Fino al 1969, il Methyltrienolone era considerato lo steroide anabolizzante più potente sintetizzato in laboratorio. In seguito ai lavori di due ricercatori americani emerse un nuovo AAS dal potenziale maggiore del Methyltrienolone, il 2-oxa-methyldienolone.(1)
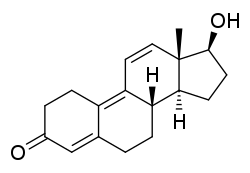
Methyltrienolone
Negli anni ’60, venne sintetizzato il Methyltrienolone, il quale ottenne una cattiva reputazione per via dei suoi marcati effetti collaterali mostrati nel primo studio su esseri umani.(2) In questo studio, l’AAS non mostrò soltanto un effetto anabolizzante estremamente accentuato ma anche un grado di tossicità epatica significativamente alto.
Il Methyltrienolone rimase l’AAS più potente mai sintetizzato in laboratorio per qualche anno, fino al 1969. Infatti, fu in quell’anno che due ricercatori della società farmaceutica americana Searle, pubblicarono uno studio svolto su animali nel quale era stato utilizzato un AAS con potenzialità addirittura maggiori del Methyltrienolone.
I ricercatori hanno svolto esperimenti con una classe di AAS largamente utilizzati in ambito veterinario, i dienolini. Due degli AAS testati sono stati commercializzati come designer steroid all’inizio del XXI Secolo. Si tratta del Dienolone e del Methyldienolone, composti I e II presenti nella figura seguente.
Nella tabella seguente, i ricercatori hanno confrontato gli effetti anabolizzanti e androgeni degli AAS testati con quelli del Methyltestosterone. I ricercatori hanno somministrato le molecole per via orale, sciolte in olio di mais. Hanno usato ratti come animali da test. Puoi approfondire il metodo della ricerca cliccando qui.
Come si può vedere, il Dienolone ha mostrato effetti leggermente più anabolici e leggermente meno androgeni rispetto al Methyltestosterone. Il Methyldienolone (aka Composto II) ha mostrato una potenza maggiore con effetti androgeni e anabolici rispettivamente di 6 e 80 volte maggiori del Methyltestosterone.
*La tabella seguente è stata semplificata.
Tuttavia, gli AAS più interessanti che i ricercatori hanno testato erano i composti III e IV. Queste molecole non avevano un atomo di carbonio nella posizione C2 nel loro scheletro carbossilico, ma un atomo di ossigeno. Per comodità, chiameremo questi due AAS 2-oxa-dienolone e 2-oxa-methyldienolone.
La sostituzione ha causato una modifica dell’effetto androgeno e anabolico del 2-oxa-dienolone [composto III] superiore a quello del Methyltestosterone di un fattore rispettivamente di 6 e 93.
Tetrahydrogestrinone (THG)
Il 2-oxa-methyldienolone [composto IV] ha mostrato effetti ancora più significativi. Questo AAS ha avuto un effetto androgeno circa 30 volte maggiore rispetto al Methyltestosterone e con un effetto anabolico non inferiore alle 550 volte maggiore. Questo lo rende potenzialmente anche più potente dell’AAS sintetizzato da Patrick Arnold, il Tetrahydrogestrinone (THG).
Gli autori dello studio riportarono che gli aumenti nell’attività, sia androgena che miotrofica, con cambiamenti nella struttura della molecola madre in questa serie di composti erano stati drammatici. La più alta attività biologica è stata raggiunta nel derivato 17a-metil-2-oxa (IV), che si rivelò la sostanza androgeno-miotrofica più potente fino a quel momento segnalata e avente anche un’ampia separazione delle proprietà anaboliche e androgene.
Una nota interessante riportata dai ricercatori impegnati nello studio è la citazione della (allora) recente sintesi del Methyltrienolone [17 alfa-metil-4,9 (10), ll-trien-17-beta-ol-3-one] e della segnalata attività miotrofica molto elevata.(3) I ricercatori aggiunsero anche che mancava l’alto grado di stimolazione preferenziale del muscolo levatore ani che era caratteristico del derivato 17a-metil-2-oxa (IV) di loro sintesi.
Nel 1960, la società farmaceutica svizzera CIBA iniziò a commercializzare lo steroide anabolizzante sintetizzato nei suoi laboratori nella seconda metà degli anni cinquanta, il noto Methandrostenolone che venne commercializzato sotto il nome di Dianabol. Con la diffusione del Methandrostenolone nel Bodybuilding e nel Weightlifting iniziò ufficialmente l’era degli AAS in ambiente sportivo. Negli anni successivi a questo diffuso successo nell’utilizzo del Dianabol, la CIBA avviò degli studi su un altro composto steroideo anabolizzante, del quale però non si sa quasi nulla. L’AAS in questione è il 36644-Ba.(1)
La struttura chimica del 36644-Ba è senza dubbio particolare, anche se sarebbe corretto di “le strutture” . Infatti, il 36644-Ba è in realtà una molecola con una ciclicità metabolica che ne caratterizza la farmacodinamica. La molecola progenitore (il 36644-Ba) si converte in un’altra struttura molecolare la quale ritorna alla forma di partenza ciclicamente.
La prima formula strutturale mostrata di seguito è quella ufficiale, la seconda è il metabolita in cui il composto progenitore 36644-Ba si converte per poi ritornare alla forma di partenza.
Gli unici studi sull’attività biologica del 36644-Ba reperibili sono stati svolti su animali. In questi studi le cavie esaminate non sono soltanto state trattate con il 36644-Ba, ma anche con un corticosteroide. E’ superfluo sottolineare che i corticosteroidi hanno un azione catabolica a livello del tessuto muscolare.
Nel miocita, i corticosteroidi riducono la sintesi proteica da parte dei ribosomi. In alcuni studi, i ricercatori della CIBA hanno dimostrato che il 36644-Ba può mantenere l’attività dei ribosomi durante la somministrazione di corticosteroidi. Uno di questi studi, pubblicato nel 1968, può essere trovato integralmente su PubMed Central.
Nel 1970, i ricercatori hanno pubblicato uno studio simile in cui il 36644-Ba ha mostrato di avere effetti più potenti dello Stanozololo. Non è stato però possibile reperire il suddetto studio. Ciò che è possibile sapere a riguardo proviene dalle relazioni annuali di chimica farmaceutica, pubblicate da Academic Press nel 1971.(2)
Nella precedentemente citata pubblicazione del 1968, gli autori spiegano come il 36644-Ba ha agito sugli animali trattati, dando un’idea delle proprietà anaboliche desiderate e delle proprietà androgene indesiderate dell’AAS in questione.
Questi dati provengono dagli Atti del Secondo Congresso Internazionale sugli Steroidi Ormonali, Milano, 23-28 maggio 1966.(3) Non è stato possibile reperire nemmeno questa pubblicazione.
Il più recente riferimento scientifico al 36644-Ba risale al 2009. In quell’anno, il “doping hunter” australiano Ray Kazlauskas menzionò il 36644-Ba nel suo capitolo sui designer steroids in Doping in Sports collection: Biochemical Principles, Effects and Analysis, pubblicato da Academic nel 2009.(4)
Il 36644-Ba sembra essere molto attivo, ma non sembra essere stato usato al di fuori della sfera sperimentale, come sottolineato da Kazlauskas a pagina 168 della sua pubblicazione.
Nel 2019 dovrebbe essere ormai chiaro a tutti che la risposta agli androgeni da parte delle donne è molto più accentuata rispetto agli uomini ma, purtroppo, non sembra essere così. La cosa si manifesta nel dilagare di fenomeni da baraccone che, sebbene aventi condizioni ipertrofiche di un certo livello, mostrano gradi di mascolinizzazione imbarazzanti, da cambio di sesso. Ora, non ho intenzione di fare una disamina dei disturbi di identità che portano certi membri del gentil sesso ad affidarsi a soggetti incompetenti nel campo della farmacologia applicata al miglioramento delle prestazioni sportive con l’intento di diventare il “lui” con qualcosa di “lei”, ma è, come sempre, mia intenzione far riflettere sull’oggettività delle cose. Se è sufficiente 1/5 del dosaggio di un uomo per avere una spiccata risposta ipertrofica con un buon grado di conservazione della femminilità, qual senso denaturare l’estetica del BodyBuilding femminile con dosaggi che sarebbero alti anche per molti uomini? Il diventare “più grosse”? Naaa, non regge più come scusa…
Recentemente, è stato pubblicato uno studio sul “British Journal of Sports Medicine” da parte di ricercatori svedesi i quali hanno sottoposto delle donne a trattamento topico a base di 10mg di Testosterone al giorno. Tale dosaggio lungi dal poter far raggiungere la soglia ematica di Testosterone riscontrabile in un uomo, ma è stata sufficiente a portare un aumento della massa muscolare, un calo della massa grassa e il miglioramento della resistenza.(1)
Alcune donne (non parlo dei soggetti con PCO) producono molto più Testosterone del normale a causa di un difetto congenito. Tra i migliori atleti è possibile trovare queste donne con un rapporto 140 volte superiore rispetto a quello riscontrabile nel resto della popolazione. (2) I ricercatori svedesi del Karolinska Institutet si sono chiesti se tali anomalie possano effettivamente dare vantaggi prestativi negli sport alle donne.
I ricercatori hanno diviso 48 donne normali, sane e attive di età compresa tra 18 e 35 anni in 2 gruppi. Durante le 10 settimane di durata dell’esperimento, le donne prese in esame hanno applicato 1ml di soluzione topica (crema) sull’esterno coscia ogni notte. Le donne del gruppo placebo hanno usato una soluzione senza sostanze bioattive, mentre le donne del gruppo sperimentale hanno applicato quotidianamente sulla cute una soluzione contenente 10mg di Testosterone.
I ricercatori hanno utilizzato AndroFeme, un prodotto realizzato dall’azienda Australian Lawley Pharmaceuticals.(3) AndroFeme è destinato alle donne in post-menopausa.
Nelle donne che avevano applicato la soluzione contenente Testosterone, il livello ematico dell’ormone era aumentato di un fattore 4,8. Questo livello è inferiore a quello che ci si aspetterebbe di trovare negli uomini. Gli uomini sani hanno un livello minimo di Testosterone di almeno 12nanomoli per litro.
Nell’immagine in alto a destra è possibile osservare come la dose di Testosterone somministrata giornalmente non ha avuto effetti sulla forza espressa alla Leg-Extension. Inoltre, le donne trattate non erano in grado di saltare più in alto [vedi in alto a sinistra].
I vantaggi della somministrazione topica di 10mg di Testosterone si sono manifestati in un aumento della resistenza aerobica delle donne trattate. Durante un test in cui le donne dovevano correre su un tapis roulant con una pendenza sempre più elevata [in basso a sinistra] e un test in cui le donne dovevano pedalare per 30 secondi il più velocemente possibile [in basso a destra], la somministrazione di Testosterone ha portato ad un aumento rispettivamente il tempo di esaurimento e la velocità massima esprimibile.
La massa muscolare totale nelle donne trattate con Testosterone ha subito un aumento del 2%, anche se i soggetti non hanno subito un incremento di peso. L’aumento della massa muscolare è stato compensato da una diminuzione della massa grassa (ricomposizione corporea).
Nel gruppo Testosterone, i cambiamenti corporei che potrebbero essere il risultato del trattamento ormonale, come l’aumento dell’acne e dei peli nel viso e nel corpo, erano tre volte più comuni rispetto al gruppo placebo. Più del 70% delle donne del gruppo Testosterone ha riportato questi cambiamenti. Ovviamente, vista la dose, non sono stati particolarmente marcati.
I ricercatori concludono con il dire che il loro studio supporta un effetto causale del Testosterone sulle prestazioni fisiche, misurato in base al tempo di esaurimento, in giovani donne sane. L’effetto ergogenico dovuto alla concentrazione di Testosterone moderatamente aumentata nel breve termine sembrava applicarsi solo sulla prestazione aerobica.
Il Testosterone ha anche promosso un miglioramento della composizione corporea con un aumento della massa muscolare sebbene il peso corporeo fosse invariato.
Vi ricordo che 10mg di Testosterone somministrato per via topica hanno una biodisponibilità molto contenuta. Ma nonostante ciò, gli effetti positivi si sono manifestati in modo statisticamente significativo.
La dose può fare il veleno o una differenza tutto sommato positiva…
Secondo uno studio su animali svolto da fisiologi rumeni presso l’Università di Medicina e Farmacia Carol Davila e pubblicato su “Medicina”, la supplementazione di Taurina potrebbe offrire protezione cardiovascolare agli utilizzatori di AAS.(1)
Per lo svolgimento del loro studio, i ricercatori hanno diviso dei ratti maschi in 4 gruppi:
Un gruppo di controllo [C] al quale non è stato somministrato alcun farmaco;
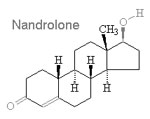
Un gruppo al quale veniva somministrata un’iniezione settimanale di Nandrolone Decanoato [A];
Un gruppo al quale veniva somministrata della Taurina miscelata all’acqua [T];
Un gruppo al quale veniva somministrata un’iniezione settimanale di Nandrolone Decanoato e Taurina miscelata all’acqua [AT].
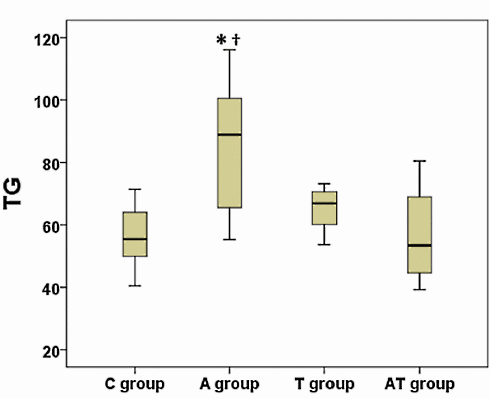
Nei tre mesi di durata dell’esperimento, la concentrazione dei Trigliceridi nel sangue dei ratti trattati con Nandrolone era ovviamente peggiorata. L’ipertrigliceridemia è un noto fattore di rischio per malattie cardiovascolari. La supplementazione di Taurina, tuttavia, ha impedito il verificarsi di questo aumento dei livelli di Trigliceridi, come mostrato nella figura seguente.
La somministrazione di Nandrolone decanoato ha portato anche ad una riduzione delle concentrazioni di HDL, una condizione nota per essere un fattore di rischio cardiovascolare. La figura seguente mostra l’impatto “contenitivo” sulla riduzione del HDL che la Taurina ha esercitato nei topi trattati con Nandrolone Decanoato e la supplementazione dell’aminoacido.
Nel 2016, i ricercatori rumeni avevano pubblicato un altro studio svolto su animali, condotto esattamente nelle stesse modalità, nel Journal of Medical and Biological Research. (2) In quello studio, la somministrazione di Nandrolone Decanoato aveva causato un aumentato della pressione sanguigna dei ratti trattati, ma la co-somministrazione con Taurina aveva ridotto notevolmente questo incremento pressorio.
I ricercatori affermano che, alla luce delle evidenti prove relative alla provata sicurezza della somministrazione di Taurina nell’uomo, il loro studio solleva il presupposto che la Taurina potrebbe essere utile in determinate circostanze associate ad alti livelli di androgeni circolanti in cronico, come nei disturbi endocrini o nell’abuso di AAS da parte degli atleti.
Bisognerebbe comunque prendere i risultati del presente studio con la dovuta cautela. Si tratta pur sempre di uno studio svolto su animali (ratti) e le variabili nella risultante che l’applicazione di questo protocollo integrativo potrebbero avere sull’uomo potrebbero essere assai diverse.
La dose di Nandrolone Decanoato somministrata ai ratti rapportata al dosaggio umano ammonterebbe a 200mg a settimana. Dosaggio decisamente minimale rispetto ai livelli (stupidamente) raggiunti dalla media degli utilizzatori odierni.
Per quanto modesto possa essere il dosaggio di Nandrolone Decanoato utilizzato nel presente studio, la dose di Taurina che i ricercatori somministravano ai ratti era decisamente elevata. Il dosaggio rapportato all’essere umano equivarrebbe a circa 15g di Taurina al giorno. In questo caso la dose è nettamente inferiore agli standard di utilizzo per questo aminoacido (dose massima raccomandata 8g/die). Giova ricordare che vi sono studi in attesa di conferma che indicano come un eccesso di Taurina negli adulti provochi ipertensione (il che andrebbe ad annullare i presunti benefici sulla pressione in co-somministrazione con AAS) e problemi gastrointestinali (diarrea e ulcera peptica). Sembra inoltre che l’assunzione di alti dosaggi di Taurina contribuiscano ad aggravare la psoriasi (comparsa di prurito, squame e diffusione delle lesioni cutanee).
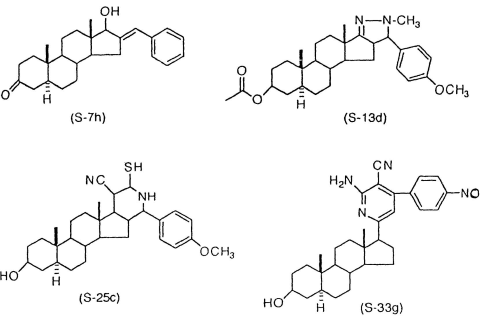
Nel mio quotidiano scartabellare la letteratura scientifica, mi sono imbattuto in un articolo pubblicato da due chimici egiziani nel 2000 su “Scientia Pharmazeutica”. I ricercatori hanno sintetizzato nuovi steroidi anabolizzanti modificando la struttura molecolare del DHT. Uno di questi AAS – l’S7h – risulta essere molto interessante .(1)
Nella pubblicazione, i ricercatori hanno descritto i quattro AAS la cui struttura è riportata di seguito. Tutte e quattro le molecola condividono la struttura simile al DHT con modifiche nell’anello D dello scheletro carbonioso.
La molecola più interessante sintetizzata dai ricercatori – l’S7h – è anche quella con meno modifiche.
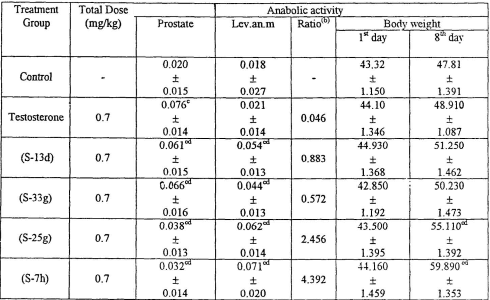
I ricercatori hanno testato l’effetto biologico dei loro nuovi AAS con una variante del test di Hershberger, utilizzando ratti Wistar maschi di 21 giorni di età pre-pubescente. I ricercatori hanno iniettato quotidianamente agli animali Testosterone, S7h, S13d, S25c o S33g alla dose di 0,7mg per Kg al giorno. Un gruppo di controllo è stato trattato con iniezioni senza sostanze attive.
Tutte le molecole hanno mostrato di avere un effetto anabolizzante, ma l’S7h ha mostrato gli effetti migliori nello studio. La somministrazione di S7h ha aumentato il peso corporeo degli animali trattati del 296% rispetto al gruppo di controllo. L’effetto del S7h sul muscolo levator ani [Lev.an.m] è stato del 246% maggiore di quello riscontrato con il Testosterone.
Allo stesso tempo, l’S7h ha avuto un effetto minore sulla dimensione della prostata rispetto a tutte le altre molecole somministrate. Ciò suggerisce che l’S7h ha un potenziale di effetti collaterali androgeni relativamente basso.
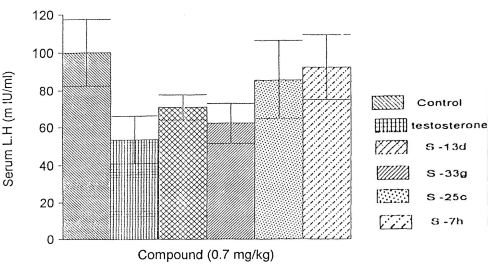
Infine, l’S7h ha esercitato un impatto contenuto sulla soppressione della sintesi di LH e Testosterone, sebbene l’effetto si è presentato e non è trascurabile.
Non ho al momento alcun dato su una sua applicazione sull’uomo, e dato che sono passati diciannove anni dallo studio dubito che ciò possa avvenire nell’ordinario ambiente accademico. E’ più probabile che questa molecola possa comparire nel mercato “grigio” come Designer steroid e/o nel mercato “nero”…
Sembrerebbe che gli utilizzatori di AAS soffrano maggiormente di Candida albicans rispetto ai non utilizzatori. Grazie a uno studio svolto in vitro dai ricercatori della Pontificia Università Cattolica di Parana, e pubblicato su “Steroids”, si ha una visione più chiara del perché ciò avvenga. Lo studio ha mostrato che gli AAS aumentano l’”aggressività” della Candida albicans.(1)
Esiste un’associazione tra l’uso di AAS e gengive infiammate – o parodontite. La parodontite può essere il risultato di un’infezione cronica da funghi come la Candida albicans.
In un recente studio, ricercatori argentini hanno scoperto che la Candida tropicalis, la Candida spp. La Candida parapsilosi hanno un incidenza rispettivamente di 4.3, 5.6 e 14.8 volte maggiore nelle gengive degli utilizzatori di AAS rispetto ai non utilizzatori.(2)
Funghi come la Candida albicans hanno recettori sterolici. Si ipotizza, quindi, che gli AAS possano alterare la biochimica di questa classe di funghi attraverso l’interazione con i suddetti recettori.
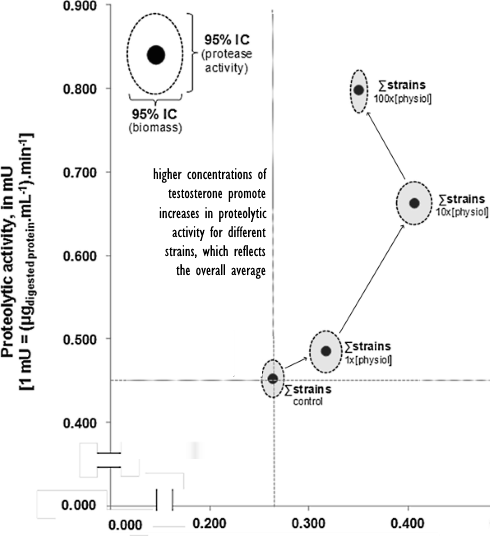
I ricercatori hanno svolto l’esperimento in vitro su 3 ceppi di Candida albicans – ceppi ATCC90028, SC5314 e 15A2. Essi hanno determinato la crescita dei funghi sulla base della loro biomassa e la loro capacità di degradare le proteine nel loro ambiente di coltura- la loro capacità proteolitica. Ed è proprio a causa di questa capacità proteolitica che la Candida albicans è dannosa per la salute dell’organismo ospite. Questi funghi causano il deterioramento dei tessuti sui quali proliferano.
I ricercatori hanno osservato i funghi in un ambiente privo di ormoni [Ctrl] e in uno con una concentrazione elevata di Testosterone, dalle 10 alle 100 volte superiore ai livelli normali riscontrabili in un essere umano. Tali concentrazioni possono essere presenti nel corpo degli utilizzatori di AAS.
I ricercatori hanno esposto i funghi anche al Nandrolone ed ad altri AAS.
La risultante fu che maggiori erano le concentrazione di Testosterone o Nandrolone (e altri AAS) nelle Piastre di Petri, maggiore era la capacità proteolitica dei funghi Candida. Le seguenti figure si riferiscono a tutti i ceppi studiati.
Gli AAS hanno contemporaneamente aumentato la biomassa e l’attività proteolitica con caratteristiche dose-dipendenti.
La combinazione di questi due fattori può essere considerata a ragione un effetto collaterale ulteriormente dannoso per gli utilizzatori aumentando la possibilità di marcata degradazione locale dei tessuti e la persistenza del fungo nelle aree infettate.
Se non avete ancora letto le precedenti parti componenti questa serie di articoli vi invito a farlo prima di procedere con la lettura di questa quarta ed ultima parte: 1° Parte – 2° Parte – 3° Parte.
