
Introduzione:
Durante l’esercizio fisico, il sistema endocrino secerne ormoni nel sangue per regolare il metabolismo (Mckeever, 2002; Mastorakos et al., 2005) e mantenere l’integrità dell’ambiente interno del corpo. Pertanto, il controllo della secrezione ormonale deve essere complesso e sensibile per adattarsi rapidamente al cambiamento delle sollecitazioni biologiche dell’organismo durante l’esercizio fisico. Il Testosterone è un ormone steroideo anabolizzante che si trova nel sangue in tre forme: fortemente legato alla globulina legante gli ormoni sessuali (SHBG) (~70%), debolmente legato all’albumina (~30%) e non legato (~0,5-3%). Tradizionalmente, la funzione fisiologica del Testosterone nel tessuto muscolare scheletrico è il mantenimento e l’aumento della massa muscolare scheletrica (ipertrofia) attraverso meccanismi genomici (a lungo termine, trascrizionali) e il conseguente aumento indiretto della forza muscolare (Cardinale e Stone, 2006; Griggs et al., 1989). Tuttavia, è stato dimostrato che gli ormoni steroidei, tra cui il Testosterone, possono esercitare un’azione rapida (da pochi secondi a pochi minuti) in diversi tipi di cellule (Benten et al., 1997; 1999a; Ceballos et al., 1999; Estrada et al., 2003; Furukawa e Kurokawa, 2008; Hamdi e Mutungi, 2010; Jones et al., 2004; Waldkirch et al., 2008) attraverso meccanismi non genomici (a breve termine, non trascrizionali) (Benten et al., 1999b). Tuttavia, la complessa interazione tra l’adattamento acuto e quello a lungo termine degli steroidi non è ancora stata descritta. Mentre la significativa azione genomica del Testosterone nel muscolo scheletrico è ben descritta (Bhasin et al., 2005; Sinha-Hikim et al., 2002; 2003; Urban et al., 1995), poca attenzione è stata rivolta alle azioni non genomiche del Testosterone nel muscolo scheletrico. Questo articolo discuterà una modalità di azione non genomica nel muscolo scheletrico e le implicazioni proposte per le prestazioni sportive, in particolare per le atlete.
Cosa sono le azioni non genomiche degli steroidi?
Le azioni non genomiche degli ormoni steroidei sono quelle azioni mediate dagli steroidi in cui la trascrizione genica non è direttamente coinvolta (come dimostrato dall’insensibilità agli inibitori della trascrizione e della sintesi proteica), coinvolgono la partecipazione di secondi messaggeri e sono rapide nell’azione (da pochi secondi a pochi minuti). Come descritto più avanti in questo articolo, le azioni non genomiche si differenziano dai meccanismi genomici in primo luogo per il legame dello steroide a un recettore per gli androgeni situato sulla membrana cellulare o per il legame con un recettore della membrana plasmatica associato a una proteina G sensibile alla tossina di Pertussis (PTX) (Vicencio et al., 2006), anziché legarsi al tradizionale recettore per gli androgeni nel citoplasma della cellula prima di essere traslocato nel nucleo. A differenza degli effetti genomici degli ormoni steroidei, gli effetti non genomici richiedono la presenza costante dell’ormone. Una volta che l’ormone viene eliminato dal tessuto, anche gli effetti non genomici vengono meno.

Le prove di un’azione non tradizionale del Testosterone sono state documentate regolarmente in tessuti diversi dal muscolo scheletrico. All’inizio degli anni ’90, è stato riportato un rapido effetto del Testosterone sulla mobilità del calcio nelle cellule T, iniziato a livello della membrana cellulare (Benten et al., 1999b), suggerendo l’esistenza di una rapida risposta biologica alternativa al Testosterone. Analogamente, i miociti cardiaci di ratti adulti esposti al Testosterone hanno indotto rapidamente (1-7 minuti) un aumento dei livelli di calcio intracellulare rilasciato dai depositi intracellulari attraverso i recettori dell’inositolo trisfosfato (IP3). Questa risposta al calcio non era legata al recettore intracellulare degli androgeni, ma all’attivazione di un recettore di membrana plasmatica associato a una proteina G (PTX) sensibile (Vicencio et al., 2006). Analogamente, il Testosterone stimola rapidamente l’aumento della concentrazione di calcio negli osteoblasti di ratto (entro 5 s attraverso un maggiore afflusso di calcio) e una maggiore mobilizzazione di calcio dal reticolo endoplasmatico, oltre ad aumentare la formazione di IP3 entro 10 s (Lieberherr e Grosse, 1994). L’IP3 viene utilizzato per la trasduzione del segnale nelle cellule biologiche attraverso il rilascio di calcio dal reticolo endoplasmatico tramite il recettore IP3 (IP3R). Il Testosterone sembra esercitare queste azioni rapide attraverso una proteina G non identificata (le proteine che legano i nucleotidi alla guanina comunicano i segnali degli ormoni extracellulari che poi regolano i cambiamenti intracellulari) situata sulla membrana cellulare (Benten et al., 1999a; Vicencio et al., 2006).
