Se non avete letto ancora la prima, la seconda, la terza, la quarta e la quinta parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte – 3° Parte– 4° Parte– 5° Parte.
Vanadio: caratteristiche e possibili applicazioni.
Vanadio Solfato
Il Vanadio è noto per essere un minerale “ultratraccia” nell’alimentazione umana. (1) Non fa parte delle classiche 24 vitamine e minerali, anche se è necessario per alcune reazioni nel corpo, come la formazione di un Pervanadato richiesto per la fosforilazione di alcuni recettori proteici. (2) Normalmente vengono consumati giornalmente dai 6 ai 20mcg di Vanadio, principalmente attraverso il consumo di peperoni e frutti di mare. (2) Altre fonti alimentare che presentano una concentrazione significativa di questo minerale sono i cereali integrali, la birra, la frutta secca e alcune qualità di funghi. Nei cereali, e in modo particolare nella farina e nel pane, le concentrazioni di Vanadio possono superare i 10mcg/kg.
Il Vanadio ha una storia di utilizzo nel trattamento del diabete, patologia nella quale la somministrazione orale di Vanadio di Sodio rappresentava l’intervento farmacologico di base prima della scoperta dell’Insulina avvenuta nel 1921. (3)
Il Vanadio (Vanadato) può anche formare complessi legandosi al Perossido di Idrogeno (H202) e formando il Perossivanadio (Pervanadato), biologicamente più attivo nell’attività inibitoria nei confronti delle Proteine Fosfotirosina-Fosfatasi (PTP) rispetto al Vanadato. (4) (5)
Il Vanadio, come Vanadato in vivo, inibisce la fosforilazione delle ATPasi di tipo P (6) (7) e delle Proteine Fosfotirosina-Fosfatasi (PTP). (8) La dose per inibire le ATPasi, tuttavia, è clinicamente significativa solo a livelli tossici. L’inibizione delle PTP aumenta i vari processi di fosforilazione della Tirosina (poiché le fosfatasi sono enzimi inibitori nella fosforilazione).
Vari effetti derivano dall’inibizione delle PTP, come l’inibizione dell’assorbimento amminoacidico cellulare del sistema di trasporto A degli AA neutri (5), l’aumento dell’assorbimento del glucosio e la mobilizzazione dei GLUT4 attraverso una via indipendente dal fosfoinositolo-3-chinasi (PI3K) [a differenza dell’Insulina]. (9) Il meccanismo attraverso il quale il Vanadio esplica i suoi effetti si trova nella sua azione insulino-mimetica e nell’aumento della fosforilazione generale che agisce sulla Tirosina Chinasi del recettore insulinico (la quale viene fosforilata sotto l’influenza dell’Insulina, oltre all’inibizione delle PTP).
Potenziali siti di azione del Vanadio nella cascata di segnalazione dell’Insulina. Il legame del recettore insulinico sulla superficie della cellula determina i cambiamenti conformazionali nel recettore, principale attivazione della via PI3-K, la via principale implicata nella mediazione degli effetti metabolici. I potenziali siti di azione del Vanadio in questo percorso sono riassunti in questa figura. V ()), V (+), denotano effetti negativi, positivi o nulli, rispettivamente. IRS, substrato del recettore dell’insulina; PI3-K, fosfatidilinositolo-3-chinasi; PDK, chinasi 3-fosfoinositide-dipendente; PKB, protein chinasi B; GSK- glicogeno sintasi chinasi-3; PFK-2, fosfofuctokinasi-2; GLUT 4, trasportatore di glucosio di tipo 4; 4E- 4 proteina legante; PTP1B, proteina tirosina fosfatasi 1B; PP-1, proteina fosfatasi-1.Sodio Ortovanadato
Composti contenenti Vanadio sono noti per inibire le PTP (10) (11) poiché essi tendono a formare una struttura bipirramidale trigonale che può agire come un inibitore competitivo del trasferimento di fosforile (come nel caso del Sodio Ortovanadato) o un ossidante del residuo di cisteina del ciclo PTP (complessi Perossivanadati). (12) Similmente ad altri composti inibitori delle PTP (gli inibitori PTP1B includono Berberina e Acido Ursolico) il Vanadio può prolungare la segnalazione attraverso il Recettore dell’Insulina prevenendo la regolazione negativa della degradazione dell’IRS-1, che è indotta a valle del segnale del recettore insulinico (tramite mTOR [13] S6K1 (14) ) e funziona tramite le PTP per sopprimere (15) e stimolare la degradazione (16) (17) delle IRS-1 come forma di feedback negativo.
IRS-1
L’Ortovanadato di Sodio a 1mmol/L sembra migliorare la segnalazione (miocita L6) di circa il 25% ed è associato all’inibizione del declino dell’attività complessa PI3K/IRS-1, ma è stato osservato che non inibisce la perdita di massa della proteina IRS-1. (18)
In uno studio nel quale sono stati presi in esame diabetici di tipo II e soggetti obesi ma ancora considerabili come sani (gruppo di controllo) è emerso che 100mg di Vanadio per 3 settimane sono stati in grado di sopprimere la gluconeogenesi epatica in entrambi i gruppi, ma solo il gruppo dei diabetici ha riscontrato un aumento della sensibilità all’Insulina mentre i soggetti del gruppo di controllo non hanno avuto cambiamenti significativi nei parametri della glicemia. (19)
Uno studio svolto su esseri umani (n = 14) di età compresa tra i 40 e i 50 anni con alterata tolleranza al glucosio, ha mostrato che la somministrazione di Vanadio ad un dosaggio di 50mg due volte al giorno (100mg al giorno) per un periodo di un mese (30 giorni), in risposta ad una condizione iperinsulinemica euglicemica, non ha prodotto miglioramenti della sensibilità all’Insulina e ha portato ad un piccolo ma significativo aumento dei Trigliceridi. (20) In questo studio non si sono osservate modifiche del LDL-C, HDL-C o del Colesterolo totale mentre il prima citato aumento dei Trigliceridi consisteva in un incremento delle concentrazioni ematiche da 1,4 +/- 0,6mmol/L a 1,7 +/- 0,5mmol/L.
Uno studio condotto su diabetici di tipo II (n = 6) ha osservato che dopo la somministrazione di Vanadio a 100mg al giorno durante 3-5 settimane di un periodo complessivo di 7 settimane (con placebo somministrato all’inizio e nell’ultimo fase dello studio) ha osservato che dopo 3 settimane di somministrazione di Vanadio si assisteva ad un aumento dell’assorbimento di glucosio e della sensibilità all’Insulina che era ancora presente due settimane dopo la cessata somministrazione del minerale; questi miglioramenti sono stati accompagnati da una diminuzione dell’HbA1c da 9,6 + 0,6% a 8,8 ± 0,6%. (21) L’utilizzo del Vanadio (come Solfato) ad un dosaggio di 150mg per 6 settimane è stato associato a una riduzione del 20% della glicemia a digiuno e ad una diminuzione del HbA1c da 8,1 ± 0,4 a 7,6 ± 0,4%. (22) Sebbene questo studio abbia rilevato una riduzione della produzione endogena di glucosio del 20% nei diabetici, la correlazione di questa riduzione alla diminuzione della glicemia a digiuno è stata di r = 0.6. (22)
Tuttavia, la qualità di questi studi è stata messa in discussione. E’ stata svolta una review sistemica la quale mirava a valutare tutti gli studi in doppio cieco controllati con placebo sul Vanadio e la ricerca preliminare terminò quando non ne trovarono nessuno. (23) Gli studi esistenti e quelli citati sopra tendono a non avere alcun gruppo placebo e sono limitati a campioni di dimensioni ridotte; i loro risultati significativi hanno perciò una potenza statistica molto inferiore rispetto ad altri composti.
Uno studio nel quale è stata utilizzata la Rapamicina (un inibitore dell’mTOR, utilizzato in questo studio per inibire la degradazione dell’IRS-1 che è un regolatore negativo della segnalazione dell’Insulina) ha osservato che la cosomministrazione di questa con l’Ortovanadato di Sodio (1mmol/L) risultava sinergica nell’aumento dell’assorbimento di glucosio in un miocita L6 in presenza di Insulina da 26,42 +/- 3,73% (osservata con il solo Vanadio) a 97,83 +/- 8,54% (in combinazione: la sola Rapamicina ha causato un assorbimento del 39,9 ± 3,39%). (18) Ciò era dovuto all’inibizione della degradazione dell’IRS-1 e al prolungamento della segnalazione attraverso il Recettore dell’Insulina, che promuoveva l’assorbimento del glucosio indotto dal PI3K, ed era probabilmente legato al fatto che il Vanadio poteva inibire la degradazione dell’associazione PI3K / IRS-1 (Rapamicina inefficace) mentre la Rapamicina impediva l’effettiva degradazione delle concentrazioni della proteina IRS-1 (Vanadio inefficace). (18)
La carenza di Vanadio non è stata descritta nell’uomo. Un suo scarso apporto, o mancanza, negli animali causa sterilità, riduzione della eritropoiesi (con conseguente anemia), difetti del metabolismo del ferro, alterata formazione ossea, dentale e cartilagginea. Non esiste quindi un RDA per il Vanadio: si è ipotizzato che un’assunzione giornaliera nel range dei 10 – 100mcg sia con tutta probabilità sufficiente.
Il Vanadio viene commercializzato come integratore alimentare sotto forma di Vanadilsolfato [VOSO4]. La biodisponibilità del Vanadio supplementare è scarsa (meno del 5%) e la maggior parte di esso viene espulso attraverso le feci.
Alcuni prodotti conteneti Vanadilsolfato riportano in etichetta che il composto “aumenta i livelli plasmatici di Insulina”, ma, da quanto è possibile estrapolare dalle informazioni sopra riportate, e cioè da quanto la letteratura scientifica ci dice sull’azione del composto, il Vanadio facilita il meccanismo d’azione dell’ormone peptidico, con una probabile azione (parzialmente dimostrata) insulino-mimetica.
Prendendo in attenta considerazione le informazioni derivanti dalla letteratura scientifica sulle possibili potenzialità del Vanadio nei confronti del metabolismo glucidico si evince, e non è un caso isolato tra i GDA, che la sua efficacia sia di grado significativo nei soggetti patologici (vedi soggetti con diabete di tipo II) e non negli individui sani. Ipoteticamente si potrebbero avere dei vantaggi dal suo utilizzo nei soggetti con una insulino-resistenza di base genetica, nei periodi di alimentazione ipercalorica prolungata (vedi peggiormanto dell’insulino-resistenza in tale contesto) e/o durante e dopo l’uso dell’Insulina esogena: durante, il suo potenziale effetto insulino-mimetico potrebbe permettere di per se di utilizzare dosi inferiori dell’ormone mentre, nel periodo successivo, potrebbe agevolare il ripristino di un ottimale insulino-sensibilità in concerto con altri GDA. L’effetto del Vanadio sulla riduzione della gluconeogenesi epatica potrebbe rappresentare un vantaggio nei periodi di restrizione calorica, in specie durante le diete Ketogeniche o simil tali. Sicuramente, l’impatto negativo del Vanadio sui Trigliceridi potrebbe causare un ulteriore peggioramento della dislipidemia indotta dall’uso di AAS.
E’ bene tenere a mente che le dosi efficaci per ottenere un qualche effetto positivo dall’integrazione con Vanadio (Vanadilsolfato) sono considerevoli per via della bassa biodisponibilità che, però, non elimina la possibilità di intossicazione proprio a causa di queste. I possibili effetti derivanti dall’assunzione di livelli tossici di Vanadio comprendono nausea, mal di stomaco, diarrea, ipertrigliceridemia, disfunzioni epatiche, danni renali, ipoglicemia, leucopenia, ritardo dello sviluppo ed inappetenza.
La dose tossica di Vanadio è quantificata essere di 25mg al giorno. Quindi, considerando anche la bassa biodisponibilità del composto, una dose potenzialmente efficace ma che garantisca comunque un buon margine di sicurezza è di circa 100mg/die di Vanadilsolfato (20mg di Vanadio) divisa in due-tre somministrazioni prima dei pasti principali.
La supplementazione di Vanadio può comportare delle interazioni farmacologiche con Warfarin e Coumadin (difficoltà di coagulazione) o con farmaci con azione ipoglicemizzante diretta e/o indiretta come Aspirina e Exubera (riduzione eccessiva della glicemia ematica).
Quindi, nel caso si decidesse di testare l’efficacia di questo minerale, il potenziale utilizzatore dovrebbe prestare particolare attenzione a quanto appena esposto partendo dalla dose giornaliera assunta.
L’estratto dei funghi Cordyceps sinensis e Ganoderma lucidum sembra poter aumentare i livelli di Testosterone e abbassare i livelli di Cortisolo negli atleti sottoposti ad allenamenti di resistenza. Ciò è emerso in seguito ad uno studio svolto dai ricercatori dell’Università di Pavia su sette ciclisti trattati con l’estratto di entrambi i funghi. (1)
Durante l’esperimento, i ricercatori hanno somministrato a sette ciclisti dilettanti, di età compresa tra i 30 ed i 40 anni, capsule contenenti un placebo per un paio di settimane e capsule contenenti estratto di Cordyceps sinensis e Ganoderma lucidum per tre mesi. Durante entrambi i periodi i ciclisti hanno preso parte a una competizione ciclistica su 85 km.
I ciclisti hanno assunto tre capsule al giorno, ciascuna delle quali contenente 445 mg di Cordyceps sinensis. Le capsule consistevano di un terzo di polisaccaridi e 0,5% di Adenosina. Inoltre, le capsule contenevano la Cordicepina, un derivato del nucleoside Adenosina, differente da essa per l’assenza di un atomo di ossigeno nella posizione 3 dell’anello di ribosio. L’Adenosina e la Cordicepina probabilmente svolgono entrambi un ruolo chiave nell’effetto ergogenico legato all’uso del Cordyceps sinensis. Tre giorni prima di ciascuna gare, i ciclisti hanno aumentato la loro assunzione a sei capsule al giorno.
Precedenti studi avevano dimostrato che la supplementazione con estratto di Cordyceps sinensis, ad un dosaggio di 3g, non aveva alcun effetto sull’assorbimento dell’ossigeno o sulla capacità di resistenza dei ciclisti (2), ma aumentava la capacità di resistenza in soggetti di età superiore ai 50 anni. (3)
Nello studio qui discusso, i ciclisti hanno assunto anche due capsule al giorno contenenti 390mg di estratto di Ganoderma lucidum. Questo estratto era composto da un terzo di polisaccaridi e dall’1,5% di Triterpeni.
Quando i ricercatori hanno analizzato la saliva prelevata dai ciclisti trattati con placebo poco prima e subito dopo la gara, hanno osservato che le concentrazioni di Testosterone di due di questi non erano cambiate durante la gara. Mentre le concentrazioni di Cortisolo risultavano più basse. Quindi, è probabile che questi ciclisti avessero un grado di adattamento ottimale alla performance richiesta.
Quando trattati con Cordyceps sinensis e Ganoderma lucidum, i ciclisti, per via esame salivare, hanno mostrato un aumento dei livelli di Testosterone, come mostrato nella figura seguente.
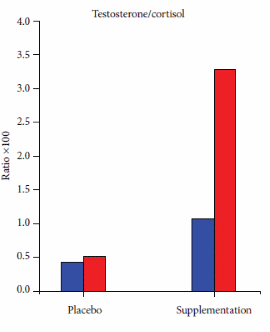
In ciclisti, la gara condotta in condizioni di trattamento con placebo ha portato ad un moderato abbassamento delle concentrazioni di Testosterone e ad un aumento del Cortisolo, il che suggerirebbe che questi atleti erano vicini alla soglia dell’overreacting. La figura riportata qui sopra mostra che l’integrazione con Cordyceps sinensis e Ganoderma lucidum ha aumentato la concentrazione di Testosterone nei ciclisti prima e dopo la gara inibendo anche l’aumento del Cortisolo post-gara.
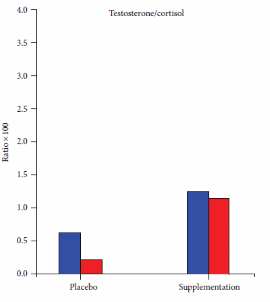
La supplementazione con estratti di questi due funghi ha migliorato la Testosterone/Cortisolo ratio sia nei ciclisti ben allenati [prima figura sotto] che nei ciclisti meno allenati [seconda figura sotto].
I ricercatori hanno affermato che, in seguito ai risultati ottenuti, si possa concludere che un periodo di tre mesi di supplementazione con estratti di Cordyceps sinensis e Ganoderma lucidum possa proteggere gli atleti di resistenza dall’overreaching / overtraining. Inoltre, i ricercatori hanno sostenuto la ovvia necessità di svolgere ulteriori ricerche in follow-up con un gruppo più ampio di atleti.
Un interessante sviluppo futuro di questa ricerca sarebbe l’analisi dei parametri infiammatori al fine di comprendere il ruolo svolto dall’integrazione fungina sul sistema immunitario. Lo studio dovrebbe continuare a selezionare integratori alimentari fungini standardizzati, ma dovrebbe essere esteso in modo da includere un numero maggiore di atleti di resistenza, a causa della variabilità delle loro condizioni atletiche e dell’impatto di queste nella risposta alla supplementazione presa in analisi.
A seguito di uno studio in vitro svolto da tossicologi dell’Università di Basilea e pubblicato nel 2012, è emerso che il Fluoxymesterone può causare un aumento significativo dei livelli di Cortisolo. (1) Argomento da me accennato nell’articolo dedicato alla molecola in questione.
Come risaputo, il Fluoxymesterone è un AAS orale metilato in C-17, con un potere androgeno elevato e non soggetto all’enzima aromatasi.
Nonostante quest’ultimo punto, la casa produttrice (Pfizer) riporta nelle avvertenze del prodotto una caratteristica che non ci si aspetta da una molecola priva di attività estrogenica diretta e indiretta: “L’edema, con o senza insufficienza cardiaca congestizia, può essere una seria complicanza in pazienti con preesistente malattia cardiaca, renale o epatica”.(2)
I ricercatori hanno scoperto il meccanismo attraverso il quale il Fluoxymesterone può causare edema e quindi aggravare ulteriormente la sua influenza sulla salute del sistema cardiovascolare. Il Fluoxymesterone si lega all’enzima 11-beta-HSD2, enzima preposto alla conversione del Cortisolo in Cortisone (inattivo). Di conseguenza il gruppo 11-idrossile del Fluoxymesterone viene convertito in un gruppo 11-oxo. Poiché l’attività dell’11-beta-HSD2 subisce una riduzione, si osserva un aumento della concentrazione di Cortisolo.
I ricercatori hanno esteso la loro ricerca ad altri composti al fine di valutarne una possibile azione sui meccanismi di conversione del Cortisolo in Cortisone. L’esito è stata la scoperta della marcata attività inibitoria dell’11-beta-HSD2 da parte del Fluoxymesterone. Durante l’esperimento è stato constatato che il Fluoxymesterone esplica una potenza maggiore sull’alterazione dei livelli di Cortisolo dell’Acido Glicirretico, sostanza presente nella liquirizia che causa un aumentano della produzione endogena di corticosteroidi, attraverso l’inibizione degli enzimi 4 e 5-beta-reduttasi che inattivano gli steroidi.(3)
L’Oxymesterone – o 4-idrossi-17-metil-testosterone – e, in misura minore, l’Oxymetholone inibiscono l’11-beta-HSD2 quasi quanto il Fluoxymesterone.
I ricercatori hanno scoperto anche che il Fluoxymesterone non può interagire direttamente con i Recettori del Cortisolo. Ma, le concentrazioni aumentate di Cortisolo portano, ovviamente, ad un consequenziale aumento dell’attività di quest’ultimo.
Un eccesso di Cortisolo altera l’equilibrio elettrolitico data l’interazione dello steroide con i Recettori Mineralocorticoidi. Induce il corpo a trattenere più sodio e quindi aumenta la ritenzione idrica extracellulare. Ciò significa che la quantità di plasma nel sangue aumenta e di conseguenza aumenta la pressione sanguigna. Oltre a ciò, un livello elevato di Cortisolo ha un effetto restringente sui vasi sanguigni, cosa che, a sua volta, causa un aumento della pressione sanguigna. Infine, un livello elevato di Cortisolo rende i vasi sanguigni più suscettibili ai danni causati dall’accumulo di colesterolo nelle loro pareti.
Pertanto, gli AAS con azione inibitoria nei confronti dell’11-beta-HSD2, come il Fluoxymesterone, possono avere un azione avversa maggiore nel causare effetti cardiovascolari avversi.
Si è ipotizzata anche una differenza nell’impatto sull’11-beta-HSD2 e i livelli di Cortisolo tra assunzione orale e somministrazione per iniezione con maggiore influenza data da quest’ultima. La cosa potrebbe con molta probabilità essere legata alle modifiche che la farmacocinetica subisce con la somministrazione tramite iniezione rispetto alla classica somministrazione orale.
Per avere una visione d’insieme più completa riguardo al Fluoxymesterone vi rimando all’artico ad esso dedicato.
Se non avete letto ancora la prima, la seconda e la terza parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte – 3° Parte.
Cannella: caratteristiche e possibili applicazioni.
La Cannella è una spezia nella quale sono contenuti diversi agenti bioattivi. L’aldeide cinnamica conferisce alla Cannella il suo aroma (1), la Cumarina (una tossina) contribuisce al gusto (2) e diversi composti, tra cui il polifenolo MHCP (methylhydroxychalcone polymer), sembrano contribuire ai suoi benefici sistemici sulla sensibilizzazione all’Insulina.(3) Oltre ai tre composti unici elencati, la Cannella contiene anche tannini, flavonoidi, glicosidi, terpenoidi e antrachinoni. (4)
La Cannella sembra poter esercitare effetti di controllo benefici in contesti dietetici “pro-diabetici” attraverso diversi meccanismi.
La Cannella può inibire numerosi enzimi digestivi, come l’α-glucosidasi (4), il Saccarasi (5) e, potenzialmente, l’Amilasi Pancreatica.(5) Attraverso l’inibizione di questi enzimi, la Cannella può ridurre l’afflusso di glucosio nella circolazione sistemica ed evitare così picchi insulinici eccessivamente significativi.
MHCP
Anche nella circolazione sistemica (oltre il fegato) la Cannella sembra possedere effetti anti-diabetici. Un composto presente in essa, il prima citato polifenolo MHCP, agisce come un insulino-mimetico a livello adipocitario.(3)(6)(7) Gli effetti insulino-mimetici del MHCP sono dose-dipendenti e agiscono trasfosforizzando il Recettore dell’Insulina sulla membrana citoplasmatica (lo stesso meccanismo d’azione esplicato dalla molecola d’Insulina). I suoi effetti sull’assorbimento del glucosio e sullo stoccaggio del glicogeno, anche se dose-dipendenti, sembrano avvenire lentamente ( l’Insulina ha un tempo di risposta che si manifesta entro 10 minuti dal raggiungimento della cellula, mentre con il MHCP i tempi di risposta variano da 30 a 60 minuti, suggerendo, appunto, un ritardo temporale intracellulare).(3)
In vitro, la Cannella ha mostrato di potenziare l’azione insulinica di 20 volte.(8)
Quando ingerita da modelli umani durante gli esperimenti, la Cannella ha mostra molte potenzialità nella riduzione dei livelli di glucosio ematico (9)(10)(11) e, talvolta, dei marker del metabolismo lipidico (LDL, Trigliceridi, Colesterolo totale).(12) Ci sono anche studi nei quali è stato osservato un miglioramento dell’insulino-sensibilità con l’assunzione dell’estratto di Cannella, risposta probabilmente legata in modo indiretto alla riduzione dei livelli di glucosio nel sangue.(13)(14)
Cumarina
Come accennato in precedenza, la Cumarina è una sostanza fitochimica epatotossica e cancerogena presente in alcune piante e ad alti livelli in alcune varianti della Cannella. La Cumarina non è il composto attivo che riduce la glicemia ematica, ma è uno dei principi attivi presenti nella Cannella. Inizialmente, questa sostanza aveva un TDI (dose giornaliera tollerabile) di massimo 2mg/kg di peso corporeo, ma tale limite è stato abbassato a 0,5 e attualmente è pari a 0,1mg/kg di peso corporeo.(2) Sebbene in quest’ultima raccomandazione sia incluso un margine di sicurezza, alcuni sottogruppi della popolazione umana sono più sensibili alla tossicità della Cumarina a causa della ridotta capacità di metabolizzarla.(2)
Ciò è rilevante dal momento che la maggior parte dei benefici antidiabetici riscontrati con l’uso della Cannella sono dose-dipendenti, nell’intervallo di 300mg/kg di peso corporeo.(5) A questa dose, è particolarmente facile oltrepassare il TDI della Cumarina.