Farmacocinetica, Farmacodinamica e Feedback Negativi
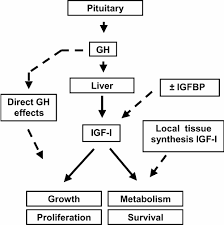
Come già detto, il fegato rappresenta il principale bersaglio del GH, il quale è il principale regolatore della sintesi epatica di IGF-1. Per causare tale effetto, il GH si lega con i GHR localizzati nel dominio extracellulare degli epatociti stimolando successivamente la produzione di IGF-1 endocrino tramite la trascrizione genica, utilizzando la via di segnalazione JAK-STAT. Inoltre, è stato dimostrato che la somministrazione di GH causa una rapida sovraregolazione dell’mRNA del IGF-1 nel fegato.[338]
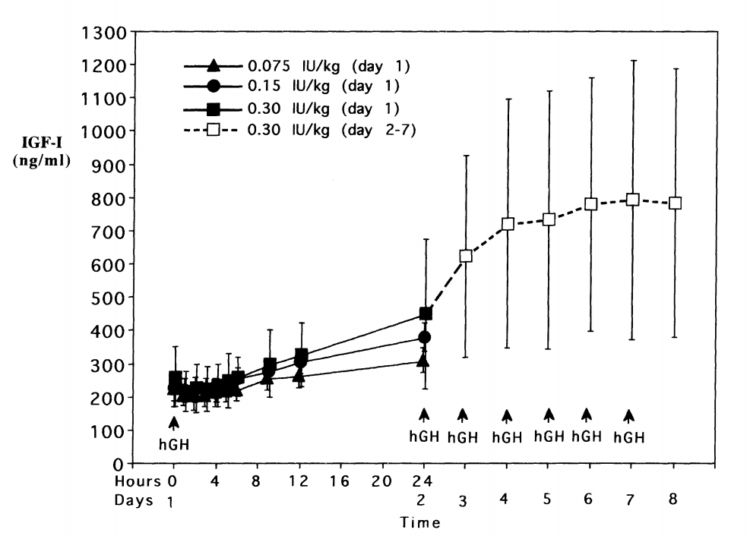
Aumenti dei livelli serici di IGF-1 si verificano molto rapidamente anche in presenza di un grande bolo di rHGH. Incrementi significativi di IGF-1 sono già osservabili dopo 6-12h dall’iniezione.[339] Questi livelli serici di IGF-1 continuano ad aumentare fino a raggiungere il loro punto di saturazione dose-dipendente entro 4-7 giorni, anche quando si utilizzano dosi estremamente elevate che ammontano a 20-30UI al giorno di rHGH.[340] In particolare, il punto di saturazione si è rivelato essere compreso nell’intervallo dei 700-800 ng/mL e sembra suggerire che i livelli endocrini di IGF-1 hanno un tetto massimo negli adulti sani. I meccanismi esatti devono ancora essere chiariti, ma sono probabilmente il risultato dei complessi meccanismi di controllo intrinseci all’Asse GH/IGF-1. Coloro i quali desiderano elevare i livelli endocrini di IGF-1 al fine di ottenerne un vantaggio sull’ipertrofia dovrebbe tenerlo a mente, in quanto vi è un punto in cui l’uso di dosi maggiori di rHGH semplicemente non si traducono in elevati livelli serici di IGF-1. Qui di seguito ho riportato il grafico dello studio di Tanaka il quale mostra la relazione tra l’rhGH e i livelli serici di IGF-:
Ora, vorrei dedicarmi brevemente all’analisi dell’azione del IGF-1autocrino e del perché esso rappresenti un mediatore cruciale del processo ipertrofico, prima di tornare nuovamente a discutere su questioni inerenti alla farmacodinamica e farmacocinetica. La segnalazione recettoriale del IGF-1 è unica nel suo genere, e questo lo si deve al fatto che utilizza due percorsi distinti per stimolare la proliferazione o la differenziazione.[341-343] Questo è un comportamento abbastanza interessante, poiché nessun altro membro della famiglia dei fattori di crescita ha dimostrato di agire in tal modo. Poiché la proliferazione e la differenziazione sono processi opposti, inizialmente era difficile per i ricercatori capire come un singolo fattore di crescita, attraverso un singolo recettore, potesse inviare un segnale che attivasse entrambi.[294] Da quando sono state fatte queste prime scoperte, è stato ulteriormente chiarito che l’IGF-1 non svolge simultaneamente queste azioni. Test su varie linee di coltura cellulare hanno dimostrato che gli effetti proliferativi arrivano prima, durando tra le 24 e le 36 ore. È solo dopo questa fase proliferativa iniziale che si verifica la differenziazione miogenica.[344]
Gli effetti proliferativi mediati dall’IGF-1 sui mioblasti sono noti sin dagli anni ’70, quando vennero osservati per la prima volta nelle cellule epatiche di ratto.[345] Questa stimolazione proliferativa del IGF-1 si traduce in un aumento del numero di cellule, nei livelli di proteine, nella sintesi del DNA, nell’assorbimento di aminoacidico, nell’assorbimento del glucosio e nella soppressione della proteolisi.[346] Nelle colture cellulari umane, l’IGF-1 ha anche dimostrato di aumentare la dimensione dei miotubi indipendentemente dal fatto che i mioblasti proliferino attivamente o che la proliferazione sia cessata. Regola la dimensione dei miotubi attivando la sintesi proteica, inibendo la degradazione proteica e inducendo la fusione delle cellule di riserva.[347-348] La capacità dell’IGF di sopprimere la proteolisi nel muscolo scheletrico, la scomposizione delle proteine in aminoacidi, è stata dimostrata innumerevoli volte nel corso degli anni.[349-352] È stato anche dimostrato che l’IGF-1 induce la proliferazione e la differenziazione delle cellule satelliti in miociti maturi, come determinato da un aumento del numero di miofibre nucleate a livello centrale rispetto a quelle periferiche.[148,353-354]
La capacità dell’IGF-1 autocrino di causare la differenziazione dei mioblasti è stata in realtà una scoperta che potremmo definire quasi “ibrida” dal momento che degli studi svolti negli anni ’60 avevano mostrato che questo effetto si verifica con alti livelli di Insulina.[355] Successivamente è stato dimostrato che gli IGF sono stimolatori molto più potenti nella differenziazione miogenica rispetto all’Insulina e si è concluso che la stessa Insulina agisce realmente come un analogo dell’IGF-1 in questo sistema.[356-357] Gli effetti di differenziazione dati dall’IGF-1 autocrino sono bifasici, con basse concentrazioni che stimolano progressivamente la differenziazione dei mioblasti mentre concentrazioni molto elevate mostrano una cessazione dell’attività di differenziazione. Il limite massimo per la differenziazione sembra attestarsi a circa 100ng/mL per l’IGF-1 e 300ng/mL per IGF-2.[358] Questo effetto non è legato alla proliferazione, poiché non si osservano ulteriori aumenti nel numero complessivo delle cellule.[294] È possibile che le molecole di segnalazione coinvolte nella regolazione negativa del sistema miogenico siano aumentate, ma questa è una affermazione puramente speculativa.[359-360]
La somministrazione di rHGH eleva l’espressione dell’mRNA dell’IGF-1 nel muscolo scheletrico in numerosi modelli cellulari, umani e animali.[127,150,361-364] Ciò avviene abbastanza rapidamente, entro 60 minuti dall’iniezione sottocutanea di rHGH ed i picchi sono segnalati tra le 6 e le 12 ore post iniezione.[363] In questo particolare modello animale citato, il raddoppio della dose di GH non ha portato ad ulteriori aumentati dei livelli di mRNA dell’IGF-1, il che suggerisce che esiste un sistema di regolazione che determina quanto GH sia necessario per stimolare al massimo l’espressione locale dell’IGF-1 nel muscolo scheletrico. In precedenza si è potuto appurare che la differenziazione dei miociti mediata dall’IGF-1 si arresta quando le concentrazioni locali raggiungono circa i 100ng/mL, ma quanto GH è necessario per raggiungere il punto di saturazione dell’espressione dell’mRNA dell’IGF-1?
Gli studi sui miociti umani mostrano che il GH aumenta l’espressione dell’mRNA dell’IGF-1 entro 30-60 minuti con picchi molto più rapidi rispetto a quelli osservati negli studi sugli animali, entro 1-2 ore, usando la via di segnalazione JAK / STAT5b.[365] Questi livelli elevati di mRNA hanno dimostrato di durare fino a 48 ore dopo una singola esposizione al GH. La quantità di GH necessaria per stimolare al massimo l’espressione dell’mRNA dell’IGF-1 è risultata essere una dose compresa tra i 7,5ng/mL e 30ng/ml [366], con una dose media efficace che si attesta a 3ng/ml. Questi numeri sono in linea con gli intervalli di dose fisiologica osservati negli animali, che sono effettivamente compresi tra i 2-100 ng/mL.[367] Inoltre, si collocano esattamente in linea con quanto si osserva endogenamente nell’uomo, con concentrazioni normali di picco comprese tra i 22,4 e 32,4ng/mL.[368-369,436] Ci sono stati casi in cui gli uomini presi in esame hanno mostrato concentrazioni di picco leggermente più alte, ma questi devono essere considerati valori anomali.[370] In ogni caso, ciò che questi dati tendono a suggerire è che il corpo umano è particolarmente adatto a gestire livelli naturali di picco della secrezioni di GH endogeno. Cercare di incidere ulteriormente il sistema elevando i livelli di GH oltre quelli endogeni, unicamente per tentare di potenziare i processi ipertrofici, potrebbe in realtà non tradursi nell’effetto desiderato.
Gli studi che mettono a confronto le infusioni locali con le infusioni sistemiche di GH o IGF-1 sono un po’ più difficili da trovare di quanto si vorrebbe. Le poche sperimentazioni sugli animali che sono riuscito a trovare indicano che l’infusione diretta di GH o IGF-1 nei tessuti bersaglio determina un aumento della massa muscolare. Questo aumento dell’ipertrofia si verifica anche senza che il muscolo bersaglio sia stato sottoposto ad attività motoria.[371-372] Gli studi dimostrano anche che le iniezioni locali di GH portano a livelli sostanzialmente più alti nell’espressione dell’mRNA dell’ IGF-1 locale rispetto alle iniezioni locali di IGF-1, di un fattore di oltre venti.[127] Sono riuscito a trovare uno studio nel quale si confrontavano le risposte dei ratti (attivi e non) all’infusione locale di IGF-1. Il gruppo “IGF-1 plus training” ha mostrato un aumento sia della massa muscolare che della forza locale maggiore rispetto al semplice trattamento in isolamento.[373] Quindi, anche se limitata, la letteratura disponibile è apparentemente in grado di dimostrare che le iniezione locali di GH o IGF-1 hanno effettivamente valore.
Ne ho già parlato diverse volte ma, nel tentativo di imprimere ulteriormente questo concetto, è necessario ricordarsi che i livelli autocrini di IGF-1 sembrano essere molto più importanti dei livelli endocrini di IGF-1 in relazione alla regolazione della massa muscolare. Oltre a questo punto, la sovraespressione dell’IGF-1 autocrino nel muscolo provoca l’ipertrofia delle fibre.[374] La sovraespressione dell’IGF-1 autocrino ha anche mostrato effetti anti-catabolici, con modelli animali tendenti a mostrare una resistenza generale all’atrofia muscolare normalmente osservata con l’invecchiamento.[375] L’IGF-1 localizzato fornisce anche capacità rigenerative indipendenti dall’età nelle cellule muscolari.[376]
Vi sono anche alcune prove convincenti che suggeriscono che l’IGF-1 endocrino agisce direttamente come un regolatore di feedback negativo sulla produzione di IGF-1 autocrino. Questo meccanismo di feedback negativo è dipendente dal pathway PI3K/Akt [377-378]. Inoltre, elevati livelli di IGF-1 endocrino possono anche agire indirettamente per sopprimere la produzione di IGF-1 autocrino. Quindi, in altre parole, non solo l’IGF-1 endocrino ha un impatto diretto minore sulla regolazione della massa muscolare, ma può anche sopprimere l’IGF-1 autocrino che ha impatti maggiori sull’ipertrofia.
Elevati livelli di IGF-1 circolante e, nello specifico, di IGF-1 libero elevati agiscono in modo negativo sul GH determinando un tasso di soppressione della produzione di IGF-1 autocrino a valle.[379] Non è del tutto chiaro, tuttavia, se la regolazione negativa dell’IGF-1 modifichi l’emivita dell’mRNA dell’IGF-1 o influenzi direttamente l’espressione del gene IGF-1. Oltre a questo, è stato anche dimostrato che l’espressione dell’IGF-1 autocrino è sottoregolata nelle cellule muscolari dopo trattamento con IGF-1.[366] È stato anche dimostrato che l’espressione epatica dell’mRNA dell’IGF-1 è sottoregolata dall’esposizione acuta all’IGF-1.[127] Quindi, mantenere livelli endocrini il più possibile soppressi con rispettiva dose di rHGH, elevando contemporaneamente i livelli autocrini, dovrebbe essere un fattore prioritario in un protocollo di GH volto all’ipertrofia.
Il GH è pulsatile per natura sia nell’uomo che nelle specie animali. Quindi, sarebbe logico pensare che molti dei processi intrinseci del corpo saranno tarati in modo tale da rispondere in maniera ottimale all’esposizione al GH in modo simile. In accordo con questa affermazione è stato dimostrato che solo la somministrazione di GH pulsatile, e non l’infusione continua, ha la capacità di stimolare massimamente l’espressione dell’mRNA dell’IGF-1 nel muscolo scheletrico.[366,380-381] È stato anche dimostrato che la somministrazione pulsatile porta ad un aumento del potenziale di crescita postnatale complessivo rispetto all’infusione continua.[89,382] La somministrazione pulsatile può anche portare a livelli endocrini di IGF-1 serici comparabili, o addirittura diminuiti [383], il che è vantaggioso a causa delle potenziali capacità di regolazione negativa che possiede sull’espressione dell’IGF-1 autocrino e che sono state discusse in precedenza. L’evidenza suggerisce anche che il picco stesso, e non necessariamente il numero di picchi, potrebbe essere della massima importanza per i tessuti bersaglio.[384] Per la massima crescita e potenziale ipertrofico, l’evidenza tende a suggerire che creare picchi di GH elevati, e quindi tornare ai livelli basali più volte al giorno, può essere preferibile rispetto a mantenerli elevati per periodi di tempo più lunghi. Questo pratica permette di riprodurre gli schemi secretori in vivo.
I pathways del GH coinvolti nell’anabolismo sono anche suscettibili alla desensibilizzazione, che è parte della fisiologia del GH endogeno.[385] A causa della natura intrinsecamente pulsatile del GH in vivo, l’attività dei recettori e dei pathways sono regolati da un impulso seguito da un periodo di inattività.[386] L’esposizione continua o ripetuta al GH senza un adeguato lasso di tempo refrattario comporterà livelli di attività fortemente soppressi. In effetti, nel corso degli anni numerosi studi hanno dimostrato che tale effetto si verifica. Le cellule ed il tessuto muscolare richiedono un periodo refrattario piuttosto lungo prima che la loro piena risposta al GH venga recuperata. Dopo l’esposizione al GH, le cellule muscolari non sono nemmeno in grado di rispondere alle successive dosi di GH. In realtà, occorrono due ore complete per riprendere parzialmente la reattività nei modelli cellulari, con un totale di 6-8 ore di astinenza dall’uso di GH necessarie per ripristinare la piena sensibilità.[366] Viceversa, quando il GH è micro-dosato in impulsi di dieci minuti, seguiti da intervalli di otto ore, è stato mostrato aumentare progressivamente l’mRNA dell’IGF-1 con ogni impulso successivo.[386]
Questo fenomeno è potenzialmente il risultato di una desensibilizzazione complessiva all’interno della via JAK-STAT5, poiché è stato dimostrato che l’esposizione al GH negli studi sulle cellule epatiche causa resistenza alla successiva attivazione della via STAT5 per 4-8 ore.[387-388] Questo lasso di tempo è sufficiente per sincronizzarsi abbastanza bene con ciò che è stato visto nei modelli di cellule miocitarie citati in precedenza. Nei modelli di cellule epatiche, il GH ha stimolato un significativo aumento dell’espressione del SOCS3, che è un potente inibitore dell’azione del GH.[389]. Poiché il GH non ha avuto alcun effetto sull’espressione del SOCS3 nelle cellule muscolari, questo deve essere un altro meccanismo causante il periodo refrattario. Questo meccanismo può essere dipeso dalla sottoregolazione dei GHR, dall’inibizione mediata da un’altra proteina SOCS, o dall’induzione di una tirosina fosfatasi che semplicemente inattiva la via JAK / STAT.[390] La via JAK-STAT5b, che come ricorderete è intimamente associata al muscolo scheletrico e all’espressione dell’IGF-1, è di natura transitoria – con attivazione massima raggiunta entro 10-30 minuti, seguita da un prolungato periodo di inattivazione.
Una scoperta piuttosto nuova di Xu et al. [391] ha dimostrato che anche distanziare le esposizioni al GH di cinque ore lasciava entrambi i percorsi a valle MEK1/2 e ERK1/2 significativamente soppressi rispetto a tutti i percorsi a monte, a causa di una potenziale disconnessione nella trasduzione del segnale . Ciò è di particolare interesse in quanto questi stessi due percorsi a valle sono stati coinvolti in modo significativo sia nella crescita sia nella proliferazione.[392-393] È stato anche scoperto che l’attivazione indotta da GH di STAT1 e STAT3 è stata desensibilizzata, ma l’esposizione all’Insulina inverte la desensibilizzazione osservata in tutti i percorsi interessati. Anche se non sto per trattare approfonditamente l’Insulina, ci sono un paio di importanti punti da dovere prendere in considerazione. Bisogna comprendere innanzitutto che ci sono molti obiettivi a valle del recettore del GH e molti di questi hanno il potenziale per essere desensibilizzati dopo l’esposizione al GH. Bisogna comprendi anche che l’Insulina possiede l’abilità unica di risensibilizzare molti di questi percorsi. Ciò ha un senso vista la relazione tipo yin-yang tra i due composti. È noto che il GH e l’Insulina possiedono una relazione anabolica sinergica a causa di molti effetti che esercitano l’uno sull’altro. Questo sembra essere soltanto un’anteprima di uno di questi effetti.
Asse GH/IGF-1 – Relazione con altri ormoni
Prima di passare alle note conclusive, vorrei trattare brevemente alcuni altri ormoni connessi a diverso grado con l’Asse GH/IGF-1. Per prima cosa, voglio trattare brevemente l’Asse Tiroideo dal momento che l’inserimento di composti tiroidei insieme al GH è una pratica comune anche durante i protocolli di massa.
Il muscolo scheletrico è il principale bersaglio di segnalazione dell’ormone tiroideo, con trasportatori degli ormoni tiroidei e enzimi di conversione espressi localmente.[394] È ben noto che il GH potenzia la deiodinazione periferica che converte il T4 in T3, riducendo così il T4 e il reverse T3, aumentando contemporaneamente i livelli di T3.[395-398] Ciò che molte persone non riescono a capire è che questo è un effetto transitorio, e studi a lungo termine sembrano indicare che gli effetti mediati dal GH sulla conversione periferica si stabilizzino con il tempo.[399-402]
Via ubiquitina/proteasoma
Invece di proseguire ulteriormente su questo, avendo già trattato la questione nel dettaglio in un mio vecchio articolo, preferirei concentrarmi su alcune pubblicazioni relative alla tiroide che non vengono discusse abbastanza spesso. Gli ormoni tiroidei, per loro natura, sono composti tendenzialmente catabolici in quanto stimolano la disgregazione proteica dell’intero corpo in misura maggiore rispetto alla sintesi proteica.[403] A livello locale, nel muscolo scheletrico stimolano un aumento dell’attività all’interno della via ubiquitina/proteasoma, che è ampiamente coinvolta nella proteolisi.[404-406] Il risultato di questo è un tasso accelerato del turnover proteico e una perdita netta complessiva degli aminoacidi situati all’interno dei muscoli scheletrici.
Inoltre, negli esseri umani, sia gli stati di ipertiroidismo che di ipotiroidismo sono stati associati a livelli di IGF-1 soppressi con una tendenza alla normalizzazione quando viene ristabilita una condizione di eutiroidismo. L’ipertiroidismo è anche associato ad una bassa attività di legame recettoriale del GH, che si ipotizza essere il risultato di una ridotta capacità di elaborazione dei recettori del GH.[407] E’ stato anche ipotizzato che l’ipertiroidismo sia in grado anche di accelerare la clearance del GH urinario.[408] Inoltre, studi su animali hanno dimostrato che gli ormoni tiroidei possono avere importanti effetti soppressivi sulla sintesi di IGF-1 stimolata con il GH.[409] Ovviamente, a causa della complessa relazione che l’Asse Tiroideo ha con l’Asse GH/IGF-1, raggruppando tutte le interazioni che hanno tra loro in pochi paragrafi, trattare l’argomento diventerebbe poco pratico. Tuttavia, quando il corpo della letteratura scientifica viene esaminato nella sua interezza, ci sono molte prove che suggeriscono che la supplementazione con composti tiroidei esogeni potrebbe non essere l’ideale quando l’obiettivo di un individuo è l’ipertrofia, anche se la regolazione del dosaggio dei tiroidei in tale contesto rimane la misura di “sicurezza” più intelligente visto l’impatto negativo sulla funzionalità tiroidea dato dal GH. Comunque, per tutti coloro che sono interessati ad approfondire questo argomento, consiglio di iniziare con la review nella nota seguente.[410]
Miostatina
Mi piacerebbe trattare anche la Miostatin, che rappresenta un argomento molto discusso nei vari forum di BodyBuilding presenti in rete. La sua fama proviene dai risultati ipertrofici espressi dai bovini privi per mutazione del gene della Miostatina, i quali mostrano una massa muscolare significativamente maggiore rispetto ai loro simili non mutati.[411] La Miostatina, un fattore di crescita e differenziazione appartenente alla superfamiglia dei TGF-beta, ha dimostrato di inibire selettivamente la miogenesi, in gran parte tramite il suo effetto soppressivo sulla proliferazione dei mioblasti.[412] È espressa e secreta prevalentemente dal muscolo scheletrico. Come molti sanno, se riesci a sopprimere o inibire la Miostatina, di conseguenza il potenziale ipertrofico aumenta significativamente.
Le mutazioni della Miostatina sono state osservate sia negli animali che nell’uomo. Queste mutazioni del gene della Miostatina portano ad un fenotipo ipertrofico negli animali, come accennato in precedenza.[413-415] L’Asse GH/IGF-1 e la Miostatina sembrano avere una relazione regolativa diretta tra loro, come osservato nei pazienti affetti contemporaneamente da GHD e HIV che mostrano marcati aumenti nell’espressione dell’mRNA della Miostatina.[416] Quindi, è possibile che attraverso una supplementazione di dosi sovrafisiologiche di rHGH si possa indurre una diminuzione dell’mRNA della Miostatina [209,417-419]? Sfortunatamente, nonostante la presenza di alcuni casi studio selezionati, non credo che si abbiano abbastanza dati in questo momento per sapere se ciò possa dare risultati apprezzabili in seguito alla sua applicazione.
Quello che sappiamo è che aumenti dell’espressione dell’mRNA dell’IGF-1 e le concentrazioni circolanti di IGF-1 sono state osservati dopo inibizione della Miostatina.[419-421] Sappiamo anche che l’inibizione della Miostatina tende a causare l’ipertrofia attraverso molte delle stesse modalità osservate con l’IGF-1 autocrino, cioè l’aumento della sintesi proteica e l’attivazione delle cellule satelliti.[422-425] E sappiamo anche che l’ipertrofia indotta dalla sovraespressione dell’IGF-1 o dall’inibizione della Miostatina utilizza la stessa identica via – PI3K/Akt/mTOR.[426-428] Tuttavia, l’IGF-1 non è un requisito per l’ipertrofia indotta dalla Follistatina, tranne nel caso di livelli di Insulina estremamente bassi – come ben sappiamo, la Follistatina è un inibitore della Miostatina [429]. E l’esposizione cronica al GH può in realtà portare ad un’espressione sovrastimolata della Miostatina e del suo recettore.[209]
Quindi quello che possiamo dire, con certezza, è che l’espressione della Miostatina non sarà un fattore diretto o indiretto per quanto riguarda il potenziamento dei processi ipertrofici, né dell’attività contrattile, nei muscoli scheletrici umani.[430] Proprio per questo motivo, non ritengo che sia un fattore sul quale gli atleti debbano eccessivamente concentrarsi, al di fuori di un utile arricchimento delle proprie conoscenze in materia.
Applicazioni pratiche e pensieri conclusivi
Arrivati a questo punto è mia intenzione unire tutto ciò che è stato esposto in questa serie di articoli e esporlo sotto forma di alcuni suggerimenti pratici rivolti a tutti coloro i quali vogliono semplicemente massimizzare la loro capacità ipertrofica.