Le azioni rapide del Testosterone nel muscolo scheletrico sono state meno studiate rispetto ad altri tessuti come il muscolo cardiaco e l’osso (Benten et al., 1997; 1999a), come già detto. Tuttavia, i pochi lavori che hanno studiato l’azione non genomica del Testosterone nel muscolo scheletrico suggeriscono che il Testosterone è in grado di produrre effetti rapidi simili (entro 2 minuti) nelle cellule muscolari scheletriche (Estrada 2000; 2003; Hamdi e Mutungi, 2010). Ad oggi, l’azione non genomica del Testosterone nel muscolo scheletrico è stata studiata solo in cellule e fasci di fibre isolate di roditori, con la maggior parte delle prove derivanti da due studi di Estrada et al (Estrada et al., 2000; 2003). Questi due studi hanno isolato miotubi e hanno ottenuto una risposta a 100 nM di Testosterone nel 70% delle cellule. Livelli sovrafisiologici di Testosterone (100 nM) a livello di singola cellula in miotubi primari hanno indotto un aumento transitorio relativamente rapido (<2 min) del calcio intracellulare, spesso accompagnato da oscillazioni e da un aumento transitorio della massa di IP3 fino a triplicare i livelli basali entro 45 s. Sia il Testosterone non legato che quello legato all’albumina hanno avviato questa azione non genomica. Questi risultati sono simili a quelli precedentemente identificati in altre cellule tissutali (miociti cardiaci, cellule T, osteoblasti). Questi risultati hanno fornito supporto per un recettore legato a una proteina G sulla membrana plasmatica e per una via mediata da IP3/calcio, Ras/MEK/ERK. ERK1/2 è aumentato in risposta al Testosterone in modo transitorio e dose-dipendente (50-100nM), mentre gli inibitori della proteina G hanno bloccato il rapido aumento del calcio e dell’IP3 e quindi l’effetto rapido del Testosterone. Pertanto, i meccanismi di trasduzione del segnale delle azioni non genomiche nel muscolo scheletrico sono probabilmente regolati da secondi messaggeri come il calcio intracellulare e l’IP3 (Estrada et al., 2000; Estrada et al., 2003). Queste vie di segnale non genomiche sono state precedentemente identificate nell’azione del Testosterone, dell’Aldosterone (Estrada et al., 2000) e del 17-β-estradiolo (Morley e Whitfield et al., 1992) e sono delineate nella figura seguente.
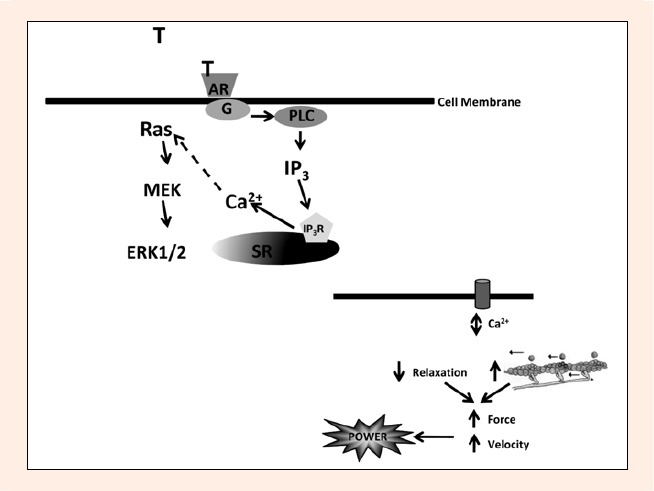
Il Testosterone (T) si lega a un recettore androgeno (AR) situato sulla membrana cellulare, accoppiato a una proteina G sensibile al PTX che attiva la fosfolipasi C (PLC). Questa a sua volta aumenta i livelli di IP3, che vengono liberati e diffusi ai recettori (IP3R) sul reticolo sarcoplasmatico (SR). Questo aumenta a sua volta i livelli di calcio intracellulare attraverso il rilascio di calcio dal SR. È possibile che si verifichi un’attivazione indotta dal calcio della cascata di fosforilazione Ras/ERK, con conseguente trascrizione del DNA, che viene poi espresso sotto forma di proteine.