Il modo migliore per evitare di assumere dosi rilevanti di Cumarina è optare per la giusta fonte di Cannella. La Cannella di Ceylon presenta i livelli più bassi di Cumarina con meno di 190mg/kg (alcuni campioni sono al di sotto dei livelli di rilevazione) mentre la Cassia contiene tra i 700mg/kg ed i 12.230mg/Kg.(15) La Ceylon può essere riconosciuta in forma integra attraverso le sue sottili e numerose pieghe, mentre la Cassia, sempre in forma integra, ha meno pieghe e un aspetto più spesso. Non possono essere distinte in forma di polvere e la Cassia è più frequentemente utilizzata nella realizzazione di prodotti alimentari per via della sua elevata disponibilità e del basso costo.(2)(16)
Tramite i numeri di cui sopra, un essere umano di 90Kg può ingerire 47,8g di Cannella di Ceylon e arrivare, nella peggiore delle ipotesi, ad assumere 0,1 mg/kg di peso corporeo del TDI della Cumarina (assumendo, quindi, la punta massima di assunzione della Cumarina). Diversamente, utilizzando la Cannella Cassia può facilmente essere ingerito un dosaggio di Cumarina al di sopra del TDI con un apporto decisamente inferiore di prodotto.
L’assorbimento della Cumarina non sembra dipendere dalla forma di Cannella ingerita. Livelli serici simili e livelli escreti sono stati raggiunti con l’assunzione di cumarina isolata, di pillole contenenti cannella, tè e budino di riso (cibo solido).(17) Questi risultati sono stati standardizzati alla dose X di Cumarina, quindi la fonte di Cannella è irrilevante.
Lo studio di cui sopra, tuttavia, ha notato un tasso di estrazione del 38,5% di Cumarina dalla polvere al liquido una volta entrati in contatto (appena bollente per 30 minuti); suggerendo che si può ribaltare la bilancia a favore dei polifenoli e del MHCP solubili in acqua rispetto alla Cumarina se la Cannella viene messa in acqua e servita nel tè o usando l’acqua di infusione per mescolare frullati proteici, poiché i componenti idrosolubili hanno un tasso di estrazione molto più alto.
Come spesso accade, però, sono stati gli studi iniziali svolti sulla Cannella ha mostrare una efficacia marcata dei composti ivi contenuti.(12)(18) Ma, nonostante le possibili e promettenti potenzialità rilevate nelle prime ricerche, al momento, l’utilizzo della Cannella ha mostrato soltanto una riduzione della glicemia ematica in acuto con ridotti e ben poco rilevanti risultati in cronico, come la mancata influenza sulla emoglobina glicata (HbA1c) o su un reale miglioramento della insulino resistenza.
I meccanismi attraverso i quali l’assunzione di Cannella porta ad un abbassamento della glicemia post-prandiale in modo dose dipendente sono quindi legati al rallentamento dello svuotamento gastrico, all’inibizione di numerosi enzimi digestivi tra cui i prima citati alfa-glicosidasi, maltasi , sucrasi e, anche, da una sorta di azione “carb-blocker” data dalla possibile inibizione del enzima amilasi pancreatico.
Come già accennato, i risultati più importanti riscontrati sono legati per lo più ad una riduzione della risposta glicemica in acuto dopo trenta minuti dal pasto senza modifiche profonde degli altri marker del controllo glicemico. Nonostante ciò, non sono da escludere possibili vantaggi di questa natura con un assunzione di Cannella sul lungo periodo, sebbene, come ormai risaputo, l’Indice Glicemico non è un parametro rilevante sulla qualità del dimagrimento e, quindi, della composizione corporea.
Acarbosio
Anche nei soggetti diabetici, l’uso in monoterapia della cannella è risultato fallimentare mostrando semplicemente una validità additiva in concomitanza con l’assunzione di farmaci ipoglicemizzanti. Ancora una volta, tale risultato potrebbe trovare un nesso causale nella possibile attività “carb-blocker” simile a quella osservata in seguito ad assunzione di Acarbosio, farmaco utilizzato nel trattamento del diabete di tipo II e dell’obesità avente azione ritardante sulla digestione e l’assorbimento di zuccheri alimentari in quanto è un inibitore dell’alfa-glucosidasi intestinale e, attraverso tale meccanismo, ha un effetto nel ridurre la glicemia post-prandiale. Riduce inoltre i livelli di trigliceridi, di emoglobina glicata (HbA1) e la resistenza periferica all’insulina.(19)
Nei soggetti sani, quindi, sembrerebbe maggiormente plausibile che la riduzione della glicemia ematica dopo il pasto sia da attribuirsi quasi totalmente all’inibizione della digestione e assorbimento dei carboidrati ma non ad un effettivo miglioramento dell’insulino sensibilità.
Alfa Amilasi
La questione prima trattata sulla lenta risposta ipoglicemizzante osservata in seguito all’assunzione di Cannella è oggetto di dibattito per quanto riguarda il suo meccanismo. Sembrerebbe, infatti, che tale effetto sia dovuto quasi esclusivamente all’inibizione dell’alfa amilasi e, quindi, ad una rallentata assimilazione dei carboidrati ingeriti, e non dal azione insulino-mimetica del MHCP. Se così fosse, questo tipo di risultati sarebbero ottenibili con una semplice ridistribuzione dei macronutrienti e, in particolare, con una riduzione del carico glucidico del singolo pasto.
A questo punto, dopo aver letto le informazioni presenti in questo articolo, si potrebbe giungere facilmente alla conclusione secondo cui l’uso della Cannella sia relegabile al solo insaporire cibi o bevande, vista la limitatezza della sua azione sul metabolismo glucidico. Nonostante ciò, la sua possibile applicazione come GDA non è completamente da escludere, sebbene con forti limitazioni legate alle sue caratteristiche e a quelle del soggetto al quale viene somministrata a tal fine.
L’uso della Cannella potrebbe apportare dei vantaggi, anche se minimi, se abbinata ad altri GDA (vedi Berberina, ALA e Cromo) in soggetti con una marcata insulino resistenza, sia di base genetica e/o “situazionale” (legata ad una condizione di sovrappeso/obesità e/o per via di una alimentazione ipercalorica/iperglucidica). Tali soggetti, avendo difficoltà nella gestione di carichi glicemici (nel singolo pasto) anche di entità moderata, possono sfruttare l’effetto di riduzione/rallentamento dell’assorbimento glucidico post prandiale dato dalla Cannella, così da avere una migliore gestione del picco glicemico, gestione ulteriormente migliorata dagli altri GDA co-assunti e da alcune accortezze alimentari come la scelta della fonte glucidica. 1g di Cannella di Ceylon appena prima del pasto sembra essere un dosaggio sufficiente ad esplicare tali effetti.
I risultati addizionali non saranno particolarmente evidenti ma, nei casi sopra citati, l impatto potrebbe facilitare la gestione del carico glucidico.
Ovviamente, soggetti nella norma in salute possono benissimo evitare di usare la Cannella con tali finalità e limitarsi, secondo palato, al suo uso come semplice spezia.
Un basso dosaggio di Witaferina A, un lattone steroideo presente nella pianta indiana Ashwagandha, potrebbe facilitare la perdita di peso nei soggetti sovrappeso o obesi. I ricercatori della Harvard Medical School hanno ipotizzato ciò in seguito allo svolgimento di uno studio effettuato su topi resi grassi i quali, in seguito alla somministrazione di Witaferina A, avevano perso ¼ del loro peso in tre settimane.(1)
Per lo svolgimento dell’esperimento, i ricercatori hanno usato topi normopeso fatti ingrassare attraverso una dieta ipercalorica.
I ricercatori hanno somministrato la Witaferina A direttamente nell’intestino tenue di una parte degli animali presi in esame. Se al posto dei topi ci fossero stati soggetti umani di 100Kg, la dose somministrata di Witaferina A (in caps) sarebbe stata di circa 25mg/die.
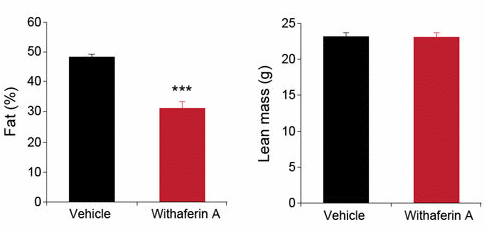
La somministrazione di Witaferina A ha portato ad una riduzione del peso corporeo dei topi [in basso a sinistra]. Ciò era dovuto principalmente al fatto che la Witaferina A induceva gli animali trattati a consumare meno cibo [in basso a destra].
La Witaferina A ha ridotto la massa grassa. Sebbene i topi del gruppo trattato con il lattone steroideo abbiano perso peso, la loro massa magra è rimasta praticamente intatta.
Gli animali trattati con Witaferina A mostravano una normalizzazione dei livelli serici di Leptina.
I ricercatori hanno ripetuto l’esperimento utilizzando topi magri con un normale livello di Leptina. In essi, la Witaferina A non ha avuto alcun effetto sul peso e sulla composizione corporea. I ricercatori hanno ripetuto l’esperimento anche con i topi ob/ob che non producono Leptina a causa di un difetto genetico, e con topi db/db con un difetto del recettore della Leptina. In entrambi i casi gli animali non avevano un controllo sul consumo di cibo e, di conseguenza, avevano subito un marcato aumento della massa grassa. Il trattamento con la Witaferina A in questi animali non ha mostrato quasi alcun effetto.
La sintesi di Leptina aumenta in risposta ad un aumento delle riserve adipose o, più precisamente, ad un aumento del metabolismo glucidico adipocitario. Questo effetto riduce l’appetito. Quando le riserve adipose diminuiscono, la sintesi di Leptina diminuisce e l’appetito aumenta. Tuttavia, se si consuma un eccesso calorico nel lungo termine, questo meccanismo viene alterato. Si sviluppa resistenza alla Leptina. Si ipotizza, quindi, che la supplementazione di Witaferina A aumenti la sensibilità alla Leptina.
I ricercatori scrivono che, sono passati più di due decenni dalla scoperta storica della Leptina da parte di Friedman e colleghi (2), ma finora non è stato sviluppato un trattamento leptino-centrico applicabile per il trattamento dell’obesità. Subito dopo le prime pubblicazioni sulla Leptina, è stato suggerito che l’obesità sia una condizione correlata alla leptino-resistenza.
Nel corso degli ultimi vent’anni, le speranze per lo sviluppo di un trattamento orientato verso la manipolazione/attività della Leptina nell’obesità sono progressivamente diminuite, poiché molti tentativi di ri-sensibilizzare il cervello degli individui obesi al peptide erano falliti. Questi sforzi infruttuosi per aumentare la sensibilità alla Leptina e trattare lo stato iperleptinemico nell’obesità per trattare tale condizione hanno anche contribuito in modo sostanziale al dibattito sulla presenza o meno della leptino-resistenza.
Il trattamento di topi obesi e iperleptinemici con Witaferina A ha portato ad una forte riduzione dell’assunzione di cibo e del peso corporeo. Entrambi le risposte osservate sono legate ai livelli di Leptina serica. Poiché i livelli di Leptina diminuiscono gradualmente durante il periodo di trattamento, parallelamente alla stabilizzazione del peso e della percentuale di massa grassa degli animali trattati, anche l’effetto della Witaferina A diminuisce gradualmente. Inoltre, nei topi magri, con bassi livelli di Leptina circolante, non sono stati osservati cambiamenti nel consumo di cibo o nel peso corporeo.
Gli estratti di Withania somnifera, che contengono anche Witaferina A, sono stati usati dall’uomo per secoli, e sembrano possedere un effetto fortemente conservativo sul sistema della Leptina nei mammiferi tra cui topi e umani. Considerando tutte queste informazioni, i ricercatori ritengono che l’uso della Witaferina A per il trattamento dell’obesità negli esseri umani sia una grande promessa per il futuro.
Se non avete letto ancora la prima e la seconda parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte.
Cromo: caratteristiche e possibili applicazioni.
Cromo Picolinato
Il Cromo è un minerale essenziale nella dieta umana ed è comunemente utilizzato come integratore alimentare (es. Picolinato o Polinicotinato) per migliorare la sensibilità all’insulina nei soggetti sani o nei soggetti diabetici. (1)
Il Cromo può essere trovato nel:
Colostro bovino (sotto forma di un oligopeptide di cromodulina ricco di zinco, con un atomo di Cromo per quattro amminoacidi (2) (3)) che fornisce 220mcg di Cromo per 1.035g di proteine (193ng/g di proteine) (2)
• Latte scremato, ad una concentrazione di 252mcg di Cromo per 1.172g di proteine (215ng/g di proteine) (2)
Il Cromo è sia un minerale dietetico che un elemento (Cr) con più valenze. La forma completamente ossidata di Cromo (Cr (VI)), che è esavalente (+6 stato di ossidazione), è altamente tossica e impiegata in una varietà di applicazioni industriali.(4) Dato l’alto grado di tossicità, il Cromo esavalente non viene mai usato come integratore. Le forme supplementari di Cromo comprendono il bivalente (Cr (II)) o il trivalente (Cr (III)), quest’ultima è la forma più stabile.(1)
Il quantitativo di Cromo assunto con la dieta dovrebbe essere almeno di 0,005-0,2mg(5-20mcg) al giorno al fine di prevenirne il deficit, e l’assunzione giornaliera raccomandata è di 21-25mcg per le donne e di 25-35mcg per gli uomini con la fascia di età tra i 18 ed i 45 anni che richiede quantità verso il punto più alto dell’intervallo riportato.(5) Le donne di tutte le età che stanno allattando richiedono un’assunzione giornaliera di Cromo pari a 45mcg.(5) La dose raccomandata per i bambini da 1 a 3 anni è 11mcg/die mentre dai 4 agli 8 anni il dosaggio sale a 15mcg/die.(5)
Le concentrazioni standard di Cromo circolante in uno stato non carente sono state misurate nell’intervallo di 2,8-45mcg/L nel sangue intero e 0,12-2,1mcg/L nel siero.(6)
Una carenza di Cromo può essere indotta con una nutrizione parenterale totale a lungo termine (TPN) priva del minerale, e può essere invertita con una supplementazione di 150mcg di Cromo al giorno aggiunti al TPN come riscontrato attraverso un caso studio (7) e 250mcg al giorno per 2 settimane seguite da una dose di mantenimento pari a 20μg al giorno per 18 mesi in un altro.(8) I principali sintomi da carenza di Cromo in questi particolari casi si manifestavano attraverso un compromessa tolleranza al glucosio e una riduzione dell’insulino-sensibilità associata alla perdita di peso, così come la neuropatia e l’encefalopatia che erano reversibili con il reintegro del minerale. (8)(7)
Quindi, una grave carenza di Cromo è associata a sintomi simili a quelli riscontrati nel diabete di tipo I (alterata tolleranza al glucosio e perdita di peso) e nella neuropatia, e può essere invertita con la somministrazione del minerale.
Le carenze subcliniche di Cromo sono associate all’insulino resistenza, poiché le concentrazioni di questo minerale sono risultate inferiori nei diabetici rispetto ai soggetti di controllo (9) (tuttavia, l’evidenza è eterogenea per il diabete gestazionale (10)(11)). Le diete con un assunzione cronica di zuccheri (35% delle calorie giornaliere) sono state associate ad una accelerata perdita di Cromo attraverso le urine (Cromo urinario) (12) sebbene le diete composte da cibi ad alto indice glicemico non abbiano influenzato in modo significativo l’eliminazione del Cromo attraverso le urine in soggetti sani, pur mostrando una tendenza nell’arco di sei giorni.(13)
Cromodulina
Si ritiene che questa perdita accelerata di Cromo attraverso le urine si verifichi per via del rilascio di Cromodulina (LMWCr; Low-molecular-weight chromium-binding substance) nel flusso ematico da parte delle cellule insulino-sensibili, con conseguente eliminazione urinaria.(14) La Cromodulina è un peptide che esiste all’interno delle cellule. Quando combinato con il Cromo immesso nelle cellule dal flusso sanguigno, amplifica la segnalazione dell’insulina legandosi ai recettori insulinici stimolati dall’ormone.(14) La Cromodulina lega lo ione cromo ad altissima affinità, formando un complesso che può essere separato solo in condizioni non fisiologiche. Una volta che i livelli di insulina scendono, tuttavia, i recettori dell’insulina non hanno più bisogno di essere sensibilizzati, quindi l’intero complesso deve essere eliminato nel suo insieme.(14) (15)Questa ipotesi è supportata dal rilevamento della Cromodulina nelle urine (16) e dalla sua stretta correlazione con i tassi di secrezione dell’Insulina e l’esposizione in condizioni non complementari.(16) (17) (18)
Le concentrazioni urinarie di Cromo risultano elevate in seguito ad allenamenti di resistenza (con un aumento di cinque volte dopo due ore di corsa, ma con solo un aumento di due volte nel corso delle ventiquattro ore) in un modo che non è correlato ad un aumento dell’insulina serica o ad un aumento di qualsiasi altro ione urinario.(18) Questa condizione, nonostante l’assenza di significativi livelli di Insulina, è nota per richiedere un maggiore assorbimento di glucosio nel tessuto muscolare sostenuto da un maggiore rilascio di glucosio da parte del fegato.(19)
Il Cromo trivalente (che si trova negli integratori) sembra avere effetti tossici a concentrazioni superiori a 20mcg/mL nel siero o nelle cellule; questa tossicità è associata al danno ossidativo al DNA.(20) Questo è lo stesso meccanismo mediante il quale il cromo esavalente esprime la sua tossicità, con l’unica differenza che quest’ultimo è tossico a concentrazioni molto più basse (21), in particolare dopo inalazione durante un impiego che comporta la sua manipolazione. (22) (4)
Con il termine Cromo Picolinato ci si riferisce al Cromo nello stato trivalente (Cr (III)) il quale è legato a tre molecole di acido picolinico, un analogo strutturale della Niacina. Questa forma di Cromo è altamente stabile (23), a parte una possibile degradazione indotta dall’acido, che rimuove una molecola di picolinato e porta a due ioni di cromo che si legano insieme. (24) I ligandi picolinati sono in una posizione tale che il Cr (III) può essere ridotto in Cr (II) nella coltura cellulare senza perdere il picolinato (25), una proprietà che sembra essere unica per il picolinato rispetto ad altre forme supplementari (Cloruro e Nicotinato) e si pensa che sia alla base delle possibili proprietà cancerogene indotte da alte concentrazioni.(26)
Si ritiene che il Cromo Picolinato sia fisiologicamente inattivo fino ad avvenuta liberazione della molecola di Cromo (26), suggerendo che esso funga da “pro farmaco” al Cromo.
Il Cromo è noto per essere presente nel lievito, dove svolge un ruolo fisiologico importante.(27) (28) All’interno delle cellule del lievito si trova il “Fattore di Tolleranza al Glucosio” (GTF) (29), che è stato inizialmente derivato dal lievito di birra.(30) Il GTF può essere purificato dai lieviti dopo l’estrazione metanolica e la successiva filtrazione, ottenendo un insieme di molecole di dimensioni variabili da 1.000 a 3.500 Da. (31)(32) I principali componenti attivi in questo set di molecole sono considerati l’acido trivalente al cromo nicotinico insieme ad alcuni aminoacidi (Glicina, L-cisteina e Acido Glutammico).(33) L’apporto alimentare del lievito sembra conferire alcuni dei benefici dati dall’integrazione di Cromo, probabilmente a causa dell’ingestione di GTF e Cromo.(29)
Si ritiene che il Cromo presente nel lievito sia acido cromo-nicotinico, sebbene possano esistere altre forme di Cromo nel lievito che non sono state ancora rilevate.
Cromo Polinicotinato
L’Acido Nicotinico di Cromo (noto anche come Cromo Polinicotinato), forma altamente assimilabile di Cromo, è composto da Cromo legato all’Acido Nicotinico (Niacina o Vitamina B3) e si dice che abbia effetti sulla riduzione del Colesterolo.(34)(35) Negli studi in cui il Colesterolo è stato ridotto in seguito all’assunzione di Cromo Polinicotinato, non è stato trovano necessariamente un nesso benefico dato dal miglioramento del metabolismo glucidico (34)(35), suggerendo che è la Niacina a causare questi effetti.
Il Cromo Dinicocisteinato (CDNC) è un complesso dello ione Cromo con l’aminoacido L-cisteina. Uno studio che ha confrontato l’effetto di 400mcg di CDNC con 400mcg di Cromo Picolinato ha rilevato miglioramenti nei livelli di Insulina e della sensibilità a questa solo con il CDNC. (36)
Uno dei principali meccanismi che si ritiene correlato all’integrazione con Cromo comporta la modulazione della via di segnalazione dell’Insulina.(37) [38] Questo è stato scoperto per la prima volta quando è stato identificato un oligopeptide legante il Cromo a basso peso molecolare che ha aumentato gli effetti dell’Insulina e l’ossidazione del glucosio. (38)[39] Chiamato anche LMCr o Cromodulina (39), questo oligopeptide viene sintetizzato nel fegato dei ratti dopo iniezioni di Cromo (40) e ha una massa di circa 1500 kDa. (39)(41)
È stato rilevato che la Cromodulina ha aumentato la segnalazione di Insulina in presenza di un livello di quest’ultima pari a 5-8 volte superiore rispetto all’attività basale, senza influenzarne la segnalazione in assenza di Insulina.(42) La deplezione di Cromo da parte della Cromodulina ne blocca l’attività (42) che si correla positivamente con il contenuto del minerale nel peptide. Inoltre, altri minerali non sono riusciti a replicarne gli effetti. (43)
Recettore dell’Insulina
In definitiva, la Cromodulina aumenta l’autofosforilazione del Recettore dell’Insulina. La segnalazione del Recettore dell’Insulina richiede che l’Insulina o un mimetico (qualcosa che si comporti come l’Insulina) si leghi alla subunità α extracellulare del recettore (44) che consente alla subunità β intracellulare di essere autofosforilata. (44) La Cromodulina sembra agire intracellularmente nella subunità β del Recettore dell’Insulina.(14)
Le funzioni cromo-dipendenti della Cromodulina sono probabilmente la ragione biologica per la quale il Cromo è un minerale essenziale (45), sebbene la natura essenziale del Cromo sia stata recentemente contestata.(46)
L’Adenosina Monofosfato Chinasi (AMPK) è un sensore chiave dello stato energetico cellulare, il quale monitora costantemente i livelli di ATP al fine di mantenere l’omeostasi metabolica. L’AMPK si attiva durante gli stati di carenza energetica (caratterizzato da un aumento della AMP:ATP ratio) dove coordina il metabolismo degli acidi grassi e del glucosio in modo anti-obesità e anti-diabetico.(47) Quando attivato, l’AMPK sopprime le vie anaboliche come la sintesi proteica, di trigliceridi e di acidi grassi attivando contemporaneamente percorsi catabolici come la glicolisi e l’ossidazione degli acidi grassi per aumentare la produzione di ATP. (48)
È stato notato che il Cromo (trivalente con D-fenilalanina) attiva l’AMPK nel suo sito catalitico (Thr172) nei cardiomiociti e nelle cellule muscolo-scheletriche a 25μM, suggerendo che i complessi organici del Cromo possono essere nuovi attivatori della via dell’AMPK.(49)
L’assorbimento del Cromo alimentare è inversamente correlato all’assunzione, variando dallo 0,4% al 2,0%, con l’assorbimento più efficiente (2%) a un apporto dietetico inferiore di circa 10mcg negli uomini adulti.(50) Questo diminuisce a circa lo 0,5% quando l’assunzione con il cibo raggiunge i 40mcg che sembra essere il limite, dato che l’assunzione di Cromo nel range di 40-240mcg ha un assorbimento di circa lo 0,4%. (50) (51)
Acido Fitico
L’assorbimento del Cromo è influenzato da una serie di fattori dietetici. Nei ratti, l’assorbimento del Cromo sembra essere ostacolato dalla coingestione dei fitati, che impedisce il trasporto e l’assorbimento attraverso l’intestino. (52) È stato dimostrato che il deficit di Zinco aumenta l’assorbimento del Cromo, che è aumentato nei ratti carenti di Zinco e ridotto dallo Zinco supplementare (53), suggerendo che questi due minerali possono competere per l’assorbimento. L’assorbimento del Cromo nei ratti è anche potenziato dall’ossalato, un acido organico presente in molte verdure e cereali. (52) Sebbene sia informativo, occorre prestare attenzione quando si estrapolano i risultati dagli studi sui ratti rapportandoli all’uomo, poiché studi recenti hanno rilevato che l’assorbimento di Cromo alimentare nell’uomo è significativamente maggiore rispetto a quanto avviene nei ratti per numerosi complessi di cromo testati.(54)[55]
Gli amminoacidi sembrano migliorare l’assorbimento del Cromo alimentare poiché formano complessi che migliorano l’assorbimento riducendo la tendenza del Cromo a precipitare nel liquido intestinale alcalino.(1) L’assorbimento del Cromo negli esseri umani è anche significativamente aumentato in presenza di Acido Ascorbico e Acido Nicotinico.(1)
Nei diabetici di tipo II, un integrazione giornaliera di Cromo (come cromo Picolinato) pari a 1.000mcg è risultata sufficiente a portare i livelli del minerale a digiuno nel siero da 2,40 ± 0,19 vs 0,16 ± 0,05ng/dL al basale dopo 12 settimane e 2,62 ± 0,09ng/ dL vs 0,17 +/- 0,04ng /dL al basale dopo l’integrazione di 24 settimane.(55)
Transferrina
La transferrina è una proteina di trasporto del siero nota per legarsi ai minerali (in particolare il Ferro). È stato notato che presenta affinità per il Cromo trivalente.(56) Per ogni molecola di transferrina si legano due ioni di cromo. (57)(58) Si pensa che la transferrina doni il Cromo all’oligopeptide Cromodulina.(59) Anche se studi precedenti hanno suggerito che la Cromodulina dona il Cromo alla transferrina. Questo lavoro però è stato condotto a temperature più elevate, che potrebbero aver causato la degradazione della Cromodulina.(60) Tuttavia, studi più recenti hanno dimostrato che la Cromodulina non rilascia il Cromo alla transferrina. (59) Poiché la transferrina rilascia ioni all’interno di una cellula dopo l’endocitosi (61), sembra che la Cromodulina accetti e trattenga questi ioni dalla transferrina.
