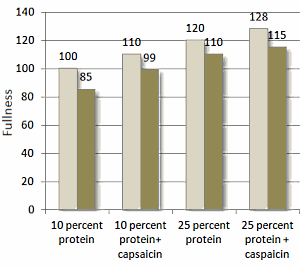Autore: Gabriel Bellizzi [also known as Ružička, The Biochemist] - CEO BioGenTech -
Negli anni trenta del ventesimo secolo si è verificata una febbre dell’oro scientifica di proporzioni inaudite nel campo della nascente endocrinologia. Questa impresa è stata portata avanti con tanta celerità grazie al pionieristico lavoro di biochimici Adolf Friedrich Johann Butenandt e Lavoslav Stjepan Ružička, entrambi premi Nobel per la chimica nel 1939 grazie proprio alla pubblicazione dell’articolo “Sulla preparazione artificiale dell’ormone testicolare testosterone (androstene-3-one--17-olio)”.
Il potenziale del Testosterone e dei suoi primi derivati che videro la luce nella seconda metà degli anni trenta del 900, arrivo’ all’orecchio degli sportivi d’élite tanto che nel 1938 vi fu una prima pubblicazione che parlava del potenziale uso del Testosterone nel Bodybuilding.
Grazie agli abbattimenti dei costi di produzione delle molecole di sintesi, resi possibili dal genio della chimica Russell Earl Marker e dalla sua “Marker degradation”, nella seconda metà degli anni quaranta l’uso di AAS si è diffuso nelle squadre olimpiche di molti paesi. Successivamente tocco’ al pubblico amatoriale. E' nel 1976 che vi fu una nuova svolta, cioè la nascita della società di biotecnologie “Genetech” nata dall’incontro tra l’imprenditore Robert Swanson e Herbert Boyer, biochimico dell’Università della California. I due decisero di fondare questa società per lo sfruttamento commerciale delle tecniche del DNA ricombinante messe a punto da Boyer. Insulina e hGH divennero parte del corollario di farmaci utilizzati dai bodybuilder, e l’era dei “Freak” venne inaugurata.
Purtroppo, lo “scandalo DOPING” negli anni 80’, e le successive restrizioni di “facciata” hanno smantellato massivamente quella nicchia di ricercatori che lavoravano a stretto contatto con gli atleti e facevano ricerca sul campo. Essi non sono “estinti” ma sono obliati da una certa narrativa di comodo. Da qui il problema presente: l’atleta è in balia di leggende e metodiche partorite da menti non avvezze alla complessità della farmacologia partendo dalle basi della biochimica.
La BioGenTech è un laboratorio di ricerca che opera direttamente sul campo dapprima della sua fondazione grazie al lavoro del CEO Amedeo Gabriel Bellizzi. Nel 2021, ha visto la luce e ha preso concretezza un idea: fornire informazioni valide e affidabili su una scienza multidisciplinare. Nessun circo delle pulci, ma qualcosa che si può vedere e constatare.
Noi alla BioGenTech, la quale è una realtà collaborativa sebbene diretta da una mente, siamo scienziati puri con un atteggiamento snobistico nei confronti dei soli affari. Riteniamo la sola corsa al denaro una cosa da bottegai, poco stimolante dal punto di vista intellettuale. E la ricerca al servizio del commerciale, quindi resa scientismo, può andare bene solo per chi non e’ dotato di etica o è limitato nella materia.
Quindi il nostro atteggiamento nei confronti di chi e’ impegnato nello scientismo speculativo, e’ essenzialmente di critica e avversione. Il Nostro tradizionale antagonismo fa sì che non subiamo la contaminazione del marketing e, ogniqualvolta si scatena un dibattito su questioni biotecnoiogiche, non manchiamo di porci al di sopra delle parti discutendo dei problemi ai massimi livelli.
Contattaci per informazioni su coaching, anti-aging e TRT/HRT sulle piattaforme Instagram, Telegram, Whatsapp o all’indirizzo mail teamympus86@outlook.it
CEO Amedeo Gabriel Bellizzi [Biochimico, esperto in nutrizione sportiva, coach di BodyBuilding, PEDs consulter, esperto in tecniche Anti-Aging, TRT e HRT, ricercatore e divulgatore scientifico indipendente]
Questione ormai conosciuta è l’effetto delle proteine sulla sazietà percepita. E’ infatti risaputo che, dopo il consumo di proteine (ma anche di grassi) nel duodeno e nella prima parte dell’intestino si libera un ormone, la Colecistochinina (CCK), che segnala al cervello di smettere di mangiare. Inoltre, nell’intestino e nel colon, le proteine ingerite determinano la produzione di un altro ormone saziante, il PYY. Il suo livello si alza dopo 1-2 ore dal pasto e rimane alto per circa 6 ore, limitando così l’insorgenza dell’appetito in questo periodo di tempo. Quindi, le proteine, da un lato, grazie al CCK e all’effetto del PYY, favoriscono la sazietà. Altrettanto conosciuto è il maggiore effetto sulla TID (Termogenesi Indotta dalla Dieta) dato dall’azione dinamica specifica delle proteine le quali, con variabili date dalla fonte, per essere digerite ed assimilate richiedono un dispendio energetico tra il 10 ed il 35% (media del 22,5%).
Da quanto riportato in uno studio pubblicato sette anni fa (2013) sul Journal of Nutrition [1], sembrerebbe che l’aggiunta di Capsaicina, uno degli alcaloidi responsabili della maggior parte della “piccantezza” dei peperoncini, ad una dieta con alto apporto proteico possa migliorarne l’effetto, migliorando ulteriormente il tempo e grado di efficacia di una dieta ipocalorica.
Dettagli dello studio
I ricercatori che hanno svolto lo studio in questione, hanno reclutato 28 soggetti sani facendoli permanere per otto periodi di 24 ore in una camera di respirazione, dove potevano essere osservati e misurate le quantità esatte di calorie ossidate dagli individui in osservazione. In ogni occasione i soggetti hanno ricevuto pasti diversi.
Ai soggetti è stato somministrato il 100% della quantità calorica ossidata giornalmente. Il 10% della quota calorica dei pasti proveniva dalle proteine.
Ai soggetti è stato somministrato l’80% della quantità calorica ossidata giornalmente. Il 10% della quota calorica dei pasti proveniva dalle proteine.
Ai soggetti è stato somministrato il 100% della quantità calorica ossidata giornalmente. Il 10% della quota calorica dei pasti proveniva dalle proteine. Inoltre ai soggetti sono state somministrate 2 capsule contenenti 40.000 unità di calore Scoville dal pepe ad ogni pasto (Capsaicina).
La sostanza bioattiva più importante del pepe è la Capsaicina. I nutrizionisti hanno utilizzato un prodotto realizzato dalla Solaray. [solarayuk.co.uk]
Ai soggetti è stato somministrato l’80% della quantità calorica ossidata giornalmente. Il 10% della quota calorica dei pasti proveniva dalle proteine. Inoltre, i soggetti hanno assunto 2 capsule contenenti 40.000 unità di calore Scoville dal pepe ad ogni pasto.
Ai soggetti è stato somministrato il 100% della quantità calorica ossidata giornalmente. Il 25% dell’introito calorico dei pasti proveniva dalle proteine. I ricercatori hanno ridotto la quantità di carboidrati sostituendola con proteine.
Ai soggetti è stato somministrato l’80% della quantità calorica ossidata giornalmente. Il 25% dell’introito calorico dei pasti proveniva dalle proteine.
Ai soggetti è stato somministrato il 100% della quantità calorica ossidata giornalmente. Il 25% dell’introito calorici dei pasti proveniva dalle proteine. Inoltre i soggetti hanno assunto 2 capsule contenenti 40.000 unità di calore Scoville dal pepe ad ogni pasto (Capsaicina).
Ai soggetti è stato somministrato l’80% della quantità calorica ossidata giornalmente. Il 25% dell’introito calorico dei pasti proveniva dalle proteine. Inoltre, i soggetti hanno assunto 2 capsule contenenti 40.000 unità di calore Scoville dal pepe ad ogni pasto (Capsaicina).
Risultato dello studio
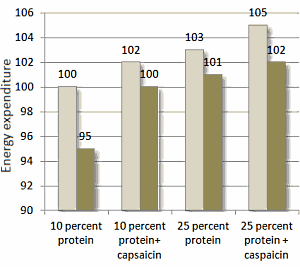
Come mostrato nella figura seguente, il ridotto apporto calorico ha portato ad una riduzione del 5% del dispendio energetico dei soggetti osservati. La riduzione del dispendio energetico a seguito della riduzione dell’apporto calorico non si è verificata quando i soggetti assumevano la Capsaicina o aumentavano il loro apporto proteico. La combinazione di una dieta ricca di proteine con la componente supplementare di Capsaicina ha portato a migliori risultati.
Colonne chiare: apporto calorico = 100% delle calorie ossidate; Colonne scure = apporto calorico = 80% delle calorie ossidate .
La Capsaicina integrativa di per se ha mostrato effetti anoressizzanti causando, consequenzialmente, una migliore compliance del protocollo alimentare. I ricercatori hanno riportato risultati migliori nei soggetti che seguivano una dieta ricca di proteine combinata con l’integrazione di Capsaicina. Indi, si è verificato un effetto additivo sulla soppressione della fame.
Sembrerebbe, quindi, che una combinazione di Capsaicina con la sostituzione di una parte dell’introito calorico giornaliero proveniente dai Carboidrati con Proteine, ed un apporto calorico pari al 20% in meno del totale di mantenimento, possa portare ad un maggiore dispendio energetico e sazietà rispetto a una dieta di controllo del solo bilancio energetico.
In conclusione, i ricercatori sottolineano che l’efficacia della combinazione di Capsaicina e Proteine dovrebbe essere ulteriormente valutata in studi sulla perdita di peso ben progettati su individui in sovrappeso e obesi.
Nota: è utile aggiungere che se si è intenzionati a testare l’effetto della Capsaicina sulla perdita di peso, bisogna tenere in considerazione la tollerabilità individuale alla molecola. Infatti, alle dosi di Capsaicina utilizzate dai ricercatori, specie sul lungo termine, possono causare problemi gastrointestinali (stomaco e tratto digerente).[2]
Riporto qui di seguito un recente studio il quale si è occupato dell’osservazione dello stimolo indotto dalla variabile di inclinazione della Panca sul reclutamento del capo clavicolare del grande pettorale. Lo studio è stato svolto dallo scienziato dello sport David Rodriguez-Ridao dell’Università di Almeria, ed è stato pubblicato il corrente mese (ottobre 2020) su Special Issue Monitoring and Evaluation of Training in Sport and Exercise.[1]
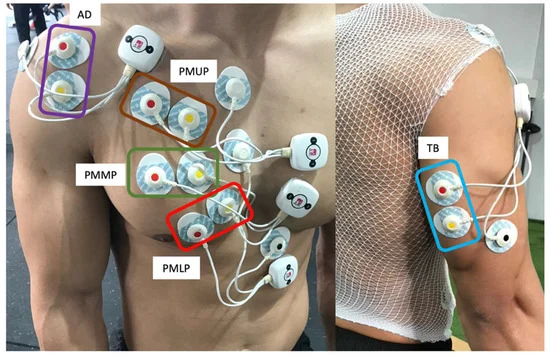
Per lo svolgimento dello studio qui trattato, David Rodriguez-Ridao ha reclutato trenta giovani esperti in sport contro-resistenza sottoponendoli a sessioni di spinte su panca a diversa inclinazione: 0°, 15°, 30°, 45° e 60° (gradi). Ai soggetti è sono stati applicati degli elettrodi i quali dovevano rilevare sulla quanto i gruppi muscolari interessati dall’esercizio venivano reclutati e a che grado.
Nell’immagine sono visibili le zone anatomiche di posizionamento degli elettrodi durante lo studio.
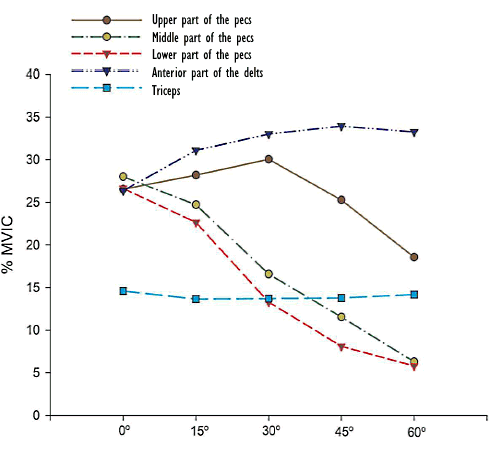
Per il capo sterno-costale ed il capo addominale, la distensione su Panca Piana è risultata superiore alla distensione su Panca Inclinata. Per il capo clavicolare, la stimolazione è stata ottimale con un angolo di 30°.
Se l’angolo della Panca diventava maggiore di 30°, la Panca Inclinata si trasformava in un esercizio che stimolava principalmente i Deltoidi anteriori.
Per i Tricipiti, l’angolo della Panca non era un fattore influente.
Uno studio che può senz’altro essere d’aiuto per atleti e preparatori intenti ad enfatizzare al meglio lo stimolo di questo capo del grande pettorale.
Lo so, è un argomento che avrete sicuramente sentito trattare e ritrattare un infinità di volte, molte delle quali si sono rivelate sterili esposizioni spesso e volentieri inducenti al solito marketing, ma volevo dare il mio contributo per mettere maggiormente in chiaro la questione “volume” nel miglioramento dei parametri prestazionali ed estetici in ambito BodyBuilding. Ovviamente, il mio unico fine a tal proposito è quello di svolgere una sana divulgazione scientifica. Per farlo citerò i lavori di Brad Schoenfeld et al. i quali, ad una analisi oggettiva, dimostrano di possedere i requisiti di design corretti per poter servire da punti di partenza per la gestione del volume all’interno di una programmazione nell’allenamento contro-resistenza.
HIT Vs. HVT – l’infinito scontro decontestualizzato tra due estremi –
Nessuno può negare il fatto che non vi è argomento nel campo dell’esercizio contro resistenza più dibattuto negli ultimi decenni della quantità di volume necessaria per massimizzare l’ipertrofia muscolare. Da un lato della controversia ci sono i partigiani dell’allenamento ad alta intensità (HIT), che generalmente predicano che una singola serie, o poche più, di un esercizio sia tutto ciò che è necessario per avere un ottimale miglioramento nello sviluppo del tessuto muscolare; essi affermano che il superamento di questa quantità di lavoro non solo non è benefico, ma che in realtà può causare una regressione a causa del sovrallenamento. Il creatore dei macchinari Nautilus Arthur Jones è spesso indicato come il capostipite di questo movimento che il Bodybuilder Mike Mentzer ha contribuito a rendere popolare alla fine degli anni ’80 e inizi ’90 con il suo famosissimo metodo “Heavy Duty”.
Dall’altra parte del fronte ci sono i sostenitori dell’alto volume (HVT) i quali affermano che l’esecuzione di più serie è il fattore essenziale per stimolare nel modo migliore lo sviluppo muscolare. La stragrande maggioranza dei Bodybuilder competitivi aderisce a quest’ultima filosofia, come mostrato in un sondaggio di qualche anno fa in cui il 95% degli intervistati ha riferito di allenarsi con protocolli in multi-frequenza.
Nel tentativo di ideare linee guida basate sull’evidenza, il team di ricerca del quale fa parte Brad Schoenfeld ha effettuato una meta-analisi sugli effetti del volume nella crescita muscolare e che è stata pubblicata nel 2017. [1] Nel caso in cui non ne siate a conoscenza, una meta-analisi riunisce i risultati di tutti gli studi su un determinato argomento al fine di confrontarne i dati come se fosse un unico grande studio invece di più piccoli studi. Ciò aiuta a estrapolare una maggiore precisione statistica e quindi a trarre conclusioni pratiche ed esaustive dalla ricerca corrente. I risultati dell’analisi di cui sopra hanno mostrato una relazione dose-risposta abbastanza chiara, con volumi più elevati che hanno portato a maggiori guadagni muscolari.
Sebbene i risultati della meta-analisi sembrano convincenti, sono presenti alcune lacune. Per esempio, la stragrande maggioranza degli studi presi in esame hanno coinvolto soggetti non allenati; solo un paio di studi hanno preso in esame persone con esperienza nell’allenamento contro-resistenza. Questo è importante perché la prima fase dell’allenamento di un principiante è associata a una diversa risposta adattativa rispetto alle ultime fasi precedenti alla maturità necessaria per essere considerati “intermedi”, per esempio; quindi, non è correttamente possibile generalizzare i risultati ottenuti con atleti ben allenati. Inoltre, i realizzatori della meta-analisi non sono stati in grado di determinare gli effetti oltre le 10 serie per muscolo a settimana; semplicemente non era sufficiente una ricerca che guardasse ai volumi di allenamento più elevati per trarre conclusioni rilevanti su quale potesse essere una quantità superiore di volume per l’ottenimento di guadagni più o meno regolari.
In quest’ottica, Schoenfeld et al. si sono proposti di colmare le lacune presenti in letteratura effettuando uno studio sull’argomento, pubblicato nel 2018 su Medicine and Science in Sports and Exercise.[2] Ecco i dettagli:
Studio, metodo e risultati ottenuti
45 giovani soggetti di sesso maschile allenati contro-resistenza sono stati assegnati in modo casuale ad allenarsi con 1, 3 o 5 serie per esercizio; Alla fine, 11 soggetti si sono ritirati per vari motivi, lasciando un totale di 34 partecipanti che hanno completato lo studio. In ogni sessione sono stati eseguiti un totale di 7 esercizi pianificati al fine di allenare tutti i principali gruppi muscolari (chest press, shoulder press, lat pulldown, seated row, squat, leg press e leg extension). I soggetti hanno eseguito 8-12 ripetizioni per serie, con tutte le serie eseguite fino al cedimento muscolare. L’allenamento è stato effettuato su 3 giorni non consecutivi a settimana per 8 settimane. Le routine sono state completamente supervisionate dal team di ricerca per garantire la piena conformità. Le misurazioni includevano: 1RM forza nello squat e nella panca; resistenza muscolare della parte superiore del corpo nella distensione su panca al 50% dell’1RM; e ipertrofia del bicipite, tricipite, metà coscia e coscia laterale mediante ultrasuoni.
Tutti i gruppi hanno ottenuto guadagni sostanziali nella forza massima sia nello squat che nella distensione su panca durante il periodo di studio di 8 settimane, ma l’entità dei guadagni era sorprendentemente simile tra i gruppi senza differenze statistiche rilevate. I risultati hanno leggermente favorito le condizioni di volume più elevato per la resistenza muscolare della parte superiore del corpo su base assoluta, ma le differenze non hanno raggiunto la significatività statistica e probabilmente non avevano rilevanza pratica.