L’unico studio sull’azione rapida degli steroidi nelle fibre muscolari scheletriche intatte è stato pubblicato dodici anni fa da Hamdi e Mutangi (2010). Questo studio ha utilizzato fibre muscolari scheletriche intatte isolate dei muscoli extensor digitorum longus (prevalentemente a contrazione rapida) e soleus (prevalentemente a contrazione lenta) di topi adulti maschi e femmine. Come gli studi precedenti, questo studio ha utilizzato metodi già pubblicati per indagare gli effetti degli ormoni steroidei sulla forza isometrica massima. Le concentrazioni fisiologiche di Diidrotestosterone (DHT) (630 pg-ml) hanno aumentato significativamente la forza prodotta sia nelle contrazioni twitch che in quelle tetaniche nelle fibre a contrazione rapida. Un aumento significativo del 24% della tensione isometrica massima nelle fibre a contrazione rapida nei topi maschi e un aumento del 30% nei topi femmine, sebbene non statisticamente diversi tra loro, suggerisce che potrebbe essere opportuno indagare sulle potenziali differenze di genere nell’azione non genomica degli steroidi. Il Testosterone, tuttavia, non ha avuto alcun effetto né sulle contrazioni tetaniche né su quelle veloci o lente in fasci di fibre muscolari intatti di roditori (Hamdi e Mutungi, 2010). Parallelamente all’aumento della forza isometrica massima osservato con l’esposizione al DHT, le concentrazioni fisiologiche di quest’ultimo hanno aumentato la fosforilazione di ERK1/2 di 2-3 volte in entrambi i tipi di fibre, aggiungendo un supporto per una via mediata da RAS/MAP/ERK non genomica degli androgeni. Il Testosterone ha aumentato la fosforilazione di ERK1/2 solo nelle fibre a contrazione lenta.
Sebbene i suddetti studi sugli animali forniscano prove di un meccanismo non genomico, la capacità di dedurre effetti sull’uomo dagli studi sui roditori non è chiara e destinata ad errori di valutazione. È quindi importante intraprendere una ricerca sull’uomo per fornire prove di un’azione non genomica degli steroidi nel muscolo scheletrico umano. Sulla base dell’attuale comprensione dell’azione non genomica del Testosterone, si suggerisce che il Testosterone possa essere in grado di produrre un aumento dei livelli di calcio intracellulare e della mobilità del calcio all’interno della cellula muscolare scheletrica umana. Ciò potrebbe aumentare la sensibilità degli elementi contrattili al calcio, il che potrebbe aumentare la velocità di legame della testa di miosina e/o la forza di trazione della testa di miosina, in modo da produrre più forza per contrazione. La combinazione di questi effetti porterebbe probabilmente a una maggiore produzione di potenza da parte del muscolo intero (vedi figura precedente). Sebbene la ricerca attuale fornisca una base sostanziale per questa teoria (Estrada et al., 2000; 2003; Hamdi e Mutungi, 2010), è fondamentale intraprendere ricerche sul muscolo scheletrico umano per fornire prove a sostegno delle sfumature sia molecolari che fisiologiche di questa teoria. Finché non saranno intraprese ulteriori ricerche, le implicazioni per le prestazioni atletiche rimarranno speculative.

E’ interessante notare come, indipendentemente da recettori, canali o vie di secondo messaggero, gli androgeni possono mediare alcune azioni non genomiche attraverso le loro proprietà strutturali. È stato riscontrato che i metaboliti degli androgeni acquisiscono cariche aggiuntive dai residui di solfato e, a loro volta, raggiungono la carica necessaria per penetrare nel complesso lipidico/proteico della membrana cellulare, diminuendo così la flessibilità della membrana e modulando le azioni degli enzimi necessari per l’idrolisi dell’ATP. Ad esempio, Verbist et al (1991) hanno dimostrato un’interazione diretta dei fosfolipidi con carica negativa con le pompe di calcio ATPasi di membrana attraverso il trasferimento di energia per risonanza fluorescente. Queste osservazioni possono avere conseguenze fisiologiche, perché la sintesi locale di steroidi potrebbe consentire una regolazione permanente e indipendente dal calcio dell’attività della Ca2+-ATPasi nelle membrane plasmatiche neuronali. A sostegno di questa ipotesi, è stato dimostrato che gli steroidi idrofobici, tra cui T e DHT, interagiscono con i fosfolipidi di membrana per influenzarne la fluidità [Van Bommel T et al.].