La supplementazione con Cromo determina un aumento dell’eliminazione urinaria del minerale.(55)
Cromo Cloride
Uno studio svolto su ratti ha osservato che i livelli tossici di Cromo (100mcg/kg assunto con il cibo) sembrano bioaccumularsi di più con il Cromo Cloruro rispetto al Cromo Picolinato, in parte dipendente da un più alto tasso di escrezione osservato con il Picolinato.(62) Ciò è stato ipotizzato essere dovuto all’Acido Picolinico, che è stato osservato aumentare l’eliminazione di minerali come lo Zinco.(63)
Diversi studi hanno suggerito che la supplementazione con Cromo può promuovere una riduzione dell’appetito, con conseguente diminuzione del consumo di cibo, sia negli animali che negli esseri umani. Una recente meta-analisi di 10 studi randomizzati, in doppio cieco, controllati con placebo ha concluso che il Cromo Picolinato ha un effetto sulla riduzione del peso relativamente modesto, ma significativo rispetto al placebo (64), suggerendo un possibile effetto sulla soppressione dell’appetito. I meccanismi associati all’effetto anoressizzante dato dall’uso del Cromo sono attualmente sconosciuti, sebbene sia stato ipotizzato che si verifichi attraverso l’azione di specifici neurotrasmettitori nel cervello deputati al controllano l’appetito e il comportamento alimentare. (65)(66)
Ciò è stato confermato in un recente studio condotto su donne in sovrappeso adulte che hanno riportato voglie di carboidrati intense (almeno due volte a settimana). La supplementazione giornaliera con 1.000mcg di Cromo (come Picolinato) nel corso di otto settimane ha comportato una maggiore riduzione dell’assunzione di cibo (25%) rispetto al placebo (8%).(67) La riduzione dell’assunzione di cibo era associata ad una diminuzione della fame e dell’appetito, tuttavia la composizione dei macronutrienti non era influenzata e questi cambiamenti erano indipendenti da qualsiasi effetto sulla sensibilità all’insulina.(67) In uno studio parallelo condotto dallo stesso gruppo di ricerca, è stato riscontrato che la somministrazione periferica di Cromo nei ratti (tramite iniezione IP) ha comportato solo una modesta diminuzione dell’assunzione di cibo, rispetto a una significativa riduzione dose-dipendente dell’assunzione di cibo quando somministrato a livello centrale (direttamente nel cervello). (67) Nel suo insieme, questo lavoro suggerisce che, come detto pocanzi, la supplementazione con Cromo può promuovere una riduzione dell’apporto di cibo attraverso l’azione di neurotrasmettitori nel cervello che controllano l’appetito e il comportamento alimentare.
Nei pazienti con depressione atipica (che è un particolare sottogruppo di depressione associato a maggiore assunzione di cibo, sonnolenza e reattività dell’umore (68)), 600mcg di Cromo Picolinato per otto settimane non hanno influenzato significativamente la maggior parte dei sintomi depressivi. Tuttavia, ci sono stati significativi miglioramenti nella voglia di carboidrati e nell’assunzione di cibo con un effetto maggiore in coloro i quali il desiderio di carboidrati era maggiore al basale. (66) Nelle persone con disturbo da alimentazione incontrollata, il tasso di declino della frequenza di binging era maggiore con 1.000mcg di Cromo rispetto al placebo e 600mcg, sebbene la riduzione complessiva non abbia raggiunto la significatività statistica.(69)
La supplementazione con 1.000mcg di Cromo (come Picolinato) in due dosi suddivise per 24 settimane nei diabetici di tipo II non ha influenzato significativamente la gluconeogenesi epatica rispetto al placebo. (55) (La gluconeogenesi epatica è spesso patologicamente elevata nei diabetici (70)).
Quando i diabetici consumavano 200mcg di Cromo (come Cloruro) al giorno aggiunto a un prodotto di latte in polvere per 16 settimane, i livelli di glucosio e di insulina nel sangue erano significativamente ridotti mentre la sensibilità all’insulina migliorava. (71) I risultati di questo studio erano tuttavia specifici per genere, in quanto miglioramenti significativi nei suddetti marker dell’omeostasi del glucosio si sono verificati solo in soggetti di sesso maschile. (71)
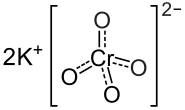
Cromato di Potassio
Come detto in precedenza, la Cromodulina è un oligopeptide endogeno (41) contenente Cromo che media positivamente la segnalazione del recettore dell’insulina in presenza di Insulina.(40) L’iniezione di Cromo (come Cromato di Potassio) nei ratti aumenta le concentrazioni urinarie e fecali di questo oligopeptide. (72) Tuttavia, la Cromodulina urinaria non sembra essere saturata in condizioni basali, il che implica che più Cromo potrebbe essere legato all’oligopeptide.(72) Poiché la potenza della Cromodulina nel potenziare la segnalazione dell’Insulina è correlata con la quantità di Cromo legata ad esso (43), e le iniezioni di Cromato di Potassio nei ratti determinano una rapida associazione con la Cromodulina (60)(73), è possibile che l’aumento di Cromo alimentare possa aumentare l’attività di questo oligopeptide.
Operando partendo dal presupposto che le assunzioni tipiche di Cromo nella dieta sono insufficienti per saturare la Cromodulina, la supplementazione con Cromo potrebbe teoricamente migliorare la segnalazione dell’Insulina tramite l’aumento del legame cromo-cromodulina.
Negli studi nei quali è stato utilizzato il Cromo trivalente, sembra esserci un aumento dell’attività della chinasi del recettore insulinico (in presenza di Insulina) quando il cromo 1-10μM viene aggiunto alla coltura di cellule di mammifero.(74) Questo aumento è indipendente da qualsiasi influenza diretta sulla fosforilazione o autofosforilazione (74) e distinta da quella della Cromodulina, che influenza l’autofosforilazione. (42)
Cromo Propionato
Alcuni complessi con Cromo trivalente hanno interazioni minori con il Recettore dell’Insulina, con complessi legati a piccole molecole endogene come Istidinato, Lattato, Acetato o Propionato che mostrano effetti inibitori minori a concentrazioni intorno a 100μM. Di questi complessi, il Cromo Propionato sembra essere il più potente, mostrando effetti inibitori a concentrazioni fino a 1μM. (75)
Lo stesso Cromo è stato implicato nel potenziare la segnalazione dell’Insulina, sebbene il meccanismo con gli ioni Cromo sembra differire da quello osservato con la Cromodulina e richiede una concentrazione significativamente più alta. Lo stesso Cromo non sembra influenzare direttamente il Recettore dell’Insulina come la lattina di Cromodulina.
La Fosfo-tirosin–fosfatasi 1B (PTP1B) è un regolatore negativo del segnale del Recettore dell’Insulina (76) che può essere soppresso dal Cromo endogeno. Anche se la Cromodulina è stata osservata promuovere l’attività della PTP della membrana in uno studio precedente (77), ci sono molti enzimi PTP endogeni e il PTP1B non è stato specificamente esaminato in questo studio. È stato dimostrato che il Cromo trivalente inibisce il PTP1B del 21-33% nelle cellule di epatoma umano e di ratto (78), suggerendo che il Cromo può potenziare la segnalazione dell’Insulina sopprimendo la defosforilazione mediata dal PTB1B nel Recettore dell’Insulina. Al contrario, uno studio più recente ha osservato che il Cromo non è riuscito a inibire l’attività della fosfatasi PTP1B umana ricombinante in un sistema in vitro puro, suggerendo che il Cromo può potenziare la segnalazione dell’Insulina da meccanismi distinti da qualsiasi effetto sul PTP1B. (74)
In uno studio in vivo, ratti obesi diabetici trattati con 80mcg/kg di Cromo (come Picolinato) hanno subito una diminuzione complessiva dell’attività della PTP1B e dell’espressione proteica correlata ad un aumento della segnalazione dell’Insulina nel muscolo scheletrico.(79) Questa diminuzione non è stata osservata nei ratti magri ai quali è stato somministrato il Cromo alla stessa dose.(79)
IRS-1
La fosforilazione dell’IRS-1, un importante trasduttore della segnalazione dell’Insulina che è inibito dalla fosforilazione a Serine307 (80), non è influenzato dal Cromo a 10μM in varie forme trivalenti.(75) Inoltre, l’espressione della proteina IRS è rimasta inalterata con una supplementazione di Cromo fino a 80mcg /kg nei ratti.(79) In assenza di Insulina, tuttavia, la segnalazione basale del IRS-1 è leggermente aumentata a 10 μM di Cromo, che si pensa sia dovuta alla diminuzione della fosforilazione del Serine307 (75) dal Jun NH (2) -terminal kinase (JNK). (37) Il JNK regola negativamente la segnalazione del IRS tramite fosforilazione a Serine307 (80) (81), che è aumentata nei topi obesi, (82) (83) causando insulino-resistenza. In particolare, l’attenuazione mediata dal JNK della segnalazione dell’Insulina nei ratti obesi è soppressa dal Cromo. (82)(83)
L’attivazione del JNK sopra riportata potrebbe essere ricondotta teoricamente allo stress del reticolo endoplasmatico (ER) (84), e gli agenti che riducono lo stress del ER attenuano anche i sintomi diabetici. (85)(86) È noto che lo stress del ER aumenta nelle cellule degli animali obesi e diabetici ed è curabile con il Cromo.(82)
Per riassumere i concetti esposti, sappiamo che l’interazioni del Cromo con il PTP1B, un regolatore negativo dell’attività del Recettore dell’Insulina, non sono ben compresi. Alcuni studi suggeriscono che il Cromo potrebbe non avere effetti apprezzabili sulla segnalazione del PTP1B. È possibile, tuttavia, che il Cromo sopprima l’attenuazione JNK-mediata della segnalazione dell’Insulina nel contesto di uno stato di insulino-resistenza preesistente.
Il Cromo non sembra aumentare l’espressione del Recettore dell’Insulina in presenza o assenza di Insulina, suggerendo che i suoi effetti sulla segnalazione dell’Insulina avvengono indipendentemente da eventuali cambiamenti nei livelli dei recettori insulinici. (87) (75) Inoltre, quando incubato con Insulina, il Cromo non influenza l’interazione dell’Insulina con il suo recettore.(74) Ciò suggerisce che il Cromo non influisce sulla sensibilità all’Insulina aumentando l’affinità del Recettore dell’Insulina.
Uno studio preliminare condotto nel 1992 ha rivelato che il Cromo aumenta l’internalizzazione dell’Insulina a 1μM (418ng/ml), effetto associato ad una maggiore fluidità della membrana e non replicato con altre chelazioni di Cromo o Zinco Picolinato. (88) La scoperta che l’Insulina è internalizzata nella cellula è stata successivamente rivelata come un importante meccanismo di feedback negativo per la segnalazione del Recettore dell’Insulina. Dopo che l’Insulina si lega con il suo recettore, il complesso del recettore insulinico viene internalizzato dall’endocitosi (89), innescando la degradazione dell’Insulina (90) e riducendo efficacemente il numero di recettori dell’insulina presenti sulla superficie cellulare come meccanismo per attenuare la risposta insulinica.(91)
In breve, dopo avvenuto legame con il suo recettore sulla superficie della cellula, l’Insulina innesca il movimento del complesso del Recettore dell’Insulina all’interno della cellula. Questo riduce il numero di recettori insulinici presenti sulla superficie cellulare e funziona come un meccanismo di feedback negativo per limitare la risposta della segnalazione insulinica.
In risposta a un test orale di tolleranza al glucosio, una supplementazione di 200mcg di Cromo per otto settimane non ha aumentato la risposta all’Insulina in soggetti diabetici di tipo II quando misurata dopo 10 minuti (71) mentre ad un dosaggio di 1.000mcg (come Picolinato) in soggetti non diabetici con sindrome metabolica per oltre 16 settimane ha aumentato la risposta all’Insulina nonostante non sia stato rilevato altro cambiamento nei biomarcatori del diabete. (92)
Uno studio ha osservato che, nonostante l’incapacità di trovare miglioramenti statisticamente significativi nella sensibilità all’Insulina per l’intero gruppo di soggetti presi in esame, il 46% degli individui che avevano un grado di insulino resistenza più elevato presentavano un miglioramento della sensibilità all’Insulina del 10%. (55) In particolare, non vi era alcuna differenza nell’assorbimento o cinetica del Cromo tra responder e non responder (55), suggerendo che la supplementazione con Cromo può aumentare la sensibilità all’Insulina in soggetti con insulino resistenza.
Inoltre, è stato osservato che una supplementazione di 1.000mcg di Cromo (come Picolinato) per 24 settimane in soggetti con diabete di tipo II riduce leggermente le concentrazioni di lipidi intramuscolari rispetto al placebo. (55) Poiché l’accumulo cronico di lipidi nel tessuto muscolare è una delle numerose cause patologiche dell’insulino-resistenza (93), anche questo lavoro suggerisce che la supplementazione con Cromo può aumentare la sensibilità all’Insulina in coloro che sono già insulino-resistenti.
Una meta-analisi di studi condotti su diabetici di tipo II trattati con >250mcg di Cromo per un periodo superiore ai tre mesi non ha rilevato alcuna influenza sul HbA1c rispetto al trattamento con placebo. (94) Ciò è in contrasto con precedenti revisioni che valutato solamente studi condotti su diabetici con un HbA1c basale superiore al 7%, in cui la supplementazione con Cromo ha determinato una riduzione dell’HbA1c dello 0,34% rispetto al placebo. (95) Altre revisioni hanno rilevato riduzioni dello 0,6% (96), e fino allo 0,9% quando sono state incluse tutte le forme di diabete e gradi di insulino resistenza.(97) Va notato, tuttavia, che alcune di queste analisi comprendevano prove della durata inferiore a tre mesi (96), che potrebbero non essere sufficienti per misurare i cambiamenti nel HbA1c. (94)
A seconda della popolazione studiata e del tipo e della qualità degli studi osservati, è dimostrato che il Cromo influisce in modo eterogeneo sui livelli di emoglobina A1C.
La supplementazione con 400 o 800mcg di Cromo (come Picolinato) insieme a un pasto di prova in adulti sani ha ridotto l’area del glucosio sotto la curva (AUC) del 30-36% nei responder, con la dose bassa più efficace.(97) In particolare, i responder sono stati classificati come soggetti aventi un consumo di carne e latte relativamente più basso (97), suggerendo che il Cromo può influenzare il metabolismo del glucosio postprandiale negli individui con livelli di Cromo basali inferiori. La riduzione del glucosio non è stata associata ad alcun cambiamento nell’Insulina, escludendo un effetto insulinogeno, e si è verificata in persone senza un metabolismo del glucosio alterato.(97)
Nella meta-analisi dove sono stati vagliati gli studi che valutavano la supplementazione di cromo oltre ai 250mcg nei diabetici di tipo II per un periodo di tre mesi (o più lungo)(94), i sette studi inclusi nella meta-analisi (55)(98)(99)(100)(101)(102)(103) non hanno mostrato una riduzione dei livelli di HbA1c nel siero nonostante una lieve riduzione della glicemia (RR di -0,95 e un IC 95% da -1,4 a -0,5).(94)
Un’analisi dei dati osservazionali del National Health and Nutrition Examination Survey (NHANES) ha rilevato che le persone che hanno consumato un integratore alimentare contenente Cromo avevano una probabilità inferiore di sviluppare il diabete (OR = 0,73), definito avendo un livello di HbA1c superiore a 6,5. L’uso di integratori in generale non ha avuto un effetto statisticamente significativo sulle probabilità di sviluppare diabete in questo studio.(104)
L’aggiunta di 400mcg di Cromo (come Picolinato) a una bevanda contenente carboidrati prima di un shuttle run test in uomini sani e attivi non ha modificato i benefici della bevanda contenente carboidrati rispetto al controllo, suggerendo che non vi è alcun beneficio aggiuntivo. (105)
Uno studio nel quale è stata somministrata una dose di 600mcg di Cromo (come Picolinato) ogni giorno per un mese prima di un esercizio di deplezione del glicogeno ha rilevato che immediatamente dopo l’esercizio e nell’ora successiva il gruppo trattato aveva livelli di lattato significativamente più alti rispetto al placebo. (106) In un altro studio che utilizzava un modello shuttle-run exercise, questo aumento di lattato non si è verificato con oltre 75 minuti di test a seguito del consumo di 400mcg di Cromo Picolinato o carboidrati o acqua (gruppo di controllo). (105) Inoltre, la concentrazione di lattato e il grado di fatica era simile in questo studio tra i due gruppi e il controllo. (105)
Glicogeno Sintasi
Il Glicogeno Sintasi è l’enzima responsabile della conversione del glucosio in glicogeno, la forma di deposito dei carboidrati nel corpo. Allo stesso modo, la fosforilasi di glicogeno è coinvolta nello scomporre queste riserve di carboidrati in glucosio per produrre energia. A causa dei suoi effetti sul metabolismo del glucosio, il Cromo è stato studiato per il suo impatto sulle riserve di glicogeno. Prove preliminari hanno rivelato che i ratti supplementati con Cromo avevano una minore dispersione del glicogeno epatico rispetto al gruppo di controllo durante il digiuno. (107) Successivamente, si è notato che il Cromo aumentava l’attività dell’enzima glicogeno sintasi nel muscolo e nel fegato dei ratti addestrati rispetto al gruppo di controllo non supplementato, ma la glicogeno fosforilasi non era influenzata.(108)
Negli adulti sovrappeso e leggermente allenati o sedentari la somministrazione di 600mcg di Cromo (come Picolinato) per un mese insieme ad una dieta standardizzata con gli ultimi due giorni progettati per esaurire il glicogeno, la supplementazione non ha modificato i livelli di glicogeno o il tasso di risintesi (da un carboidrato contenuto in una bevande) rispetto al placebo.(106)
Secondo la meta-analisi sul peso nei diabetici di tipo II supplementati con Cromo (oltre 250mcg) per oltre tre mesi, non vi è stata alcuna alterazione significativa del peso rispetto al placebo, nonostante una modesta riduzione del glucosio nel sangue.(94) Al contrario, un’altra meta-analisi ha rilevato che gli adulti sovrappeso e obesi che hanno integrato con il cromo picolinato hanno ridotto il peso corporeo nell’intervallo di dosaggio di 200-1.000 μg, indipendentemente dallo stato diabetico. La perdita di peso è stata tuttavia molto modesta, per un totale di soli 1,1 kg (IC del 95% nell’intervallo 0,4-1,7 kg).(109) Da notare, quest’ultima meta-analisi ha ritenuto la qualità delle prove non ottimale, mettendo in discussione gli effetti mediati dal cromo sulla perdita di peso.(109)
Uno studio ha rilevato che l’aumento di peso associato alla terapia con Sulfonilurea (0,9 kg su 10 mesi) nei diabetici è stato mitigato dalla cosomministrazione di 1.000mcg di Cromo. (102) È importante sottolineare che questi risultati possono essere limitati a coloro che sono sottoposti a terapia Sulfonilurea. Quando i soggetti diabetici che non erano stati trattati con il medicinale ricevevano istruzioni per seguire una dieta di mantenimento del peso, la supplementazione di 1.000mcg di Cromo Picolinato non modificavano l’assunzione di cibo, l’appetito o il peso corporeo. (55)
Il Cromo è stato anche usato nel tentativo di mitigare l’aumento di peso associato alla cessazione del fumo, poiché le persone che smettono di fumare spesso tendono ad aumentare di peso.(110) Questo studio ha utilizzato l’Hypericum perforatum (900 mg) come primo aiuto anti-fumo e poi ha diviso i soggetti dello studio in gruppi trattati con Cromo o placebo. Sfortunatamente, la tendenza del Cromo ad attenuare l’aumento di peso non ha potuto essere testata con sufficiente potenza, a causa dei bassi tassi di successo con l’erba di cui sopra. Tuttavia, gli effetti del Cromo erano promettenti, con una probabilità di attenuare l’aumento di peso da 5,76 kg a 2,7kg dopo sei mesi.(111)
Il Cromo può anche ridurre la perossidazione lipidica in alcune popolazioni. Sono necessari però ulteriori studi per determinarne la dose appropriata e chi potrebbe trarne reale beneficio.
Come accennato in precedenza, Il Cromo Picolinato, più di altre forme trivalenti di Cromo, ha la capacità di formare proossidanti che possono potenzialmente causare danni al DNA. La rilevanza per la supplementazione orale standard non è nota, poiché la concentrazione richiesta per danneggiare il DNA (livello alto di 50μM) è significativamente più alta di quella osservata nel sangue dopo l’ingestione orale di integratori. Inoltre, gli studi su soggetti umani non hanno notato danni al DNA con dosi supplementari standard (200-400mcg/die).
Il Cromo si accumula nei testicoli dei ratti quando iniettato, anche se i possibili benefici o danni nei testicoli con integrazione orale di Cromo non sono stati studiati. Il Cromo esavalente, la forma tossica non presente negli integratori, è noto per essere tossico per i testicoli.
Esistono diverse altre azioni potenziali legate all’uso del Cromo che sono state scientificamente documentate. Per ovvie ragioni, legate soprattutto all’argomento principale trattato in questa serie di articoli, ho omesso diversi studi di un certo interesse. Per chiunque volesse approfondire cliccare qui.
