Nonostante decenni di “lotta al doping” esso rimane assai diffuso, e non solo nelle competizioni di alto livello. L’errore alla base di questa campagna mediatico-salutistica è stata la generalizzazione; ossia fornire informazioni imprecise, accentuando i possibili sides senza però premurarsi di una vera e propria informativa preventiva chiara, veritiera ed efficace. In poche parole, quello che non si è fatto è dire: “l’uso di PEDs ha una serie di possibili effetti collaterali di gravità dipendente dal tipo di molecola, dal tempo e dalle modalità di assunzione”. Tutto ciò accompagnato da un manuale scientificamente corretto e di facile comprensione, contenente informazioni utili riguardanti la materia PEDs tale da permettere una migliore comprensione della questione che, a sua volta, renda possibile una più consapevole scelta individuale. Ma ciò non è stato fatto. Con l’unica eccezione di alcuni esperti indipendenti che nel corso degli anni hanno pubblicato libri e scritto articoli di una certa utilità.
Lo scopo di questa serie di articoli sarà quello di arginare il fenomeno dell’abuso dei PEDs, cosa che sta degenerando e che sta mostrando i suoi peggiori effetti su atleti di ambo i sessi.
Per la prima pubblicazione di questa nuova serie iniziamo con l’Oxymetholone…
Una (sempre utile) introduzione alla molecola di Oxymetholone:
L’Oxymetholone, noto anche come 2-idrossimetilene-17α-metil-4,5α-diidrotestosterone (2-idrossimetilene-17α-metil-DHT) o come 2-idrossimetilene-17α-metil-5α-androstan-17β-ol-3-one, è uno steroide androstano sintetico e un derivato 17α-alchilato del DHT.[1][2][3]
Le informazioni disponibili sulla farmacocinetica di questo AAS sono limitate.[4] Sembra essere ben assorbito con la somministrazione orale.[4] L’Oxymetholone ha affinità molto bassa per le globuline leganti gli ormoni sessuali nel siero umano (SHBG), meno del 5% di quella del Testosterone e meno dell’1% di quella del DHT. [5] Il farmaco viene metabolizzato nel fegato tramite ossidazione in posizione C2, riduzione in posizione C3, idrossilazione in posizione C17 e coniugazione. [4][6] Il gruppo C2 idrossimetilene del Oxymetholone può essere scisso per formare il Mestanolone (17α-metil-DHT), che può contribuire agli effetti della molecola precursore.[3] L’emivita del Oxymetholone è sconosciuta sebbene vi siano alcune ipotesi a riguardo.[6] L’Oxymetholone e suoi metaboliti vengono eliminati attraverso le urine.[5][6]
Come altri AAS, l’Oxymetholone è un agonista del recettore degli androgeni (AR).[3] Non è un substrato per la 5α-reduttasi (dal momento che è già 5α-ridotto) ed è uno substrato scarso per il 3α-idrossisteroide deidrogenasi (3α-HSD), e quindi mostra un alto rapporto di attività anabolizzante rispetto all’effetto androgenico.[3]
Data la sua derivanza dal DHT, l’Oxymetholone non è un substrato per l’enzima Aromatasi e quindi non può essere aromatizzato in metaboliti estrogenici.[3] Tuttavia, caratteristica unica tra i derivati del DHT, l’Oxymetholone è comunque associato a un’estrogenicità relativamente elevata ed è noto per avere il potenziale di produrre effetti collaterali estrogenici come ginecomastia (raramente) e ritenzione idrica. [3][7][8][9] È stato suggerito che questo può essere una conseguenza del legame diretto a l’attivazione del recettore degli estrogeni da parte dell’Oxymetholone (estrogenicità intrinseca).[3] L’Oxymetholone non possiede alcuna attività progestinica significativa.[3]
A causa della sua struttura 17α-alchilata, l’Oxymetholone è epatotossico.[3] L’uso a lungo termine del farmaco può causare una varietà di disturbi gravi, tra cui l’epatite, il cancro al fegato e la cirrosi; pertanto si raccomandano test periodici di funzionalità epatica per coloro che assumono l’Oxymetholone a fini terapeutici.[10] Questa molecola ha ottenuto, infatti, la nomea di essere uno tra gli AAS più epatotossici. Ciò deriva da i dosaggi comunemente, ed erroneamente, utilizzati in contesto culturistico. Si parla di dosaggi che facilmente sforano i 100-150mg/die. Ma tali dosaggi sono realmente vantaggiosi in termini di guadagni ipertrofici specie se messi in rapporto con gli effetti collaterali possibilmente verificabili? Questa domanda può ottenere una risposta sufficientemente esaustiva attraverso i risultati di uno studio che ha messo a confronto gli effetti di una dose di Oxymetholone da 50mg/die e una da 100mg/die.[11]
Oxymetholone – 50mg Vs. 100mg:
In questo studio, possiamo vedere i cambiamenti nel peso corporeo, nella massa magra, e la perdita di grasso in risposta a un dosaggio moderato e alto di Oxymetholone (50 mg vs 100 mg).
I cambiamenti nella composizione corporea sono mostrati per i gruppi placebo (barre nere), 50mg di Oxymetholone al giorno (barre bianche) e 100mg al giorno (barre grigie). I numeri sopra le barre rappresentano i cambiamenti assoluti medi e le barre di errore sono ± 1 SE. Per la massa corporea magra totale (LBM) e il grasso totale, le differenze tra i 3 gruppi erano significative (P <0,0001, ANOVA a una via). * Differenze significative rispetto al placebo, P ≤ 0,001.
Come ci si aspetterebbe, il gruppo placebo non ha guadagnato massa magra, né ha perso grasso corporeo.
Il gruppo trattato con 50mg di Oxymetholone ha guadagnato 3,3Kg di massa magra e ha perso 2,6kg di grasso.
Il gruppo trattato con 100mg di Oxymetholone ha guadagnato 4,2Kg di massa magra e ha perso 2,5kg di grasso.
I cambiamenti nella composizione regionale (n = 16) sono mostrati per i gruppi placebo, 50mg/die e 100mg/die. A: i numeri sopra le barre rappresentano i cambiamenti assoluti medi per il grasso del tronco mediante assorbimetria a raggi X a doppia energia (DEXA). B: le barre rappresentano i cambiamenti assoluti medi (kg) per la LBM dell’arto superiore (braccio destro più braccio sinistro) mediante DEXA. C: area della sezione trasversale del muscolo totale prossimale (barre grigie) e posteriore (barre nere) dei muscoli della coscia tramite risonanza magnetica. Le barre di errore sono ± 1 SE. * Differenza significativa rispetto al placebo, P ≤ 0,005. .
Guardando la massa corporea magra, è possibile vedere che quando si confrontano i due gruppi di dosaggio, il gruppo da 100mg ha guadagnato solo 0,9kg di massa corporea magra in più rispetto al gruppo da 50mg.
Questo dopo tre mesi di esposizione al doppio della quantità di farmaco.
Se si confrontano i biomarcatori tra i due gruppi, è possibile vedere che l’effetto di 100mg di Oxymetholone ha avuto sui livelli di ALT e AST era molto più deleterio rispetto al gruppo di 50 mg.
Caratteristiche di base della popolazione dello studio
Come molti di voi già sapranno, l’alanina aminotransferasi (ALT) e l’aspartato aminotransferasi (AST) sono biomarcatori comunemente usati per valutare i danni al fegato.
La somministrazione di un dosaggio di Oxymetholone doppio rispetto al basale di 50mg ha prodotto un ulteriore 27% di crescita muscolare relativa (la massa magra non è composta solo dal muscolo scheletrico!), ma ha provocato un picco 3.4x più alto di ALT e un picco 2.7x più alto nei livelli di AST.
Il calo del HDL è stato simile in entrambi i gruppi 50mg/die e 100mg/die.
Quelli sono solo biomarcatori con valore diagnostico per un eventuale danno epatico ma non sono indicativi di ciò che comporta la variabile del dosaggio sull’ipertrofia ventricolare, o altri fattori comunemente trascurati che dovrebbero essere utilizzati per valutare la salute cardiovascolare.
Anche se è possibile che gli aumenti di massa magra misurati dalla DEXA fossero legati in buona parte alla ritenzione idrica causata dalla terapia con Oxymetholone, i notevoli aumenti di forza muscolare misurati con il metodo 1-RM nei gruppi da 50 e 100mg/die (8,2-18,4%) suggeriscono che gli aumenti di massa magra erano probabilmente dovuti all’accrescimento di proteine miofibrillari oltre che alla semplice massa magra totale, poiché la forza è in una certa misura legata alle dimensioni dei muscoli. Inoltre, i membri del gruppo di ricerca hanno riferito che i cambiamenti nella massa magra appendicolare tramite DEXA sono quantitativamente correlati ai cambiamenti nella forza muscolare scheletrica in risposta a stimoli anabolici. In effetti, nel presente studio, sono stati in grado di corroborare questa relazione dimostrando che gli aumenti significativi del tessuto magro della parte superiore del corpo mediante scansione DEXA appendicolare erano altamente correlati con i cambiamenti nella forza della parte superiore del corpo come valutato da esercizi di Chest Press e Lat Pull-Down. Inoltre, i cambiamenti nella forza muscolare massima volontaria per gli esercizi della parte superiore del corpo hanno mostrato una risposta legata alla dose.
I cambiamenti relativi (%) nella forza sono mostrati per i gruppi placebo (barre nere), 50mg/giorno Oxymetholone (barre bianche) e 100mg/giorno Oxymetholone (barre grigie). I numeri sopra le barre rappresentano il cambiamento relativo (%) dal basale alla settimana 12 per le prove di forza massima a 1 ripetizione. Le barre di errore rappresentano ± 1 SE dalla media. * Differenza significativa rispetto al placebo, P < 0,05; † differenza significativa rispetto al placebo con il test di Wilcoxon, P < 0,02.
Al contrario, c’erano guadagni non significativi tra i tre gruppi di trattamento per la forza degli arti inferiori (3,9-12,0%), coerentemente con la mancanza di un aumento significativo della massa magra degli arti inferiori mediante scansione DEXA. Tuttavia, c’era una differenza quasi significativa (P = 0,052) tra i gruppi per il cambiamento del area della sezione trasversale del muscolo (CSA) dei muscoli della coscia tramite la risonanza magnetica, suggerendo che la terapia dello studio può aver influenzato positivamente i muscoli degli arti inferiori. È possibile che i test di forza di gruppi muscolari multipli e di grandi dimensioni, come quelli utilizzati con l’esercizio Leg Press, siano meno sensibili ai modesti cambiamenti nella massa muscolare, e lo studio potrebbe non aver avuto sufficiente potenza per rilevare piccoli ma significativi guadagni nelle estremità inferiori. Si ipotizza che ciò sia dovuto al fatto che i grandi muscoli delle gambe sono abitualmente utilizzati più frequentemente per sostenere il carico (ad esempio, camminare, alzarsi da una sedia) rispetto ai muscoli dell’estremità superiore negli adulti più anziani. Piccoli ma significativi guadagni nella forza e nella massa muscolare della parte inferiore del corpo possono essere meno dimostrabili che per i muscoli della parte superiore del corpo, che possono essere utilizzati meno per il lavoro ad alto volume e più inclini alla sarcopenia nelle persone anziane. Inoltre, i muscoli degli arti superiori, rispetto ai muscoli degli arti inferiori, hanno proporzioni maggiori di fibre a contrazione rapida di tipo II, che possono essere perse preferibilmente con l’invecchiamento. Inoltre, uno studio longitudinale in uomini anziani ha mostrato che le fibre di tipo I sono state perse principalmente nel vasto laterale della gamba, portando all’ipotesi che ci potrebbe essere una maggiore perdita di fibre di tipo II nelle braccia con l’invecchiamento. Così la risposta agli stimoli anabolici può essere più facilmente dimostrabile nelle estremità superiori di questa popolazione.
C’erano anche significative ma simili diminuzioni del grasso corporeo totale di 2,6 ± 1,2 e 2,5 ± 1,6 kg nei gruppi di 50 e 100mg al giorno, rispettivamente. Una parte importante del miglioramento dell’adiposità riguardava la diminuzione del grasso del tronco (1,7 ± 1,0 e 2,2 ± 0,9 kg nei due rispettivi gruppi di trattamento attivo). Una riduzione significativa del grasso del tronco potrebbe influenzare favorevolmente i fattori di rischio per le malattie cardiovascolari. Anche se ci aspetteremmo che la riduzione del grasso addominale si rifletta in una migliore sensibilità all’insulina, le misure indirette (HOMA-IR e QUICKI) potrebbero non essere state abbastanza sensibili. È anche possibile che ci fossero troppo pochi soggetti in ogni gruppo per rilevare cambiamenti piccoli ma significativi.
Ci sono ragioni teoriche per temere che l’eccesso di androgeni possa provocare o essere associato all’insulino-resistenza, anche se questa relazione è stata dimostrata solo in donne con sindrome dell’ovaio policistico. Non è stata misurata direttamente la sensibilità all’insulina né con il clamp euglicemico iperinsulinemico né con test di tolleranza al glucosio endovena a campionamento frequente. Tuttavia, le misure indirette della sensibilità insulinica (insulina a digiuno, HOMA-IR, QUICKI) non hanno mostrato prove di resistenza insulinica.
Cosa estrapolare?
Questo studio però presenta alcune limitazioni che possono averne influenzato i risultati. In primo luogo, la piccola dimensione del campione di meno di una dozzina di soggetti per gruppo può aver limitato la capacità di rilevare piccoli ma importanti cambiamenti in variabili come la massa magra (LBM) delle estremità inferiori e il CSA della muscolatura della coscia. Allo stesso modo, è possibile che le differenze osservate per i cambiamenti nella LBM totale e nella forza avrebbero potuto essere significative tra i gruppi di trattamento con dimensioni del campione maggiori. Quest’ultimo avrebbe fornito ulteriore supporto alla nostra supposizione di una risposta dose-dipendente con l’Oxymetholone. In secondo luogo, la popolazione rappresentava uomini adulti più anziani, che sono stati caratterizzati come a rischio di sarcopenia legata all’età sulla base dei rapporti che mostrano la perdita di massa e forza muscolare con l’invecchiamento. Tuttavia, i soggetti non sono stati reclutati per la perdita di peso, la fragilità o l’ipogonadismo palese di per sé, dal momento che è stato dimostrato che gli uomini più giovani con concentrazioni di Testosterone normali possono ottenere aumenti apprezzabili della massa muscolare e della forza dopo l’integrazione di androgeni. Inoltre, ci sono prove che la sintesi proteica miofibrillare nelle persone anziane può essere significativamente aumentata a livelli paragonabili a quelli raggiunti nelle persone più giovani in risposta a un potente stimolo anabolico. Infine, poiché l’Oxymetholone è un AAS 17-metilato che provoca un elevato effetto di primo passaggio nel fegato, e che nel presente studio non sono state prese misure di contenimento per l’epatotossicità potenziale, i risultati di AST e ALT ottenuti rappresentano solamente modelli privi di ancillari volti ad una epatoprotezione.
Conclusioni sul dosaggio “ottimale” di Oxymetholone:
Evidenziati i limiti dello studio, pur prendendo i dati ivi riportati universalmente rapportabili al basale d’uso della molecola (es. vedi epatotossicità), possiamo giungere, grazie all’ausilio di dati empirici raccolti negli anni attraverso indagini svolte sulle preparazioni di svariati atleti di medio e alto livello, ad identificare un dosaggio con una ratio “efficacia:rischio (E:R)” favorevole per l’atleta.
Un dato è emerso preponderante nel corso delle indagini svolte: quale fosse il peso dell’atleta e il suo condizionamento atletico, nonché l’utilizzo di una adeguata epatoprotezione e controllo della dislipidemia, il margine della ratio E:R diveniva evidentemente sfavorevole oltre i 150mg/die. Indi per cui, i dosaggi elevati raggiunti da certi atleti, arrivando a picchi di 200-300mg/die, sono risultati inutili al miglioramento delle risposte anabolizzanti complessive e inficianti per il corretto svolgimento della stessa preparazione (vedi, ad esempio, marcata inappetenza e nausea).
Dosaggi standard per un atleta di sesso maschile non dovrebbero discostarsi dal range 50-100mg/die, considerando che la taratura del “dosaggio ideale” si è ottenuta calcolando la dose individuale con la formula 1mg/Kg di peso corporeo. Ovviamente, l’assicurarsi una adeguata protezione epatica e lipidica è il punto parallelo da raggiungere.
Nelle atlete, invece, vista la loro maggiore sensibilità agli aumenti degli androgeni circolanti, la “dose ideale” si è attestata a 25mg/die con punte massime (anche se non necessarie) di 50mg/die. A tal proposito, vorrei ricordare che l’Oxymetholone è risultato essere una molecola più vantaggiosa nel controllo degli effetti collaterali androgenizzanti rispetto a composti quali Methenolone e Boldenone.
La linea tra abuso e uso è spesso molto sottile, ma nel caso del Oxymetholone essa si mostra sufficientemente marcata…
Pavlatos AM, Fultz O, Monberg MJ, Vootkur A (June 2001). “Review of oxymetholone: a 17alpha-alkylated anabolic-androgenic steroid”. Clinical Therapeutics. 23 (6): 789–801, discussion 771.
Saartok T, Dahlberg E, Gustafsson JA (June 1984). “Relative binding affinity of anabolic-androgenic steroids: comparison of the binding to the androgen receptors in skeletal muscle and in prostate, as well as to sex hormone-binding globulin”. Endocrinology. 114 (6): 2100–6.
Hengge UR, Stocks K, Wiehler H, Faulkner S, Esser S, Lorenz C, et al. (March 2003). “Double-blind, randomized, placebo-controlled phase III trial of oxymetholone for the treatment of HIV wasting”. AIDS. 17 (5): 699–710.
Cortesgallegos V, Castaneda G, Alonso R, Perezpasten E, Reyeslugo V, Barron C, Mondragon L, Villalpando S (January 1982). “Spontaneous and Oxymetholone-Induced Gynecomastia”. Journal of Andrology. C/O Allen Press, Inc Po Box 368, Lawrence, Ks 66044: Amer Soc Andrology, Inc. 3 (1): 33.
Villalpando S, Mondragon L, Barron C, Reyeslugo U, Perezpasten E, Alonso R, Castaneda G, Gallegos V (January 1982). “5-Alpha Reductase Blockade May Be Responsible for Spontaneous and Oxymetholone-Induced Gynecomastia”. Archivos de Investigacion Medica. Social Apdo Postal 73-032, Mexico Df 03020, Mexico: Inst Mexicano Seguro. 13 (2): s13.
Il fegato è un organo importante ed è vitale per la sopravvivenza del soggetto. È responsabile di diverse e importanti funzioni nel corpo umano. Produce acidi biliari e proteine plasmatiche, immagazzina glicogeno e produce glucosio attraverso la gluconeogenesi, gioca un ruolo nel sistema immunitario, metabolizza un numero elevato di molecole, ecc. Quindi, si, avete capito bene: è importante. Quando qualcosa risulta dannosa per il fegato, essa si indica come epatotossico (dal greco hêpar-atos, fegato). Un chiaro esempio è l’alcol. Gli alcolisti tendono a sviluppare una malattia del fegato a un certo punto della loro vita. Tuttavia, molti farmaci da prescrizione, o anche over-the-counter, possono essere epatotossici, come l’Acetaminofene. E, come è ben dimostrato, anche gli AAS possono essere epatotossici, anche se specifici. Come sembra, solo quelli con una specifica alterazione chimica sembrano essere maggiormente epatotossici – in particolare, quelli che presentano una metilazione in pozione C-17α.
Modifica della struttura carbossilica del Testosterone (sinistra) in posizione C-17α (destra).
In questo articolo tratterò principalmente ciò che sembra causare questa epatotossicità indotta da AAS. L’effetto epatotossico può essere riscontrato attraverso l’osservazione dei cambiamenti nei marcatori ematici del danno epatico, come Alanina Transaminasi (ALAT), Aspartato Transaminasi (ASAT), γ-glutamiltransferasi (GGT) e la Fosfatasi Alcalina (ALP). Una nota di cautela deve essere presa in considerazione quando si interpretano gli aumenti di ALAT e ASAT, poiché entrambi aumenteranno anche a causa del intyenso lavoro muscolare [1]. È bene sapere che in questi casi, ASAT sarà di solito più alto del ALAT, mantenendo un rapporto ASAT/ALAT superiore a 1. Quindi, quando questi aumentano con un rapporto inferiore a 1, si può essere più sicuri che il danno muscolare non è il colpevole dell’alterazione. Idealmente, nessun esercizio (contro-resistenza) viene svolto 1-2 settimane prima dell’esame del sangue per escludere il danno muscolare muscolare come causa dell’innalzamento, sebbene ciò dipenda anche dall’intensità del allenamento. In rari casi, il danno al fegato potrebbe avanzare clinicamente fino allo sviluppo di ittero colestatico [2]. In questo caso, un prodotto della degradazione dei globuli rossi (bilirubina) si accumula nel corpo. L’ittero può essere osservato visivamente (tono giallo della pelle e della sclera degli occhi), e si possono sviluppare sintomi come nausea, vomito, dolore allo stomaco e prurito. Inoltre, alcuni rari casi di peliosis hepatis (Peliosi Epatica) sono stati segnalati verificarsi come risultato dell’uso di AAS orali ad alte dosi [3]. Questa è una condizione nella quale si vengono a formare cisti piene di sangue nel fegato. La sospensione dell’AAS in questione è solitamente sufficiente e porterà alla scomparsa di queste caratteristiche cliniche entro pochi mesi. In casi più gravi, tuttavia, potrebbero richiedere un intervento chirurgico. Infine, alcuni casi in letteratura hanno riportato un’associazione tra uso di AAS e carcinoma epatico [4] e adenoma [5].
Ho già trattato in passato tale problematica legata all’uso di AAS, ma questa volta voglio trattare la questione più nello specifico, analizzando le due ipotesi che ruotano intorno all’epatotossicità AAS-dipendente: “ipotesi dello stress ossidativo” e “ipotesi di coniugazione dell’anello D”.
L’ipotesi dello stress ossidativo:
L’ipotesi dello stress ossidativo che tratterò qui si basa su un documento che William Llewellyn, Peter Van Mol e Peter Bond hanno pubblicato [6]. Lo stress ossidativo è qualcosa che si pensa possa risultare nell’epatotossicità osservata con l’uso di AAS, e se l’ipotesi è vera, dà qualche opportunità per contrastarla in modo migliore. Quindi, cominciamo con spiegare quello che è lo stress ossidativo. Lo stress ossidativo è descritto da Helmut Sies come un disturbo nell’equilibrio pro-ossidante-antiossidante a favore del primo [7], che si riduce a molecole contenenti ossigeno, che sono altamente reattive (specie reattive dell’ossigeno [ROS]), sopraffacendo il sistema antiossidante. Poiché le ROS sono così altamente reattive, possono reagire con molecole come lipidi, proteine, carboidrati e acidi nucleici (elementi costitutivi del DNA). Quando si dice “reagire con queste molecole”, si intende che danneggia queste molecole (estremamente semplificato, ma è sufficiente per far comprendere il processo). Questi ROS provengono da varie reazioni catalizzate da enzimi come la respirazione cellulare (l’ossidazione dei macronutrienti per fornire energia), altri processi metabolici e radiazioni. La fonte primaria di ROS all’interno di una cellula sono i mitocondri, il che non è sorprendente dato che i mitocondri sono le “centrali energetiche” della cellula. È il posto nella cellula dove i carboidrati alimentari, gli acidi grassi e le proteine (o, meglio, gli amminoacidi che le compongono) finiscono per essere ossidate per produrre energia in un processo chiamato fosforilazione ossidativa. Come suggerisce il nome, la fosforilazione ossidativa ossida e richiede ossigeno per farlo. Questo processo, tuttavia, non è perfetto. Per non complicare troppo le cose al lettore, non mi addentrerò nelle complessità delle reazioni chimiche, ma fondamentalmente, questo processo può produrre ROS come sottoprodotto (superossido in particolare). Le cellule del corpo sono dotate di meccanismi per tenere a bada questi ROS generati (la parte antiossidante dell’equazione). In circostanze normali questo porta ad un sottile equilibrio tra i due. Avere qualche ROS qua e là nelle cellule è normale. Essi giocano un ruolo essenziale nel normale funzionamento di vari processi vitali [8]. Tuttavia, il problema nasce quando questo equilibrio si altera a favore della parte proossidante dell’equazione: lo stress ossidativo. Questo è il momento in cui i ROS prendono il sopravvento, per così dire, e possono iniziare a creare il caos nella cellula. Quanto sopra è un quadro un po’ troppo semplificato. Ci sono diversi tipi di ROS (radicali liberi e non radicali). Ciò che conta è dove si trovano questi ROS nella cellula e come evolvono nel tempo. Inoltre, questo interagisce con il sistema antiossidativo delle cellule, il che complica ulteriormente il quadro. Ma credo che quanto sopra sia sufficiente per dare una buona comprensione di tutto questo. Ciò che conta è che l’epatotossicità indotta da AAS è stata ripetutamente dimostrata essere associata allo stress ossidativo nelle cellule epatiche (fegato) di modelli animali [9]. Questo fa sorgere la domanda: è solo un’associazione, o c’è una relazione causale con l’epatotossicità indotta da AAS? Dopo aver scavato nella letteratura, sono emersi alcuni studi che sembrano sostenere una relazione causale. Uno studio svolto su un carcinoma prostatico umano epiteliale (22Rv1) ha collegato l’attivazione del recettore degli androgeni (AR) a un aumento dei ROS basali [10]. Più tardi, lo stesso gruppo ha pubblicato una ricerca applicando un disegno di studio simile. Questo studio ha confermato i precedenti risultati e ha anche dimostrato che l’aumento dei ROS è dovuto a un aumento indotto dall’AAS nella β-ossidazione mitocondriale degli acidi grassi [11]. Quindi, l’attivazione di l’AR porta a una maggiore ossidazione degli acidi grassi nei mitocondri, con conseguente maggiore produzione di ROS come sottoprodotto. Da notare che questo studio ha anche trovato un aumento dell’mRNA della carnitina palmitoiltransferasi (CPT1). Tutto quello che dovete sapere è che la CPT1 è considerata essere l’enzima che regola la velocità nel processo di ossidazione mitocondriale degli acidi grassi. Quindi, se si aumenta la CPT1, si aumenta l’ossidazione mitocondriale degli acidi grassi. Ora, le cellule del cancro alla prostata non sono cellule del fegato, ovviamente. Ma ciò che è interessante è che l’AAS 17α-alchilato Fluoxymesterone e Metilandrostanolone hanno dimostrato di aumentare l’attività del CPT1 nel fegato di ratto [12]. Inoltre, se si guardano agli epatociti di ratto (cellule epatiche) trattati con AAS 17α-alchilati, si vedrà il gonfiore dei mitocondri e solo cristae leggermente definite [13]. (Le criste sono quelle pieghe caratteristiche della membrana interna dei mitocondri). Infatti, la produzione di ROS è una causa nota di gonfiore mitocondriale, e il gonfiore è un fattore importante che porta alla successiva morte cellulare [14]. Quindi, apparentemente, suggerisce un potenziale ruolo dello stress ossidativo. Questo non vuol dire che qualsiasi aumento nella produzione di energia di una cellula sia negativo. Usando i muscoli aumenta anche la produzione di energia nelle cellule muscolari. Di conseguenza, più ROS vengono prodotti anche in queste cellule. In contrasto con l’aumento di ROS indotto dall’AAS nelle cellule del fegato, questi aumenti sono transitori invece che continui. Inoltre, le cellule muscolari differiscono nei loro meccanismi antiossidanti per gestire questa condizione. Quindi, normalmente, questo non è assolutamente un problema. Tuttavia, l’esercizio intenso e prolungato può anche provocare danni ossidativi alle molecole delle cellule muscolari [15].
L’ipotesi dello stress ossidativo nella epatotossicità indotta da AAS come descritto da Bond et al. [49]. 1 Un androgeno si lega a, e attiva, il recettore degli androgeni (AR) nelle cellule epatiche. Questo porta a 2 la sovra-regolazione della Carnitina Palmitoiltransferasi 1 (CPT1), l’enzima che regola il tasso di β-ossidazione degli acidi grassi (FA). Si pensa che questo porti a 3 un aumento della β-ossidazione degli acidi grassi nei mitocondri. Di conseguenza, 4 la produzione di specie reattive dell’ossigeno (ROS) è aumentata. L’aumento dei ROS poi danneggia i mitocondri, il che sembra essere alla base dell’epatotossicità indotta dall’AAS.
Ora, se si integrassero gli antiossidanti (mitocondriali), si allevierebbe questo danno? Può darsi. Mentre non c’è un trial di buona qualità che valuti questo, uno studio osservazionale su 320 atleti dimostra qualcosa del genere [16]. In breve, gli utilizzatori di AAS che hanno preso un supplemento contenente alcuni composti antiossidanti non ha mostrato alcun aumento dei marcatori di danno epatico dopo il ciclo rispetto a quelli che non hanno assunto quel supplemento. Ancora una volta, questo sarebbe in linea con lo stress ossidativo che gioca un ruolo causale nell’epatotossicità indotta da AAS. Infine, sembra che l’epatotossicità indotta da AAS potrebbe essere legata all’attivazione del AR nelle cellule epatiche. In un vecchio studio del 1964, Marquardt et al. non sono riusciti a dimostrare che l’AAS non 17α-alchilato produce test di funzionalità epatica anormali [17]. Infatti, gli AAS 17α-alchilati mostrano segni di epatotossicità in diversi studi, mentre non si vede questo con AAS non-17αalchilati, nemmeno con un alto dosaggio di 600 mg di Testosterone Enantato settimanale [18]. La 17α-alchilazione sembra quasi necessaria per rendere epatotossico un AAS, probabilmente perché è l’unica alterazione che lo rende sufficientemente biodisponibile per via orale. E, di conseguenza, porta ad alte concentrazioni del composto nel fegato. Ma possiamo individuare le differenze tra i vari AAS 17α-alchilati che riguardano la loro capacità di attivare l’AR? Certamente sembra così. In generale, sembra che sia vero quanto segue:
Epatotossicità = resistenza alla decomposizione epatica×potenza di attivazione del AR
Quindi, facciamo un esempio. Il Methyltrienolone (R1881) ha un’affinità molto alta per l’AR, ha un’alta potenza per la transattivazione dell’AR [19], ed è fortemente resistente al metabolismo epatico. Come tale, è un composto ideale per un saggio dei siti di legame agli androgeni [20]. Infatti, un studio clinico che impiega un basso dosaggio dello steroide (≤1 mg al giorno) ha dimostrato un significativo aumento dei marcatori di danno epatico entro due settimane [21]. Gli autori lo hanno definito “(…) attualmente lo steroide più epatotossico”. Lo steroide 17α-alchilato meno epatotossico è solitamente considerato l’Oxandrolone. Anche con alti dosaggi fino a 80mg al giorno, mostra solo deboli segni di epatotossicità [22]. Mentre lo steroide è abbastanza resistente al metabolismo epatico [23], ha una bassa affinità per il AR [23]. La sua potenza relativa in termini di transattivazione AR è anche quasi 100 volte inferiore a quella del Methyltrienolone [19]. Allo stesso modo, anche l’Oxymetholone ha una bassa affinità per l’AR [23] e la sua potenza in termini di transattivazione AR è molto simile a quella dell’Oxandrolone [19]. Non sorprende che mostri segni di epatotossicità solo in una minoranza di pazienti, nonostante gli alti dosaggi (100-150 mg al giorno) [24].
L’ipotesi di coniugazione dell’anello D:
Avete mai sfogliato il libro Doping in Sports di Thieme e Hemmersbach? [25] In questo libro gli autori notano che non c’è correlazione tra la tossicità epatica e gli effetti farmacologici primari (cioè gli effetti anabolizzanti) – il che è sufficientemente ovvio perché gli AAS non 17α-alchilati sono rapidamente metabolizzati nel fegato, quindi la loro concentrazione in loco non sarebbe come quella dei 17α-alchilati. Naturalmente, non si troverà una correlazione se si guarda solo a questo fattore. Bisogna anche prendere in considerazione la sua resistenza al metabolismo epatico come è stato fatto con l’ipotesi dello stress ossidativo descritta sopra.
In ogni caso, questo ha portato gli autori a formulare un’alternativa ipotesi di ciò che causa l’epatotossicità indotta da AAS. E sembrava essere l’unica. Essi suggeriscono che l’epatotossicità è probabilmente dovuta alla coniugazione dell’anello D con l’acido glucuronico. Questo processo è chiamato glucuronidazione ed è una cosiddetta comune reazione di fase 2 nel metabolismo del farmaco. Rende la molecola madre più solubile in acqua, facilitando così la sua escrezione nelle urine.
Il gruppo 17β-glucuronide (in blu) attaccato al anello D di uno steroide 17α-metilato (gruppo 17α-metilico in rosso).
È semplicemente l’attaccamento (coniugazione) dell’acido glucuronico alla molecola madre (vedi figura sopra). Quando il Testosterone con un gruppo 17β-glucuronide (così come diversi estrogeni con questa modifica) viene iniettato nel ratto, il flusso biliare è inibito [521]. Presumibilmente, perché questi composti condividono somiglianze strutturali con gli acidi biliari, questi composti competono con gli acidi biliari per legarsi a certi recettori. Tuttavia, a parte questo, non c’è molta sostanza per sostenere questa ipotesi come la ragione per l’epatotossicità indotta da AAS, soprattutto perché molti degli AAS non 17α-alchilati, compreso il Testosterone, subiscono la glucuronizzazione del loro gruppo 17β-idrossi. Eppure questi non sono sensibilmente epatotossici. Infatti, la 17βglucuronidazione è stata identificata solo per alcuni AAS 17α-alchilati, e sembra che essi subiscono questo processo solo in piccola misura [26]. Così, ironicamente, se questa ipotesi fosse vera, o significativa, ci si aspetterebbe l’epatotossicità con il Testosterone ma non con gli AAS 17α-alchilati.
Conclusioni sulle ipotesi esposte:
Non è sicuramente una novità per l’utilizzatore medio, ma anche per il semplice soggetto interessato all’argomento PEDs, che gli AAS metilati in C-17 (17α-alchilati) abbiano un effetto epatotossico con lievi variabili tra molecole aventi la stessa modifica strutturale. E non è nemmeno una rivelazione che la supplementazione con antiossidanti (vedi NAC e Silimarina) possa ridurre tale effetto. Di conseguenza, l’ipotesi dello stress ossidativo sembra essere la principale causa del epatotossicità AAS-indotta. Ma non l’unico fattore.
Nell’ultimo decennio si è aggiunto ai classici composti antiossidanti l’uso di acidi biliari come l’Acido Ursodesossicolico e l’Acido Tauroursodesossicolico assunti oralmente.
L’Acido Ursodesossicolico è un acido biliare secondario che deriva dal metabolismo dell’acido colico da parte del microbiota umano intestinale. Il suo nome deriva dal fatto che è il principale acido biliare negli orsi (dal latino ursus). In biologia e biochimica lo si etichetta con l’acronimo UDCA. Il nome completo del UDCA è Acido 3α,7β-diidrossi-5β-colanoico.[27]
Acido Ursodesossicolico (UDCA)
L’Acido Tauroursodesossicolico (TUDCA) è un acido biliare ambifilico. È la forma coniugata di Taurina ed il precedentemente citato Acido Ursodeossicolico (UDCA). Il nome completo del TUDCA è 2-{(4R)-4-[(1R,3aS,3bR,4S,5aS,7R,9aS,9bS,11aR)-4,7-Dihydroxy-9a,11a-dimethylhexadecahydro-1H-cyclopenta[a]phenanthren-1-yl]pentanamido} acido etan-1-sulfonico.[28]
Acido Tauroursodesossicolico (TUDCA)
l’UDCA è approvato per il trattamento della cirrosi biliare primaria.[1][2] Di conseguenza, l’Acido Ursodesossicolico (UDCA) ha mostrato effetti epatoprotettivi. Tuttavia, i suoi meccanismi molecolari sottostanti rimangono poco chiari. Per tale motivazione, sono stati condotti alcuni studi come quello di Da Jung Kim et al. nel quale è stato osservato l’effetto epatoprotettivo dell’UDCA e della vitamina E utilizzando la metabolomica e l’analisi metagenomica. In questo studio, sono stati analizzati campioni di sangue e urine di pazienti con obesità e disfunzione epatica. Nove pazienti sono stati assegnati in modo casuale a ricevere UDCA (300 mg due volte al giorno), e 10 soggetti hanno ricevuto la vitamina E (400 UI due volte al giorno) per 8 settimane. L’UDCA ha migliorato significativamente i punteggi della funzionalità epatica dopo 4 settimane di trattamento e ha ridotto efficacemente i livelli epatici di acido Desossicolico e di microRNA-122 nel siero. Per comprendere meglio il suo meccanismo protettivo, è stato condotto uno studio di metabolomica globale ed è stato scoperto che l’UDCA ha regolato le tossine uremiche (acido ippurico, solfato di p-cresolo e metaboliti derivati dall’indolo), gli antiossidanti (solfato di ascorbato e N-acetil-L-cisteina) e il percorso fenilalanina/tirosina. Inoltre, il coinvolgimento del microbioma, in particolare di Lactobacillus e Bifidobacterium, è stato dimostrato attraverso l’analisi metagenomica delle vescicole extracellulari derivate dai batteri. Nel frattempo, il trattamento con vitamina E non ha portato a tali alterazioni, tranne che ha ridotto le tossine uremiche e la disfunzione epatica. I nostri risultati hanno suggerito che entrambi i trattamenti erano efficaci nel migliorare la funzione epatica, anche se attraverso meccanismi diversi.
Schema dei potenziali meccanismi terapeutici del trattamento con UDCA. L’analisi metabolomica ha rivelato che l’UDCA riduce i principali composti nei percorsi fenilalanina/tirosina e triptofano, tra cui fenilalanina, fenilacetato, acetilfenilalanina, aldeide 3,4-idrossifenilacetato, dopamina-3-O-solfato, idrossibenzaldeide, p-cresolo solfato, idrossicynurenamina, idrossindolo e acido ippurico, nel plasma e nelle urine. I metaboliti intermedi degli aminoacidi aromatici come l’idrossimelatonina, l’acido benzoico e l’acido salicilico sono stati aumentati. I forti antiossidanti come l’ascorbato, l’acetiltriptofano e la N-acetil-L-cisteina erano elevati. Inoltre, la disintossicazione delle tossine uremiche tramite glucuronidazione (idrossimetossiindolo glucuronide e p-cresolo glucuronide) è stata osservata dopo il trattamento UDCA. Tuttavia, la vitamina E ha ridotto l’acido indolo-propionico, il solfato di indoxile, la 3-ketosphinganina e la sfingosina, che non sono stati regolati dall’UDCA. Il colore blu indica una diminuzione del livello del metabolita, e il colore rosso indica un aumento del livello del metabolita dopo il trattamento UDCA. I metaboliti che sono cambiati dopo il trattamento con vitamina E sono contrassegnati da un asterisco (*). I metaboliti che sono stati possibilmente regolati da modifiche batteriche sono contrassegnati da un colore viola.