Ora è chiaro che il GH possiede pochissimi, se non nulli, effetti diretti sull’ipertrofia. Pertanto, qualsiasi protocollo di massa che contempli il suo uso dovrà tenere in considerazione questo punto includendo gli AAS, i quali, per l’appunto, hanno anche una proficua sinergia con il GH. Sia la letteratura scientifica che i dati aneddotici dimostrano chiaramente che l’uso combinato di entrambi i composti ha un massimale ipertrofico significativamente più alto rispetto all’uso singolo. Personalmente, penso che i BodyBuilder dovrebbero sempre optare per l’utilizzo di una base di Testosterone e Boldenone (correttamente rapportati in base a contesto e alle caratteristiche individuali) anche in una fase “Bulk” nella quale viene inserito l’uso del GH. Il Trenbolone può essere considerato come parte “accessoria” di un protocollo di massa, a causa della sua intrinseca difficoltà gestazionale come sostanza anabolizzante. Dovrebbe essere usato con parsimonia e con cautela poiché, insieme ai suoi numerosi punti di forza come composto anabolizzante, presenta alcune limitazioni. Queste limitazioni derivano per lo più, come già accennato, dalla difficile gestione del composto in quanto esso non è facilmente tollerato da una buona parte degli individui. Quindi, se viene usato il Trenbolone, dovrebbe essere inserito calcolando con attenzione il dosaggio e la tolleranza individuale, anche per quanto concerne la tolleranza temporale individuale all’uso di tale composto.
Dopo un periodo d’uso prolungato di dosi sovrafisiologico di AAS (Ciclo+Bridge), è buona cosa procedere con l’interruzione o con una marcata riduzione del numero e degli AAS utilizzati. Detta in parole semplici, questa interruzione può contemplare una completa astinenza dagli AAS, svolgendo un adeguata PCT al fine di ristabilire una omeostasi ormonale fisiologica, o una transizione ad una TRT, metodologia comunemente chiamata “blast and cruise”. La struttura del protocollo di supplementazione farmacologica dovrebbe sempre seguire i principi del dosaggio minimo efficace con aumenti nei dosaggi degli AAS solo nel caso si sia raggiunto il limite di crescita con il precedente dosaggio, assicurandosi che tutte le altre variabili nello stile di vita siano correttamente regolate. L’utilizzo di questo approccio limita il rischio che si sviluppino effetti collaterali indesiderati sul lungo termine.
Quando si decide di usare il GH, la dove ce ne sia la possibilità economica, esso dovrebbe provenire da uno dei prodotti presenti nel mercato farmaceutico. Questi prodotti approvati devono superare anni di studi strettamente controllati per dimostrare la loro sicurezza, purezza ed efficacia su soggetti umani. I progressi tecnologici nel corso degli anni hanno reso molto più facile la produzione di rHGH. Per questo motivo, i produttori ora provengono da tutto il mondo. Spesso questi produttori realizzano ciò che viene definito “GH generico” sui forum, ma tale termine non mi piace molto. Definire qualcosa come “generico” implica che sia una replica perfetta di prodotti legati a specifici marchi farmaceutici approvati dell’agenzia del farmaco che hanno perso il cui brevetto è scaduto, il che non è il caso del GH. Infatti, a causa della natura estremamente complessa del processo di produzione del rHGH, per esempio, la FDA non consente nemmeno l’uso del termine “generico” quando si tratta di rHGH e utilizza invece il termine ” follow-on protein product ” o FOPP.
Spesso questi marchi off-label sono venduti ad un costo molto ridotto, ed è qui che sta il dilemma, in quanto questo può essere molto allettante. Tuttavia, con questo costo ridotto per il consumatore, non ci sarà nemmeno la garanzia del produttore su cosa ci sia realmente nella fiala o persino su come è stato prodotto. Il problema di fondo è che il processo di produzione del rHGH è estremamente complessa, ed è molto facile che nelle fasi di questo processo si commettano errori con conseguenti variazioni nella catena proteica che potenzialmente portano ad effetti indesiderati, o anche a risposte autoimmuni.
Spesso gli atleti si affidano semplicemente ai test serici per misurare i livelli di GH e/o IGF-1 al fine di concludere che un prodotto contenete GH sia “buono”, ma dobbiamo ricordarci che raggiungere livelli ematici ormonali elevati è la parte relativamente facile. Anche le molecole di GH che sono state alterate o danneggiate durante la produzione possono dare questo esito. Tuttavia, queste stesse molecole di GH danneggiate o mutate possono spesso stimolare risposte autoimmuni. Ciò potrebbe indurre il corpo ad avere una risposta recettoriale degradata, che può anche riflettersi sulla secrezione endogena nel tempo. [431-432] Rimane poi il problema del reale contenuto della vial o fiala, la quale può non presentare nessun principio attivo al suo interno.
Il GH dovrebbe essere usato in modo pulsatile, per mimare le condizioni in vivo. Tra queste iniezioni, deve esserci un periodo di refrattarietà o si deve consumare un pasto che abbia un buon stimolo sull’Insulina. Può anche essere utilizzata l’Insulina esogena al fine di bypassare molte delle limitazioni del periodo refrattario, ma questo va oltre lo scopo di questo articolo. Anche se il cumulo delle dosi giornaliere dovrebbe essere sovrafisiologico, le dosi individuali non hanno bisogno di essere ad alto dosaggio, poiché la massima stimolazione dell’IGF-1 autocrino nel tessuto muscolo scheletrico si verifica ben all’interno delle concentrazioni fisiologiche di GH. Aneddoticamente, sembra anche esserci un limite con il quale l’uso di rHGH diventa additivo in presenza di AAS. Potrebbe essere necessario un po’ di ponderata (e supportata da personale qualificato) auto-sperimentazione per scoprire dove si trova questa singola dose di saturazione, ma la maggior parte dei soggetti troverà questo limite tra le 4 e le 8 UI/die. Oltre questo dosaggio, la maggior parte degli utilizzatori tenderà a scoprire che la giustificazione dei costi e il rapporto rischio /beneficio tendono a diminuire rapidamente.
Non bisognerebbe passare troppo tempo a riflettere sul tempo delle iniezioni di GH, dal momento che gli aumenti dei livelli di IGF-1 autocrino avvengono rapidamente e possono rimanere elevati per giorni. Bisognerebbe concentrati invece sul programma di iniezione che si addice meglio al contesto della giornata, tenendo contemporaneamente presenti le linee guida per il periodo di refrattarietà del GH. Si possono anche prendere in considerazione piccole o grandi iniezioni, poiché alcuni potrebbero trovare più pratiche e funzionali iniezioni con un dosaggio inferiore e somministrate più frequenti mentre altri potrebbero preferire iniezioni con un dosaggio maggiore e somministrazioni meno frequenti. Naturalmente, maggiore è il contenuto dell’iniezione, maggiore è la probabilità che si superi la soglia massima di espressione dell’IGF-1 autocrino.
Massimizzare l’espressione dell’IGF-1 autocrino, mentre contemporaneamente si sopprimono i livelli di IGF-1 endocrino, sarà una priorità. Esistono prove a sostegno dell’ipotesi secondo cui un iniezione locale di GH possa aiutare a raggiungere questo obiettivo, con una conseguente minore possibilità di feedback negativo. Sono stati osservati aumenti significativi della massa muscolare in appena due settimane di iniezioni locali di IGF-1.[441]
La dove cause di forza maggiore non lo impediscano, è consigliabile evitare o comunque regolare attentamente l’uso di tutti quei composti che possono avere interazioni negative con l’uso del GH a fini ipertrofici. Inibitori della Aromatasi, Modulatori Selettivi del Recettore degli Estrogeni e T3 hanno tutti dimostrato di avere un potenziale effetto negativo sul processo globale ipertrofico legato al GH e per tale motivo dovrebbero essere usati con parsimonia, se non omessi del tutto dove possibile (vedi in particolare il T3).
Questo non dovrebbe sorprendere nessuno e non dovrebbe nemmeno essere troppo difficile da tenere a mente: allenarsi duramente, allenarsi in modo intelligente e allenarsi in modo coerente. Sebbene non sia stato affrontato direttamente nell’articolo, bisogna capire che l’allenamento contro resistenza ha impatti unici e additivi sull’ipertrofia. In effetti, alcuni di questi meccanismi non sono nemmeno mediati dall’Asse AR e/o GH/IGF-1. [433] Bisogna anche comprendere che non esiste una “magica” routine allenante universalmente applicabile, la chiave di volta sarà la coerenza nel garantire un carico di lavoro adeguato, con elementi di sovraccarico progressivo nel tempo assicurando un adeguato stimolo meccanico gestendo al meglio le variabili allenanti (intensità, volume, densità e intensità percepita). La logica composizione del piano allenante servirà a garantire lo stimolo ipertrofico di base che sarà coadiuvato dall’azione dei composti utilizzati.
Nonostante la mole e l’importanza delle informazioni presentate in questa serie di articoli, è necessario ricordare che i meccanismi d’azione ormonali sono governati da innumerevoli fattori. Anche esaminando l’intero corpo della letteratura scientifica equivarrebbe a poco più che accumulare una serie di utili nozioni garanti di assicurare una solida base conoscitiva sull’argomento la quale rappresenterà un punto di partenza intelligente per l’applicazione pratica, rimanendo sempre soggetti alle variabili di risposta individuale. Seguendo questa linea, i migliori risultati nella pratica spesso provengono da coloro i quali posseggono una buona conoscenza dei principi scientifici e la capacità innata di saperli applicare non solo su se stessi ma, soprattutto, su terzi. Infatti, molto raramente due persone rispondono in modo identico alla supplementazione di ormoni esogeni (e non solo), quindi, non bisogna assolutamente pensare che basti semplicemente applicare su se stessi o su terzi un protocollo che ha portato benefici realmente apprezzabili su un soggetto per ottenere la medesima risposta.
A tal fine, invito atleti e Preparatori a utilizzare queste pubblicazioni come punto di partenza per essere in grado di gestire l’applicazione pratica in modo più consapevole e produttivo. Inoltre, chi ne fosse in grado, può consultare il vasto numero di riferimenti riportati nel corso di queste pubblicazioni è tentare di ragionare sulle mie conclusioni. Ad ogni citazione presente in questa serie di articoli, assicuratevi che il riferimento elencato supporti effettivamente le affermazioni fatte. Mantenere sempre una mente aperta ma con i giusti “filtri”, e cercare di non credere per partito preso ad una singola opinione, specialmente di fronte alle nuove evidenze scientifiche. Infine, è buona cosa verificare sempre la veridicità di quanto è stato affermato.
Punti conclusivi per un corretto utilizzo della chimica e del GH a fini ipertrofici:
Usare il GH in combinazione con gli AAS
Usare GH e AAS di grado farmaceutico la dove ciò è possibile
Assicurarsi una base di Testosterone correttamente rapportata al Boldenone aggiungendo (in base a maturità e tolleranza) il Trenbolone
Iniettare il GH in modo pulsatile, considerare l’opzione delle iniezioni locali nei gruppi carenti
Mantenere un dosaggio complessivo ottimale di GH il quale si attesta tra le 4 e le 8UI/die
Evitare o regolare attentamente l’uso dei composti che possono interagire negativamente con i processi ipertrofici legati al uso di GH (vedi AI, SERM e T3)
Dopo un periodo di tempo (variabile) nel quale si è stati sottoposti a dosaggi ormonali sovrafisiologici optare per una PCT o per una TRT (a seconda delle proprie necessità e priorità)
Essere a conoscenza dei potenziali effetti collaterali legati all’auso/abuso di GH (nausea, vomito, cefalea, ritenzione idrica e sodica, edemi, parestesie, sindrome del tunnel carpale, rigidità articolare, dolori articolari, artrite, dolori muscolari, ipertensione, insulino-resistenza, diabete di tipo II, acromegalia, dilatazione addominale, ipertrofia cardiaca ecc…)
Svolgere regolarmente esami del sangue; sia durante i periodi di picco nell’uso della farmacologia sia nel periodo successivo (vedi PCT/OCT o TRT)
Gestire al meglio le variabili legate agli stressor ambientali, all’allenamento, all’alimentazione e al sonno.
Gabriel Bellizzi
Riferimenti:
338. Mathews LS, Norstedt G, Palmiter RD. Regulation of insulin-like growth factor I gene expression by growth hormone. Proc Natl Acad Sci U S A. 1986 Dec;83(24):9343-7.
339. Keller A, Wu Z, Kratzsch J, Keller E, Blum WF, Kniess A, Preiss R, Teichert J, Strasburger CJ, Bidlingmaier M. Pharmacokinetics and pharmacodynamics of GH: dependence on route and dosage of administration. Eur J Endocrinol. 2007 Jun;156(6):647-53.
340. Tanaka T, Seino Y, Fujieda K, Igarashi Y, Yokoya S, Tachibana K, Ogawa Y. Pharmacokinetics and metabolic effects of high-dose growth hormone administration in healthy adult men. Endocr J. 1999 Aug;46(4):605-12.
341. Quinn LS, Steinmetz B, Maas A, Ong L, Kaleko M. Type-1 insulin-like growth factor receptor overexpression produces dual effects on myoblast proliferation and differentiation. J Cell Physiol. 1994 Jun;159(3):387-98.
342. Coolican SA, Samuel DS, Ewton DZ, McWade FJ, Florini JR. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J Biol Chem. 1997 Mar 7;272(10):6653-62.
343. Foulstone EJ, Huser C, Crown AL, Holly JM, Stewart CE. Differential signalling mechanisms predisposing primary human skeletal muscle cells to altered proliferation and differentiation: roles of IGF-I and TNFalpha. Exp Cell Res. 2004 Mar 10;294(1):223-35.
344. Ewton DZ, Roof SL, Magri KA, McWade FJ, Florini JR. IGF-II is more active than IGF-I in stimulating L6A1 myogenesis: greater mitogenic actions of IGF-I delay differentiation. J Cell Physiol. 1994 Nov;161(2):277-84.
345. Florini JR, Nicholson ML, Dulak NC. Effects of peptide anabolic hormones on growth of myoblasts in culture. Endocrinology. 1977 Jul;101(1):32-41.
346. Laviola L, Natalicchio A, Giorgino F. The IGF-I signaling pathway. Curr Pharm Des. 2007;13(7):663-9. Review.
347. Jacquemin V, Furling D, Bigot A, Butler-Browne GS, Mouly V. IGF-1 induces human myotube hypertrophy by increasing cell recruitment. Exp Cell Res. 2004 Sep 10;299(1):148-58.
348. Jacquemin V, Butler-Browne GS, Furling D, Mouly V. IL-13 mediates the recruitment of reserve cells for fusion during IGF-1-induced hypertrophy of human myotubes. J Cell Sci. 2007 Feb 15;120(Pt 4):670-81. Epub 2007 Jan 30.
349. Ballard FJ, Francis GL. Effects of anabolic agents on protein breakdown in L6 myoblasts. Biochem J. 1983 Jan 15;210(1):243-9.
350. Ewton DZ, Falen SL, Florini JR. The type II insulin-like growth factor (IGF) receptor has low affinity for IGF-I analogs: pleiotypic actions of IGFs on myoblasts are apparently mediated by the type I receptor. Endocrinology. 1987 Jan;120(1):115-23.
351. Hembree JR, Hathaway MR, Dayton WR. Isolation and culture of fetal porcine myogenic cells and the effect of insulin, IGF-I, and sera on protein turnover in porcine myotube cultures. J Anim Sci. 1991 Aug;69(8):3241-50.
352. Hong D, Forsberg NE. Effects of serum and insulin-like growth factor I on protein degradation and protease gene expression in rat L8 myotubes. J Anim Sci. 1994 Sep;72(9):2279-88.
353. Florini JR, Ewton DZ, Roof SL. Insulin-like growth factor-I stimulates terminal myogenic differentiation by induction of myogenin gene expression. Mol Endocrinol. 1991 May;5(5):718-24.
354. Musarò A, McCullagh KJ, Naya FJ, Olson EN, Rosenthal N. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature. 1999 Aug 5;400(6744):581-5.
355. Haba GDL, Cooper GW, Elting V. HORMONAL REQUIREMENTS FOR MYOGENESIS OF STRIATED MUSCLE IN VITRO: INSULIN AND SOMATOTROPIN. Proceedings of the National Academy of Sciences of the United States of America. 1966;56(6):1719-1723.
356. Florini JR, Ewton DZ. Insulin acts as a somatomedin analog in stimulating myoblast growth in serum-free medium. In Vitro. 1981 Sep;17(9):763-8.
357. Schmid C, Steiner T, Froesch ER. Preferential enhancement of myoblast differentiation by insulin-like growth factors (IGF I and IGF II) in primary cultures of chicken embryonic cells. FEBS Lett. 1983 Sep 5;161(1):117-21.
358. Florini JR, Ewton DZ, Falen SL, Van Wyk JJ. Biphasic concentration dependency of stimulation of myoblast differentiation by somatomedins. Am J Physiol. 1986 May;250(5 Pt 1):C771-8.
359. Quinn LS, Ehsan M, Steinmetz B, Kaleko M. Ligand-dependent inhibition of myoblast differentiation by overexpression of the type-1 insulin-like growth factor receptor. J Cell Physiol. 1993 Sep;156(3):453-61.
360. Olson EN. Signal transduction pathways that regulate skeletal muscle gene expression. Mol Endocrinol. 1993 Nov;7(11):1369-78. Review.
361. Murphy LJ, Bell GI, Friesen HG. Growth hormone stimulates sequential induction of c-myc and insulin-like growth factor I expression in vivo. Endocrinology. 1987 May;120(5):1806-12.
362. Turner JD, Rotwein P, Novakofski J, Bechtel PJ. Induction of mRNA for IGF-I and -II during growth hormone-stimulated muscle hypertrophy. Am J Physiol. 1988 Oct;255(4 Pt 1):E513-7.
363. Isgaard J, Nilsson A, Vikman K, Isaksson OG. Growth hormone regulates the level of insulin-like growth factor-I mRNA in rat skeletal muscle. J Endocrinol. 1989 Jan;120(1):107-12.
364. Bichell DP, Kikuchi K, Rotwein P. Growth hormone rapidly activates insulin-like growth factor I gene transcription in vivo. Mol Endocrinol. 1992 Nov;6(11):1899-908.
365. Sadowski CL, Wheeler TT, Wang LH, Sadowski HB. GH regulation of IGF-I and suppressor of cytokine signaling gene expression in C2C12 skeletal muscle cells. Endocrinology. 2001 Sep;142(9):3890-900.
366. Frost RA, Nystrom GJ, Lang CH. Regulation of IGF-I mRNA and signal transducers and activators of transcription-3 and -5 (Stat-3 and -5) by GH in C2C12 myoblasts. Endocrinology. 2002 Feb;143(2):492-503.
367. MacLeod JN, Pampori NA, Shapiro BH. Sex differences in the ultradian pattern of plasma growth hormone concentrations in mice. J Endocrinol. 1991 Dec;131(3):395-9.
368. Rochiccioli P, Messina A, Tauber MT, Enjaume C. Correlation of the parameters of 24-hour growth hormone secretion with growth velocity in 93 children of varying height. Horm Res. 1989;31(3):115-8.
369. Hansen TK, Gravholt CH, ØRskov H, Rasmussen MH, Christiansen JS, Jørgensen JO. Dose dependency of the pharmacokinetics and acute lipolytic actions of growth hormone. J Clin Endocrinol Metab. 2002 Oct;87(10):4691-8.
370. Baum WF, Klöditz E, Hesse V, Jahreis G, Schneyer U, Giebler H. [Increase in spontaneous growth hormone secretion in asthmatic children–a symptom of atopic disposition?]. Kinderarztl Prax. 1993 Nov;61(9):323-8.
371. Adams GR, McCue SA. Localized infusion of IGF-I results in skeletal muscle hypertrophy in rats. J Appl Physiol (1985). 1998 May;84(5):1716-22.
372. Alzghoul MB, Gerrard D, Watkins BA, Hannon K. Ectopic expression of IGF-I and Shh by skeletal muscle inhibits disuse-mediated skeletal muscle atrophy and bone osteopenia in vivo. FASEB J. 2004 Jan;18(1):221-3. Epub 2003 Nov 3.
373. Lee S, Barton ER, Sweeney HL, Farrar RP. Viral expression of insulin-like growth factor-I enhances muscle hypertrophy in resistance-trained rats. J Appl Physiol (1985). 2004 Mar;96(3):1097-104. Erratum in: J Appl Physiol. 2004 Jun;96(6):2343.
374. Coleman ME, DeMayo F, Yin KC, Lee HM, Geske R, Montgomery C, Schwartz RJ. Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J Biol Chem. 1995 May 19;270(20):12109-16.
375. Barton-Davis ER, Shoturma DI, Musaro A, Rosenthal N, Sweeney HL. Viral mediated expression of insulin-like growth factor I blocks the aging-related loss of skeletal muscle function. Proc Natl Acad Sci U S A. 1998 Dec 22;95(26):15603-7.
376. Musarò A, McCullagh K, Paul A, Houghton L, Dobrowolny G, Molinaro M, Barton ER, Sweeney HL, Rosenthal N. Localized Igf-1 transgene expression sustains hypertrophy and regeneration in senescent skeletal muscle. Nat Genet. 2001 Feb;27(2):195-200
377. Lewis MI, Bulut Y, Biring MS, Da X, Fournier M. (1999) IGF-I administration prevents corticosteroids-induced diaphragm atrophy in emphysema . Am J Respir Crit Care Med 159:A580
378. Fournier M, Huang ZS, Cercek B, Li H, Bykhovskaya I, Lewis MI. (2000) Administration of insulin-like growth factor-1 (IGF-I) and corticosteroids in emphysematous hamsters: influences on diaphragm IGF-I . Am J Respir Crit Care Med 161:A18
379. Shavlakadze T, Grounds M. Of bears, frogs, meat, mice and men: complexity of factors affecting skeletal muscle mass and fat. Bioessays. 2006 Oct;28(10):994-1009. Review.
380. Maiter D, Underwood LE, Maes M, Davenport ML, Ketelslegers JM. Different effects of intermittent and continuous growth hormone (GH) administration on serum somatomedin-C/insulin-like growth factor I and liver GH receptors in hypophysectomized rats. Endocrinology. 1988 Aug;123(2):1053-9.
381. Isgaard J, Carlsson L, Isaksson OG, Jansson JO. Pulsatile intravenous growth hormone (GH) infusion to hypophysectomized rats increases insulin-like growth factor I messenger ribonucleic acid in skeletal tissues more effectively than continuous GH infusion. Endocrinology. 1988 Dec;123(6):2605-10.
382. Clark RG, Jansson JO, Isaksson O, Robinson IC. Intravenous growth hormone: growth responses to patterned infusions in hypophysectomized rats. J Endocrinol. 1985 Jan;104(1):53-61.
383. Bick T, Hochberg Z, Amit T, Isaksson OG, Jansson JO. Roles of pulsatility and continuity of growth hormone (GH) administration in the regulation of hepatic GH-receptors, and circulating GH-binding protein and insulin-like growth factor-I. Endocrinology. 1992 Jul;131(1):423-9.
384. Weltman A, Weltman JY, Schurrer R, Evans WS, Veldhuis JD, Rogol AD. Endurance training amplifies the pulsatile release of growth hormone: effects of training intensity. J Appl Physiol (1985). 1992 Jun;72(6):2188-96.
385. Flores-Morales A, Greenhalgh CJ, Norstedt G, Rico-Bautista E. Negative regulation of growth hormone receptor signaling. Mol Endocrinol. 2006 Feb;20(2):241-53. Epub 2005 Jul 21. Review.
386. Hartman ML, Veldhuis JD, Thorner MO. Normal control of growth hormone secretion. Horm Res. 1993;40(1-3):37-47. Review.
387. Fernández L, Flores-Morales A, Lahuna O, Sliva D, Norstedt G, Haldosén LA, Mode A, Gustafsson JA. Desensitization of the growth hormone-induced Janus kinase 2 (Jak 2)/signal transducer and activator of transcription 5 (Stat5)-signaling pathway requires protein synthesis and phospholipase C. Endocrinology. 1998 Apr;139(4):1815-24.
388. Gebert CA, Park SH, Waxman DJ. Termination of growth hormone pulse-induced STAT5b signaling. Mol Endocrinol. 1999 Jan;13(1):38-56.
389. Ram PA, Waxman DJ. SOCS/CIS protein inhibition of growth hormone-stimulated STAT5 signaling by multiple mechanisms. J Biol Chem. 1999 Dec 10;274(50):35553-61.
390. Ram PA, Waxman DJ. Role of the cytokine-inducible SH2 protein CIS in desensitization of STAT5b signaling by continuous growth hormone. J Biol Chem.2000 Dec 15;275(50):39487-96.
391. Xu J, Keeton AB, Franklin JL, Li X, Venable DY, Frank SJ, Messina JL. Insulin enhances growth hormone induction of the MEK/ERK signaling pathway. J Biol Chem. 2006 Jan 13;281(2):982-92. Epub 2005 Nov 4.
392. Lewis TS, Shapiro PS, Ahn NG. Signal transduction through MAP kinase cascades. Adv Cancer Res. 1998;74:49-139. Review.
394. Mebis L, Paletta D, Debaveye Y, Ellger B, Langouche L, D’Hoore A, Darras VM, Visser TJ, Van den Berghe G. Expression of thyroid hormone transporters during critical illness. Eur J Endocrinol. 2009 Aug;161(2):243-50.
