Più le cellule sono sensibili all’insulina, più si è sani a livello metabolico e sistemico. Basta domandarlo ad un diabetico. O ai Bodybuilder supplementati chimicamente che hanno abusato dell’Insulina esogena a tal punto da aggravare marcatamente la loro insulino resistenza sfociando, a volte, nel diabete di tipo II. Scienziati molecolari australiani hanno scoperto che la sensibilità all’insulina subisce un miglioramento significativo in rapporto alle concentrazioni del coenzima Q10 nei mitocondri cellulari. (1)
Nell’esperimento svolto su esseri umani dove sono stati presi in esame soggetti sani [Ins.Sens] e soggetti con insulino-resistenza [Ins.Res], i ricercatori hanno somministrato ai partecipanti glucosio e insulina osservando successivamente la diminuzione della concentrazione di acidi grassi liberi [NEFA] nel sangue. Maggiore è la diminuzione, maggiore è la sensibilità all’insulina.
Ovviamente, la concentrazione di acidi grassi liberi è diminuita maggiormente nei soggetti sani [in basso a sinistra].
I ricercatori hanno anche determinato la quantità di coenzima Q10 presente nei mitocondri dei soggetti dello studio. E sia nei soggetti sani che in quelli insulino-resistenti, la concentrazione di acidi grassi liberi è diminuita maggiormente quando i loro mitocondri contenevano più coenzima Q10 [in alto a destra].
Ma è possibile migliorare l’insulino sensibilità attraverso l’integrazione con il coenzima Q10? Con uno studio sugli animali, i ricercatori hanno cercato di rispondere a questa domanda. Hanno dato a un gruppo di topi cibo normale [Chow] e hanno fatto ingrassare un altro gruppo di topi dando loro del cibo con grasso e zucchero extra [HFHSD]. Quest’ultimo gruppo di animali ha sviluppato una forma di diabete.
I ricercatori hanno quindi somministrato a metà degli esemplari di entrambi i gruppi una dose di coenzima Q10 ogni due giorni per due settimane. I ricercatori hanno utilizzato il coenzima Q10 sotto forma di LiQsorb, un prodotto della Tishcon Corporation.(2) Ogni due giorni i ricercatori hanno iniettato il coenzima Q10 direttamente nell’intestino tenue degli animali del test.
Questa modalità di somministrazione è di circa due volte più efficace della somministrazione orale. Se invece dei topi fossero stati presi in esame degli esseri umani, la dose di coenzima Q10/LiQsorb alla quale sarebbero stati sottoposti sarebbe stata di circa 160mg/die.
Dopo il periodo di supplementazione, i topi hanno ricevuto una forma di glucosio [2DOG]. Nei topi diabetici-ingrassati la supplementazione con coenzima Q10 ha portato ad una più rapida diminuzione dei livelli ematici dell’analogo del glucosio [in basso a sinistra]. La supplementazione ha anche causato la diminuzione della concentrazione di acidi grassi liberi dopo la somministrazione della variante glucidica [in basso a destra]. Da quanto emerso, l’assunzione del coenzima Q10 ha parzialmente migliorato l’insulino-sensibilità.
Il coenzima Q10 si trova nei mitocondri, le centrali energetiche delle cellule del corpo, dove è richiesto per il flusso di elettricità al “motore” cellulare che è responsabile della produzione di energia, come affermato durante un comunicato stampa dal primo autore dello studio qui trattato Daniel Fazakerley della University of Sydney. (3)
La produzione di energia può anche generare specie chimiche reattive – spesso indicate come” specie reattive dell’ossigeno” o ” ossidanti “- come sottoprodotto, le quali possono essere dannose per le cellule. Studi precedenti hanno dimostrato che questi ossidanti possono causare insulino-resistenza. Si è scoperto che una riduzione del coenzima Q10 mitocondriale provoca un aumento della formazione di ossidanti da parte dei mitocondri.
È importante sottolineare che reintegrando il coenzima Q10 nei mitocondri, sia nelle cellule in vitro che negli animali, i ricercatori sono stati in grado di ripristinare gli ossidanti mitocondriali portandoli a livelli “normali” migliorando di conseguenza la sensibilità all’insulina.
Il ricercatore capo dello studio qui esposto, David James, anch’esso affiliato alla University of Sydney, afferma che la supplementazione di coenzima Q10 potrebbe rivelarsi una misura preventiva inestimabile per le malattie legate all’insulino-resistenza o a malattie pre-diabetiche come il diabete di tipo II, le malattie cardiovascolari, il cancro e la demenza.
Gli AAS possono alterare la sensibilità all’insulina, un indicatore importante della salute metabolica. Tuttavia, l’effetto di questi farmaci può essere variabile. Ad esempio, la somministrazione di Testosterone può migliorare la sensibilità all’insulina in uomini ipogonadici.(1) L’Oxandrolone (a 20 mg al giorno) ha dimostrato di migliorare la sensibilità all’insulina in uomini anziani (dai 60 agli 87 anni).(2) Questi benefici effetti metabolici sono stati correlati con la riduzione del tessuto adiposo viscerale (VAT). Come ben sappiamo si tratta di uno strato di grasso profondo che circonda gli organi addominali e che è associato all’insulino resistenza.(3) Uno stato di insulino resistenza può portare al manifestarsi di altri problemi di salute come ipertensione, ipercolesterolemia, ipertrigliceridemia e l’aumento del rischio di sviluppare diabete e malattie cardiovascolari. Riducendo il VAT, il Testosterone e gli altri AAS possono migliorare la sensibilità all’insulina e potenzialmente la salute metabolica.
Al contrario, l’abuso di AAS è stato associato ad alterazione del metabolismo glucidico. (4) In uno studio, i Powerlifter che avevano abusato di AAS in dosi elevate per un massimo di sette anni hanno mostrato di avere una ridotta tolleranza al glucosio e un peggioramento dell’insulino resistenza.(5) Nonostante gli atleti presi in esame avessero una decennale esperienza nella pratica di allenamenti contro resistenza, essi mostrarono una secrezione insulinica maggiore in risposta all’ingestione di glucosio rispetto al gruppo di controllo composto da soggetti obesi sedentari. Ulteriori studi, nei quali è stato preso in esame il Methandrostenolone, hanno mostrato un aumento significativo della secrezione insulinica e una potenziale resistenza ad essa.(6) Ciò nonostante, non sono stati osservati risultati similari con tutti gli AAS. Ad esempio, la somministrazione di Testosterone Enantato a dosi fino a 600mg a settimana per 20 settimane non ha portato a cambiamenti nella sensibilità all’insulina in giovani uomini sani.(7) Anche il Nandrolone Decanoato (a 300 mg a settimana) non ha alterato la tolleranza al glucosio e ha migliorato la gestione di quest’ultimo in modo insulino-indipendente.(8) I dati relativi agli effetti degli AAS sulla sensibilità all’insulina sono difficili da interpretare. Sembra che quando questi farmaci vengono utilizzati, almeno inizialmente, la riduzione della percentuale del grasso corporeo è cosa comune, in particolare per quanto riguarda la riduzione del tessuto adiposo viscerale, come riportato in precedenza. Ciò può effettivamente migliorare la sensibilità all’insulina e lo stato metabolico generale e ridurre determinati fattori di rischio specifici per diabete e malattie cardiovascolari. Oltre a ciò, gli effetti degli AAS sul metabolismo glucidico non sono completamente compresi e, di conseguenza, sono difficili da prevedere. Gli studi nei quali sono state utilizzate dosi sovraterapeutiche di Testosterone e Nandrolone non hanno mostrato cambiamenti negativi del metabolismo glucidico, suggerendo che l’abuso moderato di AAS non è probabilmente associato ad alterazioni della sensibilità all’insulina. Allo stesso tempo, gli studi suggeriscono che potrebbero esserci problemi con l’abuso di AAS (dosi elevate per protratti periodi di tempo). Sono quindi necessarie ulteriori ricerche per valutare l’impatto dell’abuso di AAS sulla salute metabolica.
Gabriel Bellizzi
Riferimenti:
ANABOLICS 11th Edition [William Llewellyn]
Androgen treatment of middle-aged, obese men: effects on metabolism, muscle and adipose tissues. Mårin P, Krotkiewski M, Björntorp P. Eur J Med. 1992 Oct;1(6):329-36.
Effects of androgen therapy on adipose tissue and metabolism in older men. Schroeder ET, Zheng L, Ong MD, Martinez C, Flores C, Stewart Y, Azen C, Sattler FR. J Clin Endocrinol Metab. 2004 Oct;89(10):4863-72.
Insulin sensitivity, insulin secretion, and abdominal fat: the insulin resistance atherosclerosis study (IRAS) family study. Wagenknecht LE, Langerfeld CD et al. Diabetes 52:2490-2494.