Inoltre, si sa che l’UDCA a livello epatico stimola la secrezione di ATP da parte degli epatociti[29]; sebbene il significato di quest’azione non è ancora noto. Si sa però che interagisce col sistema dei citocromi P450 e che riduce la Glicuronazione degli estrogeni sintetici e non solo.[30] Vi ricorda qualcosa? Esatto! L’ipotesi di coniugazione dell’anello D e la sua potenzialità di essere parte dell’effetto epatotossico AAS-indotto! Se a ciò aggiungiamo che l’UDCA possiede la capacità di attivare direttamente il recettore per i glucocorticoidi, che contribuirebbe ad allargare i meccanismi della sua azione anticolestatica ed antinfiammatoria sul parenchima epatico [31], e che stimola la sintesi del glutatione (GSH), potente antiossidante endogeno, attraverso l’intervento delle chinasi dipendenti dai fosfoinositidi (PI-3K e PKB) [32], ciò fa si che l’UDCA risulti la chiave di volta nella protezione epatica durante l’uso di AAS con marcata resistenza al metabolismo epatico in abbinamento ai largamente utilizzati NAC (precursone ad alta biodisponibilità del Glutatione) e Silimarina.
Quanto detto non rappresenta ne un consiglio medico ne una scusa per abusare di AAS di qualsiasi tipo! Si tratta semplicemente della divulgazione di informazioni che la seria ricerca scientifica ha permesso di estrapolare, per il momento…
Gabriel Bellizzi
Riferimenti:
W. J. Meyer, A. Webb, C. A. Stuart, J. W. Finkelstein, B. Lawrence, and P. A. Walker. Physical and hormonal evaluation of transsexual patients: a longitudinal study. Archives of sexual behavior, 15(2):121–138, 1986.
A. M. Elsharkawy, S. McPherson, S. Masson, A. D. Burt, R. T. Dawson, and M. Hudson. Cholestasis secondary to anabolic steroid use in young men. Bmj, 344, 2012.
J. Nadell and J. Kosek. Peliosis hepatis. twelve cases associated with oral androgen therapy. Archives of pathology & laboratory medicine, 101(8):405–410, 1977.
F. L. Johnson, K. Lerner, M. Siegel, J. Feagler, P. Majerus, J. Hartmann, and E. D. Thomas. Association of androgenic-anabolic steroid therapy with development of hepatocellular carcinoma. The Lancet, 300(7790):1273–1276, 1972.
L. Hernandez-Nieto, M. Bruguera, J. A. Bombi, L. Camacho, and C. Rozman. Benign liver-cell adenom associated with long-term administration of an androgenic-anabolic steroid (methandienone). Cancer,40(4):1761–1764, 1977.
P. Bond, W. Llewellyn, and P. Van Mol. Anabolic androgenic steroid-induced hepatotoxicity. Medical Hypotheses, 93:150–153, 2016.
H. Sies et al. Oxidative stress: introductory remarks. Oxidative stress, 501:1–8, 1985.
K. Brieger, S. Schiavone, F. J. Miller Jr, and K.-H. Krause. Reactive oxygen species: from health to disease. Swiss medical weekly, 142:w13659, 2012.
S. P. Frankenfeld, L. P. Oliveira, V. H. Ortenzi, I. C. Rego-Monteiro, E. A. Chaves, A. C. Ferreira, A. C. Leitáo, D. P. Carvalho, and R. S. Fortunato. The anabolic androgenic steroid nandrolone decanoate disrupts redox homeostasis in liver, heart and kidney of male wistar rats. PloS one, 9(9):e102699, 2014.
J. H. Pinthus, I. Bryskin, J. Trachtenberg, J.-P. Luz, G. Singh, E. Fridman, and B. C. Wilson. Androgen induces adaptation to oxidative stress in prostate cancer: implications for treatment with radiation therapy. Neoplasia, 9(1):68–80, 2007.
H. Lin, J.-P. Lu, P. Laflamme, S. Qiao, B. Shayegan, I. Bryskin, L. Monardo, B. C. Wilson, G. Singh, and J. H. Pinthus. Inter-related in vitro effects of androgens, fatty acids and oxidative stress in prostate cancer: a mechanistic model supporting prevention strategies. International journal of oncology, 37(4):761–766, 2010.
M. Guzmán, A. Saborido, J. Castro, F. Molano, and A. Megias. Treatment with anabolic steroids increases the activity of the mitochondrial outer carnitine palmitoyltransferase in rat liver and fast-twitch muscle. Biochemical pharmacology, 41(5):833–835, 1991.
R. Gragera, A. Saborido, F. Molano, L. Jimenez, E. Muñiz, and A. Megias. Ultrastructural changes induced by anabolic steroids in liver of trained rats. Histology and histopathology, 1993.
X. Chapa-Dubocq, V. Makarov, and S. Javadov. Simple kinetic model of mitochondrial swelling in cardiac cells. Journal of cellular physiology, 233(7):5310–5321, 2018.
S. K. Powers, L. L. Ji, A. N. Kavazis, and M. J. Jackson. Reactive oxygen species: impact on skeletal muscle. Comprehensive Physiology, 1(2):941–969, 2011.
T. A. Pagonis, G. N. Koukoulis, C. S. Hadjichristodoulou, P. N. Toli, and N. V. Angelopoulos. Multivitamins and phospholipids complex protects the hepatic cells from androgenic-anabolic-steroids-induced toxicity. Clinical Toxicology, 46(1):57–66, 2008.
G. H. Marquardt, C. E. Logan, W. G. Tomhave, and R. M. Dowben. Failure of non-17-alkylated anabolic steroids to produce abnormal liver function tests. The Journal of Clinical Endocrinology & Metabolism, 24(12):1334–1336, 1964.
S. Bhasin, L. Woodhouse, R. Casaburi, A. B. Singh, D. Bhasin, N. Berman, X. Chen, K. E. Yarasheski, L. Magliano, C. Dzekov, et al. Testosterone dose-response relationships in healthy young men. American Journal of Physiology-Endocrinology And Metabolism, 281(6):E1172–E1181, 2001.
C. J. Houtman, S. S. Sterk, M. P. Van de Heijning, A. Brouwer, R. W. Stephany, B. Van der Burg, and E. Sonneveld. Detection of anabolic androgenic steroid abuse in doping control using mammalian reporter gene bioassays. Analytica chimica acta, 637(1-2):247–258, 2009.
C. Bonne and J.-P. Raynaud. Assay of androgen binding sites by exchange with methyltrienolone (r 1881). Steroids, 27(4):497–507, 1976.
H. L. Krüskemper and G. Noell. Liver toxicity of a new anabolic agent: methyltrienolone (17α-methyl-4, 9, 11-estratriene-17β-ol-3-one). Steroids, 8(1):13–24, 1966.
C. Grunfeld, D. P. Kotler, A. Dobs, M. Glesby, S. Bhasin, O. S. Group, et al. Oxandrolone in the treatment of hiv-associated weight loss in men: a randomized, double-blind, placebo-controlled study. JAIDS Journal of Acquired Immune Deficiency Syndromes, 41(3):304–314, 2006.
J. A. Kemppainen, E. Langley, C.-i. Wong, K. Bobseine, W. R. Kelce, and E. M. Wilson. Distinguishing androgen receptor agonists and antagonists: distinct mechanisms of activation by medroxyprogesterone acetate and dihydrotestosterone. Molecular Endocrinology, 13(3):440–454, 1999.
U. R. Hengge, K. Stocks, S. Faulkner, H. Wiehler, C. Lorenz, W. Jentzen, D. Hengge, and G. Ringham. Oxymetholone for the treatment of hiv-wasting: a double-blind, randomized, placebo-controlled phase iii trial in eugonadal men and women. HIV clinical trials, 4:150–163, 2003.
A. Sansone, F. Romanelli, M. Sansone, A. Lenzi, and L. Di Luigi. Gynecomastia and hormones. Endocrine, 55(1):37–44, 2017.
W. Schänzer. Metabolism of anabolic androgenic steroids. Clinical chemistry, 42(7):1001–1020, 1996.
Boatright, Jeffrey H.; Nickerson, John M.; Moring, Anisha G.; Pardue, Machelle T. (2009). “Bile acids in treatment of ocular disease”. Journal of Ocular Biology, Diseases, and Informatics. 2 (3): 149–159.
Nathanson MH et al. Stimulation of ATP secretion in the liver by therapeutic bile acids. Biochem J. 2001; 358(Pt 1):1-5.
Weitzel C et al. Ursodeoxycholic acid induced activation of the glucocorticoid receptor in primary rat hepatocytes. Eur J Gastroenterol Hepatol. 2005 Feb; 17(2):169-77.
Sanchez Pozzi EJ et al. Ursodeoxycholate reduces ethinylestradiol glucuronidation in the rat: role in prevention of estrogen-induced cholestasis. J Pharmacol Exp Ther. 2003 Jul; 306(1):279-86.
Arisawa S et al. Ursodeoxycholic acid induces glutathione synthesis through activation of PI3K/Akt pathway in HepG2 cells. Biochem Pharmacol. 2009 Mar 1;77(5):858-66.
Di AAS/SARM e saturazione recettoriale se ne parla spesso negli ambienti del culturismo “Enhancement“, nei social e nelle community online. Il problema è sempre il medesimo però, il quale colpisce altre argomentazioni le quali richiedono un certo livello culturale per essere trattate: se ne parla in modo confuso e male. Fortunatamente, però, su “Reddit” si tengono discussioni valide, e con letteratura al seguito, riguardo questo argomento, con persone “addette ai lavori”.
Quindi, l’obiettivo di questo breve articolo è principalmente quello di riportare i chiarimenti scientificamente supportati per ciò che concerne l’uso di AAS/SARM e la saturazione dei Recettori degli Androgeni.
Prima di proseguire, è giusto ricordare che ho una vasta conoscenza di biochimica e genetica e faccio ricerca e divulgazione scientifica da anni. Di conseguenza, le mie affermazioni non sono in alcun modo un “punto di vista” dal momento che, ed i miei lavori lo testimoniano già a sufficienza, ho una comprensione alquanto decente di ciò che viene riportato nelle pubblicazioni scientifiche.[1]
Vi espongo di seguito i “punti chiave” necessari per comprendere la questione AAS/SARM e saturazione AR:
I Recettori degli Androgeni nella maggior parte dei tessuti sono saturi all’estremità inferiore del normale intervallo fisiologico di Testosterone.
Nonostante questa saturazione, la crescita muscolare e la diminuzione della massa grassa è ancora legata al Testosterone in modo dipendente dalla dose, anche a livelli sovrafisiologici.
L’aumento della sintesi proteica non è l’unico (e forse non il principale) meccanismo attraverso il quale il Testosterone causa la crescita del muscolo-scheletrico.
Gli Androgeni sembrano causare un aumento delle cellule satelliti e dei mioonuclei nei muscoli. L’aggiunta di mionuclei alle fibre muscolari è uno dei meccanismi principali con cui essi crescono in dimensione. Questo aumento delle cellule satelliti e dei mionuclei avviene attraverso un percorso dipendente dal Recettore degli Androgeni.
In molti tessuti, l’aumento della concentrazione di Androgeni porta a un aumento della densità dei Recettori degli Androgeni. Questo può aiutare a dare una spiegazione alla possibilità di crescita potenziale maggiore “off cycle” attraverso il precedente uso di anabolizzanti. A tal proposito ricordiamoci anche della così detta “Memoria Muscolare”.[2]
L’aumento delle cellule satellite deriva dalla differenziazione delle cellule staminali mesodermiche pluripotenti. Queste sono le stesse cellule che si differenziano in adipociti (cellule del tessuto adiposo, quindi grasso). L’aumento della differenziazione di queste cellule in cellule satellite (che generano mionuclei) spiega il perché dosi più elevate di Androgeni portano a una diminuzione della massa grassa.
L’aumento delle cellule satelliti e dei mionuclei nella fibra muscolare è più che raddoppiato quando si confronta la somministrazione di 300mg vs. 600mg di Testosterone Enantato. Queste, ovviamente, sono già dosi sovrafisiologiche e questo dimostra l’opposto dei rendimenti decrescenti; tuttavia c’è ancora probabilmente un “collo di bottiglia” sconosciuto a questa differenziazione.
Notare le frecce nella figura C, che denotano fibre muscolari divise in un PowerLifter che aveva usato AAS nei precedenti 10 anni. Le fibre più piccole contenevano una isoforma in via di sviluppo della miosina (cioè miosina fetale), suggerendo che erano in realtà fibre di nuova formazione da iperplasia. La teoria qui esposta è che le fibre hanno una certa soglia di crescita, e che una volta raggiunta questa soglia, alla fine si dividono per formare nuove fibre. Con le tradizionali pratiche di allenamento “Natty”, non sembra che i PL raggiungano questa soglia; ma con l’uso di AAS, la crescita può diventare così accentuata che si verifica l’iperplasia (si noti la differenza di dimensioni delle fibre tra il PL “juiced” in Figura A e il sollevatore”Natty” in Figura B). Anche se mancano prove oggettive e inconfutabili, è logico supporre che le fibre aggiunte (e AR sovraespressi) vengano mantenute, anche se il sollevatore interrompe l’uso di AAS. La questione della possibile ipotrofia di queste nuove fibre una volta cessato l’uso di AAS è un altra possibilità.
Conclusioni:
Dosi più elevate di AAS/SARM o abbinamento di questi porteranno a risultati migliori? Ancora non lo sappiamo con certezza, sebbene i dati empirici ci portino ad una parziale conclusione favorevole al quesito posto. Per esempio, sappiamo che il Ki (con tale sigla ci si riferisce al potenziale di legame/saturazione del AR dose-dipendente) del RAD-140 è di 7nM (rispetto a 29nM del Testosterone e i 10nM del DHT).[3] Questo però non ci dà l’efficacia del ligando, ne il tasso di dissociazione (il testosterone si dissocia dal recettore degli androgeni a un tasso 5x rispetto al DHT nonostante abbia un Ki 2,9x maggiore [4]), ma se dovessimo usarlo come unico parametro di misurazione dell’efficacia, sembrerebbe così. Prendendo il tasso di biodisponibilità proposto del 65-75% (vedi riferimento Ki) del RAD nelle scimmie come punto di riferimento per gli esseri umani, un ciclo proposto di 10mg/die (concentrazione stabile intorno ai 25mg), sembrerebbe poter dare ancora dei benefici (e dei danni in termini di effetti collaterali) da dosi più elevate.
Un altro aspetto che non conosciamo è legato agli effetti AR-indipendenti del testosterone. Ci sono state proposte che collegano alcuni degli effetti del testosterone al suo antagonismo degli effetti dei glucocorticoidi attraverso il legame a bassa affinità con il recettore dei glucocorticoidi. Per quanto ne so, non abbiamo alcun indizio circa l’affinità di cui qualsiasi SARMs legano questo recettore.
Non sappiamo in termini assoluti se abbinare AAS/SARM apporti vantaggi superiori alla monoterapia, sebbene, e lo ripeto, i risultati empirici ci portano verso una risposta almeno parzialmente positiva. Ciò che bisogna evitare di fare, è smettere di usare affermazioni semplicistiche e riduttive come “la saturazione dei AR è il fattore principale che determina il tasso soggettivo di ipertrofia muscolare ottenibile”.
Esiste una interessantissima pubblicazione la quale suggerisce che sono le concentrazioni di Recettori degli Androgeni e non i livelli ormonali il fattore limitante della crescita muscolare a livelli fisiologici. Per l’appunto, LIVELLI FISIOLOGICI! Ancora una volta, vi ricordo di tenere a mente che gli androgeni sovraregolano i Recettori degli Androgeni in modo dose dipendente.[5]
Con il nuovo anno riprendo la pubblicazione degli articoli e lo faccio trattando un argomento che spesso, direttamente o indirettamente, è emerso nelle discussioni tra clienti e colleghi.
Il Testosterone è senza dubbio l’ormone simbolo per l’uomo della strada, preso dalla frenesia del mondo moderno e dal raggiungimento di obbiettivi tanto futili quanto irrealistici. Lo so che ve lo state domandando e la risposta è “si”. Questa entrata filosofica è perfetta per introdurre una questione legata al Androgeno per eccellenza.
Tanto per fare un esempio: quanti rimedi da banco vi sono stati proposti per migliorare i livelli plasmatici di Testosterone? Tra Tribulus Terrestris, Maca e Boro il conto è presto perso. E quanti di questi supplementi OTC hanno dato reali risultati? Misurabili, quantificabili con i livelli di partenza e che si sono tradotti in significativi miglioramenti della composizione corporea? …
La necessità di un ottimale apporto di Zinco, Vitamina D e altri macro e microelementi implicati nella biosintesi androgena, nella Testosterone:Estradiolo ratio ecc… non sono di certo messi in dubbio. Ad esserlo è il marketing, è l’affermazione sensazionalistica che va sempre con cura soppesata e valutata in concreto.
Ma, ipotizzando un miglioramento dei livelli di Testosterone endogeno rispetto al basale di partenza, ed entro l’intervallo di riferimento standard (es. per gli uomini dai 240 ai 950ng/dl dopo i 18 anni), garantite da trattamenti iatrogeni, quanto può incidere ciò nel miglioramento della composizione corporea?…
L’articolo che segue si basa sulle informazioni raccolte nella Research Review di James Krieger.
Iniziamo dalla letteratura scientifica
È assodato che l’uso AAS, che comporta la somministrazione di dosi sovrafisiologiche di Testosterone o ormoni correlati, provoca marcati aumenti delle dimensioni muscolari, ben oltre ciò che può essere ottenuto di base fisiologica dal soggetto, anche se questi guadagni addizionali sono limitati geneticamente. Nonostante ciò, mentre è assodato che dosi sovrafisiologiche di Testosterone, suoi derivati e analoghi aumenteranno significativamente i potenziali guadagni ipertrofici, questo non ci dice se le variazioni del Testosterone all’interno di un normale intervallo fisiologico possano avere qualche impatto. C’è una vasta gamma di livelli ematici di Testosterone da un uomo all’altro. Ad esempio, in uno studio nel quale sono stati presi in esame 456 uomini sani e non obesi di età compresa tra 19 e 39 anni, l’intervallo delle concentrazioni di Testosterone nel sangue (misurato al mattino dopo un digiuno notturno) era il seguente:
Lo studio di cui sopra è stato eseguito su un campione di individui della Framingham Heart Study Generation 3. Tuttavia, gli intervalli di concentrazione di Testosterone possono variare a seconda della popolazione e del dosaggio utilizzato per misurare il Testosterone. Anche i laboratori variano molto nei loro intervalli di riferimento. Travison et al. hanno estrapolato i dati da quattro importanti studi di coorte e hanno utilizzato modelli statistici per stabilire intervalli di riferimento che potrebbero essere applicati in diversi laboratori. Ecco la gamma di concentrazioni di Testosterone nel sangue che hanno stabilito:
Indipendentemente dall’intervallo di riferimento utilizzato, non c’è dubbio che vi sia un’ampia variazione nei livelli di Testosterone tra gli uomini, anche tra gli uomini sani e non obesi (poiché, come ben sappiamo, l’obesità è associata a un livello di Testosterone inferiore). Ciò solleva la questione se le variazioni in un intervallo normale possano influire sensibilmente sui guadagni muscolari. Un uomo con livelli di Testosterone naturalmente più alti riesce ad avere un maggior margine ipertrofico muscolare rispetto ad un uomo con livelli più bassi, anche se entrambi gli uomini sono all’interno di un intervallo normale? La risposta a questa domanda può avere particolare rilevanza per gli uomini che invecchiano. Il Testosterone diminuisce con l’età, ed è un altro dato di fatto, sebbene l’attività contro resistenza e una alimentazione sana possono rallentarne il declino. Ad esempio, ecco i dati dello studio French Telecom, che mostra il calo del Testosterone in tutti i percentili con l’età degli uomini.
Percentili di distribuzione plasmatica del Testosterone in un campione di 1.408 uomini caucasici dello studio Telecom, Parigi, Francia, 1985-1987
Ancora una volta, si può vedere l’ampia variazione nei livelli fisiologici di Testosterone, che vanno da 350-400ng/dL nel 5° percentile 850-1000ng/dL nel 95° percentile. Il declino continua negli anni ’60, ’70 e oltre. Ecco i dati che mostrano i livelli medi di Testosterone nei decenni di durata della vita; questi dati sono tratti da sei studi:
Testosterone totale (ng/ml) per fascia di età (moltiplicare per 100 per ottenere ng/dL); dati da 6 diversi studi.
Poiché anche la massa muscolare diminuisce con l’età e poiché gli uomini con bassi livelli di Testosterone mostrano tassi di perdita muscolare più rapidi rispetto agli uomini con livelli più alti, potremmo ipotizzare che gli uomini più anziani potrebbero trarre beneficio dal portare il Testosterone nell’intervallo fisiologico medio-alto.
Pertanto, tutti questi dati sollevano una serie di domande importanti:
I livelli di Testosterone di base sono correlati alla risposta all’allenamento?
Il Testosterone estremamente basso compromette la massa muscolare e i guadagni muscolari?
Le variazioni all’interno del normale range fisiologico influiscono sulla massa muscolare?
Se i livelli di Testosterone sono bassi o al limite, portare i livelli fino alla fascia media o superiore aiuta a migliorare la massa muscolare?
Se le variazioni nel normale range fisiologico hanno un impatto sulle condizioni muscolari negli uomini, hanno lo stesso impatto anche nelle donne?
Diamo un’occhiata alla ricerca per poter cercare di dare una risposta a queste domande.
I livelli basali di Testosterone sono correlati con la risposta all’allenamento?
Un modo per esaminare se esiste una relazione tra Testosterone in range fisiologico e guadagni muscolari è quello di guardare le risposte all’allenamento di un insieme di individui e vedere se i livelli di Testosterone di base sono correlati alla quantità di muscoli guadagnata da ciascuna persona. McCall et al. non hanno trovato alcuna correlazione tra i livelli basali di Testosterone e i cambiamenti nella dimensione muscolare in giovani uomini allenati a livello amatoriale. Tuttavia, Ahtiainen et al. hanno trovato una forte correlazione tra i livelli di Testosterone di base e il miglioramento della forza isometrica massima in 21 settimane. Ma la correlazione con l’ipertrofia non è stata affrontata.
Data la relazione tra l’ipertrofia e l’espressione di forza isometrica, potremmo ipotizzare che ci fosse una relazione tra il Testosterone di base e l’ipertrofia in questo studio, ma non è possibile saperlo con certezza.
In uno studio di Bhasin et al., uomini con infezione da HIV con Testosterone basso (<349ng/dL) sono stati assegnati in modo casuale a gruppo placebo, solo allenamento contro-resistenza, solo iniezioni di Testosterone o Testosterone e allenamento contro-resistenza combinati.
I livelli di Testosterone al basale non erano correlati con la variazione della massa magra (FFM) e non c’erano differenze significative nel guadagno assoluto di FFM tra uomini che avevano livelli di Testosterone <275ng/dL e uomini che avevano livelli di 275-350ng/dL .
E’ possibile anche confrontare i guadagni muscolari tra maschi e femmine, poiché gli uomini hanno 10 volte più Testosterone delle donne. Se i livelli di Testosterone di base fossero correlati con la risposta all’allenamento, ci aspetteremmo che gli uomini abbiano maggiori guadagni rispetto alle donne. Tuttavia, quando uomini e donne vengono sottoposti a programmi di allenamento contro-resistenza, mentre i guadagni muscolari assoluti sono maggiori negli uomini, i guadagni muscolari relativi (cioè i guadagni percentuali) sono per lo più simili.
Nel complesso, questi dati limitati suggerirebbero che i livelli di Testosterone non influiscono realmente sui guadagni. Tuttavia, si tratta di dati trasversali e non sono realmente progettati per affrontare la questione se le variazioni del Testosterone fisiologico abbiano un impatto sui guadagni di massa muscolare.
Pertanto, è necessario esaminare alcune ricerche in cui i livelli di Testosterone vengono direttamente manipolati.
Livelli di Testosterone estremamente bassi compromettono la massa muscolare e i guadagni muscolari?
Un modo per esaminare l’impatto del Testosterone sui guadagni muscolari è vedere cosa succede quando si sopprime la produzione di Testosterone. Maura et al. ha somministrato a giovani uomini il Lupron, un farmaco antiandrogeno che sopprime la produzione naturale di Testosterone. I livelli di Testosterone sono scesi da 535ng/dL a 31ng/dL dopo 10 settimane. Pertanto, la media dei soggetti trattati con Lupron aveva livelli di Testosterone simili a quelli di una donna.
La massa magra è diminuita di 2,1 kg e la sintesi proteica dell’intero corpo è diminuita del 13%. Naturalmente, non c’era alcun tipo di allenamento in questo studio. Forse l’allenamento con i pesi potrebbe interagire con questa risposta.
Kvorning et al. hanno somministrato a giovani uomini il Goserelin, che sopprime la produzione naturale di Testosterone, o un placebo. Gli uomini, che avevano una minima esperienza di allenamento contro-resistenza, si sono impegnati in un programma di allenamento della forza di 8 settimane. I livelli di Testosterone sono scesi da 651ng/dL a 57ng/dL, e poi 31ng/dL nel gruppo che ha ricevuto il Goserelina.
La soppressione del Testosterone non ha compromesso i miglioramenti nelle prestazioni del 10-RM rispetto al placebo. Tuttavia, i miglioramenti nella forza isometrica erano significativamente inferiori con il Goserelina.
I miglioramenti nella massa magra delle gambe erano significativamente inferiori per il gruppo Goserelina e anche la massa corporea magra totale tendeva verso quella direzione (valore P di 0,07, dove 0,05 è considerato significativo). L’aumento medio della massa magra è stato di 1kg maggiore nel gruppo placebo rispetto al gruppo Goserelina. La differenza nella massa magra della gamba era di 0,2kg.
Pertanto, questo studio ha dimostrato che la soppressione della produzione di Testosterone ha compromesso i guadagni di massa magra, ma la differenza non era marcata, pari a circa 1kg di differenza complessiva nei guadagni di massa magra in 8 settimane.
Non sono state eseguite misurazioni dirette della dimensione muscolare, sebbene le grandi differenze nei guadagni di forza isometrica probabilmente indichino che i guadagni muscolari erano inferiori con la soppressione del Testosterone. Quindi, questi dati suggerirebbero che c’è un impatto del Testosterone sul guadagno muscolare, pur essendo di piccola entità.
Molecola di Goserelina. La Goserelina è un agonista delle gonadotropine iniettabile (agonista GnRH), conosciuta anche come agonista dell’Ormone di Rilascio dell’Ormone Luteinizzante (LHRH).
Quindi le variazioni all’interno dell’intervallo fisiologico normale influiscono sulla massa muscolare?
Nessuna delle ricerche discusse finora può davvero dirci se le variazioni all’interno del normale range fisiologico possono avere un impatto sulla massa muscolare. Ci sono tre modi in cui è possibile rispondere a questa domanda. Il primo modo è guardare ai dati trasversali. Più semplicemente si tratta di prendere grandi gruppi di uomini e di dividerli in categorie in base ai loro livelli di Testosterone. Quindi si osserva se la massa muscolare differisce tra gli uomini in diverse categorie o se i livelli di Testosterone sono correlati ai livelli di massa muscolare.
He et al. hanno esaminato 270 uomini sedentari dell’HERITAGE Family Study. Dopo aver controllato per età e ascendenza, il Testosterone non era correlato alla massa magra. È interessante notare, tuttavia, che era correlato negativamente con l’indice di massa magra (FFM diviso per altezza al quadrato, simile all’IMC), il che significa che le persone con un indice FFM più elevato avevano livelli più bassi di Testosterone. Questo significa che avere più Testosterone significa in realtà avere meno muscoli? No! Questi dati sono confusi dal fatto che alcuni degli uomini erano obesi e che l’indice di massa corporea più elevato e le percentuali di grasso corporeo più elevate erano associate a un livello di Testosterone più basso. Ecco i livelli di testosterone per quartili di BMI; è possibile notare che i livelli di Testosterone diminuiscono all’aumentare dell’IMC.
Poiché gli uomini obesi hanno anche più FFM, questo può far credere che ci sia una relazione negativa tra FFM e livelli di Testosterone. Ciò di cui si ha bisogno per comprendere la questione è una ricerca che esamini la relazione negli individui non obesi.
Testosterone per quartili di BMI negli uomini. 1 nmol/L = 0,0347 ng/dL
Poiché gli uomini obesi hanno anche più FFM, questo può far sembrare che ci sia una relazione negativa tra FFM e livelli di Testosterone. Ciò di cui si necessita per comprendere la questione è una ricerca che esamini la relazione negli individui non obesi.
Van Den Beld et al. non hanno trovata alcuna relazione tra Testosterone e massa magra negli uomini anziani (età 73-94 anni).
Mouser et al. hanno raccolto dati sul Testosterone e sulla composizione corporea di 252 uomini nel National Health And Nutrition Examination Survey (NHANES) del 1999-2000 di età compresa tra 18 e 85 anni. Uomini che non rientravano nell’intervallo normale per il Testosterone (da 240 a 950ng/dL). ) non sono stati inclusi nell’analisi. Gli uomini sono stati suddivisi in quartili in base ai loro livelli di Testosterone. Gli uomini hanno mostrato quantità progressivamente più elevate di massa magra nella parte inferiore del corpo con livelli crescenti di Testosterone, anche dopo aver aggiustato la media per età, razza, presenza di diabete, partecipazione auto-riferita all’attività fisica, proteina C-reattiva e assunzione di proteine nella dieta. I quartili 3 e 4 erano statisticamente significativi rispetto al quartile 1. Un modello in qualche modo simile è emerso per la parte superiore del corpo, sebbene non vi fosse alcuna differenza tra il quartile 3 e 4.
Questi dati hanno mostrato che gli uomini nel 3° quartile avevano il 14,2% in più di massa magra nell’area inferiore e il 5,6% in più di massa magra in quella superiore rispetto agli uomini nel 1° quartile. Gli uomini del 4° quartile avevano il 22,1% in più di massa magra nell’area inferiore e il 5,6% in più di massa magra in quella superiore rispetto agli uomini del 1° quartile. Se si prendesse un ipotetico uomo nel 1° quartile con 17kg di massa magra nell’area inferiore, si potrebbe prevedere che un uomo nel 3° quartile possa avere 19,4kg e un uomo nel 4° quartile 20,6kg. Pertanto, questi dati hanno mostrato che gli uomini nell’estremità superiore dell’intervallo fisiologico del Testosterone avevano una massa corporea magra maggiore rispetto agli uomini nell’estremità inferiore, anche tenendo conto di altre variabili che potrebbero influenzare il Testosterone.
I dati trasversali di Mouser indicano che esiste potenzialmente una relazione tra i livelli di Testosterone nell’intervallo fisiologico e la massa magra di cui si dispone. Tuttavia, un problema con i dati trasversali è che non possono stabilire causa ed effetto. Un altro modo in cui è possibile affrontare la questione se le variazioni all’interno di un intervallo fisiologico influiscano sulla massa muscolare è sopprimere la produzione naturale di Testosterone usando farmaci, quindi somministrare dosi diverse di Testosterone e osservare se c’è un effetto dose-risposta. Ci sono quattro studi che hanno fatto questo.
Shalendar Bhasin et al. hanno somministrato a giovani uomini sani un agonista dell’ormone di rilascio delle gonadotropine (GnRH) per sopprimere la secrezione endogena di Testosterone. Hanno quindi somministrato agli uomini iniezioni settimanali di 25, 50, 125, 300 o 600mg di Testosterone Enantato per 20 settimane. Ecco i livelli ematici medi di Testosterone per le diverse dosi; come prevedibile, i livelli ematici sono aumentati con l’aumentare delle dosi e le dosi da 300 e 600mg hanno ovviamente portato a livelli di Testosterone al di sopra del normale intervallo fisiologico.
C’è stato un aumento dose-dipendente della massa magra; maggiori livelli ematici di Testosterone hanno portato a maggiori aumenti della FFM.
Anche il volume muscolare della coscia è aumentato in modo dose-dipendente.
La variazione della massa magra e la variazione del volume muscolare del quadricipite erano significativamente correlate con i livelli ematici di Testosterone.
Nel complesso, questo studio ha mostrato un effetto dose-risposta del Testosterone sulla dimensione muscolare, anche all’interno dell’intervallo fisiologico. Infatti, il solo passaggio dalla fascia bassa del fisiologico (306ng/dL) alla fascia media (542ng/dL) ha comportato un aumento della massa magra di 2,8 kg.
Bhasin ha ripetuto lo stesso esperimento in uomini più anziani di età compresa tra 60 e 75 anni. I risultati erano molto simili; i grafici seguenti mostrano gli effetti dose-risposta negli uomini più anziani.
Un terzo studio di Shalendar Bhasin ha coinvolto un design simile. La secrezione naturale di Testosterone è stata soppressa utilizzando il Lupron in uomini sani di età compresa tra 18 e 50 anni. Agli uomini sono state quindi somministrate dosi di 50, 125, 300 o 600 mg/settimana di Testosterone Enatnato, con o senza un inibitore della 5α-reduttasi (un farmaco che blocca la conversione del Testosterone in Diidrotestosterone [DHT]). I risultati sono stati ancora una volta simili, con una maggiore massa magra all’aumentare dei livelli ematici di Testosterone.
Finkelstein et al. hanno somministrato la Goserelina a 198 uomini sani di età compresa tra 20 e 50 anni per sopprimere i loro livelli di Testosterone. Sono stati quindi assegnati in modo casuale a ricevere giornalmente un gel placebo, o 1,25g, 2,5g, 5g o 10g di un gel contenente Testosterone per 16 settimane. Altri 202 uomini sono stati sottoposti allo stesso protocollo, tranne per il fatto che hanno ricevuto anche un inibitore dell’Aromatasi (Anastrozolo) per sopprimere la conversione del Testosterone in Estradiolo. C’è stato un effetto dose-risposta delle diverse dosi di Testosterone sui livelli ematici del ormone in questione, che vanno da al di sotto dell’intervallo normale fisiologico per le dosi di 0 e 1,25g, fino all’estremità superiore dell’intervallo fisiologico per la dose di 10g. Le barre nere rappresentano il gruppo trattato con Anastrozolo, mentre le barre rosse rappresentano il gruppo non trattato con Anastrozolo.
Livelli di Testosterone nel sangue con diverse dosi di un gel contenente Testosterone, dopo la soppressione del Testosterone endogeno con Goserelina. Le barre rosse rappresentano un gruppo che ha ricevuto Anastrozolo, un inibitore dell’aromatasi, per ridurre la conversione del Testosterone in Estradiolo. Dati da Finkelstein et al., NEJM, 2013
I cambiamenti nella massa magra e nell’area muscolare della coscia hanno mostrato un po’ di effetto dose-risposta, anche se non così chiaro come gli quanto osservato negli articoli di Bhasin che hanno utilizzato somministrazione per iniezioni. Nessuna dose di Testosterone ha provocato una significativa perdita di massa magra, mentre la dose più alta ha portato al guadagno maggiore di questa, sebbene molte delle differenze non fossero statisticamente significative. I numeri uguali non indicano differenze statisticamente significative rispetto ad altre barre.
C’era un’enorme quantità di variazione nel modo in cui gli individui rispondevano al Testosterone, come si può vedere in questo grafico a dispersione.
Un terzo modo per esaminare se le variazioni in un intervallo fisiologico influiscono sui guadagni muscolari è vedere se portare il +stosterone al limite molto superiore dell’intervallo normale (come quello che si verifica negli studi sui contraccettivi maschili di Testosterone) influisce sulla massa magra. Herbst et al. ha studiato l’impatto del testosterone esogeno (100 mg di testosterone enathnato a settimana) su uomini sani con normali livelli di testosterone. I livelli di testosterone sono aumentati da 570 ng/dL a 734 ng/dL (il livello subito prima dell’iniezione successiva), con un picco di 1196 ng/dL (24 ore dopo l’iniezione). Pertanto, il livello di picco era al limite molto superiore del range di normalità e il minimo era nella parte superiore del normale. Massa magra aumentata di 2,5 kg. Nel complesso, questi tre corpi di prove (dati trasversali, dati sulla risposta alla dose e dati sui contraccettivi maschili) indicano che le variazioni all’interno dell’intervallo fisiologicamente normale influiscono sulla massa magra che si trasporta. Ora, qui c’è una differenza tra quanta massa magra porti e quanto guadagnerai da un programma di allenamento (ne parleremo più avanti), ma sembra esserci un effetto. Il che ci porta alla nostra prossima domanda…
Se i livelli di Testosterone sono bassi o al limite del limite basso, portare i livelli fino al livello medio o superiore aiuta a migliorare la condizione della massa muscolare?
Partendo dal precedente quesito, cosa succede se si prendono delle persone con bassi livelli di Testosterone e li si aumenta i livelli con iniezioni di Testosterone esogeno? Fortunatamente c’è la ricerca a darci una risposta.
Urban et al. hanno reclutato 6 uomini sani e anziani con un’età media di 67 anni. I loro livelli di Testosterone erano di 480ng/dL o meno e sono stati somministrati loro iniezioni di Testosterone per 4 settimane per raggiungere livelli simili a quelli degli uomini più giovani. Sia la forza muscolare che la sintesi proteica muscolare sono migliorate, suggerendo che aumentare i livelli all’interno dell’intervallo fisiologico può aiutare a migliorare le condizioni della massa e la forza muscolare. Una limitazione è che la massa muscolare non è stata direttamente misurata; sono state determinate solo la sintesi proteica muscolare e la forza.
Sullivan et al. hanno reclutato 71 uomini di età compresa tra 65 e 93 anni e li hanno assegnati in modo casuale a uno di 4 gruppi:
Esercizio a bassa resistenza (3 x 8 con 20% del 1-RM) + Placebo
Esercizio a bassa resistenza + 100 mg/settimana di Testosterone
Esercizio di resistenza ad alta intensità (3 x 8 all’80% del 1-RM) + Placebo
Esercizio di resistenza ad alta intensità + 100 mg/settimana di Testosterone
Le iniezioni di Testosterone hanno più che raddoppiato i livelli del ormone rispetto al placebo, portando i livelli alla fascia alta del normale (804 ng/dL contro 304 ng/dL). Anche i guadagni nell’area della sezione trasversale dei muscoli a metà coscia sono stati più del doppio con le iniezioni di Testosterone rispetto al placebo. I guadagni di forza erano molto più alti nella condizione di esercizio a bassa resistenza quando veniva somministrato Testosterone rispetto al placebo. Tuttavia, quando l’allenamento era ad alta intensità, non c’era più un beneficio significativo del Testosterone, indicando che il carico di allenamento aveva un impatto maggiore sulla forza rispetto al Testosterone in questo studio.
Bhasin et al. hanno trattato uomini ipogonadici (età 19 – 47 anni) con 100mg di Testosterone Enantato a settimana per 10 settimane. I livelli medi di Testosterone al basale sono aumentati da 72ng/dL (leggermente al di sopra dell’intervallo per una donna media) a 767ng/dL alla settimana 10. La massa magra è aumentata di 5kg, la dimensione del tricipite è aumentata del 12% e la dimensione del quadricipite è aumentata del 8%.
Bhasin et al. hanno reclutato uomini con infezione da HIV con bassi livelli di Testosterone e li hanno trattati con una crema topica di Testosterone per 12 settimane. I livelli di Testosterone sono migliorati da 258ng/dL a 367ng/dL. La massa magra è aumentata di 1,4kg.
