di carico di glicogeno previsto, ma non sono riusciti a elevare i livelli di IMT al di sopra del basale. Negli esseri umani, le diete ad alto contenuto di CHO/a basso contenuto di grassi possono effettivamente far precipitare le riserve di IMT [92,93,94], forse perché i IMT vengono utilizzati preferenzialmente per coprire i costi energetici della riparazione cellulare post-esercizio e dell’assemblaggio di glicogeno-glicogenina [94, 99]. Considerando che un bodybuilder di grandi dimensioni (ad es. un uomo di categoria pesi massimi) può trasportare oltre 60kg di muscoli [100, 101], aumentare le riserve di IMT da uno stato relativamente “esaurito” a uno “carico” potrebbe concepibilmente aumentare il volume muscolare di > 1 % [85 ]; ipoteticamente, questo si traduce nell’aggiunta di ≥ 0,6kg di massa magra. Quindi, il carico di grasso sembra essere una strategia promettente da utilizzare in combinazione con il carico dei CHO durante la Peak Week per i bodybuilder, e quindi merita studi futuri in un ambiente controllato.
Proteine alimentari:
Insieme all’assunzione di carboidrati e grassi durante la Peak Week, l’ottimizzazione dell’assunzione di proteine merita di essere trattata, poiché è una componente importante e indispensabile della dieta. La dose dietetica raccomandata negli Stati Uniti (RDA) per le proteine per gli adulti è di 0,8g/kg [102] ed è rimasta invariata dal ~ 1980, nonostante la continua esposizione della sua inadeguatezza. In un invito a rivalutare e rivedere la RDA, Layman [103] ha sostenuto che il fabbisogno proteico è inversamente proporzionale all’assunzione di energia. Quest’ultimo punto si applica alle persone a dieta in generale, ma ha un significato speciale per gli atleti in condizioni ipocaloriche prolungate, incarnate dai bodybuilder agonisti nel pre-gara. Alla luce di prove crescenti, un’assunzione giornaliera di 1,2-1,6g/kg è stata proposta come ottimale per la popolazione generale che mira a ottimizzare la salute e la longevità all’interno di uno stile di vita fisicamente attivo [104]. Verso l’estremità più atletica dello spettro, nella meta-analisi più completa del suo genere, Morton et al. [105] hanno scoperto che un apporto proteico di ~ 1,6g/kg (IC al 95 % superiore di 2,2 g/kg) massimizzava l’ipertrofia muscolare e la forza negli atleti di resistenza amatori non a dieta. In uno studio più rappresentativo dei bodybuilder, Bandegan et al. [106] hanno valutato la sintesi proteica dell’intero corpo tramite il metodo degli indicatori dell’ossidazione degli aminoacidi (IAAO) e hanno determinato un fabbisogno medio stimato di 1,7g/kg/giorno con un intervallo di confidenza superiore del 95% di 2,2 g/kg/giorno vicino al loro massimo muscolare raggiungibile. In un protocollo simile utilizzando il metodo IAAO, Mazzulla et al. [107] hanno stimato che il fabbisogno proteico degli uomini allenati contro-resistenza è di 2,0-2,38g/kg.
In una review sistematica di Helms et al. [108] è stato riferito che 2,3-3,1g/kg di massa magra (FFM) erano appropriati per soggetti allenati contro-resistenza in condizioni ipocaloriche. Tuttavia, dei sei studi inclusi nella review, solo due hanno coinvolto atleti competitivi altamente allenati e solo uno studio ha esaminato i bodybuilder agonisti. Quest’ultimo studio è stato condotto da Mäestu et al. [109], che hanno monitorato la composizione corporea e il profilo ormonale di bodybuilder di livello nazionale e internazionale durante le ultime 11 settimane di preparazione al contest. I concorrenti hanno dichiarato che non stavano utilizzando steroidi da un minimo di due anni prima dello studio. L’assunzione di proteine era di 2,68g/kg (2,97 g/kg FFM) al basale e 2,48 g/kg (2,66 g/kg FFM) al punto di valutazione finale (3 giorni prima della gara).
Chappell et al. [2] hanno riportato che nei bodybuilder di alto livello natural, l’assunzione di proteine di fine preparazione di uomini e donne che si sono posizionati tra i primi 5 è stata rispettivamente di 3,3g/kg e 2,8g/kg. La composizione corporea non è stata riportata in questo studio. Sulla base dei tipici intervalli percentuali di grasso corporeo alla fine della preparazione, l’aggiunta del 4-6% all’assunzione degli uomini e del 13-15% all’assunzione delle donne fornirebbe una stima dei grammi di proteine consumati per kg di FFM. Un caso studio di Kistler et al. [3] su un campione di bodybuilder natural di alto livello ha riportato un apporto proteico di 3,4g/kg (3,6 g/kg FFM). Sebbene la natura descrittiva di questi studi precluda la capacità di trarre conclusioni sul fatto che il livello di assunzione osservato fosse benefico, neutro o dannoso da un punto di vista fisico, sembrano convergere su un dosaggio proteico simile nella fase finale del periodo pre-gara.
Una possibile considerazione per il dosaggio delle proteine durante la Peak Week è se mantenere l’assunzione di proteine statica o modificarla durante le fasi di esaurimento e carico dei carboidrati. Sebbene attualmente non esistano prove concrete su ciò che è ottimale per la nostra conoscenza, lo studio di de Moraes et al. [15] che ha riportato un aumento del volume muscolare e un miglioramento dell’aspetto fisico come risultato di un protocollo di carico di carboidrati fornisce alcune prove che i bodybuilder alterano il loro apporto proteico durante la Peak Week. In questo studio, il protocollo di esaurimento/carico prevedeva tre giorni di dieta a basso contenuto di carboidrati (1,1g/kg) e ad alto contenuto proteico (3,2g/kg) seguiti da un solo giorno di dieta ad alto contenuto di carboidrati (9,0g/kg) e dieta ipoproteica (0,6g/kg). Sembra probabile che si sarebbero verificati aumenti simili del volume muscolare se le proteine fossero state mantenute statiche. Tuttavia, nonostante il ridotto apporto proteico (46,6g nel giorno durante il carico di carboidrati rispetto ai 252,4g nei giorni di scarica dei carboidrati), il disagio gastrointestinale era ancora significativamente maggiore rispetto al gruppo di controllo senza carico di carboidrati. Ciò indica la possibilità che mantenere alta l’assunzione di proteine durante il giorno di carico avrebbe ulteriormente peggiorato i sintomi gastrointestinali, potenzialmente a causa di un’eccessiva assunzione di cibo. Un’alternativa sarebbe quella di mantenere le proteine statiche, ma ridurre il carico di carboidrati (che in questo caso era ~714g), tenendo più di 1 giorno per il carico di carboidrati. Questo sembra un approccio più pratico (vedi sopra), in modo tale che un apporto totale di carboidrati ancora maggiore possa essere consumato ma con meno rischio di problemi gastrointestinali.
Una strategia potenzialmente praticabile per alterare l’assunzione di proteine durante la Peak Week è mantenere l’assunzione di proteine relativamente alta a ~ 2,5–3,5g/kg/giorno durante i primi ~ 3 giorni di esaurimento del glicogeno di una strategia di supercompensazione del glicogeno, seguita da una apporto proteico di ~ 1,6g/kg/giorno durante una dieta ricca di carboidrati per 1-3 giorni (vedi sopra), terminando almeno 24 ore prima della competizione programmata. Successivamente, potrebbe essere impiegata una strategia per indurre la diuresi e (ulteriormente) aumentare le riserve di IMT durante il giorno precedente la competizione seguendo una dieta ricca di proteine e povera di carboidrati (riccha di grassi) per un breve periodo (~12-24 h). Come discusso in precedenza, quando si caricano i carboidrati utilizzando un approccio a basso contenuto di grassi, i livelli di IMT possono diminuire, ma livelli elevati di glicogeno persistono per diversi giorni senza contrazioni impegnative che riducono il glicogeno (ad esempio, esercizio contro-resistenza o eccessiva attività fisica). Alti livelli di glicogeno intramuscolare e dell’acqua intracellulare associata impedirebbero quindi la perdita di ICW che tipicamente accompagna la diuresi. Aumentare l’assunzione di proteine consumate il giorno prima del contest, o semplicemente consumare proteine ad alti livelli tipicamente impiegati dai bodybuilder pre-gara (~ 3,0-3,5g/kg/giorno; vedi sopra) e recentemente dimostrato di essere generalmente sicuro per periodi più lunghi [ 110], incoraggerà una maggiore deaminazione ossidativa degli amminoacidi e l’ureagenesi [111] che si avvicinano ai tassi massimi osservati in individui sani [112, 113]. La clearance dell’urea ematica a sua volta richiede un gradiente osmotico durante la sua escrezione renale, causando così diuresi [114, 115]. Inoltre, il ritorno a una dieta a basso contenuto di carboidrati (ad esempio, una simile a quella utilizzata all’inizio della settimana per il carico di grassi in preparazione al carico di carboidrati) promuoverebbe anche la perdita di acqua corporea [116, 117]. Pertanto, aumentare o mantenere un’assunzione elevata di proteine mentre si riduce l’assunzione di carboidrati e contemporaneamente aumenta l’assunzione di grassi durante il giorno prima della competizione, annullerebbe i guadagni indesiderati di acqua extracellulare/sottocutanea sperimentati durante il carico di carboidrati [118]. Sarebbe inoltre complementare ad altre misure strategiche progettate per indurre la diuresi come la manipolazione dell’assunzione di acqua/sodio/potassio, l’integrazione alimentare e il posizionamento del corpo (ad es. HDT) che offrirebbe anche una seconda opportunità per il carico dei grassi durante la Peak Week. Nell’incertezza sull’efficacia della modifica di de Moraes et al. e altri protocolli può essere mitigata solo per tentativi ed errori, come verrà ulteriormente discusso nella sezione delle applicazioni pratiche, e giustificano ulteriori indagini scientifiche.
Supplementazione alimentare:
Il consumo di integratori è comune tra i bodybuilder ed è spesso manipolato durante le loro fasi preparatorie (cioè off-season e pre-gara) [2, 3, 5]. Sebbene sia ben noto che i bodybuilder utilizzino integratori come proteine in polvere, polimeri del glucosio, stimolanti pre-allenamento, sostanze adattogene/nootrope, creatina, vitamine/minerali, omega-3, termogenici, diuretici erboristici e molto altro [2, 7], c’è una scarsità di dati su come questi integratori influenzino il processo di picco dell’atleta per migliorare la propria condizione fisica. Quindi, discuteremo i potenziali benefici dell’utilizzo di integratori alimentari (cioè polveri di proteine / carboidrati, acidi grassi), creatina ed erbe durante la settimana di punta.
Integratori alimentari che forniscono un substrato energetico come proteine e carboidrati sono stati regolarmente segnalati da altri ricercatori che esaminano i bodybuilder [2, 3, 5]. Chappell et al. [2] hanno esaminato cinquantuno (35 uomini e 16 donne) bodybuilder natural e hanno scoperto che ~ 75 % degli uomini e ~ 89 % delle donne sono stati integrati con polveri proteiche. L’integrazione di carboidrati era meno popolare, con solo il ~37 % dei concorrenti uomini e nessuna concorrente donna che ne segnalava l’uso. I bodybuilder possono utilizzare questi integratori alimentari come mezzo per manipolare e consumare quantità specifiche di macronutrienti. Come accennato in precedenza nelle sezioni carboidrati e acqua/sodio, i bodybuilder cercano di massimizzare il glicogeno muscolare e il suo effetto osmotico associato come mezzo per aumentare il volume muscolare totale. Pertanto, è comune integrare con varie polveri di carboidrati (ad esempio Destrosio, Ciclo-Destrine altamente ramificata, ecc.). Le caratteristiche dei carboidrati come l’osmolalità, il tasso di clearance gastrica e l’indice glicemico sono alcune delle variabili fisiche che gli atleti dovrebbero prendere in considerazione poiché possono variare significativamente tra le fonti e possono influire sui sintomi gastrointestinali (ad es. gonfiore, crampi, diarrea, stitichezza, ecc.) [ 119,120,121]. Inoltre, è stato dimostrato che l’indice glicemico di diverse fonti di carboidrati influisce sui tassi di sintesi del glicogeno [122, 123]. Questo può essere di maggiore importanza per i bodybuilder che mirano a riempire le riserve di glicogeno in una finestra temporale breve (ad esempio dopo aver preso peso), poiché i carboidrati ad alto indice glicemico hanno dimostrato tassi di risintesi del glicogeno superiori [122]. Tuttavia, in un arco di tempo più lungo (cioè 8 + ore), le riserve di glicogeno possono essere reintegrate in modo simile, indipendentemente dalla frequenza di alimentazione [124], quando si consuma una quantità totale adeguata di carboidrati [125]. Inoltre, i dati hanno dimostrato che la combinazione di proteine e carboidrati può migliorare la risintesi del glicogeno [126]. Tuttavia, sembra prudente che gli atleti non “sperimentino” durante la Peak Week con nuovi CHO, fonti proteiche o altri integratori non integrati nelle strategie specifiche della Peak Week per ridurre il rischio di manifestare sintomi gastrointestinali negativi o altre conseguenze deleterie.
Esistono prove sostanziali a sostegno dell’uso della supplementazione di creatina per i bodybuilder. Chappell et al. hanno riferito che ~ 48 % degli uomini e ~ 51 % delle donne hanno integrato con creatina durante la preparazione del contest [2]. È stato dimostrato che la creatina migliora la composizione corporea (cioè aumenta la massa corporea magra, diminuisce la massa grassa) [127, 128] e aumenta lo stato di idratazione intracellulare [129, 130]. Ziegenfuss et al. [129] hanno dimostrato che una fase di carico di creatina di tre giorni ha aumentato il volume del fluido intracellulare di ~ 3 % senza influire sul fluido extracellulare. L’uso dell’analisi dell’impedenza bioelettrica multifrequenza (MBIA) ha indotto alcuni a interpretare inizialmente i dati con un certo scetticismo. Tuttavia, uno studio di follow-up che impiega lo stesso schema di carico di creatina di tre giorni ha osservato un aumento del 6,6% del volume muscolare della coscia tra gli atleti di potenza NCAA d’élite, come determinato dalla risonanza magnetica standard [131]. È stato anche dimostrato che l’integrazione di creatina aiuta nella sintesi del glicogeno e nella sua supercompensazione [132]. Inoltre, il consumo di CHO con creatina aumenta il carico di creatina [133], che aumenta l’idratazione cellulare come detto sopra [32, 129]. Infine, i livelli di creatina muscolare diminuiscono molto lentamente dopo il carico [134], quindi l’assunzione di creatina dopo il carico di glicogeno nella settimana di picco non è necessaria, tranne forse in piccole quantità per accelerare potenzialmente l’apporto di carboidrati dell’ultimo minuto, il giorno della competizione, nel muscolo scheletrico. Pertanto, l’integrazione di creatina può essere uno strumento potenzialmente efficace durante la Peak Week per l’espansione acuta della massa muscolare. Tuttavia, va notato che non tutti gli individui risponderanno all’assunzione di creatina esogena a fronte di un aumento significativo del contenuto di creatina muscolare [135, 136]. In particolare, i “responder” tendono ad essere quelli che hanno un’area di fibre muscolari di tipo II più ampia (cioè quelli con una propensione innata per lo sprint e/o gli sport di forza/potenza) [137, 138] e/o quelli con creatina iniziale inferiore ai livelli basali, forse a causa della mancanza di assunzione (p. es., coloro che non hanno integrato con creatina o che sono vegetariani che non integrano) [139].
L’integrazione di acidi grassi omega-3 [acido eicosapentaenoico (EPA), acido docosaesaenoico (DHA)] è stata osservata anche nei bodybuilder [2, 3]. Chappell et al. hanno riferito che il 39% degli uomini e il 47% delle donne consumavano un integratore di omega-3 (ad esempio olio di pesce, krill, olio di lino) [2]. Sebbene dati sostanziali in molti dati demografici della popolazione supportino l’uso di EPA e DHA come mezzo per ridurre l’infiammazione sistemica e migliorare la sensibilità all’insulina [140, 141], rimane sconosciuto se ciò possa migliorare il processo di picco.
Come discusso in precedenza, l’uso di diuretici è stato comunemente riportato nel bodybuilding agonistico [8, 19,20,21, 34, 35]. I bodybuilder usano spesso diuretici (sia estratti vegetali che farmaci) per aumentare la produzione di urina ed espellere il sodio nel tentativo di alterare il volume dei liquidi, migliorare la composizione corporea e presentare un fisico più “qualitativo”[142]. Inoltre, alcuni possono utilizzare i diuretici per ridurre la massa corporea totale con l’obiettivo di stabilire una specifica classe di peso [8, 19,20,21, 34, 35, 143]. Ad esempio, Caldwell et al. [143] hanno studiato gli effetti di un diuretico da prescrizione (Furosemide 1,7mg/kg) su atleti di vari sport (ad es. sollevatori di pesi e artisti marziali) e hanno riportato una significativa riduzione della massa corporea totale (-3,1 α 0,8kg) in un periodo di 24 ore. Tuttavia, a causa dei potenziali effetti collaterali e della loro capacità di mascherare l’uso di farmaci per il miglioramento delle prestazioni, i diuretici soggetti a prescrizione sono stati vietati dall’Agenzia mondiale antidoping [144]. Sebbene questi farmaci non siano presumibilmente utilizzati dai bodybuilder natural, sono stati impiegati da agonisti non sottoposti a test [19, 20]. È interessante notare che alcuni integratori a base di erbe che non sono vietati hanno dimostrato un effetto diuretico e possono essere impiegati allo stesso modo da culturisti doped e natural. Ad esempio, è stato dimostrato che il Taraxacum Officinale (Dente di Leone) aumenta significativamente la frequenza di urinazione e la produzione di escrezione in modo acuto (cioè entro una finestra di 10 ore) [145]; tuttavia, per quanto ne sappiamo, nessuna ricerca ha esaminato direttamente il suo impatto sugli spostamenti dei fluidi intracellulari rispetto a quelli extracellulari o sulla sua efficacia durante la Peak Week.
Attività del Furosemide
La vitamina C (acido ascorbico) è idrosolubile e considerata non tossica anche in quantità elevate [146]. Poiché richiede la filtrazione renale per l’escrezione, provoca anche diuresi osmotica [147]. La ricerca supporta un effetto diuretico della vitamina C sia orale che endovenosa [148], con dosi giornaliere di appena 11mg/kg che producono diuresi nei bambini [149], sebbene una dose endovenosa di 500mg non sia riuscita a indurre diuresi nei maschi adulti [150] . Uno studio sia su soggetti sani che su pazienti con carenza di vitamina C ha dimostrato che le perdite urinarie di vitamina C (e la diuresi concomitante) si verificano solo al di sopra della soglia di concentrazioni ematiche di ~ 14mg/L (che corrisponde ai livelli di saturazione dei tessuti). Questi dati suggeriscono che il raggiungimento di concentrazioni ematiche di vitamina C che promuovono la diuresi varia in funzione dei tassi di assorbimento e assorbimento/deposito nei tessuti [151] (3). Dato il suo uso comune, la relativa sicurezza e la potenziale efficacia come diuretico non farmacologico, l’uso dell’acido ascorbico in uno scenario di picco della preparazione (compresi i modelli di dosaggio per ridurre al minimo il disagio gastrointestinale e ottimizzare le concentrazioni ematiche nel contesto dei tempi dei pasti e di altri fattori che possono influenzare l’assorbimento) giustificano ulteriori ricerche. Infatti, a causa della scarsità di ricerche disponibili sull’argomento, è difficile formulare raccomandazioni definitive sull’uso e sul dosaggio durante la Peak Week. Tuttavia, sulla base delle prove disponibili, il dosaggio ripetuto (ogni poche ore) di 500-1000mg di vitamina C è una strategia praticabile da utilizzare durante le 12-24 ore prima della competizione per accelerare potenzialmente la perdita di acqua corporea con effetti collaterali minimi (ad es. disturbi gastrointestinali). Si prega di notare che è necessaria cautela poiché un consumo eccessivo di vitamina C può causare diarrea osmotica [152].
L’uso di un integratore di Caffeina è di menzione speciale per le sue proprietà diuretiche. Dosi di almeno ~ 250-300mg di Caffeina (2-3 tazze di caffè) possono essere assunte per promuovere la diuresi acuta in coloro che non sono tolleranti alla caffeina a causa dell’uso cronico [153]. D’altra parte, diversi giorni di astinenza possono ripristinare la sensibilità agli effetti diuretici della caffeina (sebbene l’effetto diuretico sia ancora presente solo a queste dosi maggiori) [154]. Anche gli effetti diuretici, migliorativi dell’umore [155] e delle prestazioni della caffeina [156] dovrebbero essere considerati nel contesto di potenziali disturbi del sonno se assunta in modo acuto per promuovere la diuresi per ridurre il peso la notte prima della competizione, così come l’effetto di astinenza se l’uso viene interrotto bruscamente [157]. Una potenziale strategia della Peak Week sarebbe quella di limitare la caffeina all’inizio della settimana (specialmente nei consumatori cronici, per ripristinare la sensibilità), impiegarla all’inizio della giornata come diuretico (p. es., il giorno prima della competizione) per limitare gli effetti negativi su qualità del sonno e continuare il suo uso in seguito (p. es., il giorno della competizione) per prevenire gli effetti di astinenza sia sull’omeostasi dei liquidi che sull’umore e sull’eccitazione [157]. È stato notato che la caffeina può essere impiegata (3-8mg/kg) come agente per accelerare il carico di glicogeno [158], sebbene i dati siano scarsi ed equivoci su questo effetto [159]. Pertanto, gli atleti che potrebbero scegliere di includere la caffeina per aumentare il carico di carboidrati a metà della Peak Week potrebbero anche perdere la sua utilità come diuretico durante i giorni successivi (ad esempio, quando tentano di “asciugarsi” ~ 24h prima di salire sul palco ).
