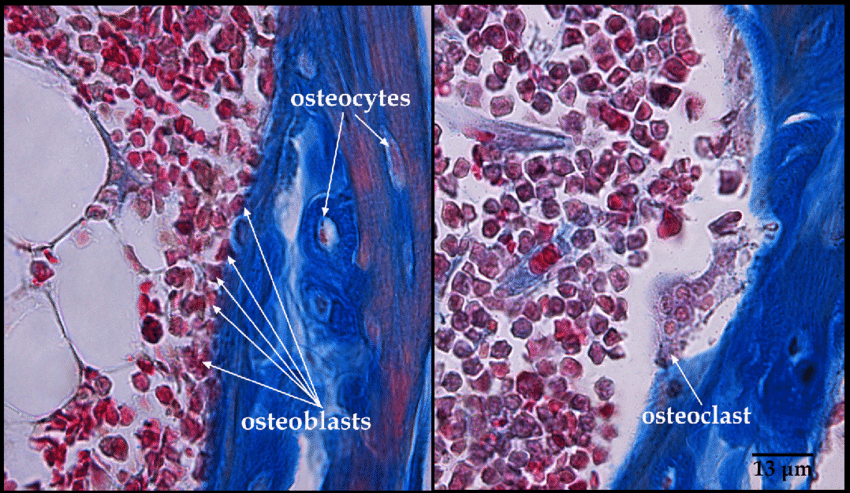
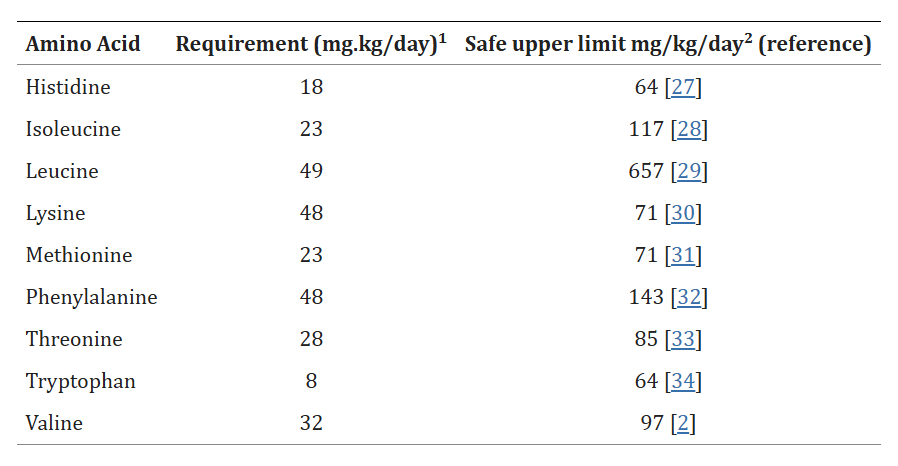
I capisaldi della gestione del peso sono gli interventi sullo stile di vita con il ruolo aggiuntivo di supporto dei farmaci anti-obesità e delle procedure bariatriche. Attualmente, i farmaci disponibili approvati per il trattamento dell’obesità agiscono sul bilancio energetico riducendo l’assunzione di cibo e il comportamento di ricompensa alimentare nel sistema nervoso centrale (ad esempio, sopprimendo l’appetito) o riducendo l’assorbimento dei grassi nell’intestino.[1,2,3] Finora non sono disponibili farmaci con un effetto diretto sull’aumento del dispendio energetico attraverso un’influenza sul tessuto adiposo [4,6]. Nell’uomo esistono due tipi di tessuto adiposo con funzioni fisiologiche distinte: il tessuto adiposo bianco (WAT), specializzato nell’immagazzinamento di trigliceridi in eccesso quando l’assunzione di energia supera il dispendio energetico, e il tessuto adiposo bruno (BAT) – con i relativi adipociti “beige”/”bruni” (derivati dal WAT) – che svolge un ruolo centrale nel metabolizzare il glucosio, gli acidi grassi e altre sostanze chimiche per produrre calore attraverso l’attivazione della proteina di disaccoppiamento 1 (UCP1) specifica del tessuto termogenico [4,7]. Alcuni dati suggeriscono che il BAT possa essere funzionale nell’uomo adulto [1]. I recettori β3-adrenergici (AR) sono espressi non solo nella vescica urinaria, ma anche sulla superficie degli adipociti bruni e bianchi [1]. I tessuti adiposi bruni e “beige”, contenenti cellule grasse termogeniche, possono essere attivati da agonisti dei recettori β3-adrenergici (β3-AR) [8]. È stato riportato che il Mirabegron, un agonista β3-AR umano selettivo, può stimolare il BAT e il processo di imbrunimento degli adipociti derivati dal WAT [9,10]. Il fatto che l’attivazione del BAT e degli adipociti “beige” possa aumentare il dispendio energetico rende i tessuti adiposi bruni e “beige” nuovi e promettenti bersagli per il trattamento dell’obesità [4,11].
Il ruolo del tessuto adiposo nella termogenesi e nei processi metabolici associati all’obesità:
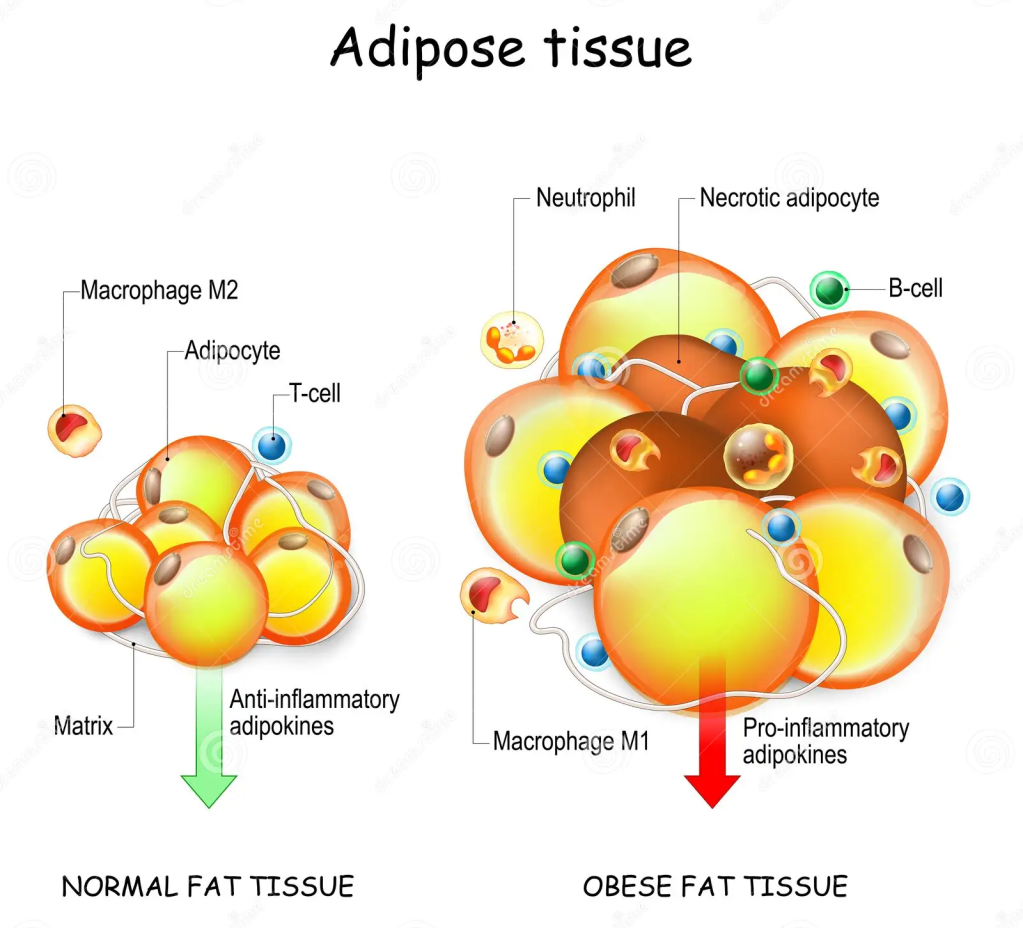
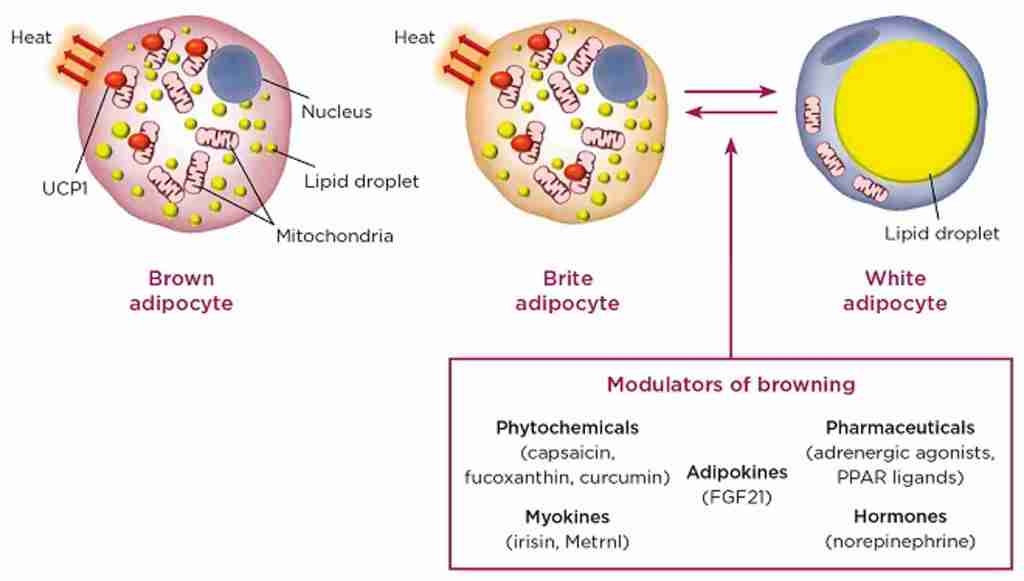
Nell’uomo esistono due tipi principali di tessuto adiposo, che svolgono funzioni diverse: il tessuto adiposo bianco e il tessuto adiposo bruno. Oltre al WAT e al BAT, sono state distinte anche cellule adipose “brune”, definite cellule adipose “beige”. Esse derivano dal WAT, ma la loro funzione metabolica è simile a quella del BAT [4,7]. Il WAT è responsabile dell’immagazzinamento di energia sotto forma di trigliceridi, del rilascio di lipidi e della funzione di ghiandola endocrina, secernendo adipochine, come l’adiponectina e la leptina, per promuovere l’omeostasi metabolica [9,12]. Nell’obesità, gli adipociti bianchi si ipertrofizzano, seguiti da fibrosi, necrosi degli adipociti e infiltrazione di cellule immunitarie, che portano a infiammazione locale e sistemica, insulino-resistenza e disfunzione metabolica [9].
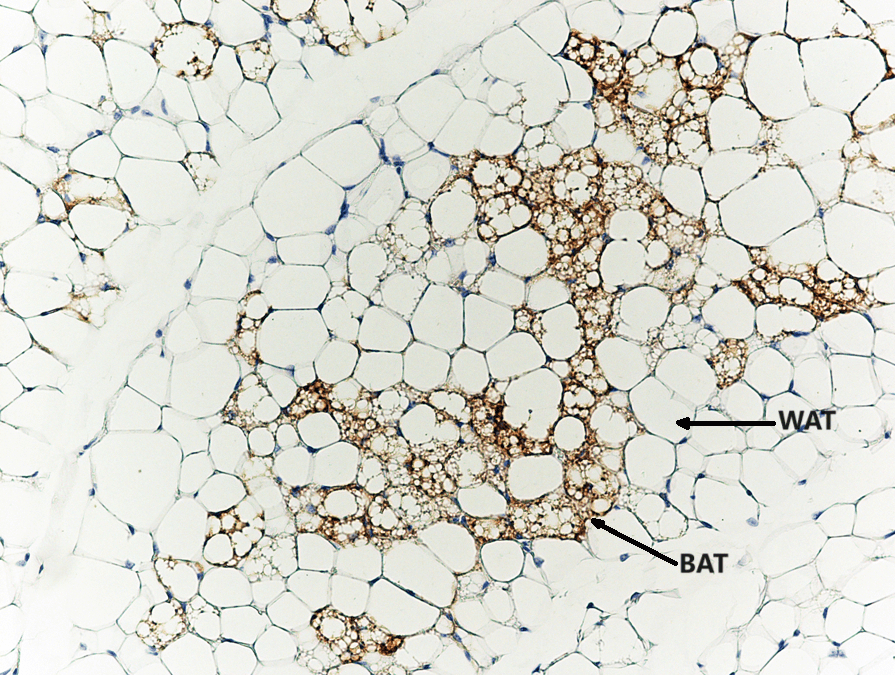
Cellule del BAT (colorate di marrone con anticorpi contro la proteina specifica del grasso bruno Ucp1) annidate tra le cellule del WAT grasso bianco.
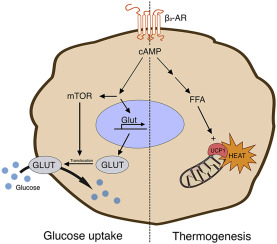
Il BAT è stato descritto per la prima volta nel 1981 in finlandesi che lavoravano all’aperto e che erano stati esposti a basse temperature ambientali [6]. Il BAT metabolicamente attivo è stato identificato negli adulti mediante imaging PET/CT focalizzato principalmente sulla fossa sopraclavicolare, sull’area succlavia e sull’ascella, seguito dalle aree mediastiniche, paraspinali, perinefriche e sopradrenali [10,12]. Sebbene il BAT sia presente nell’uomo, la sua prevalenza diminuisce con l’età e nelle persone in sovrappeso o obese rispetto ai soggetti magri [6,9,13,14]. Gli anelli mancanti nel trattamento dell’obesità sono i farmaci che possono aumentare la quantità o l’attività del BAT. È stato riportato che il volume del BAT può essere aumentato dopo la chirurgia bariatrica [12]. Il BAT è il principale organo termogenico dei mammiferi, con lo scopo di aumentare il dispendio energetico in risposta al freddo o ad altre stimolazioni nervose simpatiche, rilasciando noradrenalina dai terminali nervosi per attivare i recettori β3-adrenergici attraverso il processo definito termogenesi senza brividi [2,10,11,12,13]. La capacità termogenica del BAT è stata stimata in circa 500 W/kg [6]. Gli adipociti del BAT sono arricchiti di mitocondri (i loro livelli sono più alti di quelli del WAT), nei quali la proteina di disaccoppiamento 1 (UCP1) è altamente espressa. La UCP1 dissipa l’energia in eccesso sotto forma di calore in un processo noto come termogenesi [2,15]. L’attivazione adrenergica della lipolisi stimola l’attività termogenica della UCP1 [2,10]. L’attivazione dell’UCP1 sulla membrana mitocondriale interna disaccoppia la respirazione mitocondriale, separando il trasporto di elettroni dalla produzione di ATP per ossidare il substrato e generare calore [4,8,16]. Gli acidi grassi a catena lunga, generati dai pool lipidici intracellulari, sono trasportati ai mitocondri attraverso la carnitina palmitoiltransferasi 1 (CPT1) e utilizzati come fonte di carburante dagli adipociti bruni per produrre calore. Inoltre, è stato proposto che gli acidi grassi liberi agiscano come attivatori allosterici di UCP1. Oltre agli acidi grassi, anche il glucosio circolante può essere utilizzato dal BAT attivo per alimentare la termogenesi [2,10,17]. In sintesi, il BAT consuma glucosio e lipidi per generare calore attraverso la respirazione disaccoppiata mediata da UCP1, con conseguente miglioramento dell’omeostasi glucidica e lipidica [9,13,18].
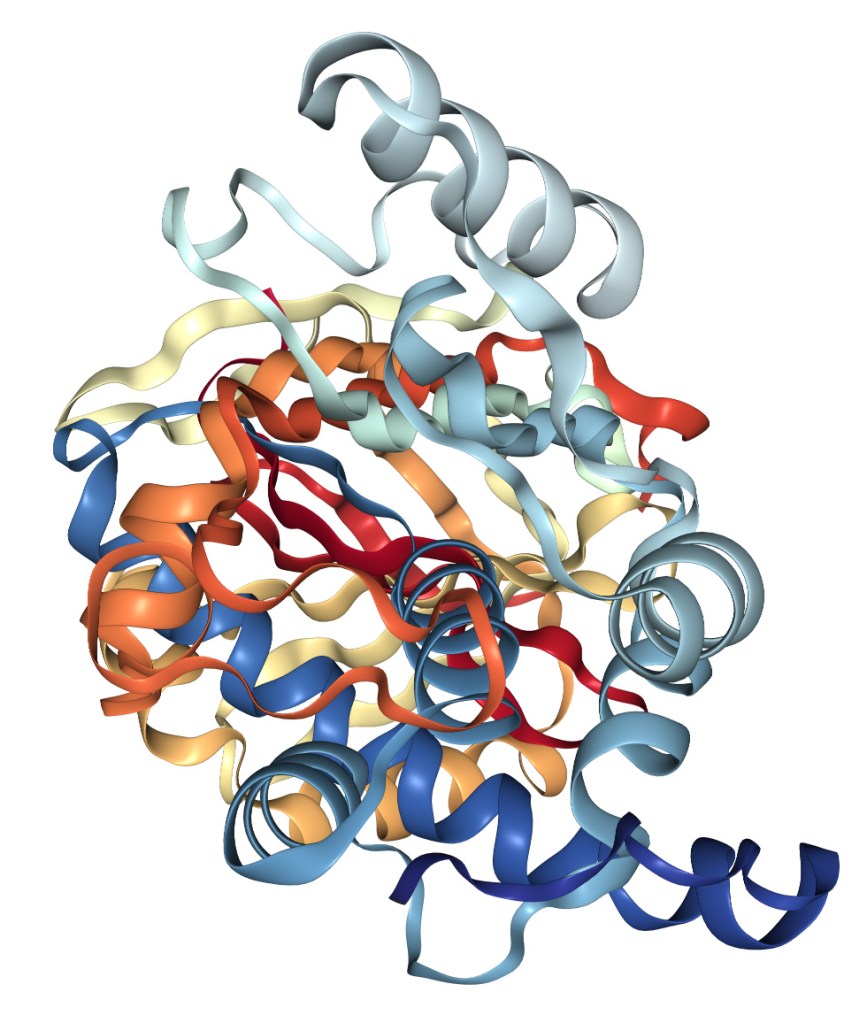
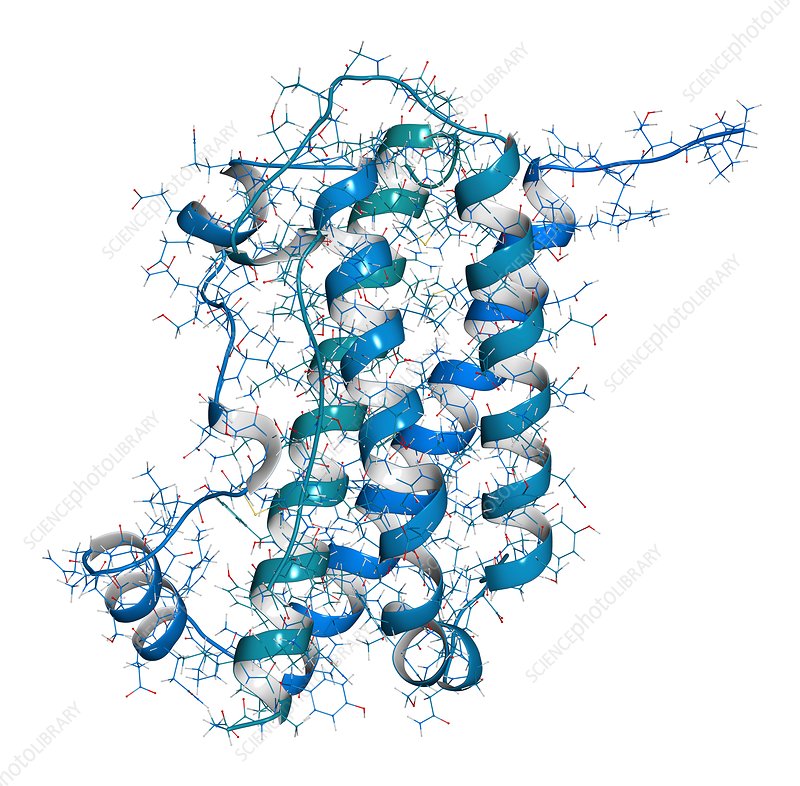
Struttura della proteina disaccoppiante umana UCP1
Gli adipociti termogenici umani possono originare da due lignaggi distinti, non solo da adipociti bruni costitutivi ma anche da cellule “beige” reclutabili, definite adipociti “bruni” o “bruno-simili” [5]. Gli adipociti “beige” sono localizzati prevalentemente nei depositi di WAT [16]. Le cellule adipose del WAT possono essere convertite in adipociti “beige” termogenici in un processo chiamato “browning” o “beiging” [12]. Da un lato, è stato dimostrato un sostanziale “beiging” del WAT sottocutaneo umano in alcuni disturbi, come la cachessia da cancro, le ustioni e le condizioni con alti livelli di catecolamine, ad esempio il feocromocitoma [8,13,14]. I pazienti con tumori che secernono catecolamine hanno anche più tessuto adiposo bruno rispetto alla maggior parte delle persone [19]. D’altra parte, gli adipociti “bruni” possono essere attivati con l’induzione dell’espressione di UCP1 da parte di stimoli ambientali, come l’esposizione al freddo e agli agonisti β-adrenergici, mediata dalla via di segnalazione p38-MAPK [9,12,14,16]. La risposta “beiging” dei soggetti obesi al freddo è simile a quella dei soggetti magri [14]. Sebbene queste cellule differiscano dagli adipociti bruni convenzionali – in quanto si sviluppano da una cellula precursore di adipociti bianchi e non da una cellula precursore di adipociti bruni, simile agli adipociti bruni classici nel BAT – gli adipociti “beige” possiedono goccioline lipidiche multiloculari, un gran numero di mitocondri e marcatori unici di espressione genica del grasso bruno, come UCP1, aumentando la capacità del tessuto di ossidazione del carburante e il dispendio energetico [4,16,18]. Inoltre, è stato dimostrato che il “beiging” è associato a una riduzione della fibrosi del tessuto adiposo e della disfunzione adiposa. Questi risultati suggeriscono che l’induzione del tessuto adiposo “beige” può migliorare l’omeostasi metabolica aumentando la capacità del WAT sottocutaneo di funzionare come serbatoio metabolico per il glucosio e i lipidi o riducendo la disfunzione del WAT che si verifica con l’obesità [13]. Così, oltre alla termogenesi e al dispendio energetico, i tessuti adiposi bruni e “beige” sono associati a un miglioramento dell’omeostasi del glucosio e dei lipidi, nonché a una maggiore sensibilità all’insulina nell’uomo e nel topo [14].
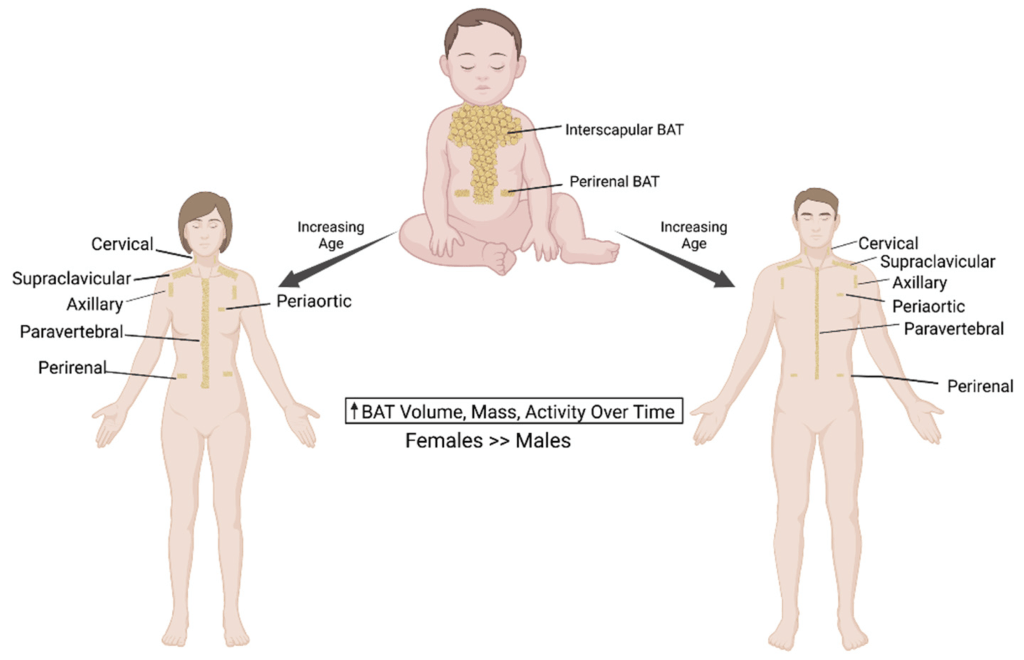
Distribuzione del BAT nei neonati, nelle donne e negli uomini. Il BAT è immagazzinato in un deposito interscapolare separato nei neonati che perdono il loro tessuto adiposo bruno con l’avanzare dell’età. Negli esseri umani adulti, la maggior parte degli adipociti bruni si trova nei depositi di BAT sopraclavicolari nella regione del collo. Quantità minori di BAT si trovano nell’aorta, nelle vertebre, nelle aree ascellari e renali. C’è una distribuzione simile del tessuto adiposo bruno sia nelle donne che negli uomini. Tuttavia, le donne hanno una maggiore quantità di massa e attività di BAT.
Considerando il fatto che nelle persone obese adulte c’è meno BAT rispetto ai soggetti magri, il WAT in eccesso, che può essere stimolante e in fase di “beigezzazione/imbrunimento”, può svolgere un ruolo aggiuntivo rispetto al BAT nei processi metabolici [13]. Pertanto, il tessuto adiposo bruno e il tessuto adiposo “beige” sono stati riconosciuti come regolatori critici del metabolismo e del dispendio energetico dell’intero corpo e sono considerati bersagli promettenti per la terapia anti-obesità [2,12,15].
L’Irisina è un ormone sintetizzato in grande quantità dal tessuto muscolare umano durante le attività sportive. La molecola è in grado di operare il meccanismo molecolare detto “browning” [“imbrunimento”], ovvero di conversione del WAT in BAT.
Il grasso bruno, il grasso “beige” e i β3-adrenocettori nel contesto dell’obesità:
Struttura del recettore β3-adrenergico
La famiglia dei recettori β-adrenergici (AR) umani è composta dai recettori β1, β2 e β3, in cui il β1-AR è altamente espresso in tutto il sistema cardiovascolare, il β2-AR si trova nelle vie aeree polmonari, in tutta la vascolarizzazione e nel muscolo scheletrico e l’espressione del β3-AR è limitata soprattutto alla vescica urinaria e alla cistifellea, oltre che al BAT e al WAT [4,10]. Il β3-AR umano, identificato nel 1989, è un recettore a 7 membrane, con una coda N-terminale extracellulare e una coda C-terminale intracellulare, composta da 408 aminoacidi. Si accoppia principalmente a Gs per attivare l’adenilato ciclasi, con conseguente aumento dei livelli intracellulari di cAMP, sebbene sia stato riportato un accoppiamento promiscuo con altri effettori, come Gi [4,16].
La β3-AR svolge un ruolo critico nel tessuto adiposo, nella regolazione della termogenesi, della glicolisi e della lipolisi [16]. Studi sugli animali hanno dimostrato che la stimolazione cronica del BAT porta a un miglioramento della tolleranza al glucosio e della sensibilità all’insulina e a una riduzione dell’obesità, oltre che al rilascio di adipochine che regolano beneficamente il metabolismo [1,8,10,12,20]. Inoltre, l’attivazione β3-AR-mediata del WAT può aumentare la secrezione insulinica delle cellule β pancreatiche [5]. È stato anche riportato che una parte significativa della termogenesi non da brivido ha luogo nel tessuto adiposo bruno ed è mediata principalmente dal β3-adrenocettore [19]. Nei topi alimentati con dieta a base di chow e ad alto contenuto di grassi, il trapianto di BAT ha ridotto il peso corporeo, aumentato il metabolismo del glucosio e la sensibilità all’insulina e incrementato l’assorbimento di glucosio nel BAT e nel WAT [15].
Oltre alla funzione metabolica, la β3-AR svolge un ruolo nel cervello, essendo coinvolta nei processi di memoria, apprendimento e regolazione dell’appetito, nel tratto gastrointestinale, dove partecipa alla regolazione della motilità, e nel sistema genitourinario, dove svolge un ruolo nella regolazione della funzione vescicale [16].
Durante la termogenesi, i β3-adrenocettori aumentano il dispendio energetico, che può portare alla perdita di grasso, in risposta alla stimolazione simpatica [19]. È dimostrato che la stimolazione cronica dell’attività nervosa simpatica e dei β3-AR può attivare il BAT [6]. È stato dimostrato che l’esposizione al freddo stimola il sistema nervoso simpatico a rilasciare noradrenalina dalle terminazioni nervose simpatiche per attivare i β-AR sulle membrane delle cellule del BAT, promuovendo la termogenesi. In questo modo, il BAT umano è in grado di avviare la termogenesi attraverso il consumo di acidi grassi e glucosio e, successivamente, di generare calore [1,3,8]. Inoltre, l’attivazione dei β3-ARs da parte dell’esposizione al freddo o di agenti farmacologici induce un programma di “beiging” nel WAT [18]. Un modo per aumentare la quantità effettiva di tessuto adiposo bruno può essere quello di somministrare l’agonista β3-adrenoccettore in modo cronico [19]. Una singola dose di agonista dei β3-adrenocettori può almeno raddoppiare il dispendio energetico in un modello murino a circa 21 °C [19].
Struttura del gene ADRB3
Il ruolo dei β3-AR nel metabolismo energetico umano è supportato da studi clinici che riportano associazioni tra polimorfismi specifici nel gene umano ADRB3 (il gene che codifica i β3-AR) e tassi più elevati di obesità, insulino-resistenza e diabete [10]. Inoltre, le mutazioni nel gene ADRB3 sono state correlate all’insulino-resistenza, all’aumento del rischio di obesità e diabete e alla malattia del fegato grasso non alcolico negli individui obesi [10]. I dati indicano che il silenziamento di ADRB3 negli adipociti umani “marroni”/”beige” altera il macchinario termogenico cellulare e causa una riduzione dei livelli di espressione dei geni associati al metabolismo degli acidi grassi, alla massa mitocondriale e alla termogenesi, senza compromettere il fenotipo “marrone”/”beige” [10].
Attività agonista dei β3-AR del Mirabegron:
Struttura molecolare del Mirabegron
Mirabegron è una nuova generazione di agonisti dei β3-adrenocettori con una buona biodisponibilità [21]. Gli effetti dell’agonista selettivo dei β3-AR mirabegron sul rilassamento della vescica sono stati scoperti nel 2007. Per la prima volta, la selettività β3 del mirabegron (YM-178) nel contesto della funzione vescicale è stata descritta da Takasu et al. [22]. YM-178 ha aumentato l’accumulo di AMP ciclico in cellule ovariche di criceto cinese che esprimono il β3-adrenocettore umano. Mirabegron ha dimostrato valori di EC50 nanomolari contro il β3-AR umano in saggi biochimici, con una potente selettività rispetto ai β1- e β2-AR [22]. Studi in vivo hanno dimostrato che la somministrazione di mirabegron ha ridotto la pressione intravescicale e le contrazioni spontanee della vescica in modo dose-dipendente [23]. Mirabegron è stato approvato dalla Food and Drug Administration (FDA) statunitense nel 2012 come nuovo tipo di trattamento farmacologico per la vescica iperattiva (OAB) [6,21,24]. Cinquanta milligrammi di mirabegron è la dose raccomandata a tutti i pazienti con OAB [24]. Il farmaco è generalmente ben tollerato e gli effetti collaterali più comuni includono ipertensione, rinofaringite e infezione del tratto urinario [6].
La selettività β3 di Mirabegron è stata confermata in molti studi con l’uso di linee cellulari che esprimono il β3-adrenocettore sia animale che umano [22,23,25]. Mirabegron ha mostrato una selettività per il β3-AR umano superiore di oltre 400 volte rispetto al β1-AR o al β2-AR umano [26]. Ad esempio, Brucker et al. [27] hanno utilizzato cellule di ovaio di criceto cinese (CHO)-K1, cellule di rene embrionale umano 293 esprimenti stabilmente recettori β1-, β2- o β3-adrenergici umani e recettori α1D- e α2B-adrenergici umani per valutare la selettività di mirabegron. A una concentrazione di 10 μM, l’attività β3-adrenergica rispetto all’isoproterenolo (agonista β-adrenergico completo) era dell’88% per mirabegron. A sua volta, l’attività β1- e β2-adrenergica di mirabegron era rispettivamente del 3% e del 15% [27]. In questo studio mirabegron non ha soddisfatto il criterio di significatività per l’inibizione dei recettori α1D- o α2B-adrenergici [27]. Tuttavia, alcuni studi hanno indicato che mirabegron potrebbe svolgere un ruolo come antagonista degli α1-adrenergici [28,29]. Alexandre et al. [28] hanno ipotizzato che mirabegron rilassasse la muscolatura liscia uretrale nei topi attraverso un duplice meccanismo che coinvolge l’attivazione dei β3-adrenocettori e il blocco degli α1-adrenocettori. In un altro studio, mirabegron ha indotto una vasorilassazione endotelio-indipendente nelle arterie del tessuto adiposo viscerale attraverso l’antagonismo degli α1-adrenocettori. Questa azione ha suggerito che mirabegron potrebbe migliorare efficacemente la perfusione del tessuto adiposo viscerale, favorendo così un sano rimodellamento del tessuto adiposo e prevenendo alcune delle conseguenze cardiometaboliche indesiderate dell’obesità e dell’invecchiamento [29]. Resta ancora difficile stabilire in che misura l’antagonismo degli α1-adrenocettori possa contribuire agli effetti clinici di mirabegron [28,29].
I cambiamenti metabolici benefici causati dal trattamento cronico con mirabegron potrebbero derivare dalla stimolazione della β3-AR nel BAT e nel WAT umani [5,12,18]. È stato suggerito che mirabegron potrebbe migliorare le malattie metaboliche legate all’obesità aumentando la termogenesi del BAT, la lipolisi del WAT e la stimolazione del processo di “brunimento” degli adipociti derivati dal WAT [4,5,9,10]. Il trattamento acuto con mirabegron ha aumentato il dispendio energetico [10,15]. Dopo il silenziamento dell’espressione dei β3-AR, il mirabegron non è stato in grado di stimolare la lipolisi e la termogenesi del BAT [10]. Molti studi hanno dimostrato che il trattamento con mirabegron ha aumentato l’assorbimento del glucosio negli adipociti bruni e “beige”, ha migliorato l’omeostasi del glucosio e ha aumentato la sensibilità all’insulina e la funzione delle cellule β [1,9]. Inoltre, è stato dimostrato che il trattamento cronico con agonisti β3-AR nell’uomo può rilasciare adipochine benefiche [1]. Il modo in cui mirabegron migliora il metabolismo del glucosio non è stato finora chiarito [5]. Tuttavia, sono stati ipotizzati alcuni meccanismi. In primo luogo, mirabegron stimola la secrezione di adiponectina, nota adipochina derivata dal WAT e associata a una maggiore sensibilità all’insulina nel muscolo scheletrico e nel fegato. In secondo luogo, mirabegron aumenta la concentrazione di polipeptide inibitore gastrico (GIP), l’incretina collegata alla secrezione di insulina. Infine, il meccanismo di mirabegron potrebbe coinvolgere le stesse cellule β [5].
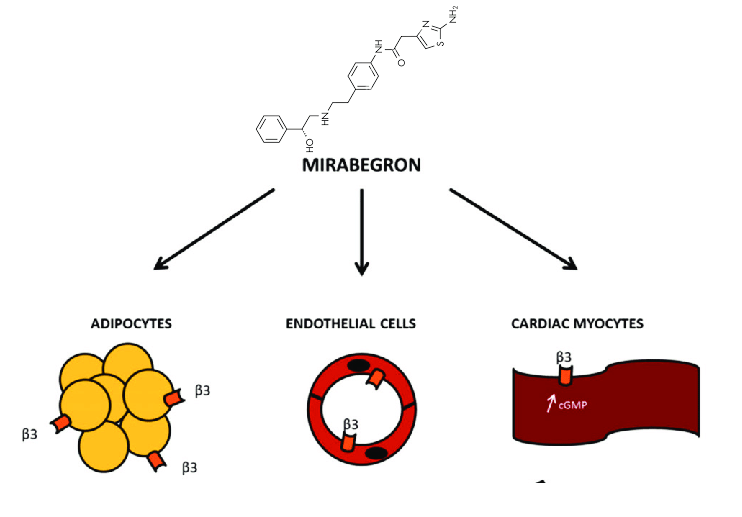
L’agonista dei recettori β3-adrenergici è un ottimo candidato per il trattamento dell’obesità, poiché l’isoforma β3 è espressa esclusivamente negli adipociti e l’azione su altri tipi di cellule, come i cardiomiociti e le cellule muscolari lisce, attraverso le altre isoforme β – β1 e β2 – è minima e dose-dipendente [11]. Pertanto, come agonista β3-AR, Mirabegron attiverebbe la termogenesi nel tessuto adiposo, stimolando l’ossidazione dei lipidi e il consumo di glucosio per produrre calore, senza causare gravi effetti collaterali cardiovascolari [13].
Mirabegron come agente antiobesità negli studi sperimentali:
Adipocita del BAT
Il trattamento dei roditori con agonisti β3-AR ha attivato il BAT, con conseguente aumento del dispendio energetico, perdita di peso e miglioramento del metabolismo del glucosio e dei lipidi. Inoltre, ha ripristinato l’equilibrio NO/redox, migliorato la funzione endoteliale e, quindi, esercitato effetti protettivi vascolari [4,6,13,17]. L’aumento dell’attività del BAT ha impedito lo sviluppo e la gravità dell’obesità e del diabete di tipo 2, mentre i topi privi di BAT erano inclini all’obesità [16]. È stato riportato che una riduzione della massa del BAT nei topi indotta da un transgene produce obesità e che questi topi presentano un’ulteriore maggiore suscettibilità all’obesità a causa di diete obesitogene [8,30,31].
Adipocita “Beige”
Come si è detto, il Mirabegron può essere efficace come attivatore del BAT, stimolatore delle cellule “beige” e controllore dell’omeostasi metabolica. L’influenza benefica di mirabegron sul metabolismo è stata confermata da studi in vitro e in vivo [2,4,15,18]. Nello studio condotto da Dehvari et al. [15], sono stati riportati gli effetti di mirabegron negli adipociti bruni, bianchi e “beige” in vitro e i suoi effetti sull’utilizzo del glucosio e sulla termogenesi in vivo. È stato dimostrato che mirabegron aumenta l’assorbimento di glucosio e la glicolisi negli adipociti bruni di topo in vitro e promuove l’assorbimento di glucosio nel BAT in vivo. Il mirabegron ha aumentato i livelli di cAMP e l’mRNA di UCP1, con conseguente aumento del consumo di ossigeno mediato da UCP1, nonché l’assorbimento di glucosio e la glicolisi cellulare negli adipociti bruni e “beige” (tale azione è mancata nelle colture cellulari primarie di adipociti bruni provenienti da topi knockout per il β3-adrenocettore), mentre questi effetti erano assenti o ridotti negli adipociti bianchi. In vivo, mirabegron ha aumentato il consumo di ossigeno nell’intero corpo e l’assorbimento di glucosio nel tessuto adiposo bruno e bianco inguinale e ha migliorato la tolleranza al glucosio. Nei topi knockout per il β3-adrenorecettore, mirabegron non è riuscito a indurre l’assorbimento di glucosio nel tessuto adiposo, né ad aumentare il consumo di ossigeno corporeo, il che dimostra che la segnalazione del β3-adrenorecettore è una via principale delle azioni metaboliche di mirabegron [15]. Analogamente a Dehvari et al. [15], Hao et al. [4] hanno studiato gli effetti anti-obesità di mirabegron utilizzando modelli in vitro e in vivo. In entrambe le linee cellulari – preadipociti bruni di topo e preadipociti bianchi 3T3-L1 – mirabegron ha stimolato l’espressione di UCP1. I topi trattati con mirabegron, alimentati con una dieta ad alto contenuto di grassi, presentavano una riduzione del peso corporeo e dell’adiposità, nonché un miglioramento della tolleranza al glucosio e della sensibilità all’insulina. Le goccioline lipidiche nel BAT dei topi trattati con mirabegron erano meno numerose e di dimensioni inferiori rispetto ai controlli. La colorazione H&E e l’immunoistochimica hanno indicato che mirabegron ha aumentato l’abbondanza di cellule “beige” nel WAT [4]. Si è concluso che mirabegron ha aumentato l’espressione di UCP1 e ha promosso la “brunitura” del WAT, che è stata accompagnata da un miglioramento della tolleranza al glucosio, della sensibilità all’insulina e della prevenzione dell’obesità indotta da una dieta ad alto contenuto di grassi [4]. In un altro studio su animali, Valgas da Silva et al. [18] hanno riferito che un trattamento di 2 settimane con mirabegron ha ridotto l’infiammazione, migliorato il metabolismo, impedito l’accumulo di grasso ectopico nel BAT e nel fegato e diminuito l’insulino-resistenza nei topi obesi (riduzione dell’indice HOMA e dei livelli di insulina). Mirabegron ha aumentato l’espressione di UCP1 nel BAT e il dispendio energetico, oltre a ridurre l’adiposità nei topi obesi. Inoltre, mirabegron ha ridotto i livelli circolanti di acidi grassi liberi, glicerolo e TNF-α. È noto che l’aumento dei livelli di FFA circolanti causa insulino-resistenza negli organi bersaglio dell’insulina ed è emerso come uno dei principali collegamenti tra l’obesità e lo sviluppo della sindrome metabolica. È noto anche che il TNF-α ha un effetto lipolitico, che determina un aumento dei livelli di FFA e glicerolo in circolo, contribuendo all’insulino-resistenza. Tuttavia, a differenza dello studio condotto da Dehvari et al. non sono stati riscontrati cambiamenti nel WAT inguinale: il mirabegron non ha indotto il “beiging” del WAT inguinale dei topi obesi. Inoltre, l’obesità indotta dalla dieta ha aumentato significativamente i depositi lipidici nel fegato e nel BAT, ma mirabegron ha parzialmente invertito questi cambiamenti, il che potrebbe indicare un ruolo protettivo di mirabegron nello sviluppo della steatosi epatica e dell’insulino-resistenza [18]. La conferma che mirabegron può essere utile come agente anti-obesità è stata trovata anche nello studio di Hao et al. [4]. È stato dimostrato che mirabegron provoca un aumento di 14 volte dell’espressione genica di UCP1 e può determinare una perdita di peso del 12% e una riduzione dell’adiposità nei topi obesi rispetto all’attività fisica.
Struttura molecolare della Metformina
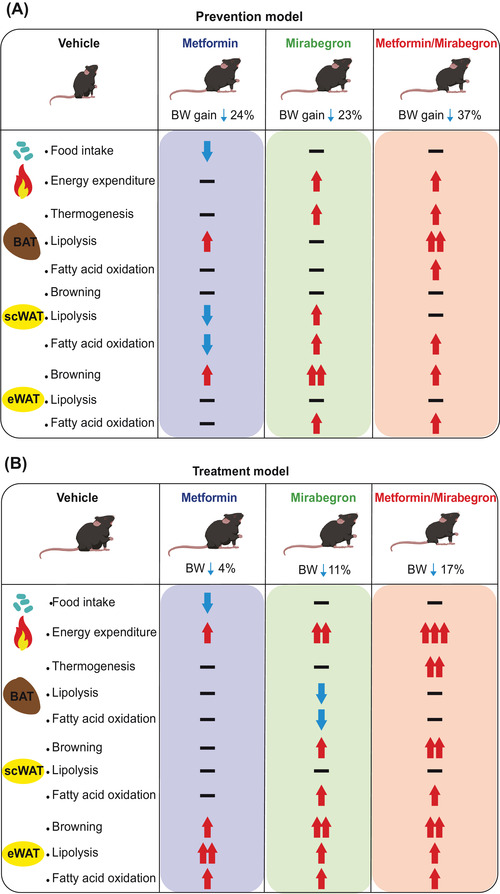
La terapia combinata, composta da Mirabegron e Metformina, è stata verificata nel modello murino di prevenzione e nel modello murino di trattamento dell’obesità [2]. La metformina, un derivato della biguanide, è uno dei farmaci più comunemente utilizzati per il trattamento del diabete di tipo 2. Inibisce il complesso mitocondriale I, vitale per il trattamento dell’obesità. Inibisce il complesso mitocondriale I, vitale per il trasporto di elettroni, che porta all’attivazione dell’AMPK (proteina chinasi attivata dall’adenosina 5′-monofosfato). Di conseguenza, la produzione di ATP (adenosina trifosfato) diminuisce e la concentrazione intracellulare di ADP (adenosina difosfato) aumenta. Di conseguenza, i livelli cellulari di AMP (adenosina monofosfato) aumentano, attivando infine l’AMPK. L’AMPK è un regolatore chiave di numerose vie metaboliche, tra cui il metabolismo del glucosio e dei lipidi e l’omeostasi energetica. La metformina svolge anche un ruolo importante inibendo la segnalazione dei recettori dell’insulina e dell’IGF, con conseguenti cambiamenti nell’omeostasi metabolica [32]. Zhao et al. [2] hanno indicato che questa terapia complessa potrebbe essere un approccio promettente per la prevenzione e il trattamento dell’obesità, agendo contemporaneamente sull’assunzione e sul dispendio energetico, senza effetti collaterali sulla funzione cardiovascolare. Nel modello di prevenzione, metformina e mirabegron hanno provocato un’ulteriore riduzione del 12% e del 14% dell’aumento di peso corporeo indotto da una dieta ad alto contenuto di grassi, rispetto a metformina o mirabegron da soli, rispettivamente. Nel modello di trattamento, metformina e mirabegron hanno promosso in modo additivo una perdita di peso corporeo del 17% nei topi obesi indotti dalla dieta, superiore del 13% e del 6% rispetto a metformina e mirabegron da soli, rispettivamente. La terapia combinata ha avuto un effetto additivo sulla perdita di peso nei topi, associato a una significativa perdita di grasso, soprattutto nel WAT sottocutaneo [2]. I ricercatori hanno suggerito che l’effetto additivo di metformina e mirabegron sull’aumento del dispendio energetico abbia contribuito in modo determinante alla riduzione del peso corporeo e della massa grassa nei topi [2]. La terapia con metformina e mirabegron ha avuto un effetto additivo sulla termogenesi del BAT e sulla doratura del WAT sottocutaneo. La terapia combinata ha aumentato significativamente l’espressione di UCP1 nel BAT e nel WAT sottocutaneo [2]. Inoltre, metformina e mirabegron hanno migliorato la tolleranza al glucosio e la sensibilità all’insulina, e l’effetto era indipendente dall’assunzione di cibo. Tuttavia, la co-somministrazione di metformina e mirabegron non ha migliorato l’omeostasi del glucosio nei topi in misura maggiore rispetto alla metformina o al mirabegron da soli [2].
Un diagramma che riassume gli effetti combinati di Metformina (Met)/Mirabegron (Mir) sull’obesità nei modelli di prevenzione e trattamento. (A) Nel modello di prevenzione in cui una dieta ricca di grassi (HFD) e farmaci venivano somministrati simultaneamente, il trattamento con Met/Mir ha ridotto l’aumento di peso in modo additivo. Ciò è dovuto principalmente a un miglioramento della spesa energetica (EE) che era accompagnato da un’espressione sovraregolata di marcatori critici nella lipolisi, nell’ossidazione degli acidi grassi e nella termogenesi nel tessuto adiposo bruno (BAT). (B) Nel modello di trattamento, è stato prima stabilito un fenotipo di obesità indotta dalla dieta (DIO), seguito da 5 settimane di trattamenti terapeutici con Met e/o Mir. Il trattamento con Met/Mir ha causato una marcata perdita di peso, derivante dall’aumento di EE
E’ interessante notare che la p-Sinefrina, una agonista selettivo dei β3-Adrenocettoiri di origine naturale, ha mostrato in studi su animali effetti positivi sull’imbrunimento del WAT, sopprimendo così l’obesità e la steatosi epatica.
Mirabegron come farmaco antiobesità: i dati degli studi sull’uomo:
Oltre agli studi sperimentali, esistono numerosi studi clinici in cui è stata dimostrata l’influenza di mirabegron sull’attività del BAT e sulla massa corporea. Gli autori hanno riferito che mirabegron ha portato a un aumento dell’attività del BAT e del dispendio energetico a riposo [1,3,5,10,17,21]. Prove preliminari suggeriscono che gli effetti del mirabegron sul metabolismo del glucosio, sul colesterolo HDL e sugli acidi biliari assomigliano a quelli ottenuti con un lieve esercizio fisico [1,5]. Nel primo gruppo di studi sono state testate soprattutto dosi elevate di mirabegron (100 mg, 150 mg o 200 mg) [1,3,5,17,21].
Cypess et al. [1] hanno usato, per la prima volta, Mirabegron per studiare il BAT umano e hanno confrontato la sua azione in un grado che corrispondeva alle risposte all’esposizione al freddo. La somministrazione di 200mg al giorno di Mirabegron orale per 12 settimane a 12 uomini sani è stata associata a una maggiore attività del BAT (misurata tramite tomografia a emissione di positroni 18F-fluorodesossiglucosio combinata con tomografia computerizzata) e all’aumento del tasso metabolico a riposo di 203 ± 40 kcal/die, rispetto agli individui che hanno ricevuto il placebo. È stato ipotizzato che la perdita di peso calcolata, associata al dispendio energetico, dovrebbe raggiungere i 5 kg nel primo anno e i 10 kg entro la fine dei 3 anni [1]. In questo studio, il dosaggio di 200 mg di mirabegron, una dose molto più alta di quelle attualmente approvate per ridurre i sintomi della vescica iperattiva, è stato generalmente ben tollerato, anche dopo 12 settimane di somministrazione orale giornaliera [1]. L’effetto collaterale più comune era la tachicardia [1].
L’alta dose di Mirabegron [100mg al giorno] è stata testata da O’Mara et al. durante un programma di terapia di 4 settimane su 14 donne sane di varie etnie [5]. Nell’endpoint primario, i ricercatori hanno riferito che la terapia cronica con Mirabegron ha aumentato il volume del BAT e l’attività metabolica, misurati tramite PET/CT con 18F-fluorodesossiglucosio [5]. Inoltre, le donne che avevano avuto principalmente meno BAT hanno finalmente raggiunto un aumento maggiore del volume e dell’attività del BAT dopo il trattamento [5]. Gli endpoint secondari hanno rivelato che la spesa energetica a riposo dell’intero corpo era più alta dopo il trattamento con mirabegron; tuttavia, non sono state riscontrate modifiche nel peso corporeo o nella composizione. Questi risultati dovrebbero essere associati a un intervallo di BMI ristretto e alla partecipazione di donne non obese. Inoltre, è stato riscontrato che la terapia con Mirabegron aumenta i biomarcatori delle lipoproteine come HDL e apolipoproteina A1, apolipoproteina E e peptide inibitorio gastrico (GIP), nonché i livelli di adiponectina, adipochina antidiabetica e antinfiammatoria. Dopo il trattamento con mirabegron, è stata osservata una riduzione del rapporto ApoB100/ApoA1, un biomarcatore del rischio cardiovascolare. Infine, dopo il trattamento cronico con mirabegron, un test di tolleranza al glucosio per via endovenosa ha rivelato una maggiore sensibilità all’insulina, efficacia del glucosio e secrezione di insulina [5]. Tuttavia, il cambiamento nella valutazione del modello omeostatico della resistenza all’insulina (HOMA-IR), una misura della resistenza all’insulina, non è stato significativo dopo il trattamento cronico con mirabegron. Gli autori hanno suggerito che la ragione principale dovrebbe essere il livello HOMA-IR quasi normale all’inizio dello studio [5]. Come è una preoccupazione comune nel trattamento cronico con agonisti adrenergici, 100mg di Mirabegron hanno portato a una variazione diurna della frequenza cardiaca tale che Mirabegron l’ha aumentata di più durante la notte rispetto a quando i soggetti erano svegli e in movimento. D’altra parte, il trattamento con Mirabegron non ha avuto alcun effetto sulla tolleranza all’esercizio [5].
Loh et al. [21] hanno riportato l’efficacia di varie dosi singole di mirabegron (50, 100, 150 e 200 mg) in un gruppo di 17 individui sani (11 uomini, 6 donne) che hanno assunto il farmaco in quattro giorni separati, con 3-14 giorni di wash-out tra ogni dose. Hanno riferito che la spesa energetica (misurata tramite calorimetria indiretta) è aumentata significativamente dopo le dosi da 100 mg e 200 mg e ha mostrato una tendenza all’aumento dopo le dosi da 150 mg, ma non era significativamente diversa dal basale in risposta a 50 mg di mirabegron. La temperatura cutanea sopraclaveare (come indicatore surrogato dell’attività BAT), è aumentata dopo le dosi di mirabegron da 50 mg, 100 mg e 150 mg, ma non era significativamente diversa dal basale in risposta a 200 mg. Considerando gli effetti collaterali, il cambiamento nella pressione sanguigna sistolica è stato significativo dopo le dosi da 150 mg e 200 mg rispetto alla dose da 50 mg e alla dose da 100 mg. Tuttavia, non c’era alcuna differenza nella pressione sanguigna diastolica tra le dosi da 50 mg, 100 mg, 150 mg e 200 mg. Il cambiamento nella frequenza cardiaca è stato maggiore dopo 200 mg rispetto alle dosi rimanenti. Hanno concluso che una dose da 100 mg di mirabegron può essere efficace per aumentare il dispendio energetico e la temperatura cutanea sopraclaveare in modo specifico per il recettore β3-adrenergico, senza gli aumenti significativi della pressione sanguigna o della frequenza cardiaca osservati a dosi più elevate [21].
Baskin et al. [17] hanno studiato le implicazioni cliniche del mirabegron in 12 uomini sani e magri a cui è stata somministrata la dose approvata di 50 mg e una dose elevata di 200 mg. Si è verificato un aumento più che proporzionale alla dose nell’attività metabolica del BAT (misurata tramite PET/CT). Rispetto al placebo, 50 mg di mirabegron hanno aumentato l’attività del BAT nella maggior parte dei soggetti. Tuttavia, l’attivazione del BAT con 50 mg è stata significativamente inferiore rispetto a quella con 200 mg. Solo la dose da 200 mg ha aumentato la spesa energetica a riposo (5,8%). La stimolazione cardiovascolare è stata coerente con studi precedenti, poiché 200 mg di mirabegron hanno aumentato sia la frequenza cardiaca che la pressione sanguigna.
Uno studio randomizzato, in doppio cieco, cross-over costituito da tre interventi (esposizione al freddo a breve termine (~2 h), mirabegron (dose singola da 200 mg) e placebo) in un gruppo di 10 uomini magri olandesi sud asiatici e 10 uomini magri europei, condotto da Nahon et al. [3], ha rivelato che l’esposizione al freddo e il mirabegron hanno indotto effetti metabolici benefici, tra cui un aumento della spesa energetica a riposo (misurata mediante calorimetria indiretta), livelli di acidi grassi liberi nel siero e ossidazione dei lipidi. Il mirabegron ha aumentato la frequenza cardiaca sia nei sud asiatici (+10 battiti/min) che nei caucasici bianchi (+7 battiti/min), mentre la pressione sanguigna sistolica e diastolica non sono cambiate in modo significativo [3]. È stato osservato che una singola dose di mirabegron ha aumentato i livelli di insulina nel siero senza influenzare i livelli di glucosio. Il mirabegron può stimolare il rilascio di insulina direttamente agendo sul β3-AR del pancreas o indirettamente attraverso un aumento degli FFA che possono stimolare il pancreas a rilasciare insulina [3].
L’azione dose-dipendente del mirabegron sul tessuto adiposo, inclusa l’influenza sull’attività BAT e sul dispendio energetico, può essere analoga all’effetto del mirabegron sulla vescica urinaria. L’attivazione dei recettori β3-adrenergici con mirabegron ha determinato risposte dei recettori β3-adrenergici dipendenti dalla concentrazione [27]. Per quanto riguarda la funzione della vescica, negli studi in vivo, la somministrazione di mirabegron ha ridotto la pressione intravescicolare e le contrazioni vescicali spontanee in modo dose-dipendente [23].
È stato riportato che dosi elevate di mirabegron (in particolare 200 mg al giorno), molto più elevate di quelle approvate dalla FDA per l’iperattività della vescica (50 mg al giorno), possono essere associate a effetti collaterali cardiovascolari come mal di testa, tachicardia e pressione sanguigna elevata (per lo più solo pressione sanguigna sistolica) [1,3,5,17,21]. L’aumento della pressione sanguigna sistolica può raggiungere ~10 mm Hg alla dose di 200 mg al giorno [21]. Questo è il risultato della perdita di selettività per il β3-adrenocettore a questa dose, tale che mirabegron attiva indirettamente i β1-adrenocettori che sono ampiamente espressi in vari organi, in particolare il sistema cardiovascolare. Questo meccanismo coinvolge l’assorbimento del trasportatore di noradrenalina del mirabegron nei terminali nervosi simpatici cardiaci, causando successivamente un rilascio di noradrenalina, che attiva i β1-adrenocettori [21]. Tuttavia, il trattamento con mirabegron non ha avuto effetti sulla tolleranza all’esercizio [5]. L’attivazione dei β1-adrenocettori può essere attenuata dalla co-somministrazione di propranololo o bisoprololo [16]. D’altro canto, gli studi clinici hanno rivelato che dosi di mirabegron fino a 100 mg al giorno per almeno 12 mesi hanno mostrato un buon profilo di sicurezza e non hanno determinato un aumento dell’incidenza di tachicardia, pressione sanguigna, alterazioni dell’ECG o eventi cardiovascolari [21]. Dosi terapeutiche inferiori (50 mg) nei pazienti con OAB hanno determinato piccole variazioni della frequenza cardiaca (1 battito al minuto) e della pressione sanguigna (1 mm Hg o meno). Considerando gli effetti collaterali cardiovascolari, il mirabegron non è raccomandato nei pazienti con grave ipertensione incontrollata (pressione sanguigna sistolica ≥ 180 mm Hg e/o pressione sanguigna diastolica ≥ 110 mm Hg) [16].
Nel secondo gruppo di studi, condotto da Finlin et al. [9,13,14], è stata testata una bassa dose di mirabegron, una che è stata approvata per il trattamento dell’OAB. In un gruppo di 13 pazienti obesi di mezza età, 50 mg di mirabegron al giorno durante una terapia di 12 settimane hanno indotto il “beiging” del tessuto adiposo bianco sottocutaneo, nonché un miglioramento della funzione delle cellule β. Mirabegron ha aumentato l’espressione proteica dei marcatori adiposi “beige” UCP1 (2,4 volte), della proteina transmembrana 26 (TMEM26) (4,2 volte) e dell’effettore A simile al DFFA che induce la morte cellulare (CIDEA) (2,4 volte) [13]. Il “beiging” del tessuto adiposo bianco sottocutaneo da parte di mirabegron può ridurre la disfunzione del tessuto adiposo, il che può migliorare la capacità ossidativa muscolare e può migliorare la funzione delle cellule β [13]. Prendendo in considerazione l’omeostasi del glucosio, il trattamento con mirabegron ha migliorato la tolleranza orale al glucosio, portando a convertire il prediabete in una normale concentrazione di glucosio, ha ridotto i livelli di emoglobina A1c e ha migliorato la sensibilità all’insulina e la funzione delle cellule β, senza influenzare la glicemia a digiuno o i livelli di insulina a digiuno e HOMA-IR. Tuttavia, i risultati delle pinze euglicemiche, che sono il gold standard per misurare la sensibilità all’insulina, hanno rivelato che il trattamento con mirabegron ha aumentato in modo coerente e significativo la velocità di infusione del glucosio di circa il 12% [13]. I livelli di lipidi plasmatici sono cambiati in modo significativo, ma, dopo il trattamento con mirabegron, è stata riscontrata una tendenza verso una riduzione del colesterolo totale [13]. Sfortunatamente, una terapia di 12 settimane non ha determinato un aumento significativo della quantità di BAT e del dispendio energetico a riposo, della perdita di peso o dei cambiamenti nella composizione corporea in tali pazienti [13].
L’effetto benefico del mirabegron, simile all’effetto dell’esposizione al freddo, sull’induzione del tessuto adiposo “beige” nel tessuto adiposo sottocutaneo umano è stato riportato anche in un altro studio condotto da Finlin et al. [14]. Hanno esposto al freddo i partecipanti alla ricerca magri e obesi o li hanno trattati con mirabegron. Il trattamento cronico con mirabegron (10 settimane; 50 mg/giorno) ha indotto UCP1 (3 volte) e TMEM26 (8,7 volte) nei soggetti obesi. Inoltre, l’espressione di UCP1 e dei marcatori degli adipociti “beige” è aumentata più che dopo 10 giorni di ripetuta esposizione al freddo [14].
Nello studio successivo, composto da 12 partecipanti obesi insulino-resistenti, Finlin et al. [9] hanno valutato la capacità del trattamento con pioglitazone (30 mg/giorno) o del trattamento con mirabegron (50 mg/giorno) in monoterapia, così come una combinazione di trattamento con pioglitazone (30 mg/giorno) e mirabegron (50 mg/giorno), di aumentare il grasso “beige” o migliorare ulteriormente il metabolismo del glucosio durante 12 settimane di terapia. Il pioglitazone è un attivatore PPARγ che può stimolare il BAT o “grasso beige”. Il trattamento con pioglitazone o la combinazione di pioglitazone e mirabegron hanno aumentato l’espressione del marcatore proteico del tessuto adiposo “beige” e migliorato la sensibilità all’insulina (misurata tramite clamp euglicemico, più efficace nella terapia combinata) e l’omeostasi del glucosio (inclusi test di tolleranza al glucosio migliorati, più efficaci nella terapia combinata), ma nessuno dei due trattamenti ha indotto il BAT o influenzato la spesa energetica nei soggetti obesi. Inoltre, non si è verificato alcun cambiamento significativo nel peso corporeo dopo il trattamento. Nonostante il fatto che mirabegron e pioglitazone somministrati separatamente abbiano indotto il “beiging” del tessuto adiposo, l’aggiunta di pioglitazone a mirabegron non ha migliorato il “beiging”, poiché il trattamento combinato ha prodotto un “beiging” inferiore rispetto a entrambi i farmaci somministrati singolarmente [9].
Sebbene i risultati preliminari degli studi sugli animali abbiano mostrato i benefici della co-somministrazione di mirabegron e metformina nella prevenzione e nel trattamento dell’obesità [2], a nostra conoscenza, l’influenza di tale terapia combinata non è stata verificata in relazione all’attività BAT, al dispendio energetico e alla perdita di peso negli esseri umani. È noto solo che non ci sono interazioni clinicamente significative tra metformina e mirabegron. Nello studio con 32 soggetti maschi sani (BMI: 18–30 kg/m2), mirabegron (160 mg somministrati una volta al giorno) non ha mostrato alcun effetto sulla farmacocinetica di metformina (500 mg somministrati due volte al giorno). La co-somministrazione di mirabegron con metformina ha determinato piccole modifiche nell’esposizione a mirabegron (AUC e Cmax diminuite del 21%). Le modifiche farmacocinetiche osservate non sono state considerate clinicamente rilevanti. Pertanto, non è necessario alcun aggiustamento del dosaggio di mirabegron quando viene co-somministrato con metformina [33].
Sebbene i dati confermino che una bassa dose di mirabegron può indurre il “beiging” del WAT sottocutaneo, è stato riportato che 50 mg di mirabegron durante il trattamento a breve termine (circa 12 settimane di terapia) non hanno alcun effetto sulla quantità di BAT, sul dispendio energetico a riposo e sulla perdita di peso. Pertanto, sono necessari studi clinici di lunga durata, con partecipanti obesi con una dose inferiore di mirabegron, per valutare se il “beiging” del tessuto adiposo si tradurrebbe in un miglioramento del dispendio energetico a riposo e in una significativa perdita di peso.
Conclusioni:
Il BAT metabolicamente attivo è stato correlato positivamente al miglioramento dell’energia, del glucosio e del metabolismo dell’intero corpo [34]. L’attivazione del BAT e l’induzione del processo di “browning” nel WAT sembrano essere un’interessante strategia terapeutica per aumentare la spesa energetica e migliorare il metabolismo. Il mirabegron, come agonista del recettore β3-adrenergico, si è rivelato efficace come attivatore del BAT, stimolatore delle cellule “beige” e regolatore dell’omeostasi metabolica sia negli studi sugli animali che negli esseri umani. Sebbene negli studi sugli animali la somministrazione di mirabegron abbia portato a un miglioramento dell’obesità, non è stata ancora dimostrata una significativa perdita di peso nei pazienti obesi dopo dosi elevate o basse del farmaco. Ciò può essere spiegato dalla durata troppo breve degli studi e dal numero esiguo di partecipanti agli studi. Inoltre, negli esseri umani, il trattamento più efficace per la stimolazione del BAT e del WAT è stato quello con dosi elevate di mirabegron; tuttavia, gli effetti collaterali cardiovascolari possono limitare l’uso di dosi superiori a quelle approvate dalla FDA per il trattamento della vescica iperattiva. Da un lato, considerando l’uso di dosi elevate di mirabegron, deve essere valutata la sicurezza a lungo termine in relazione al sistema cardiovascolare. In caso di attivazione aggravata dei recettori β1 miocardici, la somministrazione concomitante di 100-200 mg di mirabegron con un bloccante β1-AR può essere una strategia terapeutica utile per evitare effetti collaterali cardiovascolari. D’altro canto, dovrebbe essere valutato se dosi più piccole di mirabegron, ad esempio quelle approvate per la vescica iperattiva (50 mg al giorno), assunte per un periodo di tempo più lungo, saranno sufficienti a stimolare la crescita del BAT, l’imbrunimento del WAT e la termogenesi che può portare alla perdita di peso. Negli studi clinici riguardanti l’efficacia e la sicurezza del mirabegron nei pazienti con vescica iperattiva, l’influenza del mirabegron sul peso corporeo non è stata verificata. A nostra conoscenza, l’efficacia del mirabegron in relazione ai disturbi metabolici, inclusa l’obesità, nei soggetti trattati per vescica iperattiva, non è stata finora valutata.
Si potrebbe quindi ipotizzare che, il potenziale ruolo del Mirabegron nel trattamento o nella prevenzione dell’obesità dipenderebbe dai risultati della sua efficacia determinati da studi clinici a lungo termine. In caso di mancanza o insoddisfacente effetto dimagrante (rispetto ai farmaci attualmente disponibili approvati per il trattamento dell’obesità), il mirabegron potrebbe essere utilizzato per migliorare il profilo metabolico nei pazienti obesi. Se l’effetto dimagrante del mirabegron venisse confermato, il farmaco diventerebbe un’opzione alternativa agli attuali agenti anti-obesità, specialmente nei pazienti con controindicazioni o intolleranza ad altri farmaci. Inoltre, un aspetto interessante da valutare negli studi clinici sarebbe se la co-somministrazione di mirabegron e altri farmaci, come metformina, pioglitazone o altri farmaci anti-obesità attualmente utilizzati, potrebbe essere una strategia più efficace rispetto alla somministrazione di tali farmaci da soli per migliorare i profili metabolici o per trattare l’obesità. I benefici della co-somministrazione di mirabegron e metformina nella prevenzione e nel trattamento dell’obesità, dimostrati in studi sugli animali, devono essere confermati in ulteriori studi clinici. Sebbene i risultati preliminari della co-somministrazione di mirabegron e pioglitazone in partecipanti obesi non abbiano indicato alcuna influenza di tale terapia sul peso corporeo, devono essere eseguiti ulteriori studi per confermare questi risultati. Pertanto, l’introduzione di agonisti del recettore β3-adrenergico nel trattamento dell’obesità in futuro richiederà studi a lungo termine con un numero maggiore di soggetti per valutarne l’efficacia, la tollerabilità e la sicurezza.