È noto che gli androgeni sono intimamente coinvolti nel sistema riproduttivo, più precisamente nel controllo neuroendocrino dell’ormone di rilascio delle gonadotropine (GnRH). È noto da tempo che gli androgeni inibiscono la secrezione dell’ormone luteinizzante, che è sotto il diretto controllo dell’ipotalamo attraverso la secrezione del GnRH. Sebbene sia noto che gli androgeni influenzano la sensibilità ipofisaria al GnRH, vi sono risultati che suggeriscono fortemente una componente neuronale per la regolazione androgenica della secrezione di LH, lo specifico sito (o siti) neurale di azione degli androgeni rimane in gran parte sconosciuto. Ad esempio, la castrazione non ha alcun effetto sui livelli di GnRH mRNA almeno nelle prime 7 settimane dopo la castrazione. Tuttavia, alcuni studi hanno dimostrato che il Testosterone aumenta i livelli di GnRH mRNA quando viene somministrato a ratti maschi castrati. Inoltre, è stato dimostrato che il Testosterone aumenta i livelli di proteine del GnRH nell’eminenza mediana di ratti castrati e scimmie. Tuttavia, poiché il Testosterone può essere aromatizzato in metaboliti estrogenici e questi risultati non sono stati replicati con l’androgeno non aromatizzabile, il DHT da solo, o in assenza di attivazione dei recettori estrogenici, non è noto se un metabolita estrogenico del Testosterone sia in grado di aumentare l’espressione del GnRH. Allo stesso modo, gli effetti degli androgeni sulla secrezione di GnRH possono essere mediati indirettamente dagli oppioidi, poiché il Naloxone, un antagonista generale dei recettori oppioidi, può bloccare il feedback negativo indotto dagli androgeni in vivo [ne ho parlato già nel dettaglio]. L’aspetto forse più importante è che i neuroni del GnRH non contengono AR. Pertanto, si ritiene che gli androgeni agiscano attraverso una via trans-sinaptica che coinvolge gli interneuroni per influenzare la secrezione di GnRH [Herbison AE et al.].
Testosterone e prestazioni atletiche:
Dagli studi sulla somministrazione di Testosterone nei maschi (3 mg-kg-settimana per 12 settimane) (Griggs et al., 1989) e (600 mg-settimana per 10 settimane), (Bhasin et al., 1996) l’azione tradizionale (trascrizionale) del Testosterone dimostra la capacità di aumentare la sintesi proteica (27%), (Griggs et al., 1989) la massa grassa libera (6 ± 0,6 kg), (Bhasin et al., 1996) la massa muscolare (20%), (Griggs et al., 1989) e (Bhasin et al., 1996), 1989) la massa grassa (6,1 ± 0,6 kg), (Bhasin et al., 1996) la massa muscolare (20%), (Griggs et al., 1989) le dimensioni dei muscoli (tricipiti brachii 501 ± 104 mm2, quadricipiti 1174 ± mm2) (Bhasin et al., 1996) e la forza (bench press 22 ± 2 kg, squat 38 ± 4 kg) (Bhasin et al., 1996). Successivamente, il Testosterone è stato ufficialmente inserito nell’elenco delle sostanze vietate negli eventi sportivi. Mentre la somministrazione cronica di Testosterone può aumentare la forza (Bhasin et al., 1996), è stato dimostrato che la potenza o l’altezza del salto in contromovimento sono correlate positivamente con i livelli naturali acuti di Testosterone (0,62 ± 0,06 ng-ml-1 e 6,49 ± 0,37 ng-ml-1) (r = 0,061, p<0,001) rispettivamente in atleti d’élite di sesso femminile e maschile (Cardinale e Stone, 2006). I livelli naturali di Testosterone basale e l’altezza del salto in contromovimento sono più elevati negli atleti esplosivi, come i velocisti, e più bassi negli atleti di resistenza, come gli sciatori di fondo (Bosco, 1998). Considerati i doppi meccanismi del Testosterone (azione genomica e non genomica), i livelli naturali di Testosterone potrebbero rivelarsi più importanti di quanto si pensasse in precedenza nella capacità di produrre potenza esplosiva acuta, una variabile determinante per le prestazioni negli sprint, nei salti e nei lanci (Bourdin et al., 2010; Hori et al., 2008; Sleivert e Taingahue, 2004; Van Ingen Schenau et al., 1990).
Ipotesi della fatica – come l’azione non genomica del Testosterone può contrastare la fatica -:
Il calcio è un metabolita importante nella contrazione muscolare: sia la concentrazione intorno ai miofilamenti sia la sensibilità dei miofilamenti al calcio sono importanti per la produzione di forza da parte dei singoli ponti trasversali (forza massima attivata dal calcio). La contrazione intensa e ripetitiva del muscolo scheletrico provoca un declino delle prestazioni di picco (cioè l’affaticamento) caratterizzato da una ridotta produzione di forza, da una diminuzione della velocità di accorciamento e da un ritardo nel rilassamento del muscolo dopo la contrazione (Bigland-Ritchie et al., 1979; Edman e Mattiazzi, 1981; Haan et al., 1989; Jones et al., 1979; Milner-Brown e Miller, 1986; Westerblad e Lännergren, 1991; Cheng e Rice, 2010). Una delle principali teorie sull’affaticamento del muscolo scheletrico è rappresentata dai cambiamenti caratteristici nella regolazione e nella sensibilità al calcio che si verificano durante il declino delle prestazioni (Kabbara e Allen, 1999).