Recent developments in the toxicology of anabolic steroids. Graham S, Kennedy M. Drug Saf. 1990 Nov-Dec;5(6):458-76.
Insulin resistance and diminished glucose tolerance in powerlifters ingesting anabolic steroids. Cohen JC, Hickman R. J Clin Endocrinol Metab. 1987 May;64(5):960-3.
Insulin action and dynamics modelled in patients taking the anabolic steroid methandienone (Dianabol). Godsland IF, Shennan NM, Wynn V. Clin Sci (Lond). 1986 Dec;71(6):665-73
The effects of varying doses of T on insulin sensitivity, plasma lipids, apolipoproteins, and C-reactive protein in healthy young men. Singh AB, Hsia S. et al. J Clin Endocrinol Metab. 2002 Jan;87(1):136-43.
Nandrolone, a 19-nortestosterone, enhances insulin-independent glucose uptake in normal men. Hobbs CJ, Jones RE, Plymate SR. et al. J Clin Endocrinol Metab. 1996 Apr;81(4):1582-5.
Se si hanno livelli glicemici ematici eccessivamente alti, il corpo sintetizza meno Testosterone. Il meccanismo coinvolto è stato mostrato in uno studio in vitro svolto da endocrinologi italiani presso l’Università di Firenze e pubblicato sul International Journal of Endocrinology. (1) Se si desidera ottimizzare i livelli di Testosterone endogeno in modo naturale, bisogna assicurarsi di non aggravare l’insulino-resistenza (e di non far sfociare questa condizione in diabete di tipo II) oltre ad evitare picchi glicemici estremamente alti in cronico.
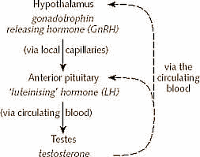
GNRH, Testosterone e Glucosio
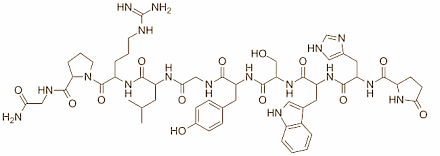
I ricercatori italiani hanno svolto il presente studio al fine di comprendere meglio le cause che portano spesso i diabetici ad avere bassi livelli di Testosterone, il quale si traduce nello sviluppo di problemi della funzionalità sessuale e di infertilità. Hanno svolto esperimenti su cellule umane volte alla produzione di ormone di rilascio delle gonadotropine [GnRH]. Il GnRH svolge un ruolo chiave nel rilascio di Testosterone nel corpo.
Ormone di rilascio delle gonadotropine [GnRH]
Nel cervello l’ipotalamo rilascia GnRH un paio di volte al giorno. Di conseguenza questo ormone stimola il rilascio di altri due ormoni ipofisari: LH e FSH. Questi ormoni a loro volta sono responsabili della produzione di Testosterone nei testicoli e della produzione di Testosterone, Progesterone ed Estradiolo nelle ovaie. E’ corretto specificare che nell’uomo l’FSH attiva la produzione della proteina legante gli androgeni (ABP) nelle cellule del Sertoli, nei tubuli seminiferi, ed è fondamentale per la spermatogenesi. E’ l’LH che nell’uomo ricopre principalmente il ruolo di stimolante l’attività endocrina delle cellule interstiziali del testicolo con produzione di Testosterone. Inoltre, a causa del suo ruolo nella produzione di Testosterone, ormone necessario per la maturazione delle cellule germinali, l’ormone luteinizzante stimola anche, in maniera indiretta, la spermatogenesi. Nella donna, invece, l’FSH e l’LH agiscono sinergicamente nella riproduzione. L’FSH è rilasciato nel circolo portale dell’adenoipofisi e poi nel circolo sanguineo sistemico dove rimane attivo per 3-4 ore. La sua funzione è principalmente di regolazione dello sviluppo dei follicoli ovarici e, quindi, della stimolazione della produzione di Estrogeno e Progesterone.
Studio
I ricercatori hanno esposto le cellule deputate alla produzione di GnRH a una concentrazione normale di glucosio [5 millimoli] [NG], ad un’alta concentrazione di glucosio [22 millimoli] [HG], ad una concentrazione molto elevata di glucosio [40 millimoli] [VHG] e al carboidrato mannitolo ad una concentrazione di 20 millimoli [M].
Se la regolazione del glucosio ematico e della secrezione e funzionalità dell’Insulina non sono compromesse, i livelli di glucosio non raggiungeranno mai gli alti livelli utilizzati dai ricercatori nel loro esperimento. D’altra parte solo i diabetici possono sperimentare questi livelli. E gli alimenti che contengono grandi quantità di carboidrati a rapido assorbimento possono aumentare i livelli di glucosio delle persone sane mediamente tra la concentrazione normale [5 millimoli] [NG] e l’alta concentrazione di glucosio [20 millimoli] [HG] utilizzate nello studio.
Risultati
Le figure sottostanti mostrano che alte concentrazioni di glucosio riducono il rilascio di GnRH. Ciò è probabilmente dovuto al fatto che, in grandi quantità, il glucosio riduce la produzione del recettore per il peptide KISS-1 e il conseguente rilascio di KISS-1. Il KISS-1 è un ormone che aumenta il rilascio di GnRH.
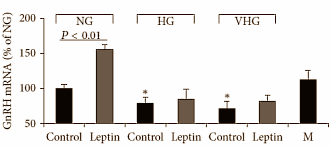
Un’alta concentrazione di glucosio riduce anche la produzione di recettori per la Leptina [LEPR], come mostrato sopra. Come ben sappiamo, la Leptina è un ormone rilasciato dalle cellule adipose. Tra le sue azioni vi è anche quella di stimola il rilascio di LH, e con sequenzialmente anche di Testosterone, tramite il KISS-1.
I ricercatori hanno voluto dimostrare se la loro teoria fosse vera esponendo le cellule deputate alla sintesi di GnRH alla Leptina sintetica. La figura seguente mostra che alte concentrazioni di glucosio hanno inibito la produzione di GnRH attraverso la Leptina.
Conclusioni
Anche se ottenute in vitro, le scoperte fatte in seguito allo studio qui riportato supportano l’idea di un deleterio contributo diretto dell’iperglicemia sui neuroni del GnRH umano, fornendo così nuove informazioni sui meccanismi patogenetici che collegano l’ipogonadismo ipogonadotropico ai disordini metabolici.
I ricercatori affermano però di non essere pienamente sicuri delle loro scoperte: e quindi se ciò che hanno osservato nelle cellule che hanno usato è rappresentativo di ciò che accade a tutte le cellule che producono il GnRH nel cervello.
Nota: La ricerca svolta non è stata finanziata da un produttore di prodotti dietetici a basso contenuto di carboidrati, ma dal governo italiano.
Quello che mi accingo a riportare in questo articolo è il protocollo di GH per la perdita di grasso ideato da Chest Rockwell.
Storia del GH e perdita di grasso
Cos’è la lipolisi?
Relazione tra ormone dello stress / GH e digiuno
Meccanismi d’azione – Lipolisi GH-mediata
Tasso massimo di lipolisi
Farmacocinetica / Farmacodinamica
Sinergie tra i composti
Applicazione pratica e progettazione di uno stack di esempio
Storia del GH e perdita di grasso
Ipotalamo
L’Ormone della Crescita (GH) è un potente agente di mobilizzazione dei grassi i cui effetti, se bene all’epoca non ancora imputabili all’ormone in se, vennero osservati dai ricercatori fin dagli anni ’20, quando primitivi studi sugli animali dimostrarono che gli esemplari trattati con l’ipofisi erano più magri rispetto agli animali di controllo non trattati [1-2]. Tuttavia, si dovette attendere fino al 1945, quando il GH fu estratto per la prima volta dall’ipofisi [3], affinché gli scienziati iniziassero veramente a isolare gli effetti di mobilizzazione dei grassi dell’ipofisi su questo specifico polipeptide.
Cos’è la lipolisi?
Triacilglicerolo
La lipolisi è il processo fisiologico che fornisce all’organismo un substrato energetico sotto forma di acidi grassi attraverso il catabolismo del triacilglicerolo immagazzinato. Il triacilglicerolo immagazzinato all’interno delle goccioline lipidiche degli adipociti viene idrolizzato in acidi grassi e glicerolo e successivamente rilasciato nel circolo ematico per ossidazione e produzione di ATP [4-5]. Sarà importante capire le differenze tra mobilitazione e ossidazione dei grassi; tuttavia una revisione completa della lipolisi va oltre lo scopo di questo articolo.