I tessuti adiposi bruni e “beige” rimangono un bersaglio attraente per combattere le malattie metaboliche. Sono necessari ulteriori studi per confermare se la combinazione di agenti attivatori di BAT e “beige”, esercizi fisici e una dieta ipocalorica sana sarebbe una strategia di successo per ottenere la perdita di peso nei pazienti con obesità.
E per l’uso off-label nella ricomposizione corporea? I test in tal senso sono ancora scarsi e dal design spesso pessimo. Vi sono stati riscontri positivi, almeno preliminarmente parlando, con protocolli di 8-12 settimane a dosaggi di 75-100mg/die. Il dosaggio era stato settato partendo da 25mg/die per poi mantenere e osservare le risposte al dosaggio per qualche giorno [pressione, battito cardiaco ecc…]. L’uso del Mirabegron in ambito sportivo è tanto pionieristico come lo è quello dei tireomimetici. Ci vorranno ancora diversi studi per poter essere maggiormente certi di concreti vantaggi applicativi di questa molecola come PEDs.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Elia, E.F.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of human brown adipose tissue by a β3-adrenergic receptor agonist. Cell. Metab.2015, 21, 33–38. [Google Scholar] [CrossRef]
Zhao, X.Y.; Liu, Y.; Zhang, X.; Zhao, B.C.; Burley, G.; Yang, Z.C.; Luo, Y.; Li, A.Q.; Zhang, R.X.; Liu, Z.Y.; et al. The combined effect of metformin and mirabegron on diet-induced obesity. Med. Comm.2023, 4, e207. [Google Scholar] [CrossRef]
Nahon, K.J.; Janssen, L.G.M.; Sardjoe Mishre, A.S.D.; Bilsen, M.P.; van der Eijk, J.A.; Botani, K.; Overduin, L.A.; Ruiz, J.R.; Burakiewicz, J.; Dzyubachyk, O.; et al. The effect of mirabegron on energy expenditure and brown adipose tissue in healthy lean South Asian and Europid men. Diabetes Obes. Metab.2020, 22, 2032–2044. [Google Scholar] [CrossRef]
Hao, L.; Scott, S.; Abbasi, M.; Zu, Y.; Khan, M.S.H.; Yang, Y.; Wu, D.; Zhao, L.; Wang, S. Beneficial Metabolic Effects of Mirabegron In Vitro and in High-Fat Diet-Induced Obese Mice. J. Pharmacol. Exp. Ther.2019, 369, 419–427. [Google Scholar] [CrossRef]
Waki, H.; Yamauchi, T. Body-weight-independent glucose-lowering effect of the b3-adrenergic receptor agonist mirabegron in humans. J. Diabetes Investig.2021, 12, 689–690. [Google Scholar] [CrossRef]
Harb, E.; Kheder, O.; Poopalasingam, G.; Rashid, R.; Srinivasan, A.; Izzi-Engbeaya, C. Brown adipose tissue and regulation of human body weight. Diabetes Metab. Ress. Rev.2023, 39, e3594. [Google Scholar] [CrossRef]
Finlin, B.S.; Memetimin, H.; Zhu, B.; Confides, A.L.; Vekaria, H.J.; El Khouli, R.H.; Johnson, Z.R.; Westgate, P.M.; Chen, J.; Morris, A.J.; et al. The β3-adrenergic receptor agonist mirabegron improves glucose homeostatsis in obese humans. J. Clin. Investig.2020, 130, 2319–2331. [Google Scholar] [CrossRef]
Finlin, B.S.; Memetimin, H.; Confides, A.L.; Kasza, I.; Zhu, B.; Vekaria, H.J.; Harfmann, B.; Jones, K.A.; Johnson, Z.R.; Westgate, P.M.; et al. Human adipose beiging in response to cold and mirabegron. JCI Insight2018, 3, e121510. [Google Scholar] [CrossRef]
Dehvari, N.; Sato, M.; Bokhari, M.H.; Kalinovich, A.; Ham, S.; Gao, J.; Nguyen, H.T.M.; Whiting, L.; Mukaida, S.; Merlin, J.; et al. The metabolic effects of mirabegron are mediated primarily by β3-adrenoceptors. Pharmacol. Res. Perspect.2020, 8, e00643. [Google Scholar] [CrossRef]
Dehvari, N.; da Silva Junior, E.D.; Bengtsson, T.; Hutchinson, D.S. Mirabegron: Potential off target effects and uses beyond the bladder. Br. J. Pharmacol.2018, 175, 4072–4082. [Google Scholar] [CrossRef]
Baskin, A.S.; Linderman, J.D.; Brycha, R.J.; McGehee, S.; Anflick-Chames, E.; Cero, C.; Johnson, J.W.; O’Mara, A.E.; Fletcher, L.A.; Leitner, B.P.; et al. Regulation of Human Adipose Tissue Activation, Gallbladder Size, and Bile Acid Metabolism by a b3-Adrenergic Receptor Agonis. Diabetes2018, 67, 2113–2125. [Google Scholar] [CrossRef]
Pereas Valgas da Silva, C.; Calmasini, F.; Alexandre, E.C.; Raposo, H.F.; Delbin, M.A.; Monica, F.Z.; Zanesco, A. The effects of mirabegron on obesity-induced inflammation and insulin resistance are associated with brown adipose tissue activation but not beiging in the subcutaneous white adipose tissue. Clin. Exp. Pharmacol. Physiol.2021, 48, 1477–1487. [Google Scholar] [CrossRef]
Arch, J.R.S. Challenges in β3-adrenoceptor agonist drug development. Ther. Adv. Endocrinol. Metab.2011, 2, 59–64. [Google Scholar] [CrossRef]
Liu, X.; Pérusse, F.; Bukowiecki, L.J. Mechanisms of the antidiabetic effects of the beta 3-adrenergic agonist CL-316243 in obese Zucker-ZDF rats. Am. J. Physiol.1998, 274, R1212–R1219. [Google Scholar]
Takasu, T.; Ukai, M.; Sato, S.; Matsui, T.; Nagase, I.; Maruyama, T.; Sasamata, M.; Miyata, K.; Uchida, H.; Yamaguchi, O. Effect of (R)-2-(2-aminothiazol-4-yl)-4′-{2-[(2-hydroxy-2-phenylethyl)amino]ethyl} acetanilide (YM178), a novel selective beta3-adrenoceptor agonist, on bladder function. J. Pharmacol. Exp. Ther.2007, 321, 642–647. [Google Scholar] [CrossRef]
Igawa, Y.; Michel, M.C. Pharmacological profile of β3-adrenoceptor agonists in clinical development for the treatment of overactive bladder syndrome. Naunyn. Schmiedebergs. Arch. Pharmacol.2013, 386, 177–183. [Google Scholar] [CrossRef] [PubMed]
Krhut, J.; Martan, A.; Zachoval, R.; Hanus, T.; Svabik, K.; Zvara, P. Impact of body mass index on treatment efficacy of mirabegron for overactive bladder in females. Eur. J. Obstet. Gynecol. Reprod. Biol.2016, 196, 64–68. [Google Scholar] [CrossRef] [PubMed]
Hatanaka, T.; Ukai, M.; Watanabe, M.; Someya, A.; Ohtake, A.; Suzuki, M.; Ueshima, K.; Sato, S.; Sasamata, M. In vitro and in vivo pharmacological profile of the selective β3-adrenoceptor agonist mirabegron in rats. Naunyn. Schmiedebergs. Arch. Pharmacol.2013, 386, 247–253. [Google Scholar] [CrossRef]
Nagiri, C.; Kobayashi, K.; Tomita, A.; Kato, M.; Kobayashi, K.; Yamashita, K.; Nishizawa, T.; Inoue, A.; Shihoya, W.; Nureki, O. Cryo-EM structure of the β3-adrenergic receptor reveals the molecular basis of subtype selectivity. Mol. Cell.2021, 81, 3205–3215. [Google Scholar] [CrossRef] [PubMed]
Brucker, B.M.; King, J.; Mudd, P.N., Jr.; McHale, K. Selectivity and Maximum Response of Vibegron and Mirabegron for β3-Adrenergic Receptors. Curr. Ther. Res. Clin. Exp.2022, 96, 100674. [Google Scholar] [CrossRef]
Alexandre, E.C.; Kiguti, L.R.; Calmasini, F.B.; Silva, F.H.; da Silva, K.P.; Ferreira, R.; Ribeiro, C.A.; Mónica, F.Z.; Pupo, A.S.; Antunes, E. Mirabegron relaxes urethral smooth muscle by a dual mechanism involving β3 -adrenoceptor activation and α1 -adrenoceptor blockade. Br. J. Pharmacol.2016, 173, 415–428. [Google Scholar] [CrossRef]
De Stefano, A.; Schinzari, F.; Di Daniele, N.; Sica, G.; Gentileschi, P.; Vizioli, G.; Cardillo, C.; Tesauro, M. Mirabegron relaxes arteries from human visceral adipose tissue through antagonism of α1-adrenergic receptors. Vascul. Pharmacol.2022, 146, 107094. [Google Scholar] [CrossRef]
Lowell, B.B.; S-Susulic, V.; Hamann, A.; Lawitts, J.A.; Himms-Hagen, J.; Boyer, B.B.; Kozak, L.P.; Flier, J.S. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature1993, 366, 740–742. [Google Scholar] [CrossRef]
Hamann, A.; Flier, J.S.; Lowell, B.B. Decreased brown fat markedly enhances susceptibility to diet-induced obesity, diabetes, and hyperlipidemia. Endocrinology1996, 137, 21–29. [Google Scholar] [CrossRef] [PubMed]
Lv, Z.; Guo, Y. Metformin and its benefits for various diseases. Front. Endocrinol.2020, 11, 191. [Google Scholar] [CrossRef] [PubMed]
Groen-Wijnberg, M.; van Dijk, J.; Krauwinkel, W.; Kerbusch, V.; Meijer, J.; Tretter, R.; Zhang, W.; van Gelderen, M. Pharmacokinetic Interactions Between Mirabegron and Metformin, Warfarin, Digoxin or Combined Oral Contraceptives. Eur. J. Drug Metab. Pharmacokinet.2017, 42, 417–429. [Google Scholar] [CrossRef] [PubMed]
Bel, J.C.; Tai, T.C.; Khaper, N.; Lees, S.J. Mirabegron: The most promising adipose tissue beiging agent. Physiol. Rep.2021, 9, e14779. [Google Scholar] [CrossRef]
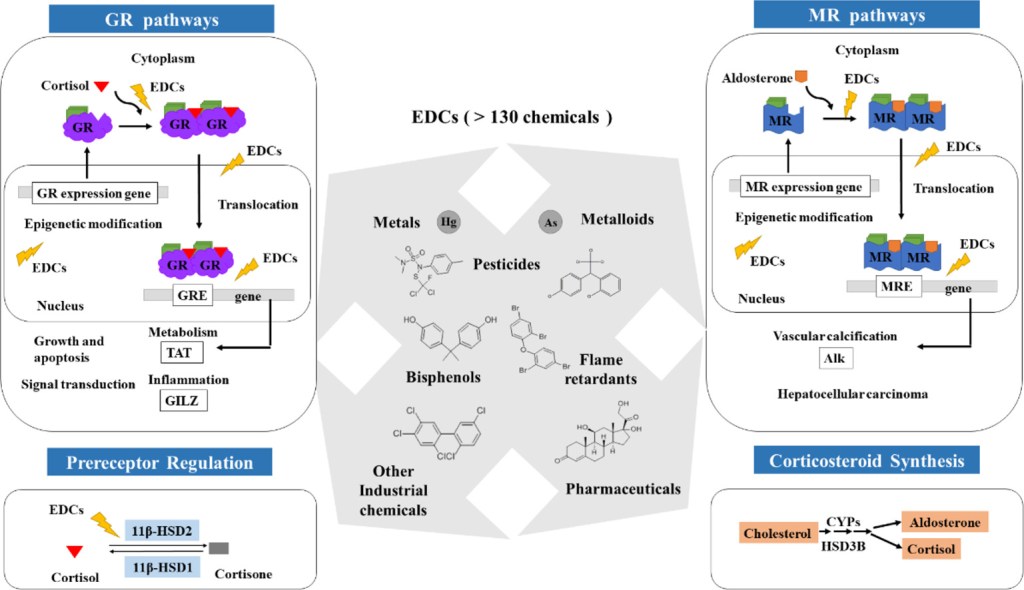
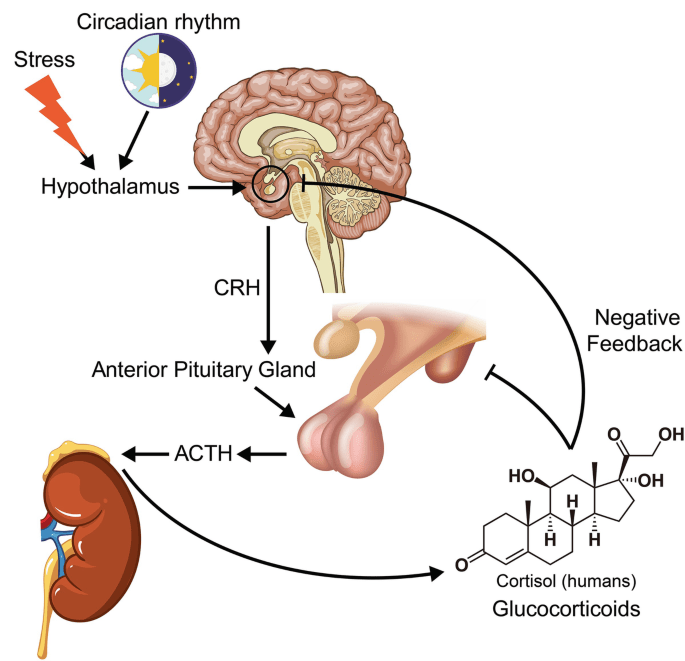
I Glucocorticoidi (o, meno comunemente, glucocorticosteroidi) sono una classe di corticosteroidi, ovvero una classe di ormoni steroidei. I Glucocorticoidi sono corticosteroidi che si legano al recettore dei glucocorticoidi[1], presente in quasi tutte le cellule animali vertebrate. Il nome “Glucocorticoide” è un portmanteau (glucosio + corteccia + steroide) ed è composto dal suo ruolo nella regolazione del metabolismo del glucosio, dalla sintesi nella corteccia surrenale e dalla sua struttura steroidea.
I Glucocorticoidi fanno parte del meccanismo di feedback del sistema immunitario, che riduce alcuni aspetti della funzione immunitaria, come l’infiammazione. Sono quindi utilizzati in medicina per trattare le malattie causate da un sistema immunitario iperattivo, come allergie, asma, malattie autoimmuni e sepsi. I Glucocorticoidi hanno molti effetti diversi, come la pleiotropia, tra cui effetti collaterali potenzialmente dannosi.[2] Inoltre, interferiscono con alcuni meccanismi anomali delle cellule tumorali, per cui vengono utilizzati in dosi elevate per il trattamento del cancro. Ciò include effetti inibitori sulla proliferazione dei linfociti, come nel trattamento di linfomi e leucemie, e l’attenuazione degli effetti collaterali dei farmaci antitumorali.
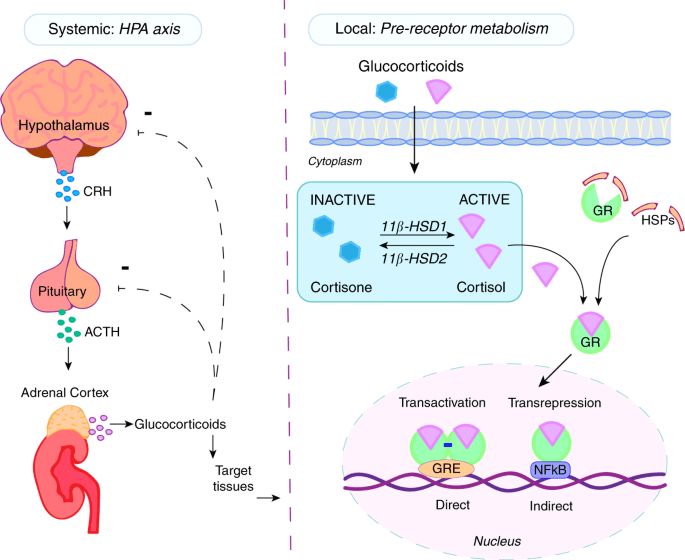
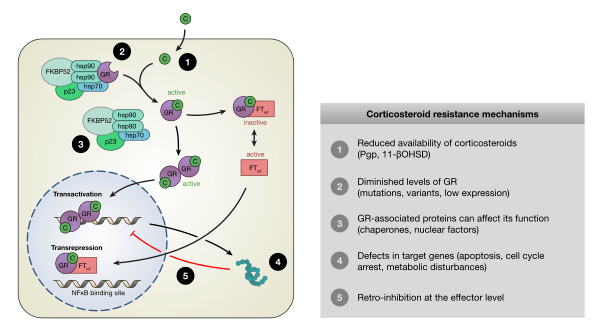
I glucocorticoidi agiscono sulle cellule legandosi al Recettore dei Glucocorticoidi (GR). Il complesso recettore glucocorticoide-glucocorticoide attivato regola l’espressione di proteine antinfiammatorie nel nucleo (processo noto come transattivazione) e reprime l’espressione di proteine pro-infiammatorie nel citosol impedendo la traslocazione di altri fattori di trascrizione dal citosol al nucleo (transrepressione).[2]
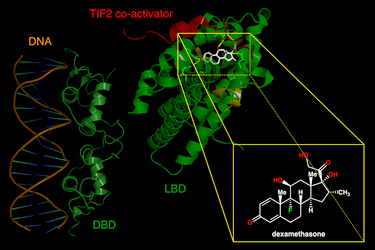
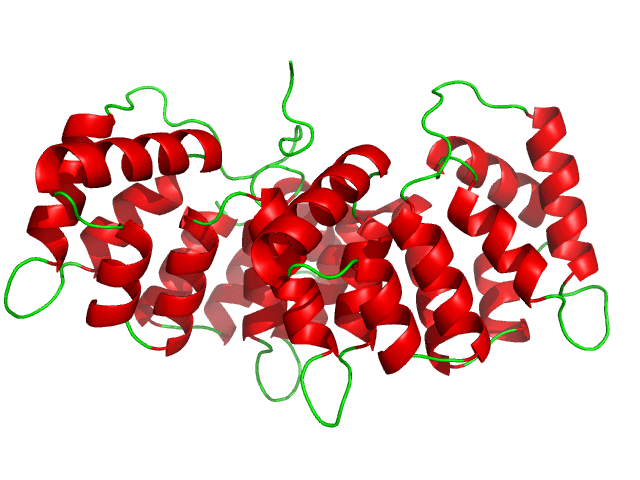
Il Recettore dei Glucocorticoidi (GR o GCR), noto anche come NR3C1 (sottofamiglia 3 del recettore nucleare, gruppo C, membro 1), è il recettore a cui si legano il Cortisolo e altri Glucocorticoidi.
I glucocorticoidi si distinguono dai mineralocorticoidi e dagli steroidi sessuali per i loro specifici recettori, cellule bersaglio ed effetti. In termini tecnici, il termine “corticosteroide” si riferisce sia ai glucocorticoidi che ai mineralocorticoidi (poiché entrambi sono mimici degli ormoni prodotti dalla corteccia surrenale), ma è spesso usato come sinonimo di “glucocorticoide”. I glucocorticoidi sono prodotti principalmente nella zona fascicolata della corteccia surrenale, mentre i mineralocorticoidi sono sintetizzati nella zona glomerulosa.
Il Cortisolo (o idrocortisone) è il più importante glucocorticoide umano. È essenziale per la vita e regola o supporta una serie di importanti funzioni cardiovascolari, metaboliche, immunologiche e omeostatiche. Gli aumenti delle concentrazioni di glucocorticoidi sono parte integrante della risposta allo stress e sono i biomarcatori più comunemente utilizzati per misurare lo stress.[3] I glucocorticoidi hanno anche numerose funzioni non correlate allo stress e le concentrazioni di glucocorticoidi possono aumentare in risposta al piacere o all’eccitazione.[4] Sono disponibili diversi glucocorticoidi sintetici, ampiamente utilizzati nella pratica medica generale e in numerose specialità, come terapia sostitutiva in caso di carenza di glucocorticoidi o per sopprimere il sistema immunitario dell’organismo.
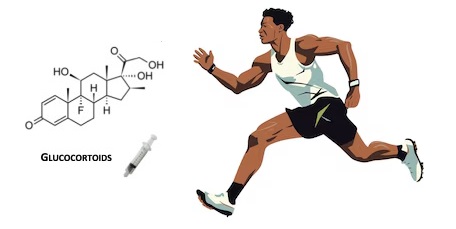
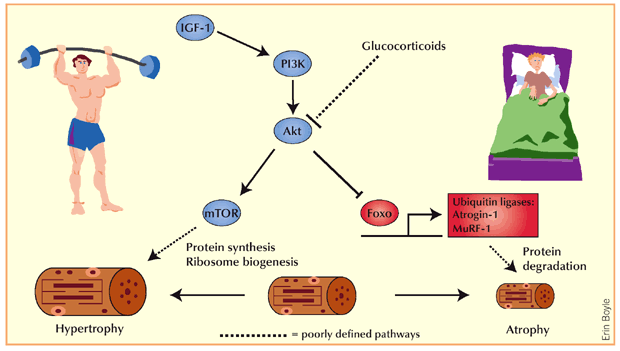
Si sospetta che gli atleti utilizzino i Glucocorticoidi per migliorare le prestazioni atletiche fin dagli anni ’60 (5). I glucocorticoidi possono migliorare le prestazioni fisiche attraverso diverse vie, tra cui una maggiore disponibilità di substrati metabolici attraverso un aumento della lipolisi (6), della proteolisi (7) e della disponibilità di glucosio (5). I Glucocorticoidi possono anche avere effetti immunosoppressivi e antinfiammatori che possono impedire al sistema immunitario di reagire in modo eccessivo a seguito di danni muscolari indotti dall’esercizio fisico (8), e il Cortisolo sembra preparare l’organismo per l’esercizio successivo (9). Inoltre, i glucocorticoidi possono stimolare i recettori cerebrali dei glucocorticoidi, determinando una riduzione del dolore muscolare durante l’esercizio, un aumento della soglia della fatica e risposte edoniche positive, che possono tradursi in un aumento delle prestazioni fisiche (10). I glucocorticoidi hanno un elevato potenziale di effetti avversi, tra cui il deperimento muscolare, e questi effetti dipendono da molteplici fattori come il tipo di glucocorticoide, la durata del trattamento, la dose e la via di somministrazione (11). Pertanto, gli atleti che assumono glucocorticoidi per migliorare le prestazioni probabilmente preferiscono periodi di somministrazione più brevi.
L’Associazione Mondiale Antidoping (WADA) aggiorna annualmente la sua lista proibita, un elenco di sostanze e metodi vietati negli sport d’élite. La lista proibita, in vigore dal 1° gennaio 2022, vieta tutti i glucocorticoidi nelle competizioni quando sono somministrati per via orale, rettale o iniettabile, poiché queste forme di somministrazione sono considerate ad effetto sistemico. Altre vie di somministrazione, come l’applicazione topica per inalazione e le iniezioni locali, sono approvate nelle competizioni, poiché si ritiene che abbiano un minore potenziale di miglioramento delle prestazioni. L’uso dei glucocorticoidi al di fuori delle competizioni è approvato (12). Per aiutare i medici a curare gli atleti e a rispettare le norme antidoping, dal 2022 la WADA raccomanda specifici periodi di wash out (tempo dall’ultima dose al giorno prima della competizione) per i diversi tipi di glucocorticoidi e le diverse vie di somministrazione. Dal 2022 la WADA ha anche introdotto livelli di segnalazione urinaria specifici per i diversi tipi di glucocorticoidi, in quanto i diversi glucocorticoidi hanno un’ampia variazione nel tempo di eliminazione (13).
Dagli anni 90 si è studiato l’effetto dei glucocorticoidi sulla frequenza cardiaca e sul consumo di ossigeno durante intervalli di corsa ad alta intensità. Da allora diversi studi RCT hanno analizzato l’effetto dei glucocorticoidi sulla prestazione fisica submassimale e massimale. Poiché le modalità di somministrazione, il tipo di glucocorticoide, la durata del trattamento, la dose, la popolazione e il protocollo di esercizio possono variare tra gli studi, essi giungono a conclusioni diverse per quanto riguarda l’effetto dei glucocorticoidi sulla prestazione fisica, tuttavia sono soprattutto gli studi che indagano l’effetto di alte dosi orali somministrate nel tempo sulla resistenza in bicicletta fino all’esaurimento a mostrare effetti. L’effetto dei glucocorticoidi sulla prestazione fisica e sul metabolismo dei soggetti sani è stato esaminato in precedenza (5, 10), ma senza una ricerca sistematica della letteratura e senza combinare i risultati degli studi inclusi nell’analisi statistica. Pertanto, in tale sede, cercherò anche di fare chiarezza sull’effetto dei glucocorticoidi sulla prestazione massimale o submassimale in soggetti sani.
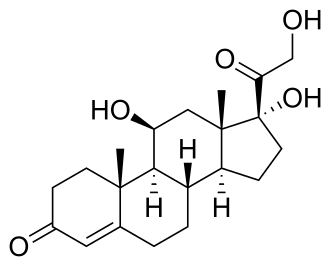
Steroidogenesi dei Glucocorticoidi:
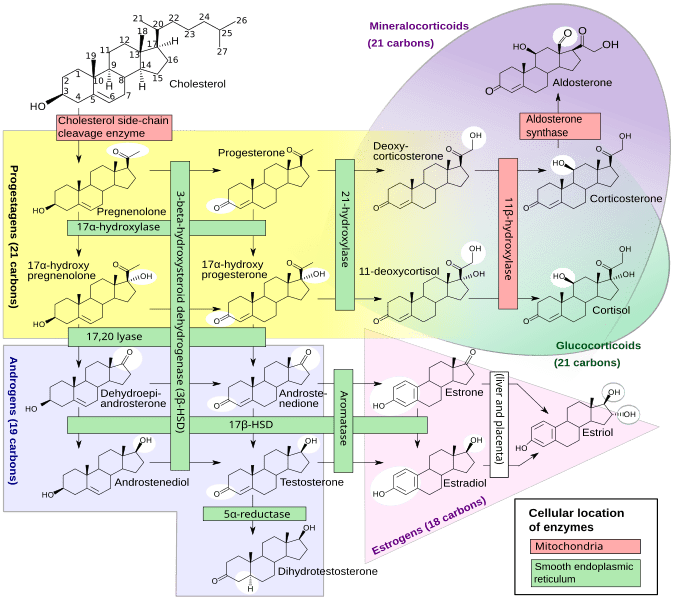
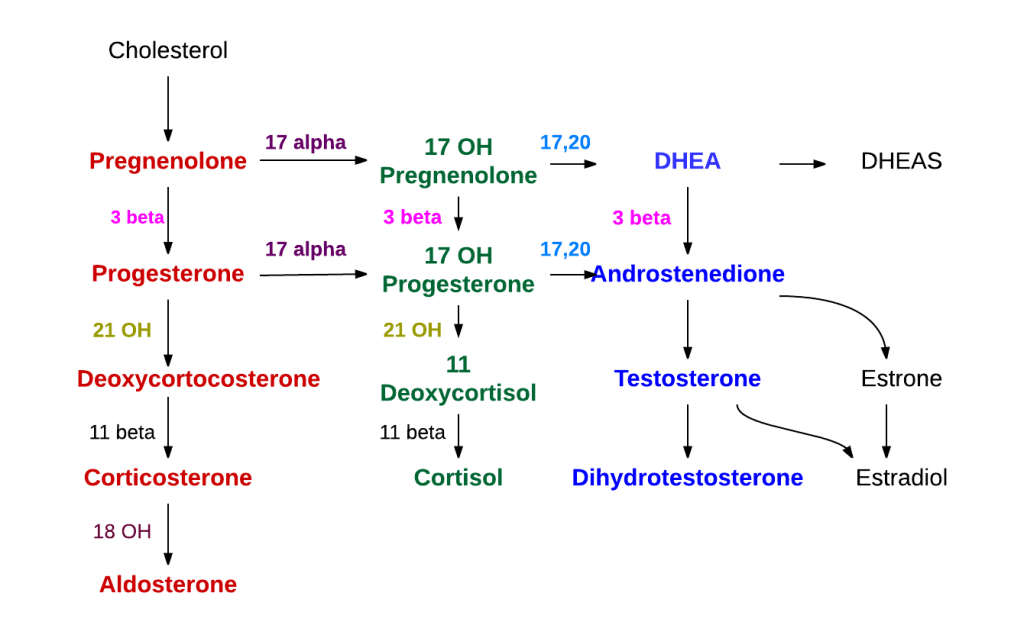
La steroidogenesi è il processo biologico attraverso il quale gli steroidi vengono generati dal colesterolo e trasformati in altri steroidi.[14] Le vie della steroidogenesi differiscono tra le specie. Le principali classi di ormoni steroidei, con i loro membri e le loro funzioni principali, sono i progestinici, i corticosteroidi (corticoidi), gli androgeni e gli estrogeni.[15][16] La steroidogenesi umana dei corticosteroidi avviene nella Corteccia Surrenale.
Steroidogenesi che mostra i Glucocorticoidi nell’ellisse verde a destra con l’esempio primario del Cortisolo. Non è un gruppo strettamente delimitato, ma un continuum di strutture con effetto glucocorticoide crescente.
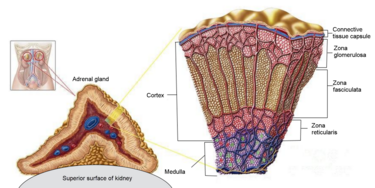
La Corteccia Surrenale è la regione più esterna e anche la parte più grande della ghiandola surrenale. È divisa in tre zone distinte: zona glomerulosa, zona fascicolata e zona reticolare. Ogni zona è responsabile della produzione di ormoni specifici. È anche un sito secondario di sintesi degli androgeni.[17]
Corteccia Surrenale
Zona Glomerulosa
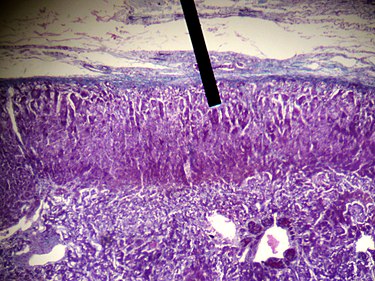
Colorazione H&E della Corteccia Surrenale. La Zona Glomerulare è lo strato più esterno, sotto la capsula renale (vicino all’indicatore)
Lo strato più esterno, la zona glomerulosa, è il sito principale per la produzione di aldosterone, un mineralcorticoide. La sintesi e la secrezione di aldosterone sono regolate principalmente dal sistema renina-angiotensina-aldosterone. Le cellule della zona glomerulosa esprimono un enzima specifico, l’aldosterone sintasi (noto anche come CYP11B2).[18][19] L’aldosterone è ampiamente responsabile della regolazione a lungo termine della pressione sanguigna. [Gli effetti dell’aldosterone si manifestano nel tubulo contorto distale e nel dotto collettore del rene, dove provoca un aumento del riassorbimento del sodio e una maggiore escrezione di potassio (da parte delle cellule principali) e di ioni idrogeno (da parte delle cellule intercalate del dotto collettore).[20] La ritenzione di sodio è anche una risposta del colon distale e delle ghiandole sudoripare alla stimolazione dei recettori dell’aldosterone. Sebbene la produzione sostenuta di aldosterone richieda un ingresso persistente di calcio attraverso canali del Ca2+ attivati a basso voltaggio, le cellule isolate della zona glomerulosa sono considerate non eccitabili, con tensioni di membrana registrate troppo iperpolarizzate per consentire l’ingresso di canali del Ca2+.[21]
La secrezione di aldosterone è stimolata anche dall’ormone adrenocorticotropo (ACTH).[22]
Le cellule della zona glomerulosa non esprimono l’11β-idrossilasi e la 17α-idrossilasi. Per questo motivo la zona glomerulosa non può sintetizzare cortisolo, corticosterone o ormoni sessuali (androgeni). [23] L’espressione di proteine specifiche per i neuroni nelle cellule della zona glomerulosa dei tessuti adrenocorticali umani è stata prevista e riportata da diversi autori [24][25][26] ed è stato suggerito che l’espressione di proteine come la molecola di adesione delle cellule neuronali (NCAM) nelle cellule della zona glomerulosa rifletta la caratteristica rigenerativa di queste cellule, che perderebbero l’immunoreattività della NCAM dopo essersi spostate nella zona fascicolata. [24][27] Tuttavia, insieme ad altri dati sulle proprietà neuroendocrine delle cellule della zona glomerulosa, l’espressione di NCAM potrebbe riflettere una differenziazione neuroendocrina di queste cellule.[24]
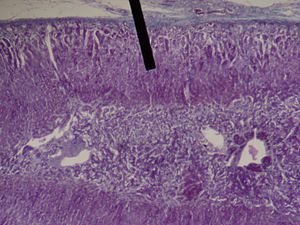
Zona Fasciculata
Zona Fasciculata
Situate tra la glomerulosa e la reticolare, le cellule della zona fascicolata sintetizzano e secernono glucocorticoidi (come l’11-deossicorticosterone, il corticosterone e il cortisolo), oltre a piccole quantità di androgeni ed estrogeni surrenalici.[28] La zona fascicolata ha una maggiore attività di 3β-idrossisteroide deidrogenasi rispetto alla zona reticolare. Pertanto, la zona fascicolata produce più 11-deossicorticosterone, corticosterone e cortisolo.[23] Il principale ormone che stimola la secrezione di cortisolo nell’uomo è l’ACTH, rilasciato dall’ipofisi anteriore.[22] È stato dimostrato che la capacità steroidogenica della zona fascicolata aumenta durante la malattia nei neonati.[22]
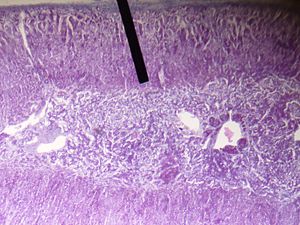
Zona Reticolare
Zona Reticolare
La zona reticolare, lo strato corticale più interno, produce gli androgeni surrenalici, oltre a piccole quantità di estrogeni e alcuni glucocorticoidi.[28] La zona reticolare possiede una quantità maggiore di cofattori necessari per l’attività della 17,20-liasi della 17α-idrossilasi rispetto alla zona fascicolata. Pertanto, la zona reticolare produce più androgeni,[23] soprattutto deidroepiandrosterone (DHEA), DHEA solfato (DHEA-S) e androstenedione (il precursore del Testosterone e DHT) nell’uomo. La secrezione di DHEAS è stimolata anche dall’ACTH.[22]
Come abbiamo visto, I glucocorticoidi sono prodotti principalmente nella Zona Fascicolata.[23]
Il precursore degli steroidi sintetizzati nella corteccia surrenale è il colesterolo che viene immagazzinato nelle vescicole. Il colesterolo può essere sintetizzato de novo nella corteccia surrenale. Tuttavia, la fonte principale di colesterolo sembra essere il colesterolo assunto dalle lipoproteine circolanti. [29]
I passaggi fino a questo punto avvengono in molti tessuti produttori di steroidi. Le fasi successive per generare aldosterone e cortisolo, tuttavia, avvengono principalmente nella corteccia surrenale:
Progesterone → (idrossilazione a C21) → 11-Deossicorticosterone → (due ulteriori idrossilazioni a C11 e C18) → Aldosterone
Progesterone → (idrossilazione a C17) → 17-alfa-idrossiprogesterone → (idrossilazione a C21) → 11-Deossicortisolo → (idrossilazione a C11) → Cortisolo
Fasi della sintesi dell’ormone steroideo surrenale
Effetto sistemico dei Glucocorticoidi:
Gli effetti dei glucocorticoidi possono essere ampiamente classificati in due categorie principali:
immunologici
metabolici.
Inoltre, i glucocorticoidi svolgono ruoli importanti nello sviluppo fetale e nell’omeostasi dei fluidi corporei.
Immunità
Come già accennato, i glucocorticoidi funzionano anche attraverso l’interazione con il recettore dei glucocorticoidi:
Aumentano l’espressione di proteine antinfiammatorie.
Riducono l’espressione di proteine pro-infiammatorie.
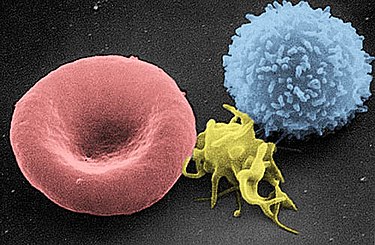
Micrografia elettronica a scansione di un globulo rosso (sinistra), una piastrina (centro) e un linfocita T (destra); colorato
È stato dimostrato che i glucocorticoidi svolgono un ruolo nello sviluppo e nell’omeostasi dei linfociti T. Questo è stato dimostrato in transgenici. Ciò è stato dimostrato in topi transgenici con una maggiore o minore sensibilità della linea delle cellule T ai glucocorticoidi.[30]
Metabolismo
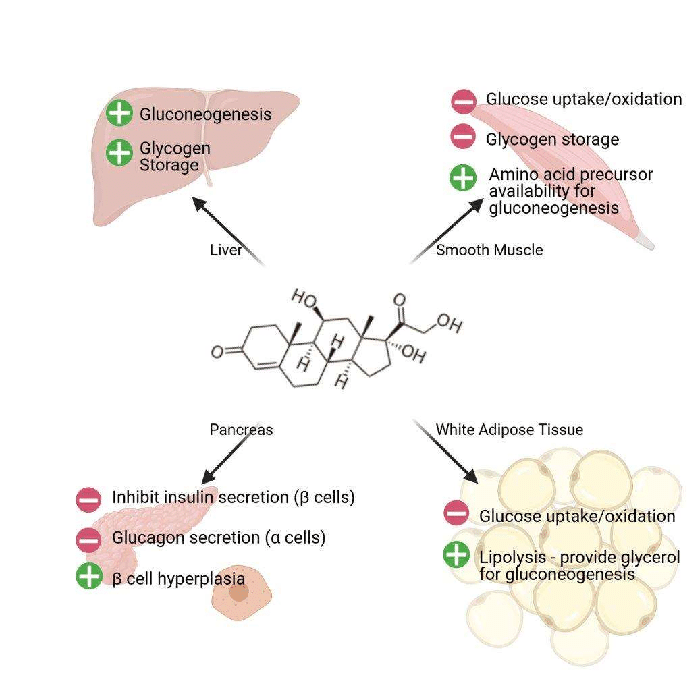
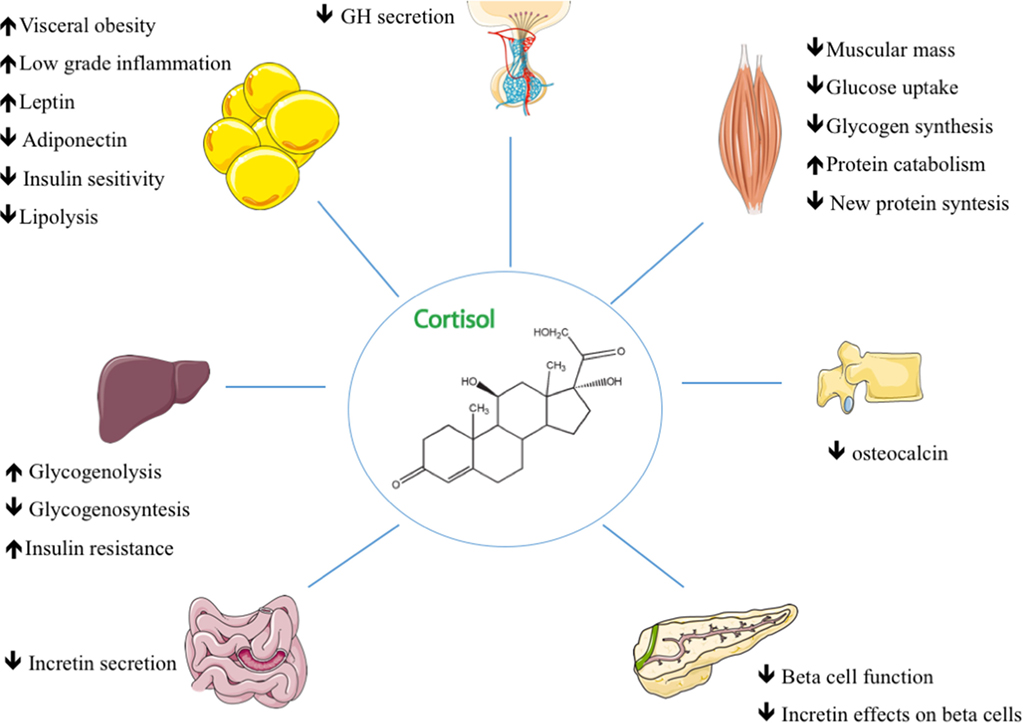
Nello stato di digiuno, il cortisolo stimola diversi processi che servono collettivamente ad aumentare e mantenere le normali concentrazioni di glucosio nel sangue.
Effetti metabolici:
Stimolazione della gluconeogenesi, in particolare nel fegato: Questa via porta alla sintesi del glucosio a partire da substrati non esosi, come gli aminoacidi e il glicerolo proveniente dalla scissione dei trigliceridi, ed è particolarmente importante nei carnivori e in alcuni erbivori. L’aumento dell’espressione degli enzimi coinvolti nella gluconeogenesi è probabilmente la funzione metabolica più nota dei glucocorticoidi.
Mobilitazione di aminoacidi dai tessuti extraepatici: Questi servono come substrati per la gluconeogenesi.
Inibizione della captazione del glucosio nel tessuto muscolare e adiposo: Un meccanismo per conservare il glucosio
Stimolazione della demolizione dei grassi nel tessuto adiposo: Gli acidi grassi rilasciati dalla lipolisi vengono utilizzati per la produzione di energia in tessuti come il muscolo e il glicerolo rilasciato fornisce un altro substrato per la gluconeogenesi.
L’aumento della ritenzione di sodio e dell’escrezione di potassio porta a ipernatremia e ipokaliemia[31].
Aumento della concentrazione di emoglobina, probabilmente dovuto all’ostacolo dell’ingestione di globuli rossi da parte di macrofagi o altri fagociti[32].
Aumento dell’acido urico urinario[33]
Aumento del calcio urinario e ipocalcemia[34]
Alcalosi[35]
Leucocitosi[36]
Il Cortisolo innesca una cascata di eventi che influenzano l’omeostasi del glucosio. Il fegato, i muscoli scheletrici, i tessuti adiposi bianchi e il pancreas svolgono un ruolo chiave nell’assicurare un apporto continuo di energia utilizzabile per la risposta di lotta/fuga.
Livelli eccessivi di glucocorticoidi derivanti dalla somministrazione di farmaci o dall’iperadrenocorticismo hanno effetti su molti sistemi. Alcuni esempi includono l’inibizione della formazione ossea, la soppressione dell’assorbimento del calcio (entrambi possono portare all’osteoporosi), il ritardo nella guarigione delle ferite, la debolezza muscolare e l’aumento del rischio di infezioni. Queste osservazioni suggeriscono una moltitudine di ruoli fisiologici meno drammatici per i glucocorticoidi.[30]
Eccitazione e sfera cognitiva
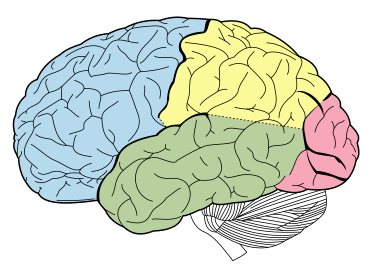
Principali scissure e lobi del cervello visti lateralmente (il lobo frontale è mostrato in blu).
I glucocorticoidi agiscono sull’ippocampo, sull’amigdala e sui lobi frontali. Insieme all’adrenalina, favoriscono la formazione di ricordi flashbulb di eventi associati a forti emozioni, sia positive che negative.[36] Ciò è stato confermato da studi in cui il blocco dell’attività dei glucocorticoidi o della noradrenalina ha compromesso il richiamo di informazioni emotivamente rilevanti. Ulteriori fonti hanno dimostrato che i soggetti il cui apprendimento della paura è stato accompagnato da alti livelli di cortisolo hanno avuto un migliore consolidamento di questa memoria (questo effetto è stato più importante negli uomini). L’effetto che i glucocorticoidi hanno sulla memoria può essere dovuto a un danno specifico all’area CA1 della formazione dell’ippocampo.
In diversi studi sugli animali, lo stress prolungato (che causa aumenti prolungati dei livelli di glucocorticoidi) ha mostrato la distruzione dei neuroni nell’area dell’ippocampo del cervello, che è stata collegata a prestazioni di memoria inferiori.[32][37][33]
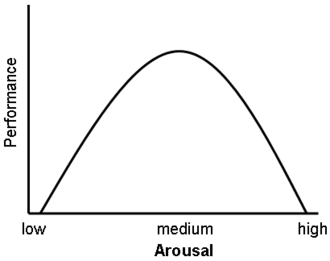
Una rappresentazione grafica della curva di Yerkes-Dodson
È stato inoltre dimostrato che i glucocorticoidi hanno un impatto significativo sulla vigilanza (disturbo da deficit di attenzione) e sulla cognizione (memoria). Questo sembra seguire la curva di Yerkes-Dodson, in quanto gli studi hanno dimostrato che i livelli circolanti di glucocorticoidi rispetto alle prestazioni della memoria seguono un andamento a U rovesciata, proprio come la curva di Yerkes-Dodson. Ad esempio, il potenziamento a lungo termine (LTP, il processo di formazione dei ricordi a lungo termine) è ottimale quando i livelli di glucocorticoidi sono leggermente elevati, mentre si osserva una significativa riduzione dell’LTP dopo la surrenalectomia (stato di basso livello di glucocorticoidi) o dopo la somministrazione di glucocorticoidi esogeni (stato di alto livello di glucocorticoidi). Livelli elevati di glucocorticoidi migliorano la memoria per gli eventi emotivamente eccitanti, ma portano più spesso a una scarsa memoria per il materiale non correlato alla fonte di stress/eccitazione emotiva.[38] In contrasto con gli effetti di potenziamento dose-dipendenti dei glucocorticoidi sul consolidamento della memoria, è stato dimostrato che questi ormoni dello stress inibiscono il recupero di informazioni già memorizzate. [È stato dimostrato che l’esposizione a lungo termine a farmaci glucocorticoidi, come quelli contro l’asma e gli antinfiammatori, crea deficit di memoria e attenzione sia durante che, in misura minore, dopo il trattamento,[39][40] una condizione nota come “demenza da steroidi”.[41]
Omeostasi dei fluidi corporei
I glucocorticoidi potrebbero agire a livello centrale e periferico per contribuire alla normalizzazione del volume dei liquidi extracellulari regolando l’azione dell’organismo nei confronti del peptide natriuretico atriale (ANP). A livello centrale, i glucocorticoidi potrebbero inibire l’assunzione di acqua indotta dalla disidratazione;[42] a livello periferico, i glucocorticoidi potrebbero indurre una potente diuresi.[43]
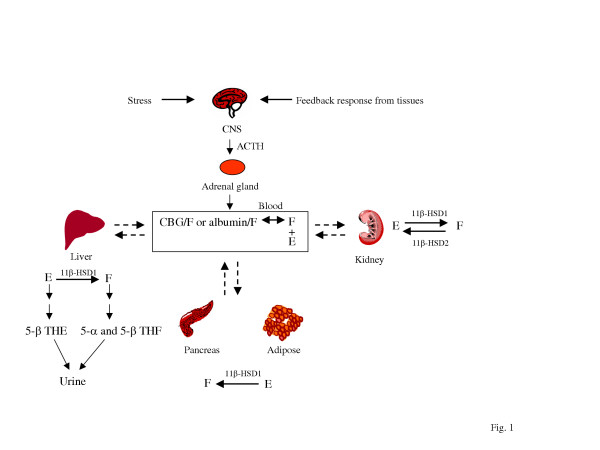
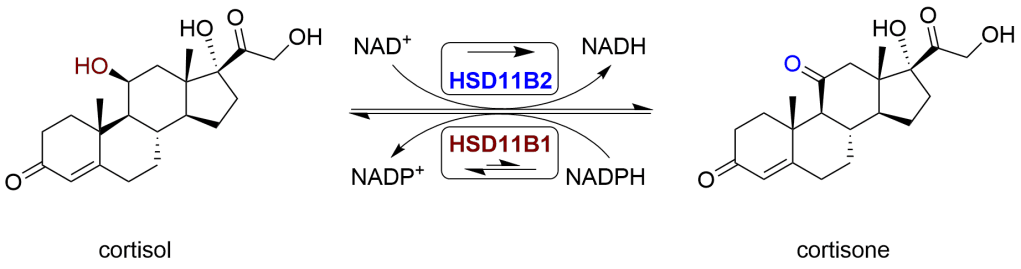
Metabolismo dei Glucocorticoidi. La secrezione di Glucocorticoidi da parte della ghiandola surrenale è regolata dall’asse HPA tramite secrezione di ACTH. Il cortisolo plasmatico principale (F) è legato alle proteine con una frazione libera del 4-5%. Il Cortisone plasmatico (E) è nella forma libera non legata. L’equilibrio di cortisolo e cortisone tra plasma e tessuti è illustrato con le frecce bidirezionali tratteggiate. È anche raffigurato il metabolismo dei GC tessuto-specifici. I GC sono metabolizzati principalmente nel fegato e i metaboliti sono escreti nelle urine. Sono mostrati solo i tessuti rilevanti per la sindrome metabolica. THE, tetraidrocortisone; THF, tetraidrocortisolo.
Meccanismi d’azione dei Glucocorticoidi:
Transattivazione
I glucocorticoidi si legano al recettore citosolico dei glucocorticoidi, un tipo di recettore nucleare che viene attivato dal legame con il ligando. Dopo che un ormone si lega al recettore corrispondente, il complesso appena formato si trasloca nel nucleo della cellula, dove si lega agli elementi di risposta ai glucocorticoidi nella regione promotrice dei geni bersaglio, determinando la regolazione dell’espressione genica. Questo processo viene comunemente definito attivazione trascrizionale o transattivazione.[44][45]
Le proteine codificate da questi geni regolati hanno un’ampia gamma di effetti, tra cui, ad esempio:[45]
L’Annessina [Lipocortina I]
Antinfiammatori – lipocortina I, proteina legante p11/calpactina, inibitore secretorio della proteasi leucocitaria 1 (SLPI) e fosfatasi della proteina chinasi attivata dal mitogeno (MAPK fosfatasi).
Aumento della gluconeogenesi – glucosio 6-fosfatasi e tirosina aminotransferasi
Transrepressione
Il meccanismo opposto è chiamato repressione trascrizionale o transrepressione. Secondo la concezione classica di questo meccanismo, il recettore dei glucocorticoidi attivato si lega al DNA nello stesso sito in cui si legherebbe un altro fattore di trascrizione, impedendo la trascrizione di geni che vengono trascritti tramite l’attività di quel fattore.[44][45] Sebbene ciò avvenga, i risultati non sono coerenti per tutti i tipi di cellule e per tutte le condizioni; non esiste un meccanismo generale e generalmente accettato per la transrepressione.[45]
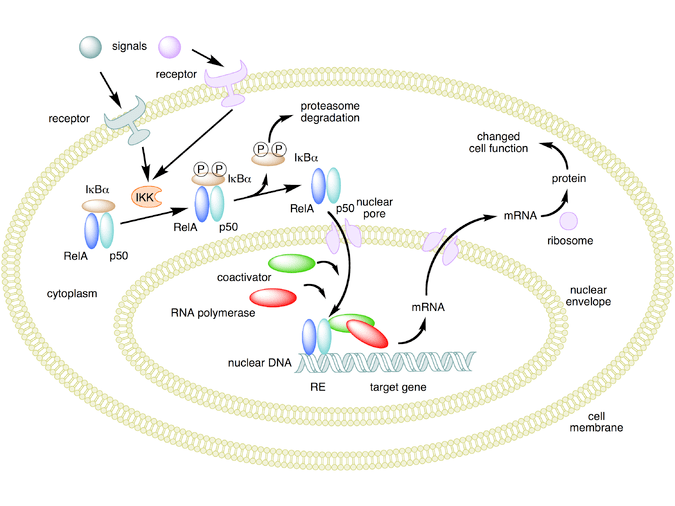
Meccanismo d’azione di NF-κB.
Si stanno scoprendo nuovi meccanismi in cui la trascrizione viene repressa, ma il recettore dei glucocorticoidi attivato non interagisce con il DNA, bensì direttamente con un altro fattore di trascrizione, interferendo con esso, o con altre proteine che interferiscono con la funzione di altri fattori di trascrizione. Quest’ultimo meccanismo sembra essere il modo più probabile in cui il recettore glucocorticoide attivato interferisce con NF-κB, ossia reclutando istone deacetilasi, che deacetilano il DNA nella regione del promotore portando alla chiusura della struttura cromatinica in cui NF-κB deve legarsi.[44][45]
Attività non-genomica
Il recettore glucocorticoide attivato ha effetti che, come è stato dimostrato sperimentalmente, sono indipendenti da qualsiasi effetto sulla trascrizione e possono essere dovuti solo al legame diretto del recettore glucocorticoide attivato con altre proteine o con l’mRNA.[44][45]
Effetti genomici e non genomici dei Glucocorticoidi. Trans-attivazione: l’effetto genomico del GC dopo il legame del GR al suo elemento di risposta positiva causa una maggiore trascrizione di proteine antinfiammatorie, ad esempio, lipocortina-1, IL-10, IL-12, MAPK fosfatasi I e IκB. Trans-repressione: l’interazione molecola-molecola tra GR attivato e fattori di trascrizione pro-infiammatori, ad esempio, AP-1 o NF-κB causa una riduzione della trascrizione di mediatori pro-infiammatori, ad esempio, Il-2, IL-3, IL-4, IL-5, IL-6, IL-13, IL-15, TNF-α e VCAM-a.
Ad esempio, la chinasi Src, che si lega al recettore glucocorticoide inattivo, viene rilasciata quando un glucocorticoide si lega al recettore glucocorticoide e fosforila una proteina che a sua volta sposta una proteina adattatrice da un recettore importante nell’infiammazione, il fattore di crescita epidermico, riducendone l’attività, che a sua volta si traduce in una riduzione della creazione di acido arachidonico, una molecola proinfiammatoria chiave. Questo è uno dei meccanismi con cui i glucocorticoidi hanno un effetto antinfiammatorio.[44]
Farmacologia dei Glucocorticoidi:
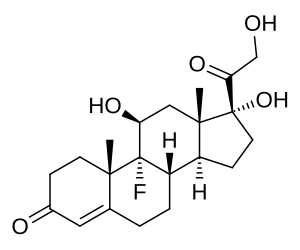
Fludrocortisone Acetato
Per uso terapeutico sono stati creati diversi Glucocorticoidi sintetici, alcuni molto più potenti del Cortisolo. Si differenziano sia per la farmacocinetica (fattore di assorbimento, emivita, volume di distribuzione, clearance) che per la farmacodinamica (ad esempio la capacità di attività mineralcorticoide: ritenzione di sodio (Na+) e acqua; fisiologia renale). Poiché permeano facilmente l’intestino, vengono somministrati principalmente per os (per bocca), ma anche con altri metodi, ad esempio per via topica sulla pelle. Oltre il 90% di essi lega diverse proteine plasmatiche, anche se con una diversa specificità di legame. I glucocorticoidi endogeni e alcuni corticoidi sintetici hanno un’elevata affinità con la proteina Transcortina (detta anche globulina legante i corticosteroidi), mentre tutti legano l’albumina. Nel fegato, vengono rapidamente metabolizzati mediante coniugazione con un solfato o un acido glucuronico e vengono secreti nelle urine.
La potenza dei Glucocorticoidi, la durata dell’effetto e la sovrapposizione della potenza dei mineralocorticoidi variano. Il cortisolo è lo standard di confronto per la potenza dei glucocorticoidi. Idrocortisone è il nome utilizzato per le preparazioni farmaceutiche di cortisolo.
I dati riportati di seguito si riferiscono alla somministrazione orale. La potenza orale può essere inferiore a quella parenterale perché quantità significative (fino al 50% in alcuni casi) possono non raggiungere la circolazione. Il Fludrocortisone Acetato e il desossicorticosterone acetato sono, per definizione, mineralocorticoidi piuttosto che glucocorticoidi, ma hanno una potenza glucocorticoide minore e sono inclusi in questa tabella per fornire una prospettiva sulla potenza dei mineralocorticoidi.
Usi terapeutici:
I Glucocorticoidi possono essere utilizzati a basse dosi nell’insufficienza surrenalica. A dosi molto più elevate, i glucocorticoidi per via orale o inalatoria sono utilizzati per sopprimere vari disturbi allergici, infiammatori e autoimmuni. I glucocorticoidi per via inalatoria sono il trattamento di seconda linea per l’asma. Sono anche somministrati come immunosoppressori post-trapianto per prevenire il rigetto acuto del trapianto e la malattia del trapianto contro l’ospite. Tuttavia, non prevengono un’infezione e inibiscono anche i successivi processi riparativi. Nuove evidenze hanno dimostrato che i glucocorticoidi potrebbero essere utilizzati nel trattamento dell’insufficienza cardiaca per aumentare la responsività renale ai diuretici e ai peptidi natriuretici. I glucocorticoidi sono storicamente utilizzati per alleviare il dolore nelle condizioni infiammatorie.[46][47][48] Tuttavia, i corticosteroidi mostrano un’efficacia limitata nell’alleviare il dolore e potenziali eventi avversi per il loro uso nelle tendinopatie.[49]
Terapia Sostitutiva
Cortisolo
Qualsiasi glucocorticoide può essere somministrato in una dose che fornisce all’incirca gli stessi effetti glucocorticoidi della normale produzione di cortisolo; si parla di dosaggio fisiologico, sostitutivo o di mantenimento. Si tratta di circa 6-12mg/m2/die di Idrocortisone (m2 si riferisce all’area di superficie corporea (BSA), ed è una misura delle dimensioni del corpo; la BSA di un uomo medio è di 1,9 m2).
Gli usi clinici dei glucocorticoidi comprendono quindi:
Terapia Immunosoppressiva: I glucocorticoidi causano immunosoppressione e la componente terapeutica di questo effetto è principalmente la diminuzione della funzione e del numero di linfociti, compresi i linfociti B e i linfociti T.
Terapia Anti-Infiammatoria: I glucocorticoidi sono potenti antinfiammatori, indipendentemente dalla causa dell’infiammazione; il loro meccanismo antinfiammatorio primario è la sintesi della lipocortina-1 (annexin-1). La lipocortina-1 sopprime la fosfolipasi A2, bloccando così la produzione di eicosanoidi, e inibisce vari eventi infiammatori dei leucociti (adesione epiteliale, emigrazione, chemiotassi, fagocitosi, esplosione respiratoria, ecc.) In altre parole, i glucocorticoidi non solo sopprimono la risposta immunitaria, ma inibiscono anche i due principali prodotti dell’infiammazione, le prostaglandine e i leucotrieni. Inibiscono la sintesi delle prostaglandine a livello della fosfolipasi A2 e a livello della cicloossigenasi/PGE isomerasi (COX-1 e COX-2),[50] quest’ultimo effetto è molto simile a quello dei FANS, potenziando così l’effetto antinfiammatorio. Inoltre, i glucocorticoidi sopprimono anche l’espressione della ciclossigenasi.[51]
Trattamento del Iperaldosteronismo: I glucocorticoidi possono essere utilizzati nella gestione dell’iperaldosteronismo familiare di tipo 1. Non sono efficaci, tuttavia, per l’uso nella condizione di tipo 2.
Trattamento insufficienza cardiaca: I glucocorticoidi possono essere utilizzati nel trattamento dell’insufficienza cardiaca scompensata per potenziare la reattività renale ai diuretici, in particolare nei pazienti con insufficienza cardiaca con resistenza diuretica refrattaria a dosi elevate di diuretici dell’ansa.[52][53][54][55][56][57][58]
Meccanismi di resistenza ai corticosteroidi
La resistenza agli usi terapeutici dei glucocorticoidi può verificarsi in un certo numero di pazienti e presentare delle difficoltà; ad esempio, il 25% dei casi di asma grave può non rispondere agli steroidi. Questo può essere il risultato di una predisposizione genetica, dell’esposizione continua alla causa dell’infiammazione (come gli allergeni), di fenomeni immunologici che bypassano i glucocorticoidi, di disturbi farmacocinetici (assorbimento incompleto o escrezione o metabolismo accelerati) e di infezioni respiratorie virali e/o batteriche.[59][60]
Glucocorticoidi e Sport:
Come abbiamo visto, i Glucocorticoidi sono una delle classi di farmaci più ampiamente utilizzate ed efficaci nella popolazione generale e sono disponibili in una varietà di formulazioni farmaceutiche (ad esempio, iniezioni, compresse, creme, colliri, gocce auricolari, inalatori e spray nasali). Somministrati sia per i loro effetti sistemici che locali, i Glucocorticoidi sono utilizzati a livello globale in una vasta gamma di specialità cliniche, principalmente per le loro proprietà antinfiammatorie e immunosoppressive. In alcuni contesti, l’uso medico dei Glucocorticoidi orali sembra essere aumentato negli ultimi anni poiché questi sono un’alternativa accessibile e conveniente ai farmaci mirati ma più costosi. La prevalenza dell’uso sistemico prevalentemente per uso a breve termine varia tra l’1% e il 3%, sebbene abbia raggiunto il 17,1% in un recente studio sugli adulti in Francia.(https://bjsm.bmj.com) Nelle popolazioni di atleti, vi è una maggiore prevalenza di lesioni muscoloscheletriche e asma, e pertanto un frequente uso legittimo di Glucocorticoidi terapeutici non sarebbe sorprendente. Tuttavia, vi è una scarsità di stime di prevalenza nelle popolazioni di atleti. Un’analisi di TUE abbreviate in cui il CIO è stato informato dell’uso di Glucocorticoidi da parte degli atleti prima dei Giochi olimpici negli anni ’90 e nei primi anni 2000 suggerisce che almeno il 5% al 12% degli atleti d’élite competitivi è stato trattato con Glucocorticoidi tramite tutte le vie, prevalentemente inalatoria. In un recente sondaggio internazionale non pubblicato di medici che lavorano con atleti d’élite, oltre l’85% ha riferito di aver somministrato almeno occasionalmente Glucocorticoidi iniettabili come parte della loro normale pratica (comunicazione personale, Dr David Hughes, Australian Institute of Sport).
I Glucocorticoidi, somministrati tramite determinate vie, sono stati proibiti per la prima volta nello sport dal CIO nel 1985 e sono stati proibiti dalla WADA sin dalla sua Lista iniziale, pubblicata nel 2004. Le sostanze o i metodi sono considerati per l’inclusione nella Lista se soddisfano due dei tre criteri seguenti come stabilito dal Codice mondiale antidoping:
potenziale di migliorare o migliorano le prestazioni sportive;
rappresentano un rischio effettivo o potenziale per la salute dell’atleta;
violano lo spirito dello sport. I Glucocorticoidi sono proibiti in competizione quando somministrati tramite vie “sistemiche” (orali, rettali, intramuscolari o endovenose).[https://www.wada-ama.org/] La somministrazione tramite tutte le altre vie (incluse le iniezioni intra-articolari e altre periarticolari) è considerata somministrazione locale e non è proibita in competizione. La somministrazione di Glucocorticoidi tramite qualsiasi via non è proibita fuori competizione (OOC).
Indipendentemente dalla sostanza specifica del Glucocorticoidi e dalle sue singole caratteristiche farmacologiche, un presunto riscontro analitico avverso (AAF) viene segnalato dai laboratori accreditati WADA quando i livelli urinari dei campioni in gara superano un livello di segnalazione di 30ng/mL. La farmacocinetica dei Glucocorticoidi è complessa e influenzata dalla formulazione, dal tipo di esterificazione e sale, dalla via di somministrazione, dal sito e dal metodo di somministrazione. Di conseguenza, mentre il limite di segnalazione del laboratorio può dimostrare la presenza di un Glucocorticoidi , non può necessariamente indicare se la somministrazione è avvenuta in gara o OOC o se è probabile che abbia un effetto farmacologico o ergogenico. Qualsiasi medico o atleta non sarà sicuro di quando interrompere l’uso di GC sistemici prima del periodo in gara per evitare di superare il limite di segnalazione. Per complicare ulteriormente il quadro farmacocinetico, le iniezioni intra-articolari possono dare origine a livelli sistemici e i medici possono inavvertitamente caratterizzare erroneamente il sito di iniezione in assenza di guida radiologica o ecografica. La definizione di limiti di segnalazione specifici per sostanza è un’area di discussione e ricerca attiva tra gli esperti nominati dalla WADA e va oltre lo scopo del presente documento.
Glucocorticoidi sistemici e performance
Alcuni atleti hanno indubbiamente tentato di sfruttare i presunti effetti di miglioramento delle prestazioni dei Glucocorticoidi sistemici che ritengono benefici nella loro particolare disciplina sportiva. Tuttavia, i meccanismi complessi e pleiotropici dell’azione dei Glucocorticoidi suggeriscono che questi farmaci sono uno strumento poco maneggevole per l’atleta che cerca di ottenere un vantaggio nelle prestazioni e sono considerati una componente meno popolare dei regimi di doping rispetto al passato.[ https://cyclingtips.com/] Alcuni pazienti e atleti hanno riferito di aver sperimentato euforia dopo la somministrazione sistemica.[ http://www.cyclingnews.com] Tuttavia, le prove scientifiche a supporto dell’euforia misurabile nelle popolazioni cliniche sono ambigue e l’interpretazione dei dati è complicata dall’associazione del dolore cronico confondente.[https://bjsm.bmj.com/]
Sembrerà starno, ma non vi è alcuna prova incontrovertibile di effetti di miglioramento delle prestazioni derivanti dall’uso a breve termine di Glucocorticoidi sistemici.[https://bjsm.bmj.com/] Esistono studi randomizzati in doppio cieco cross-over che suggeriscono che gli atleti possono sfruttare cicli di Glucococrticoidi orali ad alto dosaggio della durata di una settimana per migliorare le loro prestazioni di esercizio di intensità submassimale per brevi periodi di tempo.[https://bjsm.bmj.com/] Questi dosaggi sarebbero facilmente rilevati durante i test antidoping, se assunti in gara. Il meccanismo preciso di questo effetto non è chiaro, ma si suggerisce che derivi da una combinazione di effetti sul metabolismo energetico, sui muscoli, sull’infiammazione e sul sistema nervoso. Questo effetto del farmaco è stato dimostrato in uno studio su atleti maschi il cui allenamento era strettamente periodizzato insieme all’uso di Glucocorticoidi orali.[https://bjsm.bmj.com/] Sfruttare questo tipo di regime di miglioramento delle prestazioni evitando efficacemente l’insufficienza surrenalica e il rilevamento tramite controlli antidoping standard in gara richiederebbe una meticolosa supervisione medica. Potrebbe anche richiedere una manipolazione farmacologica più complessa ed esotica dell’asse ipotalamo-ipofisi-surrene rispetto a quella offerta dai Glucocorticoidi prescritti.[https://bjsm.bmj.com/]
Atleti e dottori hanno descritto metodi inappropriati con cui l’uso sistemico di Glucocorticoidi, un’alimentazione limitata e un allenamento a bassa intensità potrebbero essere combinati OOC per perdere peso e preservare la massa muscolare.[ https://www.nytimes.com/]Tuttavia, date le funzioni cataboliche proteiche ampiamente riconosciute dei Glucocorticoidi,[https://bjsm.bmj.com/] questo meccanismo di doping rimane speculativo e controverso. Inoltre, l’efficacia potrebbe dipendere dall’uso di Glucocorticoidi come parte di un cocktail complesso che include altri ormoni proibiti ma scarsamente rilevati come l’insulina.[https://bjsm.bmj.com/]
Schema esemplificativo sull’uso di Glucocorticoidi sistemici e perdita di peso/ricomposizione corporea:la base teorica sulla quale si sostiene la suddetta pratica farmacologica è il ciclo di feedback negativo dell’Asse HPA per via d’uso di Glucocorticoidi esogeni. Tale pratica dovrebbe portare a 1) soppressione del rilascio di —> CRH>ACTH>Cortisone<>Cortisolo con consequenziale prevenzione di 2) aumento della fame/appetito con il procedere del regime ipocalorico 3) prevenzione del aumento del catabolismo muscolare 4) prevenzione di un aumento della ritenzione idrica e 5) prevenzione dell’alterazione del metabolismo lipidico correlato ad un incremento significativo del Cortisolo. Da notare che il momento della somministrazione del Glucocorticoide esogeno può influenzare il grado di soppressione surrenalica. Per esempio, il Prednisone in una dose di 5mg somministrato la sera prima di coricarsi e 2,5mg al mattino produrrà una soppressione dell’Asse HPA più marcata rispetto a 2,5mg la sera e 5mg al mattino. 5mg è un dosaggio basso e generalmente non è promotore di insonnia sebbene in soggetti sensibili può manifestarsi. Il dosaggio comunemente utilizzato varia da 15 a 25mg/die diviso in due dosi dopo i pasti. Tale pratica comunemente è parte di protocolli PEDs più complessi e contenenti uno o più agenti anabolizzanti.