In seguito alle numerose informazioni riportate nel presente articolo, possiamo con una certa sicurezza concludere che una supplementazione di Cromo risulta maggiormente incisiva in caso di carenza del minerale a causa di una insufficiente assunzione con gli alimenti o in condizioni di insulino resistenza (sia “pre-diabetica” che nella condizione diabetica). Il potenziale anoressizzante del Cromo rappresenta sicuramente un elemento di vantaggio durante una dieta ipocalorico (specie se ipoglucidica). Il Cromo sembrerebbe avere anche una certa azione sul miglioramento dello stoccaggio del glicogeno e sulla sua preservazione, anche se la cosa, in realtà, non è mai stata riscontrata nell’uomo. Se tale azione fosse possibile o significativa, si potrebbe riflettere positivamente a livello prestativo ma che in un contesto di “scarico del glicogeno” potrebbe risultare limitante.
A questo punto la domanda che si ripresenta è “come si possono utilizzare queste informazioni per pianificare l’uso del Cromo”?
Vista la sua efficacia in soggetti patologici e/o in condizioni di insulino resistenza non patologica, l’uso temporalmente ridotto di 1mg (stand alone) o protratto di 400-600mcg (in combinazione con altri GDA; vedi possibile azione additiva con la Berberina e ALA) di Cromo Picolinato al giorno può portare a dei vantaggi in quei soggetti con una insulino-resistenza di base genetica, cioè individui con una tolleranza glucidica limitata rispetto alla media, o durante regimi ipercalorici (vedi peggioramento dell’insulino-resistenza durante regimi ipercalorici).
Dosi contenute di Cromo Picolinato (200-400mcg/die) assunte durante periodi ipocalorici possono aiutare il soggetto trattato a tollerare la riduzione calorica per via dell’effetto anoressizzante dato dall’uso di questo composto.
In combinazione con altri GDA durante e nel periodo successivo (periodo “protocollare” di 4 settimane) all’uso di Insulina esogena.
Alcuni effetti collaterali comuni riscontrati con l’uso del Cromo Picolinato (dose correlato) possono includere insonnia, cambiamenti di umore, irritabilità e mal di testa.
Altri effetti collaterali possibilmente riscontrabili con l’uso di alte dosi di Cromo Picolinato includono problemi di coordinamento o di equilibrio, problemi di concentrazione o difficoltà di pensiero, e sintomi legati a problemi epatici (che comprendono: nausea; mal di stomaco nella zona superiore; prurito; stanchezza; perdita di appetito; urina di colore scuro; ingiallimento della pelle o degli occhi (ittero)).
Chiedere prontamente assistenza medica di emergenza se si verificano segni di anafilassi, una reazione allergica grave che può includere orticaria, difficoltà di respirazione o gonfiore del viso, delle labbra, della lingua o della gola.
Esiste un caso studio di una donna che in seguito all’ingestione di 1200-2400mcg di Cromo (come Picolinato) per 4-5 mesi mostrava sintomi di danno renale. (112) In un altro caso studio, un Bodybuilder aveva sviluppato rabdomiolisi associata all’assunzione di 1.200mcg di Cromo Picolinato per due giorni.(113)
La possibile comparsa di questi effetti avversi può essere evitata con una attenta calibrazione della dose giornaliera di Cromo. Se ne sconsiglia quindi un assunzione superiore a 1mg/die (dose quest’ultima comunque relegabile a periodi d’uso brevi). Una supplementazione giornaliera di 200-400mcg di Cromo Picolinato è generalmente ben tollerata con una punta massima di dosaggio di 600mcg/die.
Per ottenere una migliore biodisponibilità del composto, il Cromo andrebbe assunto lontano dalla somministrazione di integratori di Zinco o con pasti contenenti fonti ricche di fitati (vedi cereali integrali e legumi; il cui contenuto di fitati può comunque essere ridotto con, ad esempio, l’ammollo e la adeguata cottura).
Come detto la volta scorsa per l’Acido Alfa Lipoico, anche con l’uso di Cromo Picolinato (o altra forma) è essenziale la cura del dosaggio e la ponderatezza nell’utilizzo.
Con il termine GDA (Glucose Disposal Agent) ci si riferisce ad una serie di composti (non farmaci) aventi la capacità di ottimizzare il metabolismo glucidico cellulare agendo sul miglioramento del insulino-resistenza. Questi integratori alimentari stanno avendo un largo consumo nel BodyBuilding e nel Fitness in generale data la, ormai, conosciuta importanza che la sensibilità all’insulina ha sulla composizione corporea e sulla salute generale. I GDA, quindi, agendo sul miglioramento della sensibilità all’Insulina contribuiscono ad una migliore ripartizione calorica che si traduce in un maggiore trasporto dei nutrienti verso il miocita (cellula muscolare) rispetto all’adipocita. Lo scopo di questa serie di articoli è, quindi, sostanzialmente quello di analizzare nel dettaglio i principali GDA valutandone il potenziale attraverso ciò che la letteratura scientifica ad oggi disponibile riporta analizzandone le possibili migliori applicazioni.
Principali GDA e loro caratteristiche
Berberina: caratteristiche e possibili applicazioni.
Berberina
La Berberina è un alcaloide, più precisamente un sale di ammonio quaternario appartenente al gruppo protoberberino degli alcaloidi benzilisochinolinici trovati in piante del genere Berberis (ad es. Berberis vulgaris – crespino, Berberis aristata – albero curcuma, Mahonia aquifolium – uva-Oregon, Hydrastis canadensis – goldenseal, Xanthorhiza simplicissima – yellowroot, Phellodendron amurense (1) – Albero di sughero Amur, Coptis chinensis – Goldthread cinese, Tinospora cordifolia, Argemone mexicana – papavero spinoso ed Eschscholzia californica – Papavero californiano). La Berberina si trova solitamente nelle radici, nei rizomi, nei gambi e nella corteccia delle piante precedentemente elencate.
Come accennato pocanzi, la Berberina è un alcaloide estratto da diverse piante utilizzate nella medicina tradizionale cinese.
Methoxyhydnocarpina
La Berberina è attualmente in fase di studio al fine di determinarne l’efficacia d’uso per il trattamento del aritmia, del diabete, (2) del iperlipidemia, (3) del infiammazione (4) e del cancro. La Berberina esplica azione antiaritmica di classe III.(5) Vi sono alcune prove che la Berberina possa avere proprietà Anti-Aging (gero-soppressive).(6)(7) Alcune ricerche sono state condotte su un possibile uso contro l’infezione da Staphylococcus aureus (MRSA) resistente alla meticillina.(8) La Berberina è considerata un antibiotico.(9)(10) Quando applicata in vitro e in combinazione con Methoxyhydnocarpina, un inibitore di pompe a resistenza multipla, la Berberina inibisce la crescita dello Staphylococcus aureus (11) e Microcystis aeruginosa (12), un cianobatterio tossico. La Berberina è utilizzata principalmente per i suoi effetti anti-infiammatori e anti-diabetici. Può anche migliorare la salute intestinale e abbassare il colesterolo. La Berberina è in grado di ridurre la gluconeogenesi epatica. La ricerca svolta su esseri umani e animali ha dimostrato che un dosaggio di Berberina pari a 1500mg/die, assunti in tre dosi eguali distribuite nell’arco della giornata, ha la medesima efficacia riscontrata con lo stesso dosaggio di Metformina o con 4mg di Glibenclamide, due farmaci per il trattamento del diabete di tipo II. L’efficacia è stata misurata attraverso l’impatto dei composti sulla riduzione dei biomarcatori del diabete di tipo II.
La Berberina sembra avere anche un effetto sinergico con i farmaci antidepressivi e coadiuvare la perdita del grasso corporeo. Entrambi questi benefici hanno bisogno di ulteriori prove a loro sostegno prima che la Berberina possa essere raccomandata per l’uso in tali circostanze.
Il principale meccanismo responsabile degli effetti anti-infiammatori e anti-diabetici della Berberina è rappresentato dalla sua capacità di attivare un enzima chiamato proteina kinasi dipendente dall’adenosina monofosfato (Adenosine Monophospate-activated Protein Kinase o AMPK) inibendo al contempo la proteina-tirosina fosfatasi 1B (PTP1B).
Nelle cellule vive, la Berberina si localizza nei mitocondri. La sua localizzazione mitocondriale è coerente con l’inibizione del complesso I della catena respiratoria, la diminuzione della produzione di ATP e la successiva attivazione del AMPK, che porta alla soppressione della segnalazione dell’mTOR.(6) Una caratteristica quest’ultima non positiva, specie in soggetti “Natural”, ma con gravità d’effetto dose dipendente e non significativa, in base ai dati aneddotici, ai dosaggi medi utilizzati. La biodisponibilità della Berberina è bassa.(13)
La Berberina sembra sopprimere debolmente il consumo di glucosio in acuto, (14) con 72 ore di incubazione che sopprimono l’assorbimento del glucosio ad un grado statisticamente insignificante in vitro.(15)
Acarbosio
L’enzima saccarasi è inibito in modo concentrazione-dipendente con un IC50 di 1,83 mg/L (potenza piuttosto bassa) (14) e di 0,28 mg/ml.(16) Il complesso dell’enzima saccarosio-isomaltasi (SI) sembra avere un mRNA aumentato nella condizione diabetica. Questo aumento di espressione dell’enzima viene ridotto (fino al 62% rispetto a quello dei ratti di controllo) con una dose di 100-200 mg/kg di Berberina per 35 giorni, effetto riscontrato anche nei ratti non diabetici.(17) In un altro studio nel quale è stata utilizzata una dose di Berberina pari a 125 mg/kg per 33 giorni è stato osservato che, in risposta ad un test di tolleranza al saccarosio assunto oralmente, il composto ha prodotto il 43% in meno nella AUC del glucosio serico (meno efficace del Acarbosio a 20mg/kg come controllo attivo) correlato con una minore attività del sucrasi in tutte le parti dell’intestino.(16) In un altro studio, una dose di 100mg/Kg di Berberina ha dimostrato di avere effetti analoghi. (18)
L’enzima maltasi sembra essere inibito ma non in modo dose-dipendente, (14) con uno studio che mostra le alterazioni dell’attività del maltasi nei ratti diabetici (aumento di 1,45-2,56 volte) normalizzate con 35 giorni di integrazione di Berberina (100-200 mg / kg) (17) con una normalizzazione significativa in tutte le aree dell’intestino dopo la somministrazione di 125mg /kg per 33 giorni. (16) Non sembra esserci alcuna inibizione diretta degli enzimi attivi del maltasi fino a 50μM di Berberina (sebbene contestato, un altro studio suggerisce un IC50 di 0,11 mg / ml (16)), ma 5 giorni di esposizione a questa concentrazione riducono l’attività del 48% in vitro.(17)
L’enzima lattasi (media la digestione del lattosio) è risultato essere aumentato nella condizione diabetica e attenuato, ma non normalizzato, in seguito all’ingestione di 125mg/kg di Berberina per 33 giorni nei ratti.(16)
Per quanto riguarda l’enzima alfa-amilasi (media la digestione dell’amido), la Berberina è stata testata in uno studio in vitro per l’inibizione della crescita dipendente dalla amilasi fungina con valori di Ki simili all’Acido Clorogenico e all’Acido Caffeico suggerendo una sua azione inibitoria non competitiva dell’enzima stesso. (19)
La sottoregolazione dell’attività enzimatica (complesso maltasi e SI) sembra essere in parte dipendente dalla PKA e l’inibizione della PKA con l’inibitore H89 attenua (ma non inibisce totalmente) questi effetti.(17)
L’effetto ipoglicemizzante della Berberina fu scoperto per la prima volta nel 1988 quando una condizione ipoglicemica venne osservata accidentalmente nei pazienti diabetici trattati con Berberina per i suoi effetti anti-diarroici.(20)
Una meta-analisi è stata condotta sulla Berberina al fine di valutarne le potenzialità nel trattamento del diabete di tipo II.(21) Questa meta-analisi ha rilevato 14 studi (tutti provenienti dalla Cina), che includevano l’osservazione di 1068 pazienti tra il 2007 ed il 2011, nei quali si è osservato che l’assunzione di Berberina ad un dosaggio di 0,5-1,5 g al giorno, abbinata ad un intervento correttivo dello stile di vita del paziente, per 12 settimane era associata a miglioramenti della glicemia a digiuno (0,87 mmol / Riduzione L, CI 0,54-1,20), postprandiale (riduzione di 1,72 mmol / L, CI 1,11-2,32) e del HbA1c (riduzione dello 0,72%; CI 0,47-0,97%) con miglioramenti del metabolismo lipidico e la riduzione dei livelli di Insulina a digiuno ( 0,5 mU / L; CI 0,03-0,96).(21)
Metformina
Sette studi (su 448 pazienti) hanno utilizzato una valutazione comparativa con gli agenti ipoglicemizzanti orali e, sebbene non sia possibile eseguire una meta-analisi a causa dell’eterogeneità dei dati, non sembrano esserci differenze significative quando la Berberina è stata confrontata con Metformina, Glipizide, o Rosiglitazone. (21) In 4 studi su 6 che hanno usato la Berberina come trattamento adiuvante insieme ai farmaci ipoglicemizzanti orali, i benefici additivi sono risultati significativi per la glicemia a digiuno (riduzione di 0,59 mmoli / L, CI 0,35-0,83), postprandiale (riduzione di 1,05 mmoli / L, CI 0,48-1,62 ) e l’HbA1c (riduzione dello 0,53%, CI 0,11-0,95%) che sono stati osservati ridursi maggiormente nella terapia combinata rispetto all’uso dei soli farmaci ipoglicemizzanti orali.(21)
La metodologia degli studi inclusi è ritenuta scadente (punteggio Jadad minore di 3) ma non sembra esserci il rischio di bias come valutato mediante il grafico a imbuto (anche se in 10 studi utilizzati il grafico a imbuto non può essere stato accurato in modo desiderabile (22)).(21) Questa meta-analisi ha escluso tre studi (nessuno dei quali è indicizzato online) a causa delle differenze al basale o dell’incertezza nella randomizzazione.(21)
Un altro studio sulla Berberina ha mostrato che l’assunzione di 0,3g tre volte al giorno (900mg in totale) per 12 settimane su 37 soggetti con sindrome metabolica di nuova diagnosi causano una riduzioni significativa della glicemia (17%), del HbA1c (15%), dell’Insulina a digiuno (26%), e della sensibilità all’insulina valutata con l’indice HOMA-R (41%).(23) I soggetti con diabete di tipo II che avevano assunto 1g di Berberina per un mese hanno sperimentato una riduzione del 20% e del 26% rispettivamente del glucosio ematico a digiuno e post-prandiale insieme a una riduzione del 12% del HbA1c, ma solamente con una lieve tendenza al miglioramento della sensibilità all’insulina.(24) L’assunzione di 1g di Berberina per 2 mesi ha ridotto la glicemia a digiuno (25,9%), l’HbA1c (18,1%) e i Trigliceridi (17,6%).(25) Un altro studio sugli esseri umani con steatosi epatica non alcolica a cui sono stati applicati solo interventi sullo stile di vita o interventi sullo stile di vita più 15 mg di Pioglitazone al giorno o 0,5g di Berberina al giorno per 16 settimane non ha riscontrato differenze tra i 3 gruppi sul HbA1C, ma ha osservato un miglioramento nel punteggio del HOMA-IR nel gruppo che ha subito interventi sullo stile di vita insieme all’inserimento della Berberina rispetto agli interventi sullo stile di vita senza supplementazione, con nessuna differenza rispetto agli interventi sullo stile di vita in combinazione con la somministrazione di Pioglitazone.(26) L’area sotto la curva glicemica dopo un test di tolleranza al glucosio orale è stata ridotta anche nel gruppo trattato con Berberina rispetto ai soli interventi sullo stile di vita (con ancora nessuna differenza rispetto al Pioglitazone), principalmente a causa di una maggiore riduzione del glucosio a 120 e 180 minuti.(27) Miglioramenti simili sono stati riscontrati in soggetti con la sindrome metabolica, in cui 0,5g di Berberina tre volte al giorno per tre mesi hanno portato ad un miglioramento della sensibilità all’Insulina come misurato dagli indici insulinogenici e di Matsuda, dalla glicemia della AUC del glucosio e della AUC dell’Insulina rispetto al placebo.(27)
Rosiglitazone
Studi comparativi sull’uso della Berberina hanno mostrato che l’assunzione di 1g al giorno dell’alcaloide presenta la medesima efficacia nel miglioramento dei parametri misurati (solitamente glicemia a digiuno, Insulina, HbA1c e Trigliceridi) riscontrata con la Metformina (28) (25) e il Rosiglitazone (25) quando utilizzati entro il range di dosaggio standard di 1,5g (Metformina) o 4mg (Rosiglitazone).
Adiponectina
La Berberina ha mostrato di poter aumentare i livelli di Adiponectina, una adipochina (molecola di segnalazione derivata da cellule adipose) che svolge un ruolo positivo nella sensibilità all’Insulina (viene secreta e quindi agisce sul tessuto attraverso i suoi recettori per attivare l’AMPK (29)) e che si riduce nei diabetici, in particolare nella struttura ad alta attività. (30) L’Adiponectina si trova in tre forme strutturali: trimero, esamero e ad alto peso molecolare. Quest’ultima è maggiormente correlata alla sensibilità all’Insulina (31); la Berberina (2-4uM) agisce attraverso l’attivazione del AMPK, in particolare il sottoinsieme AMPKα1, per aumentare la percentuale di Adiponectina nella sua struttura ad alta attività; un processo noto come multimerizzazione della Adiponectina. (32) Questo è stato anche notato con l’uso del AICAR, un noto farmaco sperimentale utilizzato per attivare l’AMPK, suggerendo un effetto generale che non è unico per la Berberina; (32) questo intricato ciclo (Attivazione dell’Adiponectina tramite l’AMPK che promuove l’Adiponectina ad alta attività) è un meccanismo di autoregolazione dell’Adiponectina. (33)
Va notato che gli studi sull’uso della Berberina in adipociti predifferenziati hanno rilevato una minore secrezione di Adiponectina che era la conseguenza naturale della soppressione della differenziazione.(23) Questo è stato anche replicato nello studio sopra menzionato sull’aumento della funzione dell’Adiponectina, con entrambi i fenomeni che si verificano a concentrazioni simili. (32)
La Berberina ha dimostrato di stimolare l’uptake del glucosio nel muscolo scheletrico (34) in modo parzialmente mediato dal AMPK.(35)
L’attivazione del AMPK può aumentare la biogenesi mitocondriale nelle cellule del muscolo scheletrico, cosa che è stata dimostrata in seguito ad assunzione di Berberina; è stata stabilita l’inattività della Berberina nelle cellule prive del SIRT-1 (un intermedio richiesto).(36)
2,4-thiazolidinedione
La Berberina è nota anche per migliorare l’uptake del glucosio nelle cellule adipose con una concentrazione di 25μm dimostratasi altrettanto potente di una concentrazione di 15uM di 2,4-thiazolidinedione (TZD, un farmaco antidiabetico, di 3,3 volte) e leggermente sovraperformato sia rispetto alla Arecolina (3,2 volte) che all’Acido Vanillico (2,9 volte), entrambi prodotti naturali. (37) Questo studio ha anche rilevato che Berberina ha agito sinergicamente sia con il TZD che con la Metformina. (37) La Berberina ha anche dimostrato di essere più efficace nell’aumentare l’assorbimento del glucosio rispetto ai polisaccaridi del Astragalus Membranaceus. (38)
Sebbene l’aumento dell’attività del AMPK dato dalla Berberina sia noto per migliorare l’uptake del glucosio negli adipociti, (39) la Berberina sembra agire indipendentemente dal AMPK per aumentare l’assorbimento del glucosio di 5 volte nelle cellule dei fibroblasti L929 che esprimono solo trasportatori GLUT1; si è scoperto che la Berberina aumenta l’attività dei GLUT1 (un trasportatore del glucosio normalmente poco attivo) tramite un parziale pathway del MAPK e ERK p38.(40) Questo aumento dell’attività dei GLUT1 è stato osservato negli adipociti 3T3-L1, sebbene sia classificato come secondario all’attivazione del AMPK.(41)
La Berberina può anche inibire l’enzima PTP1B e promuovere l’assorbimento del glucosio negli adipociti (e nei miociti) preservando l’attività dell’Insulina. A concentrazioni di Berberina pari a 1,25-2,5uM, la fosforilazione del Recettore dell’Insulina è aumentata senza alterare il contenuto proteico.(39) L’IC50 della Berberina sul PTP1B sembra essere di 156,9 nM con un valore Ki di 91,3 nM, un valore significativo. (42)
Si è scoperto che la Berberina normalizza parzialmente la diminuzione del consumo del glucosio indotto dal palmatato (un acido grasso), e ciò avviene attraverso gli effetti anti-infiammatori di inibizione dell’aumento dell’attività di IKKβ e NF-kB; che successivamente aumentano l’IRS-1 e riducono l’assorbimento del glucosio attraverso il Recettore dell’Insulina.(43) Questo effetto antinfiammatorio è stato notato anche quando la misurazione delle citochine (44), e della resistenza all’insulina indotta dagli acidi grassi è stata replicata in altre cellule in relazione al NF-kB.(45)
Lo studio svolto prendendo in esame persone con sindrome metabolica ha rilevato che l’assunzione di 300mg di Berberina tre volte al giorno (900 mg totali) per un totale di 12 settimane è associata ad una significativa riduzione del BMI da 31.5 +/- 3.6 a 27.4 +/- 2.4 (diminuzione media del 13%) con una diminuzione significativa della circonferenza vita del 5,5%; la massa magra e la massa grassa non sono state misurate. (23) Persone sane ma sovrappeso che assumono 500mg di Berberina per tre volte al giorno (1500 mg in totale) per 12 settimane senza aggiustamenti nell’esercizio fisico hanno notato una riduzione del peso corporeo di circa 2,26Kg (2,3% peso corporeo, 3,6% grasso corporeo); l’assunzione di cibo non è stata modificata nel complesso, ma due soggetti hanno riportato una diminuzione dell’appetito. (46)
Pioglitazone
Uno studio svolto su esseri umani con steatosi epatica non alcolica a cui sono stati applicati solo interventi sullo stile di vita o interventi sullo stile di vita più 15mg di Pioglitazone al giorno o 0,5g di Berberina al giorno per 16 settimane hanno riscontrato una riduzione significativa del BMI di 1,51 nel gruppo con aggiunta di Berberina rispetto alla riduzione di 0,72 in BMI riscontrata nel gruppo di solo interventi sullo stile di vita; il Pioglitazone ha causato una riduzione simile ai soli interventi sullo stile di vita.(47)
Per quanto riguarda la gluconeogensi epatica (la produzione di glucosio da fonti non glucidiche, la quale tende ad essere drasticamente aumentata nello stato di diabete contribuendo all’aumento del glucosio ematico a digiuno), la somministrazione di Berberina ad alte dosi (380mg/kg al giorno) per 5 settimane nei ratti diabetici è in grado di ridurre l’attività dei due enzimi implicati nella limitazione della velocità della gluconeogenesi epatica (PEPCK e G6Pase), senza che vi sia un aumento correlato dell’Insulina (normalmente soppressiva di questi due geni). (48) Questo sembra essere correlato alla normalizzazione dell’attività del FOXO1, che è stata aumentata nei ratti diabetici, (48) e alla riduzione del contenuto proteico e dell’mRNA (trascrizione genomica). (49)
Recettore dell’Insulina
Nelle cellule epatiche umane (studio in vitro), 10mcg/mL in 12 ore possono causare un aumento del contenuto di Recettori dell’Insulina negli epatociti; questo si estende anche ad altre linee cellulari testate (pancreas, colon, linfociti, fibroblasti) tra un aumento di 1,6 e 1,9 volte (25) e sembra funzionare sia in modo dose-dipendente che e dose-indipendente. (50) Anche la segnalazione dell’insulina è stata migliorata in modo secondario con la riduzione dello stress del reticolo endoplasmatico, (51) che è secondario al Recettore dell’Insulina.