In un altro studio del Dr. Bhasin, uomini con infezione da HIV con bassi livelli di Testosterone (<349 ng/dL) sono stati assegnati in modo casuale a uno dei 4 seguenti gruppi:
Placebo
Testosterone Enatnato (100 mg/settimana)
Allenamento di resistenza
Testosterone + allenamento di resistenza
Il trattamento con Testosterone ha aumentato i livelli ematici da una media di 201 – 205ng/dL a 311 – 337ng/dL. La massa magra è aumentata di 4kg nel gruppo solo Testosterone, 2kg nel gruppo solo allenamento e 1,6kg nel gruppo allenamento + Testosterone. Il volume muscolare della coscia è aumentato di 40cm³ nel gruppo solo Testosterone, 62cm³ nel gruppo solo allenamento e 44cm³ nel gruppo combinato. Non è chiaro il motivo per cui non vi è stato alcun effetto combinato nel gruppo allenamento + Testosterone.
Sattler e colleghi hanno somministrato a uomini anziani (età media 71 anni) 5 o 10g al giorno di Testosterone transdermico (formulazione per somministrazione sulla pelle). Gli uomini trattati avevano livelli ematici di Testosterone di 550ng/dL o meno (la concentrazione media era 385 nel gruppo 5g/die e 350 nel gruppo 10g/die). Le concentrazioni medie di Testosterone sono aumentate di 150ng/dl nel gruppo 5g/die (aumentando i livelli a circa 535ng/dl) e 500ng/dl nel gruppo 10g (aumentando i livelli a circa 850ng/dl). La massa corporea magra è aumentata di 1kg nel gruppo 5g/die e di 1,6kg nel gruppo 10g/die.
Basaria et al. hanno reclutato uomini con Testosterone basso (<350 ng/dL) a causa dell’abuso di oppiacei e hanno somministrato loro un 5g/die di gel contenente Testosterone. Il testosterone medio è aumentato da 243ng/dL a 790ng/dL. La massa magra aumentata di 1kg.
Storer et al. hanno reclutati uomini di età superiore ai 59 anni con livelli di Testosterone tra 100 e 400ng/dL. Agli uomini è stato somministrato 7,5g di un gel contenente Testosterone o un placebo al giorno per 3 anni. Il Testosterone ematico è aumentato da 307 ng/dL a 567 ng/dL nel gruppo gel. La massa corporea magra è aumentata di 0,7kg.
Brodsky e colleghi hanno osservato gli effetti della somministrazione di Testosterone negli uomini con livelli di questo ormone inferiori a 200 ng/dL. I livelli di Testosterone sono aumentati gradualmente da 106 ng/dL a 576 ng/dL in 4 mesi. E da 432 ng/dL in 6 mesi. Queste erano le concentrazioni più basse osservate prima di ogni iniezione. Le iniezioni sono state somministrate ogni 2 settimane; l’ultimo livello di Testosterone misurato è stato una settimana dopo l’ultima iniezione ed era di 1277ng/dL, appena al di sopra del range fisiologico. La massa magra è aumentata di 8,7kg. La sintesi proteica muscolare mista è aumentata del 56% e la sintesi proteica miofibrillare è aumentata del 46%. La sintesi proteica muscolare totale in tutto il corpo è aumentata del 71-87% (da circa 2,4 grammi all’ora a 4,3 grammi all’ora).
Snyder et al. hanno somministrato a uomini con Testosterone basso (a causa di una malattia) un cerotto cutaneo con Testosterone per 3 anni. I livelli di Testosterone sono aumentati da 78ng/dL a 407ng/dL. La massa magra è aumentata di 3,1kg.
Wang e colleghi hanno somministrato a uomini con bassi livelli di Testosterone un cerotto o uno dei due diversi gel (50mg o 100 mg/giorno) per 90 giorni. Il Testosterone è aumentato da 236 ng/dL a 417 ng/dL nel gruppo cerotto, da 236 ng/dL a 552 ng/dL nel gruppo del gel da 50mg e da 248 ng/dL a 791 ng/dL nel gruppo del gel da 100mg. La massa corporea magra è aumentata rispettivamente di 1,2 kg, 1,3 kg e 2,7 kg in questi gruppi.
In un altro studio di Wang, agli uomini con bassi livelli di Testosterone (<300 ng/dL) sono state somministrate varie dosi di gel di Testosterone per un massimo di 42 mesi. I livelli totali sono aumentati di circa 260 ng/dL e sono rimasti nell’intervallo medio-basso normale per la durata dello studio. La massa magra è aumentata di 2,9kg.
Tenover ha reclutato uomini di età compresa tra 57 e 76 anni, con livelli di Testosterone inferiori a 400 ng/dL, e ha iniettato loro 100 mg di Testosterone Enantato a settimana. I livelli ematici medi sono aumentati da 334 ng/dl a 568 ng/dl. La massa magra è aumentata di 1,8kg.
Snyder e colleghi hanno assegnato casualmente a uomini di età superiore ai 65 anni un cerotto di Testosterone o a un placebo. I livelli di Testosterone sono aumentati da 367 ng/dL a 625 ng/dL in 6 mesi. La massa magra è aumentata di 1,6 kg in 6 mesi.
Ferrando et al. hanno reclutato uomini di età pari o superiore a 60 anni e con livelli di Testosterone nel sangue inferiori a 480 ng/dL somministrando loro Testosterone Enatnato o un placebo su base settimanale per 6 mesi. Il livello medio basale di Testosterone era 363 ng/dL. Le iniezioni di Testosterone sono state regolate individualmente per cercare di mantenere un livello ematico tra 490 e 807 ng/dL, sebbene ciò non abbia avuto un successo totale e molti individui abbiano riscontrato livelli leggermente superiori a tale obiettivo. Il livello medio a 6 mesi era di 882 ng/dL. La massa magra è aumentata di 4,2 kg, mentre è diminuita di 2 kg nel gruppo placebo. Il volume muscolare delle gambe è aumentato di 488 ml, mentre è diminuito di 96 ml nel gruppo placebo. Il Testosterone ha anche determinato un aumento dell’equilibrio netto delle proteine muscolari, a causa di una diminuzione del catabolismo delle proteine muscolari.
Dias et al. hanno reclutato uomini di età compresa tra 65-82 anni e livelli di Testosterone <350 ng/dL assegnandoli in modo casuale a un placebo, Anastrozolo (un inibitore dell’Aromatasi) o un gel di Testosterone. Inibendo l’enzima Aromatasi, l’enzima che converte il Testosterone in Estradiolo, è possibile aumentare efficacemente i livelli di Testosterone, ed è una cosa largamente risaputa. L’Anastrozolo ha aumentato il Testosterone da 272 ng/dL a circa 500 ng/dL a 6 mesi; la massa magra è aumentata di 1,5kg. Il Testosterone somministrato attraverso il gel ha aumentato i livelli dell’ormone da 300 ng/dL a circa 650 ng/dL e la massa magra non ha raggiunto un aumento statisticamente significativo (0,9 kg).
Magnusson e colleghi hanno randomizzato soggetti diabetici di tipo 2 di età compresa tra 50 e 70 anni trattandoli con un Testosterone gel o un placebo per 6 mesi. I livelli di Testosterone sono aumentati da 205 ng/dL a 637 ng/dL e la massa magra è aumentata di 1,9 kg.
Ribeiro e Abucham hanno somministrato a uomini ipogonadici Clomifene Citrato, il quale causa un aumento del Testosterone endogeno legandosi ai recettori degli estrogeni ipotalamici. Legandosi ai recettori degli estrogeni, induce il cervello a percepire che non ci siano così tanti estrogeni nel corpo per garantire l’omeostasi. Questo porta ad un aumento di GnRH seguito da LH e FSH. L’LH (Ormone Luteinizzante), stimola le cellule di Leydig nei testicoli a sintetizzare più Testosterone. I livelli di Testosterone nei “responder” sono aumentati da 201 ng/dL a 435 ng/dL dopo 3 mesi e la massa magra è aumentata di 1 kg.
Liu et al. hanno reclutato uomini più anziani con bassi livelli di Testosterone e somministrato loro iniezioni di gonadotropina corionica umana (HCG). L’HCG, mimando l’LH, stimola i testicoli a sintetizzare Testosterone. Il Testosterone è aumentato da 320 ng/dL a circa 720 ng/dL e la massa magra è aumentata di 2 kg.
Bayram et al. hanno somministrato a uomini ipogonadici iniezioni di HCG. Il Testosterone è aumentato da 39 ng/dl (nell’intervallo) a 512 ng/dl, con un aumento di 473 ng/dl. La massa magra è aumentata di 2,8 kg.
In uno studio di Casaburi et al., uomini con BPCO e basso livello di Testosterone (≤400 ng/dL) sono stati assegnati in modo casuale a uno dei 4 gruppi:
-Placebo -Testosterone Enantato (100 mg/week) -Resistance Training -Testosterone + Resistance Training
Le iniezioni di Testosterone hanno aumentato i livelli ematici da 302 ng/dL nel gruppo senza allenamento a 595 ng/dL e da 408 ng/dL a 656 ng/dL nel gruppo con allenamento. La massa magra è aumentata di 2,3 kg nel gruppo solo Testosterone, 0,2 kg nel gruppo solo resistance training e 3,29 kg nel gruppo Testosterone + resistance training. Va notato che l’allenamento è stato eseguito solo nella parte inferiore del corpo. Se si osservano i guadagni di massa magra delle gambe, erano 1,07kg nel gruppo solo Testosterone, 0,49 kg nel gruppo solo resistance training e 1,41 kg nel gruppo combinato.
Ecco un riassunto di tutti questi studi appena discussi, in cui i livelli bassi o al limite del livello di Testosterone sono stati aumentati in un intervallo fisiologico utilizzando iniezioni o sistemi di somministrazione transdermica. Puoi vedere che tutti hanno mostrato impatti positivi sulla massa magra. Alcuni hanno avuto aumenti relativamente piccoli del Testosterone (come circa 100-250 ng/dL) e hanno mostrato aumenti significativi della massa magra di circa 1-2 kg. In alcuni di questi studi, i soggetti si trovavano nella fascia più bassa del range di normalità (piuttosto che al di sotto del range di riferimento), e anche con quei soggetti, portare i livelli fino alla fascia medio-alta del range fisiologico ha avuto benefici positivamente apprezzabili.
E’ possibile vedere che i guadagni di massa magra con le iniezioni ( righe arancioni) tendono ad essere maggiori rispetto alla somministrazione transdermica ( righe grigie). Ciò è probabilmente legato al fatto che le iniezioni causano un picco iniziale di Testosterone che può essere all’estremità superiore dell’intervallo fisiologico, se non superare leggermente l’intervallo fisiologico.
Tipicamente, in questi studi, il Testosterone viene misurato 1-2 settimane dopo l’iniezione, rappresentando il minimo o il livello più basso di Testosterone. Pertanto, i livelli finali di Testosterone non rappresentano i livelli di picco raggiunti. Si può vedere questa differenza quando si guarda lo studio di Brodsky et al, dove il livello di picco, misurato 1 settimana dopo l’iniezione, era tre volte superiore al livello più basso, misurato 2 settimane dopo l’iniezione. Pertanto, tutti questi studi sulla terapia sostitutiva del Testosterone (TRT) dimostrano un beneficio nella massa magra dallo spostamento al di sotto dell’intervallo fisiologico, o dall’estremità inferiore dell’intervallo fisiologico, all’intervallo fisiologico medio o alto. Pertanto, sembra che anche il passaggio da una fascia bassa della gamma fisiologica a quella superiore abbia un vantaggio.
Testosterone più alto = Muscolo basale più alto, solo tassi di guadagno leggermente maggiori.
Nel complesso, i dati trasversali, gli studi dose-risposta, gli studi sui contraccettivi maschili e gli studi su TRT (compresi quelli in cui il Testosterone al basale era ancora normale) mostrano che le variazioni del Testosterone all’interno dell’intervallo fisiologico hanno un impatto sulla massa magra e muscolare. Questo significa che qualcuno con un livello di Testosterone più alto guadagnerà più velocemente di qualcuno con un livello più basso? Non proprio. Quando si guarda il corpo delle prove, il maggiore impatto del Testosterone sembra essere sul mantenimento di un certo livello di base della massa muscolare, piuttosto che sul tasso di guadagno muscolare. Ad esempio, i malati di cancro alla prostata sono spesso sottoposti a terapia di deprivazione di androgeni, in cui i loro livelli di Testosterone sono stati soppressi. In questo studio, il Testosterone medio era 45,7 ng/dL (all’interno dell’intervallo di una donna), rispetto a 430 ng/dL per i controlli. La sintesi proteica muscolare a riposo e a stomaco pieno era più bassa nei pazienti deprivati di androgeni. Tuttavia, quando l’alimentazione è stata combinata con l’allenamento contro-resistenza, la risposta alla sintesi proteica muscolare non era statisticamente diversa dai controlli (sebbene la media grezza fosse ancora leggermente inferiore).
Tassi di sintesi proteica muscolare in soggetti di controllo rispetto a pazienti in terapia di deprivazione androgenica (ADT). La sintesi proteica muscolare è significativamente più bassa a riposo e anche dopo un pasto (FED). Tuttavia, dopo l’allenamento contro-resistenza, la sintesi proteica muscolare non è significativamente diversa dai controlli dopo un pasto (EX-FED).
Mentre il guadagno percentuale nello studio di cui sopra era leggermente favorito negli uomini, non lo era di molto. Tuttavia, il guadagno assoluto è stato quasi il doppio di quello negli uomini rispetto alle donne. Pertanto, un aumento del 15% della massa muscolare negli uomini sarà generalmente maggiore su base assoluta rispetto alle donne, poiché gli uomini hanno una linea di base più ampia.
Una terza linea di supporto a questo concetto viene dal famoso studio Bhasin del 1996 sugli steroidi anabolizzanti. In questo studio di 10 settimane, uomini normali sono stati assegnati in modo casuale a uno dei quattro gruppi:
Placebo senza allenamento
Testosterone senza allenamento
Placebo con allenamento contro-resistenza
Testosterone con allenamento contro-resistenza.
Il testosterone è stato somministrato in dosi sovrafisiologiche (600 mg/settimana). I livelli di testosterone sono stati elevati a 2828 – 3244 ng/dL con le iniezioni, rispetto ai livelli normali di 453 – 667 ng/dL nel gruppo placebo. L’iniezione di testosterone, senza allenamento, ha comportato un aumento della massa magra di 3,2 kg. Il solo allenamento ha comportato un aumento della massa magra di 2 kg. Quando l’allenamento è stato combinato con l’iniezione di testosterone, l’aumento di massa magra è stato di 6,1 kg. La dimensione del muscolo quadricipite è aumentata in modo simile nel gruppo testosterone + nessun allenamento e nel gruppo solo allenamento, mentre i guadagni sono stati raddoppiati nel gruppo testosterone + allenamento.
La cosa interessante qui è che, quando guardi questi dati, puoi vedere che c’era principalmente un effetto additivo, piuttosto che sinergico, del testosterone e dell’allenamento. La FFM è aumentata di 3,2 kg con il solo testosterone. È aumentato di 2 kg solo con l’allenamento. Quando sommi queste due quantità, ottieni 3,2 + 2 = 5,2 kg, che è ragionevolmente vicino al guadagno di 6,1 kg osservato nel gruppo combinato. In altre parole, solo circa 0,9 kg potrebbero essere spiegati da un effetto sinergico tra testosterone e allenamento. Puoi anche vedere un effetto simile per l’aumento delle dimensioni del quadricipite. Il cambiamento nella dimensione del quadricipite era più o meno lo stesso nei gruppi solo testosterone e solo allenamento, ed era per lo più additivo nel gruppo combinato. Ancora una volta, solo una piccola parte del cambiamento nella dimensione del quadricipite potrebbe essere spiegata da un effetto sinergico tra testosterone e allenamento.
Questo è simile ai dati osservati quando osserviamo uomini e donne. I maggiori livelli di testosterone negli uomini non aumentano molto il tasso di guadagno rispetto alle donne; è solo che dà agli uomini una linea di base più alta per cominciare, e quindi i guadagni assoluti sono maggiori. Se ci fosse un forte effetto sinergico tra testosterone e allenamento, allora i guadagni relativi negli uomini sarebbero significativamente maggiori rispetto alle donne, ma ovviamente non è così.
Una quarta evidenza viene dallo studio di Casaburi e colleghi sugli uomini con BPCO, di cui si è brevemente accennato in precedenza. In questo studio è stato eseguito solo l’allenamento delle gambe. I guadagni di massa magra delle gambe nel gruppo combinato allenamento+testosterone erano 1,41 kg, che è vicino alla somma dei guadagni sperimentati dal gruppo solo testosterone (1,07 kg) e il gruppo solo allenamento (0,49 kg). Questo suggerisce ancora una volta che gli effetti del testosterone e dell’allenamento di resistenza sono per lo più additivi e non sinergici.
Pertanto, i tuoi livelli di testosterone influenzano la quantità di muscoli che porti in giro, indipendentemente dal fatto che ti alleni o meno. Quindi, quando inizi ad allenarti, la tua reattività all’allenamento è per lo più simile indipendentemente dal fatto che tu abbia livelli di testosterone bassi o alti. Potrebbe essere un po’ meno con un testosterone più basso, ma l’impatto maggiore è sulla tua linea di base.
Ad esempio, supponiamo che tu abbia una massa magra di base di 50 kg e che tu abbia un livello di testosterone nella fascia bassa (diciamo circa 300 ng/dL). Guadagni il 10% in 6 mesi, ovvero 5 kg.
Ora, prendi la stessa situazione, ma la persona ha un testosterone di base a 600 ng/dL. La tua massa magra di base ora potrebbe essere di 52 kg. Guadagni ancora il 10% in 6 mesi, ovvero 5,2 kg. Pertanto, il guadagno relativo è simile. Tuttavia, il punto di partenza e il guadagno assoluto sono maggiori a causa del testosterone più alto.
Consideriamo un altro esempio. Diciamo che una persona segue un qualche tipo di sostituzione del testosterone, aumentando il testosterone da 250 ng/dL a 500 o 600 ng/dL. Quella persona inizialmente sperimenterà alcuni guadagni relativi superiori al normale, mentre si muove verso la sua nuova linea di base per il suo nuovo livello di testosterone. Ti sembrerà di guadagnare da “principiante”. Tuttavia, una volta che quella persona ha raggiunto la sua nuova linea di base, i suoi guadagni relativi saranno simili a quando aveva un testosterone più basso.
Quantificare l’impatto delle variazioni del Testosterone fisiologico sulla massa magra Quindi sappiamo che le variazioni del testosterone fisiologico influiscono sulla quantità di muscoli che hai. Ma quanto? Se passi da 300 ng/dL a 600 ng/dL, quanta massa magra in più puoi aspettarti di avere?
Per rispondere a questa domanda, torniamo agli studi dose-risposta di Shalendar Bhasin di cui abbiamo discusso in precedenza. Possiamo prendere i dati dai tre studi ed eseguire una regressione su di essi per vedere come cambia la massa magra al variare dei livelli di testosterone all’interno dell’intervallo fisiologico.
Ecco la linea di regressione per i dati di Bhasin et al. 2001, 2005 e 2012. Ho usato solo punti dati in cui il testosterone si trovava all’interno di un intervallo fisiologico o appena al di fuori di esso (da 176 ng/dL a 1345 ng/dL). Sono 11 punti dati. Per ogni aumento di 100 ng/dL di testosterone, la massa magra aumenta di 0,6 kg. L’R al quadrato per la vestibilità del modello era 0,85, il che è molto buono.
Variazione della massa magra in relazione al cambiamento del Testosterone rispetto al basale, entro un intervallo di variazione da -340 ng/dL a +691 ng/dL. Pendenza della linea = 0,006, il che significa un ulteriore 0,6 kg in FFM per ogni aumento di 100ng/dL del Testosterone. R-Quadrato = 0,85. Dati di Bhasin et al. 2001, 2005 e 2012.
Il valore di 0,6 kg di FFM per ogni aumento di 100 ng/dL è in accordo con uno studio dose-risposta di Huang e colleghi su donne isterectomizzate. Hanno anche scoperto che la FFM aumenta di 0,6 kg per ogni aumento di 100 ng/dL di testosterone.
Possiamo anche eseguire una regressione sui dati che ho discusso in precedenza da Finkelstein e colleghi, in cui i livelli di testosterone sono stati soppressi e quindi ai soggetti sono state somministrate diverse dosi di un gel di testosterone. Sebbene i numeri effettivi della massa magra non siano stati riportati da questo studio, possiamo stimarli dai valori di base riportati e dalle variazioni percentuali. Per ogni aumento di 100 ng/dL di testosterone, la massa magra aumenta di 0,3 kg. L’R al quadrato per la vestibilità del modello è 0,70, il che è buono.
Cambiamento nella massa magra in relazione al cambiamento nel Testosterone dal basale. Pendenza della linea = 0,003, il che significa un ulteriore 0,3 kg in FFM per ogni aumento di 100ng/dL del Testosterone. R-Quadrato = 0,70. Dati da Finkelstein et al. 2013.
Mentre potremmo anche provare a eseguire una regressione su alcuni degli studi discussi in cui i livelli di testosterone erano bassi e portati in un intervallo normale fisiologico, il problema con questi è che tutti usavano diversi metodi di somministrazione per il testosterone (iniezione vs gel vs. . patch), che possono avere dinamiche diverse in termini di come vengono modificati i livelli ematici. Inoltre, il problema con il tentativo di aggregare diversi studi di iniezione è che variano nel tempo in cui misurano il testosterone e variano anche in termini di frequenza delle iniezioni. Puoi avere una misurazione del testosterone molto diversa se misuri 1 settimana dopo un’iniezione, rispetto a 2 settimane.
Tuttavia, possiamo dare un’occhiata a studi in cui è stato somministrato un farmaco che ha stimolato la produzione naturale di testosterone; tali studi possono imitare meglio il modo in cui la massa magra risponde alle variazioni dei livelli di testosterone endogeno. Dias et al. dato agli uomini Anastrozolo, un inibitore dell’Aromatasi. Inibendo l’Aromatasi, l’enzima che converte il Testosterone in Estrogeno, puoi aumentare efficacemente i livelli di Testosterone. In teoria, questi livelli di Testosterone sarebbero relativamente stabili, poiché rappresenterebbero livelli di testosterone endogeno piuttosto che testosterone esogeno da iniezione o somministrazione transdermica. L’Anastrozolo ha aumentato il testosterone da 272 ng/dL a circa 500 ng/dL a 6 mesi e la massa magra è aumentata di 1,5 kg. Sono circa 0,6 kg di FFM per ogni aumento di 100 ng/dL, il che è in accordo con le nostre analisi precedenti. Ribeiro e Abucham hanno somministrato agli uomini ipogonadici Clomifene Citrato, che aumenta il Testosterone endogeno (i livelli all’interno del corpo, rispetto a quello esogeno come da un’iniezione o da una crema) legandosi ai recettori degli ipotalamici degli Estrogeni. Legandosi ai recettori degli estrogeni, induce il cervello a pensare che non ci siano così tanti estrogeni nel corpo. Questo porta il cervello a pompare più ormone Luteinizzante (LH), che poi stimola i testicoli a produrre più testosterone. I livelli di testosterone nei soggetti responsivi sono aumentati da 201 ng/dL a 435 ng/dL dopo 3 mesi e la massa magra è aumentata di 1 kg. Sono 0,4 kg per ogni aumento di 100 ng/dL di testosterone. Liu et al. iniettato HCG in uomini che avevano bassi livelli di testosterone al limite; L’HCG è un ormone che stimola i testicoli a produrre più testosterone. Il testosterone è aumentato da 320 ng/dL a circa 720 ng/dL, un aumento di 400 ng/dL. Massa magra aumentata di 2 kg. Sono 0,5 kg di massa magra per ogni 100 ng/dL di aumento del testosterone, che è ancora una volta in accordo con il range che abbiamo stabilito. Infine, Bayram et al. uomini ipogonadici iniettati con HCG. Il testosterone è aumentato da 39 ng/dL a 512 ng/dL, un aumento di 473 ng/dL. Massa magra aumentata di 2,8 kg. Sono 0,6 kg per ogni aumento di 100 ng/dL. Pertanto, questi 4 studi suggeriscono un aumento di 0,5 – 0,6 kg di massa magra per ogni aumento di 100 ng/dL di testosterone, che è in accordo con le regressioni di Bhasin e Huang.
Mettendo insieme tutto questo, i dati suggeriscono che la massa magra aumenterà di 0,7 – 1,3 libbre (0,3 – 0,6 kg) per ogni aumento di 100 ng/dL dei livelli ematici di testosterone all’interno dell’intervallo fisiologico. Quindi, se passassi da 300 ng/dL a 600 ng/dL, questo sarebbe 0,9 – 1,8 kg o circa 2,1 – 4 libbre.
Ora, tieni presente che ci sono dei limiti a questa analisi. In primo luogo, si basa su medie; i risultati individuali possono essere diversi. Ad esempio, come accennato in precedenza, Finkelstein et al. hanno mostrato un’ampia variazione nel modo in cui i soggetti hanno risposto a diversi livelli di testosterone. In secondo luogo, si basa su analisi tra soggetti; ciò che accade all’interno delle persone può essere diverso da ciò che si osserva tra le persone. Terzo, parte di esso si basa su iniezioni di testosterone esogeno. Il problema è che, con le iniezioni, i livelli medi di testosterone nel sangue saranno superiori a quelli misurati. Questo perché, quando inietti il testosterone, ottieni un grande picco nei livelli ematici e poi decade lentamente nell’arco di 1-2 settimane. I ricercatori di solito misurano il testosterone alla depressione del decadimento dopo l’iniezione, di solito 1-2 settimane dopo. Questa limitazione diventa evidente quando si osservano alcuni dei dati dose-risposta di Bhasin. Ad esempio, nello studio Bhasin 2001, gli uomini che hanno ricevuto 125 mg di testosterone iniettato hanno guadagnato 3,4 kg di massa magra, ma il livello di testosterone nel sangue misurato di 542 ng/dL era simile al livello di base naturale dei soggetti prima che avessero il loro testosterone livelli soppressi. Pertanto, i loro livelli medi di testosterone erano probabilmente molto più alti di 542 ng/dL.
Sebbene questo sia certamente un grosso limite nell’analisi, va anche ricordato che, con la regressione, stiamo valutando principalmente le differenze nella massa magra tra diversi livelli di testosterone, piuttosto che la relazione con un particolare livello assoluto. In altre parole, stiamo osservando come la massa magra cambia per un cambiamento di 100 ng/dl nel testosterone, piuttosto che come la massa magra si riferisce, ad esempio, a un livello ematico di 500 ng/dl. Quindi, anche se i livelli di testosterone sono dovuti a iniezioni esogene, e anche se i livelli medi sono molto più alti di quelli misurati alla depressione, il rapporto tra i livelli non dovrebbe cambiare drasticamente. Va anche notato che il rapporto di 0,6 kg/100 ng/dL riscontrato negli studi Bhasin ha retto nelle donne a cui sono state somministrate dosi molto più basse. Tuttavia, è ancora una limitazione che deve essere considerata.
Una cosa interessante da notare è che le variazioni all’interno di un intervallo fisiologico possono avere un impatto maggiore rispetto alle variazioni al di fuori dell’intervallo fisiologico. In altre parole, la massa magra non aumenta in modo lineare con l’aumento dei livelli di testosterone. Quando superi l’intervallo fisiologico, la pendenza della relazione diminuisce e il testosterone non ha lo stesso impatto. Ciò diventa evidente quando guardiamo di nuovo alla nostra regressione dei tre studi Bhasin, ma questa volta includiamo i dati che sono ben al di sopra dell’intervallo normale fisiologico.
Puoi vedere che la curva si adatta meglio ai dati rispetto alla linea retta. La pendenza per la linea retta è 0,002, il che significa che la massa magra è aumentata di 0,2 kg per ogni 100 ng/dL di testosterone, che è inferiore agli 0,6 kg che abbiamo osservato con gli stessi dati in precedenza. Anche l’adattamento del modello non è altrettanto buono (R-quadrato = 0,77). Questo perché i livelli estremamente elevati di testosterone attenuano la relazione. Ogni aumento di 100 ng/dL di testosterone non ha un effetto così forte a intervalli soprafisiologici rispetto al normale intervallo fisiologico. Questo è supportato anche quando diamo un’occhiata allo studio Bhasin del 1996 sul testosterone ad alte dosi che ho menzionato prima. Il testosterone nel sangue è aumentato di circa 2326 ng/dL nel gruppo con solo testosterone e la massa magra è aumentata di 3,2 kg. Questo è un aumento di 0,13 kg per ogni aumento di 100 ng/dL di testosterone, che non è lontano dall’aumento di 0,2 kg menzionato in precedenza quando abbiamo incluso alte dosi di testosterone.
E le donne?
Le donne hanno livelli di Testosterone molto più bassi rispetto agli uomini e ci sono dati limitati per stabilire intervalli di riferimento sulle donne. Uno dei problemi è che alcuni test di laboratorio tradizionali per il Testosterone, come i radioimmunodosaggi (RIA), non sono abbastanza sensibili da misurare con precisione il Testosterone nelle donne. Misurazioni accurate del testosterone nelle donne richiedono tecniche sensibili come la cromatografia liquida-spettrometria di massa tandem (LC-MS/MS). Esistono alcuni dati che stabiliscono intervalli di riferimento utilizzando questa tecnica. Ecco i dati di Haring e colleghi che mostrano i percentili più bassi e più alti per le donne in premenopausa di età compresa tra 20 e 49 anni (si noti che sto mescolando alcuni dei dati di distribuzione effettivi per ottenere il 25° e il 75° percentile, con i loro modelli di regressione quantile per ottenere il 2,5° e 97,5° percentile, ma per i nostri scopi va benissimo).
Simile agli uomini, il testosterone diminuisce con l’età.
Testosterone misurato da GC-MS / MS in 985 donne. Dati da Haring et al., J Clin Endocrinol Metab, 2012. 1 nmol/L = 0,0347 ng/dL
È stato anche riscontrato che le donne che assumevano contraccettivi orali o terapia ormonale sostitutiva avevano in media livelli di testosterone più bassi, sebbene l’intervallo percentile superiore fosse più alto. Il 25° percentile per queste donne era 10 ng/dL (vs 13) e il 75° percentile era 56 ng/dL (vs 47).
Testosterone per età nelle donne, confrontando le donne trattate con contraccettivi orali o HRT a quelle che non lo sono. Dati da Haring et al., J Clin Endocrinol Metab, 2012. 1 nmol/L = 0,0347 ng/dL
Rari et al. non ha riscontrato alcuna relazione tra testosterone totale e massa magra nelle donne anziane (età 67-94 anni), ma ha osservato una relazione significativa tra testosterone libero e massa magra (il testosterone libero è la forma che non è legata ad alcuna proteina, da qui il termine “libero”).
Possiamo anche esaminare i dati in cui alle donne con bassi livelli di testosterone è stato somministrato testosterone esogeno. Anche in questo caso, i dati sono limitati, ma ci sono alcuni studi.
Gruber e colleghi hanno somministrato alle donne in postmenopausa (età media di 51 anni) testosterone topico o un placebo per 6 mesi. Ora, va notato che in questo studio, le donne non avevano bassi livelli di testosterone (i livelli medi erano 29 ng/dL, che rientra in un intervallo normale). Il testosterone è aumentato appena al di sopra dell’intervallo normale (72 ng/dL) e non vi è stato alcun impatto sulla massa magra.
Cambiamento nella massa magra con diverse dosi di Testosterone nelle donne in menopausa isterectomizzate. Dati da Huang et al, Menopausa, 2014. Solo la dose di 25mg ha determinato un aumento statisticamente significativo. La massa magra è aumentata di 0,6kg per ogni aumento di 100ng/dL del Testosterone, che è al di fuori del normale intervallo fisiologico per le donne (13 – 56 ng/dL).
Questi dati suggeriscono che è necessario aumentare i livelli ematici di testosterone ben al di fuori di un intervallo normale fisiologico (verso la fascia molto bassa di un maschio) nelle donne in post-menopausa per ottenere aumenti misurabili della massa magra e della funzione sessuale. Nel complesso, questi dati indicano che le variazioni del testosterone nelle donne, all’interno del normale range fisiologico, hanno un impatto misurabile molto piccolo, se non nullo, sul muscolo. Ciò non sorprende se consideriamo l’aumento di 0,3 – 0,6 kg per ogni aumento di 100 ng/dL di testosterone di cui abbiamo discusso. L’intervallo normale per le femmine va da 13 a 56 ng/dL, un intervallo di soli 43 ng/dL. Ciò equivale solo a un quarto di chilogrammo (mezza libbra) o meno nelle donne.
Tiriamo le somme
Quando esaminiamo l’intero corpo di prove, è chiaro che le variazioni nei livelli fisiologici di Testosterone influiscono sulla quantità di muscoli che il soggetto ha, ma avranno un impatto minimo sui guadagni relativi (%). Riassumendo: I dati trasversali, gli studi dose-risposta e gli studi TRT supportano tutti variazioni all’interno dell’intervallo normale in quanto hanno un impatto sulla massa magra e sui muscoli-scheletrici. Le variazioni nei livelli ematici di Testosterone influiscono sul livello “base” dei muscoli, ma hanno un impatto minimo sui guadagni relativi (%). Quindi, avere livelli di Testosterone più alti significa avere un livello base più alto di massa muscolare. Mentre i guadagni relativi saranno per lo più simili, i guadagni assoluti saranno più alti a causa della linea di base più elevata. Gli impatti del Testosterone e dell’allenamento contro-resistenza sulla massa magra e sui muscoli-scheletrici sono principalmente additivi piuttosto che sinergici. La massa magra al basale aumenta di circa 0,7 – 1,3 libbre o 0,3 – 0,6 kg per ogni aumento di 100 ng/dL del Testosterone fisiologico; questo si basa su medie e dati tra soggetti, quindi i risultati individuali possono variare considerevolmente. Ci sono anche limitazioni a questa analisi, come il fatto che parte di essa si basa su dati dose-risposta provenienti da iniezioni, che potrebbero non riflettere accuratamente i cambiamenti nei livelli endogeni. L’impatto del Testosterone sulla FFM è attenuato a livelli sovrafisiologici (>1500 ng/dL); La FFM di base aumenta di circa 0,1-0,2 kg per ogni 100 ng/dL per quei livelli. Le variazioni del Testosterone nelle donne, all’interno del normale range fisiologico, hanno un impatto minimo o nullo sul muscolo-scheletrico. Quindi, sì, i soggetti di sesso maschile con un livello di Testosterone più alto hanno un vantaggio in termini assoluti sulla massa muscolare. Questi dati indicano anche che gli uomini che invecchiano, con livelli di Testosterone al limite o bassi, possono ottenere un beneficio nella costruzione muscolare da terapie progettate per aumentare i livelli di Testosterone in range fisiologici, sia da fonti esogene (come iniezioni o gel) sia da fonti che stimolano la produzione di Testosterone (come il Clomifene). , HCG o inibitori dell’Aromatasi), anche se si allenano già con i pesi.
Però, attenzione a fare comparazioni fuori luogo: la fisiologia e quello che può manifestare non è paragonabile alle sue alterazioni fuori range… nel bene e nel male…
Qualche giorno fa, scartabellando come di mio solito la letteratura scientifica, mi è capitato tra le mani uno studio che definirei tutto sommato interessante. Lo studio in questione prendeva in esame la possibilità di riscontrare esito negativo al classico Test Antidoping Testosterone:Epitestosterone ratio, in seguito al consumo di Té verde o bianco per via delle catechine in essi presenti.[1]
Ho quindi sfruttato l’occasione per sottolineare quando e come il più elementare dei test antidoping possa essere superato dall’atleta…
Tra chatechine e differenze genetiche di popolazione
I ricercatori della Kingston University di Londra hanno scoperto che bere tè verde o tè bianco può aiutare gli atleti che usano AAS a superare un test antidoping comunemente usato. Declan Naughton e colleghi hanno riferito che i composti noti come catechine possono consentire agli atleti di utilizzare dosi sovrafisioliche di Testosterone ed evitarne il rilevamento.
Il test del rapporto Testosterone:Epitestosterone (T:E ratio) viene spesso utilizzato per lo screening degli atleti sottoposti a test Anti-Doping per rilevare la somministrazione esogena di dosi sovrafisiologiche di Testosterone. Determina il rapporto tra Testosterone Glucuronide e Epitestosterone Glucoronide nelle urine di un atleta.
La somministrazione esogena di Testosterone non influenza i livelli di Epitestosterone. Pertanto, un atleta che utilizza Testosterone dovrebbe avere un rapporto più elevato di metaboliti del Testosterone nelle urine rispetto all’Epitestosterone. Le catechine presenti nel tè verde inibiscono un enzima chiamato UGT2B17. Questo enzima è responsabile del legame molecolare tra l’acido Glucuronico ad il Testosterone. Inibendo l’UGT2B17, un atleta presenterà un livello minore di Testosterone Glucuronide nelle urine. Quindi, questo atleta che usa Testosterone esogeno avrà una Testosterone:Epitestosterone ratio normale.
In pratica, l’UDP-glucuronosiltransferasi (UGT2B17) è l’enzima chiave coinvolto nella glucuronidazione del Testosterone a Testosterone Glucuronide, che funge anche da marker per la Testosterone/Epitestosterone (T/E) ratio per rilevare l’abuso di Testosterone nello sport. Gli inibitori della glucuronidazione del Testosterone potrebbero avere un impatto sui livelli di Testosterone circolante, favorendo così le prestazioni, oltre a influenzare potenzialmente il rapporto T/E urinario e quindi mascherare l’abuso di Testosterone. Rapporti precedenti hanno rivelato che i farmaci antinfiammatori non steroidei, il Diclofenac e l’Ibuprofene, inibiscono l’enzima UGT2B17. In questo studio l’analisi della glucuronidazione del Testosterone è stata condotta eseguendo i saggi UGT2B17 con il rilevamento del Testosterone non glucuronidato mediante cromatografia liquida ad alte prestazioni. I risultati dello studio hanno mostrato che la glucuronidazione del Testosterone è stata inibita dagli estratti di tè verde e bianco, insieme a specifici composti di catechina, in particolare: epicatechina, epigallocatechina gallato (EGCG) e catechina gallato. Il valore di inibizione IC50 per EGCG è stato determinato, utilizzando un diagramma Dixon, pari a 64μM, pari all’inibitore dei FANS più attivo, il Diclofenac. Pertanto, gli alimenti comuni e i loro costituenti, per la prima volta, sono stati identificati come inibitori di un enzima chiave coinvolto nella glucuronidazione del Testosterone. Sebbene questi composti comuni non siano substrati dell’enzima UGT2B17, è stato dimostrato che inibiscono la glucuronidazione del Testosterone che può avere implicazioni sull’attuale controllo del doping nello sport.
Struttura chimica del EGCG
La maggior parte degli individui ha un rapporto Testosterone:Epitestosterone di 1: 1. Ma rapporti fino a 4:1 non sono rari. L’Agenzia mondiale antidoping (WADA) considera un rapporto di 4:1 come un indicatore putativo di doping soggetto a conferma da un’altra procedura antidoping nota come test del rapporto degli isotopi di carbonio (CIR).
Il test della T:E ratio non può, ovviamente, indicare la differenza tra il Testosterone endogeno ed il Testosterone esogeno somministrato per via di iniezione, compressa o soluzione topica. I test CIR possono rilevare il Testosterone esogeno di origine vegetale. Tuttavia, se gli atleti riescono a superare con successo il test della T:E ratio, non sono soggetti al più preciso test CIR. Di conseguenza, se gli atleti possono evitare il rilevamento nel test della T:E ratio, possono evitare di essere scoperti per il loro uso di Testosterone esogeno.