Precedenti ricerche hanno dimostrato che durante la contrazione muscolare da affaticamento si verifica un calo del trasporto del calcio, un calo del rilascio di calcio da parte del reticolo sarcoplasmatico (Kabbara e Allen, 1999; Ward et al., 1998; Westerblad e Allen, 1991) e/o una riduzione della sensibilità dell’apparato contrattile al calcio (Godt e Nosek, 1989; Westerblad e Allen, 1993). Entrambi gli scenari porterebbero a un’alterazione dell’accoppiamento eccitazione-contrazione, in modo da generare meno forza per ogni singola eccitazione di membrana.
Un possibile effetto protettivo del Testosterone contro l’affaticamento del muscolo scheletrico è stato suggerito da Bosco et al., 2000, che hanno analizzato l’attività neuromuscolare e il profilo ormonale in seguito a una sessione acuta di esercizio contro-resistenza in atleti di sprint maschi e femmine. La potenza dello squat completo è diminuita del 10% alla fine della sessione solo nei maschi. Il rapporto EMG/potenza calcolato nel test di mezzo squat è diminuito sia nei maschi che nelle femmine, ma ha raggiunto la significatività solo nei maschi (p <0,05). I livelli di Testosterone, Cortisolo e Ormone Luteinizzante circolanti erano significativamente più bassi dopo l’esercizio solo nei maschi, mentre è stata riscontrata una correlazione negativa (r = -0,61) tra la variazione della concentrazione di Testosterone e il rapporto EMG/potenza nella prestazione di mezzo squat in entrambi i gruppi. Bosco et al., 2000 hanno suggerito che livelli adeguati di Testosterone possono compensare o offrire protezione contro l’effetto della fatica nelle fibre muscolari a contrazione rapida, garantendo una migliore efficienza neuromuscolare (Bosco et al., 2000).
Si suggerisce quindi, sulla base dei risultati di precedenti ricerche sull’attività del Testosterone non genomica, che gli aumenti acuti della concentrazione di Testosterone (come quelli che si verificano durante il ciclo mestruale femminile) possano essere in grado di ridurre o compensare gli effetti della fatica nelle fibre a contrazione rapida. A causa di rapidi aumenti non genomici dei livelli di calcio intracellulare e di una maggiore mobilizzazione del calcio dal reticolo sarcoplasmatico, il Testosterone può ridurre o proteggere da una compromissione dell’accoppiamento eccitazione-contrazione durante la contrazione muscolare ripetuta ad alta intensità. Tuttavia, le differenze di genere nei cambiamenti ormonali, in particolare del Testosterone dopo l’esercizio di squat, riportate da Bosco et al. (Bosco et al., 2000) suggeriscono che la ricerca futura dovrebbe indagare se esistono effettivamente risposte specifiche per genere nell’azione non genomica del Testosterone.
Importanza specifica del Testosterone per le atlete:
I livelli di Testosterone sono stati spesso difficili da misurare accuratamente nelle donne a causa della sfida combinata dei bassi livelli naturali di Testosterone circolante e della bassa sensibilità e precisione dei test. Studi precedenti hanno tuttavia dimostrato che il Testosterone circolante fluttua durante il ciclo mestruale (Judd e Yen, 1973; Sinha-Hikim et al., 1998). I livelli sierici di Testosterone totale e libero nelle fasi luteale e follicolare non sono significativamente diversi l’uno dall’altro, ma un aumento pre-ovulatorio del 30-45% circa sia del Testosterone totale che di quello libero, circa tre giorni prima del picco dell’Ormone Luteinizzante, è stato registrato in due studi distinti che hanno analizzato i livelli di Testosterone durante l’intero ciclo mestruale (Judd e Yen, 1973; Sinha-Hikim et al., 1998). Tuttavia, anche se in questi due studi è stato evidenziato un chiaro picco di Testosterone totale e libero, nessuno studio ha ancora analizzato in modo specifico i cambiamenti nella forza muscolare, nella potenza esplosiva o nella fatica al momento del picco dei livelli di Testosterone, che potrebbe rivelarsi una misura importante, dato che studi precedenti hanno dimostrato forti correlazioni positive con la concentrazione naturale acuta di Testosterone e le prestazioni acute di potenza, come l’altezza del salto in contromovimento.
Influenza dei contraccettivi orali:
I contraccettivi ormonali combinati impediscono l’ovulazione come meccanismo d’azione primario (Rivera e Yacobson et al., 1999). L’eliminazione dell’ovulazione elimina anche il picco naturale di Testosterone che precede l’aumento dell’Ormone Luteinizzante. Gli estrogeni orali ingeriti nelle comuni varietà della pillola contraccettiva orale possono avere conseguenze significative sui livelli di Testosterone libero circolante (Edwards e O’neal, 2009; Raj et al., 1983; Rickenlund et al., 2004; Thorneycroft et al., 1999; Van Der Vange et al., 1990; Wiegratz et al., 1995; 2003a). Gli estrogeni orali possono aumentare i livelli di globulina legante gli ormoni sessuali (Campagnoli et al., 1993; Thorneycroft et al., 1999; Wiegratz et al., 1995; 2003a), che si lega al Testosterone rendendolo biologicamente indisponibile, riducendo così il rapporto tra Testosterone libero e Testosterone totale circolante.