395. Jørgensen JO, Pedersen SA, Laurberg P, Weeke J, Skakkebaek NE, Christiansen JS. Effects of growth hormone therapy on thyroid function of growth hormone-deficient adults with and without concomitant thyroxine-substituted central hypothyroidism. J Clin Endocrinol Metab. 1989 Dec;69(6):1127-32.
396. Jørgensen JO, Pedersen SB, Børglum J, Møller N, Schmitz O, Christiansen JS, Richelsen B. Fuel metabolism, energy expenditure, and thyroid function in growth hormone-treated obese women: a double-blind placebo-controlled study. Metabolism. 1994 Jul;43(7):872-7.
397. Wolthers T, Grøftne T, Møller N, Christiansen JS, Orskov H, Weeke J, Jørgensen JO. Calorigenic effects of growth hormone: the role of thyroid hormones. J Clin Endocrinol Metab. 1996 Apr;81(4):1416-9.
398. Feldt-Rasmussen U. Interactions between growth hormone and the thyroid gland — with special reference to biochemical diagnosis. Curr Med Chem. 2007;14(26):2783-8. Review.
399. Kalina-Faska B, Kalina M, Koehler B. Effects of recombinant growth hormone therapy on thyroid hormone concentrations. Int J Clin Pharmacol Ther. 2004 Jan;42(1):30-4.
400. Hubina E, Mersebach H, Rasmussen AK, Juul A, Sneppen SB, Góth MI, Feldt-Rasmussen U. Effect of growth hormone replacement therapy on pituitary hormone secretion and hormone replacement therapies in GHD adults. Horm Res. 2004;61(5):211-7. Epub 2004 Jan 30.
401. Seminara S, Stagi S, Candura L, Scrivano M, Lenzi L, Nanni L, Pagliai F, Chiarelli F. Changes of thyroid function during long-term hGH therapy in GHD children. A possible relationship with catch-up growth? Horm Metab Res. 2005 Dec;37(12):751-6.
402. Losa M, Scavini M, Gatti E, Rossini A, Madaschi S, Formenti I, Caumo A, Stidley CA, Lanzi R. Long-term effects of growth hormone replacement therapy on thyroid function in adults with growth hormone deficiency. Thyroid. 2008 Dec;18(12):1249-54.
403. Müller MJ, Seitz HJ. Thyroid hormone action on intermediary metabolism. Part III. Protein metabolism in hyper- and hypothyroidism. Klin Wochenschr. 1984 Feb 1;62(3):97-102.
404. Tawa NE Jr, Odessey R, Goldberg AL. Inhibitors of the proteasome reduce the accelerated proteolysis in atrophying rat skeletal muscles. J Clin Invest. 1997 Jul 1;100(1):197-203. PubMed PMID: 9202072
405. Dace A, Zhao L, Park KS, et al. Hormone binding induces rapid proteasome-mediated degradation of thyroid hormone receptors. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(16):8985-8990.
406. Clément K, Viguerie N, Diehn M, Alizadeh A, Barbe P, Thalamas C, Storey JD, Brown PO, Barsh GS, Langin D. In vivo regulation of human skeletal muscle gene expression by thyroid hormone. Genome Res. 2002 Feb;12(2):281-91.
407. Miell JP, Taylor AM, Zini M, Maheshwari HG, Ross RJ, Valcavi R. Effects of hypothyroidism and hyperthyroidism on insulin-like growth factors (IGFs) and growth hormone- and IGF-binding proteins. J Clin Endocrinol Metab. 1993 Apr;76(4):950-5.
408. Murao K, Takahara J, Sato M, Tamaki M, Niimi M, Ishida T. Relationship between thyroid functions and urinary growth hormone secretion in patients with hyper- and hypothyroidism. Endocr J. 1994 Oct;41(5):517-22.
409. Wolf M, Ingbar SH, Moses AC. Thyroid hormone and growth hormone interact to regulate insulin-like growth factor-I messenger ribonucleic acid and circulating levels in the rat. Endocrinology. 1989 Dec;125(6):2905-14.
410. Laron Z. Interactions between the thyroid hormones and the hormones of the growth hormone axis. Pediatr Endocrinol Rev. 2003 Dec;1 Suppl 2:244-9-discussion 250. Review.
411. Fiems LO. Double Muscling in Cattle: Genes, Husbandry, Carcasses and Meat. Animals : an Open Access Journal from MDPI. 2012;2(3):472-506.
412. Langley B, Thomas M, Bishop A, Sharma M, Gilmour S, Kambadur R. Myostatin inhibits myoblast differentiation by down-regulating MyoD expression. J Biol Chem. 2002 Dec 20;277(51):49831-40. Epub 2002 Sep 18.
413. McPherron AC, Lee SJ. Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci U S A. 1997 Nov 11;94(23):12457-61.
414. Schuelke M, Wagner KR, Stolz LE, Hübner C, Riebel T, Kömen W, Braun T, Tobin JF, Lee SJ. Myostatin mutation associated with gross muscle hypertrophy in a child. N Engl J Med. 2004 Jun 24;350(26):2682-8.
415. Clop A, Marcq F, Takeda H, Pirottin D, Tordoir X, Bibé B, Bouix J, Caiment F, Elsen JM, Eychenne F, Larzul C, Laville E, Meish F, Milenkovic D, Tobin J, Charlier C, Georges M. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat Genet. 2006 Jul;38(7):813-8.
416. Gonzalez-Cadavid NF, Taylor WE, Yarasheski K, Sinha-Hikim I, Ma K, Ezzat S, Shen R, Lalani R, Asa S, Mamita M, Nair G, Arver S, Bhasin S. Organization of the human myostatin gene and expression in healthy men and HIV-infected men with muscle wasting. Proc Natl Acad Sci U S A. 1998 Dec 8;95(25):14938-43.
417. Liu W, Thomas SG, Asa SL, Gonzalez-Cadavid N, Bhasin S, Ezzat S. Myostatin is a skeletal muscle target of growth hormone anabolic action. J Clin Endocrinol Metab. 2003 Nov;88(11):5490-6.
418. Oldham JM, Osepchook CC, Jeanplong F, Falconer SJ, Matthews KG, Conaglen JV, Gerrard DF, Smith HK, Wilkins RJ, Bass JJ, McMahon CD. The decrease in mature myostatin protein in male skeletal muscle is developmentally regulated by growth hormone. J Physiol. 2009 Feb 1;587(3):669-77.
419. Williams NG, Interlichia JP, Jackson MF, Hwang D, Cohen P, Rodgers BD. Endocrine actions of myostatin: systemic regulation of the IGF and IGF binding protein axis. Endocrinology. 2011 Jan;152(1):172-80.
420. Winbanks CE, Weeks KL, Thomson RE, Sepulveda PV, Beyer C, Qian H, Chen JL, Allen JM, Lancaster GI, Febbraio MA, Harrison CA, McMullen JR, Chamberlain JS, Gregorevic P. Follistatin-mediated skeletal muscle hypertrophy is regulated by Smad3 and mTOR independently of myostatin. J Cell Biol. 2012 Jun 25;197(7):997-1008.
421. Lach-Trifilieff E, Minetti GC, Sheppard K, Ibebunjo C, Feige JN, Hartmann S, Brachat S, Rivet H, Koelbing C, Morvan F, Hatakeyama S, Glass DJ. An antibody blocking activin type II receptors induces strong skeletal muscle hypertrophy and protects from atrophy. Mol Cell Biol. 2014 Feb;34(4):606-18.
422. Bark TH, McNurlan MA, Lang CH, Garlick PJ. Increased protein synthesis after acute IGF-I or insulin infusion is localized to muscle in mice. Am J Physiol. 1998 Jul;275(1 Pt 1):E118-23.
423. Barton-Davis ER, Shoturma DI, Sweeney HL. Contribution of satellite cells to IGF-I induced hypertrophy of skeletal muscle. Acta Physiol Scand. 1999 Dec;167(4):301-5.
424. Suryawan A, Frank JW, Nguyen HV, Davis TA. Expression of the TGF-beta family of ligands is developmentally regulated in skeletal muscle of neonatal rats. Pediatr Res. 2006 Feb;59(2):175-9.
425. Gilson H, Schakman O, Kalista S, Lause P, Tsuchida K, Thissen JP. Follistatin induces muscle hypertrophy through satellite cell proliferation and inhibition of both myostatin and activin. Am J Physiol Endocrinol Metab. 2009 Jul;297(1):E157-64.
426. Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, Zlotchenko E, Scrimgeour A, Lawrence JC, Glass DJ, Yancopoulos GD. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol. 2001 Nov;3(11):1014-9.
428. Kalista S, Schakman O, Gilson H, Lause P, Demeulder B, Bertrand L, Pende M, Thissen JP. The type 1 insulin-like growth factor receptor (IGF-IR) pathway is mandatory for the follistatin-induced skeletal muscle hypertrophy. Endocrinology. 2012 Jan;153(1):241-53.
429. Barbé C, Kalista S, Loumaye A, Ritvos O, Lause P, Ferracin B, Thissen JP. Role of IGF-I in follistatin-induced skeletal muscle hypertrophy. Am J Physiol Endocrinol Metab. 2015 Sep 15;309(6):E557-67. doi: 10.1152/ajpendo.00098.2015. Epub 2015 Jul 28.
430. Coffey VG, Shield A, Canny BJ, Carey KA, Cameron-Smith D, Hawley JA. Interaction of contractile activity and training history on mRNA abundance in skeletal muscle from trained athletes. Am J Physiol Endocrinol Metab. 2006 May;290(5):E849-55.
431. Moore WV, Leppert P. Role of aggregated human growth hormone (hGH) in development of antibodies to hGH. J Clin Endocrinol Metab. 1980 Oct;51(4):691-7
432. Dannies PS. Protein folding and deficiencies caused by dominant-negative mutants of hormones. Vitam Horm. 2000;58:1-26. Review.
433. DeVol DL, Rotwein P, Sadow JL, Novakofski J, Bechtel PJ. Activation of insulin-like growth factor gene expression during work-induced skeletal muscle growth. Am J Physiol. 1990 Jul;259(1 Pt 1):E89-95.
434. Hermansen K, Bengtsen M, Kjær M, Vestergaard P, Jørgensen JOL. Impact of GH administration on athletic performance in healthy young adults: A systematic review and meta-analysis of placebo-controlled trials. Growth Horm IGF Res. 2017 Jun;34:38-44.
435. de Souza GL, Hallak J. Anabolic steroids and male infertility: a comprehensive review. BJU Int. 2011 Dec;108(11):1860-5.
436. Kraemer WJ, Marchitelli L, Gordon SE, Harman E, Dziados JE, Mello R, Frykman P, McCurry D, Fleck SJ. Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol (1985). 1990 Oct;69(4):1442-50.
437. Pfeffer LA, Brisson BK, Lei H, Barton ER. The insulin-like growth factor (IGF)-I E-peptides modulate cell entry of the mature IGF-I protein. Mol Biol Cell. 2009 Sep;20(17):3810-7.
438. Mills P, Dominique JC, Lafrenière JF, Bouchentouf M, Tremblay JP. A synthetic mechano growth factor E Peptide enhances myogenic precursor cell transplantation success. Am J Transplant. 2007 Oct;7(10):2247-59.
439. Brisson BK, Barton ER. Insulin-like growth factor-I E-peptide activity is dependent on the IGF-I receptor. PLoS One. 2012;7(9):e45588.
440. Brisson BK, Spinazzola J, Park S, Barton ER. Viral expression of insulin-like growth factor I E-peptides increases skeletal muscle mass but at the expense of strength. Am J Physiol Endocrinol Metab. 2014 Apr 15;306(8):E965-74.
441. Goldspink G, Harridge S. Mechanism for adaptation in skeletal muscle In: Komi P, editor. Strength and power in sport: Olympic encyclopedia of sports medicine. Oxford: Blackwell; 2002. p. 231–51.
442. Janssen JA, Hofland LJ, Strasburger CJ, van den Dungen ES, Thevis M. Potency of Full-Length MGF to Induce Maximal Activation of the IGF-I R Is Similar to Recombinant Human IGF-I at High Equimolar Concentrations. PLoS One. 2016 Mar 18;11(3):e0150453.
Se non avete ancora letto la prima e la seconda parte di questa serie di articoli vi invito a farlo prima di procedere con la lettura di questa terza parte: 1° Parte – 2° Parte.
Effetti diretti di GH e IGF-1 sull’ipertrofia muscolare
Andiamo dritti al punto – di per sé, il GH non causa direttamente ipertrofia muscolare. Questa caratteristica è stata ampiamente osservata per decenni e, finora, nessuno studio attendibile è stato in grado di mostrare un chiaro effetto della somministrazione di rHGH a medio-lungo termine sull’ipertrofia – anche in dosi sovrafisiologiche somministrate ad atleti di alto livello sottoposti ad intensi allenamenti contro resistenza.
Il fatto che questa correlazione non sia stata dimostrata non è certamente dovuto alla mancanza di tentativi in tal senso. Diversi gruppi di ricerca nel corso degli anni hanno tentato di identificare un ipotetica ipertrofia GH-mediata in soggetti adulti sani [48,195-199] o in soggetti anziani [188,198,200-203] senza successo, o in modo inconcludente. Inoltre, è stato dimostrato che gli aumenti della secrezione di GH indotta dall’esercizio in acuto non hanno prodotto cambiamenti nella MPS o nell’ipertrofia [204-205]. È interessante notare che, sebbene la quantità di studi presenti in letteratura siano significativamente meno comuni di quelli in cui è stato somministrato GH, anche la somministrazione sistemica di rhIGF-1 non ha prodotto alcun effetto ipertrofico misurabile sia in soggetti giovani [63,206] che negli anziani [207- 208].
Vi sono prove recenti che suggeriscono che l’esposizione cronica al GH aumenta l’espressione delle vie intramuscolari responsabili dell’atrofia.[209] È ovvio che molte delle caratteristiche anaboliche dimostrate dal GH possono essere compensate da questa maggiore espressione del catabolismo, che potrebbe essere il motivo per cui l’esposizione cronica al GH non porta all’ipertrofia. Potrebbe anche essere responsabile del perché l’esposizione cronica al GH può produrre muscoli meno efficienti e più deboli.[210] Molto semplicemente, questo potrebbe essere ancora un altro fattore nella lunga serie di effetti regolatori negativi intrinseci nell’Asse GH/IGF-1, ma ulteriori studi dovranno essere condotti per chiarire maggiormente questa ipotesi.
Per quelli che difficilmente accettano le informazioni riportate in questa serie di articoli, e vogliono addentrarsi loro stessi nella letteratura scientifica sperando di trovare informazioni che confutino quanto da me riportato, voglio esporre un concetto molto importante da me già espresso nella prima parte di questa serie di articoli. La maggior parte degli studi che hanno preso in esame il GH segnalano un aumento della massa magra nei soggetti dei gruppi trattati con tale ormone. Quindi, ad un lettore inesperto, potrebbero risultare infondate le affermazioni sulla mancanza di un effetto ipertrofico diretto del GH. Bisogna ricordare, però, che il GH causa una non indifferente ritenzione idrica, oltre ad aumentare la massa dei tessuti molli della quale parlerò brevemente. Nello specifico, il GH aumenta la ritenzione di sodio e, di conseguenza, dell’acqua extracellulare, in modo dose-dipendente, attraverso i suoi effetti sul sistema renina-angiotensina.[211] Questi incrementi nella ritenzione idrica e sodica sono stati osservati anche con la somministrazione di IGF-1, poiché l’IGF-1 stesso sembra essere un regolatore chiave del tasso di escrezione renale di sodio.[83,212-213] Quindi il punto della questione è che bisogna stare molto attenti nel trarre conclusioni quando non si ha ben chiara la differenza tra aumento della massa magra e crescita effettiva del tessuto muscolare.
Effetti indiretti di GH e IGF-1 sull’ipertrofia muscolare
Nella sezione precedente si è dimostrato che il GH e l’IGF-1, da soli, non hanno alcun impatto diretto sull’ipertrofia. Tuttavia, questo non significa che questi peptidi non abbiano un ruolo nei processi ipertrofici. L’obiettivo principale di questa sezione è quello di spiegare alcuni dei molti meccanismi che interessano in modo indiretto il GH e l’IGF-1 nei processi ipertrofici, molti dei quali saranno fattori importanti per gli atleti interessati a massimizzare il loro potenziale ipertrofico.
L’Ormone della Crescita è un potente stimolatore della sintesi di collagene sia nei tendini che nei muscoli. Questo effetto è probabilmente mediato dalla capacità dell’IGF-1 autocrino di stimolare i fibroblasti per sintetizzarlo.[214-215] In realtà, questo processo avviene senza che il peptide influenzi la sintesi proteica muscolare, nonostante i livelli di IGF-1 circolante e di quello locale siano significativamente più alti. Questo effetto è anche indotto indipendentemente dall’allenamento contro resistenza ed è stato persino osservato in soggetti immobilizzati hai quali è stato somministrato il GH.[216] La componente connettiva del muscolo scheletrico è vitale per la trasmissione della forza, che è prodotta dalle fibre muscolari, ai tendini e alle ossa affinché si verifichi il movimento. In particolare, il collagene è un importante componente nella trasmissione della forza della matrice extracellulare, che viene continuamente sottoposta a carichi intesi durante i movimenti.
A causa dei potenti effetti del GH sui componenti della matrice extracellulare, si può chiaramente iniziare a capire il perché gli aneddoti nel corso degli anni suggerivano che l’aggiunta del GH in una preparazione farmacologica produceva impatti positivi sulla riduzione del dolore tendineo-articolare. D’altra parte però, questo potrebbe anche essere un fattore che contribuisce in primo luogo al motivo per cui vari effetti collaterali sono segnalati dagli utilizzatori di GH come l’edema dei tessuti molli, il dolore articolare e la sindrome del tunnel carpale.[217-219] Ci sono anche molti che credono che il GH possa accelerare i tempi di recupero dagli infortuni, ma questo è un argomento complesso che verrà discusso in futuro.
Gli impatti del GH sulla sintesi di collagene potrebbero anche essere di grande interesse per gli atleti di forza che non sono necessariamente interessati all’ipertrofia, ma il cui obiettivo principale è la creazione di una condizione favorevole allo spostamento del carico massimale. Stimolare la sintesi del collagene potrebbe aiutare a rafforzare l’intero sistema di supporto dei muscoli scheletrici. Ora, vale la pena aggiungere una piccola nota di chiarimento. Nonostante questo suoni positivo in linea di principio, la supplementazione di GH non ha mai portato direttamente a guadagni di forza in nessuno degli studi svolti su soggetti sani, coprendo vari gruppi.[48,50,184,196-198,200-201,203,220-221] Naturalmente, se il GH fosse usato insieme ad un composto con la capacità di aumentare direttamente la forza, non è difficile supporre che in questo caso l’effetto addizionale potrebbe fare del GH un prezioso componente della preparazione.
Decorina
La Decorina è una proteina strutturale, che risiede principalmente nella matrice extracellulare del muscolo scheletrico e il cui ruolo è correlato alla crescita e alla riparazione dei muscoli.[222-223] Qualche decennio fa è stato dimostrato per la prima volta che la somministrazione di GH potrebbe aumentare direttamente l’espressione del gene della Decorina negli animali [224]. Recentemente, è stato dimostrato che questo effetto si manifesta anche in soggetti umani sottoposti ad allenamenti ricreativi.[225] Nello studio più recente, i livelli di Decorina sono fortemente correlati a quelli di PIIINP, che induce la stimolazione del Decorina GH-mediata che può essere coinvolta nel processo di assemblaggio della matrice di collagene osseo. Questo effetto è più pronunciato negli uomini rispetto alle donne e può essere un sottoprodotto dei livelli più alti di IGF-1 osservati negli uomini, sebbene questa affermazione sia speculativa. L’aumento dell’espressione di Decorina non è stato alterato dall’aggiunta di Testosterone, quindi questo effetto è indipendente dagli androgeni. Dopo aver visto gli effetti che il GH ha sulla sintesi di collagene e Decorina, sta diventando piuttosto chiaro che l’asse GH/IGF-1 è molto più importante per rafforzare la matrice extracellulare di supporto piuttosto che contribuire direttamente alla crescita del tessuto muscolare.
Adenosina Trifosfato (ATP)
La somministrazione acuta di GH in soggetti sani ha anche dimostrato di causare una maggiore produzione di ATP mitocondriale e una maggiore attività della citrato sintasi nel muscolo scheletrico, con una maggiore abbondanza di mRNA muscolari codificanti l’IGF-1.[226] Non solo questo potrebbe essere un fattore che contribuisce al motivo per cui il GH potrebbe avere la capacità di promuovere un aumento dei tassi di spesa energetica giornaliera ma potrebbe anche essere coinvolto nello spostamento verso la preferenza dei lipidi come substrato energetico. Anche se, come abbiamo visto in precedenza, il corpo della letteratura non supporta questa pratica, l’aumento della produzione di ATP mitocondriale potrebbe avere un ruolo nella capacità aerobica.[227]
È stato dimostrato che il GH promuove la fusione dei mioblasti con i miotubi in modelli cellulari [84], un effetto completamente indipendente dalla sovraregolazione locale del IGF-1. Per capire perché questo possa essere importante, bisogna approfondire maggiormente la questione dei fattori cellulari coinvolti nell’ipertrofia del muscolo scheletrico. L’ipertrofia dei muscoli scheletrici negli esseri umani si basa sulle cellule satelliti, che sono cellule dormienti situate all’interno delle miofibre, proprio sotto lo strato di lamina basale nella matrice extracellulare.[148] Una volta attivate queste cellule satellite, spesso con l’esercizio fisico o il danno muscolare, proliferano. Dopo la proliferazione, queste cellule satellite migrano verso siti ove si trova il danno differenziandosi e fondendosi con le miofibre esistenti che forniscono nuovi nuclei per l’ipertrofia e la riparazione. [228] Non è ancora del tutto chiaro se il GH abbia un effetto diretto sulla proliferazione e la differenziazione delle cellule satelliti.[229-230] Vale anche la pena di affermare che ciò che si osserva nelle colture cellulari potrebbe non essere del tutto indicativo di ciò che accade in vivo a causa di vari fattori esterni che non possono essere spiegati in condizioni di laboratorio.
Ci sono due fasi distinte in questa fusione dei mioblasti che si verifica.[85] Il primo sarebbe lo stadio iniziale della differenziazione in cui un sottoinsieme di cellule mononucleate si fondono per formare miotubi nascenti (fusione mioblasto/mioblasto). Questo è seguito dal secondo stadio che coinvolge ulteriori cellule disponibili che si fondono con questi miotubi nascenti e dove avviene la crescita muscolare effettiva (fusione mioblasto/miotubo). È all’interno di quest’ultimo stadio in cui il GH esercita i suoi effetti.
Questa è una scoperta piuttosto interessante, ma ancora una volta, numerosi studi sugli esseri umani non sono riusciti a dimostrare un effetto ipertrofico del GH in condizioni reali. Quindi, possiamo probabilmente dedurre da ciò che gli effetti che il GH ha sulla fusione dei nascenti miotubi non si traducono direttamente nell’ipertrofia. Tuttavia, cosa accadrebbe se si aggiungesse un’altra variabile nell’equazione in grado di creare un ambiente in cui esistessero numeri di celle satelliti potenziati, creando più materiale sul quale l’azione del GH possa manifestarsi?[231]
Introduzione degli AAS
A differenza del GH e del IGF, l’uso degli AAS ha mostrato di avere un impatto pronunciato su ipertrofia e forza. Questo è stato ben noto per decenni anche perché sono stati usati e abusati da atleti fin dalla loro creazione negli anni ’30 (con buona pace dei “puritani” della “old school”).[232] La famiglia degli AAS consiste in un gruppo di potenti composti sintetici che sono simili nella struttura chimica al Testosterone e/o al suo derivato 5α-ridotto (DHT). Vari tipi di singoli composti AAS sono stati creati nel corso degli anni usando come base la molecola naturale di Testosterone manipolandola, quindi, attraverso l’aggiunta di un gruppo etile, metile, idrossile o benzile in uno o più siti lungo la sua struttura.[233-234 ] Alcune delle varianti AAS più facilmente riconoscibili includono i composti metilati in C-17, i quali sono notoriamente dotati di un elevata biodisponibilità orale, e le varianti del 19-nortestosterone nelle quali viene rimosso il gruppo 19-metilico dalla molecola di Testosterone nel tentativo di aumentare la sua attività anabolica diminuendo al contempo la sua androgenicità e tendenza all’aromatizzazione. Conosciuti anche come “19-norsteroidi”, questa famiglia include ben note varianti di AAS come il Nandrolone ed il Trenbolone.