Il tè verde può fornire un altro metodo per gli atleti al fine di superare indenni il test della T:E ratio. Il professor Charles Yesalis, un noto esperto di AAS ed epidemiologo presso la Pennsylvania State University, è stato a lungo schietto sulla cosiddetta “scappatoia del Testosterone”. Anche Don Catlin, l’ex direttore dell’Olympic Analytical Testing Lab dell’UCLA e fondatore dell’Anti-Doping Research Institute, ha riconosciuto che gli atleti possono usare il Testosterone senza essere scoperti anche se i tester antidoping sanno come farlo. “Potrei capire come assumere una buona quantità di Testosterone senza farmi scoprire, e se posso dirlo, anche molti altri possono farlo”, ha ammesso Catlin.
Gli atleti aggiungono semplicemente Epitestosterone al loro protocollo farmacologico per mantenere il rapporto 4:1. Tuttavia, devono mantenere i livelli assoluti di Testosterone Gluconoride ed Epitestosterone Gluconoride urinario al di sotto del limite consentito dalla WADA pari a 200ng/mL.
“La crema” utilizzata dalla BALCO più di un decennio fa era semplicemente una variazione del cocktail di Testosterone ed Epitestosterone che era stato storicamente utilizzato dagli atleti per decenni al fine di ingannare i test antidoping.
Gli “atleti BALCO” hanno utilizzato un esclusivo sistema di somministrazione transdermica per assumere una formula personalizzata di Testosterone ed Epitestosterone.
Un grammo di “Crema” conteneva 5mg di Epitestosterone per ogni 100mg di Testosterone in un rapporto di 1:20 secondo Victor Conte. Ciò ha permesso agli “atleti BALCO” di usare il Testosterone senza essere scoperti. Anche dopo l’indagine governativa da oltre 60 milioni di dollari sulla BALCO, il rapporto Mitchell da 20 milioni di dollari sull’uso di steroidi da parte dei giocatori di MLB e le sensazionali udienze del Congresso sugli steroidi nel baseball e in altri sport professionistici, la “scappatoia del Testosterone” non è stata chiusa.
La “crema” è efficace come sempre per un giocatore di baseball professionista.
Come se il test della T:E ratio non fosse abbastanza efficace come strumento antidoping, quattordici anni fa i ricercatori del Karolinska University Hospital di Stoccolma hanno scoperto che alcuni atleti si possono somministrare quantità impressionanti di Testosterone e non risultare positivi al test antidoping della T:E ratio per via della mancanza del gene per la sintesi dell’enzima UGT2B17.
Circa il 40% di questi atleti “dotati geneticamente” potrebbe iniettarsi 500mg di Testosterone Enantato (360mg di Testosterone effettivi) senza risultare positivi al test della T:E ratio della WADA.
E questa anomalia genetica è relativamente comune ed è più comune in alcuni gruppi etnici. Pertanto, l’etnia di un atleta può dargli un vantaggio sul test antidoping “base”:
78,0% – Mulatto (brasiliano)
66,7% – Asia orientale (coreano)
57,3% – Cape Coloured (Città del Capo, Sudafrica)
37,6% – Meticcio messicano
30,4% – Asia del Pacifico (Asia sudorientale/cinese meridionale, indiana asiatica, giapponese)
Adesso sappiamo che il tè verde o il tè bianco, per azione delle catechine, o alcuni farmaci FANS, possono bloccare efficacemente l’enzima UGT2B17.
L’uso del Testosterone rimane uno dei metodi più popolari utilizzati dagli atleti che usano steroidi anabolizzanti per evitare il rilevamento. La “scappatoia del Testosterone” continuerà ad essere sfruttata finché il test della T:E ratio verrà utilizzato come schermo principale per la rilevazione d’uso del Testosterone esogeno.
Gabriel Bellizzi
Riferimenti:
1- Jenkinson, C. et al. (2012). Dietary green and white teas suppress UDP-glucuronosyltransferase UGT2B17 mediated testosterone glucuronidation. Steroids. http://dx.doi.org/10.1016/j.steroids.2012.02.023.
Verso la fine del primo decennio del presente secolo, una “presunta” nuova classe di farmaci con attività anabolizzante ha iniziato a diffondersi in diverse discipline sportive , dal ciclismo a, ovviamente, il Bodybuilding. Sto parlando ovviamente dei SARMs, acronimo di Selective Androgen Receptor Modulators (in italiano, Modulatori Selettivi del Recettore degli Androgeni, SARM).
Essendo molecole sperimentali e non ancora commercializzate come farmaci da prescrizione per uso umano, i SARM si sono diffusi rapidamente in tutto il mondo grazie anche alla vendita da parte degli store online UK e USA (dove la vendita di supplementi contenenti tali molecole è legale).
Non ci volle molto tempo prima che un “alone leggendario” avvolgesse i SARM ed i loro presunti o reali effetti. I SARM vennero in breve pubblicizzati come il “doping ideale” con tutti gli effetti positivi degli steroidi anabolizzanti, pur non avendo alcun svantaggio o effetto collaterale legato a questi ultimi.
In generale, gli effetti positivi principali degli AAS sono considerati essere l’effetto anabolizzante sulla massa muscolare e l’effetto stimolante sul miglioramento della densità minerale ossea. Tutti gli altri effetti cosi detti androgeni sono generalmente considerati indesiderati. Anche se, ovviamente, ciò dipende in gran parte dal grado con il quale essi si verificano (ma anche dal sesso e dalla disciplina praticata dall’utilizzatore).
Ad esempio, gli AAS inducono l’Eritropoiesi, il processo di biosintesi degli Eritrociti (globuli rossi). Questo porta ad un aumento dell’Ematocrito che, quando diventa troppo alto, ossia oltre la soglia del 53-54%, vede arrestati i suoi effetti benefici sulla resistenza vedendo aumentato sensibilmente il rischio di trombosi venosa. Tuttavia, se si eliminasse completamente qualsiasi effetto stimolante sull’eritropoiesi, l’ematocrito potrebbe diventare troppo basso, in specie se viene a mancare un fattore compensativo alla riduzione indotta. Di conseguenza, si finirebbe per essere anemici. Quindi anche alcuni di quegli effetti indesiderati degli AAS sono “voluti” in una certa misura. Ma i paradossi della selettività non terminano con questo, ovviamente. Per semplicità, tuttavia, tratterò il discorso più avanti nel presente articolo.
Il punto della questione è: i SARM danno veramente un vantaggio in quanto a rapporto tra effetti positivi e collaterali rispetto agli AAS? La risposta richiede una spiegazione dettagliata della storia, delle caratteristiche e degli effetti, constatati sia in ambito clinico che “off-label”, legati ai SARM.
Nozioni iniziali sui SARM.
Come la maggior parte di voi saprà, SARM sono una classe di ligandi selettivi del recettore degli androgeni (AR).[1]
Nonostante un certo numero di persone sia convinta che i SARM siano stati sintetizzati circa venti anni fa, e che non abbiano nulla a che vedere nel loro sviluppo con gli AAS, la realtà è che il termine si riferisce ad un macrogruppo di molecole affini al AR con un valore terapeutico (vedi potenziale androgeno e anabolizzante) superiore a 1, cioè al Testosterone. Per questa ragione esistono due gruppi di SARM: i SARM steroidei ed i SARM non-steroidei. Di conseguenza, tutti i derivati del Testosterone, del DHT, compresi i 19-Norsteroidi, che sono stati modificati strutturalmente al fine di accentuarne le caratteristiche anabolizzanti e ridurne quelle androgene sono considerabili quali SARM steroidei.
Due esempi tipici di SARM steroideo e non-steroideo
Gli sforzi iniziali per sviluppare SARM steroidei, basati su modifiche della molecola di Testosterone, risalgono agli anni ’40. L’era moderna dei SARM non steroidei è stata scatenata da un lavoro indipendente presso la Ligand Pharmaceuticals (2, 3) e l’Università del Tennessee.(4, 5) Gli scienziati della Ligand Pharmaceuticals sono stati i primi a sviluppare una serie di Chinolinoni ciclici con attività anabolica sul muscolo scheletrico e un certo grado di selettività tissutale.(2, 6, 7, 8) La scoperta di Dalton e Miller che le Aril Propionammidi con somiglianze strutturali con il Bicalutamide e l’Idrossiflutammide potrebbero innescare l’attività trascrizionale AR-dipendente ha fornito la prima guida per lo sviluppo della classe di SARM diaril propionammidi.(4, 5) Il decennio successivo a questi primi sforzi ha visto l’emergere di un gran numero di SARM non steroidei praticamente da tutte le principali aziende farmaceutiche.(9)
Fondamenti logici nella ricerca dei SARM non-steroidei
Il Testosterone, il principale ligando per il Recettore degli Androgeni, svolge una varietà di funzioni fisiologiche nell’uomo (10): è essenziale, anche per via della sua conversione in DHT, al fine di mantenere una corretta funzione sessuale, lo sviluppo delle cellule germinali e gli organi sessuali accessori. Il Testosterone interagisce ovviamente anche con il muscolo scheletrico, grasso, ossa, emopoiesi, coagulazione, metabolismo dei lipidi, proteine e carboidrati e comportamenti psicosessuali e cognitivi. Sebbene la carenza di androgeni negli uomini adulti sia il disturbo più diffuso della alterazione nella segnalazione AR (11), il principale impulso per lo sviluppo dei SARM è legato allo sfruttamento dei potenziali effetti anabolici di questi composti sul muscolo scheletrico e sull’osso.
Come ben sappiamo, man mano che uomini e donne invecchiano, perdono massa muscolare scheletrica, forza, potenza (12, 13), principalmente a causa della perdita preferenziale delle fibre muscolari di tipo 2 (14), e la densità ossea. La perdita di massa muscolare e forza associata all’età aumenta il rischio di cadute, fratture, limitazione della mobilità, disabilità fisica e scarsa qualità della vita (15, 16). Il declino funzionale e la dipendenza negli anziani gravano pesantemente sui servizi e sui costi sanitari. Nonostante l’elevata prevalenza di limitazioni funzionali e disabilità tra gli individui più anziani, i geriatri praticanti hanno poche scelte terapeutiche per il trattamento degli individui più anziani con limitazioni funzionali e disabilità fisica. Allo stesso modo, il decorso di molte malattie croniche, come la malattia polmonare ostruttiva cronica, la malattia renale allo stadio terminale, l’insufficienza cardiaca congestizia e alcuni tipi di cancro, è punteggiato da perdita di massa muscolare e limitazioni funzionali fisiche, che contribuiscono indipendentemente a sintomi, limitazione della mobilità e disabilità. Pertanto, c’è un enorme bisogno insoddisfatto di funzioni che promuovano terapie anabolizzanti che possano migliorare la funzione fisica e ridurre il peso della disabilità.
Tra le varie terapie anabolizzanti candidate ad applicazione in fase di sviluppo, quella con SARM non steroidei è la più recente in corso di sviluppo. La somministrazione di Testosterone aumenta la massa muscolare scheletrica e la massima forza volontaria in uomini sani, con carenza di androgeni (17-18) ed eugonadici (19, 20) e anziani (21), e negli uomini con molti disturbi cronici (22, 23). Gli effetti anabolizzanti del Testosterone sulla massa e sulla forza dei muscoli scheletrici sono correlati alla dose di Testosterone e alle sue concentrazioni ematiche (20, 21, 24, 25). Pertanto, il potenziale per ottenere il rimodellamento del muscolo scheletrico e l’aumento della massa e della forza del muscolo scheletrico con la somministrazione di androgeni è notevole. Tuttavia, la somministrazione di dosi sovrafisiologiche di androgeni è associata ad un’elevata frequenza di effetti avversi dose-dipendenti, come eritrocitosi, edema delle gambe ed eventi prostatici (21, 26). Pertanto, agenti terapeutici come i SARM non steroidei con la cui somministrazione possono far ottenere effetti anabolizzanti sul muscolo scheletrico e sull’osso senza gli effetti avversi limitanti riscontrati con dosaggi di Testosterone aventi il medesimo effetto terapeutico sarebbero attraenti come terapie anabolizzanti d’elezione (27, 28, 29). Il riconoscimento di queste potenziali opportunità per lo sviluppo di nuove terapie per le limitazioni funzionali e disabilità associate a disturbi cronici, invecchiamento e osteoporosi ha guidato gli sforzi farmaceutici per sviluppare SARM non steroidei.
Il raggiungimento della selettività dei tessuti
Storicamente sono stati utilizzati due approcci generali per ottenere la selettività tissutale dell’azione degli Androgeni. Il primo approccio consiste nello sviluppare un SARM con un profilo di attività desiderato e la selettività tissutale. Il secondo approccio è quello di chiarire i meccanismi di azione degli androgeni sul muscolo scheletrico e sulla Prostata e di identificare le molecole di segnalazione che sono a valle del recettore degli androgeni e che attivano le vie coinvolte nell’ipertrofia del muscolo scheletrico, ma non della Prostata.
SARM steroidei: relazioni struttura-attività
Come accennato in precedenza, strutturalmente, i SARM possono essere classificati in SARM steroidei e non steroidei. I SARM steroidei si formano modificando la struttura chimica della molecola di Testosterone (vedi figura seguente).
È stato riconosciuto negli anni ’40 che la sostituzione di un metile in posizione C-17 ritarda il metabolismo presistemico del Testosterone, estendendone l’emivita e rendendolo attivo per via orale. Pertanto, un certo numero di androgeni orali, come il Methylterstosterone, hanno una metilazione in C-17. Tuttavia, gli androgeni 17-alfa alchilati somministrati per via orale, sono potenzialmente epatotossici e abbassano notevolmente il colesterolo HDL plasmatico.
La rimozione del gruppo 19-metile aumenta l’attività anabolizzante del Testosterone (Figura sopra). Pertanto, il 19-nortestosterone ha costituito la base della serie di molecole derivate del Nandrolone. Il Nandrolone è ridotto dalla 5-α reduttasi nei tessuti bersaglio a un androgeno meno potente, il Diidronandrolone (DHN), ma è meno suscettibile all’aromatizzazione in estrogeni convertendo primariamente nel poco attivo Estrone.
Le sostituzioni alchiliche 7-alfa rendono il Testosterone meno suscettibile alla 5-α riduzione e ne aumentano la selettività tissutale rispetto alla Prostata. Pertanto, il 7-alfa metil, 19-nortestosterone ha attività anabolica teoricamente superiore all’attività androgena, sebbene i test fatti sono stati svolti su topi attraverso il ben poco affidabile se rapportato all’uomo “test di Hershberger” (per approfondimenti clicca qui). Comunque, altre molecole di questa serie con gruppi alchilici variabili sono state studiate per la loro attività anabolica.
Il Testosterone viene eliminato rapidamente dalla circolazione e ha una breve emivita. L’esterificazione del gruppo ossidrile 17-β rende la molecola più idrofoba; più lunga è la catena laterale dell’estere, maggiore è l’idrofobicità. Quando gli esteri idrossilici 17-β del Testosterone vengono somministrati attraverso un iniezione intramuscolare in una sospensione oleosa, vengono rilasciati lentamente dal deposito oleoso nella circolazione. Il lento rilascio di esteri idrossilici 17-β dal deposito oleoso estende la loro durata d’azione. Tuttavia, la de-esterificazione degli esteri di Testosterone non limita la velocità della metabolizzazione molecolare; in breve, l’emivita del Testosterone Enantato nel plasma non è significativamente diversa da quella del Testosterone non esterificato una volta scissa l’esterificazione. Allo stesso modo, l’esterificazione del Nandrolone per formare il Nandrolone Decanoato aumenta la sua emivita.
Molecola di Testosterone legata ad un estere Enantato.
L’Oxandrolone è un AAS orale derivato dal DHT che ha un sostituente metilico 17-alfa. La sostituzione del secondo carbonio con l’ossigeno aumenta la stabilità del 3-cheto gruppo e ne aumenta l’attività anabolizzante. Non aromatizza in estrogeno e ha mostrato una bassa attività androgena. Indi, esso è un altro esempio di SARM steroideo.
Struttura molecolare del Oxandrolone
SARM non-steroidei
Gli sforzi pionieristici degli scienziati della Ligand Pharmaceuticals e dell’Università del Tennessee hanno fornito le prime basi della scoperta dei SARM non-steroidei. Da allora, sono state esplorate una serie di categorie strutturali di SARM farmacofori: aril-propionamide (GTX, Inc.), idantoina biciclica (BMS), chinolinoni (Ligand Pharmaceuticals), analoghi della tetraidrochinolina (Kaken Pharmaceuticals, Inc.), benizimidazolo, imidazolopirazolo. , indolo e derivati pirazolina (Johnson e Johnson), derivati azasteroidali (Merck) e derivati anilina, diaril anilina e bezoxazepinoni (GSK) (vedi figura seguente). Poiché è stata pubblicata solo una parte della ricerca sulla scoperta, è probabile che esistano categorie strutturali aggiuntive. Una recente review di Narayanan et al fornisce un eccellente trattato delle strutture dei SARM (28).
Le modifiche strutturali degli analoghi dell’aril propionammide bicalutamide e idrossiflutamide hanno portato alla scoperta della prima generazione di SARM. I composti S1 e S4 in questa serie si legano al AR con elevata affinità e dimostrano selettività tissutale nel impreciso test di Hershberger che utilizza un modello di ratto castrato (30, 31). In questo modello di ratto castrato, sia S1 che S4 hanno prevenuto l’atrofia indotta dalla castrazione del muscolo levat ani e hanno agito come deboli agonisti nella Prostata (30, 31, 32). Alla dose di 3 mg/kg/die, S4 ha parzialmente ripristinato il peso della prostata a < 20% di quello intatto, ma ha ripristinato completamente il peso del levator ani, la forza dei muscoli scheletrici, la densità minerale ossea, la forza ossea e la massa corporea magra e ha soppresso LH e FSH (33, 34). S4 ha anche prevenuto la perdita ossea indotta dall’ovariectomia nel modello di osteoporosi femminile di ratto (35). La capacità dei SARM di promuovere sia la forza muscolare che la forza meccanica ossea costituisce un vantaggio unico rispetto ad altre terapie per l’osteoporosi che aumentano solo la densità ossea.
S1 e S4 sono agonisti parziali; quindi, in ratti maschi intatti (31), S1 e S4 competono con gli androgeni endogeni (o esogeni) e agiscono come antagonisti nella Prostata, tali SARM con attività antagonista o bassa attività intrinseca nella Prostata potrebbero essere utili nel trattamento dell’IPB o del cancro alla Prostata. Gli effetti soppressivi di questa classe di SARM sulla secrezione di gonadotropine nei ratti suggeriscono una potenziale applicazione per la contraccezione maschile.(31)
SARM non-steroideo S4 (Andarina)
Il legame etereo e la sostituzione della posizione-para dell’anello B sono fondamentali per l’attività agonista dei SARM aril propionammidi (30). Sulla base delle strutture cristalline, i composti con legame etereo sembrano adattare una conformazione più compatta rispetto alla bicalutamide a causa della formazione di un legame H intramolecolare, consentendo all’anello B di evitare il conflitto sterico con la catena laterale di W741 nel AR e potenzialmente spiegando l’attività agonista.(36)
I derivati dell’idantoina, sviluppati dal gruppo BMS (37), hanno una struttura ad anello A simile a quella della bicalutamide. Il gruppo ciano o nitro di queste molecole interagisce con Q711 e R752 (38, 39). L’anello benzenico o gruppo naftile, insieme all’anello idantoico, si sovrappone al piano steroideo, mentre l’azoto dell’anello idantoinico forma un legame H con N705. BMS-564929 lega al AR con alta affinità e alta specificità. BMS-564929 ha dimostrato attività anabolizzante nel muscolo levator ani e un alto grado di selettività tissutale, come indicato da una ED50 sostanzialmente più elevata per la Prostata. I derivati dell’idantoina sono potenti soppressori dell’LH. BMS-564929 è disponibile per via orale nell’uomo, con un’emivita di 8-14 ore. L’emivita prolungata di questi ligandi nei ratti può spiegare la dose più bassa necessaria per ottenere effetti farmacologici; differenze nelle attività in vivo di SARM che condividono affinità di legame e attività in vitro simili possono essere correlate alle differenze nella farmacocinetica e nell’esposizione al farmaco.(40)
Hanada et al (41) della Kaken Pharmaceutical Co. hanno riportato una serie di derivati della tetraidrochinolina come agonisti dell’AR nell’osso. Sebbene questi composti mostrino un’elevata affinità per l’AR e una forte attività agonista nella Prostata e nel levator ani, hanno dimostrato una scarsa selettività tra i tessuti androgeni e anabolici (41). Una significativa attività farmacologica in vivo è stata osservata solo ad alte dosi sottocutanee.(28, 41)
I composti ligandi LGD2226 e LGD 2941 che sono derivati biciclici del 6-anilino chinolinone hanno mostrato attività anabolica sul muscolo levator ani, nonché sulla massa ossea e sulla forza, pur avendo scarso effetto sulla dimensione della Prostata in un modello preclinico di roditori (42, 43, 44). È stato anche dimostrato che LGD2226 mantiene il comportamento riproduttivo maschile nel modello di roditore castrato (42). Gli scienziati della Johnson e Johnson hanno sostituito il legante propionammidico con elementi ciclici come pirazoli, benzimidazoli, indoli e mimetici propionanilidi ciclici (45). Gli scienziati della Merck hanno sviluppato una serie di derivati 4-azasteroidali e butanammidi (28). Ulteriori composti sono stati sviluppati da altre aziende farmaceutiche, ma una discussione dettagliata di ciascun composto esula dallo scopo di questo articolo.
Meccanismi di selettività tissutale dei SARM
Narayanan et al hanno confrontato le vie attivate da un aril propionamide SARM, S-22, con quelle attivate dal DHT (46) e hanno scoperto che S-22 e DHT attivavano diverse vie di segnalazione distinte. S-22 e DHT differivano significativamente nel reclutamento del AR e dei suoi co-regolatori come potenziatore del PSA. L’S-22 differiva anche dal DHT nell’induzione della rapida fosforilazione di diverse chinasi (46). Tuttavia, i meccanismi che contribuiscono all’attivazione trascrizionale tessuto-specifica e alla selettività degli effetti biologici dei SARM rimangono poco compresi. Sono state proposte tre ipotesi generali, anche se queste ipotesi non si escludono a vicenda. L’ipotesi del co-attivatore presuppone che il repertorio di proteine co-regolatrici che si associa al AR legato al SARM differisce da quello associato al AR legato al Testosterone che porta all’attivazione trascrizionale di un insieme di geni regolati in modo differenziale.
Antigene Prostatico Specifico (Prostate Specific Antigen, PSA)
L’ipotesi conformazionale afferma che le differenze funzionali nelle classi di ligandi (agonisti, antagonisti e SARM) si riflettono in stati conformazionalmente distinti con partizionamento termodinamico distinto. Il legame con il ligando induce specifici cambiamenti conformazionali nel dominio di legame del ligando, che potrebbe modulare la topologia di superficie e le successive interazioni proteina-proteina tra AR e altri co-regolatori coinvolti nell’attivazione trascrizionale genomica o proteine citosoliche coinvolte nella segnalazione non genomica. Le differenze nella conformazione del recettore ligando-specifico e le interazioni proteina-proteina potrebbero portare a una regolazione genica tessuto-specifica, a causa di potenziali cambiamenti nelle interazioni con ARE, co-regolatori o fattori di trascrizione. Le interazioni proteina-proteina indotte dal ligando contribuiscono alle interazioni tra le estremità amminiche e carbossiliche del AR (cioè l’interazione N/C) e il reclutamento di co-attivatori (47). Entrambe le interazioni sono mediate dall’interazione tra la regione AF2 del AR ed i motivi di legame FXXLF o LXXLL (48). Il solco idrofobo presente nella regione AF2 del AR LBD sembra essere più favorevole per il legame della fenilalanina, il che suggerisce che l’interazione N/C è preferita. Sebbene la conformazione AR-LBD legata al SARM non steroideo non sia stata ben caratterizzata, Sathya et al (49) hanno riportato che alcuni SARM steroidei che hanno attività agonista in vitro inducono un cambiamento conformazionale attivante senza facilitare le interazioni N/C. Questi dati suggeriscono che il cambiamento conformazionale specifico del ligando è ottenibile con ligandi sintetici.
(A) Il gene AR consiste di 8 esoni che codificano per il recettore degli androgeni con un prodotto genico della dimensione tipica di 919 amminoacidi. Il AR è composto da un dominio N-terminale (NTD), un dominio di legame al DNA centrale (DBD), una regione a cerniera corta e un LBD C-terminale. (B) LBD comprende una struttura elicoidale 12 che racchiude una tasca centrale di legame dell’ormone (HBP), un secondo dominio della funzione di attivazione (AF2) che si trova all’estremità carbossi-terminale dell’LBD e un sito di legame scoperto di recente, funzione di legame 3 (BF3). La conformazione adottata dell’H12 è inequivocabilmente associata al meccanismo d’azione molecolare dei ligandi legati all’HBP. (C) Come mostrato nella struttura complessa di Diidrotestosterone (DHT) e AR-LBD, l’AR HBP è composto principalmente da residui idrofobici (palla verde) che possono formare forti interazioni non polari con il DHT. L’ancoraggio proteina-ligando può essere ulteriormente stabilizzato da una rete di legami idrogeno (linea tratteggiata blu) che coinvolge i residui polari R752, Q711, N705 e T877.[fonte immagine https://www.researchgate.net/%5D
Bohl et al (36) hanno riportato che la bicalutamide adotta una conformazione molto piegata nel AR. Sebbene l’anello A e il legame ammidico della molecola di bicalutamide si sovrappongano al piano steroideo, l’anello B della bilcautammide si piega lontano dal piano, puntando verso la parte superiore della tasca di legame del ligando (LBP), che costituisce una caratteristica strutturale unica di questo classe di leganti (36). Il gruppo ciano dell’anello A forma legami H con Q711 e R752, simile al 3-cheto gruppo nel 5α-DHT (36). Il gruppo idrossile chirale forma legami H con L704 e N705, imitando l’anello C e il gruppo 17β-OH nel 5α-DHT (36). Queste interazioni di legame H sono fondamentali per un’elevata affinità di legame. Lievi modifiche strutturali possono cambiare il ligando da antagonista AR ad agonista. Il legame idrogeno favorevole tra il ligando e la catena laterale T877, le caratteristiche strutturali che imitano il 3-cheto gruppo del Testosterone e le interazioni idrofobiche sono fondamentali affinché il ligando si leghi con alta affinità e stimoli l’azione del AR. La struttura cristallina a raggi X del AR legato a S-1 ha rivelato che la catena laterale W741 è spostata dall’anello B per espandere la tasca di legame in modo che il composto si orienti verso la regione AF2 (50). Il ripiegamento proteico del AR legato al SARM è lo stesso che si tratti di un SARM steroideo e non steroideo (50). Non è chiaro come l’interazione ligando-recettore determini l’attività agonista o antagonista del ligando.
La selettività tissutale dei SARM potrebbe anche essere correlata a differenze nella loro distribuzione tissutale, potenziali interazioni con la 5α-reduttasi o l’aromatasi CYP19, o l’espressione tessuto-specifica di co-regolatori (51). Tuttavia, studi di autoradiografia con derivati di bicalutamide e idantoina (52) hanno mostrato che non si accumulano preferenzialmente nei tessuti “anabolizzanti”. L’azione del Testosterone in alcuni tessuti androgeni è amplificata dalla sua conversione in 5α-DHT (53); i SARM non steroidei non fungono da substrati per la 5α-reduttasi. La selettività tissutale dei SARM potrebbe essere correlata all’espressione tessuto-specifica delle proteine co-regolatorie. Allo stesso modo, alcune differenze delle azioni dei SARM rispetto al Testosterone potrebbero essere correlate all’incapacità dei SARM non steroidei di subire l’aromatizzazione.
Esperienza di studi preclinici e clinici con i SARM di prima generazione
Un gran numero di SARM candidati sono stati sottoposti a studi preclinici di verifica teorica e tossicologici e sono entrati in studi clinici di fase I e II (27, 28). Gli studi preclinici hanno rivelato una promettente selettività dei tessuti; tuttavia, poiché molti di questi dati generati dalle aziende farmaceutiche sono rimasti inediti, i confronti della potenza relativa e della selettività dei tessuti tra i diversi SARM sono difficili da convalidare.
Un certo numero di SARM di prima generazione sono stati testati in prove di fase I. Questi composti sono stati posizionati per studi di efficacia precoci per il trattamento dell’osteoporosi, la fragilità ossea, la cachessia del cancro e le limitazioni funzionali associate all’invecchiamento. Inoltre, i SARM che inibiscono potentemente le gonadotropine, ma risparmiano l’attività a livello della Prostata, hanno suscitato una certa attrattiva come candidati per la contraccezione maschile. È stato proposto l’uso di SARM per il trattamento delle sindromi da carenza di androgeni negli uomini; i vantaggi relativi ai SARM rispetto al Testosterone per questa indicazione non sono immediatamente evidenti e risultano limitati. Molte funzioni biologiche del Testosterone, in particolare i suoi effetti sulla libido e sul comportamento, sulle ossa e sui lipidi plasmatici, richiedono la sua aromatizzazione in estrogeni; poiché i SARM attualmente disponibili non sono né aromatizzabili né 5-alfa riducibili, questi composti risultano fortemente limitati come base terapica di sostituzione androgena in andropausa e dovrebbero affrontare una barra normativa in salita per l’approvazione in quanto sarebbero tenuti a dimostrare efficacia e sicurezza in molti più domini di azione degli androgeni rispetto a quanto richiesto dalle formulazioni di Testosterone la quale si conosce per effetti diretti ed indiretti in condizione terapeutica sostitutiva degli androgeni endogeni.
Alle dosi che sono state testate, i SARM di prima generazione inducono modesti guadagni di massa corporea magra in volontari sani, che non sono affatto vicini ai guadagni molto maggiori nella massa muscolare scheletrica riportati con dosi sovrafisiologiche di Testosterone. I modesti guadagni da 1,0 a 1,5 kg di massa magra con i SARM di prima generazione in 4-6 settimane dovrebbero essere confrontati con i guadagni di 5-7 kg di massa magra con dosi da 300 e 600mg di Testosterone Enantato (pari approssimativamente a 216mg e 432mg di Testosterone effettivo rispettivamente). Tuttavia, è possibile che la prossima generazione di molecole SARM avrà maggiore potenza e selettività rispetto ai SARM di prima generazione, ma ad oggi non sussiste ancora dimostrazione a riguardo.
Raggiungimento della selettività e spiegazione dei meccanismi d’azione
Un altro approccio per ottenere la selettività d’azione è chiarire i meccanismi dell’azione del Testosterone sulla Prostata e identificare le molecole a valle associate all’attivazione della segnalazione AR nel muscolo scheletrico, ma non nella Prostata. Attraverso la comprensione di questi meccanismi, potrebbe essere possibile identificare molecole candidate che prendono di mira aspetti specifici della cascata di segnalazione AR.
Le analisi delle biopsie muscolari di uomini trattati con dosi graduate di testosterone hanno rivelato che la somministrazione di testosterone induce ipertrofia delle fibre muscolari sia di tipo I che di tipo II (54, 55); I cambiamenti nelle aree trasversali di entrambe le fibre di tipo I e II sono correlati alla dose di Testosterone e alle concentrazioni di Testosterone totale e libero (54). Tuttavia, né il numero assoluto né la proporzione relativa delle fibre di tipo I e II cambiano durante la somministrazione di Testosterone.
Poiché le cellule satellite muscolari sono state implicate nell’ipertrofia del muscolo scheletrico e nell’aumento del numero mionucleare (56), sono state quantificate le cellule satellite e il numero mionucleare mediante microscopia elettronica, utilizzando metodi di conteggio diretto e orientamento spaziale nelle biopsie del vasto laterale ottenute al basale e dopo 20- settimane di trattamento con un agonista del GnRH e dosi graduate di Testosterone Enantato. Il numero assoluto e percentuale di cellule satellite a 20 settimane era significativamente maggiore del basale negli uomini che ricevevano dosi sovrafisiologiche di Testosterone (57). La variazione del numero di cellule satellite era correlata alle variazioni dei livelli di Testosterone totale e libero (57). Quindi, l’ipertrofia delle fibre muscolari indotta dal Testosterone è associata ad un aumento delle cellule satellite e del numero di mionuclei.
Il Testosterone e il DHT promuovono la differenziazione delle cellule staminali mesenchimali multipotenti in linea miogenica e inibiscono la loro differenziazione in linea adipogenica (58, 59). Il Testosterone inibisce anche la differenziazione dei pre-adipociti in adipociti (59, 60). Altri hanno suggerito che l’ipertrofia indotta dal Testosterone sia causata dalla stimolazione della sintesi proteica e dall’inibizione della degradazione proteica (61, 62). Testosterone e DHT promuovono l’associazione del ligando AR con il suo co-attivatore, β-catenina; questa interazione stabilizza la β-catenina, promuove la sua traslocazione nel nucleo e l’associazione con TCF-4, e l’attivazione trascrizionale di un certo numero di geni bersaglio Wnt (63). La β-catenina svolge un ruolo essenziale nel mediare gli effetti del Testosterone sulla differenziazione miogenica. Il Testosterone sovra-regola l’espressione della Follistatina in vivo e in vitro (63); l’infusione della proteina Follistatina ricombinante aumenta la massa muscolare e diminuisce la massa grassa nei topi castrati. Il Testosterone sovra-regola l’SMAD 7 e sotto-regola la segnalazione del SMAD mediata dal TGFβ e i geni bersaglio del TGFβ (63). La Follistatina inibisce l’azione di diversi membri della famiglia del TGFβ. Questi studi supportano l’ipotesi che gli effetti del Testosterone siano trasmessi in modo incrociato dalla via Wnt alla via TGFβ-SMAD attraverso la Follistatina. Pertanto, è possibile che molecole candidate come la Follistatina che sono a valle del AR e β-catenina e che mediano gli effetti del Testosterone sul muscolo possano fornire la selettività desiderata degli effetti anabolici. La via di segnalazione mediata dal AR a valle della β-catenina può essere un interessante serbatoio di bersagli candidati per lo sviluppo di farmaci anabolizzanti selettivi.
Molecola di Follistatina
Ostacoli normativi allo sviluppo dei SARM
Negli studi di fase I e II, i SARM di prima generazione hanno mostrato riduzioni significative delle concentrazioni di colesterolo HDL e SHBG e lievi aumenti transitori di AST e ALT. Non è chiaro se gli aumenti delle transaminasi riflettano la tossicità epatica di primo passaggio tipica degli androgeni somministrati per via orale o un effetto di classe sulla trascrizione del gene AST. Allo stesso modo, la soppressione del colesterolo HDL potrebbe riflettere gli effetti combinati della via di somministrazione orale e la mancanza di aromatizzazione. È possibile che una via di somministrazione sistemica – transdermica o intramuscolare – possa attenuare il potenziale di aumento delle transaminasi e riduzioni di HDL-C.
Globulina Legante gli Ormoni Sessuali (in inglese sex hormone-binding globulin o SHBG)
Mentre il percorso normativo per l’approvazione dei farmaci per l’osteoporosi è stato ben delineato a causa della precedenza stabilita dai farmaci precedentemente approvati, il percorso per l’approvazione delle terapie anabolizzanti che promuovono la suddetta funzione non è stato chiaramente stabilito. Sono in corso sforzi considerevoli per generare un consenso su indicazioni, risultati di efficacia negli studi cardine e differenze clinicamente importanti minime nei risultati di efficacia chiave; questi sforzi dovrebbero facilitare le prove di efficacia delle molecole candidate. Ma il risultato, ad oggi, non è molto promettente.
Allora i SARM non-steroidei sono tessuto-selettivi?
Ammetto che quanto esposto fino ad ora non è propriamente “masticabile” da tutti, ed è per questo che vi renderò la comprensione più facile.
Allora, un modo per ottenere la selettività tissutale è tramite un fapping molecolare che implica l’attivazione del recettore degli androgeni (AR) specificamente nel tessuto muscolare. Mentre l’AR è lo stesso in tutti i tessuti, il contesto cellulare è diverso: puoi immaginare che il contenuto di una cellula muscolare sia abbastanza diverso da quello di una cellula della ghiandola sebacea. Quando l’AR viene attivato per indurre la trascrizione genica, che alla fine porterà ai guadagni muscolari, entrano in gioco molte altre proteine. Queste proteine coinvolte nella trascrizione sono i cosiddetti coregolatori trascrizionali. Chiamiamoli cofattori in breve. Questi possono aiutare nella trascrizione (coattivatori) o reprimerla (corepressori). Quei cofattori, e le loro proporzioni, che vengono reclutati da un AR attivato, possono variare da un tessuto all’altro. Questo dipende, in parte, da quale molecola è legata all’AR. In quanto tale, un SARM potrebbe essere in grado di reclutare un gruppo di cofattori che porteranno a una trascrizione genica minima o nulla nel tessuto A (Prostata), mentre portano alla trascrizione genica completa nel tessuto B (Muscolo).
Quanto detto sopra sembra comunque piuttosto complesso, e lo è, ma non mi è possibile comunicare a gesti per spiegarvi una cosa che è di base complessa. Comunque sia, come si fa a sapere quale tipo di ligando per l’AR interagisce con quali cofattori e in che misura? Non lo fa, si dovrebbero eseguire test quasi infiniti sul composto in questione per determinarlo effettivamente. E questo processo sembra richiedere molto tempo. Tuttavia, questo è attualmente pubblicizzato come uno dei motivi per cui i SARM – in sostanza avendolo scoperto per “caso” – esercitano i loro effetti specifici sui tessuti. Ad esempio, è stato dimostrato che l’antiandrogeno steroideo TSAA-291 esercita un’attività tessuto-specifica che coincide con profili di reclutamento di coregolatori differenziali rispetto al Diidrotestosterone (DHT) [64]. Tuttavia, poiché non hanno confrontato altri AAS, potrebbe anche essere che avrebbero visto diversi profili di reclutamento di coregolatori con altri AAS. Pertanto, è difficile vedere quanto sia effettivamente rilevante per le proprietà specifiche dei SARM. Dopotutto, la correlazione non implica la causalità.
Oxendolone (TSAA-291)
Andando avanti con la semplificazione pratica del concetto di selettività specifica, un altro modo in cui un SARM potrebbe esercitare tale specificità tissutale è attraverso il la sua via di metabolizzazione. Una molecola viene metabolizzata dall’azione degli enzimi. E la presenza di tali enzimi metabolizzanti può differire da un tessuto all’altro. Ad esempio, questo è molto evidente con la metabolizzazione del Testosterone. Il Testosterone è suscettibile di metabolizzazione per riduzione sul suo quinto atomo di carbonio. Questa riduzione è catalizzata dall’enzima 5α-reduttasi. Il risultato di questa riduzione è il più potente androgeno Diidrotestosterone (DHT). Pertanto, l’effetto del testosterone viene amplificato nei tessuti che esprimono questo enzima. Sfortunatamente, il muscolo scheletrico non è uno di quei tessuti. E, in effetti, il DHT viene degradato nel molto debole androgeno 3α-Androstanediolo dall’enzima 3α-HSD nel muscolo [65], diminuendo così il suo effetto in loco.