Recenti resoconti sulla presunta potenza dei Glucocorticoidi sistemici provengono da atleti che hanno anche confessato l’uso concomitante di altri metodi e sostanze per migliorare le prestazioni, tra cui agenti anabolizzanti come il testosterone.[https://bjsm.bmj.com/] Tali regimi di Glucocorticoidi potrebbero avere rilevanza solo in un piccolo sottoinsieme di discipline sportive, come nelle ripide tappe di montagna dei Grandi Giri del ciclismo, dove gli atleti potrebbero essere disposti ad accettare compromessi nei loro regimi di allenamento o potenza assoluta in uscita nel perseguimento di un rapporto potenza/peso superiore. L’uso di OOC richiederebbe comunque una continuazione prolungata dell’uso di Glucocorticoidi nel periodo di gara per evitare l’insufficienza surrenalica dovuta a meccanismi di feedback. L’uso prolungato di Glucocorticoidi comporta rischi medici ben noti, alcuni dei quali potrebbero ridurre in modo permanente le prestazioni atletiche.[https://bjsm.bmj.com/]
Rischi per la salute, eventi avversi ed effetti negativi sulle prestazioni
Il trattamento con Glucocorticoidi per molte condizioni ha una lunga storia e un profilo di sicurezza ragionevole. Dosi elevate o uso cronico di Glucocorticoidi sistemici presentano un certo rischio per la salute dell’atleta. Un esame attento, una diagnosi e una deliberazione da parte del medico sono fondamentali e i benefici del trattamento devono essere soppesati rispetto ai potenziali rischi ed effetti avversi. L’uso potenziale per migliorare le prestazioni, descritto sopra e ritenuto limitato a contesti sportivi specifici con uso di GC ad alto dosaggio, è anche potenzialmente associato a rischi significativi per la salute di un atleta.
Gli eventi avversi con associazioni causali ben consolidate all’uso clinicamente appropriato di GC toccano praticamente ogni sistema umano, vanno da esiti negativi sulla salute acuti a cronici e includono insufficienza surrenalica, immunodeficienza, osteoporosi, atrofia muscolare, cedimento di tendini/fasce, necrosi avascolare della testa femorale, vari squilibri elettrolitici, nutrizionali e metabolici, glaucoma e cataratta. Forse perché i GC sono farmaci così comuni e clinicamente versatili, alcuni medici potrebbero sopravvalutare il loro valore terapeutico e sottostimare la gravità degli eventi avversi associati.[https://bjsm.bmj.com/] Anche una singola iniezione intra-articolare potrebbe causare un’insufficienza surrenalica clinicamente significativa che porta a malessere, squilibrio elettrolitico e immunosoppressione per diverse settimane.[https://bjsm.bmj.com/]
È importante sottolineare che l’eziologia di questi sintomi potrebbe non essere riconosciuta dall’atleta e dal personale medico, in particolare in un contesto sportivo in cui gli atleti si allenano ad alta intensità e i sintomi possono mascherarsi da affaticamento correlato al sovrallenamento. Inoltre, un atleta che subisce un trauma o un infortunio grave potrebbe essere a maggior rischio di crisi surrenalica a causa della soppressione ipotalamo-ipofisi-surrene dovuta al precedente utilizzo di GC. Ciò potrebbe essere particolarmente problematico se l’atleta non rivela questo precedente utilizzo.
Sia l’efficacia che il potenziale danno delle iniezioni intra-articolari sono ampiamente dibattuti. Le prove di un recente studio prospettico controllato con placebo su pazienti con osteoartrite hanno suggerito che frequenti iniezioni di triamcinolone al ginocchio, somministrate secondo un programma prestabilito, non sono riuscite a gestire efficacemente il dolore a lungo termine e hanno portato a una riduzione statisticamente significativa dello spessore della cartilagine.[https://bjsm.bmj.com/]Tuttavia, le raccomandazioni della società medica, così come una meta-analisi completa, supportano l’efficacia e la sicurezza dello stesso intervento,[https://bjsm.bmj.com/l] suggerendo fortemente che un uso giudizioso di iniezioni intra-articolari in pazienti e circostanze appropriate può produrre risultati positivi. Vi è una mancanza di prove pubblicate sulla sicurezza o il danno dell’uso di GC intra-articolari nelle popolazioni di atleti e sono urgentemente necessarie ulteriori ricerche a causa dell’uso onnipresente di GC intra-articolari.
Politiche per garantire l’uso appropriato dei GC
Nonostante le preoccupazioni di un possibile abuso per un vantaggio competitivo o potenziali effetti dannosi sulla salute degli atleti, i GC sono ampiamente utilizzati nello sport per legittime ragioni terapeutiche. Considerando che l’elenco è armonizzato in tutti gli sport, dal tiro con l’arco al wakeboard, il doping con i GC non è un problema laddove i presunti benefici dell’uso di GC ad alto dosaggio (potenza prolungata a intensità di esercizio submassimali o gestione aggressiva del peso catabolico) difficilmente miglioreranno le prestazioni. Pertanto, un AAF per i GC non sarebbe probabilmente associato a nessun intento di doping. L’uso di GC sistemici in molti sport deve essere considerato sotto una luce diversa rispetto agli sport ad alto rischio come il ciclismo, dove l’abuso è ben documentato e le prove scientifiche forniscono un certo supporto.
Consapevoli delle sfide specifiche poste dall’uso di GC nello sport, le organizzazioni sportive e antidoping hanno introdotto politiche innovative e stanno rafforzando le normative esistenti per affrontare l’uso terapeutico ragionevole dei GC.
Conclusioni su Glucocorticoidi e prestazioni sportive:
Da recenti review, sappiamo che l’uso dei Glucocorticoidi sistemici può migliorare la prestazione fisica massima rispetto al placebo (SDM 0,300, 95% CI 0,080-0,520). In una recente review [https://www.frontiersin.org/], l’SDM per i 13 confronti inclusi non era eterogeneo (I2 = 35%, p = 0,099). L’analisi di sensibilità escludendo i due studi con alto rischio di bias ha mostrato un effetto simile (SDM 0,349, 95% CI 0,071-0,626). Con la meta-regressione si è scoperto che la durata del trattamento, la via di somministrazione e il tipo di esercizio non hanno influenzato (p > 0,124) l’SDM. Nell’analisi stratificata il trattamento prolungato e l’ingestione orale hanno migliorato la prestazione fisica (p = 0,003). Il trattamento acuto e l’inalazione non hanno avuto alcun effetto sulla prestazione fisica (p > 0,564), l’analisi di sensibilità con studi ad alto rischio di bias rimossi o solo un trattamento per gruppo di controllo, ha mostrato un effetto simile all’analisi completa con SDM 0,334, 95% CI 0,075-0,592 e SDM 0,296 0,059-0,532, rispettivamente. L’analisi di sensibilità escludendo i sei confronti con meno di 10 coppie di dati non ha indicato alcun effetto dei Glucocorticoidi sulla prestazione fisica (p = 0,070). I Glucocorticoidi hanno migliorato la prestazione aerobica (SDM 0,348, 95% CI 0,129-0,567). Tre confronti hanno testato l’effetto dei Glucocorticoidi sulla prestazione anaerobica massima e la meta-analisi dei confronti non ha mostrato alcun effetto (p = 0,573) sulla prestazione fisica. L’effetto è rimasto non statisticamente significativo dopo aver incluso i due studi che misuravano la prestazione anaerobica all’interno di un test di prestazione aerobica (p = 0,491) e quando tutti i risultati della prestazione anaerobica negli studi inclusi (anche più risultati dello stesso studio) sono stati meta-analizzati (p = 0,177). Non è stato trovato alcun effetto dei Glucocorticoidi sulla spesa energetica durante la prestazione submassimale (SDM 0,-332, 95% CI −0,785 a 0,121).
In una review narrativa del 2016, Collomp et al. hanno mirato a riassumere le attuali conoscenze sugli effetti ergogenici dei glucocorticoidi negli esseri umani. Hanno riferito che un effetto ergogenico (sull’esercizio di endurance) dei Glucocorticoidi sistemici a breve termine è stato chiaramente dimostrato e che gli effetti a breve termine (4,5 giorni) e a lungo termine (4 settimane) dell’assunzione di Glucocorticoidi non hanno avuto alcun effetto sul VO2-max o sulla potenza massima in uscita durante protocolli di esercizio graduati. Hanno anche il test sul campo come categoria di test delle prestazioni, ma fanno riferimento solo allo studio di Casuso et al. che riporta un miglioramento delle prestazioni nella corsa a navetta, ma nessun cambiamento nelle prestazioni nello sprint. I risultati successivi supportano e rafforzano la conclusione di Collomp et al. ( per quanto riguarda l’esercizio di endurance poiché scopriamo anche che il trattamento prolungato con glucocorticoidi migliora le prestazioni aerobiche. Tuttavia, a differenza di Collomp et al. nelle ultime review sono stati inclusi sia test di esercizio graduati che test sul campo (della durata di 1 minuto o più) nella definizione di prestazione aerobica e quindi, più studi che hanno aggiunto potenza statistica alla analisi. Non è stato trovato alcun effetto dei Glucocorticoidi sulla prestazione anaerobica quando è stato analizzato secondo il protocollo di studio, tuttavia sono stati inclusi solo tre studi che testavano la prestazione anaerobica. Per aumentare la potenza statistica, è stato anche incluso la prestazione anaerobica all’interno di test aerobici e più test anaerobici dallo stesso studio, ma ancora non era evidente alcun effetto dei glucocorticoidi. Questo approccio può diminuire la validità dell’analisi poiché la prestazione anaerobica all’interno di test aerobici può testare altre abilità rispetto ai test anaerobici e la meta-analisi di più risultati dagli stessi soggetti e l’intervento non è raccomandato dal Cochrane Handbook. Collomp et al. concludono che non è chiaro se i Glucocorticoidi migliorino la prestazione durante l’esercizio breve-intenso. Questa conclusione è ancora valida poiché solo pochi studi hanno indagato l’effetto dei glucocorticoidi sull’esercizio anaerobico/breve intenso. Tuttavia, quando meta-analizziamo tutte le prove disponibili, sembra che i Glucocorticoidi non migliorino le prestazioni anaerobiche. I Glucocorticoidi non hanno influenzato le prestazioni submassimali aumentando l’energia totale spesa e/o il VO2max a un carico fisso, ma questa conclusione dovrebbe anche essere interferita con cationi poiché l’analisi include solo 35 soggetti, il che fornisce una potenza statistica limitata.
L’uso di Glucocorticoidi nello sport è una questione altamente complessa a causa del loro uso diffuso in medicina, delle numerose formulazioni e vie di somministrazione con farmacocinetica variabile, effetti negativi sulla salute e potenziali associazioni di doping.
In definitiva, da quanto recentemente emerso attraverso il riassunto delle migliori prove scientifiche disponibili, i Glucocorticoidi migliorano le prestazioni aerobiche e massime, ma non influenzano le prestazioni anaerobiche nei soggetti sani.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Pelt AC (2011). Glucocorticoids: effects, action mechanisms, and therapeutic uses. Hauppauge, N.Y.: Nova Science. ISBN978-1617287589.[page needed]
Ralph CR, Tilbrook AJ (February 2016). “INVITED REVIEW: The usefulness of measuring glucocorticoids for assessing animal welfare”. Journal of Animal Science. 94 (2): 457–470. doi:10.2527/jas.2015-9645. PMID27065116.
Collomp K, Arlettaz A, Buisson C, Lecoq AM, Mongongu C. Glucocorticoid administration in athletes: performance, metabolism and detection. Steroids. (2016) 115:193–202. doi: 10.1016/j.steroids.2016.09.008 PubMed Abstract | CrossRef Full Text | Google Scholar
Fain JN, Cheema P, Tichansky DS, Madan AK. Stimulation of human omental adipose tissue lipolysis by growth hormone plus dexamethasone. Mol Cell Endocrinol. (2008) 295(1):101–5. doi: 10.1016/j.mce.2008.05.014PubMed Abstract | CrossRef Full Text | Google Scholar
Thomasson R, Rieth N, Jollin L, Amiot V, Lasne F, Collomp K. Short-term glucocorticoid intake and metabolic responses during long-lasting exercise. Horm Metab Res. (2011) 43(3):216–22. doi: 10.1055/s-0030-1269919 PubMed Abstract | CrossRef Full Text | Google Scholar
Duclos M, Guinot M, Le Bouc Y, Cortisol GH. Odd and controversial ideas. Appl Physiol Nutr Metab. (2007) 32(5):895–903. doi: 10.1139/h07-064PubMed Abstract | CrossRef Full Text | Google Scholar
Sapolsky RM, Romero LM, Munck AU. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev. (2000) 21(1):55–89. doi: 10.1210/edrv.21.1.0389PubMed Abstract | CrossRef Full Text | Google Scholar
Duclos M. Evidence on ergogenic action of glucocorticoids as a doping agent risk. Phys Sportsmed. (2010) 38(3):121–7. doi: 10.3810/psm.2010.10.1817PubMed Abstract | CrossRef Full Text | Google Scholar
Strehl C, Bijlsma JWJ, de Wit M, Boers M, Caeyers N, Cutolo M, et al. Defining conditions where long-term glucocorticoid treatment has an acceptably low level of harm to facilitate implementation of existing recommendations: viewpoints from an EULAR task force. Ann Rheum Dis. (2016) 75(6):952–7. doi: 10.1136/annrheumdis-2015-20891PubMed Abstract | CrossRef Full Text | Google Scholar
Ventura R, Daley-Yates P, Mazzoni I, Collomp K, Saugy M, Buttgereit F, et al. A novel approach to improve detection of glucocorticoid doping in sport with new guidance for physicians prescribing for athletes. Br J Sports Med. (2021) 55(11):631–42. doi: 10.1136/bjsports-2020-103512
Zhou M, Gomez-Sanchez CE (July 1993). “Cloning and expression of a rat cytochrome P-450 11 beta-hydroxylase/aldosterone synthase (CYP11B2) cDNA variant”. Biochem. Biophys. Res. Commun. 194 (1): 112–7. doi:10.1006/bbrc.1993.1792. PMID8333830.
Lefebvre H, Cartier D, Duparc C, et al. (March 2002). “Characterization of serotonin(4) receptors in adrenocortical aldosterone-producing adenomas: in vivo and in vitro studies”. J. Clin. Endocrinol. Metab. 87 (3): 1211–6. doi:10.1210/jcem.87.3.8327. PMID11889190.
Lupien SJ, McEwen BS, Gunnar MR, Heim C (Jun 2009). “Effects of stress throughout the lifespan on the brain, behaviour and cognition”. Nature Reviews. Neuroscience. 10 (6): 434–445. doi:10.1038/nrn2639. PMID19401723. S2CID205504945.
Belanoff JK, Gross K, Yager A, Schatzberg AF (2001). “Corticosteroids and cognition”. Journal of Psychiatric Research. 35 (3): 127–145. doi:10.1016/S0022-3956(01)00018-8. PMID11461709.
Lupien SJ, Maheu F, Tu M, Fiocco A, Schramek TE (Dec 2007). “The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition”. Brain and Cognition. 65 (3): 209–237. doi:10.1016/j.bandc.2007.02.007. PMID17466428. S2CID5778988.
Wolkowitz OM, Lupien SJ, Bigler ED (Jun 2007). “The ‘steroid dementia syndrome’: a possible model of human glucocorticoid neurotoxicity”. Neurocase. 13 (3): 189–200. doi:10.1080/13554790701475468. PMID17786779. S2CID39340010.
Liu C, Chen Y, Kang Y, Ni Z, Xiu H, Guan J, et al. (Oct 2011). “Glucocorticoids improve renal responsiveness to atrial natriuretic peptide by up-regulating natriuretic peptide receptor-A expression in the renal inner medullary collecting duct in decompensated heart failure”. The Journal of Pharmacology and Experimental Therapeutics. 339 (1): 203–209. doi:10.1124/jpet.111.184796. PMID21737535. S2CID1892149.
Newton R, Holden NS (Oct 2007). “Separating transrepression and transactivation: a distressing divorce for the glucocorticoid receptor?”. Molecular Pharmacology. 72 (4): 799–809. doi:10.1124/mol.107.038794. PMID17622575. S2CID52803631.
Rado JP, Blumenfeld G, Hammer S (Nov 1959). “The effect of prednisone and 6-methylprednisolone on mercurial diuresis in patients with refractory cardiac edema”. The American Journal of the Medical Sciences. 238 (5): 542–551. doi:10.1097/00000441-195911000-00003. PMID14435747. S2CID38687480.
Riemer AD (Apr 1958). “Application of the newer corticosteroids to augment diuresis in congestive heart failure”. The American Journal of Cardiology. 1 (4): 488–496. doi:10.1016/0002-9149(58)90120-6. PMID13520608.
Newman DA (Feb 1959). “Reversal of intractable cardiac edema with prednisone”. New York State Journal of Medicine. 59 (4): 625–633. PMID13632954.
Massari F, Mastropasqua F, Iacoviello M, Nuzzolese V, Torres D, Parrinello G (Mar 2012). “The glucocorticoid in acute decompensated heart failure: Dr Jekyll or Mr Hyde?”. The American Journal of Emergency Medicine. 30 (3): 517.e5–10. doi:10.1016/j.ajem.2011.01.023. PMID21406321.
Il collagene (o collageno) è la principale proteina fibrosa del tessuto connettivo negli animali. È la proteina più abbondante nei mammiferi (circa il 25% della massa proteica totale), rappresentando nell’uomo circa il 6% del peso corporeo.
Il collagene, quindi, è il componente strutturale primario di tutti i tessuti connettivi (principalmente collagene di tipo I per ossa, tendini e legamenti; e principalmente collagene di tipo II per la cartilagine). L’attività del collagene di tipo III riflette la crescita e il turnover dei tessuti molli; è ampiamente distribuito nelle fibrille interstiziali dei tessuti molli, compresi tendini e legamenti, fascia, nonché nella matrice extracellulare (ECM) e nei suoi strati (ad esempio, l’endomisio) che avvolgono le parti costitutive dei gruppi muscolari e che contengono vari fattori di crescita coinvolti nella funzione e nella crescita del muscolo scheletrico.
Il metabolismo del collagene comprende i processi biochimici che regolano la sintesi, la degradazione e il ricambio dei tessuti molli che sono costituiti principalmente da collagene. I tessuti connettivi del nostro corpo sono in uno stato costante di equilibrio e flusso. Il metabolismo del collagene mantiene l’integrità strutturale e la funzione di articolazioni, ossa, tendini e legamenti del corpo. Gli enzimi e le vie di segnalazione regolano questi processi di sintesi, scomposizione e ricambio.
Il procollagene, la proteina madre del collagene, è sintetizzata e secreta dai fibroblasti. Le molecole di procollagene sono costituite da 3 filamenti proteici disposti a tripla elica. Il procollagene lascia la cellula con estensioni protettive alle estremità per prevenire la formazione prematura di collagene. La scissione dei prolungamenti tramite enzimi porta alla formazione di collagene attivo che si allinea con altre molecole di collagene.
Punto chiave: La misurazione di questi enzimi (cioè dei marcatori) fornisce un’indicazione del metabolismo del collagene. Un aumento o una diminuzione dei livelli di alcuni enzimi indica la sintesi netta di collagene (ad esempio, di tipo I), mentre una diminuzione o un aumento dei livelli di alcuni enzimi indica la degradazione netta di collagene (ad esempio, di tipo I).
Una microfibrilla è un’unità di filamenti di collagene disposti in parallelo. Sono le subunità delle fibre di collagene. Le fibre sono disposte in fasci. I legami incrociati tra le molecole di collagene adiacenti nei fasci di collagene sono legami chimici forti che garantiscono l’integrità e un robusto reticolo di tessuti connettivi che supportano lo scheletro nella locomozione.
Il collagene di tipo I è particolarmente importante per le modifiche del contenuto minerale osseo e della densità minerale ossea (BMC/BMD).
Il collagene di tipo III è particolarmente importante per i cambiamenti nella ECM, nei tendini, nei legamenti e nei tessuti connettivi:
Trasmissione della forza dal sarcomero all’osso (aumento della forza)
Recupero da lesioni muscoloscheletriche, in particolare quelle che coinvolgono la matrice di collagene.
Prevenzione di lesioni muscoloscheletriche da uso eccessivo o acute (aumentando il rapporto forza-fatica).
In generale gli AAS influenzano il metabolismo del collagene. AAS sovrafisiologici:
↑ PIIINP sierica [in maniera dose-dipendente]
↑ urina HP:LP – Dosi elevate di AAS aumentano il metabolismo del collagene dei tessuti molli, senza variazioni nel riassorbimento osseo. [1].
Cessazione (“cycling-off”) d’uso degli AAS:
↓ ICTP sierico [in funzione del tempo]. [1].
Marker del metabolismo del collagene:
Tendine ( marker metabolici del collagene)
Il propeptide N-terminale del procollagene di tipo III (PIIINP) è un marker della biosintesi delle fibrille interstiziali nei tessuti molli. Il PIIINP è stimolato dall’allenamento pliometrico nei tendini e dagli AAS in generale, dove la rigidità va a vantaggio della velocità, ma l’aumento della forza muscolare deve compensare l’aumento del rischio di strappi muscolari dovuto all’eccessiva rigidità dei tendini.
-Aumenta in modo dose-dipendente con gli AAS a concentrazione sovrafisiologica.
Ossa e tendini (“marker metabolici del collagene”)
Procollagene di tipo I C-terminale propeptide (PICP): rilasciato in circolo dagli osteoblasti proliferanti durante la biosintesi del collagene, è quindi in gran parte un biomarcatore della formazione di collagene osseo, sebbene vi sia anche un certo contributo da parte del collagene di tipo I nei tessuti molli. [2].
Telopeptide reticolato del collagene di tipo I (ICTP): rilasciato in circolo durante la fase osteoclastica della modellazione e del rimodellamento osseo (disgregazione dell’osso/collagene). [2].
-Diminuisce con la sospensione di AAS sovrafisiologici (“cycling-off”) in modo dipendente dal tempo.
Dpyr: marcatore urinario della degradazione del collagene di tipo I. [2].
Sia l’ICTP che il Dpyr costituiscono legami incrociati piridinolinici formati nel collagene maturo di tipo I e le loro concentrazioni nel siero e nelle urine riflettono il riassorbimento osseo. [2].
Ossa (“marker metabolici del collagene”)
Fosfatasi alcalina ossea (ALP): proteina presente nelle cellule ossee della placca di crescita epifisaria e negli osteoblasti maturi. È irrilevante per le dimensioni o la forza muscolare. [2].
HP (PYD): idrossilisilpiridinolina; riflette il turnover dei collageni di tipo I (osso), II, III e IX; presente in tendini, cartilagini, ossa, pareti dei vasi e dentina. [2].
LP (DPD): lisilpiridinolina; riflette il turnover del collagene di tipo I (osso); presente nell’osso e nella dentina.
Esiste un declino associato all’età di HP (↓), LP (↓) e del rapporto HP/LP (↓).
HP:LP (urina) esclude il metabolismo cutaneo (utile). [2].
Idrossiprolina
L’HP e l’LP urinari sono marcatori potenzialmente più utili del catabolismo delle fibre di collagene dei tessuti scheletrici rispetto all’idrossiprolina urinaria. Quest’ultima ha un profilo di specificità inferiore poiché si trova in tutti i tipi di collagene di tutti i tessuti connettivi (compresa la pelle). Inoltre, può anche essere rilasciata dalle molecole di collagene prima della loro incorporazione nelle fibrille e una grande percentuale di idrossiprolina viene metabolizzata nel fegato, eludendo così l’analisi quantitativa del riassorbimento del collagene maturo mediante misurazioni urinarie. [3].
Un rapporto HP:LP più basso può riflettere una maggiore proporzione relativa di riassorbimento osseo rispetto al turnover del collagene e della cartilagine, poiché il collagene osseo è la fonte primaria di LP, mentre l’HP riflette i tessuti molli in generale (eccetto la pelle)… suggerisce che anche il riassorbimento del collagene di tipo III (più HP rispetto al tipo I) diminuisce con l’età. [3].
-Aumenta con gli AAS sovrafisiologici.
Il picco di massa ossea e di HP:LP si verifica all’età di 27 anni. [3].
Ossa:
Cellule ossee
Osteoprogenitori: Cellule osteogeniche che si sviluppano in osteoblasti.
Osteoblasti: Formano l’osso; producono ECM ossea e mineralizzano l’osso (mononucleati).
Osteociti: Cellule ossee mature; secernono enzimi per mantenere l’osso.
La formazione ossea osteoblastica è associata alla deposizione di collagene di tipo I, seguita dalla mineralizzazione e dalla maturazione, durante le quali si formano legami incrociati stabili tra le fibrille di collagene. [4].
Osteocalcina (siero): Riflette la formazione ossea (così come il procollagene di tipo I [siero]).
Deossipiridinolina (urina): Riflette il riassorbimento osseo. [4].
Tendini e legamenti:
Il tendine è un tessuto connettivo che collega l’osso al muscolo, mentre il legamento è un tessuto connettivo che collega l’osso all’osso. In entrambi, circa ¾ del peso secco è costituito da collagene: la maggior parte è di tipo I: 60% (tendine) e fino all’85% (legamento).
La struttura relativamente (quasi totalmente) avascolare e collagena di legamenti e tendini limita il loro potenziale rigenerativo, con conseguenti complicazioni mediche sostanziali, che spesso rendono necessario un intervento chirurgico dopo una lesione traumatica.
I tendini e i legamenti maturi contengono relativamente poche cellule. Il numero ridotto di cellule metabolicamente attive comporta un fabbisogno di ↓O₂ e di nutrienti. I legamenti contengono fibre di elastina e collagene.
Le proprietà meccaniche di tendini e legamenti sono funzione di:
Densità delle fibre di collagene
Diametro
Orientamento e
Reticolazione -Legami incrociati enzimatici, formati dall’ossido di lisile (LOX) -Legami incrociati non enzimatici attraverso gli AGE (advanced glycation end-products), formati da una reazione di Maillard senza enzimi specifici tra uno zucchero e un amminoacido. -Entrambi i legami incrociati aumentano la rigidità di tendini e legamenti. [5].
Adattamenti dell’allenamento
Il metabolismo del tendine è molto più lento di quello del muscolo a causa della sua ridotta vascolarizzazione e circolazione, e l’aumento del flusso sanguigno al muscolo scheletrico attraverso l’esercizio fisico non è parallelo alla stessa perfusione del flusso nel tendine. [6].
L’ipertrofia muscolare è correlata a un aumento del numero e delle dimensioni dei fibroblasti, con conseguente aumento dell’apporto totale di collagene. L’attivazione dei fibroblasti e la successiva crescita del tessuto connettivo sono i prerequisiti dell’ipertrofia [7], in modo che il contenuto di collagene sia mantenuto in proporzione alla massa muscolare.
La rigidità del tendine si riferisce alla trasmissione della forza per unità di sforzo, o allungamento del tendine. L’aumento della rigidità del legamento è una buona cosa (✓), poiché la rigidità è utile per mantenere la stabilità dell’articolazione e il rischio di lesioni. Al contrario, poiché il tendine collega l’osso rigido al muscolo cedevole, un tendine più rigido non è sempre vantaggioso:
In termini di prestazioni: ↑La rigidità (tendine) trasmette più velocemente le forze muscolari all’osso, con conseguente ↑ prestazione; tuttavia, questo interesse deve essere bilanciato dal potenziale di concentrazione delle deformazioni all’interno del muscolo:
La maggiore deformazione (“stiramento”) prodotta in un determinato movimento si concentra nel muscolo collegato a un tendine rigido ⇒ contrazione isometrica piuttosto che nel tendine che si allunga in modo flessibile mentre il muscolo si contrae.
Un tendine rigido non si allunga; piuttosto, è costretto ad allungarsi durante la contrazione (eccentrica), pertanto un muscolo collegato a un tendine rigido subisce un carico eccentrico maggiore per un determinato movimento e presenta un rischio maggiore di lesioni. [5].
Effetti degli androgeni sul tendine:
Tabella che descrive le prove degli effetti meccanici, strutturali e biologici degli AAS sul tendine. [9].
Il risultato di questa tabella mostra che gli effetti degli AAS sul tendine, e soprattutto sull’unità muscolo-tendinea, non si prestano a una descrizione univoca o a una conclusione univoca, ma presentano sfumature dovute all’eterogeneità dei dati. Gli AAS migliorano e ostacolano diversi elementi della struttura e della funzione del tendine.
In sintesi, le conclusioni pratiche che si possono trarre dalla letteratura sugli effetti degli AAS sul tendine sono le seguenti:
Gli effetti biomeccanici potenzialmente deleteri degli AAS possono essere transitori.
Gli AAS somministrati per via sistemica e locale possono avere effetti simili sui tendini.
Esistono notevoli lacune nelle conoscenze relative a: -Lesioni/patologia del tendine -Risposta alla dose -Risposta al farmaco (la maggior parte degli studi utilizza il nandrolone; alcuni il metandienone) -Cosomministrazione -Tempistica e -Popolazione di risposta (differenze legate al sesso e all’età).
Effetti specifici sui tessuti: -È probabile che gli AAS influenzino in modo diverso il metabolismo dei tendini, dei muscoli e della fibrocartilagine (sintesi/degradazione netta), ma finora non sono stati condotti studi per caratterizzare queste differenze. [9].
Articolazioni:
Le articolazioni del corpo sono i punti in cui le ossa si incontrano (articolazione) e che consentono la locomozione umana (i muscoli tirano le ossa sulle articolazioni per muovere il corpo). Le articolazioni sono costituite da ossa, tendini (che collegano le ossa ai muscoli) e legamenti (che collegano le ossa alle ossa). È possibile classificare le articolazioni secondo vari schemi (ad esempio, funzionale, strutturale). Questo articolo si concentrerà sulle articolazioni sinoviali dell’organismo, che comprendono l’anca, il ginocchio, la spalla e il gomito. Oltre a essere composte da ossa, tendini e legamenti, queste articolazioni contengono cellule sinoviali, un tipo di cellula che contiene una membrana di rivestimento che produce il liquido sinoviale. Il liquido sinoviale lubrifica e nutre l’articolazione, riduce l’attrito e fornisce ammortizzazione tra le superfici articolari. È importante notare che questa lubrificazione e questo nutrimento dipendono dalla produzione di proteine e fattori di crescita.
Gli effetti degli AAS sulle articolazioni non sono descritti in modo esaustivo, ma esistono studi sui singoli composti che verranno esaminati in seguito per approfondire i particolari AAS e i loro effetti sulle articolazioni. I potenziali meccanismi con cui gli AAS influenzano le articolazioni riguardano in generale gli effetti sul metabolismo, sul ricambio, sulla scomposizione e sulla sintesi del collagene, gli effetti sul C1-INH e la funzione delle articolazioni sinoviali.
C1-INH:
Inibitore della C1-esterasi; SERPING1
Struttura molecolare del C1-INH
Il C1-INH è un inibitore della proteasi multi-serina che controlla diverse vie catalitiche, tra cui l’attivazione dei componenti classici.
Gli androgeni attenuati – AAS che possiedono una potenza androgena relativamente ridotta – qui discussi, l’Oxandrolone e lo Stanozololo, entrambi 17AA, inducono la produzione intrinseca di C1-INH e il ↑catabolismo (cioè la scomposizione) della bradichinina.
La bradichinina, attraverso la sua azione sul B₂R, media la vasodilatazione e aumenta la permeabilità con conseguente angioedema. L’angioedema ereditario (HAE), per estensione, deriva da ↓C1-INH. L’HAE è una condizione che può essere trattata con androgeni attenuati.
Quando il C1-INH è ridotto, si verifica una permeabilità vascolare (sottocutanea e sottomucosa) (“angioedema”) dovuta alla ↑bradichinina (che il C1-INH attenua) [a causa degli effetti sul sistema di contatto classico e sull’attivazione del complemento che esulano dallo scopo di questo articolo]. [10].
Gli androgeni regolano l’espressione genica della C1-INH e l’aminopeptidasi P plasmatica (che catabolizza le chinine) (55, 56). [10].
Gli androgeni attenuati (17AA) aumentano in modo più potente la produzione epatica di C1-INH [23] per azione diretta a livello epatico piuttosto che per azione dell’AR di per sé ⇒ ↑C1-INH e C4 (a causa dell’inversione dei livelli secondariamente depressi di C4). [11].
I complessi C1/C1-INH si formano quando C1-INH si combina e rimuove C1r e C1s dal C1 attivato e questi complessi – rappresentativi dell’attivazione della via del complemento classica – sono associati a condizioni artritiche e reumatologiche. [12]. Ciò può contribuire alla reputazione di Stanozolol, in particolare, di causare dolori articolari.
Punto chiave: L'”angioedema acquisito senza focolai” può essere causato da farmaci, di cui gli ACE-inibitori (nello 0,1-2,2% dei consumatori, più numerosi negli africani) sono i responsabili più comuni. L’ACE è necessario per la degradazione della bradichinina e la sua inibizione può provocare un accumulo di bradichinina che causa l’angioedema. Gli androgeni possono in qualche misura sopprimere l’angioedema grazie ai loro effetti sulla C1-INH. In un articolo di prossima pubblicazione di questo autore si discutono i rischi della diffusione di farmaci antipertensivi, soprattutto ACE inibitori e ARB, per i bodybuilder sani che fanno uso di AAS in assenza di ipertensione cronica, e si affronta la questione particolare (e apparentemente controversa) della diminuzione (blunting) dell’ipertrofia indotta dagli ACE inibitori.
Molecole:
Nandrolone
Scheletro carbossilico del Nandrolone
Meccanismi putativi che migliorano i sintomi del dolore articolare:
Aumento della sintesi e del deposito di collagene nei tendini e nei legamenti.
Ritenzione del liquido sinoviale articolare.
Entrambi i meccanismi (aumento della produzione di collagene, ad esempio nei tendini flessori) e la ritenzione di liquido articolare sinoviale possono essere influenzati positivamente dal sistema renina-angiotensina (RAS). Il RAS regola l’equilibrio idrico ed elettrolitico, la crescita delle cellule del tessuto connettivo e il metabolismo del tessuto connettivo lasso e denso e dei siti di riparazione dei tessuti. [13]. Dal punto di vista patologico, l’attivazione del RAS aumenta la vascocostrizione, l’ipertrofia cardiaca e la fibrosi (con conseguente infarto del miocardio e fibrosi del fegato). [13]. Pertanto, è importante considerare la dualità del potenziale aumento dell’articolazione (cioè del tendine estensore del ginocchio) con l’uso di nandrolone: si può avere un beneficio transitorio nel rimodellamento del tendine (cioè del tendine estensore), ma attraverso questo stesso meccanismo, si possono accumulare disadattamenti fibrotici o cardiaci.
Quindi, per quanto riguarda i potenziali benefici transitori: L’enzima di conversione dell’angiotensina I (ACE) è un marcatore positivamente correlato all’attività dell’mRNA del collagene di tipo I e può riflettere il rimodellamento della matrice extracellulare (ECM) in cui la sintesi di collagene supera la degradazione.
Il Nandrolone aumenta l’attività ACE e incrementa la deposizione di collagene di tipo I nella matrice. In un modello di allenamento (ad esempio, pliometrico), Nandrolone + allenamento per i salti >> allenamento per i salti > sedentario per quanto riguarda l’attività ACE nel tendine (ad esempio, estensore del bicipite femorale), suggerendo una potenziale sinergia tra allenamento e Nandrolone a questo proposito. Si consideri, tuttavia, che questa stessa via è implicata nel rimodellamento del tessuto cardiaco e nell’azione patologica. Inoltre, l’aumento della rigidità tendinea rappresenta un rischio se non si aumenta la forza per ridurre la probabilità di lesioni gravi dovute a uno sforzo concentrato. Pertanto, le considerazioni sull’allenamento devono essere pianificate in modo rigoroso, soprattutto se si pratica l’allenamento pliometrico. [13].
Oxandrolone
Scheletro carbossilico del Oxandrolone
L’aumento significativamente maggiore della velocità di crescita in altezza ottenuto con il trattamento con GH più oxandrolone rispetto al solo GH si è riflesso in differenze simili nella risposta precoce dell’ALP ossea e del PICP, entrambi associati alla formazione dell’osso, ma non nel PIIINP, un marcatore del turnover dei tessuti molli, o nell’ICTP, un marcatore della degradazione del collagene osseo. Ciò suggerisce che l’oxandrolone può, direttamente o indirettamente, influenzare la proliferazione degli osteoblasti e la proliferazione e maturazione dei condrociti, con un effetto additivo rispetto a quello del solo GH. [2].
La reputazione del nandrolone di migliorare la funzione articolare durante le fasi di allenamento con carichi pesanti è rafforzata da questi risultati, secondo cui, aumentando l’attività dell’ACE e influenzando il RAS, serve ad aumentare la sintesi netta di collagene e l’equilibrio dei fluidi nelle articolazioni.
I risultati di Crofton et al. suggeriscono che GH+oxandrolone > GH+test > GH > placebo nel ΔPIINP, ma le differenze significative tra i gruppi potrebbero non essere misurabili a causa delle ridotte dimensioni del campione (un potenziale errore di tipo 2). [2].
La somministrazione di oxandrolone fino a 24 mesi a pazienti pediatrici gravemente ustionati ha migliorato significativamente il contenuto minerale osseo dell’intero corpo (WB BMC), il contenuto minerale osseo della colonna lombare (LS BMC), la densità minerale ossea della colonna lombare (LS BMD) e la velocità in altezza. [14].
Una grave ustione induce una risposta ipermetabolica e ipercatabolica caratterizzata da un aumento del lavoro cardiaco, del dispendio energetico a riposo e della degradazione delle proteine muscolari (1-6). Questa risposta compensatoria è accompagnata da un’elevata produzione epatica di glucosio e da insulino-resistenza (4, 5, 7-10). I pazienti in genere subiscono una perdita di massa magra e nei bambini la crescita è ostacolata. Nel tempo si verifica una significativa riduzione del contenuto minerale osseo (BMC), della densità minerale ossea (BMD) e del tessuto adiposo. [14].
Gli effetti sulla BMC/BMD sono diventati significativi in questi pazienti solo dopo più di un anno di trattamento continuo. [14]. Nei pazienti pediatrici è stata riscontrata una sinergia in questo effetto modulato dalla fase di maturazione della crescita, che potrebbe avere ramificazioni per l’uso in età adulta di rhGH e androgeni aromatizzanti, associati all’impennata puberale. L’ipotesi che si potrebbe trarre è che l’uso a lungo termine di androgeni aromatizzanti (ad esempio, T, nandrolone) in combinazione con dosi elevate di rhGH possa imitare alcuni aspetti della fase di maturazione della crescita nei bambini, potenzialmente aumentando la BMC e la BMD in modo sinergico in combinazione con l’oxandrolone se usato per lunghi periodi.
I particolari effetti dell’oxandrolone sul contenuto e sulla densità minerale ossea nei bambini in crescita suggeriscono che sia particolarmente utile nella fase iniziale (cioè i primi sei mesi) dell’uso continuo di RhGH, quando il turnover osseo è aumentato.
Stanozololo
Scheletro carbossilico dello Stanozololo
Lo Stanozololo, popolarmente associato al dolore articolare (“articolazioni doloranti e secche”), agisce sui fibroblasti sinoviali, precursori delle cellule che compongono le articolazioni sinoviali (ad esempio, anca, ginocchia, spalle), inibendo la sintesi del DNA. [10]. Mentre lo Stanozololo è considerato particolarmente potente nello stimolare l’attività della procollagenasi nel cuoio capelluto, un effetto che è associato all’aumento della secrezione di TGF-β1 [15] – stimolando così l’attività procollagene, ma inducendo perversamente, con questo stesso meccanismo, l’alopecia androgenica nel cuoio capelluto – l’effetto sulla cellula sinoviale è qualitativamente diverso.
L’inibizione della sintesi del DNA da parte dello Stanozololo nella cellula sinoviale, insieme ai suoi effetti sul C1-INH, fornisce diverse modalità esplicative per il fatto che provoca dolore nelle articolazioni sinoviali.
Alla fine del 1980 ricercatori britannici hanno scoperto che le cellule della pelle producono più collagene quando viene usato lo Stanozololo, ma che le cellule delle articolazioni non lo fanno [https://www.ncbi.nlm.nih.gov/pubmed/2556901].
Testosterone
Scheletro carbossilico del Testosterone
Il testosterone (250 mg a settimana) ha causato modesti aumenti dei biomarcatori del metabolismo del collagene, aumentando il PINP del 28%, l’ICTP del 22% e il PIINP del 70%. [16]. Il testosterone ha aumentato il tipo I (osso; PINP, ICTP) e il tipo III (tendini e legamenti, tessuti molli; PIINP) e ha potenziato in particolare l’effetto dell’rhGH sui marcatori di tipo III (PIINP), suggerendo un forte effetto modulante del testosterone sul collagene di tipo III che comprende tendini, legamenti e probabilmente fasce in risposta al GH. [16].
rhGH
Struttura peptidica del rhGH.
La somministrazione di rhGH nel tendine o nel legamento determina un marcato aumento del metabolismo del collagene con conseguente deposito netto. [17]. L’RhGH aumenta l’attività dell’osteocalcina e del procollagene di tipo I nel siero, riflettendo un aumento della BMD/BMC. Nel muscolo scheletrico e nel tendine la matrice extracellulare (ECM) conferisce importanti proprietà di trazione ed è di fondamentale importanza per la rigenerazione dei tessuti dopo una lesione. [10]. La somministrazione di rhGH promuove la sintesi di collagene ECM nel tessuto muscolo-tendineo di giovani adulti sani.
L’RhGH ha aumentato l’mRNA del collagene I (osso) di 2,3 volte e del collagene III (ECM, tendini, legamenti) di 2,5 volte. [10]. È stata osservata una tendenza ad un aumento di 5,8 volte della sintesi proteica del collagene muscolare. [10]. Il dosaggio utilizzato è stato di 33,3 µg * kg-¹ * giorno-¹ nei primi sette giorni e di 50 µg * kg-¹ * giorno-¹ in giovani uomini sani. [10]. Ciò equivale a una dose giornaliera di 3 – 4,5 UI di rhGH per un uomo di 90 kg.
Nella popolazione GHD si osserva una variazione bifasica (a due punte) della BMD in risposta alla somministrazione di rhGH, con una diminuzione iniziale a circa sei mesi dall’inizio della terapia, seguita da un successivo aumento dopo almeno un anno di trattamento. [18]. L’ipotesi prevalente è che il GH stimoli sia la formazione che il riassorbimento osseo, con conseguente aumento del turnover osseo. [18]. Questo effetto è evidente almeno nei primi sei mesi di somministrazione di rhGH, con conseguente diminuzione della BMD e del contenuto minerale osseo. [18]. Non è stata stabilita una relazione dose-risposta a causa dell’ampia variazione dei dosaggi utilizzati. [18]. Sembra che l’osso trabecolare (colonna vertebrale lombare) abbia una diversa sensibilità al GH rispetto all’osso corticale (collo del femore). [18].
L’RhGH si combina almeno in alcuni aspetti con gli androgeni (ad esempio, oxandrolone, testosterone) e in altri in modo sinergico per quanto riguarda il metabolismo dei tessuti molli. L’RhGH e il testosterone, se usati in combinazione, stimolano particolarmente la PIINP, suggerendo un effetto sinergico su tendini, legamenti e probabilmente fasce.
Conclusioni
Gli AAS hanno effetti di classe e specifici sulle articolazioni e sui tessuti connettivi che le compongono. In generale, gli AAS migliorano il metabolismo dei tessuti molli senza alcun effetto sul riassorbimento osseo, ma è importante considerare i vari effetti positivi e negativi degli AAS nei confronti dei tendini.
Gli androgeni aromatizzanti, come il testosterone, hanno effetti sinergici (più che additivi) in combinazione con il GH su alcuni aspetti del metabolismo del collagene, in particolare per quanto riguarda gli aspetti legati al metabolismo dei tessuti molli (ad esempio, la fascia), ed effetti additivi per altri aspetti.
I 17AA non aromatizzabili Oxandrolone (un 5α-androstan-3-one) e Stanozololo (un AAS il cui anello A ha una giunzione anulare pirazolica) condividono alcuni effetti di classe, ad esempio la stimolazione della produzione intrinseca di C1-INH che è associata a condizioni reumatologiche e artritiche, ma lo Stanozololo si distingue per la sua dimostrabile inibizione dose-dipendente della sintesi di DNA nelle cellule sinoviali che comprendono le principali articolazioni del corpo, modulando la disgregazione del tessuto connettivo. È stato dimostrato che l’Oxandrolone aumenta in modo additivo la densità minerale ossea (BMD/BMC) e può essere particolarmente utile nei primi sei mesi di inizio di un ciclo di androgeni aromatizzanti e rhGH per migliorare la conservazione della massa ossea.
Punto chiave: Un approccio sfumato al processo decisionale sul carico dell’allenamento (ad esempio, progressione, intensità, modalità) e sugli effetti dei farmaci (ad esempio, AAS, rhGH) in relazione alle articolazioni, ai tendini, ai legamenti e alle ossa, è reso possibile da una lettura attenta di questo articolo. Per i professionisti, le aree particolari da leggere con attenzione riguardano la rigidità dei tendini e le implicazioni per coloro che si impegnano in allenamenti pliometrici o anche con carichi leggeri; i pro e i contro degli AAS sugli aspetti biologici, strutturali e meccanici dei tendini; e la scarsità di dati solidi relativi agli effetti sui tessuti molli da parte di diversi AAS.
[1] Pärssinen, M., Karila, Kovanen, and Seppälä. (2000). The Effect of Supraphysiological Doses of Anabolic Androgenic Steroids on Collagen Metabolism. International Journal of Sports Medicine, 21(6), 406–411. doi:10.1055/s-2000-3834
[2] Crofton, P. M., Stirling, H. F., Schönau, E., and Kelnar, C. J. H. (1996). Bone alkaline phosphatase and collagen markers as early predictors of height velocity response to growth-promoting treatments in short normal children. Clinical Endocrinology, 44(4), 385–394. doi:10.1046/j.1365-2265.1996.706cn527.x
[3] Açil, Y., Brinckmann, J., Notbohm, H., Müller, P. K., and Bätge, B. (1996). Changes with age in the urinary excretion of hydroxylysylpyridinoline (HP) and lysylpyridinoline (LP). Scandinavian Journal of Clinical and Laboratory Investigation, 56(3), 275–283. doi:10.3109/00365519609088617
[4] Aerssens, J., Van Audekercke, R., Geusens, P., Schot, L. P. C., Osman, A. A.-H., and Dequeker, J. (1993). Mechanical properties, bone mineral content, and bone composition (collagen, osteocalcin, IGF-I) of the rat femur: Influence of ovariectomy and nandrolone decanoate (anabolic steroid) treatment. Calcified Tissue International, 53(4), 269–277. doi:10.1007/bf01320913
[5] Chidi-Ogbolu N, Baar K. Effect of Estrogen on Musculoskeletal Performance and Injury Risk. Front Physiol. 2019;9:1834. Published 2019 Jan 15. doi:10.3389/fphys.2018.01834
[6] Kjaer, M.J. (2008). Role of extracellular matrix in adaptation of tendon and skeletal muscle to mechanical loading. Physiol. Rev. 84:649-698.
[7] Joblecki, C.K., Heuser, J.E., Kaufman, S. (1973). Autoradiographic localization of new RNA synthesis in hypertrophying skeletal muscles. Journal of Cellular Biology. 57:743-759.
[8] Spadari, A., Romagnoli, N., Predieri, P. G., Borghetti, P., Cantoni, A. M., and Corradi, A. (2013). Effects of intraarticular treatment with stanozolol on synovial membrane and cartilage in an ovine model of osteoarthritis. Research in Veterinary Science, 94(3), 379–387. doi:10.1016/j.rvsc.2012.11.020
[9] Jones, I. A., Togashi, R., Rick Hatch, G. F., Weber, A. E., and Vangsness JR, C. T. (2018). Anabolic Steroids and Tendons: A Review of their Mechanical, Structural and Biologic Effects. Journal of Orthopaedic Research®. doi:10.1002/jor.24116
[10] Ellis AJ, Cawston TE, Mackie EJ. The differential effects of stanozolol on human skin and synovial fibroblasts in vitro: DNA synthesis and receptor binding. Agents Actions. 1994 Mar;41(1-2):37-43. doi: 10.1007/BF01986391. PMID: 8079819.
[11] Wright JK, Smith AJ, Cawston TE, Hazleman BL. The effect of the anabolic steroid, stanozolol, on the production of procollagenase by human synovial and skin fibroblasts in vitro. Agents Actions. 1989 Nov;28(3-4):279-82. doi: 10.1007/BF01967415. PMID: 2556901.
[12] Oleesky DA, Daniels RH, Williams BD, Amos N, Mor gan BP. Terminal complement complexes and C1/C1 inhibitor complexes in rheumatoid arthritis and other arthritic conditions. Clin Exp Immunol. 1991;84(2):250-255.
[13] Marqueti, R. de C., Hashimoto, N. Y., Durigan, J. L. Q., Batista e Silva, L. L., Almeida, J. A. de, Silva, M. da G. da, … Araújo, H. S. S. de. (2015). Nandrolone increases angiotensin-I converting enzyme activity in rats tendons. Revista Brasileira de Medicina Do Esporte, 21(3), 173–177. doi:10.1590/1517-869220152103143667
[14] Reeves, P. T., Herndon, D. N., Tanksley, J.D., Jennings, K., Klein, G. L., Mlcak, R. P., … Finnerty, C. C. (2016). FIVE-YEAR OUTCOMES AFTER LONG-TERM OXANDROLONE ADMINISTRATION IN SEVERELY BURNED CHILDREN. Shock, 45(4), 367–374. doi:10.1097/shk.0000000000000517
[15] Falanga, V., Greenberg, A. S., Zhou, L., Ochoa, S. M., Roberts, A. B., Falabella, A., and Yamaguchi, Y. (1998). Stimulation of Collagen Synthesis by the Anabolic Steroid Stanozolol. Journal of Investigative Dermatology, 111(6), 1193–1197. doi:10.1046/j.1523-1747.1998.00431.x
[16] Nelson, A. E., Meinhardt, U., Hansen, J. L., Walker, I. H., Stone, G., Howe, C. J., … Ho, K. K. (2008). Pharmacodynamics of Growth Hormone Abuse Biomarkers and the Influence of Gender and Testosterone: A Randomized Double-Blind Placebo-Controlled Study in Young Recreational Athletes. The Journal of Clinical Endocrinology and Metabolism, 93(6), 2213–2222.
[17] Vestergaard, P., Jørgensen, J. O. L., Olesen, J. L., Bosnjak, E., Holm, L., Frystyk, J., … Hansen, M. (2012). Local administration of growth hormone stimulates tendon collagen synthesis in elderly men. Journal of Applied Physiology, 113(9), 1432–1438. doi:10.1152/japplphysiol.00816.2012
[18] Barake, M., Klibanski, A., and Tritos, N. A. (2014). Effects of Recombinant Human Growth Hormone Therapy on Bone Mineral Density in Adults With Growth Hormone Deficiency: A Meta-Analysis. The Journal of Clinical Endocrinology and Metabolism, 99(3), 852–860. doi:10.1210/jc.2013-3921
Nelle precedenti parti di questo lungo viaggio alla scoperta e comprensione degli aminoacidi [Parte 1°, 2°, 3°, 4° e 5°] abbiamo capito chiaramente cosa sono, quali sono le loro funzioni biologiche ed abbiamo analizzato quegli AA che sono maggiormente utilizzati in campo sportivo. In questa parte conclusiva il viaggio raggiungerà il culmine con una trattazione approfondita degli Aminoacidi Essenziali/EAA.
Introduzione agli EAA:
Gli aminoacidi “essenziali” (EAA) della dieta – istidina, isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptofano e valina – sono detti “essenziali” perché non possono essere prodotti endogenamente e quindi devono essere consumati per la sopravvivenza umana. Inoltre, l’arginina è considerata un aminoacido “condizionatamente” essenziale, il che significa che in alcune circostanze la produzione endogena di arginina non riesce a soddisfare le esigenze fisiologiche. La necessità di consumare tutti gli EAA è stata ben stabilita negli ultimi 100 anni [1], ed esiste un fabbisogno giornaliero accettato per ciascun EAA come parte del normale apporto dietetico [2]. Il fabbisogno giornaliero si basa sulla quantità minima di ciascun EAA che deve essere consumata per evitare sintomi clinici di carenza. Un consumo inadeguato di uno solo dei nove EAA provoca sintomi di carenza, tra cui un’alterata sintesi proteica [3]. I requisiti per il consumo giornaliero di ciascun EAA sono convenzionalmente soddisfatti come componenti dell’apporto proteico dietetico di routine. La quantità e il profilo degli EAA nelle singole proteine alimentari, insieme alla digeribilità degli EAA legati alle proteine, costituiscono la base per la valutazione quantitativa della qualità della proteina pura [4]. Le proteine che contengono una quantità abbondante di tutti gli EAA in un formato altamente digeribile sono considerate proteine di “alta qualità” [4].
Sono stati fatti vari tentativi per esprimere la “qualità” o il “valore” di vari tipi di proteine. Le misure includono il valore biologico, l’utilizzo netto delle proteine, il rapporto di efficienza proteica, il punteggio aminoacidico corretto per la digeribilità delle proteine e il concetto di proteine complete. Questi concetti sono importanti nell’industria zootecnica, perché la mancanza relativa di uno o più aminoacidi essenziali negli alimenti per animali avrebbe un effetto limitante sulla crescita e quindi sul rapporto di conversione degli alimenti. Pertanto, diversi mangimi possono essere somministrati in combinazione per aumentare l’utilizzo netto delle proteine, oppure si può aggiungere al mangime un supplemento di un singolo aminoacido (metionina, lisina, treonina o triptofano).
Mentre l’importanza di soddisfare il fabbisogno minimo di ciascun EAA attraverso il consumo di proteine alimentari di alta qualità è riconosciuta da molti decenni [5], i benefici ottenibili dal consumo di EAA in forma libera in quantità superiori al fabbisogno minimo sono stati pienamente apprezzati solo negli ultimi 25 anni. Sono disponibili prodotti a base di singoli aminoacidi liberi, come la leucina o la lisina, e composizioni di piccoli gruppi di EAA, in particolare gli aminoacidi a catena ramificata (leucina, valina e isoleucina; BCAA), ma molti studi hanno documentato che si ottengono maggiori benefici da composizioni contenenti tutti gli EAA. L’integrazione giornaliera con composizioni di tutti gli EAA in forma libera ha dimostrato di essere benefica in molti modi [6]. In particolare, le composizioni di EAA in forma libera stimolano la sintesi proteica e il ricambio proteico nell’organismo, compresa la sintesi di nuove proteine muscolari. La stimolazione della sintesi proteica muscolare (MPS) da parte degli EAA può produrre un aumento della massa e della qualità muscolare, che si traduce in un miglioramento delle prestazioni fisiche e dei risultati funzionali [7].
La valutazione dei benefici degli EAA si differenzia da quella di molti altri integratori alimentari valutati in quanto esistono requisiti ben accettati per il consumo giornaliero di EAA. Inoltre, piuttosto che integratori contenenti un solo composto, come la creatina, esistono combinazioni quasi illimitate dei nove EAA che possono essere realizzate a seconda della richiesta fisiologica.
Meccanismo d’azione degli EAA
L’importanza del turnover delle proteine muscolari
Il continuo rinnovamento delle proteine muscolari degradate e danneggiate è importante per mantenere la massa e la funzione proteica del muscolo. Nello stato post-assorbitivo, la ripartizione netta delle proteine muscolari mantiene una fornitura costante di EAA plasmatici che forniscono precursori per la sintesi proteica in altri tessuti e organi. Gli EAA assunti con la dieta ripristinano la perdita netta di proteine muscolari stimolando la MPS. In condizioni normali, i tassi di MPS e di degradazione delle proteine muscolari sono uguali nel corso della giornata. Se la MPS supera il tasso di degradazione delle proteine muscolari, la massa muscolare aumenterà nel tempo, con un potenziale aumento della forza. L’accelerazione del turnover proteico muscolare (cioè la sintesi e la degradazione delle proteine aumentano in egual misura), senza un aumento della massa proteica muscolare netta, può anche giovare alla funzione muscolare, sostituendo le fibre muscolari più vecchie e danneggiate con nuove fibre altamente funzionanti [8]. Pertanto, la stimolazione del MPS/turnover è la principale base metabolica per l’aumento della forza e della funzione fisica. Sebbene anche i cambiamenti nella degradazione delle proteine giochino un ruolo nel controllo del metabolismo proteico muscolare, la stimolazione della MPS è la base principale degli effetti benefici degli integratori di EAA. Inoltre, le prime ricerche hanno dimostrato che l’effetto degli aminoacidi sul muscolo scheletrico si esplica principalmente attraverso la stimolazione della MPS, dato che la ripartizione delle proteine muscolari è rimasta invariata in uno studio acuto [9]. È importante notare che, poiché la misurazione della disgregazione delle proteine muscolari non è semplice e problematica in caso di assunzione esogena, la MPS rappresenta anche un indicatore surrogato del turnover proteico.
Controllo della sintesi proteica muscolare
Il modo più definitivo per valutare l’effetto degli integratori di EAA sulle prestazioni fisiche è quello di misurare le risposte metaboliche e funzionali nel tempo quando vengono fornite quantità e profili diversi di EAA, a condizione che tutte le altre variabili siano mantenute costanti. Tuttavia, per ottenere risultati affidabili possono essere necessari mesi di trattamento, a causa del lento tasso di turnover delle proteine muscolari e della difficoltà di controllare tutte le altre variabili (dieta, assunzione totale di EAA, attività, ecc.). Di conseguenza, l’uso della metodologia dei traccianti isotopici stabili per quantificare la risposta acuta delle MPS a una singola dose di EAA in soggetti umani è diventato il surrogato accettato per prevedere la risposta anabolica nel muscolo. Aspetti della sintesi proteica, trascrizione e traduzione, possono essere potenzialmente influenzati dal consumo di EAA: in particolare l’iniziazione e l’allungamento traslazionale (vedi Figura 1a). La trascrizione dell’RNA messaggero (mRNA) dal DNA comporta l’attivazione dei relativi geni. I cambiamenti nell’attivazione dei geni si riflettono nel numero di mRNA specifici nella cellula. L’espressione dell’mRNA è importante perché l’assemblaggio fisico di nuove proteine avviene sull’mRNA. Il complesso processo di iniziazione consiste in diverse fasi collegate tra loro e mediate da fattori di iniziazione eucariotici (eIF). Il complesso mammalian target of rapamycin 1 (mTORC1) è un regolatore chiave dell’attivazione degli eIF a valle che sono mediatori dell’iniziazione delle MPS (vedi Figura 1b). Sia la trascrizione che la traslazione del processo di sintesi proteica possono essere stimolate dagli aminoacidi e dall’esercizio fisico [13-16]. Tuttavia, sia la trascrizione dell’mRNA [17] che lo stato di fosforilazione di mTORC1 [18] sono generalmente poco correlati con i tassi di MPS, il che significa che nessuno dei due processi è probabilmente limitante per la MPS nella maggior parte delle circostanze. Il controllo traslazionale della sintesi proteica da parte della disponibilità di EAA è stato riconosciuto fin dal 1958 [19]. La traduzione comporta il collegamento successivo degli aminoacidi nell’ordine dettato dal codice dell’mRNA. Gli aminoacidi intracellulari liberi sono legati ai corrispondenti RNA di trasferimento (tRNA), formando molecole di tRNA cariche. Le molecole di tRNA cariche a loro volta trasferiscono in sequenza gli amminoacidi legati ai siti dell’mRNA che corrispondono al codice del tRNA carico. L’allungamento traslazionale può procedere fino al completamento solo se sono disponibili quantità adeguate di tutti i precursori amminoacidici richiesti. Una carenza relativa di un qualsiasi EAA lo renderà limitante e l’allungamento traslazionale verrà interrotto prima del completamento del processo. Il controllo traslazionale della MPS richiede che siano disponibili quantità adeguate di tutti gli EAA per sostenere un aumento dei tassi di MPS. Oltre a fornire i precursori necessari per la sintesi proteica, gli EAA aumentano i geni associati al rilevamento, al trasporto e alla regolazione di mTORC1 degli aminoacidi [20].
Segnalazione mTOR: regolazione di mTORC1 da parte di stimoli a monte; insulina, esercizio fisico (resistenza, endurance), glucosio e aminoacidi (Aa). L’esercizio fisico porta a un deficit energetico (aumento dell’AMP) che stimola l’AMPK, inibendo l’mTORC1, mentre il consumo di glucosio aumenta l’ATP, inibendo l’AMPK. L’insulina e l’esercizio di resistenza attivano la via PI3K, regolando positivamente mTORC1, mentre l’esercizio di resistenza attiva CaMK, promuovendo soprattutto la biogenesi mitocondriale. Gli AA stimolano principalmente mTORC1 promuovendo la fosforilazione e la de-fosforilazione delle GTPasi rag, rag A/B e rag C/D, rispettivamente. Gli AA stimolano generalmente FNIP1/FLCN, promuovendo la de-fosforilazione di rag C/D, tuttavia, alcuni EAA (leucina, istidina, valina, treonina, isoleucina, metionina) agiscono per promuovere la fosforilazione di rag A/B; ciò porta a un’upregulation di mTORC1 e della traduzione a valle, nonché del metabolismo del glucosio e dei lipidi. Le figure sono tratte da [10-12]. Abbreviazioni: Akt, proteina chinasi B; AMPK, proteina chinasi attivata dall’AMP; PI3K, fosfoinositide 3-chinasi; Ca2 +, ione calcio; CaMK, proteina chinasi calcio/calmodulina-dipendente; FNIP1, proteina folliculina-interagente 1; FLCN, folliculina (FLCN); mTORC1, complesso 1 del bersaglio mammifero della rapamicina; FOX-O, fattori di trascrizione forkhead box-O; PGC-1α, peroxisome proliferator-activated gamma coactivator-1 alpha; MuRF-1, muscle ring-finger protein-1; eEF-2K, eukaryotic elongation factor-2 kinase; eEF2, eukaryotic elongation factor-2; TSC1/2, Tuberous sclerosis proteins 1 (hamartin) +2 (tuberin); Rheb, Ras homolog enriched in brain; LRS, leucil-tRNA sintetasi; SESN2, Sestrin-2; GATOR1/2, GAP (GTPase-activating protein) activity toward Rags 1+2; SAM, s-adenosyl methionine; SAMTOR, s-adenosyl methionine sensor for mTORC1; 4E-BP1, eukaryotic translation initiation factor 4E-binding protein 1; P70-S6K, (S6K1) proteina ribosomiale S6 chinasi beta-1; SREBP, sterol regulatory element binding protein; HIF-1α, hypoxia-inducible factor-1 alpha; PDCD4, Programmed cell death protein 4; SKAR, S6K1 Aly/REF-like substrate; eIF4E/B, eukaryotic translation initiation factor 4E+B.Sintesi proteica: iniziazione, allungamento e terminazione trascrizionale che portano alla produzione di mRNA nel nucleo, poi esportato nel citosol per essere sottoposto a iniziazione, allungamento e terminazione traslazionale; produce un polipeptide che viene ripiegato in una proteina.
Importanza del protocollo utilizzato per misurare la risposta delle MPS al consumo di aminoacidi
Il metodo più comunemente utilizzato per misurare la MPS nei soggetti umani consiste nel determinare il tasso di incorporazione di un aminoacido tracciante con isotopi stabili nelle proteine muscolari nel corso del tempo, diviso per l’arricchimento del precursore [21]. Questo approccio porta al calcolo del tasso di sintesi frazionale delle proteine muscolari (FSR). Poiché la massa muscolare è relativamente costante nell’arco di diversi giorni, le variazioni del FSR sono convenzionalmente considerate un riflesso diretto della MPS [21]. Un approccio alternativo alla misurazione della MPS si basa sulla differenza artero-venosa di traccianti aminoacidici non marcati e marcati e sull’arricchimento isotopico del pool libero intracellulare [22]. Questi due metodi danno risultati comparabili per la MPS in soggetti umani [23]. Infatti, è stato dimostrato che la stimolazione acuta della MPS da parte del consumo di EAA si riflette nel bilancio proteico delle 24 ore della gamba [24]. L’evidenza supporta la traduzione delle differenze nella risposta acuta della MPS al consumo di EAA in parametri di risultato misurati per settimane o mesi. Ad esempio, l’effetto del consumo giornaliero di una formula a base di EAA in soggetti sani giovani e normali è stato confrontato con un placebo per 28 giorni di riposo completo a letto [7]. Il consumo quotidiano della formula a base di EAA per tutti i 28 giorni di riposo a letto ha migliorato la perdita di massa muscolare osservata nei soggetti che consumavano il placebo di una quantità prevista dallo studio con tracciante prima del riposo a letto [7]. L’accuratezza predittiva del metodo del tracciante in acuto in questo paradigma è particolarmente significativa perché l’attività e l’assunzione di cibo sono state completamente controllate durante i 28 giorni di intervento [7].
Il periodo di tempo in cui viene determinata la MPS è importante per interpretare il significato fisiologico dei cambiamenti acuti in risposta all’assunzione di aminoacidi. Un aumento transitorio della MPS ha meno probabilità di predire un aumento a lungo termine della massa e della funzione muscolare rispetto a una risposta che rimane al di sopra del valore di base per tre ore o più. Ad esempio, il consumo di leucina da sola può suscitare una risposta transitoria (1-2 ore) nella MPS muscolare, ma questa risposta deve essere interpretata con cautela [25]. Il consumo di una quantità sufficiente di leucina da sola può attivare mTORC1 e le molecole associate coinvolte nell’avvio del processo di sintesi proteica e si riflette in un aumento transitorio delle proteine muscolari MPS. Tuttavia, la sintesi delle proteine muscolari richiede un’adeguata disponibilità di tutti gli aminoacidi componenti, compresi i nove EAA. In assenza di apporto dietetico, gli EAA necessari per produrre proteine muscolari complete devono provenire da fonti endogene. Inizialmente, gli EAA aggiuntivi necessari per la sintesi di proteine muscolari complete possono provenire da pool di EAA liberi nel fluido intracellulare ed extracellulare. Tuttavia, il conseguente esaurimento degli EAA liberi in questi pool limiterà la sintesi proteica muscolare a causa dell’inadeguata disponibilità di precursori (EAA disponibili nei pool di aminoacidi). L’unica altra fonte potenziale degli EAA necessari per mantenere la MPS in questa circostanza è la degradazione proteica accelerata, che limiterà qualsiasi guadagno netto di proteine muscolari che ci si potrebbe aspettare in base alle variazioni acute della MPS. Pertanto, la risposta anabolica (cioè MPS – MPB) delle proteine muscolari al consumo di un singolo EAA, come la leucina, o di piccoli gruppi di EAA (BCAA) sarà limitata dalla scarsa disponibilità degli altri EAA.
Il lavoro di Fuchs e collaboratori [26] fornisce prove sull’interpretazione dell’importanza dell’intervallo di campionamento sulla risposta della FSR muscolare all’assunzione di aminoacidi. In questo studio, la FSR muscolare è stata determinata in risposta al consumo di BCAA, proteine del latte o chetoacidi a catena ramificata, corrispondenti a leucina, valina e isoleucina. La FSR delle proteine muscolari è stata stimolata nelle prime due ore dopo il consumo di tutti e tre gli integratori alimentari. Tuttavia, 2-5 ore dopo l’ingestione di ciascun integratore, la FSR delle proteine muscolari è rimasta stimolata solo dopo il consumo di proteine del latte. In altre parole, l’ingestione dei BCAA o dei chetoacidi associati non è riuscita a stimolare la FSR delle proteine muscolari rispetto al valore di base dopo il consumo [26]. Il tasso di FSR delle proteine muscolari è stato limitato nelle ore 2-5 dopo il loro consumo da una diminuzione della disponibilità degli EAA non forniti dall’integratore alimentare, come risulta dalla diminuzione della concentrazione plasmatica di fenilalanina. La diminuzione degli EAA plasmatici è stata probabilmente attenuata in qualche misura da un aumento del tasso di degradazione delle proteine muscolari, limitando così l’effetto anabolico netto della stimolazione della FSR delle proteine muscolari. Al contrario, la disponibilità di fenilalanina (come riflesso degli EAA) era elevata 2-5 ore dopo il consumo di proteine del latte a causa della continua digestione e assorbimento di tutti gli EAA, nonché della disponibilità di aminoacidi non essenziali di supporto. Gli autori hanno concluso che “questi dati suggeriscono che, oltre all’aumento postprandiale delle concentrazioni plasmatiche di BCAA, è necessario fornire altri aminoacidi (essenziali) per consentire un aumento postprandiale più prolungato del tasso di sintesi proteica muscolare” [26]. È quindi ragionevole basarsi principalmente su dati provenienti da studi in cui la MPS è stata determinata in un intervallo di 3 ore o più dopo il consumo di EAA per aspettarsi una traduzione dei risultati in risultati funzionali.
Sicurezza
Il consumo di EAA non è stato segnalato come causa di reazioni avverse. I soggetti affetti da rare malattie genetiche che comportano un’alterazione della capacità di metabolizzare alcuni EAA, come la malattia delle urine a sciroppo d’acero (incapacità di metabolizzare i BCAA), potrebbero avere una risposta avversa agli integratori contenenti tutti gli EAA. Tuttavia, gli errori innati del metabolismo che influenzano il metabolismo di un EAA sono evidenti in età precoce e l’adattamento alla dieta è necessario per la salute ed eventualmente per la sopravvivenza. È quindi improbabile che un adulto con un errore innato del metabolismo che limita il consumo sicuro di EAA non sia consapevole di tale condizione. È anche possibile che un individuo con una malattia renale possa reagire male all’integrazione di EAA, poiché una dieta a basso contenuto proteico è spesso raccomandata nelle malattie renali a causa dell’accumulo di urea e ammoniaca nel sangue. Tuttavia, un’integrazione a base di EAA generalmente non contribuisce ad aumentare la produzione di urea o ammoniaca a causa del maggiore riutilizzo di aminoacidi non essenziali per la sintesi proteica. Tuttavia, non sono disponibili dati sufficienti su individui con funzionalità renale compromessa per determinare la sicurezza degli integratori alimentari a base di EAA.
Sono disponibili pochi dati su cui basare il limite massimo di sicurezza del consumo dei singoli EAA. La Tabella 1 elenca i livelli di consumo di ciascun EAA che si sono dimostrati sicuri. I limiti massimi di sicurezza riportati nella Tabella 1 sono espressi come quantità di ciascun EAA consumata al di sopra dell’assunzione abituale. Pertanto, se si considerano le quantità sicure di ciascun EAA, questi dati indicano che più di 100 g di EAA supplementari possono essere consumati in modo sicuro al giorno in un adulto americano che già consuma l’apporto alimentare medio abituale di circa 40 grammi al giorno. Il dosaggio ragionevole di un integratore di EAA non supera i 15 g, il che significa che anche tre dosi massime al giorno sono in linea con il normale consumo giornaliero di EAA attraverso le fonti alimentari di proteine. I seguenti dati sugli effetti dell’integrazione di EAA sono stati ricavati in popolazioni con un’adeguata assunzione di proteine alimentari, a meno che non sia indicato diversamente.
Fabbisogno alimentare di aminoacidi essenziali e limite massimo di consumo sicuro per gli adulti:
Riferimenti [2,4], basati sui DRI; 2Al di sopra dell’assunzione abituale.
Punti chiave:
Il turnover proteico assicura il continuo rinnovo delle proteine muscolari degradate e danneggiate ed è importante per mantenere la massa e la funzione proteica muscolare.
Il surrogato accettato per la misurazione del turnover proteico è la determinazione della sintesi proteica muscolare con la metodologia dei traccianti isotopici stabili. Sebbene la degradazione delle proteine sia importante in questo processo, la risposta acuta principale dell’assunzione di EAA sul muscolo scheletrico è la stimolazione della sintesi proteica.
Il limite massimo di sicurezza dell’assunzione giornaliera di EAA consente un’integrazione sostanziale.
Consenso dei risultati della ricerca
EAA e sintesi proteica muscolare a riposo
La MPS è stimolata dal consumo di composizioni di EAA [35] e inibita da una ridotta disponibilità di EAA nel plasma [36]. L’entità dell’aumento della MPS in seguito al consumo di EAA è funzione della quantità ingerita. A riposo, è stato riportato che una dose orale di EAA pari a 1,5 g stimola la MPS [37], mentre la dose massima efficace, dopo la quale non si ottiene un’ulteriore stimolazione della sintesi in una singola dose, è ritenuta pari a 15-18 grammi di EAA [38]. La stimolazione delle MPS attraverso il consumo di EAA non richiede il consumo simultaneo di aminoacidi non essenziali (NEAA) [35,38]. L’inclusione di NEAA in una miscela di 18 g di EAA nel profilo di proteine di manzo non ha avuto alcun effetto sulla stimolazione mediata dagli EAA della MPS [35]. Mentre il consumo di NEAA non ha alcun effetto sulla MPS quando si consumano meno di 18 g di EAA, è possibile che quando si consumano più di 18 g di EAA i NEAA siano limitati dai tassi massimi di produzione endogena; tuttavia, sono necessarie ulteriori ricerche per esplorare questa possibilità.
Gli integratori di EAA stimolano le MPS più di una pari quantità di proteine di alta qualità, sia come isolato [39] sia come componente di un pasto [40]. È stato riscontrato che una dose orale di 3 g di EAA stimola le MPS in misura simile a 20 g di proteine isolate del siero di latte, che contengono circa 10 g di EAA [41]. Inoltre, è stato dimostrato che l’aggiunta di EAA alle proteine del siero di latte aumenta significativamente la risposta delle MPS rispetto alle sole proteine del siero di latte [42]. L’effetto stimolante superiore degli EAA in forma libera (solo singoli EAA) è legato alla maggiore quantità di EAA/grammo rispetto a una fonte proteica alimentare [43]. A causa dell’elevato tasso di assorbimento intestinale degli EAA in forma libera [44], il rapido aumento delle concentrazioni plasmatiche di EAA in circolo favorisce il trasporto verso l’interno del muscolo [23,45], con conseguente raggiungimento più rapido del picco di concentrazione intramuscolare di EAA rispetto ad altre fonti proteiche alimentari.
L’importanza delle concentrazioni plasmatiche di EAA e della velocità di aumento fino al picco di concentrazione sulla risposta MPS non è chiara, ma alcuni studi hanno trovato una relazione tra le concentrazioni plasmatiche di EAA e la MPS [46] e l’analisi di dati consolidati mostra una correlazione tra la velocità di aumento fino al picco di concentrazione di EAA e la MPS [43]. D’altra parte, non è stata riscontrata alcuna differenza nella risposta alla MPS quando la stessa dose di EAA è stata somministrata in un unico bolo o in cinque dosi più piccole nel tempo [47]. Pertanto, si concorda sull’esistenza di una relazione tra la dose di EAA e la risposta alle MPS, ma non si concorda sui meccanismi di regolazione che collegano dose e MPS.
L’impatto complessivo di un integratore alimentare sulla MPS nell’arco delle 24 ore dipende non solo dalla risposta acuta al consumo della composizione, ma anche dalle risposte anaboliche ai pasti normali. È ormai assodato che la risposta anabolica a un pasto è ridotta in seguito a un pre-carico con un isolato proteico [48]. Al contrario, una dose di 15 g di EAA non ha avuto alcun impatto sulla risposta anabolica al pasto successivo [49]. Tuttavia, è importante notare che l’integrazione di EAA in forma libera determina un aumento delle concentrazioni ematiche di EAA molto maggiore rispetto a un pasto con una quantità maggiore di EAA, poiché non richiedono digestione e vengono assorbiti rapidamente. La Figura 2 mostra gli effetti di 15 g di EAA in forma libera [50] sulle concentrazioni plasmatiche rispetto a un pasto misto contenente 70 grammi di proteine di manzo [51]. A causa della lenta digestione e del rilascio degli EAA alimentari dopo il consumo di un pasto misto, i livelli ematici aumentano solo in minima parte, rispetto a un aumento rapido e robusto dopo l’ingestione di EAA cristallini/liberi. Inoltre, questo rapido aumento (3 volte o più) predice una maggiore risposta della MPS [43].
L’effetto di 15 g di EAA in forma libera rispetto a 70 g di proteine magre di manzo e all’ingestione di un pasto misto sulla cinetica plasmatica degli EAA. Adattato dalle referenze [50,51].
Lo stato fisiologico può influenzare la risposta delle MPS agli EAA. L’invecchiamento è lo stato non clinico più comunemente studiato in cui la risposta agli EAA può essere alterata, definita resistenza anabolica. Una minore reattività delle MPS al consumo è stata ben documentata [52,53], ma non osservata in modo coerente [54]. Le diverse risposte delle MPS degli individui anziani al consumo di EAA possono essere spiegate da differenze nel profilo EAA della composizione. Ad esempio, in un’occasione Katsanos et al. hanno fornito a soggetti anziani una miscela di 6,7 g di EAA con il profilo presente nelle proteine del siero di latte (27% di leucina) e in una seconda occasione hanno fornito la stessa quantità di una miscela di EAA con la leucina che comprendeva circa il 40% del totale degli EAA [55]. In questo studio, la composizione al 40% di leucina ha stimolato la sintesi proteica muscolare circa il 50% in più rispetto al profilo di leucina inferiore, nonostante contenesse la stessa quantità di EAA totali [55]. Questi risultati dimostrano la potenziale importanza del profilo EAA in una composizione. Tuttavia, le combinazioni dei nove EAA sono pressoché illimitate e ci sono pochi dati che confrontano direttamente l’efficacia di diversi profili di EAA nella stessa circostanza. A causa della mancanza di dati comparativi sufficienti, l’impatto dei diversi profili di EAA sulla MPS non sarà discusso in questo documento. A questo proposito sono necessarie ulteriori ricerche per determinare i profili ottimali per i vari requisiti fisiologici.
EAA e bilancio proteico ed energetico dell’intero organismo
Un bilancio netto negativo delle proteine in tutto il corpo diverse da quelle muscolari (cioè nei tessuti e negli organi; riflesso dai tassi di sintesi e di degradazione delle proteine nell’intero organismo) influisce negativamente sulle proteine muscolari e quindi sulle prestazioni fisiche. Se l’assunzione di precursori di EAA con la dieta non è sufficiente a soddisfare il fabbisogno di tutto l’organismo, la scomposizione delle proteine muscolari e il rilascio di aminoacidi nel sangue forniranno gli EAA necessari. Un bilancio energetico negativo influirà indirettamente anche sulle proteine muscolari, poiché gli EAA ingeriti saranno almeno in parte destinati all’ossidazione per la produzione di energia, anziché essere incanalati verso la sintesi proteica muscolare [56]. Una discussione sugli effetti degli EAA sulla sintesi proteica muscolare deve quindi essere considerata nel contesto dello stato dell’equilibrio proteico ed energetico dell’intero organismo.
I periodi di deficit calorico sono comuni nelle categorie di peso e negli sport di resistenza come la maratona e il nuoto di distanza, dove i periodi di allenamento intenso e il desiderio di avere un peso corporeo ridotto possono limitare l’apporto calorico [57]. Il deficit calorico aumenta il fabbisogno di EAA dell’intero organismo [58]. Ad esempio, cinque giorni di deficit calorico del 30% hanno richiesto un aumento di 3 volte dell’assunzione di EAA per produrre un bilancio proteico corporeo positivo [58]. Se non si riesce a soddisfare l’aumento del fabbisogno di EAA dell’intero corpo, si verifica una scomposizione netta delle proteine muscolari per fornire gli EAA necessari e non si può invertire completamente la tendenza fino a quando non si soddisfa il fabbisogno dell’intero corpo.
Molti stati clinici inducono cambiamenti nel metabolismo proteico dell’intero corpo che influenzano il fabbisogno di EAA e l’equilibrio proteico muscolare. Possono esserci nuove richieste di precursori di EAA per funzioni quali la riparazione dei tessuti danneggiati, la guarigione delle ferite e la produzione di proteine della fase acuta. In queste condizioni è probabile che la normale risposta anabolica del muscolo scheletrico agli EAA assunti con la dieta diminuisca (resistenza anabolica). Di conseguenza, la rapida perdita di massa muscolare è una complicazione comune di gravi malattie e lesioni [59]. La stessa risposta può verificarsi, anche se in misura minore, in seguito a un allenamento intenso o a un evento agonistico, in particolare negli atleti che si trovano volontariamente o involontariamente in deficit calorico.
Punti chiave: Effetti degli EAA sul muscolo e sulle proteine dell’intero corpo
Esiste una dose-risposta degli EAA orali sulla sintesi proteica del muscolo scheletrico che raggiunge un plateau a circa 15-18 g.
Esiste una relazione tra la cinetica degli EAA plasmatici e la stimolazione della sintesi proteica.
Gli EAA orali stimolano la sintesi proteica muscolare in misura maggiore rispetto a una pari quantità di proteine di alta qualità.
La riduzione della risposta anabolica con l’invecchiamento richiede un diverso profilo di EAA, in particolare una maggiore proporzione di leucina.
Il fabbisogno di EAA dell’intero organismo aumenta con il deficit calorico. Se questo fabbisogno non è soddisfatto, si verifica una disgregazione netta delle proteine muscolari per fornire gli EAA necessari.
EAA e funzione fisica in assenza di esercizio fisico
Diversi studi documentano che la stimolazione acuta delle MPS da parte di composizioni libere di EAA si traduce in un aumento a lungo termine della massa e della funzione muscolare, anche in assenza di un controllo dell’assunzione di proteine con la dieta. Gli studi sui risultati sono stati generalmente condotti su individui anziani. Utilizzando un disegno randomizzato, in doppio cieco e controllato con placebo, donne anziane sono state assegnate a ricevere placebo o 15 g di EAA al giorno per tre mesi [60]. L’ingestione di 7,5 g di EAA ha stimolato acutamente la FSR delle proteine muscolari in entrambi i gruppi al basale [60]. La FSR basale a tre mesi era aumentata solo in coloro che ricevevano un’integrazione giornaliera di EAA, e l’entità della risposta acuta agli EAA era inalterata dopo tre mesi di consumo di EAA. Coerentemente con i dati sulla FSR delle proteine muscolari, la massa corporea magra è aumentata significativamente nei soggetti che ricevevano EAA ma non il placebo [36]. In uno studio simile, 12 soggetti intolleranti al glucosio hanno ingerito 11 g di EAA due volte al giorno tra i pasti per 16 settimane [61]. La dieta e l’attività fisica non sono state modificate in altro modo. Il consumo di EAA ha aumentato la massa magra e, soprattutto, ha migliorato una serie di parametri della funzione fisica [61]. In un gruppo di 38 donne anziane (≥75 anni), l’integrazione quotidiana con 3 g di EAA due volte al giorno per tre mesi ha migliorato significativamente la velocità di camminata [62]. Risultati simili sono stati osservati in uno studio che ha incluso 92 persone anziane a bassa funzionalità a cui sono stati somministrati per 12 settimane integratori di 15 g di proteine isolate del siero di latte, EAA (12 g di EAA più 3 g di aromi) o educazione alimentare [63]. I soggetti che hanno ricevuto gli EAA hanno migliorato significativamente la distanza percorsa in 6 minuti, la forza di presa e la forza delle gambe (coppia di picco misurata con il Cybex). Anche i soggetti che hanno ricevuto le proteine del siero del latte hanno migliorato significativamente la distanza percorsa in 6 minuti, ma il miglioramento è stato significativamente inferiore rispetto a quelli che hanno ricevuto gli EAA. La forza delle gambe non è migliorata nel gruppo del siero di latte. È interessante notare che la distanza percorsa dal gruppo di educazione alimentare è diminuita nel corso delle 12 settimane di intervento [63]. A riprova del potenziale impatto positivo dell’integrazione di EAA sui miglioramenti funzionali, l’entità del miglioramento osservato nella distanza di cammino di 6 minuti nei soggetti che ricevevano EAA era approssimativamente la stessa riportata in una revisione sistematica di studi che riportavano i risultati di 2-6 mesi di allenamento di resistenza in 241 individui [64]. Questi risultati sono coerenti con l’estrapolazione degli effetti acuti della somministrazione di EAA sul controllo della MPS [46] attraverso la farmacocinetica ematica [43]. In uno studio condotto su persone anziane e sane costrette al riposo a letto per 10 giorni, il consumo di tre dosi di 15 g di EAA al giorno ha attenuato il declino della funzione fisica evidente nei soggetti che avevano ricevuto un placebo equivalente dal punto di vista calorico [65]. In sintesi, gli studi esistenti sugli effetti degli EAA sui risultati funzionali in assenza di allenamento si sono concentrati in gran parte su popolazioni anziane o compromesse. Poiché gli EAA sono potenti stimolatori dell’anabolismo proteico muscolare e dell’intero corpo, queste popolazioni sono logiche destinatarie di indagini primarie in quanto manifestano una resistenza anabolica nel muscolo scheletrico che porta alla perdita muscolare, alla debolezza funzionale, alle comorbidità e ad altri esiti clinici negativi. Pertanto, è opportuno raccomandare ulteriori studi per determinare l’effetto anabolico degli EAA in giovani individui sani in assenza di allenamento.
Interazione degli EAA con l’esercizio fisico
I primi esami incrociati degli effetti degli aminoacidi sul muscolo scheletrico hanno dimostrato che l’aumento della somministrazione di aminoacidi a riposo determina una stimolazione del trasporto di aminoacidi verso l’interno del muscolo scheletrico (dal 30 al 100%, a seconda dei singoli EAA), una stimolazione della sintesi proteica (dal 30 al 300%) e un miglioramento del bilancio netto di aminoacidi [9]. Quando l’esercizio di resistenza della parte inferiore del corpo è stato eseguito prima dell’infusione di aminoacidi, si sono verificati effetti maggiori nel trasporto verso l’interno di alcuni aminoacidi, ma soprattutto si sono registrati aumenti ancora maggiori nella sintesi proteica e nel bilancio proteico muscolare netto [9]. In ogni caso, non si sono verificati cambiamenti nella degradazione delle proteine muscolari. È importante notare che gli effetti combinati dell’esercizio di resistenza e dell’aumento dell’apporto di aminoacidi sono interattivi. Quando lo stesso esercizio di resistenza è stato eseguito senza la somministrazione di aminoacidi, si è registrato un aumento del trasporto interno di aminoacidi e della sintesi proteica muscolare; tuttavia, il bilancio muscolare netto è rimasto negativo [66]. L’assenza di miglioramento del bilancio netto è dovuta all’aumento della sintesi e della degradazione delle proteine dopo il solo esercizio di resistenza [66]. Pertanto, l’esercizio di resistenza da solo non porta all’anabolismo muscolare (il bilancio netto delle proteine muscolari è negativo). L’anabolismo si verifica solo se supportato dai precursori amminoacidici necessari. La Figura 3 rappresenta graficamente il bilancio trasversale degli arti della fenilalanina (un surrogato del bilancio degli aminoacidi, poiché non viene metabolizzata nel muscolo scheletrico) a digiuno, dopo il solo esercizio di resistenza, dopo l’infusione dei soli aminoacidi e con l’infusione combinata di aminoacidi ed esercizio di resistenza. Gli effetti interattivi degli EAA somministrati per via orale e dell’esercizio di resistenza hanno rivelato risultati simili. L’esercizio di resistenza della parte inferiore del corpo è stato seguito dalla somministrazione di EAA per via orale o di una miscela completa di aminoacidi. I risultati hanno indicato che gli EAA dopo l’esercizio fisico hanno migliorato il bilancio netto muscolare nella stessa misura della miscela completa, fornendo ulteriori prove [38] del fatto che, se somministrati insieme all’esercizio fisico, solo gli EAA sono necessari per stimolare l’anabolismo muscolare [67]. Anche l’ingestione orale in bolo di EAA dopo l’esercizio di resistenza della parte inferiore del corpo ha aumentato la sintesi proteica e il bilancio netto muscolare, indipendentemente dal fatto che la bevanda sia stata consumata una o tre ore dopo l’esercizio [68]. Vi sono indicazioni che i benefici degli EAA sul muscolo scheletrico potrebbero non dipendere interamente dalla stimolazione delle MPS. L’integrazione di EAA ha migliorato l’evidenza istologica del danno muscolare e ha ridotto la perdita di forza muscolare, anche in assenza di cambiamenti nella MPS [69].
Bilancio netto muscolare di fenilalanina (umol/kg/min) durante il digiuno, l’esercizio di resistenza da solo (RE), la somministrazione/infusione completa di aminoacidi da sola (AA) e con la combinazione di AA e RE. È stato dimostrato un effetto interattivo tra la somministrazione di RE e AA. Dati derivati dalle referenze [9,66].
Gli effetti interattivi degli EAA e dell’esercizio di resistenza si riflettono nella segnalazione dell’avvio della traduzione nel muscolo. Ai volontari è stata somministrata una soluzione di placebo, leucina, BCAA o EAA dopo l’esercizio contro resistenza. I risultati hanno indicato che 90 minuti dopo il recupero dell’esercizio, l’attivazione della proteina ribosomiale S6K1 e del fattore di iniziazione della traduzione eucariotica 4E-BP1, nonché una riduzione sostenuta dell’interazione 4E-BP1:eIF4E, erano maggiori con gli EAA [70]. Mentre nello studio sugli EAA è stato osservato un aumento di 9 volte dell’espressione di S6K1, la stimolazione complessiva dell’iniziazione della traduzione è stata più efficace con gli EAA, con conseguente aumento progressivo dell’iniziazione della traduzione (placebo < leucina < BCAA < EAA) [70]. Uno studio condotto su giovani uomini ha confermato l’aumento della via di segnalazione mammalian target of rapamycin complex 1 (mTORC1) dopo l’esercizio di resistenza, ma ha anche osservato che un integratore di EAA mantiene mTORC1 nelle regioni periferiche delle fibre muscolari, più vicino al suo attivatore diretto Rheb [71]. Gli autori hanno ipotizzato che “la localizzazione intracellulare di mTOR può servire a innescare la chinasi per futuri stimoli anabolici” [71]. Gli effetti degli EAA e dell’esercizio di resistenza sull’aumento della segnalazione anabolica sono coerenti con l’invecchiamento. Gli EAA e l’esercizio di resistenza aumentano le concentrazioni periferiche di EAA in egual misura sia nei soggetti giovani che in quelli più anziani [72]. Indipendentemente dall’età, con il trattamento combinato sono stati dimostrati aumenti di mTOR (Ser2481) e della proteina ribosomiale S6 (Ser235/236) [72], il che suggerisce una maggiore sensibilità muscolare agli stimoli combinati. inoltre, la somministrazione di EAA dopo l’esercizio fisico in uomini anziani porta a un aumento della proliferazione delle cellule satelliti [73]. Il prodotto metabolico di questa segnalazione è un miglioramento della sintesi proteica muscolare. Inoltre, donne anziane hanno eseguito un’estensione unilaterale del ginocchio e poi hanno consumato 1,5 o 6 g di una formula EAA con quantità variabili di leucina, oppure 20 o 40 g di proteine del siero di latte. I risultati hanno indicato che le dosi di 1,5 g e 6 g di EAA erano efficaci quanto 40 g di proteine del siero del latte nella stimolazione della sintesi proteica muscolare acuta (miofibrillare) [37].
Strategie di integrazione: Tempistica degli EAA
È necessario considerare la tempistica della somministrazione di EAA in relazione all’esercizio di resistenza. I soggetti a cui sono stati somministrati EAA immediatamente prima o dopo l’esercizio di resistenza hanno entrambi registrato un aumento del 130% delle concentrazioni di fenilalanina arteriosa e muscolare [74]. Tuttavia, l’effetto sull’assorbimento netto di fenilalanina (una misura indiretta della sintesi proteica netta) è stato molto maggiore quando la bevanda è stata consumata subito prima dell’esercizio [74]. È importante notare che ogni trattamento ha portato a un bilancio netto di fenilalanina positivo; tuttavia, il migliore apporto di aminoacidi (flusso sanguigno X concentrazione arteriosa di EAA) quando è stato consumato immediatamente prima dell’esercizio ha portato a un apporto di aminoacidi circa 3 volte superiore [74]. L’effetto stimolante dell’esercizio di resistenza sul flusso sanguigno muscolare, se combinato con il maggiore apporto di aminoacidi derivante dal consumo prima dell’esercizio, determina una maggiore risposta anabolica nel muscolo scheletrico. Ciò è coerente con una recente revisione che denota la relazione tra l’aumento degli EAA periferici e la stimolazione della sintesi proteica muscolare e dell’intero corpo [43]. Altri lavori non confermano questi risultati, poiché una soluzione di carboidrati/EAA somministrata prima dell’esercizio di resistenza non ha migliorato la sintesi proteica muscolare post-esercizio [75]; tuttavia, le differenze metodologiche/interpretative e l’assunzione di carboidrati possono complicare la coerenza dell’interpretazione. Inoltre, è stato dimostrato che un secondo bolo di EAA un’ora dopo il primo duplica l’anabolismo muscolare della prima somministrazione, indicando che i meccanismi di sintesi non sono inattivi dopo una prima stimolazione [76].
Interazione degli EAA con altre modalità di esercizio
A causa dei forti effetti interattivi dimostrati con gli EAA e l’esercizio di resistenza, la maggior parte del lavoro si è concentrata su questa combinazione. Anche i dati sulla combinazione di EAA ed esercizio aerobico sono coerenti con i suoi effetti sull’anabolismo proteico del muscolo scheletrico. Nei giovani adulti che eseguivano 90 minuti di esercizio in cicloergometria o con carico ponderato (30% della massa corporea) su tapis roulant, con o senza EAA (consumati ogni 30 minuti per tutta la durata dell’esercizio), la sintesi proteica muscolare era maggiore durante ciascuna modalità di esercizio con gli EAA [77]. Tuttavia, il trasporto del carico e gli EAA hanno determinato un aumento maggiore della MPS sia durante che dopo l’esercizio [77]. Questi risultati indicano che il carico del muscolo scheletrico fornisce uno stimolo più forte per gli effetti combinati di EAA ed esercizio fisico. L’interazione tra EAA ed esercizio aerobico è consistente anche nell’invecchiamento. Volontari anziani (72 ± 1 anno) sono stati randomizzati a ricevere 15 g/d di EAA o 15 g/d di EAA più 3 giorni/settimana di allenamento aerobico supervisionato. Coerentemente con i risultati precedenti, l’assunzione acuta di EAA prima dell’intervento ha aumentato la MPS [78]. Tuttavia, dopo 24 settimane di intervento, il gruppo che combinava EAA ed esercizio aerobico presentava una maggiore risposta sintetica del muscolo agli EAA rispetto al gruppo dei soli EAA [78], indicando che l’esercizio fisico costante sensibilizza ulteriormente il muscolo scheletrico agli effetti anabolici degli EAA. Soprattutto, la maggiore sensibilità del muscolo scheletrico agli EAA si è tradotta in una maggiore qualità muscolare e in una maggiore velocità di cammino sui 400 m nel gruppo EAA più esercizio aerobico [78].
L’esercizio/allenamento ad alta intensità e intermittente (HIIT) e la maggior parte degli sport di squadra (calcio, basket, hockey, tennis, ecc.) richiedono componenti di allenamento sia aerobico che anaerobico/di resistenza. Gli studi condotti sulle proteine suggeriscono che l’esercizio fisico ad alta intensità intrapreso con una maggiore disponibilità di proteine (cioè eseguito in uno stato di alimentazione con proteine) può migliorare sinergicamente l’ipertrofia muscolare [79]. Altri potenziali benefici includono l’aumento della biogenesi mitocondriale, il recupero dell’esercizio, la capacità aerobica e il miglioramento delle prestazioni di sprint [80]. Ad oggi, le poche ricerche che utilizzano EAA e HIIT non sono ancora definitive. In adulti sovrappeso/obesi non allenati, 8 settimane di allenamento a intervalli ad alta intensità (HIIT; 6-10 × 1 min@90% W max: 1 min di riposo) hanno prodotto un aumento significativo delle dimensioni, della sezione trasversale, del volume e della qualità del muscolo della coscia, ma non è stato potenziato sinergicamente da una bassa dose di EAA (3,6 g due volte al giorno) [81]. Inoltre, anche il VO2 è aumentato con l’HIIT, ma non è stato potenziato sinergicamente dagli EAA [82]. Nella stessa coorte, non sono stati riscontrati effetti acuti (3,6 g prima dell’esercizio) o cronici (dopo 4 e 8 settimane di 3,6 g due volte al giorno) dell’integrazione di EAA sul tempo di esaurimento o sulla progressione del carico di lavoro [83]. Sebbene non vi siano prove che suggeriscano che l’integrazione di EAA sia dannosa o attenui gli adattamenti fisiologici associati all’HIIT, non vi sono prove sufficienti per trarre conclusioni sui benefici in termini di adattamento e prestazioni. Tuttavia, c’è motivo di aspettarsi un’interazione degli EAA con l’esercizio aerobico. La sola camminata moderata su tapis roulant (45 minuti al 40% di VO2 di picco) in uomini giovani e anziani ha aumentato la sintesi proteica muscolare subito dopo l’esercizio, con la risposta dei più giovani mantenuta fino a un’ora dopo l’esercizio [84]. Anche la sintesi di fibrinogeno è stata elevata in entrambi i gruppi fino a tre ore dopo l’esercizio [84]. L’apporto di aminoacidi è stato maggiore subito dopo l’esercizio, il che denota ancora una volta l’effetto dell’esercizio sul flusso sanguigno degli arti. Pertanto, se l’aumento del flusso sanguigno si combina con un maggiore apporto di aminoacidi per via orale, ci si aspetta un anabolismo muscolare.
Per quanto riguarda le interazioni con l’esercizio fisico, gli EAA in forma libera possono essere presi in considerazione rispetto alle proteine intatte (siero di latte) sulla base della facilità di assunzione in prossimità e durante l’esercizio. Le formule orali di EAA in forma libera richiedono una digestione minima, comportano un carico gastrico minimo e sono rapidamente assorbite e trasportate in periferia. Per questo motivo, sono ideali per il consumo prima dell’esecuzione di un esercizio fisico rigoroso.
EAA e condizioni cliniche ed esiti
Gli effetti benefici dell’integrazione della dieta con EAA sono stati dimostrati in un’ampia varietà di condizioni cliniche. Le condizioni in gran parte associate all’invecchiamento sono state un obiettivo frequente della terapia con EAA, tra cui la sarcopenia [62,85], le infezioni contratte durante l’assistenza a lungo termine [86], la scarsa funzionalità fisica [63] e l’insufficienza cardiaca [40,87,88]. Gli effetti benefici degli EAA sono stati riportati anche nelle seguenti condizioni o situazioni: riabilitazione [89-92]; ictus [93,94]; riposo a letto/immobilizzazione [8,65,95-97]; malattia arteriosa periferica [98]; insufficienza renale [99-103]; infiammazione [104,105]; malattia critica [106]; cancro del polmone [107]; fibrosi cistica [108]; broncopneumopatia cronica ostruttiva [109-111]; guarigione delle ferite [112]; lesioni cerebrali [113,114]; sindrome metabolica e fattori di rischio cardiovascolare [115-117]; obesità [118,119]; grasso epatico [115,120-122]; e diabete [123-127]. È importante notare che in tutti questi studi gli effetti benefici sono stati osservati nonostante l’assenza di controllo del consumo di EAA contenuti nelle proteine alimentari, il che implica l’importanza di un assorbimento rapido e completo degli EAA liberi in circostanze cliniche in cui la digestione può essere compromessa e la resistenza anabolica è prevalente. I risultati ottenuti nelle popolazioni cliniche evidenziano la necessità e il potenziale impatto dell’integrazione di EAA nell’ambito degli sforzi di riabilitazione da lesioni ortopediche e interventi chirurgici associati. Questi scenari rappresentano gli ambienti clinici più probabili in cui si troveranno gli atleti agonisti e questa rimane un’area di ricerca poco esplorata.
Punti chiave: EAA, esercizio fisico e loro funzione
Gli effetti anabolici degli EAA e dell’esercizio fisico sono interattivi, in quanto la risposta alla combinazione è maggiore della somma delle singole risposte. L’aumento del flusso sanguigno degli arti indotto dall’esercizio contribuisce a questa risposta aumentando il trasporto di EAA nel muscolo scheletrico.
La combinazione di EAA ed esercizio aerobico, così come altre modalità di carico muscolare, è anche anabolizzante per il muscolo scheletrico.
Gli EAA somministrati prima dell’esercizio fisico determinano un anabolismo maggiore rispetto a quelli assunti dopo il completamento dell’esercizio; tuttavia, un effetto anabolico minore può essere realizzato entro un’ora dalla cessazione dell’esercizio.
In assenza di uno stimolo all’esercizio, la somministrazione di EAA in popolazioni anabolicamente resistenti, come quelle affette da invecchiamento e patologie cliniche, si è dimostrata benefica per i risultati clinici ed efficace nel ripristino della forza e delle prestazioni funzionali.
Domande rimanenti
Gli effetti metabolici degli EAA sono stati elegantemente articolati. Inoltre, è stata stabilita la loro interazione metabolica con l’esercizio di resistenza e aerobico. Esiste quindi una base metabolica per la traduzione dell’integrazione di EAA in risultati di performance. Come accennato in precedenza, il sostanziale effetto anabolico dell’integrazione di EAA in forma libera è coerente con la loro efficacia nelle popolazioni che trarrebbero maggior beneficio dal loro utilizzo. Per questo motivo, la stragrande maggioranza dei lavori longitudinali che hanno esaminato l’integrazione di EAA e i risultati funzionali si è svolta in popolazioni anziane o cliniche caratterizzate da insensibilità anabolica muscolare, perdita muscolare e/o debolezza muscolare. Meno lavori hanno documentato gli effetti/risultati a lungo termine associati all’uso di EAA in popolazioni di atleti. Inoltre, sarebbe utile un ulteriore lavoro che definisca il profilo ottimale degli EAA, nonché il dosaggio e la tempistica ottimali per circostanze specifiche.
Stechiometria degli EAA – l’importanza della corretta AA ratio
La stechiometria è il calcolo delle quantità relative di reagenti e prodotti nelle reazioni chimiche. La stechiometria si basa sulla legge di conservazione della massa, secondo la quale la massa totale dei reagenti è uguale alla massa totale dei prodotti, il che porta a capire che le relazioni tra le quantità di reagenti e prodotti formano tipicamente un rapporto di numeri interi positivi. Ciò significa che se le quantità dei singoli reagenti sono note, è possibile calcolare la quantità del prodotto. Al contrario, se un reagente ha una quantità nota e la quantità di prodotto può essere determinata empiricamente, è possibile calcolare anche la quantità degli altri reagenti.
In questo modo si sviluppano metodi per determinare le quantità di composti prodotti o consumati nelle reazioni chimiche e si descrivono alcuni tipi fondamentali di reazioni chimiche. Applicando i concetti e le competenze introdotte, si è in grado di spiegare cosa succede allo zucchero contenuto in una barretta di cioccolato che si mangia, o agli AA contenuti in una fonte proteica e/o integratore, tanto per fare un esempio.
Justus von Liebig
Il concetto di reagente limitante è stato utilizzato dal chimico tedesco Justus von Liebig (1803-1873) nel XIX secolo per ricavare un’importante legge biologica ed ecologica. La legge del minimo di Liebig afferma che la sostanza essenziale disponibile nella quantità più piccola rispetto a un minimo critico controllerà la crescita e la riproduzione di qualsiasi specie vegetale o animale. Quando un gruppo di organismi esaurisce il reagente limitante essenziale, le reazioni chimiche necessarie per la crescita e la riproduzione devono arrestarsi. Vitamine, proteine e altri nutrienti sono essenziali per la crescita del corpo umano e delle popolazioni umane.
Alcuni di voi conosceranno già la “complementarietà degli AA”. In breve, e per fare un esempio, essa riguarda il modo più efficace per ottenere tutti i 9 EAA nella dieta di un vegetariano. La complementarietà degli AA consiste nel combinare due proteine vegetali (ad esempio, legumi e cereali) per ottenere tutti i 9 aminoacidi essenziali per l’organismo. La ripartizione della complementarietà degli AA è la seguente:
Quindi, l’idea della complementarità degli AA, non è altro che l’abbinamento di diversi alimenti della dieta al fine di fornire il giusto equilibrio di aminoacidi per costruire le proteine umane. Le proteine umane richiedono quantità stechiometriche di circa 9 aminoacidi “essenziali”; se ne manca solo uno, la proteina non può essere sintetizzata e ne deriva una malnutrizione proteica. Questo illustra il concetto di “reagente limitante”, ovvero un reagente presente in quantità inferiore a quella stechiometrica rispetto agli altri reagenti.
Un gruppo eterogeneo di culture sopravvissute in tutto il mondo ha adottato diete, per quanto diverse, che forniscono il corretto equilibrio di aminoacidi. Gli antropologi ritengono che l’adozione fortuita di queste diete abbia fornito alle culture un valore di sopravvivenza. Nelle aree in cui il cibo scarseggia, le culture che non adottano diete con un corretto equilibrio di aminoacidi potrebbero non sopravvivere.
In Sud America sono comuni le diete che combinano le tortillas di mais o altri prodotti a base di mais con i fagioli, come la tostada. I fagioli mangiati da soli forniscono quantità limitate di aminoacidi contenenti zolfo, come la metionina e la cisteina, quindi questi aminoacidi limitano la quantità di proteine umane che possono essere sintetizzate. I fagioli contengono grandi quantità di aminoacidi come la lisina e il triptofano, che sono quindi “reagenti in eccesso” quando vengono utilizzati per sintetizzare le proteine umane e vengono degradati in urea e sprecati. Se il grano, il riso o il mais vengono consumati da soli, in genere forniscono quantità di lisina e triptofano che limitano la quantità di proteine umane che possono essere sintetizzate. Ma se i fagioli vengono mangiati insieme ai cereali o al mais, i reagenti in eccesso dei fagioli completano i reagenti limitanti dei cereali (e viceversa). Le proteine umane possono quindi essere sintetizzate in modo efficiente e quantità molto ridotte di aminoacidi vengono semplicemente espulse come urea.
In India, il riso o il chapati vengono consumati con le lenticchie per fornire un equilibrio di aminoacidi.
Per capire meglio il concetto di stechiometria, i questa sede analizzeremo gli esperimenti che riguardano il mais e la malnutrizione proteica che ne può derivare se viene mangiato da solo. È noto che il mais è un alimento povero di proteine di per sé, essendo povero di lisina e triptofano [8], e ancora di più quando viene nixtamalizzato. Queste carenze hanno spinto i ricercatori a sviluppare il QPM (Quality Protein Maze) per aumentare le concentrazioni di questi aminoacidi essenziali nelle sue proteine.
Esperimenti sui ratti, il cui fabbisogno di aminoacidi è simile a quello degli esseri umani, forniscono una base per alcuni calcoli stechiometrici [9]. Come mostra la tabella sottostante, l’aumento di peso è stato minimo quando LYS o TRP sono stati aggiunti separatamente a una dieta “base”. Ciò significa che nessuno dei due, da solo, è un aminoacido limitante e impedisce la sintesi delle proteine dei topi. Ma quando sono stati aggiunti entrambi, è stato misurato un aumento significativo, insieme a una diminuzione dell’urea sierica. Ciò significa che entrambi erano limitanti e che, quando sono stati aggiunti entrambi, è stata sprecata una quantità molto minore di aminoacidi. Gli aminoacidi sono stati destinati alla sintesi proteica, anziché essere semplicemente metabolizzati ed escreti come urea. Mentre l’aggiunta di isoleucina nella dieta 5 ha fatto poca differenza (quindi deve essere fornita adeguatamente dal mais), l’aggiunta della sola treonina (THR) alla dieta 4 ha aumentato l’aumento di peso corporeo, quindi deve essere limitante in una dieta di mais + LYS + TRP. L’aggiunta di ILE, metionina (MET), istidina (HIS) e valina (VAL) costituisce infine una dieta quasi bilanciata, come dimostrano i grandi aumenti di peso e la bassa urea sierica della dieta 8.
La dieta di base consisteva in 920,2 g di mais, 30,0 g di olio di mais, 35,0 g di miscela di minerali, 10 g di miscela di vitamine, 2,5 g di calcare e 2,3 g di colina. b Questo forte aumento è dovuto al raddoppio delle proteine totali della dieta, piuttosto che all’inefficienza proteica.
Lo stesso documento[10] fornisce la sequenza, dalla più limitante alla meno, degli aminoacidi del mais e il fabbisogno del ratto (simile a quello umano) pubblicato dal NRC[11]. Abbiamo aggiunto le quantità di aminoacidi nelle lenticchie per rappresentare i fagioli[12].
Corretta stechiometria o ratio degli aminoacidi:
In base ai dati della Tabella II, calcolare il rapporto molare ottimale tra lisina (LYS, C6H14N2O2, massa molare 146,19 g/mol) e arginina (ARG, C6H14N4O2, massa molare 174,2 g/mol) nella dieta.
Aminoacido limitante:
Dai dati della tabella precedente, calcolare quale, LYS o ARG, è il reagente limitante in una dieta a base di mais?
c. Quanto aminoacido in eccesso verrà sprecato in 1kg di dieta a base di mais?
Soluzione
a. L’equazione bilanciata avrà dei coefficienti che dobbiamo determinare:
x LYS + y ARG + altri amminoacidi → Proteina + acqua
Secondo la teoria atomica, per ogni y mole di ARG sono necessarie x mol di LYS per produrre proteine utili. Se la dieta è ottimale quando si consumano 7,0 g di LYS per ogni 6,0 g di ARG (questo rapporto potrebbe essere fornito dalle uova, per esempio), possiamo calcolare il rapporto stechiometrico ottimale:
Questo rapporto esatto di aminoacidi potrebbe non essere presente in una particolare proteina del ratto, ma è il rapporto medio per tutte le proteine del ratto. L’equazione chimica (parziale) è
b. Vediamo qual è il rapporto molare nella dieta a base di mais.
Chiaramente, la LYS è l’amminoacido limitante perché il rapporto iniziale tra LYS e ARG (0,55:1) è molto inferiore al rapporto stechiometrico richiesto (1,4:1).
Dall’esempio della dieta a base di mais si può iniziare a capire cosa bisogna fare per determinare quale dei due reagenti, X o Y, è limitante. Dobbiamo confrontare il rapporto stechiometrico S(X/Y) con il rapporto effettivo delle quantità di X e Y inizialmente mescolate.
La regola generale corrispondente, per qualsiasi reagente X e Y, è
Naturalmente, quando le quantità di X e Y sono esattamente nel rapporto stechiometrico, entrambi i reagenti vengono consumati completamente nello stesso momento e nessuno dei due è in eccesso.
Possiamo verificare che LYS è limitante calcolando la quantità di LYS che sarebbe necessaria se tutta la ARG reagisse, utilizzando un rapporto stechiometrico:
Allo stesso modo, se tutta la LYS reagisce, la quantità di ARG richiesta è
Questa quantità è molto inferiore a quella presente, quindi ARG è il reagente in eccesso.
c. Possiamo facilmente utilizzare le masse molari per convertire le quantità di ciascun amminoacido in masse.
Prepariamo una tabella:
Al termine della reazione, rimangono 3,14 g dei 5,0 g originali (63%), che saranno metabolizzati in urea ed escreti. Dell’intera dieta di 2,4 g di LYS + 5,0 g di ARG, vediamo che 3,14 g o il 42% viene sprecato. Che spreco di cibo e di risorse per produrlo! Si noti che anche la porzione di mais dovrà essere maggiore rispetto alla porzione di dieta ottimale per ottenere la stessa quantità di proteine, perché il mais ha solo 5 g/kg di ARG, mentre la dieta ottimale ha 6,0 g/kg. La dieta ottimale potrebbe essere fornita dalla carne o dalle uova, ma sono impegnative per l’ambiente e presentano problemi di salute.
Una dieta di sussistenza a base di fagioli può anche portare a una malnutrizione proteica, come mostra la tabella II. Sebbene il contenuto di lisina sia molto superiore a quello dei fagioli, i livelli di metionina (MET) e cistina (CYS) sono bassi, così come quelli di fenilalanina (PHE) e tirosina (TYR).
Ma supponiamo che fagioli e mais vengano consumati nello stesso giorno, in una dieta che mescola in egual misura mais e lenticchie. Ricalcolate ora l’amminoacido limitante in una dieta con 2,4 + 6,3 = 8,7 g di LYS e 5,0 + 6,97 = 11,97 g di ARG.
b. In questo caso, quanta parte dell’amminoacido in eccesso viene sprecata?
Soluzione
Ora vediamo che il rapporto tra le quantità presenti è
0,060 mol LYS / 0,069 mol ARG = 0,869,
che è ancora inferiore al rapporto stechiometrico,
7 mol LYS / 5 mol ARG = 1,4
Quindi LYS è ancora una volta l’amminoacido limitante. Ricalcolando i valori in una tabella come sopra, otteniamo
In questo caso, da una porzione di 11,97 g rimangono 4,61 g di ARG in eccesso, ovvero il 38% dell’ARG. La dieta totale è di 8,7 g di LYS + 11,97 g di ARG = 20,7 g, di cui solo il 22% viene sprecato, un grande miglioramento rispetto al caso del solo mais di cui sopra.
Per progettare una dieta adeguata, gli alimenti proteici (o singoli AA) complementari dovrebbero essere scelti dalle tabelle dei contenuti di aminoacidi degli alimenti.
Il processo di Nixtamalizzazione converte parte del già limitante triptofano in 2-amminoacetofenone, aggravando lo scarso valore nutrizionale e di sopravvivenza del mais. Ma la Nixtamalizzazione rende anche più biodisponibile la Niacina contenuta nei chicchi, riducendo l’incidenza della Pellagra e più che compensando la perdita di aminoacidi se nella dieta sono inclusi anche molti fagioli. La dieta a base di fagioli e mais, anche con la Nixtamalizzazione, ha un valore di sopravvivenza.
Ninidrina
Per rilevare un amminoacido (anche in un’impronta digitale nella chimica forense), si utilizza spesso il test della ninidrina.
Nel test della ninidrina, due molecole di ninidrina (C9H6O4, mostrato a sinistra) vengono legate dalla N attaccata al primo carbonio della catena aminoacidica, producendo lo ione blu/viola mostrato di seguito.
Se si utilizzano 2,00 mg di ninidrina (Nin) per rilevare 2 mg di TRP, è stata aggiunta abbastanza ninidrina da reagire con tutta il TRP? Qual è il reagente limitante e quale massa di H2O si formerà?
La soluzione
Il rapporto stechiometrico che collega Nin e TRP è
b) La quantità di prodotto acquoso che si forma in una reazione può essere calcolata attraverso un appropriato rapporto stechiometrico dalla quantità di un reagente consumato. Rimarrà una parte del reagente in eccesso TRP, ma tutta la quantità iniziale di Nin sarà consumata. Pertanto, utilizziamo nNin (iniziale) per calcolare la quantità di H2O ottenuta.
Si tratta di 0,302mg di acqua.
Questi calcoli possono essere organizzati come una tabella, con le voci sotto i rispettivi reagenti e prodotti dell’equazione chimica. I calcoli sono mostrati per ogni possibile caso, ipotizzando che un reagente sia completamente consumato e determinando se è presente una quantità sufficiente di altri reagenti per consumarlo. In caso contrario, lo scenario viene scartato.
Come si può notare dall’esempio, nel caso in cui vi sia un reagente limitante, per calcolare la quantità di prodotto formato si deve utilizzare la quantità iniziale del reagente limitante. Utilizzare la quantità iniziale di un reagente presente in eccesso non sarebbe corretto, perché tale reagente non viene consumato interamente.
Sulla traccia della “complementarietà AA” potremmo prendere come esempio i piselli che sono poveri di Metionina ma ricchi di Lisina e, al contrario, il riso integrale che è ricco di Metionina ma povero di Lisina.
Se dovessimo prendere come esempio principale al quale applicare una stechiometria con l’aggiunta del reagente limitante da AA integrativi.
Vediamo dalla tabella sopra esposta che il reagente limitante nelle proteine del pisello è la Metionina mentre il reagente in eccesso è la Lisina.
In base ai dati precedentemente riportati, calcolare il rapporto molare ottimale tra Metionina (MHT, C5H11NO2S, massa molare 149,21 g/mol) e Lisina (LYS, C6H14N2O2, massa molare 146,19 g/mol) nella dieta.
Quindi, procedendo rapidamente…
x MHT + y LYS + altri amminoacidi → Proteina + acqua
Secondo la teoria atomica, per ogni y mole di LYS sono necessarie x mol di MHT per produrre proteine utili. Se la dieta è ottimale quando si consumano 7,0 g di LYS per ogni 2,1 g di MHT (questo rapporto potrebbe essere fornito dalle uova, per esempio), possiamo calcolare il rapporto stechiometrico ottimale:
Accelerando la procedura di calcolo, sempre sulla linea prima esposta, abbiamo 1.5g di MHT e 11.42g di LYS con mole rispettivamente di 149.21 e 146.19 = 0.010 MHT e 0.078 LYS = 0.13/1
Anche qui, la MHT è l’amminoacido limitante perché il rapporto iniziale tra MHT e LYS (0,13:1) è inferiore al rapporto stechiometrico richiesto (0.29:1).
Di conseguenza, per ogni 100g di proteine del pisello andrebbero addizionati 140mg di MHT al fine di migliorare la ratio stechiometrica.
Nel riso “bianco”, comunemente consumato dai Bodybuilder, abbiamo la presenza dello stesso reagente limitante del pisello; ossia la MHT con il reagente in eccesso LYS.
Punto della situazione
Dovremmo aver compreso, ora, che un prodotto contenete EAA andrebbe valutato in base alla sua formulazione stechiometrica la quale, per essere considerata ottimale, non deve presentare ne carenze ne eccessi per uno o più dei 9 EAA.
Sul mercato attuale sono presenti poche formulazioni stechiometriche di EEA valide. Tra queste vi è certamente la MAP® . Il MAP [Master Amino Acid Pattern®], è una combinazione stechiometrica di EAA usata anche in campo clinico. MAP fornisce, con il minor peso e volume, il maggior valore nutritivo proteico/AA in assoluto.
Clinicamente, il MAP è particolarmente consigliabile per coloro che soffrono di nausea o di inappetenza sia per ragioni fisio-patologiche (AIDS, cancro, anemia, denutrizione, ecc.) sia per ragioni psicologiche (anoressia, bulimia, ecc.)
Il MAP fornisce, in soli 10g, un valore nutritivo proteico/AA pari a circa 350g di carne rossa, pesce o pollame, con un valore energetico di sole 50Kcal. Per via della sua stechiometria, 10g di MAP, a differenza dei 350g di carne, pesce o pollame a cui equivale, rilascia una quantità minima (meno dell’1%) di cataboliti azotati. Quindi, il MAP, o altra formulazione stechiometricamente corretta, è particolarmente consigliabile per l’alimentazione di coloro che hanno deficit di funzionalità epatica o renale o per coloro i quali desiderano gravarne il meno possibile per via del già presente impatto stressorio d’organo dato da terapia iatrogena.
Il MAP viene completamente assorbito in meno di 23 minuti dalla sua ingestione, poiché è assimilabile ad una proteina alimentare digerita (ossia scomposta nelle sue singole unità AA). Il MAP non necessita quindi dell’azione dell’ enzima peptidasi, ed il suo assorbimento, stimola la secrezione biliare, pancreatica ed intestinale in misura minima. Quindi, il MAP viene particolarmente consigliato per tutti coloro che soffrono di disturbi gastrointestinali anche per bisogno di consumare grosse quantità di cibo [vedi bodybuilder].
Punti chiave del MAP:
Il MAP fornisce un 99% di Net Nitrogen Utilization (NNU): vale a dire che il 99% dei suoi aminoacidi costitutivi agiscono come precursori della sintesi proteica corporea.
Il MAP fornisce una quantità minima (meno dell’ 1%) di cataboliti azotati.
Il MAP é digerito e assorbito in meno di 23 minuti.
Sintesi e conclusioni finali:
I 15 punti seguenti costituiscono al momento “le linee guida” sugli EEA avallati dalla ISSN:
L’integrazione di EAA in forma libera (non derivati da proteine esogene intatte) è un robusto stimolatore della sintesi e del turnover delle proteine muscolari.
Gli EAA stimolano la sintesi proteica muscolare più di un isolato proteico isonitrogeno.
L’ingestione di EAA produce un rapido aumento delle concentrazioni periferiche e il trasporto degli aminoacidi nel muscolo scheletrico.
La stimolazione degli EAA della sintesi proteica muscolare può avvenire con dosaggi multipli e non interferisce con gli effetti dei pasti.
Singoli o gruppi di EAA possono avviare il processo di stimolazione; tuttavia, una stimolazione significativa e prolungata si verifica quando vengono consumati tutti gli EAA.
La stimolazione della sintesi proteica a riposo da parte degli EAA avviene con dosaggi che vanno da 1,5 g a 18 g.
Una percentuale maggiore di leucina (%/g) contenuta nelle composizioni di EAA ingerite è necessaria per stimolare al massimo la sintesi proteica muscolare nelle popolazioni (invecchiamento, patologie cliniche) che dimostrano resistenza anabolica.
Nelle popolazioni anabolicamente resistenti, l’integrazione longitudinale di EAA migliora i risultati funzionali.
Gli effetti degli EAA e dell’esercizio fisico sono interattivi, tanto da amplificare gli effetti combinati. Questa interazione è dovuta a un maggiore apporto di EAA al muscolo in esercizio, grazie all’aumento del flusso sanguigno e alle più alte concentrazioni di EAA nel sangue.
Le risposte anaboliche sono costantemente riportate con la combinazione dell’ingestione di EAA con l’esercizio di resistenza o aerobico. Questo effetto si mantiene con l’invecchiamento.
L’integrazione di EAA in forma libera rientra nel limite massimo di sicurezza del consumo giornaliero abituale.
L’integrazione di EAA è efficace nella maggior parte degli studi clinici e delle condizioni.
Numerosi studi longitudinali che prevedono l’integrazione di EAA in popolazioni anziane riportano costantemente miglioramenti favorevoli sia a livello metabolico che funzionale.
Sono necessarie ulteriori ricerche per esaminare il potenziale impatto della somministrazione di EAA in popolazioni atletiche sottoposte intenzionalmente o meno a privazione energetica sui cambiamenti del metabolismo proteico muscolare e sui cambiamenti associati in termini di prestazioni e composizione corporea.
Sono necessarie ulteriori ricerche per esaminare il ruolo della somministrazione di EAA a popolazioni di atleti che attraversano periodi inaspettati e improvvisi di inattività, probabilmente secondari a lesioni acute e a periodi di riabilitazione che seguono abitualmente interventi chirurgici.
L’integrazione con EAA vede le sue migliori applicazioni in:
Presenza di una carenza proteica, o comunque una condizione alimentare che non da modo di apportare la giusta quantità di EAA con gli alimenti;
Bodybuilder in ipercalorica che vogliono ridurre il carico di cibo e massimizzare al meglio digestione e assorbimento AA.
Soggetto che non tollera le proteine in polvere;
Inserimento “compensativo” per ridurre il consumo di fonti proteiche intere.
Soggetto con problematiche digestive di base.
Assicuratevi di acquistare prodotti di alta qualità altrimenti i vantaggi ottenibili e sopra esposti non si verificheranno.
Parliamo comunque di un integratore “compensativo” e “migliorativo” dello stato e compliance alimentare del utilizzatore.
Gli EEA non sostituiscono tutti i 21 AA; utilizzarli come substrato di sintesi per la matrice mancante è uno spreco di risorse e di soldi. Dovreste aver capito che è tutta una questione di equilibrio funzionale.
1. Bradford, H, Schmidt, V.. The history of the discovery of the amino acids. Chem Rev. 1931;9:169–864. doi: 10.1021/cr60033a001 [CrossRef] [Google Scholar]
2. Dietary reference intakes for energy, carbohydrates, fiber, fat, protein and amino acids (macronutrients). Washington, DC: Institute of Medicine of the National Academies, 2005. 2002/2005. Report No. [Google Scholar]
3. Elango, R, Ball, RO, Pencharz, PB. Recent advances in determining protein and amino acid requirements in humans. British Journal Of Nutrition. 2012;108(2):S22–30. Epub 2012/10/31. PubMed PMID: 23107531. doi: 10.1017/S0007114512002504 [PubMed] [CrossRef] [Google Scholar]
4. Food and Agriculture Organization of the United Nations Dietary protein quality evaluation in human nutrition. Rome: FAO Food and Nutrition. 2013. 31 March – 2 April 2011. Report No. [Google Scholar]
5. CE, B, JS, A, DT, H. Protein quality in humans: assessment and in vitro estimation. Westport Connecticut: AVI Publishing, Inc; 1981. [Google Scholar]
6. Bifari, F, Ruocco, C, Decimo, I, et al. Amino acid supplements and metabolic health: a potential interplay between intestinal microbiota and systems control. Genes Nutr. 2017;12:27. Epub 2017/10/19. PubMed PMID: 29043007; PMCID: PMC5628494. doi: 10.1186/s12263-017-0582-2 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Paddon-Jones, D, Sheffield-Moore, M, Urban, RJ, et al. Essential amino acid and carbohydrate supplementation ameliorates muscle protein loss in humans during 28 days bedrest. J Clin Endocrinol Metab. 2004;89(9):4351–4358. PubMed PMID: 15356032. doi: 10.1210/jc.2003-032159 [PubMed] [CrossRef] [Google Scholar]
8. Fitts, RH, Romatowski, JG, Peters, JR, et al. The deleterious effects of bed rest on human skeletal muscle fibers are exacerbated by hypercortisolemia and ameliorated by dietary supplementation. Am J Physiol Cell Physiol. 2007;293(1):C313–20. PubMed PMID: 17409123. doi: 10.1152/ajpcell.00573.2006 [PubMed] [CrossRef] [Google Scholar]
9. Biolo, G, Tipton, KD, Klein, S, et al. An abundant supply of amino acids enhances the metabolic effect of exercise on muscle protein. Am J Physiol. 1997;273(1 Pt 1):E122–9. PubMed PMID: 9252488. doi: 10.1152/ajpendo.1997.273.1.E122. [PubMed] [CrossRef] [Google Scholar]
10. Magaway, C, Kim, E, Jacinto, E. Targeting mTOR and metabolism in cancer: lessons and innovations. Cells. 2019;8(12). Epub 2019/12/11. PubMed PMID: 31817676; PMCID: PMC6952948. doi: 10.3390/cells8121584 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
15. Drummond, MJ, Rasmussen, BB. Leucine-enriched nutrients and the regulation of mammalian target of rapamycin signalling and human skeletal muscle protein synthesis. Current opinion in clinical nutrition and metabolic care. 2008;11(3):222–226. Epub 2008/04/12. PubMed PMID: 18403916; PMCID: PMC5096790. doi: 10.1097/MCO.0b013e3282fa17fb [PMC free article] [PubMed] [CrossRef] [Google Scholar]
16. Dreyer, HC, Fujita, S, Glynn, EL, et al. Resistance exercise increases leg muscle protein synthesis and mTOR signalling independent of sex. Acta Physiol (Oxf). 2010;199(1):71–81. Epub 2010/01/15. PubMed PMID: 20070283; PMCID: PMC2881180. doi: 10.1111/j.1748-1716.2010.02074.x [PMC free article] [PubMed] [CrossRef] [Google Scholar]
17. Maier, T, Guell, M, Serrano, L. Correlation of mRNA and protein in complex biological samples. FEBS Lett. 2009;583(24):3966–3973. Epub 2009/10/24. PubMed PMID: 19850042. doi: 10.1016/j.febslet.2009.10.036. [PubMed] [CrossRef] [Google Scholar]
18. Greenhaff, PL, Karagounis, LG, Peirce, N, et al. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am J Physiol. 2008;295(3):E595–604. PubMed PMID: 18577697. doi: 10.1152/ajpendo.90411.2008 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Crick, FH. On protein synthesis. Symp Soc Exp Biol. 1958;12:138–163. Epub 1958/01/01. PubMed PMID: 13580867. [PubMed] [Google Scholar]
20. Graber, TG, Borack, MS, Reidy, PT, et al. Essential amino acid ingestion alters expression of genes associated with amino acid sensing, transport, and mTORC1 regulation in human skeletal muscle. Nutri Metabol. 2017;14:35. Epub 2017/05/16. PubMed PMID: 28503190; PMCID: PMC5426042. doi: 10.1186/s12986-017-0187-1 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Wolfe, RR, Chinkes, D. Isotope tracers in metabolic research: principles and practice of kinetic analysis. New York, New Yor: Wiley-Liss; 2004. p. 274. [Google Scholar]
22. Biolo, G, Chinkes, D, Zhang, X, et al. Harry M. Vars research award: a new model to determine in vivo the relationship between amino acid transmembrane transport and protein kinetics in muscle. JPEN J Parenter Enteral Nutr. 1992;16(4):305–315. doi: 10.1177/0148607192016004305 [PubMed] [CrossRef] [Google Scholar]
23. Biolo, G, Fleming, RY, Maggi, SP, et al. Transmembrane transport and intracellular kinetics of amino acids in human skeletal muscle. Am J Physiol. 1995;268(1 Pt 1):E75–84. PubMed PMID: 7840186. doi: 10.1152/ajpendo.1995.268.1.E75. [PubMed] [CrossRef] [Google Scholar]
24. Tipton, KD, Borsheim, E, Wolf, SE, et al. Acute response of net muscle protein balance reflects 24-h balance after exercise and amino acid ingestion. Am J Physiol (Endocrinol Metab). 2003;284(1):E76–89. PubMed PMID: 12388164. doi: 10.1152/ajpendo.00234.2002. [PubMed] [CrossRef] [Google Scholar]
25. Plotkin, DL, Delcastillo, K, Van Every, DW, et al. Isolated leucine and branched-chain amino acid supplementation for enhancing muscular strength and hypertrophy: a narrative review. Int J Sport Nutr Exercise Metab. 2021;31(3):292–301. Epub 2021/03/21. PubMed PMID: 33741748. doi: 10.1123/ijsnem.2020-0356 [PubMed] [CrossRef] [Google Scholar]
26. Fuchs, CJ, Hermans, WJH, Holwerda, AM, et al. Branched-chain amino acid and branched-chain ketoacid ingestion increases muscle protein synthesis rates in vivo in older adults: a double-blind, randomized trial. Am J Clin Nutr. 2019;110(4):862–872. Epub 2019/06/30. PubMed PMID: 31250889; PMCID: PMC6766442. doi: 10.1093/ajcn/nqz120. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Pinals, RS, Harris, ED, Burnett, JB, et al. Treatment of rheumatoid arthritis with L-histidine: a randomized, placebo-controlled, double-blind trial. J Rheumatol. 1977;4(4):414–419. Epub 1977/01/01. PubMed PMID: 342692. [PubMed] [Google Scholar]
28. Jackman, SR, Witard, OC, Jeukendrup, AE, et al. Branched-chain amino acid ingestion can ameliorate soreness from eccentric exercise. Med & Sci In Sports & Ex. 2010;42(5):962–970. PubMed PMID: 19997002. doi: 10.1249/MSS.0b013e3181c1b798 [PubMed] [CrossRef] [Google Scholar]
29. Cynober, L, Bier, DM, Kadowaki, M, et al. A proposal for an upper limit of leucine safe intake in healthy adults. J Nutr. 2012;142(12):2249S–2250S. Epub 2012/10/26. PubMed PMID: 23096009. doi: 10.3945/jn.112.160853 [PubMed] [CrossRef] [Google Scholar]
30. Kim, IY, Williams, RH, Schutzler, SE, et al. Acute lysine supplementation does not improve hepatic or peripheral insulin sensitivity in older, overweight individuals. Nutri Metabol. 2014;11(1):49. Epub 2014/10/18. PubMed PMID: 25324894; PMCID: PMC4198625. doi: 10.1186/1743-7075-11-49 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Report of the expert advisory committee on amino acids. Ottawa, Canada: Minister of Supply and Services Canada, 1990. [Google Scholar]
32. Ryan-Harshman, M, Leiter, LA, Anderson, GH. Phenylalanine and aspartame fail to alter feeding behavior, mood and arousal in men. Physiol Behav. 1987;39(2):247–253. Epub 1987/01/01. PubMed PMID: 3575461. doi: 10.1016/0031-9384(87)90017-5. [PubMed] [CrossRef] [Google Scholar]
33. Growdon, JH, Nader, TM, Schoenfeld, J, et al. L-threonine in the treatment of spasticity. Clin Neuropharmacol. 1991;14(5):403–412. Epub 1991/10/01. PubMed PMID: 1742749. doi: 10.1097/00002826-199110000-00003 [PubMed] [CrossRef] [Google Scholar]
34. Cynober, L, Bier, DM, Kadowaki, M, et al. Proposals for upper limits of safe intake for arginine and tryptophan in young adults and an upper limit of safe intake for leucine in the elderly. J Nutr. 2016;146(12):2652S–2654S. Epub 2016/12/10. PubMed PMID: 27934658. doi: 10.3945/jn.115.228478 [PubMed] [CrossRef] [Google Scholar]
35. Volpi, E, Kobayashi, H, Sheffield-Moore, M, et al. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78(2):250–258. PubMed PMID: 12885705. doi: 10.1093/ajcn/78.2.250 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. Kobayashi, H, Borsheim, E, Anthony, TG, et al. Reduced amino acid availability inhibits muscle protein synthesis and decreases activity of initiation factor eIF2B. Am J Physiol. 2003;284(3):E488–98. PubMed PMID: 12556349. doi: 10.1152/ajpendo.00094.2002 [PubMed] [CrossRef] [Google Scholar]
37. Wilkinson, DJ, Bukhari, SSI, Phillips, BE, et al. Effects of leucine-enriched essential amino acid and whey protein bolus dosing upon skeletal muscle protein synthesis at rest and after exercise in older women. Clin Nutr. 2018;37(6 Pt A):2011–2021. Epub 2017/10/17. PubMed PMID: 29031484; PMCID: PMC6295981. doi: 10.1016/j.clnu.2017.09.008 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
38. Tipton, KD, Gurkin, BE, Matin, S, et al. Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. J Nutr Biochem. 1999;10(2):89–95. PubMed PMID: 15539275. doi: 10.1016/S0955-2863(98)00087-4. [PubMed] [CrossRef] [Google Scholar]
39. Paddon-Jones, D, Sheffield-Moore, M, Katsanos, CS, et al. Differential stimulation of muscle protein synthesis in elderly humans following isocaloric ingestion of amino acids or whey protein. Exp Gerontol. 2006;41(2):215–219. PubMed PMID: 16310330. doi: 10.1016/j.exger.2005.10.006 [PubMed] [CrossRef] [Google Scholar]
40. Kim, IY, Park, S, Smeets, E, et al. Consumption of a specially-formulated mixture of essential amino acids promotes gain in whole-body protein to a greater extent than a complete meal replacement in older women with heart failure. Nutrients. 2019;11(6). Epub 2019/06/20. PubMed PMID: 31212940; PMCID: PMC6627910. doi: 10.3390/nu11061360 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
41. Bukhari, SS, Phillips, BE, Wilkinson, DJ, et al. Intake of low-dose leucine-rich essential amino acids stimulates muscle anabolism equivalently to bolus whey protein in older women at rest and after exercise. Am J Physiol. 2015;308(12):E1056–65. Epub 2015/04/02. PubMed PMID: 25827594. doi: 10.1152/ajpendo.00481.2014 [PubMed] [CrossRef] [Google Scholar]
42. Park, S, Church, DD, Azhar, G, et al. Anabolic response to essential amino acid plus whey protein composition is greater than whey protein alone in young healthy adults. J Int Soc Sports Nutr. 2020;17(1):9. Epub 2020/02/12. PubMed PMID: 32041644; PMCID: PMC7011510. doi: 10.1186/s12970-020-0340-5 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
43. Church, DD, Hirsch, KR, Park, S, et al. Essential amino acids and protein synthesis: insights into maximizing the muscle and whole-body response to feeding. Nutrients. 2020;12(12). Epub 2020/12/06. PubMed PMID: 33276485; PMCID: PMC7760188. doi: 10.3390/nu12123717 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
44. Adibi, SA, Fogel, MR, Agrawal, RM. Comparison of free amino acid and dipeptide absorption in the jejunum of sprue patients. Gastroenterology. 1974;67(4):586–591. Epub 1974/10/01. PubMed PMID: 4412534. doi: 10.1016/S0016-5085(19)32783-0. [PubMed] [CrossRef] [Google Scholar]
45. Drummond, MJ, Glynn, EL, Fry, CS, et al. An increase in essential amino acid availability upregulates amino acid transporter expression in human skeletal muscle. Am J Physiol. 2010;298(5):E1011–8. Epub 2010/03/23. PubMed PMID: 20304764; PMCID: PMC2867366. doi: 10.1152/ajpendo.00690.2009 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
46. Bohe, J, Low, A, Wolfe, RR, et al. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: a dose-response study. Journal Of Physiology. 2003;552(Pt 1):315–324. PubMed PMID: 12909668. doi: 10.1113/jphysiol.2003.050674. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
47. Mitchell, WK, Phillips, BE, Williams, JP, et al. A dose- rather than delivery profile-dependent mechanism regulates the “muscle-full” effect in response to oral essential amino acid intake in young men. J Nutr. 2015;145(2):207–214. Epub 2015/02/04. PubMed PMID: 25644339; PMCID: PMC4304023. doi: 10.3945/jn.114.199604 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Anderson, GH, Moore, SE. Dietary proteins in the regulation of food intake and body weight in humans. J Nutr. 2004;134(4):974S–979S. PubMed PMID: 15051857. doi: 10.1093/jn/134.4.974S. [PubMed] [CrossRef] [Google Scholar]
49. Paddon-Jones, D, Sheffield-Moore, M, Aarsland, A, et al. Exogenous amino acids stimulate human muscle anabolism without interfering with the response to mixed meal ingestion. Am J Physiol. 2005;288(4):E761–7. PubMed PMID: 15572657. doi: 10.1152/ajpendo.00291.2004 [PubMed] [CrossRef] [Google Scholar]
50. Abdulla, H, Bass, JJ, Stokes, T, et al. The effect of oral essential amino acids on incretin hormone production in youth and ageing. Endocrinol Diabetes Metab. 2019;2(4):e00085. Epub 2019/10/09. PubMed PMID: 31592446; PMCID: PMC6775449. doi: 10.1002/edm2.85 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
51. Park, S, Jang, J, Choi, MD, et al. The anabolic response to dietary protein is not limited by the maximal stimulation of protein synthesis in healthy older adults: a randomized crossover trial. Nutrients. 2020;12(11). Epub 2020/10/30. PubMed PMID: 33114585; PMCID: PMC7693481. doi: 10.3390/nu12113276 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
52. Fry, CS, Drummond, MJ, Glynn, EL, et al. Aging impairs contraction-induced human skeletal muscle mTORC1 signaling and protein synthesis. Skelet Muscle. 2011;1(1):11. Epub 2011/07/30. PubMed PMID: 21798089; PMCID: PMC3156634. doi: 10.1186/2044-5040-1-11 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
53. Katsanos, CS, Kobayashi, H, Sheffield-Moore, M, et al. Aging is associated with diminished accretion of muscle proteins after the ingestion of a small bolus of essential amino acids. Am J Clin Nutr. 2005;82(5):1065–1073. PubMed PMID: 16280440. doi: 10.1093/ajcn/82.5.1065 [PubMed] [CrossRef] [Google Scholar]
54. Paddon-Jones, D, Sheffield-Moore, M, Zhang, XJ, et al. Amino acid ingestion improves muscle protein synthesis in the young and elderly. Am J Physiol. 2004;286(3):E321–8. PubMed PMID: 14583440. doi: 10.1152/ajpendo.00368.2003 [PubMed] [CrossRef] [Google Scholar]
55. Katsanos, CS, Kobayashi, H, Sheffield-Moore, M, et al. A high proportion of leucine is required for optimal stimulation of the rate of muscle protein synthesis by essential amino acids in the elderly. Am J Physiol. 2006;291(2):E381–7. PubMed PMID: 16507602. doi: 10.1152/ajpendo.00488.2005 [PubMed] [CrossRef] [Google Scholar]
56. Carbone, JW, McClung, JP, Pasiakos, SM. Skeletal muscle responses to negative energy balance: effects of dietary protein. Adv nutri (Bethesda, Md). 2012;3(2):119–126. Epub 2012/04/21. PubMed PMID: 22516719; PMCID: PMC3648712 conflicts of interest. doi: 10.3945/an.111.001792 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
57. Trappe, TA, Gastaldelli, A, Jozsi, AC, et al. Energy expenditure of swimmers during high volume training. Medi and sci sports and exerci. 1997;29(7):950–954. Epub 1997/07/01. PubMed PMID: 9243495. doi: 10.1097/00005768-199707000-00015 [PubMed] [CrossRef] [Google Scholar]
58. Gwin, JA, Church, DD, Hatch-McChesney, A, et al. Effects of high versus standard essential amino acid intakes on whole-body protein turnover and mixed muscle protein synthesis during energy deficit: a randomized, crossover study. Clin Nutr. 2021;40(3):767–777. Epub 2020/08/10. PubMed PMID: 32768315. doi: 10.1016/j.clnu.2020.07.019 [PubMed] [CrossRef] [Google Scholar]
59. Biolo, G, Fleming, RY, Maggi, SP, et al. Inverse regulation of protein turnover and amino acid transport in skeletal muscle of hypercatabolic patients. J Clin Endocrinol Metab. 2002;87(7):3378–3384. PubMed PMID: 12107253. doi: 10.1210/jcem.87.7.8699 [PubMed] [CrossRef] [Google Scholar]
60. Dillon, EL, Sheffield-Moore, M, Paddon-Jones, D, et al. Amino acid supplementation increases lean body mass, basal muscle protein synthesis, and insulin-like growth factor-I expression in older women. J Clin Endocrinol Metab. 2009;94(5):1630–1637. PubMed PMID: 19208731. doi: 10.1210/jc.2008-1564 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Borsheim, E, Bui, QU, Tissier, S, et al. Effect of amino acid supplementation on muscle mass, strength and physical function in elderly. Clin Nutr. 2008;27(2):189–195. PubMed PMID: 18294740. doi: 10.1016/j.clnu.2008.01.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
62. Kim, HK, Suzuki, T, Saito, K, et al. Effects of exercise and amino acid supplementation on body composition and physical function in community-dwelling elderly Japanese sarcopenic women: a randomized controlled trial. J Am Geriatr Soc. 2012;60(1):16–23. PubMed PMID: 22142410. doi: 10.1111/j.1532-5415.2011.03776.x [PubMed] [CrossRef] [Google Scholar]
63. Azhar, G, Wei, JY, Schutzler, SE, et al. Daily consumption of a specially formulated essential amino acid-based dietary supplement improves physical performance in older adults with low physical functioning. J Gerontol A Biol Sci Med Sci. 2021;76(7):1184–1191. Epub 2021/01/22. PubMed PMID: 33475727; PMCID: PMC8202157. doi: 10.1093/gerona/glab019 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
64. Hwang, CL, Chien, CL, Wu, YT. Resistance training increases 6-minute walk distance in people with chronic heart failure: a systematic review. J Physiother. 2010;56(2):87–96. Epub 2010/05/21. PubMed PMID: 20482475. doi: 10.1016/s1836-9553(10)70038-2. [PubMed] [CrossRef] [Google Scholar]
65. Ferrando, AA, Paddon-Jones, D, Hays, NP, et al. EAA supplementation to increase nitrogen intake improves muscle function during bed rest in the elderly. Clin Nutr. 2010;29:18–23. PubMed PMID: 19419806. doi: 10.1016/j.clnu.2009.03.009. [PubMed] [CrossRef] [Google Scholar]
66. Biolo, G, Maggi, SP, Williams, BD, et al. Increased rates of muscle protein turnover and amino acid transport after resistance exercise in humans. Am J Physiol. 1995;268(3 Pt 1):E514–20. PubMed PMID: 7900797. doi: 10.1152/ajpendo.1995.268.3.E514. [PubMed] [CrossRef] [Google Scholar]
67. Tipton, KD, Ferrando, AA, Phillips, SM, et al. Postexercise net protein synthesis in human muscle from orally administered amino acids. Am J Physiol. 1999;276(4Pt1):E628–34. PubMed PMID: 10198297. doi: 10.1152/ajpendo.1999.276.4.E628. [PubMed] [CrossRef] [Google Scholar]
68. Rasmussen, BB, Tipton, KD, Miller, SL, et al. An oral essential amino acid-carbohydrate supplement enhances muscle protein anabolism after resistance exercise. J Appl Physiol. 2000;88(2):386–392. PubMed PMID: 10658002. doi: 10.1152/jappl.2000.88.2.386 [PubMed] [CrossRef] [Google Scholar]
69. Waskiw-Ford, M, Hannaian, S, Duncan, J, et al. Leucine-enriched essential amino acids improve recovery from post-exercise muscle damage independent of increases in integrated myofibrillar protein synthesis in young men. Nutrients. 2020;12(4). Epub 2020/04/16. PubMed PMID: 32290521; PMCID: PMC7231404. doi: 10.3390/nu12041061 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
70. Moberg, M, Apro, W, Ekblom, B, et al. Activation of mTORC1 by leucine is potentiated by branched-chain amino acids and even more so by essential amino acids following resistance exercise. Am J Physiol Cell Physiol. 2016;310(11):C874–84. Epub 2016/04/08. PubMed PMID: 27053525. doi: 10.1152/ajpcell.00374.2015 [PubMed] [CrossRef] [Google Scholar]
71. Hannaian, SJ, Hodson, N, Abou Sawan, S, et al. Leucine-enriched amino acids maintain peripheral mTOR-Rheb localization independent of myofibrillar protein synthesis and mTORC1 signaling postexercise. J Appl Physiol. 2020;129(1):133–143. Epub 2020/06/12. PubMed PMID: 32525432; PMCID: PMC7469228. doi: 10.1152/japplphysiol.00241.2020 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
72. Lees, MJ, Wilson, OJ, Webb, EK, et al. Novel essential amino acid supplements following resistance exercise induce aminoacidemia and enhance anabolic signaling irrespective of age: a proof-of-concept trial. Nutrients. 2020;12(7). Epub 2020/07/16. PubMed PMID: 32664648; PMCID: PMC7400893. doi: 10.3390/nu12072067 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Tipton, KD, Rasmussen, BB, Miller, SL, et al. Timing of amino acid-carbohydrate ingestion alters anabolic response of muscle to resistance exercise. Am J Physiol. 2001;281(2):E197–206. PubMed PMID: 11440894. doi: 10.1152/ajpendo.2001.281.2.E197 [PubMed] [CrossRef] [Google Scholar]
75. Fujita, S, Dreyer, HC, Drummond, MJ, et al. Essential amino acid and carbohydrate ingestion before resistance exercise does not enhance postexercise muscle protein synthesis. J Appl Physiol. 2009;106(5):1730–1739. Epub 2017/06/10. PubMed PMID: 18535123; PMCID: PMC2681328. doi: 10.1152/japplphysiol.90395.2008 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
76. Borsheim, E, Tipton, KD, Wolf, SE, et al. Essential amino acids and muscle protein recovery from resistance exercise. Am J Physiol. 2002;283(4):E648–57. PubMed PMID: 12217881. doi: 10.1152/ajpendo.00466.2001 [PubMed] [CrossRef] [Google Scholar]
77. Pasiakos, SM, McClung, HL, Margolis, LM, et al. Human muscle protein synthetic responses during weight-bearing and non-weight-bearing exercise: a comparative study of exercise modes and recovery nutrition. Plos One. 2015;10(10):e0140863. PubMed PMID: 26474292; PMCID: PMC4608805. doi: 10.1371/journal.pone.0140863. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
78. Markofski, MM, Jennings, K, Timmerman, KL, et al. Effect of aerobic exercise training and essential amino acid supplementation for 24 weeks on physical function, body composition, and muscle metabolism in healthy, independent older adults: a randomized clinical trial. J Gerontol A Biol Sci Med Sci. 2019;74(10):1598–1604. Epub 2018/05/12. PubMed PMID: 29750251; PMCID: PMC6748753. doi: 10.1093/gerona/gly109 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
79. Callahan, MJ, Parr, EB, Hawley, JA, et al. Can high-intensity interval training promote skeletal muscle anabolism? Sports Med (Auckland, NZ). 2021;51(3):405–421. Epub 2017/10/17. PubMed PMID: 33512698. doi: 10.1007/s40279-020-01397-3. [PubMed] [CrossRef] [Google Scholar]
80. Forbes, SC, Candow, DG, Smith-Ryan, AE, et al. Supplements and nutritional interventions to augment high-intensity interval training physiological and performance adaptations—A narrative review. Nutrients. 2020;12(2):390. Epub 2020/02/07. PubMed PMID: 32024038; PMCID: PMC7071320. doi: 10.3390/nu12020390. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
81. Hirsch, KR, Greenwalt, CE, Saylor, HE, et al. High-intensity interval training and essential amino acid supplementation: effects on muscle characteristics and whole-body protein turnover. Physiol Rep. 2021;9(1):e14655. Epub 2020/12/29. PubMed PMID: 33369879; PMCID: PMC7769174. doi: 10.14814/phy2.14655 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
82. Hirsch, KR, Greenwalt, CE, Cabre, HE, et al. Metabolic effects of high-intensity interval training and essential amino acids. Eur J Appl Physiol. 2021;121(12):3297–3311. Epub 2020/12/29. PubMed PMID: 34427732. doi: 10.1007/s00421-021-04792-4 [PubMed] [CrossRef] [Google Scholar]
83. Hirsch, KR, Cabre, HE, Gould, LM, et al. Effects of essential amino acids on high-intensity interval training performance, fatigue outcomes, and workload progression. J Am Nutr Assoc. 2023;42(4):411–417. Epub 2020/12/29. PubMed PMID: 35512775. doi: 10.1080/07315724.2022.2060373 [PubMed] [CrossRef] [Google Scholar]
84. Sheffield-Moore, M, Yeckel, CW, Volpi, E, et al. Postexercise protein metabolism in older and younger men following moderate-intensity aerobic exercise. Am J Physiol. 2004;287(3):E513–22. Epub 2004/05/20. PubMed PMID: 15149953. doi: 10.1152/ajpendo.00334.2003 [PubMed] [CrossRef] [Google Scholar]
85. Aquilani, R, D’Antona, G, Baiardi, P, et al. Essential amino acids and exercise tolerance in elderly muscle-depleted subjects with chronic diseases: a rehabilitation without rehabilitation? Biomed Res Int. 2014;2014:341603. Epub 2014/07/11. PubMed PMID: 25009815; PMCID: PMC4070286. doi: 10.1155/2014/341603 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
86. Aquilani, R, Zuccarelli, GC, Dioguardi, FS, et al. Effects of oral amino acid supplementation on long-term-care-acquired infections in elderly patients. Arch Gerontol Geriatr. 2011;52(3):e123–8. Epub 2010/10/12. PubMed PMID: 20934757. doi: 10.1016/j.archger.2010.09.005 [PubMed] [CrossRef] [Google Scholar]
87. Aquilani, R, Viglio, S, Iadarola, P, et al. Oral amino acid supplements improve exercise capacities in elderly patients with chronic heart failure. Am J Cardiol. 2008;101(11A):104E–110E. Epub 2008/07/02. PubMed PMID: 18514618. doi: 10.1016/j.amjcard.2008.03.008 [PubMed] [CrossRef] [Google Scholar]
88. Scognamiglio, R, Piccolotto, R, Negut, C, et al. Oral amino acids in elderly subjects: effect on myocardial function and walking capacity. Gerontology. 2005;51(5):302–308. Epub 2005/08/20. PubMed PMID: 16110231. doi: 10.1159/000086366 [PubMed] [CrossRef] [Google Scholar]
89. Aquilani, R, Zuccarelli, GC, Condino, AM, et al. Despite inflammation, supplemented essential amino acids may improve circulating levels of albumin and haemoglobin in patients after hip fractures. Nutrients. 2017;9(6). Epub 2017/06/22. PubMed PMID: 28635634; PMCID: PMC5490616. doi: 10.3390/nu9060637 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
90. Aquilani, R, Zuccarelli Ginetto, C, Rutili, C, et al. Supplemented amino acids may enhance the walking recovery of elderly subjects after hip fracture surgery. Aging Clin Exp Res. 2019;31(1):157–160. Epub 2018/04/19. PubMed PMID: 29667153. doi: 10.1007/s40520-018-0941-x [PubMed] [CrossRef] [Google Scholar]
91. Baldissarro, E, Aquilani, R, Boschi, F, et al. The hip functional retrieval after elective surgery May be enhanced by supplemented essential amino acids. Biomed Res Int. 2016;2016:9318329. Epub 2016/04/26. PubMed PMID: 27110573; PMCID: PMC4823478. doi: 10.1155/2016/9318329 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
92. Ferrando, A, Bamman, MM, Schutzler, S, et al. Increased nitrogen intake following hip arthroplasty expedites muscle strength recovery. J Aging: Res And Clin Pract. 2013;2(4):369. [Google Scholar]
93. Aquilani, R, Boselli, M, D’Antona, G, et al. Unaffected arm muscle hypercatabolism in dysphagic subacute stroke patients: the effects of essential amino acid supplementation. Biomed Res Int. 2014;2014:964365. Epub 2016/04/26. PubMed PMID: 25431770; PMCID: PMC4241696. doi: 10.1155/2014/964365 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
94. Aquilani, R, Emilio, B, Dossena, M, et al. Correlation of deglutition in subacute ischemic stroke patients with peripheral blood adaptive immunity: essential amino acid improvement. Int J Immunopathol Pharmacol. 2015;28(4):576–583. Epub 2015/10/07. PubMed PMID: 26437899. doi: 10.1177/0394632015608249 [PubMed] [CrossRef] [Google Scholar]
95. Brooks, N, Cloutier, GJ, Cadena, SM, et al. Resistance training and timed essential amino acids protect against the loss of muscle mass and strength during 28 days of bed rest and energy deficit. J Appl Physiol. 2008;105(1):241–248. Epub 2008/05/17. PubMed PMID: 18483167; PMCID: PMC2494840. doi: 10.1152/japplphysiol.01346.2007 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Cholewa, JM, Dardevet, D, Lima-Soares, F, et al. Dietary proteins and amino acids in the control of the muscle mass during immobilization and aging: role of the MPS response. Amino Acids. 2017;49(5):811–820. Epub 2017/02/09. PubMed PMID: 28175999. doi: 10.1007/s00726-017-2390-9 [PubMed] [CrossRef] [Google Scholar]
97. Holloway, TM, McGlory, C, McKellar, S, et al. A novel amino acid composition ameliorates short-term muscle disuse atrophy in healthy young men. Front Nutr. 2019;6:105. Epub 2019/07/30. PubMed PMID: 31355205; PMCID: PMC6636393. doi: 10.3389/fnut.2019.00105 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
98. Killewich, LA, Tuvdendorj, D, Bahadorani, J, et al. Amino acids stimulate leg muscle protein synthesis in peripheral arterial disease. J Vasc Surg. 2007;45(3):554–559. discussion 9-60. Epub 2007/02/27. PubMed PMID: 17321342. doi: 10.1016/j.jvs.2006.11.033 [PubMed] [CrossRef] [Google Scholar]
99. Bauerdick, H, Spellerberg, P, Lamberts, B. Therapy with essential amino acids and their nitrogen-free analogues in severe renal failure. Am J Clin Nutr. 1978;31(10):1793–1796. Epub 1978/10/01. PubMed PMID: 707334. doi: 10.1093/ajcn/31.10.1793 [PubMed] [CrossRef] [Google Scholar]
101. Ferrando, AA, Raj, D, Wolfe, RR. Amino acid control of muscle protein turnover in renal disease. J Ren Nutr. 2005;15(1):34–38. PubMed PMID: 15648004. doi: 10.1053/j.jrn.2004.09.014. [PubMed] [CrossRef] [Google Scholar]
102. Hecking, E, Kohler, H, Zobel, R, et al. Treatment with essential amino acids in patients on chronic hemodialysis: a double blind cross-over study. Am J Clin Nutr. 1978;31(10):1821–1826. Epub 1978/10/01. PubMed PMID: 360819. doi: 10.1093/ajcn/31.10.1821 [PubMed] [CrossRef] [Google Scholar]
103. Unverdi, S, Ceri, M, Uz, E, et al. The effectiveness of oral essential aminoacids and aminoacids containing dialysate in peritoneal dialysis. Ren Fail. 2014;36(9):1416–1419. Epub 2014/09/24. PubMed PMID: 25246343. doi: 10.3109/0886022X.2014.950933 [PubMed] [CrossRef] [Google Scholar]
104. Kato, H, Miura, K, Nakano, S, et al. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids. 2016;48(9):2145–2155. Epub 2016/05/12. PubMed PMID: 27168073; PMCID: PMC4989025. doi: 10.1007/s00726-016-2240-1 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
105. Rittig, N, Bach, E, Thomsen, HH, et al. Amino acid supplementation is anabolic during the acute phase of endotoxin-induced inflammation: a human randomized crossover trial. Clin Nutr. 2016;35(2):322–330. Epub 2015/04/22. PubMed PMID: 25896101. doi: 10.1016/j.clnu.2015.03.021. [PubMed] [CrossRef] [Google Scholar]
106. Jones, C, Eddleston, J, McCairn, A, et al. Improving rehabilitation after critical illness through outpatient physiotherapy classes and essential amino acid supplement: a randomized controlled trial. J Crit Care. 2015;30(5):901–907. Epub 2016/05/12. PubMed PMID: 26004031. doi: 10.1016/j.jcrc.2015.05.002 [PubMed] [CrossRef] [Google Scholar]
107. Engelen, M, Safar, AM, Bartter, T, et al. High anabolic potential of essential amino acid mixtures in advanced nonsmall cell lung cancer. Ann Oncol. 2015;26(9):1960–1966. Epub 2015/06/27. PubMed PMID: 26113648; PMCID: PMC4551158. doi: 10.1093/annonc/mdv271 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
109. Dal Negro, RW, Aquilani, R, Bertacco, S, et al. Comprehensive effects of supplemented essential amino acids in patients with severe COPD and sarcopenia. Monaldi Arch Chest Dis. 2010;73(1):25–33. Epub 2010/05/27. PubMed PMID: 20499791. doi: 10.4081/monaldi.2010.310 [PubMed] [CrossRef] [Google Scholar]
110. Dal Negro, RW, Testa, A, Aquilani, R, et al. Essential amino acid supplementation in patients with severe COPD: a step towards home rehabilitation. Monaldi Arch Chest Dis. 2012;77(2):67–75. Epub 2012/12/01. PubMed PMID: 23193843. doi: 10.4081/monaldi.2012.154 [PubMed] [CrossRef] [Google Scholar]
111. Jonker, R, Deutz, NE, Erbland, ML, et al. Effectiveness of essential amino acid supplementation in stimulating whole body net protein anabolism is comparable between COPD patients and healthy older adults. Metabolism. 2017;69:120–129. Epub 2017/03/14. PubMed PMID: 28285641; PMCID: PMC5351771. doi: 10.1016/j.metabol.2016.12.010 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
112. Corsetti, G, Romano, C, Pasini, E, et al. Diet enrichment with a specific essential free amino acid mixture improves healing of undressed wounds in aged rats. Exp Gerontol. 2017;96:138–145. Epub 2017/07/04. PubMed PMID: 28669821. doi: 10.1016/j.exger.2017.06.020 [PubMed] [CrossRef] [Google Scholar]
113. Borsheim, E, Bui, QU, Wolfe, RR. Plasma amino acid concentrations during late rehabilitation in patients with traumatic brain injury. Archives of physical medicine and rehabilitation. Arch Phys Med Rehabil. 2007;88(2):234–238. Epub 2007/02/03. PubMed PMID: 17270522. doi: 10.1016/j.apmr.2006.11.003. [PubMed] [CrossRef] [Google Scholar]
114. Boselli, M, Aquilani, R, Baiardi, P, et al. Supplementation of essential amino acids may reduce the occurrence of infections in rehabilitation patients with brain injury. Nutr Clin Pract. 2012;27(1):99–113. Epub 2012/02/07. PubMed PMID: 22307494. doi: 10.1177/0884533611431068 [PubMed] [CrossRef] [Google Scholar]
115. Borsheim, E, Bui, QU, Tissier, S, et al. Amino acid supplementation decreases plasma and liver triacylglycerols in elderly. Nutrition. 2009;25:281–288. PubMed PMID: 19041223. doi: 10.1016/j.nut.2008.09.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
116. Coker, RH, Deutz, NE, Schutzler, S, et al. Nutritional supplementation with essential amino acids and phytosterols may reduce risk for metabolic syndrome and cardiovascular disease in overweight individuals with Mild Hyperlipidemia. J Endocrinol Diabetes Obes. 2015;3(2):Epub 2016/01/05. PubMed PMID: 26726312; PMCID: PMC4696774. [PMC free article] [PubMed] [Google Scholar]
117. Marquis, BJ, Hurren, NM, Carvalho, E, et al. Skeletal muscle acute and chronic metabolic response to essential amino acid supplementation in hypertriglyceridemic older adults. Curr Dev Nutr. 2017;1(11):e002071. Epub 2018/06/30. PubMed PMID: 29955688; PMCID: PMC5998789. doi: 10.3945/cdn.117.002071 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
118. Coker, RH, Miller, S, Schutzler, S, et al. Whey protein and essential amino acids promote the reduction of adipose tissue and increased muscle protein synthesis during caloric restriction-induced weight loss in elderly, obese individuals. Nutr J. 2012;11:105. Epub 2018/06/30. PubMed PMID: 23231757; PMCID: PMC3546025. doi: 10.1186/1475-2891-11-105 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
119. Coker, RH, Shin, K, Scholten, K, et al. Essential amino acid-enriched meal replacement promotes superior net protein balance in older, overweight adults. Clin Nutr. 2019;38(6):2821–2826. Epub 2015/04/22. PubMed PMID: 30638738; PMCID: PMC6588419. doi: 10.1016/j.clnu.2018.12.013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
121. Jegatheesan, P, Beutheu, S, Ventura, G, et al. Effect of specific amino acids on hepatic lipid metabolism in fructose-induced non-alcoholic fatty liver disease. Clin Nutr. 2016;35(1):175–182. Epub 2015/03/05. PubMed PMID: 25736031. doi: 10.1016/j.clnu.2015.01.021 [PubMed] [CrossRef] [Google Scholar]
122. Theytaz, F, Noguchi, Y, Egli, L, et al. Effects of supplementation with essential amino acids on intrahepatic lipid concentrations during fructose overfeeding in humans. Am J Clin Nutr. 2012;96(5):1008–1016. Epub 2012/10/05. PubMed PMID: 23034968. doi: 10.3945/ajcn.112.035139 [PubMed] [CrossRef] [Google Scholar]
123. Brocca, L, D’Antona, G, Bachi, A, et al. Amino acid supplements improve native antioxidant enzyme expression in the skeletal muscle of diabetic mice. Am J Cardiol. 2008;101(11A):57E–62E. Epub 2008/07/02. PubMed PMID: 18514628. doi: 10.1016/j.amjcard.2008.03.002 [PubMed] [CrossRef] [Google Scholar]
124. Natarajan Sulochana, K, Lakshmi, S, Punitham, R, et al. Effect of oral supplementation of free amino acids in type 2 diabetic patients– a pilot clinical trial. Med Sci Monit. 2002;8(3): CR131–7. Epub 2002/03/12. PubMed PMID: 11887024. [PubMed] [Google Scholar]
125. Pellegrino, MA, Patrini, C, Pasini, E, et al. Amino acid supplementation counteracts metabolic and functional damage in the diabetic rat heart. Am J Cardiol. 2008;101(11A):49E–56E. Epub 2008/07/02. PubMed PMID: 18514627. doi: 10.1016/j.amjcard.2008.03.001 [PubMed] [CrossRef] [Google Scholar]
126. Solerte, SB, Fioravanti, M, Locatelli, E, et al. Improvement of blood glucose control and insulin sensitivity during a long-term (60 weeks) randomized study with amino acid dietary supplements in elderly subjects with type 2 diabetes mellitus. Am J Cardiol. 2008;101(11A):82E–88E. Epub 2008/07/02. PubMed PMID: 18514633. doi: 10.1016/j.amjcard.2008.03.006 [PubMed] [CrossRef] [Google Scholar]
127. Solerte, SB, Gazzaruso, C, Schifino, N, et al. Metabolic effects of orally administered amino acid mixture in elderly subjects with poorly controlled type 2 diabetes mellitus. Am J Cardiol. 2004;93(8A):23A–29A. Epub 2008/07/02. PubMed PMID: 15094102. doi: 10.1016/j.amjcard.2003.11.006 [PubMed] [CrossRef] [Google Scholar]
Nella 3° parte abbiamo analizzato le caratteristiche e funzioni biochimiche della L-Citrullina e della L-Arginina. In questa quarta parte, invece, andremo ad analizzare uno degli integratori a base di AA più discussi in assoluto: gli Aminoacidi a Catena Ramificata/BCAA.
BCAA – caratteristiche biochimiche:
Un aminoacido a catena ramificata (BCAA) è un aminoacido con una catena laterale alifatica con un ramo (un atomo di carbonio centrale legato a tre o più atomi di carbonio). Tra gli aminoacidi proteinogenici, vi sono tre BCAA: leucina, isoleucina e valina.[1] Tra i BCAA non proteinogenici vi sono l’acido 2-aminoisobutirrico e l’alloisoleucina.
I tre BCAA proteinogenici sono tra i nove aminoacidi essenziali per l’uomo, rappresentando il 35% degli aminoacidi essenziali nelle proteine muscolari e il 40% degli aminoacidi preformati richiesti dai mammiferi.[2] La sintesi dei BCAA avviene in tutti i luoghi delle piante, all’interno dei plastidi della cellula, come determinato dalla presenza di mRNA che codificano per gli enzimi della via metabolica.[3][4][5] L’ossidazione dei BCAA può aumentare l’ossidazione degli acidi grassi e svolgere un ruolo nell’obesità. Fisiologicamente, i BCAA svolgono un ruolo nel sistema immunitario e nella funzione cerebrale. I BCAA vengono scomposti efficacemente dagli enzimi deidrogenasi e decarbossilasi espressi dalle cellule immunitarie e sono necessari per la crescita e la proliferazione dei linfociti e per l’attività dei linfociti T citotossici.[4] Infine, i BCAA condividono con gli aminoacidi aromatici (Trp, Tyr e Phe) la stessa proteina di trasporto nel cervello. Una volta nel cervello, i BCAA possono avere un ruolo nella sintesi proteica, nella sintesi dei neurotrasmettitori e nella produzione di energia.[4]
Aspartato transaminasi [aminotrasferasi] da E. coli con cofattore Piridossal 5′ Fosfato
Cinque enzimi partecipano alle vie di sintesi parallele di isoleucina, valina e leucina: la treonina deidrogenasi, l’acetoidrossiacido sintasi, la chetoacido reduttasi, la diidrossiacido deidrogenasi e l’aminotransferasi.[3] La treonina deidrogenasi catalizza la deaminazione e la disidratazione della treonina a 2-chetobutirrato e ammoniaca. L’isoleucina forma un circuito di feedback negativo con la treonina deidrogenasi. L’acetoidrossiacido sintasi è il primo enzima della via parallela che esegue la reazione di condensazione in entrambe le fasi: condensazione del piruvato ad acetolattato nella via della valina e condensazione del piruvato e del 2-chetobutirrato per formare acetoidrossibutirrato nella via dell’isoleucina. Successivamente, la chetoacido-riduttasi riduce gli acetoidrossiacidi della fase precedente per produrre diidrossiacidi in entrambe le vie della valina e dell’isoleucina. La diidrossiacido deidrogenasi converte i diidrossiacidi nella fase successiva. La fase finale della via parallela è condotta dall’amino-transferasi, che produce i prodotti finali di valina e isoleucina.[3] Una serie di altri quattro enzimi – isopropilmalato sintasi, isopropilmalato isomerasi, isopropilmalato deidrogenasi e aminotransferasi – è necessaria per la formazione della leucina dal 2-ossolsovalerato.[3]
La degradazione degli aminoacidi a catena ramificata coinvolge il complesso della deidrogenasi degli alfa-chetoacidi a catena ramificata (BCKDH). Una carenza di questo complesso porta a un accumulo di aminoacidi a catena ramificata (leucina, isoleucina e valina) e dei loro sottoprodotti tossici nel sangue e nelle urine, dando alla condizione il nome di malattia delle urine a sciroppo d’acero. D’altra parte, l’attività incontrollata di questo complesso causa un deficit di chetoacido deidrogenasi a catena ramificata.
La degradazione di leucina, isoleucina e valina. Viene illustrata anche la via di degradazione della metionina.
Il complesso BCKDH converte gli aminoacidi a catena ramificata in derivati dell’acil-CoA, che dopo successive reazioni vengono convertiti in acetil-CoA o succinil-CoA che entrano nel ciclo dell’acido citrico.[6]
Gli enzimi coinvolti sono l’aminotransferasi a catena ramificata e la 3-metil-2-ossobutanoato deidrogenasi.
Mentre la maggior parte degli aminoacidi viene ossidata nel fegato, i BCAA vengono ossidati principalmente nel muscolo scheletrico e in altri tessuti periferici.[4] Sono stati testati gli effetti della somministrazione di BCAA sulla crescita muscolare del diaframma di ratto e si è concluso che non solo una miscela di BCAA da sola ha lo stesso effetto sulla crescita di una miscela completa di aminoacidi, ma che una miscela di aminoacidi con tutti i BCAA tranne i BCAA non influisce sulla crescita muscolare del diaframma di ratto. [7] La somministrazione di isoleucina o valina da sole non ha influenzato la crescita muscolare, anche se la somministrazione di leucina da sola sembra essere quasi altrettanto efficace della miscela completa di BCAA. La leucina attiva indirettamente la p70 S6 chinasi e stimola l’assemblaggio del complesso eIF4F, essenziali per il legame dell’mRNA nell’avvio della traslazione.[7] La p70 S6 chinasi fa parte della via di segnalazione del complesso mammalian target of rapamycin (mTOR) ed è stato dimostrato che consente l’ipertrofia adattiva e il recupero del muscolo di ratto. [8] A riposo, l’infusione di proteine stimola la sintesi proteica 30 minuti dopo l’inizio dell’infusione e la sintesi proteica rimane elevata per altri 90 minuti.[9] L’infusione di leucina a riposo produce un effetto stimolante di sei ore e un aumento della sintesi proteica attraverso la fosforilazione della p70 S6 chinasi nei muscoli scheletrici.[10] Dopo l’esercizio di resistenza, senza la somministrazione di BCAA, una sessione di esercizio di resistenza non influisce sulla fosforilazione di mTOR e produce addirittura una diminuzione della fosforilazione di Akt. È stata rilevata una certa fosforilazione della p70 S6 chinasi. Quando sono stati somministrati BCAA dopo una sessione di allenamento, una fosforilazione sufficiente di p70 S6 chinasi e S6 ha indicato l’attivazione della cascata di segnalazione.[10]
Metabolismo degli aminoacidi a catena ramificata nel muscolo scheletrico. (1) I BCAA sono transaminati con α-chetoglutarato dalla BCAA transaminasi per generare glutammato. (2) La deaminazione del glutammato produce α-chetoglutarato e ammoniaca. (3) Durante l’esercizio fisico, l’AMP viene generato dalla degradazione dell’ATP nel muscolo scheletrico. La reazione dell’AMP deaminasi muscolare forma anche ammoniaca. (4) La glutammina si forma da ammoniaca e glutammato, una reazione catalizzata dalla glutammina sintetasi. (5) L’α-chetoglutarato formato dalla glutammato deidrogenasi può entrare anapleroticamente nel ciclo TCA. Tratto e modificato da Groper e Smith (2013).
Oltre alla segnalazione cellulare, la via mTOR svolge anche un ruolo nella crescita delle cellule beta che porta alla secrezione di insulina.[11] L’elevata presenza di glucosio nel sangue avvia il processo della via di segnalazione mTOR, in cui la leucina svolge un ruolo indiretto.[9][12] La combinazione di glucosio, leucina e altri attivatori fa sì che mTOR inizi a segnalare la proliferazione delle cellule beta e la secrezione di insulina. Concentrazioni più elevate di leucina causano un’iperattività della via mTOR e l’attivazione della chinasi S6 porta all’inibizione del substrato del recettore dell’insulina attraverso la fosforilazione della serina.[11][12] Nella cellula l’aumento dell’attività del complesso mTOR causa l’eventuale incapacità delle cellule beta di rilasciare insulina e l’effetto inibitorio della chinasi S6 porta all’insulino-resistenza nelle cellule, contribuendo allo sviluppo del diabete di tipo 2.[12]
La metformina è in grado di attivare l’AMP chinasi che fosforila le proteine coinvolte nella via mTOR e porta alla progressione del complesso mTOR dallo stato inattivo a quello attivo.[12] Si suggerisce che la Metformina agisca come inibitore competitivo dell’amminoacido leucina nella via mTOR.
BCAA e Sport:
Sappiamo che la Leucina, in particolare, attiva mTOR, un segnale anabolico che media la sintesi proteica muscolare [13], a sua volta correlata agli adattamenti della forza e dell’ipertrofia [14]. A tal fine, si è ipotizzato che i BCAA siano utili per le prestazioni, il recupero e la composizione corporea [15].
Una recente pubblicazione sostiene che l’assunzione di livelli accettabili di BCAA dovrebbe essere prioritaria rispetto alla partecipazione all’esercizio fisico [16]. Tuttavia, l’ingestione orale di BCAA è molto discutibile per quanto riguarda l’ottimizzazione delle prestazioni e della sintesi proteica [14,17,18]. Recenti review sistematiche mostrano che l’integrazione di BCAA tende ad attenuare l’indolenzimento muscolare, che è un indicatore di danno muscolare [17,18]. Gli effetti ergogenici dei BCAA sono stati esaminati principalmente negli adulti e pochi studi hanno incluso partecipanti allenati, che possono differire sostanzialmente in termini di prestazioni, danno muscolare e composizione corporea. Inoltre, l’Australian Institute of Sport ha classificato i BCAA nel gruppo C, che comprende gli integratori privi di supporto scientifico tra gli atleti o studi non conclusivi. Tuttavia, i BCAA non sono raccomandati nei programmi di integrazione [15]. La panoramica del Comitato Olimpico Internazionale non ha menzionato i BCAA tra i diversi argomenti trattati, ovvero gli integratori utilizzati per prevenire o trattare le carenze di nutrienti, gli integratori utilizzati per fornire energia e gli integratori che migliorano le prestazioni sportive [19]. Per quanto riguarda i tre BCAA, sono stati descritti solo gli effetti della leucina sul turnover proteico, anche se gli studi citati non includevano partecipanti allenati [20,21].
Negli studi che hanno valutato l’impatto dell’integrazione di BCAA sulle prestazioni, i partecipanti erano in genere atleti di ciclismo e corsa [21,22,24,26], atleti impegnati in allenamenti contro-resistenza [23,25,27], pallavolisti [28] e calciatori [29,30]. Tenendo conto delle variazioni nei protocolli di integrazione, i risultati suggeriscono che i BCAA hanno effetti insignificanti sulle prestazioni. In un altro studio, i partecipanti hanno seguito una dieta ipocalorica con carboidrati o BCAA e un pesante programma di allenamento contro-resistenza per 8 settimane [26]. Gli autori hanno riscontrato che l’integrazione con BCAA ha aumentato significativamente la forza della parte superiore (15,1 ± 2,2 kg) e inferiore del corpo (7,1 ± 1,6 kg), mentre il gruppo CHO ha mostrato cambiamenti trascurabili nella forza della parte superiore del corpo (4,8 ± 1,8 kg). Uno studio separato, che ha esaminato gli effetti dell’integrazione di BCAA a lungo termine per 10 settimane in 18 ciclisti, ha riscontrato un’interazione tra gruppo e tempo per il picco di potenza nel gruppo BCAA [27]. Tuttavia, gli autori non hanno controllato l’apporto nutrizionale e gli atleti sono stati istruiti a mantenere le loro abitudini alimentari durante l’indagine [27]. Altri studi hanno esaminato gli effetti dell’integrazione di BCAA per 7-8 giorni [25,31,32] immediatamente prima, durante o dopo l’esercizio fisico [21,28,29,30]. Per questi studi, pochi manoscritti hanno testato l’integrazione di BCAA in riferimento agli indicatori di prestazione.
L’efficacia dell’integrazione di BCAA sulla composizione corporea è stata esaminata in 50 corridori amatoriali [25], 17 atleti contro-resistenza [26] e 18 ciclisti [27]. Dopo 7 giorni di integrazione orale, sono stati osservati cambiamenti comparabili nel peso corporeo nel gruppo BCAA e nel gruppo di controllo [25]. Sono stati osservati effetti trascurabili per il gruppo BCAA nel tessuto magro e nella massa grassa [26,27].
L’indolenzimento o il recupero muscolare sono stati valutati mediante l’uso di scale o esaminando i cali di prestazione post-esercizio attraverso l’uso di test specifici sul campo o in laboratorio. Negli sport di resistenza (corsa o ciclismo), i risultati degli studi estratti non sono conclusivi. Le valutazioni dello sforzo percepito e della fatica mentale sono risultate significativamente ridotte nel gruppo BCAA tra sette ciclisti di resistenza [21], mentre in 50 maratoneti non sono state riportate differenze nelle valutazioni dello sforzo percepito [25]. Al contrario, l’indolenzimento muscolare è risultato inferiore nel gruppo BCAA rispetto al placebo in 16 corridori di distanza [33]. Gli studi che hanno incluso partecipanti allenati contro-resistenza e che hanno esaminato gli effetti dei BCAA sull’indolenzimento muscolare o sul recupero hanno indicato potenziali benefici di leucina, isoleucina o valina nell’attenuazione dell’indolenzimento muscolare dopo l’esercizio [34,35]. Due studi hanno riportato che il consumo di un’integrazione di BCAA ha attenuato i cali di prestazione [36,37]. Una serie di parametri biochimici, ormonali e molecolari sono stati analizzati dopo un intervento di esercizio fisico combinato con l’integrazione di BCAA. I risultati principali di questi studi estratti rivelano che l’assunzione di BCAA ha causato un sostanziale miglioramento del rapporto BCAA:triptofano [21,27] e della risposta immunitaria [27,38,39]. Tenendo conto delle variazioni metodologiche tra gli studi, i cambiamenti ormonali indicano che l’integrazione di BCAA ha favorito una risposta ormonale anabolizzante. Il livello di cortisolo è diminuito dopo l’esercizio fisico e, parallelamente, il testosterone tendeva ad aumentare nei partecipanti impegnati nell’allenamento contro-resistenza [40]. Gli studi che hanno esaminato i meccanismi biochimici si sono concentrati principalmente su segnali metabolici specifici e sulla loro dipendenza dal meccanismo del complesso target della rapamicina 1 (mTORC1).
BCAA e sintesi proteica muscolare:
Le proteine muscolari sono in costante stato di turnover, il che significa che vengono continuamente prodotte nuove proteine mentre quelle più vecchie vengono degradate. Lo stato anabolico non ha una definizione specifica, ma in generale si riferisce alla circostanza in cui il tasso di sintesi delle proteine muscolari supera il tasso di degradazione delle proteine muscolari. Il risultato è un aumento della massa muscolare. Convenzionalmente si ritiene che lo stato anabolico sia guidato da una stimolazione della sintesi proteica muscolare, ma teoricamente potrebbe anche derivare da un’inibizione della degradazione delle proteine muscolari.
L’obiettivo metabolico principale del consumo di integratori di BCAA è quello di massimizzare lo stato anabolico. È opinione diffusa che i BCAA inducano uno stato anabolico stimolando la sintesi proteica muscolare. Un’abbondante disponibilità di tutti gli EAA è un requisito per una stimolazione significativa della sintesi proteica muscolare [41]. La sintesi proteica muscolare sarà limitata dalla mancanza di disponibilità di uno qualsiasi degli EAA, mentre una carenza di NEAA può essere compensata da una maggiore produzione de novo dei NEAA carenti [41]. Nello stato postprandiale successivo a un pasto contenente proteine, tutti i precursori degli EAA necessari per la sintesi di nuove proteine muscolari possono essere ricavati dalle elevate concentrazioni plasmatiche derivanti dalla digestione delle proteine consumate o dal riciclo dalla scomposizione delle proteine. In questa circostanza di abbondante disponibilità di EAA, il tasso di sintesi proteica muscolare supera il tasso di degradazione, producendo così uno stato anabolico. Nello stato post-assorbitivo i livelli plasmatici di EAA scendono al di sotto dei valori post-prandiali perché gli aminoacidi non vengono più assorbiti. Di conseguenza, gli EAA non vengono più assunti dal muscolo, ma rilasciati dal muscolo nel plasma [42]. Questo stato catabolico della proteina muscolare nello stato post-assorbitivo consente di mantenere la disponibilità di EAA per altri tessuti per mantenere il tasso di sintesi proteica a spese della proteina muscolare, che può essere considerata come una riserva di EAA a cui attingere per il resto del corpo.
Poiché gli EAA non possono essere prodotti nell’organismo e vi è un rilascio netto di EAA dal muscolo, nello stato post-assorbitivo l’unica fonte di precursori di EAA per la sintesi proteica muscolare è costituita dagli EAA intracellulari derivati dalla degradazione delle proteine muscolari [42]. Oltre a essere reincorporati nelle proteine muscolari attraverso la sintesi, alcuni EAA rilasciati dalla disgregazione delle proteine muscolari possono essere parzialmente ossidati all’interno del muscolo, rendendoli così indisponibili per la reincorporazione nelle proteine muscolari. Gli EAA rilasciati dalla degradazione delle proteine muscolari che non vengono reincorporati nelle proteine muscolari o ossidati all’interno del tessuto muscolare vengono rilasciati nel plasma, dove possono essere assorbiti da altri tessuti come precursori per la sintesi proteica o ossidati irreversibilmente [43]. Pertanto, il tasso di sintesi proteica muscolare sarà sempre inferiore al tasso di degradazione delle proteine muscolari nello stato post-assorbitivo, a causa del flusso netto di EAA dalla degradazione delle proteine al plasma e alle vie ossidative. In altre parole, è impossibile che la sintesi proteica muscolare superi il tasso di degradazione delle proteine muscolari quando i precursori derivano interamente dalla degradazione delle proteine, e quindi non si può verificare uno stato anabolico in assenza di assunzione di aminoacidi esogeni.
Tutti i precursori EAA per la sintesi proteica muscolare nello stato post-assorbitivo derivano dalla disgregazione delle proteine muscolari. È stato costantemente riportato che negli esseri umani normali in fase post-assorbitiva il tasso di degradazione delle proteine muscolari supera il tasso di sintesi delle proteine muscolari di circa il 30% [44]. Il consumo dei soli BCAA (cioè senza gli altri EAA) può aumentare la sintesi proteica muscolare nello stato post-assorbitivo solo aumentando l’efficienza del riciclo degli EAA dalla disgregazione proteica alla sintesi proteica, invece di essere rilasciati nel plasma o ossidati. Questo perché tutti i 9 EAA (e gli 11 NEAA) sono necessari per produrre proteine muscolari e gli EAA non possono essere prodotti dall’organismo. Se si consumano solo 3 EAA, come nel caso del consumo di BCAA, la ripartizione proteica è l’unica fonte dei restanti EAA necessari come precursori per la sintesi proteica muscolare. È quindi teoricamente impossibile che il consumo di soli BCAA crei uno stato anabolico in cui la sintesi proteica muscolare superi la degradazione delle proteine muscolari. Se si ipotizza generosamente che il consumo di BCAA migliori del 50% l’efficienza del riciclo degli EAA dalla disgregazione delle proteine muscolari alla sintesi delle proteine muscolari, ciò si tradurrebbe in un aumento del 15% del tasso di sintesi delle proteine muscolari (30% riciclato allo stato basale X 50% miglioramento del riciclo = 15% aumento della sintesi). Inoltre, una riduzione del 50% del rilascio di EAA nel plasma dal muscolo ridurrebbe anche i pool plasmatici e intracellulari di EAA liberi. La figura seguente illustra schematicamente questi principi. Poiché un miglioramento del 50% nell’efficienza del riciclo sarebbe circa il limite massimo ragionevole, ciò significa che la stimolazione massima della sintesi proteica muscolare non potrebbe superare il 15%. Ciò corrisponderebbe a un aumento del tasso di sintesi frazionale del muscolo da un valore di circa 0,050%/h allo stato basale a 0,057%/h, e questa differenza nel tasso di sintesi frazionale (FSR) delle proteine sarebbe difficile da misurare con precisione [45].
Rappresentazione schematica del riciclo degli aminoacidi essenziali (EAA) dalla disgregazione delle proteine muscolari alla sintesi delle proteine muscolari nello stato post-assorbitivo. Le unità arbitrarie sono utilizzate per semplicità e si basano sui tassi misurati di ciascuna via in soggetti umani in fase post-assorbitiva in circostanza normale nello stato post-assorbitivo. Circa il 70% degli EAA provenienti dalla disgregazione delle proteine muscolari viene riciclato nella sintesi proteica. Esiste un efflusso netto di circa l’85% degli EAA rilasciati dalla disgregazione proteica, che possono essere assunti e incorporati nelle proteine di altri tessuti oppure ossidarsi. Circa il 15% degli EAA provenienti dalla degradazione delle proteine viene parzialmente ossidato nel muscolo e non è disponibile per la sintesi proteica. Le cifre relative al flusso verso l’esterno e all’ossidazione intracellulare degli EAA sono medie, poiché alcuni EAA, come la fenilalanina, non vengono ossidati affatto nel muscolo. b Rappresentazione di un aumento del 50% dell’efficienza del riciclo degli EAA dalla disgregazione delle proteine muscolari alla sintesi proteica. In questo esempio si avrebbe un aumento della sintesi da 70 a 80 unità, ovvero del 20%. La sintesi proteica non può mai superare la ripartizione proteica nello stato post-assorbitivo, poiché la ripartizione proteica è l’unica fonte di EAA.
I BCAA sono stati somministrati per via endovenosa negli unici studi che hanno determinato la risposta del metabolismo proteico muscolare in soggetti umani ai soli BCAA. Sebbene l’infusione di BCAA non sia la modalità convenzionale di assunzione di un integratore alimentare, è stato dimostrato che gli aminoacidi infusi per via endovenosa e quelli ingeriti per via orale producono effetti comparabili sulla sintesi proteica muscolare in altre circostanze [46]. Di conseguenza, è ragionevole valutare i documenti in cui viene descritta la risposta della sintesi proteica muscolare all’infusione endovenosa di BCAA in soggetti umani.
Louard et al. [47] hanno utilizzato il metodo dell’equilibrio dell’avambraccio per quantificare la risposta all’infusione endovenosa di una miscela di BCAA per 3 ore in 10 soggetti in fase post-assorbitiva. Il metodo dell’equilibrio dell’avambraccio prevede la misurazione dell’assorbimento e del rilascio di singoli EAA (leucina e fenilalanina in questo caso) e delle loro controparti marcate isotopicamente. Vengono calcolati i tassi di scomparsa (Rd) e di comparsa (Ra) di fenilalanina e leucina. Partendo dal presupposto che il bilancio di leucina e fenilalanina nel muscolo è rappresentativo di tutti gli EAA, il Rd. della fenilalanina è considerato un riflesso della sintesi proteica muscolare, poiché la sintesi proteica è l’unico destino della fenilalanina assunta dal plasma nel muscolo. La Rd. della leucina non può essere interpretata in relazione alla sintesi proteica, poiché la leucina assunta dal muscolo può essere ossidata oltre che incorporata nelle proteine. L’infusione di 3 ore di BCAA ha aumentato le concentrazioni plasmatiche di tutti e 3 i BCAA di quattro volte, mentre le concentrazioni di altri EAA sono diminuite [47]. Invece di essere stimolata dall’infusione di BCAA, la sintesi proteica muscolare è diminuita da 37+/- 3 a 21 +/- 2 nmol/min/100 ml di gamba (statisticamente significativo, p < 0,05) [47]. Non si sono verificate variazioni significative nel bilancio netto della fenilalanina, il che indica che anche la degradazione delle proteine muscolari è stata ridotta in misura simile alla riduzione della sintesi proteica muscolare. Il bilancio tra la sintesi e la degradazione delle proteine muscolari è rimasto negativo, il che significa che lo stato catabolico è persistito e non si è prodotto uno stato anabolico. La diminuzione simultanea della sintesi e della degradazione delle proteine muscolari durante l’infusione di BCAA può essere descritta come una diminuzione del turnover proteico muscolare.
Risultati simili sono stati ottenuti dagli stessi ricercatori quando hanno esteso l’infusione di BCAA a 16 ore in 8 volontari normali e hanno determinato se l’aumento cronico di BCAA stimolasse la sintesi proteica muscolare [48]. Per calcolare la sintesi e la ripartizione delle proteine muscolari è stata utilizzata la stessa metodologia di bilanciamento dell’avambraccio dello studio precedente. L’infusione di 16 ore ha aumentato le concentrazioni di BCAA da 5 a 8 volte [48], ovvero il doppio dei livelli raggiunti con una dose normale di BCAA ingeriti per via orale [49]. Come nello studio precedente, la sintesi proteica muscolare (riflessa dalla fenilalanina Rd) si è ridotta nei soggetti che hanno ricevuto i BCAA rispetto all’infusione di soluzione salina, passando da 36 +/- 5 a 27 +/-2 nmol/min/100 ml. Anche la disgregazione proteica muscolare si è ridotta, il che significa che anche il turnover proteico muscolare è stato ridotto e che è persistito uno stato catabolico.
Da questi due studi possiamo concludere che l’infusione di BCAA non solo non aumenta il tasso di sintesi proteica muscolare nei soggetti umani, ma anzi riduce il tasso di sintesi proteica muscolare e il tasso di turnover proteico muscolare. Lo stato catabolico non è stato invertito in uno stato anabolico in nessuno dei due studi. Inoltre, una riduzione prolungata del tasso di turnover delle proteine muscolari dovrebbe avere un effetto negativo sulla forza muscolare, anche se la massa muscolare viene mantenuta. Il ricambio delle proteine muscolari rinnova le fibre muscolari e determina una maggiore efficienza della contrazione a livello di singola fibra [50], che si riflette in un aumento della forza in vivo, indipendentemente dalla massa muscolare [51, 52].
Il mancato aumento significativo della sintesi proteica muscolare in risposta all’infusione dei soli BCAA è atteso in base alle considerazioni teoriche discusse in precedenza e illustrate nella figura sopra esposta per quanto riguarda il requisito di tutti gli EAA per sostenere un aumento. Invece, poiché la disgregazione delle proteine muscolari è diminuita, è diminuita anche la disponibilità di EAA, che a sua volta ha ridotto il tasso di sintesi proteica muscolare.
L’affermazione che la sintesi proteica muscolare sia stimolata dai BCAA deriva, almeno in parte, dall’osservazione dell’aumento della segnalazione anabolica intracellulare, compreso lo stato di attivazione di fattori chiave coinvolti nell’avvio della sintesi proteica [53]. La teoria secondo cui l’attivazione dei fattori di segnalazione anabolica intracellulare provoca un aumento del tasso di sintesi proteica muscolare si è radicata nei moderni concetti di regolazione della sintesi proteica muscolare. L’aumento della segnalazione anabolica in risposta ai BCAA è stato citato come prova di una stimolazione della sintesi proteica muscolare, anche in assenza della misurazione della sintesi proteica muscolare (ad esempio, [53]). Tuttavia, l’attivazione delle vie di segnalazione anabolica può coincidere con un aumento della sintesi proteica muscolare solo in presenza di un’ampia quantità di EAA che forniscano i precursori necessari per produrre proteine complete.
La dissociazione tra lo stato di fosforilazione dei fattori di segnalazione e la sintesi proteica muscolare nell’uomo è stata dimostrata in diverse circostanze quando la disponibilità di tutti gli EAA è limitata. Ad esempio, un aumento della concentrazione di insulina (ad esempio in seguito all’assunzione di glucosio) è un potente attivatore delle vie di segnalazione anabolica, ma non riesce ad aumentare la FSR muscolare a causa della carenza di EAA [54]. Al contrario, il consumo di una piccola quantità (3 g) di EAA stimola la sintesi proteica muscolare senza influenzare l’attività dei fattori di iniziazione, come Akt, S6 chinasi e 4E-BP1 [55]. Un piccolo aumento delle concentrazioni plasmatiche di EAA non avrebbe alcun effetto se la sintesi proteica fosse limitata dallo stato di attivazione dei fattori di iniziazione. Negli studi citati in precedenza, in cui i BCAA sono stati infusi per via endovenosa, è ragionevole presumere che un aumento così consistente delle concentrazioni di BCAA avrebbe attivato i fattori di segnalazione, eppure la sintesi proteica muscolare è effettivamente diminuita a causa della mancanza di disponibilità di EAA derivante da una diminuzione della disgregazione proteica. Pertanto, nei soggetti umani la somministrazione di EAA può aumentare la sintesi proteica muscolare in assenza di qualsiasi cambiamento nell’attivazione dei fattori di iniziazione, mentre l’attivazione dei fattori di iniziazione in assenza del consumo di tutti gli EAA non ha alcun effetto sulla sintesi proteica muscolare. Questi risultati possono essere interpretati solo come la dimostrazione che il controllo limitante della sintesi proteica muscolare basale nell’uomo è la disponibilità di tutti gli EAA e non l’attività dei fattori di segnalazione anabolica. Questa conclusione mette ulteriormente in dubbio il ruolo dell’integrazione alimentare dei soli BCAA come stimolatori della sintesi proteica muscolare.
Se si considerano tutte le prove e le teorie, è ragionevole concludere che non esistono prove credibili che l’ingestione di un integratore alimentare di BCAA determini da solo una stimolazione fisiologicamente significativa delle proteine muscolari. Anzi, le prove disponibili indicano che i BCAA in realtà diminuiscono la sintesi proteica muscolare. Tutti gli EAA devono essere disponibili in abbondanza perché l’aumento della segnalazione anabolica si traduca in un’accelerazione della sintesi proteica muscolare.
A differenza della mancanza di un effetto interattivo tra BCAA e carboidrati, i BCAA possono potenziare l’effetto anabolico di un pasto proteico. Ad esempio, l’aggiunta di 5 g di BCAA a una bevanda contenente 6,25 g di proteine del siero di latte ha aumentato la sintesi proteica muscolare a un livello paragonabile a quello indotto da 25 g di proteine del siero di latte [56]. Questo risultato suggerisce che uno o più BCAA potrebbero essere limitanti per la stimolazione della sintesi proteica muscolare da parte delle proteine del siero di latte, oppure che i BCAA in più inducono un maggiore potenziale di risposta anabolica del muscolo alle proteine del siero di latte attivando i fattori di iniziazione. In entrambi i casi, la risposta dei BCAA in combinazione con le proteine intatte è una questione diversa rispetto all’effetto dei soli BCAA, poiché le proteine intatte forniscono tutti gli EAA necessari per produrre una proteina intatta.
Le risposte ai singoli BCAA (cioè leucina, valina o isoleucina) potrebbero differire dalla combinazione dei tre per diversi motivi. È dimostrato che la leucina da sola può esercitare una risposta anabolica (ad esempio, [57]), mentre non esistono dati simili per l’isoleucina o la valina. Pertanto, ci si potrebbe aspettare che la leucina da sola sia più efficace della combinazione di tutti i BCAA. Tuttavia, un’integrazione alimentare di sola leucina presenta due limitazioni significative. In primo luogo, gli stessi problemi che limitano l’entità della stimolazione della sintesi proteica muscolare da parte dei soli BCAA, relativi alla disponibilità degli altri EAA necessari per la produzione di proteine muscolari intatte, limitano anche la risposta alla sola leucina. In secondo luogo, l’aumento della concentrazione plasmatica di leucina attiva la via metabolica che ossida tutti i BCAA. Di conseguenza, l’ingestione della sola leucina determina una diminuzione delle concentrazioni plasmatiche di valina e isoleucina. La disponibilità di valina e isoleucina può quindi diventare limitante per la sintesi proteica muscolare quando si consuma solo leucina. Questo potrebbe essere il motivo per cui gli studi sui risultati a lungo termine con l’integrazione dietetica di leucina non hanno dato risultati positivi [58]. Il motivo principale per cui un integratore alimentare contiene tutti i BCAA rispetto alla sola leucina è quello di superare le diminuzioni delle concentrazioni plasmatiche di valina e isoleucina che si verificherebbero quando la leucina viene somministrata da sola.
Sebbene un integratore alimentare con tutti i BCAA superi le diminuzioni di concentrazione derivanti dal consumo della sola leucina, l’aggiunta di valina e isoleucina può comunque limitare l’efficacia della sola leucina a causa della competizione per il trasporto nelle cellule muscolari. I BCAA sono tutti trasportati attivamente nelle cellule, comprese quelle muscolari, dallo stesso sistema di trasporto. Pertanto, se forniti insieme, i BCAA competono tra loro per il trasporto nelle cellule. Se uno dei BCAA (ad esempio, la leucina) è limitante per la sintesi proteica, l’aggiunta degli altri due BCAA potrebbe limitare la stimolazione della sintesi proteica a causa del ridotto ingresso della leucina nella cellula. I BCAA competono anche con altri aminoacidi per il trasporto, compresa la fenilalanina, e questa competizione potrebbe influenzare la disponibilità intramuscolare di altri EAA. A causa della competizione per i trasportatori, è possibile che la leucina da sola, ad esempio, abbia un effetto stimolante transitorio sulla sintesi proteica muscolare (ad esempio, [59]) laddove i BCAA non riescono a suscitare tale risposta [60, 61].
Conclusioni:
Come abbiamo visto, la ricerca ha evidenziato che l’integrazione di BCAA non sembra avere un impatto significativo sulle prestazioni. D’altra parte, l’ingestione orale di BCAA isolati riduce l’indolenzimento muscolare. I BCAA sono disponibili anche in diversi prodotti di integrazione (ad esempio, proteine del siero del latte) e sono spesso combinati con altri nutrienti (ad esempio, carboidrati). Pertanto, i potenziali benefici dell’integrazione di BCAA isolati negli atleti per attenuare l’indolenzimento muscolare e ritardare l’affaticamento devono essere interpretati con cautela.
Rimane il fatto che un aumento fisiologicamente significativo del tasso di sintesi proteica muscolare richiede un’adeguata disponibilità di tutti i precursori aminoacidici. La fonte di EAA per la sintesi proteica muscolare nello stato post-assorbitivo è il pool libero intracellulare. Gli EAA liberi intracellulari disponibili per l’incorporazione nelle proteine derivano dalla degradazione delle proteine muscolari. In condizioni normali, circa il 70% degli EAA rilasciati dalla degradazione delle proteine muscolari viene reincorporato nelle proteine muscolari. L’efficienza della reincorporazione degli EAA provenienti dalla degradazione delle proteine nelle proteine muscolari può essere aumentata solo in misura limitata. Per questo motivo fondamentale, un’integrazione alimentare di soli BCAA non può sostenere un aumento del tasso di sintesi proteica muscolare. La disponibilità degli altri EAA diventerà rapidamente limitante per la sintesi proteica accelerata. Coerentemente con questa prospettiva, i pochi studi condotti su soggetti umani hanno riportato una diminuzione, piuttosto che un aumento, della sintesi proteica muscolare dopo l’assunzione di BCAA. Si può concludere, quindi, che gli integratori alimentari di BCAA da soli non promuovono l’anabolismo muscolare.
Direi che le informazioni riportate siano sufficienti a far desistere nell’acquisto di questo integratore chiunque sia dotato di un minimo di capacità cognitiva…
Jump up to:ab Scaini, G.; Jeremias, I. C.; Morais, M. O.; Borges, G. D.; Munhoz, B. P.; Leffa, D. D.; Andrade, V. M.; Schuck, P. F.; Ferreira, G. C.; Streck, E. L. (2012). “DNA damage in an animal model of maple syrup urine disease”. Molecular Genetics and Metabolism. 106 (2): 169–174. doi:10.1016/j.ymgme.2012.04.009. PMID22560665.
Jump up to:ab Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, et al. (November 2001). “Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo”. Nature Cell Biology. 3 (11): 1014–9. doi:10.1038/ncb1101-1014. PMID11715023. S2CID16284975.