Molte di questi composti sono nati come risultato di un desiderio di aumentare le caratteristiche anaboliche del Testosterone a livello muscolare, abbassando allo stesso tempo gli effetti collaterali androgenici intrinsecamente inerenti alla molecola di Testosterone.[235] In generale, i rischi complessivi degli effetti collaterali derivanti dall’uso cronico di AAS sembrano essere relativamente bassi rispetto a molte sostanze socialmente accettate come l’alcol, il tabacco e vari farmaci da prescrizione.[233,236] Ovviamente, l’uso e l’abuso sono termini che si escludono a vicenda e abusare di qualsiasi sostanza tende a creare un ambiente a più alto rischio di effetti collaterali. A meno che non sia diversamente specificato, da questo punto in poi, parlerò specificamente del Testosterone in quanto è l’ormone sessuale endogeno maschile nativo, nonché l’androgeno più approfonditamente studiato in letteratura. Tanto per avere un riferimento, i maschi adulti sani producono una media di 14-77mg/settimana di Testosterone endogeno. [435]
Testosterone
Il Testosterone è un noto regolatore della massa muscolare, e gli aumenti della massa muscolare mediati dal Testosterone sono associati all’ipertrofia delle fibre, così come ad un aumento delle cellule satelliti e del numero mio nucleare.[231,237-240] Il muscolo scheletrico sembra essere uno dei tessuti più reattivi al Testosterone e si stima che i livelli circolanti di Testosterone rappresentino il fattore causale significativo (incidenza del 40-75%) dei guadagni nella massa muscolare osservati in studi di controllo randomizzati. Se ricordate ciò che è stato affermato e dimostrato nella precedente sezione, il GH possiede un’abilità molto particolare in quanto può aumentare la fusione dei mioblasti durante il processo ipertrofico. La massimizzazione di questa capacità si basa sull’avere un numero adeguato di cellule satellitari disponibili.
I risultati mostrano costantemente che i trattamenti con Testosterone determinano aumenti dose-dipendenti della massa e della forza del muscolo scheletrico, indipendentemente dal fatto che i soggetti presi in esame siano maschi più giovani o più anziani.[241-243] Al contrario, negli studi con soggetti sani giovani in cui il Testosterone endogeno è stato soppresso artificialmente, la forza e la composizione corporea hanno entrambi subito peggioramenti significativi.[244] Per ribadire il nostro precedente punto, gli aumenti della massa muscolare mediati dal Testosterone sono basati sull’ipertrofia e non sono il risultato di una transizione della fibra (cambiando le fibre di tipo I in fibre di tipo II o viceversa) o di una sua scissione.[245] A causa dei loro meccanismi anabolici unici e non sovrapposti, la somministrazione di Testosterone e l’allenamento contro resistenza hanno anche mostrato effetti sinergici e additivi l’uno sull’altro per quanto riguarda lo stimolo dell’aumento della massa muscolare.[246]
Recettore degli Androgeni (AR)
Gli Androgeni mediano principalmente i loro effetti attraverso il gene del recettore degli androgeni (AR) che è espresso in mioblasti, miofibre e cellule satelliti.[247-248] I AR sono stati rilevati anche nelle cellule che supportano i muscoli, come i fibroblasti e le cellule endoteliali. La densità dei AR sembra essere specifica per i gruppi muscolari, con l’allenamento contro resistenza e l’uso di AAS che hanno la capacità di influenzare il numero di AR presenti in questi gruppi muscolari. Oltre ai suoi effetti sulla densità dei AR, l’uso di AAS ha anche dimostrato la capacità di influenzare i livelli di attività dei AR sia in modo acuto che sul lungo termine.[249-250] Questi sono fattori piuttosto importanti da considerare quando ci si imbatte in individui che sostengono che il precedente utilizzo di AAS non offre necessariamente un vantaggio competitivo permanente.
A causa della complessità dell’argomento, ci sono state diverse ipotesi riguardo ai meccanismi con cui l’AAS esercita le sue azioni anaboliche sul muscolo scheletrico.[251] È stato dimostrato che i trattamenti con Testosterone aumentano i tassi di sintesi proteica muscolare (MPS) [252], riducono i tassi di degradazione delle proteine [253] facendo persino in modo che il corpo utilizzi più efficientemente gli amminoacidi prontamente disponibili. Quindi, ancora una volta, questo è un sistema abbastanza complesso che può anche essere semplificato ricordando che gli AAS promuovono l’anabolismo muscolare attraverso la loro capacità di incidere positivamente sull’equilibrio degli aminoacidi.
Via Wnt/B-catenina
È generalmente accettato che gli AAS esercitino i loro effetti anabolizzanti attraverso il legame e attivazione del AR che successivamente attiva cascate di segnalazione a valle che coinvolgono la via Wnt/β-catenina.[254-256]. I Wingless-INT (Wnt) sono una famiglia di glicoproteine secrete che regolano la proliferazione e la differenziazione cellulare.[257-258] I modelli cellulari hanno mostrato che il AR forma un complesso con la beta-catenina che si potenzia in presenza di AAS. [259-260] Una volta attivato questo complesso, migra nei nuclei in cui regola l’espressione dei geni target e la differenziazione delle cellule satelliti.[261-262] Questo è anche il pathway degli AAS in gran parte responsabile della miogenesi, la formazione del tessuto muscolare.[263-265]
Vale la pena notare che l’AAS possiede anche caratteristiche non genomiche che possono influenzare rapidamente numerosi processi ormonali e metabolici al di fuori del classico legame con il recettore. In letteratura sono stati riportati casi di maschi adulti con disturbi di insensibilità agli androgeni, causati da mutazioni del AR, che hanno risposto al Testosterone in modo molto simile ai soggetti sani. Questi casi studio rafforzano l’ipotesi che gli effetti anabolici dell’AAS possano essere mediati indipendentemente dal AR.[266] Le azioni non genomiche degli Androgeni possono in realtà essere un argomento piuttosto affascinante, ma che va oltre lo scopo previsto per questo articolo. Per coloro che vogliono approfondire l’argomento, consiglio di iniziare con le seguenti note di riferimento.[267-268]
AAS e potenziale sinergico con l’Asse GH/IGF-1
Ho riportato molte informazioni utili fino a questo punto, ma è qui che le cose cominciano davvero a farsi interessanti. A questo punto una domanda logica sarebbe: esistono studi svolti su soggetti sani nei quali si sono confrontate le differenze tra trattamenti singoli con GH o Androgeni e trattamenti combinati? Fortunatamente per noi, la risposta è “sì” in quanto vi sono stati alcuni studi, principalmente utilizzando soggetti anziani, sia maschi che femmine. I risultati di ognuno di questi studi dimostrano chiaramente che il GH ha un effetto additivo sui benefici consolidati che la terapia con ormoni sessuali fornisce: l’ipertrofia, la lipolisi, la sintesi del collagene, la funzione fisica, la qualità della vita e altri vari indicatori di prestazione.[187-188,269-270] Dato che è abbastanza chiaro che esiste un effetto additivo, vediamo se è possibile approfondire ulteriormente l’argomento al fine di scoprire alcuni dei meccanismi sottostanti che operano nella sinergia tra Androgeni e GH.
GHRH
Deve essere chiaro che il Testosterone, di per sé, ha un effetto additivo sull’intero Asse GH/IGF-1. Questo è stato osservato sia in modelli umani che animali, con la somministrazione di Testosterone che porta ad un aumento dei livelli circolanti di GH e IGF-1.[241,271-276] Viceversa, il deficit di Testosterone è comunemente associato a livelli significativamente ridotti di IGF-1.[277] L’effetto stimolante del Testosterone sull’Asse GH/IGF-1 sembra essere mediato a livello ipotalamico da una promozione della funzionalità del GHRH.[278]
Inoltre, e questo è bene ricordarlo, gli AAS non aromatizzabili sembrano non possedere lo stesso effetto stimolante sull’Asse GH/IGF-1.[279] Gli inibitori dell’aromatasi (AI), progettati per sopprimere il processo di aromatizzazione degli Androgeni, hanno dimostrato di attenuare direttamente la stimolazione del GH seguente la somministrazione di Testosterone. Questi indizi forniscono prove abbastanza convincenti sul fatto che gli estrogeni locali, derivanti dall’aromatizzazione, svolgono un ruolo fondamentale nella regolazione della secrezione di GH nei maschi.[280-281] Poiché l’aromatasi non è espressa nel fegato, gli AI non influenzano l’azione epatica del GH, mentre influenzano la sua azione a livello del sistema centrale.[282-283] Tuttavia i modulatori selettivi del recettore degli estrogeni (SERM) sono ancora più soppressivi in quanto agiscono ad entrambi i livelli a causa del loro meccanismo d’azione.[284-285]
Anche gli Androgeni che aumentano i livelli serici di Estrogeni, come il Nandrolone (Nor-Estrogeni), mostrano un effetto minimo sui livelli sistemici di GH e IGF-1 rispetto al Testosterone.[286] Ciò è legato ipoteticamente sia al fatto che il Nandrolone è poco soggetto all’aromatizzazione e sia al fatto che la forma estrogenica derivata (Nor-Estrogeno) abbia un ridotto potenziale di attività.[287] Ricordiamoci che l’aromatizzazione sembra essere il passo più cruciale nella stimolazione ipotalamica mediata dagli Androgeni. Ora, è ovvio che un utilizzatore di rHGH deve preoccuparsi parzialmente di questo effetto rispetto ad un non utilizzatore, considerando il fatto che, in tal caso, i livelli ormonali sono controllati quasi esclusivamente da mezzi esogeni. Detto questo, è sempre utile analizzare il quadro generale, soprattutto se l’obiettivo è massimizzare l’ipertrofia.
GHR
Un altro potenziale motivo legato agli aumenti dei livelli di GH e IGF osservati durante i trattamenti con Testosterone, è rappresentato dagli effetti diretti di quest’ultimo sui GHR. Sia gli studi sull’uomo che sugli animali hanno fornito prove del fatto che il Testosterone modifica i GHR sia nel fegato che nei tessuti periferici, migliorandone l’espressione.[288-289] Inoltre, soggetti umani ipopituitari e ipogonadici sottoposti a trattamenti con GH hanno mostrato una aumentata risposta sia all’IGF-1 locale che all’espressione del gene del recettore degli androgeni quando sottoposti anche a trattamento con Testosterone.[187,290-291] Inoltre, i maschi ipopituitari sottoposti a trattamenti con Testosterone hanno mostrato effetti notevoli sull’anabolismo proteico in presenza di GH, con il sito primario di interazione ormonale a livello epatico.[292] Quindi anche quando i livelli ormonali sono carenti, c’è ancora un’interazione molto importante tra il Testosterone e l’Asse GH/IGF-1.
Come accennato in precedenza, il GHR è espresso in quasi tutti i principali tipi di tessuto. Vale la pena sottolineare che il GHR è espresso in quantità molto bassa nel muscolo scheletrico – solo circa il 4-33% dei livelli osservati in altri tessuti. D’altra parte, il recettore del IGF-1 è espresso in maniera molto più elevata nei muscoli scheletrici, proprio come nel tessuto epatico.[293-294] Detto questo, l’aumento della sensibilità del GHR in un ambiente con livelli sovra fisiologici di GH andrà certamente a beneficio del BodyBuilder. I BodyBuilder cercano continuamente di utilizzare quantità elevate di rHGH nel tentativo di massimizzare i processi ipertrofici, e se il GH vede potenziata la sua azione e, quindi, il suo stimolo diretto sulla sintesi e rilascio del IGF-1, il quale opera direttamente sul tessuto muscolare, venendosi a creare quindi una vantaggiosa alterazione dell’Asse GH/IGF-1, l’effetto ipertrofico potenziato sarà indubbio.
IGFBP-4
Gli Androgeni hanno mostrato di avere la capacità di aumentare l’espressione del mRNA del IGF-1 locale nel muscolo scheletrico. Possiamo pertanto ipotizzare che gli Androgeni, in particolare a dosi più elevate, creino un ambiente all’interno del muscolo scheletrico estremamente favorevole alla gestione di livelli più alti di IGF-1 seguenti alla somministrazione di dosi sovrafisioogiche di rHGH. Ci sono persino stati studi sull’uomo che hanno mostrato livelli ridotti di IGFBP-4 locale nei campioni di tessuto muscolare, oltre a maggiori livelli di mRNA del IGF-1. Ciò potrebbe indicare che i cambiamenti hanno avuto luogo in quei muscoli per liberare più IGF-1 locale affinché esso possa legarsi ai suoi recettori.[277,295] Non ho molto approfondito la questione delle proteine leganti l’IGF, ma l’IGFBP-4 inibisce l’azione dell’IGF-1 e, quindi, minore è il livello della proteina legante, maggiori saranno i livelli di IGF-1 libero e attivo.[149,296]
È stato anche dimostrato che il Testosterone promuove l’ipertrofia negli stati deficitari di GH/IGF-1. [297-298] Questo è interessante in quanto dimostra che il Testosterone possiede sia vie metaboliche IGF-indipendenti che IGF-indipendenti nel tessuto muscolare.[299] Inoltre, i modelli cellulari hanno dimostrato che il Testosterone può sovraregolare l’espressione delle varie isoforme di IGF nei muscoli scheletrici, anche in assenza di GH/IGF-1.[298] E, anche se questo è stato dimostrato nei fibroblasti, il Testosterone ha mostrato di aumentare l’espressione del IGFBP-3 – un effetto che è stato ulteriormente migliorato dalla somministrazione di IGF-1.[300] È abbastanza chiaro, comunque, che il Testosterone eserciti effetti sinergici e additivi sull’anabolismo mediato da GH/IGF-1.
Fino ad ora mi sono concentrato sul Testosterone, tuttavia, però, sono stati osservati comportamenti leggermente diversi in relazione alle varianti androgene e al loro impatto sull’espressione sistemica e locale del IGF-1. Volevo analizzare un paio di specifici composti che sono spesso presenti nei protocolli farmacologici: sto parlando del Trenbolone e del Nandrolone. Salvo diverse indicazione, i risultati in seguito esposti provengono da studi svolti su animali.
Nandrolone
La somministrazione di Nandrolone ha costantemente dimostrato di non causare cambiamenti nei livelli di IGF-1 endocrino, nonostante produca simultaneamente un’espressione del IGF-1 muscolare significativamente più alta e un aumento della CSA delle fibre muscolari.[286,301-302] Inoltre, i livelli di IGFBP-3 locali sono stati riportati come significativamente più alti mentre i livelli di IGFBP-4 sono stati significativamente soppressi, il che ci rimanda a quanto esposto in precedenza – cioè ad un fattore che determina un aumento dei livelli di IGF-1 locale libero. In tutti gli studi, la somministrazione di Nandrolone ha portato direttamente ad un aumento dell’ipertrofia, nonostante non abbia avuto alcun impatto sui livelli di IGF-1 sistemici. Ciò rafforza ulteriormente l’ipotesi secondo cui l’IGF-1 endocrino non sia un fattore primario nell’ipertrofia dei muscoli scheletrici e quindi i livelli elevati non sono un prerequisito per l’aumento della massa muscolare.[303-305]
Trenbolone
È stato inoltre universalmente dimostrato che il Trenbolone aumenta i tassi di crescita muscolare in tutte le varie specie nelle quali è stato testato. A differenza del Testosterone e del Nandrolone, non si converte in estrogeno ed è stato suggerito fin dagli anni ’70 che l’aggiunta di Estradiolo al Trenbolone sembra migliorare gli effetti anabolici del composto.[306-307] Ci sono stati anche effetti potenziati sull’ipertrofia quando il Trenbolone è stato somministrato insieme ad un fattore di rilascio dell’Ormone della Crescita (GHRF).[308] Come accennato in precedenza, l’aumento di GH/IGF-1 necessita di livelli adeguati di estrogeni (derivanti principalmente dall’aromatizzazione) affinché questi esercitino uno stimolo dell’asse GH/IGF-1. Poiché la somministrazione di Trenbolone diminuisce intrinsecamente i livelli di Estradiolo, mediante inibizione a feedback negativo del Testosterone attraverso l’Asse Ipotalamo-Ipofisi-Gonadi (HPG) [309], la somministrazione di Estradiolo, tecnicamente, dovrebbe migliorare la funzionalità dell’asse GH/IGF-1. Questo dovrebbe quindi favorire ulteriormente la sinergia anabolica che avrebbe con l’androgeno. Questa ipotesi è in linea con ciò che vari studi hanno dimostrato nel corso degli anni.
Nelle colture cellulari, è stato anche dimostrato che gli Estrogeni alterano direttamente i tassi di MPS e MPB del Trenbolone attraverso meccanismi che coinvolgono sia il recettore estrogenico che il recettore del IGF-1.[310-311] In effetti, in linea di massima, i trattamenti con solo Trenbolone non mostrano aumenti significativi dei livelli endocrini o autocrini del IGF-1. Tuttavia, la co-somministrazione con Estradiolo ha tradizionalmente mostrato aumenti nei livelli di IGF-1 autocrini, similmente a quanto osservato con il Testosterone.[312-314] Questa è solo un’ulteriore prova che suggerisce che l’Estrogeno, sia di origine sistemica che derivata dall’aromatasi, è un componente chiave sia della massima stimolazione dell’Asse GH/IGF-1 che delle capacità anaboliche massimali degli Androgeni.
Proprio come il suo “cugino” 19-nor, il Trenbolone ha anche mostrato una maggiore espressione del fattore di crescita nei tessuti muscolo scheletrici, oltre ad una maggiore reattività dei muscoli a tali fattori di crescita.[315] Il Trenbolone ha anche mostrato di operare un aumento dell’attivazione e della proliferazione delle cellule satelliti in varie specie, in misura simile al Testosterone.[316-317] Ora, essendo a conoscenza delle interazioni legate al GH, si può facilmente giungere alla conclusione che entrambi questi effetti sarebbero vantaggiosi in un protocollo che includesse entrambi i composti.
PIIINP
Tornando al Testosterone, sia il GH che questo Androgeno aumentano i marker di sintesi del collagene come il PIIINP. Inoltre, il Testosterone ha anche dimostrato di potenziare le capacità del GH di aumentare la sintesi di collagene sia nei muscoli che nei tendini.[318] A sostegno di ciò, la somministrazione concomitante di GH e Testosterone in soggetti umani allenati (a livello amatoriale) ha causato aumenti significativi sia dei marker del IGF-1 che del collagene.[319] E’ interessante notare come alcuni di questi stessi marker del collagene dei quali si sta discutendo sono indicatori esatti che vengono esaminati come parte dei test antidoping per la rilevazione del GH.[320-321]
In precedenza ho solo brevemente toccato il percorso JAK-STAT ma, arrivati a questo punto, è necessario trattare l’argomento in maniera leggermente più approfondita. La via JAK-STAT è un componente critico del GH e riguarda sia la trascrizione del gene del IGF-1 che la crescita postnatale. Una delle proteine STAT in particolare, la STAT5, sembra essere intimamente coinvolta anche nella regolazione del muscolo scheletrico.[322] Ci sono due sottoproteine nella famiglia STAT5 e sono indicate come STAT5a e STAT5b. Sebbene siano identiche al 96%, è la variante STAT5b che è abbondante nei muscoli e nel tessuto epatico ed è quindi la proteina specifica su cui mi concentrerò da questo punto in avanti.[323-324]
Una revisione completa del percorso di segnalazione renderebbe questo articolo estremamente prolisso, ma ritengo importante che si comprenda che il percorso JAK/STAT5b è stato ripetutamente dimostrato avere una relazione diretta con l’espressione dell’IGF-1 locale nel tessuto muscolare e sul’ipertrofia.[248,325-332] Per questo motivo, se ci fossero modi per migliorare o ottimizzare questo specifico percorso, sembrerebbe che ciò si possa tradurre non solo in un aumento dell’attivazione del gene del IGF-1 [333-335] ma anche in un maggiore potenziale ipertrofico.
Fortunatamente, alcuni nuovi studi svolti su animali hanno già mostrato come i percorsi AR e JAK-STAT siano intimamente correlati.[248,336] Per essere precisi, il percorso STAT5a/b è a monte e l’AR è un bersaglio diretto a valle tramite la regolazione dell’espressione genica del AR. Studi su esseri umani hanno anche dimostrato che questo si verifica anche nell’uomo, con l’attività della STAT5 correlata positivamente all’espressione AR nelle linee cellulari del cancro alla prostata. [337]
Nella prossima parte di questa serie di articoli, parlerò dei modi in cui è possibile tentare di garantire che la via JAK-STAT5b-AR sia massimamente sensibilizzata, assicurando così che il potenziale ipertrofico sia potenziato quando Androgeni e GH vengono co-somministrati. Inoltre tratterò le applicazioni pratiche dell’uso del GH a fini ipertrofici esponendo infine le mie conclusioni in merito.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
Yarasheski KE, Campbell JA, Smith K, Rennie MJ, Holloszy JO, Bier DM. Effect of growth hormone and resistance exercise on muscle growth in young men. Am J Physiol. 1992 Mar;262(3 Pt 1):E261-7.
Yarasheski KE, Zachwieja JJ, Campbell JA, Bier DM. Effect of growth hormone and resistance exercise on muscle growth and strength in older men. Am J Physiol. 1995 Feb;268(2 Pt 1):E268-76.
Fryburg DA. Insulin-like growth factor I exerts growth hormone- and insulin-like actions on human muscle protein metabolism. Am J Physiol. 1994 Aug;267(2 Pt 1):E331-6.
Ohlsson C, Mohan S, Sjögren K, Tivesten A, Isgaard J, Isaksson O, Jansson JO, Svensson J. The role of liver-derived insulin-like growth factor-I. Endocr Rev. 2009 Aug;30(5):494-535.
Wakelam MJ. The fusion of myoblasts. Biochem J. 1985 May 15;228(1):1-12. Review.
Crist DM, Peake GT, Egan PA, Waters DL. Body composition response to exogenous GH during training in highly conditioned adults. J Appl Physiol (1985). 1988 Aug;65(2):579-84.
Deyssig R, Frisch H, Blum WF, Waldhör T. Effect of growth hormone treatment on hormonal parameters, body composition and strength in athletes. Acta Endocrinol (Copenh). 1993 Apr;128(4):313-8.
Yarasheski KE, Zachweija JJ, Angelopoulos TJ, Bier DM. Short-term growth hormone treatment does not increase muscle protein synthesis in experienced weight lifters. J Appl Physiol (1985). 1993 Jun;74(6):3073-6.
Lange KH, Andersen JL, Beyer N, Isaksson F, Larsson B, Rasmussen MH, Juul A, Bülow J, Kjaer M. GH administration changes myosin heavy chain isoforms in skeletal muscle but does not augment muscle strength or hypertrophy, either alone or combined with resistance exercise training in healthy elderly men. J Clin Endocrinol Metab. 2002 Feb;87(2):513-23.
Ehrnborg C, Ellegård L, Bosaeus I, Bengtsson BA, Rosén T. Supraphysiological growth hormone: less fat, more extracellular fluid but uncertain effects on muscles in healthy, active young adults. Clin Endocrinol (Oxf). 2005 Apr;62(4):449-57.
Rudman D, Feller AG, Nagraj HS, Gergans GA, Lalitha PY, Goldberg AF, Schlenker RA, Cohn L, Rudman IW, Mattson DE. Effects of human growth hormone in men over 60 years old. N Engl J Med. 1990 Jul 5;323(1):1-6.
Taaffe DR, Pruitt L, Reim J, Hintz RL, Butterfield G, Hoffman AR, Marcus R. Effect of recombinant human growth hormone on the muscle strength response to resistance exercise in elderly men. J Clin Endocrinol Metab. 1994 Nov;79(5):1361-6.
Taaffe DR, Jin IH, Vu TH, Hoffman AR, Marcus R. Lack of effect of recombinant human growth hormone (GH) on muscle morphology and GH-insulin-like growth factor expression in resistance-trained elderly men. J Clin Endocrinol Metab. 1996 Jan;81(1):421-5.
Hennessey JV, Chromiak JA, DellaVentura S, Reinert SE, Puhl J, Kiel DP, Rosen CJ, Vandenburgh H, MacLean DB. Growth hormone administration and exercise effects on muscle fiber type and diameter in moderately frail older people. J Am Geriatr Soc. 2001 Jul;49(7):852-8.
West DW, Kujbida GW, Moore DR, Atherton P, Burd NA, Padzik JP, De Lisio M, Tang JE, Parise G, Rennie MJ, Baker SK, Phillips SM. Resistance exercise-induced increases in putative anabolic hormones do not enhance muscle protein synthesis or intracellular signalling in young men. J Physiol. 2009 Nov 1;587(Pt 21):5239-47.