Relazione tra ormone dello stress / GH e digiuno
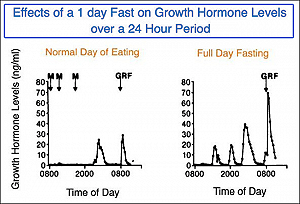
Il GH è definibile come “ormone dello stress”, per sua natura, con l’aumento della sua secrezione durante eventi stressanti come il digiuno e l’esercizio fisico [6]. Per il resto di questo articolo, tuttavia, ci concentreremo maggiormente sul ruolo che il GH svolge durante il digiuno e su come sia applicabile a coloro che cercano di massimizzare il potenziale lipolitico dei loro protocolli di GH esogeno.
Il periodo di digiuno (post-riassorbimento) inizia circa sei ore dopo l’assunzione di cibo e gli obiettivi primari del corpo durante questa fase sono di fornire, convertire e conservare i substrati energetici. Durante questo periodo, vi è un aumento significativo del tasso di secrezione di GH che può durare per 48-72 ore [7-8]. L’innalzamento del GH secreto è direttamente correlato all’ampiezza dell’impulso, mentre la frequenza degli impulsi e i livelli di intervallo basali rimangono sostanzialmente invariati [9]. Durante il digiuno, il GH è l’unico ormone anabolico ad aumentare mentre aumentano i livelli degli ormoni catabolici (ad esempio Glucagone, Cortisolo, Epinefrina ecc.).
Meccanismi d’azione – Lipolisi GH-mediata
Le elevate percentuali di secrezione di GH portano con sé numerosi cambiamenti metabolici, che sono importanti da comprendere. La prima priorità del corpo durante il digiuno è quella di mantenere l’omeostasi del glucosio, in modo da fornire sufficiente glucosio al cervello e ad altri tessuti (ad esempio globuli rossi) che dipendono, principalmente o tassativamente (tessuti glucosio-prefereziali e glucosio-dipendenti), da questo substrato energetico [10]. Per raggiungere questo obiettivo, si verifica uno shift metabolico a favore del metabolismo lipidico in modo che vengano salvaguardate le preziose riserve glucidiche e aminoacidiche. Parallelamente a questo cambiamento shift metabolico a favore del metabolismo lipidico da parte del tessuto muscolare ed epatico, viene attuata la mobilizzazione del glicogeno in quanto non viene rilevato alcun apporto dietetico di glucosio. Grandi quantità di glucosio vengono anche rilasciate dal fegato nel flusso ematico per aiutare a mantenere i livelli di glucosio nel sangue in assenza di un apporto di glucosio con la dieta. Ciò è possibile, in gran parte, grazie al calo simultaneo dei livelli sierici di insulina che impedisce al glucosio rilasciato di entrare nei muscoli e nel tessuto adiposo.
Oltre a questo, con l’aumento del GH si verifica uno stato di insulino-resistenza, vitale per la conservazione delle preziose riserve di glucosio. Questi effetti antagonisti all’insulina che il GH porta con sé riducono l’ossidazione del glucosio e, al contrario, la necessità di precursori gluconeogenici delle riserve di proteine muscolari [11]. Ci sarebbero alcune riflessioni da fare sull’eventualità che sia lo stesso GH o l’aumento degli FFA i principali responsabili dell’instaurarsi di una accentuata insulino-resistenza, tuttavia, la questione potrà essere trattata in un altro articolo. Quindi, per legare tutto ciò che è stato fino ad ora detto, durante il digiuno l’aumento della secrezione di GH porta ad un aumento della mobilitazione degli FFA dagli adipociti, sotto-regola i GLUT-1 per inibire l’assorbimento di glucosio nei tessuti periferici, previene l’ossidazione del glucosio aumentando l’insulino-resistenza e preserva le riserve amminiche sia in modo diretto che indiretto.
È ben noto che il GH influenza la lipolisi, tuttavia i meccanismi esatti con cui lo fa rimangono in qualche modo elusivi. È stato ipotizzato che questo potrebbe essere multifattoriale con il GH il quale mostra la capacità di ridurre la lipoproteina lipasi del tessuto adiposo (LPL), stimolare la lipasi ormono-sensibile (HSL) e antagonizzare gli effetti antilipolitici dell’insulina. L’aumentata espressione del HSL negli adipociti aumenta il potenziale lipolitico in quanto l’HSL è intimamente coinvolto nel processo di idrolisi del triacilglicerolo. Una volta attivato, l’HSL viene trasportato alla periferia della goccia lipidica intracellulare dove idrolizza il triacilglicerolo a FFA e glicerolo. È anche riconosciuto da molti come l’enzima che determina la velocità della lipolisi [12]. Vale la pena notare che non tutti gli studi hanno universalmente dimostrato che il GH aumenta i livelli di mRNA del HSL negli adipociti [13-14].
Come già accennato, è stato anche dimostrato che il GH ha un impatto diretto sulla soppressione dell’attività della LPL nei tessuti adiposi umani, sebbene ciò non sia stato dimostrato nei tessuti muscoli scheletrici [15-17]. Il motivo per il quale ciò è potenzialmente rilevante per chi è interessato alla perdita di grasso è legato al fatto che la LPL è direttamente coinvolta nella liberazione degli acidi grassi nel flusso ematico, nel loro successivo immagazzinamento negli adipociti e / o renderli disponibili per la beta-ossidazione da parte dei tessuti muscolo scheletrici e del muscolo cardiaco. Quindi, se la LPL può essere soppressa nei tessuti adiposi, si potrebbe ipotizzare che meno acidi grassi potranno essere immagazzinati attivamente (riesterificati) mentre altri saranno disponibili per alimentare i processi metabolici.
Studi condotti su adipociti umani in coltura hanno dimostrato che il GH in realtà non ha effetti lipolitici diretti [18], ma aumenta significativamente la sensibilità alle catecolamine in queste cellule, suggerendo che il GH attivi la lipolisi in uno stadio successivo al coinvolgimento dei beta-adrenorecettori e / o Proteine-G. In effetti, è ragionevole supporre che il GH possa aumentare la densità dei beta-adrenorecettori, rendendo questo fattore di grande interesse per un protocollo volto a massimizzare la lipolisi. È stato precedentemente dimostrato che esistono beta-adrenocettori “di riserva” sugli adipociti umani e un aumento acuto del numero di recettori accoppiati aumenterebbe la sensibilità e, in definitiva, il potenziale lipolitico [19]. E, in modelli animali, è stato dimostrato che il GH aumenta specificamente l’espressione dei recettori β3-adrenergici negli adipociti, seguita dall’attivazione del HSL [20]. Quindi, oltre agli effetti direttamente mediati che il GH possiede, è ovvio che l’uso di un agonista beta-adrenergico potrebbe molto probabilmente creare un effetto additivo sul processo lipolitico.
Tasso massimo di lipolisi
Ormone della Crescita (GH)
Quindi, mentre ci si adopera per creare uno stack che massimizzi il potenziale lipolitico del GH, esiste un limite effettivo (o teorico) alla velocità con cui ciò si verifichi? In realtà esiste una risposta; almeno per quanto riguarda esclusivamente il tasso massimo in cui il GH con somministrazione endovenosa possa provocare la lipolisi [21]. Si è constatata una dose di circa 3 mcg / kg (corrispondente a un picco medio di GH di 32,4 mcg / litro). La dose non era dipendente dall’età o dal sesso e si aggira intorno all’equivalente di 1,2-1,5 UI per un maschio magro di 100 kg. Una dose più elevata di questa non provoca in realtà un maggiore impatto sulla lipolisi. E’ interessante notare come questo sia anche essenzialmente il limite superiore dei picchi secretori endogeni naturali [22]. Si ipotizza che questo potrebbe essere un limite, o un collo di bottiglia, causato almeno in parte dai tassi di clearance extra-renale in concomitanza con i livelli di GHBP circolanti [23].
Ci sono alcune prove che questo sia un collo di bottiglia specifico del GH, e che i trattamenti combinati con varianti delle catecolamine producono un effetto additivo sulla lipolisi, maggiore di entrambi i singoli trattamenti [24-27] che supportano ulteriormente l’idea che il GH sia un mediatore lipolitico di mediazione. Alcuni hanno definito questo come un effetto permissivo sulla lipolisi indotta da catecolamine [28]. Aneddoticamente, sembra che un associazione di GH e composti catecolamine-stimolanti [29] abbia un effetto sinergico. Ma di questo se ne parlerà più approfonditamente proseguendo con questo articolo.
Farmacocinetica / Farmacodinamica
La farmacocinetica è una branca della farmacologia che studia gli effetti che i processi dell’organismo hanno sul farmaco (assorbimento, distribuzione, metabolismo, eliminazione). La farmacodinamica, invece, è lo studio degli effetti biochimici e fisiologici dei farmaci sull’organismo, ed il loro meccanismo d’azione [30]. È abbastanza importante capire entrambi, in relazione al GH, al fine di massimizzare il suo potenziale lipolitico e garantire che il GH somministrato sia ottimizzato così da prevenire la stesura di protocolli non corretti. Il GH lo sappiamo bene che è un composto costoso, quindi sarebbe nell’interesse dell’utilizzatore cercare di non sprecarlo.
