
Quello che mi accingo a riportare in questo articolo è il protocollo di GH per la perdita di grasso ideato da Chest Rockwell.
- Storia del GH e perdita di grasso
- Cos’è la lipolisi?
- Relazione tra ormone dello stress / GH e digiuno
- Meccanismi d’azione – Lipolisi GH-mediata
- Tasso massimo di lipolisi
- Farmacocinetica / Farmacodinamica
- Sinergie tra i composti
- Applicazione pratica e progettazione di uno stack di esempio
Storia del GH e perdita di grasso

L’Ormone della Crescita (GH) è un potente agente di mobilizzazione dei grassi i cui effetti, se bene all’epoca non ancora imputabili all’ormone in se, vennero osservati dai ricercatori fin dagli anni ’20, quando primitivi studi sugli animali dimostrarono che gli esemplari trattati con l’ipofisi erano più magri rispetto agli animali di controllo non trattati [1-2]. Tuttavia, si dovette attendere fino al 1945, quando il GH fu estratto per la prima volta dall’ipofisi [3], affinché gli scienziati iniziassero veramente a isolare gli effetti di mobilizzazione dei grassi dell’ipofisi su questo specifico polipeptide.
Cos’è la lipolisi?

La lipolisi è il processo fisiologico che fornisce all’organismo un substrato energetico sotto forma di acidi grassi attraverso il catabolismo del triacilglicerolo immagazzinato. Il triacilglicerolo immagazzinato all’interno delle goccioline lipidiche degli adipociti viene idrolizzato in acidi grassi e glicerolo e successivamente rilasciato nel circolo ematico per ossidazione e produzione di ATP [4-5]. Sarà importante capire le differenze tra mobilitazione e ossidazione dei grassi; tuttavia una revisione completa della lipolisi va oltre lo scopo di questo articolo.
Relazione tra ormone dello stress / GH e digiuno
Il GH è definibile come “ormone dello stress”, per sua natura, con l’aumento della sua secrezione durante eventi stressanti come il digiuno e l’esercizio fisico [6]. Per il resto di questo articolo, tuttavia, ci concentreremo maggiormente sul ruolo che il GH svolge durante il digiuno e su come sia applicabile a coloro che cercano di massimizzare il potenziale lipolitico dei loro protocolli di GH esogeno.
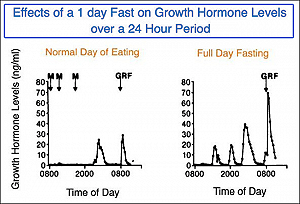
Il periodo di digiuno (post-riassorbimento) inizia circa sei ore dopo l’assunzione di cibo e gli obiettivi primari del corpo durante questa fase sono di fornire, convertire e conservare i substrati energetici. Durante questo periodo, vi è un aumento significativo del tasso di secrezione di GH che può durare per 48-72 ore [7-8]. L’innalzamento del GH secreto è direttamente correlato all’ampiezza dell’impulso, mentre la frequenza degli impulsi e i livelli di intervallo basali rimangono sostanzialmente invariati [9]. Durante il digiuno, il GH è l’unico ormone anabolico ad aumentare mentre aumentano i livelli degli ormoni catabolici (ad esempio Glucagone, Cortisolo, Epinefrina ecc.).
Meccanismi d’azione – Lipolisi GH-mediata
Le elevate percentuali di secrezione di GH portano con sé numerosi cambiamenti metabolici, che sono importanti da comprendere. La prima priorità del corpo durante il digiuno è quella di mantenere l’omeostasi del glucosio, in modo da fornire sufficiente glucosio al cervello e ad altri tessuti (ad esempio globuli rossi) che dipendono, principalmente o tassativamente (tessuti glucosio-prefereziali e glucosio-dipendenti), da questo substrato energetico [10]. Per raggiungere questo obiettivo, si verifica uno shift metabolico a favore del metabolismo lipidico in modo che vengano salvaguardate le preziose riserve glucidiche e aminoacidiche. Parallelamente a questo cambiamento shift metabolico a favore del metabolismo lipidico da parte del tessuto muscolare ed epatico, viene attuata la mobilizzazione del glicogeno in quanto non viene rilevato alcun apporto dietetico di glucosio. Grandi quantità di glucosio vengono anche rilasciate dal fegato nel flusso ematico per aiutare a mantenere i livelli di glucosio nel sangue in assenza di un apporto di glucosio con la dieta. Ciò è possibile, in gran parte, grazie al calo simultaneo dei livelli sierici di insulina che impedisce al glucosio rilasciato di entrare nei muscoli e nel tessuto adiposo.
Oltre a questo, con l’aumento del GH si verifica uno stato di insulino-resistenza, vitale per la conservazione delle preziose riserve di glucosio. Questi effetti antagonisti all’insulina che il GH porta con sé riducono l’ossidazione del glucosio e, al contrario, la necessità di precursori gluconeogenici delle riserve di proteine muscolari [11]. Ci sarebbero alcune riflessioni da fare sull’eventualità che sia lo stesso GH o l’aumento degli FFA i principali responsabili dell’instaurarsi di una accentuata insulino-resistenza, tuttavia, la questione potrà essere trattata in un altro articolo. Quindi, per legare tutto ciò che è stato fino ad ora detto, durante il digiuno l’aumento della secrezione di GH porta ad un aumento della mobilitazione degli FFA dagli adipociti, sotto-regola i GLUT-1 per inibire l’assorbimento di glucosio nei tessuti periferici, previene l’ossidazione del glucosio aumentando l’insulino-resistenza e preserva le riserve amminiche sia in modo diretto che indiretto.

È ben noto che il GH influenza la lipolisi, tuttavia i meccanismi esatti con cui lo fa rimangono in qualche modo elusivi. È stato ipotizzato che questo potrebbe essere multifattoriale con il GH il quale mostra la capacità di ridurre la lipoproteina lipasi del tessuto adiposo (LPL), stimolare la lipasi ormono-sensibile (HSL) e antagonizzare gli effetti antilipolitici dell’insulina. L’aumentata espressione del HSL negli adipociti aumenta il potenziale lipolitico in quanto l’HSL è intimamente coinvolto nel processo di idrolisi del triacilglicerolo. Una volta attivato, l’HSL viene trasportato alla periferia della goccia lipidica intracellulare dove idrolizza il triacilglicerolo a FFA e glicerolo. È anche riconosciuto da molti come l’enzima che determina la velocità della lipolisi [12]. Vale la pena notare che non tutti gli studi hanno universalmente dimostrato che il GH aumenta i livelli di mRNA del HSL negli adipociti [13-14].
Come già accennato, è stato anche dimostrato che il GH ha un impatto diretto sulla soppressione dell’attività della LPL nei tessuti adiposi umani, sebbene ciò non sia stato dimostrato nei tessuti muscoli scheletrici [15-17]. Il motivo per il quale ciò è potenzialmente rilevante per chi è interessato alla perdita di grasso è legato al fatto che la LPL è direttamente coinvolta nella liberazione degli acidi grassi nel flusso ematico, nel loro successivo immagazzinamento negli adipociti e / o renderli disponibili per la beta-ossidazione da parte dei tessuti muscolo scheletrici e del muscolo cardiaco. Quindi, se la LPL può essere soppressa nei tessuti adiposi, si potrebbe ipotizzare che meno acidi grassi potranno essere immagazzinati attivamente (riesterificati) mentre altri saranno disponibili per alimentare i processi metabolici.
Studi condotti su adipociti umani in coltura hanno dimostrato che il GH in realtà non ha effetti lipolitici diretti [18], ma aumenta significativamente la sensibilità alle catecolamine in queste cellule, suggerendo che il GH attivi la lipolisi in uno stadio successivo al coinvolgimento dei beta-adrenorecettori e / o Proteine-G. In effetti, è ragionevole supporre che il GH possa aumentare la densità dei beta-adrenorecettori, rendendo questo fattore di grande interesse per un protocollo volto a massimizzare la lipolisi. È stato precedentemente dimostrato che esistono beta-adrenocettori “di riserva” sugli adipociti umani e un aumento acuto del numero di recettori accoppiati aumenterebbe la sensibilità e, in definitiva, il potenziale lipolitico [19]. E, in modelli animali, è stato dimostrato che il GH aumenta specificamente l’espressione dei recettori β3-adrenergici negli adipociti, seguita dall’attivazione del HSL [20]. Quindi, oltre agli effetti direttamente mediati che il GH possiede, è ovvio che l’uso di un agonista beta-adrenergico potrebbe molto probabilmente creare un effetto additivo sul processo lipolitico.
Tasso massimo di lipolisi

Quindi, mentre ci si adopera per creare uno stack che massimizzi il potenziale lipolitico del GH, esiste un limite effettivo (o teorico) alla velocità con cui ciò si verifichi? In realtà esiste una risposta; almeno per quanto riguarda esclusivamente il tasso massimo in cui il GH con somministrazione endovenosa possa provocare la lipolisi [21]. Si è constatata una dose di circa 3 mcg / kg (corrispondente a un picco medio di GH di 32,4 mcg / litro). La dose non era dipendente dall’età o dal sesso e si aggira intorno all’equivalente di 1,2-1,5 UI per un maschio magro di 100 kg. Una dose più elevata di questa non provoca in realtà un maggiore impatto sulla lipolisi. E’ interessante notare come questo sia anche essenzialmente il limite superiore dei picchi secretori endogeni naturali [22]. Si ipotizza che questo potrebbe essere un limite, o un collo di bottiglia, causato almeno in parte dai tassi di clearance extra-renale in concomitanza con i livelli di GHBP circolanti [23].
Ci sono alcune prove che questo sia un collo di bottiglia specifico del GH, e che i trattamenti combinati con varianti delle catecolamine producono un effetto additivo sulla lipolisi, maggiore di entrambi i singoli trattamenti [24-27] che supportano ulteriormente l’idea che il GH sia un mediatore lipolitico di mediazione. Alcuni hanno definito questo come un effetto permissivo sulla lipolisi indotta da catecolamine [28]. Aneddoticamente, sembra che un associazione di GH e composti catecolamine-stimolanti [29] abbia un effetto sinergico. Ma di questo se ne parlerà più approfonditamente proseguendo con questo articolo.
Farmacocinetica / Farmacodinamica
La farmacocinetica è una branca della farmacologia che studia gli effetti che i processi dell’organismo hanno sul farmaco (assorbimento, distribuzione, metabolismo, eliminazione). La farmacodinamica, invece, è lo studio degli effetti biochimici e fisiologici dei farmaci sull’organismo, ed il loro meccanismo d’azione [30]. È abbastanza importante capire entrambi, in relazione al GH, al fine di massimizzare il suo potenziale lipolitico e garantire che il GH somministrato sia ottimizzato così da prevenire la stesura di protocolli non corretti. Il GH lo sappiamo bene che è un composto costoso, quindi sarebbe nell’interesse dell’utilizzatore cercare di non sprecarlo.
Le vie di somministrazione modificano notevolmente la farmacocinetica del GH. Come discusso in precedenza, il GH endogeno viene secreto in maniera pulsatile e la sua presenza nel siero viene rapidamente eliminata attraverso i naturali feedback negativi di regolazione del corpo. Per simulare più strettamente questo comportamento secretorio, è necessario ricorrere alla somministrazione endovenosa una volta ogni 2-3 ore, che è il tempo necessario affinché il feedback ultra-corto (GHRH che inibisce la sua stessa secrezione) conduca al calo ematico dell’ormone. Tuttavia, a meno che non si è sotto la supervisione di un medico, questa non è una pratica che un qualsiasi preparatore degno di tale appellativo consiglierebbe e, infatti, in questo articolo ci si concentrerà invece sui due metodi di somministrazione del GH più popolari; sottocutaneo e intramuscolare.
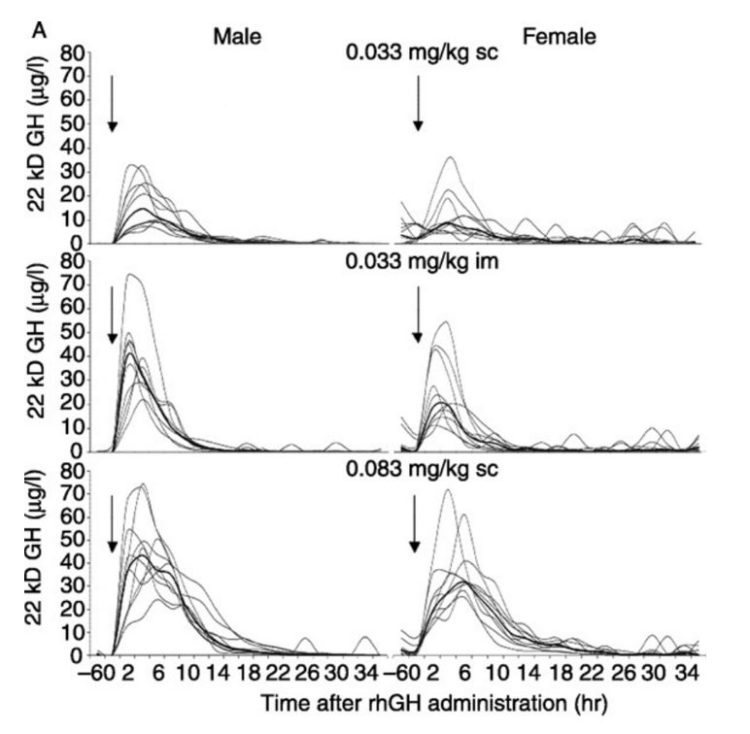
A seguito di una iniezione SC o IM di GH, i livelli plasmatici di FFA e glicerolo aumentano dopo un breve periodo di latenza con un picco nei tassi di mobilizzazione dei grassi raggiunto intorno ai 150-160 minuti.
L’altro punto importante che vale la pena di sottolineare riguarda la tempistica d’iniezione del GH. Lo stato di digiuno è associato non solo a un aumento pronunciato della secrezione di GH endogeno, come precedentemente descritto, ma anche a un aumento della sensibilità lipolitica data dalla somministrazione di GH esogeno [33]. Infatti, tutti i marker lipolitici sono migliorati quando il GH viene somministrato a digiuno rispetto a quando viene somministrato in stato digestivo / postprandiale [34]. Anche il tasso di clearance globale è aumentato in modo significativo rimanendo nello stato di digiuno.
Vale anche la pena di notare che la somministrazione serale di GH ha dimostrato di avere una maggiore biodisponibilità, almeno quando somministrata a soggetti con deficit dell’Ormone della Crescita (GHD) [35]. Ciò potrebbe essere dovuto al fatto che tale somministrazioni mimi in modo ravvicinato il rilascio endogeno di GH nelle ore serali, ma questa è in gran parte solo una semplice speculazione.
Sinergie tra i composti

Quando si progetta uno stack finalizzato alla perdita di grasso che massimizza il potenziale lipolitico, bisogna scegliere con logica l’inserimento di composti con una potenziale sinergia, o con effetti additivi l’uno sull’altro. In precedenza, abbiamo discusso alcuni dei meccanismi mediante i quali il GH esercita i suoi effetti lipolitici. Uno dei suoi percorsi chiave coinvolge i recettori beta-adrenergici. Quindi sembrerebbe plausibile che se si è in grado di aumentare la sensibilità e / o l’espressione dei recettori beta-adrenergici, si possa ulteriormente ottimizzare la risposta lipolitica complessiva dello stack.

Gli androgeni inducono potenti effetti lipolitici direttamente attraverso i recettori degli androgeni (AR) espressi nel tessuto adiposo [36]. Questo è interessante, in sé e per sé, perché è un percorso diverso rispetto a quello utilizzato dal GH e, anche, perché è stato dimostrato che gli androgeni aumentano l’espressione dei recettore beta-adrenergici [37-38]. Si è già discusso del’aumento del numero dei recettori beta-adrenergici e della loro sensibilità e, in definitiva, come questo si rifletta positivamente sul potenziale lipolitico. Quindi questa potrebbe essere una potente sinergia nella metodologia di progettazione di uno stack.
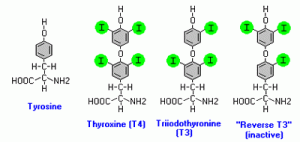
L’asse tiroideo e l’asse GH / IGF hanno una relazione molto interessante, anche se complicata, l’una con l’altra. Di particolare interesse per il design di uno stack lipolitico sono i suoi impatti sull’mRNA del recettore beta-adrenergico e più specificamente i suoi impatti sull’espressione dei β3, che ha dimostrato di essere un passo critico nella lipolisi GH-mediata [39-41]. Inoltre gli ormoni tiroidei stimolano la sintesi, la degradazione e la mobilizzazione dei lipidi con conseguente aumento dei livelli degli FFA circolanti [42]. E l’ultima caratteristica sinergica che vale la pena di menzionare è che essi, proprio come il GH, sembrano regolare la sensibilità alle catecolamine dei processi metabolici.
Per approfondire la questione GH e ormoni tiroidei vi rimando al mio articolo dedicato
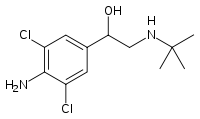
E una volta aumentata la sensibilità e la densità dei recettori beta-adrenergici, è ovvio che un agonista beta-adrenergico sarebbe la ciliegina sulla torta per massimizzare il potenziale di mobilizzazione del grasso. Il Clenbuterolo [43] o il Salbutamolo sono entrambi candidati eccellenti per far parte di uno stack lipolitico sinergico; tuttavia una revisione completa degli agonisti beta-adrenergici va oltre lo scopo di questo articolo.
Applicazione pratica e progettazione di uno stack di esempio
Ora che si hanno le informazioni che stanno alla base di una progettazione oculata di uno stack lipolitico sinergico e avente come colonna portante l’uso del GH, come fare a progettare lo stack ed a gestire i composti facenti parte di esso?
Penso che sia abbastanza evidente il fatto che la somministrazione di GH (a fini lipolitici) debba essere fatta a digiuno. Sebbene gli effetti lipolitici del GH non siano completamente attenuati in presenza di cibo (diversamente dal GH endogeno), somministrandolo a stomaco pieno si eserciterebbe un serio antagonismo nei confronti dell’azione ricercata.

Utilizzando le informazioni esposte in precedenza riguardo al digiuno, insieme a una dose ben calibrata per ottenere la massima risposta lipolitica, si giunge all’applicazione di un’unica iniezione da 2UI di GH. Svolgere tale somministrazione al mattino appena svegli soddisfacerebbe molto bene i fini della pratica, poiché ci si troverà in uno stato di digiuno, più che probabile. Poiché le iniezioni sottocutanee hanno un tempo di eliminazione più lungo, questo sarebbe un buon metodo di somministrazione, soprattutto se si prevede di rimanere a digiuno per molte ore dopo l’iniezione. Per un potenziale effetto additivo sui tassi di mobilizzazione del grasso, vorrei anche prendere in considerazione l’esecuzione di attività strutturate (sia sotto forma di LISS che di allenamento di resistenza) durante questa finestra di digiuno [44].

Seguendo le stesse linee guida, si potrebbe molto probabilmente somministrare una seconda iniezione di 2UI di GH prima di andare a letto, sempre se si è in grado di consumare tutti i pasti della giornata in un lasso di tempo nel complesso ridotto ed entrare così nelle ore serali in uno stato di semi-digiuno. Come illustrato in precedenza, vi è anche evidenza che la somministrazione serale di GH per via sottocutanea fornisca una maggiore biodisponibilità. Quindi, se uno è costretto a scegliere dovrebbe farlo prendendo in considerazione una somministrazione AM o PM, in quanto questo sarebbe il tempo di somministrazione raccomandato. In caso contrario, l’utilizzo di una strategia che comprenda due iniezione da 2IU di GH, una somministrazione AM ed una PM, può teoricamente massimizzare il potenziale lipolitico nel corso delle 24 ore, assumendo che aderisca alle linee guida sul digiuno/semi-digiuno corrette.

L’uso di questo protocollo di dosaggio di GH, in combinazione con una co-somministrazione di androgeni, può aumentare il potenziale complessivo sulla perdita di grasso. Per questo motivo si dovrebbe prendere in considerazione l’uso di dosi sovra fisiologiche di AAS, oltre al fatto che tale inserimento previene il rischio di atrofia del tessuto magro durante periodi sostenuti di deficit calorico. Ci possono essere piccole differenze nel tasso di perdita di grasso prodotto dall’utilizzo di diversi tipi di androgeni, ed esso è strettamente legato al potenziale androgeno del composto, ma questo va oltre lo scopo di questo articolo. Anche il semplice inserimento di Testosterone nello stack funzionerebbe molto bene per lo scopo prefissato.
Aggiungendo sia composti tiroidei che Clenbuterolo, o Salbutamolo, nello stack si massimizzerà ulteriormente il potenziale lipolitico globale basato sulle loro proprietà sinergiche addizionate a quelle tra androgeni e GH spiegate in precedenza. A causa delle loro rispettive proprietà, il Clenbuterolo può essere somministrato una volta al giorno come pure il T4. Per quelli che usano il T3, si raccomanda un minimo di due dosi al giorno, distribuite a distanza di 12 ore. Chi sceglie di usare il Salbutamolo è raccomandabile distribuire la dose totale in più somministrazioni durante il giorno (4-5h di distanza l’una dall’atra).
Per riunire quanto detto fino ad ora, e al fine di renderlo comprensibile ai più, qui di seguito viene esposto uno schema breve ma decisamente esaustivo:
- Cosomministrazione di AAS soprafisiologico
- Cosomministrazione di Clenbuterolo o Salbutamolo + tiroidei
- GH somministrato per via sottocutanea a una dose non superiore a 2 UI a digiuno due volte al giorno (AM / PM) o una volta al giorno (PM) se si è costretti a scegliere.
- Attività strutturata eseguita durante lo stato di digiuno, post-iniezione per aumentare il potenziale di mobilizzazione del grasso.
Nota conclusiva
L’articolo ” The Most Effective Growth Hormone Protocol for Fat Loss”, il quale è stato utilizzato per la stesura del presente articolo, è un estratto sintetico ed abbreviato della sezione GH presente nel libro “The Growth Hormone Handbook” (titolo provvisorio) di Chester “Chest” Rockwell.
Gabriel Bellizzi
Riferimenti:
- National Research Council (US) Committee on Technological Options to Improve the Nutritional Attributes of Animal Products. Designing Foods: Animal Product Options in the Marketplace. Washington (DC): National Academies Press (US); 1988. The Role of Growth Hormone in Fat Mobilization
- Lee, M. O., and Schaffer, N. K.: Anterior Pituitary Growth Hormone and the Composition of Growth , J. Nutrition 7: 337 ( (March 10) ) 1934.
- Li CH, Evans HM, Simpson ME. Isolation and properties of the anterior hypophyseal growth hormone. J Biol Chem. 1945;159:353–366.
- Schoemaker RC, Buijs MM, Pijl H, Burggraaf J, Cohen AF. Modeling the influence of growth hormone on lipolysis. J Pharmacokinet Pharmacodyn. 2002 Apr;29(2):157-70
- Lass A, Zimmermann R, Oberer M, Zechner R. Lipolysis – A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Progress in Lipid Research. 2011;50(1-4):14-27.
- ROTH J, GLICK SM, YALOW RS, BERSON SA. Secretion of human growth hormone:physiologic and experimental modification. Metabolism. 1963 Jul;12:577-9
- Hartman ML, Veldhuis JD, Johnson ML, Lee MM, Alberti KG, Samojlik E, Thorner MO. Augmented growth hormone (GH) secretory burst frequency and amplitude mediate enhanced GH secretion during a two-day fast in normal men. J Clin Endocrinol Metab. 1992 Apr;74(4):757-65.
- Ho PJ, Friberg RD, Barkan AL. Regulation of pulsatile growth hormone secretion by fasting in normal subjects and patients with acromegaly. J Clin Endocrinol Metab. 1992 Sep;75(3):812-9.
- Goldenberg N, Barkan A. Factors regulating growth hormone secretion in humans. Endocrinol Metab Clin North Am. 2007 Mar;36(1):37-55. Review
- Berg JM, Tymoczko JL, Stryer L. Biochemistry. 5th edition. New York: W H Freeman; 2002. Section 30.3, Food Intake and Starvation Induce Metabolic Changes.
- Nørrelund H. The metabolic role of growth hormone in humans with particular reference to fasting. Growth Horm IGF Res. 2005 Apr;15(2):95-122. Review.
- Birnbaum MJ. Lipolysis: more than just a lipase. J Cell Biol. 2003 Jun 23;161(6):1011-2. Epub 2003 Jun 16. Review.
- Pedersen, S.B. & Børglum, Jens & Jorgensen, Jens & Richelsen, Bjørn. (1995). Growth hormone treatment of obese premenopausal women: Effects on isolated adipocyte metabolism. 2. 251-258.
- Slavin BG, Ong JM, Kern PA. Hormonal regulation of hormone-sensitive lipase activity and mRNA levels in isolated rat adipocytes. J Lipid Res. 1994 Sep;35(9):1535-41.
- Ottosson M, Vikman-Adolfsson K, Enerbäck S, Elander A, Björntorp P, Edén S. Growth hormone inhibits lipoprotein lipase activity in human adipose tissue. J Clin Endocrinol Metab. 1995 Mar;80(3):936-41.
- Richelsen B. Effect of growth hormone on adipose tissue and skeletal muscle lipoprotein lipase activity in humans. J Endocrinol Invest. 1999;22(5 Suppl):10-5. Review.
- Richelsen B, Pedersen SB, Kristensen K, Børglum JD, Nørrelund H, Christiansen JS, Jørgensen JO. Regulation of lipoprotein lipase and hormone-sensitive lipase activity and gene expression in adipose and muscle tissue by growth hormone treatment during weight loss in obese patients. Metabolism. 2000 Jul;49(7):906-11.
- Marcus C, Margery V, Kamel A, Brönnegård M. Effects of growth hormone on lipolysis in humans. Acta Paediatr Suppl. 1994 Dec;406:54-8; discussion 59. Review.
- Arner P, Hellmér J, Wennlund A, Ostman J, Engfeldt P. Adrenoceptor occupancy in isolated human fat cells and its relationship with lipolysis rate. Eur J Pharmacol. 1988 Jan 27;146(1):45-56
- Heffernan M, Summers RJ, Thorburn A, Ogru E, Gianello R, Jiang WJ, Ng FM. The effects of human GH and its lipolytic fragment (AOD9604) on lipid metabolism following chronic treatment in obese mice and beta(3)-AR knock-out mice. Endocrinology. 2001 Dec;142(12):5182-9.
- Hansen TK, Gravholt CH, ØRskov H, Rasmussen MH, Christiansen JS, Jørgensen JO. Dose dependency of the pharmacokinetics and acute lipolytic actions of growth hormone. J Clin Endocrinol Metab. 2002 Oct;87(10):4691-8
- Hartman ML, Faria AC, Vance ML, Johnson ML, Thorner MO, Veldhuis JD. Temporal structure of in vivo growth hormone secretory events in humans. Am J Physiol. 1991 Jan;260
- Haffner D, Schaefer F, Girard J, Ritz E, Mehls O 1994 Metabolic clearance of recombinant human growth hormone in health and chronic renal failure. J Clin Invest 93:1163–1171
- Fain JN. Effect of dibutyryl-3′,5′-AMP, theophylline and norepinephrine on lipolytic action of growth hormone and glucocorticoid in white fat cells. Endocrinology. 1968 Apr;82(4):825-30.
- Goodman HM. Effects of growth hormone on the lipolytic response of adipose tissue to theophylline. Endocrinology. 1968 May;82(5):1027-34.
- Yip RG, Goodman HM. Growth hormone and dexamethasone stimulate lipolysis and activate adenylyl cyclase in rat adipocytes by selectively shifting Gi alpha2 to lower density membrane fractions. Endocrinology. 1999 Mar;140(3):1219-27.
- Fain JN, Cheema P, Tichansky DS, Madan AK. Stimulation of human omental adipose tissue lipolysis by growth hormone plus dexamethasone. Mol Cell Endocrinol. 2008 Nov 25;295(1-2):101-5.
- Goodman HM 1970 Permissive effects of hormones on lipolysis. Endocrinology 86:1064–1074
- Davis E, Loiacono R, Summers RJ. The rush to adrenaline: drugs in sport acting on the β-adrenergic system. British Journal of Pharmacology. 2008;154(3):584-597
- Greenblatt, D. J. and Abourjaily, P. N. (2016), Pharmacokinetics and Pharmacodynamics for Medical Students: A Proposed Course Outline. The Journal of Clinical Pharmacology, 56: 1180–1195.
- Keller A, Wu Z, Kratzsch J, Keller E, Blum WF, Kniess A, Preiss R, Teichert J, Strasburger CJ, Bidlingmaier M. Pharmacokinetics and pharmacodynamics of GH:dependence on route and dosage of administration. Eur J Endocrinol. 2007 Jun;156(6):647-53
- Laursen T. Clinical pharmacological aspects of growth hormone administration. Growth Horm IGF Res. 2004 Feb;14(1):16-44. Review.
- Møller N, Pørksen N, Ovesen P, Alberti KG. Evidence for increased sensitivity of fuel mobilization to growth hormone during short-term fasting in humans. Horm Metab Res. 1993 Mar;25(3):175-9.
- Moller L, Dalman L, Norrelund H, Billestrup N, Frystyk J, Moller N, Jorgensen JO. Impact of fasting on growth hormone signaling and action in muscle and fat. J Clin Endocrinol Metab. 2009 Mar;94(3):965-72
- Jørgensen JO, Møller N, Lauritzen T, Alberti KG, Orskov H, Christiansen JS. Evening versus morning injections of growth hormone (GH) in GH-deficient patients: effects on 24-hour patterns of circulating hormones and metabolites. J Clin Endocrinol Metab. 1990 Jan;70(1):207-14.
- Blouin K, Veilleux A, Luu-The V, Tchernof A. Androgen metabolism in adipose tissue: recent advances. Mol Cell Endocrinol. 2009 Mar 25;301(1-2):97-103
- Xu X, De Pergola G, Björntorp P. The effects of androgens on the regulation of lipolysis in adipose precursor cells. Endocrinology. 1990 Feb;126(2):1229-34
- De Pergola G. The adipose tissue metabolism: role of testosterone and dehydroepiandrosterone. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S59-63. Review
- Viguerie N, Millet L, Avizou S, Vidal H, Larrouy D, Langin D. Regulation of human adipocyte gene expression by thyroid hormone. J Clin Endocrinol Metab. 2002 Feb;87(2):630-4
- Rubio A, Raasmaja A, Silva JE. Thyroid hormone and norepinephrine signaling in brown adipose tissue. II: Differential effects of thyroid hormone on beta 3-adrenergic receptors in brown and white adipose tissue. Endocrinology. 1995 Aug;136(8):3277-84.
- Ghosh M, Das S. Increased beta(2)-adrenergic receptor activity by thyroid hormone possibly leads to differentiation and maturation of astrocytes in culture. Cell Mol Neurobiol. 2007 Dec;27(8):1007-21. Epub 2007 Sep 8
- Pucci E, Chiovato L, Pinchera A. Thyroid and lipid metabolism. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S109-12. Review
- Kim HK, Della-Fera MA, Hausman DB, Baile CA. Effect of clenbuterol on apoptosis, adipogenesis, and lipolysis in adipocytes. J Physiol Biochem. 2010 Sep;66(3):197-203
- Taaffe DR, Thompson JL, Butterfield GE, Hoffman AR, Marcus R. Recombinant human growth hormone, but not insulin-like growth factor-I, enhances central fat loss in postmenopausal women undergoing a diet and exercise program. Horm Metab Res. 2001 Mar;33(3):156-62.