West DW, Burd NA, Tang JE, Moore DR, Staples AW, Holwerda AM, Baker SK, Phillips SM. Elevations in ostensibly anabolic hormones with resistance exercise enhance neither training-induced muscle hypertrophy nor strength of the elbow flexors. J Appl Physiol (1985). 2010 Jan;108(1):60-7.
Fryburg DA, Jahn LA, Hill SA, Oliveras DM, Barrett EJ. Insulin and insulin-like growth factor-I enhance human skeletal muscle protein anabolism during hyperaminoacidemia by different mechanisms. J Clin Invest. 1995 Oct;96(4):1722-9
Butterfield GE, Thompson J, Rennie MJ, Marcus R, Hintz RL, Hoffman AR. Effect of rhGH and rhIGF-I treatment on protein utilization in elderly women. Am J Physiol. 1997 Jan;272(1 Pt 1):E94-9.
Friedlander AL, Butterfield GE, Moynihan S, Grillo J, Pollack M, Holloway L, Friedman L, Yesavage J, Matthias D, Lee S, Marcus R, Hoffman AR. One year of insulin-like growth factor I treatment does not affect bone density, body composition, or psychological measures in postmenopausal women. J Clin Endocrinol Metab. 2001 Apr;86(4):1496-503.
Consitt LA, Saneda A, Saxena G, List EO, Kopchick JJ. Mice overexpressing growth hormone exhibit increased skeletal muscle myostatin and MuRF1 with attenuation of muscle mass. Skelet Muscle. 2017 Sep 4;7(1):17.
Wolf E, Wanke R, Schenck E, Hermanns W, Brem G. Effects of growth hormone overproduction on grip strength of transgenic mice. Eur J Endocrinol. 1995 Dec;133(6):735-40.
Ho KY, Weissberger AJ. The antinatriuretic action of biosynthetic human growth hormone in man involves activation of the renin-angiotensin system. Metabolism. 1990 Feb;39(2):133-7.
Blazer-Yost BL, Cox M. Insulin-like growth factor 1 stimulates renal epithelial Na+ transport. Am J Physiol. 1988 Sep;255(3 Pt 1):C413-7.
Giordano M, DeFronzo RA. Acute effect of human recombinant insulin-like growth factor I on renal function in humans. Nephron. 1995;71(1):10-5.
Ehrnborg C, Lange KH, Dall R, Christiansen JS, Lundberg PA, Baxter RC, Boroujerdi MA, Bengtsson BA, Healey ML, Pentecost C, Longobardi S, Napoli R, Rosén T; GH-2000 Study Group. The growth hormone/insulin-like growth factor-I axis hormones and bone markers in elite athletes in response to a maximum exercise test. J Clin Endocrinol Metab. 2003 Jan;88(1):394-401.
Doessing S, Heinemeier KM, Holm L, Mackey AL, Schjerling P, Rennie M, Smith K, Reitelseder S, Kappelgaard AM, Rasmussen MH, Flyvbjerg A, Kjaer M. Growth hormone stimulates the collagen synthesis in human tendon and skeletal muscle without affecting myofibrillar protein synthesis. J Physiol. 2010 Jan 15;588(Pt 2):341-51.
Boesen AP, Dideriksen K, Couppé C, Magnusson SP, Schjerling P, Boesen M, Kjaer M, Langberg H. Tendon and skeletal muscle matrix gene expression and functional responses to immobilisation and rehabilitation in young males: effect of growth hormone administration. J Physiol. 2013 Dec 1;591(23):6039-52.
Cohn L, Feller AG, Draper MW, Rudman IW, Rudman D. Carpal tunnel syndrome and gynaecomastia during growth hormone treatment of elderly men with low circulating IGF-I concentrations. Clin Endocrinol (Oxf). 1993 Oct;39(4):417-25.
Sullivan DH, Carter WJ, Warr WR, Williams LH. Side effects resulting from the use of growth hormone and insulin-like growth factor-I as combined therapy to frail elderly patients. J Gerontol A Biol Sci Med Sci. 1998 May;53(3):M183-7.
Dickerman RD, Douglas JA, East JW. Bilateral median neuropathy and growth hormone use: a case report. Arch Phys Med Rehabil. 2000 Dec;81(12):1594-5.
Papadakis MA, Grady D, Black D, Tierney MJ, Gooding GA, Schambelan M, Grunfeld C. Growth hormone replacement in healthy older men improves body composition but not functional ability. Ann Intern Med. 1996 Apr 15;124(8):708-16.
Zachwieja JJ, Yarasheski KE. Does growth hormone therapy in conjunction with resistance exercise increase muscle force production and muscle mass in men and women aged 60 years or older? Phys Ther. 1999 Jan;79(1):76-82. Review.
Kishioka Y, Thomas M, Wakamatsu J, Hattori A, Sharma M, Kambadur R, Nishimura T. Decorin enhances the proliferation and differentiation of myogenic cells through suppressing myostatin activity. J Cell Physiol. 2008 Jun;215(3):856-67.
Kanzleiter T, Rath M, Görgens SW, Jensen J, Tangen DS, Kolnes AJ, Kolnes KJ, Lee S, Eckel J, Schürmann A, Eckardt K. The myokine decorin is regulated by contraction and involved in muscle hypertrophy. Biochem Biophys Res Commun. 2014 Jul 25;450(2):1089-94.
Zhang CZ, Li H, Bartold PM, Young WG, Waters MJ. Effect of growth hormone on the distribution of decorin and biglycan during odontogenesis in the rat incisor. J Dent Res. 1995 Oct;74(10):1636-43.
Bahl N, Stone G, McLean M, Ho KKY, Birzniece V. Decorin, a growth hormone regulated protein in humans. Eur J Endocrinol. 2017 Nov 14. pii: EJE-17-0844.
Short KR, Moller N, Bigelow ML, Coenen-Schimke J, Nair KS. Enhancement of muscle mitochondrial function by growth hormone. J Clin Endocrinol Metab. 2008 Feb;93(2):597-604. Epub 2007 Nov 13.
Lange KH, Isaksson F, Juul A, Rasmussen MH, Bülow J, Kjaer M. Growth hormone enhances effects of endurance training on oxidative muscle metabolism in elderly women. Am J Physiol Endocrinol Metab. 2000 Nov;279(5):E989-96.
Chargé SB, Rudnicki MA. Cellular and molecular regulation of muscle regeneration. Physiol Rev. 2004 Jan;84(1):209-38. Review.
Halevy O, Hodik V, Mett A. The effects of growth hormone on avian skeletal muscle satellite cell proliferation and differentiation. Gen Comp Endocrinol. 1996 Jan;101(1):43-52.
Kim H, Barton E, Muja N, Yakar S, Pennisi P, Leroith D. Intact insulin and insulin-like growth factor-I receptor signaling is required for growth hormone effects on skeletal muscle growth and function in vivo. Endocrinology. 2005 Apr;146(4):1772-9. Epub 2004 Dec 23.
Sinha-Hikim I, Roth SM, Lee MI, Bhasin S. Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men. Am J Physiol Endocrinol Metab. 2003 Jul;285(1):E197-205. Epub 2003 Apr 1.
Ruzicka, L., Wettstein, A. and Kägi, H. (1935), Sexualhormone VIII. Darstellung von Testosteron unter Anwendung gemischter Ester. HCA, 18: 1478–1482.
Haupt HA, Rovere GD. Anabolic steroids: a review of the literature. Am J Sports Med. 1984 Nov-Dec;12(6):469-84. Review.
Shahidi NT. A review of the chemistry, biological action, and clinical applications of anabolic-androgenic steroids. Clin Ther. 2001 Sep;23(9):1355-90. Review.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7
Hartgens F, Kuipers H. Effects of androgenic-anabolic steroids in athletes. Sports Med. 2004;34(8):513-54. Review.
Kadi F, Eriksson A, Holmner S, Thornell LE. Effects of anabolic steroids on the muscle cells of strength-trained athletes. Med Sci Sports Exerc. 1999 Nov;31(11):1528-34.
Herbst KL, Bhasin S. Testosterone action on skeletal muscle. Curr Opin Clin Nutr Metab Care. 2004 May;7(3):271-7. Review.
Eriksson A, Kadi F, Malm C, Thornell LE. Skeletal muscle morphology in power-lifters with and without anabolic steroids. Histochem Cell Biol. 2005 Aug;124(2):167-75.
Kadi F. Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle. A basis for illegal performance enhancement. Br J Pharmacol. 2008 Jun;154(3):522-8. Epub 2008 Apr 14. Review.
Bhasin S, Woodhouse L, Casaburi R, Singh AB, Bhasin D, Berman N, Chen X, Yarasheski KE, Magliano L, Dzekov C, Dzekov J, Bross R, Phillips J, Sinha-Hikim I, Shen R, Storer TW. Testosterone dose-response relationships in healthy young men. Am J Physiol Endocrinol Metab. 2001 Dec;281(6):E1172-81.
Woodhouse LJ, Reisz-Porszasz S, Javanbakht M, Storer TW, Lee M, Zerounian H, Bhasin S. Development of models to predict anabolic response to testosterone administration in healthy young men. Am J Physiol Endocrinol Metab. 2003 May;284(5):E1009-17.
Bhasin S, Woodhouse L, Casaburi R, Singh AB, Mac RP, Lee M, Yarasheski KE, Sinha-Hikim I, Dzekov C, Dzekov J, Magliano L, Storer TW. Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle. J Clin Endocrinol Metab. 2005 Feb;90(2):678-88. Epub 2004 Nov 23.
Kvorning T, Andersen M, Brixen K, Madsen K. Suppression of endogenous testosterone production attenuates the response to strength training: a randomized, placebo-controlled, and blinded intervention study. Am J Physiol Endocrinol Metab. 2006 Dec;291(6):E1325-32. Epub 2006 Jul 25.
Sinha-Hikim I, Artaza J, Woodhouse L, Gonzalez-Cadavid N, Singh AB, Lee MI, Storer TW, Casaburi R, Shen R, Bhasin S. Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. Am J Physiol Endocrinol Metab. 2002 Jul;283(1):E154-64.
Bhasin S, Storer TW, Berman N, Callegari C, Clevenger B, Phillips J, Bunnell TJ, Tricker R, Shirazi A, Casaburi R. The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. N Engl J Med. 1996 Jul 4;335(1):1-7.
Sinha-Hikim I, Taylor WE, Gonzalez-Cadavid NF, Zheng W, Bhasin S. Androgen receptor in human skeletal muscle and cultured muscle satellite cells: up-regulation by androgen treatment. J Clin Endocrinol Metab. 2004 Oct;89(10):5245-55.
Klover P, Chen W, Zhu BM, Hennighausen L. Skeletal muscle growth and fiber composition in mice are regulated through the transcription factors STAT5a/b: linking growth hormone to the androgen receptor. FASEB J. 2009 Sep;23(9):3140-8.
Sheffield-Moore M, Urban RJ, Wolf SE, Jiang J, Catlin DH, Herndon DN, Wolfe RR, Ferrando AA. Short-term oxandrolone administration stimulates net muscle protein synthesis in young men. J Clin Endocrinol Metab. 1999 Aug;84(8):2705-11.
Kadi F, Bonnerud P, Eriksson A, Thornell LE. The expression of androgen receptors in human neck and limb muscles: effects of training and self-administration of androgenic-anabolic steroids. Histochem Cell Biol. 2000 Jan;113(1):25-9.
de Rooy C, Grossmann M, Zajac JD, Cheung AS. Targeting muscle signaling pathways to minimize adverse effects of androgen deprivation. Endocr Relat Cancer. 2016 Jan;23(1):R15-26.
Brodsky IG, Balagopal P, Nair KS. Effects of testosterone replacement on muscle mass and muscle protein synthesis in hypogonadal men–a clinical research center study. J Clin Endocrinol Metab. 1996 Oct;81(10):3469-75.
Ferrando AA, Sheffield-Moore M, Paddon-Jones D, Wolfe RR, Urban RJ. Differential anabolic effects of testosterone and amino acid feeding in older men. J Clin Endocrinol Metab. 2003 Jan;88(1):358-62.
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Singh R, Artaza JN, Taylor WE, Gonzalez-Cadavid NF, Bhasin S. Androgens stimulate myogenic differentiation and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinology. 2003 Nov;144(11):5081-8. Epub 2003 Jul 24.
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
Mulholland DJ, Dedhar S, Coetzee GA, Nelson CC. Interaction of nuclear receptors with the Wnt/beta-catenin/Tcf signaling axis: Wnt you like to know? Endocr Rev. 2005 Dec;26(7):898-915. Epub 2005 Aug 26. Review.
Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006 Nov 3;127(3):469-80. Review.
Singh R, Bhasin S, Braga M, Artaza JN, Pervin S, Taylor WE, Krishnan V, Sinha SK, Rajavashisth TB, Jasuja R. Regulation of myogenic differentiation by androgens: cross talk between androgen receptor/ beta-catenin and follistatin/transforming growth factor-beta signaling pathways. Endocrinology. 2009 Mar;150(3):1259-68.
Zhao JX, Hu J, Zhu MJ, Du M. Trenbolone enhances myogenic differentiation by enhancing β-catenin signaling in muscle-derived stem cells of cattle. Domest Anim Endocrinol. 2011 May;40(4):222-9.
Armstrong DD, Esser KA. Wnt/beta-catenin signaling activates growth-control genes during overload-induced skeletal muscle hypertrophy. Am J Physiol Cell Physiol. 2005 Oct;289(4):C853-9. Epub 2005 May 11.
Takada I, Kouzmenko AP, Kato S. Wnt and PPARgamma signaling in osteoblastogenesis and adipogenesis. Nat Rev Rheumatol. 2009 Aug;5(8):442-7.
Cossu G, Borello U. Wnt signaling and the activation of myogenesis in mammals. EMBO J. 1999 Dec 15;18(24):6867-72. Review.
Buckingham M. Skeletal muscle formation in vertebrates. Curr Opin Genet Dev. 2001 Aug;11(4):440-8. Review.
Polesskaya A, Seale P, Rudnicki MA. Wnt signaling induces the myogenic specification of resident CD45+ adult stem cells during muscle regeneration. Cell. 2003 Jun 27;113(7):841-52.
Tincello DG, Saunders PT, Hodgins MB, Simpson NB, Edwards CR, Hargreaves TB, Wu FC. Correlation of clinical, endocrine and molecular abnormalities with in vivo responses to high-dose testosterone in patients with partial androgen insensitivity syndrome. Clin Endocrinol (Oxf). 1997 Apr;46(4):497-506.
Foradori CD, Weiser MJ, Handa RJ. Non-genomic Actions of Androgens. Frontiers in neuroendocrinology. 2008;29(2):169-181.
Lucas-Herald AK, Alves-Lopes R, Montezano AC, Ahmed SF, Touyz RM. Genomic and non-genomic effects of androgens in the cardiovascular system: clinical implications. Clin Sci (Lond). 2017 Jul 1;131(13):1405-1418.
Münzer T, Harman SM, Hees P, Shapiro E, Christmas C, Bellantoni MF, Stevens TE, O’Connor KG, Pabst KM, St Clair C, Sorkin JD, Blackman MR. Effects of GH and/or sex steroid administration on abdominal subcutaneous and visceral fat in healthy aged women and men. J Clin Endocrinol Metab. 2001 Aug;86(8):3604-10.
Sattler FR, Castaneda-Sceppa C, Binder EF, Schroeder ET, Wang Y, Bhasin S, Kawakubo M, Stewart Y, Yarasheski KE, Ulloor J, Colletti P, Roubenoff R, Azen SP. Testosterone and growth hormone improve body composition and muscle performance in older men. J Clin Endocrinol Metab. 2009 Jun;94(6):1991-2001.
Illig R, Prader A. Effect of testosterone on growth hormone secretion in patients with anorchia and delayed puberty. J Clin Endocrinol Metab. 1970 May;30(5):615-8.
Pfeilschifter J, Scheidt-Nave C, Leidig-Bruckner G, Woitge HW, Blum WF, Wüster C, Haack D, Ziegler R. Relationship between circulating insulin-like growth factor components and sex hormones in a population-based sample of 50- to 80-year-old men and women. J Clin Endocrinol Metab. 1996 Jul;81(7):2534-40.
Erfurth EM, Hagmar LE, Sääf M, Hall K. Serum levels of insulin-like growth factor I and insulin-like growth factor-binding protein 1 correlate with serum free testosterone and sex hormone binding globulin levels in healthy young and middle-aged men. Clin Endocrinol (Oxf). 1996 Jun;44(6):659-64.
van Kesteren P, Lips P, Deville W, Popp-Snijders C, Asscheman H, Megens J, Gooren L. The effect of one-year cross-sex hormonal treatment on bone metabolism and serum insulin-like growth factor-1 in transsexuals. J Clin Endocrinol Metab. 1996 Jun;81(6):2227-32.
Veldhuis JD, Keenan DM, Mielke K, Miles JM, Bowers CY. Testosterone supplementation in healthy older men drives GH and IGF-I secretion without potentiating peptidyl secretagogue efficacy. Eur J Endocrinol. 2005 Oct;153(4):577-86.
Lewis MI, Fournier M, Storer TW, Bhasin S, Porszasz J, Ren SG, Da X, Casaburi R. Skeletal muscle adaptations to testosterone and resistance training in men with COPD. J Appl Physiol (1985). 2007 Oct;103(4):1299-310.
Mauras N, Hayes V, Welch S, Rini A, Helgeson K, Dokler M, Veldhuis JD, Urban RJ. Testosterone deficiency in young men: marked alterations in whole body protein kinetics, strength, and adiposity. J Clin Endocrinol Metab. 1998 Jun;83(6):1886-92.
Bondanelli M, Ambrosio MR, Margutti A, Franceschetti P, Zatelli MC, degli Uberti EC. Activation of the somatotropic axis by testosterone in adult men: evidence for a role of hypothalamic growth hormone-releasing hormone. Neuroendocrinology. 2003 Jun;77(6):380-7.
Veldhuis JD, Metzger DL, Martha PM Jr, Mauras N, Kerrigan JR, Keenan B, Rogol AD, Pincus SM. Estrogen and testosterone, but not a nonaromatizable androgen, direct network integration of the hypothalamo-somatotrope (growth hormone)-insulin-like growth factor I axis in the human: evidence from pubertal pathophysiology and sex-steroid hormone replacement. J Clin Endocrinol Metab. 1997 Oct;82(10):3414-20.
Weissberger AJ, Ho KK. Activation of the somatotropic axis by testosterone in adult males: evidence for the role of aromatization. J Clin Endocrinol Metab. 1993 Jun;76(6):1407-12.
Veldhuis JD, Mielke KL, Cosma M, Soares-Welch C, Paulo R, Miles JM, Bowers CY. Aromatase and 5alpha-reductase inhibition during an exogenous testosterone clamp unveils selective sex steroid modulation of somatostatin and growth hormone secretagogue actions in healthy older men. J Clin Endocrinol Metab. 2009 Mar;94(3):973-81.
Yamamoto T, Sakai C, Yamaki J, Takamori K, Yoshiji S, Kitawaki J, Fujii M, Yasuda J, Honjo H, Okada H. Estrogen biosynthesis in human liver–a comparison of aromatase activity for C-19 steroids in fetal liver, adult liver and hepatoma tissues of human subjects. Endocrinol Jpn. 1984 Jun;31(3):277-81.
Hata S, Miki Y, Saito R, Ishida K, Watanabe M, Sasano H. Aromatase in human liver and its diseases. Cancer Med. 2013 Jun;2(3):305-15.
Riggs BL, Hartmann LC. Selective estrogen-receptor modulators — mechanisms of action and application to clinical practice. N Engl J Med. 2003 Feb 13;348(7):618-29. Review. Erratum in: N Engl J Med. 2003 Mar 20;348(12):1192.
Löfgren L, Wallberg B, Wilking N, Fornander T, Rutqvist LE, Carlström K, von Schoultz B, von Schoultz E. Tamoxifen and megestrol acetate for postmenopausal breast cancer: diverging effects on liver proteins, androgens, and glucocorticoids. Med Oncol. 2004;21(4):309-18.
Centrella M, McCarthy TL, Chang WZ, Labaree DC, Hochberg RB. Estren (4-estren-3alpha,17beta-diol) is a prohormone that regulates both androgenic and estrogenic transcriptional effects through the androgen receptor. Mol Endocrinol. 2004 May;18(5):1120-30.
Yu YM, Domené HM, Sztein J, Counts DR, Cassorla F. Developmental changes and differential regulation by testosterone and estradiol of growth hormone receptor expression in the rabbit. Eur J Endocrinol. 1996 Nov;135(5):583-90.
Zung A, Phillip M, Chalew SA, Palese T, Kowarski AA, Zadik Z. Testosterone effect on growth and growth mediators of the GH-IGF-I axis in the liver and epiphyseal growth plate of juvenile rats. J Mol Endocrinol. 1999 Oct;23(2):209-21.
Hayes VY, Urban RJ, Jiang J, Marcell TJ, Helgeson K, Mauras N. Recombinant human growth hormone and recombinant human insulin-like growth factor I diminish the catabolic effects of hypogonadism in man: metabolic and molecular effects. J Clin Endocrinol Metab. 2001 May;86(5):2211-9.
Sculthorpe N, Solomon AM, Sinanan AC, Bouloux PM, Grace F, Lewis MP. Androgens affect myogenesis in vitro and increase local IGF-1 expression. Med Sci Sports Exerc. 2012 Apr;44(4):610-5.
Birzniece V, Meinhardt UJ, Umpleby MA, Handelsman DJ, Ho KK. Interaction between testosterone and growth hormone on whole-body protein anabolism occurs in the liver. J Clin Endocrinol Metab. 2011 Apr;96(4):1060-7.
Mertani HC, Delehaye-Zervas MC, Martini JF, Postel-Vinay MC, Morel G. Localization of growth hormone receptor messenger RNA in human tissues. Endocrine. 1995 Feb;3(2):135-42.
Florini JR, Ewton DZ, Coolican SA. Growth hormone and the insulin-like growth factor system in myogenesis. Endocr Rev. 1996 Oct;17(5):481-517. Review.
Urban RJ, Bodenburg YH, Gilkison C, Foxworth J, Coggan AR, Wolfe RR, Ferrando A. Testosterone administration to elderly men increases skeletal muscle strength and protein synthesis. Am J Physiol. 1995 Nov;269(5 Pt 1):E820-6.
Ewton DZ, Coolican SA, Mohan S, Chernausek SD, Florini JR. Modulation of insulin-like growth factor actions in L6A1 myoblasts by insulin-like growth factor binding protein (IGFBP)-4 and IGFBP-5: a dual role for IGFBP-5. J Cell Physiol. 1998 Oct;177(1):47-57.
Venken K, Movérare-Skrtic S, Kopchick JJ, Coschigano KT, Ohlsson C, Boonen S, Bouillon R, Vanderschueren D. Impact of androgens, growth hormone, and IGF-I on bone and muscle in male mice during puberty. J Bone Miner Res. 2007 Jan;22(1):72-82.
Serra C, Bhasin S, Tangherlini F, Barton ER, Ganno M, Zhang A, Shansky J, Vandenburgh HH, Travison TG, Jasuja R, Morris C. The role of GH and IGF-I in mediating anabolic effects of testosterone on androgen-responsive muscle. Endocrinology. 2011 Jan;152(1):193-206.
Spangenburg EE, Le Roith D, Ward CW, Bodine SC. A functional insulin-like growth factor receptor is not necessary for load-induced skeletal muscle hypertrophy. J Physiol. 2008 Jan 1;586(1):283-91.
Yoshizawa A, Clemmons DR. Testosterone and insulin-like growth factor (IGF) I interact in controlling IGF-binding protein production in androgen-responsive foreskin fibroblasts. J Clin Endocrinol Metab. 2000 Apr;85(4):1627-33.
Gayan-Ramirez G, Rollier H, Vanderhoydonc F, Verhoeven G, Gosselink R, Decramer M. Nandrolone decanoate does not enhance training effects but increases IGF-I mRNA in rat diaphragm. J Appl Physiol (1985). 2000 Jan;88(1):26-34.
Lewis MI, Horvitz GD, Clemmons DR, Fournier M. Role of IGF-I and IGF-binding proteins within diaphragm muscle in modulating the effects of nandrolone. Am J Physiol Endocrinol Metab. 2002 Feb;282(2):E483-90
Salmons S. Myotrophic effects of an anabolic steroid in rabbit limb muscles. Muscle Nerve. 1992 Jul;15(7):806-12.
Bisschop A, Gayan-Ramirez G, Rollier H, Dekhuijzen PN, Dom R, de Bock V, Decramer M. Effects of nandrolone decanoate on respiratory and peripheral muscles in male and female rats. J Appl Physiol (1985). 1997 Apr;82(4):1112-8
Lewis MI, Fournier M, Yeh AY, Micevych PE, Sieck GC. Alterations in diaphragm contractility after nandrolone administration: an analysis of potential mechanisms. J Appl Physiol (1985). 1999 Mar;86(3):985-92.