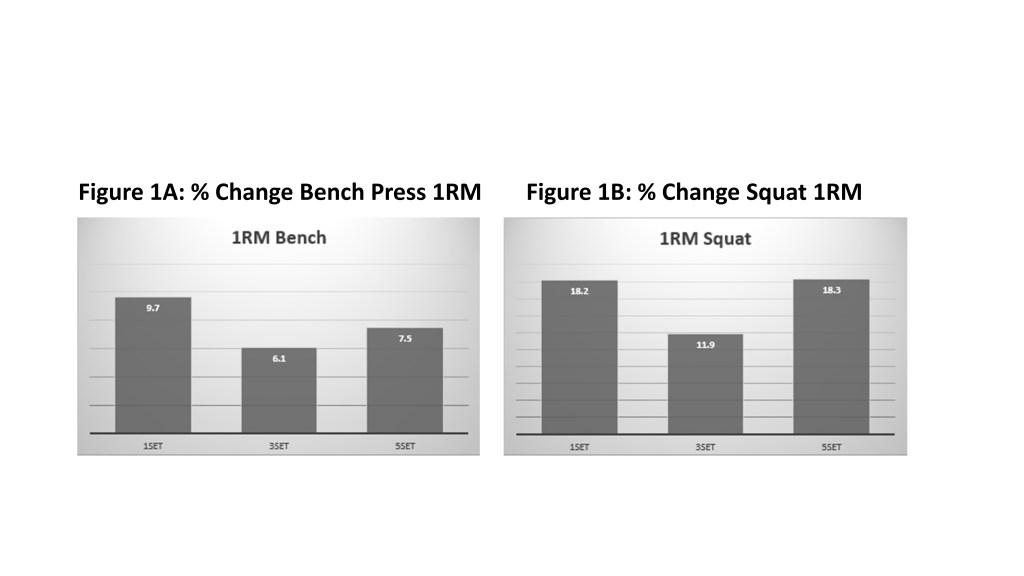
In alternativa, c’era una chiara relazione dose-risposta per l’ipertrofia muscolare, con 5 serie che mostravano i maggiori guadagni, seguite da 3 serie e poi 1 serie. Questi risultati erano generalmente coerenti per tutti i muscoli studiati ed erano più pronunciati nella muscolatura inferiore del corpo. Il grafico seguente presenta una rappresentazione visiva dei risultati.
I guadagni di forza nel gruppo da 1 set erano altrettanto buoni come per i gruppi a volume più alto (vedi Figura 1). Inoltre, questi risultati sono stati raggiunti in una frazione del tempo in rapporto alle routine con volume più elevato poiché il gruppo da 1 set si è allenato per circa 13 minuti a sessione mentre le sessioni del gruppo a 5 set sono durate circa 68 minuti! Quindi, una conclusione importante sarebbe che, se aumentare la forza è l’obiettivo principale, è possibile adottare vantaggiosamente un approccio di allenamento a basso volume e alta intensità raggiungendo il risultato prefissato in meno di 45 minuti a settimana. Ora questa osservazione necessita di una precisazione: l’allenamento è stato effettuato in un intervallo di ripetizioni moderato (8-12 RM). Direi che se usassimo un vero range da powerlifting (3-5 RM), probabilmente si dovrebbero aggiungere alcune serie addizionali poiché il basso numero di ripetizioni per serie limiterebbe la quantità di “stimolo” e quindi richiederebbe un volume più alto.
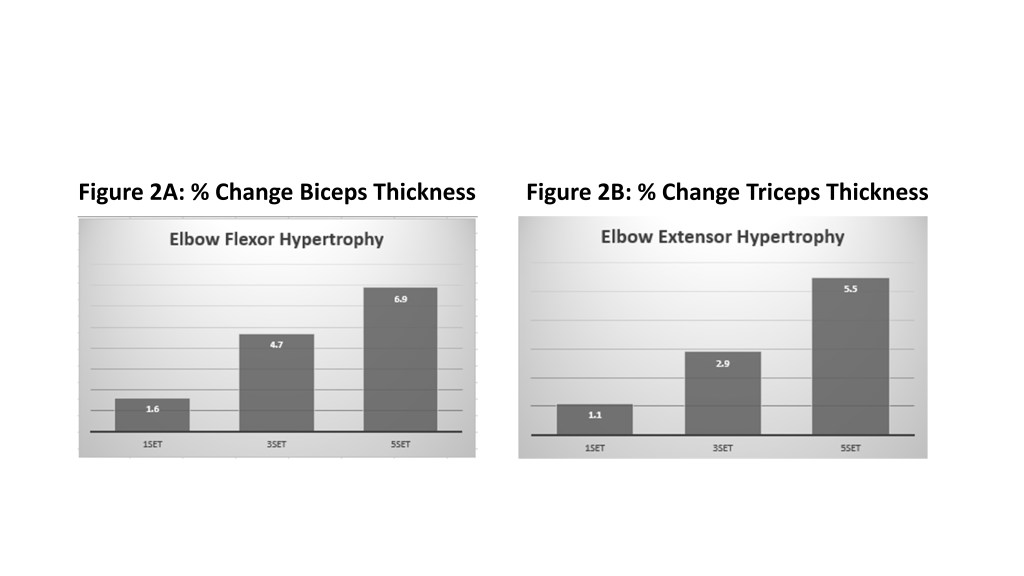
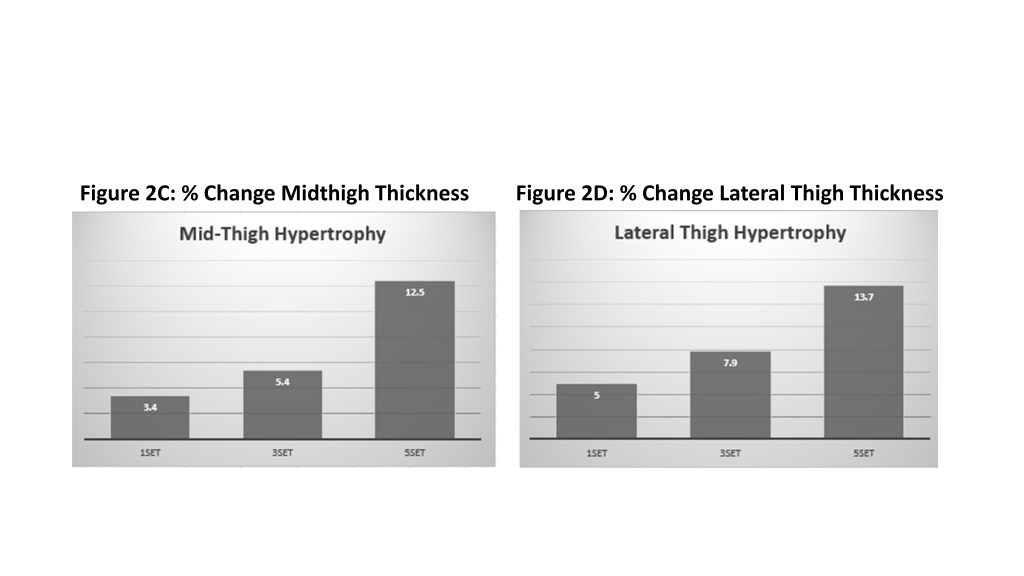
D’altra parte, i risultati per l’ipertrofia erano coerenti con la meta-analisi argomentativa iniziale, dimostrando che sono necessari volumi più elevati per massimizzare la crescita muscolare. Questi risultati erano coerenti e ricorrenti in tutti i muscoli studiati (bicipiti, tricipiti e quadricipiti). Inoltre, è stata generalmente osservata una relazione dose-risposta, per cui 3 set hanno prodotto maggiori aumenti di dimensioni muscolari rispetto a 1 set e 5 set erano superiori a 3 set. Come mostrato nel grafico allegato (Figure 2), l’entità delle differenze era piuttosto netta. Ad esempio, la crescita del bicipite è stata dell’1,6%, 4,7% e 6,9% rispettivamente per i gruppi 1, 3 e 5. Il volume ha avuto un effetto ancora maggiore sulla crescita del quadricipite, con aumenti del 5,0%, 7,9 e 13,7% per la coscia laterale rispettivamente nei gruppi 1, 3 e 5 serie. Spread simili sono stati osservati in relazione alla crescita nella regione mediale della coscia.
Quantificando il volume in serie per muscolo a settimana, come spesso si fa nel Bodybuilding, la quantità di volume più alto del gruppo in questione ha eseguito 30 serie / muscolo / settimana per bicipiti e tricipiti e 45 serie / muscolo / settimana per le cosce. E’ stato notato che la maggior parte delle serie è stata eseguita con esercizi composti. Pertanto, i tricipiti sono stati stimolati con movimenti di spinta (chest press e shoulder press) mentre i bicipiti con movimenti di trazione (lat pulldown e rows). Sebbene tricipiti e bicipiti ricevano chiaramente molto lavoro con questi movimenti, non è chiara la misura in cui sono stimolati; Rimane discutibile se i risultati sarebbero cambiati se gli autori avessero incluso esercizi a mono-articolari come triceps pushdown e arm curls. Allo stesso modo, la parte inferiore del corpo coinvolge le prestazioni dello squat e del leg press, che non solo stimolano i quadricipiti femorali ma comportano anche una sostanziale attivazione degli estensori dell’anca. Ancora una volta, non è chiaro quanta stimolazione riceva il quadricipite femorale con questi movimenti rispetto agli altri muscoli che lavorano in sinergia. E’ stata inclusa la Leg-Estension per la parte inferiore del corpo, la quale fornisce un targeting più diretto nello stimolo dei quadricipiti femorali.
Ora, vanno considerati alcuni punti riguardo ai risultati che devono essere presi in esame al fine di tentare una estrapolazione di risultati applicabili nella pratica. Innanzi tutto , i soggetti esaminati erano giovani uomini allenati contro resistenza che apparentemente sarebbero stati in grado di riprendersi bene dagli effetti di grandi volumi allenanti. È ragionevole ipotizzare che gli individui di mezza età e gli anziani probabilmente non sarebbero stati in grado di riprendersi anche da volumi più elevati di allenamento, e quindi potrebbero non rispondere in modo favorevole o forse anche avere una risposta ridotta.
Inoltre, la durata dello studio è stata relativamente breve, comprendendo 8 settimane di allenamento pianificato. Il corpo umano è molto resistente e sopporta bene alti livelli di stress a breve termine. Quando questi fattori di stress sono adeguatamente gestiti, c’è una risposta adattativa positiva; nel caso di volumi di allenamento ad alta resistenza, il risultato è una maggiore crescita muscolare. Tuttavia, l’esposizione persistente a tali fattori di stress alla fine sovraccarica la capacità di risposta del corpo, portando a uno stato di sovrallenamento. Ciò solleva la possibilità che la periodizzazione di brevi periodi di volume maggiore con periodi di allenamento di volume da inferiore a moderato possa essere la migliore strategia per massimizzare i guadagni di massa muscolare.
In aggiunta, non sono stati effettuati test a metà dello studio. È possibile che la forza possa essere stata maggiore per il gruppo sottoposto a volume più alto dopo 4 settimane e che i soggetti si siano avvicinati (o abbiano addirittura raggiunto) ad una condizione di sovrallenamento in seguito, in modo tale da causare uno stallo o una regressione dei risultati. Di conseguenza è possibile che l’ipertrofia si sia stabilizzata in precedenza nello studio per via delle condizioni di volume più elevato e che continuare ad allenarsi con tale volume fosse superfluo o addirittura dannoso. Queste possibilità richiedono ulteriori studi. È interessante notare che le interviste di fine studio fatte ai soggetti non hanno indicato alcun segno percettivo di sovrallenamento anche nel gruppo con il volume più alto, sebbene ciò non precluda la prospettiva che possa essersi verificato.
È anche importante notare che è stata misurata solo l’ipertrofia delle braccia e delle cosce. Resta da determinare se diversi gruppi muscolari possono avere risposte diverse a quantità variabili di volume. Sembra probabile che questo possa essere il caso e che gli effetti differenziali sarebbero specifici in risposta alla genetica individuale, per cui alcuni muscoli rispondono meglio a volumi più elevati di allenamento mentre altri no. C’è ancora molto da ricercare e apprendere.
Conclusioni
Ricapitolando:
I guadagni di forza sono simili tra il gruppo a 1 serie per esercizio che in quello con 5 serie per esercizio, indicando che se l’obiettivo è semplicemente quello di aumentare la forza, questo può essere ottenuto con una quantità minima di volume. Questi risultati sono specifici per allenamenti con una gamma moderata di ripetizioni (8-12 ripetizioni); sembra probabile che l’allenamento in più di un range di powerlifting (3-5 ripetizioni) richiederebbe l’esecuzione di più serie per massimizzare la forza.
Il volume è il fattore principale della crescita muscolare in sala pesi, con più serie che si traducono in maggiori guadagni. L’ipertrofia della parte superiore del corpo ha continuato a mostrare effetti benefici con 30 serie per muscolo a settimana e sono stati osservati continui guadagni della parte inferiore del corpo con 45 serie per muscolo a settimana. Questi numeri non devono essere necessariamente considerati come raccomandazioni definitive, ma piuttosto come indicatori del fatto che volumi più elevati possono provocare guadagni muscolari superiori in tempi relativamente brevi. Inoltre, sembra probabile che l’allenamento ripetuto con volumi elevati porterà inevitabilmente a un superamento della soglia adattativa non funzionale e quindi a compromettere i risultati. Pertanto, il volume dovrebbe essere manipolato in maniera ondulatoria in modo che i periodi di volumi relativi più elevati vengano ciclati con periodi di volume inferiori; inoltre, dovrebbero essere impiegati scarichi regolari per promuovere un corretto recupero.
È essenziale ricordare che gli studi si limitano a fornire le risposte medie su gruppi di persone. Pertanto, questi risultati possono solo fornire linee guida generali su quanto volume sia benefico per la forza e l’ipertrofia; la risposta per una determinata persona varierà in base alla genetica e ai fattori dello stile di vita e quindi la prescrizione deve essere calibrata individualmente. Una buona raccomandazione generale per il volume sarebbe quella di eseguire ~ 10-20 serie / muscolo / settimana; alcuni atleti si troveranno bene con volumi inferiori a questo intervallo, mentre altri progrediranno con volumi leggermente più alti. I risultati dello studio qui riportato suggeriscono che si possono impiegare cicli di allenamento ad alto volume nel breve termine per stimolare maggiormente lo sviluppo di gruppi muscolari carenti che rispondono male all’allenamento a volume più conservativo o hanno raggiunto un plateau nello sviluppo.
Come mi sono già ritrovato a sottolineare nel mio profilo Instagram, è essenziale che ogni atleta o professionista del settore Fitness o Bodybuilding sappia che la dimensione e la composizione del muscolo scheletrico è determinata dai cambiamenti nei tassi di sintesi proteica muscolare (MPS) rispetto a quelli della degradazione delle proteine muscolari. Nello stato di riposo, a digiuno, il tasso di MPS è inferiore a quello della degradazione delle proteine muscolari, determinando uno stato di equilibrio proteico negativo. L’ingestione di proteine di alta qualità, ricche di amminoacidi essenziali, stimola un aumento transitorio del tasso di MPS con conseguente stato di equilibrio proteico muscolare positivo. È anche noto che un singolo periodo di esercizio contro resistenza eseguito a digiuno indurrà un aumento sia della MPS che del catabolismo; tuttavia, il tasso di MPS è elevato 48h dopo l’esercizio, mentre il tasso di degradazione proteica ritorna al valore basale 48h dopo l’esercizio (curve incrociate e inversamente proporzionali). Fondamentalmente, l’assunzione di un adeguato apporto proteico e l’esercizio contro resistenza conferiscono effetti additivi sulla MPS e sul bilancio proteico netto. Quindi quando ripetuti esercizi contro resistenza sono abbinati con una adeguata quantità di proteine nella dieta si verifica uno stato prolungato di equilibrio proteico muscolare positivo il quale porta ad un aumento graduale nelle dimensioni del muscolo scheletrico.
Morale della “favola”? Multi-frequenza ondulatoria sui parametri di volume e intensità con periodi di scarico basati sui feedback di resa dell’atleta. E, ovviamente, il fattore alimentare dovrà essere correttamente gestito in funzione delle esigenze soggettive-preparatorie del soggetto interessato (adeguato apporto proteico, glucidico e lipidico su base calorica giornaliera/settimanale).
Sono ormai diversi anni che in nutrizione si discute della questione “dolcificanti artificiali” e se essi siano o meno deleteri nel contesto dell’alimentazione umana. Molti studi hanno “assolto” dalla loro presunta pericolosità dolcificati ipocalorici molto diffusi come l’Aspartame, con le corrette modalità d’uso ovviamente (vedi dosaggio totale giornaliero). Mentre altri dolcificanti artificiali sono decisamente posizionati nella “zona grigia”, come l’Acesulfame-K. Il peggiore, secondo quanto emerso dalle ultime ricerche, sembrerebbe essere il Sucralosio. Il Sucralosio, un dolcificante sintetico mille volte più dolce dello zucchero da cucina (Saccarosio), sembra che possa causare sintomi pre-diabetici nelle persone sane. I ricercatori dell’Università di Yale hanno riportato della comparsa di questi sintomi in un articolo comparso recentemente su “Cell Metabolism”.[1] Sebbene i soggetti dello studio non fossero effettivamente patologici, i risultati sono stati così preoccupanti che l’università ha consigliato ai ricercatori di interrompere lo studio.
Caratteristiche del Sucralosio:
La maggior parte del Sucralosio (E-955) ingerito non viene enzimaticamente scomposto, quindi non apporta calorie. [2] È prodotto dalla clorurazione del saccarosio. Il Sucralosio è da 320 a 1.000 volte più dolce del Saccarosio [3], tre volte più dolce dell’Aspartame e dell’Acesulfame-K, e due volte più dolce della Saccarina Sodica.
Sebbene il Sucralosio è ampiamente considerato stabile e sicuro per l’uso a temperature elevate (come nei prodotti da forno), ci sono alcune prove che mostrano un iniziale degradazione a temperature superiori a 119 gradi Celsius. [4][5] Il successo commerciale dei prodotti a base di Sucralosio deriva semplicemente dal confronto favorevole con altri dolcificanti ipocalorici in termini di gusto, stabilità e sicurezza nelle prima citate circostanze.[6]
Lo studio in questione e risultati emersi…
Per lo svolgimento dello studio che qui andiamo trattando, i ricercatori hanno diviso 45 soggetti sani in tre gruppi. Ogni gruppo si recava al laboratorio di controllo sette volte durante un periodo di due settimane. Li, ai soggetti veniva data una bevanda analcolica da 355ml.
