
Continua la disamina dei principali PEDs utilizzati e del confine che delimita l’uso dall’abuso. In questo terzo articolo della serie tratteremo due ormoni, o meglio un precursore poco attivo [T4] ed il suo derivato molto attivo [T3], che non rientrano pienamente nella categoria PEDs, ma che, volenti o nolenti, si sono diffusi da decenni nel mondo del BodyBuilding, in ambo i sessi. Inutile dire che l’abuso con questa classe di farmaci è alquanto facile e spesso praticato.
Tanto per ribadirlo, questo articolo non rappresenta ne un incitamento all’uso di farmaci fuori dalla prescrizione medica ne tantomeno un indicazione medica. Si tratta di divulgazione scientifica.
Introduzione agli ormoni tiroidei [T4 e T3]:
Gli ormoni tiroidei sono ormoni secreti dalla tiroide. La tiroide è una ghiandola endocrina situata nella parte anteriore del collo, direttamente sotto la laringe (pomo d’Adamo), ed ha un peso di circa 20g. I due principali ormoni tiroidei che secerne sono la Triiodotironina (T3) e la Tiroxina (T4). Quest’ultima ha soprattutto una attività da pro-ormone, poiché la maggior parte dei suoi effetti dipende dalla conversione in T3. Questa conversione da T4 a T3, chiamata anche deiodinazione dell’anello esterno, avviene principalmente al di fuori della tiroide, nei tessuti periferici. Complessivamente, ciò porta a una produzione giornaliera di circa 88mcg (113 nmol) di T4 e 28mcg (43 nmol) di T3 [2]. Circa un quinto della T3 deriva dalla tiroide, mentre gli altri quattro quinti sono prodotti dalla conversione extratiroidea di T4 in T3 [3].

Come nel caso degli steroidi anabolizzanti, gli ormoni tiroidei sono trasportati nel flusso sanguigno da proteine trasportatrici. La maggior parte è legata alla globulina legante la tiroxina (TBG), mentre la parte restante è legata alla transtiretina, all’albumina e ad alcune lipoproteine. Nel complesso, esse legano oltre il 99% degli ormoni tiroidei in circolazione. Si ritiene che la frazione non legata sia disponibile per i tessuti per l’assorbimento e sia responsabile dei suoi effetti [4]. Sebbene vi siano alcune riserve sulle prove a sostegno di questa tesi, non intendo addentrarmi in una discussione sull’ipotesi dell’ormone libero (se non ricordare che nella sua forma più rigorosa è sbagliata, ma le misurazioni dell’ormone tiroideo libero sono comunque utili).
Una volta che raggiunge i tessuti periferici e attraversa la membrana plasmatica di una cellula, esso esplica la sua attività. Nel caso del T4, deve prima essere convertito in T3, come già detto, in quanto il T4 può essere considerato un pro-ormone. Questa conversione avviene all’interno della cellula, o vicino alla membrana plasmatica (dopo di che si equilibra rapidamente con il plasma sanguigno), o vicino al nucleo della cellula, il sito d’azione [5].Il T3, invece, può continuare direttamente il suo viaggio entrando nel nucleo della cellula. Il nucleo cellulare è l’organello della cellula dove avviene la trascrizione dei geni. Proprio come gli steroidi anabolizzanti, gli ormoni tiroidei esercitano i loro effetti principalmente attraverso la modulazione della trascrizione genica. Lo fanno legandosi ai recettori degli ormoni tiroidei che si trovano principalmente all’interno del nucleo cellulare, legati al DNA.
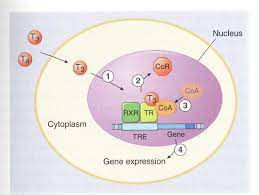
Gli ormoni tiroidei agiscono su una vasta gamma di tessuti e hanno un’infinità di effetti, ma in questo articolo mi concentrerò sull’effetto che essi hanno sul metabolismo energetico e sul turnover delle proteine (muscolo scheletrico). Con tutta probabilità sono i due aspetti che più interessano le persone che leggono questo articolo per quanto riguarda la sua efficacia.