L’uso del farmaco Metformina (500mg-1.5g/die) è utilizzato durante i giorni di ricarica dei CHO della Peak Week. Essa aumenta la sensibilità all’insulina e il miglioramento del ripartizionamento calorico. Nonostante riduca in parte l’assorbimento glucidico e possa alterare la glicogenolisi epatica, molti atleti hanno riferito di usarla con successo aggiustando la quantità di carboidrati la dove necessario e senza compromissioni della supercompensazione del glicogeno muscolare. La pratica d’uso della Metformina ha di gran lunga sostituito il malsano utilizzo dell’Insulina la quale mostra una quantità secreta più che sufficiente dall’alimentazione. L’aggiunta di un GDA (farmacologico o OCT) ne migliora l’effetto. Si sconsiglia un suo inserimento nella Peak Week se non già testata in precedenza dal momento che può causare disturbi gastrointestinali con conseguenti eventi diarroici.
Come sostituto alla Metformina, l’uso della Berberina è stato applicato da alcuni atleti durante la Peak Week nei giorni di ricarica dei carboidrati. L’attività della Berberina, similmente alla Metformina, si esplica attraverso i PPAR–γ e l’attivazione indiretta del AMPK, con conseguente aumento del uptake del glucosio (migliore insulino sensibilità) sia da parte del miocita che del adipocita (quest’ultimo, in una condizione di bassa concentrazione e deplezione epatico-muscolare di glicogeno risulta limitato per ciò che concerne la ripartizione calorica).
Meccanismo d’azione della Metformina
I dosaggi di Berberina che hanno dimostrato un impatto statisticamente significativo, anche alla luce della ricerca scientifica svolta fino ad oggi, vanno dai 500mg a 1.5g al giorno assunti in 2-3 somministrazioni di uguale portata distribuite durante la giornata (preferibilmente prima dei pasti principali). L’abbinamento con la Silimarina ha mostrato di aumentarne la biodisponibilità. L’emivita della Berberina è stata stimata essere di circa 5-6 ore.(The 5-minute Herb and Dietary Supplement Consult – a cura di Adriane Fugh-Berman (pag. 158)) Ai dosaggi comunemente usati nella medicina tradizionale e nel limite di 1.5g/die la Berberina è ben tollerata e sicura; a dosaggi più alti può determinare: disturbi gastrointestinali, dispnea, diminuzione pressoria, sintomi simil-influenzali e danno cardiaco. (https://www.ncbi.nlm.nih.gov/pubmed/10767672)
Effetto della Berberina sul metabolismo glucidico
I “NO-booster” vengono utilizzati da alcuni atleti il giorno del contest. Alcuni si limitano ad assumere 8g di Citrullina Malato 30 minuti-1h prima di salire sul palco. Altri, invece, aggiungono, al fine di potenziare l’effetto “pump”, il Sildenafil assumendolo ad un dosaggio di 25-100mg 1h prima di salire sul palco.
Alcuni “intrepidi” con poca conoscenza su farmacologia ed emivita, somministrano per via intramuscolare nei gruppi carenti la PGF1-α per aumentare il “pump” in questi gruppi. Si tratta di una pratica di nicchia e, in definitiva, poco efficace per via della breve vita attiva della molecola (pochi minuti) e la scomodità di utilizzo durante un contest.
Sono anche utilizzate le soluzioni topiche anti-infiammatorie e drenanti per aumentare il flusso dei liquidi extracellulari dalle zone critiche come le gambe. Molecole come il Glicosaminoglicanopolisolfato che viene applicato a partire dall’ultimo post workout del “Leg Day” fino alla sera prima del contest con applicazione in 3 somministrazioni. La sua efficacia è risultata apprezzabile e additiva con le altre pratiche per il controllo dei liquidi extracellulari, anche se vanno comunque valutate le risposte individuali.
I bodybuilder utilizzatori di PEDs usano in vista della Peak Week anche pratiche per la riduzione marcata degli estrogeni per via farmacologica (vedi inibitori dell’Aromatasi) nel tentativo di ridurre lo spessore della pelle. L’Estradiolo, come il GH, è implicato nella sintesi di collagene, una riduzione ulteriore di queste due variabili potrebbe ridurre nel giro di 14 giorni lo spessore cutaneo. La pratica, per essere oggettiva, deve basarsi sugli esami ematici di controllo per valutare i livelli di E2. Non esiste alcuna letteratura in merito ma semplici dati aneddotici raccolti negli anni. E’ scontato dire che gli effetti negativi degli estrogeni sullo spessore della pelle possono richiedere diversi mesi affinché vangano eliminati completamente, quindi il mantenimento di un elevato livello di estrogeni durante i primi mesi di preparazione alla gara per poi farli calare fino al livello minimo solo un paio di settimane prima dell’esibizione non è l’ideale. Per tutti coloro che usano grandi dosi di AAS aromatizzabili per tutta la maggior parte della preparazione, tenete questo bene in mente.
Fibre e FODMAP:
La fibra alimentare è materia vegetale indigeribile proveniente da fonti di carboidrati che possono essere classificate come idrosolubili o insolubili (cioè fermentabili) e svolge un ruolo vitale nella salute gastrointestinale e nella regolarità del movimento intestinale [160]. I culturisti che mirano a ridurre la massa corporea totale durante la Peak Week come mezzo per fare una particolare classe di peso possono trarre beneficio dalla riduzione intenzionale dell’assunzione di fibre. Ad esempio, Reale et al. [55] hanno studiato l’effetto delle manipolazioni dietetiche (cioè macronutrienti, fibre, sodio e assunzione di acqua) sulla perdita di peso acuta per gli atleti da combattimento e hanno prescritto 10-13g di fibre per ridurre il contenuto intestinale totale e la massa corporea. Diverse fonti di cibo influiscono sulle caratteristiche di massa fecale e quelle ricche di fibre tendono ad aumentare l’acqua nello spazio interstiziale e la massa delle feci [161]. I dati hanno dimostrato che una relazione diretta tra l’assunzione di fibre e il contenuto intestinale con periodi di restrizione acuta (di appena due giorni) è efficace nello svuotamento/pulizia del tratto gastrointestinale [162]. Pertanto, la logica per ridurre l’assunzione di fibre prima della competizione è in genere quella di ridurre al minimo il rischio di gonfiore/ritenzione idrica [11] e, per alcuni, può essere una strategia efficace per stabilire una classe di peso.
Sebbene la ricerca sull’argomento sia limitata, Chappell et al. [11] hanno riferito che i bodybuilder che hanno osservato hanno ridotto gravemente il loro apporto di fibre principalmente riducendo/omettendo le verdure fibrose durante la Peak Week. Inoltre, è ben noto che oligosaccaridi, disaccaridi, monosaccaridi e polioli fermentati (FODMAP) sono scarsamente assorbiti, attirano fluidi all’interno del tratto gastrointestinale e aumentano la probabilità di gonfiore/gas [163]. Pertanto, potrebbe essere consigliabile per i bodybuilder limitare le fonti di cibo ad alto contenuto di FODMAP durante la Peak Week. Questo potrebbe essere uno dei motivi per cui anche le fonti alimentari come latticini e fonti ricche di lattosio e glutine sono aneddoticamente limitate in questo periodo. D’altra parte, fonti di fibre come la gomma di guar [164] e lo psillio [165], che hanno dimostrato di ridurre i sintomi della sindrome dell’intestino irritabile dominata sia da stitichezza che da diarrea, potrebbero essere impiegate su base individuale per compensare il disagio gastrointestinale, come notato sopra nello studio di de Moraes et al. [15]. Nonostante la mancanza di dati all’interno di questo gruppo demografico, la fibra alimentare è probabilmente una variabile che può influire sul processo di picco di un bodybuilder e dovrebbe essere considerata su base individuale nel contesto con gli altri aspetti dell’approccio della Peak Week.
Allenamento:
Poiché i bodybuilder si allenano invariabilmente e principalmente con esercizi contro-resistenza (RE), la misura in cui questo tipo di allenamento in particolare riduce il glicogeno e i IMT merita una considerazione. In uno studio iniziale sull’uso di substrati energetici durante un RE in bodybuilder allenati, Essen-Gustavsson e Tesch [96] hanno scoperto che una sessione di RE ad alto volume nella parte inferiore del corpo riduceva sia il glicogeno che i IMT del vasto laterale di ~ 30%, e che sia i livelli a riposo che l’entità della deplezione era correlata rispettivamente agli enzimi energeticamente connessi come la esochinasi e il 3-idrossi-Co-A deidrogenasi. In un altro studio, solo tre serie di Curl delle braccia (80% 1RM o ~ 12RM) erano sufficienti per ridurre il glicogeno del bicipite brachiale del 24 % e aumentare il lattato muscolare ~ 20 volte nei bodybuilder allenati [166]. Allo stesso modo, Roberg et al. [167] hanno scoperto che 6 serie di estensioni del ginocchio (~ 13 ripetizioni/serie; intervalli di riposo di 2 minuti) hanno ridotto il glicogeno muscolare di circa il 40% negli uomini allenati contro-resistenza, ma i livelli di glicogeno hanno recuperato il 50% delle perdite durante le 2 ore di riposo a digiuno, presumibilmente a causa dell’immediata assimilazione post-esercizio dei metaboliti glicogenolitici (ad esempio, lattato) [168]. Lo stesso gruppo ha anche scoperto che un regime di corrispondenza del carico di lavoro esterno (impiegando il doppio del carico in modo tale da impostare una media di solo ~ 6 ripetizioni ad esaurimento) produceva un modello quasi identico di uso di glicogeno e recupero immediato post-esercizio. Pertanto, il RE eseguito con intervalli di ripetizioni comunemente impiegati tra i bodybuilder riduce sostanzialmente le riserve di glicogeno muscolare in un modo correlato al carico di lavoro/volume di una data seduta.
In linea con la ricerca precedente che suggeriva che l’ossidazione del grasso è maggiore nelle donne così come in quelle con livelli di grasso corporeo più elevati [85, 169], uno studio su donne obese non allenate ha rilevato che il 42% delle riserve IMT miste a riposo sono state utilizzate solo durante 6 serie di 10 ripetizioni di estensione del ginocchio [170]. Mentre i IMT erano tornati al 33% al di sotto della linea di base 2 ore dopo l’esercizio, nonostante l’assenza di assunzione di cibo, le riserve di glicogeno muscolare sono diminuite solo del 25% nel corso della seduta, ma non sono riuscite a recuperare significativamente in assenza di consumo di cibo [170]. I dati di cui sopra suggeriscono che il ripristino dei IMT può procedere lentamente in luogo delle fonti alimentari [171], mentre per i CHO è necessario per ripristinare sostanzialmente i livelli di glicogeno una deplezione acuta post allenamento contro-resistenza con riassorbimento degli intermedi glicolitici.
Pertanto, il potenziale di modificare le riserve di glicogeno intramuscolare e di IMT attraverso la dieta (vedi sopra) e l’esercizio è chiaro, ma gli effetti corrispondenti possono essere variabili tra i bodybuilder in funzione della dieta pre-gara (la composizione e il contenuto dei macronutrienti possono influenzare le riserve a riposo), attività enzimatica muscolare e sesso, tra le altre variabili non studiate. Anche il danno muscolare indotto dall’esercizio può essere importante nell’interpretazione dei dati di cui sopra poiché è altamente variabile [172,173,174], una funzione dello stato di allenamento [175] e noto per compromettere la sensibilità all’insulina muscolare [176] così come il rifornimento di glicogeno [177] . Evitare un danno muscolare eccessivo può quindi essere importante quando si considera una strategia di allenamento contro-resistenza durante la Peak Week non solo per massimizzare le riserve di glicogeno e IMT, ma anche per prevenire indolenzimento muscolare a insorgenza ritardata indesiderato che potrebbe impedire la capacità di attivare i muscoli [178] durante la posa sul palco. In effetti, le richieste energetiche di recupero da un allenamento apportante danno muscolare possono essere così grandi che in casi estremi i livelli di glicogeno possono continuare a diminuire dopo l’esercizio e non recuperare completamente in 24h nonostante un elevato consumo di CHO (10 g/kg/giorno) [179 ]. La variabilità nell’entità dell’infiammazione post-esercizio [180, 181] può anche spiegare la sopra menzionata variabilità nell’entità dell’idratazione che accompagna il carico di glicogeno. I livelli di IMT a riposo e di glicogeno sono più elevati e vengono utilizzati più facilmente in soggetti allenati che impiegano un carico di lavoro assoluto maggiore. Tuttavia, il ripristino post-esercizio di entrambe le riserve di energia è correlato alla sensibilità all’insulina e procede in modo simile rispetto ai depositi a riposo, indipendentemente dallo stato di allenamento [182]. Pertanto, l’elevata sensibilità all’insulina generalmente osservata nei bodybuilder nel pre-gara [5, 7, 90, 183, 184] conferisce un vantaggio per i depositi di IMT e il ripristino del glicogeno dopo sessioni di allenamento ad alto fabbisogno di substrato [185], ma la loro maggiore massa e capacità muscolare per ridurre le riserve di energia muscolare impongono che l’assunzione di grassi alimentari e CHO debba essere proporzionalmente grande per garantire un effetto super compensativo.
Applicazioni pratiche per la Peak Week:
È evidente che i bodybuilder implementano una varietà di strategie per la Peak Week nonostante la scarsità di ricerche specifiche sulla sicurezza e l’efficacia sui bodybuilder. Poiché ci sono molte variabili correlate da considerare durante il processo di picco che si influenzano direttamente a vicenda, non sono possibili raccomandazioni specifiche per la Peak Week. Inoltre, ci sono significative risposte interindividuali alla manipolazione di queste variabili e i bodybuilder potrebbero dover adottare approcci diversi durante la Peak Week a seconda delle loro circostanze, obiettivi e come il loro corpo risponde alle alterazioni delle variabili. Ad esempio, gli approcci alla Peak Week potrebbero differire sostanzialmente in base alle circostanze di un bodybuilder che deve raggiungere un peso per una specifica classe rispetto a un bodybuilder che non è vincolato da un limite di peso. Allo stesso modo, gli atleti che gareggiano nelle varie suddivisioni di categoria del bodybuilding potrebbero dover adottare approcci diversi (ad es. women’s physique/figure/wellness/bikini/fitness e men’s physique/classic physique) in cui gli standard di giudizio possono differire da quelli del bodybuilding tradizionale.
Mentre una discussione approfondita degli standard di giudizio sfumati e in qualche modo fluidi (che variano tra le numerose federazioni/organizzazioni di bodybuilding) delle varie categorie del culturismo competitivo esula dallo scopo di questo articolo, le seguenti considerazioni generali possono essere applicate nella costruzione di una strategia della Peak Week per queste altre categorie:
(1) Lo standard per la magrezza nelle divisioni femminili non bodybuilding spesso richiede livelli di grasso corporeo più elevati e meno muscolosità rispetto al bodybuilding femminile, e può anche quindi richiedere poche o nessuna delle manipolazioni della Peak Week descritte fino a qui ;
(2) Aneddoticamente, le concorrenti femminili (tipicamente nelle divisioni Bikini o Figure) possono ridurre intenzionalmente il grasso corporeo totale per ottenere livelli di grasso corporeo inferiori e competitivi e, invece di applicare procedure diuretiche, “carico d’acqua”, nel tentativo di ridurre la comparsa di eccessivo magrezza, mantengono l’aspetto desiderato di una distribuzione del grasso corporeo più uniformemente e omogenea;
(3) I concorrenti fitness, in cui vengono giudicate le prestazioni fisiche e l’aspetto fisico, potrebbero dover creare approcci altamente individualizzati al ripristino dell’acqua e dei depositi energetici che ottimizzino la competitività, minimizzino il rischio di lesioni e tengano conto della tempistica relativa dei set di routine e fisici nel corso di una gara.
Date le attuali prove discusse nel corso di questo articolo, offro le seguenti raccomandazioni generali per i bodybuilder al fine di aiutare i lettori a sviluppare strategie personalizzate per la Peak Week che coordinino l’assunzione di macronutrienti, le strategie di idratazione e di gestione degli elettroliti, l’integrazione e la routine di allenamento contro-resistenza/endurance(cardio). È importante sottolineare che queste raccomandazioni non dovrebbero essere considerate “regole” concrete in quanto esiste una significativa variabilità individuale di come gli atleti possono rispondere alla manipolazione di queste variabili. Infatti, a causa del numero di variabili che possono essere manipolate e degli scenari espansi che possono verificarsi, presento linee guida più specifiche di Peaking per:
(1) Un concorrente Physique femminile (60kg che non è vincolata da un limite di peso (BB1);
(2) un bodybuilder supermassimo (105 kg) che non è vincolato da un limite di peso (BB2);
(3) un concorrente Classic Physique che deve essere sotto un limite di peso (85 kg) in base alla sua classe di altezza (BB3)
In tutte le circostanze, si presumerà che i concorrenti effettuino il check-in (e il peso, se applicabile) il venerdì pomeriggio per competere il sabato mattina per il giudizio preliminare e il sabato sera per le finali. Si prega di notare che, nonostante queste circostanze specifiche, le raccomandazioni presentate nella Fig. 1 e nelle Tabelle 1, 2 e 3 dovrebbero essere viste come punti di partenza raccomandati che probabilmente richiederanno aggiustamenti basati sulle risposte dell’individuo all’alterazione delle variabili. La strategia simulata della Peak Week nella Fig. 1 è presentata solo come illustrativo esemplificativa e non devono essere considerati consigli dietetici, di esercizio fisico e/o medici prescrittivi. Si prega di fare riferimento al testo per una spiegazione razionale dettagliata per la manipolazione di macronutrienti, acqua, sodio e potassio presentata nella Fig. 1 e nelle Tabelle 1, 2 e 3. A tal fine, le strategie della Peak Week includerebbero le seguenti considerazioni:
1) Durante un protocollo di esaurimento/supercompensazione indotta da allenamenti contro-resistenza, l’attività fisica dovrebbe coinvolgere tutti i principali gruppi muscolari e impiegare una varietà di esercizi per garantire una riduzione diffusa dei livelli di IMT e di glicogeno nell’intera massa muscolare.
2) Utilizzando uno schema di ripetizione relativamente alto (>12 ripetizioni) con un approccio a volume più basso o più alto [167], ed esercitando uno sforzo e/o un carico sufficienti per impegnare la maggior parte dei tipi di fibre [186,187,188] ma fermandosi prima del cedimento applicando un rapporto tra volume/intensità ben delineato e non eccessivo, evitando nuovi esercizi, sembra un approccio prudente per garantire che il danno muscolare sia ridotto al minimo.
3) Gli esercizi che sovraccaricano il muscolo nella fase di allungamento/o eccentrico dominante (es. Stacco da terra rumeno, DB Lat Pullover, DB Fly) dovrebbero essere ridotti al minimo poiché è stato dimostrato che l’allenamento con modalità di allungamento aumenta il danno muscolare [189].
4) L’esercizio cardiovascolare dovrebbe essere ridotto o eliminato preferibilmente prima di entrare nella fase compensatoria delle riserve energetiche nei giorni precedenti la competizione.
5) L’allenamento contro-resistenza durante la Peak Week dovrebbe generalmente avvenire all’inizio della settimana, distribuito su 3-4 giorni a seconda della suddivisione di allenamento abituale dell’atleta, per consentire un tempo adeguato per la supercompensazione durante i giorni prima di salire sul palco di gara. Allenare le gambe per prime in questa serie di allenamenti della Peak Week consente il massimo tempo per il recupero in questi gruppi muscolari.
6) Il potenziale per il carico di glicogeno di compromettere la conservazione dei IMT suggerisce che la separazione dei periodi di carico di glicogeno e di grassi può essere prudente, con una dieta ad alto contenuto di CHO che precede gli sforzi per il carico dei grassi [92]. Ridurre la coingestione dei grassi con grandi quantità di carboidrati può anche evitare gli effetti negativi degli acidi grassi liberi sulla formazione di glicogeno [190], ridurre la distensione gastrica accelerando lo svuotamento gastrico, nonché migliorare il carico di glicogeno aumentando ulteriormente i livelli di glucosio nel sangue e di insulina [191,192,193] . Se consumate in giorni diversi, le diete contenenti grassi a 2g/kg/giorno [92] e CHO a 10g/kg/giorno [100] possono ripristinare e potenzialmente sovracompensare i rispettivi depositi energetici entro 24 ore. La variabilità individuale e gli obiettivi/bisogni dell’atleta possono richiedere strategie diverse, incluso consentire >24h per il carico di glicogeno [194] se le circostanze lo consentono.
7) Piuttosto che introdurre nuovi alimenti, consumare principalmente gli stessi costituenti dietetici durante la Peak Week che si consumano durante le settimane/mesi precedenti può anche essere utile per evitare disturbi gastrici. Poiché le fonti di carboidrati di frutta e fruttosio stimolano meglio il ripristino del glicogeno epatico, mentre il glucosio lo fa per il glicogeno muscolare [195], si raccomanda che la maggior parte dei carboidrati consumati provenga da fonti a base di amido/glucosio. Da notare, tuttavia, che è stato dimostrato che combinazioni di glucosio, fruttosio e saccarosio con bevande sportive aumentano la velocità di assorbimento dei liquidi dall’intestino tenue prossimale [196]. Pertanto, si consiglia agli atleti di sperimentare prima della settimana di picco su quali fonti di carboidrati funzionano meglio per loro.
8) Garantire che le proteine siano co-ingerite, anche se in quantità inferiori, con i CHO durante la ricarica può aumentare il rilascio di insulina e facilitare il carico di glicogeno [197, 198].