Quando testato in cellule CEM, la coincubazione di 0,5nM di Insulina con 10mcg/mL di Berberina induce l’attivazione del Akt allo stesso grado dell’Insulina a 10nM. (25)
Uno studio sui ratti ha evidenziato che la Berberina è associata ad un aumento della secrezione di GLP-1 misurata sia nel siero che nell’intestino, in seguito all’aumento dell’mRNA del proglucagone nell’intestino; questi effetti si aggiungono alle qualità anti-diabetiche della Berberina. (52)
L’inibizione del PTP1B, che promuove la segnalazione del Recettore dell’Insulina con un IC50 di 156.9nM, (42) sembra anche avvenire nelle cellule muscolari. (39)
La Berberina sembra indurre l’assorbimento del glucosio nelle cellule muscolari di per sé indipendentemente dal fatto che la cellula sia in uno stato di aggravata insulino-resistenza (53) o di ottimale insulino-sensibile (54), e l’interazione sinergica tra Insulina e Berberina esiste solo quando la cellula muscolare è in una condizione di insulino-resistenza, mentre in presenza di cellule insulino-sensibili l’interazione è appena additiva (effetti additivi non statisticamente significativi, sembra esserci un crossover nei meccanismi). (53) (54)
La sovraregolazione del numero dei recettori insulinici (quantità di recettori insulinici espressi sulla superficie cellulare) sembra estendersi ai miociti diìel ratto L6 a 7,5 ÷ 2,5 volte quella del gruppo di controllo, con benefici significativi ma minori rilevati a 2,5uM.(50) Ciò era dovuto all’aumento della trascrizione del recettore a livello genomico, ed è dipendente dalla PKC, che la Berberina sembra attivare in modo dose-dipendente.
A causa delle interazioni con il CYP3A4 (inibizione attraverso cui può aumentare il Testosterone) e il CYP1A2 (Aromatasi), (55) in teoria la Berberina può aumentare i livelli circolanti di Testosterone; questo, però, non è ancora stato testato in sistemi viventi.
L’incubazione di Tamoxifene (1,5uM) e Berberina (16ug / mL) nelle cellule del cancro al seno MCF-7 estrogeno-sensibili è in grado di aumentarne sinergicamente l’apoptosi. (56) Questo sinergismo sembra essere correlato ai SERM in generale, sebbene il meccanismo(i) esercitato dalla Berberina non sia attualmente noto. (56)
L’intervento con Berberina su soggetti con sindrome metabolica di nuova diagnosi ad un dosaggio di 300mg assunti tre volte al giorno (900 mg totali) per 12 settimane è stato in grado di ridurre i livelli circolanti di Leptina del 36% mentre aumentava significativamente l’Adiponectina, ma il rapporto Leptina/Adiponectina è migliorato passando da 0,76 a 0,58.(23)
GLP-1
Il GLP-1 (glucagon-like peptide-1) è un ormone peptidico noto per essere secreto dall’intestino con proprietà ipoglicemizzanti, esplicate in parte attraverso la stimolazione della secrezione dell’Insulina (57)(58) e può anche avere un ruolo nella proliferazione delle cellule β del pancreas. (59) È stato osservato che la Berberina, alla dose orale di 120mg/kg per 5 settimane, aumenta le concentrazioni di GLP-1 e di Insulina nei ratti con diabete indotto da streptozotocina (misurazioni eseguite dopo somministrazione postprandiale). (60) Questo studio ha anche rilevato un aumento della popolazione delle cellule β (460% nel controllo diabetico, ma ancora meno della metà nel controllo non diabetico) che è stato attribuito all’azione del GLP-1.(60)
Le azioni della Berberina scientificamente documentate sono svariate e non implicano soltanto le interazioni di questo composto con il metabolismo glucidico. Per motivi di praticità, e per motivi legati all’argomento principale del presente articolo, ho dovuto omettere alcuni studi di un certo interesse. Per chiunque volesse approfondire cliccate qui.
Adesso sappiamo che la Berberina può migliorare la sensibilità all’Insulina in modo parzialmente dipendente dall’attivazione del AMPK, con conseguente aumento del uptake del glucosio sia da parte del miocita che del adipocita (sebbene, in questo ultimo caso, la cosa non è negativa come sembra dal momento che un ottimizzazione del metabolismo glucidico adipocitario porta ad un miglioramento della secrezione di Leptina con i benefici ad essa correlati), e che può ridurre l’assorbimento glucidico intestinale e ridurre i tassi di gluconeogenesi epatica. Sappiamo anche che l’impatto della Berberina in sinergia con l’Insulina risulta non significativo in condizioni di un ottimale insulino-sensibilità. A questo punto la domanda è “come si possono utilizzare queste informazioni per pianificare l’uso della Berberina”? La risposta è, ovviamente, dipendente dal vantaggio che le suddette caratteristiche possono apportare in una data fase e su determinati soggetti.
Soggetti con una insulino-resistenza di base genetica, e cioè individui con una tolleranza glucidica limitata rispetto alla media, possono trarre vantaggi dall’uso della Berberina sia in contesti di alimentazione ipercalorica (con un aumento del carico glucidico) che ipocalorica, sfruttando, nel primo caso, l’effetto sull’insulino-sensibilità dato anche dalla sinergia Berberina/Insulina in presenza di cellule con una condizione di insulino-resistenza (condizione peggiorata già di per se da una dieta ipercalorica) e, nel secondo caso, mantenendo una insulino-sensibilità migliorata rispetto al basale con una parallela riduzione dell’assorbimento glucidico intestinale e della gluconeogenesi epatica che risultano di particolare vantaggio durante diete ipocaloriche “low carb” nonché in regimi alimentari chetogenici. L’impatto della Berberina sull’attività mitocondriale nel muscolo-scheletrico non è di certo da trascurare specie in tali contesti.
Soggetti con una buona affinità con il glucosio possono trarre vantaggi dall’uso della Berberina durante fasi ipercaloriche, mantenendo una discreta sensibilità all’Insulina con una, e già citata, sinergia additiva tra la Berberina e l’Insulina in situazioni di insulino-resistenza cellulare (indotta e/o peggiorata dalla dieta ipercalorica). L’alterazione dell’assorbimento glucidico potrebbe dare alcuni problemi se si sta cercando di aumentare la massa magra dal momento che questo meccanismo riduce l’apporto calorico. E’ vero anche, però, che il miglioramento della ripartizione calorica dato dall’uso della Berberina aumenta l’efficienza dei nutrienti assunti il che, di per se, compensa l’eventuale riduzione dell’assorbimento glucidico intestinale. Questo effetto, invece, risulta vantaggioso, parallelamente all’applicazione della Berberina in soggetti con insulino-resistenza genetica basale, durante fasi ipocaloriche. L’uso durante “refeed” dovrebbe essere evitato per le possibili “complicazioni” relative all’assorbimento glucidico (tranne nei casi di insulino-resistenza genetica basale).
L’uso della Berberina durante e dopo protocolli d’uso dell’Insulina esogena permette di utilizzare dosaggi del peptide inferiori con il mantenimento di una discreta insulino-sensibilità (durante) con una azione “rigenerativa” a livello pancreatico (dopo).
I dosaggi che hanno dimostrato un impatto statisticamente significativo, anche alla luce della ricerca scientifica svolta fino ad oggi, vanno dai 500mg a 1.5g al giorno assunti in 2-3 somministrazioni di uguale portata distribuite durante la giornata (preferibilmente prima dei pasti). L’emivita della Berberina è stata stimata essere di circa 5-6 ore.(61)
Ai dosaggi comunemente usati nella medicina tradizionale e nel limite dei 1.500mg/die la Berberina è ben tollerata e sicura; a dosaggi più alti può determinare: disturbi gastrointestinali, dispnea, diminuzione pressoria, sintomi simil-influenzali e danno cardiaco. (62)
Fine 1° Parte…
Gabriel Bellizzi
Rifermenti:
1- Zhang Q, Cai L, Zhong G, Luo W (2010). “Simultaneous determination of jatrorrhizine, palmatine, berberine, and obacunone in Phellodendri Amurensis Cortex by RP-HPLC”. Zhongguo Zhong yao za zhi = Zhongguo zhongyao zazhi = China journal of Chinese materia medica. 35 (16): 2061–4. doi:10.4268/cjcmm20101603. PMID 21046728.
2- Dong H, Wang N, Zhao L, Lu F (2012). “Berberine in the treatment of type 2 diabetes mellitus: a systemic review and meta-analysis”. Evid Based Complement Alternat Med. 2012: 591654. doi:10.1155/2012/591654. PMC 3478874 . PMID 23118793.
3- Dong H, Zhao Y, Zhao L, Lu F (2013). “The effects of berberine on blood lipids: a systemic review and meta-analysis of randomized controlled trials”. Planta Med. 79 (6): 437–46. doi:10.1055/s-0032-1328321. PMID 23512497.
4- Mohan MC, Abhimannue AP, B PK. Identification and Characterization of Berberine in Tinospora cordifolia by Liquid Chromatography Quadrupole Time of Flight Mass Spectrometry (LC MS/MS Q-tof) and Evaluation of its anti Inflammatory Potential. Pharmacognosy Journal. 2017;9(3):350–355.
5- Huang WW, Xu SZ, Xu YQ. A study of the antiarrhythmic mechanism of berberine on delayed activation potassium current by voltage clamp. Zhonghua Xin Xue Guan Bing Za Zhi, 20 (5): 310–2, 1992 | PMID 1306830
6- Zhao H, Halicka HD, Li J, Darzynkiewicz Z. Berberine suppresses gero-conversion from cell cycle arrest to senescence. Aging (Albany) 2013; 6: 623–636. PMID 23974852, doi:10.18632/aging.100593
7- Darzynkiewicz Z, Zhao H, Halicka HD, Li J, Lee Y-S, Hsieh T-C, Wu J. In search of anti-aging modalities: evaluation of mTOR- and ROS/DNA damage- signaling by cytometry. Cytometry A 2014;85A:386-99. PMID 24677687, doi:10.1002/cyto.a.22452
8- Yu HH, Kim KJ, Cha JD, Kim HK, Lee YE, Choi NY, You YO (2005). “Antimicrobial activity of berberine alone and in combination with ampicillin or oxacillin against methicillin-resistant Staphylococcus aureus”. Journal of Medicinal Food. 8 (4): 454–61. doi:10.1089/jmf.2005.8.454. PMID 16379555.
9- “Poster Presentations”. FEBS Journal. 277: 37–271. 2010. doi:10.1111/j.1742-4658.2010.07680.x.
10- Li Y., Zuo G.-Y. ‘Advances in studies on antimicrobial activities of alkaloids” Chinese Traditional and Herbal Drugs 2010 41:6 (1006–1014)
11- Stermitz FR, Lorenz P, Tawara JN, Zenewicz LA, Lewis K (February 2000). “Synergy in a medicinal plant: antimicrobial action of berberine potentiated by 5′-methoxyhydnocarpin, a multidrug pump inhibitor”. Proceedings of the National Academy of Sciences of the United States of America. 97 (4): 1433–7. Bibcode:2000PNAS…97.1433S. doi:10.1073/pnas.030540597. PMC 26451 . PMID 10677479.
12- Zhang S, Zhang B, Xing K, Zhang X, Tian X, Dai W (2010). “Inhibitory effects of golden thread (Coptis chinensis) and berberine on Microcystis aeruginosa”. Water Science & Technology. 61 (3): 763–9. doi:10.2166/wst.2010.857. PMID 20150713.
13- Liu CS, Zheng YR, Zhang YF, Long XY (2016). “Research progress on berberine with a special focus on its oral bioavailability”. Fitoterapia (Review). 109: 274–82. doi:10.1016/j.fitote.2016.02.001. PMID 26851175.
14- Pan GY, et al. Inhibitory action of berberine on glucose absorption. Yao Xue Xue Bao. (2003)
15- Pan GY, et al. The antihyperglycaemic activity of berberine arises from a decrease of glucose absorption. Planta Med. (2003)
16- Liu SZ, et al. Antihyperglycemic effect of the traditional Chinese scutellaria-coptis herb couple and its main components in streptozotocin-induced diabetic rats. J Ethnopharmacol. (2012)
17- Liu L, et al. Berberine suppresses intestinal disaccharidases with beneficial metabolic effects in diabetic states, evidences from in vivo and in vitro study. Naunyn Schmiedebergs Arch Pharmacol. (2010)
18- Liu L, et al. Berberine attenuates intestinal disaccharidases in streptozotocin-induced diabetic rats. Pharmazie. (2008)
19- Tintu I, et al. An isoquinoline alkaloid, berberine, can inhibit fungal alpha amylase: enzyme kinetic and molecular modeling studies. Chem Biol Drug Des. (2012)
20- Ni YX. Therapeutic effect of berberine on 60 patients with type II diabetes mellitus and experimental research. Zhong Xi Yi Jie He Za Zhi. (1988)
21- Dong H, et al. Berberine in the treatment of type 2 diabetes mellitus: a systemic review and meta-analysis. Evid Based Complement Alternat Med. (2012)
22- Sterne JA, Gavaghan D, Egger M. Publication and related bias in meta-analysis: power of statistical tests and prevalence in the literature. J Clin Epidemiol. (2000)
23- Yang J, et al. Berberine improves insulin sensitivity by inhibiting fat store and adjusting adipokines profile in human preadipocytes and metabolic syndrome patients. Evid Based Complement Alternat Med. (2012)
24- Zhang Y, et al. Treatment of type 2 diabetes and dyslipidemia with the natural plant alkaloid berberine. J Clin Endocrinol Metab. (2008)
25- Zhang H, et al. Berberine lowers blood glucose in type 2 diabetes mellitus patients through increasing insulin receptor expression. Metabolism. (2010)
26- Yan HM, et al. Efficacy of Berberine in Patients with Non-Alcoholic Fatty Liver Disease. PLoS One. (2015)
27- Pérez-Rubio KG, et al. Effect of berberine administration on metabolic syndrome, insulin sensitivity, and insulin secretion. Metab Syndr Relat Disord. (2013)
28- Efficacy of Berberine in Patients with Type 2 Diabetes.
29- Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr Rev. (2005)
30- Basu R, et al. Selective downregulation of the high molecular weight form of adiponectin in hyperinsulinemia and in type 2 diabetes: differential regulation from nondiabetic subjects. Diabetes. (2007)
31- Wang Y, et al. Post-translational modifications of adiponectin: mechanisms and functional implications. Biochem J. (2008)
32- Li Y, et al. Activation of AMPK by berberine promotes adiponectin multimerization in 3T3-L1 adipocytes. FEBS Lett. (2011)
33- Lin H, Li Z. Adiponectin self-regulates its expression and multimerization in adipose tissue: an autocrine/paracrine mechanism. Med Hypotheses. (2012)
34- Berberine-stimulated glucose uptake in L6 myotubes involves both AMPK and p38 MAPK.
35- Ma X, et al. Berberine-induced activation of 5’-adenosine monophosphate-activated protein kinase and glucose transport in rat skeletal muscles. Metabolism. (2010)
36- Gomes AP, et al. Berberine protects against high fat diet-induced dysfunction in muscle mitochondria by inducing SIRT1-dependent mitochondrial biogenesis. Biochim Biophys Acta. (2012)
37- Prabhakar PK, Doble M. Effect of Natural Products on Commercial Oral Antidiabetic Drugs in Enhancing 2-Deoxyglucose Uptake by 3T3-L1 Adipocytes. Ther Adv Endocrinol Metab. (2011)
38- Wang SH, et al. Effect of Astragalus polysaccharides and berberine on carbohydrate metabolism and cell differentiation in 3T3-L1 adipocytes. Zhongguo Zhong Xi Yi Jie He Za Zhi. (2004)
39- Chen C, Zhang Y, Huang C. Berberine inhibits PTP1B activity and mimics insulin action. Biochem Biophys Res Commun. (2010)
40- Cok A, et al. Berberine acutely activates the glucose transport activity of GLUT1. Biochimie. (2011)
41- Kim SH, et al. Berberine activates GLUT1-mediated glucose uptake in 3T3-L1 adipocytes. Biol Pharm Bull. (2007)
42- Bustanji Y, et al. Berberine potently inhibits protein tyrosine phosphatase 1B: investigation by docking simulation and experimental validation. J Enzyme Inhib Med Chem. (2006)
43- Yi P, et al. Berberine reverses free-fatty-acid-induced insulin resistance in 3T3-L1 adipocytes through targeting IKKbeta. World J Gastroenterol. (2008)
44- Choi BH, et al. Berberine reduces the expression of adipogenic enzymes and inflammatory molecules of 3T3-L1 adipocyte. Exp Mol Med. (2006)
45- Yi P, Lu FE, Chen G. Molecular mechanism of berberine in improving insulin resistance induced by free fatty acid through inhibiting nuclear trascription factor-kappaB p65 in 3T3-L1 adipocytes. Zhongguo Zhong Xi Yi Jie He Za Zhi. (2007)
46- Hu Y, et al. Lipid-lowering effect of berberine in human subjects and rats. Phytomedicine. (2012)
47- Yan HM, et al. Efficacy of Berberine in Patients with Non-Alcoholic Fatty Liver Disease. PLoS One. (2015)
48- Li H, et al. Hepatocyte nuclear factor 1alpha plays a critical role in PCSK9 gene transcription and regulation by the natural hypocholesterolemic compound berberine. J Biol Chem. (2009)
49- Zhang M, et al. Sodium caprate augments the hypoglycemic effect of berberine via AMPK in inhibiting hepatic gluconeogenesis. Mol Cell Endocrinol. (2012)
50- Kong WJ, et al. Berberine reduces insulin resistance through protein kinase C-dependent up-regulation of insulin receptor expression. Metabolism. (2009)
51- Wang ZS, et al. Berberine reduces endoplasmic reticulum stress and improves insulin signal transduction in Hep G2 cells. Acta Pharmacol Sin. (2010)
52- Sack RB, Froehlich JL. Berberine inhibits intestinal secretory response of Vibrio cholerae and Escherichia coli enterotoxins. Infect Immun. (1982)
53- Liu LZ, et al. Berberine modulates insulin signaling transduction in insulin-resistant cells. Mol Cell Endocrinol. (2010)
54- Liu LZ, et al. The pivotal role of protein kinase C zeta (PKCzeta) in insulin- and AMP-activated protein kinase (AMPK)-mediated glucose uptake in muscle cells. Cell Signal. (2010)
55- Zhao Y, et al. The in vitro inhibition of human CYP1A2, CYP2D6 and CYP3A4 by tetrahydropalmatine, neferine and berberine. Phytother Res. (2012)
56- Liu J, et al. Coptis extracts enhance the anticancer effect of estrogen receptor antagonists on human breast cancer cells. Biochem Biophys Res Commun. (2009)
57- Doyle ME, Egan JM. Mechanisms of action of glucagon-like peptide 1 in the pancreas. Pharmacol Ther. (2007)
58- Meier JJ, Nauck MA. Glucagon-like peptide 1(GLP-1) in biology and pathology. Diabetes Metab Res Rev. (2005)
59- Holst JJ, Gromada J. Role of incretin hormones in the regulation of insulin secretion in diabetic and nondiabetic humans. Am J Physiol Endocrinol Metab. (2004)
60- Li J1, et al. Berberine represses DAXX gene transcription and induces cancer cell apoptosis. Lab Invest. (2013)
61- The 5-minute Herb and Dietary Supplement Consult – a cura di Adriane Fugh-Berman (pag. 158).
Con il termine Antiestrogeni ci si riferisce genericamente ad una classe di farmaci aventi azione diretta o indiretta sull’attività tissutale e/o concentrazione ematica degli estrogeni. Agiscono bloccando il recettore dell’estrogeno (ER) e/o riducendo o sopprimendo la sintesi estrogenica.(1)(2) Una recente categoria di agenti facenti parte di questa classe di farmaci, i SERD (Selective Estrogen Receptor Degrader), esplicano la loro azione antiestrogena degradando/sottoregolando il recettore dell’estrogeno. Gli Antiestrogeni sono una delle tre classi di farmaci antagonisti dell’ormone sessuale, insieme agli Antiandrogeni e agli Antiprogestinici.(3)
Largamente utilizzati in ambito sportivo, in special modo nell’ambiente culturistico, con il fine di controllare l’attività estrogenica durante l’uso di AAS aromatizzabili, o aventi attività estrogenica intrinseca, e durante la PCT con lo scopo aggiunto di stimolare la ripresa dell’HPTA, questi farmaci hanno un discreto carico di effetti collaterali tra i quali, quelli che destano maggior preoccupazione nell’atleta previdente, vi sono la dislipidemia (aumento dell’LDL, dei Trigliceridi, riduzione del HDL e alterazione delle loro ratio), l’atralgia (dolore articolare), il calo della libido/disfunzione erettile e l’affaticamento/letargia. Non sono di certo da meno le preoccupazioni legate all’alterazione dell’Asse GH/IGF-1 o la riduzione delle potenzialità di induzione ipertrofica di un ciclo in seguito ad una eccessiva soppressione dell’attività e/o delle concentrazioni estrogeniche. Ma esiste un’altra preoccupazione legata all’uso di composti antiestrogeni, ed è la possibilità che si verifichi un rebound estrogenico in seguito al l’oro uso. Purtroppo, la letteratura a disposizione è al quanto scarsa e poco chiara nella specifica del problema. E’ possibile, però, fare maggiore chiarezza sulla questione analizzando le caratteristiche dei composti antiestrogeni e il loro impatto, passando in rassegna tutti i componenti dell’addizione (Recettori Estrogeni e enzima Aromatasi). In questo articolo cercherò di esporre un ragionamento logico grazie al quale, seppur non avendo una risposta definitiva, sarà possibile avere un idea, la più concreta possibile, sul binomio antiestrogeni/rebound estrogenico.
Una analisi della questione…
Recettori dell’Estrogeno, SERM e “rebound estrogenico”
Un dimero della regione legame-ligando del ERa.
I Recettori degli Estrogeni (ER) sono un gruppo di proteine presenti all’interno delle cellule. Sono recettori attivati dall’ormone estrogeno (con maggiore attività del 17β-estradiolo).(4) Esistono due classi di ER: i Recettori degli Estrogeni nucleari (ERα e ERβ), che sono membri della famiglia dei recettori nucleari e dei recettori intracellulari, ed i Recettori degli Estrogeni di Membrana (MR) (GPER (GPR30), ER-X e Gq-mER), che sono per lo più recettori accoppiati alla proteina G. In questa sede ci si riferirà ai primi (ER).
Una volta attivato dall’estrogeno, l’ER è in grado di traslocare nel nucleo e legarsi al DNA per regolare l’attività di diversi geni (ciò significa che è un fattore di trascrizione del DNA). Tuttavia, ha anche funzioni aggiuntive indipendenti dal legame con il DNA.(5)
Poiché l’estrogeno è un ormone steroideo, può passare attraverso le membrane fosfolipidiche della cellula, e pertanto i recettori non hanno bisogno di essere legati alla membrana per potersi legare a loro volta con l’estrogeno.
L’estrogeno esplica la sua attività cellulare attraverso un azione Genomica e Non-Genomica.
• Genomica
In assenza di ormoni, i ER si trovano in gran parte nel citosol. Il legame dell’ormone al recettore innesca un numero di eventi che iniziano con la migrazione del recettore dal citosol nel nucleo, la dimerizzazione del recettore e il successivo legame del dimero del recettore a specifiche sequenze di DNA conosciute come elementi di risposta ormonale. Il complesso DNA / recettore quindi recluta altre proteine che sono responsabili della trascrizione del DNA a valle in mRNA e, infine, in una proteina la quale porta a dei cambiamenti nella funzione cellulare. I recettori degli estrogeni si trovano anche all’interno del nucleo della cellula, ed entrambi i sottotipi del recettore dell’estrogeno hanno un dominio di legame con il DNA e possono funzionare come fattori di trascrizione per regolare la produzione di proteine.