Il contenuto della suddetta bevanda differiva nei tre gruppi esaminati come segue:
Contenuto 1° gruppo [LCS]: 60mg di Sucralosio;
Contenuto 2° gruppo [Sugar]: 30g di Saccarosio [normale zucchero da tavola];
Contenuto 3° gruppo [Combi]: 60mg di Sucralosio + 31g di Maltodestrine.
Come già accennato, la struttura chimica del Sucralosio è molto simile a quella del Saccarosio. In tre punti, tuttavia, il Sucralosio presenta gruppi cloro che mancano nel Saccarosio. A causa di questi gruppi cloro, secondo alcuni studi, il Sucralosio è mille volte più dolce del Saccarosio.
Le bevande analcoliche assunte dai partecipanti dei gruppi 1 e 2 non hanno avuto alcun effetto sulla farmacocinetica del glucosio negli individui esaminati. Quando i ricercatori hanno somministrato a questi soggetti un lotto di glucosio dopo 2 settimane, la glicemia ematica si è ridotta con la stessa velocità osservata prima del periodo di due settimane dello studio. A questo proposito, le bevande analcoliche erano sicure.
Il quadro è cambiato quando i ricercatori hanno esaminato la quantità di insulina che era presente nel sangue dei soggetti dopo la somministrazione del glucosio. Questa quantità era significativamente maggiore nei soggetti che avevano ricevuto bevande analcoliche contenenti Maltodestrine più Sucralosio.
Ciò implica che la combinazione di Sucralosio con un carboidrato ad assorbimento altera il metabolismo glucidico peggiorando, sebbene in acuto, la sensibilità all’Insulina.
I ricercatori hanno anche osservato che in un certo numero di soggetti, la combinazione di Sucralosio e un carboidrato a rapido assorbimento portava ad un aumento dell’Insulina basale, misurata al mattino prima che i soggetti consumassero il loro primo pasto della giornata. Ciò suggerisce anche una possibile ridotta sensibilità all’insulina in cronico.
In bocca, nell’intestino e in altri punti del corpo, i dolcificanti come il Sucralosio interagiscono con i recettori del dolce T1R2 / T1R3. Questi recettori sono in realtà destinati al glucosio e ad altri zuccheri naturali. Regolano l’assorbimento degli zuccheri da parte dell’intestino tenue.
I ricercatori ipotizzano che, tramite questi recettori, il Sucralosio possa indurre il corpo ad assorbire rapidamente i carboidrati assimilandoli ancora più velocemente, interrompendo l’equilibrio tra glucosio e insulina e riducendo la sensibilità all’insulina non solo in acuto ma anche, potenzialmente, in cronico.
I ricercatori hanno scritto che questi risultati suggeriscono che il consumo di Sucralosio altera il metabolismo del glucosio consumato simultaneamente producendo rapidamente effetti deleteri sulla salute metabolica.
Durate di esposizione simili quasi certamente si verificano negli esseri umani nella quotidianità, soprattutto se si considera il consumo di una bevanda dietetica insieme ad un pasto. Ciò solleva la possibilità che l’effetto combinato possa essere un importante contributo all’aumento dell’incidenza del diabete di tipo 2 e l’obesità, in senso indiretto o induttivo.
In tal caso, l’aggiunta di dolcificanti a basso contenuto calorico per aumentare la dolcezza di cibi e bevande già contenenti carboidrati dovrebbe essere scoraggiato e il consumo di bevande dietetiche durante i pasti dovrebbe essere sconsigliato.
Nota:Il Sucralosio risulta particolarmente deleterio anche sul microbiota intestinale. Il primo studio che ha valutato il Sucralosio sul microbiota intestinale è stato eseguito nel 2008 con l’uso di campioni fecali di ratti Sprague-Dawley che hanno ricevuto il dolcificante per 12 settimane. Il consumo di Sucralosio ha ridotto il numero totale di batteri anaerobici e aerobici, bifidobatteri, lattobacilli, Bacteroides e Clostridium.(7) La somministrazione di 15mg di Sucralosio/kg ha influenzato l’abbondanza relativa del Clostridium cluster XIVa nei topi.(8) Più recentemente, la somministrazione di Sucralosio nei topi ha prodotto modifiche nel microbiota intestinale a 14 diversi livelli tassonomici, tra cui Turicibacteraceae, Lachnospiraceae, Ruminococcaceae, Verrucomicrobiaceae, Staphylococcaceae, Streptococcaceae, Dehalobacteriaceae, Dehalobacterium, Lachnospiraceae, Lachnospiraceae ordine Bacillales e cambiamenti nella sintesi e regolazione degli amminoacidi. Queste variazioni erano correlate all’infiammazione nell’ospite.(9)
Nonostante lo studio sia di piccole dimensioni e non sia controllato (non vi è sicurezza nel comportamento alimentare seguito dai soggetti esaminati al di fuori di quanto emergesse durante i controlli), esso rappresenta un forte incentivo verso la ricerca sugli effettivi vantaggi e svantaggi del consumo di dolcificanti in soggetti sani e non.
Nota:Mancano ad oggi prove di un possibile beneficio per la perdita di peso a lungo termine con alcuni dati che supporto il rischio di un aumento di peso e di sviluppo di malattie cardiache con l’uso di questo dolcificante.[10]
Splenda alters gut microflora and increases intestinal p-glycoprotein and cytochrome p-450 in male rats.Abou-Donia MB, El-Masry EM, Abdel-Rahman AA, McLendon RE, Schiffman SSJ Toxicol Environ Health A. 2008; 71(21):1415-29.
Effects of Consuming Xylitol on Gut Microbiota and Lipid Metabolism in Mice.Uebanso T, Kano S, Yoshimoto A, Naito C, Shimohata T, Mawatari K, Takahashi ANutrients. 2017 Jul 14; 9(7):.
Gut Microbiome Response to Sucralose and Its Potential Role in Inducing Liver Inflammation in Mice.Bian X, Chi L, Gao B, Tu P, Ru H, Lu KFront Physiol. 2017; 8():487.
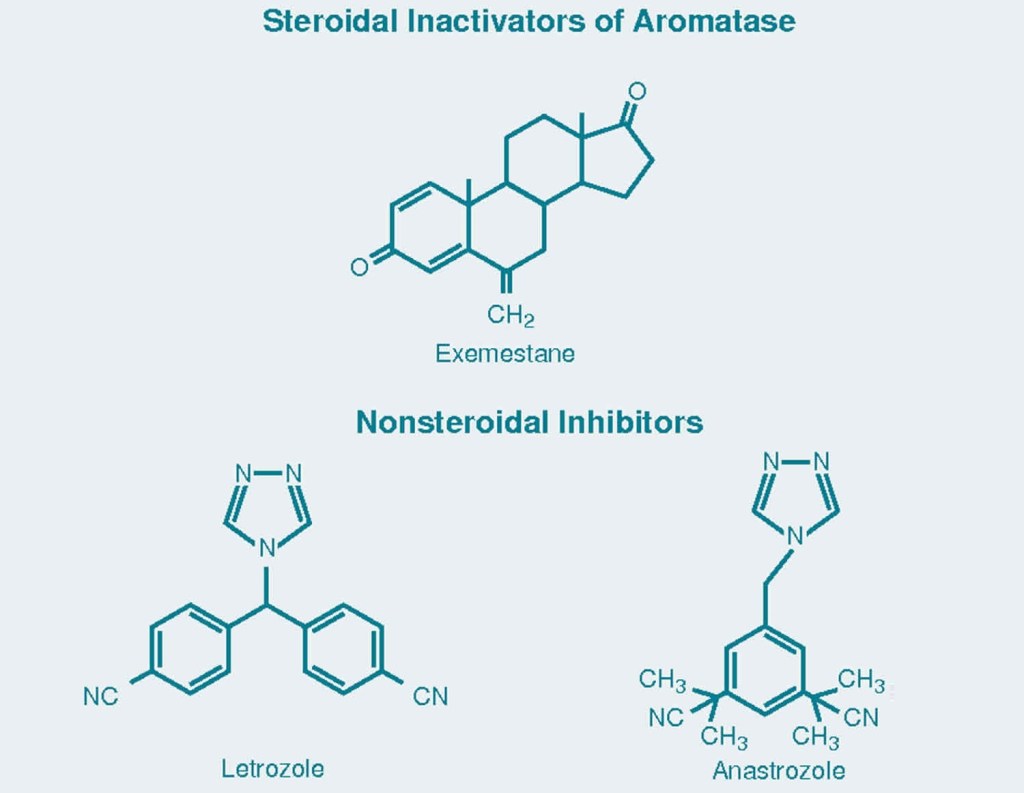
Come ben sappiamo, gli Inibitori dell’Aromatasi (AI) sono farmaci che possono abbassare il livello degli estrogeni nel flusso ematico riducendone di conseguenza l’attività tissutale-metabolica. Ovviamente, la loro azione inibitoria è esplicata attraverso l’inattivazione di uno specifico enzima. Si tratta infatti dell’Enzima Aromatasi. Detto in modo semplicistico e riduttivo, l’Enzima Aromatasi converte gli Androgeni in Estrogeni. Ad esempio, converte il Testosterone in Estradiolo.
Ci sono attualmente in circolazione tre AI divenuti estremamente popolari in ambito sportivo, e soprattutto nel BodyBuilding. Questi sono: Exemestane (Aromasin), Anastrozolo (Arimidex) e Letrozolo (Femera). Sono comunemente usati nel trattamento del cancro al seno. Sono state quindi svolte molte ricerche sulle donne. Nelle donne, gli AI hanno mostrato un grado di soppressione estrogenica significativamente marcato. Ad esempio, l’Exemestane sopprime i livelli di Estradiolo del 92% nei pazienti con carcinoma mammario in postmenopausa.[1] Allo stesso modo, il Letrozolo e l’Anastrozolo riducono i livelli di Estradiolo di quasi il 90%, anche in questo caso nei pazienti con carcinoma mammario in postmenopausa.[2]
Per questo motivo, nel presente articolo non mi accingerò ad esporre una semplice disamina degli effetti generali o specifici degli inibitori del Aromatasi (cosa già fatta, e nemmeno molto semplicisticamente, qualche anno fa). E’ mia intenzione, invece, trattare con il supporto della letteratura scientifica ad oggi disponibile il reale impatto che questi farmaci hanno sugli individui di sesso maschile, sia in fisiologia che durante l’uso di AAS esogeni soggetti all’Enzima Aromatasi.
Effetto degli AI negli individui di sesso maschile in fisiologia:
Pochi studi hanno osservato l’efficacia degli Inibitori dell’Aromatasi per ridurre i livelli di estrogeni negli uomini. I loro risultati differiscono sensibilmente da quelli riscontrati negli studi sulle donne.
Uno studio condotto su giovani uomini ha rilevato che 25mg di Exemestane sopprimevano i livelli di Estradiolo del 62%, 12 ore dopo l’assunzione.[3] Dopo una somministrazione regolare protratta per dieci giorni, è stata riscontrata una soppressione di solo il 38% nelle 24 ore seguenti l’ultima dose somministrata. In particolare, raddoppiare il dosaggio a 50mg/die non ha portato a una maggiore diminuzione dei livelli di Estradiolo.
Risultati simili si osservano in giovani uomini trattati con Anastrozolo per 10 giorni.[4] La somministrazione giornaliera di Anastrozolo a 0,5 e 1 mg porta ad una diminuzione dei livelli di Estradiolo di circa il 50%. Sebbene di dubbia rilevanza, gli autori non hanno menzionato quante ore dopo l’ultima dose sono state effettuate le misurazioni.
Infine, il Letrozolo ha mostrato la stessa efficacia dell’Exemestane e dell’Anastrozolo. Dopo 28 giorni di trattamento con Letrozolo alla dose di 2,5mg/die, i livelli di Estradiolo hanno subito una riduzione del 46% negli uomini giovani e del 62% negli uomini anziani.[5] Non è chiaro se la differenza tra giovani uomini e uomini anziani sia una differenza reale. Gli autori non hanno eseguito alcun test statistico per valutare matematicamente ciò. In linea con questi risultati, un altro studio ha rilevato che i livelli di Estradiolo erano stati ridotti del 56% negli uomini trattati con 2,5mg di Letrozolo al giorno per 4 settimane.[6]
In sintesi, si potrebbe affermare che i tre popolari AI sono dotati di pari efficaci nel diminuire gli estrogeni negli individui di sesso maschile in fisiologia.
Effetto degli AI nei soggetti di sesso maschile sottoposti a somministrazioni sovrafisiologiche di AAS:
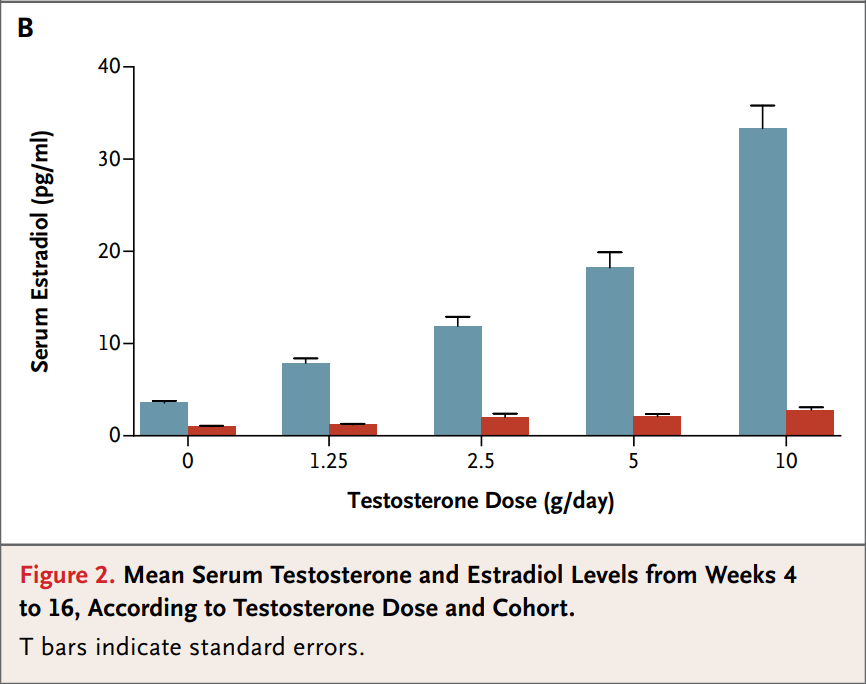
Gli studi di cui sopra sono stati condotti su uomini in fisiologia con i loro livelli endogeni di Testosterone nel range di normalità. Ma la situazione sembra essere diversa quando il Testosterone endogeno viene sostituito con il Testosterone esogeno. Finkelstein et al. ha esaminato gli effetti di quantità crescenti di Testosterone con e senza Inibitori dell’Aromatasi su diversi fattori.[7] Un totale di 198 uomini sani sono stati assegnati in modo casuale a ricevere un placebo, 1,25g, 2,5g, 5g o 10g/die di Testosterone in gel per 16 settimane, e altri 202 hanno ricevuto l’Anastrozolo in combinazione con la dose di Testosterone. La produzione endogena di Testosterone era stata soppressa da iniezioni di Goserelina Acetato.
Meccanismo d’azione schematizzato della Goserelina Acetato
L’immagine seguente mostra l’effetto sui livelli di Testosterone ed Estradiolo in riferimento alle diverse somministrazioni. Si noti che le barre blu si riferiscono agli uomini che ricevevano solo Testosterone mentre le barre rosse quelli che lo ricevevano in combinazione con Anastrozolo.
Tratto da Finkelstein et al. [7]
Come si può vedere, i livelli di Estradiolo sono maggiormente diminuiti rispetto ai numeri discussi in precedenza. Ciò suggerisce che la soppressione dell’Estradiolo da parte degli Inibitori dell’Aromatasi è più marcata negli uomini trattati con Testosterone esogeno in modo “dose dipendente”, e non solamente attribuibile ad una ratio derivante da livelli sovrafisiologici di AAS soggetti all’aromatizzazione.
Qual’è il nesso causale che determina la differenza di risposta tra soggetti in fisiologia e soggetti trattati con Testosterone esogeno?
Sfortunatamente, non esiste una ricerca diretta che risponda in modo chiaro e inequivocabile a questa domanda. Ma se si dovesse ipotizzare, per i dati emersi, sembrerebbe che, banalmente, l’efficacia degli AI negli uomini sia direttamente proporzionata al livello di AAS circolanti soggetti all’Aromatasi il quale causerebbe un incremento dell’espressione enzimatica a livello testicolare e adiposo. Una parte sostanziale dell’Estradiolo è prodotta dall’attività dell’Aromatasi nei testicoli. Nei testicoli, le concentrazioni di Testosterone arrivano a livelli circa 100 volte superiori a quelli presenti nel circolo ematico. Poiché gli AI devono inibire in modo competitivo l’Aromatasi, i dosaggi potrebbero dover essere molto più alti per portare a una significativa inibizione enzimatica nei testicoli. Ma, ovviamente, questa è un’ipotesi al momento non scientificamente dimostrata, sebbene rimanga una delle più plausibile.
Riferimenti:
1- J. Geisler, N. King, G. Anker, G. Ornati, E. Di Salle, P. Lønning, and M. Dowsett. In vivo inhibition of aromatization by exemestane, a novel irreversible aromatase inhibitor, in postmenopausal breast cancer patients. Clinical Cancer Research, 4(9):2089–2093, 1998. 2- J. Geisler, B. Haynes, G. Anker, M. Dowsett, and P. Lonning. Influence of letrozole and anastrozole on total body aromatization and plasma estrogen levels in postmenopausal breast cancer patients evaluated in a randomized, cross-over study. Journal of Clinical Oncology, 20(3):751–757, 2002. 3- N. Mauras, J. Lima, D. Patel, A. Rini, E. di Salle, A. Kwok, and B. Lippe. Pharmacokinetics and dose finding of a potent aromatase inhibitor, aromasin (exemestane), in young males. The Journal of Clinical Endocrinology & Metabolism, 88(12):5951–5956, 2003. 4- N. Mauras, K. O. O’Brien, K. O. Klein, and V. Hayes. Estrogen suppression in males: metabolic effects. The Journal of Clinical Endocrinology & Metabolism, 85(7):2370–2377, 2000. 5- G. G. T’sjoen, V. A. Giagulli, H. Delva, P. Crabbe, D. De Bacquer, and J.-M. Kaufman. Comparative assessment in young and elderly men of the gonadotropin response to aromatase inhibition. The Journal of Clinical Endocrinology & Metabolism, 90(10):5717–5722, 2005. 6- G. Raven, F. H. de Jong, J.-M. Kaufman, and W. de Ronde. In men, peripheral estradiol levels directly reflect the action of estrogens at the hypothalamo-pituitary level to inhibit gonadotropin secretion. The Journal of Clinical Endocrinology & Metabolism, 91(9):3324–3328, 2006. 7- J. S. Finkelstein, H. Lee, S.-A. M. Burnett-Bowie, J. C. Pallais, E. W. Yu, L. F. Borges, B. F. Jones, C. V. Barry, K. E. Wulczyn, B. J. Thomas, et al. Gonadal steroids and body composition, strength, and sexual function in men. New England Journal of Medicine, 369(11):1011–1022, 2013.