Heitzman RJ. The effectiveness of anabolic agents in increasing rate of growth in farm animals; report on experiments in cattle. Environ Qual Saf Suppl. 1976;(5):89-98. Review.
Buttery, P., Vernon, B., & Pearson, J. (1978). Anabolic agents—some thoughts on their mode of action. Proceedings of the Nutrition Society, 37(3), 311-315.
Hongerholt DD, Crooker BA, Wheaton JE, Carlson KM, Jorgenson DM. Effects of a growth hormone-releasing factor analogue and an estradiol-trenbolone acetate implant on somatotropin, insulin-like growth factor I, and metabolite profiles in growing Hereford steers. J Anim Sci. 1992 May;70(5):1439-48.
Tan RS, Scally MC. Anabolic steroid-induced hypogonadism–towards a unified hypothesis of anabolic steroid action. Med Hypotheses. 2009 Jun;72(6):723-8.
Kamanga-Sollo E, White ME, Hathaway MR, Weber WJ, Dayton WR. Effect of Estradiol-17beta on protein synthesis and degradation rates in fused bovine satellite cell cultures. Domest Anim Endocrinol. 2010 Jul;39(1):54-62.
Kamanga-Sollo E, Thornton KJ, White ME, Dayton WR. Role of G protein-coupled estrogen receptor-1, matrix metalloproteinases 2 and 9, and heparin binding epidermal growth factor-like growth factor in estradiol-17β-stimulated bovine satellite cell proliferation. Domest Anim Endocrinol. 2014 Oct;49:20-6.
Dunn JD, Johnson BJ, Kayser JP, Waylan AT, Sissom EK, Drouillard JS. Effects of flax supplementation and a combined trenbolone acetate and estradiol implant on circulating insulin-like growth factor-I and muscle insulin-like growth factor-I messenger RNA levels in beef cattle. J Anim Sci. 2003 Dec;81(12):3028-34.
Pampusch MS, Johnson BJ, White ME, Hathaway MR, Dunn JD, Waylan AT, Dayton WR. Time course of changes in growth factor mRNA levels in muscle of steroid-implanted and nonimplanted steers. J Anim Sci. 2003 Nov;81(11):2733-40.
Pampusch MS, White ME, Hathaway MR, Baxa TJ, Chung KY, Parr SL, Johnson BJ, Weber WJ, Dayton WR. Effects of implants of trenbolone acetate, estradiol, or both, on muscle insulin-like growth factor-I, insulin-like growth factor-I receptor, estrogen receptor-{alpha}, and androgen receptor messenger ribonucleic acid levels in feedlot steers. J Anim Sci. 2008 Dec;86(12):3418-23.
Thompson SH, Boxhorn LK, Kong WY, Allen RE. Trenbolone alters the responsiveness of skeletal muscle satellite cells to fibroblast growth factor and insulin-like growth factor I. Endocrinology. 1989 May;124(5):2110-7.
Johnson BJ, Halstead N, White ME, Hathaway MR, DiCostanzo A, Dayton WR. Activation state of muscle satellite cells isolated from steers implanted with a combined trenbolone acetate and estradiol implant. J Anim Sci. 1998 Nov;76(11):2779-86.
Dalbo VJ, Roberts MD, Mobley CB, Ballmann C, Kephart WC, Fox CD, Santucci VA, Conover CF, Beggs LA, Balaez A, Hoerr FJ, Yarrow JF, Borst SE, Beck DT. Testosterone and trenbolone enanthate increase mature myostatin protein expression despite increasing skeletal muscle hypertrophy and satellite cell number in rodent muscle. Andrologia. 2017 Apr;49(3).
Bhasin S, He EJ, Kawakubo M, Schroeder ET, Yarasheski K, Opiteck GJ, Reicin A, Chen F, Lam R, Tsou JA, Castaneda-Sceppa C, Binder EF, Azen SP, Sattler FR. N-terminal propeptide of type III procollagen as a biomarker of anabolic response to recombinant human GH and testosterone. J Clin Endocrinol Metab. 2009 Nov;94(11):4224-33.
Nelson AE, Meinhardt U, Hansen JL, Walker IH, Stone G, Howe CJ, Leung KC, Seibel MJ, Baxter RC, Handelsman DJ, Kazlauskas R, Ho KK. Pharmacodynamics of growth hormone abuse biomarkers and the influence of gender and testosterone: a randomized double-blind placebo-controlled study in young recreational athletes. J Clin Endocrinol Metab. 2008 Jun;93(6):2213-22.
Holt RI. Detecting growth hormone misuse in athletes. Indian J Endocrinol Metab. 2013 Oct;17(Suppl 1):S18-22.
Tan SH, Lee A, Pascovici D, Care N, Birzniece V, Ho K, Molloy MP, Khan A. Plasma biomarker proteins for detection of human growth hormone administration in athletes. Sci Rep. 2017 Aug 30;7(1):10039.
Jørgensen JO, Jessen N, Pedersen SB, Vestergaard E, Gormsen L, Lund SA, Billestrup N. GH receptor signaling in skeletal muscle and adipose tissue in human subjects following exposure to an intravenous GH bolus. Am J Physiol Endocrinol Metab. 2006 Nov;291(5):E899-905.
Liu X, Robinson GW, Gouilleux F, Groner B, Hennighausen L. Cloning and expression of Stat5 and an additional homologue (Stat5b) involved in prolactin signal transduction in mouse mammary tissue. Proc Natl Acad Sci U S A. 1995 Sep 12;92(19):8831-5.
Hennighausen L, Robinson GW. Interpretation of cytokine signaling through the transcription factors STAT5A and STAT5B. Genes Dev. 2008 Mar 15;22(6):711-21.
Eshet R, Laron Z, Pertzelan A, Arnon R, Dintzman M. Defect of human growth hormone receptors in the liver of two patients with Laron-type dwarfism. Isr J Med Sci. 1984 Jan;20(1):8-11.
Teglund S, McKay C, Schuetz E, van Deursen JM, Stravopodis D, Wang D, Brown M, Bodner S, Grosveld G, Ihle JN. Stat5a and Stat5b proteins have essential and nonessential, or redundant, roles in cytokine responses. Cell. 1998 May 29;93(5):841-50.
Hwa V, Little B, Adiyaman P, Kofoed EM, Pratt KL, Ocal G, Berberoglu M, Rosenfeld RG. Severe growth hormone insensitivity resulting from total absence of signal transducer and activator of transcription 5b. J Clin Endocrinol Metab. 2005 Jul;90(7):4260-6. Epub 2005 Apr 12.
Rowland JE, Lichanska AM, Kerr LM, White M, d’Aniello EM, Maher SL, Brown R, Teasdale RD, Noakes PG, Waters MJ. In vivo analysis of growth hormone receptor signaling domains and their associated transcripts. Mol Cell Biol. 2005 Jan;25(1):66-77. Erratum in: Mol Cell Biol. 2005 Mar;25(5):2072.
Klover P, Hennighausen L. Postnatal body growth is dependent on the transcription factors signal transducers and activators of transcription 5a/b in muscle: a role for autocrine/paracrine insulin-like growth factor I. Endocrinology. 2007 Apr;148(4):1489-97.
Barclay JL, Kerr LM, Arthur L, Rowland JE, Nelson CN, Ishikawa M, d’Aniello EM, White M, Noakes PG, Waters MJ. In vivo targeting of the growth hormone receptor (GHR) Box1 sequence demonstrates that the GHR does not signal exclusively through JAK2. Mol Endocrinol. 2010 Jan;24(1):204-17.
Hwa V, Nadeau K, Wit JM, Rosenfeld RG. STAT5b deficiency: lessons from STAT5b gene mutations. Best Pract Res Clin Endocrinol Metab. 2011 Feb;25(1):61-75.
Varco-Merth B, Feigerlová E, Shinde U, Rosenfeld RG, Hwa V, Rotwein P. Severe growth deficiency is associated with STAT5b mutations that disrupt protein folding and activity. Mol Endocrinol. 2013 Jan;27(1):150-61.
Davey HW, Xie T, McLachlan MJ, Wilkins RJ, Waxman DJ, Grattan DR. STAT5b is required for GH-induced liver IGF-I gene expression. Endocrinology. 2001 Sep;142(9):3836-41.
Woelfle J, Chia DJ, Rotwein P. Mechanisms of growth hormone (GH) action. Identification of conserved Stat5 binding sites that mediate GH-induced insulin-like growth factor-I gene activation. J Biol Chem. 2003 Dec 19;278(51):51261-6. Epub 2003 Oct 7.
Woelfle J, Billiard J, Rotwein P. Acute control of insulin-like growth factor-I gene transcription by growth hormone through Stat5b. J Biol Chem. 2003 Jun 20;278(25):22696-702. Epub 2003 Apr 7.
MacLean HE, Chiu WS, Notini AJ, Axell AM, Davey RA, McManus JF, Ma C, Plant DR, Lynch GS, Zajac JD. Impaired skeletal muscle development and function in male, but not female, genomic androgen receptor knockout mice. FASEB J. 2008 Aug;22(8):2676-89.
Tan SH, Dagvadorj A, Shen F, Gu L, Liao Z, Abdulghani J, Zhang Y, Gelmann EP, Zellweger T, Culig Z, Visakorpi T, Bubendorf L, Kirken RA, Karras J, Nevalainen MT. Transcription factor Stat5 synergizes with androgen receptor in prostate cancer cells. Cancer Res. 2008 Jan 1;68(1):236-48.
de Souza GL, Hallak J. Anabolic steroids and male infertility: a comprehensive review. BJU Int. 2011 Dec;108(11):1860-5.
Alcuni di voi potrebbero essersi già imbattuti nell’articolo di Chest Rockwell intitolato “The Science of Trenbolone” il quale si trova da qualche tempo su alcuni siti dedicati al BodyBuilding. L’intento dell’autore è stato quello di creare un articolo approfondito sul Trenbolone. Tale articolo è stato progettato per essere una guida di riferimento rapida volta a rispondere a molte domande riguardanti questo AAS. La mole di informazioni presenti in esso ha reso questo articolo un punto di riferimento per comprendere le caratteristiche e le potenziali applicazioni del Trenbolone. Il mio intento è quello di realizzare una “guida” sul Trenbolone, sul modello di quella realizzata da Chest Rockwell, in grado di fornire una serie di importanti informazioni riguardanti questo AAS e utilizzabili da atleti e preparatori.
II. Introduzione
Il Trenbolone è senza dubbio l’AAS con una reputazione quasi mitica all’interno del mondo del BodyBuilding. Poiché i dati sugli esseri umani sono molto limitati, spesso dobbiamo fare affidamento sugli aneddoti nel tentativo di formulare ipotesi su di esso. Come si può leggere in quasi tutti i forum dedicati al BodyBuilding, le esperienze con il Trenbolone variano ampiamente – con alcuni utenti che adorano assolutamente il composto mentre altri ne consigliano estrema cautela nell’uso o dicono agli altri utenti di evitarlo a tutti i costi. Nonostante questa grande divergenza di opinioni, non si può contestare la sua popolarità poiché numerosi studi hanno dimostrato negli anni che tale molecola è uno dei composti anabolizzanti più frequentemente usati, con una percentuale compresa tra il 20 e il 25% dei culturisti supplementari chimicamente che hanno riferito di averlo usato nell’arco degli ultimi dodici mesi. (1)(2)(3)
Il mio obiettivo con questo articolo sarà quello di utilizzare quante più informazioni disponibili per cercare di formulare alcune solide conclusioni riguardo al funzionamento e al potenziale utilizzo del composto. Allo stesso tempo spero di contribuire a dissipare alcuni miti che vengono ancora propagandati troppo spesso.
Come precedentemente accennato, esistono solo un paio di studi sull’uomo di cui sono a conoscenza, quindi la maggior parte del materiale citato proverrà da studi svolti su animali o in vitro. La domanda che dovrebbe essere posta è la seguente: possiamo prendere questi dati e applicarli concretamente sui BodyBuilder? Personalmente, ritengo che ci sono dei dati molto concreti e universalmente applicabili sugli esseri umani e poi ce ne sono altri che potrebbero richiedere un disclaimer. Cercherò di fare del mio meglio per indicarli nel corso di questo articolo.
III. Nozioni di base sul Trenbolone
Trenbolone
Il Trenbolone è considerabile quale Modulatore Selettivo del Recettore degli Androgeni (SARM), non progettato per l’uso umano (4), anche se venne commercializzato come farmaco da prescrizione per uso umano dalla Negma Laboratoires in Francia sotto il nome di Parabolan (Trenbolone Hexahydrobenzylcarbonato). Nonostante la sua primaria designazione, questo AAS continua ad essere pesantemente utilizzato dai Bodybuilder per promuovere la crescita muscolare, la riduzione del grasso e, di conseguenza, migliorare la composizione corporea.(5)(6) Quando sentiamo parlare di SARM colleghiamo tale termine ad una classe di agenti anabolizzanti non steroidei (vedi Ostarina, Andarina, LGD4033, ecc…). Però, tale definizione “particolaristica” non è del tutto corretta. Infatti, tutti gli AAS aventi un indice terapeutico (dato dalla Anabolico:Androgeno ratio) superiore a “1” sono considerabili quali SARM (l’indice terapeutico del Trenbolone è di circa 3,4). Si sa perfettamente che i SARM sono analoghi modificati degli ormoni sessuali maschili che normalmente esibiscono attività anabolica favorevole mentre, contemporaneamente, presentano una attività androgenica da moderata a minima in vivo rispetto agli androgeni endogeni.(7)(8)(9) Sono composti in fase di sviluppo da parte di molte aziende farmaceutiche nel tentativo di creare mezzi alternativi per il trattamento di condizioni come l’ipogonadismo e gli stati di deperimento muscolare e osseo. In sostanza, l’obiettivo è quello di ricreare gli aspetti positivi dati da dosi sovrafisiologiche di testosterone eliminando al contempo il rischio di eventi avversi che tendono a verificarsi quando si utilizzano tali dosaggi.(10) Questo obbiettivo lo si ritrova, appunto, concretizzato in tutti gli AAS che presentano modifiche dello scheletro carbossilico tali da permettere tale effetto.
La maggior parte dei SARM steroidei hanno come base di partenza una molecola di Testosterone. La struttura chimica della molecola di Testosterone viene quindi tradizionalmente modificata in uno dei seguenti tre modi (11)(12):
Esterificazione del gruppo 17β-idrossile che aumenta l’idrofobicità o la probabilità che la molecola possa essere respinta da una massa d’acqua;
Alchilazione in posizione 7α la quale riduce l’affinità di legame con l’enzima 5α-reduttasi;
Modifica strategica dei legami di carbonio in C1, C2, C9, C11 o C19 per ottenere una vasta gamma di effetti terapeutici.
Il Trenbolone è un C19-norsteroide (19-nor), derivato dal Nandrolone (Nortestosterone). La rimozione del gruppo metilico nella posizione C19 dello scheletro carbossilico steroideo riduce significativamente la suscettibilità dei 19-norsteroidi all’azione dell’enzima aromatasi e a quella dell’enzima 5α-reduttasi.(4) Entreremo più nel dettaglio dei meccanismi sopracitati in seguito ma, per ora, è necessario che si comprenda semplicemente come le sottili modifiche allo scheletro carbossilico della molecola di Testosterone possano tradursi direttamente in cambiamenti significativi del comportamento della nuova molecola derivata. Alcuni di questi cambiamenti possono includere l’affinità di legame del composto per il recettore degli androgeni e la sua affinità di legame con numerosi enzimi in grado di convertirlo in altri steroidi.(13)
Il Trenbolone differisce dal suo precursore (Nandrolone) per:
1- Il doppio legame inserito in C9– C10, che inibisce totalmente l’aromatizzazione e aumenta la resistenza al passaggio epatico;
2- l’insaturazione in C11-C12 che aumenta l’affinità per il recettore androgeno, rendendo il Trenbolone uno degli anabolizzante con la più forte affinità AR. (14)
Il valore Anabolico/Androgeno del Trenbolone rispetto al Testosterone (100/100) è pari a 625:185.
Quindi, il Trenbolone ha proprietà SARM-simili in quanto presenta una affinità significativamente minore ai percorsi metabolici ai quali è soggetto il Testosterone. Ma di questo ne parleremo in seguito.
IV. Storia del Trenbolone
Metribolone
L’enorme potenziale anabolizzante del Trenbolone, così come dei suoi analoghi, è stato riportato fin dagli anni ’60. Come molti di voi già sapranno, venne sintetizzata anche una versione orale denominata Metribolone (conosciuta anche come Methyltrienolone o Methyltrenbolone), tuttavia non è mai stata commercializzata come agente anabolizzante a causa della sua tossicità epatica estrema – causando colestasi intraepatica a quantità somministrate per via orale pari a 1 mg/giorno.(15)
A parte la parentesi francese del Parabolan (Trenbolone Hexahydrobenzylcarbonato), commercializzato dalla Negma fino al 1997, il Trenbolone non è mai stato approvato per l’uso umano e il suo utilizzo è stato (ed è) principalmente quello di agente per la promozione della crescita nel bestiame.(16)(17) Per tale scopo viene usato sia singolarmente che in combinazione con Estradiolo (E2).(18) L’uso di impianti per il bestiame contenenti la combinazione di Trenbolone ed Estradiolo è stata approvata dalla FDA nel 1992 (19), e ora circa il 90% dei bovini da carne negli Stati Uniti viene trattato con un mix di estrogeni, androgeni e/o progestinici volto a promuoverne la crescita.(20) Gli impianti per il bestiame sono un grande business con fino a 20 milioni di bovini all’anno impiantati con Trenbolone e un reddito annuo probabilmente superiore a un miliardo di dollari.(21)
Trenbolone Acetato
Nonostante l’approvazione da parte della FDA, sussistono ancora problemi di sicurezza poiché il Trenbolone Acetato (TBA), estere utilizzato per trattare il bestiame, ei suoi metaboliti sono stati identificati come potenziali sostanze chimiche dannose per il sistema endocrino (EDC). Gli EDC sono molecole esogene che possono imitare o inibire l’azione dei recettori ormonali come i recettori degli estrogeni, degli androgeni e degli ormoni tiroidei. Questi EDC possono anche alterare la sintesi, l’azione, il metabolismo e la secrezione di ormoni endogeni la quale può portare a problemi gravi come obesità, diabete e persino il cancro. (22)(23)
Meccanismi d’azione degli EDC. A: Interazione diretta del EDC con un recettore nucleare dell’ormone che porta alla stimolazione (agonismo) o inibizione (antagonismo) della sua attività trascrizionale. B: Stimolazione o inibizione della biosintesi degli ormoni endogeni. C: Stimolazione o inibizione della degradazione degli ormoni endogeni. D: Stimolazione o inibizione della proteina legante gli ormoni endogeni che porta ad un aumento o diminuzione della disponibilità degli ormoni circolanti.
A causa della potenziale gravità degli EDC, negli ultimi due decenni si è registrata una maggiore attenzione internazionale sull’esposizione ambientale e sugli effetti degli EDC nell’uomo e nella fauna selvatica.(24)(25) Come precedentemente accennato, il TBA ei suoi metaboliti sono stati identificati come EDC attraverso numerosi studi, possono essere diffusi in ambienti agricoli e sono associati a tossicità riproduttiva. (26)(27)(28) Il problema è che è necessaria soltanto un’esposizione a concentrazioni molto basse per causare potenziali problemi, come è stato dimostrato negli animali come i pesci, i quali hanno mostrato disordini nel comportamento sessuale e una diminuzione della fertilità. (29)
Sarà anche importante essere in grado di distinguere i vari tipi di impianti contenenti TBA, dal momento che molti degli studi che esamineremo in seguito utilizzeranno tipi diversi sugli animali presi in esame. Quello che segue è un elenco dei tipi di impianti comuni usati negli Stati Uniti, con specificazione delle loro concentrazioni ormonali:
Revalor-XS (200mg TBA / 40mg E2)
Revalor-200 (200mg TBA / 20mg E2)
Revalor-H (140mg TBA / 14mg E2)
Revalor-S (120mg TBA / 24mg E2)
Revalor-IS (80mg TBA / 16mg E2)
Revalor-IH (80mg TBA / 8mg E2)
Revalor-G (40mg TBA / 8mg E2)
Synovex PLUS (200mg TBA / 28mg E2)
Synovex-C (100mg Progesterone / 10mg E2)
Synovex-ONE Grass (150mg TBA / 15mg E2)
Synovex-S (200mg Progesterone / 20mg E2)
Synovex-H (200mg Testosterone / 20mg E2)
V. Metabolismo e fisiologia
Precedentemente ho fatto breve menzione ai metaboliti del TBA, quindi, ora, per completezza è giusto entrare nei dettagli. E’ corretto rammentare che la maggior parte delle informazioni riguardanti il metabolismo del Trenbolone in vivo provengono da osservazioni su bestiame e roditori (30)(31)(32). È fondamentale anche comprendere che vi sono differenze marcate nella quantità di vari metaboliti osservati nei modelli di ratti e mucche, i due mammiferi più intensamente studiati. (33) Torneremo a trattare questo argomento dopo aver esaminato alcuni dei principi fondamentali.
Trendione
Il nome chimico del TBA è 17ß-idrossi-estra-4,9,11-trien-3-one-17-acetato, talvolta abbreviato in 17β-TBOH-acetato. Dopo un’iniezione intramuscolare, viene rapidamente idrolizzato nel metabolita biologicamente attivo noto come 17β-idrossi-estra-4,9,11-trien-3-one o 17β-TBOH.(34) Da lì viene ulteriormente convertita in metaboliti inclusi i glucuronidi (per esempio Trendione / TBO) e altri cinque metaboliti idrossilati polari.(35) La serie di questi processi metabolici può essere riassunta come segue:
Il 17β-TBOH ha una maggiore affinità per gli AR rispetto a qualsiasi altro dei suoi metaboliti primari suggerendo che la biotrasformazione del Trenbolone riduce l’attività biologica dello steroide.(26)(27)(35) Per mettere questo in prospettiva, in uno studio l’elevata affinità del 17β-TBOH nei confronti del recettore degli androgeni umani e del recettore del progesterone bovino è stata ridotta dopo che la molecola è stata metabolizzata in 17α-TBOH e TBO a meno di 1/24 della dose originale del composto.(36) Questo comportamento è in netto contrasto con quello osservato nel Testosterone la cui conversione in DHT e in estrogeni porta a composti più potenti in relazione all’affinità del legame con il recettore bersaglio.(37)(38) Tuttavia, il comportamento del TBA è simile per natura ad altri composti 19-norsteroidei (come il Nandrolone), la cui affinità AR diminuisce una volta 5α-ridotta.(39)
Come accennato in precedenza, vi è una certa variazione nel metabolismo del 17-TBOH tra i mammiferi poiché i metaboliti primari sono il 17ß-idrossi-estra-4,9,11-trien-3-one e l’Estra-4,9,11-triene- 3,17-dione con, a loro volta, metaboliti 16α e 16ß-idrossilati nel ratto. Nella mucca questi metaboliti sono trascurabili e il 17α-TBOH è il prodotto principale insieme a piccole quantità di 16α e 16ß-idrossi-17α-TBOH (30)(31). Segue un grafico dettagliato che confronta le differenze tra gli animali:
Confronto dei metaboliti biliari del 17beta-trenbolone acetato nel ratto e nella mucca.
Fortunatamente per noi, esiste uno studio su esseri umani che ci può aiutare a chiarire come l’uomo metabolizzi il Trenbolone – almeno dopo che questo è stato ingerito per via orale. (35) Lo studio è stato progettato per indagare sugli effetti dell’ingestione di cibo contaminato e quindi il team di ricerca ha iniettato il 17β-TBOH in un pezzo di hamburger fritto da 5g, con una dose di 0,04 mg/kg di peso corporeo. Dopo un singolo consumo, il 63% della dose somministrata è stata escreta tramite l’urina entro 72 ore; a 24 ore il 50% della dose somministrata è stata osservata nei campioni di urina.
I risultati hanno anche rivelato che, negli esseri umani, il 17β-TBOH ingerito è principalmente escreto intatto come 17β-TBOH, come Epitrenbolone (17α-TBOH) o come Trendione (TBO) – con la maggior parte in forma 17α-TBOH. In questo senso, la biotrasformazione del 17ß-TBOH negli esseri umani assomiglia più da vicino a quella delle mucche che a quella dei roditori. Inoltre, nell’urina umana sono stati rilevati numerosi metaboliti polari di 17β-TBOH, sebbene in concentrazione molto inferiore rispetto a quei metaboliti precedentemente menzionati. (40)
Il 17β-TBOH presenta una bassa biodisponibilità orale dal momento che non presenta una metilazione in posizione 17α. I risultati di due saggi di Hershberger dimostrano che il Trenbolone è circa 80-100 volte meno efficace assunto per via orale rispetto a somministrazione tramite iniezione (comportamento legato appunto alla biodisponibilità).(26) Nonostante questo, il TBA e, ovviamente, 17β-TBOH hanno ancora dimostrato di alterare il sistema riproduttivo di umani, maiali, topi, ratti e altre specie di mammiferi a livelli di dosaggio relativamente bassi quando somministrati per via orale.