9) Un apporto proteico più elevato (es. 3,0g/kg) può essere combinato con un apporto più elevato di grassi durante i periodi di deplezione dei CHO per avviare il carico di grassi seguito da un carico di CHO con un apporto proteico inferiore (es. 1,6 g/kg) per compensare le riserve di glicogeno . Una volta completato il carico di carboidrati, può essere implementata una dieta ad alto contenuto proteico (3,0 g/kg)/alto contenuto di grassi/basso contenuto di CHO. Ancora una volta, la variabilità individuale e gli obiettivi/bisogni dell’atleta possono richiedere strategie diverse per raggiungere il massimo della forma fisica.
10) Varie strategie di carico dei CHO sono state riportate nel bodybuilding. Ad esempio, Roberts et al. [199] discussero la pratica della ricarica dei CHO a caricamento frontale (l’assunzione è maggiore all’inizio della settimana e poi ridotta per mantenere la pienezza muscolare fino alla competizione) e il carico dei CHO a caricamento posteriore (l’assunzione avviene più tardi nella settimana ma può comportare meno tempo per apportare modifiche al fisico). In alternativa, potrebbe essere utilizzato anche un modello in cui i CHO vengono esaurite all’inizio della settimana (7 − 4 giorni out), caricati a metà settimana (3 − 2 giorni out), e quindi regolati/mantenuti (1 giorno out). Nello studio di de Moraes et al. [15], è stato utilizzato un metodo di carica a ritroso, ma sono necessarie ulteriori prove prima di formulare raccomandazioni più concrete. Sulla base delle prove attuali, si raccomanda il terzo modello discusso, come presentato nella Tabella 1 per la concorrente Physique femminile di 60kg e il bodybuilder di sesso maschile di 105kg, per ottenere i benefici del carico frontale e del carico posteriore; tuttavia, devono essere prese in considerazione le risposte/preferenze individuali al carico dei CHO e le esigenze dell’individuo (ad es., fare una classe di peso può richiedere il carico a ritroso).
11) La precedente dieta pre-gara può influenzare la tolleranza del concorrente alla manipolazione dietetica, nonché l’entità della restrizione dietetica di grassi e CHO durante i giorni di allenamento della Peak Week necessari per accelerare un successivo effetto super-compensativo. Ad esempio, quei concorrenti che seguono una dieta ricca di CHO/basso contenuto di grassi, ma molto ipocalorica (lasciando i livelli di glicogeno cronicamente bassi) potrebbero evitare di eliminare completamente i CHO durante l’allenamento della Peak Week. Tuttavia, coloro che hanno utilizzato un approccio a basso contenuto di carboidrati potrebbero continuare a utilizzare una dieta di questo tipo durante la Peak Week, ma potrebbero diffidare di applicare un allenamento eccessivo (invece di un approccio tapering) se i livelli di glicogeno sono già probabilmente diminuiti all’inizio della Peak Week.
12) In generale, ridurre i CHO e aumentare l’assunzione di grassi (in base alla tollerabilità individuale) durante i giorni di allenamento (“esaurimento”) della Peak Week può facilitare il carico di glicogeno durante i giorni successivi all’allenamento e, contemporaneamente, garantire che i livelli di IMT non vengano abbassati eccessivamente. Dopo 1-2 giorni di carico di glicogeno a metà/fine settimana come raccomandato nel prima citato approccio al carico dei CHO, i livelli di IMT potrebbero essere aumentati il giorno prima della competizione con un approccio ad alto contenuto di grassi/basso di CHO che servirebbe anche a ridurre l’acqua corporea in eccesso [117]. Ancora una volta, la variabilità individuale e gli obiettivi/bisogni dell’atleta possono richiedere strategie diverse con queste linee guida generali come base.
13) La pratica del carico d’acqua seguita dalla restrizione idrica è stata documentata come una strategia di perdita di peso sicura ed efficace per perdere TBW negli atleti da combattimento [55]. Sebbene il rapporto tra ECW e ICW perso non sia stato riportato in questo studio, Costill et al. [40] (come affermato in precedenza) hanno riportato che il rapporto tra ECW e perdita di ICW rimane vicino a 1:1 quando i livelli di glicogeno si stabilizzano nel tempo e vengono raggiunti livelli più elevati di disidratazione. Pertanto, sembra che la ritenzione del glicogeno muscolare, evitando l’esercizio che fa molto affidamento sul glicogeno, possa essere importante se i metodi di perdita di acqua devono essere effettuati per una perdita favorevole di ECW rispetto a ICW (ECW > ICW) in modo tale che la dimensione muscolare venga mantenuta mentre L’ECW interstiziale viene preferibilmente perso, migliorando potenzialmente l’aspetto della “definizione” muscolare.
14) Molte variabili possono alterare l’approccio utilizzato per il carico/esaurimento idrico (cioè quanta acqua l’atleta è abituato a bere regolarmente), ma i partecipanti allo studio di Reale et al. hanno perso con successo la TBW bevendo una grande quantità di acqua (100ml/kg) per tre giorni, seguita da una significativa riduzione dell’acqua a 15ml/kg il quarto giorno [55] senza effetti deleteri. In alternativa, l’assunzione di acqua può essere mantenuta relativamente costante (ad eccezione di qualche ora prima di gareggiare per prevenire qualsiasi distensione addominale) per ridurre al minimo le variabili manipolate; in effetti, questo potrebbe essere l’approccio migliore se non vengono eseguite prove pratiche prima della competizione. Mantenere una ratio Sodio:Acqua pari a 1:0.4 (per semplificare, 1L d’acqua = 1g di Cloruro di Sodio (Na)= 400mg di Sodio) fino al giorno precedente alla gara aggiungendo nelle ultime 24h il Glicerolo (generalmente 3 dosi da 10-20ml) il quale ha un noto effetto osmotico intracellulare.
15) Poiché il glicogeno muscolare crea un effetto osmotico, attirando l’acqua nella cellula mentre il glicogeno viene immagazzinato [26], il carico di CHO dovrebbe essere effettuato insieme all’assunzione di acqua [199] in modo che l’ICW muscolare possa essere massimizzato mentre l’assunzione di CHO è alta. Dopo circa tre giorni di carico d’acqua con un apporto di CHO più alto (se si utilizza il metodo di carico d’acqua), l’assunzione di acqua può diminuire a ~ 15ml/kg per 24h, il che aiuterà a indurre la diuresi entro ~ 24h prima della competizione. Si noti che questa raccomandazione si basa su quanto studiato e riportato; tuttavia, gli autori riconoscono che maggiori assunzioni di acqua possono essere preferenziali, come 30-40ml/kg, ma non sono state studiate e quindi richiedono ulteriori ricerche.
16) Aumentare o mantenere un’assunzione elevata di proteine mentre si riduce il consumo di carboidrati e contemporaneamente aumentare l’assunzione di grassi durante il giorno prima della competizione può invertire i guadagni indesiderati di acqua extracellulare/sottocutanea sperimentati durante il carico dei carboidrati [118].
17) È stato riportato che l’assunzione di sodio è significativamente ridotta dai bodybuilder durante la Peak Week [11, 14, 20], ma i tempi di questa pratica dovrebbero essere attentamente implementati e l’assunzione di sodio non dovrebbe essere ridotta contemporaneamente al carico dei CHO poiché l’evidenza suggerisce che la somministrazione di CHO è limitata dalla capacità di trasporto di SGLT1 [72,73,74,75]. Una volta che l’assunzione di CHO è diminuita dopo il carico di glicogeno, l’assunzione di sodio può essere temporaneamente ridotta poiché la ricerca indica che l’attivazione del RAAS è evidente entro 24 ore e sono necessarie ~ 48 ore per osservare un forte aumento dei livelli di Aldosterone plasmatico [53]. Questo ritardo temporale nello stabilire l’omeostasi di fluidi ed elettroliti, se programmato correttamente, può essere implementato per indurre la diuresi prima che i meccanismi omeostatici protettivi si manifestino completamente per arrestare la perdita di acqua. A seconda delle esigenze del bodybuilder prima della competizione (ad esempio, necessità di stabilire una classe di peso), nella Tabella 2 sono presentati vari scenari di assunzione di sodio. In alternativa, il sodio può essere mantenuto come costante per ridurre al minimo le variabili manipolate; in effetti, questo potrebbe essere l’approccio migliore se non vengono eseguite prove pratiche prima della competizione. Il mantenimento di una corretta ratio Sodio:Acqua, accennata in precedenza, garantisce una migliore escrezione di liquidi e impedisce la comparsa di stati di iponatriemia. L’assunzione di Potassio (2g netti al giorno) garantisce all’atleta una migliore omeostasi qualitativa dei liquidi corporei.
18) I bodybuilder utilizzatori di PEDs che presentano nel proprio protocollo il GH, dovrebbero cessarne l’uso almeno 14-7 giorni prima della competizione dal momento che il peptide ha una attività di alterazione del RAAS.
18) L’immagazzinamento e la ritenzione del glicogeno muscolare dipendono fortemente dalla disponibilità di potassio (un catione intracellulare primario) [41,42,43,44,45,46]. Pertanto, garantire un’adeguata assunzione di potassio durante le procedure sia di carico dei carboidrati che di riduzione dell’acqua (se aggiunta) è probabilmente fondamentale per ottimizzare l’aspetto della pratica attraverso lo stoccaggio e la ritenzione di glicogeno muscolare e quindi stimolare una perdita più favorevole di ECW rispetto a ICW quando si utilizzano strategie di disidratazione.
19) Ridurre l’assunzione di fibre durante la Peak Week sembra offrire alcuni potenziali benefici. Rale et al. [55] hanno riferito che la riduzione dell’assunzione di fibre a 10-13 g/giorno per ~ 5 giorni ha ridotto con successo il contenuto intestinale totale e la massa corporea nei contact fighters. I dati hanno dimostrato che una relazione diretta tra l’assunzione di fibre e il contenuto intestinale con periodi di restrizione acuta (di appena due giorni) è efficace nello svuotamento/pulizia del tratto gastrointestinale [162]. Pertanto, la logica per ridurre l’assunzione di fibre prima della competizione è tipicamente quella di ridurre al minimo il rischio di gonfiore/ritenzione idrica [11] e, per alcuni, parte del loro processo per creare una classe di peso.
20) L’utilizzo di alcuni integratori durante la Peak Week può rivelarsi vantaggioso per gli atleti. È stato dimostrato che l’integrazione di creatina aiuta nella sintesi del glicogeno e nella sua supercompensazione [132]. Inoltre, il consumo di CHO con creatina aumenta il carico di creatina [133], che aumenta l’idratazione intracellulare [32, 129]. Insieme alla creatina, possono essere prese in considerazione anche polveri di carboidrati (ad es. destrosio, ciclo-destrine altamente ramificate, ecc.). Le caratteristiche dei carboidrati come l’osmolalità, il tasso di clearance gastrica e l’indice glicemico sono alcune delle variabili che i bodybuilder dovrebbero prendere in considerazione poiché questi fattori possono variare significativamente tra le fonti e possono influire sui sintomi gastrointestinali (ad es. gonfiore, crampi, diarrea, costipazione, ecc.) [ 119,120,121]. Sia le polveri di proteine del siero di latte idrolizzate, gli EAA, le classiche proteine che le polveri di carboidrati possono essere utilizzate come mezzi per manipolare e consumare quantità specifiche di macronutrienti senza dover consumare grandi volumi di cibo. Anche l’uso di diuretici erboristici e farmacologici, con le dovute precauzioni, possono essere inseriti negli ultimi giorni prima del contest per facilitare l’eliminazione dell’acqua extracellulare. Come detto in precedenza, l’uso di Metformina o Berberina nei giorni di ricarica dei CHO ha il potenziale di migliorare la “qualità” di questa pratica per via di un migliore ripartizionamento calorico. L’uso di “NO-booster” come Citrullina Malato e/o Sildenafil possono dare un affetto “pump” dando un aspetto muscolare più “pieno”. Soluzioni topiche con attività anti-infiammatoria/drenante, come quelle contenenti Glicosaminoglicanopolisolfato, possono agevolare la perdita di liquidi extracellulari in aree critiche come le gambe (applicazione dal post workout dell’ultimo Leg Day fino alla sera precedente il contest). Sempre per i bodybuilder utilizzatori di PEDs, la riduzione estrema dei livelli estrogenici nelle ultime due settimane pre-contest può facilitare l'”assottiogliamento” della cute per via del legame tra sintesi di Collagene e Estrogeni.
21) Situazioni emotivamente stressanti possono evocare polidipsia e alterare l’omeostasi dei fluidi in appena 48 ore [78, 79]. Quindi, lo stress psicologico può contrastare i tentativi del bodybuilder agonista di ridurre l’acqua corporea, specialmente nei casi estremi di ansia pre-gara. Come notato in precedenza, si raccomanda di eseguire una prova pratica della strategia della Peak Week ~ 2–4 settimane prima della competizione effettiva, in parte per ridurre l’ansia e assicurare al concorrente che la strategia della Peak Week è sia gestibile che efficace.
22) Riposare e/o dormire con una posizione di “inclinazione a testa in giù” (HDT) (tipicamente da − 4 a -6˚ per cui l’intera superficie del corpo durante il sonno è inclinata verso il basso [57, 58] simula l’aumento del ritorno venoso cardiaco (e la perdita di pressione ortostatica) che si verifica durante la microgravità e si traduce in diuresi e risposte cardiovascolari [57, 59]. Pertanto, i concorrenti potrebbero plausibilmente impiegare l’HDT quando riposano e dormono durante le 12-24 ore prima della competizione per stimolare ulteriormente la diuresi. Questo potenziale beneficio dovrebbe essere bilanciato con possibili effetti dannosi della pratica sui modelli di sonno, che potrebbero interferire con le prestazioni in gara.
23) Il peso della bilancia può essere utilizzato durante la Peak Week per valutare e confermare i livelli di idratazione (vedere la seguente sezione “Considerazioni pratiche per il giorno della gara”).
24) Dal momento che sono coinvolte una moltitudine di variabili e una sostanziale interindividualità biologica, una pratica o “mock” Peak Week durante le ~ 2-4 settimane prima della competizione può fornire informazioni preziose sull’entità e sui tempi appropriati delle alterazioni della dieta e dell’allenamento durante la Peak Week. Inoltre, può attenuare i livelli di stress che un bodybuilder può avere prima della competizione, il che può facilitare il modo in cui il corpo risponde al protocollo della Peak Week.
25) Gli atleti che possono partecipare a una serie di competizioni in successione relativamente rapida, in genere su base settimanale, dovrebbero costruire strategie per le Peak Week (come negli esempi qui riportati) che possono essere replicate, con aggiustamenti aggiuntivi se necessario, durante il periodo di tempo tra le gare . Ciò può richiedere ai concorrenti di mantenere uno stretto controllo dietetico e di stabilire rapidamente l’omeostasi dei fluidi post-gara in modo da ripristinare le condizioni iniziali di base (ad esempio, i livelli di glicogeno muscolare) su cui può fare affidamento una data strategia della Peak Week. Inoltre, oltre ai rischi medici rilevati citati in precedenza, l’uso non oculato di diuretici farmacologici durante la Peak Week può probabilmente interrompere l’omeostasi dei liquidi e diminuire l’affidabilità e quindi il successo delle strategie diuretiche impiegate in una serie di competizioni in stretta prossimità temporale.
26) Come detto in precedenza, i bodybuilder utilizzatori di PEDs utilizzano in vista della Peak Week anche pratiche per la riduzione marcata degli estrogeni per via farmacologica (vedi inibitori dell’Aromatasi) nel tentativo di ridurre lo spessore della pelle. L’Estradiolo, come il GH, è implicato nella sintesi di collagene, una riduzione di queste due variabili potrebbe ridurre nel giro di 14 giorni lo spessore cutaneo. La pratica, per essere oggettiva, deve basarsi sugli esami ematici di controllo per valutare i livelli di E2. Non esiste alcuna letteratura in merito ma semplici dati aneddotici raccolti negli anni.
È essenziale capire che nessuna delle suddette strategie della Peak Week fornirà un restyling fisico per compensare la mancanza di una preparazione corretta o della aderenza durante le fasi di preparazione al contest off-season o pre-gara. Il grasso corporeo deve essere ridotto al minimo ~ 2–3 settimane prima della competizione, in modo che il concorrente possa concentrarsi sulla riduzione al minimo dell’acqua sottocutanea per mostrare al meglio la muscolatura e sull’ottimizzazione delle dimensioni muscolari aumentando le riserve intramuscolari di glicogeno e trigliceridi. Pertanto, l’utilizzo di strategie per la Peak Week è semplicemente un mezzo per ottenere un aspetto migliore durante il giorno della competizione sul palco “sintonizzando” il corpo rispetto al semplice mantenimento della dieta e delle strategie di allenamento pre-gara (cioè quelle focalizzate principalmente sulla riduzione del grasso corporeo e sul mantenimento della massa muscolare).
Considerazioni pratiche per il giorno della gara:
Idealmente il fisico presentato sul palco rappresenta il miglior aspetto possibile dell’atleta, superando quello delle settimane e dei mesi precedenti. Garantire che il picco si verifichi il giorno della competizione spesso richiede un approccio su misura con almeno le seguenti considerazioni:
Programma della giornata di gara: quando viene giudicato l’atleta e quante volte? Molte organizzazioni competitive includono più turni di valutazione [200,201,202] e categorie tali che la competizione può svolgersi nel corso di un’intera giornata (o più).
Strategie (pre-programmate o meno) per mettere a punto l’aspetto del fisico il giorno della competizione manipolando l’assunzione di acqua, cibo e integratori alimentari secondo necessità.
Aspetto personale e percezione del fisico (quanto detto sopra), ed altri mezzi per valutare la fase preparazione. Naturalmente, gli obiettivi della Peak Week settimana di punta per ridurre al minimo l’acqua sottocutanea e garantire che i IMTG e le riserve di glicogeno del muscolo scheletrico siano massimizzate, mettendo i muscoli in pieno rilievo e mostrando la massima “muscolarità” dovrebbero essere ampiamente raggiunti prima del risveglio il giorno della competizione. Nel gergo del bodybuilding, queste componenti della muscolatura potrebbero essere considerate “secchezza” (mancanza di fluido sottocutaneo) e “pienezza” (le riserve energetiche delle cellule muscolari sono completamente riempite / supercompensate). Tuttavia, spesso è necessaria una messa a punto per ottimizzare l’aspetto del fisico quando viene giudicato.
A mia conoscenza, non vi è ricerca che esamini la misura in cui i mezzi soggettivi o altri mezzi pratici al fine di garantire la buona preparazione al giorno della competizione di bodybuilding siano associati al presunto fluido sottocutaneo e alle misure istologiche. Tuttavia, i seguenti sono modi comunemente accettati e suggeriti in precedenza [36] per valutare la prestanza per il giorno del contest:
Le riserve di glicogeno muscolare sono “piene” e l’atleta può ottenere un buon “pump”? I metaboliti glicolitici (ad esempio, lattato e fosfato inorganico) derivati dall’uso del glicogeno producono una risposta di iperemia reattiva post-esercizio nota come “pump” [203] che gonfia il tessuto muscolare, aumentandone lo spessore fino a ~ 10% [204, 205]. Ciò rappresenta un vantaggio per l’aumento acuto della dimensione muscolare prima di salire sul palco e spostare il fluido in specifici ventri muscolari (idealmente anche riducendo così il volume del fluido sottocutaneo interstiziale per migliorare ulteriormente l’aspetto della muscolosità, tale che un atleta può preferibilmente “pompare” la muscolatura per migliorare l’equilibrio dell’espansione muscolare.
L’atleta è “secco”? L’acqua corporea è stata ridotta abbastanza da ridurre al minimo il fluido sottocutaneo per evidenziare notevolmente la muscolatura sottostante?
L’atleta è “piatto”? Creare una situazione di pienezza muscolare e con un aspetto fisico “asciutto” richiede uno stretto atto di riequilibrio fisiologico. La “pompa” iperemica richiede un fluido corporeo adeguato per spostarsi nel ventre muscolare; tuttavia, un atleta con alti livelli di glicogeno muscolare ma acqua corporea eccessivamente ridotta può sperimentare la “piattezza muscolare”, cioè la mancanza di un “pump” muscolare solitamente associata a un aspetto sciupato a causa dell’eccessiva disidratazione. D’altra parte, la colpa potrebbe essere anche della mancanza di glicogeno muscolare come fonte di osmoliti metabolici per l’effetto “pump” [203].
Sia le riserve di glicogeno (“pienezza”) che la disidratazione (aspetto “asciutto”) dipendono dall’omeostasi dei fluidi che cambia rapidamente. Pertanto, propongo che il peso della bilancia possa essere impiegato come indicatore rudimentale, ma pratico e oggettivo dell’idratazione corporea nel contesto del “pump” muscolare e dell’aspetto visivo, nonché delle perdite di liquidi urinari [notare che il colore delle urine è una misura di campo adeguata dello stato di idratazione, ma può essere alterata dal consumo di integratori alimentari [206, 207]. Pertanto, misurare il peso corporeo durante la Peak Week e il suo tasso di variazione può aiutare a determinare la misura in cui l’acqua corporea è stata ridotta al minimo il giorno della competizione. Le misurazioni per un ipotetico concorrente sono fornite nella Tabella 4. Presumiamo qui che il glicogeno del muscolo-scheletrico sia stato adeguatamente super compensato (aumentando il contenuto di acqua intramiocellulare e aumentando il peso corporeo) dopo un periodo di ridotta assunzione di carboidrati che riduce il contenuto di acqua corporea (e peso corporeo) all’inizio della Peak Week (vedi sopra). Se le strategie di disidratazione determinano una riduzione del peso della bilancia che si avvicina o è inferiore ai livelli di pre ricarica dei carboidrati, ipotizziamo che ciò rifletta che i cambiamenti desiderati nell’ECF (riduzione del fluido sottocutaneo) e negli spazi ICF (aumento del fluido intramiocellulare e del glicogeno) siano stati raggiunti.