Al principio del mese di giugno di quest’anno ho riportato alcuni casi studio i quali facevano emergere il potenziale effetto epatotossico dato dall’uso dei SARM RAD-140 e LGD-4033. Il caso studio riguardante LGD-4033 non era di per sé convincente, poiché il Bodybuilder in questione era solito consumare discrete quantità di alcol. Di recente, i medici del Baylor College of Medicine negli Stati Uniti hanno segnalato un altro caso di danno epatico legato all’uso di LGD-4033.[1] E in questo nuovo caso studio, non ci sono altri fattori esplicativi del problema.
Il soggetto protagonista del caso studio è un Bodybuilder di 32 anni che ha riferito ai medici di aver usato 10mg/die di LGD-4033 in forma liquida per 15 giorni. Dopo di che aveva cominciato a lamentare malessere e interruppe il suo ciclo con il suddetto SARM. L’uomo aveva dolori di stomaco e nausea, oltre a prurito e ittero. Le sue feci erano grigie, e aveva perso l’appetito. Quando si è rivolto ai medici, l’uomo aveva già perso 18Kg.
Questi sono classici sintomi da danno epatico. Infatti, quando i medici hanno scansionato la cavità addominale del Bodybuilder, hanno notato che il fegato dell’uomo era più grande del normale. Una biopsia ha mostrato che il fegato del aveva cicatrici in alcuni punti. I dotti biliari, che trasportano i sali biliari all’intestino, erano ostruiti.
Nelle settimane successive, i medici hanno monitorato quattro marker del danno epatico nel sangue del Bodybuilder. La figura seguente mostra che le condizioni del fegato dell’uomo sono lentamente migliorate.
Come avevo già riportato nell’articolo di giugno, secondo uno studio del 2012 condotto dai produttori del LGD-4033, questo SARM non è significativamente dannoso per il fegato. Ma in questo studio, i soggetti non hanno ricevuto più di 1mg/die di LGD-4033.[2]
Le aziende che vendono SARM online e alcuni guru del bodybuilding raccomandano dosi significativamente più elevate di 1mg/die. Ad esempio, il paziente del quale si è parlato in questo articolo ha assunto dieci volte la dose più alta testata nello studio del 2012. Con tutta probabilità, un dosaggio di LGD-4033 di tale entità o superiore rappresenta uno stress epatico eccessivo, in specie se l’utilizzatore presenta una marcata sensibilità e manca di una efficace epatoprotezione (comunque non garante di immunità da effetti collaterali a livello epatico).
Alcuni utilizzatori di LGD-4033 hanno pubblicato il loro esame ematico sui forum presenti in rete. Sembra che non mostrino segni di danno epatico, ma l’affidabilità di certi dati è assai scarsa.
Forse il Bodybuilder in questione stava usando un prodotto contaminato o fake. Non tutti i SARM negli store online sono prodotti con le giuste misure di controllo, come riportato da una recente ricerca inglese e americana.[3]
E’ anche possibile che il Bodybuilder del caso studio stava usando altre sostanze oltre al solo LGD-4033 e non ne ha fatto menzione ai medici che lo hanno preso in cura. Le possibilità sono diverse ma ciò che è sufficientemente certo è che l’uso di LGD-4033 ad alte dosi, per vie metaboliche intrinseche, è un potenziale fattore causale per stress e danno epatico.
Nonostante il terrorismo diffuso da alcuni scientisti riguardo alla “malvagità assoluta” del IGF-1 e della sua correlazione con il cancro (esistente ma contestualizzabile), la terapia sostitutiva dell’Ormone della Crescita (GH) sembra non aumentare il rischio di cancro; sembrerebbe che, addirittura, ne riduca il rischio. Ciò è stato evidenziato in una meta-analisi pubblicata da endocrinologi cinesi della Zhejiang University College of Medicine nell’Open Journal of Endocrine and Metabolic Diseases.[1]
I ricercatori hanno esaminato 10 studi pubblicati in precedenza che hanno seguito adulti trattati con Ormone della Crescita per diversi anni. Ai soggetti è stato diagnosticato un deficit dell’ormone della crescita negli adulti. Ovviamente, e mi riferisco al lettore nella media che troppo velocemente trae conclusioni inesatte, gli studi potrebbero non dire molto sugli effetti che gli atleti supplementati farmacologicamente possano incorrere nel contesto dell’incidenza di sviluppo del cancro.
Nella tabella sottostante troverete maggiori informazioni sugli studi utilizzati.
Negli studi esaminati, i soggetti trattati con il GH non hanno sviluppato il cancro in maniera maggiore rispetto ai soggetti dei gruppi di controllo. Da quanto è emerso, per lo meno dagli studi presi in esame, l’Ormone della Crescita ha persino ridotto il rischio di cancro.
I ricercatori hanno però trovato presenza di bias. Ciò significa che sembra che non siano stati pubblicati studi con risultati meno interessanti. Tuttavia, il bias sembrava essere modesto ed i ricercatori sospettano che l’inclusione degli studi potenzialmente mancanti nella loro meta-analisi non ne altererebbe realmente il risultato.
I ricercatori concludono dicendo che la loro analisi corrobora le prove di studi precedenti che dimostrano che la terapia sostitutiva dell’Ormone della Crescita nei pazienti con deficit di questo ormone in età adulta non vedrebbero aumentare il loro rischio di sviluppare il cancro; invece, potrebbe addirittura diminuire il rischio. I risultati hanno suggerito che la terapia sostitutiva dell’Ormone della Crescita nei pazienti con deficit dell’ormone in età adulta era sicura.
Nota: i dosaggi terapici variano nettamente a seconda dell’età e delle finalità della terapia. Sono generalmente dosaggi inferiori a quelli utilizzati a scopo estetico-prestativo. Pertanto, il presente articolo non ha assolutamente lo scopo di far passare un dato che ad oggi non è statisticamente comprovato.
A differenza del trattamento pediatrico con GH, spesso dosato in microgrammi / chilogrammi di peso corporeo / giorno, il dosaggio sostitutivo del GH attualmente raccomandato negli adulti è individualizzato indipendentemente dal peso, tenendo conto dell’età del paziente, del sesso e dello stato degli estrogeni (Johannsson et al., 1997a; Hoffman et al., 2004b). L’inizio della terapia a basse dosi (dose totale 0,2-0,4 mg / die SC) riduce la probabilità di sviluppare effetti collaterali comuni come rigidità articolare, artralgie, mialgie, parestesie ed edema periferico, con ritenzione di liquidi. La dose deve essere titolata a intervalli di 6-8 settimane in base alla risposta clinica, evitando effetti collaterali e monitorando i livelli sierici di IGF-I. Si desidera raggiungere un obiettivo nella metà superiore dell’intervallo normale del IGF-I corretto per l’età del paziente. È ragionevole iniziare con dosi più elevate (0,4-0,5 mg / die) nei pazienti di età inferiore a 30 anni, ma i pazienti più anziani (di età superiore a 60 anni) dovrebbero iniziare con dosi più basse (0,1-0,2 mg / die) e titolato più lentamente per ridurre al minimo il verificarsi di effetti collaterali. Alcuni pazienti con AGHD ad esordio infantile con IGF-I pretrattamento molto basso possono sviluppare effetti collaterali da GH ma non raggiungere livelli medi normali nonostante dosi elevate di quest’ultimo.
Le donne che assumono una terapia sostitutiva degli estrogeni per via orale possono richiedere dosi più elevate per la GHRT, presumibilmente perché l’estrogeno orale inibisce la produzione e la secrezione di IGF-I nel fegato con effetto di primo passaggio (Weissberger et al., 1991). Di solito non è necessario alcun aggiustamento della dose nei pazienti che assumono dosi moderate di estrogeni transdermici.
La Niacina è largamente utilizzata dagli atleti supplementati chimicamente, in special modo da coloro i quali usano molecole con un potenziale negativo marcato sui lipidi ematici. Ma come spesso capita, gli utilizzatori non conoscono a sufficienza le caratteristiche di ciò che assumono, e questa essenziale vitamina del gruppo B (B3) non è da meno. Per la maggior parte degli individui tanto basta sapere che una sua integrazione si traduce in livelli migliorati di Colesterolo e Trigliceridi. Purtroppo, però, si trascurano caratteristiche importanti la cui conoscenza può fare la differenza tra un uso più o meno funzionale per la salute sistemica. Infatti, un effetto collaterale dell’integrazione di Niacina è un peggioramento della resistenza all’insulina, cosa che limita i benefici di tale supplementazione sulla salute cardiovascolare se non vengono prese adeguate precauzioni.
Prima di correre a defenestrare in preda al panico la vostra Niacina, leggete con attenzione (e comprendete) le informazioni che seguono…
Introduzione alla Niacina (vitamina B3)
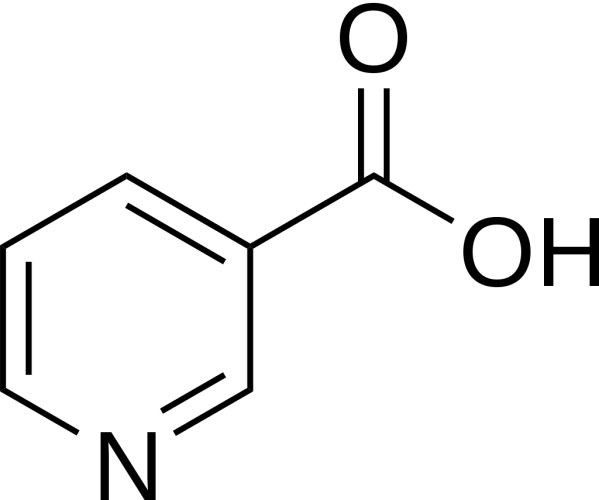
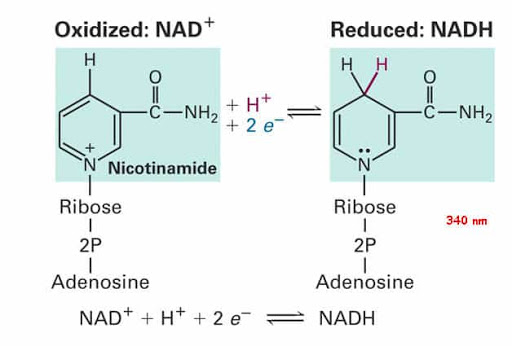
Niacina
La Niacina, nota anche come Acido Nicotinico, è un composto organico e una forma di vitamina B3, un micronutriente essenziale per l’essere umano. [1] La Niacina ha formula bruta C6H5NO2 e appartiene al gruppo dell’acido piridinecarbossilico.[1] Come precursore di NAD e NADP, la Niacina è coinvolta nella riparazione del DNA.[2] La Niacina viene assunta attraverso la dieta da una varietà di alimenti interi e trasformati, con il più alto contenuto in alimenti confezionati fortificati, carne, pollame, pesce rosso come tonno e salmone, con minori quantità nelle noci, legumi e semi. [1] [3] La Niacina come integratore alimentare viene anche utilizzata per trattare la pellagra, una malattia causata da una sua carenza. Segni e sintomi includono lesioni della pelle e della bocca, anemia, mal di testa e stanchezza.[4] Molti paesi richiedono la sua aggiunta alla farina di grano o ad altri cereali, riducendo così il rischio di pellagra.[1][5] Come vitamina, le raccomandazioni di dosaggio giornaliero indicate in diversi paesi sono 14-18mg/die per gli adulti, quota sufficiente per soddisfare le esigenze delle persone sane. [6] [7] [8]
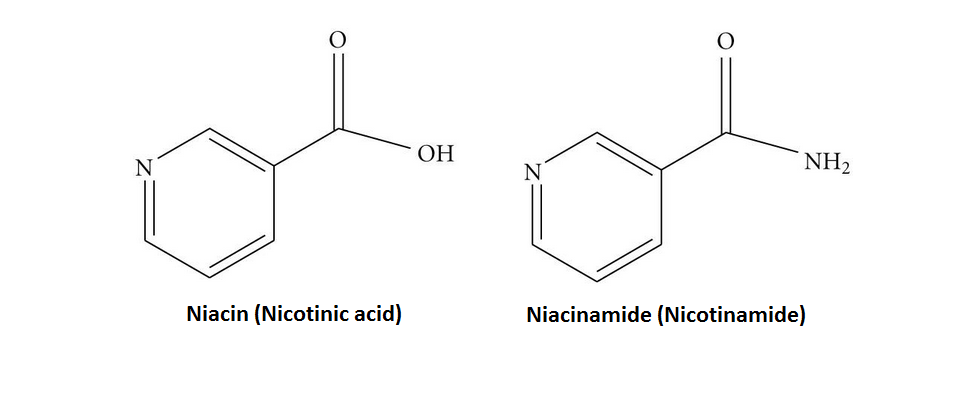
Sebbene la Niacina e la Nicotinamide (Niacinamide) siano identiche nella loro attività vitaminica, la Nicotinamide non ha gli stessi effetti farmacologici, modificanti i lipidi o gli effetti collaterali della Niacina, cioè quando la Niacina assume il gruppo -amide, non riduce il Colesterolo né causa vampate di calore.[9][10] La Nicotinamide è raccomandata come trattamento per la carenza di Niacina poiché può essere somministrata in quantità correttive senza causare l’effetto negativo del rossore.[11]
La Niacina è anche un farmaco di prescrizione. Quantità molto superiori all’assunzione dietetica raccomandata per le funzioni vitaminiche ridurranno i Trigliceridi nel sangue e le lipoproteine a bassa densità (LDL-C) e aumenteranno le lipoproteine ad alta densità (HDL-C). Ne esistono due forme: Niacina a rilascio immediato e a rilascio prolungato. Le quantità iniziali di prescrizione sono di 500mg/die, con possibilità di essere aumentate nel tempo fino a raggiungere l’effetto terapeutico ricercato. Le dosi a rilascio immediato possono arrivare fino a 3g/die; quelle a rilascio prolungato fino a 2g/die. [12] Nonostante i comprovati cambiamenti lipidici, la Niacina non è stata trovata utile per ridurre il rischio di malattie cardiovascolari nei soggetti già in trattamento con statine. [13] Una review del 2010 aveva concluso che l’efficacia della Niacina si osservava in mono-terapia, [14] ma una review del 2017 che incorporava il doppio del numero degli studi ha concluso che la Niacina su prescrizione, pur influenzando i livelli lipidici, non riduceva la mortalità per tutte le cause, la mortalità cardiovascolare, gli infarti del miocardio, né ictus fatali o non fatali. [15] È stato dimostrato che la Niacina da prescrizione provoca epatotossicità [16] e aumenta il rischio di diabete di tipo 2. [17] [18] Le prescrizioni di Niacina negli Stati Uniti avevano raggiunto il picco nel 2009, a 9,4 milioni, in calo a 1,3 milioni entro il 2017.[19]
Niacina, flusso ematico, pressione e vasodilatazione
Uno studio sulla supplementazione di Niacina che ha valutato il flusso sanguigno dell’avambraccio non è riuscito a trovare un effetto significativo fino a 1g al giorno somministrati nel corso di due settimane in soggetti altrimenti sani, [20] e 1.5g di Niacina a rilascio prolungato negli uomini con sindrome metabolica non sono riusciti a influenzare la dilatazione flusso- mediata (FMD). [21] Un altro studio non è riuscito a trovare un effetto significativo in un intero gruppo di pazienti affetti da afta epizootica, mentre in un gruppo di pazienti con malattia coronarica ha riscontrato un miglioramento in un sottogruppo con bassi livelli HDL-C. [22]
In soggetti con bassi livelli di HDL-C, è stato osservato che 1g di Niacina a rilascio prolungato per una settimana aumenta il flusso sanguigno (via FMD) del 4,5%; questo meccanismo non era correlato alle Prostaglandini, poiché il Laropiprant (un inibitore della Prostaglandine D2) non ha influenzato l’effetto. [23] Questo effetto ha anche coinciso con un aumento della bilirubina indiretta (ma non totale) del 62%. [23] Poiché la bilirubina del acido biliare è un antiossidante endoteliale, [24] e poiché i benefici della niacina sulla funzione endoteliale in questo studio sono stati ritenuti dipendenti dall’ossido nitrico, [23] è stato ipotizzato che un effetto conservativo della bilirubina sulla biodisponibilità dell’ossido nitrico sia alla base della beneficio osservato. Sia l’aumento della bilirubina che il miglioramento del flusso sanguigno si sono dissipati una settimana dopo l’interruzione della Niacina.[23]
Laropiprant
I soggetti che in precedenza avevano subito infarto del miocardio, a seguito del trattamento con Niacina (con Laropiprant) hanno riscontrato un aumento del flusso sanguigno dipendente dall’ossido nitrico (FMD) dopo dodici settimane di terapia insieme a un miglioramento della vasodilatazione indotta da nitroglicerina, entrambe non correlate con alterazioni dei trigliceridi. [25] Miglioramenti simili nel flusso sanguigno sono stati osservati in pazienti con infezione da HIV e con bassi livelli di HDL-C trattati con la sola Niacina. [26]
Prostaglandine D2 (PGD2)
È noto che la Niacina influenza il diametro dei vasi sanguigni, in particolare per via della sua reazione vasodilatativa cutanea (allargamento dei vasi nella pelle), che ha portato a ipotizzare che potrebbe influenzare la pressione sanguigna aumentando il diametro delle arterie e vene. Tuttavia, una review [27] ha notato che un possibile effetto di riduzione della pressione arteriosa della Niacina è indipendente dalla Prostaglandine che media il rossore, nota come PGD2.