Le vie di somministrazione modificano notevolmente la farmacocinetica del GH. Come discusso in precedenza, il GH endogeno viene secreto in maniera pulsatile e la sua presenza nel siero viene rapidamente eliminata attraverso i naturali feedback negativi di regolazione del corpo. Per simulare più strettamente questo comportamento secretorio, è necessario ricorrere alla somministrazione endovenosa una volta ogni 2-3 ore, che è il tempo necessario affinché il feedback ultra-corto (GHRH che inibisce la sua stessa secrezione) conduca al calo ematico dell’ormone. Tuttavia, a meno che non si è sotto la supervisione di un medico, questa non è una pratica che un qualsiasi preparatore degno di tale appellativo consiglierebbe e, infatti, in questo articolo ci si concentrerà invece sui due metodi di somministrazione del GH più popolari; sottocutaneo e intramuscolare.
A seguito di una iniezione SC o IM di GH, i livelli plasmatici di FFA e glicerolo aumentano dopo un breve periodo di latenza con un picco nei tassi di mobilizzazione dei grassi raggiunto intorno ai 150-160 minuti.
L’altro punto importante che vale la pena di sottolineare riguarda la tempistica d’iniezione del GH. Lo stato di digiuno è associato non solo a un aumento pronunciato della secrezione di GH endogeno, come precedentemente descritto, ma anche a un aumento della sensibilità lipolitica data dalla somministrazione di GH esogeno [33]. Infatti, tutti i marker lipolitici sono migliorati quando il GH viene somministrato a digiuno rispetto a quando viene somministrato in stato digestivo / postprandiale [34]. Anche il tasso di clearance globale è aumentato in modo significativo rimanendo nello stato di digiuno.
Vale anche la pena di notare che la somministrazione serale di GH ha dimostrato di avere una maggiore biodisponibilità, almeno quando somministrata a soggetti con deficit dell’Ormone della Crescita (GHD) [35]. Ciò potrebbe essere dovuto al fatto che tale somministrazioni mimi in modo ravvicinato il rilascio endogeno di GH nelle ore serali, ma questa è in gran parte solo una semplice speculazione.
Sinergie tra i composti
Recettore Beta-2 adrenergico
Quando si progetta uno stack finalizzato alla perdita di grasso che massimizza il potenziale lipolitico, bisogna scegliere con logica l’inserimento di composti con una potenziale sinergia, o con effetti additivi l’uno sull’altro. In precedenza, abbiamo discusso alcuni dei meccanismi mediante i quali il GH esercita i suoi effetti lipolitici. Uno dei suoi percorsi chiave coinvolge i recettori beta-adrenergici. Quindi sembrerebbe plausibile che se si è in grado di aumentare la sensibilità e / o l’espressione dei recettori beta-adrenergici, si possa ulteriormente ottimizzare la risposta lipolitica complessiva dello stack.
Recettore degli Androgeni
Gli androgeni inducono potenti effetti lipolitici direttamente attraverso i recettori degli androgeni (AR) espressi nel tessuto adiposo [36]. Questo è interessante, in sé e per sé, perché è un percorso diverso rispetto a quello utilizzato dal GH e, anche, perché è stato dimostrato che gli androgeni aumentano l’espressione dei recettore beta-adrenergici [37-38]. Si è già discusso del’aumento del numero dei recettori beta-adrenergici e della loro sensibilità e, in definitiva, come questo si rifletta positivamente sul potenziale lipolitico. Quindi questa potrebbe essere una potente sinergia nella metodologia di progettazione di uno stack.
L’asse tiroideo e l’asse GH / IGF hanno una relazione molto interessante, anche se complicata, l’una con l’altra. Di particolare interesse per il design di uno stack lipolitico sono i suoi impatti sull’mRNA del recettore beta-adrenergico e più specificamente i suoi impatti sull’espressione dei β3, che ha dimostrato di essere un passo critico nella lipolisi GH-mediata [39-41]. Inoltre gli ormoni tiroidei stimolano la sintesi, la degradazione e la mobilizzazione dei lipidi con conseguente aumento dei livelli degli FFA circolanti [42]. E l’ultima caratteristica sinergica che vale la pena di menzionare è che essi, proprio come il GH, sembrano regolare la sensibilità alle catecolamine dei processi metabolici.
E una volta aumentata la sensibilità e la densità dei recettori beta-adrenergici, è ovvio che un agonista beta-adrenergico sarebbe la ciliegina sulla torta per massimizzare il potenziale di mobilizzazione del grasso. Il Clenbuterolo [43] o il Salbutamolo sono entrambi candidati eccellenti per far parte di uno stack lipolitico sinergico; tuttavia una revisione completa degli agonisti beta-adrenergici va oltre lo scopo di questo articolo.
Applicazione pratica e progettazione di uno stack di esempio
Ora che si hanno le informazioni che stanno alla base di una progettazione oculata di uno stack lipolitico sinergico e avente come colonna portante l’uso del GH, come fare a progettare lo stack ed a gestire i composti facenti parte di esso?
Penso che sia abbastanza evidente il fatto che la somministrazione di GH (a fini lipolitici) debba essere fatta a digiuno. Sebbene gli effetti lipolitici del GH non siano completamente attenuati in presenza di cibo (diversamente dal GH endogeno), somministrandolo a stomaco pieno si eserciterebbe un serio antagonismo nei confronti dell’azione ricercata.
Utilizzando le informazioni esposte in precedenza riguardo al digiuno, insieme a una dose ben calibrata per ottenere la massima risposta lipolitica, si giunge all’applicazione di un’unica iniezione da 2UI di GH. Svolgere tale somministrazione al mattino appena svegli soddisfacerebbe molto bene i fini della pratica, poiché ci si troverà in uno stato di digiuno, più che probabile. Poiché le iniezioni sottocutanee hanno un tempo di eliminazione più lungo, questo sarebbe un buon metodo di somministrazione, soprattutto se si prevede di rimanere a digiuno per molte ore dopo l’iniezione. Per un potenziale effetto additivo sui tassi di mobilizzazione del grasso, vorrei anche prendere in considerazione l’esecuzione di attività strutturate (sia sotto forma di LISS che di allenamento di resistenza) durante questa finestra di digiuno [44].
Seguendo le stesse linee guida, si potrebbe molto probabilmente somministrare una seconda iniezione di 2UI di GH prima di andare a letto, sempre se si è in grado di consumare tutti i pasti della giornata in un lasso di tempo nel complesso ridotto ed entrare così nelle ore serali in uno stato di semi-digiuno. Come illustrato in precedenza, vi è anche evidenza che la somministrazione serale di GH per via sottocutanea fornisca una maggiore biodisponibilità. Quindi, se uno è costretto a scegliere dovrebbe farlo prendendo in considerazione una somministrazione AM o PM, in quanto questo sarebbe il tempo di somministrazione raccomandato. In caso contrario, l’utilizzo di una strategia che comprenda due iniezione da 2IU di GH, una somministrazione AM ed una PM, può teoricamente massimizzare il potenziale lipolitico nel corso delle 24 ore, assumendo che aderisca alle linee guida sul digiuno/semi-digiuno corrette.
Testosterone
L’uso di questo protocollo di dosaggio di GH, in combinazione con una co-somministrazione di androgeni, può aumentare il potenziale complessivo sulla perdita di grasso. Per questo motivo si dovrebbe prendere in considerazione l’uso di dosi sovra fisiologiche di AAS, oltre al fatto che tale inserimento previene il rischio di atrofia del tessuto magro durante periodi sostenuti di deficit calorico. Ci possono essere piccole differenze nel tasso di perdita di grasso prodotto dall’utilizzo di diversi tipi di androgeni, ed esso è strettamente legato al potenziale androgeno del composto, ma questo va oltre lo scopo di questo articolo. Anche il semplice inserimento di Testosterone nello stack funzionerebbe molto bene per lo scopo prefissato.
Aggiungendo sia composti tiroidei che Clenbuterolo, o Salbutamolo, nello stack si massimizzerà ulteriormente il potenziale lipolitico globale basato sulle loro proprietà sinergiche addizionate a quelle tra androgeni e GH spiegate in precedenza. A causa delle loro rispettive proprietà, il Clenbuterolo può essere somministrato una volta al giorno come pure il T4. Per quelli che usano il T3, si raccomanda un minimo di due dosi al giorno, distribuite a distanza di 12 ore. Chi sceglie di usare il Salbutamolo è raccomandabile distribuire la dose totale in più somministrazioni durante il giorno (4-5h di distanza l’una dall’atra).