Anche i progestinici possono influenzare i livelli di Testosterone circolante nelle femmine (Gordon et al, 1970) Il Medrossiprogesterone, una versione sintetica del Progesterone umano sintetizzato naturalmente (spesso usato nei contraccettivi orali), diminuisce il tasso di produzione del Testosterone, probabilmente a causa dell’inibizione della secrezione ipofisaria dell’Ormone Luteinizzante e può aumentare il tasso di rimozione del Testosterone dalla circolazione (Gordon et al., 1970) (Palatsi et al., 1984; Wiegratz et al., 2003b). Oltre all’eliminazione del picco di Testosterone durante il ciclo e alla riduzione della concentrazione di Testosterone totale, i contraccettivi orali possono, con buona probabilità, influenzare le azioni genomiche e non genomiche del Testosterone, riducendo la fisiologia ormonale ottimale per le prestazioni atletiche femminili d’élite.
Il potenziale impatto del ciclo mestruale sulle prestazioni anaerobiche ha ricevuto meno attenzione rispetto all’impatto sulle variabili aerobiche. Non c’è consenso sul fatto che le fluttuazioni degli ormoni sessuali abbiano un’influenza sulla prestazione anaerobica, con alcuni studi che concludono che la prestazione non è influenzata dalla fase del ciclo mestruale (Doolittle e Engebretsen, 1972; Giacomoni et al., 2000; Lebrun, 1993; 1994; Lebrun et al., 1995) e altri che riportano differenze nelle variabili della prestazione anaerobica con la fase del ciclo mestruale (Davies et al., 1991; Masterson, 1999; Wearing et al., 1972). La forza della ricerca sulle prestazioni anaerobiche e gli ormoni sessuali è compromessa dalla mancanza di controlli sperimentali, tra cui la variazione nella determinazione della fase del ciclo (dosaggio ormonale rispetto alla temperatura corporea), criteri di selezione dei soggetti poco chiari e misure diverse delle prestazioni (sprint, salto, nuoto e sforzi acuti o ripetuti). Non sono ancora state pubblicate ricerche sull’effetto degli ormoni del ciclo mestruale sulle variabili anaerobiche/di potenza in popolazioni di atlete d’élite.
Studi in cui le utilizzatrici di contraccettivi orali sono state confrontate con femmine eumenorroiche hanno riportato una tendenza alla riduzione della forza durante il ciclo contraccettivo orale rispetto al ciclo naturale. Inoltre, nelle donne che utilizzano un contraccettivo orale è stata dimostrata una completa riduzione della naturale fluttuazione del Testosterone e della forza, spesso riscontrata con un ciclo naturale (Phillips et al., 1996; Sarwar et al., 1996). Ci sono poche ricerche che hanno esaminato specificamente l’effetto del ciclo mestruale o dei contraccettivi orali sulla potenza esplosiva nelle atlete. Tuttavia, uno studio ha esaminato le variabili delle prestazioni negli sport di squadra durante un ciclo di contraccettivi orali (Rechichi e Dawson, 2009), con l’unica differenza significativa riscontrata nell’altezza del salto in alto nella fase di sospensione tardiva (fine delle pillole di zucchero), dove la riduzione dell’altezza del salto in alto coincideva con l’aumento dei livelli sierici di estrogeni (Rechichi e Dawson, 2009). È possibile che si siano verificati cambiamenti nelle variabili di prestazione nelle donne con un ciclo mestruale naturale a causa delle fluttuazioni del testosterone circolante; tuttavia, sebbene sia noto che il Testosterone raggiunga un picco durante il ciclo mestruale, nessuno studio ha esaminato specificamente le prestazioni atletiche in questo periodo. Inoltre, non sono disponibili studi che abbiano condotto test di performance ogni giorno del ciclo mestruale per garantire che tutte le fluttuazioni ormonali siano esaminate. Nonostante ciò, data la correlazione del Testosterone con la potenza esplosiva e l’evidenza di un’azione non genomica del Testosterone nel muscolo scheletrico, le singole atlete i cui eventi richiedono forza o potenza possono trarre beneficio dall’uso di metodi contraccettivi non ormonali.
Sindrome dell’ovaio policistico e altri disturbi mestruali: Un vantaggio fisiologico?