Nella prossima parte di questa serie di articoli, esploreremo l’impatto del Trenbolone su vari percorsi anabolici e marker della salute metabolica. Vedremo anche come il Trenbolone influenzi la produzione endogena di ormoni e inizieremo ad analizzare in maniera approfondita i suoi effetti su anabolismo e ipertrofia.
Stay tuned!
Gabriel Bellizzi
Riferimenti:
The Science of Trenbolone – By Chest Rockwell
Perry PJ, Lund BC, Deninger MJ, Kutscher EC, Schneider J. Anabolic steroid use in weightlifters and bodybuilders: an internet survey of drug utilization. Clin J Sport Med. 2005 Sep;15(5):326-30.
Parkinson AB, Evans NA. Anabolic androgenic steroids: a survey of 500 users. Med Sci Sports Exerc. 2006 Apr;38(4):644-51.
Ip EJ, Barnett MJ, Tenerowicz MJ, Perry PJ. The Anabolic 500 survey: characteristics of male users versus nonusers of anabolic-androgenic steroids for strength training. Pharmacotherapy. 2011 Aug;31(8):757-66.
Donner DG, Beck BR, Bulmer AC, Lam AK, Du Toit EF. Improvements in body composition, cardiometabolic risk factors and insulin sensitivity with trenbolone in normogonadic rats. Steroids. 2015 Feb;94:60-9.
Daniels JM, van Westerloo DJ, de Hon OM, Frissen PH. [Rhabdomyolysis in a bodybuilder using steroids]. Ned Tijdschr Geneeskd. 2006 May 13;150(19):1077-80. Dutch.
Geraci MJ, Cole M, Davis P. New onset diabetes associated with bovine growth hormone and testosterone abuse in a young body builder. Hum Exp Toxicol. 2011 Dec;30(12):2007-12.
Omwancha J, Brown TR. Selective androgen receptor modulators: in pursuit of tissue-selective androgens. Curr Opin Investig Drugs. 2006 Oct;7(10):873-81. Review.
Gao W, Dalton JT. Expanding the therapeutic use of androgens via selective androgen receptor modulators (SARMs). Drug Discov Today. 2007 Mar;12(5-6):241-8. Review.
Thevis M, Schänzer W. Synthetic anabolic agents: steroids and nonsteroidal selective androgen receptor modulators. Handb Exp Pharmacol. 2010;(195):99-126.
Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci. 2005 Nov;60(11):1451-7.
Kicman AT. Pharmacology of anabolic steroids. British Journal of Pharmacology. 2008;154(3):502-521.
Haendler B, Cleve A. Recent developments in antiandrogens and selective androgen receptor modulators. Mol Cell Endocrinol. 2012 Apr 16;352(1-2):79-91.
Fragkaki AG, Angelis YS, Koupparis M, Tsantili-Kakoulidou A, Kokotos G, Georgakopoulos C. Structural characteristics of anabolic androgenic steroids contributing to binding to the androgen receptor and to their anabolic and androgenic activities. Applied modifications in the steroidal structure. Steroids. 2009 Feb;74(2):172-97.
Krüskemper HL, Noell G. Liver toxicity of a new anabolic agent: methyltrienolone (17-alpha-methyl-4,9,11-estratriene-17 beta-ol-3-one). Steroids. 1966 Jul;8(1):13-24.
Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci. 1991 Jun;69(6):2452-62.
Chung KY, Johnson BJ. Application of cellular mechanisms to growth and development of food producing animals. J Anim Sci. 2008 Apr;86(14 Suppl):E226-35.Epub 2007 Oct 26. Review.
Metzler M, Pfeiffer E. Genotoxic potential of xenobiotic growth promoters and their metabolites. APMIS. 2001 Feb;109(2):89-95. Review.
Chung KY, Baxa TJ, Parr SL, Luqué LD, Johnson BJ. Administration of estradiol, trenbolone acetate, and trenbolone acetate/estradiol implants alters adipogenic and myogenic gene expression in bovine skeletal muscle. J Anim Sci. 2012 May;90(5):1421-7.
Balter M. Scientific cross-claims fly in continuing beef war. Science. 1999 May 28;284(5419):1453, 1455.
Lawrence JD, Ibarburu MA. Proceedings of the NCCC-134 Conference on Applied Commodity Price Analysis, Forecasting, and Market Risk Management; 16 and 17 April 2007; Chicago.
Diamanti-Kandarakis E, Bourguignon JP, Giudice LC, Hauser R, Prins GS, Soto AM, Zoeller RT, Gore AC. Endocrine-disrupting chemicals: an Endocrine Society scientific statement. Endocr Rev. 2009 Jun;30(4):293-342. Review.
Rachoń D. Endocrine disrupting chemicals (EDCs) and female cancer: Informing the patients. Rev Endocr Metab Disord. 2015 Dec;16(4):359-64.
Birnbaum LS. State of the science of endocrine disruptors. Environ Health Perspect. 2013 Apr;121(4):A107.
Baynes RE, Dedonder K, Kissell L, Mzyk D, Marmulak T, Smith G, Tell L, Gehring R, Davis J, Riviere JE. Health concerns and management of select veterinary drug residues. Food Chem Toxicol. 2016 Feb;88:112-22.
Wilson VS, Lambright C, Ostby J, Gray LE Jr. In vitro and in vivo effects of 17beta-trenbolone: a feedlot effluent contaminant. Toxicol Sci. 2002 Dec;70(2):202-11.
Durhan EJ, Lambright CS, Makynen EA, Lazorchak J, Hartig PC, Wilson VS, Gray LE, Ankley GT. Identification of metabolites of trenbolone acetate in androgenic runoff from a beef feedlot. Environ Health Perspect. 2006 Apr;114 Suppl 1:65-8.
Gall HE, Sassman SA, Lee LS, Jafvert CT. Hormone discharges from a midwest tile-drained agroecosystem receiving animal wastes. Environ Sci Technol. 2011 Oct 15;45(20):8755-64.
Jensen KM, Makynen EA, Kahl MD, Ankley GT. Effects of the feedlot contaminant 17alpha-trenbolone on reproductive endocrinology of the fathead minnow. Environ Sci Technol. 2006 May 1;40(9):3112-7.
Pottier J, Busigny M, Grandadam JA. Plasma kinetics, excretion in mild and tissue levels in the cow following implantation of trenbolone acetate. J Anim Sci. 1975 Sep;41(3):962-8.
Pottier J, Cousty C, Heitzman RJ, Reynolds IP. Differences in the biotransformation of a 17 beta-hydroxylated steroid, trenbolone acetate, in rat and cow. Xenobiotica. 1981 Jul;11(7):489-500.
Evrard P, Maghuin-Rogister G, Rico AG. Fate and residues of trenbolone acetate in edible tissues from sheep amd calves implanted with tritium-labeled trenbolone acetate. J Anim Sci. 1989 Jun;67(6):1489-96
Metzler M. Metabolism of some anabolic agents: toxicological and analytical aspects. J Chromatogr. 1989 Apr 7;489(1):11-21.
Dorts J, Richter CA, Wright-Osment MK, Ellersieck MR, Carter BJ, Tillitt DE. The genomic transcriptional response of female fathead minnows (Pimephales promelas) to an acute exposure to the androgen, 17beta-trenbolone. Aquat Toxicol.2009 Jan 18;91(1):44-53.
Spranger B, Metzler M. Disposition of 17 beta-trenbolone in humans. J Chromatogr. 1991 Apr 5;564(2):485-92.
Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. APMIS. 2000 Dec;108(12):838-46.
Wilson JD. The role of 5alpha-reduction in steroid hormone physiology. Reprod Fertil Dev. 2001;13(7-8):673-8. Review.
Santen RJ, Brodie H, Simpson ER, Siiteri PK, Brodie A. History of aromatase: saga of an important biological mediator and therapeutic target. Endocr Rev. 2009 Jun;30(4):343-75.
Sundaram K, Kumar N, Monder C, Bardin CW. Different patterns of metabolism determine the relative anabolic activity of 19-norandrogens. J Steroid Biochem Mol Biol. 1995 Jun;53(1-6):253-7.
Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids. 2010 Jun;75(6):377-89.
Androgenico: nessun dato disponibile Anabolico: nessun dato disponibile Standard: // Nome chimico: 18a-Homo-pregna-4,9,11-trien-17b-ol-3-one
Attività Estrogenica: nessuna Attività Progestinica: molto alta
Il Tetrahydrogestrinone (THG), detto “The Clear“, è uno steroide anabolizzante derivato dal Nandrolone. Questo agente è un “designer steroid”, il che significa che non è mai stato commercializzato come farmaco da prescrizione, ma è stato sviluppato specificamente per l’uso da parte degli atleti per il fatto che non era rintracciabile tramite test anti doping. Il THG è molto simile nella struttura all’agente anti-gonadotropico da prescrizione Gestrinone. (1) Il Gestrinone non è uno steroide anabolizzante attivo, ma ha una stretta relazione strutturale con il Trenbolone. L’aggiunta di quattro atomi di idrogeno (tetra-idro) al Gestrinone rompe il suo gruppo etinilico 17-aIpha (un tratto che interferisce notevolmente con il legame con il recettore androgeno) ad un etile, creando uno steroide capace di forte azione anabolica e androgena. Il THG è semplicemente Gestrinone 17-alfa-metilato, un “nuovo” derivato del potente AAS Trenbolone. L’AAS risultante è altamente anabolico, moderatamente androgeno, non estrogenico e altamente progestinico.
Differenze strutturali tra una molecola di Trenbolone, una di Gestrinone e una di Tetrahydrogestrinone.
Il Tetrahydrogestrinone è stato descritto per la prima volta nel 2004. (2) È stato portato alla luce in quello che è stato chiamato “il più grande fallimento organizzato per l’uso di steroidi nella storia degli sport competitivi”. Queste erano le parole di Terry Madden, Chief Executive Officer della u.s. Anti-Doping Agency (USADA), affrontando un gruppo di giornalisti e reporter televisivi in una conferenza stampa che ha reso noto lo scandalo doping internazionale. Il suo comunicato stampa allegato parlava di un allenatore olimpico anonimo, che girò una siringa contenente il “designer steroid” coinvolgendo Victor Conte della BALCO Labs in California, accreditato da molti coaching dei migliori atleti al mondo, come fonte per l’agente. La fonte anonima fu poi identificata come l’allenatore dell’atleta Trevor Graham. La USADA è stata presto messa in mezzo alla frenesia dei media, con molti grandi nomi danneggiati per essere risultati positivi all’uso del THG.
Patrick Arnold, creatore del THG.
Per un breve periodo di tempo, il THG fu un AAS ideale per gli atleti soggetti a test anti doping, in quanto non rilevabili con l’esame delle urine. La sua detenzione era tecnicamente legale negli Stati Uniti, essendo esso sconosciuto ai legislatori al momento della stesura del Anabolic Sterc Control Act. Ma con lo scandalo doping internazionale che avrebbe circondato la BALCO qualsiasi valore che il THG possedeva per gli atleti sottoposti a controlli scomparve. Questo AAS è stato aggiunto alla lista delle sostanze controllate nel gennaio 2005. Victor Conte trascorse diversi mesi in carcere per il suo ruolo nello scandalo BALCO, dovuto al fatto che, pur essendo un farmaco sconosciuto, il THG poteva essere posseduto legalmente, ma non era legale venderlo. Patrick Arnold, il chimico organico creatore del THG, né subì le conseguenze penali, e dovette scontare 3 mesi di carcere alla fine del 2006.
Il Tetrahydrogestrinone è stato un opportunità tra i “designer steroid”, creato da un composto facilmente disponibile con processi chimici intermedi non regolamentati e con un semplice metodo.
Nandrolone
Come già accennato, il Tetrahydrogestrinone è una forma modificata del Nandrolone. Esso differisce dal suo precursore di base per 1) l’aggiunta di un gruppo metile in posizione C-17 per proteggere l’ormone durante la somministrazione orale, 2) l’aggiunta di un doppio legame in C-9 e C-11, che aumentano notevolmente l’attività relativa dello steroide, e 3) la presenza di un gruppo 18a-homo, che conferisce allo steroide l’attività progestinica.
Il Tetrahydrogestrinone non è soggetto ad aromatizzazione, e non possiede attività estrogenica misurabile. La ginecomastia rimane comunque una preoccupazione durante la somministrazione, a causa della sua natura progestinica (vedi sotto), specialmente quando questo AAS viene combinato con altri farmaci soggetti ad aromatizzazione.
Allo stesso tempo la ritenzione idrica e l’accumulo di grasso con modello femminile possono diventare un problema durante l’uso del Tetrahydrogestrinone: alcuni, nel tentativo di eliminare tali effetti collaterali, inseriscono Nolvadex o antiprogestinici specifici; pratica poco consigliabile visti i numerosi effetti collaterali riscontrabili con l’uso degli antiprogestinici, anche sui livelli di Progesterone e sul numero dei suoi siti recettori (problema riscontrato anche con l’uso del Nolvadex).
Come già accennato, il Tetrahydrogestrinone è un progestinico estremamente attivo. (3) E’ stato dimostrato avere un’attività progestinica più forte del Nandrolone, del Trenbolone e del Gestrinone, un ormone steroideo di sintesi che ha attività sia progestinica che anti-progestinica (agonista parziale del recettore del progesterone (PR) o Modulatore Selettivo del Recettore del Progesterone (SPRM) ) e lievemente androgena. Il THG è risultato essere 7 volte più potente del progesterone stesso. Gli effetti collaterali associati con il Progesterone sono simili a quelli degli estrogeni, compresa il feedback negativo di inibizione della produzione di Testosterone e una maggiore velocità di accumulo di grasso. I progestinici aumentano anche l’effetto stimolante degli estrogeni sulla crescita del tessuto mammario. Sembra che ci sia una forte sinergia tra questi due ormoni, in modo tale che la ginecomastia potrebbe anche verificarsi con l’aiuto dei progestinici, senza eccessivi livelli di estrogeni. L’uso di un anti-estrogeno, che inibisce la componente estrogenica di questa alterazione, è spesso sufficiente per mitigare la ginecomastia causata da AAS con attività progestinica. Il rialzo della Prolattina è un altra possibile conseguenza. L’uso di anti-prolattinici andrebbe preso in considerazione solo quando, attraverso appositi esami ematici, si è appurata una iperprolattinemia. Molto spesso, il controllo adeguato degli estrogeni risulta più che sufficiente.
Il THG possiede una spiccata attività androgena. Effetti collaterali androgeni sono comuni con questa molecola, e possono includere pelle grassa, acne, e alopecia androgenetica. Le atlete di sesso femminile possono incorrere in effetti collaterali androgenici molto forti tra i quali troviamo approfondimento della voce, irregolarità mestruale, cambiamenti nella struttura della pelle, crescita di peli sul viso, e allargamento del clitoride.
Come già detto, il Tetrahydrogestrinone è un composto metilato in C-17. Questa alterazione protegge il farmaco dalla disattivazione epatica, consentendo ad una percentuale molto elevata della molecola di entrata nel flusso ematico dopo somministrazione orale. E come ben sappiamo, è la metilazione in C-17 che rende il THG un composto epatotossico. L’esposizione prolungata nel tempo o ad alte dosi può causare danni al fegato. In rari casi può svilupparsi disfunzione epatica. Si consiglia pertanto di effettuare regolari monitoraggi della funzione epatica e della salute generale durante l’uso di questo AAS. L’assunzione di AAS metilati in C-17 è comunemente limitata a 6-8 settimane, nel tentativo di evitare il più possibile un eccessivo stress epatico. L’uso di un integratore disintossicante epatico come il Liv-52 o l’Essentiale Forte è consigliato durante l’assunzione di AAS epatotossici.
Come risaputo, gli AAS possono avere effetti deleteri sul colesterolo sierico. Questo include una tendenza alla riduzione delle concentrazioni di colesterolo HDL (buono) e un aumento delle concentrazioni di colesterolo LDL (cattivo), cosa che comporta uno sbilanciamento dell’equilibrio HDL/LDL che si traduce in un rischio maggiore di sviluppare arteriosclerosi. L’impatto relativo all’assunzione di un AAS nei confronti dei lipidi ematici dipende dalla dose, dalla via di somministrazione (per via orale o iniettabile), dal tipo di steroide (aromatizzabile o non aromatizzabile), e dal livello di resistenza al metabolismo epatico. Il Tetrahydrogestrinone ha un forte effetto sulla gestione epatica del colesterolo a causa della sua resistenza strutturale alla disattivazione epatica, alla sua natura non aromatizzabile e alla sua tipica via di somministrazione (esiste anche la versione iniettabile). Gli AAS possono anche influenzare negativamente la pressione del sangue e i livelli dei trigliceridi, riducendo il rilassamento endoteliale, e promuovendo l’ipertrofia ventricolare sinistra, tutti fattori con un potenziale nel aumentare il rischio di malattie cardiovascolari e infarto del miocardio.
Per contribuire a ridurre lo sforzo cardiovascolare si consiglia di mantenere un programma di esercizio cardiovascolare attivo e di ridurre al minimo l’assunzione di grassi saturi, colesterolo e carboidrati semplici in ogni momento durante la somministrazione di AAS.
La supplementazione con oli di pesce (4 grammi al giorno) e un integratore alimentare di Niacina per il controllo del colesterolo è anche raccomandata.
Tutti gli AAS se assunti in dosi sufficienti per promuovere l’aumento della massa muscolare causano una soppressione del Testosterone endogeno. Senza l’intervento con sostanze Testosterone-stimolante, e una adeguata PCT, i livelli di Testosterone dovrebbero tornare alla normalità entro 1-4 mesi dalla cessione del farmaco. Si noti che un ipogonadismo ipogonadotropo prolungato può svilupparsi secondariamente all’abuso di steroidi, cosa che richiede un intervento medico.
Anche per il THG (quando assunto oralmente) se ne consiglia l’assunzione lontano dai pasti in quanto studi hanno dimostrato che l’assunzione di uno steroide anabolizzante orale con cibo può diminuirne la sua biodisponibilità.(4) Questo è causato dalla natura liposolubile degli ormoni steroidei, che può permettere ad una parte del farmaco di sciogliersi con i grassi alimentari non digeriti, riducendo l’assorbimento dal tratto gastrointestinale. Per la massima biodisponibilità , questo steroide deve essere assunto a stomaco vuoto.
Il THG non è mai stato approvato per l’uso negli esseri umani. Linee guida di prescrizione non sono disponibili. Per scopi dopanti in ambito maschile la dose giornaliera tipicamente efficace per via orale si aggira nel range dei 2-5mg al giorno. Possiamo anche notare che il THG privo della sua modificazione delta-11 (di-ene invece di tri-ene) ha mostrato essere oltre 14 volte più attivo del Methyltestosterone. (5) Anche se non esistono schemi che lo dimostrino, questo “designer steroid” dovrebbe essere molto più potente di qualsiasi AAS disponibile in commercio, e probabilmente sarebbe leggermente meno potente del Methyltrienolone. Per gli atleti che utilizzano il THG, i risultati si manifestano in un miglioramento misurabile nella forza, nella massa muscolare e delle prestazioni generali.
Il THG non è generalmente raccomandato per le atlete a causa della sua natura androgena molto forte e alla tendenza a produrre effetti collaterali virilizzanti. Questo composto potrebbe essere comunque utilizzato con successo in ambito femminile utilizzando dosi inferiori ad 1mg/die: quindi a dosaggi nell’ordine dei microgrammi.
L’emivita del THG è di circa 24-48 ore, con variabili a seconda del metodo di somministrazione.
Il THG non è generalmente disponibile sul mercato nero.
Gabriel Bellizzi
Riferimenti:
–William Llewellyn’s ANABOLICS, 9th ed.
1- Hormonal therapy of endometriosis. Metzger DA, Luciano AA. Obstet Gynecol Clin North Am. 1989 Mar;16(1 ):105-22.
2- Tetrahydrogestrinone: discovery, synthesis, and detection in urine. Catlin et al. Rapid Commun Mass Spectrom. 2004;18(12):1245-049.
3- Tetrahydrogestrinone Is a Potent Androgen and Progestin. Death A, McGrath K et al. J Clin Endocrinol and Metab. 89: 2498-2500, 2004.
4-Anabolic Steroids and Sports Volume II. James E. Wright. Sports Science Consultants, Natick, MA 1982.



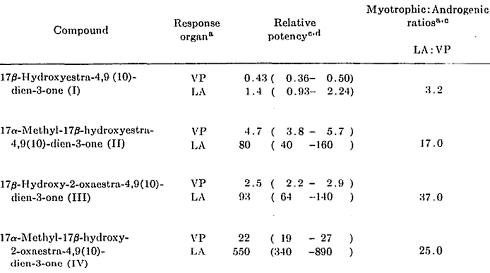


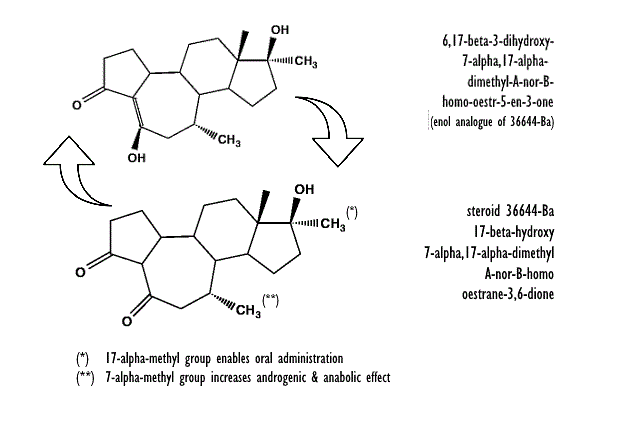 La prima formula strutturale mostrata di seguito è quella ufficiale, la seconda è il metabolita in cui il composto progenitore 36644-Ba si converte per poi ritornare alla forma di partenza.
La prima formula strutturale mostrata di seguito è quella ufficiale, la seconda è il metabolita in cui il composto progenitore 36644-Ba si converte per poi ritornare alla forma di partenza.



 Recentemente, è stato pubblicato uno studio sul “British Journal of Sports Medicine” da parte di ricercatori svedesi i quali hanno sottoposto delle donne a trattamento topico a base di 10mg di Testosterone al giorno. Tale dosaggio lungi dal poter far raggiungere la soglia ematica di Testosterone riscontrabile in un uomo, ma è stata sufficiente a portare un aumento della massa muscolare, un calo della massa grassa e il miglioramento della resistenza.(1)
Recentemente, è stato pubblicato uno studio sul “British Journal of Sports Medicine” da parte di ricercatori svedesi i quali hanno sottoposto delle donne a trattamento topico a base di 10mg di Testosterone al giorno. Tale dosaggio lungi dal poter far raggiungere la soglia ematica di Testosterone riscontrabile in un uomo, ma è stata sufficiente a portare un aumento della massa muscolare, un calo della massa grassa e il miglioramento della resistenza.(1)
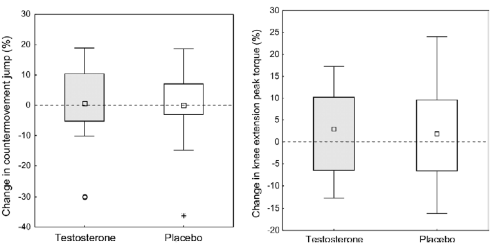
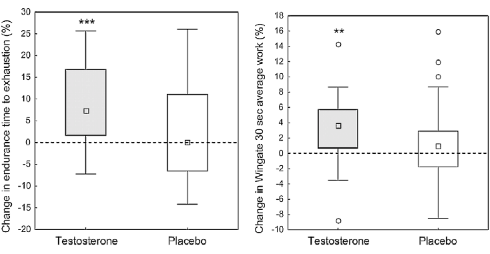
 Secondo uno studio su animali svolto da fisiologi rumeni presso l’Università di Medicina e Farmacia Carol Davila e pubblicato su “Medicina”, la supplementazione di Taurina potrebbe offrire protezione cardiovascolare agli utilizzatori di AAS.(1)
Secondo uno studio su animali svolto da fisiologi rumeni presso l’Università di Medicina e Farmacia Carol Davila e pubblicato su “Medicina”, la supplementazione di Taurina potrebbe offrire protezione cardiovascolare agli utilizzatori di AAS.(1)