Effetto sul metabolismo energetico (Parte 1):
Quando è presente una quantità insufficiente di ormoni tiroidei, si parla di ipotiroidismo. Una delle caratteristiche dell’ipotiroidismo è l’aumento di peso. Al contrario, quando la quantità di ormoni tiroidei è eccessiva, si parla di ipertiroidismo. Una delle sue caratteristiche è la perdita di peso. Queste variazioni di peso sono probabilmente il risultato di cambiamenti nel tasso metabolico basale. È noto che gli ormoni tiroidei aumentano il dispendio energetico.
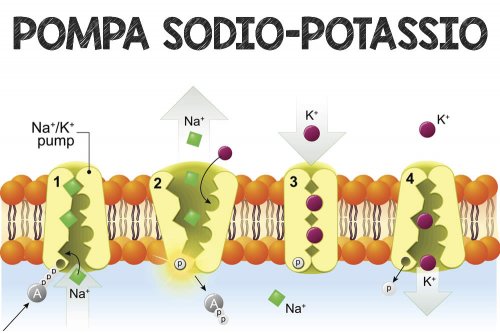
Sono stati proposti alcuni meccanismi che spiegano come gli ormoni tiroidei riescano a ottenere questo risultato. In questo articolo tratterò i tre più interessanti (o forse semplicemente quelli che si incontrano di più nella letteratura scientifica). I primi due meccanismi si basano sull’energia necessaria per mantenere i gradienti ionici all’interno della cellula. Ad esempio, le cellule mantengono una bassa concentrazione intracellulare di sodio e un’alta concentrazione intracellulare di potassio rispetto all’esterno della cellula. Il mantenimento di questa condizione è assicurato da pompe incorporate nella membrana plasmatica, che richiedono energia per funzionare. Esse pompano ioni sodio fuori dalla cellula e ioni potassio dentro la cellula. Queste pompe sono note come Na+/K+-ATPasi, o semplicemente pompe sodio-potassio. L’energia necessaria al funzionamento di queste pompe deriva dalla molecola portatrice di energia adenosina trifosfato (ATP). L’ATP è utilizzato da molti processi cellulari per alimentare il proprio fabbisogno energetico e l’energia contenuta in queste molecole deriva dai macronutrienti che mangiamo: carboidrati, acidi grassi e proteine (aminoacidi). Ed ora d’obbligo descrivere il modo principale in cui le cellule producono queste molecole di ATP attraverso un processo chiamato fosforilazione ossidativa.
Uno dei modi in cui gli ormoni tiroidei potrebbero aumentare il dispendio energetico è simile al modo in cui il famoso DNP ottiene questo risultato: “sabotando” la fosforilazione ossidativa.
Fosforilazione ossidativa: ottenere energia dal passaggio degli elettroni.
È inutile ribadire che è sempre un piacere per me trattare di biochimica in un articolo. Ritengo che questi principi di base tolgano un po’ di magia agli effetti dei farmaci, e forniscano quindi un quadro più chiaro di come funzionano le cose. Con un po’ di fortuna potrei anche, forse, interessare qualcuno di voi che sta leggendo questo articolo ad approfondire l’argomento. La biochimica e la biologia cellulare sono campi di studio estremamente interessanti.
Le cellule del vostro corpo svolgono continuamente ogni sorta di funzione per, essenzialmente, mantenervi in vita. Molti di questi processi consumano energia. Questa energia deriva, in ultima analisi, dagli alimenti che mangiamo. Carboidrati, grassi e proteine, persino l’alcol, hanno tutti energia immagazzinata nei loro legami chimici. È compito dell’organismo estrarre questa energia e trasformarla in qualcosa di utile. Come il motore della vostra auto non funziona con il petrolio grezzo, questi processi cellulari non funzionano direttamente con i macronutrienti. Al contrario, la maggior parte di questi processi richiede energia da una molecola chiamata adenosina trifosfato (ATP), proprio come il motore di un’automobile richiede specificamente la benzina.