3α-idrossisteroide deidrogenasi ( 3α-HSD o aldo-cheto reduttasi famiglia 1 membro C4)
Tuttavia, questo aspetto è leggermente diverso per i SARM. Gli enzimi steroidogeni, come la 5α-reduttasi e la 3α-HSD, non hanno effetto sui SARM non steroidei. Gli enzimi che metabolizzano i SARM variano da una classe di SARM all’altra. Come tale, deve essere studiato per ogni SARM, analizzandone il modo in cui viene metabolizzato e con quale velocità ciò si verifica nei vari tessuti. Questo risulta essere più banale per la maggior parte degli AAS sui quali possiamo ampiamente prevederlo. Contrariamente, risulta difficile per lo sviluppo dei SARM non steroidei.
3α-Androstanediolo
Infine, è noto che gli AAS possono esercitare anche effetti non genomici [66]. Come suggerisce il nome, questi sono effetti che non sono mediati dalla trascrizione genica. Pertanto, questi effetti si verificano molto rapidamente (entro secondi/minuti dopo l’esposizione di una cellula ad essa). Alcune ricerche indicano che il recettore degli androgeni localizzato nella membrana plasmatica, così come altri recettori legati alla membrana, mediano questi effetti. Ipoteticamente è possibile che AAS – e per estensione SARM – siano in grado di influenzare le vie di segnalazione a seconda del contesto cellulare, cioè gli effetti potrebbero differire da una cellula all’altra: specificità del tessuto.
Più di 20 anni di ricerca sui SARM ma nessuna approvazione clinica
Sapere queste cose è interessante e utile per comprendere l’attività di tali molecole, ma tali attività ci mostrano di essere ben lungi (ancora) dal possedere la chiave di volta nello sviluppo di SARM terapeuticamente e pienamente efficaci. Ma almeno abbiamo una base attraverso la quale i SARM potrebbero effettivamente funzionare. Tuttavia, dopo oltre 2 decenni di ricerca sui SARM [67], nessuno è stato approvato dalla Food and Drug Administration (FDA). E no, non c’entra “bIg PhaRma”, complottaro da tastiera.
Parte del motivo per cui ciò avviene può essere ricondotto al modo in cui i ricercatori hanno esaminato i potenziali SARM. Come ho riportato in un mio precedente articolo, la anabolico:androgeno ratio, come valutato dal test di Hershberger, è pressoché inutile. Eppure questo test è stato utilizzato dalle aziende farmaceutiche per decidere se perseguire o meno la ricerca su determinati SARM di particolare interesse, queste aziende includono la GTx, Inc. con lo sviluppo del Enobosarm (GTx-024) [68], la GlaxoSmithKline con lo sviluppo del GSK2881078 [69 ], la Takeda Pharmaceutical Company con lo sviluppo del SARM-2f [70], la Aska Pharmaceuticals con lo sviluppo del S42 [71], e la Merck & Co, Inc con lo sviluppo del MK-4541 [72], ecc.
Non si sono forse già visti risultati ridicolmente buoni con AAS convenzionali in passato utilizzando questi test? Si, e non per una molecola. Ad esempio, si dice che lo Stanozololo abbia un rapporto anabolico/androgeno circa 10 volte superiore a quello del Testosterone, mentre il Methyldrostanolone ha circa un rapporto anabolico/androgeno 20 volte superiore [73]. Tuttavia, come sappiamo, queste molecole non sono considerate SARM sito-specifici e non sono scevre da eventuali effetti androgenizzanti. Perché? Perchè uno studio con molteplici variabili svolto su roditori non può essere rapportato correttamente all’uomo, come ho spiegato nell’articolo dedicato alla anabolico:androgeno ratio.
Un ulteriore problema con la ricerca sui SARM emerge quando si esaminano gli studi clinici. Poiché i SARM vengono sviluppati per superare gli AAS convenzionali, non ci si aspetterebbe forse che essi vengano confrontati con gli AAS convenzionali negli studi clinici? Per qualche ragione, in tutti gli studi clinici con i SARM, questi vengono confrontati con un placebo. Se si vuole valutare l’efficacia reale di una molecola rispetto ad un altra, non lo si fa confrontandola solo ad un placebo, o forse solo inizialmente lo si farebbe, come in una sperimentazione pilota per risparmiare sui costi, e per valutare se ne vale la pena o meno. Questi studi mostrano comunemente guadagni marginali (nell’ordine di 1kg) di LBM in un periodo di diverse settimane/mesi con una corrispondente buona tollerabilità. Anche gli AAS convenzionali sono generalmente ben tollerati e aumentano marginalmente l’LBM quando vengono somministrati a basso dosaggio, niente di sconvolgente in questo. La Ligand Pharmaceuticals ha persino trovato la necessità di menzionare quanto segue nella conclusione del loro abstract di studio che copre gli effetti del loro SARM LGD-4033: “LGD-4033 era sicuro, aveva un profilo farmacocinetico favorevole e un aumento della massa corporea magra anche durante questo breve periodo senza cambiamento nell’antigene prostatico specifico”. Cosa si aspettavano in poche settimane di trattamento con il loro SARM? Anche 600mg di Testosterone Enantato a settimana per 20 settimane non aumentano l’antigene prostatico specifico (PSA) negli uomini giovani [74, 75] o negli uomini più anziani [76].
LGD-4033
Se l’unico requisito ricercato è che un SARM non steroideo sia più efficace di un placebo pur essendo ben tollerato, ce l’hanno fatta. Ma praticamente tutti gli AAS convenzionali sono anche più efficaci di un placebo pur essendo ben tollerati. Superare il placebo non è mai stato l’obiettivo dello sviluppo dei SARM, quindi perché gli studi testa a testa sono ancora gravemente carenti? Forse perchè non vi è superiorità ne negli effetti benefici e nel rapporto tra benefici e rischi sistemici? …
Conclusioni:
I SARM si basano sulla selettività dei tessuti per esercitare i loro effetti anabolici (costruzione muscolare), mantenendo gli effetti collaterali al minimo assoluto. Dopotutto, gli effetti collaterali si riducono in gran parte, ma non totalmente, all’azione androgena nei tessuti diversi dai muscoli. I SARM possono esercitare questi effetti tessuto-specifici attraverso circa tre diversi meccanismi. Uno sfrutta le differenze nelle molecole tra i diversi tipi di cellule che “aiutano” un SARM ad avviare la trascrizione genica. Un altro si basa su enzimi di espressione tessuto-specifici che metabolizzano il SARM. Un terzo si basa sugli effetti non genomici che potrebbero essere mediati da un SARM che, ancora una volta, potrebbe variare da un tipo di cellula all’altro.
Poiché questi processi biochimici sono estremamente difficile da prevedere in anticipo, le aziende farmaceutiche devono esaminare molte molecole per vedere quale potrebbe essere la soluzione migliore. Nessun SARM è stato ancora approvato e credo che ciò sia in parte dovuto a questo processo di screening che si basa su metodi obsoleti e imperfetti come il test di Hershberger e all’incapacità di sopperire all’attività fisiologica del DHT e dell’Estradiolo, i quali subiscono una marcata soppressione consequenziale al abbassamento dei livelli di Testosterone endogeno. Questo punto deve essere sicuramente migliorato. Ed è quindi questa la strada che dovrebbe intraprendere la ricerca sui SARM.
Negli sport, ed in particolare nel Bodybuilding, l’uso dei SARM non steroidei, dopo l’iniziale eccitazione per le promesse commerciali affiancate al loro uso da parte dei rivenditori e brand, sono caduti in un uso più che altro amatoriale, da parte di persone poco informate in materia e dalla mente facilmente manipolabile dalla pubblicità e informazioni incomplete se non del tutto errate.
L’unico ambito in cui i SARM non steroidei hanno visto un certo potenziale è nel culturismo femminile. In questa circostanza, le molecole più testate, prima su tutte l’Ostarina, ha mostrato un certo vantaggio se l’obbiettivo era quello di aumenti contenuti del tessuto muscolare e la mancanza di possibili effetti mascolinizzanti alle dosi comprese tra 5 e 10mg/die.
Nell’uso maschile i SARM hanno lasciato una serie di delusioni e promesse non mantenute. In monoterapia il loro uso ha portato ad atleti con problemi non indifferenti nella sfera sessuale, con difficoltà di raggiungimento e mantenimento dell’erezione, letargia, stanchezza cronica, affaticabilità, depressione e stati ansiosi. Tutti sintomi legati ad un calo significativo del DHT e del Estradiolo, con conseguente riduzione o mancanza della loro, per esempio, attività a livello cerebrale (neurosteroideo).
Di conseguenza, utilizzare uno o più SARM senza una base esogena di Testosterone (o, per lo meno, di hCG) è una totale pazzia! E, comunque, l’uso dei SARM come aggiunte ad un ciclo di classici AAS iniettabili non risulta quasi mai all’altezza delle aspettative di risposta ipertrofica rispetto all’uso, per esempio, di AAS orali come starter e/o finisher. Ovviamente la valutazione si basa anche e soprattutto sul rapporto effetti collaterali:benefici in contesto preparatorio correttamente impostato.
Inoltre, gli effetti collaterali a livello epatico e della lipidemia ematica non sono estranei all’uso di SARM non steroidei, sebbene essi si mostrino a diverso grado di entità molecola-dipendente e dose-dipendente. La stessa Ostarina aveva mostrato lievi alterazioni di ALT e AST con riduzione del HDL al dosaggio di 3mg in studi clinici; la molecola in ambito “physique” viene assunta ad un dosaggio nel range di 10-20mg/die, e l’impatto sulle transaminasi, colesterolo totale, LDL e HDL osservato attraverso esami ematici mostrano variazioni significative e variabili in misura soggettiva.
Il SARM non steroideo con il più alto carico di effetti collaterali è risultato essere LGD4033, il quale, in diversi casi studio, ha mostrato di poter causare forte stress epatico oltre che alterare sensibilmente la lipidemia ematica. Nel caso di questa molecola, si è osservato anche una perdita della selettività con possibile comparsa di effetti androgenicizzanti. Complice di questi riscontri è soprattutto l’abuso che se ne fa della molecola, sforando i dosaggi efficaci e contenitivi (2-8mg/die) a favore di somministrazioni elevate (≥10mg/die).
Anche il RAD140 sembra non essere privo di effetti collaterali significativi a livello epatico, nonostante il suo potenziale effetto protettivo sulla Prostata che, a dosaggi minimi (5mg/die) potrebbe avere un riscontro terapeutico preventivo per l’ipertrofia prostatica.
SARM non-steroideo RAD140
Lascerei perdere discorsi ipotetici su altri SARM comunemente utilizzati dagli atleti (specialmente amatori) ma che alle spalle sono privi di studi clinici (vedi, per esempio, l’S23) e, quindi, di dati oggettivi sulle possibili attività nell’uomo. L’unica eccezione tra questi la fa, forse, il SARM steroideo YK11, il quale sembra essere gestibile a dosi di 5-10mg/die con un buon rapporto tra benefici ed alterazioni dei marker ematici.
SARM steroideo YK11
Per concludere, mi sembra di avervi dato sufficienti informazioni per valutare correttamente i SARM e deporli con cognizione logica dall'”altarino” di innocuità sul quale brand e venditori li hanno posti e dove una parte di voi continua a tenerli.
Gabriel Bellizzi
Riferimenti:
Mohler ML, Bohl CE, Jones A, Coss CC, Narayanan R, He Y, Hwang DJ, Dalton JT, Miller DD (June 2009). “Nonsteroidal selective androgen receptor modulators (SARMs): dissociating the anabolic and androgenic activities of the androgen receptor for therapeutic benefit”. Journal of Medicinal Chemistry. 52(12): 3597–617.
Yin D, Gao W, Kearbey JD, Xu H, Chung K, He Y, Marhefka CA, Veverka KA, Miller DD, Dalton JT (March 2003). “Pharmacodynamics of selective androgen receptor modulators”. The Journal of Pharmacology and Experimental Therapeutics. 304 (3): 1334–40.
Manfredi MC, Bi Y, Nirschl AA, Sutton JC, Seethala R, Golla R, Beehler BC, Sleph PG, Grover GJ, Ostrowski J, Hamann LG (August 2007). “Synthesis and SAR of tetrahydropyrrolo[1,2-b][1,2,5]thiadiazol-2(3H)-one 1,1-dioxide analogues as highly potent selective androgen receptor modulators”. Bioorganic & Medicinal Chemistry Letters. 17 (16): 4487–90.
Zhang X, Li X, Allan GF, Sbriscia T, Linton O, Lundeen SG, Sui Z (August 2007). “Design, synthesis, and in vivo SAR of a novel series of pyrazolines as potent selective androgen receptor modulators”. Journal of Medicinal Chemistry. 50 (16): 3857–69.
Long YO, Higuchi RI, Caferro TR, Lau TL, Wu M, Cummings ML, Martinborough EA, Marschke KB, Chang WY, López FJ, Karanewsky DS, Zhi L (May 2008). “Selective androgen receptor modulators based on a series of 7H-[1,4]oxazino[3,2-g]quinolin-7-ones with improved in vivo activity”. Bioorganic & Medicinal Chemistry Letters. 18 (9): 2967–71.
Bhasin S, Cunningham GR, Hayes FJ, Matsumoto AM, Snyder PJ, Swerdloff RS, Montori VM. Testosterone therapy in adult men with androgen deficiency syndromes: an endocrine society clinical practice guideline. J Clin Endocrinol Metab. 2006;91:1995–2010.
Baumgartner RN. Body composition in healthy aging. Annals of the New York Academy of Sciences. 2000;904:437–448.
Bassey EJ, Fiatarone MA, O’Neill EF, Kelly M, Evans WJ, Lipsitz LA. Leg extensor power and functional performance in very old men and women. Clin Sci (Lond) 1992;82:321–327.
. Lexell J, Downham D, Sjostrom M. Distribution of different fibre types in human skeletal muscles. A statistical and computational study of the fibre type arrangement in m. vastus lateralis of young, healthy males. Journal of the neurological sciences. 1984;65:353–365.
Orwoll E, Lambert LC, Marshall LM, Blank J, Barrett-Connor E, Cauley J, Ensrud K, Cummings SR. Endogenous testosterone levels, physical performance, and fall risk in older men. Arch Intern Med. 2006;166:2124–2131.
Bhasin S, Storer TW, Berman N, Yarasheski KE, Clevenger B, Phillips J, Lee WP, Bunnell TJ, Casaburi R. Testosterone replacement increases fat-free mass and muscle size in hypogonadal men. J Clin Endocrinol Metab. 1997;82:407–413.
Snyder PJ, Peachey H, Berlin JA, Hannoush P, Haddad G, Dlewati A, Santanna J, Loh L, Lenrow DA, Holmes JH, Kapoor SC, Atkinson LE, Strom BL. Effects of testosterone replacement in hypogonadal men. J Clin Endocrinol Metab. 2000;85:2670–2677.
. Bhasin S, Storer TW, Berman N, Callegari C, Clevenger B, Phillips J, Bunnell TJ, Tricker R, Shirazi A, Casaburi R. The effects of supraphysiologic doses of testosterone on muscle size and strength in normal men. N Engl J Med. 1996;335:1–7.
. Bhasin S, Woodhouse L, Casaburi R, Singh AB, Bhasin D, Berman N, Chen X, Yarasheski KE, Magliano L, Dzekov C, Dzekov J, Bross R, Phillips J, Sinha-Hikim I, Shen R, Storer TW. Testosterone dose-response relationships in healthy young men. Am J Physiol Endocrinol Metab. 2001;281:E1172–1181.
Bhasin S, Woodhouse L, Casaburi R, Singh AB, Mac RP, Lee M, Yarasheski KE, Sinha-Hikim I, Dzekov C, Dzekov J, Magliano L, Storer TW. Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle. J Clin Endocrinol Metab. 2005;90:678–688.
Casaburi R, Bhasin S, Cosentino L, Porszasz J, Somfay A, Lewis MI, Fournier M, Storer TW. Effects of testosterone and resistance training in men with chronic obstructive pulmonary disease. Am J Respir Crit Care Med. 2004;170:870–878.
Johansen KL, Mulligan K, Schambelan M. Anabolic effects of nandrolone decanoate in patients receiving dialysis: a randomized controlled trial. Jama. 1999;281:1275–1281.
Woodhouse LJ, Reisz-Porszasz S, Javanbakht M, Storer TW, Lee M, Zerounian H, Bhasin S. Development of models to predict anabolic response to testosterone administration in healthy young men. Am J Physiol Endocrinol Metab. 2003;284:E1009–1017.
Storer TW, Magliano L, Woodhouse L, Lee ML, Dzekov C, Dzekov J, Casaburi R, Bhasin S. Testosterone dose-dependently increases maximal voluntary strength and leg power, but does not affect fatigability or specific tension. J Clin Endocrinol Metab. 2003;88:1478–1485.
Calof O, Singh AB, Lee ML, Urban RJ, Kenny AM, Tenover JL, Bhasin S. Adverse events associated with testosterone supplementation of odler men. J Greontol Med Sci. 2005 in press.
Bhasin S, Calof OM, Storer TW, Lee ML, Mazer NA, Jasuja R, Montori VM, Gao W, Dalton JT. Drug insight: Testosterone and selective androgen receptor modulators as anabolic therapies for chronic illness and aging. Nature Clinical Practice Endocrinology & Metabolism. 2006;2:146–159.
Narayanan R, Mohler ML, Bohl CE, Miller DD, Dalton JT. Selective androgen receptor modulators in preclinical and clinical development. Nuclear receptor signaling. 2008;6:e010. An excellent treatise of SARM chemistry and structure-activity relationships.
Negro-Vilar A. Selective androgen receptor modulators (SARMs): a novel approach to androgen therapy for the new millennium. J Clin Endocrinol Metab. 1999;84:3459–3462.
Yin D, He Y, Perera MA, Hong SS, Marhefka C, Stourman N, Kirkovsky L, Miller DD, Dalton JT. Key structural features of nonsteroidal ligands for binding and activation of the androgen receptor. Mol Pharmacol. 2003;63:211–223.
Gao W, Kearbey JD, Nair VA, Chung K, Parlow AF, Miller DD, Dalton JT. Comparison of the pharmacological effects of a novel selective androgen receptor modulator, the 5alpha-reductase inhibitor finasteride, and the antiandrogen hydroxyflutamide in intact rats: new approach for benign prostate hyperplasia. Endocrinology. 2004;145:5420–5428.
Gao W, Reiser PJ, Coss CC, Phelps MA, Kearbey JD, Miller DD, Dalton JT. Selective Androgen Receptor Modulator (SARM) Treatment Improves Muscle Strength and Body Composition, and Prevents Bone Loss in Orchidectomized Rats. Endocrinology 2005
Gao W, Reiser PJ, Coss CC, Phelps MA, Kearbey JD, Miller DD, Dalton JT. Selective androgen receptor modulator treatment improves muscle strength and body composition and prevents bone loss in orchidectomized rats. Endocrinology. 2005;146:4887–4897.
Gao W, Reiser PJ, Kearbey JD, Phelps MA, Coss CC, Miller DD, Dalton JT. Effects of Novel Selective Androgen Receptor Modulator (SARM) on Skeletal Muscle Mass and Strength in Castrated Male Rats. The Endocrine Society; New Orleans: 2004.
Kearbey JD, Gao W, Narayanan R, Fisher SJ, Wu D, Miller DD, Dalton JT. Selective Androgen Receptor Modulator (SARM) treatment prevents bone loss and reduces body fat in ovariectomized rats. Pharmaceutical research. 2007;24:328–335.
Bohl CE, Gao W, Miller DD, Bell CE, Dalton JT. Structural basis for antagonism and resistance of bicalutamide in prostate cancer. Proc Natl Acad Sci U S A. 2005;102:6201–6206. An important paper that describes the structural basis of antagonism of bicalutamide based on the crystal structure.
Hamann LG, Manfredi MC, Sun C, Krystek SR, Jr, Huang Y, Bi Y, Augeri DJ, Wang T, Zou Y, Betebenner DA, Fura A, Seethala R, Golla R, Kuhns JE, Lupisella JA, Darienzo CJ, Custer LL, Price JL, Johnson JM, Biller SA, Zahler R, Ostrowski J. Tandem optimization of target activity and elimination of mutagenic potential in a potent series of N-aryl bicyclic hydantoin-based selective androgen receptor modulators. Bioorganic & medicinal chemistry letters. 2007;17:1860–1864.
Manfredi MC, Bi Y, Nirschl AA, Sutton JC, Seethala R, Golla R, Beehler BC, Sleph PG, Grover GJ, Ostrowski J, Hamann LG. Synthesis and SAR of tetrahydropyrrolo[1,2-b][1,2,5]thiadiazol-2(3H)-one 1,1-dioxide analogues as highly potent selective androgen receptor modulators. Bioorganic & medicinal chemistry letters. 2007;17:4487–4490.
Ostrowski J, Kuhns JE, Lupisella JA, Manfredi MC, Beehler BC, Krystek SR, Jr, Bi Y, Sun C, Seethala R, Golla R, Sleph PG, Fura A, An Y, Kish KF, Sack JS, Mookhtiar KA, Grover GJ, Hamann LG. Pharmacological and x-ray structural characterization of a novel selective androgen receptor modulator: potent hyperanabolic stimulation of skeletal muscle with hypostimulation of prostate in rats. Endocrinology. 2007;148:4–12.
Kim J, Wu D, Hwang DJ, Miller DD, Dalton JT. The para substituent of S-3-(phenoxy)-2-hydroxy-2-methyl-N-(4-nitro-3-trifluoromethyl-phenyl)-prop ionamides is a major structural determinant of in vivo disposition and activity of selective androgen receptor modulators. The Journal of pharmacology and experimental therapeutics. 2005;315:230–239.
Hanada K, Furuya K, Yamamoto N, Nejishima H, Ichikawa K, Nakamura T, Miyakawa M, Amano S, Sumita Y, Oguro N. Bone anabolic effects of S-40503, a novel nonsteroidal selective androgen receptor modulator (SARM), in rat models of osteoporosis. Biol Pharm Bull. 2003;26:1563–1569.
Miner JN, Chang W, Chapman MS, Finn PD, Hong MH, Lopez FJ, Marschke KB, Rosen J, Schrader W, Turner R, van Oeveren A, Viveros H, Zhi L, Negro-Vilar A. An orally active selective androgen receptor modulator is efficacious on bone, muscle, and sex function with reduced impact on prostate. Endocrinology. 2007;148:363–373.
van Oeveren A, Motamedi M, Mani NS, Marschke KB, Lopez FJ, Schrader WT, Negro-Vilar A, Zhi L. Discovery of 6-N,N-bis(2,2,2-trifluoroethyl)amino-4-trifluoromethylquinolin-2(1H)-one as a novel selective androgen receptor modulator. Journal of medicinal chemistry. 2006;49:6143–6146.
van Oeveren A, Motamedi M, Martinborough E, Zhao S, Shen Y, West S, Chang W, Kallel A, Marschke KB, Lopez FJ, Negro-Vilar A, Zhi L. Novel selective androgen receptor modulators: SAR studies on 6-bisalkylamino-2-quinolinones. Bioorganic & medicinal chemistry letters. 2007;17:1527–1531.
Ng RA, Lanter JC, Alford VC, Allan GF, Sbriscia T, Lundeen SG, Sui Z. Synthesis of potent and tissue-selective androgen receptor modulators (SARMs): 2-(2,2,2)-Trifluoroethyl-benzimidazole scaffold. Bioorganic & medicinal chemistry letters. 2007;17:1784–1787.
Narayanan R, Coss CC, Yepuru M, Kearbey JD, Miller DD, Dalton JT. Steroidal androgens and nonsteroidal, tissue-selective androgen receptor modulator, S-22, regulate androgen receptor function through distinct genomic and nongenomic signaling pathways. Mol Endocrinol. 2008;22:2448–2465. This paper showed that DHT and SARMs activate distinct signaling pathways.
Masiello D, Chen SY, Xu Y, Verhoeven MC, Choi E, Hollenberg AN, Balk SP. Recruitment of beta-catenin by wild-type or mutant androgen receptors correlates with ligand-stimulated growth of prostate cancer cells. Mol Endocrinol. 2004;18:2388–2401.
Song LN, Herrell R, Byers S, Shah S, Wilson EM, Gelmann EP. Beta-catenin binds to the activation function 2 region of the androgen receptor and modulates the effects of the N-terminal domain and TIF2 on ligand-dependent transcription. Mol Cell Biol. 2003;23:1674–1687.
Sathya G, Chang CY, Kazmin D, Cook CE, McDonnell DP. Pharmacological uncoupling of androgen receptor-mediated prostate cancer cell proliferation and prostate-specific antigen secretion. Cancer Res. 2003;63:8029–8036.
Sathya G, Chang CY, Kazmin D, Cook CE, McDonnell DP. Pharmacological uncoupling of androgen receptor-mediated prostate cancer cell proliferation and prostate-specific antigen secretion. Cancer Res. 2003;63:8029–8036.
Bohl CE, Wu Z, Miller DD, Bell CE, Dalton JT. Crystal structure of the T877A human androgen receptor ligand-binding domain complexed to cyproterone acetate provides insight for ligand-induced conformational changes and structure-based drug design. J Biol Chem. 2007;282:13648–13655.
Heinlein CA, Chang C. Androgen receptor (AR) coregulators: an overview. Endocr Rev. 2002;23:175–200.
Hamann LG. Discovery and preclinical profile of a highly potent and muscle selective androgen receptor modulator (SARM). 227th National Meeting of the American Chemical Society Medicinal Chemistry Division.2004.
Sinha-Hikim I, Artaza J, Woodhouse L, Gonzalez-Cadavid N, Singh AB, Lee MI, Storer TW, Casaburi R, Shen R, Bhasin S. Testosterone-induced increase in muscle size in healthy young men is associated with muscle fiber hypertrophy. Am J Physiol Endocrinol Metab. 2002;283:E154–164.
Kadi F, Eriksson A, Holmner S, Thornell LE. Effects of anabolic steroids on the muscle cells of strength-trained athletes. Medicine and science in sports and exercise. 1999;31:1528–1534.
Sinha-Hikim I, Roth SM, Lee MI, Bhasin S. Testosterone-induced muscle hypertrophy is associated with an increase in satellite cell number in healthy, young men. Am J Physiol Endocrinol Metab. 2003;285:E197–205.
Singh R, Artaza JN, Taylor WE, Gonzalez-Cadavid NF, Bhasin S. Androgens stimulate myogenic differentiation and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through an androgen receptor-mediated pathway. Endocrinology. 2003;144:5081–5088. This paper was the first to report that androgens regulate myogenic differentiation of mesenchymal multipotent cells.
Gupta V, Bhasin S, Guo W, Singh R, Miki R, Chauhan P, Choong K, Tchkonia T, Lebrasseur NK, Flanagan JN, Hamilton JA, Viereck JC, Narula NS, Kirkland JL, Jasuja R. Effects of dihydrotestosterone on differentiation and proliferation of human mesenchymal stem cells and preadipocytes. Molecular and cellular endocrinology. 2008;296:32–40.
Singh R, Artaza JN, Taylor WE, Braga M, Yuan X, Gonzalez-Cadavid NF, Bhasin S. Testosterone inhibits adipogenic differentiation in 3T3-L1 cells: nuclear translocation of androgen receptor complex with beta-catenin and T-cell factor 4 may bypass canonical Wnt signaling to down-regulate adipogenic transcription factors. Endocrinology. 2006;147:141–154.
Brodsky IG, Balagopal P, Nair KS. Effects of testosterone replacement on muscle mass and muscle protein synthesis in hypogonadal men–a clinical research center study. J Clin Endocrinol Metab. 1996;81:3469–3475.
Ferrando AA, Sheffield-Moore M, Yeckel CW, Gilkison C, Jiang J, Achacosa A, Lieberman SA, Tipton K, Wolfe RR, Urban RJ. Testosterone administration to older men improves muscle function: molecular and physiological mechanisms. Am J Physiol Endocrinol Metab. 2002;282:E601–607.
Singh R, Bhasin S, Braga M, Artaza JN, Pervin S, Taylor WE, Krishnan V, Sinha SK, Rajavashisth TB, Jasuja R. Regulation of Myogenic Differentiation by Androgens: Cross-Talk between Androgen Receptor/{beta}-Catenin and Follistatin/TGF-{beta} Signaling Pathways. Endocrinology. 2008 This paper describes the important role of beta-catenin/Wnt pathway in mediating the effects of testosterone on myogenic differentiation and the role of follistatin in cross-communicating the signal from Wnt to TGFbeta/SMAD pathway.
Hikichi, Yukiko, et al. “Selective androgen receptor modulator activity of a steroidal antiandrogen TSAA-291 and its cofactor recruitment profile.” European journal of pharmacology 765 (2015): 322-331.
Becker, H., et al. “In vivo uptake and metabolism of 3H-testosterone and 3H-5α-dihydrotestosterone by human benign prostatic hypertrophy.” European Journal of Endocrinology 71.3 (1972): 589-599.
Foradori, C. D., M. J. Weiser, and R. J. Handa. “Non-genomic actions of androgens.” Frontiers in neuroendocrinology 29.2 (2008): 169-181.
Negro-Vilar, Andres. “Selective androgen receptor modulators (SARMs): a novel approach to androgen therapy for the new millennium.” The Journal of Clinical Endocrinology & Metabolism 84.10 (1999): 3459-3462.
Kim, Juhyun, et al. “The para substituent of S-3-(phenoxy)-2-hydroxy-2-methyl-N-(4-nitro-3-trifluoromethyl-phenyl)-propionamides is a major structural determinant of in vivo disposition and activity of selective androgen receptor modulators.” Journal of Pharmacology and Experimental Therapeutics 315.1 (2005): 230-239.
Neil, David, et al. “GSK2881078, a SARM, produces dose-dependent increases in lean mass in healthy older men and women.” The Journal of Clinical Endocrinology & Metabolism 103.9 (2018): 3215-3224.
Aikawa, Katsuji, et al. “Synthesis and biological evaluation of novel selective androgen receptor modulators (SARMs) Part III: Discovery of 4-(5-oxopyrrolidine-1-yl) benzonitrile derivative 2f as a clinical candidate.” Bioorganic & medicinal chemistry 25.13 (2017): 3330-3349.
Min, Liu, et al. “A novel synthetic androgen receptor ligand, S42, works as a selective androgen receptor modulator and possesses metabolic effects with little impact on the prostate.” Endocrinology 150.12 (2009): 5606-5616.
Schmidt, Azriel, et al. “Identification of an anabolic selective androgen receptor modulator that actively induces death of androgen-independent prostate cancer cells.” The Journal of steroid biochemistry and molecular biology 143 (2014): 29-39.
Basaria, Shehzad, et al. “The safety, pharmacokinetics, and effects of LGD-4033, a novel nonsteroidal oral, selective androgen receptor modulator, in healthy young men.” Journals of Gerontology Series A: Biomedical Sciences and Medical Sciences 68.1 (2013): 87-95.
Bhasin, Shalender, et al. “Testosterone dose-response relationships in healthy young men.” American Journal of Physiology-Endocrinology And Metabolism (2001).
Bhasin, Shalender, et al. “Effect of testosterone supplementation with and without a dual 5α-reductase inhibitor on fat-free mass in men with suppressed testosterone production: a randomized controlled trial.” Jama 307.9 (2012): 931-939.
Bhasin, Shalender, et al. “Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle.” The Journal of Clinical Endocrinology & Metabolism 90.2 (2005): 678-688.
Di norma il Nandrolone viene considerato un composto con una bassa androgenicità pari a circa il 37% di quella del Testosterone. Questa sua caratteristica è stata osservata e documentata fin dai primi anni di sperimentazione clinica. Però, la questione della sua androgenicità vista in questo modo risulta incompleta e, quindi, limitante per la sua reale e totale valutazione.
Sono decenni che atleti, soprattutto di sesso femminile, optano per il Nandrolone dal momento che i suoi effetti androgeni sono rari e generalmente lievi. Gli uomini che soffrono di alopecia androgenetica e che desiderano ridurre al minimo la perdita di capelli con l’uso di AAS hanno optato per anni per l’uso del Nandrolone come parte della base dei loro cicli.
E allora qual è il problema? Beh, esso risiede nelle abitudini di trattamento iatrogeno che gli atleti con alopecia androgenetica abbinano alle loro preparazioni contenenti Nandrolone al fine di arrestare la perdita di capelli…
Ma vediamo di cosa si tratta…
C’è “androgenicità” e “androgenicità”…
Il motivo per cui il Nandrolone è abbastanza sicuro per i capelli è la sua relativa mancanza di androgenicità nel corpo.
Rispetto agli ormoni di base che normalmente vengono considerati come punto di riferimento per la caduta dei capelli (vedi Testosterone e DHT), il Nandrolone è un’opzione superiore. Tuttavia, questo non avviene perché il Nandrolone è meno androgeno del DHT ma non del Testosterone. In effetti, il Nandrolone è più androgeno del Testosterone, sebbene risulti comunque meno androgeno del DHT.
La bassa attività androgena è dovuta esclusivamente all’interazione del Nandrolone con la 5α-reduttasi. Come ben sappiamo, la 5α-riduttasi è l’enzima principalmente responsabile per la sintesi di Diidrotestosterone (DHT) nel corpo. Il Testosterone viene in parte convertito tramite la 5α-reduttasi nel suo metabolita più androgeno, il DHT. Questo stesso processo enzimatico converte anche in parte il Nandrolone in un metabolita noto come Diidronandrolone (DHN).
Il DHN è il meno androgeno dei quattro ormoni sopra citati.
Se dovessimo elencare questi quattro ormoni in ordine dal più al meno androgeno, verrebbe fuori una graduatoria così come segue:
Diidrotestosterone (DHT) – maggior attività androgena
Nandrolone
Testosterone
Diidronandrolone (DHN) – minore attività androgena.
5α-Dihydronandrolone [DHN]
Poiché la 5α-reduttasi è significativamente espressa nel cuoio capelluto, in realtà si finisce per restare in una situazione di bassa nocività del Nandrolone a livello del bulbo del capello attraverso i processi enzimatici endogeni naturali.
In effetti, questo è anche il motivo per cui l’inibizione di questo processo enzimatico attraverso l’uso degli inibitori della 5α-reduttasi può diventare problematico per gli utilizzatori di AAS.
Coloro che utilizzano il Nandrolone con Finasteride o Dutasteride vanno ad inibire effettivamente questo processo enzimatico della 5α-reduttasi che converte il Nandrolone nel metabolita DHN, molto più sicuro per i capelli. Pertanto, quando si combina il Nandrolone con Finasteride o Dutasteride, si verifica l’effetto opposto rispetto a quanto accade con il Testosterone.[1]
Effetto degli inibitori della 5α-reduttasi sulla attività androgena di Testosterone, Nandrolone e Trestolone in ratti castrati.
Come si può vedere dal grafico sopra esposto, sebbene vada considerato con un certo margine di fallibilità parziale dal momento che si tratta di uno studio svolto su animali (ratti), l’uso di un inibitore della 5α-reduttasi ha parzialmente inibito la conversione del Testosterone in DHT. Questo è il motivo per cui la stimolazione ventrale della prostata (legato all’attività androgena come alla caduta dei capelli) è diminuita così significativamente nel gruppo trattato con l’inibitore della 5α-reduttasi.
Nei gruppi trattati con Nandrolone, l’uso di un inibitore della 5α-reduttasi ha parzialmente inibito la conversione del Nandrolone in DHN. Poiché il Nandrolone è molto più androgeno del DHN, l’inibizione di questo processo enzimatico ha mantenuto concentrazioni sieriche più elevate di Nandrolone nei tessuti dove la 5α-reduttasi è maggiormente espressa e la stimolazione ventrale della prostata è aumentata in modo significativo.
La maggior parte degli AAS utilizzati nel BodyBuilding e nel PowerLifting non sono potenti substrati per la 5α-reduttasi o non lo sono affatto. Ciò significa che Finasteride e Dutasteride non aiuteranno assolutamente a prevenire la caduta dei capelli se assunti durante un protocollo contenente AAS come Methandrostenolone, Trenbolone, Oxymetholone, Stanozololo, Drostanolone, Boldenone, Trestolone, Mesterolone, Methenolone, DHB ecc… .
Inoltre, non aiuterà a prevenire la caduta dei capelli con l’uso di SARM non steroidei androgeni come l’S23.
S-23
L’S-23 è un Modulatore Selettivo del Recettore degli Androgeni (SARM) sperimentale sviluppato da GTX come potenziale contraccettivo ormonale maschile. Si lega al recettore degli androgeni in modo più forte rispetto ai farmaci più vecchi come l’Andarina con un Ki di 1,7nM, è possiede una potenziale attività androgena superiore alla maggior parte dei SARM non steroidei.
Alla fine della giostra, l’unico AAS degno di nota che può essere attenuato nei suoi effetti androgeni a livello del cuoio capelluto mediante l’uso di Finasteride o Dutasteride è il Testosterone. E anche con il DHT sostanzialmente inibito con Finasteride, o quasi completamente eliminato con Dutasteride, il Testosterone ha ancora la sua androgenicità intrinseca e atrofizzerà i follicoli piliferi, soltanto in misura molto minore e più lenta.
Questo è il motivo per cui alcuni individui utilizzatori di Dutasteride con alopecia androgenetica aggressiva perdono ancora i capelli anche con zero DHT in circolo.
La soluzione in caso di uso di Nandrolone da parte di soggetti predisposti alla alopecia androgenetica, risulta essere l’uso del RU58841.
RU-58841
Il RU-58841, noto anche come PSK-3841 o HMR-3841, è un antiandrogeno non steroideo (NSAA) inizialmente sviluppato negli anni ’80 da Roussel Uclaf, l’azienda farmaceutica francese da cui ha preso il nome. In precedenza era oggetto di indagine da parte della ProStrakan (precedentemente ProSkelia e Strakan) per un potenziale utilizzo come trattamento topico per condizioni androgeno-dipendenti tra cui acne, perdita di capelli,[2] e crescita eccessiva dei capelli.[3][4][5][6]
Il composto è simile nella struttura all’NSAA RU-58642 ma contiene una catena laterale diversa.[7] Questi composti sono simili nella struttura chimica alla Nilutamide,[8] che è correlata a Flutamide, Bicalutamide ed Enzalutamide, che sono tutte NSAA.[9]
Il RU-58841 può essere sintetizzato sia sintetizzando la porzione idantoina che mediante accoppiamento arilico con 5,5-dimetilidantoina.[10] Il RU-58841 produce Cianonilutamide (RU-56279) e RU-59416 come metaboliti negli animali.[11]
La Cianonilutamide ha un’affinità relativamente bassa per il recettore degli androgeni, ma mostra una significativa attività antiandrogena negli animali.[11] Mentre il RU-59416 ha un’affinità molto bassa per il recettore degli androgeni.[11]
Fortunatamente, sembra che il RU58841 abbia un’affinità di legame equivalente o maggiore di quella del Testosterone per l’AR, quindi in realtà è abbastanza efficace nell’inibire l’attività androgena sia del debole DHN che del Testosterone e delle quantità residue di Nandrolone nel cuoio capelluto.
Conclusioni:
Mi sembra ovvio che il messaggio principale da portarsi a casa sia quello di evitare , la dove si voglia tamponare la perdita di capelli legata alla alopecia androgenetica, l’abbinamento di inibitori della 5α-reduttasi (vedi Finasteride o Dutasteride) quando nel protocollo di supplementazione farmacologica è presente il Nandrolone.
Se si desidera utilizzare un trattamento più efficace per arginare l’alopecia androgenetica durante un ciclo contenente Nandrolone, sarebbe opportuno considerare l’uso di un anti-androgeno topico di forte affinità di legame all’AR come il RU58841.
Battmann T, Bonfils A, Branche C, Humbert J, Goubet F, Teutsch G, Philibert D (January 1994). “RU 58841, a new specific topical antiandrogen: a candidate of choice for the treatment of acne, androgenetic alopecia and hirsutism”. J. Steroid Biochem. Mol. Biol.
Van Dort ME, Jung YW (April 2001). “Synthesis and structure-activity studies of side-chain derivatized arylhydantoins for investigation as androgen receptor radioligands”. Bioorg. Med. Chem. Lett. 11 (8): 1045–7.
Poulos GA, Mirmirani P (February 2005). “Investigational medications in the treatment of alopecia”. Expert Opin Investig Drugs. 14 (2): 177–84.
Leonard, Matthew J.; Lingham, Anthony R.; Niere, Julie O.; Jackson, Neale R. C.; McKay, Peter G.; Hügel, Helmut M. (2014). “Alternative synthesis of the anti-baldness compound RU58841”. RSC Adv. 4 (27): 14143–14148.
Jump up to:abc Cousty-Berlin D, Bergaud B, Bruyant MC, Battmann T, Branche C, Philibert D (October 1994). “Preliminary pharmacokinetics and metabolism of novel non-steroidal antiandrogens in the rat: relation of their systemic activity to the formation of a common metabolite”. J. Steroid Biochem. Mol. Biol.
ATTENZIONE:quanto segue è un articolo volto alla divulgazione scientifica in campo biochimicoe non ha alcuna valenza di consiglio o parere medico favorevole all’uso di sostanza illegali.
Nota introduttiva per il lettore:
Contrariamente a quanto potreste essere indotti a pensare, questo articolo non consiste in una mera successione di eventi che hanno delineato le scoperte scientifiche nel campo dell’endocrinologia e, nella fattispecie, dell’andrologia, piuttosto quello che mi accingo a riportare è una storia dettagliata che permetterà ad ognuno di voi di avere un idea molto più concreta sugli AAS, sul loro potenziale (nel bene o nel male) e sulle loro applicazioni dagli albori fino ad oggi.
“La conoscenza della Verità permette all’uomo lo stato di concreta libertà e di libera scelta”
Gabriel Bellizzi
La storia degli AAS:
Sarà sicuramente difficile da accettare o anche soltanto da immaginare per alcuni di voi, in specie per i puristi della “Hold Scool”, che l’uso di steroidi gonadici è precedente alla loro identificazione e isolamento. Si ritiene che l’estrazione di ormoni dalle urine è iniziata in Cina nel 100 a.C. L’uso medico dell’estratto di testicolo iniziò alla fine del XIX secolo mentre i suoi effetti sulla forza erano ancora in fase di studio.[1] L’isolamento degli steroidi gonadici può essere fatto risalire al 1931, quando Adolf Butenandt, un chimico di Marburg, purificò 15 milligrammi dell’ormone maschile Androstenone da decine di migliaia di litri di urina. Questo steroide fu successivamente sintetizzato nel 1934 da Leopold Ružička, un chimico di Zurigo.[2]
Charles-Édouard Brown-Séquard (8 aprile 1817 – 2 aprile 1894), fisiologo e neurologo mauriziano che, nel 1850, fu il primo a descrivere quella che oggi è chiamata sindrome di Brown-Séquard [3][4], è stato il vero pioniere dello studio e dell’applicazione terapeutica e sportiva degli androgeni.
Charles-Édouard Brown-Séquard
Brown-Séquard era una figura piuttosto controversa ed eccentrica, ed è anche noto per aver autodichiarato, all’età di 72 anni, di essere riuscito ad ottenere “l’abilità sessuale ringiovanita dopo l’iniezione sottocutanea di estratti di testicolo di scimmia”. Migliaia di uomini hanno provato la terapia. L’endocrinologo Robert B. Greenblatt scrisse che questa terapia non avrebbe potuto funzionare perché, a differenza della tiroide, i testicoli non immagazzinano gli ormoni che producono e, quindi, ottenere una dose terapeutica di Testosterone direttamente dalle ghiandole animali “richiederebbe circa una -quarto di tonnellata di testicoli di toro.”[5] Si pensa che la risposta positiva di molti uomini sia stata nulla più che un effetto placebo, ma a quanto pare questo era “sufficiente per mettere in moto il campo dell’endocrinologia”.[6]
Molto più importante è che Brown-Séquard fu uno dei primi a postulare l’esistenza di sostanze, ora note come ormoni, secrete nel flusso sanguigno per influenzare organi distanti. In particolare, dimostrò (nel 1856) che la rimozione delle ghiandole surrenali portava alla morte, per mancanza di ormoni essenziali. Come accennato in precedenza, all’età di 72 anni, in una riunione della Societé de Biologie a Parigi, Brown-Séquard riferì che l’iniezione ipodermica di un fluido preparato dai testicoli di cavie e cani porta al ringiovanimento e al prolungamento della vita umana. Era noto, tra gli scienziati, in modo derisorio, come l’Elixir di Brown-Séquard . Una pubblicazione medica di Vienna scherzò sulle affermazioni dello scienziato in modo sprezzante: “La conferenza deve essere vista come un’ulteriore prova della necessità di mandare in pensione i professori che hanno raggiunto i loro settant’anni”.[7]
La ricerca sugli ormoni di Brown-Séquard incuriosì il futuro lanciatore di baseball della Major League e Hall of Fame “Pud” Galvin. Galvin si chiese se le iniezioni di Elixir di Brown-Séquard avrebbero migliorato le prestazioni nel baseball. Poiché l’estratto conteneva indubbiamente tracce di steroidi androgeni non ancora identificati, Galvin divenne il primo atleta conosciuto a somministrarsi un prodotto a base di steroidi usando l’Elixir in modo regolare. L’anno era il 1889. L’uso di “steroidi” da parte di Galvin precedette di oltre 100 anni il recente scandalo sugli AAS nel baseball. La ricerca di Brown-Séquard ha ispirato diversi scienziati a sviluppare la sua ricerca con estratti testicolari.
Pochi anni dopo che “Pud” Galvin divenne il primo “utilizzatore di steroidi” del baseball, il fisiologo austriaco Oskar Zoth ipotizzò che le iniezioni di estratti testicolari potessero migliorare le prestazioni atletiche. Zoth pubblicò un documento nel 1896 proponendo ulteriori ricerche sulle prestazioni da condurre su atleti. L’idea che una sostanza misteriosa nei testicoli degli animali potesse offrire benefici per il miglioramento delle prestazioni negli atleti si radicò saldamente nella comunità di ricerca da allora. I due decenni successivi hanno visto gli scienziati confermare ripetutamente gli effetti androgeni di vari estratti testicolari.
“Pud” Galvin
Nel 1927, il professore di chimica dell’Università di Chicago Fred Koch e l’assistente di ricerca Lemuel McGee ricavarono 20mg di una sostanza da 40 libbre di testicoli bovini ottenuti dai Chicago Stockyards. L’estratto testicolare ha rimascolinizzato galli, maiali e ratti castrati. Tuttavia, le strutture chimiche di potenti androgeni come il Testosterone non erano ancora state chiarite e identificate.
Tre potenti aziende farmaceutiche sono state fortemente coinvolte nella corsa allo sviluppo di steroidi anabolizzanti. Non sorprende che queste tre società abbiano avuto un effetto lungo e duraturo sulla storia e sullo sviluppo degli AAS che continua fino ad oggi.
Lo sviluppo degli steroidi era un grande business anche negli anni ’30. Grandi aziende farmaceutiche come Organon, Schering e Ciba hanno visto un notevole potenziale in questo mercato emergente. Non sorprende che le aziende che hanno lanciato la rivoluzione degli steroidi continuino ad essere fortemente associate agli steroidi anabolizzanti tra gli atleti moderni.
I chimici che lavorano per queste grandi aziende farmaceutiche hanno cambiato il mondo forse non nei modi che avrebbero potuto immaginare. Diventerebbero “gli dei degli steroidi” negli annali della storia dello sport. Gli atleti avrebbero presto utilizzato le loro creazioni durante i successivi 75 anni!
Organon, Schering e Ciba si sono precipitati per isolare e sintetizzare i potenti ormoni contenuti negli estratti testicolari.
Karoly David ed Ernst Laqueur di Organon (Paesi Bassi) sono stati il primo team farmaceutico a isolare e identificare la struttura chimica del Testosterone quando ne hanno isolato 10mg da 100kg di testicoli di toro. La scoperta del Testosterone fu annunciata per la prima volta nel classico documento intitolato “On Crystalline Male Hormone from Testes (Testosterone): More Active Than Androsterone Preparations from Urine or Cholesterol” il 27 maggio 1935.[8]
Ernst Laqueur (1880-1947)
A questo punto, erano necessarie grandi quantità di testicoli animali per estrarre il testosterone, il che rendeva l’uso del testosterone impraticabile per l’uso commerciale. Tuttavia, i team di ricerca concorrenti erano a pochi mesi dalla pubblicazione di metodi più efficienti per sintetizzare il Testosterone.
Quindi, negli anni ’30, era già noto che i testicoli contengono un androgeno più potente dell’Androstenone e tre gruppi di scienziati, finanziati da aziende farmaceutiche concorrenti nei Paesi Bassi, in Germania e in Svizzera (prima citate), si sono adoperati per isolarlo.[9][10]
I ricercatori chiamarono l’ormone Testosterone, dai testicolo e sterolo e il suffisso di chetone. La sintesi chimica del Testosterone fu raggiunta nell’agosto di quell’anno, quando Butenandt e G. Hanisch pubblicarono un articolo che descriveva “Un metodo per preparare il Testosterone dal colesterolo”.[11] Solo una settimana dopo, il terzo gruppo, Ruzicka e A. Wettstein, annunciò una domanda di brevetto in un documento “Sulla preparazione artificiale dell’ormone testicolare Testosterone (Androsten-3-one-17-ol).”[12] Ruzicka e Butenandt ricevettero il premio Nobel per la chimica nel 1939 per il loro lavoro, ma il Il governo nazional-socialista costrinse Butenandt a declinare l’onore, sebbene accettò il premio dopo la fine della seconda guerra mondiale.[9][10]
A proposito di Ruzicka, egli nel 1934, sintetizzò l’ormone maschile Androsterone e dimostrò anche “la sua relazione costituzionale e configurazionale con gli steroli”, cosa che permise il passaggio molto più economici di sintesi dal Colesterolo agli steroli vegetali. Nel periodo 1934-1939 Ruzicka aveva pubblicato 70 articoli nel campo degli ormoni sessuali steroidei importanti dal punto di vista medico e aveva depositato diverse dozzine di brevetti.[13]
Leopold Ružička nel 1939
Gli studi clinici sull’uomo, che prevedevano dosi PO (per via orale) di Methyltestosterone o iniezioni di Testosterone Propionato, iniziarono già nel 1937.[14] Il Testosterone Propionato è menzionato in una lettera all’editore della rivista Strength and Health nel 1938; questo è il primo riferimento noto a un AAS in una rivista statunitense di sollevamento pesi o bodybuilding.[14] Vengono spesso riportate voci secondo cui ai soldati tedeschi furono somministrati AAS durante la seconda guerra mondiale, con l’obiettivo di aumentare la loro aggressività e resistenza, ma queste sono ancora non provate.[15] Lo stesso Adolf Hitler, secondo il suo medico, è stato trattato con derivati del Testosterone per il trattamento di vari disturbi.[16] Gli AAS sono stati usati in esperimenti condotti dai nazional-socialisti sui detenuti dei campi di concentramento,[16] e in seguito dagli alleati nel tentativo di curare le vittime malnutrite sopravvissute ai campi di prigionia tedeschi.[15] Al presidente John F. Kennedy furono somministrati steroidi sia prima che durante la sua presidenza.[17]
Lo sviluppo delle proprietà di costruzione muscolare del Testosterone proseguì negli anni ’40, in Unione Sovietica e nei paesi del blocco orientale come la Germania dell’Est, dove sono stati utilizzati programmi di AAS per migliorare le prestazioni dei sollevatori di pesi olimpici e di altri dilettanti. In risposta al successo dei sollevatori di pesi russi, il medico della squadra olimpica statunitense John Ziegler lavorò con un equipe di chimici per sviluppare un AAS con effetti androgeni ridotti.[202] Il lavoro di Ziegler portò alla sintesi di Methandrostenolone, che la Ciba Pharmaceuticals commercializzò sotto il nome Dianabol. Il nuovo steroide venne approvato per l’uso negli Stati Uniti dalla Food and Drug Administration (FDA) nel 1958. È stato più comunemente somministrato a vittime di ustioni e anziani. Gli utilizzatori off-label del farmaco erano per lo più culturisti e sollevatori di pesi. Sebbene Ziegler prescrivesse solo piccole dosi agli atleti, scoprì presto che coloro che avevano abusato del Dianabol soffrivano di prostata ingrossata e atrofia testicolare.[18] Gli AAS sono stati inseriti nell’elenco delle sostanze vietate del Comitato Olimpico Internazionale (CIO) nel 1976, e un decennio dopo il comitato ha introdotto i test antidoping “fuori gara” perché molti atleti utilizzavano gli AAS nel loro periodo di allenamento piuttosto che durante la competizione.[19]
Il Dr. John Ziegler
Tre idee principali hanno governato le modifiche del Testosterone in una moltitudine di AAS: l’alchilazione in posizione C17α con il gruppo metile o etilico che ha creato composti POly attivi perché rallenta la degradazione del farmaco da parte del fegato; l’esterificazione del Testosterone e del Nortestosterone in posizione C17β consente la somministrazione della sostanza per via parenterale e ne aumenta la durata dell’efficacia in quanto agenti solubili in liquidi oleosi possono essere presenti nell’organismo per diversi mesi; e le alterazioni della struttura ad anello sono state applicate sia per AAS orali che per agenti parenterali per cercare di ottenere diversi rapporti di effetto anabolico-androgeno.[19]
Nel 1945, lo scrittore Paul de Kruif celebrò le proprietà anaboliche del Testosterone, quindi della forma esterificata Testosterone Propionato e di quella metilata in C17 Methytestosterone nel libro intitolato “The Male Hormone”. Si dice che questo libro ampiamente letto abbia contribuito a rendere popolare il potenziale del Testosterone (e degli altri futuri steroidi anabolizzanti) per aumentare la massa muscolare tra i bodybuilder della costa occidentale alla fine degli anni ’40 e all’inizio degli anni ’50. Questo è stato solo l’inizio del fascino del Bodybuilding per gli steroidi anabolizzanti.
La comunità del bodybuilding nel suo insieme avrebbe presto iniziato a sperimentare ampiamente con gli AAS negli anni ’50 e sarebbe diventata pioniera nell’uso degli steroidi. Rimarranno all’avanguardia dei farmaci per l’aumento delle prestazioni fino al presente secolo.
L’IFBB Mr. Olympia Larry Scott ha ammesso che lui, e praticamente tutti i migliori bodybuilder competitivi, usavano anche steroidi anabolizzanti nel 1960.
Larry Scott
La Searle avviò uno sforzo senza precedenti nella ricerca sugli steroidi al fine di scoprire ormoni steroidei sintetici superiori per l’uso in medicina. Tra il 1948 e il 1955, i chimici della Searle avevano sintetizzato più di mille diversi derivati e analoghi del Testosterone con l’obiettivo specifico di creare uno steroide anabolizzante attivo per via orale con effetti collaterali androgeni minimi. L’obbiettivo della Searle era quello di creare steroidi che evitassero qualsiasi effetto virilizzante.
Dei mille potenziali profili di steroidi creati da Searle durante questo periodo, il Nilevar (Noretandrolone) fu il candidato vincente selezionato per la commercializzazione. Il chimico della Searle Frank Colton sintetizzò il Noretandrolone nel 1953.
Il Noretandrolone divenne il primo steroide anabolizzante sintetico attivo per via orale quando venne approvato dalla Food and Drug Administration (FDA) con il marchio Nilevar nel 1956. L’unico altro androgeno attivo per via orale disponibile all’epoca era il Methytestosterone che era semplicemente un versione metilata in C17 del Testosterone per aumentarne principalmente (ma non solo) la biodisponibilità orale.
Nel 1958, il bodybuilder della West Coast e campione del Mr. Universe Bill Pearl è stato uno dei primi bodybuilder a sperimentare il nuovo steroide anabolizzante creato da Searle. Pearl ha fatto un ciclo di 12 settimane usando 30mg di Noretandrolone aumentando il suo peso corporeo di 25 libbre da 225 a 250 libbre.
Bill Pearl nel 1958
Bill Pearl ha ammesso apertamente di aver usato steroidi anabolizzanti in preparazione per il concorso Mr. Universe della National Amateur BodyBuilders Association (NABBA) del 1961. Ha rivelato che l’uso di steroidi non era più una pratica clandestina tra i migliori bodybuilder, confermando la valutazione del Mr. Olympia Larry Scott sul ruolo degli steroidi nella scena del bodybuilding.
La GD Searle non è stata l’unica azienda farmaceutica a spendere enormi risorse nello sviluppo di nuovi steroidi anabolizzanti sintetici. Diverse grandi aziende farmaceutiche sono impazzite nel creare steroidi anabolizzanti negli anni ’50 e nei primi anni ’60.
Tra il 1950 e il 1965 sono stati sviluppati praticamente tutti i più popolari steroidi attualmente utilizzati. Questi includono ma non sono limitati a: Methandrostenolone, Oxymetholone, Oxandrolone, Stanozololo, Fluoxymesterone, Boldenone, Nandrolone, Metenolone, Clorodeidromethyltestosterone, Drostanolone e Trenbolone.
Anche alcuni degli steroidi più “esotici” utilizzati dai futuri bodybuilder sono stati sviluppati durante questo periodo come il Furazabol, Formebolone, Oxymesterone, Mibolerone, Stenbolone e Etilestrenolo.
La Organon creò alcuni steroidi iniettabili incredibilmente popolari durante questo periodo, molti dei quali sono ancora ampiamente utilizzati da culturisti e atleti. La Organon sarà per sempre legata agli steroidi anabolizzanti a causa del rilascio del Durabolin e in particolare del Deca Durabolin.
La Organon ha rilasciato il Durabolin (Nandrolone Fenilpropionato) nel 1957 che divenne molto popolare. La sua popolarità è stata presto eclissata quando la Organon rilasciò il Deca Durabolin nel 1962 oltre un decennio dopo la sintesi del Nandrolone Decanoato.
Il Deca Durabolin alla fine è diventato uno degli steroidi più popolari di tutti i tempi nella storia del miglioramento delle prestazioni insieme a Dianabol, Anadrol, Anavar e Winstrol.
Russell Marker ha lasciato il segno nell’industria degli steroidi con la fondazione della Syntex. I successori di Marker alla Syntex hanno continuato la sua ricerca sugli steroidi e hanno rilasciato l’Anadrol (Oxymetholone) nel 1959 dopo che era stato sintetizzato da Howard Ringold e George Rosenkranz. Rosenkranz e Ringold avevano sintetizzato il Masteron (Drostanolone Acetato) per la Syntex un paio di anni prima.
Russell Marker
Molte delle creazioni di Ringold non sono mai state introdotte commercialmente dalla Syntex. Tuttavia, almeno uno dei suoi steroidi anabolizzanti accantonati – Methyldrostanolone o Methasteron – sarebbe stato commercializzato come un “integratore alimentare” di grande successo chiamato “Superdrol” durante l’era dei supplementi pro-ormonali inaugurato dalla Dietary Health and Supplement Education Act del 1994.
La Syntex non solo ha sintetizzato l’Anadrol, ma ha fornito l’ispirazione alla Winthrop Laboratories per sintetizzare lo Stanozololo. Il chimico Winthrop è stato in grado di sintetizzare lo Stanozololo dall’Oxymetholone nel 1959. Lo Stanozololo è stato commercializzato come Winstrol e Winstrol Depot negli Stati Uniti nel 1962.
Fortunatamente, Julius Vida ha compilato i risultati pubblicati di circa 650 steroidi androgeni anabolizzanti scoperti fino al 1967 nel suo libro di testo del 1969 “Androgens and Anabolic Agents: Chemistry and Pharmacology”. Questo in seguito divenne una guida di riferimento inestimabile, non solo per le aziende di nutrizione sportiva, ma anche per i chimici rinnegati alla ricerca di AAS efficaci, non rilevabili e che aggirassero le leggi di controllo. Divenne una miniera d’oro di informazioni per gli “imprenditori” degli integratori alla fine degli anni ’90.
Quindi, alcuni degli steroidi anabolizzanti scoperti durante il periodo 50′-60′ sono stati successivamente reintrodotti, non come prodotti farmaceutici e non come “integratori alimentari”, ma come steroidi non rilevabili utilizzati per eludere i protocolli antidoping negli sport.
Ad esempio, Patrick Arnold ha utilizzato il Norbolotone, sviluppato nel 1963 da Wyeth, per aiutare alcuni atleti a raggiungere questo obiettivo. Il Methyltrienolone è un altro di questi steroidi che non era rilevabile fino ad un certo periodo negli sport. A Patrick Arnold si deve la sintesi del Tetraidrogestrinone (THG) partendo dalla struttura del Methyltrienolone.
Patrick Arnold “The Father of Prohormones”, il creatore del Tetraidrogestrinone (THG)
Ma adesso andiamo ad immergerci nella chimica degli AAS…
La chimica degli Steroidi Anabolizzanti Androgeni:
Esistono moltissimi AAS che differiscono l’uno dall’altro in vari modi (sottili). Queste differenze, in sostanza, si riducono alle differenze sottostanti nella struttura chimica. In questa sezione tratterò alcune alterazioni chimiche che possono essere trovate nei comuni steroidi anabolizzanti. Tratterò anche brevemente cosa sono gli esteri. Sebbene non siano un’alterazione chimica della molecola madre di per sé, influenzano il “comportamento” di uno steroide anabolizzante indipendentemente dal suo effetto sulla velocità di rilascio dall’olio nel corpo.
Cos’è un estere?
La parola estere viene spesso usata quando si parla di steroidi anabolizzanti. E la maggior parte delle persone sa che è qualcosa che influenza l’emivita dello steroide anabolizzante. Ma cos’è in realtà?
Se hai una molecola, come il Testosterone, puoi legare qualcosa a questa molecola madre per influenzarne le sue proprietà biofisiche. Nel caso dell’esterificazione di uno steroide anabolizzante, si riduce al legame di un gruppo di acido carbossilico sul carbonio 17 dello scheletro carbossilico dello steroide come illustrato di seguito:
Il legame di questo gruppo influisce sulla polarità della molecola. E la polarità di una molecola si riferisce a come una carica è distribuita su di essa. Una carica su una molecola porta all’interazione con le molecole cariche circostanti. Ciò è di particolare rilevanza per quanto riguarda l’acqua. L’acqua è piuttosto polare e quindi le molecole che sono anche abbastanza polari si dissolvono facilmente in essa. Le molecole polari sono idrofile.
I lipidi, al contrario, sono apolari, o solo leggermente polari. Di conseguenza, hanno difficoltà a dissolversi in acqua. Ad esempio, se mettete dell’olio d’oliva in un bicchiere d’acqua, potete vedere come l’olio si attacca e forma uno strato sull’acqua. Non si sta dissolvendo. Questo è il risultato della mancanza di polarità dell’olio d’oliva. Gli oli sono quindi idrofobici.
E qui arriva il nocciolo della questione: le molecole non polari (o solo leggermente polari) si dissolvono facilmente nell’olio. Quando un estere è legato a una molecola steroidea, ne ridurrà la polarità e quindi lo renderà più facilmente solubile in olio e meno facilmente solubile in acqua. O in altre parole: diventa meno idrofilo (“amante dell’acqua”) e più lipofilo (“amante dell’olio”). Gli AAS esterificati sono quindi lipofili.
Quindi perchè è importante? Come forse saprete, gli steroidi esterificati hanno un’emivita più lunga rispetto alle loro controparti non esterificate. L’esterificazione ha il potenziale per estendere notevolmente l’emivita. Ad esempio, il Testosterone Undecanoato ha dimostrato di avere un’emivita di 70 giorni (!) in un piccolo gruppo di uomini ipogonadici.[20] Questa lunga emivita può essere direttamente ricondotta al grande effetto che l’Undecanoato ha sulla polarità della molecola madre. La diminuisce notevolmente. Quindi quello che succede è quanto segue. Il Testosterone esterificato (o qualsiasi altro tipo di steroide) viene solubilizzato nell’olio. Questo olio viene quindi iniettato per via intramuscolare in cui si diffonderà per tutta la lunghezza dei fasci muscolari. Questo appare come segue nell’immagine (notare come l’olio si diffonde lungo la lunghezza dei fasci muscolari):
Immagine presa da Kalicharan [21].
Il Testosterone esterificato quindi, in una certa misura, si diffonderà fuori dal deposito oleoso e si dissolverà nel tessuto circostante, che è costituito principalmente da acqua. La velocità con cui ciò accade è determinata dalla sua polarità. Se è molto apolare (e quindi lipofilo) si diffonderà molto molto lentamente dall’olio nel tessuto circostante. Dopotutto: le sostanze non polari sono lipofile e non idrofile. Quindi l’esterificazione determina l’emivita della molecola iniettata.
Una grande regola empirica è che, maggiore è la lunghezza della catena di carbonio dell’estere, maggiore sarà l’emivita del composto. Perché più lunga è la catena, più diminuirà la polarità della molecola. Oltre alla lunghezza dell’acido carbossilico, puoi anche cambiarne la struttura per influenzare la polarità. Ad esempio, questo è evidente con l’estere che vedete legato al Parabolan: Trenbolone Hexahydrobenzylcarbonate. Qui puoi trovare un gruppo cicloesano. Nell’immagine è stato evidenziato il gruppo cicloesano in blu:
Trenbolone Hexahydrobenzylcarbonate
Ad ogni modo, questo è anche il motivo per cui il Testosterone Enantato (6 atomi di carbonio) ha un’emivita significativamente più breve rispetto al Testosterone Undecanoato (11 atomi di carbonio) che ho menzionato precedentemente. Una volta che lo steroide esterificato raggiunge il flusso sanguigno, l’estere viene rapidamente idrolizzato dalle esterasi nel plasma, “liberando” la molecola madre che potrà svolgere la sua attività tissutale.[22]
In sostanza, le differenze negli effetti biologici dei vari steroidi anabolizzanti sono il risultato delle loro differenze nella struttura chimica. Esaminerò alcune di queste differenze strutturali e le loro conseguenze.
Il gruppo C-3-chetone:
Se guardate sopra l’immagine del nucleo steroideo, potete vedere il terzo atomo di carbonio in basso a sinistra. Fa parte dell’anello A ed è estremamente importante nel legame con il recettore degli androgeni (AR). Il gruppo C-3-chetone, come si trova nel Testosterone, ha questo aspetto (colorato in rosso):
È un atomo di ossigeno legato al terzo atomo di carbonio da un doppio legame. L’importanza di questo gruppo per il legame con il recettore è stata evidenziata da esperimenti di Ojasoo et al. [23]. Nel loro articolo “Towards the mapping of the progesterone and androgen receptors” hanno determinato le affinità di legame relative di 33 ligandi steroidei per il recettore degli androgeni. Ciò includeva l’androgeno estremamente potente R1881 (Methyltrienolone) e il suo “fratello chimico” R3773, a cui manca il gruppo chetonico al carbonio 3. Mentre l’R1881 ha dimostrato un’affinità di legame molto elevata per il recettore degli androgeni e del progesterone e persino una notevole affinità di legame per il recettore dei glucocorticoidi, R3773 ha mostrato la completa mancanza di qualsiasi affinità di legame come risultato di questa singola modifica strutturale.
In effetti, vediamo succedere qualcosa di simile quando il gruppo C-3-chetone viene ridotto. Cioè, l’atomo di ossigeno legato con un doppio legame diventa un gruppo ossidrile (-OH). Prendete ad esempio il Diidrotestosterone (DHT). Può essere ridotto in posizione C-3 per produrre 3α- o 3β-androstanediolo. Questa reazione è catalizzata rispettivamente dal 3α-HSD o dal 3β-HSD. Entrambi questi metaboliti ridotti del DHT hanno un’affinità di gran lunga inferiore per il recettore degli androgeni [23]. Questa alterazione appare come segue quando si osserva la struttura chimica:
Ora posso spiegare la differenza tra qualcosa che è α-orientata e qualcosa che è β-orientata. Qualcosa che è α-orientato è illustrato usando un cuneo tratteggiato, mentre qualcosa che è β-orientato è illustrato usando un cuneo solido. Dovreste vedere un cuneo tratteggiato come entrare nella carta, mentre un cuneo solido sta uscendo dalla carta. La vera molecola rappresentata dai nostri disegni bidimensionali è, in realtà, tridimensionale. Quindi disegnandolo con cunei tratteggiati e solidi, siamo in grado di incorporare la struttura tridimensionale di una molecola nei nostri disegni bidimensionali.
Ad ogni modo, questa riduzione diminuisce significativamente la sua affinità di legame per il recettore degli androgeni e questo è un processo che accade attivamente nel muscolo scheletrico. In effetti, l’iniezione endovenosa di DHT marcato ha mostrato una grande lisi (73,8 – 81,9%) entro 20-60 minuti nel muscolo scheletrico rispetto al Testosterone (20,2 – 30,5%) [24]. Il principale metabolita del DHT era il 3α-androstanediolo. Questa è la probabile spiegazione del perché, nonostante la sua affinità di legame molte volte superiore per l’AR, il DHT mostra effetti di costruzione muscolare simili a quelli del Testosterone con esposizione equimolare.
Torniamo al punto in cui eravamo: il gruppo C-3-chetone. Cosa lo rende così speciale per il legame? Il motivo è perché l’atomo di ossigeno del cheto gruppo ha una coppia solitaria di elettroni. Come tale, può funzionare come accettore di legami idrogeno per formare un legame idrogeno (legame H). E sembrano esserci due residui del recettore degli androgeni che si trovano nel suo dominio legante il ligando (LBD) che lo fanno [25]. Il LBD è la parte del recettore degli androgeni che interagisce direttamente con un ligando, come il Testosterone. È il sito del recettore degli androgeni a cui si legano gli steroidi anabolizzanti. Proprio come una chiave può inserirsi in una serratura, un ligando si inserisce nella LBD (o, dovrei dire, tasca legante il ligando) di un recettore. Senza il gruppo C-3-chetone, il ligando semplicemente non si adatta bene, poiché i legami H che mantengono il ligando nel posto giusto del LBD, non possono essere formati. Di norma, la mancanza di un gruppo chetone in C-3 rende uno steroide una chiave difettosa per la sua serratura.
Ma che dire dello Stanozololo? In effetti, lo Stanozololo è una molecola strana in quanto ha un anello pirazolico fuso con l’anello A. Per riferimento, ecco come appare e, come potete vedere, non c’è ovviamente nessun gruppo chetone in C-3:
Tuttavia, si presume che il secondo atomo di azoto formi il legame idrogeno in questo caso [26]. (Tuttavia, l’affinità è ancora significativamente inferiore.)
Il gruppo C-17β-idrossile e C-17α-alchilato:
Mentre il gruppo C-3-chetone può fungere da accettore di legami idrogeno, il gruppo C-17β-idrossile può fungere da donatore di legami idrogeno. Due residui nel LBD dell’AR possono fungere da corrispondente accettore di legami idrogeno per formare un legame H [25]. E mentre il gruppo C-3-chetone (=O) potrebbe essere ridotto a un gruppo ossidrile (-OH), il gruppo C-17 (-OH) potrebbe essere ossidato per formare un gruppo chetonico (=O). Questa reazione è catalizzata da una classe di enzimi denominata 17β-HSD [27]. Questa reazione è reversibile. Esistono diversi enzimi di questa classe, ciascuno dei quali catalizza principalmente una reazione unidirezionale. Cioè, alcuni catalizzano la reazione di ossidazione, formando il gruppo chetonico (=O), mentre altri catalizzano la reazione di riduzione, formando il gruppo ossidrile (-OH).
Proprio come con la riduzione o l’eliminazione del gruppo C-3-chetone, l’ossidazione o l’eliminazione del gruppo C-17β-idrossile riduce l’affinità di legame per l’AR. La reazione di ossidazione, ampiamente, avviene nel fegato. Questo è molto probabilmente uno dei motivi principali per cui gli steroidi anabolizzanti hanno una biodisponibilità orale così bassa. Ad esempio, solo il 3,66% del Testosterone somministrato per via orale è diventato sistematicamente disponibile in un esperimento [28]. Una volta che passa nel fegato, già una quantità notevole sarà metabolizzata. Per contrastare ciò, potrebbe essere legato ad un gruppo C-17α-alchile (comunemente un gruppo metilico). Si pensa che l’aggiunta di questo gruppo ostacoli questa ossidazione per impedimento sterico. O, in altre parole, il gruppo metilico impedisce all’enzima di agire correttamente sulla molecola e catalizzare la reazione.
Inoltre, di conseguenza, gli steroidi che possiedono questo gruppo metilico hanno un’emivita prolungata di diverse ore rispetto alle loro controparti non metilate. (Si noti che la lunga emivita degli steroidi anabolizzanti esterificati non è il risultato di un metabolismo rallentato, ma di una diffusione ritardata fuori dal deposito oleoso.) Questa resistenza contro il metabolismo epatico è probabilmente anche il motivo per cui sono intrinsecamente epatotossici [29] . Il gruppo metilico C-17α, in combinazione con diverse altre modifiche strutturali, consente anche allo steroide di legarsi fortemente a diversi recettori nucleari. Ciò è molto evidente con il composto C-17α-metilato Methyltrienolone, che dimostra un’affinità molto forte per il recettore degli androgeni [23], ma ha anche un’affinità significativa per il recettore dei glucocorticoidi [23], il recettore del progesterone [23, 30] e i recettori mineralcorticoidi [31].
Il doppio legame C-4,5:
La differenza tra Testosterone e Diidrotestosterone (DHT) è data dall’aggiunta di un atomo di idrogeno α-orientato in C-5 e, di conseguenza, la conversione del doppio legame C-4-C-5 in un singolo legame. Questa è una reazione catalizzata dalla classe di enzimi nota come 5α-reduttasi. Allo stesso modo, la 5β-reduttasi aggiunge un atomo di idrogeno β-orientato in C-5.
La riduzione di C-5α di un composto può influenzarne l’affinità di legame per il recettore degli androgeni per il metabolita risultante. Nel caso del Testosterone, aumenta l’affinità di legame. (La riduzione del C-5α nello scheletro carbossilico del Testosterone produce DHT, che ha un’affinità significativamente maggiore.) Il risultato di ciò è che gli effetti del Testosterone possono essere amplificati nei tessuti che esprimono la 5α-reduttasi, come il cuoio capelluto. Ecco perché un’importante modalità di trattamento dell’alopecia androgenetica è l’inibizione di questo enzima (per esempio, la Finasteride è un inibitore della 5α-reduttasi).
Tuttavia, quando il Nandrolone subisce una riduzione in C-5α, forma Diidronandrolone (DHN): un composto con affinità notevolmente inferiore per il recettore degli androgeni [32]. Di conseguenza, le azioni del Nandrolone possono essere smorzate nei tessuti che esprimono la 5α-reduttasi. In generale, la riduzione in C-5α appare rilevante solo per Testosterone e Nandrolone, poiché gli AAS più comunemente usati sono già ridotti in C-5α o non subiscono una significativa riduzione in C-5α nel corpo.
Una delle funzioni della 5β-reduttasi nel corpo umano è quella di disattivare il Testosterone e trasformarlo in metaboliti con quasi nessuna affinità per il recettore degli androgeni. Insieme alla 5α-reduttasi, questi enzimi catalizzano quella che si pensa sia la fase iniziale e limitante del metabolismo degli androgeni con un gruppo C-3-chetone e un doppio legame tra C-4 e C-5, ovvero i 3-cheto- Androgeni 4-eni come il Testosterone [33]. La 5β-reduttasi è prevalentemente espressa nel fegato [34]. Infine, mentre il DHT ha un’affinità molto elevata per il recettore degli androgeni, il suo fratello C-5β ridotto ha una affinità per esso minima [23].
Rimozione in C-19 (“19-Nor”):
Se si rimuove il gruppo C-19-metile dalla molecola di Testosterone, si otterrà il Nandrolone (noto anche come 19-nortestosterone). Una delle conseguenze di ciò, come descritto sopra, è che la successiva riduzione in C-5α produce un androgeno meno potente (DHN) invece di un androgeno più potente, come nel caso del Testosterone (vale a dire DHT). Inoltre, la rimozione di questo gruppo metilico sembra influenzare il tasso di aromatizzazione. Uno studio sui microsomi placentari umani ha osservato un’attività relativa di aromatizzazione che era solo il 20% di quella del Testosterone [35]. La soppressione delle gonadotropine si basa in parte sull’aromatizzazione di un androgeno nell’estrogeno che causa la soppressione a livello dell’ipofisi [36]. In effetti, la somministrazione di 200mg di Nandrolone Esilossifenilprionato ogni 3 settimane per diversi mesi non è stata in grado di sopprimere completamente le gonadotropine e il Testosterone [37]. Ciò è in linea con la premessa che si verifica relativamente poca aromatizzazione del Nandrolone. Infine, questa alterazione chimica conferisce al composto una significativa affinità per il recettore del progesterone [23, 30], ed è questo fattore che ha mostrato il vero potenziale di soppressione del HPTA, maggiormente con iniezioni settimanali.
Un altro noto steroide anabolizzante a cui manca il gruppo C-19-metile è il Trenbolone. Simile al Nandrolone, dimostra anche un’affinità significativa per il recettore del progesterone. Tuttavia, contrariamente al Nandrolone, per il composto non sono stati rilevati metaboliti ridotti di C-5α/β [33]. Pertanto, le altre due alterazioni chimiche nel Trenbolone rispetto al Nandrolone sembrano impedirlo (l’aggiunta di doppi legami negli anelli B e C). Questo sembra anche impedire l’aromatizzazione dell’anello A, dimostrando un’attività estrogenica molto bassa anche a concentrazioni estremamente elevate [38].
Adesso che abbiamo sufficienti conoscenze sulle principali caratteristiche chimiche degli AAS, passiamo ad un argomento che unisce la teoria alla potenziale pratica….
I tre gruppi di AAS: Testosterone, DHT, Nandrolone e loro principali derivati.
In generale, gli steroidi anabolizzanti più comunemente usati rientrano in una delle tre diverse famiglie/categorie: Testosterone e suoi derivati, Diidrotestosterone (DHT) e suoi derivati, e Nandrolone (19-Nortestosterone) e suoi derivati.
Presumibilmente, molti degli steroidi anabolizzanti in ogni gruppo hanno attributi simili tra loro.
Ci sono eccezioni a questo in ogni gruppo, ma in generale la suddivisione degli steroidi anabolizzanti nei tre gruppi fornisce un quadro semplificato per identificare l’azione mirata di ciascun composto, nonché il modo in cui esso influisce sulla costruzione muscolare e sulle prestazioni complessive.
Ci sono innumerevoli processi nel corpo che vengono potenziati o inibiti dagli steroidi anabolizzanti. Per evitare di complicare questa sezione, fornirò una panoramica dei principali meccanismi che direzionano il miglioramento delle prestazioni atletiche o la costruzione muscolare tramite AAS esogeni.
Quando si sceglie quali agenti anabolizzanti sono più favorevoli agli obiettivi di un atleta, dovrebbero essere presi in considerazione i seguenti meccanismi e dovrebbero essere favoriti i composti che sono maggiormente attivi verso gli effetti desiderati.
Attività anabolica esplicata tramite l’attivazione del recettore degli androgeni o per via non-genomica, l’aromatizzazione, la 5α riduzione o la conversione a valle in metaboliti che facilitano l’aumento delle dimensioni e della forza muscolare
Antagonismo dei recettori dei glucocorticoidi che porta all’inibizione della degradazione proteica e ad un aumento netto delle dimensioni e della forza muscolare attraverso il risparmio delle proteine muscolari
Effetti psicoattivi nel cervello che portano a un allenamento più spinto e successivi aumenti delle dimensioni muscolari
Stimolazione della produzione di eritropoietina (EPO) che porta a una migliore prestazione aerobica (con limite prestativo intorno al 53%).
Quando si inizia a scoprire come i diversi steroidi anabolizzanti influiscono sulle diverse funzioni del corpo al di fuori della comprensione di base minimale “farmaci = risultati”, può diventare alquanto complicato decidere quale sia la scelta di protocollo ottimale per i propri o altrui obiettivi.
Ad un livello più alto, anche una volta che in genere si comprende quali scelte composte sarebbero “sagge” solo in base agli obiettivi, ci si rende presto conto che bisogna anche considerare la tolleranza agli effetti collaterali con composti specifici, l’attuale stato di salute, l’età, la storia di uso dei PEDs e una miriade di altri fattori.
E, ovviamente, la corretta selezione dei composti per massimizzare le prestazioni varia ampiamente tra i diversi sport. Ad esempio, un protocollo ottimizzato per un bodybuilder sarà probabilmente completamente diverso da un protocollo ottimizzato per un combattente di MMA.
Questo articolo potrà servire anche come introduzione generale a come ciascuna categoria di AAS possa essere sfruttata, nonché l’approccio generale potenzialmente adottabile verso la selezione dei composti.
Ma andiamo con ordine…
Il Testosterone è l’ormone genitore di una cospicua parte di AAS ed è anche tecnicamente l’ormone genitore dell’intero macrogruppo degli steroidi anabolizzanti perché il DHT e il Nandrolone sono entrambi derivati dal Testosterone.
I derivati del Testosterone più importanti utilizzati per il miglioramento delle prestazioni sono i seguenti:
Testosterone;
Boldenone (Equipoise);
Methandrostenolone (Dianabol);
Fluoxymesterone (Halotestin);
4-Chlorodehydromethyltestosterone (Turinabol).
I tre composti di questo gruppo più comunemente usati in un contesto di Bodybuilding sono Testosterone, Boldenone (Equipoise) e Methandrostenolone (Dianabol).
Questi tre composti sono caratterizzati principalmente dai loro effetti ad ampio spettro sulle funzioni anaboliche e androgeno dipendenti, nonché dalla loro interazione con l’enzima Aromatasi.
Hanno tutti una forte influenza sulla conta degli eritrociti (globuli rossi), sui sistemi energetici e mostrano una serie di interazioni sistemiche ibride analoghe a quelle osservate con la steroidogenesi endogena nella regolazione dell’attività omeostatica.
Sono anabolizzanti certo, ma non così marcatamente.
In generale, questa classe di composti facilita ad un livello intermedio la crescita muscolare, con effetti neurologici e potenziamento dei meccanismi aerobici.
Il Testosterone e il Methandrostenolone sono entrambi substrati soggetti all’azione dell’Aromatasi e possono essere metabolizzati enzimaticamente in potenti estrogeni.
Il Testosterone può essere metabolizzato in Estradiolo (E2) e il Methandrostenolone può essere metabolizzato in Methylestradiolo (17α-methylestradiolo).
Il livello di estrogenicità del Boldenone, come tratto in un mio recente articolo dedicato, è meno chiaro in quanto non sono disponibili dati certi che forniscano dettagli sulla sua interazione con l’Aromatasi nell’uomo.
William Llewellyn ha affermato nel suo libro “Anabolics” che il Boldenone è un substrato meno soggetto all’attività dell’Aromatasi ed è metabolizzato enzimaticamente in estrogeni al 50% del tasso del Testosterone. Questa affermazione non è supportata in nessuno dei dati clinici che ho visionato personalmente, quindi sarei riluttante a presumere che il Boldenone possa facilitare un’attivazione sufficiente del recettore degli estrogeni nell’applicazione pratica come base di un ciclo.
Si ipotizza anche se alcuni metaboliti del Boldenone agiscano o meno come inibitori dell’aromatasi e attenuino l’attività estrogenica dell’ormone progenitore. Sebbene ciò sia certamente possibile e meriti ulteriori ricerche prima di poter fare affermazioni conclusive sul Boldenone, in generale con le nostre conoscenze attuali è abbastanza ben accettato nella comunità che il Boldenone sia leggermente estrogenico, ma significativamente meno del Testosterone e del Methandrostenolone.
Il Fluoxymesterone e il 4-Chlorodehydromethyltestosterone non sono substrati soggetti all’Aromatasi, quindi Testosterone e Methandrostenolone sono gli unici composti in questo gruppo di AAS che possono ricoprire il ruolo di “Base di prova” ed essere utilizzati come base di un ciclo di AAS, l’eccezione riguarda ovviamente il Boldenone per le ragioni già riportate nell’articolo dedicato e accennate pocanzi.
A differenza dei tre principali ormoni utilizzati tra i bodybuilder di questo gruppo di AAS, Fluoxymesterone e il 4-Chlorodehydromethyltestosterone sono derivati del Testosterone che inducono effetti analoghi ai potenti derivati del DHT.
Questi due anabolizzanti sono stati progettati chimicamente per non avere attività estrogenica e per amplificarne la capacità di indurre guadagni significativi di massa, ed essere maggiormente orientati verso l’espressione della sintesi proteica pura e lo stimolo neurologico.
Il successivo gruppo di AAS è rappresentato dal Diidrotestosterone (DHT) ed i suoi derivati.
Il DHT è il metabolita 5α ridotto del Testosterone che facilita la differenziazione sessuale dei genitali maschili durante l’embriogenesi e guida la maturazione sessuale durante la pubertà. È molto più androgeno del Testosterone e durante la pubertà è necessaria una quantità sufficiente di esso nei tessuti dipendenti dagli androgeni per garantire la piena maturazione sessuale.
Un malinteso comune è che gli steroidi derivati dal DHT devono essere estremamente androgeni semplicemente perché sono derivati dal sopra citato androgeno. Al contrario, i derivati del DHT sono quasi tutti più selettivi dei tessuti rispetto al Testosterone.
In generale, tutti gli steroidi anabolizzanti sono stati progettati con quell’obiettivo esatto in mente, non solo i derivati DHT.
Le poco affidabili Androgeno:Anabolico ratio sono state valutate in modelli animali preclinici prima di passare all’uso umano in un ambiente clinico.
Se uno steroide non avesse mostrato una anabolico/androgeno ratio più favorevole rispetto al Testosterone, sarebbe stato abbandonato per qualsiasi applicazione diversa dalla sostituzione pura degli androgeni nei maschi (ad es. Proviron). Ma erano i test sull’uomo la vera “prova del 9”.
I derivati del DHT più comunemente usati per il miglioramento delle prestazioni includono quanto segue:
Masteron (Drostanolone)
Primobolan (Metenolone)
Anavar (Oxandrolone)
Anadrol (Oxymetholone)
Winstrol (Stanozololo)
Proviron (Mesterolone)
Superdrol (methasterone)
DHB (Diidroboldenone, 1-Testosterone)
I derivati del DHT sono spesso percepiti come scelte sbagliate per una fase di Bulk/Off season perché non sembrano stimolare molta ipertrofia quanto i membri dei gruppi Testosterone e 19-Nor. Questo è un altro equivoco, poiché i derivati del DHT hanno semplicemente un’azione più mirata nel corpo. Non sono substrati per l’Aromatasi come i derivati del Testosterone, non agiscono come potenti agonisti di una miriade di diversi recettori nel corpo come i 19-Nor steroidi e non subiscono la 5α riduzione in metaboliti più androgeni.
I derivati del DHT eliminano gran parte dell’attività meno prevedibile associata agli altri due gruppi e si orientano maggiormente verso l’espressione proteica pura e una maggiore stimolazione di forza. Non c’è ritenzione idrica, non ci sono effetti collaterali progestinici, tutto ciò che si ottiene è pura crescita muscolare e quantità proporzionalmente più elevate di guadagni di forza rispetto ai derivati del Testosterone e 19-Nor steroidi.
I derivati del DHT mostrano più o meno un’azione mirata sul tessuto contrattile, motivo per cui sono comunemente percepiti come anabolizzanti “deboli”, quando in realtà è la mancanza di effetti collaterali percepiti il motivo per il quale vengono erroneamente considerati come “deboli”. Quando si tratta di ritenzione di azoto, abbiamo visto negli studi clinici che non c’è una differenza significativa tra gli AAS più potenti di ciascuna categoria.
“Dopo il 1935 il metodo migliore per scoprire e misurare l’azione di costruzione delle proteine da parte degli steroidi androgeni nell’uomo si è rivelato essere lo studio dell’equilibrio metabolico.
Nel 1955, quando furono sviluppati gli steroidi anabolizzanti meno androgeni, il metodo del bilancio dell’azoto fu nuovamente utilizzato per valutare e confrontare l’effetto di risparmio di azoto delle varie preparazioni. I risultati dei numerosi studi sull’equilibrio dell’azoto che sono stati eseguiti sono i seguenti: i 17 beta-esteri iniettabili, come Nandrolone Fenilpropionato, Nandrolone Decanoato e Metenolone Enantato esercitano una forte azione anabolica per diverse settimane, pari a 2-2,50g di azoto/giorno, che corrisponde a un guadagno giornaliero di 12-15g di proteine o 60-75g di massa magra. I derivati 17-alchilati oralmente attivi inducono un effetto di risparmio di azoto dose-dipendente dello stesso ordine.”
La ritenzione di azoto era più o meno la stessa tra tutti gli steroidi valutati nello studio di cui sopra.[39]
Il più delle volte sono gli effetti collaterali stessi che vengono interpretati erroneamente come indice di un composto più potente di un altro in un contesto di costruzione muscolare.
Se guadagni 2.5Kg di ritenzione idrica pura in una settimana con l’uso di Methandrostenolone, è un costruttore di muscoli più potente del Oxandrolone? No non lo è. Quando tutto è stato detto e fatto, la quantità di tessuto contrattile effettivo guadagnato sarà simile, ma ciò che accade al corpo e come vengono aumentati gli altri indicatori della composizione corporea durante l’intervallo di tempo in cui ti muovi dal punto A al punto B è ciò che differirà significativamente tra quei due composti.
Solo perché i derivati del DHT hanno un’azione più mirata, non significa che siano la scelta ideale in tutti gli scenari.
In effetti, a volte gli effetti collaterali di alcuni composti producono risultati più desiderabili, a seconda dell’obiettivo. In generale, i derivati del DHT sono costruttori di forza con livelli affidabili e prevedibili di attività anabolica di qualità. Un esempio di uno sport in cui i derivati del DHT sono particolarmente utili è l’MMA anche se atleti di altre discipline di combattimento possono trovare in questa classe di AAS un netto vantaggio prestativo.
I combattenti devono combattere con una classe di peso il più bassa possibile senza compromettere le loro prestazioni, devono avere un rapporto favorevole di produzione di forza rispetto al loro peso corporeo e traggono grandi benefici anche dall’aumento dell’aggressività e dal potenziamento neurologico. Questo è uno scenario specifico in cui usare qualcosa che è più orientato verso l’accumulo di massa e meno verso l’attività androgena sarebbe la scelta peggiore.
Ad esempio, il Nandrolone è 5α-ridotto in DHN, un metabolita molto meno androgeno, e interagisce anche con i recettori del progesterone nel corpo che possono avere un effetto anti-androgeno. Anche se questo potrebbe essere un composto ideale per chi cerca la massima ipertrofia con una relativa mancanza di effetti collaterali androgeni come la caduta dei capelli, in questo scenario è l’opposto di quello a cui vorremmo mirare.
Per un combattente di MMA vogliamo un’elevata produzione di forza e aggressività con una relativa mancanza di aumento di peso. Questo è un esempio di uno scenario in cui sarebbe saggio inserire un derivato del DHT. In alternativa, questo è un esempio di uno scenario in cui anche il 4-Chlorodehydromethyltestosterone e il Fluoxymesterone sarebbero buone opzioni, soprattutto il secondo, ma è solo perché si comportano più come i derivati del DHT che come i derivati del Testosterone quali sono.
Ci sono eccezioni alla regola in ogni categoria di AAS e i derivati del DHT non sono diversi in quanto hanno l’Oxymetholone.
Nonostante sia un derivato del DHT, l’Oxymetholone si comporta più come un 19-Nor steroide poiché determina guadagni significativamente più pronunciati nella massa pura (da non confondere con la massa muscolare magra) rispetto agli altri derivati del DHT, ed è anche un agonista dei recettori degli estrogeni.
In generale, tuttavia, i derivati del DHT più comunemente sfruttati dagli atleti sono Methenolone, Drostanolone, Oxandrolone e Stanozololo e condividono tutti effetti sovrapposti simili sulla composizione corporea e sulle prestazioni.
L’ultimo gruppo di AAS è rappresentato dai derivati del 19-Nortestosterone.
Molti non si rendono nemmeno conto che il Nandrolone Decanoato e il Nandrolone Fenilpropionato non sono AAS diversi ma semplicemente la stessa molecola legata ad esteri differenti.
Quando qualcuno si riferisce al “Deca” si riferisce al Nandrolone legato ad un estere Decanoato. E, quando qualcuno si riferisce al “NPP” si riferisce al Nandrolone legato ad un estere Fenilpropionato.
L’ormone progenitore di questo gruppo è il Nandrolone (19-Nortestosterone) e tutti gli steroidi anabolizzanti di questa categoria sono derivati da esso.
I derivati del Nandrolone sono più comunemente indicati semplicemente come “19-Nor steroidi” nella comunità del Bodybuilding. Ci sono alcuni 19-Nor stroidi che sono stati sintetizzati e documentati nel corso degli anni, la maggior parte dei quali sono tradizionalmente usati a bassi dosaggi come mezzi contraccettivi per le donne.
I 19-Nor steroidi più comunemente utilizzati per il miglioramento delle prestazioni includono:
Nandrolone (19-Nortestosterone)
Trenbolone (Trienolone)
Methyltrienolone
Trestolone (MENT)
I 19-Nor steroidi sono generalmente caratterizzati dalla loro attività anabolica e progestinica.
Sono molto anabolizzanti e, ad eccezione del Nandrolone, posseggono una spiccata attività androgena. Mostrano anche quantità significative di interazione con altri recettori nel corpo. Il più notevole è rappresentato dalla loro interazione con il recettore del progesterone.
I 19-Nor steroidi sono progestinici e da ciò né deriva una attività agonista del recettore del progesterone, il che significa che possono legarsi a questo recettore attivarlo.
Il Nandrolone non è un potente substrato per l’Aromatasi e si converte principalmente in un estrogeno più debole chiamato Estrone (l’Estradiolo è circa 10 volte più potente dell’Estrone). Il Nandrolone è anche leggermente estrogenico da solo grazie alla sua capacità di agire come agonista del recettore degli estrogeni alfa (ERα).[40] Ma, nel complesso, il Nandrolone è molto meno androgeno ed estrogenico del Testosterone.
Il Trenbolone non è un substrato soggetto all’azione dell’enzima Aromatasi, tuttavia, alcuni dati suggeriscono che potrebbe interagire con i recettori degli estrogeni in modo simile al Nandrolone.
Il Trenbolone facilita anche un maggiore risparmio proteico muscolare attraverso una serie di meccanismi anticatabolici che interessano il recettore dei glucocorticoidi (vedi recettore del Cortisolo). Il Trenbolone si lega al recettore dei glucocorticoidi e agisce come un antagonista.[41] Inoltre sopprime significativamente l’espressione dei glucocorticoidi.[42] Riduce i livelli di Corticosterone e Cortisolo, mentre contemporaneamente inibisce il legame del Cortisolo con i recettori glucocorticoidi del muscolo scheletrico.[43, 44, 45]
Attraverso questi meccanismi, il Trenbolone mostra un’inibizione molto più significativa della degradazione proteica muscolare rispetto al Testosterone. Questo è il motivo per cui il Trenbolone sembra eccellere molto più di altri steroidi anabolizzanti in un contesto di deficit calorico.
Il Trestolone (MENT) è unico rispetto agli altri 19-Nor steroidi in quanto è un substrato soggetto all’enzima Aromatasi e si comporta quasi come un ibrido tra un 19-Nor steroide e il Testosterone. A causa di questa interazione con l’Aromatasi, il Trestolone si presenta come una promessa terapeutica potenziale alternativa alla terapia ormonale sostitutiva a base di Testosterone e può ricoprire il ruolo di valida alternativa come AAS di base in un ciclo, sebbene la il deficit di DHT e il comportamento non lineare con gli effetti biochimici di questo nei tessuti bersaglio potrebbero essere un problema.
I 19-Nor steroidi sono il gruppo di AAS più soppressivo e sono in grado di mantenere la soppressione della HPTA anche a minuscole tracce e con i loro metaboliti i quali perdurano per lungo tempo nel flusso ematico.
Considerando questo, sarebbe prudente riservarne l’uso solo in atleti avanzati e decisi a gestirne le conseguenze. Anche una sola iniezione di Nandrolone può mantenere soppressa l’HPTA per mesi, indipendentemente da ciò che uno faccia in seguito.
Il gruppo dei 19-Nor steroidi tende maggiormente a indurre ipertrofia rispetto ai derivati del DHT, ma, come accennato precedentemente, ciò è anche accompagnato da una miriade di interazioni con altri recettori nel corpo che sono meno prevedibili e spesso giustificano una gestione responsabile da parte di un utilizzatore più esperto.
I 19-Nor steroidi hanno anche interazioni uniche con il percorso GH/IGF-1 che i derivati DHT non hanno, il che complica ulteriormente una loro possibile applicazione “responsabile”, ma può anche essere sfruttato per aumentare i livelli di crescita muscolare in determinati scenari.
Comprendere le applicazioni ottimali degli anabolizzanti e la gestione degli effetti collaterali può diventare molto complicato, specialmente quando si entra nelle varie sfaccettature prestazionali per ogni specifico sport, per i test antidoping, per impossibilità di reperire determinati farmaci e i limiti di budget.
Per semplificare le cose, i più previdenti iniziano le loro preparazioni farmacologiche suddivido gli AAS nei tre gruppi precedentemente esposti. Iniziando dal gruppo del Testosterone e osservando le risposte che questo ha sul soggetto, approfondendo successivamente i meccanismi metabolici del Testosterone nel corpo dell’individuo interessato e analizzando il tasso di aromatizzazione in Estradiolo e di 5α riduzione in DHT. Solo successivamente, come passo ulteriore, un soggetto potrà sperimentare abbinamenti con gli AAS del gruppo dei DHT derivati.
Molti, ancora oggi, soffrono di una sorta di “estrogenofobia”. Queste persone dovrebbero comprendere che un controllo degli estrogeni non significa il loro azzeramento, dal momento che un corretto dosaggio di E2 faciliterà indirettamente il bilanciamento dell’attività anabolica e, direttamente, dell’attività androgena, fornendo contemporaneamente neuro e cardioprotezione.
La maggior parte di coloro che leggono questo articolo non avranno applicazioni specifiche per lo sport praticato o per i test antidoping ai quali potrebbero essere sottoposti. Considerando anche questo fattore, la maggior parte dei soggetti si saprà gestire meglio imparando a fondo sull’uso di Testosterone (magari con esteri a breve vita attiva) prima ancora di passare agli altri due gruppi.
Il Testosterone è l’androgeno anabolizzante bioidentico che produciamo e su cui facciamo affidamento in modo endogeno, ed è il più delle volte l’ormone più intelligente da utilizzare per un primo ciclo. Solo una volta che si è in grado di sfruttare appieno il gruppo del Testosterone si dovrebbe passare all’aggiunta di uno o più derivati del DHT nei cicli successivi.
Se si hanno obiettivi che non possono essere raggiunti solo tramite derivati del Testosterone e del DHT, o si verificano effetti collaterali indesiderati da derivati del Testosterone e / o del DHT, sarebbe giustificato guardare al gruppo dei derivati del Nandrolone.
Per una spiegazione più elaborata di come uno dovrebbe avvicinarsi al mondo dei PEDs in generale e degli AAS in particolare, consiglio di leggere il mio articolo che descrive in dettaglio ciò che va obbligatoriamente preso in considerazione prima di fare il “salto”. Per saperne di più sui potenziali effetti collaterali da uso/abuso di AAS li trovate nell’apposita sezione di questo sito.
Ricapitolando, una volta che si è compreso nel dettaglio il gruppo del Testosterone e suoi derivati e si è in grado di implementare responsabilmente tali informazioni per raggiungere i propri obiettivi, passate a conoscere i derivati del DHT. Una volta che avrete compreso anche questo gruppo e sapete come implementare responsabilmente tali informazioni per raggiungere i vostri obiettivi, iniziate a conoscere il Nandrolone e i suoi derivati.
Cosa fare delle informazioni fino ad ora esposte?
Ciò che farete delle informazioni esposte non è affare che mi riguarda, e penso che tutti voi siate al corrente che l’uso, la detenzione e lo spaccio di sostanze dopanti è perseguibile a norma di legge.
Adesso però conoscete la “genesi” storica degli AAS, avete una base sufficiente per ciò che concerne la chimica di questa categoria di farmaci e, infine, conoscete le linee valutative che dovrebbero essere applicate in ambito sportivo, quindi nel miglioramento delle prestazione per ogni categoria sportiva.
Avete una conoscenza sufficiente ad evitarvi di cadere nel vortice delle “credenze da spogliatoio” che dilagano specialmente in ambito Bodybuilding.
Sicuramente il vostro livello di cultura biochimica è stato valorizzato… che non è poco in tempi di relativismo universale.
2- Hoberman JM, Yesalis CE (1995). “The history of synthetic testosterone”. Scientific American. 272 (2): 76–81.
3- C.-É. Brown-Séquard: De la transmission croisée des impressions sensitives par la moelle épinière. Comptes rendus de la Société de biologie, (1850) 1851, 2: 33–44.
4- Kelly, Howard A.; Burrage, Walter L. (eds.). “Brown-Séquard, Charles Edward” . American Medical Biographies . Baltimore: The Norman, Remington Company.
5- Greenblatt, Robert B. (1963). Search The Scriptures: A Physician Examines Medicine in the Bible. Philadelphia: J. B. Lippincott. p. 55.
6- The Practice of Neuroscience, pp. 199–200, John C.M. Brust (2000).
7- Segerberg, Jr., Osborn (1974). The Immortality Factor. New York: E. P. Dutton and Co., Inc. pp. 84–85.
8- David K, Dingemanse E, Freud J, Laqueur L (1935). “Uber krystallinisches mannliches Hormon aus Hoden (Testosteron) wirksamer als aus harn oder aus Cholesterin bereitetes Androsteron”. Hoppe-Seyler’s Z Physiol Chem.
9- Hoberman JM, Yesalis CE (1995). “The history of synthetic testosterone”. Scientific American.
10- Freeman ER, Bloom DA, McGuire EJ (2001). “A brief history of testosterone”. Journal of Urology.
11- Butenandt A, Hanisch G (1935). “Über die Umwandlung des Dehydro-androsterons in Δ4-Androsten-ol-(17)-0n-(3) (Testosteron); ein Weg zur Darstellung des Testosterons aus Cholesterin (Vorläuf. Mitteil.)” [On the conversion of dehydro-Δ4-androstene androsterons in-ol (17) 0n (3) (testosterone), a way to represent the testosterone from cholesterol (Vorläuf. msgs.)]. Berichte der Deutschen Chemischen Gesellschaft (A and B Series) (in German). 68 (9): 1859–62.
12- Ruzicka L, Wettstein A (1935). “Sexualhormone VII. Uber die kunstliche Herstellung des Testikelhormons. Testosteron (Androsten-3-one-17-ol.)” [Sex hormones VII About the artificial production of testosterone Testikelhormons (androstene-3-one-17-ol)]. Helvetica Chimica Acta (in German).
14- Hoberman JM, Yesalis CE (1995). “The history of synthetic testosterone”. Scientific American.
15- Lenahan P (2003). Anabolic Steroids: And Other Performance-enhancing Drugs. London: Taylor & Francis.
16- Taylor WN (January 1, 2009). Anabolic Steroids and the Athlete. McFarland & Company. p. 181. 17- Suarez R, Senior Correspondent, Kelman J, physician (2002-11-18). “President Kennedy’s Health Secrets”. PBS NewsHour. Public Broadcasting System.
18- Justin Peters The Man Behind the Juice, Slate Friday, Feb. 18, 2005. Retrieved 29 April 2008^
19- Hartgens F, Kuipers H (2004). “Effects of androgenic-anabolic steroids in athletes”. Sports Med. 34
20- Von Eckardstein, Sigrid, and Eberhard Nieschlag. “Treatment of male hypogonadism with testosterone undecanoate injected at extended intervals of 12 weeks: a phase II study.” Journal of andrology 23.3 (2002): 419-425. 21- Kalicharan, Raween Wikesh. New Insights into Drug Absorption from Oil Depots. Diss. Utrecht University, 2017. 22-Van der Vies, J. “Implications of basic pharmacology in the therapy with esters of nandrolone.” European Journal of Endocrinology 110.3_Suppla (1985): S38-S44. 23-Ojasoo, T., et al. “Towards the mapping of the progesterone and androgen receptors.” Journal of steroid biochemistry 27.1-3 (1987): 255-269. 24-Becker, H., et al. “In vivo uptake and metabolism of 3H-testosterone and 3H-5α-dihydrotestosterone by human benign prostatic hypertrophy.” European Journal of Endocrinology 71.3 (1972): 589-599. 25-Pereira de Jésus‐Tran, Karine, et al. “Comparison of crystal structures of human androgen receptor ligand‐binding domain complexed with various agonists reveals molecular determinants responsible for binding affinity.” Protein Science 15.5 (2006): 987-999. 26-Feldkoren, Boris I., and Stefan Andersson. “Anabolic-androgenic steroid interaction with rat androgen receptor in vivo and in vitro: a comparative study.” The Journal of steroid biochemistry and molecular biology 94.5 (2005): 481-487. 27-Luu-The, Van. “Analysis and characteristics of multiple types of human 17β-hydroxysteroid dehydrogenase.” The Journal of steroid biochemistry and molecular biology 76.1-5 (2001): 143-151. 28-Täuber, U., et al. “Absolute bioavailability of testosterone after oral administration of testosterone-undecanoate and testosterone.” European journal of drug metabolism and pharmacokinetics 11.2 (1986): 145-149. 29-Bond, Peter, William Llewellyn, and Peter Van Mol. “Anabolic androgenic steroid-induced hepatotoxicity.” Medical Hypotheses 93 (2016): 150-153. 30- Houtman, Corine J., et al. “Detection of anabolic androgenic steroid abuse in doping control using mammalian reporter gene bioassays.” Analytica chimica acta 637.1-2 (2009): 247-258. 31- Takeda, Armelle-Natsuo, et al. “The synthetic androgen methyltrienolone (r1881) acts as a potent antagonist of the mineralocorticoid receptor.” Molecular pharmacology 71.2 (2007): 473-482. 32-Bergink, E. W., et al. “Comparison of the receptor binding properties of nandrolone and testosterone under in vitro and in vivo conditions.” Journal of steroid biochemistry 22.6 (1985): 831-836. 33-Schänzer, Wilhelm. “Metabolism of anabolic androgenic steroids.” Clinical chemistry 42.7 (1996): 1001-1020. 34-Chen, Mo, and Trevor M. Penning. “5β-Reduced steroids and human Δ4-3-ketosteroid 5β-reductase (AKR1D1).” Steroids 83 (2014): 17-26. 35-Ryan, Kenneth J. “Biological aromatization of steroids.” Journal of Biological Chemistry 234.2 (1959): 268-272. 36-Pitteloud, Nelly, et al. “Inhibition of luteinizing hormone secretion by testosterone in men requires aromatization for its pituitary but not its hypothalamic effects: evidence from the tandem study of normal and gonadotropin-releasing hormone-deficient men.” The Journal of Clinical Endocrinology & Metabolism 93.3 (2008): 784-791. 37-Behre, H. M., et al. “Suppression of spermatogenesis to azoospermia by combined administration of GnRH antagonist and 19-nortestosterone cannot be maintained by this non-aromatizable androgen alone.” Human Reproduction 16.12 (2001): 2570-2577. 38-Le Guevel, Rémy, and Farzad Pakdel. “Assessment of oestrogenic potency of chemicals used as growth promoter by in-vitro methods.” Human reproduction 16.5 (2001): 1030-1036.
Chi segue il sito e legge con attenzione i miei lavori, si ricorderà certamente che l’argomento della Anabolico:Androgeno ratio era già stato toccato nell’articolo di analisi dettagliata sul Methenolone. Visto che la questione alzò una non indifferente reazione da parte degli “irriducibili” del “ribattere con banalità”, e che la lettura di certi validi testi in lingua inglese sembra per i più ostica, ho deciso di trattare con minuzia di dettagli questo tanto dibattuto argomento. Ovviamente, chi vive di convinzioni basate sul nulla difficilmente potrà accettare quanto mi accingerò a riportare. Per tutti gli altri sarà un altra occasione per imparare qualcosa di nuovo e potenzialmente utile.
Ma andiamo avanti…
Tutti noi sappiamo che gli Steroidi Androgeni Anabolizzanti (AAS) hanno proprietà anaboliche e androgene, da cui il nome. In generale, per proprietà anaboliche si indica l’effetto di costruzione muscolare e l’effetto stimolante sulla densità minerale ossea (BMD). Gli altri effetti sono considerati effetti Androgeni, come l’impatto sulla ipertrofia prostatica, sullo stimolo del midollo osseo, sul cuore, sull’ipotalamo e sull’ipofisi, ecc. In generale, questi effetti sono considerati indesiderati. Ad esempio, una meta-analisi ha rilevato che un aumento dell’ematocrito (la % del volume di sangue occupato dagli eritrociti) è l’effetto avverso più frequente associato alla Terapia Sostitutiva del Testosterone (TRT).[1] Un altro problema degli androgeni è che sono in grado di indurre, in particolari circostanze, la crescita del cancro alla prostata. In quanto tale, una delle terapie utilizzate per il trattamento del cancro alla prostata è la terapia di deprivazione androgenica. Tuttavia, è importante notare che esiste un limite alla capacità degli androgeni di stimolare la crescita del cancro alla prostata. Ciò significa che, fino a una certa concentrazione, gli androgeni ne stimoleranno la crescita, ma al di sopra di essa avranno poco o nessun ulteriore effetto. Da qui sono nate alcune ipotesi tra le quali quella del “modello di saturazione dei recettori degli androgeni” [2], un modello dibattuto e tutt’altro che dimostrato. Comunque sia, questo effetto sulla riduzione dell’attività ipertrofica prostatica è in realtà qualcosa che sembra già avvenire a basse concentrazioni di Testosterone, nel intervallo basso classico del soggetto ipogonadico. Avremo comunque tempo di ritornare nuovamente su questo punto più tardi, quando parlerò di un rinomato test che viene utilizzato per valutare il rapporto tra potenza anabolica e androgena.
Lo scopo della Androgeno:Anabolico ratio è quello di fornire dati numerici al fine di dividere i diversi AAS in termini di potenza anabolica e androgena. Quindi, ad esempio, si potrebbe prendere il Testosterone come AAS di “paragone”, assegnandogli una Anabolico/Androgeno ratio di 100 e 100 (o solo 1). Quindi, attraverso alcuni esperimenti, viene determinato che un altro AAS ha un rapporto pari a 400:200 (o solo 2) Ciò implicherebbe che questo AAS è 4 volte più anabolico del Testosterone, pur essendo solo due volte più androgena.
Se le cose stessero in questo modo, allora, volendo ridurre al minimo il rischio di effetti collaterali androgeni, si potrebbe semplicemente scegliere un AAS con una Anabolico-Androgeno ratio molto favorevole e il problema non sussisterebbe. Tuttavia, ci sono così tanti problemi e variabili sia con il concetto stesso di Anabolico:Androgeno, ratio sia con il modo in cui esso è determinato sperimentalmente, che tutti questi rapporti che si trovano online o in letteratura sono praticamente inutili.
Il test di Hershberger
Un test molto comune utilizzato per determinare la Anabolico:Androgeno ratio è il cosiddetto test di Hershberger. Il test è stato descritto per la prima volta nel 1953 da Hershberger e dai suoi colleghi dell’Università del Wisconsin.[3] Come già accennato nell’articolo sul Methenolone, il test funziona come segue. Si prendono dei ratti e li si castra. La castrazione assicura che si abbia pochissimo Testosterone endogeno nell’animale e che ciò possa influenzare i risultati del test. Successivamente, si somministra l’AAS di cui si vuole conoscere il rapporto tra potenza anabolico e androgena all’animale. Successivamente si attende un po’ di tempo (8 giorni nel caso del originale test di Hershberger) e si procede con l’uccisione dei ratti trattati per sezionarli e pesarne il muscolo levator ani, la prostata ventrale e le vescicole seminali. L’aumento di peso de levator ani sarebbe quindi indicativo dell’attività anabolica dell’AAS, mentre quello della prostata ventrale e delle vescicole seminali sarebbe indicativo della sua attività androgena.
LA: levator ani.
Anche se questo test può sembra un metodo ragionevolmente valido, in esso vi sono una serie di problemi. Un primo punto su cui vorrei soffermarmi riguarda il muscolo levator ani, che, appunto, è il muscolo bulbocavernoso dorsale.[4] È un muscolo che fa parte del sistema riproduttivo maschile e quindi non dovrebbe essere considerato in alcun modo rappresentativo del muscolo scheletrico. È un muscolo fortemente androgeno-dipendente e dopo la castrazione subisce un tasso di diminuzione del peso simile a quello dell’atrofia da denervazione nei muscoli scheletrici.[5] Proprio questa informazione da sola garantisce già che il lato anabolico dell'”equazione” sia imperfetto. Un altro problema è che il muscolo bulbocavernoso dorsale e le vescicole seminali rispondono in modo diverso a una diminuzione della concentrazione di AAS all’interno del range fisiologico.[6] Di conseguenza, il rapporto determinato sperimentalmente dipenderà dalla dose utilizzata e dal momento in cui vengono effettuate le misurazioni. Ciò è ben illustrato nella figura sottostante tratta da una pubblicazione di van der Vies.[6] Durante i primi 3 giorni, la concentrazione di AAS (Nandrolone in questo caso) è abbastanza alta da stimolare la crescita sia delle vescicole seminali che del muscolo bulbocavernoso dorsale. Tuttavia, dopo tre giorni la concentrazione non è abbastanza elevata da sostenere questa crescita per le vescicole seminali, che diminuiscono nuovamente di dimensioni. Tuttavia, il muscolo bulbocavernoso è ancora sufficientemente stimolato per continuare a crescere di dimensioni. Pertanto, se determinassi il rapporto anabolico/androgeno il giorno 3, esso sarebbe molto diverso rispetto al risultato che rileverei se la misurazione venisse fatta il giorno 7, nonostante sia utilizzato lo stesso composto.
Scomparsa del Nandrolone Fenilpropionato dal deposito intramuscolare ed effetti sui pesi del muscolo bulbocavernoso dorsale e delle vescicole seminali. Figura tratta da van der Vies [6].
Ciò evidenzia anche che i diversi organi rispondono semplicemente in modo diverso a seconda della concentrazione della molecola. E anche se ci fosse un modo accurato per determinare una anabolico:androgeno ratio, estrapolarlo oltre le concentrazioni fisiologiche sarebbe completamente errato.
Un altro difetto è l’ipotesi che la crescita della prostata ventrale o delle vescicole seminali sia rappresentativa di tutti gli altri effetti androgeni. Non ci sono mai state prove a sostegno di questa ipotesi. I tessuti androgeni variano molto in risposta l’uno dall’altro e non ci si deve assolutamente aspettare che un tessuto risponda nella stessa misura di un altro. In effetti, ricordate quanto menzionato nell’introduzione sulla prostata? Gli androgeni sembrano già smettere di stimolare ulteriormente la crescita della prostata oltre l’intervallo ipogonadico basso. Infatti, il volume della prostata rimane invariato quando a uomini sani vengono somministrati 600mg di Testosterone Enantato (pari a 432mg di Testosterone) settimanalmente per 20 settimane.[7] Eppure sappiamo per certo che altri effetti collaterali androgeni iniziano a comparire quando il dosaggio comincia ad essere elevato! Comunque, questo mette in luce anche la questione del concetto stesso di rapporto tra potenza anabolica e androgena. Un singolo rapporto non è mai in grado di catturare le risposte differenziali dei vari tessuti sensibili agli androgeni o la complessità della risposta androgena all’interno di un tessuto specifico, per essere di valore. Diversi tessuti rispondono in modo diverso ad un AAS, come sarà mai possibile rappresentarlo con un singolo numero?
Naturalmente, il test di Hershberger viene eseguito sui ratti, non sugli esseri umani. È un altro errore è quello di presumere che i tessuti omologhi nell’uomo rispondano allo stesso modo osservato in un ratto trattato con AAS. L’intero test di Hershberger è semplicemente pieno di falle, e nonostante ciò viene attualmente utilizzato per lo screening di potenziali Modulatori Selettivi del Recettore degli Androgeni (SARM). Ad esempio, GlaxoSmithKline ha valutato la selettività tissutale del proprio SARM GSK2881078 utilizzando il classico test di Hershberger.[8]
GSK2881078
Test di affinità di legame relativo (RBA)
Mentre il test di Hershberger viene eseguito in un organismo vivente, i saggi di affinità di legame relativa (RBA) vengono eseguiti in una sorta di piastra di Petri. Viene quindi esaminata l’affinità di legame dei composti per il Recettore degli Androgeni (AR). In questo contesto, l’affinità di legame si riferisce alla forza con cui un AAS si lega all’AR. L’RBA mostra quindi quanto fortemente un AAS si lega all’AR rispetto ad un altro. O in altre parole: relativamente l’uno all’altro.
Il principio alla base è abbastanza semplice. Si prende un AAS di riferimento, comunemente il Methyltrienolone (R1811), e si misura la sua affinità di legame. Successivamente si misura l’affinità di legame di altri AAS e si esprimono i dati relativi all’AAS di riferimento. Quindi al Methyltrienolone viene assegnato un RBA di 1, essendo il composto di riferimento, e quindi se qualche altra molecola si lega con una potenza maggiore di due volte gli viene assegnato un RBA di 2. Allo stesso modo, se un’altra molecola si lega due volte più debolmente le viene assegnato un RBA di 0,5. Si potrebbero fare queste misurazioni in diversi tipi di cellule. Una che rappresenta il muscolo scheletrico e un altra che rappresenta in qualche modo i suoi effetti androgeni (ad esempio le cellule della prostata). Come tale, anche in questo caso si possono porre alcune delle stesse obiezioni del test Hershberger descritte sopra.
Ad ogni modo, l’immagine seguente raccoglie gli RBA di una selezione di AAS popolari misurati nei tessuti di ratto e coniglio.[9] Se dovessimo ottenere questi risultati, il Testosterone avrebbe un rapporto anabolico-androgeno più favorevole rispetto al Nandrolone. È un po’ l’opposto di quello che si osserva nei test di Hershberger. È anche un po’ sorprendente, dato che gli effetti androgeni del Testosterone sono amplificati nei tessuti che esprimono la 5α-reduttasi, a causa della conversione all’androgenicamente più potente DHT. Al contrario, l’azione androgena del Nandrolone è indebolita nei tessuti che esprimono questo enzima, a causa della conversione al meno potente androgeno Dihydronandrolone (DHN).[10]
Gli RBA di una selezione di AAS presi dal lavoro di Saartok et al. [9]. Il Methyltrienolone è servito come steroide di riferimento.