È stato osservato che le infusioni di Niacina riducono acutamente la pressione sanguigna negli ipertesi senza alcun effetto nei soggetti con pressione sanguigna normale ed è stata associata ad un aumento della gittata cardiaca e della frequenza cardiaca che era simile in entrambi i gruppi. [28] Un altro studio ha confermato questo risultato, scoprendo che la pressione arteriosa ambulatoriale di 24 ore non sembra essere influenzata da un supplemento di Niacina fino a 1g nell’arco di due settimane in soggetti altrimenti sani. [20]
In termini di effetti della Niacina in cronico sulla pressione sanguigna, una review [27] che ha valutato gli studi che hanno misurato la pressione sanguigna negli ipertesi [29] [30] [31] [32] non ha notato alcun effetto statisticamente significativo nella riduzione della pressione sanguigna associata alla supplementazione di Niacina, sebbene questi studi in quanto a metodologie di misurazione sulle variazioni della pressione sanguigna non fossero ideali secondo gli autori della review. Tuttavia, la review ha osservato che in un ampio studio (il Coronary Drug Project), che inizialmente non è riuscito a trovare alcuna influenza della terapia con Niacina sulla pressione arteriosa, [32] ha osservato variazioni sensibili soltanto sui soggetti con sindrome metabolica. Questi presentavano un lieve riduzione di 2,2mmHg della pressione arteriosa sistolica con una moderata riduzione di 2,9mmHg della pressione diastolica. [33] Un’analisi post-hoc di un altro studio clinico [34] ha rilevato che la pressione arteriosa sistolica è stata abbassata di 2,2mmHg e la pressione sistolica di 2,7 rispetto al placebo nei pazienti dislipidemici trattati per 24 settimane. [35]
Niacina, Trigliceridi, Colesterolo e Aterosclerosi
Apolipoproteina B
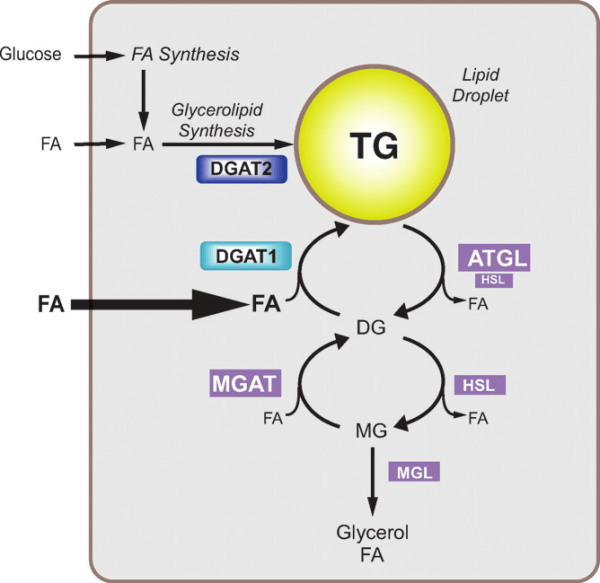
La Niacina sembra abbassare i trigliceridi nel sangue inibendo sia la sintesi degli acidi grassi sia la loro esterificazione epatica per formare i trigliceridi, il che aumenta il tasso di degradazione dell’apolipoproteina B riducendo la sua secrezione dalle cellule epatiche. [36] Un meccanismo con cui la Niacina fa questo è attraverso l’inibizione diretta e non competitiva della diacilglicerolo aciltransferasi 2 (DGAT2), l’enzima finale nella sintesi dei trigliceridi nelle cellule epatiche, senza inibizione della DGAT1. [37]
Si è visto che gli effetti della Niacina sulla sintesi dei trigliceridi influenzano i livelli sierici di lipoproteine a densità molto bassa (vLDL-C), dove la terapia con Niacina per 16 settimane in soggetti con malattia del fegato grasso non alcolica (NAFLD) sembra ridurre le vLDL-C nel siero così come i complessi con trigliceridi (vLDL-TG) e apolipoproteina B (vLDL-ApoB) rispetto al placebo e con una potenza paragonabile al fenofibrato. [38] La Niacina lo fa riducendo la secrezione epatica di vLDL-C, sebbene ciò non aumenti la quantità di trigliceridi nel fegato anche nello stato di NAFLD. [38]
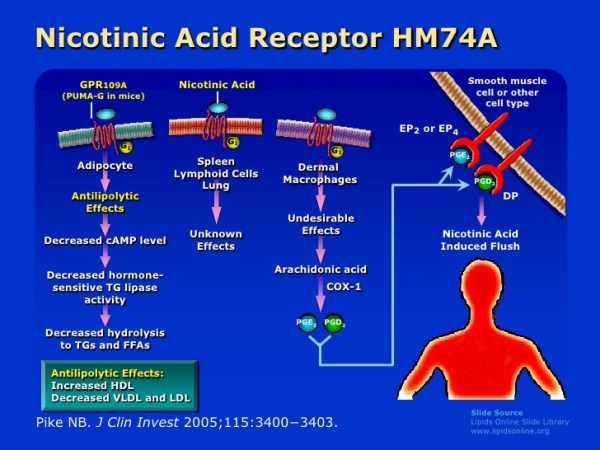
Oltre ai suoi effetti sul fegato, la Niacina può anche sopprimere il rilascio di acidi grassi liberi dal tessuto adiposo [39] che normalmente verrebbero reesterificati come trigliceridi nel fegato e quindi secreti via vLDL. [40] Tuttavia, questo meccanismo specifico, che è mediato dal recettore HM74A, [39] non sembra essere rilevante per le proprietà riducenti dei trigliceridi della Niacina. [41]
I benefici sui livelli di trigliceridi possono verificarsi entro una settimana dall’inizio della supplementazione con Niacina a rilascio prolungato (1g), sebbene in misura minore di circa il 4%. [23]
L’integrazione di 1.5-2g di Niacina a rilascio prolungato per due anni con follow-up di un anno nelle persone in terapia con statine caratterizzate da bassi livelli di HDL-C ha mostrato una riduzione dei trigliceridi del 28,6% (statina da sola dell’8,1%). [42]
Esiste un fenomeno noto come “rimbalzo degli acidi grassi” associato alla supplementazione di Niacina, in quanto l’azione iniziale del composto sul suo recettore (HM74A) nel tessuto adiposo può determinare una minore lipolisi e una minore secrezione di acidi grassi non esterificati (NEFA) nel sangue [43] e una migliore conservazione adiposa; [44] si tratta di fenomeni prontamente reversibili in quanto in un giorno di esposizione continua vi è un aumento netto del NEFA piuttosto che la sua soppressione [45] [46] [47] e alterazioni nel NEFA possono non riflette alterazioni dei trigliceridi.
Il primo meccanismo pensato per spiegare il miglioramento del profilo sierico di colesterolo in seguito alla supplementazione di Niacina è stato attraverso la riduzione del rilascio di acidi grassi non esterificati (NEFA) dai tessuti, che non è più considerato un probabile meccanismo in quanto l’integrazione di niacina in cronico è associata ad un aumento, piuttosto che alla soppressione, di NEFA mentre il recettore HM74A appare superfluo in termini di effetti della Niacina nei topi con altri ligandi del HM74A (Acipimox [48] e MK-0354 [49]) che si sono mostrati rispettivamente meno efficaci o inefficaci sul colesterolo. Attualmente si ritiene che l’influenza della Niacina sui NEFA nel siero non sia un fattore determinante nel modo in cui influenza i livelli di colesterolo nel corpo, con le teorie attuali che ipotizzano che il fattore sia determinato dalla sua sintesi e dal suo tasso di catabolismo.
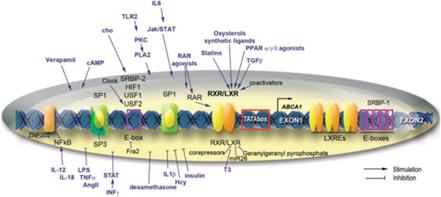
Il primo potenziale meccanismo prevede la sintesi di HDL-C nel fegato attraverso l’aumento della trascrizione del gene ABCA1 (che dipende dal legame LXRα alla regione del promotore DR4 di questo gene). [50] L’attività di ABCA1 promuove la “lipidazione” della principale proteina dell’HDL nota come apolipoproteina AI (ApoAI) aumentando il tasso che associa ai fosfolipidi e al colesterolo, [51] [52] un passaggio obbligatorio nella sintesi dell’HDL-C che è aumentato di 500-1000µM con Niacina in vitro. [50] Questo meccanismo non è stato confermato, poiché mentre l’ApoAI può essere aumentato parallelamente all’aumento dell’HDL-C in soggetti trattati con Niacina e con livelli di HDL-C bassi di base, [53] LXRα sembra richiedere un coattivatore (PPARγ) per esercitare questi effetti, [54] che è attivato dal recettore della Niacina. [55] Tuttavia, l’attività del recettore della Niacina non è stata richiesta per i suoi effetti sui livelli di colesterolo, suggerendo che altri meccanismi potrebbero essere rilevanti.
PPARγ
L’altra teoria relativa alla sintesi di HDL dalla Niacina afferma che ciò dipenda dalla proteina di trasferimento dell’estere del colesterolo (CETP) nonostante la riduzione del colesterolo totale e dei trigliceridi non richieda per entrambe questa proteina. [56] [57] CETP è una proteina che facilita il trasferimento di lipidi tra diverse lipoproteine (generalmente donando un trigliceride da vLDL a HDL e prendendo un estere di colesterolo in un processo noto come trasporto inverso di colesterolo. [58]) La Niacina riduce l’espressione di CETP nel fegato e la sua attività nel sangue dei topi; [56] una riduzione del CETP aumenta la quantità di HDL-C nel sangue poiché i tassi di catabolismo dell’HDL / LDL riflettono l’attività del trasporto inverso del colesterolo e raggiungono rapidamente l’equilibrio, [59] e se il CETP è ridotto allora sarebbe necessario più HDL per normalizzare i tassi di trasporto inverso del colesterolo. Questo meccanismo può anche essere correlato a LXRα, poiché mentre un eteromero di LXRα con il recettore nucleare di vitamina A (RXR) attiva l’elemento DR4 aumenta la CETP [60] la Niacina agevola l’eterodimerizzazione di LXRα e PPARγ che attiva ancora DR4, ma in un modo che promuove l’efflusso di colesterolo. [61-44] Questa eterodimerizzazione competitiva [62] non è stata ancora dimostrata sperimentalmente, e lo studio che ha utilizzato dosi di Niacina da 2g nell’uomo non è riuscito a trovare un’influenza sull’attività del CETP nel siero nonostante un aumento dell’HDL. [63]
L’ultimo potenziale meccanismo per l’aumento dell’HDL non consiste nel suo incremento di sintesi ma piuttosto nel preservare il colesterolo HDL già sintetizzato arricchito con apoAI, riducendo il tasso in cui la lipoproteina viene assunta nelle cellule epatiche nonostante la donazione di colesterolo dall’HDL a queste cellule sia inalterata a causa della riduzione dell’espressione del recettore (catena beta sintasi ATP) che normalmente trasporta l’HDL nella cellula. [64] Questa ipotesi funziona meglio con le osservazioni che suggeriscono che il ridotto catabolismo dell’HDL è il principale fattore determinante dei suoi livelli più elevati, [65] e influenza anche l’apoA1 poiché la sua clearance dal sangue e l’assorbimento da parte dei reni sono ridotti. [66]
Una supplementazione di Niacina a rilascio prolungato (1g) della durata di una settimana in soggetti con bassi livelli di HDL-C non sembra essere sufficiente da aumentare sensibilmente i livelli totali di HDL-C, sebbene sia stata notata una riduzione della dimensione media delle particelle; [23] le variazioni di HDL -C possono mediare un miglioramento della vasodilatazione dipendente dall’ossido nitrico, sebbene sia stato anche osservato un aumento della bilirubina indiretta. [23]
L’integrazione prolungata di Niacina nei diabetici è associata ad un aumento della quantità e delle dimensioni particellari dell’HDL-C (32,7%) mentre le particelle di dimensioni più piccole sono diminuite (8,2%). [67]
È stato osservato che la Niacina conferisce un effetto protettivo sulla mortalità cardiovascolare poiché una metanalisi [68] ha osservato che negli studi su soggetti con malattia coronarica la terapia con Niacina era associata a un minor rischio di rivascolarizzazione dell’arteria coronarica (RR di 0,31; IC al 95% di 0,15-0,63), infarto miocardico non fatale (RR di 0,72; IC al 95% di 0,60-0,86) e attacco ischemico transitorio (RR di 0,76; IC al 95% di 0,61-0,94) mentre la riduzione della mortalità complessiva non è riuscita a raggiungere significatività statistica (RR 0,883; IC 95% di 0,773-1,008). I sette studi inclusi in questa meta-analisi [32] [29] [31] [30] (e un follow-up [69]) hanno totalizzato 5137 pazienti che utilizzavano anche vari prodotti farmaceutici della classe di statine e fibrati .
In uno studio i cui partecipanti erano in terapia con statine e avevano bassi livelli di colesterolo HDL è stato rilevato che 1.5-2g di Niacina a rilascio prolungato sono stati in grado di fornire benefici additivi nel miglioramento dell’HDL-C (20%) e nella riduzione dell’LDL-C (17%) rispetto al placebo, sebbene per quanto riguarda l’endpoint clinico predeterminato (morte o ricovero in ospedale) sia la Niacina che il placebo avevano una uguale quantità di responder. [70] Questo studio ha rilevato un’alta percentuale di pazienti con sindrome metabolica (80%) e commenti [71] hanno suggerito che a causa di una possibile capacità della Niacina a rilascio prolungato di deteriorare l’insulino-resistenza [72] che i suoi benefici potrebbero essere compensati da questo effetto avverso, mentre lo studio stesso ha suggerito che i benefici delle statine hanno sostituito i benefici della Niacina.
Mentre uno studio precedente che utilizzava alte dosi di Niacina a rilascio immediato (3g) ha riscontrato una riduzione della morte del 14% rispetto al placebo insieme a una riduzioni del colesterolo totale, [32] ed è stato osservato che questa riduzione è simile per grandezza agli studi che combinano statine con placebo.
Studi in vitro suggeriscono che la Niacina potrebbe in teoria prevenire la formazione di placche aterosclerotiche riducendo l’infiammazione e il danno alla parete endoteliale attraverso diversi meccanismi. Limitate ricerche su animali hanno mostrato che la Niacina nella dieta, a concentrazioni paragonabili a quelle utilizzate per ridurre il colesterolo, riduce la deposizione della placca sulla parete dell’arteria e ritarda l’aterosclerosi.[73][74][75][76][77][78][79][80]
Niacina e sue interazioni con il metabolismo del glucosio
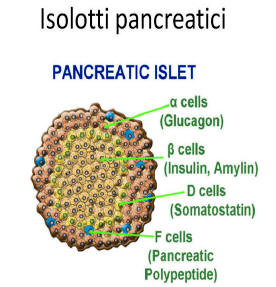
L’assunzione prolungata di Niacina è stata osservata causare una riduzione della sensibilità all’insulina, causando un aumento compensativo della produzione di insulina da parte delle cellule β del pancreas per mantenere i livelli di glucosio nel sangue. [81] La Niacina non sembra avere effetti diretti sulle cellule β pancreatiche, tuttavia, poiché la perfusione negli isolotti pancreatici (isole di Langerhans) di ratto isolati con Niacina in vitro non ha influenzato la secrezione di insulina. [82] Ciò indica che la Niacina aumenta la produzione di insulina mediante un meccanismo indiretto, secondario a causare insulino-resistenza periferica. È stato osservato che la supplementazione induce resistenza all’insulina a dosi comprese tra 500mg e 1g, che rientrano nell’intervallo di dosaggio che conferisce effetti di riduzione del colesterolo. [83]
In particolare, sembra che sia necessaria una supplementazione cronica di Niacina per aumentare la produzione di Insulina, poiché in uno studio è stato dimostrato che la supplementazione acuta riduce i livelli di questo peptide in soggetti altrimenti sani prima di un picco dopo un giorno, [84] mentre altri studi in acuto hanno notato un effetto minimo o nullo sui livelli di Insulina. [85] [86] [87] [88]
Gli effetti dell’integrazione cronica di Niacina sui livelli di Insulina possono anche dipendere dalla popolazione. È stato osservato che la Niacina provoca iperinsulinemia in soggetti che invecchiano altrimenti sani [83] (1g / die) ed è stato dimostrato che quasi raddoppiano i livelli di Insulina nei soggetti con NAFLD (2g / giorno [89] [90]). Nei pazienti con sindrome metabolica, l’integrazione di Niacina a 6 settimane di somministrazione alla dose di 1.5g / die ha aumentato i livelli di Insulina del 30%. [91]
Nei soggetti obesi con malattia del fegato grasso non alcolico (NAFLD), l’integrazione giornaliera di Niacina a rilascio prolungato (titolata fino a 2g) per 16 settimane sembrava aumentare lo stato di resistenza all’insulina nel fegato, nei muscoli e nel tessuto adiposo [89] con un effetto inibitorio sulle azioni dell’Insulina nel fegato notate negli uomini non diabetici con dislipidemia. [92]
Adiponectina
Negli uomini adulti con sindrome metabolica, è stato osservato che la Niacina a rilascio prolungato alla dose di 1.5g ostacola in modo significativo la sensibilità all’Insulina, valutata dall’HOMA-IR (42%), che è stata associata ad un aumento dell’Insulina sierica nonostante un aumento dell’Adiponectina sierica. [91] Questo è stato notato anche in un altro studio (aumento del 22% dell’HOMA-IR), in cui l’Aspirina assunta insieme alla Niacina non ha impedito la comparsa di una ridotta sensibilità all’Insulina. [93]
Questo effetto può persistere in soggetti altrimenti sani, poiché i soggetti trattati con 1g di Niacina per due settimane a cui veniva somministrato un clamp iperinsulinaemico-euglicemico richiedono meno glucosio per mantenere l’omeostasi, il che è indicativo di una riduzione dell’assorbimento del glucosio (attraverso un aumento dell’Insulino-resistenza). [94]
La resistenza all’Insulina indotta dalla Niacina è stata inizialmente attribuita a un effetto di rebound nel tessuto adiposo in cui un aumento del rilascio di acidi grassi non esterificati (NEFA) da parte della Niacina compromette gli effetti della segnalazione dell’Insulina. [95] [96] Ciò è plausibile, poiché la resistenza all’Insulina può essere indotta con infusione di NEFA in 24 ore nei roditori. [97] Altre fonti suggeriscono che la resistenza all’Insulina non è associata al rebound del NEFA, poiché i soggetti con NAFLD che sperimentano resistenza all’Insulina dalla terapia con Niacina non hanno necessariamente un aumento del NEFA nel siero. [89].