In precedenza, l’oligomenorrea nelle donne che praticano attività fisica era considerata un sintomo di disturbi mestruali, secondari a perturbazioni metaboliche dovute a un deficit energetico estremo (spesso osservato nelle atlete di resistenza e associato alla magrezza) (Rosetta et al., 1998; Sanborn et al., 1982; Torstveit e Sundgot-Borgen, 2005). Tuttavia, dato che l’oligomenorrea può essere associata a iperandrogenismo [in particolare a un aumento dei livelli di Testosterone al di fuori (o all’interno dell’intervallo superiore) dei limiti fisiologici per le donne normalmente mestruate], molte atlete che soffrono di oligomenorrea potrebbero non presentare, come ci si aspetterebbe, sintomi di deficit energetico. Pertanto, è opinione di alcuni che la percezione che le atlete che presentano l’oligomenorrea siano molto probabilmente affette da un deficit energetico estremo, possa essere infondata.
Sebbene le ricerche sulla prevalenza dell’iperandrogenismo e della sindrome dell’ovaio policistico nelle popolazioni atletiche siano scarse, i dati disponibili suggeriscono che la diagnosi più comune di disturbo mestruale nelle atlete olimpiche (Hagmar e Berglund et al., 2009) e nelle donne che praticano sport in cui la massa muscolare è vantaggiosa o non dannosa per le prestazioni (Lebrun, 1994; Masterson, 1999) è la sindrome dell’ovaio policistico. È possibile che le donne con sindrome dell’ovaio policistico o iperandrogenismo siano intrinsecamente attratte e abbiano successo nelle attività sportive. Ciò suggerisce che l’oligomenorrea sia anche un sintomo della sindrome dell’ovaio policistico o della sindrome metabolica nelle donne atletiche, piuttosto che un tratto indotto dall’esercizio fisico in sé. Si suggerisce quindi che la sindrome dell’ovaio policistico possa essere un vantaggio competitivo a causa del duplice meccanismo proposto per il Testosterone (effetti a lungo termine sulla forza e sulla dimensione muscolare e un rapido effetto acuto sull’efficienza contrattile).
Sono necessarie ulteriori ricerche per confermare se le fluttuazioni acute di Testosterone osservate durante il ciclo mestruale naturale o i livelli più elevati osservati in condizioni come la sindrome dell’ovaio policistico siano abbastanza significativi da determinare una maggiore efficienza del muscolo scheletrico nel produrre forza/potenza. La sindrome dell’ovaio policistico o l’iperandrogenismo potrebbero portare a un vantaggio fisiologico per le atlete, in particolare per quelle che gareggiano in eventi atletici che richiedono movimenti rapidi e/o di forza.
Azioni non genomiche degli estrogeni:
Sebbene questo articolo si concentri sul ruolo specifico dell’azione non genomica del Testosterone nel muscolo scheletrico e sui suoi effetti all’interno del compartimento contrattile della muscolatura scheletrica, è nell’ambito di questo articolo discutere brevemente i potenziali effetti di un’azione sussidiaria non genomica di un altro importante ormone sessuale che fluttua durante il ciclo mestruale femminile, gli estrogeni.
Sebbene l’invecchiamento, il deperimento muscolare e le ricerche sulla patologia sostengano un ruolo degli estrogeni nel muscolo scheletrico, in particolare nelle donne, è probabile che la prevenzione della perdita di forza attraverso la terapia ormonale sostitutiva con estrogeni sia un meccanismo genomico o trascrizionale piuttosto che un rapido meccanismo non trascrizionale. Poiché questo articolo si concentra anche sugli effetti del Testosterone sull’apparato contrattile, sembra pertinente discutere i risultati degli studi sui roditori che indicano che gli estrogeni possono influenzare la capacità di generare forza del muscolo scheletrico non mantenendo le dimensioni delle singole fibre, ma mantenendo l’integrità e la capacità delle singole fibre di generare forza. Questa idea è sostenuta da Wattanapermpool et al. (1999) che hanno misurato l’area della sezione trasversale (CSA) e la tensione isometrica di picco delle fibre muscolari isolate del soleo di ratto 10 e 14 settimane dopo l’ovariectomia. Mentre la CSA non era ridotta in seguito all’ovariectomia, la CSA era significativamente aumentata rispetto ai controlli non ovariectomizzati a 14 settimane, la tensione di picco era significativamente più bassa nei ratti ovariectomizzati rispetto ai controlli sham-operati sia a 10 settimane (~19%) che a 14 settimane (~20%) dall’ovariectomia. Questi risultati dimostrano che le fibre delle ratte ovariectomizzate non erano più deboli a causa delle loro dimensioni più piccole, ma piuttosto c’era un deficit nell’apparato contrattile, probabilmente dovuto all’assenza di estrogeni, che si traduceva in una ridotta capacità di produrre forza.