Il recettore interagisce anche con la proteina attivatore 1 e Sp-1 per promuovere la trascrizione, attraverso diversi coattivatori come il PELP-1.(6)
L’acetilazione diretta del recettore alfa dell’estrogeno ai residui della lisina nella regione cerniera mediante il p300 regola la transattivazione e la sensibilità ormonale.(7)
• Non-Genomica
Alcuni recettori per gli estrogeni sono presenti nelle membrana della superficie della cellula e possono essere rapidamente attivati dall’esposizione di questa agli estrogeni.(8)(9)
Inoltre, alcuni ER possono associarsi alle membrane cellulari legandole alla caveolina-1 e formarmando complessi con la proteine G, striatina, tirosina chinasi del recettore (es. EGFR e IGF-1) e tirosina chinasi non recettoriale (es. Src). (6)(8) Attraverso la striatina, alcuni di questi ER legati alla membrana possono portare a livelli aumentati di Ca2 + e ossido nitrico (NO).(10) Attraverso il recettore tirosin chinasi, i segnali vengono inviati al nucleo attraverso la via della proteina chinasi attivata dal mitogeno (MAPK / ERK) e la via del fosfoinositide 3-chinasi (Pl3K / AKT).(11) La glicogeno sintasi chinasi-3 (GSK) -3β inibisce la trascrizione dal ER nucleare inibendo la fosforilazione della serina 118 dell’ERa nucleare. La fosforilazione di GSK-3β rimuove il suo effetto inibitorio, e questo può essere ottenuto tramite il pathway PI3K / AKT e il pathway MAPK / ERK, tramite rsk.
Il 17β-estradiolo ha dimostrato di attivare il recettore GPR30 accoppiato alla proteina G.(12) Tuttavia, la localizzazione subcellulare e il ruolo di questo recettore sono ancora oggetto di controversie.(13)
ERb.
Gli estrogeni e gli ER sono implicati nel cancro al seno, nel carcinoma ovarico, nel cancro del colon, nel cancro alla prostata e nel cancro dell’endometrio. Il carcinoma del colon avanzato è associato a una perdita di ERβ, l’ER predominante nel tessuto del colon, e il tumore del colon è trattato con agonisti specifici per ERβ.(14)
Sappiamo che i recettori degli estrogeni sono sovraespressi in circa il 70% dei casi di cancro al seno, indicati come “ER-positivi”, e possono essere dimostrati in tali tessuti mediante l’immunoistochimica.(15) E’ ipotizzabile ,quindi, che gli atleti più sensibili agli effetti estrogenici presentino un espressione dei ER più elevata del normale, cosa che li porta a sviluppare con maggiore facilità effetti avversi dati da un eccesso dei livelli estrogenici e/o da un aumento della loro attività dato dalla cosomministrazione con progestinici (es. Nandrolone e Trenbolone).
I Modulatori Selettivi del Recettore dell’Estrogeno (SERM) sono composti antiestrogenici che agiscono a livello del ER. (16) Una caratteristica che distingue queste sostanze dagli agonisti e antagonisti ER puri (cioè agonisti completi e antagonisti silenti) è che la loro azione è diversa nei vari tessuti, garantendo in tal modo la possibilità di inibire selettivamente o stimolare l’azione estrogenica in diversi tessuti.
I SERM sono agonisti parziali competitivi del ER.(17) Tessuti diversi presentano differenti gradi di sensibilità all’attività degli estrogeni, pertanto i SERM esplicano effetti estrogenici o antiestrogeni a seconda del tessuto specifico con il quale interagiscono e della percentuale di attività intrinseca (IA) del composto in questione.(18) Un esempio di SERM con alta IA, e quindi di effetti prevalentemente estrogenici, è rappresentato dal Clorotrianisene, mentre un esempio di SERM con bassa IA, e quindi avente per lo più attività antiestrogenica, è rappresentato dall’Ethamoxytrifetolo. SERM come il Clomifene e il Tamoxifene, largamente utilizzati in ambito sportivo, sono considerabili come composti con valore IA intermedio essendo molecole con una azione bilanciata tra effetti estrogenici e antiestrogenici. Il Raloxifene è un SERM che presenta una azione antiestrogenica maggiore del Tamoxifene; entrambi hanno una attività estrogenica (sebbene differente) a livello osseo, ma il Raloxifene presenta una attività antiestrogenica nell’utero mentre il Tamoxifene ha un azione estrogenica nel tessuto dell’utero.(18)
Tamoxifene
Il Tamoxifene è un farmaco di prima linea per il trattamento del carcinoma mammario metastatico ER-positivo. È usato per la riduzione delle possibilità di sviluppo del cancro al seno nelle donne ad alto rischio, come trattamento adiuvante del nodo ascellare negativo e positivo, e nel carcinoma duttale in situ.(19)(20)
Il Tamoxifene è classificabile come un profarmaco, dal momento che la sua affinità per la proteina bersaglio (ER) è limitata. Il Tamoxifene viene metabolizzato nel fegato dall’isoforma del citocromo CYP2D6 e CYP3A4 in metaboliti attivi come l’Afimoxifene (4-idrossitamoxifene; 4-OHT) e l’Endoxifene (N-desmetil-4-idrossitamoxifene) (21) che presentano una affinità da 30 a 100 volte maggiore per il ER rispetto al Tamoxifene. (22) Questi metaboliti attivi competono con gli estrogeni per il legame con il recettore. Nel tessuto mammario, il 4-OHT agisce come un antagonista del ER in modo da inibire la trascrizione dei geni che reagiscono agli estrogeni. (23) Il Tamoxifene ha rispettivamente il 7% e il 6% dell’affinità dell’Estradiolo per il ERα e il ERβ, mentre il 4-OHT ha il 178% e il 338% dell’affinità dell’Estradiolo per il ERα e il ERβ.(24)
Afimoxifene (4-OHT)
Il 4-OHT si lega al ER, il complesso ER/Tamoxifene recluta altre proteine note come co-repressori e quindi si lega al DNA per modulare l’espressione genica. Alcune di queste proteine includono la NCoR e la SMRT. (25) La funzione del Tamoxifene può essere regolata da una serie di variabili diverse, compresi i fattori di crescita.(26) Il Tamoxifene deve bloccare le proteine del fattore di crescita come ErbB2/HER2 (27) perché è stato dimostrato che livelli elevati di ErbB2 si manifestano nei tumori resistenti al Tamoxifene.(28) Il Tamoxifene sembra richiedere una proteina PAX2 affinché possa esplicare il suo pieno effetto antitumorale. (27)(29) In presenza di un elevata espressione della PAX2, il complesso Tamoxifene/ER è in grado di sopprimere l’espressione della proteina pro-proliferativa del ERBB2. Al contrario, quando l’espressione del AIB-1 è superiore alla PAX2, il complesso di Tamoxifene/ER aumenta l’espressione del ERBB2 con conseguente stimolazione della crescita del cancro al seno. (27)(30)
Il 4-OHT si lega al ER in modo competitivo (rispetto all’estrogeno agonista) nelle cellule tumorali e in altri bersagli tissutali, producendo un complesso nucleare che riduce la sintesi del DNA e inibisce gli effetti degli estrogeni. È un agente non steroideo con potenti proprietà antiestrogeniche che competono con gli estrogeni per i siti di legame nel seno e in altri tessuti. Il Tamoxifene fa sì che le cellule rimangano nelle fasi G0 e G1 del ciclo cellulare. Poiché impedisce alle cellule (pre) cancerose di dividersi ma non provoca la morte cellulare, il Tamoxifene è citostatico piuttosto che citocida.
La letteratura scientifica riguardante l’attività del Tamoxifene è a dir poco complessa ed occorre prestare particolare attenzione ai dati disponibili per stabilire se il Tamoxifene, o il suo metabolita 4-idrossi, abbiano il maggiore impatto complessivo.
Norendoxifene
Il Norendoxifene (N, N-didesmetil-4-idrossitamoxifene), un altro metabolita attivo del Tamoxifene, è stato osservato agire come un potente inibitore dell’aromatasi competitivo (IC50 = 90 nM), cosa che a sua volta può amplificare l’attività antiestrogenica complessiva del Tamoxifene.(31)
Come già accennato in precedenza, e come molti sapranno, il Tamoxifene è largamente utilizzato in ambito sportivo, sia da solo che in abbinamento con altri SERM come il Clomifene ( in PCT) o con AI (“on-cycle” e/o in PCT). La sua applicazione all’interno di una preparazione che contempla l’uso di AAS aromatizzabili, alla luce di quanto esposto pocanzi, lo vede come agente preventivo o di trattamento dell’attività estrogenica a livello tissutale, in specie per quanto concerne l’attività estrogenica nel tessuto mammario al fine di evitare (o “tamponare”) la comparsa della ginecomastia. In un contesto PCT tale composto, oltre ad esercitare la funzione di regolazione dell’attività estrogenica appena esposta, agendo a livello ipotalamico stimola il rilascio di GnRH e, consequenzialmente, di LH ed FSH dall’Ipofisi che a loro volta stimoleranno la sintesi di Testosterone e la spermatogenesi.
Il suo utilizzo massivo e cronico è stato spesso collegato aneddoticamente a rebound estrogenico. Ora, conoscendo la complessità d’azione che questo composto (ed i suoi metaboliti) ha sul controllo dell’attività estrogenica, si può facilmente ipotizzare che un suo uso protratto (legato anche alla dose e, quindi, al suo impatto sulla attività estrogenica sistemica) possa innescare degli adattamenti reattivi con conseguente aumento dell’attività estrogenica attraverso l’incremento dei livelli serici di Estradiolo e dell’attività non-genomica dello steroide (ipotizzabile anche un aumento del numero dei ER). Nel corso degli anni sono state esposte diverse ipotesi volte a spiegare i meccanismi attraverso i quali un abuso di Tamoxifene possa portare ad un rebound estrogenico. Una di queste ipotesi venne riportata all’inizio del secolo dal compianto A.L. Rea il quale affermava che la causa andasse ricercata nell’aumento del rilascio di DHEA da parte delle ghiandole surrenali e dalla sua successiva (e aumentata) conversione in Androstenedione e, attraverso l’intervento dell’enzima aromatasi che lo converte in Estrone e la successiva azione del estradiolo 17beta-deidrogenasi, Estradiolo.(32) In breve, secondo questa teoria i processi innescati causerebbero l’instaurarsi di livelli di E2 cronicamente alti con conseguente impossibilità del SERM di esplicare la sua azione. Questa teoria seppur, in parte, possa dare una spiegazione logica dei possibili meccanismi implicati manca di alcuni tasselli. Il principale “tallone d’Achille” è rappresentato dai livelli di E2 che, una volta aumentati, diventano dei competitor recettoriali più aggressivi rispetto al 4-OHT (che ricordiamo avere il 178% dell’affinità dell’Estradiolo per il ERα). Ciò potrebbe avvenire in situazioni di calo delle concentrazioni di 4-OHT seguenti alla riduzione del dosaggio del farmaco o alla sua cessazione, quindi, in questo ultimo caso, esplicabili in crescendo nei 7-14 giorni successivi all’interruzione della somministrazione e con una durata indeterminata. Di conseguenza, sembra più plausibile che l’aumento delle concentrazioni di E2, durante l’uso del Tamoxifene, si affianchi ad un consequenziale incremento dell’attività Non-Genomica dell’ormone e da un aumentato numero di ER. Seguendo questa logica, una volta interrotto l’uso del Tamoxifene, queste condizioni tenderanno ad aggravarsi come gli effetti avversi a loro legati.
Consultando la bibliografia scientifica disponibile, non si trovano accenni su un possibile rebound estrogenico in seguito all’uso di Tamoxifene, ma si parla nello specifico di “resistenza al Tamoxifene” o “sottoregolazione degli ER”.(33)(34) Nel caso della “resistenza al Tamoxifene” sembra che l’aumento dell’espressione del gene MACROD2 porti ad una risposta negativa all’azione del SERM con conseguente proliferazione delle cellule cancerose estradiolo-dipendenti. La sovra espressione di tale gene sembra essere di base genetica anche se non si esclude una risposta di adattamento in seguito ad uso cronico del composto in questione.
Raloxifene
Il Raloxifene, un altro SERM discretamente utilizzato nella pratica sportiva, è un agonista-antagonista misto del ER.(35)(36)(37) Ha effetti estrogenici a livello osseo ed epatico con effetti antiestrogenici nei seni e nell’utero. Le azioni biologiche del Raloxifene sono quindi ampiamente mediate dal legame con i ER. Questo legame determina l’attivazione di percorsi estrogenici in alcuni tessuti (agonismo) e il blocco di questi in altri (antagonismo). Le sue caratteristiche d’azione similari a quelle del Tamoxifene, sembrano poter far pensare ad un medesimo e ipotetico meccanismo che possa portare ad un rebound estrogenico. Questa volta la letteratura scientifica sembra dare alcune conferme. In un caso studio (38), una paziente di 66 anni si è presentata con recidiva metastatica acuta estrogeno-positiva e progesterone-positiva, carcinoma mammario Her-2 / neu-negativo, lesioni ossee (colonna lombare, bacino), noduli polmonari, metastasi epatiche, antigene tumorale elevato 15 e enzimi epatici, dispepsia e diarrea. La paziente aveva assunto Raloxifene per circa 8 anni. Dopo la sospensione del farmaco, parametri e sintomi clinici sono migliorati rapidamente senza terapia oncologica o altre forme di trattamento. Tre mesi dopo la sospensione del Raloxifene, l’oncologo ha prescritto alla paziente l’uso della Capecitabina dato che non riteneva plausibile un effetto di rebound estrogenico (anti-estrogen withdrawal effect – AEWE). Tuttavia, la regressione duratura è stata più indicativa di un effetto rebound dato dal Raloxifene rispetto alla chemioterapia o ad altri interventi. In seguito la paziente si è mostrata asintomatica con un buono stato di prestazione. La regressione metastatica epatica è stata confermata, senza alcun trattamento oncologico somministrato negli ultimi 16 mesi e circa 23 mesi dopo il termine d’uso del Raloxifene. Questo caso evidenzia la necessità di esaminare pazienti con carcinoma mammario per la possibilità di un AEWE con l’uso di Raloxifene o con altri SERM . Ovviamente, il caso presentato non è molto comparabile, soprattutto per quanto riguarda i tempi di somministrazione, ad un BodyBuilder supplementato chimicamente nella “media” ma, ciò nonostante, ci offre un indizio sulla probabilità che si possa manifestare un rebound estrogenico con l’uso di SERM.
Enzima Aromatasi, Inibitori della Aromatasi e “rebound estrogenico”
Enzima Aromatasi
L’Enzima Aromatasi, chiamato anche estrogeno sintetasi o estrogeno sintasi, è un enzima responsabile del processo fondamentale della biosintesi degli Estrogeni. Denominato CYP19A1, questo enzima è un membro della superfamiglia del citocromo P450 (EC 1.14.14.1), che sono monoossigenasi che catalizzano molte reazioni coinvolte nella steroidogenesi. In particolare, l’Aromatasi è responsabile dell’aromatizzazione degli Androgeni in Estrogeni. L’enzima Aromatasi è sintetizzato in molti tessuti tra cui le gonadi (cellule della granulosa), cervello, tessuto adiposo, placenta, vasi sanguigni, pelle e ossa, nonché nei tessuti dell’endometriosi, dei fibromi uterini, del cancro al seno e del cancro dell’endometrio.
L’Aromatasi è localizzato nel reticolo endoplasmatico dove è regolato da promotori tissutali che sono a loro volta controllati da ormoni, citochine e altri fattori. Catalizza gli ultimi passaggi della biosintesi degli estrogeni dagli androgeni (in particolare, converte l’Androstenedione in Estrone e il Testosterone in Estradiolo). Queste fasi comprendono tre idrossilazioni successive del gruppo 19-metilico degli androgeni, seguite dall’eliminazione simultanea del gruppo metilico come formiato e aromatizzazione dell’anello A.
Reazioni generali per la conversione del Testosterone in Estradiolo catalizzata dall’Aromatasi. Gli Steroidi sono formati da quattro anelli fusi (A-B-C-D). L’Enzima Aromatasi converte l’anello “A” in uno stato aromatico.
Il gene esprime due varianti di trascrizione. (39) Nell’uomo, il gene CYP19, situato sul cromosoma 15q21.1, codifica per l’Enzima Aromatasi. (40) Il gene ha nove esoni codificanti e un numero di primi esoni non codificanti alternativi che regolano l’espressione specifica del tessuto. (41)
Il CYP19 è presente in un cordato precoce divergente, l’anfiosso cefalocordato (il Florida lancelet, Branchiostoma floridae), ma non nel precedente tunicato divergente Ciona intestinalis. Pertanto, gli evoluzionisti ipotizzano che il gene Aromatasi si sia evoluto precocemente nell’evoluzione dei cordati e non sembra essere presente negli invertebrati non-cordati (ad esempio insetti, molluschi, echinodermi, spugne, coralli). Tuttavia, gli Estrogeni possono essere sintetizzati in alcuni di questi organismi, attraverso altri percorsi sconosciuti.
I fattori noti che aumentano l’attività dell’Aromatasi includono l’età, l’obesità, l’Insulina, le gonadotropine e l’alcol. L’attività dell’Aromatasi risulta diminuita dalla Prolattina, dall’ormone anti-Mülleriano e dal glifosato , un comune erbicida.(42) L’attività dell’Aromatasi sembra essere migliorata in alcuni tessuti estrogeno-dipendenti come il tessuto mammario, nel carcinoma dell’endometrio, nell’endometriosi e nei fibromi uterini.
Gli Inibitori dell’Aromatasi (AI) sono una gruppo di farmaci usati nel trattamento del carcinoma mammario nelle donne in postmenopausa e nella ginecomastia negli uomini. Come i SERM, trovano un largo uso off-label in ambito sportivo durante la somministrazione di AAS aromatizzabili o durante la PCT. Possono anche essere utilizzati per la chemioprevenzione in donne ad alto rischio.
Esistono due tipi di Inibitori dell’Aromatasi approvati per il trattamento del carcinoma mammario e, quindi, diffusi anche per l’uso off-label: (43)
– Gli inibitori steroidei irreversibili, come l’Exemestano (nome commerciale Aromasin), formano un legame permanente e disattivante con l’Enzima Aromatasi.
– Gli inibitori non steroidei, come l’Anastrozolo (nome commerciale Arimidex) e il Letrozolo (nome commerciale Femara), inibiscono la sintesi degli Estrogeni attraverso la competizione reversibile per l’Enzima Aromatasi.
Gli inibitori dell’Aromatasi disponibili (AI) includono:
– Non selettivi:
• L’Aminoglutetimide, il quale però inibisce l’enzima P450scc agendo come inibitore della biosintesi di tutti gli ormoni steroidei (aprirò una nota a riguardo più avanti).
• Testolactone (nome commerciale Teslac) – Selettivi:
• Anastrozolo (Arimidex)
• Letrozolo (Femara)
• Exemestano (Aromasin)
• Vorozolo (Rivizor)
• Formestano (Lentaron)
• Fadrozolo (Afema)
– Non classificati:
• 1,4,6-Androstatrien-3,17-dione (ATD)
• 4-Androstene-3,6,17-trione (“6-OXO”)
Oltre agli AI farmaceutici, alcuni composti naturali hanno mostrato effetti di inibizione dell’Aromatasi, come le foglie di damiana. Il loro impatto non è stato pienamente chiarito sull’uomo.
Gli Inibitori dell’Aromatasi agiscono, proprio come suggerisce il nome, inibendo l’azione dell’enzima Aromatasi, che converte gli Androgeni in Estrogeni mediante un processo chiamato aromatizzazione. Poiché il tessuto mammario è stimolato dagli Estrogeni, diminuirne la produzione è un modo per sopprimere la recidiva del tessuto tumorale del seno. La principale fonte di Estrogeni è rappresentata dalle ovaie nelle donne in premenopausa, mentre nelle donne in post-menopausa la maggior parte degli Estrogeni del corpo viene prodotta nei tessuti periferici (al di fuori del SNC) e anche in alcuni siti del SNC in varie regioni del cervello. L’Estrogeno viene prodotto e agisce localmente in questi tessuti, ma qualsiasi estrogeno circolante, che esercita effetti estrogenici sistemici in uomini e donne, è il risultato dell’Estrogeno che sfugge al metabolismo locale e si diffonde nel sistema circolatorio.(44)
Come già accennato pocanzi, i composti AI sono anch’essi, al pari dei SERM, largamente utilizzati in ambito sportivo, sia come agenti di controllo dei livelli estrogenici durante l’uso di AAS aromatizzabili (uso preventivo della comparsa di effetti estrogenici), in caso di ginecomastia (spesso in combinazione con un SERM, specie se l’AI utilizzato è l’Exemestano) o in combinazione con i SERM in ambito PCT (specie nella fase preliminare dove viene utilizzata l’hCG).
Più che con i SERM, il rebound estrogenico è stato riportato, soprattutto aneddoticamente, con l’uso di AI, specialmente quelli reversibili (vedi Anastrozolo e Letrozolo).
L’Anastrozolo ed il Letrozolo agiscono legandosi in modo reversibile all’Enzima Aromatasi (unità eme del citocromo P450) e, attraverso l’inibizione competitiva, blocca la conversione degli Androgeni in Estrogeni nei tessuti periferici (extragonali).
Il Letrozolo ha dimostrato, attraverso studi clinici, di poter abbassare rapidamente il livello degli estrogeni fino al 65%. Il motivo principale è probabilmente legato alla capacità che la molecola ha di abbassare drasticamente gli estrogeni attraverso un legame competitivo reversibile al gruppo eme della relativa unità del citocromo P450. L’Anastrozolo, il quale agisce similmente al Letrozolo, ha mostrato una riduzione del livello estrogenico in soggetti di sesso maschile del 50%.(45) Il problema di un possibile rebound estrogenico con questi composti nasce proprio dalla loro natura “reversibile”.
Rebound estrogenici sono stati riportati sia con l’uso di Letrozolo che con l’uso di Anastrozolo, sebbene il Letrozolo, avendo un azione inibitoria più marcata, sembra causare rebound di intensità maggiore dopo la sua interruzione. La causa del rebound estrogenico indotto da cessazione d’uso di Letrozolo o di Anastrozolo è proprio legata al comportamento che queste due molecole esplicano nei confronti dell’Enzima aromatasi. Il legame tra la molecola di Letrozolo o di Anastrozolo con l’Enzima Aromatasi è solo temporanea e non decreta la completa de-attivazione dell’enzima responsabile della conversione degli Androgeni in Estrogeni. Una volta interrotta l’assunzione del composto, i livelli di Aromatasi possono salire significativamente con la possibile comparsa di un rebound estrogenico. Una pratica per evitare che ciò si verifichi consiste nell’uso limitato dei due composti e in una loro graduale sospensione. Con questi farmaci, il rebound estrogenico può essere “multifattoriale” derivando non solo dalla cessazione del farmaco in questione ma anche da un incremento dell’espressione dell’Enzima Aromatasi come risposta adattativa all’uso (specie nel lungo termine). Ciò significa che, anche durate un utilizzo cronico, i livelli di E2 possono mostrare degli aumenti, aumenti che diverranno maggiormente significativi una volta cessato l’uso del farmaco. Cessata l’azione del composto non solo viene a mancare un controllo dell’aromatizzazione ma questa risulta anche incrementata rispetto ai tassi pre-utilizzo (l’aumento dell’espressione dell’aromatasi è un comportamento adattativo che si può manifestare anche durante cicli particolarmente lunghi). Prendendo in considerazione la vita attiva del Letrozolo e dell’Anastrozolo, il possibile rebound estrogenico potrebbe manifestarsi in crescendo dopo 64-120h circa dall’ultima assunzione.
Exemestano
L’Exemestano, invece, è un inibitore dell’Aromatasi steroideo irreversibile di tipo I, strutturalmente correlato al substrato naturale 4-androstenedione. Agisce come un falso substrato per l’Enzima Aromatasi e viene trasformato in un intermedio che si lega irreversibilmente al sito attivo dell’enzima causandone l’inattivazione, un effetto noto anche come “inibizione suicida”. Essendo strutturalmente simile agli obiettivi dell’enzima, l’Exemestano si lega in modo permanente a quest’ultimo, impedendo la sua azione di conversione degli Androgeni in Estrogeni. Il tasso di soppressione degli Estrogeni da parte dell’Exemestano varia dal 35% per l’Estradiolo (E2) al 70% per l’Estrone (E1).(46)
Grazie alla sua caratteristica di “inibitore selettivo”, l’Exemestano sembra non causare un rebound estrogenico dopo la sua cessazione. Nonostante ciò, un suo uso temporalmente protratto potrebbe (teoricamente) causare, similmente a quanto accade con l’uso di Letrozolo e Anastrozolo, un aumento dell’espressione dell’Enzima Aromatasi nonché un aumento del numero di ER come risposta adattativa.