Vediamo come funziona per una molecola di glucosio, un carboidrato. Quando una molecola di glucosio viene utilizzata da una cellula per produrre ATP, subisce prima un processo chiamato glicolisi. La glicolisi è un processo composto da varie fasi enzimatiche che scindono la molecola di glucosio in 2 molecole di piruvato e producono 2 molecole di ATP (oltre ad altre molecole). In poche parole:
glucosio -> 2 piruvato + 2 ATP
Tuttavia, un processo chiamato fosforilazione ossidativa estrarrà molta più energia, cioè molecole di ATP, dalle 2 molecole di piruvato risultanti.
La fosforilazione ossidativa è un processo che avviene nei mitocondri. Quindi è qui che il piruvato è diretto. I mitocondri sono organelli della cellula che si occupano principalmente della produzione di energia. Sono piccole fabbriche di energia di dimensioni microscopiche. Sono costituiti da una membrana esterna e da una membrana interna. Lo spazio tra la membrana esterna e quella interna è chiamato spazio intermembrana. Lo spazio incapsulato dalla membrana interna è chiamato matrice mitocondriale. La membrana interna è ripiegata in modo caratteristico. Queste pieghe sono chiamate cristae. L’aspetto è questo:

Quando il piruvato si trova all’interno della matrice mitocondriale, viene convertito in acetil-CoA e successivamente subisce una serie di reazioni che vengono chiamate collettivamente ciclo dell’acido citrico o ciclo di Krebs. Durante questo processo, tutta l’energia viene estratta da quella che in origine era una molecola di piruvato. Viene ossidata. Tuttavia, l’energia non si è ancora trasformata in ATP. Prima viene trasferita ai vettori energetici NAD e FAD (e al GTP, ma non ne parlerò). I vettori energetici NAD e FAD parteciperanno al processo chiamato fosforilazione ossidativa che segue il ciclo dell’acido citrico.
L’energia viene immagazzinata in coppie di elettroni che vengono donati a NAD e FAD. Questo processo riduce queste molecole, come viene chiamato, producendo rispettivamente NADH e FADH2. Successivamente, NADH e FADH2 cedono la coppia di elettroni a grandi complessi proteici incorporati nella membrana interna. Questa è la prima fase della fosforilazione ossidativa. Quando queste coppie di elettroni vengono cedute a tali complessi proteici, parte dell’energia in essi immagazzinata viene utilizzata per pompare un protone (H+) fuori dalla matrice mitocondriale nello spazio intermembrana. Si tratta di un aspetto estremamente cruciale, di cui si capirà presto il motivo.
Successivamente, le coppie di elettroni vengono trasferite un paio di volte da un complesso all’altro, staccando ogni volta un po’ dell’energia in esse contenuta e utilizzandola per pompare fuori un protone. A ogni passaggio, gli elettroni raggiungono uno stato energetico inferiore. (Non vengono trasferiti direttamente da un complesso all’altro, ci sono alcune proteine/molecole intermedie che li trasportano tra questi complessi proteici che pompano protoni). E ogni volta una parte dell’energia sottratta viene sfruttata per pompare fuori un protone. Se si utilizza una ruota idraulica, l’aspetto è simile a questo:
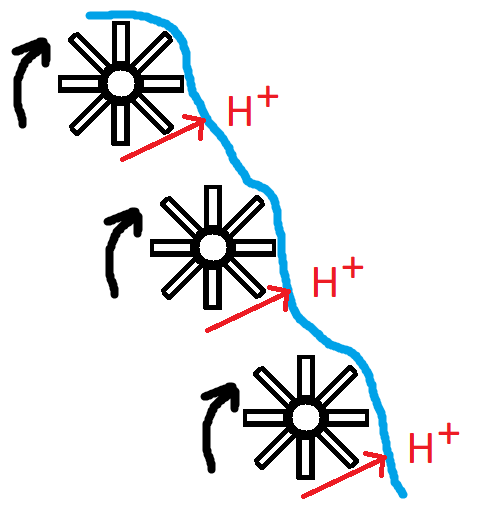
La destinazione finale degli elettroni è quella di combinarsi con l’idrogeno e l’ossigeno per formare H2O, ovvero l’acqua. Il processo di fosforilazione ossidativa ha stabilito un gradiente elettrochimico di protoni. La concentrazione di protoni nella matrice mitocondriale sarà inferiore rispetto allo spazio intermembrana. Questo gradiente contiene energia potenziale. Proprio come una ruota idraulica ruota con l’acqua che si muove in discesa, un macchinario molecolare chiamato ATP sintasi inizia a ruotare con i protoni che si muovono lungo il loro gradiente elettrochimico dallo spazio intermembrana alla matrice mitocondriale. Questa energia viene poi sfruttata per generare ATP combinando l’ADP con un gruppo fosfato inorganico. E voilà, l’intero processo di passaggio degli elettroni, di sottrazione di energia per pompare fuori i protoni e di successivo utilizzo del gradiente protonico stabilito per sintetizzare ATP, è chiamato fosforilazione ossidativa.