Modello ipotetico per i ruoli intracellulari del DGAT1 e DGAT2.
Un’altra possibile opzione è che la Niacina può inibire in modo non competitivo l’enzima noto come diacilglicerolo aciltransferasi 2 (DGAT2) con un IC50 di 100 µM (potenza simile a circa 300 µM). [98] L’inibizione di questo enzima non causa di per sé resistenza all’insulina con la somministrazione di Niacina, [92] ma poiché il DGAT catalizza il primo stadio della sintesi dei trigliceridi, la sua inibizione può favorire l’accumulo di diacilglicerolo (DAG) che è la molecola che si ritiene spieghi parzialmente la resistenza all’insulina data dalla Niacina. [92] Poiché l’aumento del DAG nelle cellule del fegato sopprime la segnalazione dell’Insulina, [99-162] l’inibizione mediata dalla Niacina del DGAT2 provoca insulino-resistenza, [98] [89] ostacolando così la capacità dell’Insulina di sopprimere la sintesi di glucosio e promuovendo indirettamente uno stato di iperglicemia.
Sebbene l’integrazione cronica di alte dosi di Niacina riduca la sensibilità all’Insulina, ciò non è associato a variazioni dei livelli di glucosio a digiuno. [90] Ciò può essere spiegato da un aumento compensativo della sintesi di Insulina che contrasta la resistenza alla stessa, lasciando sostanzialmente invariati i livelli di glucosio nel sangue. [81]
L’attivazione del recettore della Niacina (HM74A) da parte di alcuni altri agonisti sembra ridurre rapidamente il glucosio sierico nei diabetici migliorando la sensibilità all’Insulina [100] o comunque migliorando i tassi di smaltimento del glucosio. [101] Ciò indica che lo stesso recettore della Niacina può avere effetti benefici sul metabolismo del glucosio e che la resistenza all’Insulina indotta dalla Niacina non si verifica tramite l’attivazione del HM74A.
Quando si osserva il muscolo scheletrico, è stato dimostrato che la terapia con Niacina induce resistenza all’Insulina in questo tessuto in soggetti obesi con NAFLD (2g al giorno nel corso di 16 settimane). Uno studio svolto su ratti a digiuno (il digiuno aumenta la concentrazione plasmatica di acidi grassi non esterificati (NEFA), similmente alla somministrazione di Niacina [102-135] e diminuisce il glicogeno del muscolo scheletrico [103]) in cui sono stati accuratamente somministrati 20mg/kg di Niacina ha mostrato che il glicogeno nel soleo era ridotto mentre il gastrocnemius e il fegato non sono stati influenzati. [103]
Metilgliossale
Quando il processo di glicazione è testato in vitro, la Niacina ha avuto solo effetti inibitori minori sulla glicazione dell’albumina sierica bovina da un noto agente glicante (Metilgliossale [104]) nonostante altri antiossidanti testati come lo Zinco (10-25 µg / mL) avessero più potenti benefici. [105]
È importante sottolineare che qualsiasi effetto della Niacina sulla glicazione in vitro deve essere interpretato con l’avvertenza che la Niacina riduce la sensibilità all’Insulina. Mentre la resistenza all’Insulina indotta dalla Niacina è ben compensata in soggetti sani giovani, lasciando sostanzialmente invariati i livelli di glucosio nel sangue, [81] la compensazione delle cellule β del pancreas negli individui più anziani o in quelli con ridotta tolleranza al glucosio era incompleta in uno studio, [83] causando aumenti nei livelli ematici di glucosio. Pertanto, la misura in cui la Niacina possa influenzare la glicazione in vivo non è chiara e probabilmente dipendente dalla popolazione.
Obesità e massa grassa
L’Adiponectina, un’adipochina nota per migliorare la sensibilità all’Insulina, per essere cardioprotettiva e ritenuta anche antiobesogena, [106] è aumentata in risposta all’attivazione mediata dalla Niacina del recettore HM74A nei topi. [107] La produzione di Adiponectina indotta dalla Niacina è stata rapida in questo studio, aumentando i livelli di questa adipochina del 37% entro 10 minuti da una dose di 30mg / kg per iniezione. I livelli sierici hanno raggiunto il picco dopo 60 minuti e sono rimasti elevati al di sopra del basale fino a 24 ore dopo la somministrazione. [107]
Leptina
È noto anche che la Leptina è aumentata in seguito alla somministrazione di Niacina nell’uomo [91], il che si ritiene si verifichi tramite un meccanismo simile poiché l’agonista farmaceutico HM74A Acipimox induce anch’esso la secrezione di Leptina dal tessuto adiposo in vitro [108] e in vivo. [109]
È stato osservato che la supplementazione di Niacina nel corso di sei settimane negli uomini obesi aumenta l’Adiponectina sierica del 43-56%, con circa metà dell’aumento rappresentato dalla forma ad alto peso molecolare [93] [91] insieme a un aumento del 26,8% della Leptina [91 ] senza cambiamenti osservabili nella Resistina. [91] L’Adiponectina è stata osservata aumentare di circa il 30% in soggetti obesi con NAFLD in risposta alla terapia con Niacina (fino a 2g al giorno), che era correlata con un aumento dell’Insulino-resistenza, [90] portando all’ipotesi che i due meccanismi siano intrecciati, forse come risposta adattativa. [90]
Resistina
Lo “spillover” degli acidi grassi risultante da una conservazione inefficiente del grasso dopo un pasto aumenta i lipidi sierici non esterificati (NEFA), [110] che influenzano negativamente la sensibilità all’Insulina epatica, aumentando la produzione di VLDL e potenzialmente svolgono un ruolo causale nella steatosi epatica. [111] [112] La somministrazione in acuto di Niacina (285 mg per via endovenosa) nell’uomo durante l’alimentazione ha dimostrato di ridurre lo spillover degli acidi grassi, promuovendo l’assorbimento del grasso alimentare nel tessuto adiposo e riducendo i Trigliceridi sierici e i NEFA. [113]
Al contrario, è stato osservato che un trattamento prolungato con Niacina, noto per favorire la resistenza all’Insulina nell’uomo, induce la resistenza all’Insulina adipocitaria, [114] che favorirebbe lo spillover degli acidi grassi, aumentando i livelli sierici di NEFA.[115]
Glucosio-6-fosfato deidrogenasi (G6PD)
È stato osservato che la Nicotinamide sopprime la differenziazione degli adipociti 3T3-L1 in modo dipendente dalla concentrazione con un range superiore a 10mM (il valore ED50), raggiungendo la soppressione completa a 20mM dopo nove giorni. [116] Si ritiene che ciò sia correlato a un effetto inibitorio sulla poli (ADP-ribosio) sintetasi, [116] che la Nicotinamide inibisce a 50µM mentre la Niacina non lo fa. [117] Quando aggiunta dopo differenziazione e in condizioni di glucosio elevato, la Nicotinamide sembra inibire il glucosio-6-fosfato deidrogenasi (G6PD) e prevenire il normale stress ossidativo. [118]
Il recettore dell’Acido Nicotinico è espresso negli dipociti in cui la sua attivazione sopprime l’adenilato ciclasi. [119] Questo effetto sembra essere circa il 30% più efficace negli adipociti rispetto ad altre linee cellulari (milza). [120] Poiché l’attivazione di questo recettore inibisce l’adenilato ciclasi, [119] e i fenolici che agiscono su di esso riducono anch’essi i tassi di lipolisi, [35] l’effetto complessivo dell’Acido Nicotinico sarebbe quello di ridurre la lipolisi negli adipociti, almeno a breve termine.
A lungo termine, tuttavia, il recettore dell’Acido Nicotinico può essere desensibilizzato con esposizione cronica a un agonista [121] e uno studio sui topi ha evidenziato che gli adipociti che sono diventati insulino-resistenti dopo la terapia con Niacina hanno mostrato una maggiore reattività dei recettori adrenergici (β1 e β2) all’aumentare dei livelli di cAMP nella cellula adiposa, [114] (il cAMP viene normalmente soppresso dalla Niacina che agisce sul recettore GRP109A [119]). Ciò potrebbe essere stato correlato alla sottoregolazione dei geni mediata dalla Niacina nella via di segnalazione dell’Insulina incluso il PDE3B, che normalmente degrada il cAMP, [114] una potenziale risposta adattativa nelle cellule adipose che è stata osservata avere la funzione di normalizzare i tassi di lipolisi (nei ratti sotto l’infusione di Niacina) . [97]
Un piccolo studio su sette partecipanti altrimenti sani che assumevano Niacina a 500mg/die, e aumentando la dose a 2g nel corso di due settimane ha mostrato una riduzione dei tassi di ossidazione dei grassi. [94] Tuttavia, a causa di un aumento dei tassi di ossidazione dei carboidrati, non vi era alcuna differenza netta nel tasso metabolico tra Niacina e placebo [94].
I topi privi di PARP-1 sembrano avere tassi metabolici più alti e una minore massa grassa; in assenza di PARP, aumentano le concentrazioni di NAD +, attivando le SIRT1 che quindi lavorano per deacetilare varie proteine (PGC-1α e FOXO1) per promuovere il dispendio energetico attraverso un metabolismo ossidativo aumentato e un incremento dei mitocondri.[122]
La SIRT2 e la SIRT3 non sono influenzate dalla bassa attività del PARP-1, [122] e l’inibizione della ribosilazione dell’ADP con altri mezzi come il knockdown NMNAT-1 sembra conferire anche effetti antiobesità nei roditori. [182] L’alimentazione aumenta acutamente l’attività del PARP-1 nei topi e ostacola transitoriamente l’attività della SIRT1, [122] che si pensa sia correlata al PARP-1 che ha la priorità per l’uso dei donatori di NAD +.
La supplementazione orale di Nicotinamide Riboside a 400mg/kg nel topo sembra aumentare il contenuto di NAD + nel muscolo scheletrico similmente a quanto avviene alla stessa dose di Niacina (Nicotinamide Mononucleotide inefficace in questo organo) [123] e sembra aumentare il dispendio energetico nei topi nutriti con un alto contenuto di grassi insieme all’aumento dell’attività dei geni bersaglio di FOXO1, suggerendo che l’integrazione orale è efficace. [123]
Esistono prove limitate nell’uomo che valutano gli effetti della Niacina sul tasso metabolico, sebbene l’estremità inferiore del dosaggio farmacologico della niacina (1g) in soggetti altrimenti sani non sia riuscito ad aumentare il tasso metabolico rispetto al placebo. [94]
Niacina, muscolo scheletrico e prestazioni fisiche
La somministrazione di Niacina nell’uomo ha dimostrato di aumentare l’espressione dei fattori di trascrizione PPARδ e PPARγ coactivator-1α (PGC-1α) nel muscolo scheletrico. [124] Poiché questi fattori di trascrizione sono importanti regolatori del metabolismo ossidativo e della biogenesi mitocondriale, [125] [126] questo suggerisce che l’integrazione con Niacina può svolgere un ruolo nella resistenza dei muscoli scheletrici.
Gli studi sugli animali hanno supportato questa idea, in cui è stato dimostrato che l’integrazione di Niacina provoca una transizione di fibre muscolari dal tipo II (contrazione rapida) al tipo I (contrazione lenta), aumentando anche il numero complessivo di fibre di tipo I nei muscoli scheletrici nello Zucker (ratto) obeso [127] e suini in crescita [128] (750mg di Niacina/kg di dieta) e pecore (1g di Niacina al giorno). [129] Questo effetto può essere limitato a determinati modelli animali, tuttavia, poiché studi su ratti sani hanno dimostrato che la Niacina ha un effetto trascurabile sulla distribuzione del tipo di fibra muscolare o sul fenotipo metabolico. [130] Inoltre, nonostante la Niacina aumenti l’espressione dei fattori di trascrizione pro-ossidativa nell’uomo, [124] fino ad oggi nessuno studio ha dimostrato che migliora le prestazioni o la capacità di resistenza del muscolo scheletrico.
Tuttavia, come substrato per la sintesi di NAD +, un’adeguata presenza di Niacina può supportare indirettamente il metabolismo ossidativo e la resistenza muscolare. In soggetti altrimenti sani, un lieve esercizio fisico sembra essere associato ad un aumento delle concentrazioni di NAD + nel sangue rispetto a uno stato di riposo (indipendente da qualsiasi integrazione [131]) mentre, quando testato in un esercizio lieve nei roditori, portava anche ad un aumento del NAD + nel sangue prima che diminuisse durante un esercizio ad esaurimento, [131] che è stato notato anche nel muscolo scheletrico. [132] A questo livello di esaurimento c’è un concomitante aumento del contenuto di NADH nel muscolo scheletrico [133] [134] che è stato proposto [135] indicativo di una riduzione del trasferimento di elettroni dal NADH alla sintesi di ATP.
È stato inoltre proposto [135] che da quando l’esercizio aumenta l’ossidazione nei tessuti stimolati e i fattori di stress ossidativo sono noti per alterare l’attività del ciclo di Kreb (TCA) [136] e la catena di trasporto degli elettroni (compresa la NADH deidrogenasi [137]) che forniscono antiossidanti aumenterebbe la resistenza secondaria alla conservazione della cinetica intramuscolare di NAD + / NADH. Quando si forniscono 36mg di picnogenolo [135] come antiossidante durante l’esercizio fisico fino all’esaurimento, sembra che la diminuzione del NAD + nel sangue sia stata invertita in un aumento con gli effetti (sia la diminuzione che l’aumento in attesa di integrazione) più marcati negli atleti allenati. [135]
Impatto organico della Niacina e principali effetti collaterali
Indometacina
In uno studio svolto sui ratti, la Nicotinamide ad un dosaggio di 20mg/kg somministrata un’ora prima di una dose di Indometacina che induceva ulcerazioni allo stomaco ha impedito l’ulcerazione a un livello paragonabile sia al controllo (nessuna induzione di ulcere) sia al farmaco di riferimento di 400mg/kg di sucralfato, che agisce localmente per forma una superficie protettiva per lo stomaco. [138] Questo effetto si è verificato insieme alla conservazione dell’attività del glutatione, alla riduzione della perossidazione lipidica e al miglioramento del muco gastrico. [138] Simili effetti protettivi contro l’ulcerazione indotta da etanolo e stress sono stati notati altrove, con il metabolita primario della Nicotinamide (1-metilnicotinamide; MNA). [139] Questo effetto gastroprotettivo è stato associato con un aumento dell’attività delle prostaglandine, in particolare la PGI2, [139] e nicotinamide, nonché il suo metabolita MNA sono stati implicati nell’aumento del flusso sanguigno gastrico [139] e nella riduzione della permeabilità microvascolare [138] dopo l’ulcerazione.
Interleuchina 10 (IL-10)
Nel colon dei topi, il recettore della Niacina (GPR109A) è necessario per la proliferazione ottimale delle cellule T CD4 + e la produzione di IL-10, che si traduce in un effetto antiinfiammatorio. [140] Questo effetto antinfiammatorio guidato dal GPR109A è mediato dal butirrato, l’acido grasso a catena corta del colon [140], che è un agonista del GPR109A ed è prodotto attraverso la fermentazione della fibra alimentare da parte dei batteri nel colon. [141] [142]
L’effetto riducente dei Trigliceridi dato dalla Niacina sembra da doversi ricondurre al fegato, dove la secrezione di lipoproteine a bassissima densità (vLDL) è ridotta; poiché le vLDL normalmente trasportano i Trigliceridi dal fegato ad altri tessuti, riducendo la secrezione di vLDL ciò si traduce in un livello sierico di Trigliceridi inferiori. [89] La diminuzione della secrezione di vLDL può essere secondaria all’inibizione della lipolisi nel tessuto adiposo, poiché l’aumento cronico di acidi grassi liberi nel siero può regolare negativamente la secrezione di vLDL. [143]
Sembra che l’integrazione di Niacina in acuto (che riduce gli acidi grassi liberi nel siero) sopprime anche la produzione di vLDL e la sua complessazione con i trigliceridi. [144] Ciò suggerisce un altro possibile meccanismo, che può verificarsi attraverso la soppressione acuta del PGC-1β, [145] una proteina nota per promuovere la secrezione di Trigliceridi dal fegato in risposta all’ingestione di grassi nella dieta. [146] In accordo con quest’ultimo meccanismo, la somministrazione di Niacina con un pasto ad alto contenuto di grassi sembra ridurre il picco normale dei trigliceridi postprandiali. [147]
Regioni regolatorie, fattori di trascrizione e molecole di segnalazione (citochine, fattori di crescita, metaboliti, farmaci) che modulano l’espressione del gene ABCA1 nei macrofagi e in altri tessuti. Le frecce e le linee di blocco indicano rispettivamente l’attivazione e la repressione.
Non è confermato come la Niacina riduca le vLDL-C, ma la sua capacità di stimolare l’attività del gene ABCA1 e aumentare il suo contenuto proteico nelle cellule del fegato è alla base dell’aumento dell’HDL-C, [148] che è noto anche per sopprimere la secrezione di vLDL-C. [57] La Niacina (2g per 16 settimane), nonostante riduca le vLDL-C e il complesso con Trigliceridi, non sembra aumentare significativamente il contenuto di trigliceridi intraepatici in soggetti con malattia del fegato grasso non alcolica (NAFLD). [149]
La Niacina sembra anche agire sulle cellule del fegato per promuovere l’accumulo di diacilglicerolo (DAG), che è associato all’insulino-resistenza localizzata. [92] La resistenza all’insulina nelle cellule del fegato riduce l’effetto soppressivo dell’insulina sulla sintesi del glucosio, con conseguente aumento dell’efflusso di glucosio dal fegato nel sangue. [150] Poiché gli stadi iniziali dell’insulino-resistenza indotta dalla Niacina (prima degli aumenti dell’insulina basale e del glucosio) sono stati associati a un fabbisogno ridotto di glucosio per bilanciare un morsetto euglicemico iperinsulinaemico, [109] questo suggerisce che l’inizio dell’insulino-resistenza avviene a livello del fegato. Il ruolo preciso del DAG in questo processo è tuttavia incerto. Mentre il DAG promuove la resistenza all’insulina attraverso l’attivazione di PKCε, [151] l’attivazione di questa proteina non è stata osservata nelle cellule del fegato che sono diventate insulino-resistenti con la Niacina. [92]
TRANSAMINASI. Enzimi intracellulari prodotti principalmente dagli epatociti. normalmente presenti in circolo a bassi livelli nel sangue. Aumentano in caso di danno cellulare. Indici molto sensibili ma moderatamente specifici di danno epatico. ALT è un marker più specifico di danno epatocellulare. (localizzazione citoplasmatica e più lunga emivita)
Nota: La Niacina in dosi terapeutiche può causare aumenti modesti delle transaminasi sieriche e della bilirubina non coniugata, entrambi biomarcatori del danno epatico. Le modifiche vengono invertite se il trattamento farmacologico viene interrotto e di solito si risolvono anche quando si continua l’assunzione. [152] [153] [154] Tuttavia, meno comunemente, la forma di rilascio prolungato del farmaco può portare a gravi epatotossicità, con insorgenza in giorni o settimane. I primi sintomi di gravi danni al fegato includono nausea, vomito e dolore addominale, seguiti da ittero e prurito. Si ritiene che il meccanismo sia una tossicità diretta della Niacina sierica elevata. Abbassare la dose o passare alla forma a rilascio immediato può risolvere i sintomi. In rari casi la lesione è grave e progredisce fino a insufficienza epatica. [152]
È noto che la Niacina rende le cellule β pancreatiche (un tipo di cellula specializzata che secerne insulina in risposta al glucosio) meno sensibile al glucosio sierico. [81] Inoltre, la normale riduzione della sensibilità al glucosio delle cellule β del pancreas associata all’invecchiamento può essere ulteriormente esacerbata dalla supplementazione di Niacina (500mg-1g due volte al giorno). [83] Anche se sembra esserci un aumento compensativo della secrezione di insulina nella risposta alla Niacina [83], in un modello di primati con diabete di tipo I, [155] questo non è stato sufficiente a ridurre la glicemia a livelli normali, con conseguente lieve iperglicemia e iperinsulinemia dopo due settimane di integrazione.[83]
Va notato che una linea cellulare coinvolta nel rossore cutaneo tipico della Niacina, nota come Langerhans, [156] [157] è diversa dall’area del pancreas nota come “Isole di Langerhans”.
Nota: il rossore dato dalla Niacina – una dilatazione a breve termine delle arteriole della pelle, che causa il colore della pelle rossastra – di solito dura circa 15-30 minuti, anche se a volte può persistere per settimane con uso cronico e di forme a lento rilascio. In genere, il viso è maggiormente interessato, ma la reazione può estendersi al collo e alla parte superiore del torace. La causa è la dilatazione dei vasi sanguigni [158] [93] dovuta all’aumento della prostaglandina GD2 (PGD2) e serotonina. [159] [160] [161] [162] Si pensava spesso che il rossore riguardasse l’istamina, ma è stato dimostrato che l’istamina non è coinvolta nella reazione. [159] Il rossore a volte è accompagnato da una sensazione di prurito, in particolare, nelle aree coperte da indumenti. [93]
Acido Acetilsalicilico (Aspirina)
La prevenzione del rossore richiede l’alterazione o il blocco della via mediata dalle prostaglandine. [93] [163] L’Aspirina [165-325mg] assunta mezz’ora prima della Niacina riduce fortemente il rossore, così come l’Ibuprofene [200mg] (una riduzione della frequenza e intensità del rossore fino al 50%). L’assunzione di Niacina ai pasti aiuta anche a ridurre questo effetto collaterale. [93] La tolleranza acquisita aiuterà a ridurre l’effetto; dopo diverse settimane a dosaggio costante, la maggior parte delle persone non ha più esperienza di vampate di calore. [93] Sono state sviluppate forme di Niacina a rilascio lento o “prolungato” per ridurre questi effetti collaterali. [164] [165]
Conclusioni sulla supplementazione di Niacina
Le informazioni che abbiamo a disposizione sulla Niacina e la sua supplementazione, ci presentano un composto senz’altro utile per il controllo o riassesto del quadro lipidico ma allo stesso tempo questa molecola risulta di una complessità d’azione biochimica non trascurabile. Il suo peggiorare l’insulino-resistenza in cronico ma con un maggior picco in acuto, picco che sembra venire controbilanciato da altri fattori come l’aumento della Adiponectina. Lo stesso effetto sulla riduzione della lipolisi può destare preoccupazione nell’atleta, specie se questo si trova in una fase ipocalorica con il principale intento di ridurre la massa grassa. Anche in questo caso sembrerebbe che l’effetto si manifesti in acuto per poi rientrare in condizioni pre-utilizzo. Ciò che è quasi certo, è che le osservazioni sul campo non hanno fatto emergere grosse differenze nell’alterazione della composizione corporea, sia con l’uso della Niacina in regimi ipercalorici che ipocalorici. L’utilizzo di GDA (in specie Berberina) anche alle dosi base efficaci potrebbe essere un “tampone” sufficienti a compensare almeno in parte i possibili peggioramenti dei parametri dell’insulino resistenza. I controlli della glicemia basale e della insulinemia a digiuno sono indicatori da tenere sotto controllo durante l’uso di Niacina. Non è da trascurare la possibilità che la Niacina possa influenzare lo “shift” dalle fibre muscolari di tipo II a quelle di tipo I, cosa non auspicabile per un Bodybuilder o altro atleta di forza.
In definitiva, considerando i dosaggi efficaci e la migliore azione in combinazione con statine (vedi Monacolina K), l’assunzione di Niacina può essere mantenuta con un certo margine di sicurezza tra i 500mg ed 1g/die, ovviamente tarando il dosaggio in risposta agli esami ematici di controllo.
Gabriel Bellizzi
Riferimenti:
“Niacin”. Micronutrient Information Center, Linus Pauling Institute, Oregon State University, Corvallis, OR. 8 October 2018. Retrieved 16 September 2019.
Hegyi J, Schwartz RA, Hegyi V (January 2004). “Pellagra: dermatitis, dementia, and diarrhea”. International Journal of Dermatology. 43 (1): 1–5.
“Why fortify?”. Food Fortification Initiative. 2017. Retrieved 4 April 2017.
Institute of Medicine (1998). “Niacin”. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington, DC: The National Academies Press. pp. 123–149.
Bruckert E, Labreuche J, Amarenco P (June 2010). “Meta-analysis of the effect of nicotinic acid alone or in combination on cardiovascular events and atherosclerosis”. Atherosclerosis. 210 (2): 353–61.
“Niacin”. IN: LiverTox: Clinical and Research Information on Drug-Induced Liver Injury (Internet). Bethesda, MD: National Institute of Diabetes and Digestive and Kidney Diseases. February 2014.
Ricercatori australiani presso la University of New South Wales stanno testando un nuovo farmaco per la perdita di peso che agisce similmente al DNP (2,4-dinitrofenolo), ma senza i numerosi effetti collaterali connessi a quest’ultimo. Il loro lavoro è stato recentemente pubblicato su Nature Communications.[1]
BAM15?
Finché l’epidemia di obesità continuerà a crescere, la ricerca di farmaci per la perdita di peso continuerà. Parte di questa ricerca si svolge tenendo come riferimento il pericoloso ma estremamente efficace DNP. Sarebbe possibile sintetizzare una molecola che sia efficace come DNP, ma non così pericolosa?
Il DNP è un disaccoppiatore della fosforilizzazione ossidativa: interferisce con la resintesi di ATP nel mitocondrio causando una significativa dispersione di energia sotto forma di calore.
Ricercatori americani presso la Yale University stanno studiando un analogo metilato del DNP, il DNPME.[2]
La giapponese Otsuka Pharmaceutical sta conducendo esperimenti con OPC-163493.[3]
I ricercatori australiani che hanno pubblicato la ricerca che qui si sta trattando, stanno svolgendo esperimenti sul N5, N6-bis (2-fluorofenil) [1,2,5] oxadiazolo [3,4-b] pirazina-5,6-diammina. In breve: BAM15 .
Effetti del BAM15 nei topi
I ricercatori hanno somministrato ai topi dosi orali di BAM15 e hanno osservato che nelle ore successive alla somministrazione, il consumo di ossigeno degli animali – e quindi il loro consumo calorico – era aumentato di alcune decine di punti percentuale. L’effetto è stato temporaneo. Questo perché l’emivita del BAM15 nei topi è di sole 1,7 ore.
In altri esperimenti, in cui i topi sono stati alimentati con cibo contenente BAM15, i ricercatori hanno scoperto che la molecola aumentava il dispendio calorico solo di notte. Ciò non sorprende, perché i topi sono animali notturni e quindi preferiscono mangiare quando è buio. Il BAM15 non ha aumentato la quantità di attività fisica svolta.
Il dosaggio utilizzato nei topi trasposto in un essere umano adulto corrisponderebbe a circa 1g/die di BAM15.
In un altro esperimento, i ricercatori hanno nutrito i topi con zucchero e grasso addizionali [WD], rendendo gli animali più grassi. Se i topi venivano trattati con il BAM15, la loro massa grassa aumentava di meno, o per nulla, o addirittura diminuiva. L’equivalente umano delle dosi alle quali si sono verificati questi effetti è stato rispettivamente di 500mg, 1g e 1,5g di BAM15 al giorno.
Il BAM15 non ha aumentato la temperatura corporea dei topi, non ha ridotto la massa magra o aumentato l’attività dei radicali liberi. Per quanto i ricercatori hanno potuto scoprire, il BAM15 aumenta anche la sensibilità all’Insulina.
Conclusioni
Concludendo, i ricercatori riportano che il BAM15 rappresenta un raro disaccoppiatore mitocondriale che previene e inverte l’obesità senza influire sull’assunzione di cibo o sulla massa magra. Una limitazione del BAM15 è la bassa solubilità acquosa, ma questa proprietà non ha influito sulla biodisponibilità orale e infatti la bassa solubilità acquosa è un parametro importante che consente al BAM15 di penetrare nelle membrane cellulari ed entrare nei mitocondri. Un’altra limitazione del BAM15 è l’emivita di 1,7 ore e le direzioni future esamineranno le strategie di formulazione per migliorare l’esposizione. Collettivamente, i dati qui presentati supportano l’ulteriore sviluppo del BAM15 come potenziale terapeutico per l’obesità e le malattie metaboliche.
Non sono un amante delle diete “prefabbricate”, impostate su “dogmi” traballanti e letture parziali di studi mal interpretati. C’è stato un periodo della mia vita nel quale mi “innamorai” di un idea alimentare che mi portò dalla Dieta a Zona di Barry Sears alla Paleo Diet di Loren Cordain. Ma, fortunatamente, gli errori di gioventù insegnano ed è più di un decennio, da quando studio scienze della nutrizione, che mi sono totalmente discostato da una certa visione scientista dell’alimentazione.
Allora perchè dedicare un articolo ad una dieta “prefabbricata”? Beh, perchè l’onestà intellettuale non mi manca e quando c’è da riconoscere un merito, anche se può essere contornato da bislacche affermazioni, sono il primo a riconoscerlo.
Sicuramente, non scrivo questo articolo perchè penso che la “Vertical Diet” sia il must delle diete, e nemmeno perchè è opera di Stan Efferding, un Bodybuilder e PowerLifter d’élite, oppure perchè Hafthor Bjornsson, l’uomo più forte del mondo 2018, e Brian Shaw, l’uomo più forte del mondo per quattro volte, la seguono.
Ho già accennato il mio punto di vista su questa metodica alimentare in un post pubblicato qualche tempo fa su Instagram. Questa volta, però, merito del numero di caratteri qui esprimibili, avrò la possibilità di poter approfondire l’argomento in modo esaustivo.
Cos’è la “Vertical Diet”?
Stan “The White Rhino” Efferding (nato il 6 novembre 1967 a Portland, Oregon) è un Bodybuilder professionista americano IFBB ed un PowerLifter competitivo nella Southern Powerlifting Federation (SPF). Ha detenuto il record assoluto di powerlifting del mondo nella classe 275 libbre senza fasciatura del ginocchio (2.226,6 libbre) e nello Squat senza fasciatura del ginocchio (854 libbre). Grazie alla sua enorme forza fisica, partecipa regolarmente a gare di PowerLifting professionale e durante la sua carriera nel Bodybuilding competitivo professionistico, Efferding veniva spesso definito il “bodybuilder più forte del mondo”.
La “Vertical Diet” è stata sviluppata da Stan Efferding, un powerlifter d’élite, per migliorare le prestazioni di Bodybuilder, PowerLifter e atleti di alto livello.
Nel programma alimentare viene affermato anche che la presente strategia risulta ideale per i comuni frequentatori di palestra che stanno cercando di aumentare la massa muscolare o perdere peso.
A differenza delle tradizionali diete “orizzontali” che enfatizzano la varietà dietetica in numerosi gruppi alimentari, la “Vertical Diet” si concentra su un numero limitato di alimenti ricchi di nutrienti di alta qualità.
Secondo Efferding, limitare la varietà rende il corpo più efficiente nel digerire e assorbire i nutrienti, il che dovrebbe migliorare la crescita muscolare, il recupero, la salute dell’intestino e il metabolismo.
Detto questo, le sue affermazioni non sono supportate da valide prove scientifiche .
Alimenti nella “Vertical Diet”
Se c’è una cosa che rende questa metodica alimentare degna di nota è l’enfasi straordinariamente grande che l’autore da sull’ottimizzazione della digestione.
Si concentra su carne rossa, riso bianco e alimenti a basso contenuto di FODMAP.
In breve, la “Vertical Diet” riguarda il consumo di alimenti ricchi di nutrienti che sono facilmente digeribili per aiutare l’atleta perdere, guadagnare o mantenere il peso, massimizzare gli allenamenti e ottenere un migliore assorbimento dei nutrienti. L’obiettivo è quello di mantenere l’attenzione principale su micronutrienti come vitamine, minerali e antiossidanti.
Si chiama “Vertical Diet” perché quando è disposta graficamente, sembra una “T” rovesciata: alimenti base e alimenti “ancillari”. Come si può vedere nell’immagine riportata di seguito gli alimenti principali, posti in verticale, sono carne rossa e riso bianco.
Perché?
Ci sono pochi dubbi sul fatto che il riso bianco venga digerito molto facilmente dalla maggior parte delle persone, praticamente senza fibre e grassi è un “carboidrato pulito”: l’autore giustifica la sua scelta anche in base a vecchie credenze sulla “nocività” della lectina o acido fitico – chiamati “antinutrienti”, di cui parleremo di seguito – contenuti in alcuni alimenti di origine vegetale.
La carne rossa è stata scelta da Efferding perchè considerata il tipo di carne più nutriente, ricca di ferro eme, vitamine del gruppo B, Zinco, Magnesio, Creatina e Grassi sani (se grass fed).
Si può rimanere un pò sorpresi di vedere il Magnesio nell’elenco dato che le verdure a foglia verde e i legumi (sconsigliati da Efferding) sono generalmente considerati le migliori fonti. Duecento grammi ne contengono 50mg di questo importante minerale, anche se questo è solo il 12% del RDA – quindi, assumendo molti degli altri alimenti raccomandati, probabilmente è possibile raggiungere l’assunzione di 400mg al giorno.
Sotto i pilastri della dieta rappresentati da carne rossa e riso bianco, si trova la “base della “T”: frutta, patate, spinaci, peperoni rossi, carote, succo e alcuni altri prodotti animali come burro, brodo e pesce grasso.
Quindi l’enfasi è sulla carne rossa, riso e vegetali. Ma c’è un lungo elenco di cibi da “non mangiare”. E la compliance si riduce ulteriormente.
Ricapitolando, gli alimenti su cui ruota la “Vertical Diet” sono:
Riso: solo bianco
Carni rosse: manzo, agnello, bisonte e carne di cervo (grass fed)
Frutta: principalmente arance, succo d’arancia 100%, mirtilli e succo di mirtillo 100% – ma tutti i frutti sono ammessi
Patate: patate bianche e dolci
Verdure a basso contenuto di FODMAP: carote, sedano, zucchine, cetrioli, peperoni, melanzane, spinaci, zucca, ecc.
Oli e grassi: olio extra vergine di oliva, olio di cocco, olio di avocado, burro, noci
Pesce grasso: il consumo di salmone selvaggio dell’Alaska è fortemente incoraggiato
Uova: uova intere
Latticini: yogurt intero, latte intero, formaggio
Sodio: brodo di ossa, brodo di pollo, sale da tavola iodato
Pollame: pollo, tacchino
Possibili scelte solo in determinate modalità
Avena: solo se messa in ammollo e/o fermentata
Legumi: fagioli e altri legumi, solo se messi in ammollo e/o fermentati
Come si può notare, la dieta incoraggia a consumare cibi di alta qualità, come carni provenienti da animali allevati al pascolo, uova da galline ruspanti e frutta e verdura biologiche.
Cosa non mangiare nella “Vertical Diet”
Grani
L’autore, sempre condizionato da conoscenze parziali, vieta il consumo di fonti di acido fitico e glutine (che non causa alcun tipo di problema in soggetti sani), di conseguenza la dieta non include il consumo di pasta, cereali o pane, a meno che non sia pane fermentato a lievitazione naturale.
Avena
A meno che non sia messa a mollo e fermentata, sempre per la questione della riduzione degli “antinutrienti”.
Legumi
Considerata una fonte di lectine, e proteine dalla ridotta biodisponibilità per via del ricco contenuto di fibre. Possono essere consumati se messi in ammollo e fermentati.
Oli vegetali trasformati
Una scelta piuttosto non controversa: queste sono fonti ricche di acidi grassi Omega-6, già abbondantemente consumati nella dieta occidentale e che sono stati fortemente collegati a infiammazione, malattie cardiache e obesità. (1) (2) (3)
Caffè
Efferding afferma che il caffè provoca disidratazione e compromette la digestione.
Dice di non mangiare cibi che piacciono, ma di mangiare cibi che mi piacciono. Avere una ottimale salute intestinale e una digestione efficiente potrebbe far sentire meglio della soddisfazione provata consumando cibi appaganti il palato… almeno secondo l’autore.
Polialcoli (Polioli)
Dolcificanti naturali come l’Eritritolo e lo Xilitolo, piuttosto popolari nelle barrette proteiche e nei gelati dietetici. Come per i legumi, sono noti per poter causare indigestione in alcune persone, anche se non è un fenomeno così diffuso.
Verdure ad alto contenuto di Raffinosio
Forse questa è l’esclusione più controversa, che include verdure crocifere come broccoli, cavolfiori e asparagi. Il colpevole è il Raffinosio, un certo zucchero presente in alcune verdure, cereali e legumi che è resistente alla digestione e talvolta può causare gas.
Aglio e cipolla
Sono alimenti ricchi di FODMAP (sigla che sta per “Fermentable Oligosaccharides, Disaccharides, Monosaccharides and Polyols” ovvero Oligosaccaridi, Disaccaridi, Monosaccaridi Fermentabili e Polioli) .
Acqua alcalinizzata
Secondo l’autore, questa bufala commerciale, può alterare la digestione.
Bisogna tenere presente che la dieta consente il consumo di piccole quantità di alcuni di questi alimenti purché il soggetto possa digerirli senza alcun sintomo digestivo, come gas o gonfiore.
Tuttavia, gli oli vegetali trasformati non sono mai ammessi.
Quindi cos’è un alimento FODMAP?
I FODMAP sono carboidrati fermentabili il cui consumo è limitato tra le persone con disturbi digestivi
La “Vertical Diet” è fondamentalmente una dieta a basso contenuto di FODMAP.
Come già accennato, FODMAP è l’acronimo di “Fermentable Oligosaccharides, Disaccharides, Monosaccharides and Polyols” ovvero Oligosaccaridi, Disaccaridi, Monosaccaridi Fermentabili e Polioli, che si riferisce ai carboidrati a catena corta che sono spesso limitati tra le persone con disturbi digestivi come la sindrome dell’intestino irritabile. (4) (5) Sono tipi di fibre e attraversano il sistema non digerite; i batteri intestinali li usano come substrato energetico, producendo idrogeno gassoso nel processo. (6) In alcune persone, ciò può attirare liquidi nell’intestino e causare diarrea e per queste persone le diete povere o a basso contenuto di FODMAP possono essere una manna dal cielo.
Poiché la “Vertical Diet” prevede macronutrienti facilmente digeribili, i cibi ricchi di FODMAP sono limitati o preparati in modo da ridurre il disagio digestivo. Le verdure come spinaci, cetrioli, peperoni e patate sono migliori delle verdure crocifere per aumentare i risultati positivi della dieta secondo quanto affermato da Efferding.
Non ci sono molti dati su popolazioni sane che usano una dieta a basso contenuto di FODMAP, ma alcune ricerche hanno mostrato una riduzione dei gas fermentativi e miglioramenti nella salute dell’intestino, e uno studio ha persino scoperto che una dieta low-FODMAP migliora i sintomi gastrointestinali legati all’esercizio fisico. (7) (8) Questa è solo una congettura, ma questo potrebbe essere una alimentazione utile per le persone che hanno bisogno di digerire una quantità straordinariamente alta di calorie.
Lista degli alimenti ricchi in FODMAP
Nota: si ricordi che anche nella dieta FODMAP, la quale non è una classica dieta ad eliminazione di determinati alimenti ma è un regime alimentare strutturato su schemi piuttosto rigidi, prevede in seguito il reintegro, dopo un dato lasso di tempo, degli alimenti eliminati (FODMAP). Infatti non è consigliabile eliminare definitivamente tutti i cibi che contengono FODMAP, principalmente per la riduzione nella varietà della dieta e, come avvalorato da diversi studi, si registrano dei potenziali effetti benefici, soprattutto a medio-lungo termine, dei FODMAP e degli alimenti che li contengono (vedi, per esempio, i legumi).
Lectine, acido fitico e “antinutrienti”
Questi composti possono ridurre l’assorbimento di alcuni minerali
Questo può essere risolto consumandoli con vitamina C e / o tenendoli in ammollo e/o facendoli fermentare.
Lectine e acido fitico sono spesso denominati “antinutrienti” perché, a concentrazioni rilevanti, si legano e compromettono la digestione di minerali come magnesio, ferro e zinco. (9)(10)
Questo effetto viene quasi completamente eliminato se si consuma della Vitamina C nello stesso pasto, inoltre la lectina ha alcuni collegamenti per ridurre i rischi di cancro e l’acido fitico è un antiossidante che è stato associato a minori rischi di malattie cardiovascolari e calcoli renali. (11) (12) (13) (14) (15) (16) Ecco il motivo per il quale alcune persone consumano integratori di acido fitico.
E’ di facile conclusione il fatto che lectine e acido fitico non sono il “male assoluto” come propagandato, per esempio, dai fanatici della “Paleo Diet”. L’idea che questi composti siano assolutamente dannosi per tutti non è basata sui dati reali. Comunque sia, il consumo di legumi e grani nella “Vertical Diet” è consentita solo attraverso precise modalità di preparazione, come la messa in ammollo o la fermentazione, pratiche che aiutano a ridurre la presenza di “antinutrienti”. (17) (18)
Settaggio calorico nella “Vertical Diet”
Fortunatamente, questa volta non ci troviamo di fronte ad una dieta di classica foggia commerciale nella quale troneggia lo slogan “le calorie non contano”. In questo l’autore mantiene un filo logico.
Infatti, inizialmente è consigliato il calcolo della spesa calorica basale (BMR). Successivamente si addiziona al risultato del calcolo del BMR il dispendio calorico derivante dall’attività sportiva e extra-sportiva (NEAT). Quindi, una volta ottenuto il proprio TDEE, a seconda dell’obbiettivo prefissato, si aggiungono o tolgono calorie.
Viene consigliato anche di tarare la dieta in un periodo di prova nel quale il corpo si adatterà alle nuove variabili alimentari. Se in questo periodo si inizia a sentire fame tra i pasti, il soggetto dovrebbe “andare in verticale” aggiungendo più calorie. Questo processo ha lo scopo di supportare maggiori guadagni muscolari, un recupero più rapido e sessioni di allenamento più intense e/o frequenti.
Il numero esatto di calorie aggiuntive si basa sulle esigenze di allenamento e comporta l’aumento delle porzioni di riso e carne o un pasto aggiuntivo durante il giorno.
Una volta che si inizia di nuovo ad avere fame tra i pasti, va ripetuto questo processo fino a raggiungere il peso o la massa muscolare prefissata come obiettivo.
Macronutrienti nella “Vertical Diet”
di base, la “Vertical Diet” è relativamente povera di grassi e ricca di carboidrati
Le calorie totali dipendono dal metabolismo basale e dal carico di lavoro soggettivo, ma i macronutrienti di base in questa strategia alimentare sono così gestiti:
Proteine: 2g per chilo di peso corporeo
Grassi: 0,6g per chilo di peso corporeo
Carboidrati: il resto delle calorie.
Supponiamo che un soggetto pesi 80Kg e abbia una spesa calorica giornaliera di 3000Kcal. La quantità dei macronutrienti sarebbe la seguente:
E’ ovvio che il calcolo del proprio monte calorico necessario ai fini dell’obbiettivo che ci si è prefissati è un componente importante di ogni piano nutrizionale, e questa dieta non fa eccezione.
Si noti che questa è di base una dieta relativamente povera di grassi, così progettata per gli atleti, che in genere hanno bisogno di fare più spazio nei loro macronutrienti per l’inserimento di una buona quota di carboidrati rispetto alle persone sedentarie.
La “Vertical Diet” non è statica nella ripartizione calorica, tanto che risulta versatile non solo per la sua applicabilità sia nei periodi “Bulk” che in quelli +”Cut”, ma anche per quanto riguarda le percentuali di macronutrienti e la loro gestione. Infatti può essere adattata a regimi come le Low Carb, IF o Paleo.
Benefici potenziali della “Vertical Diet”
Ovviamente un surplus calorico è essenziale per aumentare la massa muscolare (5).
Concentrandosi su alimenti facilmente digeribili, la “Vertical Diet” rende più facile mangiare pasti frequenti e ipercalorici senza sperimentare effetti collaterali digestivi.
Inoltre, la dieta enfatizza l’aumento dell’assunzione di carboidrati, fattore che contribuisce ad aumentare la massa muscolare (19, 20, 21).
Gli studi dimostrano che un’adeguata assunzione di carboidrati prima dell’allenamento può migliorare le prestazioni atletiche. I carboidrati possono anche aumentare la sintesi proteica e ridurre il catabolismo muscolare (20, 21).
Le diete povere di FODMAP – alimenti limitati nella “Vertical Diet” – hanno dimostrato di ridurre significativamente i sintomi legati a problemi digestivi, come gonfiore, crampi allo stomaco, costipazione e diarrea, specie nelle persone con sindrome dell’intestino irritabile (IBS). (22, 23)
I Bodybuilder e altri atleti che hanno bisogno di consumare pasti frequenti e ipercalorici possono anche trarne beneficio, in quanto i cibi a basso contenuto di FODMAP riducono il rischio di gonfiore addominale. In caso contrario, il gonfiore può limitare il consumo di cibo compromettendo il raggiungimento della quota calorica prefissata per l’aumento della massa muscolare.
Tuttavia, alcuni alimenti ricchi di FODMAP sono ammessi nella “Vertical Diet”, tra cui latte, yogurt, mele, ciliegie, fichi e altri frutti.
Pertanto, i soggetti con IBS dovrebbero evitare anche questi alimenti.
Potenziali aspetti negativi della “Vertical Diet”
È importante notare che la “Vertical Diet” ha numerosi aspetti negativi, tra cui:
Basso contenuto di fibre. Un’adeguata assunzione di fibre aiuta a raggiungere il senso di sazietà (quando ricercato), indirettamente la salute cardiovascolare e direttamente la salute dell’apparato digerente. Può anche ridurre il rischio di comparsa di malattie croniche, come il diabete di tipo 2 e alcune forme di cancro (24, 25, 26).
Basso contenuto di prebiotici. Nonostante si affermi che questa dieta migliori la salute dell’intestino, essa esclude molte importanti fonti di prebiotici – fibra alimentare che alimenta i batteri benefici nell’intestino – tra cui, e non solo, aglio, cipolle e orzo.(24, 27)
Limitata nella varietà degli alimenti. La “Vertical Diet” è restrittiva e ripetitiva, rendendo difficile l’aderenza sul lungo termine. Può anche portare a carenze nutrizionali se non pianificata correttamente.(28, 29)
Inappropriato per vegetariani o vegani. Poiché la “Vertical Diet” enfatizza l’assunzione di carne rossa limitando l’assunzione di verdure, cereali e legumi, non è adatta per persone vegetariane o vegane.
Costosa da seguire. Mentre il riso bianco è generalmente economico, gli altri componenti della “Vertical Diet” possono essere costosi, soprattutto considerando la raccomandazione di acquistare solo alimenti di alta qualità, come carne di manzo “grass fed” e prodotti biologici.
Esempio di un piano alimentare “Vertical Diet”
Ecco un menu “Vertical Diet” di esempio di 3 giorni . Non sono specificate le quantità per alimento tranne che gli ml di succo o brodo, e il numero di pasti può variare in base al regime di allenamento e al fabbisogno calorico.
Giorno 1
Pasto 1: uova intere strapazzate con formaggio, peperoni rossi, spinaci e sale, servite con carotine crude, mandorle crude e 120ml di succo di mirtillo
Pasto 2: filetto di manzo macinato e riso bianco cotto nel brodo di pollo, più 120ml di succo d’arancia
Pasto 3: petto di pollo e patata dolce servito con 120 ml di succo d’arancia
Pasto 4: bistecca grass-fed con riso bianco cotto nel brodo di pollo e 120ml di succo di mirtillo
Spuntino: yogurt greco e carotine.
Giorno 2
Pasto 1: uova intere strapazzate con formaggio, spinaci, peperoni rossi e brodo di ossa, servite con patate lesse e 120ml di succo di mirtillo
Pasto 2: bisonte macinato con riso bianco, patata dolce e brodo di ossa, insieme a 120ml di succo d’arancia
Pasto 3: petto di pollo con riso bianco, patata dolce, brodo di ossa e un’arancia
Pasto 4: bistecca grass-fed con riso bianco, patate, zucchine e brodo di ossa, servito con 120ml di succo di mirtillo
Spuntino: latte intero e carotine.
Giorno 3
Pasto 1: uova intere strapazzate con formaggio, spinaci, peperoni rossi e sale, insieme all’avena messa in ammollo durante la notte con base di yogurt, latte e miele e noci opzionali
Pasto 2: bistecca di controfiletto macinata con riso bianco, peperoni e brodo di pollo, servito con 120ml di succo di mirtillo
Pasto 3: salmone dell’Atlantico selvaggio con riso bianco, spinaci, peperoni e brodo di pollo, oltre a carotine e 120ml di succo d’arancia
Pasto 4: bistecca grass-fed con riso bianco, patate dolci e brodo di pollo, oltre a 120ml di succo di mirtillo
Spuntino: yogurt greco e frutti di bosco.
Tiriamo le somme sulla “Vertical Diet”
Come abbiamo visto, la “Vertical Diet” ha lo scopo di aiutare i Bodybuilder ed altri atleti ad aumentare la massa muscolare e migliorare le prestazioni o a perdere peso.
Include alimenti facilmente digeribili per aiutare il corpo ad assorbire i nutrienti in modo più efficiente e prevenire effetti collaterali digestivi, come il gonfiore. Per aumentare l’assunzione di proteine e carboidrati, l’autore sottolinea di consumare porzioni sempre più grandi di carne rossa e riso bianco.
La “Vertical Diet” è severamente restrittiva, costosa da seguire e povera di fibre complessive e prebiotiche, oltre che a presentarsi come un generico piano alimentare basato su alcuni punti non adeguatamente contestualizzati. La compliance non è facilitata ed è una dieta estremamente difficili da mantenere sul lungo termine.
Ma quando si guarda a una dieta così focalizzata sull’aiutare a digerire il cibo nel modo più efficiente possibile, non è difficile capire perché possa avere delle grandi potenzialità applicative in regimi alimentari ipercalorici. Con una dieta ad elevato contenuto calorico, i cibi ricchi di FODMAP, i cibi ricchi di fibre e cibi estremamente grassi – tutti esclusi o severamente limitati nella “Vertical Diet” – potrebbero potenzialmente rendere più difficile consumare e assimilare la quantità di cibo necessaria per ottenere i risultati agognati. La maggior parte delle persone che hanno provato ad aumentare di peso con piani alimentari contenenti legumi affermeranno che ad un certo punto, la fibra smette di essere un fattore benefico a livello digestivo.
Non si può comunque negare che la maggior parte degli alimenti limitati in questa dieta hanno più benefici che svantaggi. Detto questo, la “Vertical Diet” consente una varietà abbastanza ampia di frutta, verdura, verdure a foglia verde, carne e fonti di acidi grassi Omega-3, sufficienti per rendere improbabili le carenze nutrizionali con un regime ben pianificato. E non esiste alcuna regola nel piano dietetico contro gli integratori per arginare eventuali carenze.
Personalmente considero la “Vertical Diet” un ottima opzione nutrizionale per tutti coloro i quali presentano facilmente difficoltà digestive. La sua applicazione migliore risulta essere in “Bulk” per via della considerevole quantità di cibo richiesta. In fasi “Cut” non la ritengo una dieta particolarmente vantaggiosa a meno che non si parli di soggetti con IBS o con problemi digestivi.
In conclusione, Stan Efferding ha messo insieme nozioni logicamente intuitive (vedi garantire una migliore digestione e assorbimento dei nutrienti) con una serie di limitazioni non sempre veritiere e/o applicabili vantaggiosamente (vedi, per esempio, eliminazione dei cibi contenenti glutine a prescindere dalla presenza di celiachia). Nulla di nuovo sotto il sole… nonostante una piccola ma meritata nota positiva sull’importanza della corretta assimilazione di ciò di cui ci si nutre…
Se volete acquistare l’Ebook della “Vertica Diet” cliccate qui.
Gabriel Bellizzi
Riferimenti:
Simopoulos AP, et al. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients. 2016 Mar 2;8(3):128.
Patterson E, et al. Health implications of high dietary omega-6 polyunsaturated Fatty acids. J Nutr Metab. 2012;2012:539426.
Okuyama H, et al. Omega3 fatty acids effectively prevent coronary heart disease and other late-onset diseases–the excessive linoleic acid syndrome. World Rev Nutr Diet. 2007;96:83-103.
Barrett JS, et al. Fermentable oligosaccharides, disaccharides, monosaccharides and polyols (FODMAPs) and nonallergic food intolerance: FODMAPs or food chemicals? Therap Adv Gastroenterol. 2012 Jul;5(4):261-8.
Turco R, et al. Does a low FODMAPs diet reduce symptoms of functional abdominal pain disorders? A systematic review in adult and paediatric population, on behalf of Italian Society of Pediatrics. Ital J Pediatr. 2018 May 15;44(1):53.
Ong DK, et al. Manipulation of dietary short chain carbohydrates alters the pattern of gas production and genesis of symptoms in irritable bowel syndrome. J Gastroenterol Hepatol. 2010 Aug;25(8):1366-73.
Sloan TJ, et al. A low FODMAP diet is associated with changes in the microbiota and reduction in breath hydrogen but not colonic volume in healthy subjects. PLoS One. 2018 Jul 26;13(7):e0201410.
Wiffin M, et al. Effect of a short-term low fermentable oligiosaccharide, disaccharide, monosaccharide and polyol (FODMAP) diet on exercise-related gastrointestinal symptoms. J Int Soc Sports Nutr. 2019 Jan 15;16(1):1.
Schuchardt JP, et al. Intestinal Absorption and Factors Influencing Bioavailability of Magnesium-An Update. Curr Nutr Food Sci. 2017 Nov;13(4):260-278.
Jiang QL, et al. Plant lectins, from ancient sugar-binding proteins to emerging anti-cancer drugs in apoptosis and autophagy. Cell Prolif. 2015 Feb;48(1):17-28.
Davidsson L. Approaches to improve iron bioavailability from complementary foods. J Nutr. 2003 May;133(5 Suppl 1):1560S-2S.
Siegenberg D, et al. Ascorbic acid prevents the dose-dependent inhibitory effects of polyphenols and phytates on nonheme-iron absorption. Am J Clin Nutr. 1991 Feb;53(2):537-41.
Schlemmer U, et al. Phytate in foods and significance for humans: food sources, intake, processing, bioavailability, protective role and analysis. Mol Nutr Food Res. 2009 Sep;53 Suppl 2:S330-75.
Omoruyi FO, et al. The potential benefits and adverse effects of phytic Acid supplement in streptozotocin-induced diabetic rats. Adv Pharmacol Sci. 2013;2013:172494.
Grases F, et al. Phytate (IP6) is a powerful agent for preventing calcifications in biological fluids: usefulness in renal lithiasis treatment. Anticancer Res. 1999 Sep-Oct;19(5A):3717-22.
Gupta RK, et al. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J Food Sci Technol. 2015 Feb;52(2):676-84.
Liang J, et al. Effects of soaking, germination and fermentation on phytic acid, total and in vitro soluble zinc in brown rice. Food Chem. 2008 Oct 15;110(4):821-8.