La figura 2 di seguito delinea un albero decisionale del giorno della competizione che un concorrente potrebbe utilizzare per affrontare le possibilità discusse sopra (mancanza di pienezza muscolare o condizione fisica “asciutta”, o essere “piatto”). Presumiamo una preferenza per ridurre al minimo l’acqua corporea rispetto alla pienezza muscolare. Inoltre, tenete presente che lo scenario in cui la “piattezza” è un problema potrebbe richiedere una combinazione di aggiunta di acqua, sodio, carboidrati e/o grassi alimentari a seconda delle circostanze. Le precedenti esperienze di “finta Peak Week” e di assunzione di carboidrati possono servire bene all’atleta nella scelta di un giorno strategicamente appropriato per dare il massimo sul palco. Questo stesso albero decisionale può essere applicato ripetutamente in situazioni in cui l’atleta viene giudicato in più round.
Fig. 2: Albero decisionale per le regolazioni dietetiche e l’assunzione di liquidi del giorno della competizione di Bodybuilding
Conclusioni:
Ricapitolando, l’evidenza suggerisce che i bodybuilder usano frequentemente strategie di “Peak Week” come il carico dei CHO, la manipolazione di acqua/sodio e altri approcci nel tentativo di migliorare il loro fisico durante l’ultima settimana di preparazione alla competizione. Sfortunatamente, c’è una scarsità di ricerche sull’efficacia e la sicurezza di queste strategie quando implementate individualmente o collettivamente. Poiché le variabili che vengono frequentemente manipolate dai bodybuilder sono correlate, l’alterazione di una variabile in genere influenza altre variabili. Inoltre, le risposte interindividuali all’alterazione di queste variabili rendono ancora più difficile fornire precise “regole” da seguire per la Peak Week. Data la complicata interazione di variabili fisiologiche durante la Peak Week, nonché l’interindividualità biologica e la variabilità nell’importanza attribuita alla massimizzazione dei vari aspetti della muscolarità attraverso le diverse categorie competitive, ci sono una moltitudine di vie di ricerca per studiare le strategie della Peak Week. In particolare, un esame strettamente controllato degli effetti quantificabili della supercompensazione del glicogeno, della disidratazione graduale tramite la manipolazione del sodio e/o dell’acqua e delle strategie di “pump” pre-palco, insieme alla documentazione dei cambiamenti visivi “pratici” soggettivi associati nell’aspetto fisico, potrebbero essere aree di studio rilevanti che possono aiutare a informare meglio i concorrenti e indirizzarli lontano da pratiche potenzialmente pericolose e/o meno efficaci della fase Peak Week . Pertanto, grazie alla review di Brad J. Schoenfeld e colleghi, e alla piccola aggiunta delle mie ricerche e annotazioni, viene presentato questo articolo al fine di rappresentare un approccio basato sull’evidenza alle strategie di picco pre-gara basate sullo stato attuale della letteratura scientifica (e sull’aneddotica più affidabile) nella speranza che possa stimolare il miglioramento dell’atleta e/o del preparatore i quali possono accrescere le loro conoscenze e maturare una corretta comprensione delle delicate componenti che caratterizzano la preparazione ad un contest di Bodybuilding potendo così sviluppare approcci pratici e sicuri applicabili per ottimizzare la forma fisica da mostrare sul palco.
Gabriel Bellizzi
Riferimenti:
Bamman MM, Hunter GR, Newton LE, Roney RK, Khaled MA. Changes in body composition, diet, and strength of bodybuilders during the 12 weeks prior to competition. J Sports Med Phys Fitness. 1993;33:383–91. https://www.ncbi.nlm.nih.gov/pubmed/8035587.
5.Rossow LM, Fukuda DH, Fahs CA, Loenneke JP, Stout JR. Natural bodybuilding competition preparation and recovery: a 12-month case study. Int J Sports Physiol Perform. 2013;8:582–92. https://www.ncbi.nlm.nih.gov/pubmed/23412685.
6.Tinsley GM, Trexler ET, Smith-Ryan AE, Paoli A, Graybeal AJ, Campbell BI, et al. Changes in Body Composition and Neuromuscular Performance Through Preparation, 2 Competitions, and a Recovery Period in an Experienced Female Physique Athlete. J Strength Cond Res. 2019;33:1823–39. doi:https://doi.org/10.1519/JSC.0000000000002758.ArticlePubMedGoogle Scholar
8.Kleiner SM, Bazzarre TL, Litchford MD. Metabolic profiles, diet, and health practices of championship male and female bodybuilders. J Am Diet Assoc. 1990;90:962–7. https://www.ncbi.nlm.nih.gov/pubmed/2365938.
9.Gentil P, de Lira CAB, Paoli A, dos Santos JAB, da Silva RDT, Junior JRP, et al. Nutrition, pharmacological and training strategies adopted by six bodybuilders: case report and critical review. Eur J Transl Myol. 2017;27. https://www.ncbi.nlm.nih.gov/pmc/articles/pmc5391526/.
14.Mitchell L, Hackett D, Gifford J, Estermann F, O’Connor H Do Bodybuilders Use Evidence-Based Nutrition Strategies to Manipulate Physique? Sports (Basel). 2017;5. doi:https://doi.org/10.3390/sports5040076.
15.de Moraes WMAM, de Almeida FN, Dos Santos LEA, Cavalcante KDG, Santos HO, Navalta JW, et al. Carbohydrate Loading Practice in Bodybuilders: Effects on Muscle Thickness, Photo Silhouette Scores, Mood States and Gastrointestinal Symptoms. J Sports Sci Med. 2019;18:772–9. https://www.ncbi.nlm.nih.gov/pubmed/31827362.
29.McBride JJ, Guest MM, Scott EL, Others. The storage of the major liver components; emphasizing the relationship of glycogen to water in the liver and the hydration of glycogen. J Biol Chem. 1941;139:943–52. https://www.cabdirect.org/cabdirect/abstract/19411401274.
31.Shiose K, Yamada Y, Motonaga K, Sagayama H, Higaki Y, Tanaka H, et al. Segmental extracellular and intracellular water distribution and muscle glycogen after 72-h carbohydrate loading using spectroscopic techniques. J Appl Physiol. 2016;121:205–11. doi:https://doi.org/10.1152/japplphysiol.00126.2016.CASArticlePubMedGoogle Scholar
33.Schoenfeld BJ, Alto A, Grgic J, Tinsley GM, Trexler ET. Alterations in body composition, resting metabolic rate, muscular strength, and eating behavior in response to natural bodybuilding competition preparation: A case study. Journal of Strength Conditioning Research. 2020. doi:https://doi.org/10.1519/JSC.0000000000003816. ;Publish Ahead of Print.ArticlePubMedGoogle Scholar
34.Albertson TE, Chenoweth JA, Colby DK, Sutter ME. The Changing Drug Culture: Use and Misuse of Appearance- and Performance-Enhancing Drugs. FP Essent. 2016;441:30–43. https://www.ncbi.nlm.nih.gov/pubmed/26881771.
35.Hickson JF Jr, Johnson TE, Lee W, Sidor RJ. Nutrition and the precontest preparations of a male bodybuilder. J Am Diet Assoc. 1990;90:264–7. https://www.ncbi.nlm.nih.gov/pubmed/2303663.
38.Sawka MN, Coyle EF. Influence of body water and blood volume on thermoregulation and exercise performance in the heat. Exerc Sport Sci Rev. 1999;27:167–218. https://www.ncbi.nlm.nih.gov/pubmed/10791017.
41.Gardner LI, Talbot NB, Cook CD, Berman H, Uribe RC. The effect of potassium deficiency on carbohydrate metabolism. J Lab Clin Med. 1950;35:592–602. https://www.ncbi.nlm.nih.gov/pubmed/15412290.
42.Clarke DW. The influence of potassium ion upon glucose uptake and glycogen synthesis in the isolated rat diaphragm. Can J Biochem Physiol. 1955;33:687–94. https://www.ncbi.nlm.nih.gov/pubmed/13240543.
43.Kreitzman SN, Coxon AY, Szaz KF. Glycogen storage: illusions of easy weight loss, excessive weight regain, and distortions in estimates of body composition. Am J Clin Nutr. 1992;56 1 Suppl:292S – 293S. doi:https://doi.org/10.1093/ajcn/56.1.292S.
45.King RFGJ, Cooke C, Carroll S, O’Hara J. Estimating changes in hydration status from changes in body mass: considerations regarding metabolic water and glycogen storage. J Sports Sci. 2008;26:1361–3. doi:https://doi.org/10.1080/02640410802192768.ArticlePubMedGoogle Scholar
49.Kanbay M, Aslan G, Afsar B, Dagel T, Siriopol D, Kuwabara M, et al. Acute effects of salt on blood pressure are mediated by serum osmolality. J Clin Hypertens. 2018;20:1447–54. doi:https://doi.org/10.1111/jch.13374.CASArticleGoogle Scholar
59.Convertino VA, Bisson R, Bates R, Goldwater D, Sandler H. Effects of antiorthostatic bedrest on the cardiorespiratory responses to exercise. Aviat Space Environ Med. 1981;52:251–5. https://www.ncbi.nlm.nih.gov/pubmed/7283897.
60.Mauran P, Sediame S, Pavy-Le Traon A, Maillet A, Carayon A, Barthelemy C, et al. Renal and hormonal responses to isotonic saline infusion after 3 days’ head-down tilt vs. supine and seated positions. Acta Physiol Scand. 2003;177:167–76. https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1365-201X.2003.01059.x?casa_token=aXoRJ9ceXV8AAAAA:2WA6uD_yoUfSi8ayHb30rOTAOEghdhIGhwFXqAsbNenvQL8eUUuPgEOH_xExJwt4cJmrwqzoujhcbw.
64.Reuter DA, Felbinger TW, Schmidt C, Moerstedt K, Kilger E, Lamm P, et al. Trendelenburg positioning after cardiac surgery: effects on intrathoracic blood volume index and cardiac performance. Eur J Anaesthesiol. 2003;20:17–20. doi:https://doi.org/10.1017/s0265021503000036.CASArticlePubMedGoogle Scholar
65.Kalmar AF, Foubert L, Hendrickx JFA, Mottrie A, Absalom A, Mortier EP, et al. Influence of steep Trendelenburg position and CO 2 pneumoperitoneum on cardiovascular, cerebrovascular, and respiratory homeostasis during robotic prostatectomy. Br J Anaesth. 2010;104:433–9. doi:https://doi.org/10.1093/bja/aeq018.CASArticlePubMedGoogle Scholar
66.Marshall-Goebel K, Mulder E, Bershad E, Laing C, Eklund A, Malm J, et al. Intracranial and Intraocular Pressure During Various Degrees of Head-Down Tilt. Aerosp Med Hum Perform. 2017;88:10–6. doi:https://doi.org/10.3357/AMHP.4653.2017.ArticlePubMedGoogle Scholar
70.Cohn JR. Elevation of the Head of Bed to Treat Supraesophageal Reflux: Controlling the Trigger and Reducing the “Drip.” J Allergy Clin Immunol Pract. 2015;3:362–4. https://www.jaci-inpractice.org/article/S2213-2198(15)00129-4/abstract.
74.Jeukendrup AE, McLaughlin J. Carbohydrate ingestion during exercise: effects on performance, training adaptations and trainability of the gut. Nestle Nutr Inst Workshop Ser. 2011;69:1–12. doi:https://doi.org/10.1159/000329268. discussion 13–7.CASArticlePubMedGoogle Scholar
78.Kuchel O, Cuche JL, Buu NT, Guthrie GP, Unger T, Nowaczynski W, et al. Catecholamine Excretion in “Idiopathic” Edema: Decreased Dopamine Excretion, a Pathogenic Factor? The Journal of Clinical Endocrinology Metabolism. 1977;44:639–46. doi:https://doi.org/10.1210/jcem-44-4-639.CASArticlePubMedGoogle Scholar
79.Kuchel O, Cuche JL, Hamet O, Buu NT, Nowaczynski Boucher R, Genest J. Idiopathic edema: New pathogenetic and therapeutic aspects. Mod Med Can. 1976;31:619–24.Google Scholar
86.Frayn KN, Maycock PF. Skeletal muscle triacylglycerol in the rat: methods for sampling and measurement, and studies of biological variability. J Lipid Res. 1980;21:139–44. https://www.ncbi.nlm.nih.gov/pubmed/7354251.
88.Keats TE. Reference Man. A Report Prepared by a Task Group of Committee 2 of the International Commission on Radiological ProtectionReference Man. A Report Prepared by a Task Group of Committee 2 of the International Commission on Radiological Protection. ICRP Publ. 23. Cloth, $50.00; ₤ 21.50. Pp. 480, with figures. Oxford, Pergamon Press, 1975. Radiology. 1975;117:584–584. doi:https://doi.org/10.1148/117.3.584.
89.van Loon LJC, Schrauwen-Hinderling VB, Koopman R, Wagenmakers AJM, Hesselink MKC, Schaart G, et al. Influence of prolonged endurance cycling and recovery diet on intramuscular triglyceride content in trained males. Am J Physiol Endocrinol Metab. 2003;285:E804–11. doi:https://doi.org/10.1152/ajpendo.00112.2003.ArticlePubMedGoogle Scholar
102.National Research Council, Commission on Life Sciences. Food and Nutrition Board, Subcommittee on the Tenth Edition of the Recommended Dietary Allowances. Recommended Dietary Allowances: 10th Edition. National Academies Press; 1989. https://play.google.com/store/books/details?id=gHB32IsIpu0C.
105.Morton RW, Murphy KT, McKellar SR, Schoenfeld BJ, Henselmans M, Helms E, et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. Br J Sports Med. 2018;52:376–84. doi:https://doi.org/10.1136/bjsports-2017-097608.ArticlePubMedGoogle Scholar
106.Bandegan A, Courtney-Martin G, Rafii M, Pencharz PB, Lemon PW. Indicator Amino Acid-Derived Estimate of Dietary Protein Requirement for Male Bodybuilders on a Nontraining Day Is Several-Fold Greater than the Current Recommended Dietary Allowance. J Nutr. 2017;147:850–7. doi:https://doi.org/10.3945/jn.116.236331.CASArticlePubMedGoogle Scholar
107.Mazzulla M, Sawan SA, Williamson E, Hannaian SJ, Volterman KA, West DWD, et al. Protein Intake to Maximize Whole-Body Anabolism during Postexercise Recovery in Resistance-Trained Men with High Habitual Intakes is Severalfold Greater than the Current Recommended Dietary Allowance. J Nutr. 2020;150:505–11. doi:https://doi.org/10.1093/jn/nxz249.ArticlePubMedGoogle Scholar
108.Helms ER, Zinn C, Rowlands DS, Brown SR. A Systematic Review of Dietary Protein During Caloric Restriction in Resistance Trained Lean Athletes: A Case for Higher Intakes. Int J Sport Nutr Exerc Metab. 2014;24:127–38. doi:https://doi.org/10.1123/ijsnem.2013-0054.CASArticlePubMedGoogle Scholar
109.Mäestu J, Eliakim A, Jürimäe J, Valter I, Jürimäe T. Anabolic and Catabolic Hormones and Energy Balance of the Male Bodybuilders During the Preparation for the Competition. Journal of Strength Conditioning Research. 2010;24:1074–81. doi:https://doi.org/10.1519/jsc.0b013e3181cb6fd3.ArticlePubMedGoogle Scholar
110.Antonio J, Ellerbroek A, Silver T, Vargas L, Tamayo A, Buehn R, et al. A High Protein Diet Has No Harmful Effects: A One-Year Crossover Study in Resistance-Trained Males. Journal of Nutrition Metabolism. 2016;2016:1–5. doi:https://doi.org/10.1155/2016/9104792.CASArticleGoogle Scholar
116.Gomez-Arbelaez D, Bellido D, Castro AI, Ordoñez-Mayan L, Carreira J, Galban C, et al. Body Composition Changes After Very-Low-Calorie Ketogenic Diet in Obesity Evaluated by 3 Standardized Methods. J Clin Endocrinol Metab. 2017;102:488–98. doi:https://doi.org/10.1210/jc.2016-2385.ArticlePubMedGoogle Scholar
117.Yang MU, Van Itallie TB. Composition of weight lost during short-term weight reduction. Metabolic responses of obese subjects to starvation and low-calorie ketogenic and nonketogenic diets. Journal of Clinical Investigation. 1976;58:722–30. doi:https://doi.org/10.1172/jci108519.CASArticlePubMed CentralGoogle Scholar
120.Takii H, Kometani T, Nishimura T, Kuriki T, Fushiki T. A sports drink based on highly branched cyclic dextrin generates few gastrointestinal disorders in untrained men during bicycle exercise. Food Sci Technol Res. 2007;10:428–31. https://www.jstage.jst.go.jp/article/fstr/10/4/10_4_428/_article/-char/ja/.
128.Chilibeck PD, Magnus C, Anderson M. Effect of in-season creatine supplementation on body composition and performance in rugby union football players. Appl Physiol Nutr Metab. 2007;32:1052–7. doi:https://doi.org/10.1139/H07-072.ArticlePubMedGoogle Scholar
129.Ziegenfuss TN, Lowery LM, Lemon PWR. Acute fluid volume changes in men during three days of creatine supplementation. J Exerc Physiol Online. 1998;1:1–9. http://www.asep.org/asep/asep/jan13d.htm.
132.Roberts PA, Fox J, Peirce N, Jones SW, Casey A, Greenhaff PL. Creatine ingestion augments dietary carbohydrate mediated muscle glycogen supercompensation during the initial 24 h of recovery following prolonged exhaustive exercise in humans. Amino Acids. 2016;48:1831–42. doi:https://doi.org/10.1007/s00726-016-2252-x.CASArticlePubMedPubMed CentralGoogle Scholar
133.Green AL, Hultman E, Macdonald IA, Sewell DA, Greenhaff PL. Carbohydrate ingestion augments skeletal muscle creatine accumulation during creatine supplementation in humans. American Journal of Physiology-Endocrinology Metabolism. 1996;271:E821–6. doi:https://doi.org/10.1152/ajpendo.1996.271.5.e821.CASArticleGoogle Scholar
134.Vandenberghe K, Van Hecke P, Van Leemputte M, Vanstapel F, Hespel P. INHIBITION OF MUSCLE. PHOSPHOCREATINE RESYNTHESIS BY CAFFEINE AFTER CREATINE LOADING 1417. Medicine & Science in Sports & Exercise. 1997;29 Supplement:249. doi:https://doi.org/10.1097/00005768-199705001-01416.
140.Lepretti M, Martucciello S, Burgos Aceves MA, Putti R, Lionetti L. Omega-3 Fatty Acids and Insulin Resistance: Focus on the Regulation of Mitochondria and Endoplasmic Reticulum Stress. Nutrients. 2018;10. doi:https://doi.org/10.3390/nu10030350.
142.Brunton LL, Lazo JS, Parker K, Buxton I, Blumenthal D. Book, Review: Goodman and Gilman’s The Pharmacological Basis of Therapeutics: Digital Edition, 11th Edition. Annals of Pharmacotherapy. 2006;40:1218–1218. doi:https://doi.org/10.1345/aph.1g685.
145.Clare BA, Conroy RS, Spelman K. The Diuretic Effect in Human Subjects of an Extract of Taraxacum officinale Folium over a Single Day. The Journal of Alternative Complementary Medicine. 2009;15:929–34. doi:https://doi.org/10.1089/acm.2008.0152.ArticlePubMedGoogle Scholar
147.Mydlík M, Derzsiová K, Žemberová E. Influence of Water and Sodium Diuresis and Furosemide on Urinary Excretion of Vitamin B6, Oxalic Acid and Vitamin C in Chronic Renal Failure. Miner Electrolyte Metab. 1999;25:352–6. doi:https://doi.org/10.1159/000057474.ArticlePubMedGoogle Scholar
148.Kenawy MR, El-Nabawy, El-Mohandis MM, El -D, Rohayem HK, El-Sheehy AW. Studies on the diuretic action of vitamin C in normal animals and human beings, and its clinical value in pathological retention of water. Int Z Vitaminforsch. 1952;24:40–61. https://www.ncbi.nlm.nih.gov/pubmed/12999380.
178.Vila-Chã C, Hassanlouei H, Farina D, Falla D. Eccentric exercise and delayed onset muscle soreness of the quadriceps induce adjustments in agonist–antagonist activity, which are dependent on the motor task. Exp Brain Res. 2012;216:385–95. doi:https://doi.org/10.1007/s00221-011-2942-2.ArticlePubMedGoogle Scholar
182.Décombaz J, Schmitt B, Ith M, Decarli B, Diem P, Kreis R, et al. Postexercise fat intake repletes intramyocellular lipids but no faster in trained than in sedentary subjects. Am J Physiol Regul Integr Comp Physiol. 2001;281:R760–9. doi:https://doi.org/10.1152/ajpregu.2001.281.3.R760.ArticlePubMedGoogle Scholar
183.Hocking S, Samocha-Bonet D, Milner K-L, Greenfield JR, Chisholm DJ. Adiposity and insulin resistance in humans: the role of the different tissue and cellular lipid depots. Endocr Rev. 2013;34:463–500. doi:https://doi.org/10.1210/er.2012-1041.CASArticlePubMedGoogle Scholar
186.Morton RW, Sonne MW, Zuniga AF, Mohammad IYZ, Jones A, McGlory C, et al. Muscle fibre activation is unaffected by load and repetition duration when resistance exercise is performed to task failure. The Journal of Physiology. 2019;597:4601–13. doi:https://doi.org/10.1113/jp278056.CASArticlePubMedGoogle Scholar
188.The correct interpretation of the size principle. and it’s practical appliction to resistance training – Научные статьи – Библиотека международной спортивной информации. http://bmsi.ru/doc/c33fb1e0-9e05-44fc-a4c7-ad36356db8ea. Accessed 7 Aug 2020.