Per ricapitolare ciò che è stato trattato, e che non è poco:
- Il glucosio viene scisso in due molecole di piruvato dalla glicolisi.
- il piruvato viene trasportato nella matrice mitocondriale per essere convertito in acetil-CoA
- L’acetil-CoA viene ossidato, trasferendo la sua energia nei vettori energetici NAD e FAD nelle loro forme ridotte NADH e FADH2, accettando una coppia di elettroni.
- Queste molecole di NADH e FADH2 donano le loro coppie di elettroni a un grande complesso proteico incorporato nella membrana interna, che poi viene trasferito in continuazione fino a combinarsi con idrogeno e ossigeno per formare acqua. Con questi trasferimenti, parte dell’energia viene sfruttata per pompare protoni (H+) fuori dalla matrice mitocondriale. Si stabilisce così un gradiente elettrochimico: bassa concentrazione di protoni all’interno della matrice mitocondriale, alta concentrazione di protoni all’esterno della matrice mitocondriale.
- Il flusso di protoni lungo il gradiente di concentrazione fornisce energia all’ATP sintasi per svolgere il suo lavoro e generare ATP.
Effetto sul metabolismo energetico (Parte 2):
Dopo questa dovuta parentesi, torniamo alle pompe sodio-potassio. Alcune prove suggeriscono che gli ormoni tiroidei aumentano la permeabilità della membrana plasmatica agli ioni sodio e potassio [6]. Ciò significa che una quantità maggiore di questi ioni fuoriesce lungo il gradiente di concentrazione. Pertanto, gli ioni potassio fuoriescono dalla cellula e gli ioni sodio vi entrano. Di conseguenza, le pompe sodio-potassio devono pompare maggiormente per mantenere le concentrazioni intracellulari desiderate di questi ioni e questo costa energia. Alcuni studi suggeriscono addirittura che tutti i tessuti dei mammiferi mostrano un aumento dell’attività della pompa sodio-potassio in risposta alla T3 [7].
Qualcosa di simile è stato suggerito per quanto riguarda gli ioni calcio nelle cellule muscolari [8]. Le cellule muscolari sono cellule piuttosto speciali sotto molti aspetti. Uno di questi è che contengono un organello chiamato reticolo sarcoplasmatico. Si tratta di una forma specializzata del reticolo endoplasmatico presente nelle cellule normali. Una delle caratteristiche che lo rendono speciale è che funziona come sito di stoccaggio degli ioni calcio. Questi ioni di calcio svolgono un ruolo fondamentale nella contrazione muscolare, poiché lo scarico di questi ioni di calcio dal reticolo sarcoplasmatico al resto della cellula porta alla contrazione muscolare. Quando la contrazione deve cessare, questi ioni vengono nuovamente pompati nel reticolo sarcoplasmatico. Anche questo processo, ovviamente, consuma energia. E qui viene il bello: si è visto che gli ormoni tiroidei regolano l’espressione di queste pompe del calcio in modelli animali. Inoltre, aumentano l’attività di un certo tipo di recettore nel tessuto muscolare che stimola lo scarico di questi ioni nel citosol [9]. Questo è un altro elemento che indica un potenziale aumento del dispendio energetico come risultato del mantenimento dell’accumulo di ioni calcio nel reticolo endoplasmatico.