Phillips S.M. The impact of protein quality on the promotion of resistance exercise-induced changes in muscle mass. Nutr. Metab. 2016;13:64. doi: 10.1186/s12986-016-0124-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Plotkin D.L., Delcastillo K., Van Every D.W., Tipton K.D., Aragon A.A., Schoenfeld B.J. Isolated Leucine and Branched-Chain Amino Acid Supplementation for Enhancing Muscular Strength and Hypertrophy: A Narrative Review. Int. J. Sport Nutr. Exerc. Metab. 2021;31:292–301. doi: 10.1123/ijsnem.2020-0356. [PubMed] [CrossRef] [Google Scholar]
Australian Institute of Sports . Branched-Chain Amino Acids (Bcaa) Summary Report: Consideration for Classification of a Supplement Ingredient. Australian Institute of Sport; Bruce, Australia: 2021. [Google Scholar]
Jäger R., Kerksick C.M., Campbell B.I., Cribb P.J., Wells S.D., Skwiat T.M., Purpura M., Ziegenfuss T.N., Ferrando A.A., Arent S.M., et al. International Society of Sports Nutrition Position Stand: Protein and exercise. J. Int. Soc. Sports Nutr. 2017;14:20. doi: 10.1186/s12970-017-0177-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Wolfe R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017;14:30. doi: 10.1186/s12970-017-0184-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Doma K., Singh U., Boullosa D., Connor J.D. The effect of branched-chain amino acid on muscle damage markers and performance following strenuous exercise: A systematic review and meta-analysis. Appl. Physiol. Nutr. Metab. 2021;46:1303–1313. doi: 10.1139/apnm-2021-0110. [PubMed] [CrossRef] [Google Scholar]
Hormoznejad R., Zare A.J., Mansoori A. Effect of BCAA supplementation on central fatigue, energy metabolism substrate and muscle damage to the exercise: A systematic review with meta-analysis. Sport Sci. Health. 2019;15:265–279. doi: 10.1007/s11332-019-00542-4. [CrossRef] [Google Scholar]
Maughan R.J., Burke L.M., Dvorak J., Larson-Meyer D.E., Peeling P., Phillips S.M., Rawson E.S., Walsh N.P., Garthe I., Geyer H., et al. IOC consensus statement: Dietary supplements and the high-performance athlete. Br. J. Sports Med. 2018;52:439–455. doi: 10.1136/bjsports-2018-099027. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Wilkinson D.J., Hossain T., Hill D.S., Phillips B.E., Crossland H., Williams J., Loughna P., Churchward-Venne T.A., Breen L., Phillips S.M., et al. Effects of leucine and its metabolite β-hydroxy-β-methylbutyrate on human skeletal muscle protein metabolism. J. Physiol. 2013;591:2911–2923. doi: 10.1113/jphysiol.2013.253203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Areces F., Salinero J.J., Abian-Vicen J., González-Millán C., Gallo-Salazar C., Ruiz-Vicente D., Lara B., Del Coso J. A 7-day oral supplementation with branched-chain amino acids was ineffective to prevent muscle damage during a marathon. Amino Acids. 2014;46:1169–1176. doi: 10.1007/s00726-014-1677-3. [PubMed] [CrossRef] [Google Scholar]
Dudgeon W.D., Kelley E.P., Scheett T.P. In a single-blind, matched group design: Branched-chain amino acid supplementation and resistance training maintains lean body mass during a caloric restricted diet. J. Int. Soc. Sports Nutr. 2016;13:1. doi: 10.1186/s12970-015-0112-9. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kephart W.C., Wachs T.D., Thompson R.M., Brooks Mobley C., Fox C.D., McDonald J.R., Ferguson B.S., Young K.C., Nie B., Martin J.S., et al. Ten weeks of branched-chain amino acid supplementation improves select performance and immunological variables in trained cyclists. Amino Acids. 2016;48:779–789. doi: 10.1007/s00726-015-2125-8. [PubMed] [CrossRef] [Google Scholar]
Moberg M., Apró W., Ekblom B., van Hall G., Holmberg H.C., Blomstrand E. Activation of mTORC1 by leucine is potentiated by branched-chain amino acids and even more so by essential amino acids following resistance exercise. Am. J. Physiol. Cell Physiol. 2016;310:C874–C884. doi: 10.1152/ajpcell.00374.2015. [PubMed] [CrossRef] [Google Scholar]
Samuelsson H., Moberg M., Apró W., Ekblom B., Blomstrand E. Intake of branched-chain or essential amino acids attenuates the elevation in muscle levels of PGC-1α4 mRNA caused by resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2016;311:E246–E251. doi: 10.1152/ajpendo.00154.2016. [PubMed] [CrossRef] [Google Scholar]
Smith J.W., Krings B.M., Shepherd B.D., Waldman H.S., Basham S.A., McAllister M.J. Effects of carbohydrate and branched-chain amino acid beverage ingestion during acute upper body resistance exercise on performance and postexercise hormone response. Appl. Physiol. Nutr. Metab. 2018;43:504–509. doi: 10.1139/apnm-2017-0563. [PubMed] [CrossRef] [Google Scholar]
Martín-Martínez J.P., Calleja Gonzalez J., Adsuar Sala J.C., Gómez-Pomares S., Carlos-Vivas J., Pérez-Gómez J. Short-term branched-chain amino acid supplementation does not enhance vertical jump in professional volleyball players. A double-blind, controlled, randomized study. Nutr. Hosp. 2020;37:1007–1011. [PubMed] [Google Scholar]
Pancar S. The effect of Branched Chain Amino Acids Intake Before and after Exercise on Physical Performance and Recovery. Ambient. Sci. 2020;7:243–246. doi: 10.21276/ambi.2020.07.sp1.oa
[CrossRef] [Google Scholar]33. Mor A., Acar K., Yilmaz A.K., Arslanoglu E. The effects of BCAA and creatine supplementation on anaerobic capacity and ball kicking speed in male football players. J. Mens Health. 2022;18:5. [Google Scholar]34. Koba T., Hamada K., Samurai M., Matsumoto K., Higuchi T., Zhao M., Miyata H. Effect of A Branched-chain Amino Acids Supplementation on Muscle Soreness during Intensive Training Program: 229: Board# 136: 11:00 AM–12:30 PM. Med. Sci. Sports Exerc. 2005;37:S43. [Google Scholar]
Gee T.I., Deniel S. Branched-chain aminoacid supplementation attenuates a decrease in power-producing ability following acute strength training. J. Sports Med. Phys. Fit. 2016;56:1511–1517. [PubMed] [Google Scholar]
Waldron M., Whelan K., Jeffries O., Burt D., Howe L., Patterson S.D. The effects of acute branched-chain amino acid supplementation on recovery from a single bout of hypertrophy exercise in resistance-trained athletes. Appl. Physiol. Nutr. Metab. 2017;42:630–636. doi: 10.1139/apnm-2016-0569. [PubMed] [CrossRef] [Google Scholar]
VanDusseldorp T.A., Escobar K.A., Johnson K.E., Stratton M.T., Moriarty T., Cole N., McCormick J.J., Kerksick C.M., Vaughan R.A., Dokladny K., et al. Effect of Branched-Chain Amino Acid Supplementation on Recovery Following Acute Eccentric Exercise. Nutrients. 2018;10:1389. doi: 10.3390/nu10101389. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Bassit R.A., Sawada L.A., Bacurau R.F., Navarro F., Martins E., Jr., Santos R.V., Caperuto E.C., Rogeri P., Costa Rosa L.F. Branched-chain amino acid supplementation and the immune response of long-distance athletes. Nutrition. 2002;18:376–379. doi: 10.1016/S0899-9007(02)00753-0. [PubMed] [CrossRef] [Google Scholar]
Bassit R.A., Sawada L.A., Bacurau R.F., Navarro F., Costa Rosa L.F. The effect of BCAA supplementation upon the immune response of triathletes. Med. Sci. Sports Exerc. 2000;32:1214–1219. doi: 10.1097/00005768-200007000-00005. [PubMed] [CrossRef] [Google Scholar]
Nemet D., Wolach B., Eliakim A. Proteins and amino acid supplementation in sports: Are they truly necessary? Isr. Med. Assoc. J. 2005;7:328–332. [PubMed] [Google Scholar]
Vahid I., Abdolali B., Fatemeh M., Alireza N., Mehdi S. The effects of branch-chain amino acids on fatigue in the athletes. Interv. Med. Appl. Sci. 2018;10:233–235. doi: 10.1556/1646.10.2018.10. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Lysenko E.A., Vepkhvadze T.F., Lednev E.M., Vinogradova O.L., Popov D.V. Branched-chain amino acids administration suppresses endurance exercise-related activation of ubiquitin proteasome signaling in trained human skeletal muscle. J. Physiol. Sci. 2018;68:43–53. doi: 10.1007/s12576-016-0506-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Goldberg A.L., Chang T.W. Regulation and significance of amino acid metabolism in skeletal muscle. Fed. Proc. 1978;37:2301–2307. [PubMed] [Google Scholar]
Stipanuk M.H., Claudill M.A. Biochemical, Physiological, and Molecular Aspects of Human Nutrition. Saunders; St. Louis, MO, USA: 2013. [Google Scholar]
Volpi E, Kobayashi H, Sheffield-Moore M, Mittendorfer B, Wolfe RR. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78:250–258. [PMC free article] [PubMed] [Google Scholar]
Wolfe RR. The underappreciated role of muscle in health and disease. Am J Clin Nutr. 2006;84(3):475–482. [PubMed] [Google Scholar]
Biolo G, Gastaldelli A, Zhang X-J, Wolfe RR. Protein synthesis and breakdown in skin and muscle: a leg model of amino acid kinetics. Am J Physiol Endocrinol Metab. 1994;30:E467–E474. [PubMed] [Google Scholar]
Smith GI, Patterson BW, Mittendorfer B. Human muscle protein turnover-why is it so variable? J Appl Physiol. 1985;110:480–491. doi: 10.1152/japplphysiol.00125.2010. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Rasmussen BB, Wolfe RR, Volpi E. Oral and intravenously administered amino acids produce similar effects on muscle protein synthesis in the elderly. J Nutr Health Aging. 2002;6:358–362. [PMC free article] [PubMed] [Google Scholar]
Louard RJ, Barrett EJ, Gelfand RA. Effect of infused branched-chain amino acids on muscle and whole body amino acid metabolism in man. Clin Sci. 1990;79:457–466. doi: 10.1042/cs0790457. [PubMed] [CrossRef] [Google Scholar]
Matthews DE. Observations of branched-chain amino acid administration in humans. J Nutr. 2005;135:1580S–1584S. [PMC free article] [PubMed] [Google Scholar]
Fitts RH, Ramatowski JG, Peters JR, Paddon-Jones D, Wolfe RR, Ferrando AA. The deleterious effects of bed rest on human skeletal muscle fibers are exacerbated by hypercortisolemia and ameliorated by dietary supplementation. Am J Physiol Cell Physiol. 2007;293:C313–C320. doi: 10.1152/ajpcell.00573.2006. [PubMed] [CrossRef] [Google Scholar]
Balagopal P, Rooyackers OE, Adey DB, Ades PA, Nair KS. Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans. Am J Phys. 1997;273:E790–E800. [PubMed] [Google Scholar]
Paddon-Jones D, Sheffield-Moore M, Urban RJ, Aarsland A, Wolfe RR, Ferrando AA. The catabolic effects of prolonged inactivity and acute hypercortisolemia are offset by dietary supplementation. J Clin Endocrinol Metab. 2005;90(3):1453–1459. doi: 10.1210/jc.2004-1702. [PubMed] [CrossRef] [Google Scholar]
Blomstrand E, Eliasson J, Karlsson HKR, Kohnke R. Branched-chain amino acids activate key enzymes in protein synthesis after physical exercise. J Nutr. 2006;136:269S–273S. [PubMed] [Google Scholar]
Greenhaff PL, Karagounis LG, Peirce N, Simpson EJ, Hazell M, Layfield R, Wackerhage H, Smith K, Atherton P, Selby A, Rennie MJ. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in muscle. Am J Physiol Endocrinol Metab. 2008;295:E 590–E 597. doi: 10.1152/ajpendo.90411.2008. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Bukari SS, Phillips BE, Wilinson DJ, Limb MC, Rankin D, Mitchell WK, Kobayashi H, Greenhaff PL, Smith K, Atherton PJ. Intake of low-dose leucine-rich essential amino acids stimulates muscle anabolism equivalently to bolus whey protein in older women at rest and after exercise. Am J Phys. 2015;308:E1056–E1065. [PubMed] [Google Scholar]
Churchward-Venne TA, Breen L, Di Donato DM, Hector AJ, Mitchell CJ, Moore DR, Stellingwerff T, Breuille D, Offord EA, Baker SK, Phillips SM. Leucine supplementation of a low-protein mixed macronutrient beverage enhances myofibrillar protein synthesis in young men: a double blind, randomized trial. Am J Clin Nutr. 2014;99:276–286. doi: 10.3945/ajcn.113.068775. [PubMed] [CrossRef] [Google Scholar]
Wilinson DJ, Hossain T, Hill DS, Phillips BE, Crossland H, Williams J, Loughna P, Chruchward-Venne TA, Breen L, Phillips SM, Etheridge T, Rathmacher JA, Smith K, Szewczk NJ, Atherton PJ. Effects of leucine and its metabolite β-hydroxy-β-methlybutryate on human skeletal muscle protein metabolism. J Physiol. 2013;591:2911–2923. doi: 10.1113/jphysiol.2013.253203. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Van Loon LJ. Leucine as a pharmaconutrient in health and disease. Curr Opin Clin Nutr Metab Care. 2012;15:71–77. doi: 10.1097/MCO.0b013e32834d617a. [PubMed] [CrossRef] [Google Scholar]
Ferrando AA, Williams BD, Stuart CA, Lane HW, Wolfe RR. Oral branched chain amino acids decrease whole-body proteolysis. JPEN. 1995;19:47–54. doi: 10.1177/014860719501900147. [PubMed] [CrossRef] [Google Scholar]
Louard RJ, Barrett EJ, Gelfand RA. Effect of infused branched-chain amino acids on muscle and whole body amino acid metabolism in man. Clin Sci. 1990;79:457–466. doi: 10.1042/cs0790457. [PubMed] [CrossRef] [Google Scholar]
Nella 2° parte abbiamo analizzato le caratteristiche e funzioni biochimiche della Glutammina. In questa terza parte, invece, andremo ad analizzare due AA legati tra loro per via metabolica, la L-Citrullina e la L-Arginina.
Dalla L-Citrullina alla L-Arginina – Biologia e principali attività:
Il composto organico Citrullina è un α-amminoacido (formula H2NC(O)NH(CH 2)3CH(NH2)CO2H. ).[1] Sebbene sia stato nominato e descritto dai gastroenterologi fin dalla fine del XIX secolo, è stato isolato per la prima volta dall’anguria nel 1914 dai ricercatori giapponesi Yotaro Koga e Ryo Odake [2] [3] e ulteriormente codificato da Mitsunori Wada dell’Università Imperiale di Tokyo nel 1930.[4] La L-Citrullina è un composto aminoacidico non proteico (non viene utilizzato per formare proteine strutturali come gli enzimi) e, a differenza della L-Arginina, non è ampiamente presente in tutte le proteine. Si trova in concentrazioni particolarmente alte nell’anguria (da cui deriva il suo nome, dato che i cocomeri sono conosciuti come Citrullus vulgaris[1]), dove si trova in media a 2,1mg/g di peso umido (anche se i numeri assoluti variano)[2] e si è notato che il consumo di anguria aumenta in modo acuto sia l’Arginina plasmatica che la Citrullina (3.3 kg di anguria equivalgono a 10g di L-Arginina supplementare)[3][4] e di aumentare l’Arginina e l’Ornitina a digiuno del 12-22% in seguito al consumo di 780-1560g al giorno.[5]
Altre fonti alimentari di L-Citrullina sono i meloni, i meloni amari, le zucchine, le zucche e i cetrioli.[6]
La Citrullina viene sintetizzata nell’organismo attraverso una delle due vie: riciclata dall’Arginina (la conversione dell’arginina in ossido nitrico lascia la citrullina come sottoprodotto)[7][8] o prodotta dall’azoto (e da una parte del carbonio) contenuto nella L-glutammina,[9] dove l’enzima ornitina transcarbamilasi utilizza sia l’Ornitina che il carbamoilfosfato (che richiede la Clutammina) per produrre Citrullina negli enterociti.[10][11]
Sembra che la via dell’Arginina sia responsabile di circa il 10% della Citrullina circolante, mentre la via della Glutammina ne rappresenta il 90%;[6] la riduzione dei livelli plasmatici di Glutammina può ridurre la Citrullina plasmatica.[12]
Per quanto riguarda il ciclo dell’urea (uno dei meccanismi alla base del 10%), la L-Arginina viene convertita in L-Ornitina tramite l’enzima arginasi (cedendo urea come cofattore)[13][14] e da qui l’Ornitina (utilizzando il carbamoilfosfato come cofattore) viene sottoposta all’enzima Ornitina carbamoiltransferasi per produrre L-Citrullina. In questo senso, la via metabolica dall’Arginina alla Citrullina (attraverso l’Ornitina) provoca un aumento dell’urea e una concomitante diminuzione dell’ammoniaca, utilizzata dall’enzima carbamoilfosfato sintasi per creare carbamoilfosfato.[15] Se necessario, l’arginina può essere convertita direttamente in L-Citrullina attraverso un enzima arginina deiminasi per produrre, anziché richiedere, ammoniaca.[16]
Il ciclo si forma quando la citrullina si lega con l’L-aspartato (correlato all’acido D-aspartico come isomero) per formare l’arginosuccinato attraverso l’enzima arginosuccinato sintasi, quindi l’enzima arginosuccinato lisasi degrada l’arginosuccinato in arginina libera e fumarato; l’arginina rientra quindi nel ciclo dell’urea. [Il fumarato può semplicemente entrare nel ciclo TCA (Krebs) come intermedio energetico,[17] e la citrullina regola negativamente l’enzima arginasi.[18]
Anche la conversione della citrullina in L-arginosuccinato e la successiva conversione in L-arginina è coinvolta nel ciclo dell’ossido nitrico piuttosto che nel ciclo dell’urea, con l’unica differenza che l’arginina si converte direttamente in citrullina (cedendo una molecola di ossido nitrico) piuttosto che essere convertita indirettamente tramite l’ornitina.[18][19]
Come accennato, l’Arginina entra prima in contrata con il metabolismo intestinale e splancnico, in cui una certa quantità di essa viene consumata dagli enterociti o interconvertita in L-citrullina o L-ornitina. Oltre all’elevato utilizzo dell’arginina da parte del fegato, anche l’assorbimento intestinale dell’arginina è scarso in condizioni normali e aumenta in varie patologie.[20] Sembra che una quantità minima di L-arginina arrivi ai tessuti sistemici rispetto agli altri aminoacidi del ciclo dell’urea, dato che la L-ornitina supplementare raggiunge una concentrazione sierica doppia rispetto alla L-arginina e la L-citrullina 9,3 volte superiore. Ciò sembra direttamente correlato al grado di metabolismo epatico e intestinale.[21][22][23]
L’Arginina alimentare rappresenta il 40-60% dell’arginina sierica, come evidenziato da un calo equivalente durante i periodi di assenza di arginina. Il tasso di conversione della L-citrullina in L-arginina non sembra influenzato dall’assunzione con la dieta.[24]
La citrullina di per sé è più che altro un sottoprodotto del metabolismo dell’arginina (ciclo dell’ossido nitrico) e dell’ornitina (ciclo dell’urea) e viene semplicemente riconvertita in arginina tramite l’arginosuccinato. Detto questo, l’integrazione di citrullina influisce positivamente anche sulle concentrazioni di arginina e ornitina, quindi anche la loro bioattività è rilevante.
L’arginina può essere convertita in L-citrullina attraverso gli enzimi dell’ossido nitrico sintasi (NOS), di cui esistono forme endoteliali (eNOS) e neuronali specifiche (nNOS), nonché una forma inducibile (iNOS) che risponde ai segnali infiammatori. La conversione dell’arginina attraverso gli enzimi NOS produce ossido nitrico come sottoprodotto più importante, e la Citrullina è vista come un sottoprodotto.[25] La Citrullina può poi riconvertirsi in L-arginina attraverso l’arginosuccinato, ma la L-ornitina non è coinvolta nella via dell’ossido nitrico.
La L-Citrullina viene assorbita nell’intestino in misura molto maggiore rispetto alla sua controparte L-Arginina e determina un livello plasmatico più elevato di L-Arginina attraverso il ciclo Arginina/Ornitina/Citrullina.[26] Viene assorbita attraverso numerosi trasportatori sodio-dipendenti.[27]
È stato osservato che l’integrazione orale di citrullina nell’uomo a 0,18 g/kg raddoppia l’arginina plasmatica[28], cosa che è stata replicata altrove[29], insieme a un aumento equivalente delle concentrazioni di ornitina[29], ma questi raddoppi di arginina e ornitina sono associati a un aumento di 6-11 volte della citrullina plasmatica[28][29].
Una singola dose di 6 g di citrullina malato (0,08 g/kg) in atleti prima dell’esercizio fisico ha fatto registrare aumenti della citrullina plasmatica (aumento del 173%), dell’ornitina (aumento del 152%) e dell’arginina (aumento del 123%) quando misurata dopo l’esercizio fisico, valori che si sono normalizzati con 3 ore di riposo.[30] Questa stessa dose è stata notata altrove per aumentare la citrullina plasmatica e l’arginina in misura simile.[31]
È interessante notare che gli studi sopra citati che hanno utilizzato 0,18 g/kg di citrullina hanno rilevato un aumento di 6-11 volte della citrullina a fronte di un mero raddoppio dell’arginina e dell’ornitina[28][29], mentre lo studio successivo che ha utilizzato 6 g (calcolati come 0,08 g/kg) ha registrato un aumento molto minore della citrullina, ma ha comunque più che raddoppiato sia l’arginina che l’ornitina. [30] Ciò è stato osservato anche in uno studio dose-risposta che ha utilizzato da 2 a 15 g di citrullina, in cui la citrullina nel plasma ha seguito una dipendenza lineare dalla dose, mentre sia l’arginina che l’ornitina hanno avuto una dipendenza minore dalla dose.[29] Gli autori hanno ipotizzato che, dato che l’aumento dell’arginina è stato inferiore a quello previsto e che la citrullina sierica è il principale predittore della sintesi dell’arginina[19], ciò indichi il raggiungimento di una fase di limitazione della velocità nei reni.
È stato osservato che la citrullina non influenza i livelli sierici degli aminoacidi a catena ramificata a riposo,[21] ma può accelerare la deplezione dei BCAA indotta dall’esercizio fisico prolungato (aumentandone l’utilizzo come carburante).[20]
Con l’integrazione di citrullina è stata notata una riduzione della glutammina (13% dopo 0,18 g/kg di citrullina per 7 giorni)[21], anche se un altro studio ha rilevato che l’uso acuto di 6 g di citrullina (0,08 g/kg) non ha alterato le concentrazioni di glutammina.[20]
Gli altri aminoacidi testati (acido glutammico, acido aspartico, asparagina, alanina, lisina, triptofano, fenilalanina, L-tirosina, istidina) sono per lo più inalterati.[20]
Circa l’83% della citrullina ingerita per via orale sembra essere assorbita dai reni[26][27][28] dove viene convertita in L-arginina nei tubuli prossimali (attraverso gli enzimi arginosuccinato sintasi e arginosuccinato liasi[29]); Questa conversione della citrullina in arginina (sia da citrullina supplementare che da quella prodotta come sottoprodotto della creazione di ossido nitrico da parte dell’arginina) rappresenta il 5-15% dell’arginina circolante. [11][30]
Il meccanismo principale con cui l’integrazione di arginina (e, per estensione, di L-citrullina) influisce sulla salute del sangue è quello di essere il substrato per gli enzimi dell’ossido nitrico sintasi (NOS) per la produzione di ossido nitrico, che poi segnala attraverso i recettori ciclici solubili della guanilina la produzione di cGMP. La produzione di ossido nitrico e la successiva produzione di cGMP intracellulare sono alla base di buona parte dei benefici dell’arginina.
Gli enzimi NOS si presentano in tre isoforme principali: [32][33] la NOS inducibile (iNOS), che viene creata in risposta a fattori di stress infiammatori, la NOS neuronale (nNOS), che è stata scoperta per la prima volta nei neuroni e si trova anche nelle terminazioni motorie dei muscoli scheletrici, e la NOS endoteliale (eNOS), che inizialmente si pensava si trovasse solo nell’endotelio, ma è piuttosto diffusa[34], compreso il tessuto cerebrale.[35][36]
Gli enzimi NOS lavorano in dimeri uniti testa a testa e i meccanismi catalitici dipendono da questa dimerizzazione, oltre che dall’eme, dalla tetraidrobiopterina, dalla calmodulina, dal NADPH (come donatore di elettroni) e da FMN e FAD.[37][38][39] Di conseguenza, gli enzimi NOS (tutte e tre le isoforme) sono flavoproteine che richiedono NADPH. [40][41][42] La loro struttura e funzione è complessa (esaminata qui[43]), ma esiste un sito di legame di base per l’arginina e gli elettroni donati dal NADPH fanno sì che l’arginina si converta in citrullina, rilasciando come sottoprodotto l’ossido nitrico; l’iNOS utilizza esclusivamente e l’eNOS può anche utilizzare un intermedio radicale libero chiamato Nω-idrossi-L-arginina (L-NOHA), che si degrada in citrullina e ossido nitrico in presenza di H2O2.[32][44]
L’aumento dell’ossido nitrico (solitamente misurato attraverso le concentrazioni plasmatiche di nitrato/nitrito, citrullina o cGMP urinario) sembra essere aumentato con la L-arginina in persone affette da ipertensione essenziale,[45] arterotrombosi,[46] e diabete di tipo II. [47] Gli studi condotti su atleti altrimenti sani che assumono L-arginina sono piuttosto contrastanti; ci sono casi in cui i biomarcatori del metabolismo dell’ossido nitrico sono aumentati[48] mentre altri studi non notano alcuna modifica.[49][50][51] Non sorprende che i benefici associati all’ossido nitrico non si verifichino quando i biomarcatori dell’ossido nitrico non sono aumentati.
L’inaffidabilità dell’aumento dell’ossido nitrico da parte dell’arginina può essere dovuta al fatto che le concentrazioni fisiologiche di arginina (40-100µM nello spazio extracellulare[52] e forse fino a 800µM a livello intracellulare[53]) sono sufficienti a saturare intrinsecamente l’ossido nitrico sintasi endoteliale (eNOS) (di solito si dichiara una Km di 3µM[54][55], ma a volte viene misurata fino a 29μM[56). Ciò implica che l’enzima è già al massimo dell’efficacia e che un’ulteriore integrazione non aumenta il tasso di conversione (a causa di un arretrato di arginina nel siero); l’osservazione che l’arginina aumenta ancora l’ossido nitrico a volte (anche se in modo inaffidabile) è indicata come il paradosso della L-arginina.[57][58]
Questa teoria è in linea con le osservazioni secondo cui a volte il metabolismo dell’ossido nitrico non viene influenzato nonostante aumenti fino al 300% dell’arginina plasmatica.[59]
Uno studio ha osservato un aumento transitorio della produzione di ossido nitrico che sembra essere più simile a quello di un agonista che di un substrato[60] e successivamente è stato scoperto che l’arginina ha la capacità di attivare i recettori α2-adrenergici,[61] che possono stimolare direttamente l’ossido nitrico senza richiedere la conversione in citrullina attraverso la NOS. Tuttavia, l’arginina è risultata piuttosto debole (superata dall’agmatina)[61] ma questo meccanismo non è ancora stato escluso.
Inoltre, l’arginina extracellulare sembra essere un fattore determinante per il rilascio di ossido nitrico[56] (il trasportatore CAT1 che trasporta l’arginina è altamente associato alla eNOS[62] e l’inibizione dell’afflusso extracellulare impedisce l’attivazione della eNOS[63]), mentre la concentrazione intracellulare di arginina non sembra essere associata. [58] Poiché il trasporto è necessario, ma l’arginina intracellulare non è di per sé necessaria, si ritiene che la colocalizzazione di CAT1 con eNOS[62][64] possa svolgere un ruolo nella stimolazione dell’attività di eNOS.
ADMA
L’ADMA è un metabolita metilato dell’arginina e sembra agire in opposizione all’arginina inibendo le azioni dell’enzima NOS e la conseguente produzione di ossido nitrico. Livelli eccessivi di ADMA possono essere causati da fattori di stress ossidativo che diminuiscono l’attività dell’enzima che lo degrada, mentre la riduzione dell’ADMA provoca una vasodilatazione dovuta alla produzione di ossido nitrico.
Sebbene la maggior parte delle evidenze suggerisca che l’ADMA non aumenta con l’integrazione di L-arginina (questi studi notano che il rapporto arginina:ADMA è aumentato a causa dell’aumento dell’arginina plasmatica), ci sono prove limitate che suggeriscono un aumento che richiede ulteriori indagini.[38]
L’integrazione orale di arginina (anche la citrullina si applica in questo caso perché aumenta l’arginina plasmatica) è in grado di aumentare il flusso sanguigno nelle persone con flusso sanguigno ridotto e, sebbene abbia il potenziale di ridurre la pressione sanguigna, sembra un po’ inaffidabile e può verificarsi solo negli ipertesi. Esistono prove contrastanti sugli effetti dell’integrazione di arginina sul flusso sanguigno in persone con resistenza periferica o cladicazione intermittente, con studi a breve termine che notano un beneficio e studi a più lungo termine che notano un’alterazione.[65][66]
L’integrazione di citrullina sembra ridurre la pressione sanguigna e migliorare il flusso sanguigno in situazioni in cui il flusso sanguigno è altrimenti ostacolato o la pressione sanguigna è più alta del normale, ma la citrullina non ha effetti di riduzione unidirezionali; può essere inefficace in persone normotese a riposo.[41]
In atleti allenati a cui sono stati somministrati 6 g di citrullina malato prima di un test ciclistico prolungato (137 km), l’aumento dell’ormone della crescita indotto dall’esercizio sembra essere aumentato; quando è stato misurato subito dopo l’esercizio, il gruppo con citrullina aveva concentrazioni di GH più elevate del 66,8%, che (dopo 3 ore di riposo) si sono attenuate al 28%.[20] Altrove, dosi di 2-15 g di citrullina non sono riuscite a influenzare l’ormone della crescita a riposo, se misurate nell’arco di 8 ore.[22]
Le concentrazioni di IGF-1 dopo 0,18 g/kg di citrullina per 7 giorni non sono state influenzate in modo significativo.
Durante l’esercizio fisico, sebbene uno studio che ha utilizzato 3 g di L-arginina (associata a 2.200 mg di L-ornitina e 12 mg di vitamina B12) per 3 settimane abbia rilevato un aumento del 35,7% della secrezione di ormone della crescita indotta dall’esercizio fisico (che si è normalizzata entro un’ora)[67], altri studi notano il contrario; è stato osservato che l’integrazione di arginina determina un minore picco di ormone della crescita durante l’esercizio fisico rispetto all’esercizio fisico da solo[68][69] e che, sebbene sembri influenzare maggiormente i giovani rispetto agli anziani[70], si dice che influisca su entrambi i gruppi di età. [L’entità della soppressione (supponendo che il 100% sia il valore di base) è stata notata intorno a un aumento del 300-500% visto con l’esercizio fisico, attenuato al 200%.[68]
È possibile che un aumento eccessivo dell’ormone della crescita stimoli un feedback autogeno, il che spiegherebbe come gli individui più anziani siano meno sensibili a questa soppressione, in quanto hanno intrinsecamente meno picchi di GH dovuti all’esercizio fisico rispetto ai giovani.[71] Infine, poiché i picchi dell’ormone della crescita si normalizzano comunque nel giro di poche ore[67][71], non si sa esattamente quanto sia preoccupante questa soppressione (dato che le concentrazioni di ormone della crescita nelle 24 ore sono più rilevanti).
A riposo, l’integrazione di 5-9 g di L-arginina è in grado di provocare un aumento delle concentrazioni di picco dell’ormone della crescita (aumento del 34,4-120%), mentre 13 g sono risultati inefficaci a causa della sofferenza intestinale che impedisce l’assorbimento della L-arginina.[36]
Negli studi che misurano la secrezione di GH nelle 24 ore, non sono state riscontrate alterazioni significative con la somministrazione di 2 g due volte al giorno[72] o con dosi acute di 5 g.[73] Ciò è potenzialmente legato a un noto fenomeno di feedback autonomo dell’ormone della crescita,[69][74][75] e un effetto modulatorio simile sull’ormone della crescita si riscontra anche durante la restrizione del sonno (in cui una riduzione del rilascio di ormone della crescita indotto dal sonno viene compensato durante le ore di luce). È stato osservato che l’arginina ad alte dosi (250mg/kg di arginina aspartato al giorno, circa 17,5g di arginina) aumenta l’impulso di GH nel sonno a onde lente di circa il 60%, pur non avendo un’influenza sufficiente sulle concentrazioni di GH durante la veglia.[76] Non è chiaro come questo grande aumento influisca sulle concentrazioni di ormone della crescita nell’intera giornata.
L’integrazione di arginina nei ratti è in grado di aumentare il nitrato urinario post-esercizio, indicativo della produzione di ossido nitrico.[77] Aumenti nella produzione di ossido nitrico (nitrato urinario o sierico) sono stati confermati anche nell’uomo in seguito ad assunzione orale o infusione endovenosa.[78]
Non sempre si riscontra un aumento della produzione di ossido nitrico (anche nonostante l’aumento dell’arginina plasmatica), suggerendo che l’attività dell’enzima NOS potrebbe essere un fattore limitante. Per quanto riguarda gli studi in acuto (assunzione di una singola dose di L-arginina prima dell’esercizio), 3 g di arginina (sotto forma di AAKG) non hanno apportato benefici all’allenamento con i pesi,[79] 6 g di L-arginina per 3 giorni non hanno modificato i risultati del cicloergometro in atleti di judo, mentre un protocollo simile in ciclisti allenati ha rilevato un miglioramento del tempo di esaurimento (25,8%).[80]
Alcuni studi hanno utilizzato una forma di arginina nota come GAKIC (Glycine L-Arginine α-Ketoisocaproic acid) e hanno rilevato un aumento della potenza media durante gli sprint di 10s su cicloergometro (con 11,2 g di GAKIC)[81] e un aumento del 10,5+/-0. Questi studi, tuttavia, sono confusi sia dall’inclusione della glicina sia da quella del metabolita della leucina, l’acido α-chetoisocaproico.
Per quanto riguarda gli studi più cronici, l’integrazione di L-arginina (come asparato) con 2,8 o 5,7 g di arginina al giorno per 4 settimane non è riuscita a modificare le prestazioni o altri biomarcatori[82]; anche uno studio precedente, condotto per 2 settimane con una metodologia simile, ha fallito.[83] Nel complesso, quando si esaminano le revisioni o le meta-analisi sull’argomento L-arginina e prestazioni sportive, si nota che è promettente, ma manca un consenso sufficiente per raccomandarla come ergogenico.[84]
Si ritiene che la citrullina sia un agente pro-erettile in quanto è un precursore dell’arginina, e l’arginina è il substrato da cui viene prodotto l’ossido nitrico che può poi indurre il cGMP (attraverso la via NO/cGMP/VEGF);[65] un aumento del cGMP è anche l’effetto finale degli inibitori della PDE5 come il viagra o l’icariina dall’erba cornuta.[66]
Negli uomini con disfunzione erettile, valutata come debolezza dell’erezione (valutata con il punteggio di durezza yerettile[67]), la somministrazione di 1.500 mg di citrullina al giorno (due dosi da 750 mg) per un mese è stata in grado di apportare benefici alla metà dei 24 pazienti valutati (valutati come “molto soddisfatti” del trattamento), mentre il miglioramento del placebo è stato solo dell’8,3%.[68]
La citrullina sembra interagire con il metabolismo dei BCAA nell’organismo, anche se gli studi sull’uomo sembrano avere risultati diversi a seconda del contesto dello studio.
La citrullina può mediare positivamente la segnalazione della leucina attraverso mTOR, il che suggerisce teoricamente una sinergia. L’applicazione di questa combinazione ai sollevatori di pesi non è ancora stata studiata, quindi il sinergismo è attualmente solo un’ipotesi piuttosto che un fatto dimostrato.
Il nitrato è un piccolo donatore di ossido nitrico che costituisce il principale bioattivo del succo di barbabietola. Il nitrito sierico (forma ridotta del nitrato) sembra aumentare durante l’esercizio fisico in seguito al consumo di 6 g di citrullina malato, che si ritiene sia un indicatore dell’aumento della produzione di ossido nitrico.[20]
I farmaci a base di statine possono aumentare l’espressione dell’enzima che media la conversione dell’arginina in ossido nitrico e per questo motivo è possibile che vi sia un sinergismo per tutto ciò che riguarda l’ossido nitrico. Questo non è ancora stato testato in un sistema vivente.
L-Citrullina come sostituto alla L-Arginina?
L’integrazione di L-citrullina è stata definita un’alternativa alla L-arginina, in quanto ne aggira lo scarso assorbimento e si converte in L-arginina nei reni. La L-citrullina tecnicamente segue aumenti dose-dipendenti della L-arginina sierica fino a 15 g, ma la dose orale più alta di citrullina assunta ha ritorni sempre minori (cioè per ogni 5 g in più di citrullina ingerita si aggiunge meno arginina al siero).[85]
È stato osservato che la citrullina orale a 0,18 g/kg raddoppia approssimativamente l’arginina plasmatica (aumento del 100%)[86][87] o è leggermente superiore (123%) con 0,08 g/kg.[88] Poiché le dosi più elevate di L-citrullina hanno una minore conversione in arginina[85], è improbabile che la L-citrullina supplementare possa essere utilizzata per superare l’arginina per l’aumento acuto dell’arginina sierica.
Gli studi che hanno confrontato direttamente la L-arginina con la L-citrullina hanno osservato che entrambe aumentano la Cmax a livelli comparabili a dosi orali simili (Cmax di 79+/-8μM per 3 g di citrullina e 84+/-9μM per l’arginina), ma la citrullina risulta in un’AUC complessiva maggiore (48,7% in più rispetto all’arginina). [Questo potrebbe essere dovuto al fatto che, anche fino all’ingestione di 15g di citrullina, non si verifica un aumento significativo dell’escrezione di citrullina.[85] L’assenza di un aumento dell’eliminazione di L-citrullina dal sangue nonostante l’integrazione consentirebbe di avere un pool di L-citrullina disponibile per la conversione su richiesta in L-arginina.
Citrullina Malato:
La Citrullina Malato (CM), una combinazione di L-citrullina e acido malico, è stata pubblicizzata come un aiuto ergogenico (che aumenta l’energia) per l’allenamento contro-resistenza e l’esercizio ad alta intensità.
Come abbiamo visto, la L-citrullina è un precursore dell’ossido nitrico (NO), un vasodilatatore che può migliorare l’apporto di sangue e ossigeno ai muscoli durante l’esercizio. Tuttavia, le prove finora disponibili suggeriscono che il miglioramento del flusso sanguigno non è il meccanismo attivo degli effetti ergogenici del CM. Il meccanismo potrebbe invece essere dovuto alla capacità della citrullina di favorire l’eliminazione dell’ammoniaca durante l’esercizio ad alta intensità, alla capacità del malato di aumentare la produzione di ATP, a un aumento dell’espressione genica o a una maggiore efficienza della navetta malato-aspartato.
La maggior parte delle ricerche condotte finora ha utilizzato una dose acuta di 8 grammi di CM un’ora prima dell’esercizio. Sebbene l’assunzione di CM un’ora prima dell’esercizio rimanga la migliore raccomandazione, alcuni dati suggeriscono che dosi maggiori, fino a 15 grammi, potrebbero essere più benefiche.
È stato dimostrato che l’ingestione di una serie di dosi di CM (2-15 grammi) non ha effetti negativi sui marker ematologici. Sebbene la sicurezza di un’integrazione di CM a lungo termine richieda ulteriori indagini, le ricerche condotte finora indicano che la CM è ben tollerata nella maggior parte degli individui.
Ricerche preliminari hanno suggerito che 8 grammi di CM ingeriti un’ora prima dell’esercizio fisico aumentano la resistenza muscolare (ripetizioni fino al cedimento) in uomini e donne. Tuttavia, ricerche più recenti hanno suggerito che il CM potrebbe non avere un beneficio sulle prestazioni nell’allenamento contro-resistenza, potenzialmente a causa di variazioni nei tempi e nei dosaggi.[89]
Arginina AKG:
La differenza principale tra la L-arginina e Arginina AKG è che la L-arginina è un aminoacido non essenziale che l’organismo è in grado di produrre, mentre l’arginina AKG è un sale della L-arginina e dell’acido α-chetoglutarato. Inoltre, la L-arginina regola il flusso sanguigno attraverso la produzione di ossido nitrico, mentre l’Arginina AKG dovrebbe potenzialmente aumentare il flusso sanguigno, l’energia e il recupero.
Nella nutrizione sportiva, l’AKG è stato utilizzato come integratore per migliorare la sintesi proteica muscolare e diminuire la disgregazione muscolare, ed è quindi utilizzato dagli atleti per migliorare la composizione corporea.[90][91] L’integrazione di AKG potrebbe anche migliorare le prestazioni atletiche. Uno studio ha rilevato che un integratore di arginina e alfa-chetoglutarato (AAKG) ha migliorato la forza nella panca, ma non la capacità aerobica. Sono necessarie ulteriori ricerche per sostenere le affermazioni sull’AKG come aiuto ergogenico.[92]
L’AKG viene utilizzato anche per il recupero da interventi chirurgici o traumi, perché è un precursore dell’aminoacido glutammina. Sebbene la glutammina sia un aminoacido non essenziale, viene talvolta definita “condizionatamente essenziale” perché la quantità di glutammina di cui l’organismo ha bisogno per il recupero dopo un trauma significativo può superare la quantità che l’organismo è in grado di produrre. In questo caso, un integratore di AKG può aiutare il processo di recupero.[93][94]
L’AKG è stato proposto come integratore per la longevità; alcune ricerche condotte su vermi tondi, ratti e topi suggeriscono che potrebbe aumentare la durata della vita e ritardare l’insorgenza di malattie legate all’età, anche se gli studi clinici dovranno confermare questi risultati.[95][96]
Nelle persone con malattie renali croniche, in particolare in quelle sottoposte a dialisi, la somministrazione di AKG in combinazione con il calcio ha migliorato i biomarcatori della funzione renale.[97][98]
In uno studio è stato rilevato che l’AKG aumenta l’espressione di involucrina, filaggrina e serina palmitoil transferasi. Queste molecole sono tutte importanti per la struttura dello strato esterno della pelle e per l’idratazione dello strato esterno della pelle, quindi l’uso di AKG per via topica potrebbe migliorare l’aspetto della pelle.[99][100]
Nella ricerca, i dosaggi utilizzati variano da 3,6 g a 6 g, con dosaggi più elevati nelle persone che hanno subito ustioni, ma non è ancora stata stabilita una dose giornaliera raccomandata.[101] Poiché gli effetti sono dose-dipendenti, trovare una raccomandazione di dosaggio accurata sarà una parte importante della ricerca in corso.[102]
Sicurezza e tossicità:
La citrullina sembra essere ben tollerata dai ratti in dosi fino a 3 g/kg di peso corporeo[58][46]. Negli esseri umani, 15 g di citrullina assunti acutamente non sembrano causare diarrea o disturbi intestinali[22], il che è notevolmente diverso rispetto all’ornitina e all’arginina che possono causare diarrea a dosaggi di 10 g se assunti in bolo[74][75] a causa del limitato assorbimento di questi aminoacidi che poi procedono verso il colon causando diarrea osmotica.[74]
Il limite di sicurezza osservato, ovvero la dose più alta in cui si può essere relativamente sicuri che non si verifichino effetti collaterali nel corso della vita, è stato suggerito in 20 g di arginina al giorno in forma di integratore (al di sopra dell’assunzione di cibo).[103] Dosi più elevate sono state testate e ben tollerate, ma non esistono prove che suggeriscano la loro sicurezza in tutte le popolazioni nel corso della vita.
La L-arginina ha un tasso di assorbimento gastrointestinale piuttosto scarso. Può inoltre agire come assorbente, rilasciando acqua ed elettroliti nel lume intestinale attraverso la stimolazione dell’ossido nitrico e inducendo disturbi gastrici e diarrea.[12] Questo fenomeno è noto come diarrea osmolitica e tende a verificarsi a dosi orali superiori a 10 g circa, se assunte in bolo.[36]
Si pensa che ciò avvenga attraverso la stimolazione della produzione di ossido nitrico, poiché la D-Arginina (incapace di produrre NO) non produce diarrea[104] e l’ossido nitrico stesso è noto per essere un meccanismo attraverso il quale molti lassativi osmolitici funzionano.[105]
Singoli boli di 5-9 g di L-arginina senza cibo non sembrano causare disturbi intestinali come dosi superiori a 10 g,[36] suggerendo che, almeno per uno stomaco vuoto, il dosaggio di 9 g è un limite superiore.
Conclusioni:
L’Integrazione di L-citrullina si è dimostrata più redditizia tra costi e benefici (vedi assorbimento intestinale) rispetto alla L-arginina. L’uso alternativo di Citrullina Malato può portare ad eventi gastrointestinali più frequenti rispetto alla semplice forma L-citrullina. Nonostante la ridotta biodisponibilità orale della L-arginina, questa può essere mixata con L-citrullina per un effetto additivo, anche se non rappresenta un vero e proprio vantaggio proprio di tale abbinamento.
L’assunzione di L-arginina e/o L-citrullina vede la sua miglior tempistica prima dell’allenamento al fine di aumentare il flusso sanguigno ai distretti allenati, per effetto della vasodilatazione NO indotta.
Ciò si traduce in:
Aumento dell’apporto di nutrienti e ossigeno al tessuto muscolare abbinato ad un effetto di pulizia dalle molecole di scarto, come l’acido lattico;
Esaltazione dilatatoria sul reticolo venoso sottocutaneo, che migliora la qualità estetica in definizione.
Bonus: l’abbinamento con estratto di barbabietola notoriamente ricco di nitrati.
Come effetto diretto dell’introduzione nell’organismo di estratto di barbabietola abbiamo un aumento della sintesi di ossido nitrico (NOs), dovuta, per l’appunto, ai nitrati (NO3-) contenuti in questo vegetale, convertiti rapidamente in nitriti (NO2-2) tramite enzimi che si trovano fin dal tratto orofaringeo, gastrointestinale e tracheo-bronchiale. Dato ciò, la sintesi di NO sfrutta un percorso non usuale come quello della L-Arginina, ma coadiuvante a questa e alla L-Citrullina.
Le concentrazioni di nitrati raggiungono il picco dopo circa un’ora dalla sua ingestione, per ritornare ai livelli basali dopo quasi 24h, mentre gli effetti della L-Arginina permangono per almeno 75-80 minuti, per poi iniziare a tornare ai livelli basali.
“Citrulline – Compound Summary”. PubChem Compound. USA: National Center for Biotechnology Information. 16 September 2004. Identification. Retrieved 1 May 2012.
Chin RM, Fu X, Pai MY, Vergnes L, Hwang H, Deng G, Diep S, Lomenick B, Meli VS, Monsalve GC, Hu E, Whelan SA, Wang JX, Jung G, Solis GM, Fazlollahi F, Kaweeteerawat C, Quach A, Nili M, Krall AS, Godwin HA, Chang HR, Faull KF, Guo F, Jiang M, Trauger SA, Saghatelian A, Braas D, Christofk HR, Clarke CF, Teitell MA, Petrascheck M, Reue K, Jung ME, Frand AR, Huang JThe metabolite α-ketoglutarate extends lifespan by inhibiting ATP synthase and TOR.Nature.(2014-Jun-19)
Le ricerche che vanno dalla seconda parte del XX secolo a oggi hanno dimostrato che l’allenamento contro-resistenza aumenta la crescita delle fibre muscolari scheletriche radiali (o trasversali) (rivisto in Folland e Williams, 2007; Walker et al., 2011; Haun et al., 2019c). Tuttavia, si conoscono meno gli adattamenti morfologici che si verificano nelle fibre muscolari in seguito a interventi di allenamento contro-resistenza. E’ stato recentemente ipotizzato che un allenamento contro-resistenza a volume più elevato possa facilitare la crescita delle fibre attraverso un aumento sproporzionato del volume del sarcoplasma rispetto all’accrescimento delle proteine della miofibrilla (Haun et al., 2019b; Vann et al., 2020b). L’ipertrofia sarcoplasmatica è stata una descrizione corrente di questo fenomeno e questo termine verrà utilizzato in questa sede. Non sono il primo a suggerire che l’ipertrofia sarcoplasmatica si verifichi in risposta all’allenamento contro-resistenza, e gli studi citati più avanti in questo articolo hanno sostenuto questo meccanismo. Tuttavia, è generalmente accettato dalla comunità scientifica che l’ipertrofia convenzionale si verifica in risposta all’allenamento contro-resistenza; in particolare, le fibre muscolari subiscono una crescita radiale indotta dall’allenamento attraverso un accrescimento proporzionale di proteine miofibrillari e di spazio nel sarcoplasma. In base a questo presupposto, se un individuo presenta un aumento del 20% dell’area trasversale media delle fibre (fCSA) e supponendo che le miofibrille costituiscano il ∼85% dello spazio intracellulare, un’aggiunta del 17% di proteine miofibrillari e un aumento del 3% del volume del sarcoplasma accompagnerebbero la crescita delle fibre. Questa modalità di ipertrofia delle fibre muscolari è certamente ragionevole. Ciononostante, ad oggi esiste un solo studio sull’uomo che supporta vagamente questo meccanismo (Luthi et al., 1986), e questo studio viene discusso in modo più dettagliato più avanti nell’articolo.
Numerosi studi hanno riportato che i tassi di sintesi delle proteine miofibrillari aumentano ore dopo un allenamento in persone non allenate (reviewed in Walker et al., 2011; Haun et al., 2019c) e allenate (reviewed in Damas et al., 2015). Queste osservazioni sono state coerenti e danno certamente credito al modello convenzionale di ipertrofia discusso in precedenza. Pur essendo informativi, gli studi sui traccianti non hanno chiarito in che misura l’accumulo di proteine miofibrillari a lungo termine contribuisca all’ipertrofia delle fibre muscolari. In effetti, gli aumenti post-esercizio dei tassi di sintesi proteica miofibrillare ore dopo un allenamento non sono correlati con i risultati ipertrofici a lungo termine (reviewed in Mitchell et al., 2015). Gli studi che esaminano i tassi integrati di sintesi proteica miofibrillare a distanza di giorni o settimane dall’allenamento hanno prodotto associazioni migliori con i risultati ipertrofici (Brook et al., 2015; Damas et al., 2016a; Bell et al., 2019). Tuttavia, rimane possibile che la varianza esistente tra questi dati del tracciante e i risultati ipertrofici sia legata alla crescita di componenti non miofibrillari. Inoltre, nessuna indagine sull’uomo ha determinato se l’accrescimento proteico delle miofibrille indotto dall’allenamento avvenga attraverso l’allargamento di miofibrille preesistenti, la creazione di nuove miofibrille (cioè la miofibrillogenesi de novo) o l’allargamento delle miofibrille seguito da una scissione delle stesse per mantenere una dimensione miofibrillare conservata.Si tratta certamente di lacune nella letteratura sull’allenamento contro-resistenza. Tuttavia, piuttosto che concentrarsi su questi inconvenienti, è più utile fornire una review dettagliata e approfondita degli scenari in cui l’ipertrofia sarcoplasmatica è stata e non è stata osservata nella letteratura sull’allenamento contro-resistenza nell’uomo. Voglio anche spiegare perché questo fenomeno può verificarsi da una prospettiva teorica e meccanicistica. Infine, partendo dal presupposto che l’ipertrofia sarcoplasmatica è un adattamento all’allenamento contro-resistenza, discuterò le implicazioni più ampie di questo meccanismo.
Morfologia intracellulare delle fibre muscolari:
Prima di discutere le diverse modalità di ipertrofia, è fondamentale che i lettori comprendano la morfologia intracellulare delle fibre muscolari. La figura seguente illustra la sezione trasversale di una fibra muscolare come appare al microscopio elettronico a trasmissione (TEM). Da un punto di vista molecolare, le osservazioni in laboratorio hanno riportato che il tessuto muscolare è composto per il ∼75% da acqua, per il ∼10-15% da proteine miofibrillari e per il ∼5% da proteine non miofibrillari (o sarcoplasmatiche) (Vann et al., 2020b). Da un punto di vista spaziale, tuttavia, la maggior parte della fibra muscolare è occupata dalle miofibrille; secondo alcune stime, le miofibrille occupano ∼85% dello spazio intracellulare (Macdougall et al., 1982; Alway et al., 1988; Claassen et al., 1989). Le miofibrille sono costituite da varie proteine, tra cui le isoforme della catena pesante e della catena leggera della miosina, varie isoforme dell’actina, varie isoforme della troponina e della tropomiosina, proteine della linea z (ad esempio, alfa-actinina, proteina LIM specifica del muscolo, miopalladina e teletonina), proteine della linea m (ad esempio, miomesina e proteina M tra le altre) e altre proteine che servono a mantenere l’integrità strutturale delle miofibrille (ad esempio, titina e nebulina tra le altre) (Haun et al., 2019c; Vann et al., 2020b). Nel complesso, questo pool di proteine è solitamente indicato come la frazione miofibrillare del tessuto muscolare. Lo spazio tra le miofibrille è scarso (cioè, secondo le stime sopra riportate, il ∼15% dello spazio intracellulare non è occupato dalle miofibrille). Questi spazi contengono organelli e componenti cellulari, tra cui i mitocondri (probabilmente sotto forma di reticolo), il reticolo sarcoplasmatico, macromolecole (ad esempio, ribosomi, glicogeno e goccioline lipidiche) e varie proteine ed enzimi. Il TEM ha dimostrato che i mitocondri occupano il 5-6% dello spazio all’interno delle fibre muscolari, mentre il sarcoplasma ne occupa il 9% (Claassen et al., 1989). Il mezzo acquoso in cui risiedono le miofibrille e i componenti non miofibrillari è chiamato sarcoplasma. Come il citoplasma di altre cellule, il sarcoplasma è fondamentale per mantenere l’equilibrio degli ioni e del pH all’interno delle fibre muscolari. I componenti non miofibrillari delle fibre muscolari e il sarcoplasma costituiscono la cosiddetta frazione sarcoplasmatica del tessuto muscolare.
Schema semplificato della morfologia delle fibre muscolari. L’inserto di sinistra è una figura ricreata di ciò che si osserverebbe al TEM se il tessuto fosse preparato per l’imaging in sezione. L’immagine dell’inserto di destra è una pseudocolorata immagine TEM in sezione trasversale all’interno della fibra muscolare scheletrica di un ratto Wistar che mostra i diversi componenti cellulari (15.000×). In particolare, gran parte dell’attenzione in questa figura è dedicata alle miofibrille. Tuttavia, è importante notare che i mitocondri costituiscono il ∼5% dell’area della cellula muscolare in sezione trasversale. L’immagine è stata concessa in licenza da sciencesource.com (per gentile concessione di Jose Luis Calvo) dagli autori per scopi di pubblicazione scientifica.
La figura qui sopra non illustra come le differenze nel tipo di fibra influenzino la morfologia cellulare. Gli studi sugli animali hanno dimostrato che le differenze principali tra le fibre a contrazione lenta (tipo I) e quelle a contrazione rapida (tipo II) sono le seguenti:
1- le fibre a contrazione lenta contengono più mitocondri (o un reticolo mitocondriale espanso) rispetto alle fibre a contrazione rapida (Schiaffino et al., 1970);
2- le fibre a contrazione rapida possiedono vescicole del reticolo sarcoplasmatico più grandi che circondano le miofibrille (Lee et al., 1991) e
3- le vescicole del reticolo sarcoplasmatico circondano le miofibrille, e i diametri delle miofibrille sembrano essere leggermente più grandi nelle fibre a contrazione rapida, sebbene i sarcomeri siano leggermente più lunghi e le linee z siano leggermente più spesse nelle fibre a contrazione lenta (Schiaffino et al., 1970).
Alcune di queste caratteristiche specifiche del tipo di fibra sono state evidenziate anche nel muscolo scheletrico umano con metodi TEM (Alway et al., 1988). La figura seguente è un’immagine TEM che mostra visivamente alcune di queste differenze specifiche del tipo di fibra.
Immagine TEM pseudocolorata in sezione trasversale di un ratto Wistar (8.000×) che mostra le differenze morfologiche tra i diversi tipi di fibre muscolari. L’immagine è stata concessa in licenza da sciencesource.com (per gentile concessione di Jose Luis Calvo) dagli autori per scopi di pubblicazione scientifica.
Anche il sarcolemma, o membrana cellulare di una fibra muscolare, è un importante argomento di discussione. Il sarcolemma contiene varie proteine, tra cui recettori e trasportatori legati alla membrana. Il sarcolemma contiene anche proteine transmembrana e queste proteine formano complessi che collegano proteine citoscheletriche intracellulari a proteine che risiedono nella matrice extracellulare (Henderson et al., 2017). Dati recenti proteomici indicano che alcune proteine associate al sarcolemma sono arricchite nella frazione miofibrillare (ad esempio, la distrofina), mentre altre sono arricchite nella frazione sarcoplasmatica (ad esempio, la vinculina) (Vann et al., 2020b). Il processo di omogeneizzazione del tessuto porta probabilmente a questa compartimentazione differenziale delle proteine associate al sarcolemma. In particolare, sebbene queste proteine siano arricchite in una o in entrambe le frazioni, non rientrano tecnicamente in nessuna delle due classificazioni. Questo argomento verrà rivisto più avanti.
Metodi utilizzati per valutare l’ipertrofia sarcoplasmatica:
Esistono quattro metodi pubblicati che sono stati utilizzati per segnalare che l’ipertrofia sarcoplasmatica può verificarsi in risposta all’allenamento di resistenza. Tre di questi metodi includono la TEM, la colorazione con falloidina e la colorazione SDS-PAGE e Coomassie. Anche la valutazione della tensione specifica nelle fibre muscolari isolate (o spellate) può fornire una prova indiretta dell’ipertrofia sarcoplasmatica. Di seguito vengono illustrati i meccanismi, i vantaggi e gli svantaggi di ciascun metodo.
TEM
Microscopio Elettronico a Trasmissione (TEM).
Questo metodo prevede di sottoporre un piccolo pezzo di tessuto muscolare (∼1-2 mg) a lunghe fasi di fissazione e disidratazione, seguite da un’inclusione in resina. Vengono quindi generate sezioni di spessore nanometrico con l’ausilio di un ultramicrotomo; queste sezioni vengono posizionate su piccole griglie e trattate chimicamente con acetato di uranile e citrato di piombo per generare contrasto tra le strutture cellulari. Le griglie contenenti una o più sezioni possono quindi essere visualizzate in un TEM. Supponendo che questi metodi siano eseguiti correttamente, le immagini ottenute da questa tecnica forniscono miofibrille con una buona risoluzione a ingrandimenti di 1.000-20.000×. I ricercatori possono quindi utilizzare un software di analisi delle immagini manuale o automatico per dimensionare le miofibrille, i mitocondri e il vuoto spaziale che esiste tra queste strutture (cioè il sarcoplasma). I filamenti di miosina e di actina possono anche essere fotografati a ingrandimenti di ∼40.000× per esaminare la spaziatura e le proprietà reticolari. Uno dei principali svantaggi della TEM è che la tecnica fornisce solo un’immagine bidimensionale per una porzione di una o due fibre muscolari. Sebbene sia possibile acquisire più immagini casuali su più fibre muscolari per ottenere una rappresentazione più olistica del tessuto, l’ottenimento di 20 immagini per soggetto fornirebbe (nella migliore delle ipotesi) solo dati relativi a porzioni di 20-40 cellule. Un secondo limite della TEM è che i metodi di fissazione, disidratazione, inclusione, colorazione e ultramicrotomo richiedono giorni di lavoro in laboratorio. Questo non solo è problematico dal punto di vista della produttività, ma l’elevato numero di passaggi potrebbe alterare artificialmente la spaziatura miofibrillare, confondendo le misure del volume sarcoplasmatico. A questo proposito, sono stati segnalati artefatti TEM (ad esempio, spaziatura delle strutture cellulari o contrazione degli organelli) dovuti a errori di elaborazione (Mollenhauer, 1993), e questi fenomeni possono portare a conclusioni errate sulle proprietà cellulari. Infine, i dispositivi GST sono raramente presenti nei laboratori di fisiologia dell’esercizio, dati i costi e le competenze necessarie per il funzionamento e la manutenzione delle apparecchiature.
Colorazione con falloidina
Si tratta di un metodo istologico in cui una soluzione di colorazione contenente un fluoroforo coniugato alla falloidina (ad esempio, Alexa Fluor 488 Phalloidin; Thermo Fisher, Waltham, MA, Stati Uniti) viene applicata a sezioni muscolari aderenti a vetrini da microscopio. Le proprietà chimiche della falloidina le consentono di legarsi con elevata specificità ai polimeri di actina; pertanto, un punto di forza della tecnica è la specificità della colorazione. L’abbondanza relativa di actina nelle sezioni trasversali di muscolo incubate con Alexa Fluor 488 Falloidina può essere ottenuta in ∼20-30 fibre intere con una semplice microscopia a fluorescenza con un ingrandimento di 200×. Se si ottengono più immagini di una sezione in diverse regioni del tessuto, si possono interrogare ∼50-100 fibre. Con un software di conteggio automatico dei pixel, è possibile fornire un conteggio imparziale dei pixel verdi all’interno dei bordi colorati di distrofina per ricavare l’abbondanza relativa di actina per miofibra. Oltre all’elevata specificità di legame della falloidina all’actina, un altro punto di forza della tecnica è il campionamento di un maggior numero di fibre intere rispetto all’imaging TEM. Anche le fasi di lavorazione del tessuto sono meno laboriose rispetto alla preparazione TEM. Come la TEM, tuttavia, anche questa tecnica è limitata a un’immagine bidimensionale. Un’altra limitazione è che nelle fibre muscolari sono presenti proteine di actina non contrattili che fungono da impalcatura (Stromer, 1998). Anche se queste proteine probabilmente non costituiscono gran parte della frazione proteica sarcoplasmatica, è probabile che la falloidina si leghi a queste proteine e generi segnale dal materiale non contrattile. Per eseguire con successo questa tecnica è necessaria anche un’adeguata conservazione dei tessuti mediante congelamento lento in mezzi a temperatura di taglio ottimale. Un congelamento improprio potrebbe portare a un artefatto comunemente osservato di grandi strutture simili a vacuoli, che potrebbero essere erroneamente interpretate come spazio non occupato dalle miofibrille (Meng et al., 2014). Per ulteriori dettagli su questo metodo, il lettore può fare riferimento al nostro lavoro passato (Haun et al., 2019b). Inoltre, Gokhin et al. (2008) hanno utilizzato questo metodo per quantificare le quantità relative di materiale contrattile per fibra muscolare nei roditori in crescita.
Elettroforesi su gel di sodio dodecil solfato-poliacrilammide (SDS-PAGE) con colorazione Coomassie
Si tratta di una tecnica non istologica che consente di determinare le abbondanze relative delle isoforme proteiche della catena pesante della miosina, dell’actina, della tropomiosina, della troponina e della catena leggera della miosina per milligrammo di tessuto muscolare umido o secco. Poiché abbiamo eseguito diverse iterazioni di questo saggio (Roberts et al., 2018; Haun et al., 2019b; Vann et al., 2020b), forniremo una descrizione più approfondita di questa tecnica rispetto ad altre. Nella Figura 3 è riportato anche uno schema che mostra come vengono interpretati i risultati del saggio. Va notato che anche altri ricercatori hanno eseguito questa tecnica per quantificare la quantità relativa di proteine contrattili per unità di tessuto in varie specie (Yamada et al., 1997; Thys et al., 2001; Wilkens et al., 2008). Una volta ottenuta la biopsia muscolare, il tessuto viene avvolto in una pellicola pre-etichettata, congelato in azoto liquido e trasportato a -80°C per la conservazione a lungo termine. Dato che il tessuto è congelato allo stato nativo e che le fibre muscolari contengono ∼75% di acqua, come già detto, si parla comunemente di tessuto umido. Durante il giorno della sperimentazione, il tessuto viene rimosso da -80°C e posto su un mortaio raffreddato ad azoto liquido (LN2). Il tessuto viene quindi polverizzato con un pestello raffreddato a LN2 e una frazione del tessuto (∼20 mg) viene posta in una provetta da 1,7 mL prepesata contenente un tampone a base di Tris con Triton-X 100 allo 0,5% e inibitori delle proteasi (denominato tampone 1). Questi dettagli sono fondamentali, poiché questo primo tampone contiene il detergente Triton-X, che è in grado di lisare le membrane cellulari senza solubilizzare le miofibrille. Questo tampone può anche essere uno dei motivi per cui alcune proteine del sarcolemma sono arricchite nella frazione proteica sarcoplasmatica, come indicato in precedenza nella recensione. Le provette vengono quindi ripesate per ottenere un peso muscolare umido basato sulla differenza di peso tra la provetta prima e dopo l’aggiunta di tessuto. La provetta contenente il tampone 1 e il tessuto viene quindi rimossa dalla bilancia analitica, polverizzata con pestelli a tenuta fino alla formazione di un impasto, quindi le provette vengono tappate e centrifugate. Dopo la centrifugazione, rimangono un surnatante liquido e un pellet. Il surnatante contiene principalmente il tampone 1 e i costituenti del sarcoplasma (ad esempio, proteine sarcoplasmatiche e mitocondriali). Il pellet contiene soprattutto proteine miofibrillari, dato che non sono state solubilizzate. Tuttavia, abbiamo rilevato anche alcune proteine istoniche nella frazione pellettata con la proteomica (Vann et al., 2020b). Sebbene la presenza di queste proteine nucleari sia minima rispetto ad altre proteine contrattili (ad esempio, la catena pesante della miosina, la titina, la nebulina e altre), questi dati suggeriscono che i nuclei vengono probabilmente eliminati durante la prima fase di centrifugazione. Circa il 90% del surnatante viene rimosso con una pipetta, il surnatante viene posto in una nuova provetta da 1,7 mL e questa provetta contenente la frazione sarcoplasmatica viene conservata a -80°C per le analisi successive. Alla provetta originale da 1,7 mL contenente il pellet di proteine miofibrillari viene aggiunto altro tampone 1 e il pellet viene risospeso per lavarlo. Lo slurry ottenuto viene centrifugato e il surnatante risultante viene scartato dopo la centrifugazione. Il pellet di miofibrille può essere asciugato all’aria con ghiaccio e poi risospeso in quello che abbiamo chiamato tampone 2, un tampone a base di Tris contenente inibitori delle proteasi, cloruro di potassio, glicerolo e spermidina. In particolare, tutti questi additivi favoriscono la solubilizzazione del pellet di miofibrille. Una volta risospese le miofibrille, un volume prestabilito della risospensione (ad esempio, 10 μL) può essere preparato con tampone riducente e sottoposto a SDS-PAGE. Il gel può quindi essere colorato con Coomassie, la densità delle bande prominenti esistenti a ∼220 kD (catena pesante di miosina) e 43 kD (actina) può essere quantificata e le densità delle bande possono essere divise per il peso del muscolo per ottenere le quantità relative di catena pesante di miosina e actina. In relazione alle limitazioni discusse con il TEM e l’istologia, l’uso di questo metodo presenta dei vantaggi. In primo luogo, viene campionata una quantità maggiore di muscolo rispetto alla TEM o alla colorazione con falloidina, e questo vantaggio offre una maggiore sicurezza nelle conclusioni sullo stato fisiologico del tessuto. In secondo luogo, la determinazione dell’abbondanza relativa (o delle concentrazioni) delle due principali proteine contrattili consente di estrapolare le proprietà tridimensionali del tessuto. In alternativa, se le abbondanze relative della catena pesante della miosina e dell’actina diminuiscono con un concomitante aumento dell’area del tessuto o della fCSA durante l’allenamento di resistenza, questo risultato potrebbe riflettere un aumento del volume del sarcoplasma in centinaia di fibre muscolari. In terzo luogo, abbiamo dimostrato che questa tecnica presenta una buona riproducibilità e sensibilità (Roberts et al., 2018). Tuttavia, come nel caso del Western blotting, una limitazione importante del metodo è che fornisce dati su lisati grezzi. A questo proposito, l’osservazione di diminuzioni indotte dall’allenamento nelle abbondanze relative della catena pesante della miosina e dell’actina può indicare la presenza di edema localizzato (piuttosto che di ipertrofia sarcoplasmatica). Allo stesso modo, questo metodo genera una classificazione binaria delle proteine come miofibrillari o sarcoplasmatiche. In effetti, i nostri dati di proteomica hanno dimostrato che la maggior parte delle proteine miofibrillari ottenute con questo metodo sono proteine contrattili o altre proteine associate alle miofibrille, mentre la maggior parte delle proteine sarcoplasmatiche ottenute sono enzimi che risiedono nel sarcoplasma (Vann et al., 2020b). Tuttavia, in ogni frazione si trovano tracce di proteine che potrebbero essere considerate contaminanti. Come già detto, sono state identificate proteine nucleari nella frazione miofibrillare e proteine del sarcolemma sono state trovate in entrambe le frazioni. Abbiamo anche rilevato tracce di isoforme di laminina e collagene nella frazione miofibrillare. Anche se non è una limitazione, il ricercatore deve assicurarsi che il tessuto muscolare sia visibilmente privo di sangue, grasso e tessuto connettivo subito dopo la biopsia e prima del congelamento istantaneo. A questo proposito, il tessuto congelato con sangue residuo aumenterà artificialmente il peso del muscolo umido e questo errore tecnico porterà alla falsa conclusione che le abbondanze relative delle proteine contrattili sono inferiori rispetto ai campioni di tessuto visibilmente privi di sangue. Infine, la sospensione delle miofibrille nel tampone 2 (descritto sopra) rappresenta una sfida tecnica. Sebbene i ricercatori abbiano sviluppato una tecnica che solubilizza le miofibrille (Roberts et al., 2020), gli altri ricercatori interessati all’utilizzo di questa tecnica dovrebbero eseguire un’ampia sperimentazione per garantire che la solubilizzazione sia ottimizzata prima di aumentare la scala della sperimentazione.
Concettualizzazione dell’ipertrofia convenzionale rispetto a quella sarcoplasmatica e come questi fenomeni si manifesterebbero con la tecnica di elettroforesi discussa. L’inserto di sinistra mostra varie modalità di ipertrofia delle fibre muscolari. I piccoli cerchi blu all’interno della fibra rappresentano i mitocondri, i cerchi grigi più piccoli che occupano la maggior parte dello spazio intracellulare rappresentano le miofibrille e il resto dello spazio rappresenta il sarcoplasma. Nelle situazioni in cui l’ipertrofia si verifica a causa dell’accumulo proporzionale di proteine miofibrillari (cioè l’ipertrofia convenzionale), si può verificare l’aggiunta di nuove miofibrille alla periferia che “spingono” la cellula verso l’esterno (indicate come nuovi punti gialli), oppure l’ispessimento di miofibrille preesistenti (indicate dall’alone dei cerchi grigi). In alternativa, l’ipertrofia sarcoplasmatica si verifica attraverso l’espansione sproporzionata del sarcoplasma rispetto all’aggiunta di miofibrille. Alcuni studi hanno dimostrato che questo processo può verificarsi in risposta all’allenamento contro-resistenza, anche se alcuni studi confutano questo modello di ipertrofia. Questo processo potrebbe essere evidenziato attraverso l’imaging TEM, la colorazione con falloidina o la determinazione della tensione specifica nelle fibre isolate (non rappresentata nella figura). Questo processo potrebbe anche essere riflesso analizzando i campioni bioptici prima e dopo l’allenamento mediante SDS-PAGE e colorazione di Coomassie e analizzando i cambiamenti nella densità delle bande proteiche della miosina pesante (MHC) e dell’actina (illustrati nell’inserto destro della figura).
Valutazione della tensione specifica nelle singole fibre muscolari
Questa tecnica viene eseguita utilizzando un trasduttore di forza specializzato e può fornire una valutazione indiretta del materiale contrattile per sezione trasversale di fibra muscolare (D’antona et al., 2007; Degens et al., 2010; Meijer et al., 2015). I lettori interessati a osservare questa tecnica possono fare riferimento a un eccellente articolo sui metodi visivi di Roche et al. (2015), nonché a una revisione completa sull’argomento di Canepari et al. (2010). Questo metodo prevede innanzitutto l’ottenimento di una biopsia, la crioconservazione del tessuto e il suo congelamento fino al giorno della sperimentazione. Il giorno della sperimentazione, il tessuto viene scongelato e permeabilizzato in una soluzione contenente Triton X-100. Il tessuto viene quindi posto in una soluzione rilassante, le singole fibre vengono sezionate (o spellate) e una fibra viene sospesa da un capo all’altro a perni opposti con suture di nylon che si interfacciano con un trasduttore di forza. La tensione della fibra viene regolata nella soluzione rilassante al microscopio per ottenere una lunghezza ottimale del sarcomero (∼2,6 μm). A questo punto, è possibile ottenere i diametri delle fibre per estrapolare i valori di fCSA, presumendo che le fibre scorticate adottino una sezione trasversale circolare. Le fibre vengono quindi trasferite in una soluzione fisiologica di “attivazione” (pH 7,0) contenente calcio, ATP e altri componenti. La forza isometrica sviluppata dalla fibra viene quindi monitorata fino al raggiungimento di un plateau; questo valore può essere diviso per fCSA per ottenere la tensione normalizzata per l’area della fibra (cioè la tensione specifica). Come ulteriore passo, i ricercatori possono determinare il tipo di fibra rimuovendo la fibra dall’apparato del trasduttore di forza, omogeneizzandola in tamponi specifici, eseguendo SDS-PAGE con colorazione Coomassie o argento e osservando il modello di banding dell’isotipo della catena pesante della miosina. Se si esegue questa tecnica con due fibre che presentano diametri simili, si può ragionevolmente concludere che la fibra che presenta un valore di tensione specifica inferiore possiede meno miofibrille in sezione trasversale. Il punto di forza della tecnica sono i dati funzionali acquisiti su fibre ex vivo. Per fare un esempio contestuale, può essere impegnativo disgiungere il ruolo che gli adattamenti neurali svolgono nel processo di adattamento quando un individuo presenta un aumento della forza e dell’ipertrofia muscolare durante un periodo di allenamento alla resistenza. A tal fine, le interrogazioni di singole fibre possono fornire prove più chiare per stabilire se l’ipertrofia delle fibre muscolari e, più specificamente, l’accumulo di proteine miofibrillari possano aver contribuito all’aumento della forza. Un limite di questa tecnica è che vengono analizzati solo segmenti di singole fibre piuttosto che fibre di lunghezza intera. Un’altra limitazione è che la permeabilizzazione altera le caratteristiche intrinseche delle fibre, come la sensibilità al calcio e l’equilibrio osmotico, e le fibre si espandono artificialmente a causa di questi cambiamenti, come evidenziato dai valori di fCSA sensibilmente più grandi rispetto ai valori di fCSA ottenuti tramite istologia. Una terza e significativa limitazione di questa tecnica è rappresentata dalle delicate e lunghe fasi di microdissezione e dalla delicatezza dell’interfacciamento delle fibre con il trasduttore di forza. A causa del lavoro richiesto per eseguire questa tecnica, è comune che vengano analizzate meno di 10 fibre per soggetto (Meijer et al., 2015). Infine, come nel caso della TEM, questa tecnica non viene eseguita su larga scala a causa delle attrezzature specializzate e delle competenze necessarie.
In particolare, tutti i metodi discussi in precedenza richiedono l’ottenimento di biopsie di muscolo scheletrico da partecipanti umani che, di per sé, possono rappresentare una limitazione per molti laboratori. Il campionamento bioptico del muscolo vasto laterale è inoltre limitato da una bassa resa di tessuto (∼100 mg) rispetto alle dimensioni del muscolo. A questo proposito, le analisi su cadavere mostrano che la massa dell’intero muscolo vasto laterale è in media di ∼375 g negli esseri umani di età superiore a 80 anni (Ward et al., 2009), e queste masse probabilmente superano in media i 500 g nei partecipanti più giovani e sani. Di conseguenza, una biopsia preleverebbe solo 1/2500 di un muscolo quadricipite. Questa cifra diventa più misera se si considera che la TEM interroga 1-2 mg di tessuto, gli studi sulle fibre scorticate interrogano ∼10 delle centinaia di migliaia di fibre del muscolo vasto laterale, gli studi istologici esaminano decine di fibre e gli studi biochimici interrogano 20-30 mg di tessuto. Un’altra limitazione della tecnica bioptica è che presumibilmente lo stesso fascio di fibre non viene campionato quando si esegue il metodo nei momenti precedenti e successivi all’intervento. Il lettore non esperto deve essere consapevole di queste limitazioni per apprezzare meglio le argomentazioni presentate in questa revisione.
Prove nell’uomo a sostegno dell’ipertrofia sarcoplasmatica indotta dall’allenamento contro-resistenza:
Macdougall et al. (1982) hanno pubblicato il primo studio sull’uomo che fornisce buone prove per suggerire che l’ipertrofia sarcoplasmatica può contribuire in modo significativo all’ipertrofia delle fibre muscolari. Circa un secolo prima, Morpurgo riportò i risultati del primo studio sull’ipertrofia indotta dall’allenamento in un modello animale; l’autore riteneva che l’aumento delle dimensioni delle fibre fosse principalmente legato all’espansione dei componenti non miofibrillari (Morpurgo, 1897). Un precedente rapporto di potenziale ipertrofia sarcoplasmatica con l’allenamento contro-resistenza è stato riportato anche da Penman, che ha utilizzato metodi TEM (Penman, 1969). Tuttavia, lo studio di Morpurgo riguardava animali allenati al tapis roulant, mentre lo studio di Penman era limitato a 2-3 partecipanti, il che precludeva valutazioni statistiche formali. Macdougall et al. (1982) hanno esaminato cinque partecipanti che si sono impegnati in 6 mesi di allenamento contro-resistenza della parte superiore del braccio. Sono state ottenute biopsie del muscolo tricipite prima e dopo l’intervento di allenamento. I campioni di muscolo sono stati analizzati per la morfologia con metodi TEM. I valori di fCSA di tipo II sono aumentati in seguito all’allenamento e gli autori hanno inoltre osservato una diminuzione del ∼3% dello spazio bidimensionale occupato dalle miofibrille (p < 0,05) e un aumento del ∼15% dello spazio occupato dal sarcoplasma (p < 0,05). Più convincenti sono i dati riportati nello stesso articolo relativi a sette partecipanti che avevano anni di esperienza nell’allenamento contro-resistenza. Questi individui possedevano fibre muscolari di tipo II più grandi rispetto ai partecipanti non allenati prima dell’intervento di allenamento di 6 mesi, ma mostravano valori inferiori del 30% nello spazio occupato dalle miofibrille e valori due volte superiori nello spazio occupato dal sarcoplasma. Toth et al. (2012) hanno utilizzato la TEM per riferire che 18 settimane di allenamento contro-resistenza hanno ridotto lo spazio occupato dalle miofibrille del ∼15% nelle fibre muscolari del vasto laterale di individui sani. Meijer et al. (2015) hanno successivamente utilizzato tecniche di isolamento delle fibre e hanno riportato che i body-builder, che possedevano fibre muscolari del vasto laterale di grandi dimensioni, presentavano valori di tensione specifica inferiori del ∼40% rispetto ai partecipanti non allenati. Un gruppo di ricerca ha utilizzato la SDS-PAGE e la colorazione Coomassie per determinare che 6 settimane di allenamento contro-resistenza ad alto volume hanno ridotto le abbondanze relative (per milligrammo di tessuto secco) della catena pesante della miosina e dell’actina del ∼30% nel muscolo vasto laterale di 15 partecipanti ben allenati (Haun et al., 2019b). La colorazione con falloidina è stata utilizzata anche per mostrare che la densità della proteina actina per fibra è diminuita in questi partecipanti da prima a dopo l’intervento. Questo studio è stato limitato dal fatto che la biopsia post-allenamento è stata raccolta dopo solo 24 ore di recupero dall’ultimo allenamento. Tuttavia, otto di questi partecipanti hanno effettuato una terza biopsia 8 giorni dopo l’ultimo allenamento e le caratteristiche dell’ipertrofia sarcoplasmatica erano ancora evidenti. Più recentemente, è stato pubblicato uno studio su 15 uomini di età universitaria, precedentemente allenati, in cui le biopsie del vasto laterale sono state ottenute prima di un intervento di allenamento di 10 settimane e 72 ore dopo l’ultimo allenamento (Vann et al., 2020a). Rispetto a tutti gli studi citati, i partecipanti hanno eseguito un allenamento contro-resistenza a carico più elevato in cui gli esercizi sono stati eseguiti con 3-5 serie di 3-8 ripetizioni al ∼80-90% di forza massima (1RM). In media, i valori di fCSA di tipo I non sono cambiati e quelli di fCSA di tipo II sono aumentati del 19% (p < 0,05). Mentre la SDS-PAGE e la colorazione Coomassie hanno indicato che l’allenamento ha ridotto le abbondanze relative delle proteine della catena pesante della miosina e dell’actina (per milligrammo di tessuto secco) solo del ∼3%, questi cambiamenti da prima a dopo l’allenamento sono stati statisticamente significativi (p < 0,05). In particolare, queste diminuzioni della proteina contrattile per milligrammo di tessuto non erano così robuste come quelle osservate nel nostro studio precedente di Haun et al. (2019b) (cioè, diminuzioni del ∼30%). Al posto dell’ipertrofia delle fibre di tipo II osservata nel secondo studio, questi dati suggeriscono che un’apprezzabile accrescimento di proteine miofibrillari si è probabilmente verificato con un allenamento a breve termine a carico più elevato. Ciononostante, gli studi del nostro laboratorio suggeriscono che l’ipertrofia sarcoplasmatica può essere evidente con nuovi paradigmi di allenamento in soggetti già allenati. Inoltre, i nostri dati indicano che questa modalità di ipertrofia si verifica maggiormente con un allenamento a volume più elevato.
Nel complesso, i sette studi sull’uomo sopra menzionati forniscono prove ragionevoli che suggeriscono che l’ipertrofia sarcoplasmatica può verificarsi durante l’allenamento contro-resistenza. Sebbene tutti gli studi differiscano per quanto riguarda la durata dell’allenamento e gli schemi di carico, solo lo studio più recente ha attuato quello che sarebbe comunemente considerato un allenamento con carichi più elevati (cioè l’esecuzione di meno di cinque ripetizioni per serie a >85% del 1RM). Questo punto è importante da notare in quanto si riferisce ai contenuti discussi più avanti.
Prove nell’uomo che confutano l’ipertrofia sarcoplasmatica indotta dall’allenamento contro-resistenza:
Prima di discutere i dati sull’uomo che confutano il coinvolgimento dell’ipertrofia sarcoplasmatica nella crescita delle fibre, è importante discutere uno studio cardine sui roditori che ha preceduto il lavoro sull’uomo. Goldspink (1964) pubblicò un rapporto che coinvolgeva quattro gruppi di topi femmina. Due gruppi sono stati addestrati a procurarsi il cibo azionando un apparecchio a carrucola caricato a resistenza che aveva come obiettivo il muscolo bicipite brachiale; l’unica variabile che differenziava questi gruppi era la quantità di cibo somministrata al giorno (cioè 3,5 g/d e 5,0 g/d). Gli altri due gruppi non sono stati alloggiati con dispositivi a carrucola e sono serviti come gruppi di controllo con alimentazione corrispondente. Dopo 25 giorni di sperimentazione, tutti i topi sono stati soppressi e i muscoli bicipiti sono stati esaminati istologicamente al microscopio ottico con ingrandimento 1000×. Indipendentemente dalla quantità di cibo somministrata, i topi alloggiati con sistemi a carrucola possedevano fibre più grandi del 30% rispetto agli animali di controllo. È stata inoltre rilevata una forte relazione lineare tra la fCSA media e il numero di miofibrille nei topi alloggiati con sistemi a carrucola. Le fibre grandi, e presumibilmente ipertrofizzate, nei topi sottoposti a esercizio fisico possedevano anche meno spazio occupato dal sarcoplasma rispetto alle fibre più piccole. Per spiegare i suoi risultati, l’autore ha dichiarato (pag. 215):
È stato dimostrato che il numero di miofibrille per fibra aumenta in modo lineare con l’aumento del diametro della fibra. Ciò significa che l’aumento del numero di [mio]fibrille non procede allo stesso ritmo dell’aumento dell’area della sezione trasversale della fibra. Non è stato possibile misurare con precisione il diametro delle [mio]fibrille utilizzando la microscopia ottica, ma sembra molto probabile che la loro circonferenza aumenti con l’aumentare delle dimensioni della fibra.
Pur essendo limitati ai roditori, questi dati suggeriscono che una quantità apprezzabile di accrescimento di proteine miofibrillari accompagna l’ipertrofia delle fibre muscolari indotta dal carico contro-resistenza. Inoltre, i dati di Goldspink suggeriscono che le miofibrille aumentano in circonferenza piuttosto che in numero e che un leggero grado di impacchettamento delle miofibrille – o un aumento sproporzionato dell’accumulo di proteine miofibrillari rispetto alla crescita delle fibre – può essere coinvolto nell’ipertrofia delle fibre indotta dal carico. In seguito Goldspink ha scritto un eccellente capitolo del libro in cui discute i meccanismi di addizione delle miofibrille con la crescita delle fibre durante i periodi di allenamento (Goldspink, 2011). Ha citato numerosi studi sugli animali, molti dei quali sono stati eseguiti da Goldspink (2011), che suggeriscono: (i) la creazione di nuove miofibrille (cioè la miofibrillogenesi de novo) si verifica solo durante lo sviluppo embrionale, (ii) l’accrescimento proteico delle miofibrille negli animali sessualmente maturi comporta probabilmente l’aggiunta di proteine di nuova sintesi alla periferia delle miofibrille esistenti e (iii) le miofibrille possono proliferare durante i periodi di allenamento, anche se ciò è probabilmente dovuto alla scissione delle miofibrille.
Mentre il lavoro di Goldspink ha aggiunto enormi conoscenze sul comportamento delle miofibrille, finora nessuno studio sull’uomo ha fornito questo livello di dettaglio. In accordo con i risultati di Goldspink, tuttavia, esistono pochi dati sull’uomo che confutano l’idea che l’ipertrofia sarcoplasmatica indotta dall’allenamento contro-resistenza preceda o contribuisca alla crescita delle fibre o dei tessuti. Luthi et al. (1986) hanno utilizzato la TEM per esaminare gli adattamenti morfologici nelle fibre muscolari del vasto laterale di otto uomini di età universitaria non allenati che si sono allenati contro-resistenza per 6 settimane. Mentre l’area del vasto laterale è aumentata dell’8,4%, i valori di fCSA e la densità delle miofibrille sono rimasti invariati. Questi risultati suggeriscono che l’accumulo proporzionale di proteine miofibrillari accompagna la crescita muscolare durante l’allenamento. I dati TEM sono stati combinati con quelli della tomografia computerizzata a metà coscia per stimare che il volume assoluto occupato dalle miofibrille è aumentato del 10% in tutti i muscoli estensori della gamba. In particolare, non è stato delineato se questo aumento del contenuto di miofibrille si sia verificato attraverso la creazione di nuove miofibrille, l’allargamento di miofibrille preesistenti e/o la scissione di miofibrille. Inoltre, l’ipotesi che la morfologia acquisita tramite TEM si applichi a tutti i muscoli estensori della gamba ha dei limiti, dato che la TEM esamina una quantità estremamente ridotta di tessuto, come già detto. Infine, la mancanza di aumenti di fCSA con l’allenamento al posto di aumenti dell’area muscolare è difficile da riconciliare e si riferisce a una questione più ampia di mancanza di accordo tra i metodi utilizzati per valutare l’ipertrofia, come abbiamo discusso in precedenza (Haun et al., 2019c). Uno studio successivo dello stesso gruppo di ricerca ha esaminato le immagini TEM ad alto ingrandimento degli stessi partecipanti per determinare i cambiamenti nella spaziatura dei filamenti spessi e sottili con l’allenamento (Claassen et al., 1989). La distanza tra i filamenti di miosina era simile nei momenti precedenti e successivi all’intervento. Questi risultati forniscono un’ulteriore prova che l’accumulo di proteine miofibrillari è proporzionale alla crescita del tessuto. In quest’ultima relazione sono stati utilizzati anche dati di tomografia computerizzata ottenuti in precedenza per dimostrare che la densità radiologica dei muscoli della mezza coscia aumentava con l’allenamento. È interessante notare che i ricercatori hanno attribuito quest’ultimo risultato a un potenziale aumento dell’impacchettamento delle miofibrille. Sebbene sia una conclusione ragionevole, questo risultato potrebbe anche essere legato a una diminuzione dei lipidi intramuscolari o a un aumento del contenuto di proteine non contrattili (ad esempio, proteine della matrice extracellulare). Inoltre, la presunzione degli autori che si sia verificato un impacchettamento delle miofibrille è in conflitto con i loro precedenti dati TEM che suggeriscono che, mentre il contenuto di miofibrille è aumentato in termini assoluti, la densità e la spaziatura delle miofibrille sono state conservate. Uno studio di Trappe et al. (2000) ha esaminato i cambiamenti di tensione specifici nelle fibre muscolari isolate del vasto laterale di sette uomini anziani precedentemente non allenati dopo 12 settimane di allenamento contro-resistenza. Sebbene le fibre di tipo I e IIa abbiano mostrato un aumento del 20% e del 13% dei valori di fCSA, i valori di tensione specifica in entrambi i tipi di fibre erano simili ai punti temporali precedenti e successivi. Analogamente a quanto riportato da Luthi et al. (1986), questi risultati suggeriscono che l’accrescimento proporzionale delle proteine miofibrillari accompagna la crescita delle fibre. Il gruppo di Trappe ha pubblicato uno studio di follow-up su sette donne anziane non allenate che si sono impegnate in 12 settimane di allenamento contro-resistenza (Trappe et al., 2001). A differenza dei maschi nel loro precedente studio, le fibre di tipo IIa non si sono ipertrofizzate. Inoltre, i confronti statistici hanno indicato che i valori di tensione specifica di entrambi i tipi di fibre non sono stati influenzati dall’allenamento. Questi risultati suggeriscono che l’allenamento non ha influenzato la morfologia delle fibre di tipo IIa e che la crescita radiale delle fibre di tipo I è stata accompagnata da un aumento proporzionale delle proteine miofibrillari. Risultati simili sono stati riportati da altri ricercatori in sei uomini precedentemente non allenati, sottoposti a 12 settimane di allenamento contro-resistenza (Widrick et al., 2002); in particolare, l’ipertrofia delle fibre del muscolo vasto laterale si è verificata senza alterazioni dei valori di tensione specifica delle fibre isolate.
Più di recente, un gruppo di ricerca ha esaminato gli adattamenti tissutali del vasto laterale che si sono verificati in maschi in età universitaria precedentemente non allenati, sottoposti a 12 settimane di allenamento contro- resistenza (Roberts et al., 2018). I partecipanti sono stati suddivisi in rispondenti alti e bassi in base a un punteggio composito delle variazioni da prima a dopo l’intervento della fCSA media, della massa di tessuto magro totale del corpo determinata con l’assorbimetria a raggi X a doppia energia e dello spessore del vasto laterale. I 13 soggetti ad alta risposta hanno registrato un aumento del 34% della fCSA media e del 24% dello spessore del vasto laterale. Tuttavia, questi individui non hanno registrato cambiamenti nelle abbondanze di proteine della catena pesante della miosina e dell’actina per milligrammo di tessuto umido, e lo stesso è stato riscontrato per i 12 soggetti a bassa risposta anabolica, che non hanno mostrato praticamente alcun cambiamento nella fCSA media. Analogamente a molti degli studi precedenti, questi risultati suggeriscono che l’accrescimento proteico delle miofibrille era proporzionale alla crescita delle fibre e dei tessuti nei soggetti ad alta risposta. Nel complesso, questi studi suggeriscono in larga misura che la crescita della fibra muscolare o del tessuto in risposta a 6-12 settimane di allenamento contro-resistenza avviene attraverso un’ipertrofia convenzionale piuttosto che sarcoplasmatica. In relazione alle ipotesi formulate più avanti, è importante notare che tutti questi studi hanno esaminato partecipanti non allenati in precedenza.
Anche diversi studi sui traccianti mettono in dubbio la veridicità dell’ipertrofia sarcoplasmatica. Ad esempio, Moore et al. (2009) hanno riferito che un allenamento contro-resistenza provoca forti aumenti dei tassi di sintesi proteica miofibrillare a digiuno e minime alterazioni dei tassi di sintesi proteica sarcoplasmatica in una finestra di 5 ore dopo l’esercizio. Inoltre, questi autori hanno riferito che i tassi di sintesi proteica sarcoplasmatica erano più reattivi all’apporto di nutrienti dopo l’esercizio piuttosto che all’esercizio stesso. Gasier et al. (2012) hanno pubblicato risultati simili utilizzando il metodo del tracciante D2O per dimostrare che un allenamento contro-resistenza della parte inferiore del corpo ha stimolato un aumento dei tassi di sintesi proteica miofibrillare, pur non influenzando i tassi di sintesi proteica complessiva in una finestra di 16 ore dopo l’esercizio. Anche Wilkinson et al. (2014) hanno utilizzato il metodo del tracciante D2O per dimostrare che 4 giorni di allenamento di resistenza unilaterale delle gambe per un periodo di 8 giorni hanno aumentato significativamente i tassi di sintesi proteica miofibrillare, senza influenzare i tassi di sintesi proteica sarcoplasmatica. Allo stesso modo, altri hanno utilizzato il metodo del tracciante D2O per dimostrare che i tassi di sintesi proteica miofibrillare sono elevati settimane dopo l’inizio dell’allenamento contro-resistenza (Brook et al., 2015). Questi dati suggeriscono complessivamente che il pool di proteine miofibrillari, piuttosto che quello sarcoplasmatico, potrebbe subire l’espansione più forte all’inizio dell’allenamento. Tuttavia, anche in questo caso, tutti questi studi sono stati condotti su partecipanti non allenati.
Oltre a questi interventi a breve termine, esistono anche interventi cronici e confronti trasversali che mettono legittimamente in dubbio il coinvolgimento dell’ipertrofia sarcoplasmatica nell’ipertrofia muscolare a lungo termine. Alway et al. (1988) hanno utilizzato la TEM per segnalare che lo spazio intracellulare occupato dalle miofibrille nelle fibre muscolari del gastrocnemio era simile tra soggetti allenati contro-resistenza e sedentari, nonostante i valori di fCSA fossero maggiori nei soggetti allenati contro-resistenza. Shoepe et al. (2003) hanno confrontato le caratteristiche delle fibre del vasto laterale di sei uomini di età universitaria non allenati rispetto a sei ben allenati (l’età di allenamento dichiarata era in media di ∼7 anni). Mentre i valori di fCSA di tipo I e II erano significativamente maggiori nella coorte ben allenata, i valori di tensione specifica erano simili tra i gruppi, suggerendo un’ipertrofia a lungo termine delle fibre con un accrescimento proporzionale di proteine miofibrillari. I due studi citati suggeriscono che la crescita delle fibre a lungo termine con l’allenamento contro-resistenza avviene attraverso l’ipertrofia convenzionale. Tuttavia, questi dati sono in contrasto con altri studi. Pansarasa et al. (2009) hanno esaminato le caratteristiche delle fibre del vasto laterale in cinque donne di età universitaria che si sono allenate contro-resistenza per un anno. La tensione specifica è aumentata significativamente nelle fibre di tipo I e II, mentre i valori medi di fCSA non sono cambiati da prima a dopo l’intervento. Sebbene siano difficili da conciliare, questi risultati supportano un modello di ipertrofia a lungo termine in cui si verifica un impacchettamento delle miofibrille senza un aumento dell’ipertrofia delle fibre muscolari. E’ stato recentemente esaminato lo spessore del vasto laterale e le caratteristiche del tessuto in sei uomini di età universitaria ben allenati (l’età di allenamento dichiarata era in media di ∼10 anni) rispetto a sei uomini non allenati (Vann et al., 2020b). Mentre lo spessore del muscolo era maggiore del 22% (p < 0,05) nella coorte allenata, anche le abbondanze di catena pesante di miosina e actina per milligrammo di muscolo umido erano maggiori del ∼9% (p < 0,05). I valori di fCSA di tipo I e II erano numericamente maggiori, ma statisticamente simili tra le coorti. D’Antona et al. (2006) hanno anche riportato che i valori di tensione specifica delle fibre di tipo II erano significativamente maggiori nei bodybuilder ben allenati rispetto ai controlli allenati a livello ricreativo. Anche in questo caso, i risultati di entrambi gli studi suggeriscono che l’ipertrofia a lungo termine è accompagnata dall’impaccamento delle miofibrille. Contrariamente alle implicazioni di tutti gli studi citati, due studi su body-builder ben allenati suggeriscono che sono evidenti le caratteristiche dell’ipertrofia sarcoplasmatica (Macdougall et al., 1982; Meijer et al., 2015). Nel complesso, questi risultati discordanti rendono difficile stabilire come l’allenamento contro-resistenza a lungo termine influisca sulla densità delle miofibrille e/o se l’ipertrofia sarcoplasmatica sia un processo coinvolto quando gli individui diventano più allenati. È plausibile che dosi diverse di allenamento contro-resistenza nel corso di anni di allenamento possano provocare cambiamenti diversi nella morfologia muscolare. Si tratta di un’area non risolta e sono necessarie ulteriori ricerche per comprendere meglio i risultati adattativi a livello cellulare in risposta a diversi stili di allenamento.
Creare un paradigma Considerando l’evidenza collettiva:
Considerando la letteratura citata, è difficile costruire un paradigma collettivo di crescita delle fibre muscolari che coinvolga l’ipertrofia sarcoplasmatica. Tuttavia, possiamo ipotizzare che ci siano tre scenari candidati in cui l’ipertrofia sarcoplasmatica potrebbe manifestarsi con l’allenamento contro-resistenza. In particolare, si ritiene che l’ipertrofia sarcoplasmatica sia:
un sintomo transitorio dell’edema indotto dall’allenamento,
un meccanismo transitorio di crescita delle fibre muscolari e/o
il risultato del raggiungimento di una soglia di accrescimento delle proteine miofibrillari in individui ben allenati.
L’edema tissutale è associato a un danno localizzato e si manifesta come una ritenzione di liquido interstiziale tra le cellule. Specificamente al danno muscolare indotto dall’esercizio fisico, l’edema può essere accompagnato da un danno al sarcolemma in cui si verifica un’infiltrazione di grandi proteine del siero (ad esempio, albumina) e di liquido all’interno e intorno alle fibre muscolari (Valle et al., 2013). In queste circostanze, sembra plausibile che l’edema possa elevare artificialmente i valori di fCSA attraverso la ritenzione di liquidi intracellulari, che si manifesterebbe come un’espansione del sarcoplasma. A sostegno di questa ipotesi, Yu et al. (2013) hanno esaminato l’effetto di una corsa prolungata al piano inferiore sulla fCSA media del muscolo soleo in uomini di età universitaria. Questo studio è stato limitato dal fatto che non sono state ottenute biopsie ripetute dai partecipanti; in particolare, 16 uomini hanno eseguito l’allenamento e le biopsie sono state ottenute da un sottogruppo di partecipanti a 1 ora dopo l’esercizio, 2-3 giorni dopo l’esercizio e 7-8 giorni dopo l’esercizio. Questo studio è stato inoltre limitato dal fatto che la corsa ad alto volume al piano di sotto ha poca somiglianza con l’esercizio contro-resistenza. Ciononostante, i valori medi di fCSA erano superiori del 30% nelle biopsie ottenute 7-8 giorni dopo l’esercizio rispetto a quelle ottenute nei momenti precedenti e gli autori hanno ipotizzato che ciò fosse principalmente associato a un edema localizzato. Sebbene sia informativo, è difficile determinare se l’ipertrofia sarcoplasmatica fosse evidente, dato che non sono state eseguite interrogazioni ultrastrutturali. Tuttavia, questo studio dimostra che i cambiamenti dell’fCSA nei giorni successivi al danno muscolare sono dovuti all’edema, che probabilmente si manifesta con un’espansione del sarcoplasma. Le valutazioni non invasive dell’edema possono essere eseguite anche acquisendo immagini muscolari con ultrasuoni o risonanza magnetica (RM) e quantificando l’intensità dell’eco B-mode o l’intensità del segnale nelle immagini pesate in T2, rispettivamente. Damas et al. (2016c) hanno monitorato i cambiamenti nell’intensità dell’eco B-mode del vasto laterale in uomini non allenati di età universitaria prima, dopo 3 settimane e dopo un intervento di allenamento contro-resistenza di 10 settimane. Mentre l’aumento dell’area muscolare è stato registrato a 3 settimane e dopo l’allenamento, l’aumento dell’edema (cioè l’intensità del segnale B-mode/area del vasto laterale) è stato riscontrato solo a 3 settimane. Gli autori hanno concluso che l’ipertrofia muscolare era confusa dall’edema a 3 settimane, ma non a 10 settimane dall’allenamento. Questo articolo ha anche portato a discutere se l’ipertrofia muscolare possa essere valutata accuratamente senza gli effetti confondenti dell’edema nel primo mese di allenamento in partecipanti precedentemente non allenati (Damas et al., 2016b; Defreitas et al., 2016). Quando si mettono in relazione questi risultati con la nozione di ipertrofia sarcoplasmatica indotta dall’allenamento, è plausibile che questo fenomeno possa essere osservato sia in:
individui non allenati durante il primo mese di allenamento, sia
individui allenati esposti a paradigmi di allenamento non abituali.
Il lavoro del laboratorio di Haun et al. (2019b), che ha dimostrato che l’allenamento facilita gli aspetti dell’ipertrofia sarcoplasmatica, si allinea con quest’ultima ipotesi poiché i partecipanti, pur essendo ben allenati, hanno eseguito volumi di allenamento straordinariamente elevati e non abituali. A ulteriore sostegno di questa tesi vi sono dati precedenti relativi a questi partecipanti che dimostrano che l’aumento del contenuto di fluido extracellulare nel corpo intero (valutato mediante spettroscopia di impedenza bioelettrica, o BIS) ha contribuito in modo significativo all’aumento delle variazioni della massa muscolare nel corpo intero con l’allenamento (Haun et al., 2018). Sebbene in precedenza avessimo sostenuto che l’edema non confondesse le variabili di risultato in questo studio (Haun et al., 2019b), i dati BIS sopra citati forniscono la prova del contrario. Pertanto, in futuro, i ricercatori dovrebbero considerare gli effetti confondenti dell’edema quando misurano i surrogati grossolani dell’ipertrofia muscolare e i cambiamenti morfologici o biochimici nel tessuto muscolare. Inoltre, l’influenza dello stato di idratazione sulle macro- e micro-variabili legate all’ipertrofia dovrebbe continuare a essere esplorata, dato che sono stati segnalati spostamenti di liquidi durante i periodi cronici di allenamento di resistenza (ad esempio, acqua intramuscolare e volume plasmatico) (Reidy et al., 2017).
Oltre a essere un potenziale sintomo di edema, si propone che l’ipertrofia sarcoplasmatica possa essere un meccanismo che contribuisce alle prime fasi di crescita delle fibre radiali. È comune che l’fCSA medio aumenti del ∼15-30% in risposta a mesi di allenamento contro-resistenza (rivisto in Grgic e Schoenfeld, 2018). Questi risultati suggeriscono che le fibre muscolari devono generare una quantità apprezzabile di spazio intracellulare per accumulare più proteine miofibrillari. Un potenziale meccanismo per realizzare tale crescita potrebbe coinvolgere l’ipertrofia sarcoplasmatica seguita dal “riempimento” dello spazio sarcoplasmatico appena generato con proteine miofibrillari. Questo meccanismo contrasta con la nozione avanzata da Phillips (Phillips, 2000), secondo cui l’impacchettamento delle miofibrille precede l’ipertrofia e il nuovo materiale miofibrillare aggiunto “spinge” il sarcolemma verso l’esterno. Sebbene nessuno dei due meccanismi abbia prove definitive a sostegno nell’uomo, i dati raccolti in altre linee cellulari suggeriscono che l’ipertrofia sarcoplasmatica possa essere favorita durante la crescita delle fibre muscolari. A questo proposito, Neurohr et al. (2019) hanno recentemente dimostrato che l’espansione citoplasmatica si verifica prontamente in lievito e cellule primarie di mammifero attraverso l’interruzione di un gene (Cdc28). Gli autori hanno anche notato che la biosintesi di RNA e proteine non è aumentata con l’aumento della crescita cellulare e hanno ipotizzato che questo fenomeno sia dovuto al carico trascrizionale imposto ai nuclei. Questo lavoro è limitato dal fatto che la crescita cellulare negli organismi intatti è vincolata dal tessuto connettivo, mentre questo vincolo non è applicabile nelle colture monostrato. Tuttavia, questi risultati dimostrano in linea di principio che l’espansione citoplasmatica è un meccanismo conservato di ipertrofia cellulare nel lievito e in numerose cellule di mammifero. I dati di Neurohr et al. (2019) ribadiscono inoltre la nozione che le cellule possiedono un rapporto ottimale tra DNA e volume cellulare, poiché i processi di biosintesi dei ribosomi, dell’mRNA e delle proteine sono ad alta intensità di risorse (rivisti in Gillooly et al., 2015). Sebbene le fibre muscolari siano multinucleate, la teoria del dominio mionucleare si basa su un presupposto simile (Allen et al., 1999): in particolare, un aumento delle dimensioni delle fibre muscolari richiede l’aggiunta di mionuclei dalle cellule satelliti per sostenere l’accrescimento e il mantenimento del materiale intracellulare appena aggiunto. Pertanto, le fibre muscolari potrebbero crescere preferenzialmente attraverso l’ipertrofia sarcoplasmatica all’inizio di un intervento di allenamento, dato che:
questo meccanismo sembra essere conservato in diversi tipi di cellule, come discusso in precedenza,
l’accrescimento dei mionuclei ritarda all’inizio dell’allenamento e alcuni studi hanno riportato che la crescita delle fibre avviene senza accrescimento dei mionuclei (rivisto in Murach et al,
se la crescita delle fibre avvenisse attraverso l’ipertrofia convenzionale o l’impacchettamento delle miofibrille senza l’accrescimento dei mionuclei, allora i mionuclei residenti sarebbero probabilmente sollecitati a coordinare la produzione su larga scala dei ribosomi, degli mRNA che codificano le grandi proteine delle miofibrille e di altri mRNA necessari per l’omeostasi cellulare. Questo concetto è ulteriormente illustrato nella seguente.
Due meccanismi ipotetici di ipertrofia delle fibre muscolari. Nell’ipotesi di espansione del sarcoplasma proposta dal modello di ipertrofia, le fibre muscolari crescono prima attraverso l’ipertrofia sarcoplasmatica. Ciò consente ai domini mionucleari residenti di prepararsi spazialmente, bioenergeticamente e biosinteticamente all’accumulo di ulteriori miofibrille. Sebbene i domini residenti possano sintetizzare e accumulare alcune proteine delle miofibrille durante questa fase dell’ipertrofia, ipotizziamo che la predominanza dell’ipertrofia sarcoplasmatica riduca il carico trascrizionale sui domini residenti. Una volta che l’ipertrofia sarcoplasmatica porta a una crescita cellulare apprezzabile, un ulteriore dominio mionucleare viene acquisito attraverso la fusione delle cellule satelliti. Questo nuovo dominio mionucleare può quindi contribuire in modo apprezzabile all’accrescimento delle proteine della miofibrilla, “riempiendo” lo spazio creato dall’espansione del sarcoplasma. Nell’ipotesi dell’espansione delle miofibrille nel modello dell’ipertrofia, le fibre muscolari sintetizzano e accumulano rapidamente proteine miofibrillari. Questo fenomeno porta all’impacchettamento delle miofibrille che agisce spingendo il sarcolemma verso l’esterno. Le fibre raggiungono infine una soglia dimensionale, che porta all’acquisizione di un ulteriore dominio mionucleare attraverso la fusione delle cellule satelliti. A questo punto entrambi i domini possono operare per mantenere e sintetizzare nuove miofibrille.
In relazione alla figura sopra esposta, l’ipotesi di espansione del sarcoplasma dell’ipertrofia muscolare durante l’allenamento contro-resistenza prevede quanto segue:
Prima dell’aggiunta mionucleare mediata dalle cellule satelliti, le fibre muscolari subiscono un priming spaziale attraverso l’ipertrofia sarcoplasmatica, un priming bioenergetico attraverso l’aumento della regolazione degli enzimi necessari per la generazione di ATP e un priming della biosintesi proteica attraverso la biogenesi dei ribosomi. Questi fenomeni preparano spazialmente, “bioenergeticamente” e “biosinteticamente” le fibre per l’accrescimento proteico delle miofibrille, gestendo efficacemente il carico trascrizionale sui mionuclei residenti.
Una volta che si verifica l’addizione mionucleare mediata dalle cellule satelliti, i nuovi domini mionucleari possono facilitare una significativa crescita della sintesi di mRNA associati alle miofibrille e di proteine delle miofibrille. Questo è il riempimento dello spazio del sarcoplasma con componenti contrattili.
In relazione ai fenomeni discussi sopra, è da notare che i partecipanti che hanno mostrato segni di ipertrofia sarcoplasmatica nel nostro studio sull’allenamento a volume più elevato non hanno mostrato aumenti nel numero di mionuclei per fibra (Haun et al., 2019a, b). Pertanto, anche se in via speculativa, un motivo per cui si è verificata l’ipertrofia sarcoplasmatica potrebbe essere legato al carico trascrizionale imposto ai mionuclei residenti senza l’accrescimento dei mionuclei, come discusso in precedenza. Questi partecipanti hanno anche dimostrato un aumento della biogenesi dei ribosomi e dell’espressione proteica degli enzimi sarcoplasmatici responsabili della generazione di ATP attraverso le vie ATP-PCr e glicolitica. Quest’ultima osservazione potrebbe essere correlata agli aspetti della biosintesi proteica e del priming bioenergetico dell’ipertrofia sarcoplasmatica discussi in precedenza. Sebbene questi dati siano convincenti, sono necessarie ulteriori ricerche per convalidare questo modello.
Infine, si ipotizza che un terzo scenario che coinvolge l’ipertrofia sarcoplasmatica possa comportare il raggiungimento di una soglia di accrescimento di proteine miofibrillari nelle fibre muscolari di grandi dimensioni con anni di allenamento contro-resistenza. Si ritiene che questa modalità di ipertrofia si verifichi per ragioni simili a quelle discusse in precedenza; in particolare, lo stress trascrizionale imposto ai mionuclei residenti nelle fibre muscolari di grandi dimensioni favorirebbe la crescita cellulare attraverso l’ipertrofia sarcoplasmatica. Questa ipotesi è ulteriormente illustrata nella figura seguente.
Questa figura avanza l’ipotesi che l’ipertrofia sarcoplasmatica con anni di allenamento contro-resistenza possa essere dovuta al superamento della soglia di accrescimento delle miofibrille da parte delle fibre muscolari di grandi dimensioni. Questo fenomeno è simile alla teoria dello stress trascrizionale presentata nella Figura 3. In particolare, proponiamo che le cellule muscolari crescano attraverso l’accrescimento delle miofibrille fino a una certa soglia. In seguito, le fibre non possono più aggiungere miofibrille a causa di uno stress significativo per i domini mionucleari residenti. Sulla base di prove limitate citate nel testo, proponiamo che questo meccanismo sia evidente nei soggetti che si impegnano per anni in allenamenti ad alto volume (ad esempio, i culturisti).
Anche in questo caso, si tratta di un’ipotesi speculativa, dato che, come per le altre due ipotesi citate, non esistono praticamente dati a supporto. Inoltre, attualmente non si sa se esista una soglia di accrescimento delle miofibrille nelle fibre muscolari. A sostegno di questa ipotesi, tuttavia, altri ricercatori hanno ipotizzato che nelle cellule mitoticamente attive si verifichi una soglia di accrescimento proteico, che agisce per innescare la divisione cellulare (Soltani et al., 2016). Poiché le miofibre sono cellule post-mitotiche che non possono dividersi, l’ipertrofia sarcoplasmatica potrebbe essere l’unico modo attraverso il quale le fibre di grandi dimensioni sono in grado di crescere. In effetti, questa è una potenziale spiegazione del perché alcuni degli studi citati hanno riportato caratteristiche di ipertrofia sarcoplasmatica in body-builder con anni di esperienza di allenamento (Macdougall et al., 1982; Meijer et al., 2015). Questo potrebbe anche essere il motivo per cui si sono osservate caratteristiche di ipertrofia sarcoplasmatica in individui precedentemente allenati e sottoposti a nuovi paradigmi di allenamento (Haun et al., 2019b; Vann et al., 2020a). A parte ciò, è stata avanzata un’altra teoria per spiegare il comportamento delle grandi fibre muscolari in risposta a stimoli ipertrofici. Murach et al. (2019) hanno ipotizzato che le fibre muscolari dei mammiferi (e le miofibrille all’interno delle fibre) siano in grado di dividersi in risposta a un sovraccarico, e gli autori hanno fornito dati umani e animali che supportano queste ipotesi [esempi includono riferimenti (Goldspink, 1970; Timson et al., 1985; Antonio e Gonyea, 1994)]. Non è ancora noto se una soglia di accumulo di proteine miofibrillari all’interno di una fibra muscolare inneschi la scissione delle fibre. Inoltre, al di là dell’analisi bidimensionale su sezioni seriali tramite istologia (Eriksson et al., 2005, 2006), la scissione delle fibre nell’uomo non è stata dimostrata in modo convincente. Altri ricercatori hanno anche fornito prove convincenti che suggeriscono che la scissione delle fibre percepita nei muscoli dei roditori sottoposti a sovraccarico è dovuta principalmente ad alterazioni dell’angolo di pennato e della lunghezza delle fibre piuttosto che alla scissione (Jorgenson e Hornberger, 2019). È chiaro che c’è ancora molto da risolvere per quanto riguarda la risposta fisiologica delle grandi miofibre agli stimoli ipertrofici.
Sebbene il modello di ablazione sinergica nei roditori abbia i suoi limiti (ad esempio, una crescita muscolare rapida e non fisiologica in risposta a uno stimolo di crescita costante), tali esperimenti sui roditori possono fornire informazioni utili per sostenere o confutare i meccanismi teorici proposti sopra. In particolare, i ricercatori hanno già riflettuto sullo stress trascrizionale suscitato sui mionuclei residenti durante l’ipertrofia indotta dal sovraccarico. Il gruppo di McCarthy (Kirby et al., 2016) ha somministrato 5-etinil uridina a topi wild-type con l’intento di etichettare la trascrizione di RNA nascente in risposta all’ipertrofia indotta da sovraccarico. Questi autori hanno esaminato diversi gruppi di topi, tra cui controlli sham-operated (CTL) o topi esposti a tre, sette o 14 giorni di sovraccarico delle plantari tramite ablazione sinergica (SA). Si è osservato un aumento lineare del peso delle plantari nei gruppi in cui CTL < SA di 3 giorni < SA di 7 giorni < SA di 14 giorni. Tuttavia, la trascrizione nascente era regolata solo nei topi con SA di 3 giorni rispetto a quelli con CTL. Questi dati suggeriscono che, sebbene i mionuclei residenti presentino una riserva trascrizionale durante l’ipertrofia, questa risposta è rapida, pulsatile e di durata limitata. È stato inoltre riportato che la trascrizione nascente è più bassa nelle fibre più grandi durante la SA. Questa scoperta è in linea con quella di Neurohr et al. (2019), che hanno dimostrato che la biosintesi dell’RNA non è in scala nelle cellule di mammifero che crescono attraverso l’ipertrofia citoplasmatica in vitro. Resta da stabilire se i risultati di Kirby et al. (2016) indichino o meno che le fibre grandi e ipertrofiche crescono prevalentemente attraverso l’ipertrofia sarcoplasmatica. Tuttavia, riteniamo che questi parallelismi tra gli studi siano interessanti e meritevoli di ulteriori approfondimenti. Riteniamo inoltre che il modello murino Pax7-DTA possa essere informativo per determinare se l’ipertrofia sarcoplasmatica accompagni la crescita delle fibre durante le prime fasi dell’ipertrofia indotta da sovraccarico. I topi Pax7-DTA sono geneticamente modificati in modo tale che, quando viene loro somministrato tamoxifene, mostrano un’ablazione quasi completa delle cellule satelliti a causa di eventi mediati da Cre-ER che guidano l’espressione del frammento A della tossina difterica (DTA) nelle cellule Pax7+ (Lepper et al., 2011). Mccarthy et al. (2011) hanno dimostrato che, mentre l’accrescimento mionucleare è abrogato in questi topi durante l’ipertrofia plantare indotta dall’ablazione sinergica, la crescita delle fibre non è stata compromessa. Se le ipotesi di cui sopra fossero vere per quanto riguarda lo stress dei mionuclei residenti durante la crescita delle fibre indotta dal sovraccarico, allora è possibile che l’ipertrofia sarcoplasmatica si sia verificata maggiormente nei topi a cui è stato somministrato il Tamoxifene (cioè quelli che non mostravano accrescimento di mionuclei) rispetto ai topi a cui non è stato somministrato il Tamoxifene (cioè quelli che mostravano accrescimento di mionuclei). Tuttavia, in questi animali non è stata valutata la morfologia muscolare. Allo stesso modo, altri hanno dimostrato che la crescita delle fibre plantari in risposta all’ablazione sinergica è ridotta nei topi Pax7-DTA a cui è stato somministrato Tamoxifene (Egner et al., 2016). Alla luce delle nostre ipotesi e di questi dati contrastanti sul modello Pax7-DTA, sono giustificati futuri esperimenti che esaminino gli adattamenti morfologici che si verificano in questo modello murino.
Implicazioni più ampie:
Il tema dell’ipertrofia sarcoplasmatica esiste da decenni in ambito atletico. Stone ne parlò come di un potenziale adattamento all’allenamento all’inizio degli anni ’80 (Stone et al., 1983) e Kraemer e Zatsiorsky (2006) ipotizzarono che l’ipertrofia convenzionale e quella sarcoplasmatica derivino rispettivamente da allenamenti ad alto e basso carico (Kraemer e Zatsiorsky, 2006). Sebbene queste ipotesi siano in una certa misura speculative, esistono dati a sostegno dei paradigmi di carico volumetrico che influenzano la densità miofibrillare. A questo proposito, due degli studi citati in precedenza che hanno osservato caratteristiche di ipertrofia sarcoplasmatica sono stati condotti su culturisti ben allenati (Macdougall et al., 1982; Meijer et al., 2015), e questa popolazione si allena generalmente con volumi più elevati. Al contrario, esistono caratteristiche di impacchettamento delle miofibrille negli atleti di potenza (Meijer et al., 2015; Fluck et al., 2019), e questa popolazione si allena generalmente con carichi più elevati e volumi inferiori. Anche i due studi citati del nostro laboratorio supportano questo modello (Haun et al., 2019b; Vann et al., 2020a), ed è da notare che una riduzione della densità dell’impaccamento dei miofilamenti attraverso l’ipertrofia sarcoplasmatica può anche promuovere alcuni adattamenti dell’allenamento, come l’aumento della velocità di accorciamento delle fibre muscolari (Metzger e Moss, 1987).
Anche la rilevanza mainstream dell’ipertrofia sarcoplasmatica con l’allenamento di resistenza sta prendendo piede. Dal 2014 sono stati pubblicati diversi articoli online per i non addetti ai lavori che parlano dell’ipertrofia sarcoplasmatica con l’allenamento alla resistenza. Allo stesso modo, la più grande enciclopedia online del mondo1 discute le sfumature relative all’ipertrofia miofibrillare rispetto a quella sarcoplasmatica in un articolo principale intitolato Ipertrofia muscolare, nonostante nessuno dei due meccanismi sia stato ampiamente validato. È quindi evidente che il pubblico generale è interessato a saperne di più su questo aspetto della fisiologia muscolare. Per il futuro, più laboratori dovranno perseguire questa domanda di ricerca per convalidare i risultati precedenti e fornire scoperte significative.
Determinare se l’ipertrofia sarcoplasmatica sia un “unicorno” scientifico o un adattamento all’allenamento di resistenza è fondamentale per il campo per diversi motivi. A tal fine, alcuni hanno ipotizzato che la vera ipertrofia si verifichi solo quando l’accrescimento delle proteine miofibrillari è proporzionale all’aumento della fCSA (Damas et al., 2015; Taber et al., 2019). Si tratta di un’ipotesi interessante per diversi motivi. In primo luogo, altri hanno notato che l’ipertrofia muscolare scheletrica indotta dall’allenamento contro-resistenza non è fortemente associata all’aumento della forza (Ahtiainen et al., 2016). In altre parole, muscoli più grandi dopo l’allenamento non producono un aumento proporzionale della forza. Questo è probabilmente vero per una serie di ragioni, molte delle quali sono state spiegate da Taber et al. (2019). Per ribadire un punto chiave di Taber et al. (2019), il ruolo che l’ipertrofia sarcoplasmatica può avere nella spiegazione di questa relazione imperfetta, per quanto piccolo, dovrebbe essere ulteriormente esaminato. Come già detto, alcuni risultati della nostra ricerca suggeriscono che l’allenamento ad alto volume promuove l’ipertrofia sarcoplasmatica in misura maggiore rispetto all’allenamento ad alto carico. La definizione di ciascuna modalità di allenamento è intrinsecamente difficile. Ad esempio, alcuni considerano l’esecuzione di serie contenenti più di 10 ripetizioni al ∼60-70% di 1RM come un allenamento a più alto volume, mentre altri possono considerare l’allenamento ad alto volume come l’esecuzione di serie di alte ripetizioni fino al cedimento utilizzando il 30% di 1RM. Pertinenti ai temi qui discussi sono i risultati di Mitchell et al. (2012), che hanno riferito che l’allenamento di resistenza degli estensori delle gambe all’80% dell’1RM e al 30% dell’1RM ha prodotto aumenti simili dell’area della sezione trasversale del muscolo della mezza coscia e dell’fCSA del muscolo. Sebbene le caratteristiche ipertrofiche fossero simili con ogni forma di allenamento, indagare sul modo in cui ogni forma di allenamento influisce sulla morfologia muscolare sarà utile alla luce di alcuni risultati delle nostre ricerche citati in precedenza. Altri dati interessanti che possono essere correlati all’ipertrofia sarcoplasmatica provengono da Kadi et al. (2004) che hanno ottenuto biopsie seriali del vasto laterale da partecipanti non allenati durante un intervento di allenamento contro-resistenza di 90 giorni seguito da un periodo di disallenamento di 90 giorni. Dopo 90 giorni di allenamento, la fCSA media è aumentata del 16% rispetto ai valori precedenti all’allenamento. Tre giorni di detraining hanno aumentato numericamente la fCSA del ∼3% rispetto al punto di 90 giorni e del 19% rispetto al pre-training. È sorprendente che i valori medi di fCSA siano tornati ai livelli pre-allenamento dopo soli altri sette giorni di disallenamento. In sostanza, questi dati suggeriscono che durante il detraining si verifica un rapido decadimento dei valori medi di fCSA. Abbiamo sostenuto che, se le fibre muscolari avessero subito una significativa accrescimento miofibrillare, ci sarebbe voluto più tempo per osservare l’atrofia indotta dal detraining (Roberts et al., 2018). Tuttavia, se l’ipertrofia sarcoplasmatica ha contribuito in modo significativo alla crescita delle fibre, rimane plausibile che il ripristino dello spazio sarcoplasmatico ai valori precedenti all’allenamento possa aver contribuito in larga misura al fenomeno del detraining. Quindi, ancora una volta, l’utilizzo di metodi per valutare la morfologia cellulare in questi scenari fornirà informazioni utili in relazione agli adattamenti da detraining. Infine, ci sono i dati sull’allenamento di resistenza a lungo termine pubblicati da Fatouros et al. (2005), in cui si è tentati di ipotizzare che alcuni risultati dello studio possano essere dovuti, in parte, all’ipertrofia sarcoplasmatica. Questi autori hanno diviso uomini anziani precedentemente non allenati in un gruppo di allenamento a bassa intensità (n = 18) e in un gruppo di allenamento ad alta intensità (n = 20). Entrambi i gruppi hanno eseguito un allenamento di resistenza per tutto il corpo per 6 mesi, ma il primo gruppo ha utilizzato pesi corrispondenti a ∼55% 1RM mentre il secondo gruppo ha utilizzato pesi corrispondenti a ∼82% 1RM. La forza della parte superiore e inferiore del corpo è stata misurata utilizzando gli esercizi di chest press e leg press, rispettivamente, prima e dopo l’allenamento. Tutti i partecipanti si sono poi staccati per 48 settimane e la forza è stata nuovamente valutata. I partecipanti al gruppo ad alta intensità hanno acquisito più forza con l’allenamento, come era prevedibile. Tuttavia, i partecipanti al gruppo ad alta intensità hanno mantenuto una forza maggiore rispetto a quelli del gruppo a bassa intensità dopo il lungo periodo di disallenamento. Anche le circonferenze a metà coscia e le pieghe della pelle sono state ottenute in tutti i punti temporali sopra menzionati, ma non sono state notate differenze degne di nota nella crescita o nell’atrofia muscolare tra i gruppi; ciò è probabilmente dovuto alla risoluzione relativamente scarsa che le valutazioni delle pieghe della pelle e delle circonferenze forniscono. Sebbene questi risultati possano essere in parte legati agli adattamenti neurali a lungo termine con l’allenamento a carichi più elevati, è possibile ipotizzare che l’allenamento di resistenza a carichi più elevati possa anche aver favorito una maggiore accumulazione di proteine miofibrillari rispetto all’allenamento a carichi più bassi, viste le argomentazioni esposte in precedenza. A questo proposito, è opportuno replicare studi come questi, in cui le biopsie vengono analizzate per individuare le caratteristiche dell’ipertrofia convenzionale rispetto a quella sarcoplasmatica.
Conclusioni:
Le ricerche suggeriscono che possono verificarsi diversi adattamenti morfologici con l’ipertrofia delle fibre muscolari durante i periodi di allenamento contro-resistenza. Tuttavia, la letteratura sull’uomo presenta ancora notevoli lacune per quanto riguarda il modo in cui le diverse forme di allenamento o la durata dell’allenamento influenzano gli adattamenti morfologici. Nonostante la scarsità di dati, l’intento finale nel redigere questo articolo è quello di suscitare ulteriore interesse per questo argomento di ricerca. Tali ricerche aiuteranno a confermare o a confutare il ruolo dell’ipertrofia sarcoplasmatica nel facilitare gli adattamenti dell’allenamento contro-resistenza.
Infatti, ritengo che la domanda più rilevante è se questo fenomeno abbia implicazioni pratiche significative.
Sebbene la ricerca sia ancora preliminare, la risposta breve è: dipende dagli obbiettivi individuali. L’aumento della massa sarcoplasmatica sarebbe certamente vantaggioso per chi cerca di migliorare l’estetica, poiché qualsiasi aumento delle dimensioni muscolari è un vantaggio, indipendentemente dalle implicazioni funzionali. L’aumento delle proteine sarcoplasmatiche può persino contribuire a migliorare le prestazioni negli allenamenti di tipo bodybuilding, aumentando il potenziale di costruzione muscolare con attività di sostegno alla componente miofibrillare.
D’altra parte, l’ipertrofia sarcoplasmatica sarebbe probabilmente di scarso beneficio per un atleta che cerca di migliorare la forza, come un PowerLifter. Questi ultimi, giustamente, si concentrano principalmente sul sollevamento di carichi più pesanti e di volumi più bassi per massimizzare la forza nei sollevamenti. Detto questo, alcuni PowerLifter come Layne Norton, Ph.D., combinano il lavoro di forza con modalità incentrate sul pompaggio, come l’allenamento per la restrizione del flusso sanguigno, per ottenere il meglio da entrambi i metodi.