Un’altra cosa che questi dati rivelano in modo appropriato sono le differenze interspecie dei valori RBA. Nel muscolo di ratto, l’1α-methyl DHT si lega all’AR circa 3 volte più debolmente del Testosterone. Se si osservano i dati provenienti dall’analisi del muscolo di coniglio, fondamentalmente si vede un risultato contrario: 1α-methyl-DHT si lega con una forza all’incirca 3 volte maggiore all’AR come il Testosterone. L’estrapolazione da una specie animale a un’altra è (altamente) problematica, e quindi anche l’estrapolazione dal ratto, coniglio o qualsiasi altro animale, all’uomo.
Nel caso ti stia chiedendo perché il DHT dimostri un RBA così basso nel muscolo di coniglio e ratto, questo è probabilmente dovuto alla sua rapida degradazione nel tessuto muscolare. Il DHT costituisce un eccellente substrato per l’enzima 3α-HSD. Questo enzima lo scompone in 3α-androstanediolo, il quale si lega molto debolmente all’AR.[11] Questo accade anche negli esseri umani [12], e questo è uno dei motivi per cui non si vedono protocolli basati sull’uso di DHT.
Un ultimo punto che deve essere evidenziato è che l’affinità di legame non determina la potenza del AAS nel modulare anche l’espressione genica. Che è alla fine ciò che più interessa. Tuttavia, questo è possibilmente valutabile per via sperimentale. Si tratta del test del gene reporter responsivo agli androgeni (dosaggi biologici AR). Questi test biologici, per quanto ne so, sono stati inizialmente utilizzati per lo screening di nuovi androgeni di design nei campioni di urina per contrastare l’uso di doping. Un test biologico AR è essenzialmente in grado di dimostrare se un campione contiene qualcosa che riesce ad attivare il recettore degli androgeni e avviare la trascrizione genica. Per contrastare il doping, questo è molto utile. Dopotutto, puoi dimostrare che un campione di urina contiene qualcosa che attiva l’AR senza conoscere la struttura chimica del composto utilizzato.
Ad ogni modo, uno di questi test è stato sviluppato da un team di scienziati olandesi. [13] I ricercatori hanno utilizzato un test chiamato test biologico della LUciferasi attivata da sostanze chimiche reattive agli androgeni (AR CALUX). Hanno preso una linea cellulare di osteosarcoma umano e l’hanno co-trasfettata con l’AR umano e un gene reporter della luciferasi che è sotto il controllo trascrizionale degli elementi di risposta agli androgeni (ARE). Ciò significa che quando l’AR viene attivato, l’enzima luciferasi arriva all’espressione. Questo enzima produce bioluminescenza, o per dirla semplicemente: luce. E la luce può essere misurata. Quindi il grado di bioluminescenza è il grado in cui avviene l’attivazione del recettore degli androgeni.
I ricercatori hanno quindi proceduto a testare una varietà di AAS noti con il test biologico AR CALUX. Simile all’RBA, con esso si può calcolare la potenza relativa in termini di attivazione del recettore (REP). E non è solo stato fatto per l’AR, ma lo hanno fatto anche per il recettore del progesterone (PR), entrambe le isoforme del recettore degli estrogeni (ERα e ERβ) e il recettore dei glucocorticoidi (GR). Nella tabella seguente sono elencati gli REP di alcuni (popolari) AAS.
Gli REP di una selezione di AAS tratti dal lavoro di Houtman et al. [13]. Il DHT è servito come steroide di riferimento per il AR, ORG-2058 per il PR, l’Estradiolo per ERα/β e il Desametasone per GR.
Permettetemi di evidenziare il REP del Testosterone e del DHT per l’AR. Il REP del Testosterone è circa 5 volte inferiore al REP del DHT. Cosa ci dice questo? Semplicemente che quel DHT non è stato degradato enzimaticamente nella linea cellulare che hanno usato come sarebbe successo nel mondo reale se si fosse legato al AR del muscolo scheletrico. Il metabolismo che di solito avviene nel muscolo scheletrico non sembra quindi avvenire in questa linea cellulare. Questo problema invalida i risultati di questo test biologico per quegli AAS che sono metabolizzati nel muscolo scheletrico, come il DHT, ma probabilmente anche il Methenolone (Primobolan). Un altro problema è che l’espressione genica è complessa (e dirlo è un eufemismo). L’AR regola un vasto numero di geni. Se due composti aumentano la trascrizione genica di un determinato gene in misura simile, non significa necessariamente che questi due composti modulino in modo comparabile la trascrizione genica di altri geni. Sicuramente non sarebbe sorprendente se ci fosse una correlazione in un modo o nell’altro, ma questi test biologici dipingono solo un quadro approssimativo. Anche se è probabile che questa immagine approssimativa sia più accurata di quella degli RBA. Tuttavia, fino ad oggi, i biotest AR non sono stati eseguiti in più linee cellulari di vari tessuti (sensibili agli androgeni) per fornirci “nuovi” rapporti tra il potenziale anabolico e androgeno.
Conclusioni
Sia il test di Hershberger che gli studi che valutano gli RBA dell’AAS in vari tessuti sono irrimediabilmente inaffidabili. Inoltre, il concetto di numero che cattura la complessità delle proprietà anabolizzanti e di quelle androgene dovrebbe essere abbandonato. Un singolo rapporto è semplicemente incapace di descrivere le risposte differenziali di vari tessuti agli androgeni, così come la complessità della risposta androgena all’interno di un tessuto specifico, per essere di valore. Forse sarebbe più appropriato un “profilo di attività” che descriva l’azione androgenica su base tissutale. Qualcosa di simile è stato proposto per descrivere come dovrebbe essere un SARM ideale per il trattamento di una condizione specifica. Tuttavia, è estremamente difficile quantificare l’azione androgena per tessuto, se non impossibile. Forse i biotest AR eseguiti su linee cellulari di tessuti di interesse potrebbero avere un valore clinico predittivo. Alla fine, dopo tutto, sono necessari studi clinici per dimostrare (il grado di) eventi avversi che si verificano con l’uso di un determinato composto.
Ovviamente, e lo dico per i tordi che affermano “e allora perchè con l’Oxandrolone gli effetti androgeni sono bassi come descritto dalla sua anabolico:androgeno ratio?” Bambino caro, l’Oxandrolone, come altri composti steroidei testati nel corso degli ultimi sessant’anni, hanno passato trial clinici dove gli effetti sono stati documentati anche nel caso di trattamento di donne in pre e post-menopausa. Ma non sono mai stati effettuati confronti di potenziale ed estrapolate ratio anabolico:androgeno. Semplicemente hanno osservato il miglioramento clinico dei pazienti trattati per svariate patologie e condizioni come, ad esempio, soggetti ustionati o gravemente sottopeso (vedi malati di HIV). Sono state effettuate biopsie, controlli della ritenzione d’azoto ma nessun test comparativo. Così facendo, puoi basarti sul grado di trofismo indotto dalla molecola e dal grado di espressione dei caratteri androgeni, ma, lo ribadisco, nessuna ratio di confronto!
Ricordate, inoltre, che per i soggetti sensibili l’espressione degli effetti androgeno-correlati avviene anche con le molecole con la ratio più bassa… Fatevene una ragione…
Gabriel Bellizzi
Riferimenti:
Calof, Olga M., et al. “Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials.” The Journals of Gerontology Series A: Biological Sciences and Medical Sciences 60.11 (2005): 1451-1457.
Morgentaler, Abraham, and Abdulmaged M. Traish. “Shifting the paradigm of testosterone and prostate cancer: the saturation model and the limits of androgen-dependent growth.” European urology 55.2 (2009): 310-321.
Hershberger, L. G., Elva G. Shipley, and Roland K. Meyer. “Myotrophic activity of 19-nortestosterone and other steroids determined by modified levator ani muscle method.” Proceedings of the Society for Experimental Biology and Medicine 83.1 (1953): 175-180.
Hayes, Keith J. “The so-called ‘levator ani’ of the rat.” European Journal of Endocrinology 48.3 (1965): 337-347.
Gori, Zina, C. Pellegrino, and Maria Pollera. “The castration atrophy of the dorsal bulbocavernosus muscle of rat: an electron microscopic study.” Experimental and molecular pathology 6.2 (1967): 172-198.
Van der Vies, J. “Implications of basic pharmacology in the therapy with esters of nandrolone.” European Journal of Endocrinology 110.3_Suppla (1985): S38-S44.
Bhasin, Shalender, et al. “Effect of testosterone supplementation with and without a dual 5α-reductase inhibitor on fat-free mass in men with suppressed testosterone production: a randomized controlled trial.” Jama 307.9 (2012): 931-939.
Neil, David, et al. “GSK2881078, a SARM, produces dose-dependent increases in lean mass in healthy older men and women.” The Journal of Clinical Endocrinology & Metabolism 103.9 (2018): 3215-3224.
Saartok, Tönu, Erik Dahlberg, and JAN-ÅKE GUSTAFSSON. “Relative binding affinity of anabolic-androgenic steroids: comparison of the binding to the androgen receptors in skeletal muscle and in prostate, as well as to sex hormone-binding globulin.” Endocrinology 114.6 (1984): 2100-2106.
Bergink, E. W., et al. “Comparison of the receptor binding properties of nandrolone and testosterone under in vitro and in vivo conditions.” Journal of steroid biochemistry 22.6 (1985): 831-836.
Jin, Yi, and Trevor M. Penning. “Steroid 5α-reductases and 3α-hydroxysteroid dehydrogenases: key enzymes in androgen metabolism.” Best Practice & Research Clinical Endocrinology & Metabolism 15.1 (2001): 79-94.
Becker, H., et al. “In vivo uptake and metabolism of 3H-testosterone and 3H-5α-dihydrotestosterone by human benign prostatic hypertrophy.” European Journal of Endocrinology 71.3 (1972): 589-599.
Houtman, Corine J., et al. “Detection of anabolic androgenic steroid abuse in doping control using mammalian reporter gene bioassays.” Analytica chimica acta 637.1-2 (2009): 247-258.
Chiunque segua questo sito o si sia interessato minimamente alla questione “doping”, è a conoscenza del fatto che durante l’uso di AAS e/o SARM, la produzione endogena di Testosterone subisce un calo marcato in misura maggiormente dipendente dalla molecola/e utilizzata/e e in minor parte dal tempo di utilizzo. Una volta interrotta la somministrazione di AAS e/o SARM, la produzione di Testosterone rimane (a diverso grado ma, pur sempre, significativo) soppressa per un periodo di tempo transitorio. Durante questo periodo di tempo, il soggetto si trova in una condizione di ipogonadismo, cioè sarà carente di Testosterone con importanti alterazioni di Estradiolo, DHT e Prolattina. Sappiamo allo stesso modo che è usanza comune l’utilizzo di alcuni farmaci dopo il termine d’uso di AAS e/o SARM con il fine, sperato, di accelerare il processo di recupero dell’attività dell’Asse HPT e la stabilizzazione della normale produzione di Testosterone. Questa pratica è ovviamente la conosciutissima, almeno per nome, PCT (Post-Cycle Therapy).
Tre tipi di farmaci sono frequentemente utilizzati per la PCT, e questi sono:
Modulatori Selettivi del Recettore degli Estrogeni (SERM), cioè Tamoxifene e Clomifene Citrato;
Inibitori dell’Aromatasi (IA), come Letrozolo, Anastrozolo ed Exemestane;
Gonadotropina Corionica umana (hCG).
Il ragionamento dietro l’uso di questi farmaci è abbastanza semplice. I SERM agiscono a livello del recettore degli estrogeni bloccando l’attività, principalmente, dell’Estradiolo portando ad un feedback negativo a livello ipofisario il quale, a cascata, porta ad un aumento del rilascio di GnRH e di LH ed FSH i quali, rispettivamente, andranno a stimolare la sintesi di Testosterone e la spermatogenesi. Allo stesso modo, gli Inibitori dell’Aromatasi causano una riduzione dei livelli di Estradiolo e, quindi, della sua attività portando ad un medesimo ciclo di feedback negativo stimolante il rilascio di GnRH e, consequenzialmente, di LH ed FSH. In fine, l’hCG viene usato inizialmente al fine di compensare i livelli bassi di LH e FSH, prima del loro incremento legato all’uso di SERM e AI, incrementando l’attività delle cellule di Leydig e del Sertoli stimolando la sintesi di Testosterone e la spermatogenesi.
Asse Ipotalamo-Ipofisi-Gonadi (HPGA; conosciuta anche come HPTA, Asse Ipotalamo-Ipofisi-Testicoli)
Di questi farmaci, i SERM sono solitamente il pilastro portante della PCT. E, in effetti, i SERM hanno dimostrato di aumentare il Testosterone in vari stati di ipogonadismo. Tuttavia, nessuno studio fino ad oggi aveva effettivamente esaminato in modo prospettico la sua efficacia nell’ipogonadismo indotto da AAS. Nemmeno la tanto acclamata PCT di Scally riporta scientificamente buone certezze d’efficacia. Di recente è uscito uno studio che ci mostra quanta efficacia possa avere una PCT nella “corsa al recupero” post ciclo di AAS e/o SARM. Parlo dello studio HAARLEM.[1]
Lo studio HAARLEM
Lo studio HAARLEM è uno studio prospettico e osservazionale a cui hanno partecipato 100 utilizzatori di AAS. Si tratta di un’iniziativa dell’ambulatorio per i consumatori di steroidi anabolizzanti di Haarlem, nei Paesi Bassi. L’ambulatorio nasce nel 2010 ed è gestito dai due endocrinologi dott. de Ronde e il dott. Smit.
L’obiettivo dello studio HAARLEM era quello di ottenere informazioni preziose sui rischi per la salute coinvolti nell’uso di AAS. Le caratteristiche di base di questa coorte sono state pubblicate in precedenza.[2]
In breve: nello studio sono stati inclusi un totale di 100 soggetti (tutti uomini) che intendevano iniziare un ciclo di steroidi anabolizzanti entro 2 settimane. Diverse misurazioni dello stato di salute, tra cui gli esami del sangue, sono state eseguite su tutti i partecipanti prima del ciclo (T0), durante l’ultima settimana del ciclo (T1), 3 mesi dopo la fine del ciclo (T2), e 1 anno dopo l’inizio del ciclo (T3). Per essere chiari: i soggetti stavano usando AAS che essi stessi si erano procurati, gli endocrinologi non hanno prescritto nessun AAS.
Ciò che è di particolare rilevanza per questo articolo è che i ricercatori hanno anche misurato i livelli di Testosterone e, quindi, hanno potuto osservare come potesse avvenire la ripresa dell’attività dell’Asse HPT dopo un ciclo. Inoltre, 80 dei soggetti in osservazione hanno eseguito la PCT (mentre i restanti 20 non hanno svolto alcuna PCT). Quindi, detto ciò, questo sarebbe il primo studio prospettico in cui l’efficacia della PCT potrebbe diventare evidente. Alla fine, però, i dati erano disponibili per 79 soggetti che avevano svolto la PCT e 19 soggetti che non l’avevano svolta.
Anche i farmaci per la PCT non sono stati forniti dagli endocrinologi. I soggetti interessati si sono procurati autonomamente tali farmaci. I ricercatori hanno notato che la maggior parte dei regimi PCT consisteva nell’uso di Tamoxifene Citrato (70% delle volte) e/o Clomifene Citrato (55% delle volte) per 4 settimane dopo il ciclo. Il che, in effetti, rappresenta l’esempio stereotipato di una classica PCT.
I risultati dello studio
Sono sicuro che questo darà fastidio a qualche “relativista ad oltranza”, ma i dati sono questi:
I valori di Testosterone basale (T0) erano praticamente identici e, come prevedibile, sono risultati aumentati a livelli soprafisiologici durante l’ultima settimana del ciclo (T1). Quindi, 3 mesi dopo la fine del ciclo, i valori sono stati di nuovo praticamente normalizzati in entrambi i gruppi (sebbene leggermente, ma non in modo statisticamente significativo, più bassi nel gruppo PCT).
Questa ricerca ha sicuramente delle mancanze e non arriva ad essere una “pietra miliare” ella dimostrazione scientifica in questo specifico contesto. Non si è trattato di uno studio in doppio cieco controllato con placebo. Ma è molto improbabile che un tale studio venga mai eseguito. Questo è un buon lavoro di ricerca in un frangente ben poco analizzato. Quali altre deficienze presenta lo studio HAARLEM? Qualcuno potrebbe blaterare riguardo ad improbabili bias di selezione. Cioè, i soggetti che “sanno” di recuperare più facilmente, potrebbero aver optato per non utilizzare una PCT. Dubito fortemente che ciò porterebbe a differenze significative. Un’altra ragione potrebbe essere che il dosaggio di AAS medio era più alto nel gruppo PCT, che era 1,110 contro 839mg/settimana. Tuttavia, entrambi sono ben al di sopra dei dosaggi richiesti per la massima soppressione della produzione endogena di Testosterone (il dosaggio minimo richiesto come criterio di inclusione nello studio era anche di 200mg a settimana). Inoltre, il gruppo che non ha svolto la PCT in media ha avuto una durata del ciclo più lunga (20 settimane contro 18 settimane).
In linea di principio, forse il gruppo PCT si era ripreso un po’ prima, il che sarebbe stato visibile se avessero misurato i marker specifici 2 mesi dopo aver interrotto l’uso di steroidi anabolizzanti invece che 3 mesi dopo. In effetti i controlli avrebbero dovuto essere più assidui. Comunque sia, fatte le dovute eccezioni, non ci si aspetterebbe comunque molta differenza . Se non altro perché la maggior parte di questi soggetti avrebbe impiegato probabilmente circa un mese prima che iniziasse il recupero dell’Asse HPT. Dopotutto, con alti dosaggi e molecole legate ad esteri che ne conferiscono lunghe emivite ci vorrà semplicemente più tempo prima che la soglia ematica degli AAS scenda sotto la curva del basale.
Sicuramente una buona parte di chi leggerà questo articolo dirà che (la maggior parte di) questi soggetti hanno semplicemente sbagliato la loro PCT. Ma, nonostante molti di voi considerino la “PCT di Scally” il metro di misura per valutare una PCT corretta da ciò che non lo è, purtroppo, non ci sono prove disponibili che abbiano esaminato l’efficacia dei vari tipi di PCT. Naturalmente, esiste una logica di gestione del post ciclo che andrebbe calcolata sul soggetto interessato. Tuttavia, questo studio mostra che quando si osserva un gruppo di persone che eseguono PCT come fatto nella maggior parte della pratica (SERM per circa un mese) semplicemente non si dimostra una reale efficace al fine di un recupero rapido della sintesi endogena di Testosterone. E, come si vede dai dati riportati, c’è stato uno scarso effetto accelerante se il gruppo non PCT si riprende dall’alterazione ormonale comunque in 3 mesi.
Come nota finale, gli autori chiariscono un punto chiave nel ridurre e migliorare i tempi di recupero e cioè il mantenimento della funzione gonadica per via somministrazione di hCG anche durante il ciclo. Infatti i ricercatori hanno scoperto che quando la funzione gonadica era normale al basale, c’era una probabilità del 90% di avere una normale concentrazione di Testosterone totale dopo 3 mesi di recupero e una probabilità del 100% alla fine del follow -up (in media circa 8 mesi dopo l’interruzione del ciclo).
Ma allora perché una PCT non da i risultati sperati se i SERM mostrano risultati così buoni in vari tipi di ipogonadismo?
Sfortunatamente, attualmente non sono disponibili studi di buona qualità nei quali i SERM vengano valutati come trattamento per l’ipogonadismo indotto da AAS. Principalmente il loro uso è destinato, e risultato efficace, nell’ipogonadismo dovuto ad altre cause. Di conseguenza è ovvio che bisognerebbe quindi avere cautela prima di giungere ad affrettate conclusioni, poiché attualmente non è noto quanto bene questi risultati si traducano in coloro che soffrono di ipogonadismo AAS-indotto. La causa sottostante dell’ipogonadismo è molto diversa. In linea di principio, l’ipogonadismo indotto da AAS è uno stato transitorio post-ciclo in cui l’ipotalamo e l’ipofisi non rispondono adeguatamente alla diminuzione delle concentrazioni di androgeni ed estrogeni. Dopo tutto, le concentrazioni post-ciclo di Testosterone ed Estradiolo sono di molto alterate e quindi il feedback negativo che solitamente impone all’ipotalamo e all’ipofisi il rilascio di GnRH e di LH ed FSH è notevolmente diminuito. Quindi, mentre lo stimolo (alterazione di Estradiolo e Testosterone) per produrre LH e FSH è variabilmente presente, le cellule endocrine temporaneamente non riescono a rispondere in modo adeguato a questa condizione. Non è sicuro di come l’uso di SERM possa rendere questo stimolo più marcato e aiutare nel recupero dell’HPGA. A differenza dell’ipogonadismo indotto da AAS, le popolazioni di studio sull’ipogonadismo secondario sono in uno stato stazionario di carenza di Testosterone. Qui, in quel caso, per via delle condizioni di base, avrebbe di certo senso che un soggetto possa spostare lo stato stazionario aumentando lo stimolo con un SERM per aumentare a sua volta il livello di Testosterone, ed è dimostrato. Quindi, tanto per ribadire i concetti primari quando si parla di studi, bisognerebbe essere cauti quando si traducono questi studi alla luce di una situazione ben diversa seppur simile, ossia la situazione ormonale post-ciclo.
Conclusioni e riflessioni critiche
Ricapitolando, lo studio HAARLEM è uno studio prospettico in cui è stata seguita nel tempo un’ampia coorte di utilizzatori di AAS. Diverse misurazioni, inclusi i livelli di Testosterone, sono state eseguite prima, durante e in due punti temporali dopo la cessazione dell’uso di AAS. Confrontando quei soggetti che hanno svolto una PCT con quelli che non l’hanno svolta, sono finalmente emerse alcune buone prove iniziali sulla reale efficacia della PCT. Sfortunatamente, la pratica comunemente applicata sembra essere un po’ inutile, per usare un eufemismo.
Ma quali altre critiche possono essere mosse verso questo studio? Beh, qualcuno potrebbe obbiettare che “Olivier de Hon è uno degli autori. Ed è una autorità dell’antidoping olandese”. Sì, vero, ma in che modo questo invalida i risultati esattamente? basterebbe indicare solo quale parte potrebbe essere stata influenzata da lui. Inoltre, sono sicuro che l’autorità antidoping avrebbe voluto vedere gli utilizzatori di AAS NON recuperare affatto, anche dopo 3 mesi. Ma lo hanno fatto. Sono il primo a mettere in dubbio l’onesta o meglio la lucidità di certi enti, ma sono quasi certo che avrebbero apprezzato risultati diversi da questi.
Si potrebbe anche dire che “Non hanno istruito gli utilizzatori di AAS a fare A, B e C, il che avrebbe portato a risultati migliori”. Sì, infatti è uno studio OSSERVAZIONALE, non uno studio interventistico. Se avessero istruito gli utilizzatori di AAS ad applicare determinate pratiche con i composti che stavano usando, sarebbe stato piuttosto difficile far passare la cosa al comitato etico medico in primo luogo. L’unico modo per superare l’ottenimento di un intervento è se quest’ultimo incoraggia gli utilizzatori a prendere meno AAS, o a non utilizzarli del tutto. L’obiettivo di questo studio era valutare i rischi per la salute legati all’abuso di AAS nella pratica. Una configurazione osservazionale come questa è ESATTAMENTE ciò che si vorrebbe fare in quel caso.
Un altra obbiezione potrebbe riguardare il fatto che tutti i dosaggi di AAS utilizzati non siano stati equiparati su base milligrammo per milligrammo. Ovviamente non ci sono prove che sia stato fatto diversamente. Potresti assegnare arbitrariamente qualcosa come “2mg di Testosterone = 1mg di Trenbolone” o qualsiasi altra molecola, ma sarebbe ben poco valido viste le informazioni che si hanno in materia. Cosa starebbe a significherebbe quel numero? Il Trenbolone è due volte più potente nella stimolazione dell’ipertrofia muscolare? Due volte più potente nel sopprimere l’HPGA? Due volte più potente nel causare l’acne? Da dove basi questi numeri? Medie di dosaggi degli androgeni estremamente imprecise? E in che modo questo avrebbe comunque influenzato i risultati? TUTTI gli utilizzatori hanno riscontrato una soppressione marcata dei loro livelli endogeni di Testosterone durante i loro cicli.
I soggetti potrebbero aver sbagliato la modalità delle loro PCT? Bene, in primo luogo, tornando a quanto detto in precedenza, i ricercatori non potevano dire loro di fare diversamente da quanto essi avevano previsto. E secondo, quale ricerca può dirci cosa comporta una “buon PCT”? Non ne esiste nessuna! E, sebbene la “PCT di Scally” risulti quella con il desing più logico, le prove a suo favore rimangono limitate. È per lo più tutta una ipotesi e supposizioni basate su ricerche estrapolate da popolazioni di studio con diverse cause di ipogonadismo. I soggetti di questo studio hanno semplicemente svolto una PCT come fa la maggior parte degli utilizzatori: assumere SERM per circa un mese.
Forse avrebbero dovuto iniziare la PCT più tardi? Ok, quindi che differenza ci si aspetterebbe? Il gruppo senza PCT aveva comunque gli stessi livelli di Testosterone che avevano al basale 3 mesi dopo l’ultima iniezione. Dovremmo forse aspettare 3 mesi? Sembra funzionare abbastanza bene…
I ricercatori forse hanno sbagliato a non fare una sottoanalisi basata su chi ha usato un tipo di composto e chi ne ha usato un altro? Beh, sarebbe stato alquanto arduo poterlo fare. Il motivo di ciò è che solo nel 13% dei campioni la fiala conteneva esclusivamente l’AAS che era riportato sull’etichetta e nel 47% dei casi la fiala non conteneva nemmeno l’AAS dichiarato sull’etichetta ma ne conteneva un altro (o altri).[2]
Attenzione, non sto dicendo che la PCT sia stata o sia completamente una cattiva idea. Sto semplicemente sottolineando ciò che lo studio prospettico e anni di osservazione ci suggeriscono. Anche nei casi di uso corretto di hCG durante il ciclo, uso dei SERM e hCG post ciclo secondo logica di decadenza dei livelli ematici del/gli AAS usato/i e l’inserimento di un AI quando necessariamente richiesto dagli esami ematici di controllo, la risultante è sempre soggetta a fortissime variabili legate non solo alla lunghezza del ciclo e/o al tipo di molecole usate (vedi anche tipo/i di estere) ma anche dall’età del soggetto e dal numero di cicli svolti in precedenza. Alcuni utilizzatori si attestano a livelli discreti nella metà del range di riferimento, mentre una parte non indifferente soffre per anni di variazioni estrogeno-prolattiniche con livelli di Testosterone totale verso il limite basso e il Testosterone libero sotto il limite minimo.
Non è un caso se molti utilizzatori, specie dai 30 anni in su, optino per una TRT piuttosto di tentare un recupero travagliato.
Gabriel Bellizzi
Riferimenti:
Smit, D. L., et al. “Disruption and recovery of testicular function during and after androgen abuse: the HAARLEM study.” Human Reproduction (2021).
Smit, Diederik L., et al. “Baseline characteristics of the HAARLEM study: 100 male amateur athletes using anabolic androgenic steroids.” Scandinavian journal of medicine & science in sports 30.3 (2020): 531-539.


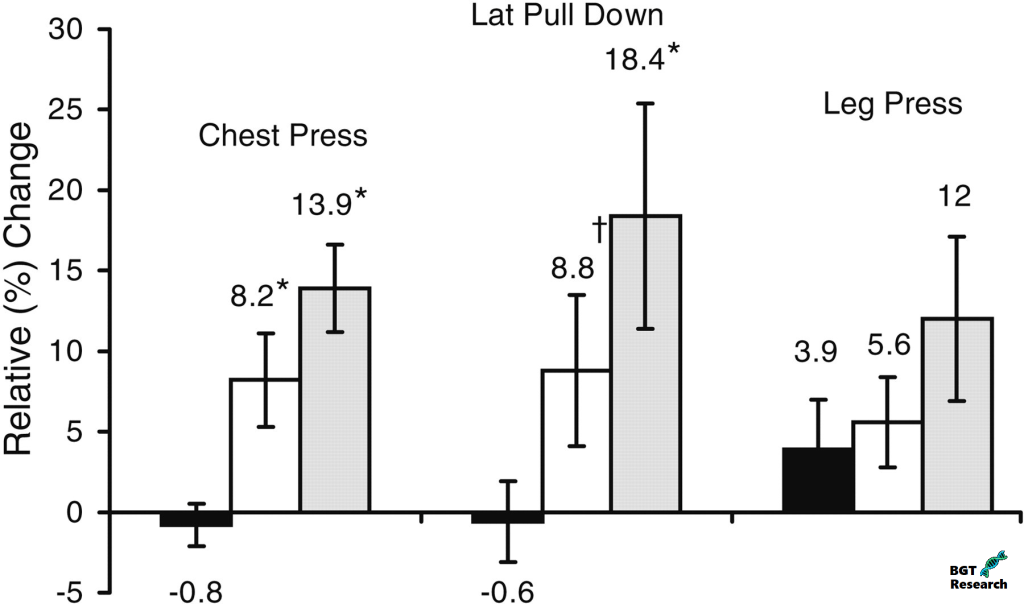

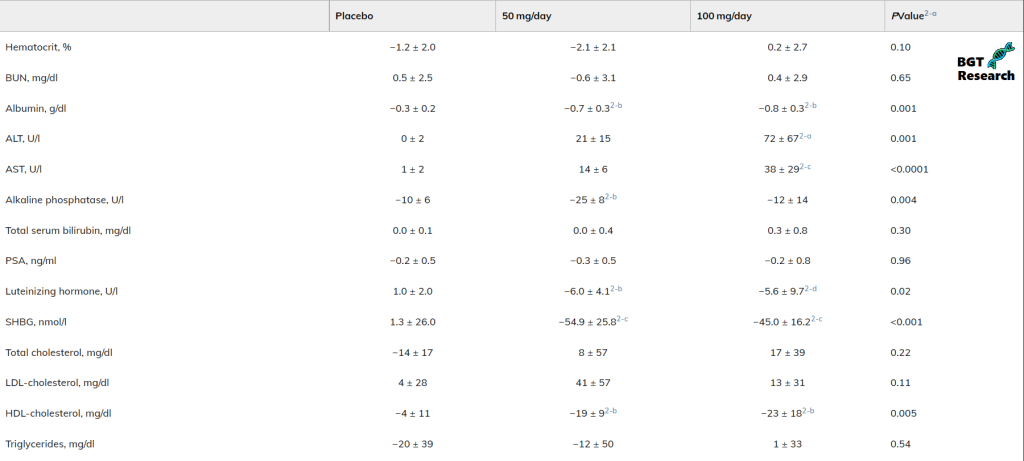

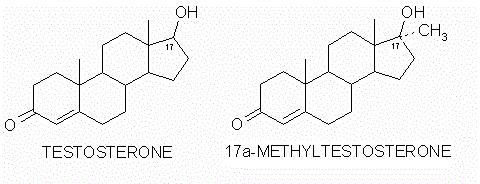
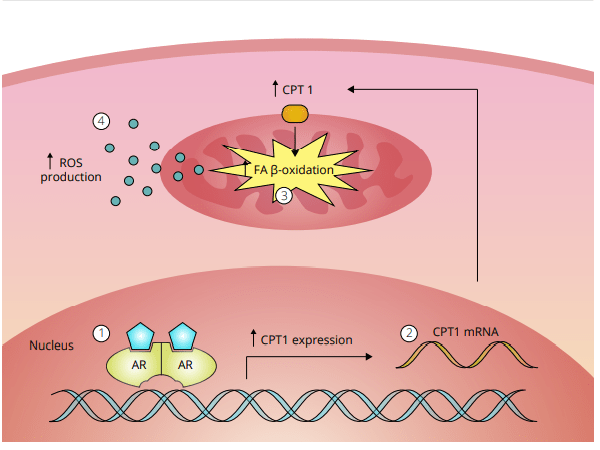


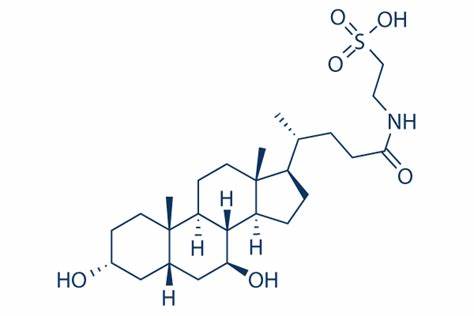
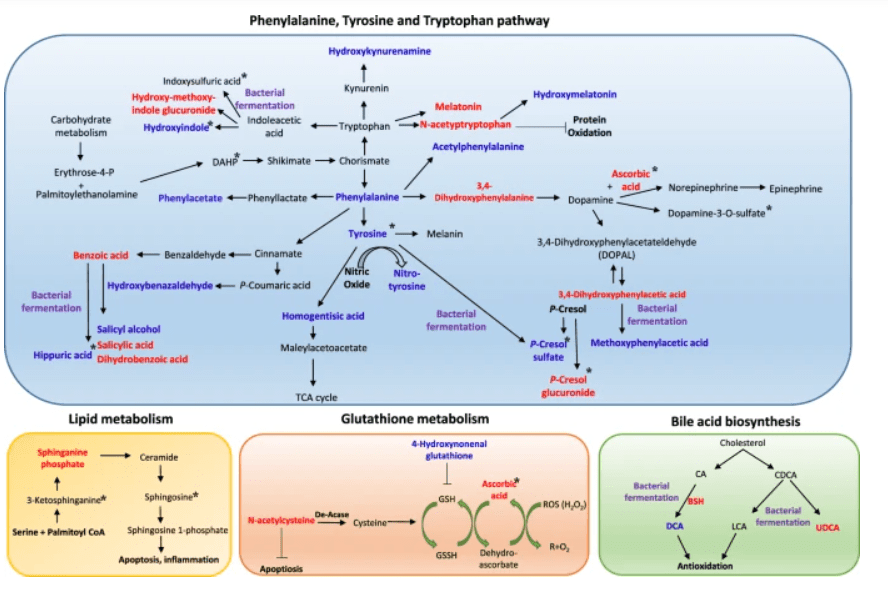
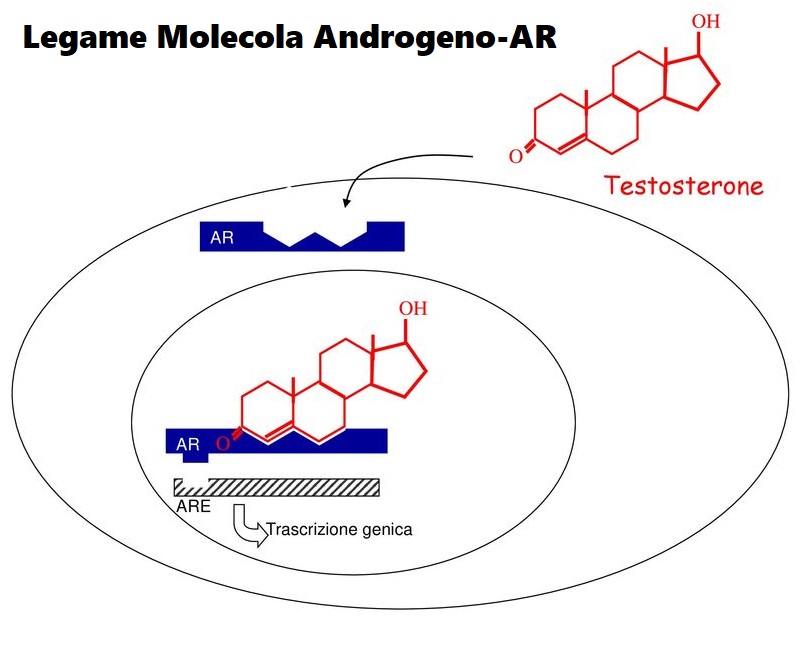
































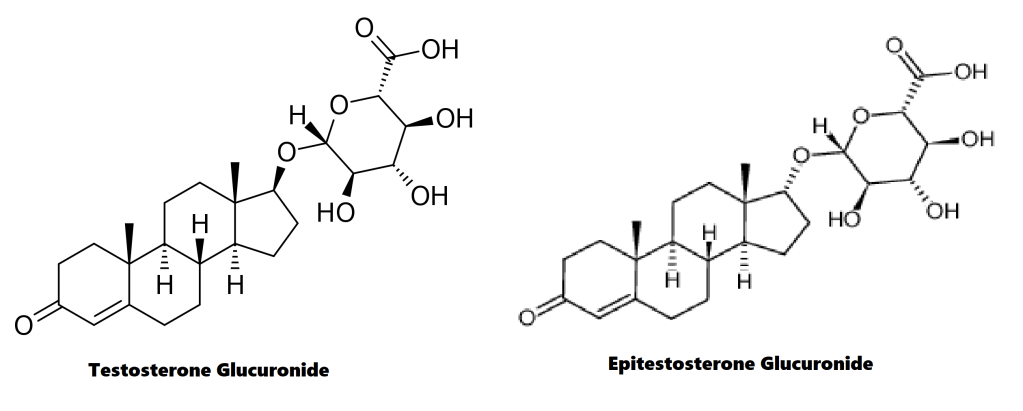





















































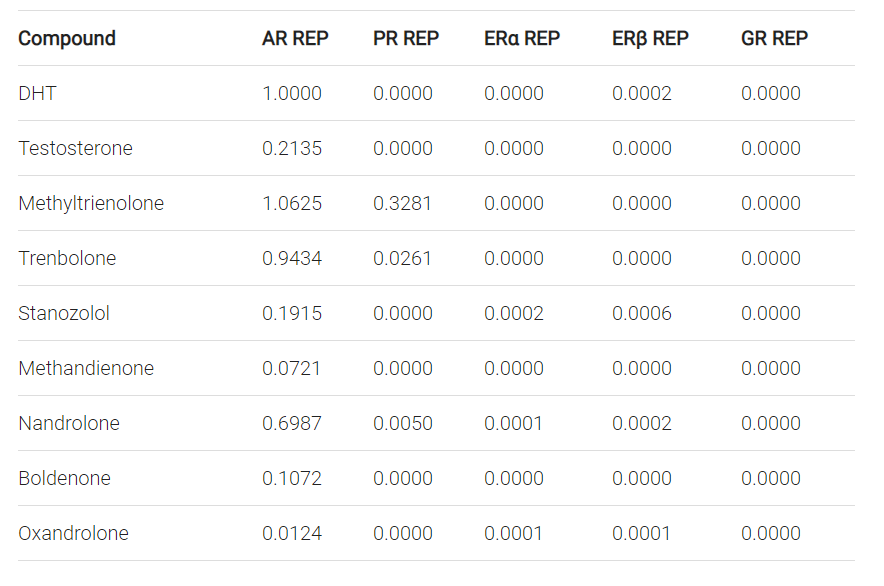



{kind=link}