Infine, ci sono buone prove che indicano che “sabota” la fosforilazione ossidativa. Come detto sopra, ma questa volta in breve, la fosforilazione ossidativa avviene in un organello cellulare chiamato mitocondrio. I macronutrienti che mangiamo vengono ulteriormente scomposti in componenti più piccoli e in questo processo viene rilasciata energia sotto forma di coppie di elettroni. Un complesso gioco molecolare nei mitocondri tra varie molecole e complessi proteici estrae l’energia da queste coppie di elettroni, utilizzandola essenzialmente per pompare protoni (H+). Questi protoni vengono pompati all’esterno del nucleo dei mitocondri, chiamato matrice mitocondriale, e nello spazio intermembrana – lo spazio tra la membrana mitocondriale interna e quella esterna (i mitocondri hanno due membrane, una che avvolge l’altra). Questo crea un gradiente protonico, con un’alta concentrazione di protoni nello spazio intermembrana e una concentrazione relativamente bassa nella matrice mitocondriale. Proprio come l’acqua che scorre dall’alto verso il basso, da cui possiamo estrarre energia con una turbina ad acqua, le cellule possono estrarre energia da questi protoni che scendono lungo il loro gradiente di concentrazione guidando questo flusso attraverso un fantastico macchinario proteico chiamato ATP sintasi. È questo che alimenta la sintesi di ATP.
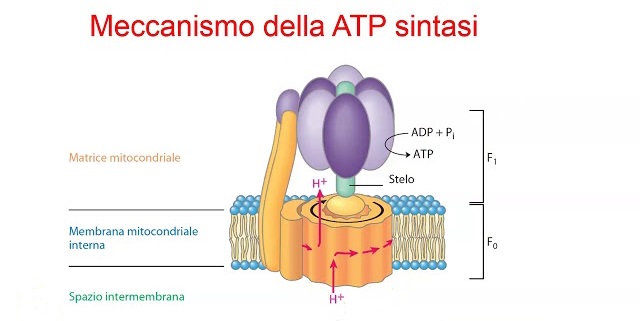
Ok, torniamo al modo in cui gli ormoni tiroidei influiscono su questo aspetto: aumentano l’espressione delle proteine di disaccoppiamento [10, 11]. Si tratta di proteine incorporate nella membrana interna dei mitocondri che lasciano fuoriuscire i protoni lungo il loro gradiente di concentrazione. I protoni passano quindi dallo spazio intermembrana alla matrice mitocondriale, senza passare per l’ATP sintasi. In questo modo, l’energia viene rilasciata come calore anziché essere destinata alla produzione di ATP.
Gli ormoni tiroidei influenzano il turnover delle proteine:
Sembrerà strano, ma non è così raro sentire qualcuno che dice di assumere T3 in Bulk nel tentativo aumentare il turnover proteico. Ma è una buona idea? No. Mentre il turnover proteico aumenta, si verifica un contemporaneo aumento sia della sintesi proteica sia della degradazione proteica, quest’ultima supera il tasso di sintesi. Di conseguenza, si verifica una degradazione netta delle proteine.
In uno studio in cui i soggetti hanno ricevuto 150mcg di T3 al giorno per 7 giorni, la degradazione proteica è aumentata notevolmente [12]. L’escrezione di azoto (un indicatore della degradazione delle proteine) è aumentata del 45% e l’ossidazione della leucina del 74%. È stato riscontrato anche un piccolo aumento della sintesi proteica corporea, ma l’entità era inferiore all’aumento della degradazione proteica. Un altro studio, nel quale è stata usata una dose di 100mcg di T3 al giorno per 2 settimane, ha ottenuto risultati simili [13]. La sintesi proteica corporea a digiuno è aumentata del 9%, anche se in modo non statisticamente significativo, mentre la degradazione proteica e l’ossidazione della leucina hanno mostrato un aumento statisticamente significativo, rispettivamente del 12 e del 24%.