Ahtiainen J. P., Walker S., Peltonen H., Holviala J., Sillanpaa E., Karavirta L., et al. (2016). Heterogeneity in resistance training-induced muscle strength and mass responses in men and women of different ages. Age 38:10. [PMC free article] [PubMed] [Google Scholar]
Allen D. L., Roy R. R., Edgerton V. R. (1999). Myonuclear domains in muscle adaptation and disease. Muscle Nerve 22 1350–1360. 10.1002/(sici)1097-4598(199910)22:10<1350::aid-mus3>3.0.co;2-8 [PubMed] [CrossRef] [Google Scholar]
Alway S. E., Macdougall J. D., Sale D. G., Sutton J. R., Mccomas A. J. (1988). Functional and structural adaptations in skeletal muscle of trained athletes. J. Appl. Physiol. 64 1114–1120. 10.1152/jappl.1988.64.3.1114 [PubMed] [CrossRef] [Google Scholar]
Antonio J., Gonyea W. J. (1994). Muscle fiber splitting in stretch-enlarged avian muscle. Med. Sci. Sports Exerc. 26 973–977. [PubMed] [Google Scholar]
Bell K. E., Brook M. S., Snijders T., Kumbhare D., Parise G., Smith K., et al. (2019). Integrated myofibrillar protein synthesis in recovery from unaccustomed and accustomed resistance exercise with and without multi-ingredient supplementation in overweight older men. Front. Nutr. 6:40. 10.3389/fnut.2019.00040 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Brook M. S., Wilkinson D. J., Mitchell W. K., Lund J. N., Szewczyk N. J., Greenhaff P. L., et al. (2015). Skeletal muscle hypertrophy adaptations predominate in the early stages of resistance exercise training, matching deuterium oxide-derived measures of muscle protein synthesis and mechanistic target of rapamycin complex 1 signaling. FASEB J. 29 4485–4496. 10.1096/fj.15-273755 [PubMed] [CrossRef] [Google Scholar]
Canepari M., Pellegrino M. A., D’antona G., Bottinelli R. (2010). Single muscle fiber properties in aging and disuse. Scand. J. Med. Sci. Sports 20 10–19. 10.1111/j.1600-0838.2009.00965.x [PubMed] [CrossRef] [Google Scholar]
Claassen H., Gerber C., Hoppeler H., Luthi J. M., Vock P. (1989). Muscle filament spacing and short-term heavy-resistance exercise in humans. J. Physiol. 409 491–495. 10.1113/jphysiol.1989.sp017509 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Damas F., Phillips S., Vechin F. C., Ugrinowitsch C. (2015). A review of resistance training-induced changes in skeletal muscle protein synthesis and their contribution to hypertrophy. Sports Med. 45 801–807. 10.1007/s40279-015-0320-0 [PubMed] [CrossRef] [Google Scholar]
Damas F., Phillips S. M., Libardi C. A., Vechin F. C., Lixandrao M. E., Jannig P. R., et al. (2016a). Resistance training-induced changes in integrated myofibrillar protein synthesis are related to hypertrophy only after attenuation of muscle damage. J. Physiol. 594 5209–5222. 10.1113/jp272472 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Damas F., Phillips S. M., Lixandrao M. E., Vechin F. C., Libardi C. A., Roschel H., et al. (2016b). An inability to distinguish edematous swelling from true hypertrophy still prevents a completely accurate interpretation of the time course of muscle hypertrophy. Eur. J. Appl. Physiol. 116 445–446. 10.1007/s00421-015-3287-5 [PubMed] [CrossRef] [Google Scholar]
Damas F., Phillips S. M., Lixandrao M. E., Vechin F. C., Libardi C. A., Roschel H., et al. (2016c). Early resistance training-induced increases in muscle cross-sectional area are concomitant with edema-induced muscle swelling. Eur. J. Appl. Physiol. 116 49–56. 10.1007/s00421-015-3243-4 [PubMed] [CrossRef] [Google Scholar]
D’Antona G., Lanfranconi F., Pellegrino M. A., Brocca L., Adami R., Rossi R., et al. (2006). Skeletal muscle hypertrophy and structure and function of skeletal muscle fibres in male body builders. J. Physiol. 570 611–627. 10.1113/jphysiol.2005.101642 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
D’antona G., Pellegrino M. A., Carlizzi C. N., Bottinelli R. (2007). Deterioration of contractile properties of muscle fibres in elderly subjects is modulated by the level of physical activity. Eur. J. Appl. Physiol. 100 603–611. 10.1007/s00421-007-0402-2 [PubMed] [CrossRef] [Google Scholar]
Defreitas J. M., Beck T. W., Stock M. S. (2016). The findings of Damas et al. have not influenced the previously proposed time course of skeletal muscle hypertrophy. Eur. J. Appl. Physiol. 116 443–444. 10.1007/s00421-015-3286-6 [PubMed] [CrossRef] [Google Scholar]
Degens H., Bosutti A., Gilliver S. F., Slevin M., Van Heijst A., Wust R. C. (2010). Changes in contractile properties of skinned single rat soleus and diaphragm fibres after chronic hypoxia. Pflugers Arch. 460 863–873. 10.1007/s00424-010-0866-5 [PubMed] [CrossRef] [Google Scholar]
Egner I. M., Bruusgaard J. C., Gundersen K. (2016). Satellite cell depletion prevents fiber hypertrophy in skeletal muscle. Development 143 2898–2906. 10.1242/dev.134411 [PubMed] [CrossRef] [Google Scholar]
Eriksson A., Kadi F., Malm C., Thornell L. E. (2005). Skeletal muscle morphology in power-lifters with and without anabolic steroids. Histochem. Cell Biol. 124 167–175. 10.1007/s00418-005-0029-5 [PubMed] [CrossRef] [Google Scholar]
Eriksson A., Lindstrom M., Carlsson L., Thornell L. E. (2006). Hypertrophic muscle fibers with fissures in power-lifters; fiber splitting or defect regeneration? Histochem. Cell Biol. 126 409–417. 10.1007/s00418-006-0176-3 [PubMed] [CrossRef] [Google Scholar]
Fatouros I. G., Kambas A., Katrabasas I., Nikolaidis K., Chatzinikolaou A., Leontsini D., et al. (2005). Strength training and detraining effects on muscular strength, anaerobic power, and mobility of inactive older men are intensity dependent. Br. J. Sports Med. 39 776–780. 10.1136/bjsm.2005.019117 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Fluck M., Kramer M., Fitze D. P., Kasper S., Franchi M. V., Valdivieso P. (2019). Cellular aspects of muscle specialization demonstrate genotype – phenotype interaction effects in athletes. Front. Physiol. 10:526. 10.3389/fphys.2019.00526 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Folland J. P., Williams A. G. (2007). The adaptations to strength training : morphological and neurological contributions to increased strength. Sports Med. 37 145–168. 10.2165/00007256-200737020-00004 [PubMed] [CrossRef] [Google Scholar]
Gasier H. G., Fluckey J. D., Previs S. F., Wiggs M. P., Riechman S. E. (2012). Acute resistance exercise augments integrative myofibrillar protein synthesis. Metabolism 61 153–156. 10.1016/j.metabol.2011.07.001 [PubMed] [CrossRef] [Google Scholar]
Gillooly J. F., Hein A., Damiani R. (2015). Nuclear DNA content varies with cell size across human cell types. Cold Spring Harb. Perspect. Biol. 7:a019091. 10.1101/cshperspect.a019091 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Gokhin D. S., Ward S. R., Bremner S. N., Lieber R. L. (2008). Quantitative analysis of neonatal skeletal muscle functional improvement in the mouse. J. Exp. Biol. 211 837–843. 10.1242/jeb.014340 [PubMed] [CrossRef] [Google Scholar]
Goldspink G. (1964). The combined effects of exercise and reduced food intake on skeletal muscle fibers. J. Cell. Comp. Physiol. 63 209–216. 10.1002/jcp.1030630211 [PubMed] [CrossRef] [Google Scholar]
Goldspink G. (1970). The proliferation of myofibrils during muscle fibre growth. J. Cell Sci. 6 593–603. [PubMed] [Google Scholar]
Goldspink G. (2011). Alterations in myofibril size and structure during growth, exercise, and changes in environmental temperature. Compr. Physiol. 2 539–554. [Google Scholar]
Grgic J., Schoenfeld B. J. (2018). Are the hypertrophic adaptations to high and low-load resistance training muscle fiber type specific? Front. Physiol. 9:402. 10.3389/fphys.2018.00402 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Haun C. T., Vann C. G., Mobley C. B., Osburn S. C., Mumford P. W., Roberson P. A., et al. (2019a). Pre-training skeletal muscle fiber size and predominant fiber type best predict hypertrophic responses to 6 weeks of resistance training in previously trained young men. Front. Physiol. 10:297. 10.3389/fphys.2019.00297 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Haun C. T., Vann C. G., Mobley C. B., Roberson P. A., Osburn S. C., Holmes H. M., et al. (2018). Effects of graded whey supplementation during extreme-volume resistance training. Front. Nutr. 5:84. 10.3389/fnut.2018.00084 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Haun C. T., Vann C. G., Osburn S. C., Mumford P. W., Roberson P. A., Romero M. A., et al. (2019b). Muscle fiber hypertrophy in response to 6 weeks of high-volume resistance training in trained young men is largely attributed to sarcoplasmic hypertrophy. PLoS One 14:e0215267. 10.1371/journal.pone.0215267 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Haun C. T., Vann C. G., Roberts B. M., Vigotsky A. D., Schoenfeld B. J., Roberts M. D. (2019c). A critical evaluation of the biological construct skeletal muscle hypertrophy: size matters but so does the measurement. Front. Physiol. 10:247. 10.3389/fphys.2019.00247 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Henderson C. A., Gomez C. G., Novak S. M., Mi-Mi L., Gregorio C. C. (2017). Overview of the muscle cytoskeleton. Compr. Physiol. 7 891–944. 10.1002/cphy.c160033 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Jorgenson K. W., Hornberger T. A. (2019). The overlooked role of fiber length in mechanical load-induced growth of skeletal muscle. Exerc. Sport Sci. Rev. 47 258–259. 10.1249/jes.0000000000000198 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kadi F., Schjerling P., Andersen L. L., Charifi N., Madsen J. L., Christensen L. R., et al. (2004). The effects of heavy resistance training and detraining on satellite cells in human skeletal muscles. J. Physiol. 558 1005–1012. 10.1113/jphysiol.2004.065904 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kirby T. J., Patel R. M., Mcclintock T. S., Dupont-Versteegden E. E., Peterson C. A., Mccarthy J. J. (2016). Myonuclear transcription is responsive to mechanical load and DNA content but uncoupled from cell size during hypertrophy. Mol. Biol. Cell 27 788–798. 10.1091/mbc.e15-08-0585 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Kraemer W. J., Zatsiorsky V. M. (2006). Science and Practice of Strength Training. Champaign, IL: Human Kinetics. [Google Scholar]
Lee Y. S., Ondrias K., Duhl A. J., Ehrlich B. E., Kim D. H. (1991). Comparison of calcium release from sarcoplasmic reticulum of slow and fast twitch muscles. J. Membr. Biol. 122 155–163. 10.1007/bf01872638 [PubMed] [CrossRef] [Google Scholar]
Lepper C., Partridge T. A., Fan C. M. (2011). An absolute requirement for Pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development 138 3639–3646. 10.1242/dev.067595 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Luthi J. M., Howald H., Claassen H., Rosler K., Vock P., Hoppeler H. (1986). Structural changes in skeletal muscle tissue with heavy-resistance exercise. Int. J. Sports Med. 7 123–127. 10.1055/s-2008-1025748 [PubMed] [CrossRef] [Google Scholar]
Macdougall J. D., Sale D. G., Elder G. C., Sutton J. R. (1982). Muscle ultrastructural characteristics of elite powerlifters and bodybuilders. Eur. J. Appl. Physiol. Occup. Physiol. 48 117–126. 10.1007/bf00421171 [PubMed] [CrossRef] [Google Scholar]
Mccarthy J. J., Mula J., Miyazaki M., Erfani R., Garrison K., Farooqui A. B., et al. (2011). Effective fiber hypertrophy in satellite cell-depleted skeletal muscle. Development 138 3657–3666. 10.1242/dev.068858 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Meijer J. P., Jaspers R. T., Rittweger J., Seynnes O. R., Kamandulis S., Brazaitis M., et al. (2015). Single muscle fibre contractile properties differ between body-builders, power athletes and control subjects. Exp. Physiol. 100 1331–1341. 10.1113/ep085267 [PubMed] [CrossRef] [Google Scholar]
Meng H., Janssen P. M., Grange R. W., Yang L., Beggs A. H., Swanson L. C., et al. (2014). Tissue triage and freezing for models of skeletal muscle disease. J. Vis. Exp. 89:51586. [PMC free article] [PubMed] [Google Scholar]
Metzger J. M., Moss R. L. (1987). Shortening velocity in skinned single muscle fibers. Influence of filament lattice spacing. Biophys. J. 52 127–131. 10.1016/s0006-3495(87)83197-1 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Mitchell C. J., Churchward-Venne T. A., Cameron-Smith D., Phillips S. M. (2015). What is the relationship between the acute muscle protein synthesis response and changes in muscle mass? J. Appl. Physiol. 118 495–497. 10.1152/japplphysiol.00609.2014 [PubMed] [CrossRef] [Google Scholar]
Mitchell C. J., Churchward-Venne T. A., West D. W., Burd N. A., Breen L., Baker S. K., et al. (2012). Resistance exercise load does not determine training-mediated hypertrophic gains in young men. J. Appl. Physiol. 113 71–77. 10.1152/japplphysiol.00307.2012 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Mollenhauer H. H. (1993). Artifacts caused by dehydration and epoxy embedding in transmission electron microscopy. Microsc. Res. Tech. 26 496–512. 10.1002/jemt.1070260604 [PubMed] [CrossRef] [Google Scholar]
Moore D. R., Tang J. E., Burd N. A., Rerecich T., Tarnopolsky M. A., Phillips S. M. (2009). Differential stimulation of myofibrillar and sarcoplasmic protein synthesis with protein ingestion at rest and after resistance exercise. J. Physiol. 587 897–904. 10.1113/jphysiol.2008.164087 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Morpurgo B. (1897). Ueber Activitäts-Hypertrophie der willkürlichen Muskeln. Arch. Pathol. Anat. Physiol. Klin. Med. 1S0 522–554. [Google Scholar]
Murach K. A., Dungan C. M., Peterson C. A., Mccarthy J. J. (2019). Muscle fiber splitting is a physiological response to extreme loading in animals. Exerc. Sport Sci. Rev. 47 108–115. 10.1249/jes.0000000000000181 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Murach K. A., Englund D. A., Dupont-Versteegden E. E., Mccarthy J. J., Peterson C. A. (2018). Myonuclear domain flexibility challenges rigid assumptions on satellite cell contribution to skeletal muscle fiber hypertrophy. Front. Physiol. 9:635. 10.3389/fphys.2018.00635 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Neurohr G. E., Terry R. L., Lengefeld J., Bonney M., Brittingham G. P., Moretto F., et al. (2019). Excessive cell growth causes cytoplasm dilution and contributes to senescence. Cell 176:1083. 10.1016/j.cell.2019.01.018 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Pansarasa O., Rinaldi C., Parente V., Miotti D., Capodaglio P., Bottinelli R. (2009). Resistance training of long duration modulates force and unloaded shortening velocity of single muscle fibres of young women. J. Electromyogr. Kinesiol. 19 e290–e300. 10.1016/j.jelekin.2008.07.007 [PubMed] [CrossRef] [Google Scholar]
Penman K. A. (1969). Ultrastructural changes in human striated muscle using three methods of training. Res. Q. 40 764–772. 10.1080/10671188.1969.10614916 [PubMed] [CrossRef] [Google Scholar]
Phillips S. M. (2000). Short-term training: when do repeated bouts of resistance exercise become training? Can J. Appl. Physiol. 25 185–193. 10.1139/h00-014 [PubMed] [CrossRef] [Google Scholar]
Reidy P. T., Borack M. S., Markofski M. M., Dickinson J. M., Fry C. S., Deer R. R., et al. (2017). Post-absorptive muscle protein turnover affects resistance training hypertrophy. Eur. J. Appl. Physiol. 117 853–866. 10.1007/s00421-017-3566-4 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Roberts M. D., Romero M. A., Mobley C. B., Mumford P. W., Roberson P. A., Haun C. T., et al. (2018). Skeletal muscle mitochondrial volume and myozenin-1 protein differences exist between high versus low anabolic responders to resistance training. PeerJ 6:e5338. 10.7717/peerj.5338 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Roberts M. D., Young K. C., Fox C. D., Vann C. G., Roberson P. A., Osburn S. C., et al. (2020). An optimized procedure for isolation of rodent and human skeletal muscle sarcoplasmic and myofibrillar proteins. J. Biol. Methods 7:e127. [PMC free article] [PubMed] [Google Scholar]
Roche S. M., Gumucio J. P., Brooks S. V., Mendias C. L., Claflin D. R. (2015). Measurement of maximum isometric force generated by permeabilized skeletal muscle fibers. J. Vis. Exp. 100:e52695. [PMC free article] [PubMed] [Google Scholar]
Schiaffino S., Hanzlikova V., Pierobon S. (1970). Relations between structure and function in rat skeletal muscle fibers. J. Cell Biol. 47 107–119. 10.1083/jcb.47.1.107 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Shoepe T. C., Stelzer J. E., Garner D. P., Widrick J. J. (2003). Functional adaptability of muscle fibers to long-term resistance exercise. Med. Sci. Sports Exerc. 35 944–951. 10.1249/01.mss.0000069756.17841.9e [PubMed] [CrossRef] [Google Scholar]
Soltani M., Vargas-Garcia C. A., Antunes D., Singh A. (2016). Intercellular variability in protein levels from stochastic expression and noisy cell cycle processes. PLoS Comput. Biol. 12:e1004972. 10.1371/journal.pcbi.1004972 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Stone M. H., Wilson D. R. R., Newton H. (1983). Physiological basis. Strength Cond. J. 5 40–64. [Google Scholar]
Stromer M. H. (1998). The cytoskeleton in skeletal, cardiac and smooth muscle cells. Histol. Histopathol. 13 283–291. [PubMed] [Google Scholar]
Taber C. B., Vigotsky A., Nuckols G., Haun C. T. (2019). Exercise-induced myofibrillar hypertrophy is a contributory cause of gains in muscle strength. Sports Med. 49 993–997. 10.1007/s40279-019-01107-8 [PubMed] [CrossRef] [Google Scholar]
Thys T. M., Blank J. M., Coughlin D. J., Schachat F. (2001). Longitudinal variation in muscle protein expression and contraction kinetics of largemouth bass axial muscle. J. Exp. Biol. 204 4249–4257. [PubMed] [Google Scholar]
Timson B. F., Bowlin B. K., Dudenhoeffer G. A., George J. B. (1985). Fiber number, area, and composition of mouse soleus muscle following enlargement. J. Appl. Physiol. 58 619–624. 10.1152/jappl.1985.58.2.619 [PubMed] [CrossRef] [Google Scholar]
Toth M. J., Miller M. S., Vanburen P., Bedrin N. G., Lewinter M. M., Ades P. A., et al. (2012). Resistance training alters skeletal muscle structure and function in human heart failure: effects at the tissue, cellular and molecular levels. J. Physiol. 590 1243–1259. 10.1113/jphysiol.2011.219659 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Trappe S., Godard M., Gallagher P., Carroll C., Rowden G., Porter D. (2001). Resistance training improves single muscle fiber contractile function in older women. Am. J. Physiol. Cell Physiol. 281 C398–C406. [PubMed] [Google Scholar]
Trappe S., Williamson D., Godard M., Porter D., Rowden G., Costill D. (2000). Effect of resistance training on single muscle fiber contractile function in older men. J. Appl. Physiol. 89 143–152. 10.1152/jappl.2000.89.1.143 [PubMed] [CrossRef] [Google Scholar]
Valle X., Til L., Drobnic F., Turmo A., Montoro J. B., Valero O., et al. (2013). Compression garments to prevent delayed onset muscle soreness in soccer players. Muscles Ligaments Tendons J. 3 295–302. [PMC free article] [PubMed] [Google Scholar]
Vann C. G., Osburn S. C., Mumford P. W., Roberson P. A., Fox C. D., Sexton C. L., et al. (2020a). Skeletal muscle protein composition adaptations to 10 weeks of high-load resistance training in previously-trained males. Front. Physiol. 11:259. 10.3389/fphys.2020.00259 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Vann C. G., Roberson P. A., Osburn S. C., Mumford P. W., Romero M. A., Fox C. D., et al. (2020b). Skeletal muscle myofibrillar protein abundance is higher in resistance-trained men, and aging in the absence of training may have an opposite effect. Sports 8:7. 10.3390/sports8010007 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Walker D. K., Dickinson J. M., Timmerman K. L., Drummond M. J., Reidy P. T., Fry C. S., et al. (2011). Exercise, amino acids, and aging in the control of human muscle protein synthesis. Med. Sci. Sports Exerc. 43 2249–2258. 10.1249/mss.0b013e318223b037 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Ward S. R., Eng C. M., Smallwood L. H., Lieber R. L. (2009). Are current measurements of lower extremity muscle architecture accurate? Clin. Orthop. Relat. Res. 467 1074–1082. 10.1007/s11999-008-0594-8 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
Widrick J. J., Stelzer J. E., Shoepe T. C., Garner D. P. (2002). Functional properties of human muscle fibers after short-term resistance exercise training. Am. J. Physiol. Regul. Integr. Comp. Physiol. 283 R408–R416. [PubMed] [Google Scholar]
Wilkens J. L., Cavey M. J., Shovkivska I., Zhang M. L., Ter Keurs H. E. (2008). Elasticity, unexpected contractility and the identification of actin and myosin in lobster arteries. J. Exp. Biol. 211 766–772. 10.1242/jeb.007658 [PubMed] [CrossRef] [Google Scholar]
Wilkinson D. J., Franchi M. V., Brook M. S., Narici M. V., Williams J. P., Mitchell W. K., et al. (2014). A validation of the application of D(2)O stable isotope tracer techniques for monitoring day-to-day changes in muscle protein subfraction synthesis in humans. Am. J. Physiol. Endocrinol. Metab. 306 E571–E579. [PMC free article] [PubMed] [Google Scholar]
Yamada A., Yoshio M., Nakayama H. (1997). Bi-directional movement of actin filaments along long bipolar tracks of oriented rabbit skeletal muscle myosin molecules. FEBS Lett. 409 380–384. 10.1016/s0014-5793(97)00558-9 [PubMed] [CrossRef] [Google Scholar]
Yu J. G., Liu J. X., Carlsson L., Thornell L. E., Stal P. S. (2013). Re-evaluation of sarcolemma injury and muscle swelling in human skeletal muscles after eccentric exercise. PLoS One 8:e62056. 10.1371/journal.pone.0062056 [PMC free article] [PubMed] [CrossRef] [Google Scholar]
A causa della natura catabolica del Cortisolo e del desiderio viscerale di molti bodybuilder di mantenere uno stato di anabolismo muscolare costante si è speso molto tempo per cercare di contenere il rilascio di Cortisolo, soprattutto quando non necessario. Gli allenamenti sono stati ridotti in volume e intensità nella sciocca speranza di tenere sotto controllo il Cortisolo nei momenti cruciali della sua funzione fisiologica.
Tuttavia, questa visione cortisolocentrica e del suo impatto in acuto è sia riduttiva che controproducente. Essa non tiene conto della differenza tra gli aumenti acuti e cronici del corticosteroide in questione.
Detto ciò, approfondiamo il ruolo dell’ormone Cortisolo nel processo di ipertrofia muscolare.
Caratteristiche principali del Cortisolo:
Il Cortisolo è un ormone steroideo, appartenente alla classe degli ormoni glucocorticoidi. Quando viene utilizzato come farmaco, è noto come Idrocortisone.
Viene sintetizzato in molti animali, principalmente dalla zona fascicolata della corteccia surrenale nella ghiandola surrenale.[1][2] Viene prodotto in altri tessuti in quantità inferiori.[3] Viene rilasciato con un ciclo diurno e il suo rilascio aumenta in risposta allo stress e a una bassa concentrazione di glucosio nel sangue. Funziona per aumentare la glicemia ematica attraverso la gluconeogenesi, per sopprimere il sistema immunitario e per coadiuvare il metabolismo di grassi, proteine e carboidrati.[4] Diminuisce anche la formazione delle ossa.[5] Molte di queste funzioni sono svolte dal Cortisolo che si lega ai recettori dei glucocorticoidi o dei mineralocorticoidi all’interno della cellula, che poi si legano al DNA per influenzare l’espressione genica.[6][7]
Asse Ipotalamo-Ipofisi-Surrene
Grazie alle proprietà immunoregolatrici del Cortisone, i derivati farmaceutici del Cortisolo, come il Prednisone, sono utilizzati per controllare forti reazioni allergiche, artrite e altre condizioni infiammatorie. I pericoli di un aumento cronico del Cortisolo sono evidenti nel modo attento in cui questi farmaci vengono dosati e nella breve durata dei trattamenti che li utilizzano.
Il Cortisolo è sintetizzato a partire dal Colesterolo. Come già accennato, la sua sintesi avviene nella zona fascicolata della corteccia surrenale (il nome Cortisolo deriva da corteccia). Sebbene la corteccia surrenale produca anche Aldosterone (nella zona glomerulosa) e alcuni ormoni sessuali (nella zona reticolare), il Cortisolo è la sua secrezione principale nell’uomo e in molte altre specie. La midollare della ghiandola surrenale si trova sotto la corteccia e secerne principalmente le catecolamine Adrenalina (Epinefrina) e Noradrenalina (Norepinefrina) sotto stimolazione simpatica.
Ormone Adrenocorticotropo (ACTH)
La sintesi di Cortisolo nella ghiandola surrenale è stimolata dal lobo anteriore dell’ipofisi con l’ACTH; la produzione di ACTH è a sua volta stimolata dal CRH, rilasciato dall’ipotalamo. L’ACTH aumenta la concentrazione di Colesterolo nella membrana mitocondriale interna, attraverso la regolazione della proteina regolatrice steroidogenica acuta. Stimola inoltre la principale fase limitante della sintesi del Cortisolo, in cui il Colesterolo viene convertito in Pregnenolone e catalizzato dal citocromo P450SCC (enzima di scissione della catena laterale).[8]
Il Cortisolo viene metabolizzato reversibilmente a Cortisone[9-89] dal sistema dell’11-beta idrossisteroide deidrogenasi (11-beta HSD), che consiste in due enzimi:11-beta HSD1 e 11-beta HSD2. Il metabolismo del Cortisolo a Cortisone comporta l’ossidazione del gruppo ossidrilico in posizione 11-beta.[10]
L’11-beta HSD1 utilizza il cofattore NADPH per convertire il Cortisone biologicamente inerte in Cortisolo biologicamente attivo.
L’11-beta HSD2 utilizza il cofattore NAD+ per convertire il Cortisolo in Cortisone.
Conversione enzimatica da Cortisone a Cortisolo [11-beta HSD1] e da quest’ultimo a Cortisone [11-beta HSD2].
Nel complesso, l’effetto netto è che l’11-beta HSD1 serve ad aumentare le concentrazioni locali di Cortisolo biologicamente attivo in un dato tessuto; l’11-beta HSD2 serve a diminuire le concentrazioni locali di Cortisolo biologicamente attivo. Se è presente l’esoso-6-fosfato deidrogenasi (H6PDH), l’equilibrio può favorire l’attività dell’11-beta HSD1. L’H6PDH rigenera NADPH, aumentando l’attività dell’11-beta HSD1 e diminuendo quella dell’11-beta HSD2.[11]
È stato ipotizzato che un’alterazione dell’11-beta HSD1 svolga un ruolo nella patogenesi dell’obesità, dell’ipertensione e dell’insulino-resistenza, note come sindrome metabolica.[12]
Un’alterazione dell’11-beta HSD2 è stata implicata nell’ipertensione essenziale ed è nota per portare alla sindrome da eccesso apparente di mineralcorticoidi (SAME).
A breve termine, l’aumento del Cortisolo è associato a una diminuzione della sintesi proteica. Il motivo è che una delle azioni del Cortisolo è quella di fornire substrati energetici alternativi all’organismo quando non c’è abbastanza glucosio. Ciò si verifica durante la restrizione calorica o il digiuno, ma anche durante l’esercizio fisico intenso. Il Cortisolo media la degradazione muscolare in modo che gli aminoacidi presenti nel tessuto muscolare possano essere utilizzati per creare glucosio, attraverso la gluconeogenesi.
Il Cortisolo svolge anche un ruolo importante, ma indiretto, nella glicogenolisi epatica e muscolare (la scissione del glicogeno in glucosio-1-fosfato e glucosio) che si verifica in seguito all’azione del Glucagone e dell’Adrenalina. Inoltre, il Cortisolo facilita l’attivazione della glicogeno fosforilasi, necessaria affinché l’Adrenalina abbia effetto sulla glicogenolisi.[13][14]
È paradossale che il Cortisolo promuova non solo la gluconeogenesi nel fegato, ma anche la glicogenesi: è quindi meglio pensare che il Cortisolo stimoli il turnover di glucosio/glicogeno nel fegato. [Questo è in contrasto con l’effetto del cortisolo nel muscolo scheletrico, dove la glicogenolisi è promossa indirettamente attraverso le catecolamine.[15] In questo modo, il Cortisolo e le catecolamine lavorano sinergicamente per promuovere la scissione del glicogeno muscolare in glucosio, che viene poi utilizzato da altri tessuti.
Il Cortisolo aumenta anche i livelli di glucosio nel sangue riducendo l’assorbimento del glucosio nei tessuti muscolari e adiposi, diminuendo la sintesi proteica e aumentando la scomposizione dei trigliceridi di deposito in grassi acidi liberi (lipolisi). Tutte queste modifiche metaboliche hanno l’effetto netto di aumentare i livelli di glucosio nel sangue, che alimentano il cervello e altri tessuti durante la risposta di lotta o fuga [16] … e i workout…
Livelli elevati di Cortisolo, se prolungati, quindi elevati in cronico, possono portare alla proteolisi (disgregazione delle proteine) protratta e al deperimento muscolare.[17] La ragione della proteolisi è quella di fornire ai tessuti interessati una materia prima per la gluconeogenesi; si vedano gli aminoacidi glucogenici.[18] Gli effetti del Cortisolo sul metabolismo lipidico sono più complicati, poiché la lipogenesi è osservata in pazienti con livelli cronici elevati di glucocorticoidi circolanti,[18] mentre un aumento acuto del Cortisolo circolante promuove la lipolisi. La spiegazione abituale di questa apparente discrepanza è anche l’aumento della concentrazione di glucosio nel sangue (per azione del Cortisolo) stimola il rilascio di Insulina. L’Insulina stimola la lipogenesi, quindi questa è una conseguenza indiretta dell’aumento della concentrazione di cortisolo nel sangue, ma si verifica solo su una scala temporale più lunga. Stiamo parlando sempre di condizioni croniche e non in range fisiologici.
Il Cortisolo è un ormone controinsulinare, contribuisce quindi all’iperglicemia stimolando la gluconeogenesi[19] e inibisce l’utilizzo periferico del glucosio (insulino-resistenza)[19] diminuendo la traslocazione dei trasportatori del glucosio (in particolare GLUT4) sulla membrana cellulare. Il Cortisolo aumenta anche la sintesi di glicogeno (glicogenesi) nel fegato, immagazzinando il glucosio in forma facilmente accessibile.[20] L’effetto permissivo del Cortisolo sull’azione dell’Insulina nella glicogenesi epatica è stato osservato in coltura di epatociti in laboratorio, anche se il meccanismo di questo fenomeno è sconosciuto.
Il Cortisolo aumenta gli aminoacidi liberi nel siero inibendo la formazione di Collagene, diminuendo l’assorbimento di aminoacidi da parte del muscolo e inibendo la sintesi proteica.[21] Il Cortisolo (sotto forma di Opticortinolo) può inibire inversamente le cellule precursori delle IgA nell’intestino dei vitelli.[22] Il Cortisolo inibisce anche le IgA nel siero, come le IgM; tuttavia, non è dimostrato che inibisca le IgE.[23]
Il Cortisolo diminuisce la velocità di filtrazione glomerulare e il flusso plasmatico renale dai reni, aumentando così l’escrezione di fosfati e aumentando la ritenzione di Sodio e acqua e l’escrezione di Potassio agendo sui recettori dei mineralocorticoidi. Aumenta inoltre l’assorbimento di Sodio e acqua e l’escrezione di Potassio nell’intestino.[24] Il Cortisolo favorisce l’assorbimento del Sodio attraverso l’intestino tenue dei mammiferi.[25] La deplezione di Sodio, tuttavia, non influisce sui livelli di Cortisolo[26] e quindi questo ormone non può essere utilizzato per regolare il Sodio sierico. Un carico di Sodio aumenta l’intensa escrezione di Potassio da parte del Cortisolo. In questo caso, il Corticosterone è paragonabile al Cortisolo.[27] Affinché il Potassio esca dalla cellula, il Cortisolo sposta un numero uguale di ioni Sodio all’interno della cellula.[28] Ciò dovrebbe facilitare la regolazione del pH (a differenza della normale situazione di carenza di Potassio, in cui due ioni Sodio si spostano all’interno per ogni tre ioni Potassio che si spostano all’esterno, il che si avvicina all’effetto del Desossicorticosterone).
Cortisolo e workout:
Nell’articolo del 1998 “Stress-Related Cortisol Secretion in Men: Relationships with Abdominal Obesity and Endocrine, Metabolic, and Hemodynamic Abnormalities”, i ricercatori del Sahlgrenska University Hospital in Svezia hanno dato diversi contributi preziosi alla nostra comprensione del Cortisolo e delle sue attività differenti in acuto e in cronico. Innanzitutto, le singole letture dei livelli di Cortisolo di un soggetto “non sono altamente informative, perché il Cortisolo viene secreto in modo molto irregolare”.
I livelli di Cortisolo in genere salgono e scendono nel corso della giornata e un livello elevato in un determinato momento non è indicativo di un problema. Al contrario, livelli di Cortisolo variabili, flessibili e reattivi riflettono un sistema endocrino sano. Se il corpo perdesse la capacità di rispondere ai fattori di stress e di regolare in modo appropriato i livelli di Cortisolo, sarebbe un problema.
Un secondo punto che lo studio svedese fornisce riguarda un aspetto che molte persone sbagliano nella loro ricerca di una body fat ridotta, soprattutto addominale. Il Cortisolo viene spesso definito “l’ormone del grasso della pancia”, ma la verità è che il Cortisolo ha il suo maggiore impatto sul grasso viscerale, che è il grasso che circonda gli organi, non il grasso sottocutaneo che copre gli addominali. Se la body fat rende poco visibile il retto addominale, il problema principale non è il Cortisolo.
Nel 2006, Stephen Bird ha pubblicato una serie di articoli che tracciano un buon quadro dei cambiamenti ormonali che si verificano in seguito al sollevamento pesi e di come i diversi interventi nutrizionali influiscano su tali cambiamenti. Nel loro insieme, questi lavori forniscono un quadro della differenza tra i cambiamenti ormonali a breve termine, ad esempio durante o dopo una sessione di allenamento, e quelli a lungo termine.
Nello studio di Bird erano presenti quattro gruppi di soggetti, suddivisi in base a ciò che potevano bere durante gli allenamenti: acqua, aminoacidi essenziali, carboidrati o aminoacidi essenziali più carboidrati. Nell’arco di 12 settimane, tutti i gruppi hanno perso all’incirca la stessa quantità di grasso corporeo, mentre il gruppo che aveva una supplementazione più completa durante l’allenamento (EAA + carboidrati) ha guadagnato più muscoli.
Esaminiamo ora i cambiamenti acuti che hanno accompagnato questa differente risposta. I ricercatori hanno misurato l’aminoacido 3-metil-istidina nelle urine come marcatore della degradazione muscolare. Come mostra il grafico sottostante, il gruppo che ha bevuto solo acqua (il placebo) ha registrato un aumento della disgregazione muscolare 48 ore dopo la sessione di allenamento. I gruppi che hanno assunto aminoacidi essenziali o carboidrati non hanno subito variazioni. Il gruppo che ha assunto la bevanda combinata per l’allenamento ha registrato una diminuzione dei livelli di 3-metil-istidina dopo l’allenamento.
Ciò che è successo, molto banalmente, è che i substrati ingeriti con la bevanda intra-workout hanno tamponato l’uso delle proteine strutturali.
cosa è successo al Cortisolo? Come si può vedere qui sotto, i livelli di Cortisolo 30 minuti dopo l’esercizio fisico sono aumentati di oltre il 50% nel gruppo che ha bevuto acqua, mentre sono rimasti praticamente invariati nel gruppo EAA. Il Cortisolo è diminuito in entrambi i gruppi che hanno assunto Carboidrati come parte dell’alimentazione peri-workout.
Pensate alla gluconeogenesi, il processo che nel fegato crea glucosio da fonti non glucidiche per fornire energia alle cellule del corpo che ne hanno essenziale bisogno. L’organismo non ha bisogno di generare glucosio – un processo ad alto costo metabolico – quando nel flusso ematico c’è glucosio extra dato da una bevanda sportiva. Pertanto, non si è verificato alcun aumento sensibile del Cortisolo in presenza di carboidrati.
Il catabolismo muscolare a breve termine e il picco di Cortisolo in acuto per il gruppo che beveva acqua possono sembrare significativi, ma non bisogna dimenticare che tutti i gruppi hanno guadagnato massa muscolare nel corso dello studio. Il gruppo che ha bevuto solo acqua ha aggiunto quasi due chili di massa muscolare in 12 settimane! Adesso cominciate ad avere chiara la differenza tra effetto in acuto e effetto in cronico?… Il catabolismo è propedeutico all’anabolismo! Eventi in acuto sono largamente compensati dai processi di recupero, in fisiologia.
I picchi di Cortisolo decrescono nel breve termine!
Quindi le persone con la più alta risposta catabolica in acuto hanno comunque guadagnato muscoli? Certo che si! Ed è piuttosto semplice, in realtà: oltre all’attività propedeutica del catabolismo per avviare i processi anabolici, la fisiologia dei soggetti osservati si è adattata allo stimolo dell’allenamento contro-resistenza nel corso del tempo e ha rilasciato sempre meno Cortisolo, anche senza alcun intervento nutrizionale. Nel gruppo che beveva acqua, i livelli di Cortisolo post-esercizio sono diminuiti del 28% nel corso delle 12 settimane dello studio.
Sì, è emerso che i livelli di Cortisolo e la degradazione muscolare acuta non hanno affatto un impatto negativo sull’aumento dei muscoli o sulla perdita di grasso per un periodo di 12 settimane.
I ricercatori della McMaster hanno analizzato la relazione tra i livelli di Cortisolo post-allenamento e i cambiamenti nella forza, nella massa magra e nella sezione trasversale delle fibre muscolari. Hanno scoperto che dopo 12 settimane di allenamento contro-resistenza, alti livelli di Cortisolo post-allenamento erano correlati (anche se debolmente) con l’aumento della massa magra e con le variazioni delle dimensioni delle fibre muscolari di tipo II.
È bene ripeterlo: Le persone con livelli di Cortisolo più elevati in acuto erano quelle che avevano maggiori probabilità di guadagnare più muscoli nel corso dello studio. Tutto il contrario di quello che i limitati detrattori del Cortisolo si sarebbero aspettati!
Conclusioni:
In definitiva, i dati della ricerca sottolineano che l’interruzione di un allenamento contro-resistenza e/o l’abbassamento del intensità e del volume per paura che i livelli di Cortisolo post-allenamento impennassero, è potenzialmente controproducente ai fini ipertrofici. Lo studio della McMaster ha lasciato intendere che potrebbe addirittura esistere una correlazione tra l’innalzamento acuto del Cortisolo e la crescita muscolare a lungo termine. Detto ciò potreste chiedervi: “Se gli innalzamenti acuti del Cortisolo riflettono una buona sessione di allenamento, allora dovrei smettere di usare i protocolli nutrizionali che riducono il Cortisolo?”. Direi di no, non è assolutamente necessaria l’eliminazione del intra-workout. Le proteine e i carboidrati assunti prima, durante e dopo l’allenamento sono comunque importanti per avviare il processo di recupero.
In queste situazioni, il Cortisolo elevato è semplicemente un indicatore di un allenamento produttivo. E, per non dimenticare, nello studio iniziale di 12 settimane il gruppo che ha assunto aminoacidi e carboidrati ha guadagnato più del doppio dei muscoli rispetto a chi ha bevuto solo acqua; per ovvie ragioni di substrati disponibili e migliore prestazione data dal consumo di CHO.
Un ultimo dubbio: se dobbiamo ignorare i livelli di cortisolo post-allenamento, questo significa che dobbiamo dimenticarci del tutto del Cortisolo e ignorare qualsiasi cambiamento a lungo termine nei nostri livelli?
Assolutamente no!
I cambiamenti a lungo termine del Cortisolo e la diminuzione della sua flessibilità circadiana dovrebbero essere monitorati. Gli effetti sistemici di questo ormone catabolico devono essere presi in considerazione quando si guarda al quadro generale dell’allenamento, della alimentazione e dello stile di vita in generale.
Sonno adeguato, calorie e attenzione al recupero sono i tre fattori più importanti su cui abbiamo il controllo quotidiano. Oltre a questi fattori, è stato suggerito l’uso di integratori come il SAMe o l’Ashwagandha per favorire l’adattamento allo stress e prevenire ulteriormente l’aumento cronico del Cortisolo, o, se atleti “enhanced” farmaci come il Trilostano o l’Aminoglutettimide che sono inibitori della biosintesi steroidea.
Indipendentemente dalla forza e dalla forma fisica, gli elevati livelli di Cortisolo indotti dallo stress cronico possono compromettere il benessere psicofisico. Aumentano il rischio di ipertensione e di malattie cardiovascolari e aggravano qualsiasi altro problema di cui si possa soffrire. Ricordate, tuttavia, di mantenere le cose in prospettiva e di guardare al lungo termine.
Jump up to:ab Chyun YS, Kream BE, Raisz LG (February 1984). “Cortisol decreases bone formation by inhibiting periosteal cell proliferation”. Endocrinology.
DeRijk RH, Schaaf M, de Kloet ER (June 2002). “Glucocorticoid receptor variants: clinical implications”. The Journal of Steroid Biochemistry and Molecular Biology.
Margioris AN, Tsatsanis C (2011). “ACTH Action on the Adrenal”. In Chrousos G (ed.). Adrenal physiology and diseases. Endotext.org. Archived from the original on 29 November 2011. Retrieved 5 June 2012.
^ Jump up to:ab Finken MJ, Andrews RC, Andrew R, Walker BR (September 1999). “Cortisol metabolism in healthy young adults: sexual dimorphism in activities of A-ring reductases, but not 11beta-hydroxysteroid dehydrogenases”. The Journal of Clinical Endocrinology and Metabolism. 84 (9): 3316–3321. doi:10.1210/jcem.84.9.6009. PMID10487705.
^ Dammann C, Stapelfeld C, Maser E (April 2019). “Expression and activity of the cortisol-activating enzyme 11β-hydroxysteroid dehydrogenase type 1 is tissue and species-specific”. Chemico-Biological Interactions. 303: 57–61. doi:10.1016/j.cbi.2019.02.018. PMID30796905. S2CID73467693.
^ Atanasov AG, Nashev LG, Schweizer RA, Frick C, Odermatt A (July 2004). “Hexose-6-phosphate dehydrogenase determines the reaction direction of 11beta-hydroxysteroid dehydrogenase type 1 as an oxoreductase”. FEBS Letters. 571 (1–3): 129–133. doi:10.1016/j.febslet.2004.06.065. PMID15280030. S2CID6360244.
Martin PA, Crump MH (2003). “The adrenal gland”. In Dooley MP, Pineda MH (eds.). McDonald’s veterinary endocrinology and reproduction (5th ed.). Ames, Iowa: Iowa State Press. ISBN978-0-8138-1106-2.
^ Coderre L, Srivastava AK, Chiasson JL (June 1991). “Role of glucocorticoid in the regulation of glycogen metabolism in skeletal muscle”. The American Journal of Physiology. 260 (6 Pt 1): E927–32.
Kuo T, McQueen A, Chen TC, Wang JC (2015). “Regulation of Glucose Homeostasis by Glucocorticoids”. In Wang JC, Harris C (eds.). Glucocorticoid Signaling: From Molecules to Mice to Man. Advances in Experimental Medicine and Biology. Vol. 872. Springer. pp. 99–126.
Khani S, Tayek JA (December 2001). “Cortisol increases gluconeogenesis in humans: its role in the metabolic syndrome”. Clin Sci (Lond).
Brown DF, Brown DD (2003). USMLE Step 1 Secrets: Questions You Will Be Asked on USMLE Step 1. Philadelphia: Hanley & Belfus. p. 63.
Baynes J, Dominiczak M (2009). Medical biochemistry. Mosby Elsevier.
Manchester, KL (1964). “Sites of Hormonal Regulation of Protein Metabolism”. In Allison, NH; Munro JB (eds.). Mammalian Protein Metabolism. New York: Academic Press.
Husband AJ, Brandon MR, Lascelles AK (October 1973). “The effect of corticosteroid on absorption and endogenous production of immunoglobulins in calves”. The Australian Journal of Experimental Biology and Medical Science.
^ Posey WC, Nelson HS, Branch B, Pearlman DS (December 1978). “The effects of acute corticosteroid therapy for asthma on serum immunoglobulin levels”. The Journal of Allergy and Clinical Immunology.
^ Sandle GI, Keir MJ, Record CO (1981). “The effect of hydrocortisone on the transport of water, sodium, and glucose in the jejunum. Perfusion studies in normal subjects and patients with coeliac disease”. Scandinavian Journal of Gastroenterology.
^ Mason PA, Fraser R, Morton JJ, Semple PF, Wilson A (August 1977). “The effect of sodium deprivation and of angiotensin II infusion on the peripheral plasma concentrations of 18-hydroxycorticosterone, aldosterone and other corticosteroids in man”. Journal of Steroid Biochemistry.
Muller AF, Oconnor CM (1958). An International Symposium on Aldosterone. Little Brown & Co. p. 58.
Knight RP, Kornfeld DS, Glaser GH, Bondy PK (February 1955). “Effects of intravenous hydrocortisone on electrolytes of serum and urine in man”. The Journal of Clinical Endocrinology and Metabolism.
Nel 1986 i ricercatori hanno svolto studi approfonditi sull’ossido nitrico (NO), un potente vasodilatatore che può migliorare la circolazione e la salute del cuore. I ricercatori della Pfizer iniziarono a sperimentare farmaci chiamati inibitori della PDE-5 che potenziano e perpetuano gli effetti di dilatazione dei vasi sanguigni dell’NO.
Il loro obiettivo, all’epoca, era quello di trovare un trattamento per l’angina. Il primo farmaco fu il Sildenafil citrato, ma le sperimentazioni dimostrarono che la sua efficacia nel trattamento della patologia era modesta. Tuttavia, i ricercatori hanno iniziato a esaminare le note che descrivevano gli effetti collaterali del farmaco. Ed ecco che molti soggetti hanno riferito di aver sperimentato erezioni durature. Pfizer cambiò rapidamente marcia e avviò studi pilota sugli effetti del Sildenafil citrato sulla disfunzione erettile. Il Viagra, nome commerciale del Sildenafil, fu presto approvato dalla FDA.
Non sono gli anziani hanno beneficiato di questo effetto. Infatti, uomini più giovani si sono affezionati al farmaco, come hanno fatto con i suoi cugini Cialis [Tadalafil] e Levitra [Vardenafil], perché i farmaci in questione aiutavano a gestire l’ansia da prestazione e riducevano i tempi morti tra un episodio sessuale e l’altro.
Confronto tra le strutture di cGMP, Sildenafil e altri inibitori della PDE5. a | Il substrato nativo, cGMP. b | Sildenafil. c | Vardenafil e Tadalafil. cGMP, guanosina monofosfato ciclico; PDE-5, fosfo-diesterasi di tipo 5.
Ma ci sono altri motivi per cui gli uomini potrebbero usare questa classe di farmaci. Non sono solo legati alla salute sessuale, ma anche al Bodybuilding. Infatti, ci sono prove sufficienti per sostenere l’idea di assumere questi farmaci ogni giorno, come qualsiasi altro integratore ritenuto “base” nella preparazione di un bodybuilder.
Caratteristiche dei PDE-5 inibitori:
Un inibitore della fosfodiesterasi di tipo 5 (inibitore della PDE-5) è un farmaco vasodilatatore che agisce bloccando l’azione degradativa della fosfodiesterasi di tipo 5 (PDE-5) specifica per il cGMP sul GMP ciclico nelle cellule muscolari lisce che rivestono i vasi sanguigni che riforniscono vari tessuti. Questi farmaci dilatano i corpi cavernosi del pene, facilitando l’erezione con la stimolazione sessuale, e sono utilizzati nel trattamento della disfunzione erettile (DE). Il Sildenafil è stato il primo trattamento orale efficace disponibile per la DE. Poiché la PDE-5 è presente anche nella muscolatura liscia delle pareti delle arteriole polmonari, due inibitori della PDE-5, il Sildenafil e il Tadalafil, sono approvati dalla FDA per il trattamento dell’ipertensione polmonare. Dal 2019 si stanno apprezzando i più ampi benefici cardiovascolari degli inibitori della PDE-5.[https://www.ncbi.nlm.nih.gov/]
Schema della via dell’Ossido Nitrico (NO)/guanosina monofosfato ciclico (cGMP)/ nucleotide ciclico fosfodiesterasi 5 (PDE-5) e del sito d’azione degli inibitori della PDE-5.
Parte del processo fisiologico di vasodilatazione prevede il rilascio di ossido nitrico (NO) da parte delle cellule endoteliali vascolari, che poi si diffonde alle vicine cellule muscolari lisce vascolari. Lì, l’NO attiva la guanilato ciclasi solubile che converte la guanosina trifosfato (GTP) in guanosina monofosfato ciclico (cGMP), il principale effettore del sistema. Ad esempio, nel pene, il rilascio di NO ad alti livelli dalle cellule endoteliali e dai nervi penieni durante la stimolazione sessuale porta al rilassamento della vascolarizzazione liscia dei corpi cavernosi, causando una vasocongestione e un’erezione prolungata.[https://www.ncbi.nlm.nih.gov/]
Gli inibitori della PDE-5 prolungano l’azione del cGMP inibendo la sua degradazione da parte dell’enzima PDE-5, presente in tutto il corpo. Nel pene, gli inibitori della PDE-5 potenziano gli effetti del cGMP per prolungare l’erezione e aumentare la soddisfazione sessuale, mentre nel muscolo scheletrico aumentano l’iperemia del tessuto per via della vasodilatazione.[https://www.nejm.org/] Tuttavia, gli inibitori della PDE-5 non provocano erezioni senza stimolazione sessuale.
Oltre agli effetti emodinamici, gli inibitori della PDE-5 hanno dimostrato in diversi esperimenti proprietà antinfiammatorie, antiossidanti, antiproliferative e metaboliche.[https://www.ncbi.nlm.nih.gov/] Ma sono ovviamente necessari studi più ampi e a lungo termine per stabilirne l’efficacia e la sicurezza rispetto ad altri farmaci in altre patologie.
Quindi l’uso di questa classe di farmaci nel Bodybuilding si limita al classico trattamento per la disfunzione erettile e il pompaggio muscolare? Non esattamente.
Sicuramente, il potenziale additivo dei PDE-5 inibitori per lo stimolo massimo del “pump”, in specie in combinazione con Citrullina, nel pre-palco può incidere positivamente sugli ultimi ritocchi del “look” dell’atleta. Ricordo inoltre che un maggiore afflusso di sangue al tessuto muscolare significa un migliore pompaggio dato dall’esercizio contro-resistenza e un maggiore afflusso di sostanze nutritive ai muscoli, il che è positivo per la performance, il recupero e la crescita muscolare.
Ma i potenziali non si fermano qui:
Tadalafil
Uno studio del 2005 ha rilevato che dosi di Tadalafil da 10 e 20mg, assunte in media 10 volte al mese, riducevano significativamente i livelli di Estradiolo, ma solo negli uomini che non avevano troppo grasso corporeo – quelli con un IMC inferiore a 27 (1). Gli uomini con più grasso corporeo hanno livelli di Aromatasi più elevati e convertono maggiormente il Testosterone in Estradiolo, indipendentemente dal Tadalafil assunto.
Uno studio sugli effetti del Sildenafil su 140 uomini con un basso livello di Testosterone, di età compresa tra i 40 e i 70 anni, ha rilevato che il farmaco ha aumentato i livelli di Testosterone di circa 100 punti (2). Sebbene una parte di questo aumento dell’ormone maschile possa essere dovuta alla mancata conversione in Estradiolo di una parte del Testosterone, una percentuale di questo aumento sembra derivare anche da una maggiore produzione di Testosterone da parte dei testicoli.
Il Sildenafil riduce lo stress ossidativo indotto dal diabete e migliora la sensibilità all’Insulina. (3) Questo esperimento, a differenza degli altri, è stato condotto sui ratti, ma è probabile che funzioni in modo simile anche nell’uomo.
L’ipotesi che i farmaci che influenzano il flusso sanguigno possano essere utili per la crescita muscolare negli adulti più anziani, ha spinto il Dipartimento di Medicina Interna dell’Università del Texas Medical Branch ha condurre uno studio.
Time-line dello studio
Secondo i ricercatori, le riduzioni della funzione muscolare scheletrica si verificano nel corso di un invecchiamento sano, ma anche con la sedentarietà o con diverse malattie come il cancro, la distrofia muscolare e l’insufficienza cardiaca. Tuttavia, non esistono terapie farmacologiche accettate per migliorare la funzione muscolare scheletrica compromessa.
L’ossido nitrico può influenzare la funzione del muscolo scheletrico attraverso effetti sull’accoppiamento eccitazione-contrazione, sulla funzione miofibrillare, sulla perfusione e sul metabolismo.
I soggetti dello studio erano di mezza età, non allenati e per lo più in sovrappeso, e dovevano assumere un’integrazione giornaliera di Sildenafil per otto giorni, mentre si è analizzato l’effetto sulla sintesi proteica muscolare (il processo che guida la crescita muscolare) e sulla funzione muscolare rispetto a un placebo.
Lo studio ha dimostrato che l’aumento della segnalazione dell’ossido nitrico-guanosina monofosfato ciclico mediante la somministrazione giornaliera a breve termine dell’inibitore della fosfodiesterasi 5, il Sildenafil, aumenta la sintesi proteica, altera l’espressione proteica e la nitrosilazione e riduce la fatica nel muscolo scheletrico umano.
Questi risultati suggeriscono che gli inibitori della fosfodiesterasi 5 rappresentano un valido intervento farmacologico per migliorare la funzione muscolare. Ciò che è stato rilevato, infatti, è che Il Sildenafil aumenta la sintesi proteica muscolare e riduce l’affaticamento muscolare.
Effetti del trattamento con Sildenafil sulla funzione muscolare scheletrica. (A) Forza isometrica degli estensori del ginocchio (percentuale media del giorno di riferimento ± errore standard (SE)) dopo 8 giorni di trattamento, determinata con la dinamometria. (B) Forza isocinetica (120° al secondo) degli estensori del ginocchio (percentuale media del giorno di riferimento ± SE) dopo 8 giorni di trattamento, determinata con la dinamometria. (C) Ripetizioni riuscite (percentuale media del giorno di riferimento ± SE) durante contrazioni isocinetiche affaticanti (120° al secondo) dopo 8 giorni di trattamento. *p = 0,016 rispetto al placebo, t-test non accoppiato, N = 6 placebo, 5 sildenafil. Il numero individuale di ripetizioni riuscite prima (pre) e dopo (post) il trattamento per i soggetti che hanno ricevuto il placebo (pannello superiore) e il Sildenafil (pannello inferiore) è mostrato a destra.Effetti del trattamento con Sildenafil sul proteoma del muscolo scheletrico. (A) Sintesi proteica del muscolo scheletrico (media ± SE) dopo 8 giorni di trattamento, determinata utilizzando l’approccio precursore-prodotto per determinare il tasso di sintesi frazionale. *p = 0,004 rispetto al placebo, t-test non accoppiato, N = 6 placebo, 5 Sildenafil. Percorsi canonici (B) e funzionali (C) influenzati in modo differenziato da sildenafil e placebo, determinati utilizzando l’Ingenuity Pathways Analysis (IPA) dell’espressione proteica in campioni di biopsia del muscolo scheletrico (sono mostrati i 6 percorsi principali). Percorsi canonici (D) e funzionali (E) influenzati in modo differenziato dal Sildenafil e dal placebo, determinati utilizzando l’IPA della S-nitrosilazione delle proteine nei campioni di biopsia del muscolo scheletrico (sono indicati i sei percorsi principali).
L’affaticamento del muscolo scheletrico nel primo giorno di trattamento non era statisticamente diverso dal basale o diverso tra i gruppi di trattamento. Tuttavia, gli scienziati hanno ammesso che dopo otto giorni di trattamento, i soggetti del gruppo Sildenafil hanno completato un numero significativamente maggiore di ripetizioni di successo rispetto al basale in rapporto a quelli che hanno ricevuto il placebo durante contrazioni isocinetiche massimali ripetute.
Essendo un farmaco già approvato e con un eccellente record di sicurezza, i risultati di questo studio suggeriscono che il Sildenafil, e possibilmente altri inibitori della fosfodiesterasi 5, rappresenta una potenziale strategia farmacologica per migliorare la funzione del muscolo scheletrico.
Jeff Nippard
Jeff Nippard, un famoso blogger di fitness su YouTube, ha cercato di mettere le cose in chiaro. Per prima cosa ha contattato Jorn Tromellen, ricercatore sul metabolismo muscolare. Tromellen ha rivelato che in realtà i risultati sono meno impressionanti di quanto sembri. Il Sildenafil non è paragonabile agli AAS, ovviamente, in quanto quando si assume il Sildenafil la sintesi proteica aumenta nel giro di un’ora o due. Mentre gli AAS la stimolano in modo significativo (vedi attività genomica) per tutta la vita attiva del farmaco.
Tuttavia, questo non era sufficiente per Nippard, così si è rivolto a colui che ha effettivamente usato il Sildenafil: il compianto John Meadows.
John Meadows
Meadows ha fatto uso di Sildenafil per tutta la sua carriera agonistica. La prima cosa che ha ammesso è che il livello “di lavoro” del Sildenafil dipendono dalla quantità di cibo presente nello stomaco.
Lui ammette che, gli atleti lo usano prevalentemente per avere più “pump” prima di salire sul palco.
Poi Meadows ha confrontato le sensazioni generali dopo l’assunzione di Testosterone e Sildenafil e, ovviamente, non sono neanche lontanamente paragonabili. Considero questa deduzione al pari dell’affermazione secondo cui l’uomo non possa respirare sottacqua senza attrezzatura apposita… banalità…
La verità è che questo studio ha lasciato ancora più domande rispetto a prima. I risultati sembrano molto promettenti, ma rimangono ancora alcune perplessità.
Lo studio è stato condotto su persone non allenate. Quindi la prima domanda è: Funzionerebbe anche su soggetti più giovani e/o allenati?.
E ancora: sappiamo che avviene un aumento della sintesi proteica, ma non è chiaro se ciò sia a carico delle fibre miofibrillari o di altri tessuti.
Possibili effetti avversi dall’uso di inibitori del PDE-5:
Tutti gli inibitori della PDE-5 sono generalmente ben tollerati.[1] La comparsa di effetti collaterali, o reazioni avverse al farmaco (ADR), con gli inibitori della PDE-5 dipende dalla dose e dal tipo di agente.[https://www.ncbi.nlm.nih.gov/] La cefalea è una ADR molto comune, che si verifica nel >10% dei pazienti. Altre ADR comuni sono: vertigini, vampate di calore, dispepsia, congestione nasale o rinite.[6] Anche il mal di schiena e i dolori muscolari sono più comuni nei pazienti che assumono Tadalafil.[https://www.ncbi.nlm.nih.gov/]
Nel 2007, la Food and Drug Administration (FDA) statunitense ha annunciato l’aggiunta di un’avvertenza sulla possibile perdita improvvisa dell’udito alle etichette dei farmaci inibitori della PDE-5.[https://www.fda.gov/]
Dal 2007 sono emerse prove che suggeriscono che gli inibitori della PDE-5 possono causare una neuropatia ottica anteriore,[https://doi.org/1] anche se l’aumento del rischio assoluto è piccolo.[https://www.ncbi.nlm.nih.gov/]
Infine, si teme che gli inibitori della PDE-5 possano aumentare il rischio di mortalità neonatale nelle donne in gravidanza, e sono stati sospesi gli studi sull’uso dei farmaci per la restrizione della crescita fetale.[https://www.ncbi.nlm.nih.gov/]
Gli inibitori della PDE5 sono metabolizzati principalmente dal sistema enzimatico del citocromo P450, in particolare dal CYP3A4. Esiste la possibilità di interazioni avverse con altri farmaci che inibiscono o inducono il CYP3A4, tra cui gli inibitori della proteasi dell’HIV, il Ketoconazolo e l’Itraconazolo,[Australian Medicines Handbook 2006.] anche se la co-somministrazione non è stata collegata a cambiamenti nella sicurezza o nell’efficacia di entrambi gli agenti. [La combinazione con nitrovasodilatatori come la nitroglicerina e il PETN è controindicata perché può verificarsi ipotensione potenzialmente pericolosa per la vita.[Haberfeld H, ed. (2009). Austria-Codex (in German) (2009/2010 ed.). Vienna: Österreichischer Apothekerverlag.] Gli inibitori della PDE5 non interagiscono sinergicamente con altri farmaci antipertensivi.[https://www.ncbi.nlm.nih.gov/]
Conclusioni:
L’uso sporadico di questi farmaci (meno di 8-10 volte al mese) potrebbe non conferire effetti duraturi sulla salute. Tuttavia, il Tadalafil è approvato per l’uso una volta al giorno ed è ragionevole pensare che i pazienti, giovani o anziani, che hanno una tale prescrizione e lo usano ogni giorno, stiano raccogliendo alcuni, se non tutti, i benefici di cui sopra.
Tuttavia, se ulteriori ricerche confermeranno o aggiungeranno ulteriori elementi positivi all’elenco, potremmo arrivare al punto in cui i medici raccomanderanno quasi universalmente l’uso pressoché quotidiano di questi farmaci.
Per il momento, l’applicazione del Sildenafil nel Bodybuilding si è dimostrata più redditizia come “NO booster” potenziato e coadiuvato dalla Citrullina sia come mezzo per aumentare marcatamente il “pump” sul palco e sia per aumentare l’afflusso ematico nei muscoli (vedi ossigeno e nutrienti) durante il workout.
Diversamente, piccole dosi di Tadalafil possono garantire un contenuto controllo estrogenico in soggetti con body fat contenute, senza il rischio di incorrere il alterazioni lipidiche ematiche.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
Greco EA et al. Testosterone:Estradiol ratio changes associated with long-term tadalafil administration: a pilot study. J Sex Med. 2006 Jul;3(4):716-722. PubMed.
Spitzer M et al. Sildenafil increases serum testosterone levels by a direct action on the testes. Andrology. 2013 Nov;1(6):913-8. PubMed.
Milani E et al. Reduction of diabetes-induced oxidative stress by phos- phodiesterase inhibitors in rats. Comp Biochem Physiol C Toxicol Pharmacol. 2005 Feb;140(2):251-5. PubMed.
Il muscolo scheletrico presenta una notevole complessità biochimica, plasticità fisiologica ed eterogeneità cellulare (Ohlendieck, 2010). È ben noto che l’allenamento contro-resistenza (RT) è la strategia interventistica non farmacologica più potente per ottenere un aumento delle dimensioni del muscolo scheletrico (American College of Sports Medicine, 2009). L’American College of Sports Medicine (2009) raccomanda l’allenamento contro-resistenza nell’intervallo delle 6-12 ripetizioni massime (RM) come ideale per aumentare l’ipertrofia muscolare. Tuttavia, Schoenfeld et al. (2017) hanno concluso che, sulla base delle evidenze attuali, il RT a basso carico (≤60% 1 RM) eseguito fino al cedimento muscolare momentaneo aumenta le dimensioni muscolari in modo simile al RT ad alto carico (>60% 1 RM). Va notato che queste conclusioni sono specifiche per le tecniche di imaging del muscolo intero (cioè ultrasuoni, risonanza magnetica e tomografia computerizzata); non è stato possibile eseguire una meta-analisi per la determinazione istologica diretta (cioè la biopsia muscolare) a causa della mancanza di studi che soddisfacessero i criteri di inclusione della review.
L’evidenza è equivoca per quanto riguarda l’accordo tra le tecniche di imaging del muscolo intero e la determinazione istologica dell’ipertrofia muscolare. Ad esempio, McCall et al. (1996) hanno riportato un aumento del 13% dell’area della sezione trasversale del muscolo (CSA) misurata tramite risonanza magnetica, insieme a un aumento del 10 e del 17% per le fibre muscolari di tipo I e II, rispettivamente. Mentre la meta-analisi di Schoenfeld et al. (2017) ha riportato cambiamenti simili nelle dimensioni del muscolo indipendentemente dagli schemi di carico, resta da chiedersi se l’ipertrofia delle fibre muscolari di tipo I e II sia simile anche tra condizioni di alto e basso carico. È possibile che il RT ad alto carico enfatizzi l’ipertrofia muscolare di tipo II (Campos et al., 2002), mentre il RT a basso carico stimoli una maggiore crescita delle fibre muscolari di tipo I (Ogborn e Schoenfeld, 2014; Grgic et al., 2018).
Diversi studi di elettromiografia (EMG) in acuto mostrano che l’ampiezza EMG è significativamente maggiore con il RT ad alto carico rispetto a quello a basso carico, suggerendo così che sono necessari carichi più elevati per stimolare completamente le unità motorie a soglia più alta associate alle fibre di tipo IIx (Schoenfeld et al., 2014, 2016). Tuttavia, una maggiore ampiezza EMG in una determinata condizione non riflette necessariamente gli adattamenti a lungo termine al RT regolato (per una rassegna dettagliata sull’argomento, si veda Vigotsky et al., 2018). L’unico modo per valutare questo argomento è analizzare gli studi che hanno confrontato direttamente il RT ad alto e basso carico e il suo impatto sull’ipertrofia delle fibre muscolari. Pertanto, il presente articolo riporta le informazioni contenute nella review di Jozo Grgic e Brad J. Schoenfeldsi nella quale sono stati discussi e interpretati gli studi che hanno valutato i cambiamenti delle fibre muscolari che si verificano longitudinalmente con schemi di RT ad alto e basso carico. Pur riconoscendo che esistono diverse fibre di tipo ibrido delineate nel muscolo scheletrico umano, in questo articolo verrà utilizzata la classificazione di base in tipo I, tipo IIa e tipo IIx.
Effetti dell’allenamento contro-resistenza ad alto o basso carico sull’adattamento all’ipertrofia delle fibre muscolari: Finora solo pochi studi hanno analizzato questo argomento (vedi tabella seguente). Jackson et al. (1990) hanno condotto uno studio crossover controbilanciato in cui i partecipanti hanno eseguito prima un mesociclo di 7,5 settimane di RT ad alto carico orientato alla forza muscolare o 7,5 settimane di RT a basso carico orientato alla resistenza muscolare. Dopo un periodo di “washout” di 5,5 settimane (cioè senza RT), i partecipanti sono passati a eseguire la routine alternativa. Dopo le prime 7,5 settimane, si è verificato un aumento delle dimensioni di tutti i tipi di fibre muscolari per entrambi i gruppi di RT. Dopo le seconde 7,5 settimane di RT, i partecipanti che hanno eseguito il mesociclo di forza muscolare come secondo trattamento hanno registrato un’ulteriore crescita delle fibre di tipo I e II. Al contrario, i partecipanti che hanno eseguito il RT orientato alla resistenza muscolare come secondo trattamento hanno mostrato una diminuzione delle dimensioni di tutti i tipi di fibre, con la diminuzione maggiore osservata per le fibre di tipo IIx. Questi risultati suggeriscono che una progressione da carichi bassi a carichi elevati potrebbe portare a guadagni più consistenti.
Sintesi dei risultati degli studi che confrontano l’allenamento ad alto e basso carico e i suoi effetti sugli adattamenti all’ipertrofia delle fibre muscolari.
Solo uno studio è stato condotto su adulti anziani. Taaffe et al. (1996) hanno cercato di confrontare gli effetti del RT ad alto carico rispetto a quello a basso carico sulle dimensioni muscolari, utilizzando un intervento della durata di 1 anno. Le donne non allenate si sono allenate 3 giorni alla settimana; il gruppo ad alto carico ha eseguito dieci ripetizioni con carichi esterni dell’80% del 1 RM, mentre il gruppo a basso carico ha eseguito 14 ripetizioni con carichi corrispondenti al 40% del 1 RM. Valutando le variazioni percentuali, il gruppo ad alto carico ha ottenuto una maggiore ipertrofia sia nelle fibre muscolari di tipo I che in quelle di tipo II. In apparenza, questi risultati suggeriscono che il RT ad alto carico è un prerequisito per massimizzare l’ipertrofia in tutti i tipi di fibre. Tuttavia, i ricercatori hanno scelto di equiparare il carico volumetrico totale tra le condizioni, facendo interrompere al gruppo a basso carico ogni serie ben al di sotto dell’affaticamento volitivo. Tali eventi hanno inavvertitamente influenzato i risultati a favore della condizione ad alto carico, dato che un elevato livello di fatica è obbligatorio per ottenere benefici ipertrofici quando ci si allena con carichi inferiori (Morton et al., 2016).
Di fondamentale importanza è il fatto che in entrambi gli studi sopra citati, il programma di RT nei gruppi a basso carico non è stato eseguito fino al cedimento muscolare momentaneo, il che indica una disparità di stimoli di allenamento tra le condizioni di alto e basso carico. L’impiego di un programma di RT a basso carico e la mancata esecuzione di ripetizioni a cedimento muscolare momentaneo possono ostacolare l’adattamento muscolare (Burd et al., 2012). Secondo il principio delle dimensioni di Henneman (1985), le unità motorie più grandi saranno reclutate in sequenza all’aumentare dei requisiti di produzione di forza, portando infine all’attivazione dell’intero pool di unità motorie. È possibile che con l’allenamento a basso carico le unità motorie a soglia più bassa siano sottoposte a carico per un periodo più lungo, il che a sua volta potrebbe aumentare la risposta ipertrofica delle fibre muscolari di tipo I. Questo effetto potrebbe non verificarsi con l’allenamento ad alto carico e, forse per questo motivo, l’ipertrofia preferenziale delle fibre muscolari di tipo II è stata comunemente riportata con il RT (Folland e Williams, 2007).
Campos et al. (2002) sono stati i primi a confrontare il RT ad alto carico con quello a basso carico, con entrambi i gruppi che si allenavano fino al cedimento muscolare momentaneo, utilizzando misure istologiche della crescita muscolare. I ricercatori hanno randomizzato 27 partecipanti non allenati in tre diversi gruppi di carico: programmi di RT ad alto carico (3-5 RM), a carico moderato (9-11 RM) e a basso carico (20-28 RM). Dopo 8 settimane di RT, tutti i tipi di fibre muscolari si sono ipertrofizzati nei gruppi ad alto e moderato carico. Tuttavia, nel gruppo a basso carico non è stato osservato un aumento significativo della fCSA muscolare prima e dopo l’intervento nelle fibre muscolari di tipo I o II. Schuenke et al. (2012) hanno riportato risultati simili a quelli di Campos et al. (2002). Dopo 6 settimane di RT per la parte inferiore del corpo, il gruppo a basso carico (40-60% 1 RM) non ha mostrato aumenti significativi della CSA delle fibre muscolari di tipo I, mentre il gruppo ad alto carico (80-85% 1 RM) ha ottenuto robusti aumenti nelle dimensioni di tutti i tipi di fibre muscolari, con i maggiori guadagni osservati nelle fibre di tipo IIx. Sebbene il tessuto muscolare sia un ottimo esempio di plasticità tissutale e sia in grado di subire drastici cambiamenti nel profilo fenotipico con il RT regolato, il tasso di aumento delle dimensioni delle fibre muscolari nella condizione di carico elevato merita di essere esaminato (Burd et al., 2013). I ricercatori hanno riportato un tasso di crescita delle fibre muscolari dello 0,66% per giorno di allenamento, che è da 4 a 5 volte superiore ai valori presentati nella review di Wernbom et al. (2007). Tuttavia, sebbene non siano così elevati come quelli rilevati da Schuenke et al. (2012), tassi di crescita simili con il RT ad alto carico per la parte inferiore del corpo sono stati rilevati in precedenza in letteratura (Staron et al., 1990; Lamas et al., 2010).
È interessante notare che alcuni studi riportano che il RT a basso carico induce una maggiore risposta ipertrofica nelle fibre muscolari di tipo I. Vinogradova et al. (2013) hanno confrontato il RT ad alto (80-85% di 1 RM) e a basso carico (50% di 1 RM) in un gruppo di giovani uomini non allenati. I risultati hanno indicato che la crescita dei tipi di fibre muscolari è direttamente correlata al carico di allenamento. In particolare, il gruppo ad alto carico ha ottenuto i maggiori aumenti delle dimensioni delle fibre muscolari di tipo II, mentre il gruppo a basso carico ha ottenuto i maggiori aumenti delle dimensioni delle fibre muscolari di tipo I. I ricercatori hanno ipotizzato che i maggiori aumenti delle fibre muscolari di tipo I nel gruppo a basso carico fossero legati a un maggiore stress metabolico (Vinogradova et al., 2013). Lo stress metabolico si riferisce all’accumulo di metaboliti, ad esempio all’aumento del flusso di calcio, del lattato, del potassio e degli ioni idrogeno, ed è un meccanismo ipotizzato per mediare l’ipertrofia muscolare attraverso un maggiore reclutamento di fibre, cambiamenti nella produzione ormonale e/o aumento del volume delle cellule per osmosi e/o re-sintesi glicogeno, tra gli altri (Schoenfeld, 2013). Risultati simili a quelli presentati da Vinogradova et al. (2013) sono stati riportati dallo stesso laboratorio, con un RT a basso carico che porta a un maggiore aumento dell’ipertrofia di tipo I e un RT ad alto carico che aumenta l’ipertrofia delle fibre di tipo II (Netreba et al., 2013).
Utilizzando un within-subject design, Mitchell et al. (2012) hanno assegnato casualmente 18 uomini in modo controbilanciato all’esecuzione di un’estensione unilaterale del ginocchio con una gamba che si allenava ad alto carico (80% di 1 RM) e l’altra a basso carico (30% di 1 RM). Dopo 10 settimane di RT, è stata osservata un’ipertrofia significativa dal basale al post-intervento in tutti i tipi di fibre muscolari per entrambi i gruppi di RT ad alto carico (80% di 1 RM) e a basso carico (30% di 1 RM). Non sono state rilevate differenze statisticamente significative tra le condizioni di carico nella crescita delle fibre muscolari. Tuttavia, è importante sottolineare che il gruppo a basso carico ha ottenuto un aumento del 23% delle dimensioni delle fibre muscolari di tipo I, rispetto a un aumento del 16% delle dimensioni delle fibre muscolari di tipo I osservato nel gruppo ad alto carico. Data l’esiguità del campione impiegato, ciò solleva la possibilità di un errore di tipo 2, in base al quale esistono effettivamente differenze significative tra le condizioni e, considerando l’entità delle differenze, tali risultati possono essere significativi dal punto di vista pratico.
Un limite comune a tutti gli studi citati è l’utilizzo di partecipanti non allenati. È stato dimostrato che gli individui con un RT più elevato sperimentano una risposta nella sintesi proteica attenuata dopo l’esercizio di resistenza, suggerendo un possibile effetto tetto del guadagno muscolare (Damas et al., 2015). Per colmare questa lacuna nella letteratura, Morton et al. (2016) hanno randomizzato giovani uomini con una media di 4 anni di esperienza di RT a un programma di esercizi total body di 12 settimane, utilizzando uno schema ad alto carico (8-12 RM) o a basso carico (20-25 RM). Dopo il protocollo di RT, sono stati osservati tassi di crescita simili in tutti i tipi di fibre muscolari, indipendentemente dalla condizione di carico. Questi risultati isolati suggeriscono che gli individui allenati possono sperimentare una crescita uguale in tutti i tipi di fibre, indipendentemente dallo schema di carico utilizzato.
Come per la maggior parte delle ricerche sulla RT, la stragrande maggioranza degli studi che hanno analizzato questo argomento sono stati di durata relativamente breve; non è chiaro se le potenziali differenze nell’ipertrofia dei tipi di fibre specifiche per un determinato schema di carico possano ampliarsi o ridursi nel tempo. Infine, tutti gli studi inclusi hanno valutato la crescita delle fibre nel muscolo vasto laterale. Come già evidenziato in precedenza, il muscolo vasto laterale è il muscolo più comunemente scelto per le biopsie a causa della sua composizione mista di fibre, della sua accessibilità e della sua allenabilità (Staron et al., 2000). Data la scarsità di dati, l’estrapolazione dei risultati ad altri gruppi muscolari è limitata.
Considerazioni sperimentali e conclusioni generali:
L’esame dei cambiamenti del tipo di fibra attraverso la biopsia muscolare presenta alcune limitazioni intrinseche. In primo luogo, la tecnica bioptica prevede l’estrazione di una piccola quantità di tessuto da un determinato muscolo, che potrebbe non riflettere necessariamente i cambiamenti specifici del tipo di fibra a livello dell’intero muscolo. Inoltre, esistono variazioni nella distribuzione dei tipi di fibre da superficiali a profonde e da prossimali a distali (Blomstrand e Ekblom, 1982), per cui è importante considerare questi fattori quando si estrapolano i risultati in termini pratici. In secondo luogo, il metodo utilizzato per determinare i tipi di fibre varia da uno studio all’altro. La maggior parte della letteratura recensita ha impiegato l’istochimica della mATPasi per delineare la tipizzazione delle fibre, mentre molti altri (Netreba et al., 2013; Vinogradova et al., 2013; Morton et al., 2016) hanno utilizzato anticorpi, che potrebbero avere una maggiore precisione nel delineare le fibre ibride. Infine, le differenze nella misurazione di fCSA possono comportare differenze nelle stime dimensionali, limitando così il confronto tra gli studi.
In conclusione si evince che, quando il RT a basso carico non viene eseguito fino al cedimento muscolare, l’allenamento ad alto carico sembra fornire uno stimolo ipertrofico superiore e quindi una maggiore crescita di tutte le fibre muscolari. Sebbene alcune prove indichino che il RT a basso carico, se effettuata fino al cedimento muscolare, possa indurre una maggiore risposta ipertrofica nelle fibre muscolari di tipo I rispetto al RT ad alto carico e che il RT ad alto carico possa indurre una crescita preferenziale delle fibre muscolari di tipo II, la letteratura rimane alquanto equivoca sull’argomento. In sintesi, attualmente non ci sono prove sufficienti per trarre conclusioni definitive sui cambiamenti che si verificano a livello delle fibre muscolari con i diversi schemi di carico.
Anche in questo caso, sembrerebbe che la formula ” in medio stat virtus” sia nuovamente la più indicata da applicare…