Ovviamente, questa possibilità può interessare tutti gli AI con legame irreversibile (es. Formestano).
Queste sono semplici ipotesi nate da una riflessione sulle possibili cause e meccanismi che potrebbero (teoricamente) portare al manifestarsi di un rebound estrogenico con l’uso di tali composti. La letteratura scientifica, purtroppo, non ci aiuta a fare molta chiarezza sulla connessione AI/rebound estrogenico, sebbene esistono alcuni studi nei quali la cosa viene accennata.(47)
Aminoglutetimide
*Nota sull’Aminoglutetimide: inibendo l’enzima P450scc e agendo, di conseguenza, come inibitore della biosintesi di tutti gli ormoni steroidei, l’abuso di Aminoglutetimide può potenzialmente causare non solo un rebound estrogenico ma anche un rebound dei livelli di cortisolo. Lo stesso vale per il farmaco Trylostano.
Conclusioni
Basarsi per la maggior parte sui dati aneddotici è un azzardo, anche perché la maggior parte delle variabili soggettive in gioco rimangono celate. Banalmente, alcuni lamentano rebound estrogenici che alla fine non risultano legati all’uso del SERM o del AI ma alla loro (o del Preparatore) ignoranza, come quando cessano l’utilizzo di AAS, e di SERM e/o AI, senza preoccuparsi di svolgere un adeguata PCT convinti, magari, che un po’ di Mesterolone (Proviron) risolvi tutto. Infatti, la maggior parte dei casi di presunti rebound estrogenici SERM o AI dipendenti sono causati da una repentina cessazione d’uso di questi e di AAS, oppure da una PCT mal pianificata e/o che non ha dato i risultati sperati (vedi anche alterazione della Testosterone:Estradiolo ratio). Queste condizioni sono legate più che altro ad una alterazione del HPTA data dall’uso di AAS e non ad una presunta azione diretta del SERM e/o AI precedentemente utilizzati.
In conclusione, l’uso ponderato e consapevole è l’unica vera arma che l’atleta supplementato chimicamente (o il Preparatore che lo segue) ha per far si che ipotetici rebound non si manifestino.
Gabriel Bellizzi
Riferimenti:
1- “Definition of antiestrogen – NCI Dictionary of Cancer Terms, Definition of antiestrogen – NCI Dictionary of Cancer Terms”.,
2- Jump up ^ “antiestrogen” at Dorland’s Medical Dictionary
3- Jump up ^ Judi Lindsley Nath (2006). Using Medical Terminology: A Practical Approach. Lippincott Williams & Wilkins. pp. 977–. ISBN 978-0-7817-4868-1.
4- Dahlman-Wright K, Cavailles V, Fuqua SA, Jordan VC, Katzenellenbogen JA, Korach KS, Maggi A, Muramatsu M, Parker MG, Gustafsson JA (Dec 2006). “International Union of Pharmacology. LXIV. Estrogen receptors”. Pharmacological Reviews. 58 (4): 773–81. doi:10.1124/pr.58.4.8. PMID 17132854.
5- Levin ER (Aug 2005). “Integration of the extranuclear and nuclear actions of estrogen”. Molecular Endocrinology. 19 (8): 1951–9. doi:10.1210/me.2004-0390. PMC 1249516 . PMID 15705661.
6- Levin ER (Aug 2005). “Integration of the extranuclear and nuclear actions of estrogen”. Molecular Endocrinology. 19 (8): 1951–9. doi:10.1210/me.2004-0390. PMC 1249516 . PMID 15705661.
7- Wang C, Fu M, Angeletti RH, Siconolfi-Baez L, Reutens AT, Albanese C, Lisanti MP, Katzenellenbogen BS, Kato S, Hopp T, Fuqua SA, Lopez GN, Kushner PJ, Pestell RG (May 2001). “Direct acetylation of the estrogen receptor alpha hinge region by p300 regulates transactivation and hormone sensitivity”. The Journal of Biological Chemistry. 276 (21): 18375–83. doi:10.1074/jbc.M100800200. PMID 11279135.
8- Zivadinovic D, Gametchu B, Watson CS (2005). “Membrane estrogen receptor-alpha levels in MCF-7 breast cancer cells predict cAMP and proliferation responses”. Breast Cancer Research. 7 (1): R101–12. doi:10.1186/bcr958. PMC 1064104 . PMID 15642158.
9- Björnström L, Sjöberg M (Jun 2004). “Estrogen receptor-dependent activation of AP-1 via non-genomic signalling”. Nuclear Receptor. 2 (1): 3. doi:10.1186/1478-1336-2-3. PMC 434532 . PMID 15196329.
10- Lu Q, Pallas DC, Surks HK, Baur WE, Mendelsohn ME, Karas RH (Dec 2004). “Striatin assembles a membrane signaling complex necessary for rapid, nongenomic activation of endothelial NO synthase by estrogen receptor alpha”. Proceedings of the National Academy of Sciences of the United States of America. 101 (49): 17126–31. doi:10.1073/pnas.0407492101. PMC 534607 . PMID 15569929.
11- Kato S, Endoh H, Masuhiro Y, Kitamoto T, Uchiyama S, Sasaki H, Masushige S, Gotoh Y, Nishida E, Kawashima H, Metzger D, Chambon P (Dec 1995). “Activation of the estrogen receptor through phosphorylation by mitogen-activated protein kinase”. Science. 270 (5241): 1491–4. doi:10.1126/science.270.5241.1491. PMID 7491495.
12- Prossnitz ER, Arterburn JB, Sklar LA (Feb 2007). “GPR30: A G protein-coupled receptor for estrogen”. Molecular and Cellular Endocrinology. 265-266: 138–42. doi:10.1016/j.mce.2006.12.010. PMC 1847610 . PMID 17222505.
13- Otto C, Rohde-Schulz B, Schwarz G, Fuchs I, Klewer M, Brittain D, Langer G, Bader B, Prelle K, Nubbemeyer R, Fritzemeier KH (Oct 2008). “G protein-coupled receptor 30 localizes to the endoplasmic reticulum and is not activated by estradiol”. Endocrinology. 149 (10): 4846–56. doi:10.1210/en.2008-0269. PMID 18566127.
14- Harris HA, Albert LM, Leathurby Y, Malamas MS, Mewshaw RE, Miller CP, Kharode YP, Marzolf J, Komm BS, Winneker RC, Frail DE, Henderson RA, Zhu Y, Keith JC (Oct 2003). “Evaluation of an estrogen receptor-beta agonist in animal models of human disease”. Endocrinology. 144 (10): 4241–9. doi:10.1210/en.2003-0550. PMID 14500559.
15- Deroo BJ, Korach KS (Mar 2006). “Estrogen receptors and human disease”. The Journal of Clinical Investigation. 116 (3): 561–70. doi:10.1172/JCI27987. PMC 2373424 . PMID 16511588.
16- Riggs BL, Hartmann LC (Feb 2003). “Selective estrogen-receptor modulators — mechanisms of action and application to clinical practice”. The New England Journal of Medicine. 348 (7): 618–29. doi:10.1056/NEJMra022219. PMID 12584371.
17- Cameron JL, Cameron AM (20 November 2013). Current Surgical Therapy. Elsevier Health Sciences. pp. 582–. ISBN 978-0-323-22511-3.
18- Huang X, Aslanian RG (19 April 2012). Case Studies in Modern Drug Discovery and Development. John Wiley & Sons. pp. 392–394. ISBN 978-1-118-21967-6.
19- Pickar JH, Komm BS (Sep 2015). “Selective estrogen receptor modulators and the combination therapy conjugated estrogens/bazedoxifene: A review of effects on the breast”. Post Reproductive Health. 21 (3): 112–21. doi:10.1177/2053369115599090. PMID 26289836.
20- Mirkin S, Pickar JH (Jan 2015). “Selective estrogen receptor modulators (SERMs): a review of clinical data”. Maturitas. 80 (1): 52–7. doi:10.1016/j.maturitas.2014.10.010. PMID 25466304.
21- Desta Z, Ward BA, Soukhova NV, Flockhart DA (Sep 2004). “Comprehensive evaluation of tamoxifen sequential biotransformation by the human cytochrome P450 system in vitro: prominent roles for CYP3A and CYP2D6”. The Journal of Pharmacology and Experimental Therapeutics. 310 (3): 1062–75. doi:10.1124/jpet.104.065607. PMID 15159443.
22- Ahmad A, Shahabuddin S, Sheikh S, Kale P, Krishnappa M, Rane RC, Ahmad I (December 2010). “Endoxifen, a new cornerstone of breast cancer therapy: demonstration of safety, tolerability, and systemic bioavailability in healthy human subjects”. Clinical Pharmacology and Therapeutics. 88 (6): 814–7. doi:10.1038/clpt.2010.196. PMID 20981001.
23- Wang DY, Fulthorpe R, Liss SN, Edwards EA (Feb 2004). “Identification of estrogen-responsive genes by complementary deoxyribonucleic acid microarray and characterization of a novel early estrogen-induced gene: EEIG1”. Molecular Endocrinology. 18 (2): 402–11. doi:10.1210/me.2003-0202. PMID 14605097.
24- Kuhl H (2005). “Pharmacology of estrogens and progestogens: influence of different routes of administration”. Climacteric. 8 Suppl 1: 3–63. doi:10.1080/13697130500148875. PMID 16112947.
25- Shang Y, Hu X, DiRenzo J, Lazar MA, Brown M (Dec 2000). “Cofactor dynamics and sufficiency in estrogen receptor-regulated transcription”. Cell. 103 (6): 843–52. doi:10.1016/S0092-8674(00)00188-4. PMID 11136970.
26- Massarweh S, Osborne CK, Creighton CJ, Qin L, Tsimelzon A, Huang S, Weiss H, Rimawi M, Schiff R (Feb 2008). “Tamoxifen resistance in breast tumors is driven by growth factor receptor signaling with repression of classic estrogen receptor genomic function”. Cancer Research. 68 (3): 826–33. doi:10.1158/0008-5472.CAN-07-2707. PMID 18245484.
27- Hurtado A, Holmes KA, Geistlinger TR, Hutcheson IR, Nicholson RI, Brown M, Jiang J, Howat WJ, Ali S, Carroll JS (Dec 2008). “Regulation of ERBB2 by oestrogen receptor-PAX2 determines response to tamoxifen”. Nature. 456 (7222): 663–6. Bibcode:2008Natur.456..663H. doi:10.1038/nature07483. PMC 2920208 . PMID 19005469.
28- Osborne CK, Bardou V, Hopp TA, Chamness GC, Hilsenbeck SG, Fuqua SA, Wong J, Allred DC, Clark GM, Schiff R (Mar 2003). “Role of the estrogen receptor coactivator AIB1 (SRC-3) and HER-2/neu in tamoxifen resistance in breast cancer”. Journal of the National Cancer Institute. 95 (5): 353–61. doi:10.1093/jnci/95.5.353. PMID 12618500.
29- “New Mechanism Predicts Tamoxifen Response: PAX2 gene implicated in tamoxifen-induced inhibition of ERBB2/HER2-mediated tumor growth”. http://www.modernmedicine.com. 2008-11-13. Archived from the original on 2011-07-14. Retrieved 2008-11-14.
30- “Study sheds new light on tamoxifen resistance”. News. CORDIS News. Archived from the original on 2009-02-20. Retrieved 2008-11-14.
31- Liu J, Flockhart PJ, Lu D, Lv W, Lu WJ, Han X, Cushman M, Flockhart DA (2013). “Inhibition of cytochrome p450 enzymes by the e- and z-isomers of norendoxifen”. Drug Metab. Dispos. 41 (9): 1715–20. doi:10.1124/dmd.113.052506. PMC 3876808 . PMID 23824607.
32- Chemical muscle enhancement. Report. B.B. desk reference. di Author L. Rea. Pag. 106.
33- https://www.rdmag.com/article/2014/12/journal-watch-tamoxifen-news-again
34- https://www.ncbi.nlm.nih.gov/pubmed/14687597
35- Bryant HU (2001). “Mechanism of action and preclinical profile of raloxifene, a selective estrogen receptor modulation”. Rev Endocr Metab Disord. 2 (1): 129–38. PMID 11704975.
36- Thiebaud D, Secrest RJ (2001). “Selective estrogen receptor modulators: mechanism of action and clinical experience. Focus on raloxifene”. Reprod. Fertil. Dev. 13 (4): 331–6. PMID 11800172.
37- Gizzo S, Saccardi C, Patrelli TS, Berretta R, Capobianco G, Di Gangi S, Vacilotto A, Bertocco A, Noventa M, Ancona E, D’Antona D, Nardelli GB (2013). “Update on raloxifene: mechanism of action, clinical efficacy, adverse effects, and contraindications”. Obstet Gynecol Surv. 68 (6): 467–81. doi:10.1097/OGX.0b013e31828baef9. PMID 23942473.
38- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5739193/
39- “Entrez Gene: CYP19A1 cytochrome P450, family 19, subfamily A, polypeptide 1”.
40- Toda K, Shizuta Y (April 1993). “Molecular cloning of a cDNA showing alternative splicing of the 5′-untranslated sequence of mRNA for human aromatase P-450”. European Journal of Biochemistry. 213 (1): 383–9. doi:10.1111/j.1432-1033.1993.tb17772.x. PMID 8477708.
41- Czajka-Oraniec I, Simpson ER (2010). “Aromatase research and its clinical significance”. Endokrynologia Polska. 61 (1): 126–34. PMID 20205115.
42- Gasnier C, Dumont C, Benachour N, Clair E, Chagnon MC, Séralini GE (August 2009). “Glyphosate-based herbicides are toxic and endocrine disruptors in human cell lines”. Toxicology. 262 (3): 184–91. doi:10.1016/j.tox.2009.06.006. PMID 19539684.
43- Mokbel K (2002). “The evolving role of aromatase inhibitors in breast cancer”. Int. J. Clin. Oncol. 7 (5): 279–83. doi:10.1007/s101470200040 (inactive 2017-01-15). PMID 12402060.
44- Simpson ER (2003). “Sources of estrogen and their importance”. J. Steroid Biochem. Mol. Biol. 86 (3–5): 225–30. doi:10.1016/S0960-0760(03)00360-1. PMID 14623515.
45- Leder BZ, Rohrer JL, Rubin SD, Gallo J, Longcope C (March 2004). “Effects of aromatase inhibition in elderly men with low or borderline-low serum testosterone levels”. J. Clin. Endocrinol. Metab. 89 (3): 1174–80. doi:10.1210/jc.2003-031467. PMID 15001605.
46- Mauras, N; Lima, J; Patel, D; Rini, A; Di Salle, E; Kwok, A; Lippe, B (2003). “Pharmacokinetics and Dose Finding of a Potent Aromatase Inhibitor, Aromasin (Exemestane), in Young Males”. The Journal of Clinical Endocrinology & Metabolism. 88 (12): 5951–6. doi:10.1210/jc.2003-031279. PMID 14671195.
47- https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3263690/
L’uso della Gonadotropina Corionica Umana (hCG) è largamente diffuso nell’ambiente culturistico. Usata principalmente per ripristinare la funzionalità gonadale in seguito all’uso di AAS, questo peptide vede la sua applicazione anche durante l’uso di questa classe di farmaci (ciclo, Bridge o TRT), o di altri composti causanti un ciclo di feedback negativo dell’HPTA (vedi SARM), al fine di prevenire l’istaurarsi di una disfunzione testicolare. Fin dai primi anni della sua applicazione su soggetti di sesso maschile, l’hCG è stato oggetto di speculazioni riguardo la possibilità o meno che il suo uso possa portare ad una desensibilizzazione delle cellule di Leydig con conseguente sviluppo di ipogonadismo ipergonadotropo. Il seguente articolo è volto a riportare le caratteristiche del hCG, le sue possibili applicazioni e, in modo approfondito, fare maggiore chiarezza sulla questione legata alla possibile desensibilizzazione hCG-dipendente.
hCG: storia, usi clinici e off-label
Gonadotropina Corionica Umana
L’hCG (Human chorionic gonadotropin) o Gonadotropina Corionica è un ormone polipeptidico prodotto dall’embrione all’inizio della seconda settimana di sviluppo, in particolare dalle cellule del sinciziotrofoblasto, un tessuto epiteliale monostratificato posto nella porzione profonda del cito-sinciziotrofoblasto, subito dopo l’impianto nell’endometrio. La molecola di hCG è un eterodimero, composto da due subunità (α e β). La subunità α ha struttura identica a quella delle altre gonadotropine (LH e FSH), mentre la subunità β è specifica di ciascun ormone. Per questo motivo, i metodi di dosaggio dell’hCG utilizzano anticorpi diretti contro la subunità β dell’hCG.
Più specificatamente, la Gonadotropina Corionica è una glicoproteina oligosaccaridica composta da 244 aminoacidi. La subunità α è lunga 92 aminoacidi ed è identica a quella dell’Ormone Luteinizzante (LH), dell’Ormone Follicolo-Stimolante (FSH) e dell’Ormone Stimolante la Tiroide (TSH). Come già accennato, la subunità beta è unica per l’hCG.
L’hCG è quindi un analogo del LH, l’ormone prodotto dall’ipofisi che stimola la produzione di ormoni sessuali nei testicoli o nelle ovaie. L’hCG si lega e attiva lo stesso recettore dell’LH ed è ugualmente efficace nello stimolare la produzione di Testosterone negli uomini e di Estrogeni nelle donne.
La Gonadotropina Corionica venne isolata ed identificata per la prima volta nel 1920 (1) venendo in seguito classificata come un ormone della gravidanza circa otto anni dopo.(2) La prima preparazione farmaceutica contenente Gonadotropina Corionica si presentava sotto forma di estratto pituitario animale, il quale venne sviluppato come prodotto commerciale dalla Organon. La Organon introdusse nel mercato l’estratto nel 1931, con il nome commerciale di Pregnon. Una controversia sui marchi obbligò la compagnia a cambiare il nome Pregnyl, che raggiunse il mercato nel 1932. Il Pregnyl è attualmente venduto dalla MSD–Organon, anche se il principio attivo non è più estratto dalla pituitaria animale. Nel 1940 furono introdotte tecniche di produzione che consentivano di ottenere l’ormone filtrando e purificando l’urina delle donne incinta, e alla fine degli anni ’60 questa tecnica di produzione fu adottata da tutti i produttori che avevano usato precedentemente gli estratti animali. Nel corso degli anni i processi di produzione sono stati perfezionati, ma l’hCG è ottenuta essenzialmente nello stesso modo oggi come lo era decenni fa. Nonostante i preparati moderni siano di origine biologica, si afferma che i rischi di contaminanti biologici siano bassi (sebbene non possano essere completamente esclusi).
Al principio della sua applicazione clinica, gli usi indicati per le preparazioni a base di Gonadotropina Corionica erano molto più ampi di quanto non lo siano attualmente. La letteratura inerente al composto degli anni ’50 e ’60 raccomandava l’uso di questo farmaco per, tra le altre cose, il trattamento del sanguinamento uterino e dell’amenorrea, la sindrome di Froehlich, il criptochismo, la sterilità femminile, l’obesità, la depressione e l’impotenza maschile. Un buon esempio degli ampi usi della Gonadotropina Corionica è illustrato nel preparato Glukor, che fu descritto nel 1958 come “Tre volte più efficace del Testosterone. Per i giovani stanchi dal climaterio maschile. Per vecchi stanchi dalla senilità maschile. Benefici nell’impotenza, angina e malattia coronarica, neuropsicosi, prostatite, [e] miocardite.” Tali raccomandazioni, tuttavia, riflettono un’era meno strettamente regolata dall’agenzia governativa e meno dipendente da studi cliniche comprovati. Oggi, le indicazioni approvate dalla FDA per l’uso del hCG sono limitate al trattamento dell’ipogonadotropismo ipogonadico e del criptocridismo negli uomini e alla sterilità anovulatoria nelle donne.
Dr. A.T.W. Simeons
L’hCG non ha alcuna attività significativa di stimolo della tiroide. Questo necessita di essere specificato dato che l’hCG è stata ampiamente usata in passato per il trattamento dell’obesità. Questa applicazione d’uso sembra che sia divenuta popolare nel 1954, dopo la pubblicazione di un articolo del Dr. A.T.W. Simeons nel quale sosteneva che la Gonadotropina Corionica era un’aggiunta efficace alla dieta. Secondo lo studio, i pazienti sono stati in grado di sopprimere efficacemente l’appetito seguendo una dieta con marcata restrizione calorica abbinata alla somministrazione di hCG. Soprannominata la dieta Simeons, le persone in tutti gli Stati Uniti si sottoposero presto a severe restrizioni caloriche (500 Kcal al giorno) e iniezioni di hCG. Poco dopo, l’ormone stesso divenne il coadiuvante principale per la perdita di grasso. Infatti, nel 1957 si diceva che l’hCG era il farmaco più comunemente prescritto per la perdita di peso. Indagini più recenti e complete, tuttavia, confutano l’esistenza di qualsiasi vantaggio anoressizzante o metabolico dato dall’uso di hCG.(3) Nel 1962, il Journal of American Medical Association aveva già avvertito i consumatori circa la dieta Simeons inclusiva di hCG, affermando che la grave restrizione calorica tipica di tale protocollo dimagrante (che si rifletteva in un accentuato catabolismo del tessuto magro) era più pericolosa dell’obesità stessa. Nel 1974, la FDA aveva raccolto abbastanza dichiarazioni sull’uso del hCG per la perdita di grasso che fece inserire una dichiarazione in merito nel bugiardino dei prodotti contenenti l’ormone, nella quale affermava che non vi erano dimostrazioni sulla presunta efficacia nella perdita di peso data dalla somministrazione di hCG in concomitanza con regimi alimentari ipocalorici. Questo avvertimento è tutt’oggi presente su tutti prodotto venduti negli Stati Uniti. Nonostante questo avvertimento e prove che confutano l’efficace di tale pratica, alcune cliniche promuovono ancora l’uso di hCG per la perdita di peso.
La Gonadotropina Corionica Umana è oggi una preparazione farmaceutica molto popolare, poiché rimane una parte indispensabile della terapia di ovulazione per molti casi di infertilità femminile. Sebbene la forma di hCG sintetizzata tramite la tecnica del DNA ricombinante sia stata introdotta sul mercato negli ultimi anni, l’ampia offerta e il basso costo dell’hCG biologico continuano a renderlo un prodotto di base per gli usi clinici e off-label.
Quando vengono somministrati AAS (o SARM), i livelli di LH diminuiscono rapidamente. Il calo o l’assenza del rilascio ipofisario di LH, e suo consequenziale segnale, induce un calo o interruzione dell’attività testicolare (la quale, ovviamente, si riflette negativamente sulla sintesi di Testosterone) che causa la rapida insorgenza dell’atrofia testicolare. Questa degenerazione testicolare inizia con una riduzione del volume delle cellule di Leydig, seguita da una riduzioni rapida del Testosterone Intra-Testicolare (ITT), dei perossisomi e del fattore insulino-simile 3 (INSL3) – Tutti bio-marcatori e fattori importanti per una corretta funzione testicolare e biosintesi di Testosterone.
Tuttavia, questa degenerazione testicolare viene trattata dai Bodybuilder supplementari chimicamente con la somministrazione di hCG, in special modo all’uscita di un ciclo e per il periodo iniziale della PCT.
Tutte, o quasi tutte, le esperienze pratiche con questo farmaco nel Bodybuilding avvengono con l’uso del hCG biologico (estratto dalle urine di donne gravide), che viene generalmente venduto in vial contenenti polvere liofilizzata da ricombinare con acqua fisiologica o batteriostatica, con un contenuto che va dalle 250 alle 10.000UI per vial.