L’aspetto forse più interessante è che i ricercatori hanno anche prelevato biopsie muscolari dal muscolo gastrocnemio. Hanno misurato una serie di elementi, tra cui l’area della sezione trasversale (CSA) delle fibre muscolari. I risultati sono stati i seguenti:

Si tratta di una situazione piuttosto drastica per sole 2 settimane. (Si noti anche il cambiamento del tipo di fibra indotto da uno stato di ipertiroidismo).
In un altro studio, sei partecipanti hanno ricevuto 2mcg/kg di peso corporeo di T4 al giorno per 6 settimane, insieme a 1mcg/kg di peso corporeo di T3 al giorno per le ultime 2 settimane [14]. Questo (le prime 4 settimane) è un po’ più alto di un dosaggio completo di ormoni tiroidei. In effetti, il TSH è stato soppresso da 1,8 a 0,3 mIU/L e sia il T4 che il T3 sono aumentati in modo significativo. La successiva aggiunta di T3 ha reso i livelli di TSH non rilevabili e ha aumentato ulteriormente i livelli di T3. In questo studio non è stata misurata la cinetica delle proteine muscolari. È stata misurata la sintesi e la degradazione delle proteine nell’intero corpo nello stato di post-assorbimento. L’integrazione di ormoni tiroidei ha portato a un aumento di entrambi, ma con un aumento sostanziale della degradazione. Sarebbe ragionevole ipotizzare che questo rifletta anche ciò che accade nel tessuto muscolare.
Infine, vale la pena sottolineare un altro studio di lunga durata, con un dosaggio relativamente basso rispetto agli altri studi. Lovejoy et al. hanno somministrato T3 per 2 mesi a un piccolo gruppo di uomini [15]. Il dosaggio è iniziato con 75 mcg di T3 al giorno, ma è stato ridotto a 50 o 62,5 mcg al giorno quando i livelli di T3 nel siero superavano i 4,6 nmol/L. Cosa che, in effetti, si è verificata per 5 dei 7 uomini partecipanti. Il bilancio dell’azoto è risultato significativamente ridotto rispetto al basale nella seconda e terza settimana, ma in seguito tendeva a tornare verso lo zero. Questo fa pensare a un meccanismo di risparmio proteico che entra in funzione dopo le prime settimane. Inoltre, hanno riscontrato una diminuzione significativa della massa magra (-1,5 kg) e della massa grassa (-2,7 kg) dopo 6 settimane. Alla 9a settimana, la massa magra non è diminuita ulteriormente (-0,1 kg rispetto alla 6a settimana), mentre la massa grassa è sembrata continuare a diminuire (-0,6 kg), anche se non si tratta di una differenza statisticamente significativa rispetto alla 6a settimana. Non sono state riscontrate differenze statisticamente significative nelle misure del turnover proteico, ma questo è stato probabilmente il risultato delle ridotte dimensioni del campione: un errore statistico di tipo 2.
Conclusioni:
Gli agenti anabolizzanti, che essi siano SARM steroidei o non steroidei, possono annullare gli effetti catabolici degli ormoni tiroidei? Dai dati aneddotici ed empirici raccolti sul campo sembrerebbe molto probabile, in una certa misura, ma non ci sono dati clinici al riguardo. La variabile di picco nella questione è il dosaggio. Si è potuto osservare che gli atleti con maggiori vantaggi dalla somministrazione di T3 in regimi ipocalorici protratti li ottenevano con dosaggi nel range tra 25 e 50mcg/die massimo! Tale dosaggio, con riscontro per via esami ematici, permette all’atleta di mantenere livelli tiroidei da normo o ipercalorica, senza sforare il range di riferimento fisiologico, nonostante la forte restrizione alimentare. Ovviamente, questi atleti sono sottoposti ad una preparazione complessa comprendente l’uso di uno o più PEDs.