189.Child RB, Saxton JM, Donnelly AE. Comparison of eccentric knee extensor muscle actions at two muscle lengths on indices of damage and anglespecific force production in humans. J Sports Sci. 1998;16:301–8. doi:https://doi.org/10.1080/02640419808559358.CASArticlePubMedGoogle Scholar
192.Collier G, O’Dea K. The effect of coingestion of fat on the glucose, insulin, and gastric inhibitory polypeptide responses to carbohydrate and protein. The American Journal of Clinical Nutrition. 1983;37:941–4. doi:https://doi.org/10.1093/ajcn/37.6.941.CASArticlePubMedGoogle Scholar
193.Gentilcore D, Chaikomin R, Jones KL, Russo A, Feinle-Bisset C, Wishart JM, et al. Effects of Fat on Gastric Emptying of and the Glycemic, Insulin, and Incretin Responses to a Carbohydrate Meal in Type 2 Diabetes. The Journal of Clinical Endocrinology Metabolism. 2006;91:2062–7. doi:https://doi.org/10.1210/jc.2005-2644.CASArticlePubMedGoogle Scholar
195.Blom PC, Høstmark AT, Vaage O, Kardel KR, Maehlum S. Effect of different post-exercise sugar diets on the rate of muscle glycogen synthesis. Med Sci Sports Exerc. 1987;19:491–6. https://www.ncbi.nlm.nih.gov/pubmed/3316904.
196.Shi X, Passe DH. Water and solute absorption from carbohydrate-electrolyte solutions in the human proximal small intestine: a review and statistical analysis. Int J Sport Nutr Exerc Metab. 2010;20:427–42. doi:https://doi.org/10.1123/ijsnem.20.5.427.ArticlePubMedGoogle Scholar
204.Freitas EDS, Miller RM, Heishman AD, Ferreira-Júnior JB, Araújo JP, Bemben MG. Acute Physiological Responses to Resistance Exercise With Continuous Versus Intermittent Blood Flow Restriction: A Randomized Controlled Trial. Front Physiol. 2020;11. doi:https://doi.org/10.3389/fphys.2020.00132.
205.Freitas EDS, Poole C, Miller RM, Heishman AD, Kaur J, Bemben DA, et al. Time Course Change in Muscle Swelling: High-Intensity vs. Blood Flow Restriction Exercise. Int J Sports Med. 2017;38:1009–16. doi:https://doi.org/10.1055/s-0043-118342.ArticlePubMedGoogle Scholar
DISCLAIMER: Il presente articolo NON intende in alcun modo consigliare il regime alimentare ivi presentato. Si tratta di semplice divulgazione scientifica e non ha nessun valore medico e/o prescrittivo.
Introduzione:
Le mode alimentari che hanno caratterizzato gli ultimi sessant’anni delle preparazioni ai contest di Bodybuilding mostrano pratiche gestionali dietetiche dal similare al totalmente opposto. Si è passati dalle “low fat” alle “higt fat” con un alternanza quasi ciclica e sempre più dipendente dalla “leggenda da spogliatoio” che non dalla ricerca scientifica. Oggi, per lo meno buona parte di noi, sa che la chiave principale della perdita di peso/grasso è il deficit calorico. L’ipotesi dell’Insulina e quella secondo la quale “una caloria non è una caloria” sono state bocciate dal metodo scientifico e dalla pratica ormai da tempo. E allora perchè dedicare un articolo ad una metodologia alimentare estrema com’è la “Carne e Acqua” se alla fine, semplicemente, basta un deficit calorico di qualsiasi tipo e con un adeguato apporto proteico per raggiungere l’agognato obbiettivo? Beh, i motivi sono principalmente 2:
1- Lo sviluppo e le caratteristiche che tale pratica ha avuto nel mondo del Culturismo agonistico, ma non solo, nel tempo;
2- La sua capacità di essere adattabile e, seppur con una sostenibilità limitata, applicabile su un discreto numero di soggetti pur presentando tagli calorici molto drastici per un numero di giorni variabili durante la settimana.
Essendo, nel mio piccolo, un ricercatore anche in campo nutrizionale, ho osservato e analizzato l’applicazione di questa pratica nelle sue diverse varianti su diversi bodybuilder (sia “Natural” che “Doped”), maturando una valutazione sufficientemente oggettiva e arrivando a “crearne” due nuove versioni.
Ma non dilunghiamoci oltre, per il momento, e partiamo dal principio…
Esiste una letteratura sulla dieta “Carne e Acqua”?
Nello specifico, la risposta è no sebbene esista una discreta letteratura su una condizione alimentare estrema osservata nelle spedizioni artiche o in altre condizioni al limite della sopravvivenza dove l’unica fonte di cibo facilmente reperibile era selvaggina dalle carni estremamente magre: si tratta del “Protein poisoning“, letteralmente “avvelenamento da proteine”.
L’avvelenamento da proteine (chiamato anche colloquialmente come rabbit starvation, morbo del caribou, o fat starvation) è una forma acuta di malnutrizione causata da una dieta carente di grassi, in cui quasi tutte le calorie consumate provengono da carni magre.[1][2 ] Il concetto è solitamente discusso nel contesto delle ipotesi paleoantropologiche sulla dieta degli antichi esseri umani, specialmente durante l’ultimo massimo glaciale e alle alte latitudini.[3][4]
Il termine “rabbit starvation” deriva dal fatto che la carne di coniglio è molto magra, con quasi la totalità delle calorie da essa provenienti apportate dalle proteine e una percentuale di grasso molto bassa. Di conseguenza, è un alimento che, se imperante nella dieta, causerebbe il così detto “avvelenamento da proteine”.[4] D’altra parte, si è osservato che alcuni animali che vivono in ambienti terrestri rigidi e freddi si presentano molto magri.[3]
In Storia Romana di Appiano, Volume I, Libro VI: Le guerre in Spagna, capitolo IX, pagina 223, l’autore osserva e descrive che una moltitudine di soldati romani morivano di dissenteria grave dopo aver mangiato quasi esclusivamente conigli, mentre assediavano la città di Intercatia nel 150 A.C. :
… strano terrore nell’accampamento romano. I soldati erano malati per la veglia e la mancanza di sonno, e per il cibo inconsueto che il paese offriva. Non avevano vino, né sale, né aceto, né olio, ma vivevano di grano e orzo, e quantità di carne di cervo e di coniglio bollite senza sale, causando la dissenteria, dalla quale molti morirono. [5]
Soldati romani consumano il rancio.
Si dice che l’esploratore Vilhjalmur Stefansson abbia vissuto per anni esclusivamente di carne di selvaggina e pesce, senza effetti negativi. Lo stesso vale per il suo compagno esploratore Karsten Anderson. Come parte della sua promozione della dieta a base di carne modellata sulla cucina Inuit, e per dimostrarne gli effetti, a New York City a partire dal febbraio 1928, Stefansson e Anderson “vissero e mangiarono nel reparto metabolismo del Russell Sage Institute of Pathology del Bellevue Hospital. , New York” per un anno, con le loro prestazioni metaboliche osservate da vicino, tutto questo in parte finanziato dall’Institute of American Meat Packers.[6] I ricercatori che speravano di replicare l’esperienza di Stefansson con la “rabbit starvation” sul campo lo hanno esortato a ridurre a zero l’assunzione di grassi nella sua dieta a base di carne. Lo ha fatto, e ha sperimentato un’insorgenza di diarrea molto più rapida rispetto a quanto osservato nel campo. Con l’aggiunta di grasso, Stefansson si è ripreso, sebbene con un periodo seguente di stitichezza di 10 giorni. Lo studio ha riferito di non aver trovato letteratura medica precedente che esaminasse gli effetti delle diete a base di sola carne, che sembrano essere sostenibili, o sulla “rabbit starvation”, che è fatale.
Stefansson scrisse:
I gruppi che dipendono dagli animali grassi sono i più fortunati nello stile di vita della caccia, perché non soffrono mai di fat starvation. Questo problema è peggiore, per quanto riguarda il Nord America, tra quegli indiani delle foreste che dipendono a volte dai conigli, l’animale più magro del Nord, e che sviluppano l’estrema fame di grasso nota come rabbit starvation. I mangiatori di conigli, se non assumono grasso da un’altra fonte – castoro, alce, pesce – svilupperanno diarrea in circa una settimana, con mal di testa, stanchezza e vago disagio. Se ci sono abbastanza conigli, la gente mangia fino a dilatare lo stomaco; ma non importa quanto mangiano si sentono insoddisfatti. Alcuni pensano che un uomo morirà prima se mangia continuamente carne senza grasso piuttosto che se non mangia nulla, ma questa è una credenza sulla quale nel Nord non sono state raccolte prove sufficienti per una conferma in merito. Le morti per rabbit starvation o per il consumo di altra carne magra sono rare; poiché tutti ne comprendono il principio, e vengono naturalmente prese tutte le misure preventive possibili.[7]
Vilhjalmur Stefansson durate una delle sue spedizioni artiche.
Nella prefazione del libro di Alden Todd “Abbandonati: la storia della spedizione artica Greely, 1881-1884” sempre lo stesso Stefansson sostiene che sia stato il cannibalismo dei compagni già morti a provocare il decesso di buona parte dell’equipaggio come conseguenza di questa inquietante e inusuale rabbit starvation. Charles Darwin nel “Viaggio del Beagle” descrisse esperienze simili relative alla fame da grassi, pur valutando che i gauchos argentini mangiavano per mesi esclusivamente chili di carne di manzo ogni giorno apparentemente senza problema alcuno. In questo ultimo caso, la risposta alla tolleranza dei gauchos è con tutta probabilità da attribuirsi alla percentuale di grasso presente nella carne di manzo che, anche quando il taglio è magro, risulta essere di circa 5g per ogni 100g di carne.
Un opuscolo sulla sopravvivenza artica dell’epoca della seconda guerra mondiale emesso dal comando di controllo di volo delle forze aeree dell’esercito degli Stati Uniti includeva questo enfatico avvertimento:
A causa dell’importanza dei grassi, in nessuna condizione limitarsi a una dieta a base di carne di coniglio solo perché si verifica essere abbondante nella regione in cui sei costretto a stanziare. Una dieta continua di coniglio produrrà la rabbit starvation – la diarrea inizierà in circa una settimana e se la dieta viene continuata POTREBBE RISULTARE LA MORTE.[8]
In Into the Wild (1996), Jon Krakauer ha ipotizzato che Chris McCandless, un avventuriero americano del XX secolo, potrebbe aver sofferto di rabbit starvation.
Chris McCandless
La revisione dell’assunzione di riferimento dietetico statunitense e canadese per le proteine menziona la “rabbit starvation”, ma ha concluso che non c’erano prove sufficienti fino al 2005 per stabilire un livello di assunzione superiore tollerabile, ovvero un limite massimo per la quantità di proteine che può essere consumata in sicurezza.[9 ] Secondo quanto riferito, gli esseri umani moderni sono in grado di ricavare solo il 20% del loro fabbisogno energetico dalle proteine.[10] Per i cacciatori-raccoglitori artici, tuttavia, la quantità può aumentare stagionalmente fino al 45%.[11] In realtà, specie negli atleti, si è osservata un ampia capacità di adattamento alla quota proteica senza ripercussioni negative sul medio termine (circa 4g/Kg). Secondo Bilsborough e Mann (2006), l’assunzione di proteine è principalmente limitata dal ciclo dell’urea. Suggeriscono, quindi, un limite di 2,5 g/kg.[10] L’errore di fondo, è quello di confondere una possibilità di adattamento funzionale (quota proteica aumentata) con la mancanza di lipidi, totale o marcata, nella dieta.
Quindi, è chiaro che la letteratura scientifica non ci fornisce moltissime informazioni in merito a questo argomento. E, a proposito di ciò, esiste uno studio [12] su di un singolo caso di decesso per rabbit starvation riportato anche dal fanatico della “Paleo Dieta” Loren Cordain il quale peraltro ritiene [1], similmente a Bilsborough e Mann, che a fronte di un carico proteico eccessivo il fegato non sia in grado di produrre enzimi sufficienti per la sintesi dell’urea. Oltre un certo limite variabile, infatti, l’organismo va in iperammonemia e iperaminoacidemia.[13] Il tratto gastro-intestinale potrebbe assorbire in teoria non oltre 1,3 – 10 gr di aminoacidi ogni ora [14], anche se il dato ci appare superato di gran lunga all’atto pratico. Lo stesso autore d’altronde nella ricerca citata sottolinea come il 75% delle comunità di cacciatori–raccoglitori ricavi da fonti animali fino al 73% (98% gli Eskimos) del proprio nutrimento. Il limite di tolleranza sarebbe raggiunto sempre secondo Cordain quando l’apporto proteico equivale a circa il 40% dell’introduzione calorica complessiva, secondo Billsborought quando si aggira sul 35%.
La “Carne e Acqua” ed il Bodybuilding:
Se vogliamo parlare di dieta “Carne e Acqua” nel Bodybuilding non possiamo esimerci dal citare Rheo Blair.
Rheo H. Blair è stato il primo uomo riconosciuto come un “mago della nutrizione” quando si trattava di dieta e integratori per il Bodybuilding. Ha regolarmente eseguito degli ottimi lavori di trasformazione fisica su centinaia di bodybuilder inferiori alla media dei competitor. Durante gli anni ’50, ’60 e ’70, abbondavano le testimonianze sulle incredibili trasformazioni fisiche che Blair aveva compiuto su centinaia di “casi senza speranza”. Si diceva trasformasse regolarmente i deboli di 97 libbre in uomini robusti con le sue speciali formule proteiche e la vigorosa routine di allenamento con i pesi basata sul volume. Anche i bodybuilder avanzati riportavano risultati simili dal sistema Blair. In un articolo del numero di maggio 1967 della rivista Iron Man, un bodybuilder scrisse:
“Dopo aver seguito il programma di Rheo per sole tre settimane, ho ottenuto più guadagni di quelli che ho avuto negli ultimi sei anni. Ho messo quasi mezzo pollice sulle mie braccia. E dopo due mesi ho messo su quasi 20 libbre di muscoli puri.”
Sotto la guida di Blair, Jim Park passò dall’essere uno sconosciuto bodybuilder con un fisico nella media ad essere Mr. America… in meno di quattro mesi! Non si esclude l’uso di AAS che, già negli anni 50, pur essendo di nicchia e non ancora dilaganti, erano presenti. Ovviamente, come vuole il politically correct, viene, e venne, affermato che questa straordinaria trasformazione era stata realizzata senza farmaci! Adducendo al fatto che, dal momento che il tutto accadeva negli anni ’50, cioè prima che l’uso degli AAS diventasse così diffuso nel bodybuilding, la probabilità d’uso non era possibile. Personalmente, lo trovo poco importante e banale come discussione. C’erano AAS disponibili all’epoca? Si (es. Methyltestosterone, Mesterolone e Testoterone Propionato). Avrebbe potuto averne accesso? Possibile. E’ importante? Non per chi valuta l’atleta e non il suo “sgabuzzino”.
Sebbene avesse la reputazione di essere un eccentrico, tutti i grandi bodybuilder degli anni ’50, ’60 e ’70 (Arnold, Frank Zane, Dave Draper, Larry Scott, ecc.) hanno seguito i consigli di Blair e sono entrati nella loro forma migliore. Uno dei segreti di Blair era la sua speciale formula proteica. La famosa polvere proteica di Blair si basava sui rapporti di aminoacidi nel latte materno. Era anni in anticipo sui tempi. Sfortunatamente, Blair è morto prematuramente nei primi anni ’80 e molti dei suoi “segreti” sul bodybuilding sono morti con lui… inclusa la formula esatta della sua polvere proteica.
Rheo Blair
Ma la sua “arma” nelle preparazioni alimentari, soprattutto nella preparazione alla gara, era l’uso elevato di proteine con un esclusione marcata di Carboidrati e Grassi.
Come abbiamo visto in precedenza, mentre Vilhjamur Stefannsson ha reso popolare la dieta a base di carne degli Inuit all’inizio del 1900, una dieta a base di carne per atleti sembra essere un nuovo sviluppo dietetico, anche se così non è. Facendo eco alla meravigliosa “serie niente di nuovo sotto il sole” prodotta da Chaos and Pain (sicuramente non sicura per il lavoro!), abbiamo precedenti per la dieta “Carne e Acqua” anche con Rheo H. Blair e, in un certo qual modo, con Vince Gironda, diete dimagranti a breve termine utilizzate dai bodybuilder prima di una competizione.
A parte Vince Gironda, Rheo H. Blair è, a mio avviso, uno dei personaggi più affascinanti del bodybuilding degli anni ’50 e ’60. Fondamentali nella divulgazione degli integratori proteici, le polveri proteiche a base di latte e le compresse vitaminiche di Blair hanno acquisito uno status quasi mitico tra la comunità del sollevamento pesi. Blair contava tra i suoi clienti bodybuilder, atleti, celebrità e individui comuni. Il suo continuo interesse per la nutrizione fu senza dubbio la ragione del suo successo. Fu questo interesse che portò al suo esperimento “carne e acqua”, una dieta simile alla “dieta di massima definizione” di Gironda. Come raccontato da Steve Davis, che ha subito un drastico cambiamento nella dieta a base di carne e acqua, Blair ha voluto stabilire se una dieta di questo genere fosse preferenziale rispetto alla dieta a base di carne e uova promossa da Gironda, quest’ultima soprannominata “maximum definition diet”. Preparandosi per un servizio fotografico, Davis, allora aspirante bodybuilder, si è dimostrato una cavia ideale.
Al momento ho prove che i bodybuilder usassero la dieta a base di carne e acqua di Blair alla fine degli anni ’60 e all’inizio degli anni ’70. Il primo, Steve Davis, ha usato la dieta per coronare un’incredibile perdita di peso. Come raccontato da Old School Bodybuilding, Davis era inizialmente un powerlifter che voleva cimentarsi nel bodybuilding. Sotto la guida di Vince Gironda e Rheo H. Blair, ha subito una notevole perdita di peso che comprendeva quasi 100 libbre (circa 45,35Kg).
Parlando con Dennis Weis in Raw Muscularity, Davis ha raccontato le sue esperienze non così felici sul programma alimentare a base di carne e acqua:
Per raggiungere la forma in queste foto mi sono reso conto che avrei dovuto perdere quel minuscolo strato di tessuto adiposo per affinare davvero il mio corpo alla condizione di un Larry Scott o di un Gable Boudreaux…
A questo punto del programma Rheo ha detto: “Steve, vogliamo aiutarti a ottenere un po’ di magrezza in più per il tuo corpo e c’è un programma dietetico che possiamo usare per farlo. Vivere solo di carne e acqua…
Rheo mi ha spiegato che, per potermi preparare appositamente per il servizio fotografico, solo per due o cinque giorni al massimo avrei dovuto vivere di nient’altro che carne più un integratore di proteine della carne …
Ho continuato questo tipo di programma per un periodo fino a dodici giorni. E questa è la parte triste della storia. Sono diventato così fisicamente esausto, così teso, così tassato e tirato che dopo che Rheo aveva scattato le foto ero sul punto di crollare.
Steve Davis prima e dopo essere diventato un atleta di Blair.
Ora, ciò che è importante sottolineare della testimonianza di Davis è che ha intrapreso una dieta a base di carne che era estremamente povera di calorie per prepararsi ad un servizio fotografico nel breve termine.
Passando ora a Heart of Steel, una meravigliosa biografia di Dan Lurie, troviamo menzione di innumerevoli altri bodybuilder che usarono la dieta di Blair, incluso il rivale di Arnold in Pumping Iron, Lou Ferrigno. Parlando all’inizio degli anni ’70, un tempo in cui Lou Ferrigno era ancora un imponente bodybuilder piuttosto che “l’incredibile Hulk”, Lurie commentò che:
Lui (Lou Ferrigno) è stato quindi sottoposto a una dieta speciale “carne e acqua” per ridurre il suo peso, per ottenere un aspetto “strappato”, da competizione. I bodybuilder che volevano preservare i muscoli mentre perdevano grasso usavano spesso la dieta a base di carne e acqua a quei tempi, e aveva funzionato a meraviglia per campioni come Vince Gironda, e per gli atleti seguiti da Rheo H. Blair e altri risalenti agli anni ’50.
Era una dieta che prevedeva carne di ogni tipo, poche verdure e acqua, il che la rendeva ricca di proteine, moderatamente grassa e con pochissimi carboidrati. E Lou ha fatto grandi progressi con questo piano.
Lou Ferrigno vincitore del Mr. Universo 1974.
Quindi Davis era in buona compagnia durante l’età dell’oro del Bodybuilding. Su questo punto sono probabilmente necessarie spendere alcune parole. La riduzione del grasso corporeo per le competizioni o i servizi fotografici è stata spesso eseguita in modo semplice: ridurre gli amidi (quindi le calorie dai carboidrati) e aumentare l’attività fisica (maggiore dispendio calorico). Non è avvenuto fino agli anni ’80 che gli atleti hanno iniziato a contare le calorie in modo ossessivo. Non che contare le calorie sia sbagliato, ma l’eccesso porta sempre a ripercussioni negative. Dalla metà del secolo, si può quindi osservare una adesione nel pre-contest molto evidente alla dieta “carne e acqua”, era semplice ed efficace. Inoltre ha avuto il sostegno di alcuni dei migliori allenatori e atleti di questo sport. Anche quella “statua vivente” quale fu Serge Nubret, in preparazione alla gara seguiva un regime “Carne e Acqua” basato su carne, pesce e amminoacidi.
Serge Nubret
Vi sono taluni che pensano che la “dieta carnivora” per gli atleti sia nata negli ultimi decenni insieme a regimi ortoressici e inutilmente restrittivi come la “Paleo Dieta”, ma come abbiamo visto non è così. Non sono estraneo alle diete restrittive – ho osservato e seguito molti soggetti sotto regime chetogenico per quasi sei anni – ma il concetto di “dieta carnivora” mi è sempre parso intrinsecamente fallimentare se non adeguatamente contestualizzato e, in definitiva, inserito in un piano di preparazione culturistica. Non sono affatto favorevole all’applicazione di questo modo di mangiare sul lungo termine, poiché risulterebbe controproducente su più aspetti della salute umana. Ma la dieta “carne e acqua”, come quella di Blair, dà a questa metodica alimentare una giusta e limitata dimensione nell’ambito della preparazione alla gara di Bodybuilding.
Attualmente, il maggior sostenitore della dieta carnivora come stile di vita è Shawn Baker, ex medico ortopedico americano.(15) Egli cita fantomatiche testimonianze di coloro che seguono la dieta carnivora come prova (non provata) che può curare la depressione, l’ansia, l’artrite, l’obesità, il diabete e altro.[15][16] Ovviamente, nessuna ricerca ha analizzato gli effetti della dieta carnivora e dimostrato quanto precedentemente asserito. Inoltre, nel 2017 la licenza medica di Baker è stata revocata dal New Mexico Medical Board a causa delle preoccupazioni sulla sua competenza.[17]
E’ chiaro, quindi, che il concetto applicativo di Blair e Baker differiscono significativamente nella loro applicazione. Baker, da quanto egli stesso fa trasparire, vede la dieta carnivora come una dieta a vita, il che significa che può essere utilizzata per anni senza problemi (secondo lui). Blair e Gironda, invece, con intelligenza preferivano un uso a breve termine. Gironda, ad esempio, aveva di logica un approccio alimentare ciclico con i suoi clienti a seconda dei loro obiettivi. Quindi, si sta parlando fondamentalmente di una forma commerciale venduta al grande pubblico (la dieta carnivora di Baker) e di una strategica per il miglioramento della composizione corporea rivolta ad atleti in preparazione ad un contest di Bodybuilding.
La mia ricerca applicata:
Dal 2016 ho iniziato a raccogliere dati sulla pratica alimentare “carne e acqua” valutandone gli effetti su diversi culturisti, sia agonisti in preparazione alla gara che amatori nella fase “Cut”.
La prima cosa che annotai, e che era del tutto presumibile già partendo dai dati preliminari in mio possesso, era che la “carne e acqua” risultava tollerabile in un numero ristretto di persone. ma questo era direttamente proporzionale al numero di giorni nei quali l’atleta rimaneva alimentato totalmente con carni magre. Classificai tre principali modalità di applicazione:
Modalità Estrema: si trattava di seguire un regime alimentare basato su carni magre e integratori di fibre (anche se non sempre) e integratori multi vitaminici-minerali per un periodo determinato totalmente sulla resistenza del soggetto sottoposto e dalla valutazione della forma fisica;
Modalità Metabolica: su stampo della famosissima dieta di Mauro di Pasquale, “La Dieta Metabolica”, l’atleta segue un regime “carne e acqua” per 5-6 giorni a settimana per poi “ricaricare” con Carboidrati e Grassi per 1-2 giorni a settimana;
Modalità Ciclica: simile alla precedente, essa tiene strettamente conto delle risposte psicofisiche dell’atleta alternando periodi a “carne e acqua” con giorni di refeed.
La più problematica e meno sostenibile risulta essere, e per ovvie ragioni, la “Modalità Estrema”. Se dovessimo elencare i principali problemi questi sarebbero:
Deficit nutrizionali;
Aumento del Cortisolo in risposta al deficit calorico e alle richieste metaboliche per la gestione del carico amminoacidico.
Risposta fisiologica e non determinante nella negativizzazione della composizione corporea nel breve termine.
Ricordiamoci inoltre che la transaminazione al fine di convertire le proteine in urea e indurre la sintesi degli aminoacidi derivati in glucosio è un processo dal costo metabolico molto elevato. E’ un processo aerobico che ha il proprio limite nella disponibilità di ossigeno del fegato. La capacità complessiva del sistema si attesta comunque sulla produzione di 250g circa. E’ vero anche che la conversione metabolica degli amminoacidi in Glucosio, per via della sua richiesta energetica, è soggettivamente limitato e dipendente dagli adattamenti metabolici in atto o all’uso di determinati farmaci.
Via della gluconeogenesi con molecole ed enzimi chiave. Molti passaggi sono opposti a quelli che si osservano nella glicolisi.
Al deficit calorico diretto va sommato, oltre a quanto sopra, quello causato dalla termogenesi indotta dal cibo che nel caso delle Proteine si aggira tra il 10 ed il 35% (22,5% in media) delle calorie ingerite.
Questo aspetto risulta positivo, almeno in parte, ma insieme ad esso, in una “carne e acqua” estrema, va considerato l’enorme deficit nutrizionale che, nonostante la possibilità di sopperire a ciò sul piano vitaminico, minerale e di fibre, colpisce duramente la richieste fisiologiche di una quantità sufficiente di Grassi. Inoltre, la privazione eccessivamente prolungata del consumo glucidico peggiora per circostanze adattative il metabolismo glucidico ed i vantaggi ad esso legati (vedi, per esempio, la qualità della prestazione, la capacità di utilizzo del substrato energetico una volta reintrodotto ecc… ).
L’aumento del Cortisolo, e so già che alcuni limitati si scandalizzeranno, è la preoccupazione minore in quanto trattasi di un adattamento fisiologico che può diventare un problema nel cronico. Stesso discorso vale per lo squilibrio della bilancia acido-base dell’organismo e conseguente catabolismo muscolare. Ciò si verifica solo in sistemi organici gravemente compromessi, come quelli osservati all’inizio del XX secolo nelle aree del Canada dove per molti mesi non ci si nutriva se non di coniglio e qualche radice.
Comunque sia, gli atleti che optavano per la versione più “bruta” della “carne e acqua” arrivavano ben presto ad un esaurimento fisico e mentale con forte nervosismo e calo della performance sportiva. Non mi dilungherò a parlare dei volumi che, e questo lo dovreste sapere più o meno tutti, venivano rapidamente supercompensati con i refeed.
Le più tollerabili tra le versioni elencate erano la “Metabolica” e la “Ciclica”. Quest’ultima, la migliore in assoluto tra le tre versioni, viene retta anche da alcuni “Natutral” che, al fine di tagliare le calorie totali della settimana tenevano due giorni in cui il loro consumo calorico era molto ridotto, e per fare ciò usavano in quei giorni una “carne e acqua”.
Personalmente, sperimentai una mia prima versione di dieta “carne e acqua ibridata” nell’estate del 2016. Si trattava di ciclicizzare 3 giorni in regime simil-Chetogenico (versione dieta Atkins Modificata) seguiti da 1 giorno di refeed seguito a sua volta da 3 giorni a “carne e acqua” per poi ripetere la sequenza per tutta la durata della programmazione. All’epoca la ribattezzai con il “pacchianissimo” nome di “Roller Coaster“.
Ripartizione macro-calorica nelle fasi della “Roller Coaster”
L’anno successivo, siamo quindi nel 2017, modificai il sopra citato schema rendendolo pianificabile sui canonici 7 giorni settimanali. Si trattava quindi di 3 giorni in regime simil-Chetogenico (versione dieta Atkins Modificata) seguito da 1 giorno di refeed seguito a sua volta da 2 giorni a “carne e acqua” e successivamente un altro giorno di refeed. Questa volta la ribattezzai “Keto Starvation”.
Ripartizione macro-calorica nelle fasi della “Keto Starvation””
Quest’ultimo schema applicativo risultò essere ben tollerato e con pochi disagi se non gli iniziali annessi a tutti i regimi low-carb (mal di testa, spossatezza, feci molli) che tendono a scomparire dopo poche settimane.
Da questa versione ne è nata una recente e decisamente più “drastica”. Infatti, la “Protein Starvation” consiste in un piano settimanale nel quale vi sono 3 giorni iniziali a “carne e acqua” seguiti da 1 giorno di refeed glucidico/low fat, successivamente vi sono altri 2 giorni a “carne e acqua” e 1 giorno di refee glucidico/lipidico.
Ripartizione macro-calorica nelle fasi della “Protein Starvation””
Questo schema è poco tollerato dalla maggior parte delle persone ma ha mostrato effetti su periodi di tempo di 8 settimane statisticamente significativi rispetto a quanto ottenuto negli stessi soggetti con il regime più “soft”. Ed è ovvio che non si tratta di una pratica propriamente da “Natural”.
Importante da ricordare è che vi è un integrazione d’obbligo in questa pratica alimentare:
Multivitaminico Multiminerale;
Potassio;
Magnesio;
Calcio;
Ferro;
EPA+DHA;
Zinco;
Fibra o in alternativa shirataki di Konjac.
Supplementazione addizionale:
Metformina: l’uso della Metformina trova la sua ragione d’essere in una “carne e acqua” per via del suo effetto su l’attività metabolica cellulare via PPAR e AMPK, con conseguente miglioramento del metabolismo energetico sia glucidico che lipidico che, sebbene ci si trovi in un regime low-carb, ipoteticamente dovrebbe indurre delle “forzature di sistema” tali da spingere l’organismo ad attingere maggiormente dai depositi adiposi e dal surplus amminoacidico come substrati di sostentamento per via della “precarietà” dei livelli di glucosio ematico. Inoltre, la Metformina riduce le concentrazioni di glucosio plasmatico a digiuno riducendo i tassi di produzione epatica di glucosio a partire dagli amminoacidi [18][19], il suo effetto sui contributi relativi della glicogenolisi epatica e della gluconeogenesi rimane comunque controverso. Alcuni studi concludono che la Metformina agisca principalmente riducendo i tassi di gluconeogenesi [20]; altri, che agisce riducendo i tassi di glicogenolisi epatica [21][22]. Comunque sia, come già precedentemente accennato, il corpo può ricavare solo 1000 calorie al giorno attraverso la gluconeogenesi in una dieta di sole proteine dal momento che il fegato è in grado di produrre solo 250g di glucosio dalle proteine, e non importa quante proteine si mangiano. Con l’aggiunta di 750mg/die di Metformina si ipotizza, almeno da considerazioni fatte per via di rapporti sui cambiamenti glicemici, per quello che valgono, che la produzione di glucosio scenda a circa 180g al giorno, al limite del mantenimento dei tessuti glucosio dipendenti.
Metformina
Nota: l’uso della Metformina può dare dissenteria e problemi gastrointestinali.
Sfortunatamente, nei piani “carne e acqua” prolungati, il fegato inizierà a non riuscire a convertire l’ammoniaca in urea (non abbastanza ATP), quindi l’ammoniaca rientrerà nel flusso sanguigno. Questo inizierà a dare problemi al sistema nervoso.
Mi sembra scontato aggiungere che la dove viene applicata una supplementazione farmacologica strategica e contestualizzata il piano da i suoi migliori risultati. Per esempio, oltre a quanto detto prima per la Metformina, l’uso del 7-Keto-DHEA, o di altro inibitore della 11β–HSD-1, riduce la risposta cortisolemica. Alcuni riducono l’assorbimento lipidico dei pasti con fonti non accuratamente pesate e che possono contenere una quantità di grassi più elevata delle fonti comunemente consumate.
Azione del 7-Keto-DHEA sull’attività dell’enziama 11 β-HSD-1
Vi ricordo, e sto parlando all’idiota che sta sempre dietro lo schermo leggendo ma non capendo una emerita ciola, che estremizzare questo tipo di dieta già di suo estrema scimmiottando quanto sofferto dagli esploratori del secolo scorso beh, gli effetti non sono affatto piacevoli e comprendono nausea e affaticamento iniziali, seguiti da diarrea continua e infine, nei casi cronici, la morte.[4]
Conclusione:
Cosa si può imparare dalla dieta “carne e acqua”?
Lo ripeto affinché sia chiaro a tutti: la dieta “carne e acqua” trova il suo motivo d’esistere per brevi periodi di tempo (vedi pre-contest), e ancora meglio se nello schema alimentare i giorni di “starvation” vengono intervallati da refeed glucidici e misti!
Come ben si comprende, sembra che la dieta “carne e acqua” abbia funzionato per un certo numero di culturisti, un punto che sottolinea la necessità di adattare il piano alimentare alle capacità adattative del soggetto. Ma questo, ad oggi, lo abbiamo ben capito, di qualsiasi piano alimentare o allenante si tratti.
Consiglio la “carne e acqua” o la mia “Protein Starvation”? Non la consiglio da un punto di vista etico e professionale dal momento che per seguire tali regimi il soggetto interessato deve per forza di cose essere attentamente seguito da un professionista onde evitare che si ritrovi collassato sul cesso a causa di una pesante disidratazione da dissenteria. Non consiglio in nessun modo di seguire la “dieta carnivora” come stile di vita promossa da Shawn Baker, una follia!
Per il resto, affidatevi a personale qualificato prima di intraprendere qualsiasi percorso alimentare.
“Not by Bread Alone”, Vilhjalmur Stefansson, Publisher, Macmillan, 1946
Jungle, Desert, and Arctic Emergencies Booklet. Flight Control Command Safety Education Division of the United States Army Air Forces. 1 January 1941. p. 116,119. Retrieved 27 July 2020.
Jump up to:ab Bilsborough, S; Mann, N (April 2006). “A review of issues of dietary protein intake in humans”. International Journal of Sport Nutrition and Exercise Metabolism. 16 (2): 129–52.
Non è la prima volta che attraverso un articolo o un post tratto di molecole con un ipotetico potenziale sulla riduzione dell’accumulo di grasso in contesto di una dieta ipercalorico. Questa volta vorrei parlare degli studi effettuati sulla Alpinia officinarum e pubblicati sul Journal of Medicinal Food tra il 2010 ed il 2012.[1][2] Sono studi su animali, quindi di puro interesse speculativo e non applicabili con validità comprovata sull’uomo, ma rimangono sempre dei lavori di discreto interesse nella ricerca di agenti anti-obesogeni.
Cos’è l’Alpinia officinarum?
L’Alpinia officinarum ( galanga minore) è una pianta appartenente alla famiglia delle Zingiberaceae [3], nativa della Cina, in particolare delle coste del sud-est (Isola di Hainan), sebbene sia comunque presente anche in India e in tutto il Sud-Est asiatico. La pianta è imparentata con lo zenzero e la curcuma ed è un parente stretto della galanga maggiore o Alpinia galangal. Sia Alpinia officinarum che Alpinia galangal hanno una lunga storia di uso medicinale e culinario in oriente.
Sebbene la composizione di entrambe le piante sia molto simile, si trova spesso l’Alpinia officinarum nelle medicine tradizionali asiatiche e l’Alpinia galangal più spesso come spezia.
Per qualunque fine siano usate, le parti più interessanti di entrambe le piante sono le radici.
Dettagli dello studio del 2010
I ricercatori della Zhejiang Chinese Medical University si sono procurati le radici essiccate di Alpinia officinarum e le hanno trasformate in un estratto alcolico. Hanno messo gli estratti ottenuti nel cibo dei ratti da laboratorio per 6 settimane.
I ricercatori hanno diviso i ratti in 4 gruppi. Il primo gruppo ricevette cibo standard. Questo era il gruppo di controllo. Un secondo gruppo ricevette cibo con zuccheri e grassi extra [HFD]. Quei ratti, come ci si potrebbe aspettare, sono ingrassati. Un terzo gruppo venne nutrito con mangime con zuccheri e grassi extra costituiti dal 3% di estratto [AOE], un quarto gruppo venne nutrito con zucchero e una dieta ricca di grassi costituita dal 5% di estratto. L’estratto ha mostrato di bloccare quasi completamente l’aumento della massa grassa nei ratti trattati.
L’alimentazione iper-glucidica e iper-lipidica aveva causato, come d’ovvia intuizione, un peggioramento dei livelli di colesterolo, ma l’estratto di Alpinia officinarum ha praticamente eliminato questo peggioramento [vedi la figura sopra].
I ricercatori hanno affermato che questo studio potrebbe avere importanti implicazioni perché è il primo rapporto che riporti gli effetti anti-obesogeni e di miglioramento della lipidemia ematica dell’estratto di Alpinia officinarum nei ratti nutriti con una dieta ricca di grassi.
Tuttavia, i ricercatori erano perfettamente consapevoli della necessità di ulteriori studi per indagare quali composti nell’estratto di Alpinia officinarum sono responsabili degli effetti osservati, nonché i meccanismi molecolari responsabili dell’attività anti-obesità e ipolipemizzante.
Se calcolassimo la dose per l’uomo da quella usata per i ratti trattati, si ottengono quantità molto elevate, pari a oltre i 10g al giorno. Tale dosaggio non è né sicuro né necessario.
Dettagli dello studio del 2012
Nel 2012, i ricercatori del Korea Food Research Institute hanno effettuato uno studio simile al precedente, ma con un dosaggio somministrato ai ratti meno elevato.
I ricercatori hanno somministrato attraverso il cibo una quantità di estratto di Alpinia officinarum pari a 5g per chilo. Come i ricercatori dello studio cinese, i coreani hanno usato un estratto autoprodotto a base alcolica 1:4 (wt/wt).
Se dovessimo rapportare il dosaggio utilizzato al dosaggio umana, esso sarebbe di circa 3-4g di estratto al giorno. È ancora un dosaggio abbastanza alto, ma decisamente inferiore a quello somministrato agli animali da laboratorio nello studio cinese.
Il design sperimentale dei ricercatori coreani assomigliava a quello dei loro colleghi cinesi. Infatti, i ricercatori hanno somministrato ad un gruppo di topi mangime standard [C], ad un altro gruppo mangime in cui era stato aggiunto zucchero e grasso extra [HFD], e ad un terzo gruppo di animali è stato somministrato cibo ipercalorico più l’estratto di Alpinia officinarum [HFD + AOE].
L’estratto ha inibito l’aumento di peso corporeo degli animali trattati durante le 8 settimane dell’esperimento.
Quando i ricercatori hanno studiato i depositi di grasso dei topi dopo 8 settimane, hanno osservato che Alpinia officinarum aveva ridotto la loro crescita.
Quando i ricercatori hanno effettuato test con cellule adipose in vitro, hanno scoperto che l’estratto di Alpinia officinarum ne bloccava la crescita.
A detta dei ricercatori, la Galangina, un importante flavonolo presente nell’Alpinia officinarum, potrebbe essere responsabile di questo effetto.
Nel tessuto adiposo dei topi trattati, l’Alpinia officinarum aveva causato una parziale disattivazione di recettori attivati da proliferatori perossisomiali come la PPAR-γ, che ostacolava la crescita delle cellule adipose. Nel fegato dei topi, invece, l’Alpinia officinarum aveva causato l’attivazione del PPAR-α, che nei roditori svolge un ruolo importante nell’ossidazione degli acidi grassi.
Conclusione
Come agffermato dagli stessi ricercatori, la prima citata Galangina, può inibire l’adipogenesi durante la differenziazione degli adipociti e può essere efficace nel migliorare lo stato di obesità.
Se la Galangina è davvero il principio attivo cardine nella Alpinia officinarum, non dovrebbe essere difficile testarlo in un numero discreto di soggetti con un dosaggio rivisto e più sicuro. Ma la momento, in mancanza di dati sull’uomo, possiamo solo ipotizzare la sua efficacia nell’uomo.
Come si potrà facilmente capire dalle prime righe di questo articolo, la Rauwolscina è una molecola molto simile nella sua struttura alla Yohimbina e probabilmente condivide simili effetti; ipoteticamente potrebbe essere più potente, ma esistono prove limitate.
Dal momento che mi è capitato molto spesso di ricevere domande su questa forma di Yohimbina, ho deciso di scrivere alcune righe in merio trattando gli effetti maggiormente ricercati dall’utilizzatore medio dell’alcaloide in questione.
Rauwolscina: caratteristiche proprie e similitudini con la Yohimbina
La Rauwolscina, nota anche come Isoyohimbina, α-yohimbina e Corynanthidina, è un alcaloide presente in varie specie del genere Rauvolfia e Pausinystalia (precedentemente noto come Corynanthe).[1] È uno stereoisomero della Yohimbina. [1] la Rauwolscina è uno stimolante del sistema nervoso centrale, un anestetico locale e possiede un certo potenziale afrodisiaco.[1] La Rauwolscina agisce prevalentemente come antagonista del recettore α2-adrenergico.[2][3] È stato anche dimostrato che agisce come agonista parziale del recettore 5-HT1A e antagonista del recettore 5-HT2A e 5-HT2B. [4] [5] [6]
la Rauwolscina, o {3H}Rauwolscina, come già accennato, è un antagonista dei recettori adrenergici alfa-1 e alfa-2 (con una maggiore selettività per quest’ultimo).[7] La Yohimbina condivide il medesimo meccanismo d’azione recettoriale.
Come ormai risaputo, la Yohimbina agisce sul sistema dei recettori adrenergici delle cellule adipose, che regolano la termogenesi. Le subunità beta dei recettori adrenergici (bersagli, per esempio, dell’Efedrina) possono essere viste come stimolanti per la perdita di grasso poiché aumentano l’attività dell’enzima adenil ciclasi e successivamente dei livelli di cAMP (principalmente attraverso le subunità b1 e b2; con la b3 che è meno attiva negli umani).[8][9] Le subunità alfa sono soppressive del metabolismo lipidico, e attraverso la loro attivazione si riduce l’attività dell’adenil ciclasi e si riducono i livelli di cAMP (in particolare per opera degli alfa-2). La Yohimbina, come la Rauwolscina, è un antagonista selettivo del recettore alfa-2 adrenergico (inattivatore), che inibisce l’attivazione del set di recettori soppressivi e preserva l’attività dell’adenil ciclasi e gli effetti mediati dai recettori beta.[10]
Osservando le interazioni della Yohimbina a livello del recettore, la molecola è risultata essere un antagonista alfa2-adrenergico selettivo con un’affinità 44 volte maggiore per la subunità alfa2 rispetto alla subunità alfa1 quando testata su ratti anoccigei e dotti deferenti; questo differisce dal relativo composto Corinantino e Rauwolscina che sono selettivi per il recettore alfa1 (33 volte) e per lo più non selettivi (3,3 volte); rispettivamente.[11] Questi valori sono stati derivati da sperimentazioni in vitro e un secondo test nel quale è stato osservato il legame competitivo in campioni di cervello notando che la selettività era ridotta da 45 a 5,7. Quando si osserva il recettore alfa2 stesso, la Yohimbina sembra avere ulteriore selettività per la subunità alfa2C piuttosto che per la A o la B; nell’intervallo di 4-15 volte la selettività,[12] mentre la Rauwolscina sembra essere non selettiva tra queste tre subunità.[13][12] La Rauwolscina sembra essere efficace a livello del recettore quanto la Yohimbina,[14] con la Coynantina che presenta la minore entità di efficacia.[11]
Oltre a ciò, la stessa Yohimbina può potenzialmente indurre la perdita di grasso per via indiretta attraverso il rilascio di Adrenalina; l’Adrenalina stessa è un attivatore dei recettori beta-adrenergici.[15] Tuttavia, questo aumento di Adrenalina può svanire con il tempo raggiungendo l’irrilevanza statistica 2 settimane dopo l’inizio dell’ingestione giornaliera.[16] L’aumento degli acidi grassi liberi plasmatici e la densità dei recettori alfa2-adrenergici rimangono simili in entrambi i momenti, suggerendo che la Yohimbina perde selettivamente il picco di Adrenalina ma non gli effetti diretti sulla sovra-regolazione della lipolisi recettore-dipendente.
La Yohimbina è stata inizialmente studiata per il suo ruolo nella riduzione degli accumuli adiposi localizzati grazie al suo utilizzo come crema topica (potendo scegliere dove applicare la Yohimbina)[17][18] ma anche secondariamente al suo utilizzo per ridurre l’adipe nelle cosce delle donne, poiché livelli elevati di Estrogeni aumentano l’attività del recettore alfa2-adrenergico.[19] A causa dell’aumento dell’attività alfa2-adrenergica nel tessuto adiposo sito nelle cosce delle donne, si pensava che l’antagonismo di questi recettori riducesse l’adiposità in modo selettivo; i risultati con la soluzione topica sono ad oggi vaghi, con uno studio con esito positivo [18] e l’altro nel quale si sono notati benefici sia con la Forskolina (da Coleus Forskohlii) che con l’Aminofillina ma non con la Yohimbina. [17]
La chetogenesi, o la produzione di corpi chetonici, è potenziata dalla presenza di Noradrenalina in condizioni normali. Il blocco dei recettori alfa adrenergici, tramite l’antagonismo della Yohimbina (e della Rauwolscina) sul recettore alfa-2 adrenergico, aumenta gli effetti chetogenici della Noradrenalina. [20]
Uno studio nel quale si è osservato un aumento della ossidazione lipidica (aumento dei biomarcatori di NEFA e glicerolo) ha anche notato che questo aumento è stato soppresso durante lo stato di alimentazione.[21] È stato ipotizzato che questa interazione con lo stato di digiuno così come gli effetti apparentemente additivi/sinergici della Yohimbina e l’esercizio sulla lipolisi[21] potrebbero portare a tempistiche di assunzione ideale della Yohimbina prima dell’esercizio mattutino.[22] Se abbinata al cibo in modo acuto, la Yohimbina può effettivamente aumentare il rilascio di Insulina indotto dal glucosio attraverso lo stimolo delle cellule pancreatiche[23][24] ma non si verifica a digiuno dopo il consumo orale di 0,2g/kg.[25][21]
Se il meccanismo avviene tramite la stimolazione del rilascio di Insulina che riduce la lipolisi, è teorico che un pasto privo di carboidrati e a basso contenuto proteico (per ridurre la stimolazione indotta dagli amminoacidi) potrebbe essere simile allo stato di digiuno; questo non è stato però studiato.
Uno studio è stato condotto con la somministrazione di Yohimbina a giocatori di calcio d’élite che assumevano 10mg della molecola due volte al giorno (20 mg in totale) per un periodo di 21 giorni. Durante lo studio si è osservato che, in seguito al controllo della dieta, la percentuale di grasso era diminuita dallo 9,3 +/- 1,1% allo 7,1 +/- 2,2% (valutato tramite calibro), mentre nel gruppo placebo è stato registrato un aumento non significativo.[26] La dose di 0,2mg/kg di Yohimbina in uomini altrimenti sani sembra aumentare gli effetti sul miglioramento del metabolismo lipidico e della beta-ossidazione della Noradrenalina endogena e sembra essere più efficace durante i periodi di esercizio e attenuarsi se somministrati dei beta-bloccanti;[21] un altro studio ha rilevato che questa attenuazione deve essere misurata al 70%.[25] Tuttavia, almeno uno studio ha rilevato risultati nulli, in quanto la Yohimbina non ha fatto diminuire il peso nei volontari sani.[27]
La {3H} Rauwolscina è meno potente della Yohimbina nel proteggere dagli aumenti della pressione sanguigna indotti dall’Adrenalina (entrambi meno potenti della Corinantina e più potenti della 3-epi-alfa-yohimbina), questo effetto sembra correlato alla loro affinità per gli alfa -2 adrenorecettori.[28]
Come la Yohimbina, la Rauwolscina è un agonista dei recettori 5-HT1a/b e induce effetti simili alla Serotonina. Mentre la Yohimbina ha più affinità per tale recettore, la Rauwolscina ha un valore IC50 più basso (il che significa che può saturare più recettori alla stessa dose) e può essere considerata leggermente più potente nell’attività serotoninergica.[29][30]
Non mi dilungherò oltre, dal momento che gli argomenti trattati fino a questo momento sono di maggiore interesse per la valutazione di supplementi contenenti Rauwolscina e del loro possibile utilizzo.
Conclusione:
Ora, sappiamo che le caratteristiche di entrambe le forme di Yohimbina sono pressoché identiche sebbene la selettività recettoriale della Rauwolscina sembri minore di quella della Yohimbina. La caratteristica che sembra spostare l’interesse sulla Rauwolscina è la sua più lunga emivita ( Yohimbine 0.25-2.5h; Rauwolscina 0.5-5h ipotetiche). Se però valutiamo quest’aspetto alla luce della sua bassa selettività e alto potenziale di saturazione recettoriale, viene facile intuire che ciò possa influire negativamente sulla percentuale di emersione di effetti collaterali tipici dell’alcaloide (tremori, insonnia, emicrania, tachicardia, ecc…). Con molta probabilità il dosaggio “ideale” con un certo margine di “sicurezza” può essere attestato a 0.1mg/Kg/die.
Gabriel Bellizzi
Riferimenti:
KOHLI JD, DE NN (June 1956). “Pharmacological action of rauwolscine”. Nature. 177 (4521): 1182. doi:10.1038/1771182a0. PMID13334509.
Perry BD, U’Prichard DC (December 1981). “[3H]rauwolscine (alpha-yohimbine): a specific antagonist radioligand for brain alpha 2-adrenergic receptors”. European Journal of Pharmacology. 76 (4): 461–4. doi:10.1016/0014-2999(81)90123-0. PMID6276200.
Arthur JM, Casañas SJ, Raymond JR (June 1993). “Partial agonist properties of rauwolscine and yohimbine for the inhibition of adenylyl cyclase by recombinant human 5-HT1A receptors”. Biochemical Pharmacology. 45 (11): 2337–41. doi:10.1016/0006-2952(93)90208-E. PMID8517875.
Kaumann AJ (June 1983). “Yohimbine and rauwolscine inhibit 5-hydroxytryptamine-induced contraction of large coronary arteries of calf through blockade of 5 HT2 receptors”. Naunyn-Schmiedeberg’s Archives of Pharmacology. 323 (2): 149–54. doi:10.1007/BF00634263. PMID6136920.
DISCLAIMER: Il presente articolo è a solo scopo educativo, di intrattenimento e informativo. Non rappresenta in alcun modo una forma di incitamento all’uso/abuso di sostanze dopanti. L’autore ed il sito, per tanto, è esentato da qualsiasi responsabilità dipendente dalla libera scelta individuale.
Introduzione ad un dilemma…
Chiunque frequenti l’ambiente del Bodybuilding e del Fitness avrà letto o sentito almeno una volta nella vita espressioni del genere “se mi dopassi sarei anche io così [indicando Flex Wheeler]” o “ho provato di tutto e senza farmaci non riuscirò ad ottenere risultati”. Andando poi ad approfondire la storia di ognuno di questi soggetti si scopre in percentuale quasi assoluta che si tratta di individui nella norma (o al di sotto) frustrati e/o con personalità deboli, speranzosi omini che attendono placidamente che accada una svolta miracolosa nella loro banale e piatta esistenza e, cosa molto importante, con il minimo dello sforzo (meglio se nessuno).
Nella mia esperienza come ricercatore e operatore nel campo della cultura fisica in qualità di Preparatore Atletico, ho assistito a innumerevoli casi in cui un soggetto aspirava al miglioramento della propria composizione corporea trascurando, consciamente o inconsciamente, le basi fondamentali rappresentate da Nutrizione e Allenamento baipassandole in vista della possibile prescrizione di una pillola miracolosa capace di renderlo/a possessore della forma fisica ambita.
Tralasciando l’ovvio ragionamento che spinge ogni essere umano dotato di un minimo d’intelletto verso la comprensione che la genetica è il blocco d’argilla sul quale si va ad operare, ma le sue qualità e difetti sono presenti in modo eterogeneo nella popolazione mondiale, e ciò non è modificabile nemmeno con la farmacologia più oculata, quando ci si trova davanti al bivio tra “pillola rossa” (PEDs) e “pillola blu” (drug free) bisogna essere pienamente consapevoli non solo del fattore illegalità ma del fattore conoscitivo. Purtroppo, la politica del terrore ha operato in modo fallimentare nel goffo intento di allontanare dalla scelta “rossa”, e ciò si è tradotto in un numero sensibile di soggetti abusatori con tutte le conseguenze cliniche derivanti.
Se un individuo non ha raggiunto un livello di maturità sportiva tale da conferirgli una gestione corretta della nutrizione e della periodizzazione allenante (gestione delle variabili volume, intensità, densità ecc…), è molto meglio per lui/lei rivedere i suoi programmi e scegliere ancora la “pillola blu”. Capita, a volte, di incontrare persone decise ad intraprendere la via del “lato oscuro” che, dopo una approfondita chiacchierata sulla gestione dei suddetti fattori, rivede le proprie posizioni.
Per tutti coloro i quali sono immersi nel dilemma della scelta, vi espongo alcuni punti per rendere l’eventuale decisione meno rischiosa anche se pur sempre illegale nel “bel paese”…
“Pillola blu o pillola rossa?” I punti da tenere in considerazione per una scelta consapevole:
#1 Raggiungere una adeguata maturità sportiva
Per “maturità sportiva”, in particolare riferimento al BodyBuilding, si intende la capacità del atleta di sapersi alimentare e allenare correttamente con piena gestione delle proprie potenzialità fisiologiche/genetiche. Questa è la base, se viene a mancare ciò non solo la vostra esperienza finirà per deludervi e rendervi ancora di più dei frustrati, ma potrebbe rovinosamente portarvi ad un abuso cronico a senso inesorabilmente negativo…
#2 I PEDs non faranno miracoli
Una cosa da tenere bene a mente, e questo non dovrebbe interessare solo gli aspiranti “doped”, è che l’uso di PEDs non renderà diversi da ciò che rientra nelle potenzialità espressive del proprio patrimonio genetico. Certamente le caratteristiche genetiche verranno “iperespresse”, nel bene e nel male, dall’uso di PEDs ma non vi sarà nessun miracolo! Migliorerete ma non sarete ne più ne meno di ciò che potete essere!
Un esempio per capire come la base genetica faccia la differenza anche con protocolli che, ad oggi, spesso non raggiungono nemmeno i livelli del “bridge” più soft..
#3 Ridurre la percentuale di grasso corporeo
Il tessuto adiposo rappresenta uno dei siti dove il Testosterone, ed altri AAS soggetti all’aromatizzazione, viene convertito in Estradiolo. Soggetti con percentuali di grasso corporeo elevate vedrebbero una alterazione marcata della Testosterone:Estradiolo ratio a favore della componente estrogenica, con conseguenze quali alterazione del comportamento sessuale (impotenza, difficoltà nel raggiungere e/o mantenere l’erezione), ritenzione idrica, accumulo di grasso con modello femminile e ginecomastia. E no, l’uso di DHT derivati o di SARM non steroidei senza una base di Testosterone non risolverebbe il problema o, per lo meno, porterebbe ad altre conseguenze negative, che pur non comprendendo, per esempio, ritenzione idrica e ginecomastia, interesserebbero l’attività sessuale e la condizione psichica del soggetto trattato. [1]
Schema esemplificato del processo di aromatizzazione degli androgeni aumentati in un soggetto con percentuale di grasso corporeo alta.
Allo stesso tempo, i rischi cardiovascolari della somministrazione di AAS- come il possibile aumento esponenziale del Ematocrito, l’aumento del LDL e Trigliceridi a discapito di una riduzione del HDL, e l’aumento della pressione sanguigna – sarebbero già presenti in certa misura quando la body fat è già alta e sarebbero quindi soggetti ad un repentino aggravamento.
Se la percentuale di grasso è relativamente alta, si dovrebbe prima di tutto considerare di migliorare la composizione corporea con una adeguata routine alimentare e allenante (senza farmaci) prima di iniziare solo a pensare all’uso di AAS. Sicuramente ciò renderà la scelta più efficace e meno rischiosa.
Nel caso fosse necessario sottolinearlo, no, non è saggio nemmeno utilizzare agenti PEDs a fini lipolitici e/o antiadipogenici e/o termogenici (compresi gli Ormoni Tiroidei). A meno che non siate affetti da ipotiroidismo, e in questo caso la terapia vi dovrebbe essere stilata dal vostro medico, per ridurre in modo sensibile la body fat non sono necessari i farmaci!
#4 Controllare se si ha una storia familiare di trombosi (o qualsiasi altra malattia cardiovascolare)
Molte malattie cardiovascolari hanno una componente di base genetica. Uno stile di vita sano può ridurne sensibilmente la loro insorgenza, ma l’uso di AAS può causare l’attivazione di specifici geni implicati nella comparsa di malattie cardio-circolatorie. Caratteristico dell’interazione tra AAS e geni specifici è un caso studio ben documentato che ricercatori americani hanno pubblicato sul “Blood Coagulation & Fibrinolysis”.[2]
Trombosi venosa
Oltre all’attivazione genica diretta dagli AAS, e nociva per il sistema cardio-circolatorio, vi sono altre condizioni negative innescate dall’uso/abuso di Steroidi Anabolizzanti, e di altri PEDs, come, per esempio, l’aumento del tasso di coagulazione, l’incremento eccessivo dell’Ematocrito con aumento pressorio, rigidità dell’endotelio vascolare con perdita di efficienza strutturale e aumento della pressione ematica con incremento delle possibilità di danno strutturale dei componenti del sistema interessato.
#5 Inserire delle sedute di allenamento cardio prima, durante e dopo l’uso di PEDs
Un moderato allenamento cardiovascolare è sicuramente una delle migliori strategie preventive contro la comparsa di malattie cardio-circolatorie. Tale tipologia di allenamento può portare un miglioramento e/o riduzione delle alterazioni lipidiche ematiche del praticante, fornendo un, seppur minimo, tampone all’azione negativa degli AAS e SARM non steroidei sui livelli di LDL (aumento), Trigliceridi (aumento) e HDL (diminuzione). Secondo quanto riportato da una interessante review del 2013, l’abbinamento di sedute cardio e in sala pesi possono avere una azione additiva benefica sui livelli di LDL, Trigliceridi e HDL.[3]
Risulta interessante anche quanto emerso da alcuni studi su animali a seguito dei quali si è osservato un significativo grado di protezione dato dall’allenamento cardio negli esemplari trattati con AAS.[4]
#6 Assicurarsi di rimanere ben idratati
Oltre ad agevolare il mantenimento di un Ematocrito migliore, una buona idratazione risulta positiva sulla pressione di lavoro renale nel filtraggio del sangue. Diversi AAS come il Trenbolone e i metilati in C-17 presentano una particolare resistenza metabolica che, oltre a causare un aumentato stress epatico, può portare ad una sofferenza renale sfociabile nel patologico. Si è osservato come una combinazione di AAS, dieta iperproteica e supplementazione di Creatina possa aumentare l’incidenza di problemi renali.[5] In un soggetto in fisiologia, la sola dieta ad altro contenuto proteico e la supplementazione di Creatina non hanno mostrato nessun grado di pericolosità, soprattutto sul breve/medio termine.
#7 Non usare “droghe ricreative”
A livello globale, il numero di decessi tra gli abusatori di AAS è in aumento. Alcuni, troppo superficialmente, dicono che questo sia dovuto al fatto che sempre più uomini e donne usano AAS, ma questa è solo una spiegazione dozzinale e limitata. Il sospetto ricade soprattutto sulle modalità di approccio dei consumatori di AAS: i dosaggi sono drammaticamente aumentati e un numero crescente di individui combina PEDs con “droghe ricreative”. Ed è su questi due ultimi punti che risiede la spiegazione principale dell’aumento statistico prima menzionato. Soprattutto la combinazione di PEDs e le così dette “droghe ricreative” risulta essere probabilmente un fattore significativo, come evidenziato alcuni anni fa da ricercatori australiani. Nel loro studio sono state analizzate tutte le morti documentate tra i consumatori di AAS a Sydney tra il 1997 e il 2012, scoprendo che le droghe ricreative come la cocaina avevano avuto un ruolo nella schiacciante molteplicità dei casi. Dagli studi sugli animali ora sappiamo della possibilità che la co-assunzione di un AAS come il Nandrolone con la cocaina vede moltiplicati gli effetti cardiotossici rispetto ai singoli composti.[6] E secondo studi in vitro la combinazione di Testosterone e cocaina aumenterebbe la possibilità di formazione di coaguli nel flusso ematico. [7]
#8 Corretta modalità di iniezione e herpes labiale
Gli utilizzatori di AAS a volte sviluppano ascessi, ma non sempre dovuti alla bassa qualità dei prodotti utilizzati.
Alcuni medici ritengono che gli utilizzatori di AAS dovrebbero effettivamente ricevere una formazione sulle tecniche di iniezione corrette, onde evitare embolie oleose o ascessi per cattiva gestione igienica della procedura. [8]
Molti utilizzatori ancora non sanno che disturbi come l’herpes labiale rendono le iniezioni ancora più rischiose. Perchè? Il virus che causa l’herpes labiale, come altri patogeni, riduce l’efficienza del sistema immunitario, fornendo così terreno fertile per infezioni batteriche i cui microorganismi scatenanti vengono inoculati nel corpo del soggetto attraverso l’iniezione in modo diretto o indiretto.
#9 Non fare affidamento sugli integratori
Secondo un buon numero di studi svolti su animali, alcuni integratori proteggono dagli effetti collaterali degli AAS. Secondo alcune ricerche, la Taurina, la Vitamina C ed E proteggono i testicoli durante un ciclo e la vitamina C e il cacao proteggono la prostata.
L’utilità dei risultati provenienti da questi studi è limitata per tre motivi:
A. gli animali da laboratorio non sono esseri umani, e
B. le dosi utilizzate e rapportate ad un essere umano sono quasi sempre molto inferiori rispetto a quelle utilizzate dai “doped”, e
C. la ricerca in campo psicologico mostra che l’uso di integratori stimola comportamenti rischiosi e malsani. I supplementi fanno pensare agli utilizzatori di essere invulnerabili e di non dover comportarsi in modo sano ed attento.[9]
Gli integratori possono aiutare a creare una mentalità che non si dovrebbe avere da utilizzatore consapevole di AAS.
Ovviamente, alcuni supplementi “protettivi” utilizzati dai soggetti meglio informati hanno un potenziale di “tamponare” in modo discreto alcune alterazioni legate all’uso di AAS e SARM come, ma non limitato a, Riso Rosso fermantato (controllo lipidico) [10], Silimarina (epatoprotezione), NAC (epatoprotezione) [11], Niacina (controllo lipidico) ecc…
#10 Ridurre al minimo (se non eliminare) il consumo di alcolici
Potrebbe sembrare un indicazione superflua ma non lo è.
L’abuso di alcol è indubbiamente uno dei problemi sociali più diffusi. Uno dei problemi correlati all’abuso di alcol e l’epatopatia alcolica. Questo stato patologico è derivante da un processo infiammatorio progressivo ai danni del fegato legato al consumo eccessivo di alcolici. È una malattia a più stadi. La steatosi provoca un ingrossamento del fegato causato da un accumulo di trigliceridi, spesso senza sintomi per molto tempo. I rischi correlati sono la steatosi (fegato grasso), l’epatite alcolica e la cirrosi epatica. Il rapporto con l’alcolismo è complesso. Non tutti i bevitori, infatti, hanno danni al fegato, anche se sono altamente probabili. La causa è da rinvenire in una trasformazione dell’alcol (etanolo) in sostanze tossiche che danneggiano il fegato in maniera irreversibile e cronica, con un rischio elevato di insufficienza epatica e di cancro, fino alla necessità di un trapianto di fegato.
In acuto, invece, l’alcol può essere una causa di alterazione delle transaminasi ma non si può sapere se e con quale modalità si potrebbero innalzare: dipende molto dalla risposta individuale dell’organismo. In caso di stress preesistente, di causa iatrogena e/o alimentare, si può presentare una alterazione significativa. [12]
Il primo caso è una consequenziale possibile se eventi stressori concomitanti si presentano in cronico. Ed è semplice giungere alla conclusione che l’uso di AAS, specie se metilati, possa comportare un aumentato stress epatico che potrebbe degenerare in peliosi epatica, cirrosi ecc…
Che siate “doped” o “natural”, per ragioni legate e non, dovreste evitare di consumare più di 25g per gli uomini, o 12,5g per le donne, di Etanolo al giorno.
#11 Sottoporsi a regolari controlli medici pre, intra e post utilizzo
Il monitoraggio della salute dovrebbe essere la base fondante del comportamento del utilizzatore consapevole e minimamente attento ai potenziali rischi nei quali potrebbe imbattersi.
Gli esami di controllo sono i seguenti:
Esami ematici e delle urine (comprendenti il quadro ormonale secondo necessità);
Elettrocardiogramma ogni 6 mesi circa;
Elettrocardiogramma sotto sforzo (prima di iniziare);
Ecocardiogramma ogni 6 mesi circa;
Coronarografia ogni 6 mesi circa;
Monitoraggio della pressione ematica;
TAC addome completa ogni 6 mesi circa.
Ovviamente, ogni accertamento , al di la degli esami ematici, deve essere gestito in base alle esigenze soggettive, caratteristiche e tipo di PEDs utilizzati.
#12 Essere seguiti da personale qualificato
Fin troppa gente è stata salutisticamente deturpata da gorilla di spogliatoio a mala pena consapevoli dell’esistenza dei macronutrienti e che, nonostante ciò, si sono improvvisati farmacisti. Donne divenuti uomini e uomini divenuti simili a cagne in calore per via di orrende ginecomastie. Evitate il fai da te e l’affidarsi a semianalfabeti … la somaticità sopra la norma è cosa diversa dall’intelligenza e alla competenza in biologia, biochimica e farmacologia… senza offesa per tutti quelli che “io mi facevo e ho vinto! Senzia scienzia!” …
#13 Pensare seriamente al post ciclo prima del ciclo
Molti aspiranti “doped” non considerano il fattore post ciclo. La maggior parte di loro è convinta che la PCT sarà una facile soluzione alla sottoregolazione dell’Asse HPT, ma in realtà non è proprio così. Esistono diversi casi studio che mostrano come gli ex utilizzatori abbiano spesso livelli di Testosterone inferiori rispetto al pre-utilizzo anche a distanza di anni dal cessato uso di AAS. Sembra che i fattori che aumentano le possibilità e il grado di tale effetto sul lungo termine siano:
Tempo di somministrazione;
Età
Molecole utilizzate (con maggiore impatto negativo dato dai19-norsteroidi come il Nandrolone per via della lunga permanenza dei metaboliti nel sistema).
Tutto ciò è indipendente dalla qualità della PCT, anche se essa può avere dei riscontri positivi specie nel primo periodo di stacco dagli AAS. Le alterazioni ormonali legate ad una alterazione dell’Asse HPT comprendono depressione, ansia, bassa libido, difficoltà nel raggiungere e mantenere l’erezione, stanchezza cronica ecc…
Per questa ragione molti scelgono di entrare in TRT (Terapia Sostitutiva del Testosterone) dopo il primo ciclo.
Quale conclusione?…
Se mai non dovesse bastare il disclaimer, questo articolo non rappresenta in alcun modo un consiglio e, ne tanto meno, un incitamento all’uso di sostanze dopanti! E’ semplicemente a fine divulgativo con l’obbiettivo di far comprendere a più persone possibili che la scelta di intraprendere coscientemente certe pratiche (illegali) necessita di una sufficiente (e veritiera) conoscenza del argomento.
Quindi? Leggete e comprendete correttamente ciò che ho riportato in sintesi fruibile ad un largo pubblico… Pensate prima di tutto ad alimentarvi e allenarvi in modo ottimale!
La conoscenza della Verità rende liberi dalla cattiva informazione, dagli strumenti commerciali e dal relativismo… Negarla è semplice e pericolosa manifestazione di profonda ignoranza… di VERO NEGAZIONISMO!
Se avete una buona conoscenza della lingua inglese e volete approfondire l’argomento PEDs e Sport, potete leggere il libro ANABOLICS 11th Edition di William Llewellyn
“Magnitude and Composition of the Energy Surplus for Maximizing Muscle Hypertrophy: Implications for Bodybuilding and Physique Athlete” [1] è una interessante review di Alan Aragon e Brad Schoenfeld nella quale viene esaminata .*la letteratura scientifica ad oggi disponibile per delineare il ruolo dell’assunzione calorica nell’ipertrofia muscolare. In particolare, questa review è rivolta ai BodyBuilder e atleti Physique.
La review è suddivisa in sezioni che coprono i seguenti argomenti connessi al tema centrale:
Condizioni ipocaloriche (deficit energetico)
Condizioni eucaloriche (un apporto energetico pari al dispendio energetico)
Condizioni ipercaloriche (un surplus energetico)
Entità del surplus
Composizione dei macronutrienti
Applicazione pratica.
Nel presente articolo evidenzierò i punti importanti di ciascuna di queste sezioni.
Condizioni ipocaloriche (deficit energetico)
È noto che un deficit energetico ostacola la crescita muscolare. In effetti, può compromettere completamente la crescita muscolare negli individui allenati (specialmente quando sono già con percentuali di grasso ridotte). Meccanicamente, questo potrebbe essere spiegato dalla sottoregolazione del percorso mTORC1 e dall’attivazione della proteina del sensore di energia AMPK.
La regolazione del mTORC1 in base allo stato energetico della cellula è descritta meno bene di quella dei fattori di crescita e appare principalmente mediata dalla chinasi attivata da AMP (AMPK). AMPK è una proteina eterotrimerica che comprende una combinazione di subunità α, β e γ. Attualmente sono note due isoforme delle subunità α (α1 e α2) e β (β1, β2). Esistono tre isoforme note della subunità γ (γ1, γ2, γ3). La subunità α funziona come la subunità catalitica del complesso, mentre le altre due subunità “rilevano” lo stato energetico della cellula. La subunità β può interagire con il glicogeno e la subunità γ con i nucleotidi adenosintrifosfato (ATP), adenosindifosfato (ADP) e adenosinmonofosfato (AMP). L’interazione tra il glicogeno e la subunità β porta all’inibizione allosterica dell’attività dell’AMPK, una diminuzione del glicogeno porterà quindi alla riduzione di questa inibizione e quindi all’attivazione del complesso. In sintesi, le subunità β e γ consentono alla chinasi di misurare lo stato energetico della cellula come riflesso dal suo contenuto di glicogeno e dal rapporto ATP/ADP o AMP. Una diminuzione del glicogeno o del rapporto ATP/ADP o AMP segnala una diminuzione dell’energia disponibile alla chinasi e la attiva. In generale, l’attivazione di AMPK promuove le vie cataboliche al fine di recuperare l’omeostasi energetica cellulare e attenua le vie anaboliche per preservare l’energia.
Teoricamente, le diverse isoforme delle subunità consentono dodici combinazioni uniche. Tuttavia, fino ad oggi sono state trovate solo tre diverse combinazioni nel muscolo scheletrico umano: α2 / β2 / γ1, α2 / β2 / γ3 e α1 / β2 / γ1. La distribuzione quantitativa di queste proteine eterotrimeriche è stata stimata al 15% α1 / β2 / γ1, 65% α2 / β2 / γ1 e 20% α2 / β2 / γ3. I tre eterotrimeri mostrano regolazione ed effetti differenziali. L’eterotrimero α2 / β2 / γ3 si attiva rapidamente a seguito dell’attività fisica, mentre gli altri due impiegano molto più tempo per attivarsi. Inoltre, solo l’eterotrimero contenente α1 sembra attenuare la crescita muscolare, mentre gli eterotrimeri contenenti α2 non sembrano farlo.
L’effetto antagonizzante di AMPK sulla crescita muscolare è mediato, almeno in parte, dall’inibizione dell’attività del mTORC1. AMPK fosforila due residui (Thr1227 e Ser1345) su TSC2 che sono importanti per la sua attivazione. TSC2 agisce quindi inibendo mTORC1 mediante la formazione del complesso TSC-TBC come descritto in precedenza. Inoltre, è stato scoperto che Raptor, una delle proteine che compromettono mTORC1, è anche un substrato del AMPK. Anche la fosforilazione di Raptor ai residui Ser722 e Ser792 inibisce l’attività di mTORC1. In sintesi, l’effetto antagonista del AMPK su mTOR è mediato dalla fosforilazione di TSC2 e Raptor.[2]
Detto ciò, quello che viene evidenziato in questa sezione, è che l’allenamento contro-resistenza in concomitanza con un elevato apporto di proteine può aumentare la massa magra nonostante un deficit energetico. Sebbene questo effetto sia chiaramente diminuito negli individui con percentuali di massa grassa basse allenati, ciò è particolarmente vero per gli individui obesi non allenati. Uno studio interessante che evidenzia questo punto è quello di Donnelly et al. [3].
Nello studio, quattordici donne obese sono state sottoposte a una dieta ipocalorica (803 kcal al giorno) per 90 giorni. Sette di loro hanno eseguito simultaneamente l’allenamento con i pesi e gli altri sette sono rimasti sedentari. Come ci si aspetterebbe con un apporto energetico così basso negli individui obesi, hanno perso molto peso. In media, hanno perso 16Kg, ovvero 35 libbre. Mentre la massa grassa e la massa magra sono diminuite nella stessa misura in entrambi i gruppi, le biopsie muscolari hanno fornito alcune informazioni interessanti. Hanno dimostrato che l’area della sezione trasversale delle fibre muscolari è aumentata in coloro che hanno eseguito l’allenamento con i pesi, ma è rimasta invariata in coloro che sono rimasti sedentari. Mentre l’ipertrofia potrebbe essere stata solo regionale per quel muscolo specifico, non era così scontato (sicuramente dato che è un muscolo della gamba ei soggetti hanno perso molto peso corporeo). Va notato che una certa perdita di massa magra non può essere attribuita alla perdita muscolare. (E allo stesso modo, se misuri un aumento della massa magra, non dovresti attribuirlo direttamente alla massa muscolare.)
Condizioni eucaloriche (un apporto energetico pari al dispendio energetico)
Quando l’apporto energetico è uguale al dispendio energetico, si parla di eucalorica. In altre parole, si sta mangiando per mantenere il livello di energia in equilibrio. Queste condizioni sono più favorevoli per la costruzione muscolare.
L’articolo sembra principalmente partire da una recente meta-analisi di Morton et al. [4]. (Aragon e Schoenfeld erano entrambi coautori di questa meta-analisi.) Gli studi che coinvolgono condizioni ipocaloriche sono stati esclusi dall’analisi. E’ stato esaminato l’effetto dell’integrazione proteica in combinazione con l’esercizio contro-resistenza sull’aumento della massa muscolare.
Sostengono che a causa della mancanza di un aumento significativo dell’assunzione di energia rispetto al basale negli studi analizzati, nonché di guadagni relativamente minori nella FFM, le condizioni possono essere interpretate come eucaloriche (Poiché questo non era uno scopo mirato dello studio.). Hanno scoperto che l’integrazione di proteine ha aumentato la massa magra di 0,75kg in media negli individui allenati contro-resistenza.
Per dare una maggiore prospettiva: gli studi con persone allenate sono durati in media 10 settimane. Onestamente, è abbastanza per un cambiamento dietetico relativamente minore in un lasso di tempo relativamente breve. Basta aggiungere un po’ di proteine in più. I guadagni sono stati complessivamente molto inferiori (0,3 kg) o inesistenti (-0,01 kg) negli anziani. Aragon e Schoenfeld concludono questa sezione con “Questi guadagni relativamente insignificanti indicano l’importanza di un surplus calorico mirato combinato con un allenamento contro-resistenza progressivo quando l’obiettivo è massimizzare l’ipertrofia muscolare”.
Non posso dire di essere pienamente d’accordo con quanto affermato, dal momento che trovo l’aumento abbastanza solido negli individui allenati contro-resistenza per un cambiamento dietetico così limitato. Anche se questo potrebbe non essere ottimale, penso che il messaggio da portare a casa per gli individui allenati contro-resistenza possa essere che è possibile guadagnare una quantità discreta di massa muscolare in condizioni eucaloriche quando l’assunzione di proteine è adeguata.
Condizioni ipercaloriche (surplus energetico)
È ovvio che un surplus di energia consente una maggiore ipertrofia muscolare. Uno studio interessante che menzionano è quello di Bray et al. [5]. Questo studio è stato svolto particolarmente bene. La ragione principale di ciò è che i soggetti sono stati messi in un’unità metabolica, potendo così controllare completamente il loro apporto energetico.
Lo studio ha esaminato 25 soggetti di ambo i sessi sani e stabili di peso che sono stati sovralimentati per 8 settimane dopo una dieta di stabilizzazione del peso iniziale di 13-25 giorni. Sono stati randomizzati con una dieta a basso contenuto di proteine (5% dell’apporto energetico), proteine normali (15%) o ad alto contenuto proteico (25%). L’apporto energetico è stato aumentato di quasi 1000 kcal al giorno.
La cosa interessante è che tutti e tre i gruppi hanno guadagnato circa la stessa quantità di grasso corporeo. Tuttavia, i gruppi con una dieta normale e ricca di proteine hanno anche guadagnato massa corporea magra (misurata con DXA). Quindi solo mangiare molte più calorie, mentre si assumono abbastanza proteine, ovviamente, provoca un aumento della massa corporea magra.
Aragon & Schoenfeld continuano a sottolineare che è importante rendersi conto che la massa magra non è uguale alla massa muscolare. Cosa su cui non si può essere che pienamente d’accordo.
Questo fattore è importante quando si interpreta la ricerca. Misurazioni aggiuntive, come ultrasuoni, risonanza magnetica, TC o area della sezione trasversale delle fibre muscolari, possono dare molta più concretezza nel fatto che un cambiamento nella massa magra sia dovuto o meno a un cambiamento nella massa muscolare.
Entità del surplus energetico
Proprio come l’assunzione energetica può influenzare l’ipertrofia muscolare, così lo possono fare l’allenamento e la composizione corporea. Aragon e Schoenfeld sostengono che gli individui non allenati (deallenati) hanno un potenziale maggiore per realizzare guadagni muscolari, e la pratica lo ha dimostrato diverse volte. Pertanto è possibile che i soggetti magri e allenati siano più inclini a guadagnare grasso con un surplus energetico eccessivamente elevato. Ed anche questo è un qualcosa di verificato molte volte nella pratica.
In generale, consiglio agli atleti più avanzati, con una bassa body fat, così come per le donne (a causa del loro potenziale di crescita relativamente ridotto) un surplus energetico inferiore rispetto ai principianti che desiderano aumentare la loro massa muscolare il più possibile.
Indubbiamente, gli atleti più avanzati guadagneranno anche un po’ più di massa muscolare con grandi eccedenze di energia. Tuttavia, quando ricominciano a ridurre le Kcal in seguito, hanno bisogno di perdere più grasso e immagino che qualsiasi massa muscolare in più guadagnata con fatica verrà persa con esso in percentuali sensibili.
Uno altro studio che menzionano è quello di Rozenek et al. [6]. I soggetti del test hanno ricevuto un surplus energetico di 2010Kcal in aggiunta alla loro dieta di mantenimento.
I soggetti presi in esame sono stati divisi in tre gruppi. Uno ha ricevuto un supplemento di 2010 kcal ad alto contenuto proteico (356g di carboidrati, 106g di proteine), uno ha ricevuto un integratore isocalorici di carboidrati da 2010 kcal e uno è servito come gruppo di controllo. Tutti i soggetti dei tre gruppi hanno eseguito un allenamento contro-resistenza con un ragionevole apporto di proteine (almeno 1,3-1,5 g/kg di peso corporeo al giorno). Alla fine delle 8 settimane di studio, tutti i gruppi avevano acquisito quantità significative di massa magra (2,9, 3,4 e 1,4 kg, rispettivamente). Ciò suggerisce che, almeno negli “allenati principianti”, un surplus calorico può aumentare sostanzialmente i guadagni di massa magra quando l’assunzione di proteine è sufficiente. Tuttavia, come sottolineano Aragon e Schoenfeld, questi risultati dovrebbero essere interpretati con cautela. Entrambi i gruppi che hanno ricevuto il supplemento hanno aumentato notevolmente la loro assunzione di carboidrati. Questo porterà sicuramente ad altro glicogeno e acqua di accompagnamento (Che può essere misurata come massa magra.).
Ora confrontate questi risultati con un altro studio menzionato, ma con bodybuilder maschi d’élite.[6] Un piccolo gruppo di soggetti è stato randomizzato a un elevato apporto energetico (6087 kcal al giorno) o ad un moderato apporto energetico (4501 kcal al giorno) per 4 settimane. L’elevato apporto energetico ha portato ad un aumento maggiore (1,0 kg) della massa magra rispetto al moderato apporto energetico (0,4 kg). Tuttavia, non c’è stato alcun cambiamento significativo nella massa grassa nel gruppo con surplus di energia moderato (+0,8%), mentre il gruppo ad alto apporto di energia ha visto un aumento significativo del 7,4%. Va notato che le misurazioni del grasso corporeo sono state effettuate utilizzando un calibro, quindi è stata utilizzata una formula per ricavare la massa magra. Dubito che sia accurata, soprattutto in un arco di tempo così breve. Ad ogni modo, questi risultati potrebbero suggerire che individui molto ben allenati per lo più ingrassano solo con grandi eccedenze caloriche.
Composizione dei macronutrienti
Immagino che questa sezione possa essere riassunta in poche frasi. Se si consuma una quantità sufficiente di proteine (da 1,6 a 2,2g/kg di peso corporeo al giorno), mangiare troppo forse potrà aiutare un po’ di più con l’ipertrofia muscolare. Inoltre, potrebbe forse avere un effetto sul portare il livello di grasso corporeo fuori da range accettabili (12-15%). L’eccesso di grassi probabilmente non è l’idea migliore, quindi i carboidrati sembrano essere il macronutriente di scelta primaria quando si punta a surplus calorici elevati.
Applicazione pratica
Gabriel Bellizzi
Riferimenti:
1- Aragon AA, Schoenfeld BJ. Magnitude and Composition of the Energy Surplus for Maximizing Muscle Hypertrophy: Implications for Bodybuilding and Physique Athletes. Strength & Conditioning Journal. Publish Ahead of Print, 2020.
3- Donnelly JE, Sharp T, Houmard J, et al. Muscle hypertrophy with large-scale weight loss and resistance training. Am J Clin Nutr 58: 561–565, 1993.
4- Morton, Robert W., et al. “A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults.” Br J Sports Med 52.6 (2018): 376-384.
5- Rozenek, R., et al. “Effects of high-calorie supplements on body composition and muscular strength following resistance training.” Journal of Sports Medicine and Physical Fitness 42.3 (2002): 340-347.
6- Ribeiro, Alex S., et al. “Effects of different dietary energy intake following resistance training on muscle mass and body fat in bodybuilders: a pilot study.” Journal of Human Kinetics 70.1 (2019): 125-134.
Con l’invecchiamento, sia gli uomini che le donne, oltre alla perdita di massa muscolare (sarcopenia) devono far fronte alla perdita della densità minerale ossea (osteopenia), condizione che si aggrava in modo significativo dopo i quarant’anni. Il ritmo della degradazione ossea e di danno ad esso correlato aumenta con l’età. L’allenamento contro-resistenza può rallentare il processo. E anche l’uso della Creatina sembra poter coadiuvare il processo protettivo aumentandolo, come osservato e riportato dai ricercatori della University of Saskatchewan in Canada.[1]
Dettagli dello studio
I ricercatori hanno svolto il loro studio utilizzando diciotto donne in post-menopausa e dividendole in due gruppi. L’età media dei soggetti osservati era di 57 anni.
Le donne reclutate per lo studio si allenavano tre volte a settimana in sala pesi, allenando i loro gruppi muscolari più grandi eseguendo esercizi base come Hack Squat, Hamstring Curl, Back Extension, Bench Press, Lat Pull-Downs e Shoulder Press. Le donne hanno eseguito 3 serie da 10 ripetizioni per ogni esercizio, con un carico dell’80% del 1RM. A un gruppo è stata somministrata Creatina, ad una dose di 0,1g per kg di peso corporeo al giorno. L’altro gruppo ha ricevuto un placebo. Prima dell’inizio dell’esperimento e dopo che le donne si erano allenate per un anno, i ricercatori hanno eseguito delle scansioni per misurare la densità ossea dei soggetti.
Risultati dello studio
La diminuzione della massa ossea nel collo del femore delle donne nel gruppo trattato con Creatina si era quasi arrestata. Nelle donne che non avevano assunto Creatina la degradazione ossea è continuata. Questa è una scoperta rilevante perché le persone con osteoporosi spesso incorrono nella rottura del collo del femore quando si lesionano all’anca. La guarigione è spesso lenta o non avviene.
Conclusioni
L’effetto della matrice ossea nel collo del femore è stato l’unico effetto benefico statisticamente significativo sullo scheletro osservato dai ricercatori.
L’aumento della forza muscolare è stato modesto in tutte i soggetti e le donne in entrambi i gruppi hanno perso massa magra. Allenarsi tre volte a settimana per un anno e perdere massa magra? Probabilmente, i soggetti dello studio potrebbero non essersi allenati molto duramente e/o non hanno seguito una dieta adeguata (è probabile una co-fattorialità).
E’ comunque possibile che l’effetto positivo dell’allenamento contro-resistenza sulla massa ossea – con o senza Creatina – sia maggiore di quanto suggerito da questo studio.