Il dosaggio clinico di hCG per trattare i casi di ipogonadismo ipogonadotropo è stato tradizionalmente di 5000UI per iniezione. Prima del 1998, la dose tipicamente utilizzata nel bodybuilding per il ripristino della funzione testicolare era la medesima. Di conseguenza, trattandosi di un quantitativo molto elevato, è stato per molto tempo considerato un farmaco di non facile gestione e dagli effetti collaterali, presunti o tali, che destavano non poca preoccupazione (vedi desensibilizzazione delle cellule di Leydig che tratterò più avanti).
Successivamente, venne introdotto l’uso di un dosaggio più basso con un limite di 1500UI per ogni singola iniezione, con una preferenza di dosaggio non superiore alle 1000UI, e con l’uso consigliato di un dosaggio pari a 500UI a somministrazione.
Molti Preparatori danno come raccomandazione quella di non superare le 500UI per ogni somministrazione, poiché non è stato riscontrato alcun vantaggio aggiuntivo nell’utilizzare un dosaggio singolo superiore a questo, a condizione che le iniezioni siano ragionevolmente frequenti (ogni 2-4 giorni).
L’intervallo di dosaggio settimanale comunemente consigliato è compreso tra circa le 700 e le 1750UI. Le dosi di esempio sono 100-250UI al giorno, 250-500 UI a giorni alterni o 250-500UI da tre volte a settimana a somministrazioni distanziate l’una dall’altra da quattro giorni.
Con tali dosaggi sono stati seguiti un numero molto elevato di individui per diversi anni e con eccellenti risultati, e la ricerca scientifica sembra aver convalidato l’utilità del mantenersi all’interno di queste dosi. Come misurato dai livelli intratesticolari di testosterone, questo livello di dosaggio massimizza i risultati. Semplicemente non risulta conveniente la somministrazione di dosi maggiori.
Si raccomandano generalmente iniezioni multiple settimanali dal momento che l’emivita del hCG è di circa 36 ore. Iniezioni meno frequenti comportano uno scarso mantenimento dei livelli ematici.
Prima del 1996, l’uso tradizionale del hCG era quello di inserirla post-ciclo con lo scopo di ripristinare una funzionalità testicolare ottimale. Ma tale pratica non risulta pienamente ottimale dal momento che rallenta comunque i processi di recupero dell’HPTA. Infatti, il tempo medio di recupero della funzionalità testicolare con l’uso del hCG risulta essere in media di 4-8 settimane. Di conseguenza, la scelta migliore, in contesti nei quali i cicli durano più di quattro settimane e/o quando il ciclo viene seguito da un “Bridge” o TRT, l’uso del hCG durante il ciclo permette di conservare una buona attività testicolare permettendo, per esempio, all’atleta in uscita da un ciclo di accelerare i processi di recupero dell’HPTA dal momento che, così facendo, evita quel periodo transitorio (e potenzialmente controproducente) tra la fine del ciclo ed il ripristino di una corretta funzionalità testicolare.
Nei contesti sopra citati, la hCG viene somministrata durante il ciclo con varianti temporali che vanno dalla seconda settimana alla quarta (dipendente dalla durata complessiva del ciclo e da ciò che l’atleta farà nel post ciclo). I dosaggi mediamente utilizzati sono 100 UI al giorno, 200 UI a giorni alterni o 250UI da 3 volte a settimana a ogni 4 giorni.
Un’altra pratica d’uso del hCG è quella di inserirla durante i cicli che non contemplano l’uso di AAS soggetti ad aromatizzazione. Con il solo uso di AAS non aromatizzabili, i livelli di estrogeni diminuiscono in modo anomalo in seguito alla sottoregolazione/soppressione del Testosterone endogeno e la consequenziale diminuzione dei substrati soggetti all’aromatizzazione. Questa condizione interferisce con l’anabolismo, la libido, l’umore, la funzione articolare e, sul lungo termine, la salute cardiovascolare. Un modo ovvio per risolvere questo problema è quello di includere almeno una piccola quantità di uno AAS aromatizzabile (vedi base terapeutica di Testosterone). In questo caso i dosaggi di hCG tipicamente utilizzati sono compresi nella fascia altra d’intervallo del dosaggio efficace suggerito (500UI a giorni alterni). La risultante sarà una sintesi di Testosterone endogeno e Estradiolo.
Terminate le dovute precisazioni sul hCG adesso possiamo trattare l’argomento centrale di questo articolo…
hCG e possibile desensibilizzazione (?)
La questione sulla possibilità secondo cui l’uso prolungato di hCG possa portare ad una condizione di ipogonadismo ipergonadotropo è tutt’ora dibattuta. L’utilizzatore deve comunque tenere a mente che il dosaggio di tale composto deve essere attentamente calibrato in specie con somministrazioni prolungate, poiché alti livelli di hCG possono anche causare un aumento dell’espressione dell’aromatasi testicolare (con conseguente innalzamento dei livelli di estrogeni), (4). Esistono studi piuttosto datati, e svolti per la maggior parte sui ratti, che riportano il verificarsi della desensibilizzazione testicolare al LH in seguito a somministrazione di alti dosaggi e per lunghi periodi di tempo.(5) Il farmaco in questione può effettivamente avere il potenziale di indurre ipogonadismo primario se usato impropriamente, peggiorando notevolmente, non migliorando, la funzionalità testicolare.
I protocolli d’uso di hCG che contemplano la somministrazione di dosi pari a 250UI per via sottocutanea ogni 3 o 4 giorni con una dose massima di 500UI, sviluppati dal Dr. John Crisler, una figura ben nota nel campo dell’Anti-Aging e della terapia ormonale sostitutiva, vengono spesso utilizzati dai soggetti in Terapia Sostitutiva del Testosterone (TRT). L’atrofia testicolare per i pazienti in TRT è un disturbo cosmetico comune. Il programma di somministrazione di hCG del Dr. Crisler è progettato per risolvere questo problema con un uso a lungo termine senza causare l’ipotetica desensibilizzazione. Coloro i quali sono interessati a gestire il timing di somministrazione del hCG con precisione in relazione ad una TRT, il dott. Crisler raccomanda quanto segue: “… i miei pazienti in TRT con Testosterone Cypionato ora somministrano la loro dose di hCG di 250IU nei due giorni precedenti l’iniezione intramuscolare (Testosterone Cypionato NdR.). Tutti i pazienti somministrano la loro dose di hCG per via sottocutanea e il dosaggio può essere aggiustato secondo necessità (devo ancora vedere una necessità di dosaggio superiore alle 350 UI per somministrazione) … Quei pazienti in TRT che preferiscono usare un Testosterone transdermico, o anche Testosterone orale (sebbene io non sia favorevole a ciò) , somministrano la loro dose di hCG ogni tre giorni. ”
Il Dr. John Crisler afferma che è importante non somministrare più di 500UI di hCG in un dato giorno. Egli infatti afferma che vi è solo una quantità massima di stimolazione, e il superamento di questo dosaggio non solo è uno spreco, ma ha conseguenze negative importanti. Dosi più elevate stimolano eccessivamente l’aromatasi testicolare, che aumenta in modo inappropriato i livelli di estrogeni portando alla comparsa di effetti collaterali tipici del iperestrogenemia. Il Dr. Crisler continua dicendo che dosi superiori a quella sopra indicata (500UI) causino anche la desensibilizzazione delle cellule di Leydig verso LH inducendo quindi all’ipogonadismo primario. Egli ribadisce che 250IU ogni 2-4 giorni sia una dose efficace e sicura. Dopotutto, stiamo semplicemente sostituendo ciò che è stato inibito.
Il Dr. Scalley, dal canto suo, critica la posizione del Dr. John Crisler affermando che, la somministrazione dell’hCG per due giorni consecutivi non ha senso, inoltre la dose è omeopatica (inutile). Inoltre, il Dr. Scalley ritiene che, nonostante il Dr. Crisler qualifichi le sue affermazione ricollegandosi a determinati studi, l’errore sta nel considerare come assodato che le dosi più elevate di quelle che consiglia causino la desensibilizzazione. Il Dr. Crisler sembra mancare di una comprensione corretta della letteratura.
Scalley riporta che la desensibilizzazione hCG-dipendente si può potenzialmente verificare in caso di somministrazione prolungata di 5.000UI (cinquemila). Ma, anche in questo caso l’incidenza non è universalmente osservata. C’è anche da aggiungere che il problema della desensibilizzazione non è quasi mai stato osservato nella pratica clinica.
Gli studi solitamente menzionati non danno in realtà alcun supporto a dimostrazione che la desensibilizzazione si verifichi con dosi superiori alle 500UI o che l’uso di 250 UI X2 volte a settimana sia una terapia utile. Se ci si pensaun attimo, qual è lo scopo dell’uso di hCG per due giorni di seguito? Questa pratica risulta completamente bizzarra. Come prima cosa, sfido chiunque a riportare la letteratura (articolo o citazioni) a sostegno del suo trattamento (del Dr. Crisler). Se Crisler è così sicuro di sé, perché non cita alcuna pubblicazione a supporto della sua terapia o, meglio, pubblichi i risultati del trattamento.
Innanzitutto, lo studio che spesso viene citato a sostegno delle tesi del Dr. Crisler (6) valuta il Testosterone Intratesticolare (ITT) e questo, di per se, non è di poca importanza. I partecipanti a questo studio sono stati trattati con Testosterone Enantato (TE), 200 mg alla settimana, per la soppressione rapida della gonadotropina in combinazione con una dose variabile di hCG, somministrata sottocute ogni 2 giorni per 3 settimane: 0 (placebo salino), 125, 250 o 500 UI hCG. Il gruppo placebo è servito come gruppo di controllo. [Nota: la differenza sostanziale è che, anche se lo studio supporta Crisler, il dosaggio è molto diverso da quello da lui raccomandato.]
Quindi, quello che lo studio ci offre sono soggetti di sesso maschile con elevati livelli di Testosterone per via di iniezioni settimanali di 200mg di Testosterone Enantato. La loro produzione endogena di Testosterone è completamente soppressa (teoricamente) come le loro gonadotropine. Il ITT risulta quindi soppresso a causa dell’inibizione delle gonadotropine date dalla somministrazione di Testosterone Enantato. I ricercatori hanno scoperto che ogni dose di hCG (125, 250 e 500 UI) riportava la concentrazione di ITT alla normalità. Si da il caso però che in un maschio normale con un normale livello di Testosterone serico il suo ITT sarà normale. Tutto questo studio è stato semplicemente prendere un maschio normale e sostituire il suo Testosterone con del Testosterone esogeno per poi somministrargli hCG come sostituto del suo LH.
L’unica cosa che può essere salvata di questo studio, è che può essere istruttiva per chi usa hCG a basse dosi in on-cicle, in “Bridge” o in TRT. Più precisamente ci dice qualcosa sulla terapia con hCG mentre si usa un dosaggio “simil-TRT”.
Nello studio risulta interessante esaminare i dati sulle variazioni seriche di Testosterone con ciascuna dose di hCG. I soggetti presi in esame hanno usato una dose contenuta (seppur fisiologicamente alta) di Testosterone Enantato, creando una situazione che per certi versi riproduce quella di un individuo che usa hCG in TRT. Il risultato è stato che la dose di hCG da 125UI a giorni alterni non ha avuto effetti sul Testosterone serico. Le due dosi più elevate (250-500UI) hanno alzato i livelli di Testosterone nel siero al di sopra del normale.
Non ci sono dati individuali (sempre motivo di sospetto quando si esamina la letteratura) e non sono riportati livelli significativi. L’analisi del grafico dello studio riportato di seguito, tuttavia, mostra che il livello di Testosterone del siero non era significativamente diverso dal controllo fino al giorno 21[altra nota a discredito delle affermazioni del Dr. Crisler].
Ci sono quindi molti possibili errori nell’analisi dello studio appena discusso. Dal momento che non ci mostra un analisi sufficientemente accurata tale da permetterci di identificare una soglia di dosaggio che porti alla desensibilizzazione delle cellule di Leydig.
Si può ipotizzare che la modalità di somministrazione dell’hCG nei due giorni precedenti l’iniezione settimanale di Testosterone (come indicato nel protocollo del Dr Crisler) serva da teorico “supporto” al calo della soglia ematica di quest’ultimo. Se si legge la letteratura disponibile sugli effetti dell’hCG, il rialzo dei livelli di Testosterone serico si manifestano in modo significativo a circa 48-72 ore dopo la somministrazione del peptide. Questo dosaggio concentrato in due giorni non da reali vantaggi sulla funzionalità testicolare. Quindi, fino a dimostrazione contraria, le ipotesi del Dr. Crisler sulla somministrazione ottimale di hCG, per effetti e sicurezza riguardo la desensibilizzazione testicolari, non sono altro che opinabili speculazioni.
Per tutti gli scopi pratici, la desensibilizzazione delle cellule di Leydig hCG-dipendente praticamente non sussiste all’interno del quadro clinico, sebbene rimanga una possibilità con l’uso di dosi elevate e per un lungo periodo di tempo (>5000UI)
17 alfa-hydroxyprogesterone
Esiste uno studio che, seppur “isolato”, ritengo sia interessante per farsi un idea delle variabili e della differenza tra possibilità universalmente riscontrate e possibilità di bassa o scarsa incidenza. Si tratta di uno studio nel quale si è osservato che la somministrazione di Tamoxifene in maschi sani ha causato una riduzione dell’accumulo del 17 α-hydroxyprogesterone hCG-indotto.(7)
In questo studio, la somministrazione per via intramuscolare di 1500 UI di hCG al giorno per 3 giorni ha indotto un accumulo transitorio di 17 α-hydroxyprogesterone (17 OHP) rispetto al Testosterone (T) in uomini normali, raggiungendo il massimo nelle 24 ore successive la prima iniezione (rapporto 17 OHP / T, 1,7 +/- 0,3 volte il basale, P <0,01). La somministrazione simultanea di hCG e Tamoxifene (20 mg due volte al giorno) ha quasi completamente soppresso il blocco steroidogenico indotto dal hCG e osservato con il 17 OHP:T ratio (rapporto 17 OHP-T a 24 ore, 1,1 +/- 0,1 volte il basale; 0,01 vs hCG da solo). Questi dati suggeriscono indirettamente che, nell’uomo, la lesione steroidogenica indotta dal hCG potrebbe essere mediata attraverso il suo effetto stimolante sugli estrogeni.
Un altro studio svolto sulla falsariga del precedente, ma ad una distanza di undici anni, ha osservato l’effetto del Tamoxifene sulla risposta testicolare al hCG. (8) Se si legge con attenzione il presente studio, anche alla luce di quanto affermato pocanzi, si riesce ad avere un quadro molto più chiaro sulla questione.
Tamoxifene
In questo studio è stato osservato l’effetto del Tamoxifene (Tx) in concomitanza con la somministrazione acuta e cronica di hCG in pazienti con ipogonadismo ipogonadotropo (HH) e in uomini normali. Un test con hCG (5000 UI hCG) è stato svolto prima e dopo due mesi di somministrazione di hCG (2000 UI di hCG tre volte a settimana) e dopo due mesi di hCG + Tx (2000 UI hCG tre volte a settimana più 20 mg/die di Tamoxifene). I campioni di sangue sono stati prelevati 24 e 72 ore prima e dopo ogni test per determinare i livelli di Testosterone , Estradiolo, 17OHP e SHBG. Il Testosterone è aumentato solo nel gruppo HH con entrambi i trattamenti (X +/- SEM: basale: 97,9 +/- 19,7; hCG: 237,7 +/- 43,2; hCG +/- Tx: 204,7 +/- 10,7 ng / 100 ml). Il 17OHP è aumentato con la somministrazione di hCG da solo, ma non con hCG + Tx in entrambi i gruppi. Il rapporto Estradiolo, SHBG e 17OHP / Testosterone non è cambiato dopo i trattamenti. In risposta al hCG il Testosterone è aumentato 24 ore dopo la somministrazione in ogni test. Il rapporto 17OHP / Testosterone è salito dopo 24 ore nel primo e nel secondo test, ma nel terzo test non è cambiato. Questi risultati supportano il ruolo del Estradiolo nella desensibilizzazione delle cellule di Leydig indotto da una somministrazione acuta di hCG. Tuttavia, l’associazione di Tx non migliora i livelli serici di Testosterone, suggerendo che l’Estradiolo potrebbe non essere il fattore unico coinvolto nei meccanismi di desensibilizzazione testicolare.
Sembrerebbe, quindi, che il fattore determinante legato ad una possibile desensibilizzazione o sottoregolazione della funzionalità testicolare (in particolar modo in riferimento alle cellule di Leydig) sia la dose in acuto e, soprattutto, in cronico. Per ciò che concerne la dose utile questa è invece determinata dal limite fisiologico di stimolo della secrezione di Testosterone che, negli uomini sani con una sensibilità testicolare normale, si è visto corrispondere ad una dose di sole 250UI, con ulteriori aumenti minimi ottenuti con 500UI a 5000UI.
Conclusioni
Sebbene, come già accennato, la questione non sia del tutto chiarita, le informazioni riportate in questo articolo possono senz’altro permettere una pratica d’uso del hCG “sicura” e, soprattutto, intelligente nel contesto di una preparazione farmacologica. Risulta abbastanza chiaro che iniezione intramuscolare o sottocutanee di hCG a dosi di 100-200UI al giorno, 200-250 UI a giorni alterni o 250-500UI da tre volte a settimana a una somministrazione ogni 4 giorni, risultano pienamente efficace per evitare la disfunzione e seguente atrofia testicolare durante l’uso di AAS (o SARM) mantenendone una buona funzionalità in mancanza di stimoli dati dal LH. Dosi superiori a quelle riportate non offrono ulteriori vantaggi. Si tenga inoltre bene a mente che la desensibilizzazione delle cellule di Leydig può manifestarsi con maggiore facilità in una situazione di mancanza di segnale dato dall’LH, condizione che viene spesso osservata in quegli atleti che non usano hCG durante i cicli. Tale desensibilizzazione, però, risulta più semplice da trattare rispetto ad una desensibilizzazione indotta da uno stimolo eccessivo delle cellule di Leydig che, in casi cronici, obbliga il soggetto colpito a doversi sottoporre a trattamento con Testosterone esogeno (TRT).
Quindi, in definitiva, le nozioni base da tenere bene a mente sono:
Uso di dosi e tempi di somministrazione utili allo scopo prefissato (evitare la disfunzione testicolare e avere un ottimale stimolo della biosintesi di Testosterone)
Iniziare la somministrazione di hCG durante il ciclo (tempo variabile dalle 2 alle 4 settimane dall’inizio del ciclo e determinato dalle scelte future al ciclo [PCT, Bridge o TRT]
Non eccedere le 500UI a giorni alterni (principalmente perché dosi più elevate non portano vantaggi considerevoli)
Un’altra nota che mi sento di aggiungere è in riferimento alla ricombinazione del contenuto delle vial e delle procedure per la sterilità del prodotto. Le vial di hCG dovrebbero essere ricostituite con una quantità di soluzione acquosa (sterile o batteriostatica) basata sul quantitativo effettivo in UI della vial. Ad esempio, una vial da 5000UI può essere convenientemente ricombinata con 2,5ml d’acqua. Ciò fornisce una soluzione di 2000IU/ml , che consente un facile calcolo del dosaggio necessario. Ad esempio, una dose di 200 UI richiederebbe quindi l’aspirazione di 0,1mL di soluzione, che sarebbe contrassegnata con “10 UI” su una siringa da insulina.
Se la capacità della vial lo consente, è possibile aggiungere 5,0ml di acqua in una vial da 5000UI. La soluzione risultante sarebbe ovviamente di 1000IU/mL, consentendo un calcolo ancora più semplice del dosaggio necessario.
L’iniezione può essere eseguita intramuscolarmente o sottocute in base alle preferenze personali.
Le vial di hCG non ricostituite devono essere conservate in frigo. Sebbene possano essere spedite a temperatura ambiente. Le vial ricostituite devono sempre essere conservate in frigo; tuttavia, se una vial viene accidentalmente lasciata a temperatura ambiente per un giorno, il principio attivo non subirà alcun deterioramento.
Cosa molto importante quando si manipola l’hCG è quella di impiegare corrette procedure per mantenere la sterilità della vial e della soluzione ivi contenuta. La membrana di gomma deve sempre essere pulita accuratamente con alcool e l’ago deve essere sterile. Il peptide acquoso, o in questo caso le soluzioni glicopeptidiche possono supportare la crescita batterica molto più di quanto possano fare le soluzioni oleose, quindi è raccomandata la massima cura della sterilità del prodotto. Se si nota un intorpidimento della soluzione è consigliabile che il prodotto non venga utilizzato.
Gabriel Bellizzi
Riferimenti:
Exogenous stimulation of corpus luteum formation in the rabbit; influence of extracts of human placenta, decidua, fetus, hydatid mole and corpus luteum on the rabbit gonad. Hirose T 1920 J Jpn Gynecol Sot 16:1055.
Die Schwangerschaftsdiagnose ausdem Ham durch Nachweis des Hypophysenvorderlappen-hormone. II. Pracktishe und theoretische Ergebnisse aus den hamuntersuchungen. Ascheim S, Zondek B 1928 Klin Wochenschr 7:1453-1457.
Un maggior utilizzo di AAS da parte di un atleta si traduce in una maggiore probabilità di avere problemi sessuali durante i periodi di non utilizzo. E no, la PCT non sembra rendere esenti da tali possibili problemi. I ricercatori del American Mayo hospital hanno osservato tale effetto in seguito ad uno studio svolto su diverse centinaia di Bodybuilder supplementati farmacologicamente.(1)
I ricercatori hanno contattato 231 utilizzatori di AAS maschi, attraverso nove forum di bodybuilding, i quali erano pronti a rispondere a domande online sulla loro salute sessuale e sul loro uso di AAS. Per rilevare eventuali disfunzioni erettili, i ricercatori hanno utilizzato una versione abbreviata del questionario IEEF-5. Potete trovarne una copia qui. Gli uomini che totalizzano un punteggio pari o maggiore di 22 non hanno nulla di cui lamentarsi sessualmente parlando, mentre gli uomini che ottengono un punteggio di 17-21 soffrono di un lieve disturbo erettile.
I partecipanti hanno ottenuto un punteggio leggermente superiore sull’IEEF-5 se erano sottoposti ad iniezioni superiori ai 600mg di Testosterone a settimana, se usavano anche un AAS orale o un anti-estrogeno e se erano in buone condizioni di salute. Non molto sorprendente come risultato.
127 utilizzatori hanno affermato che nei loro periodi “off” la libido diminuiva. Questo però non è stato riportato da altri 94 utilizzatori. Quando i ricercatori hanno messo a confronto questi due gruppi, hanno notato che il rischio che si verifichi una riduzione della libido aumentava in modo significativo qualora gli utilizzatori avessero seguito protocolli di AAS per molte settimane all’anno e avessero una lunga storia di utilizzo di questa classe di farmaci.
Lo svolgimento di una PCT era pratica diffusa nel gruppo in cui la libido tra i cicli non diminuiva rispetto al gruppo in cui la libido diminuiva. Tuttavia, questa differenza non era statisticamente significativa.
L’attuale studio rappresenta ad oggi la più grande valutazione sull’impatto dell’utilizzo di AAS ad alto dosaggio e nel lungo periodo sulla funzione sessuale. I risultati dimostrano che l’aumento della durata e della frequenza di utilizzo degli AAS sono associate a più alti tassi di disfunzione erettile de novo e diminuzione della libido dopo l’interruzione d’uso del/i composto/i.
Gli uomini con disfunzione erettile de novo avevano anche maggiori probabilità di riportare altri sintomi legati a bassi livelli di Testosterone, come riduzione della libido, diminuzione dell’energia, depressione, riduzione soggettiva della massa muscolare e aumento soggettivo della massa grassa. Diversamente a ciò, durante l’uso di un dosaggio più alto di Testosterone e l’uso (con tutta probabilità ponderato) di anti-estrogeni si sono osservati punteggi più alti sul questionario IEEF-5.