I dosaggi da 100-150mcg/die di T3 o 200mcg/die di T4 sono del tutto controproducenti, a meno che per il modesto aumento del dispendio energetico (poche centinaia di kcal, con un aumento del 10%-15% del tasso metabolico a riposo) siate disposti a ritrovarvi ipertiroidei e fortemente catabolici.
Oltretutto, in ipocalorica, il T4 subisce comunque una riduzione della conversione in T3. L’uso concomitante di GH può migliorare questa risposta.
In conclusione, ricordiamo gli effetti collaterali legati ad uno stato di ipertiroidismo:
- accelerazione della frequenza cardiaca;
- palpitazioni;
- possibili aritmie;
- forte calo di peso e perdita di massa muscolare;
- insonnia;
- ansia;
- tremori;
- sudorazione;
- debolezza muscolare;
- aumento del reverse T3 [legato ad abuso di farmaci contenenti T3 e/o T4].
Riflettete e traete le corrette conclusioni… la conoscenza per farlo ora non vi manca. Per la capacità beh, miracoli non ne faccio…
Gabriel Bellizzi
Riferimenti:
- Carlé, Allan, Anne Krejbjerg, and Peter Laurberg. “Epidemiology of nodular goitre. Influence of iodine intake.” Best practice & research Clinical endocrinology & metabolism 28.4 (2014): 465-479.
- Nicoloff, John T., et al. “Simultaneous measurement of thyroxine and triiodothyronine peripheral turnover kinetics in man.” The Journal of clinical investigation 51.3 (1972): 473-483.
- Bianco, Antonio C., et al. “Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases.” Endocrine reviews 23.1 (2002): 38-89.
- Mendel, Carl M. “The free hormone hypothesis: a physiologically based mathematical model.” Endocrine reviews 10.3 (1989): 232-274.
- Gereben, Balázs, et al. “Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling.” Endocrine reviews 29.7 (2008): 898-938.
- Silva, J. Enrique. “Thermogenic mechanisms and their hormonal regulation.” Physiological reviews 86.2 (2006): 435-464.
- Ismail-Beigi, Faramarz. “Thyroid hormone regulation of Na, K-ATPase expression.” Trends in Endocrinology & Metabolism 4.5 (1993): 152-155.
- Everts, M. E. “Effects of thyroid hormones on contractility and cation transport in skeletal muscle.” Acta Physiologica Scandinavica 156.3 (1996): 325-333.
- Mullur, Rashmi, Yan-Yun Liu, and Gregory A. Brent. “Thyroid hormone regulation of metabolism.” Physiological reviews 94.2 (2014): 355-382.
- Barbe, Pierre, et al. “Triiodothyronine‐mediated upregulation of UCP2 and UCP3 mRNA expression in human skeletal muscle without coordinated induction of mitochondrial respiratory chain genes.” The FASEB Journal 15.1 (2001): 13-15.
- de Lange, Pieter, et al. “Uncoupling protein-3 is a molecular determinant for the regulation of resting metabolic rate by thyroid hormone.” Endocrinology 142.8 (2001): 3414-3420.
- Gelfand, Robert A., et al. “Catabolic effects of thyroid hormone excess: the contribution of adrenergic activity to hypermetabolism and protein breakdown.” Metabolism 36.6 (1987): 562-569.
- Martin, WH 3rd, et al. “Mechanisms of impaired exercise capacity in short duration experimental hyperthyroidism.” The Journal of clinical investigation 88.6 (1991): 2047-2053.
- Tauveron, I. G. O. R., et al. “Response of leucine metabolism to hyperinsulinemia under amino acid replacement in experimental hyperthyroidism.” American Journal of Physiology-Endocrinology and Metabolism 269.3 (1995): E499-E507.
- Lovejoy, Jennifer C., et al. “A paradigm of experimentally induced mild hyperthyroidism: effects on nitrogen balance, body composition, and energy expenditure in healthy young men.” The Journal of Clinical Endocrinology & Metabolism 82.3 (1997): 765-770.