Per riunire quanto detto fino ad ora, e al fine di renderlo comprensibile ai più, qui di seguito viene esposto uno schema breve ma decisamente esaustivo:
Cosomministrazione di AAS soprafisiologico
Cosomministrazione di Clenbuterolo o Salbutamolo + tiroidei
GH somministrato per via sottocutanea a una dose non superiore a 2 UI a digiuno due volte al giorno (AM / PM) o una volta al giorno (PM) se si è costretti a scegliere.
Attività strutturata eseguita durante lo stato di digiuno, post-iniezione per aumentare il potenziale di mobilizzazione del grasso.
Nota conclusiva
L’articolo ” The Most Effective Growth Hormone Protocol for Fat Loss”, il quale è stato utilizzato per la stesura del presente articolo, è un estratto sintetico ed abbreviato della sezione GH presente nel libro “The Growth Hormone Handbook” (titolo provvisorio) di Chester “Chest” Rockwell.
Gabriel Bellizzi
Riferimenti:
National Research Council (US) Committee on Technological Options to Improve the Nutritional Attributes of Animal Products. Designing Foods: Animal Product Options in the Marketplace. Washington (DC): National Academies Press (US); 1988. The Role of Growth Hormone in Fat Mobilization
Lee, M. O., and Schaffer, N. K.: Anterior Pituitary Growth Hormone and the Composition of Growth , J. Nutrition 7: 337 ( (March 10) ) 1934.
Li CH, Evans HM, Simpson ME. Isolation and properties of the anterior hypophyseal growth hormone. J Biol Chem. 1945;159:353–366.
Schoemaker RC, Buijs MM, Pijl H, Burggraaf J, Cohen AF. Modeling the influence of growth hormone on lipolysis. J Pharmacokinet Pharmacodyn. 2002 Apr;29(2):157-70
Lass A, Zimmermann R, Oberer M, Zechner R. Lipolysis – A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Progress in Lipid Research. 2011;50(1-4):14-27.
ROTH J, GLICK SM, YALOW RS, BERSON SA. Secretion of human growth hormone:physiologic and experimental modification. Metabolism. 1963 Jul;12:577-9
Hartman ML, Veldhuis JD, Johnson ML, Lee MM, Alberti KG, Samojlik E, Thorner MO. Augmented growth hormone (GH) secretory burst frequency and amplitude mediate enhanced GH secretion during a two-day fast in normal men. J Clin Endocrinol Metab. 1992 Apr;74(4):757-65.
Ho PJ, Friberg RD, Barkan AL. Regulation of pulsatile growth hormone secretion by fasting in normal subjects and patients with acromegaly. J Clin Endocrinol Metab. 1992 Sep;75(3):812-9.
Goldenberg N, Barkan A. Factors regulating growth hormone secretion in humans. Endocrinol Metab Clin North Am. 2007 Mar;36(1):37-55. Review
Berg JM, Tymoczko JL, Stryer L. Biochemistry. 5th edition. New York: W H Freeman; 2002. Section 30.3, Food Intake and Starvation Induce Metabolic Changes.
Nørrelund H. The metabolic role of growth hormone in humans with particular reference to fasting. Growth Horm IGF Res. 2005 Apr;15(2):95-122. Review.
Birnbaum MJ. Lipolysis: more than just a lipase. J Cell Biol. 2003 Jun 23;161(6):1011-2. Epub 2003 Jun 16. Review.
Slavin BG, Ong JM, Kern PA. Hormonal regulation of hormone-sensitive lipase activity and mRNA levels in isolated rat adipocytes. J Lipid Res. 1994 Sep;35(9):1535-41.
Ottosson M, Vikman-Adolfsson K, Enerbäck S, Elander A, Björntorp P, Edén S. Growth hormone inhibits lipoprotein lipase activity in human adipose tissue. J Clin Endocrinol Metab. 1995 Mar;80(3):936-41.
Richelsen B. Effect of growth hormone on adipose tissue and skeletal muscle lipoprotein lipase activity in humans. J Endocrinol Invest. 1999;22(5 Suppl):10-5. Review.
Richelsen B, Pedersen SB, Kristensen K, Børglum JD, Nørrelund H, Christiansen JS, Jørgensen JO. Regulation of lipoprotein lipase and hormone-sensitive lipase activity and gene expression in adipose and muscle tissue by growth hormone treatment during weight loss in obese patients. Metabolism. 2000 Jul;49(7):906-11.
Marcus C, Margery V, Kamel A, Brönnegård M. Effects of growth hormone on lipolysis in humans. Acta Paediatr Suppl. 1994 Dec;406:54-8; discussion 59. Review.
Arner P, Hellmér J, Wennlund A, Ostman J, Engfeldt P. Adrenoceptor occupancy in isolated human fat cells and its relationship with lipolysis rate. Eur J Pharmacol. 1988 Jan 27;146(1):45-56
Heffernan M, Summers RJ, Thorburn A, Ogru E, Gianello R, Jiang WJ, Ng FM. The effects of human GH and its lipolytic fragment (AOD9604) on lipid metabolism following chronic treatment in obese mice and beta(3)-AR knock-out mice. Endocrinology. 2001 Dec;142(12):5182-9.
Hansen TK, Gravholt CH, ØRskov H, Rasmussen MH, Christiansen JS, Jørgensen JO. Dose dependency of the pharmacokinetics and acute lipolytic actions of growth hormone. J Clin Endocrinol Metab. 2002 Oct;87(10):4691-8
Hartman ML, Faria AC, Vance ML, Johnson ML, Thorner MO, Veldhuis JD. Temporal structure of in vivo growth hormone secretory events in humans. Am J Physiol. 1991 Jan;260
Haffner D, Schaefer F, Girard J, Ritz E, Mehls O 1994 Metabolic clearance of recombinant human growth hormone in health and chronic renal failure. J Clin Invest 93:1163–1171
Fain JN. Effect of dibutyryl-3′,5′-AMP, theophylline and norepinephrine on lipolytic action of growth hormone and glucocorticoid in white fat cells. Endocrinology. 1968 Apr;82(4):825-30.
Goodman HM. Effects of growth hormone on the lipolytic response of adipose tissue to theophylline. Endocrinology. 1968 May;82(5):1027-34.
Yip RG, Goodman HM. Growth hormone and dexamethasone stimulate lipolysis and activate adenylyl cyclase in rat adipocytes by selectively shifting Gi alpha2 to lower density membrane fractions. Endocrinology. 1999 Mar;140(3):1219-27.
Fain JN, Cheema P, Tichansky DS, Madan AK. Stimulation of human omental adipose tissue lipolysis by growth hormone plus dexamethasone. Mol Cell Endocrinol. 2008 Nov 25;295(1-2):101-5.
Goodman HM 1970 Permissive effects of hormones on lipolysis. Endocrinology 86:1064–1074
Davis E, Loiacono R, Summers RJ. The rush to adrenaline: drugs in sport acting on the β-adrenergic system. British Journal of Pharmacology. 2008;154(3):584-597
Greenblatt, D. J. and Abourjaily, P. N. (2016), Pharmacokinetics and Pharmacodynamics for Medical Students: A Proposed Course Outline. The Journal of Clinical Pharmacology, 56: 1180–1195.
Keller A, Wu Z, Kratzsch J, Keller E, Blum WF, Kniess A, Preiss R, Teichert J, Strasburger CJ, Bidlingmaier M. Pharmacokinetics and pharmacodynamics of GH:dependence on route and dosage of administration. Eur J Endocrinol. 2007 Jun;156(6):647-53
Laursen T. Clinical pharmacological aspects of growth hormone administration. Growth Horm IGF Res. 2004 Feb;14(1):16-44. Review.
Møller N, Pørksen N, Ovesen P, Alberti KG. Evidence for increased sensitivity of fuel mobilization to growth hormone during short-term fasting in humans. Horm Metab Res. 1993 Mar;25(3):175-9.
Moller L, Dalman L, Norrelund H, Billestrup N, Frystyk J, Moller N, Jorgensen JO. Impact of fasting on growth hormone signaling and action in muscle and fat. J Clin Endocrinol Metab. 2009 Mar;94(3):965-72
Jørgensen JO, Møller N, Lauritzen T, Alberti KG, Orskov H, Christiansen JS. Evening versus morning injections of growth hormone (GH) in GH-deficient patients: effects on 24-hour patterns of circulating hormones and metabolites. J Clin Endocrinol Metab. 1990 Jan;70(1):207-14.
Blouin K, Veilleux A, Luu-The V, Tchernof A. Androgen metabolism in adipose tissue: recent advances. Mol Cell Endocrinol. 2009 Mar 25;301(1-2):97-103
Xu X, De Pergola G, Björntorp P. The effects of androgens on the regulation of lipolysis in adipose precursor cells. Endocrinology. 1990 Feb;126(2):1229-34
De Pergola G. The adipose tissue metabolism: role of testosterone and dehydroepiandrosterone. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S59-63. Review
Viguerie N, Millet L, Avizou S, Vidal H, Larrouy D, Langin D. Regulation of human adipocyte gene expression by thyroid hormone. J Clin Endocrinol Metab. 2002 Feb;87(2):630-4
Rubio A, Raasmaja A, Silva JE. Thyroid hormone and norepinephrine signaling in brown adipose tissue. II: Differential effects of thyroid hormone on beta 3-adrenergic receptors in brown and white adipose tissue. Endocrinology. 1995 Aug;136(8):3277-84.
Ghosh M, Das S. Increased beta(2)-adrenergic receptor activity by thyroid hormone possibly leads to differentiation and maturation of astrocytes in culture. Cell Mol Neurobiol. 2007 Dec;27(8):1007-21. Epub 2007 Sep 8
Pucci E, Chiovato L, Pinchera A. Thyroid and lipid metabolism. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S109-12. Review
Kim HK, Della-Fera MA, Hausman DB, Baile CA. Effect of clenbuterol on apoptosis, adipogenesis, and lipolysis in adipocytes. J Physiol Biochem. 2010 Sep;66(3):197-203
Taaffe DR, Thompson JL, Butterfield GE, Hoffman AR, Marcus R. Recombinant human growth hormone, but not insulin-like growth factor-I, enhances central fat loss in postmenopausal women undergoing a diet and exercise program. Horm Metab Res. 2001 Mar;33(3):156-62.
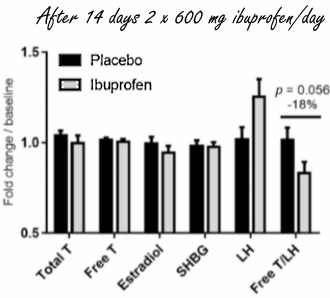
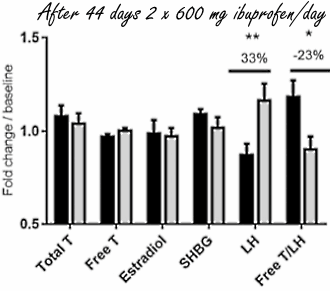
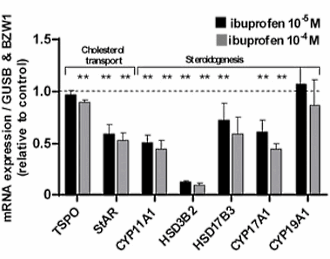
Ci sono molti atleti che assumono antidolorifici come l’Ibuprofene poco prima di un allenamento o di una competizione. Sopprimendo il dolore dato dallo sforzo a carico delle articolazioni, soprattutto di quelle che hanno subito un infortunio, l’uso degli antidolorifici permette all’atleta di ottenere prestazioni migliori – a breve termine. A lungo termine questi atleti potrebbero, in base alla ricerca danese recentemente pubblicata sul PNAS, abbassare i livelli di Testosterone endogeno. (1)
Gli endocrinologi danesi, affiliati all’Università di Copenaghen, hanno diviso 31 uomini sani di età compresa tra i 18 ed i 35 anni in 2 gruppi. Un gruppo ha assunto 2 volte al giorno una dose di 600 milligrammi di Ibuprofene per 6 settimane. Questa è la dose massima indicata nel bugiardino del farmaco. L’altro gruppo ha assunto un placebo.
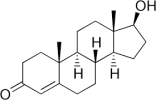
Dopo due settimane di somministrazione di Ibuprofen, la concentrazione di LH – l’ormone che stimola le cellule di Leydig a produrre Testosterone – nel flusso ematico era aumentata significativamente nei soggetti trattati, e dopo 6 settimane quell’effetto era diventato ancora più marcato. Tuttavia, la concentrazione di Testosterone non ha subito un aumento consequenziale.
Con l’avanzare dell’età, i testicoli possono avere sempre più problemi a mantenere un livello adeguato e stabile di Testosterone. Ecco perché il livello di LH nel sangue aumenta. Nei primi stadi di questo processo di invecchiamento ormonale, a causa dell’aumento del livello di LH, il rilascio di Testosterone rimane costante. Gli endocrinologi in tale circostanza parlano quindi di “ipogonadismo compensatorio”.
In una fase successiva, tuttavia, i testicoli diventano insensibili all’LH e, di conseguenza, la secrezione di Testosterone cala. Il livello di Testosterone circolante subisce un calo. Gli endocrinologi in tale circostanza parlano quindi di “ipogonadismo manifesto”.
Sembra che l’uso a lungo termine di dosi elevate di Ibuprofene imiti e forse addirittura acceleri questo processo di invecchiamento endocrinologico.
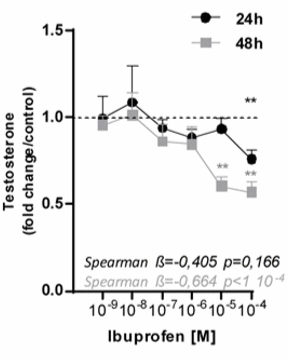
Quando i ricercatori hanno svolto esperimenti in vitro con campioni di testicoli umani, hanno osservato che la sintesi di Testosterone era ridotta dall’Ibuprofene. Questo effetto diventava più marcato quanto più le concentrazioni di Ibuprofene erano elevate e quanto più l’esposizione a tale composto era prolungata.
L’Ibuprofene ha ridotto l’effetto di letteralmente tutti gli enzimi coinvolti nella sintesi di Testosterone dal Colesterolo nei testicoli.
Come accennato poc’anzi, In ambito clinico, la compromissione della funzione delle cellule di Leydig che determina una maggiore insensibilità all’LH è definita come ipogonadismo compensatorio, un fattore associato alla mortalità per tutte le cause.(2) Pertanto, i ricercatori affermano che lo studio dell’ipogonadismo compensatorio indotto dall’Ibuprofene è cruciale, in quanto questo stato clinico è generalmente associato al fumo e all’invecchiamento.
Ulteriori caratterizzazioni dello stato di ipogonadismo compensatorio indotto dall’Ibuprofene, che era già stato stabilito dopo 14 giorni di somministrazione del farmaco, sono quindi importanti nel determinare i potenziali effetti su giovani uomini sani.
Diverse relazioni hanno sottolineato l’alto livello di uso di analgesici a lungo termine tra gli atleti dilettanti e professionisti; l’Ibuprofene è risultato un composto favorito in questo uso e abuso. Da notare, una relazione inversa è stata recentemente riportata tra l’esercizio fisico di resistenza e la libido sessuale maschile, ma la possibilità che l’assunzione di farmaci possa interferire in questa osservazione non può essere totalmente esclusa.(3)
Inoltre, l’Ibuprofene sembra essere l’analgesico farmaceutico preferito per il trattamento del dolore cronico a lungo termine e l’artrite.
Pertanto, in tale contesto, è anche preoccupante che gli uomini con ipogonadismo compensatorio possano eventualmente progredire verso l’ipogonadismo primario manifesto, caratterizzato da un basso livello di Testosterone circolante e da sintomi connessi tra i quali vi sono la riduzione della libido, la riduzione della massa e della forza muscolare, il calo dell’umore, affaticamento e depressione.
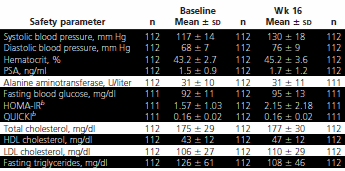
Un guadagno di massa magra di 3 kg e una perdita di grasso di 2 kg: questo è il risultato modesto seguito a uno studio di 16 settimane nelle quali uomini tra i 70 e gli 80 anni sono stati trattati con somministrazioni di Ormone della Crescita e Testosterone. I medici americani hanno pubblicato i risultati del loro studio svolto su 112 uomini sul Journal of Clinical Endocrinology & Metabolism. (1)
Tra i 70 e gli 80 anni, tra gli anziani sedentari si sperimenta una perdita del 30% della forza muscolare. Questa è la stessa perdita che si verifica tra i 50 ed i 70 anni. Ciò significa che oltre i 70 anni la forza muscolare diminuisce con il doppio della velocità. Lo scopo dello studio che qui riporto era quello di scoprire se l’Ormone della Crescita e il Testosterone avessero qualche effetto su questa condizione legata all’età. E’ bene ricordare che i soggetti dello studio non hanno fatto alcun allenamento.
I ricercatori hanno arrestato la produzione di Testosterone endogeno dei soggetti dando loro del Leuprolide. Quindi hanno diviso i soggetti in 6 gruppi.
I soggetti del gruppo A si sono applicati 5g di Testosterone in gel ogni giorno, così da portare i livelli ematici dell’ormone ai livelli normali per gli individui della loro età. Il Testosterone in gel utilizzato era stato prodotto da Solvay.
Al gruppo B, oltre al Testosterone, è stata somministrata un’iniezione di circa 0,75UI di Ormoni della Crescita 2-3 ore dopo il pasto serale. Questa quantità ha un effetto anabolico negli adulti che per ragioni mediche non producono abbastanza Ormone della Crescita. La preparazione utilizzata è stata prodotta dalla Genentech. Ai soggetti del gruppo C è stata somministrata una dose di circa 1,27 UI di Ormone della Crescita.
I soggetti del gruppo D si sono applicato giornalmente 10g di Testosterone in gel. Questo ha fatto si che i soggetti in questione raggiungessero concentrazioni ematiche dell’ormone simili ad un uomo di giovane età.
Ai soggetti del gruppo E, oltre alla somministrazione di Testosterone, è stata iniettata una dose giornaliera di 0,75 UI di Ormone della Crescita, mentre ai soggetti del gruppo F sono state somministrate 1,27 UI di Ormone della Crescita al giorno.
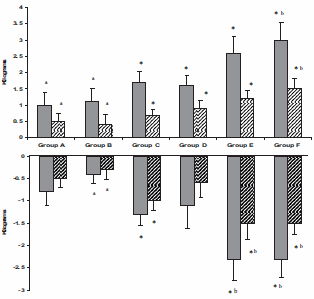
La figura seguente mostra l’aumento della massa magra nei soggetti dello studio. Le barre scure rappresentano la massa magra delle braccia e delle gambe. La figura in basso mostra la quantità di grasso che i soggetti hanno perso. Le barre scure rappresentano la massa grassa nel tronco. I risultati confermano ciò che i Bodybuilder sanno già: l’Ormone della Crescita e il Testosterone rafforzano reciprocamente i loro effetti.
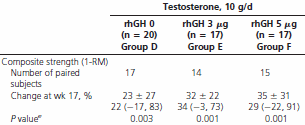
I soggetti non hanno migliorato la propria forma a causa del trattamento, ma hanno subito un aumento della forza. Ciò si è verificato soltanto nel gruppo trattato con 10g di Testosterone in gel. Quando questo quantitativo era combinato con l’Ormone della Crescita, la forza dei soggetti ha subito un aumento del 30%. Tale percentuale equivale alla perdita di forza media riscontrabile tra i 70 e gli 80 anni.
Il trattamento ha causato un aumento della pressione sanguigna aumentando la densità ematica (aumento dell’ematocrito). E’ stato osservato anche un aumento del PSA. I marker per il diabete, come il livello della glicemia al mattino, HOMA-IR e il QUICKI, sono aumentati, così come i livelli di trigliceridi. Gli effetti erano statisticamente significativi, ma non erano motivo di allarme per i medici.
Tuttavia i ricercatori non si esprimono su quali possano essere gli effetti di un trattamento simili sul lungo termine. Essi suggeriscono che le persone che intendono assumere Testosterone e Ormone della Crescita come terapia “Anti-Aging” dovrebbero essere sotto costante controllo medico.
I ricercatori concludono dicendo che, la somministrazione combinata di dosi fisiologiche di Testosterone e GH ha portato a guadagni sostanziali nella massa magra e nella forza dei muscoli volontari, insieme ad una riduzioni del grasso totale e del tronco che era di maggiore entità rispetto a quanto riscontrato con il trattamento con solo Testosterone.
Il presente studio mancava di un tassello fondamentale che da solo avrebbe potuto modificare marcatamente i risultati ottenuti, l’attività fisica! Un adeguata attività fisica, da sola, esercita un effetto contrastante considerevole contro la sarcopenia e la perdita di forza legata all’età.
Gli uomini con maggiori concentrazioni ematiche di Magnesio molto probabilmente hanno maggiori concentrazioni ematiche di Testosterone libero. Questa conclusione è stata riportata in un articolo pubblicato sul Journal of Pharmaceutical and Biomedical Analysis. (1)
Circa il 2% del Testosterone circolante nel flusso ematico è attivo: non è legato alle proteine di trasporto che impediscono al Testosterone di interagire con il recettore degli androgeni. Circa il 40% del Testosterone circolante è legato all’albumina, una proteina che non impedisce all’ormone di svolgere la sua attività. Il Testosterone libero e il Testosterone legato all’albumina sono indicati come Testosterone biodisponibile.
Circa il 60% del Testosterone circolante è legato alle Globuline leganti gli ormoni sessuali (SHBG). Gli Androgeni legati alle SHBG perdono il loro effetto anabolizzante, ma probabilmente mantengono il loro effetto androgenico. Nella prostata, per esempio, ci sono recettori per le SHBG i quali inviano segnali di “errore” alle cellule della prostata se ad essi si legano SHBG legate ad androgeni. Gli ormoni steroidei androgeni legati alle SHBG hanno quindi potenziali effetti indesiderati, ma senza effetti desiderabili.
Man mano che un uomo invecchia, aumentano le concentrazioni di SHBG e con esse il Testosterone legato. Questo avviene anche perché gli uomini più anziani mangiano meno proteine. Un basso consumo proteico aumenta le concentrazioni di SHBG nel sangue. Un apporto proteico più elevato porta a concentrazioni maggiori di albumina e aumenta la quantità di Testosterone biodisponibile. Entro certi limiti, ovviamente.
I ricercatori, associati all’Université de Franche-Comte, hanno estratto dal sangue di giovani uomini le SHBG esponendole in seguito agli ioni di Magnesio. Successivamente i ricercatori hanno misurato la velocità con cui il Testosterone si legava alle SHBG all’aumentare delle concentrazioni di Magnesio. Maggiore era la concentrazione di magnesio, minore era l’attrazione tra l’ormone e la proteina legante.
Anche se i ricercatori non hanno esaminato se maggiori concentrazioni di Magnesio portino effettivamente a maggiori concentrazioni di Testosterone libero negli esseri umani, credono che i loro risultati siano significativi a livello fisiologico.
I ricercatori scrivono che i risultati presentati forniscono evidenza per una variazione mediata dall’Mg2 + nell’associazione Testosterone-SHBG, suggerendo che un aumento della concentrazione di Mg2 + all’interno dell’intervallo di concentrazione biologica (0,75mM-1,0mM) potrebbe portare ad un miglioramento del Testosterone biodisponibile.
Venticinque anni fa dei ricercatori hanno esaminato l’effetto di concentrazioni di Magnesio estremamente elevate e biologicamente improbabili. Ciò ha portato ad un piccolo calo dei livelli di Testosterone. (2)
La ricerca negli ultimi anni si è spinta avanti prendendo in esame l’effetto delle sostanze vegetali sul legame del testosterone con le SHBG (vedere, a proposito, l’effetto del Fieno Greco sul Testosterone libero).
Il Magnesio è un minerale presente negli alimenti di origine vegetale. Buone fonti sono i cereali integrali, gli spinaci, le noci e i fagioli.
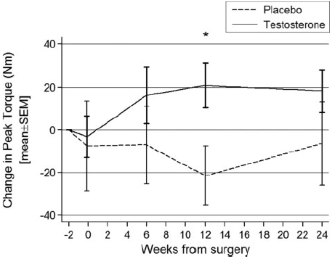
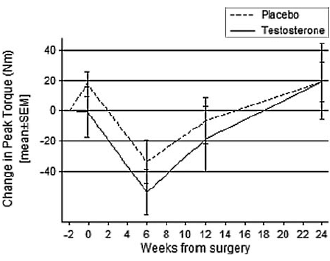
Se gli uomini assumono una modesta quantità di Testosterone nel periodo precedente e successivo alla ricostruzione del legamento crociato anteriore (LCA), guadagnano più massa magra e rafforzano i muscoli. Ma una terapia con Testosterone in tale contesto sembra non apportare miglioramenti nella forza dei muscoli della gamba che è stata operata, come riportato dai medici della Keck School of Medicine. (1)
I ricercatori hanno eseguito un’operazione del legamento crociato anteriore su 13 uomini di età compresa tra i 18 ei 50 anni. Sei uomini hanno ricevuto un’iniezione settimanale di 200 mg di Testosterone Cypionato nel periodo dell’operazione. Gli altri sette uomini hanno ricevuto un iniezioni priva di principio attivo [Placebo].
La somministrazione delle iniezioni contenenti Testosterone Cypionato è cominciata due settimane prima dell’operazione ed è proseguita per le sei settimane successive, quindi la terapia con Testosterone è durata per un totale di otto settimane. Durante questo periodo, ovviamente, il livello di Testosterone ematico degli uomini del gruppo sperimentale era più alto. Anche la quantità di massa magra degli uomini facenti parte di questo gruppo è aumentata di un paio di chilogrammi. Dopo il termine della terapia con Testosterone, i soggetti trattati riuscirono a conservare una buona quantità di massa magra extra.
Il Testosterone ha aumentato il “Torque”- la misura della forza intorno all’asse di rotazione di un’articolazione – nella gamba “buona” degli uomini, quella che non presentava i legamenti danneggiati. I ricercatori hanno misurato il “Torque” nei muscoli delle gambe dei soggetti facendoli lavorare su una Leg-Extension.
Tuttavia, la terapia con Testosterone non ha avuto alcun effetto sul “Torque” che gli uomini erano in grado di sviluppare nel ginocchio che aveva subito l’operazione, mentre i ricercatori speravano che la sua forza fosse aumentata.
Questo è il primo studio che analizza gli effetti della terapia con Testosterone perioperatoria in uomini giovani e in buona salute dopo la ricostruzione del legamento crociato anteriore.
Testosterone Cypionato
I ricercatori riportano che, i pazienti che sono stati trattati con 200mg di Testosterone Cypionato per un periodo di 8 settimane hanno riscontrato un maggiore aumento della massa magra a 6 settimane dalla ricostruzione del legamento crociato anteriore rispetto ai pazienti trattati con placebo. L’aumento della massa magra suggerisce che il Testosterone può aiutare a minimizzare gli effetti dell’atrofia muscolare associati a lesioni, interventi chirurgici e immobilità. Questo aumento di massa magra è stato osservato persistere dopo il ritorno dei livelli sierici di Testosterone al basale.
Ulteriori indagini sono necessarie per determinare il profilo di sicurezza e gli effetti della somministrazione di Testosterone perioperatorio sulla forza delle gambe e sugli esiti clinici dopo l’intervento.
Il grasso in eccesso ha la capacità di alterare negativamente i livelli di Androgeni negli uomini più di quanto si possa pensare. Gli enzimi espressi nel tessuto adiposo non solo aumentano la conversione del Testosterone in Estradiolo (vedi enzima aromatasi), ma convertono il Diidrotestosterone (DHT) in composti inattivi. (1)
Gli uomini in sovrappeso presentano più spesso una carenza di ormoni androgeni rispetto agli uomini magri. In alcuni studi i medici hanno trattato il problema con un inibitore della aromatasi come il Letrozolo (2), e ci sono anche casi di uomini in sovrappeso che hanno visto migliorare le loro condizioni di salute in conseguenza di iniezioni di Testosterone.
Ma gli uomini in sovrappeso che ripongono tutta la loro fiducia nella terapia con Testosterone senza cercare di ridurre il loro peso corporeo (e la percentuale di grasso) potrebbero fare decisamente la scelta sbagliata. Più gli uomini presentano una percentuale di grasso elevata, meno essi reagiscono positivamente alla terapia con Testosterone (3). E per di più, gli uomini in sovrappeso sono anche più propensi a sviluppare effetti collaterali durante la terapia con Testosterone (4) (vedi, appunto, aumento dell’espressione dell’enzima aromatasi con conseguente aumento dell’aromatizzazione del Testosterone in Estradiolo).
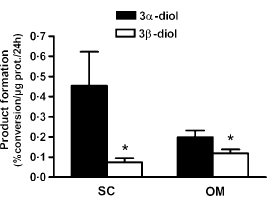
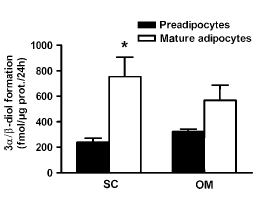
I ricercatori canadesi della Laval University hanno dimostrato che gli enzimi prodotti nel tessuto adiposo disattivano il DHT e lo fanno ad un tasso maggiore in misura direttamente proporzionale alle riserve adipose. Più un uomo è grasso, più DHT viene convertito di conseguenza in 5-alfa-androstano 3-alfa, 17-beta-diolo e 5-alfa-androstano 3-beta, 17-beta-diolo. Ovviamente, se ci fosse il bisogno di sottolinearlo, l’uso di inibitori dell’aromatasi non ha alcun effetto su tale conversione.
I ricercatori hanno esaminato campioni di tessuto adiposo sottocutaneo e intra-addominale prelevati da uomini grassi e magri. Hanno scoperto che specialmente nel grasso sottocutaneo vi è un alto livello di conversione del DHT in 5-alfa-androstano-3-alfa, 17-beta-diolo. Gli enzimi che convertono il DHT erano più attivi nelle cellule adipose mature [mature adipocytes] rispetto alle cellule adipose che non avevano ancora raggiunto la maturità [pre adipocytes].
Quindi più è alta la percentuale di grasso, più DHT viene convertito in metaboliti inattivi. Ma non finisce qui la storia, come si può vedere dal grafico qui sopra riportato. Se gli uomini aumentano di peso, quindi di kg di peso corporeo, gli enzimi disattivanti il DHT diventano più attivi.
Gli Strongman ed i Powerlifter che seguono ancora l’insana abitudine di non curanza verso la loro massa grassa, hanno un altro ottimo motivo per modificare la gestione della loro preparazione sportiva.
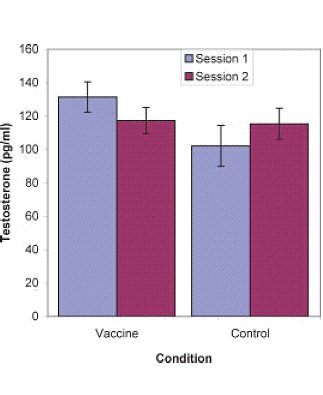
Se il sistema immunitario deve adattarsi a contrastare un agente patogeno, il livello di Testosterone cala. Questo è anche quello che accade se ci si sottopone a vaccino contro un virus, come riportato dai ricercatori dell’University of California a Santa Barbara. Gli studenti che hanno ricevuto il vaccino anti influenzale hanno sperimentato una leggera riduzione dei livelli di Testosterone per circa due settimane dopo l’inoculazione. (1)
Come tutti sanno bene, il Testosterone è un ormone anabolizzante. Quanto più alto è il livello di Testosterone, più il corpo è orientato alla riproduzione e alla crescita muscolare e meno sul sistema immunitario.
Ma il sistema immunitario ha bisogno di energia. Il sistema immunitario è alla continua ricerca di nuovi “intrusi” nel sistema e quando li trova attiva una categoria di cellule in grado di combatterli. Cosa che costa energia: un livello di androgeni elevato è negativo per il sistema immunitario in quanto gli androgeni “sottraggono” energia per il sistema immunitario.
Ecco un motivo per il quale naturalmente si producono solo quantità limitate di androgeni. La natura non è stata gentile con gli individui che sintetizzano elevate quantità di androgeni.
Questa è la teoria almeno. Ma regge con la realtà?
I ricercatori americani hanno voluto verificare la teoria svolgendo un esperimento con un gruppo di circa quaranta studenti. Gli studenti hanno ricevuto un vaccino anti influenzale. Un vaccino contiene un agente patogeno inattivato, che non fa ammalare realmente il soggetto al quale viene inoculato, ma stimola il sistema immunitario a dare il via alla produzione di un nuovo tipo di cellule immunitarie. Ciò consente alla persona di combattere la malattia se sono esposti al vero virus. La vaccinazioni è come se dessero al sistema immunitario un corso intensivo di adattamento, che costa al corpo meno energia rispetto al dover combattere un’infezione da parte del agente patogeno “originale”.
I ricercatori hanno misurato i livelli di Testosterone di venti studenti prima che questi venissero sottoposti a vaccino anti influenzale, e hanno ripetuto il controllo due settimane dopo. Hanno fatto i medesimi esami con un gruppo di controllo che non aveva ricevuto vaccinazioni. I risultati hanno mostrato un leggero calo dei livelli di Testosterone nei soggetti ai quali era stato somministrato il vaccino anti influenzale.
I ricercatori presumono che un’infezione reale avrebbe avuto un effetto maggiore sul livello di Testosterone, ma i risultati dell’esperimento hanno per lo meno confermato la loro teoria: “I risultati attuali suggeriscono che l’esposizione ad agenti patogeni può causare una diminuzione del Testosterone, presumibilmente come mezzo per dare priorità all’investimento energetico verso il miglioramento delle risposte immunitarie rispetto alla riproduzione”.






 riproduzione. L’FSH è rilasciato nel circolo portale dell’adenoipofisi e poi nel circolo sanguineo sistemico dove rimane attivo per 3-4 ore. La sua funzione è principalmente di regolazione dello sviluppo dei follicoli ovarici e, quindi, della stimolazione della produzione di Estrogeno e Progesterone.
riproduzione. L’FSH è rilasciato nel circolo portale dell’adenoipofisi e poi nel circolo sanguineo sistemico dove rimane attivo per 3-4 ore. La sua funzione è principalmente di regolazione dello sviluppo dei follicoli ovarici e, quindi, della stimolazione della produzione di Estrogeno e Progesterone.