Analogamente al Testosterone, l’azione non genomica degli estrogeni è stata regolarmente identificata in tessuti diversi dal muscolo scheletrico (Morley et al., 1992; Rubio-Gayosso e Sierra-Ramirez et al., 2000; Watson et al., 2008; Younglai et al., 2005). L’effetto non genomico più regolarmente riportato dell’esposizione agli androgeni è un rapido (entro pochi secondi) aumento della concentrazione intracellulare di calcio (Ceballos et al., 1999; Morley et al., 1992; Vicencio et al., 2006; Watson et al., 2008). Gli estrogeni non sono esenti da questo effetto ed è stato originariamente dimostrato in cellule di granulosa di pollo sottoposte a 17-β estradiolo, che vi era un aumento immediato (meno di 5 s) di 4-8 volte della concentrazione di calcio in tutte le 76 cellule esposte. È stato inoltre riscontrato che i recettori per gli estrogeni interagiscono con una proteina G sulla membrana cellulare degli osteoblasti, determinando un rapido aumento della concentrazione intracellulare di Ca2+ dovuto in modo distinto a un’aumentata mobilizzazione di Ca2+ dal reticolo endoplasmatico e alla formazione di IP3, dimostrando una via meccanicistica simile a quella mostrata durante la mobilizzazione di Ca2+ intracellulare dal reticolo sarcoplasmatico nelle cellule muscolari scheletriche a seguito di una rapida azione del Testosterone. Al momento nessuna ricerca ha studiato specificamente un’azione non genomica degli estrogeni nei muscoli scheletrici, né a livello molecolare né a livello del muscolo intero.
Anche in questo caso, come il Testosterone, gli estrogeni hanno dimostrato possibili effetti positivi nel muscolo scheletrico, in particolare producendo effetti positivi all’interno del macchinario contrattile e anche se questi effetti sono stati osservati in presenza (terapia ormonale sostitutiva) o in assenza (invecchiamento/ovariectomia) di esposizione o non esposizione costante agli estrogeni (17β-estradiolo), le azioni non genomiche degli estrogeni in tessuti diversi dal muscolo scheletrico sono simili a quelle dimostrate dal Testosterone nelle cellule muscolari scheletriche. Pertanto, è necessaria una ricerca per indagare sulla presenza di un’azione non genomica degli estrogeni nei muscoli scheletrici e determinare la loro capacità di modulare i processi di generazione della forza. È quindi probabile che sia il testosterone che gli estrogeni siano in grado di potenziare i processi di legame con la miosina e l’actina a causa di modulazioni nella mobilitazione del calcio, con conseguente aumento della forza e/o della velocità di contrazione durante le variazioni acute delle loro concentrazioni.
Conclusioni:
Attualmente, le prove dell’azione non genomica del Testosterone nel muscolo scheletrico umano sono scarse e la modalità d’azione specifica, così come l’importanza pratica del Testosterone nella competizione atletica, devono ancora essere identificate. Alcune prove indicano l’attivazione diretta non genomica di eventi mediati dal calcio nelle cellule muscolari scheletriche, che possono modulare risposte fisiologiche significative come la modulazione acuta della forza nelle singole fibre e la prevenzione o la protezione acuta dalla fatica mediata dal calcio. Queste risposte sono probabilmente complesse e mediate dall’interazione tra il Testosterone e i secondi messaggeri IP3 e calcio, che in ultima analisi possono risultare in modulazioni simultanee non genomiche e genomiche degli eventi del muscolo scheletrico. Tuttavia, sono necessarie ulteriori ricerche per chiarire il potenziale recettore di membrana coinvolto, nonché la via dei secondi messaggeri, l’azione risultante a livello di singola cellula e il trasferimento del Testosterone al significato del muscolo intero. A causa delle fluttuazioni del testosterone durante il ciclo mestruale femminile, la ricerca dovrebbe anche mirare a identificare se ci sono reali differenze di genere nella risposta o nel meccanismo di azione non genomica del Testosterone nel muscolo scheletrico. Potrebbe esserci un vantaggio fisiologico per le prestazioni atletiche femminili durante particolari fasi del ciclo mestruale. Anche la sindrome dell’ovaio policistico e simili disturbi di iperandrogenismo possono rappresentare un vantaggio fisiologico per specifiche atlete. Le ricerche condotte finora suggeriscono che la principale funzione fisiologica dell’azione del Testosterone non genomico nel muscolo scheletrico è l’aumento della produzione di forza, in particolare nelle fibre a contrazione rapida; tuttavia, le azioni sono probabilmente molteplici e sono necessarie ulteriori ricerche per fornire prove di un effetto sulle prestazioni dell’intero corpo nell’uomo.
In conclusione, l’azione non genomica del Testosterone spiega il perchè dell’efficacia di alcune pratiche dopanti pre-allenamento contro-resistenza le quali vedono l’assunzione di Testosterone in sospensione o altri AAS orali per migliorare in acuto la prestazione di forza/potenza.
Gabriel Bellizzi [CEO BioGenTech]
Letteratura scientifica di riferimento:
