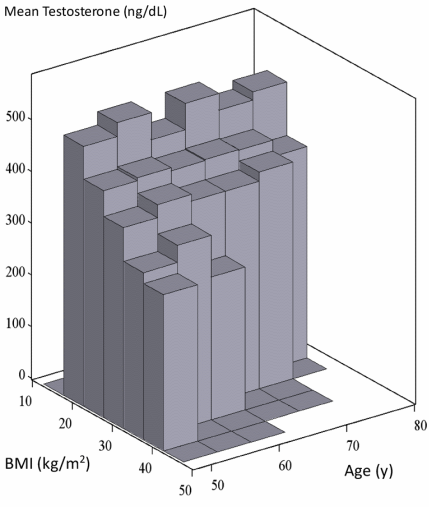
Siamo tutti al corrente del fatto che con l’invecchiamento si verifica un calo esponenziale dei livelli di Testosterone nell’uomo, e ciò è stato riportato da decine di studi. Ma secondo i ricercatori dell’American Cooper Institute, la relazione tra invecchiamento e Testosterone è un po’ più complessa di così. I ricercatori hanno preso in esame circa tremila uomini sani di età compresa tra i 50 ed i 79 anni non trovando alcuna associazione significativa tra età e Testosterone se i soggetti erano sottoposti ad allenamenti di diversa intensità e avevano un BMI inferiore. Hanno evidenziato, come già ipotizzato e riportato in altri studi, che il BMI e il livello di allenamento hanno un impatto significativo sulla concentrazione di Testosterone ematico degli uomini. (1)
I ricercatori hanno misurato i livelli di Testosterone dei partecipanti allo studio al mattino. Hanno classificato un livello di Testosterone inferiore a 250ng/dl come basso, un livello di 250-399ng/dl come normale anche se nel range basso e un livello di oltre 400ng/dl come normale.
I ricercatori hanno anche rimosso un uomo dal loro campione di studio perché il suo livello di Testosterone superava i 3000ng/dl. Sebbene l’individuo in questione non abbia riportato alcun uso di farmaci, i ricercatori hanno tuttavia deciso di non credergli.
I ricercatori hanno fatto correre gli uomini presi in esame su un tapis roulant in modo che potessero misurare il loro livello di allenamento. Hanno determinato anche il loro peso corporeo. Poi hanno osservato se vi era un’associazione tra il livello prestazionale, l’indice di massa corporea, l’età e il livello di Testosterone degli uomini.
L’età non è risultata una determinante assoluta dei livelli di Testosterone degli uomini. I ricercatori esaminando tutti i gruppi di uomini divisi per fascia d’età (età compresa tra i 50-59 anni, tra i 60-69 anni e tra 70 e 79 anni) hanno osservato la percentuale di uomini con bassi livelli di Testosterone attestarsi tra il 10 e l’11%.
Sia il BMI che il livello di allenamento hanno influenzato le concentrazioni di Testosterone. Più i soggetti presentavano un BMI inferiore e una grado di allenamento maggiore, più alto era il loro livello di Testosterone.
I ricercatori hanno scritto che questo studio non supporta la tesi secondo cui il Testosterone diminuisce in modo uniforme con l’invecchiamento in uomini sani di mezza età o anziani. L’individuazione di livelli simili di Testosterone in soggetti di fascia d’età diversa (dai 50 ai 79 anni) suggerisce che livelli di Testosterone cronicamente bassi non sono una conseguenza inevitabile dell’invecchiamento.
I livelli di Testosterone dei soggetti presi in esame erano inversamente associati al BMI e positivamente associati al livello di allenamento. Questa nuova scoperta di una forte associazione tra grado prestazionale misurato oggettivamente e Testosterone normale nelle fasce di età più anziane ha implicazioni interessanti. Sono necessarie ulteriori ricerche per comprendere meglio le implicazioni di tali scoperte, compresa la comprensione dei livelli di Testosterone nell’invecchiamento in soggetti in buona salute, se i livelli bassi di Testosterone aumentano significativamente con un incremento del livello di allenamento e se i livelli di Testosterone possono essere mantenuti entro una buona soglia nonostante l’eccesso di peso e/o la presenza di malattie croniche con un setting di attività da moderato ad alto.
Qualche tempo fa pubblicai un articolo nel quale evidenziavo, attraverso diversi studi, come l’uso di dolcificanti “low calorie” a dosaggi variabili (Aspartame da 0, 350mg a 1050 mg/die) da parte di individui sani non avesse un impatto significativo sui livelli di glucosio e insulina in risposta al test di tolleranza al glucosio, sulla massa e composizione corporea, sulla fame, pienezza e sete e sull’HbA1c (emoglobina glicata). Secondo uno studio pubblicato nel 2015 sull’American Journal of Clinical Nutrition, le persone in sovrappeso o obese che cercano di perdere peso attraverso una riduzione dell’introito calorico giornaliero e all’inserimento o aumento dell’attività fisica, ottengono migliori risultati se consumano acqua al posto di bevande light ai pasti. (1)
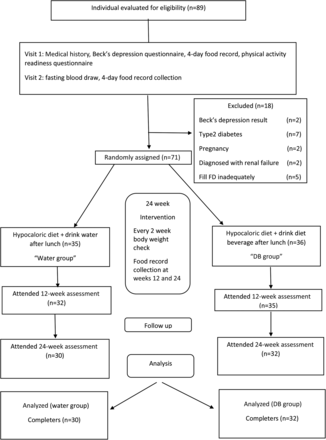
Lo studio clinico in questione era randomizzato a 2 bracci, in singolo cieco. I partecipanti eleggibili sono stati assegnati in modo casuale dopo le misure di base utilizzando un metodo sulla casualità numerica generata dal coordinatore del progetto con allocazione nascosta dai partecipanti e dai dietologi che li seguivano fino a quando la randomizzazione non è stata rivelata ai partecipanti all’inizio dello studio.
I partecipanti all’esperimento erano sessantadue donne in sovrappeso o obese (BMI 27-40) di età compresa tra i 18 ed i 50 anni.
Per 24 settimane le donne prese in esame hanno seguito una dieta ipocalorica, che creava un deficit di 500-1000Kcal rispetto al fabbisogno giornaliero. Inoltre, le donne sono state spronate ad allenarsi con intensità moderata per 60 minuti al giorno.
I partecipanti allo studio sono stati divisi in due gruppi: Il “gruppo acqua”(Water), composto al termine dello studio da 30 soggetti, nel quale è stata sostituita l’assunzione abituale di bevande light con un bicchiere d’acqua (250 mL) assunto dopo il pasto principale (pranzo), e il “gruppo bevanda dietetica”(DB), composto al termine dello studio da 32 soggetti, nel quale i componenti sono stati istruiti a bere una bevanda dietetica una volta al giorno (250 mL) dopo il pasto principale (pranzo) per 5 volte a settimana con consumo di sola acqua dopo il pasto principale (pranzo) nei restanti giorni della settimana. Le assunzioni di liquidi sono state abbinate tra i 2 gruppi nel loro pasto principale, ma entrambi i gruppi erano liberi di bere acqua come bevanda durante il resto dei giorni ma non erano autorizzati a consumare bevande dietetiche. Inoltre, a entrambi i gruppi è stato chiesto di non bere bevande dietetiche o acqua durante il pasto e di non aggiungere dolcificanti ipocalorici come l’aspartame o il sucralosio a bevande come tè o caffè. Sono state richieste visite bisettimanali al dietista per promuovere l’aderenza dei soggetti alla dieta ipocalorica e alla sostituzione delle bevande. Quelli del gruppo DB hanno ricevuto le loro bevande durante le visite di trattamento bisettimanali. Entrambi i gruppi hanno iniziato una dieta ipocalorica secondo il protocollo NovinDiet (nome della clinica dove si è svolto l’esperimento), che includeva consigli per aumentare gradualmente i livelli di attività fisica per raggiungere i 60 minuti di attività moderata per 5 giorni a settimana.
La Clinica NovinDiet è una clinica privata per la perdita di peso che utilizza un approccio integrato (dietetico, comportamentale, esercizio e trattamenti medici). Il protocollo NovinDiet si basa sullo sviluppo di un approccio di problem-solving per ogni soggetto in modo individuale e affronta la gestione sia della dieta che dell’esercizio fisico. In questo studio, il programma è stato progettato per consentire la perdita di peso del 7-10% del peso corporeo iniziale, ad una velocità di 0,5-1 kg / settimana per 24 settimane. I programmi di dieta individuali erano basati sui registri del diario alimentare dei pazienti e sulle loro preferenze alimentari, con modifiche graduali per adeguare la loro dieta al protocollo NovinDiet. La dieta ipocalorica alla quale sono stati sottoposti i soggetti dello studio era ad alto contenuto glucidico, e a basso contenuto di grassi saturi [17% delle calorie dalle proteine, il 23% dai grassi (<10% da grassi saturi) e il 60% dai carboidrati].
Alle sessioni bisettimanali sono stati discussi i problemi comportamentali riferiti dai soggetti riguardo al loro programma per la perdita di peso. Sono stati forniti opuscoli che i soggetti si portavano a casa e che utilizzavano per registrare l’aderenza al protocollo dietetico. Durante il periodo di intervento, i soggetti hanno completato il modulo di feedback in merito alla loro aderenza alla dieta assegnata e all’attività fisica svolta. I soggetti hanno anche avuto accesso a un sito Web, a riviste Internet settimanali e al supporto telefonico/online da parte di in consulente, se necessario.
Le misurazioni antropometriche di tutti i soggetti sono state prese al basale, 12 settimane e 24 settimane dopo l’inizio del programma (eccetto l’altezza, che è stata presa solo durante la visita di screening) dal dietologo.
L’assunzione macrocalorica è stata rilevata al basale e l’ultima settimana dell’intervento (settimana 24) sono stati analizzati dal software Nutritionist IV (versione 4.1; Hearst). Campioni di sangue di tutti i soggetti sono stati prelevati dopo un digiuno notturno (10-12 ore), tra le 7:00 e le 9:00, al basale e 24 settimane dopo l’inizio del programma per le misurazioni biochimiche, cellulari e ormonali. I campioni di sangue a digiuno sono stati raccolti mediante venipuntura secondo un protocollo standard.
I soggetti sono stati sottoposti a misurazione della curva glicemica e insulinica (prelievo ematico dopo 2h dall’assunzione di 75g di glucosio) secondo il metodo standard della American Diabetes Association per escludere condizioni diabetiche. Il glucosio plasmatico a digiuno (FPG) e le concentrazioni di glucosio 2h post prandiali sono state misurate utilizzando il metodo colorimetrico enzimatico. L’insulina è stata misurata utilizzando un dosaggio radioimmunologico con insulina umana marcata con 125I e un antisiero di insulina umana ad un dosaggio immunoradiometrico (IRMA; Biosource) con un sistema a contatore γ (Gamma I; Genesys). La resistenza all’insulina è stata valutata attraverso l’indice HOMA-IR, che è stato calcolato utilizzando la seguente formula:
E’ stata effettuata anche la misurazione dell’emoglobina glicata (HbA1c), mediante un metodo colorimetrico dopo una separazione iniziale mediante cromatografia a scambio ionico (Biosystem), e l’analisi biochimica del colesterolo totale sierico (TC), dei trigliceridi e del colesterolo HDL tramite un analizzatore automatico Selectra E (Vita Laboratory) seguendo le procedure standard dei kit diagnostici di Pars Azmoon. Il colesterolo LDL è stato calcolato usando la formula di Friedewald:
Degli 89 soggetti obesi e in sovrappeso reclutati per lo studio, 62 hanno completato l’intervento di 24 settimane (il 71% della popolazione originale). Dei 27 soggetti esclusi, 2 lo sono stati a causa dei risultati del Beck Depression Inventory. Cinque soggetti sono stati esclusi perché hanno smesso di tenere il registro alimentare o lo hanno riempito in modo insufficiente. I risultati degli esami del sangue al basale hanno rivelato che 11 pazienti non erano eleggibili perché avevano un criterio di esclusione ≥1.
Al basale, non c’erano differenze statisticamente significative nelle caratteristiche fisiche o nelle misurazioni biochimiche tra i gruppi di intervento o tra coloro che hanno completato o non completato lo studio una volta reclutati.
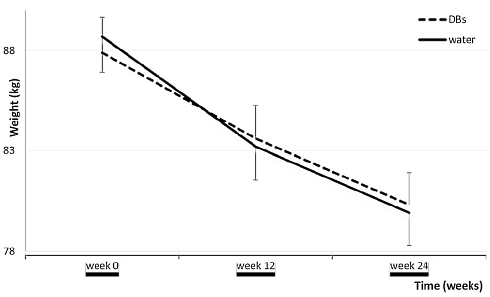
Una volta concluso il test, il “gruppo acqua” ha mostrato una perdita di peso maggiore di 1,2kg rispetto al gruppo DB. Il risultato era statisticamente significativo.
Le riduzioni della concentrazione di Colesterolo TC, LDL e Trigliceridi e l’aumento del Colesterolo HDL sono state rilevate durante le 24 settimane di studio in ciascun gruppo (P <0,001), ma non vi erano differenze significative in questi risultati tra i gruppi dopo le 24 settimane.
Il TC dopo 24 settimane era diminuito di -0,53 ± 0,16 mmol/L nel “gruppo acqua” rispetto agli -0,49 ± 0,17 mmol/L del gruppo DB.
Le concentrazioni di HDL dopo 24 settimane erano aumentate di 0,125 ± 0,045mmol/L nel gruppo acqua rispetto agli 0,124 ± 0,050mmol/L del gruppo DB, mentre il colesterolo LDL era diminuito di -0,534 ± 0,164 mmol/L (acqua) e -0,551 ± 0,191 mmol/L (DB).
Le concentrazioni di Trigliceridi dopo 24 settimane sono diminuite di -0.281 ± 0.109 mmol / L (acqua) e -0.280 ± 0.122 mmol / L (DB).
I ricercatori non hanno certezza sulla causa esatta di questa differenza, ma sospettano che possa ricercarsi nella sensibilità all’insulina. Se si osserva il grafico sopra riportato la sensibilità all’insulina è leggermente migliorata nel “gruppo acqua” rispetto al gruppo DB.
L’analisi dei dati ha mostrato che l’FPG, l’insulina a digiuno, la curva del glucosio (2hpp), l’HbA1c e l’indice HOMA-IR diminuivano nel tempo in entrambi i gruppi (P <0,001 per effetto tempo di tutte le variabili). Tuttavia, le differenze tra i gruppi erano significative solo per l’insulina, 2hpp e HOMA-IR. La riduzione media del glucosio plasmatico a digiuno su 24 settimane era di -0.767 ± 0,278 mmol / L (acqua) e -0,681 ± 0,169 mmol / L (DB).
In termini di differenze del 2hpp durante le 24 settimane di intervento, la riduzione media di questo era -1.024 ± 0.254 mmol / L (acqua) e -0.719 ± 0.275 mmol / L (DB).
Il calo dell’HbA1c su 24 settimane nel gruppo acqua era -0,54% ± 0,12%, mentre questa riduzione era pari a -0,47% ± 0,17% nel gruppo DB alla settimana 24.
Le concentrazioni sieriche di insulina a digiuno sono diminuite significativamente nel tempo, con differenze tra i 2 gruppi (P <0,001). La riduzione dell’insulina era di -2,8 ± 0,8 mU / L (acqua) e -1,8 ± 1,3 mU / L (DB) dopo 24 settimane. Inoltre, vi è stato un significativo miglioramento della resistenza all’insulina nel gruppo acqua rispetto al gruppo DB su 24 settimane. L’HOMA-IR è diminuito di -0,097 ± 0,049 nel gruppo acqua rispetto a -0,057 ± 0,042 del gruppo DB su 24 settimane.
In conclusione, i ricercatori scrivono che sostituire le bevande dietetiche con il consumo di acqua sembra avere un impatto positivo sulla perdita di peso e sulla sensibilità all’insulina nei soggetti sovrappeso e obesi che seguono una dieta ipocalorica e che ora sono necessari studi a più lungo termine.
Nonostante il presente studio mostri un ottimo design, vi sono delle “lacune” che possono alterare in parte il risultato finale, nella fattispecie per quanto riguarda le conclusioni in merito. Una di queste è rappresentata dalla “buona fede” posta nei confronti dei soggetti presi in esame e del loro impegno a svolgere l’attività fisica consigliata in modo omogeneo. Dato che la perdita di peso è determinata principalmente dall’input e dall’output calorico, è possibile che parte dei soggetti trattati non abbiano rispettato le direttive dategli soprattutto per quanto riguarda l’attività fisica. Anche la differenza d’età tra i soggetti monitorati può in parte “viziare” i risultati. Inoltre, i soggetti esaminati non possono essere considerati pienamente “sani” (mi riferisco soprattutto agli individui obesi), e le loro caratteristiche non li rendevano particolarmente avvantaggiati nel seguire una dieta ipocalorica con una percentuale glucidica del 60% (questione che andrebbe analizzata caso per caso in base alla storia clinica del paziente cosa che, nonostante la personalizzazione del programma, non sembra essere stata presa totalmente in considerazione). Nonostante ciò, i fattori elencati non possono spiegare del tutto una riduzione media di peso superiore di 1,2Kg del gruppo che consumava un bicchiere d’acqua post pranzo. Quindi, di conseguenza, allineandomi con quanto detto dai ricercatori, l’impatto della bevande dietetiche (e dei dolcificanti) nei soggetti sovrappeso o obesi va indagato ulteriormente e con controlli più rigidi sui pazienti.
Sebbene non siano legali in alcuni paesi, gli integratori per la perdita di peso contenenti l’estratto di Hoodia gordonii sono facilmente reperibili sul web. La Hoodia gordonii fa parte di un genere di piante (Hoodia) appartenenti alla famiglia delle Apocinacee (sottofamiglia Asclepiadoideae), endemiche dell’Africa sudoccidentale (Angola, Botswana, Namibia, Sudafrica e Zimbabwe), usate dai boscimani a scopi medicinali, e che hanno recentemente suscitato l’interesse delle industrie farmaceutiche per le loro possibili applicazioni per il trattamento dell’obesità. Ciò nonostante, i supplementi contenenti l’estratto di Hoodia gordonii non hanno avuto il successo previsto. E’ possibile, però, che gli integratori contenenti l’estratto di Hoodia parviflora possano avere maggiore successo rispetto a questi ultimi. Bruxelles ne ha permesso la commercializzazione in Europa entro precisi limiti: la quantità di Hoodia parviflora non deve superare i 9,4mg di estratto per dose giornaliera. (1)
Le popolazioni dell’Africa sudoccidentale utilizzano parti essiccate di queste piante per sopprimere la fame, e negli anni ’60 dei ricercatori sudafricani hanno iniziato a svolgere studi al fine di realizzare prodotti dimagranti che sfruttassero le proprietà di queste piante.
Nel primo decennio del ventunesimo secolo, la Pfizer e in seguito la Unilever, pensarono che la componente simil-steroidea p57 contenuta nella Hoodia gordonii potesse essere un componente interessante per la realizzazione di farmaci dimagranti o dei così detti alimenti funzionali per la perdita di peso, ma interruppero le loro ricerche quando risultò che il p57 causava nausea agli utilizzatori.
La Israeli Desert Labs (2) ha svolto studi sull’effetto dell’estratto di Hoodia parviflora per diversi anni iniziando a realizzare diversi prodotti contenenti l’estratto di questa pianta. I prodotti in questione comprendono integratori, gomme da masticare, tè e barrette.
Secondo la ricerca sugli esseri umani condotta dalla stessa Israeli Desert Labs, i soggetti che assumono un equivalente di 142,5mg di estratto secco di Hoodia parviflora per 40 giorni consecutivi subiscono un calo di peso corporeo di poco superiore a 0,5Kg. (3) Tale effetto è stato registrato su soggetti i quali non avevano applicato cambiamenti al proprio stile di vita. Una nota interessante è che non sono stati segnalati effetti collaterali in seguito alla somministrazione dell’estratto.
Nel 2017, l’Autorità Europea per la Sicurezza Alimentare (EFSA) ha deciso di autorizzare la vendita nel mercato europeo dei supplementi contenenti l’estratto di Hoodia parviflora con un limite massimo per dose giornaliera pari a 9,4mg.(4) Una dose decisamente inferiore ai 142,5mg utilizzati dalla Desert Labs e che hanno mostrato di poter causare un effetto coadiuvante per la perdita di peso.
Esiste comunque la possibilità che qualche lungimirante chimico trivi il modo per eludere le normative dell’UE. I principi attivi contenuti nell’estratto di Hoodia parviflora, gli hoodigosidi, sono efficaci in quantità molto piccole. In 142,5mg di estratto ve ne sono al massimo 1,43mg.
Con un buon processo di purificazione, è teoricamente possibile produrre supplementi per la perdita di peso contenenti l’estratto di Hoodia parviflora dalla significativa efficacia, rimanendo completamente conformi alle normative europee.
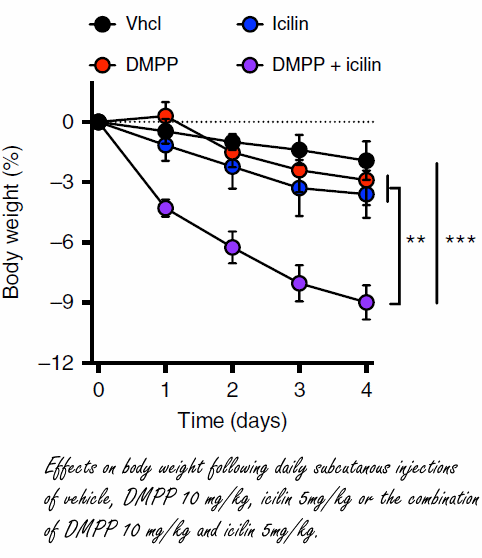
Scienziati danesi potrebbero essersi imbattuti in una strategia farmacologica completamente nuova per indurre la perdita di grasso. Essi hanno scoperto che uno stimolo simultaneo dei recettori dell’Icilina e del DMPP porta a una rapida perdita di grasso nei topi. (1)
Il recettore TRPM8 si trova negli strati esterni della pelle, dove si percepisce il freddo. Se il TRMP8 viene attivato, l’attività del metabolismo lipidico aumenta a causa, tra le altre cose, di un incremento dell’ossidazione lipidica nel tessuto adiposi marrone. Per esempio, il Mentolo stimola il recettore TRMP8.
I ricercatori hanno scoperto un composto sintetico che stimola il TRMP8 e che è quasi 200 volte più potente del Mentolo e 2,5 volte più efficace: l’Icilina. Questo composto è stato iniettato a dosaggi differenti in topi sovrappeso permettendo ai ricercatori di osservare una perdita di grasso da parte degli animali dipendente da un aumento del loro dispendio energetico.
Tuttavia, il primo autore dello studio che qui si sta trattando, Christoffer Clemmensen, affiliato all’Università di Copenhagen, ha affermato che il recettore TRMP8 non è presente nel tessuto adiposo marrone. (2) Sembra che il recettore del freddo [TRMP8] sulla superficie della pelle mandi un segnale al cervello che successivamente attiva il tessuto adiposo marrone tramite i connettori nervosi.
I topi hanno subito una riduzione della percentuale del grasso corporeo quando trattati con Icilina per via di un aumento del loro turnover energetico. Tuttavia, l’effetto riscontrato non era sufficientemente incisivo da portare ad un effetto reale su ipotetici pazienti, anche la dove l’azione della molecola venisse ottimizzata. Un punto fondamentale che i ricercatori danesi non hanno tralasciato, è che se si desidera modificare il peso corporeo di un soggetto, non è sufficiente prendere di mira solamente il turnover energetico. Come affermato da Christoffer Clemmensen, per creare davvero un bilancio energetico negativo, è necessario anche fare in modo che il soggetto mangi di meno.
E’ noto che i fumatori mangiano meno delle persone che non fumano, e ciò è dovuto in parte perché la Nicotina attiva il sottotipo del recettore Nicotinico (nAChR) alfa3beta4. I ricercatori hanno scoperto che la sostanza sintetica Dimetilfenilpiperazinio [DMPP] funziona allo stesso modo. E così ne hanno sperimentato l’effetto iniettandola nei topi sovrappeso.
Come conseguenza, gli animali trattati mangiavano di meno e perdevano peso.
Il DMPP non solo sopprime l’appetito, ma ha anche un enorme effetto positivo sul metabolismo glucidico rispetto alla Nicotina, la quale ha un effetto negativo sul grasso epatico e sulla sensibilità all’insulina.
I ricercatori hanno cosomministrato ai topi l’Icilina e il DMPP ottenendo un effetto sinergico sulla perdita del peso corporeo. Presi singolarmente, i due composti non causano effetti particolarmente significativi sulla perdita di peso, ma una loro assunzione combinata ha mostrato di poter causare una marcata perdita di peso.
I ricercatori non sono certi che tale trattamento possa essere pienamente efficace e sicuro nell’uomo. Questo studio, come affermato anche dagli stessi autori, rappresenta semplicemente una prova preliminare.
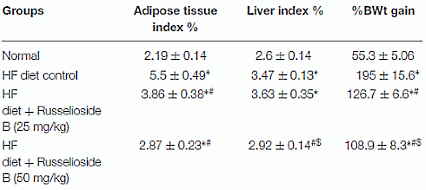
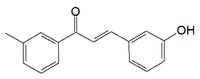
Recentemente il farmacologo egiziano Essam Abdel-Sattar ha identificato nella pianta Caralluma Quadrangula una sostanza steroidea la quale ha mostrato alcuni notevoli effetti sulla perdita di grasso negli studi svolti su animali. (1)
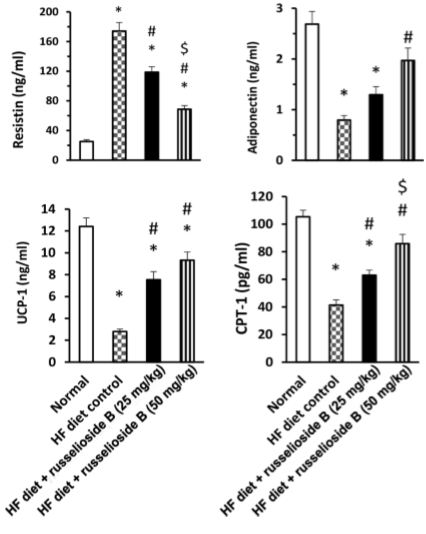
La Caralluma Quadrangula è una pianta grassa particolarmente diffusa nella penisola arabica. Da questa pianta, i ricercatori hanno estratto la sostanza steroide-simile Russelioside B che hanno somministrato ai ratti attraverso la loro alimentazione.
Se ad essere presi in esame fossero stati degli esseri umani adulti, la dose di Russelioside B ad essi somministrata sarebbe stata di circa 200-300mg [nei topi 25 mg/kg] o 400-600mg [nei topi 50 mg/kg] al giorno.
Per 16 settimane il contenuto calorico della dieta degli animali presi in esame è stato aumentato attraverso un surplus lipidico.
Al primo gruppo di controllo sono stati somministrati mangimi ricchi di grassi senza l’aggiunta di alcuna sostanza bioattiva; agli animali del secondo gruppo di controllo è stata somministrata un’alimentazione standard, anch’essa senza l’aggiunta di sostanze bioattive.
Sia gli animali trattati con la dosa bassa [25 mg/kg] che quelli trattati con la dose alta [50 mg/kg] di Russelioside-B, hanno mostrato che questa sostanza esercita un azione inibitoria considerevole sull’aumento dei depositi adiposi. Ciò nonostante, come c’era da aspettarselo, la dose alta ha esercitato una azione più marcata rispetto a quella bassa.
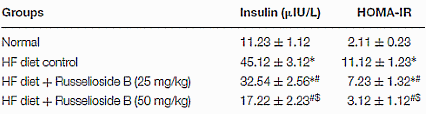
Il Russelioside B ha anche inibito la crescita degli adipociti [vedi figura seguente]. Il glicoside gravidico ha salvaguardato l’efficacia dell’Insulina limitando l’aumento del HOMA-IR [unità di misura per calcolare il grado di insulino resistenza].
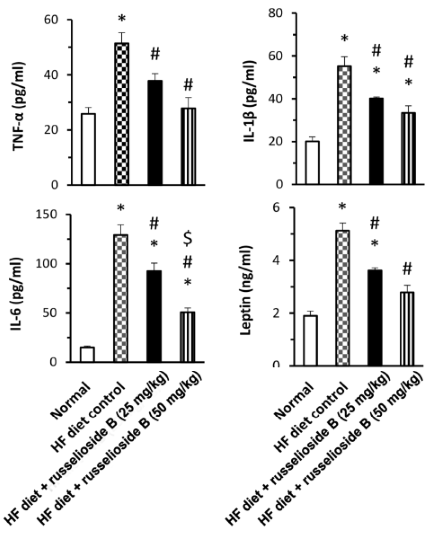
I ricercatori sospettano che il Russelioside-B funzioni attraverso molteplici meccanismi. Uno di questi è rappresentato dall’attività inibitoria sui fattori infiammatori come l’interleuchina 1-beta, l’interleuchina-6 e il TNF-alfa.
Un altro possibile meccanismo è legato alla capacità del Russelioside B sull’aumento del dispendio energetico cellulare. Il Russelioside-B ha inibito la riduzione di UCP-1 e UCP-2 causata da una dieta ipercalorica. Le UCP sono proteine disaccoppianti della membrana mitocondriale interna e sono in grado di dissipare il gradiente protonico generato dal NADH tra la matrice mitocondriale e lo spazio intermembrana mitocondriale. L’energia dissipata non viene utilizzata per lavoro biochimico e viene dispersa sotto forma di calore; difatti le UCP sono associate alla termogenesi.
I ricercatori hanno concluso che, il Russelioside B ha esercitato un controllo sull’aumento di peso, ha migliorato il profilo lipidico e il deterioramento infiammatorio che accompagna l’obesità e l’insulino-resistenza indotte dalla dieta ipercalorica. Inoltre, il Russelioside B ha modulato l’espressione delle adipochine e ha aumentato l’espressione e il livello delle proteine legate al dispendio energetico. Pertanto, l’azione antiobesgena complessiva del Russelioside B può essere, almeno in parte, attribuita alle sue attività antinfiammatorie e sulla modulazione delle adipochine, oltre al suo effetto favorevole sul dispendio energetico. Studi futuri sono giustificati per indagare le azioni farmacologiche del Russelioside B su organi importanti come il fegato e per esplorare appieno i suoi meccanismi compensatori sugli effetti metabolici di una alimentazione ad alto contenuto lipidico nei ratti.
Se non avete letto ancora la prima, la seconda, la terza, la quarta e la quinta parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte – 3° Parte– 4° Parte– 5° Parte.
Vanadio: caratteristiche e possibili applicazioni.
Vanadio Solfato
Il Vanadio è noto per essere un minerale “ultratraccia” nell’alimentazione umana. (1) Non fa parte delle classiche 24 vitamine e minerali, anche se è necessario per alcune reazioni nel corpo, come la formazione di un Pervanadato richiesto per la fosforilazione di alcuni recettori proteici. (2) Normalmente vengono consumati giornalmente dai 6 ai 20mcg di Vanadio, principalmente attraverso il consumo di peperoni e frutti di mare. (2) Altre fonti alimentare che presentano una concentrazione significativa di questo minerale sono i cereali integrali, la birra, la frutta secca e alcune qualità di funghi. Nei cereali, e in modo particolare nella farina e nel pane, le concentrazioni di Vanadio possono superare i 10mcg/kg.
Il Vanadio ha una storia di utilizzo nel trattamento del diabete, patologia nella quale la somministrazione orale di Vanadio di Sodio rappresentava l’intervento farmacologico di base prima della scoperta dell’Insulina avvenuta nel 1921. (3)
Il Vanadio (Vanadato) può anche formare complessi legandosi al Perossido di Idrogeno (H202) e formando il Perossivanadio (Pervanadato), biologicamente più attivo nell’attività inibitoria nei confronti delle Proteine Fosfotirosina-Fosfatasi (PTP) rispetto al Vanadato. (4) (5)
Il Vanadio, come Vanadato in vivo, inibisce la fosforilazione delle ATPasi di tipo P (6) (7) e delle Proteine Fosfotirosina-Fosfatasi (PTP). (8) La dose per inibire le ATPasi, tuttavia, è clinicamente significativa solo a livelli tossici. L’inibizione delle PTP aumenta i vari processi di fosforilazione della Tirosina (poiché le fosfatasi sono enzimi inibitori nella fosforilazione).
Vari effetti derivano dall’inibizione delle PTP, come l’inibizione dell’assorbimento amminoacidico cellulare del sistema di trasporto A degli AA neutri (5), l’aumento dell’assorbimento del glucosio e la mobilizzazione dei GLUT4 attraverso una via indipendente dal fosfoinositolo-3-chinasi (PI3K) [a differenza dell’Insulina]. (9) Il meccanismo attraverso il quale il Vanadio esplica i suoi effetti si trova nella sua azione insulino-mimetica e nell’aumento della fosforilazione generale che agisce sulla Tirosina Chinasi del recettore insulinico (la quale viene fosforilata sotto l’influenza dell’Insulina, oltre all’inibizione delle PTP).
Potenziali siti di azione del Vanadio nella cascata di segnalazione dell’Insulina. Il legame del recettore insulinico sulla superficie della cellula determina i cambiamenti conformazionali nel recettore, principale attivazione della via PI3-K, la via principale implicata nella mediazione degli effetti metabolici. I potenziali siti di azione del Vanadio in questo percorso sono riassunti in questa figura. V ()), V (+), denotano effetti negativi, positivi o nulli, rispettivamente. IRS, substrato del recettore dell’insulina; PI3-K, fosfatidilinositolo-3-chinasi; PDK, chinasi 3-fosfoinositide-dipendente; PKB, protein chinasi B; GSK- glicogeno sintasi chinasi-3; PFK-2, fosfofuctokinasi-2; GLUT 4, trasportatore di glucosio di tipo 4; 4E- 4 proteina legante; PTP1B, proteina tirosina fosfatasi 1B; PP-1, proteina fosfatasi-1.Sodio Ortovanadato
Composti contenenti Vanadio sono noti per inibire le PTP (10) (11) poiché essi tendono a formare una struttura bipirramidale trigonale che può agire come un inibitore competitivo del trasferimento di fosforile (come nel caso del Sodio Ortovanadato) o un ossidante del residuo di cisteina del ciclo PTP (complessi Perossivanadati). (12) Similmente ad altri composti inibitori delle PTP (gli inibitori PTP1B includono Berberina e Acido Ursolico) il Vanadio può prolungare la segnalazione attraverso il Recettore dell’Insulina prevenendo la regolazione negativa della degradazione dell’IRS-1, che è indotta a valle del segnale del recettore insulinico (tramite mTOR [13] S6K1 (14) ) e funziona tramite le PTP per sopprimere (15) e stimolare la degradazione (16) (17) delle IRS-1 come forma di feedback negativo.
IRS-1
L’Ortovanadato di Sodio a 1mmol/L sembra migliorare la segnalazione (miocita L6) di circa il 25% ed è associato all’inibizione del declino dell’attività complessa PI3K/IRS-1, ma è stato osservato che non inibisce la perdita di massa della proteina IRS-1. (18)
In uno studio nel quale sono stati presi in esame diabetici di tipo II e soggetti obesi ma ancora considerabili come sani (gruppo di controllo) è emerso che 100mg di Vanadio per 3 settimane sono stati in grado di sopprimere la gluconeogenesi epatica in entrambi i gruppi, ma solo il gruppo dei diabetici ha riscontrato un aumento della sensibilità all’Insulina mentre i soggetti del gruppo di controllo non hanno avuto cambiamenti significativi nei parametri della glicemia. (19)
Uno studio svolto su esseri umani (n = 14) di età compresa tra i 40 e i 50 anni con alterata tolleranza al glucosio, ha mostrato che la somministrazione di Vanadio ad un dosaggio di 50mg due volte al giorno (100mg al giorno) per un periodo di un mese (30 giorni), in risposta ad una condizione iperinsulinemica euglicemica, non ha prodotto miglioramenti della sensibilità all’Insulina e ha portato ad un piccolo ma significativo aumento dei Trigliceridi. (20) In questo studio non si sono osservate modifiche del LDL-C, HDL-C o del Colesterolo totale mentre il prima citato aumento dei Trigliceridi consisteva in un incremento delle concentrazioni ematiche da 1,4 +/- 0,6mmol/L a 1,7 +/- 0,5mmol/L.
Uno studio condotto su diabetici di tipo II (n = 6) ha osservato che dopo la somministrazione di Vanadio a 100mg al giorno durante 3-5 settimane di un periodo complessivo di 7 settimane (con placebo somministrato all’inizio e nell’ultimo fase dello studio) ha osservato che dopo 3 settimane di somministrazione di Vanadio si assisteva ad un aumento dell’assorbimento di glucosio e della sensibilità all’Insulina che era ancora presente due settimane dopo la cessata somministrazione del minerale; questi miglioramenti sono stati accompagnati da una diminuzione dell’HbA1c da 9,6 + 0,6% a 8,8 ± 0,6%. (21) L’utilizzo del Vanadio (come Solfato) ad un dosaggio di 150mg per 6 settimane è stato associato a una riduzione del 20% della glicemia a digiuno e ad una diminuzione del HbA1c da 8,1 ± 0,4 a 7,6 ± 0,4%. (22) Sebbene questo studio abbia rilevato una riduzione della produzione endogena di glucosio del 20% nei diabetici, la correlazione di questa riduzione alla diminuzione della glicemia a digiuno è stata di r = 0.6. (22)
Tuttavia, la qualità di questi studi è stata messa in discussione. E’ stata svolta una review sistemica la quale mirava a valutare tutti gli studi in doppio cieco controllati con placebo sul Vanadio e la ricerca preliminare terminò quando non ne trovarono nessuno. (23) Gli studi esistenti e quelli citati sopra tendono a non avere alcun gruppo placebo e sono limitati a campioni di dimensioni ridotte; i loro risultati significativi hanno perciò una potenza statistica molto inferiore rispetto ad altri composti.
Uno studio nel quale è stata utilizzata la Rapamicina (un inibitore dell’mTOR, utilizzato in questo studio per inibire la degradazione dell’IRS-1 che è un regolatore negativo della segnalazione dell’Insulina) ha osservato che la cosomministrazione di questa con l’Ortovanadato di Sodio (1mmol/L) risultava sinergica nell’aumento dell’assorbimento di glucosio in un miocita L6 in presenza di Insulina da 26,42 +/- 3,73% (osservata con il solo Vanadio) a 97,83 +/- 8,54% (in combinazione: la sola Rapamicina ha causato un assorbimento del 39,9 ± 3,39%). (18) Ciò era dovuto all’inibizione della degradazione dell’IRS-1 e al prolungamento della segnalazione attraverso il Recettore dell’Insulina, che promuoveva l’assorbimento del glucosio indotto dal PI3K, ed era probabilmente legato al fatto che il Vanadio poteva inibire la degradazione dell’associazione PI3K / IRS-1 (Rapamicina inefficace) mentre la Rapamicina impediva l’effettiva degradazione delle concentrazioni della proteina IRS-1 (Vanadio inefficace). (18)
La carenza di Vanadio non è stata descritta nell’uomo. Un suo scarso apporto, o mancanza, negli animali causa sterilità, riduzione della eritropoiesi (con conseguente anemia), difetti del metabolismo del ferro, alterata formazione ossea, dentale e cartilagginea. Non esiste quindi un RDA per il Vanadio: si è ipotizzato che un’assunzione giornaliera nel range dei 10 – 100mcg sia con tutta probabilità sufficiente.
Il Vanadio viene commercializzato come integratore alimentare sotto forma di Vanadilsolfato [VOSO4]. La biodisponibilità del Vanadio supplementare è scarsa (meno del 5%) e la maggior parte di esso viene espulso attraverso le feci.
Alcuni prodotti conteneti Vanadilsolfato riportano in etichetta che il composto “aumenta i livelli plasmatici di Insulina”, ma, da quanto è possibile estrapolare dalle informazioni sopra riportate, e cioè da quanto la letteratura scientifica ci dice sull’azione del composto, il Vanadio facilita il meccanismo d’azione dell’ormone peptidico, con una probabile azione (parzialmente dimostrata) insulino-mimetica.
Prendendo in attenta considerazione le informazioni derivanti dalla letteratura scientifica sulle possibili potenzialità del Vanadio nei confronti del metabolismo glucidico si evince, e non è un caso isolato tra i GDA, che la sua efficacia sia di grado significativo nei soggetti patologici (vedi soggetti con diabete di tipo II) e non negli individui sani. Ipoteticamente si potrebbero avere dei vantaggi dal suo utilizzo nei soggetti con una insulino-resistenza di base genetica, nei periodi di alimentazione ipercalorica prolungata (vedi peggiormanto dell’insulino-resistenza in tale contesto) e/o durante e dopo l’uso dell’Insulina esogena: durante, il suo potenziale effetto insulino-mimetico potrebbe permettere di per se di utilizzare dosi inferiori dell’ormone mentre, nel periodo successivo, potrebbe agevolare il ripristino di un ottimale insulino-sensibilità in concerto con altri GDA. L’effetto del Vanadio sulla riduzione della gluconeogenesi epatica potrebbe rappresentare un vantaggio nei periodi di restrizione calorica, in specie durante le diete Ketogeniche o simil tali. Sicuramente, l’impatto negativo del Vanadio sui Trigliceridi potrebbe causare un ulteriore peggioramento della dislipidemia indotta dall’uso di AAS.
E’ bene tenere a mente che le dosi efficaci per ottenere un qualche effetto positivo dall’integrazione con Vanadio (Vanadilsolfato) sono considerevoli per via della bassa biodisponibilità che, però, non elimina la possibilità di intossicazione proprio a causa di queste. I possibili effetti derivanti dall’assunzione di livelli tossici di Vanadio comprendono nausea, mal di stomaco, diarrea, ipertrigliceridemia, disfunzioni epatiche, danni renali, ipoglicemia, leucopenia, ritardo dello sviluppo ed inappetenza.
La dose tossica di Vanadio è quantificata essere di 25mg al giorno. Quindi, considerando anche la bassa biodisponibilità del composto, una dose potenzialmente efficace ma che garantisca comunque un buon margine di sicurezza è di circa 100mg/die di Vanadilsolfato (20mg di Vanadio) divisa in due-tre somministrazioni prima dei pasti principali.
La supplementazione di Vanadio può comportare delle interazioni farmacologiche con Warfarin e Coumadin (difficoltà di coagulazione) o con farmaci con azione ipoglicemizzante diretta e/o indiretta come Aspirina e Exubera (riduzione eccessiva della glicemia ematica).
Quindi, nel caso si decidesse di testare l’efficacia di questo minerale, il potenziale utilizzatore dovrebbe prestare particolare attenzione a quanto appena esposto partendo dalla dose giornaliera assunta.
Se non avete letto ancora la prima, la seconda e la terza parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte – 3° Parte.
Cannella: caratteristiche e possibili applicazioni.
La Cannella è una spezia nella quale sono contenuti diversi agenti bioattivi. L’aldeide cinnamica conferisce alla Cannella il suo aroma (1), la Cumarina (una tossina) contribuisce al gusto (2) e diversi composti, tra cui il polifenolo MHCP (methylhydroxychalcone polymer), sembrano contribuire ai suoi benefici sistemici sulla sensibilizzazione all’Insulina.(3) Oltre ai tre composti unici elencati, la Cannella contiene anche tannini, flavonoidi, glicosidi, terpenoidi e antrachinoni. (4)
La Cannella sembra poter esercitare effetti di controllo benefici in contesti dietetici “pro-diabetici” attraverso diversi meccanismi.
La Cannella può inibire numerosi enzimi digestivi, come l’α-glucosidasi (4), il Saccarasi (5) e, potenzialmente, l’Amilasi Pancreatica.(5) Attraverso l’inibizione di questi enzimi, la Cannella può ridurre l’afflusso di glucosio nella circolazione sistemica ed evitare così picchi insulinici eccessivamente significativi.
MHCP
Anche nella circolazione sistemica (oltre il fegato) la Cannella sembra possedere effetti anti-diabetici. Un composto presente in essa, il prima citato polifenolo MHCP, agisce come un insulino-mimetico a livello adipocitario.(3)(6)(7) Gli effetti insulino-mimetici del MHCP sono dose-dipendenti e agiscono trasfosforizzando il Recettore dell’Insulina sulla membrana citoplasmatica (lo stesso meccanismo d’azione esplicato dalla molecola d’Insulina). I suoi effetti sull’assorbimento del glucosio e sullo stoccaggio del glicogeno, anche se dose-dipendenti, sembrano avvenire lentamente ( l’Insulina ha un tempo di risposta che si manifesta entro 10 minuti dal raggiungimento della cellula, mentre con il MHCP i tempi di risposta variano da 30 a 60 minuti, suggerendo, appunto, un ritardo temporale intracellulare).(3)
In vitro, la Cannella ha mostrato di potenziare l’azione insulinica di 20 volte.(8)
Quando ingerita da modelli umani durante gli esperimenti, la Cannella ha mostra molte potenzialità nella riduzione dei livelli di glucosio ematico (9)(10)(11) e, talvolta, dei marker del metabolismo lipidico (LDL, Trigliceridi, Colesterolo totale).(12) Ci sono anche studi nei quali è stato osservato un miglioramento dell’insulino-sensibilità con l’assunzione dell’estratto di Cannella, risposta probabilmente legata in modo indiretto alla riduzione dei livelli di glucosio nel sangue.(13)(14)
Cumarina
Come accennato in precedenza, la Cumarina è una sostanza fitochimica epatotossica e cancerogena presente in alcune piante e ad alti livelli in alcune varianti della Cannella. La Cumarina non è il composto attivo che riduce la glicemia ematica, ma è uno dei principi attivi presenti nella Cannella. Inizialmente, questa sostanza aveva un TDI (dose giornaliera tollerabile) di massimo 2mg/kg di peso corporeo, ma tale limite è stato abbassato a 0,5 e attualmente è pari a 0,1mg/kg di peso corporeo.(2) Sebbene in quest’ultima raccomandazione sia incluso un margine di sicurezza, alcuni sottogruppi della popolazione umana sono più sensibili alla tossicità della Cumarina a causa della ridotta capacità di metabolizzarla.(2)
Ciò è rilevante dal momento che la maggior parte dei benefici antidiabetici riscontrati con l’uso della Cannella sono dose-dipendenti, nell’intervallo di 300mg/kg di peso corporeo.(5) A questa dose, è particolarmente facile oltrepassare il TDI della Cumarina.
Il modo migliore per evitare di assumere dosi rilevanti di Cumarina è optare per la giusta fonte di Cannella. La Cannella di Ceylon presenta i livelli più bassi di Cumarina con meno di 190mg/kg (alcuni campioni sono al di sotto dei livelli di rilevazione) mentre la Cassia contiene tra i 700mg/kg ed i 12.230mg/Kg.(15) La Ceylon può essere riconosciuta in forma integra attraverso le sue sottili e numerose pieghe, mentre la Cassia, sempre in forma integra, ha meno pieghe e un aspetto più spesso. Non possono essere distinte in forma di polvere e la Cassia è più frequentemente utilizzata nella realizzazione di prodotti alimentari per via della sua elevata disponibilità e del basso costo.(2)(16)
Tramite i numeri di cui sopra, un essere umano di 90Kg può ingerire 47,8g di Cannella di Ceylon e arrivare, nella peggiore delle ipotesi, ad assumere 0,1 mg/kg di peso corporeo del TDI della Cumarina (assumendo, quindi, la punta massima di assunzione della Cumarina). Diversamente, utilizzando la Cannella Cassia può facilmente essere ingerito un dosaggio di Cumarina al di sopra del TDI con un apporto decisamente inferiore di prodotto.
L’assorbimento della Cumarina non sembra dipendere dalla forma di Cannella ingerita. Livelli serici simili e livelli escreti sono stati raggiunti con l’assunzione di cumarina isolata, di pillole contenenti cannella, tè e budino di riso (cibo solido).(17) Questi risultati sono stati standardizzati alla dose X di Cumarina, quindi la fonte di Cannella è irrilevante.
Lo studio di cui sopra, tuttavia, ha notato un tasso di estrazione del 38,5% di Cumarina dalla polvere al liquido una volta entrati in contatto (appena bollente per 30 minuti); suggerendo che si può ribaltare la bilancia a favore dei polifenoli e del MHCP solubili in acqua rispetto alla Cumarina se la Cannella viene messa in acqua e servita nel tè o usando l’acqua di infusione per mescolare frullati proteici, poiché i componenti idrosolubili hanno un tasso di estrazione molto più alto.
Come spesso accade, però, sono stati gli studi iniziali svolti sulla Cannella ha mostrare una efficacia marcata dei composti ivi contenuti.(12)(18) Ma, nonostante le possibili e promettenti potenzialità rilevate nelle prime ricerche, al momento, l’utilizzo della Cannella ha mostrato soltanto una riduzione della glicemia ematica in acuto con ridotti e ben poco rilevanti risultati in cronico, come la mancata influenza sulla emoglobina glicata (HbA1c) o su un reale miglioramento della insulino resistenza.
I meccanismi attraverso i quali l’assunzione di Cannella porta ad un abbassamento della glicemia post-prandiale in modo dose dipendente sono quindi legati al rallentamento dello svuotamento gastrico, all’inibizione di numerosi enzimi digestivi tra cui i prima citati alfa-glicosidasi, maltasi , sucrasi e, anche, da una sorta di azione “carb-blocker” data dalla possibile inibizione del enzima amilasi pancreatico.
Come già accennato, i risultati più importanti riscontrati sono legati per lo più ad una riduzione della risposta glicemica in acuto dopo trenta minuti dal pasto senza modifiche profonde degli altri marker del controllo glicemico. Nonostante ciò, non sono da escludere possibili vantaggi di questa natura con un assunzione di Cannella sul lungo periodo, sebbene, come ormai risaputo, l’Indice Glicemico non è un parametro rilevante sulla qualità del dimagrimento e, quindi, della composizione corporea.
Acarbosio
Anche nei soggetti diabetici, l’uso in monoterapia della cannella è risultato fallimentare mostrando semplicemente una validità additiva in concomitanza con l’assunzione di farmaci ipoglicemizzanti. Ancora una volta, tale risultato potrebbe trovare un nesso causale nella possibile attività “carb-blocker” simile a quella osservata in seguito ad assunzione di Acarbosio, farmaco utilizzato nel trattamento del diabete di tipo II e dell’obesità avente azione ritardante sulla digestione e l’assorbimento di zuccheri alimentari in quanto è un inibitore dell’alfa-glucosidasi intestinale e, attraverso tale meccanismo, ha un effetto nel ridurre la glicemia post-prandiale. Riduce inoltre i livelli di trigliceridi, di emoglobina glicata (HbA1) e la resistenza periferica all’insulina.(19)
Nei soggetti sani, quindi, sembrerebbe maggiormente plausibile che la riduzione della glicemia ematica dopo il pasto sia da attribuirsi quasi totalmente all’inibizione della digestione e assorbimento dei carboidrati ma non ad un effettivo miglioramento dell’insulino sensibilità.
Alfa Amilasi
La questione prima trattata sulla lenta risposta ipoglicemizzante osservata in seguito all’assunzione di Cannella è oggetto di dibattito per quanto riguarda il suo meccanismo. Sembrerebbe, infatti, che tale effetto sia dovuto quasi esclusivamente all’inibizione dell’alfa amilasi e, quindi, ad una rallentata assimilazione dei carboidrati ingeriti, e non dal azione insulino-mimetica del MHCP. Se così fosse, questo tipo di risultati sarebbero ottenibili con una semplice ridistribuzione dei macronutrienti e, in particolare, con una riduzione del carico glucidico del singolo pasto.
A questo punto, dopo aver letto le informazioni presenti in questo articolo, si potrebbe giungere facilmente alla conclusione secondo cui l’uso della Cannella sia relegabile al solo insaporire cibi o bevande, vista la limitatezza della sua azione sul metabolismo glucidico. Nonostante ciò, la sua possibile applicazione come GDA non è completamente da escludere, sebbene con forti limitazioni legate alle sue caratteristiche e a quelle del soggetto al quale viene somministrata a tal fine.
L’uso della Cannella potrebbe apportare dei vantaggi, anche se minimi, se abbinata ad altri GDA (vedi Berberina, ALA e Cromo) in soggetti con una marcata insulino resistenza, sia di base genetica e/o “situazionale” (legata ad una condizione di sovrappeso/obesità e/o per via di una alimentazione ipercalorica/iperglucidica). Tali soggetti, avendo difficoltà nella gestione di carichi glicemici (nel singolo pasto) anche di entità moderata, possono sfruttare l’effetto di riduzione/rallentamento dell’assorbimento glucidico post prandiale dato dalla Cannella, così da avere una migliore gestione del picco glicemico, gestione ulteriormente migliorata dagli altri GDA co-assunti e da alcune accortezze alimentari come la scelta della fonte glucidica. 1g di Cannella di Ceylon appena prima del pasto sembra essere un dosaggio sufficiente ad esplicare tali effetti.
I risultati addizionali non saranno particolarmente evidenti ma, nei casi sopra citati, l impatto potrebbe facilitare la gestione del carico glucidico.
Ovviamente, soggetti nella norma in salute possono benissimo evitare di usare la Cannella con tali finalità e limitarsi, secondo palato, al suo uso come semplice spezia.
Un basso dosaggio di Witaferina A, un lattone steroideo presente nella pianta indiana Ashwagandha, potrebbe facilitare la perdita di peso nei soggetti sovrappeso o obesi. I ricercatori della Harvard Medical School hanno ipotizzato ciò in seguito allo svolgimento di uno studio effettuato su topi resi grassi i quali, in seguito alla somministrazione di Witaferina A, avevano perso ¼ del loro peso in tre settimane.(1)
Per lo svolgimento dell’esperimento, i ricercatori hanno usato topi normopeso fatti ingrassare attraverso una dieta ipercalorica.
I ricercatori hanno somministrato la Witaferina A direttamente nell’intestino tenue di una parte degli animali presi in esame. Se al posto dei topi ci fossero stati soggetti umani di 100Kg, la dose somministrata di Witaferina A (in caps) sarebbe stata di circa 25mg/die.
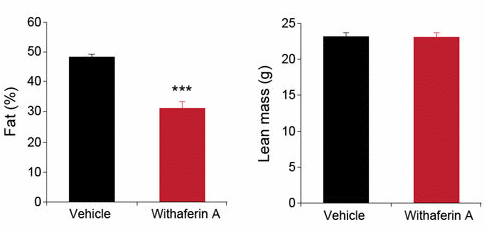
La somministrazione di Witaferina A ha portato ad una riduzione del peso corporeo dei topi [in basso a sinistra]. Ciò era dovuto principalmente al fatto che la Witaferina A induceva gli animali trattati a consumare meno cibo [in basso a destra].
La Witaferina A ha ridotto la massa grassa. Sebbene i topi del gruppo trattato con il lattone steroideo abbiano perso peso, la loro massa magra è rimasta praticamente intatta.
Gli animali trattati con Witaferina A mostravano una normalizzazione dei livelli serici di Leptina.
I ricercatori hanno ripetuto l’esperimento utilizzando topi magri con un normale livello di Leptina. In essi, la Witaferina A non ha avuto alcun effetto sul peso e sulla composizione corporea. I ricercatori hanno ripetuto l’esperimento anche con i topi ob/ob che non producono Leptina a causa di un difetto genetico, e con topi db/db con un difetto del recettore della Leptina. In entrambi i casi gli animali non avevano un controllo sul consumo di cibo e, di conseguenza, avevano subito un marcato aumento della massa grassa. Il trattamento con la Witaferina A in questi animali non ha mostrato quasi alcun effetto.
La sintesi di Leptina aumenta in risposta ad un aumento delle riserve adipose o, più precisamente, ad un aumento del metabolismo glucidico adipocitario. Questo effetto riduce l’appetito. Quando le riserve adipose diminuiscono, la sintesi di Leptina diminuisce e l’appetito aumenta. Tuttavia, se si consuma un eccesso calorico nel lungo termine, questo meccanismo viene alterato. Si sviluppa resistenza alla Leptina. Si ipotizza, quindi, che la supplementazione di Witaferina A aumenti la sensibilità alla Leptina.
I ricercatori scrivono che, sono passati più di due decenni dalla scoperta storica della Leptina da parte di Friedman e colleghi (2), ma finora non è stato sviluppato un trattamento leptino-centrico applicabile per il trattamento dell’obesità. Subito dopo le prime pubblicazioni sulla Leptina, è stato suggerito che l’obesità sia una condizione correlata alla leptino-resistenza.
Nel corso degli ultimi vent’anni, le speranze per lo sviluppo di un trattamento orientato verso la manipolazione/attività della Leptina nell’obesità sono progressivamente diminuite, poiché molti tentativi di ri-sensibilizzare il cervello degli individui obesi al peptide erano falliti. Questi sforzi infruttuosi per aumentare la sensibilità alla Leptina e trattare lo stato iperleptinemico nell’obesità per trattare tale condizione hanno anche contribuito in modo sostanziale al dibattito sulla presenza o meno della leptino-resistenza.
Il trattamento di topi obesi e iperleptinemici con Witaferina A ha portato ad una forte riduzione dell’assunzione di cibo e del peso corporeo. Entrambi le risposte osservate sono legate ai livelli di Leptina serica. Poiché i livelli di Leptina diminuiscono gradualmente durante il periodo di trattamento, parallelamente alla stabilizzazione del peso e della percentuale di massa grassa degli animali trattati, anche l’effetto della Witaferina A diminuisce gradualmente. Inoltre, nei topi magri, con bassi livelli di Leptina circolante, non sono stati osservati cambiamenti nel consumo di cibo o nel peso corporeo.
Gli estratti di Withania somnifera, che contengono anche Witaferina A, sono stati usati dall’uomo per secoli, e sembrano possedere un effetto fortemente conservativo sul sistema della Leptina nei mammiferi tra cui topi e umani. Considerando tutte queste informazioni, i ricercatori ritengono che l’uso della Witaferina A per il trattamento dell’obesità negli esseri umani sia una grande promessa per il futuro.
Se non avete letto ancora la prima e la seconda parte di questa serie di articoli vi invito a farlo: 1° Parte – 2° Parte.
Cromo: caratteristiche e possibili applicazioni.
Cromo Picolinato
Il Cromo è un minerale essenziale nella dieta umana ed è comunemente utilizzato come integratore alimentare (es. Picolinato o Polinicotinato) per migliorare la sensibilità all’insulina nei soggetti sani o nei soggetti diabetici. (1)
Il Cromo può essere trovato nel:
Colostro bovino (sotto forma di un oligopeptide di cromodulina ricco di zinco, con un atomo di Cromo per quattro amminoacidi (2) (3)) che fornisce 220mcg di Cromo per 1.035g di proteine (193ng/g di proteine) (2)
• Latte scremato, ad una concentrazione di 252mcg di Cromo per 1.172g di proteine (215ng/g di proteine) (2)
Il Cromo è sia un minerale dietetico che un elemento (Cr) con più valenze. La forma completamente ossidata di Cromo (Cr (VI)), che è esavalente (+6 stato di ossidazione), è altamente tossica e impiegata in una varietà di applicazioni industriali.(4) Dato l’alto grado di tossicità, il Cromo esavalente non viene mai usato come integratore. Le forme supplementari di Cromo comprendono il bivalente (Cr (II)) o il trivalente (Cr (III)), quest’ultima è la forma più stabile.(1)
Il quantitativo di Cromo assunto con la dieta dovrebbe essere almeno di 0,005-0,2mg(5-20mcg) al giorno al fine di prevenirne il deficit, e l’assunzione giornaliera raccomandata è di 21-25mcg per le donne e di 25-35mcg per gli uomini con la fascia di età tra i 18 ed i 45 anni che richiede quantità verso il punto più alto dell’intervallo riportato.(5) Le donne di tutte le età che stanno allattando richiedono un’assunzione giornaliera di Cromo pari a 45mcg.(5) La dose raccomandata per i bambini da 1 a 3 anni è 11mcg/die mentre dai 4 agli 8 anni il dosaggio sale a 15mcg/die.(5)
Le concentrazioni standard di Cromo circolante in uno stato non carente sono state misurate nell’intervallo di 2,8-45mcg/L nel sangue intero e 0,12-2,1mcg/L nel siero.(6)
Una carenza di Cromo può essere indotta con una nutrizione parenterale totale a lungo termine (TPN) priva del minerale, e può essere invertita con una supplementazione di 150mcg di Cromo al giorno aggiunti al TPN come riscontrato attraverso un caso studio (7) e 250mcg al giorno per 2 settimane seguite da una dose di mantenimento pari a 20μg al giorno per 18 mesi in un altro.(8) I principali sintomi da carenza di Cromo in questi particolari casi si manifestavano attraverso un compromessa tolleranza al glucosio e una riduzione dell’insulino-sensibilità associata alla perdita di peso, così come la neuropatia e l’encefalopatia che erano reversibili con il reintegro del minerale. (8)(7)
Quindi, una grave carenza di Cromo è associata a sintomi simili a quelli riscontrati nel diabete di tipo I (alterata tolleranza al glucosio e perdita di peso) e nella neuropatia, e può essere invertita con la somministrazione del minerale.
Le carenze subcliniche di Cromo sono associate all’insulino resistenza, poiché le concentrazioni di questo minerale sono risultate inferiori nei diabetici rispetto ai soggetti di controllo (9) (tuttavia, l’evidenza è eterogenea per il diabete gestazionale (10)(11)). Le diete con un assunzione cronica di zuccheri (35% delle calorie giornaliere) sono state associate ad una accelerata perdita di Cromo attraverso le urine (Cromo urinario) (12) sebbene le diete composte da cibi ad alto indice glicemico non abbiano influenzato in modo significativo l’eliminazione del Cromo attraverso le urine in soggetti sani, pur mostrando una tendenza nell’arco di sei giorni.(13)
Cromodulina
Si ritiene che questa perdita accelerata di Cromo attraverso le urine si verifichi per via del rilascio di Cromodulina (LMWCr; Low-molecular-weight chromium-binding substance) nel flusso ematico da parte delle cellule insulino-sensibili, con conseguente eliminazione urinaria.(14) La Cromodulina è un peptide che esiste all’interno delle cellule. Quando combinato con il Cromo immesso nelle cellule dal flusso sanguigno, amplifica la segnalazione dell’insulina legandosi ai recettori insulinici stimolati dall’ormone.(14) La Cromodulina lega lo ione cromo ad altissima affinità, formando un complesso che può essere separato solo in condizioni non fisiologiche. Una volta che i livelli di insulina scendono, tuttavia, i recettori dell’insulina non hanno più bisogno di essere sensibilizzati, quindi l’intero complesso deve essere eliminato nel suo insieme.(14) (15)Questa ipotesi è supportata dal rilevamento della Cromodulina nelle urine (16) e dalla sua stretta correlazione con i tassi di secrezione dell’Insulina e l’esposizione in condizioni non complementari.(16) (17) (18)
Le concentrazioni urinarie di Cromo risultano elevate in seguito ad allenamenti di resistenza (con un aumento di cinque volte dopo due ore di corsa, ma con solo un aumento di due volte nel corso delle ventiquattro ore) in un modo che non è correlato ad un aumento dell’insulina serica o ad un aumento di qualsiasi altro ione urinario.(18) Questa condizione, nonostante l’assenza di significativi livelli di Insulina, è nota per richiedere un maggiore assorbimento di glucosio nel tessuto muscolare sostenuto da un maggiore rilascio di glucosio da parte del fegato.(19)
Il Cromo trivalente (che si trova negli integratori) sembra avere effetti tossici a concentrazioni superiori a 20mcg/mL nel siero o nelle cellule; questa tossicità è associata al danno ossidativo al DNA.(20) Questo è lo stesso meccanismo mediante il quale il cromo esavalente esprime la sua tossicità, con l’unica differenza che quest’ultimo è tossico a concentrazioni molto più basse (21), in particolare dopo inalazione durante un impiego che comporta la sua manipolazione. (22) (4)
Con il termine Cromo Picolinato ci si riferisce al Cromo nello stato trivalente (Cr (III)) il quale è legato a tre molecole di acido picolinico, un analogo strutturale della Niacina. Questa forma di Cromo è altamente stabile (23), a parte una possibile degradazione indotta dall’acido, che rimuove una molecola di picolinato e porta a due ioni di cromo che si legano insieme. (24) I ligandi picolinati sono in una posizione tale che il Cr (III) può essere ridotto in Cr (II) nella coltura cellulare senza perdere il picolinato (25), una proprietà che sembra essere unica per il picolinato rispetto ad altre forme supplementari (Cloruro e Nicotinato) e si pensa che sia alla base delle possibili proprietà cancerogene indotte da alte concentrazioni.(26)
Si ritiene che il Cromo Picolinato sia fisiologicamente inattivo fino ad avvenuta liberazione della molecola di Cromo (26), suggerendo che esso funga da “pro farmaco” al Cromo.
Il Cromo è noto per essere presente nel lievito, dove svolge un ruolo fisiologico importante.(27) (28) All’interno delle cellule del lievito si trova il “Fattore di Tolleranza al Glucosio” (GTF) (29), che è stato inizialmente derivato dal lievito di birra.(30) Il GTF può essere purificato dai lieviti dopo l’estrazione metanolica e la successiva filtrazione, ottenendo un insieme di molecole di dimensioni variabili da 1.000 a 3.500 Da. (31)(32) I principali componenti attivi in questo set di molecole sono considerati l’acido trivalente al cromo nicotinico insieme ad alcuni aminoacidi (Glicina, L-cisteina e Acido Glutammico).(33) L’apporto alimentare del lievito sembra conferire alcuni dei benefici dati dall’integrazione di Cromo, probabilmente a causa dell’ingestione di GTF e Cromo.(29)
Si ritiene che il Cromo presente nel lievito sia acido cromo-nicotinico, sebbene possano esistere altre forme di Cromo nel lievito che non sono state ancora rilevate.
Cromo Polinicotinato
L’Acido Nicotinico di Cromo (noto anche come Cromo Polinicotinato), forma altamente assimilabile di Cromo, è composto da Cromo legato all’Acido Nicotinico (Niacina o Vitamina B3) e si dice che abbia effetti sulla riduzione del Colesterolo.(34)(35) Negli studi in cui il Colesterolo è stato ridotto in seguito all’assunzione di Cromo Polinicotinato, non è stato trovano necessariamente un nesso benefico dato dal miglioramento del metabolismo glucidico (34)(35), suggerendo che è la Niacina a causare questi effetti.
Il Cromo Dinicocisteinato (CDNC) è un complesso dello ione Cromo con l’aminoacido L-cisteina. Uno studio che ha confrontato l’effetto di 400mcg di CDNC con 400mcg di Cromo Picolinato ha rilevato miglioramenti nei livelli di Insulina e della sensibilità a questa solo con il CDNC. (36)
Uno dei principali meccanismi che si ritiene correlato all’integrazione con Cromo comporta la modulazione della via di segnalazione dell’Insulina.(37) [38] Questo è stato scoperto per la prima volta quando è stato identificato un oligopeptide legante il Cromo a basso peso molecolare che ha aumentato gli effetti dell’Insulina e l’ossidazione del glucosio. (38)[39] Chiamato anche LMCr o Cromodulina (39), questo oligopeptide viene sintetizzato nel fegato dei ratti dopo iniezioni di Cromo (40) e ha una massa di circa 1500 kDa. (39)(41)
È stato rilevato che la Cromodulina ha aumentato la segnalazione di Insulina in presenza di un livello di quest’ultima pari a 5-8 volte superiore rispetto all’attività basale, senza influenzarne la segnalazione in assenza di Insulina.(42) La deplezione di Cromo da parte della Cromodulina ne blocca l’attività (42) che si correla positivamente con il contenuto del minerale nel peptide. Inoltre, altri minerali non sono riusciti a replicarne gli effetti. (43)
Recettore dell’Insulina
In definitiva, la Cromodulina aumenta l’autofosforilazione del Recettore dell’Insulina. La segnalazione del Recettore dell’Insulina richiede che l’Insulina o un mimetico (qualcosa che si comporti come l’Insulina) si leghi alla subunità α extracellulare del recettore (44) che consente alla subunità β intracellulare di essere autofosforilata. (44) La Cromodulina sembra agire intracellularmente nella subunità β del Recettore dell’Insulina.(14)
Le funzioni cromo-dipendenti della Cromodulina sono probabilmente la ragione biologica per la quale il Cromo è un minerale essenziale (45), sebbene la natura essenziale del Cromo sia stata recentemente contestata.(46)
L’Adenosina Monofosfato Chinasi (AMPK) è un sensore chiave dello stato energetico cellulare, il quale monitora costantemente i livelli di ATP al fine di mantenere l’omeostasi metabolica. L’AMPK si attiva durante gli stati di carenza energetica (caratterizzato da un aumento della AMP:ATP ratio) dove coordina il metabolismo degli acidi grassi e del glucosio in modo anti-obesità e anti-diabetico.(47) Quando attivato, l’AMPK sopprime le vie anaboliche come la sintesi proteica, di trigliceridi e di acidi grassi attivando contemporaneamente percorsi catabolici come la glicolisi e l’ossidazione degli acidi grassi per aumentare la produzione di ATP. (48)
È stato notato che il Cromo (trivalente con D-fenilalanina) attiva l’AMPK nel suo sito catalitico (Thr172) nei cardiomiociti e nelle cellule muscolo-scheletriche a 25μM, suggerendo che i complessi organici del Cromo possono essere nuovi attivatori della via dell’AMPK.(49)
L’assorbimento del Cromo alimentare è inversamente correlato all’assunzione, variando dallo 0,4% al 2,0%, con l’assorbimento più efficiente (2%) a un apporto dietetico inferiore di circa 10mcg negli uomini adulti.(50) Questo diminuisce a circa lo 0,5% quando l’assunzione con il cibo raggiunge i 40mcg che sembra essere il limite, dato che l’assunzione di Cromo nel range di 40-240mcg ha un assorbimento di circa lo 0,4%. (50) (51)
Acido Fitico
L’assorbimento del Cromo è influenzato da una serie di fattori dietetici. Nei ratti, l’assorbimento del Cromo sembra essere ostacolato dalla coingestione dei fitati, che impedisce il trasporto e l’assorbimento attraverso l’intestino. (52) È stato dimostrato che il deficit di Zinco aumenta l’assorbimento del Cromo, che è aumentato nei ratti carenti di Zinco e ridotto dallo Zinco supplementare (53), suggerendo che questi due minerali possono competere per l’assorbimento. L’assorbimento del Cromo nei ratti è anche potenziato dall’ossalato, un acido organico presente in molte verdure e cereali. (52) Sebbene sia informativo, occorre prestare attenzione quando si estrapolano i risultati dagli studi sui ratti rapportandoli all’uomo, poiché studi recenti hanno rilevato che l’assorbimento di Cromo alimentare nell’uomo è significativamente maggiore rispetto a quanto avviene nei ratti per numerosi complessi di cromo testati.(54)[55]
Gli amminoacidi sembrano migliorare l’assorbimento del Cromo alimentare poiché formano complessi che migliorano l’assorbimento riducendo la tendenza del Cromo a precipitare nel liquido intestinale alcalino.(1) L’assorbimento del Cromo negli esseri umani è anche significativamente aumentato in presenza di Acido Ascorbico e Acido Nicotinico.(1)
Nei diabetici di tipo II, un integrazione giornaliera di Cromo (come cromo Picolinato) pari a 1.000mcg è risultata sufficiente a portare i livelli del minerale a digiuno nel siero da 2,40 ± 0,19 vs 0,16 ± 0,05ng/dL al basale dopo 12 settimane e 2,62 ± 0,09ng/ dL vs 0,17 +/- 0,04ng /dL al basale dopo l’integrazione di 24 settimane.(55)
Transferrina
La transferrina è una proteina di trasporto del siero nota per legarsi ai minerali (in particolare il Ferro). È stato notato che presenta affinità per il Cromo trivalente.(56) Per ogni molecola di transferrina si legano due ioni di cromo. (57)(58) Si pensa che la transferrina doni il Cromo all’oligopeptide Cromodulina.(59) Anche se studi precedenti hanno suggerito che la Cromodulina dona il Cromo alla transferrina. Questo lavoro però è stato condotto a temperature più elevate, che potrebbero aver causato la degradazione della Cromodulina.(60) Tuttavia, studi più recenti hanno dimostrato che la Cromodulina non rilascia il Cromo alla transferrina. (59) Poiché la transferrina rilascia ioni all’interno di una cellula dopo l’endocitosi (61), sembra che la Cromodulina accetti e trattenga questi ioni dalla transferrina.
La supplementazione con Cromo determina un aumento dell’eliminazione urinaria del minerale.(55)
Cromo Cloride
Uno studio svolto su ratti ha osservato che i livelli tossici di Cromo (100mcg/kg assunto con il cibo) sembrano bioaccumularsi di più con il Cromo Cloruro rispetto al Cromo Picolinato, in parte dipendente da un più alto tasso di escrezione osservato con il Picolinato.(62) Ciò è stato ipotizzato essere dovuto all’Acido Picolinico, che è stato osservato aumentare l’eliminazione di minerali come lo Zinco.(63)
Diversi studi hanno suggerito che la supplementazione con Cromo può promuovere una riduzione dell’appetito, con conseguente diminuzione del consumo di cibo, sia negli animali che negli esseri umani. Una recente meta-analisi di 10 studi randomizzati, in doppio cieco, controllati con placebo ha concluso che il Cromo Picolinato ha un effetto sulla riduzione del peso relativamente modesto, ma significativo rispetto al placebo (64), suggerendo un possibile effetto sulla soppressione dell’appetito. I meccanismi associati all’effetto anoressizzante dato dall’uso del Cromo sono attualmente sconosciuti, sebbene sia stato ipotizzato che si verifichi attraverso l’azione di specifici neurotrasmettitori nel cervello deputati al controllano l’appetito e il comportamento alimentare. (65)(66)
Ciò è stato confermato in un recente studio condotto su donne in sovrappeso adulte che hanno riportato voglie di carboidrati intense (almeno due volte a settimana). La supplementazione giornaliera con 1.000mcg di Cromo (come Picolinato) nel corso di otto settimane ha comportato una maggiore riduzione dell’assunzione di cibo (25%) rispetto al placebo (8%).(67) La riduzione dell’assunzione di cibo era associata ad una diminuzione della fame e dell’appetito, tuttavia la composizione dei macronutrienti non era influenzata e questi cambiamenti erano indipendenti da qualsiasi effetto sulla sensibilità all’insulina.(67) In uno studio parallelo condotto dallo stesso gruppo di ricerca, è stato riscontrato che la somministrazione periferica di Cromo nei ratti (tramite iniezione IP) ha comportato solo una modesta diminuzione dell’assunzione di cibo, rispetto a una significativa riduzione dose-dipendente dell’assunzione di cibo quando somministrato a livello centrale (direttamente nel cervello). (67) Nel suo insieme, questo lavoro suggerisce che, come detto pocanzi, la supplementazione con Cromo può promuovere una riduzione dell’apporto di cibo attraverso l’azione di neurotrasmettitori nel cervello che controllano l’appetito e il comportamento alimentare.
Nei pazienti con depressione atipica (che è un particolare sottogruppo di depressione associato a maggiore assunzione di cibo, sonnolenza e reattività dell’umore (68)), 600mcg di Cromo Picolinato per otto settimane non hanno influenzato significativamente la maggior parte dei sintomi depressivi. Tuttavia, ci sono stati significativi miglioramenti nella voglia di carboidrati e nell’assunzione di cibo con un effetto maggiore in coloro i quali il desiderio di carboidrati era maggiore al basale. (66) Nelle persone con disturbo da alimentazione incontrollata, il tasso di declino della frequenza di binging era maggiore con 1.000mcg di Cromo rispetto al placebo e 600mcg, sebbene la riduzione complessiva non abbia raggiunto la significatività statistica.(69)
La supplementazione con 1.000mcg di Cromo (come Picolinato) in due dosi suddivise per 24 settimane nei diabetici di tipo II non ha influenzato significativamente la gluconeogenesi epatica rispetto al placebo. (55) (La gluconeogenesi epatica è spesso patologicamente elevata nei diabetici (70)).
Quando i diabetici consumavano 200mcg di Cromo (come Cloruro) al giorno aggiunto a un prodotto di latte in polvere per 16 settimane, i livelli di glucosio e di insulina nel sangue erano significativamente ridotti mentre la sensibilità all’insulina migliorava. (71) I risultati di questo studio erano tuttavia specifici per genere, in quanto miglioramenti significativi nei suddetti marker dell’omeostasi del glucosio si sono verificati solo in soggetti di sesso maschile. (71)
Cromato di Potassio
Come detto in precedenza, la Cromodulina è un oligopeptide endogeno (41) contenente Cromo che media positivamente la segnalazione del recettore dell’insulina in presenza di Insulina.(40) L’iniezione di Cromo (come Cromato di Potassio) nei ratti aumenta le concentrazioni urinarie e fecali di questo oligopeptide. (72) Tuttavia, la Cromodulina urinaria non sembra essere saturata in condizioni basali, il che implica che più Cromo potrebbe essere legato all’oligopeptide.(72) Poiché la potenza della Cromodulina nel potenziare la segnalazione dell’Insulina è correlata con la quantità di Cromo legata ad esso (43), e le iniezioni di Cromato di Potassio nei ratti determinano una rapida associazione con la Cromodulina (60)(73), è possibile che l’aumento di Cromo alimentare possa aumentare l’attività di questo oligopeptide.
Operando partendo dal presupposto che le assunzioni tipiche di Cromo nella dieta sono insufficienti per saturare la Cromodulina, la supplementazione con Cromo potrebbe teoricamente migliorare la segnalazione dell’Insulina tramite l’aumento del legame cromo-cromodulina.
Negli studi nei quali è stato utilizzato il Cromo trivalente, sembra esserci un aumento dell’attività della chinasi del recettore insulinico (in presenza di Insulina) quando il cromo 1-10μM viene aggiunto alla coltura di cellule di mammifero.(74) Questo aumento è indipendente da qualsiasi influenza diretta sulla fosforilazione o autofosforilazione (74) e distinta da quella della Cromodulina, che influenza l’autofosforilazione. (42)
Cromo Propionato
Alcuni complessi con Cromo trivalente hanno interazioni minori con il Recettore dell’Insulina, con complessi legati a piccole molecole endogene come Istidinato, Lattato, Acetato o Propionato che mostrano effetti inibitori minori a concentrazioni intorno a 100μM. Di questi complessi, il Cromo Propionato sembra essere il più potente, mostrando effetti inibitori a concentrazioni fino a 1μM. (75)
Lo stesso Cromo è stato implicato nel potenziare la segnalazione dell’Insulina, sebbene il meccanismo con gli ioni Cromo sembra differire da quello osservato con la Cromodulina e richiede una concentrazione significativamente più alta. Lo stesso Cromo non sembra influenzare direttamente il Recettore dell’Insulina come la lattina di Cromodulina.
La Fosfo-tirosin–fosfatasi 1B (PTP1B) è un regolatore negativo del segnale del Recettore dell’Insulina (76) che può essere soppresso dal Cromo endogeno. Anche se la Cromodulina è stata osservata promuovere l’attività della PTP della membrana in uno studio precedente (77), ci sono molti enzimi PTP endogeni e il PTP1B non è stato specificamente esaminato in questo studio. È stato dimostrato che il Cromo trivalente inibisce il PTP1B del 21-33% nelle cellule di epatoma umano e di ratto (78), suggerendo che il Cromo può potenziare la segnalazione dell’Insulina sopprimendo la defosforilazione mediata dal PTB1B nel Recettore dell’Insulina. Al contrario, uno studio più recente ha osservato che il Cromo non è riuscito a inibire l’attività della fosfatasi PTP1B umana ricombinante in un sistema in vitro puro, suggerendo che il Cromo può potenziare la segnalazione dell’Insulina da meccanismi distinti da qualsiasi effetto sul PTP1B. (74)
In uno studio in vivo, ratti obesi diabetici trattati con 80mcg/kg di Cromo (come Picolinato) hanno subito una diminuzione complessiva dell’attività della PTP1B e dell’espressione proteica correlata ad un aumento della segnalazione dell’Insulina nel muscolo scheletrico.(79) Questa diminuzione non è stata osservata nei ratti magri ai quali è stato somministrato il Cromo alla stessa dose.(79)
IRS-1
La fosforilazione dell’IRS-1, un importante trasduttore della segnalazione dell’Insulina che è inibito dalla fosforilazione a Serine307 (80), non è influenzato dal Cromo a 10μM in varie forme trivalenti.(75) Inoltre, l’espressione della proteina IRS è rimasta inalterata con una supplementazione di Cromo fino a 80mcg /kg nei ratti.(79) In assenza di Insulina, tuttavia, la segnalazione basale del IRS-1 è leggermente aumentata a 10 μM di Cromo, che si pensa sia dovuta alla diminuzione della fosforilazione del Serine307 (75) dal Jun NH (2) -terminal kinase (JNK). (37) Il JNK regola negativamente la segnalazione del IRS tramite fosforilazione a Serine307 (80) (81), che è aumentata nei topi obesi, (82) (83) causando insulino-resistenza. In particolare, l’attenuazione mediata dal JNK della segnalazione dell’Insulina nei ratti obesi è soppressa dal Cromo. (82)(83)
L’attivazione del JNK sopra riportata potrebbe essere ricondotta teoricamente allo stress del reticolo endoplasmatico (ER) (84), e gli agenti che riducono lo stress del ER attenuano anche i sintomi diabetici. (85)(86) È noto che lo stress del ER aumenta nelle cellule degli animali obesi e diabetici ed è curabile con il Cromo.(82)
Per riassumere i concetti esposti, sappiamo che l’interazioni del Cromo con il PTP1B, un regolatore negativo dell’attività del Recettore dell’Insulina, non sono ben compresi. Alcuni studi suggeriscono che il Cromo potrebbe non avere effetti apprezzabili sulla segnalazione del PTP1B. È possibile, tuttavia, che il Cromo sopprima l’attenuazione JNK-mediata della segnalazione dell’Insulina nel contesto di uno stato di insulino-resistenza preesistente.
Il Cromo non sembra aumentare l’espressione del Recettore dell’Insulina in presenza o assenza di Insulina, suggerendo che i suoi effetti sulla segnalazione dell’Insulina avvengono indipendentemente da eventuali cambiamenti nei livelli dei recettori insulinici. (87) (75) Inoltre, quando incubato con Insulina, il Cromo non influenza l’interazione dell’Insulina con il suo recettore.(74) Ciò suggerisce che il Cromo non influisce sulla sensibilità all’Insulina aumentando l’affinità del Recettore dell’Insulina.
Uno studio preliminare condotto nel 1992 ha rivelato che il Cromo aumenta l’internalizzazione dell’Insulina a 1μM (418ng/ml), effetto associato ad una maggiore fluidità della membrana e non replicato con altre chelazioni di Cromo o Zinco Picolinato. (88) La scoperta che l’Insulina è internalizzata nella cellula è stata successivamente rivelata come un importante meccanismo di feedback negativo per la segnalazione del Recettore dell’Insulina. Dopo che l’Insulina si lega con il suo recettore, il complesso del recettore insulinico viene internalizzato dall’endocitosi (89), innescando la degradazione dell’Insulina (90) e riducendo efficacemente il numero di recettori dell’insulina presenti sulla superficie cellulare come meccanismo per attenuare la risposta insulinica.(91)
In breve, dopo avvenuto legame con il suo recettore sulla superficie della cellula, l’Insulina innesca il movimento del complesso del Recettore dell’Insulina all’interno della cellula. Questo riduce il numero di recettori insulinici presenti sulla superficie cellulare e funziona come un meccanismo di feedback negativo per limitare la risposta della segnalazione insulinica.
In risposta a un test orale di tolleranza al glucosio, una supplementazione di 200mcg di Cromo per otto settimane non ha aumentato la risposta all’Insulina in soggetti diabetici di tipo II quando misurata dopo 10 minuti (71) mentre ad un dosaggio di 1.000mcg (come Picolinato) in soggetti non diabetici con sindrome metabolica per oltre 16 settimane ha aumentato la risposta all’Insulina nonostante non sia stato rilevato altro cambiamento nei biomarcatori del diabete. (92)
Uno studio ha osservato che, nonostante l’incapacità di trovare miglioramenti statisticamente significativi nella sensibilità all’Insulina per l’intero gruppo di soggetti presi in esame, il 46% degli individui che avevano un grado di insulino resistenza più elevato presentavano un miglioramento della sensibilità all’Insulina del 10%. (55) In particolare, non vi era alcuna differenza nell’assorbimento o cinetica del Cromo tra responder e non responder (55), suggerendo che la supplementazione con Cromo può aumentare la sensibilità all’Insulina in soggetti con insulino resistenza.
Inoltre, è stato osservato che una supplementazione di 1.000mcg di Cromo (come Picolinato) per 24 settimane in soggetti con diabete di tipo II riduce leggermente le concentrazioni di lipidi intramuscolari rispetto al placebo. (55) Poiché l’accumulo cronico di lipidi nel tessuto muscolare è una delle numerose cause patologiche dell’insulino-resistenza (93), anche questo lavoro suggerisce che la supplementazione con Cromo può aumentare la sensibilità all’Insulina in coloro che sono già insulino-resistenti.
Una meta-analisi di studi condotti su diabetici di tipo II trattati con >250mcg di Cromo per un periodo superiore ai tre mesi non ha rilevato alcuna influenza sul HbA1c rispetto al trattamento con placebo. (94) Ciò è in contrasto con precedenti revisioni che valutato solamente studi condotti su diabetici con un HbA1c basale superiore al 7%, in cui la supplementazione con Cromo ha determinato una riduzione dell’HbA1c dello 0,34% rispetto al placebo. (95) Altre revisioni hanno rilevato riduzioni dello 0,6% (96), e fino allo 0,9% quando sono state incluse tutte le forme di diabete e gradi di insulino resistenza.(97) Va notato, tuttavia, che alcune di queste analisi comprendevano prove della durata inferiore a tre mesi (96), che potrebbero non essere sufficienti per misurare i cambiamenti nel HbA1c. (94)
A seconda della popolazione studiata e del tipo e della qualità degli studi osservati, è dimostrato che il Cromo influisce in modo eterogeneo sui livelli di emoglobina A1C.
La supplementazione con 400 o 800mcg di Cromo (come Picolinato) insieme a un pasto di prova in adulti sani ha ridotto l’area del glucosio sotto la curva (AUC) del 30-36% nei responder, con la dose bassa più efficace.(97) In particolare, i responder sono stati classificati come soggetti aventi un consumo di carne e latte relativamente più basso (97), suggerendo che il Cromo può influenzare il metabolismo del glucosio postprandiale negli individui con livelli di Cromo basali inferiori. La riduzione del glucosio non è stata associata ad alcun cambiamento nell’Insulina, escludendo un effetto insulinogeno, e si è verificata in persone senza un metabolismo del glucosio alterato.(97)
Nella meta-analisi dove sono stati vagliati gli studi che valutavano la supplementazione di cromo oltre ai 250mcg nei diabetici di tipo II per un periodo di tre mesi (o più lungo)(94), i sette studi inclusi nella meta-analisi (55)(98)(99)(100)(101)(102)(103) non hanno mostrato una riduzione dei livelli di HbA1c nel siero nonostante una lieve riduzione della glicemia (RR di -0,95 e un IC 95% da -1,4 a -0,5).(94)
Un’analisi dei dati osservazionali del National Health and Nutrition Examination Survey (NHANES) ha rilevato che le persone che hanno consumato un integratore alimentare contenente Cromo avevano una probabilità inferiore di sviluppare il diabete (OR = 0,73), definito avendo un livello di HbA1c superiore a 6,5. L’uso di integratori in generale non ha avuto un effetto statisticamente significativo sulle probabilità di sviluppare diabete in questo studio.(104)
L’aggiunta di 400mcg di Cromo (come Picolinato) a una bevanda contenente carboidrati prima di un shuttle run test in uomini sani e attivi non ha modificato i benefici della bevanda contenente carboidrati rispetto al controllo, suggerendo che non vi è alcun beneficio aggiuntivo. (105)
Uno studio nel quale è stata somministrata una dose di 600mcg di Cromo (come Picolinato) ogni giorno per un mese prima di un esercizio di deplezione del glicogeno ha rilevato che immediatamente dopo l’esercizio e nell’ora successiva il gruppo trattato aveva livelli di lattato significativamente più alti rispetto al placebo. (106) In un altro studio che utilizzava un modello shuttle-run exercise, questo aumento di lattato non si è verificato con oltre 75 minuti di test a seguito del consumo di 400mcg di Cromo Picolinato o carboidrati o acqua (gruppo di controllo). (105) Inoltre, la concentrazione di lattato e il grado di fatica era simile in questo studio tra i due gruppi e il controllo. (105)
Glicogeno Sintasi
Il Glicogeno Sintasi è l’enzima responsabile della conversione del glucosio in glicogeno, la forma di deposito dei carboidrati nel corpo. Allo stesso modo, la fosforilasi di glicogeno è coinvolta nello scomporre queste riserve di carboidrati in glucosio per produrre energia. A causa dei suoi effetti sul metabolismo del glucosio, il Cromo è stato studiato per il suo impatto sulle riserve di glicogeno. Prove preliminari hanno rivelato che i ratti supplementati con Cromo avevano una minore dispersione del glicogeno epatico rispetto al gruppo di controllo durante il digiuno. (107) Successivamente, si è notato che il Cromo aumentava l’attività dell’enzima glicogeno sintasi nel muscolo e nel fegato dei ratti addestrati rispetto al gruppo di controllo non supplementato, ma la glicogeno fosforilasi non era influenzata.(108)
Negli adulti sovrappeso e leggermente allenati o sedentari la somministrazione di 600mcg di Cromo (come Picolinato) per un mese insieme ad una dieta standardizzata con gli ultimi due giorni progettati per esaurire il glicogeno, la supplementazione non ha modificato i livelli di glicogeno o il tasso di risintesi (da un carboidrato contenuto in una bevande) rispetto al placebo.(106)
Secondo la meta-analisi sul peso nei diabetici di tipo II supplementati con Cromo (oltre 250mcg) per oltre tre mesi, non vi è stata alcuna alterazione significativa del peso rispetto al placebo, nonostante una modesta riduzione del glucosio nel sangue.(94) Al contrario, un’altra meta-analisi ha rilevato che gli adulti sovrappeso e obesi che hanno integrato con il cromo picolinato hanno ridotto il peso corporeo nell’intervallo di dosaggio di 200-1.000 μg, indipendentemente dallo stato diabetico. La perdita di peso è stata tuttavia molto modesta, per un totale di soli 1,1 kg (IC del 95% nell’intervallo 0,4-1,7 kg).(109) Da notare, quest’ultima meta-analisi ha ritenuto la qualità delle prove non ottimale, mettendo in discussione gli effetti mediati dal cromo sulla perdita di peso.(109)
Uno studio ha rilevato che l’aumento di peso associato alla terapia con Sulfonilurea (0,9 kg su 10 mesi) nei diabetici è stato mitigato dalla cosomministrazione di 1.000mcg di Cromo. (102) È importante sottolineare che questi risultati possono essere limitati a coloro che sono sottoposti a terapia Sulfonilurea. Quando i soggetti diabetici che non erano stati trattati con il medicinale ricevevano istruzioni per seguire una dieta di mantenimento del peso, la supplementazione di 1.000mcg di Cromo Picolinato non modificavano l’assunzione di cibo, l’appetito o il peso corporeo. (55)
Il Cromo è stato anche usato nel tentativo di mitigare l’aumento di peso associato alla cessazione del fumo, poiché le persone che smettono di fumare spesso tendono ad aumentare di peso.(110) Questo studio ha utilizzato l’Hypericum perforatum (900 mg) come primo aiuto anti-fumo e poi ha diviso i soggetti dello studio in gruppi trattati con Cromo o placebo. Sfortunatamente, la tendenza del Cromo ad attenuare l’aumento di peso non ha potuto essere testata con sufficiente potenza, a causa dei bassi tassi di successo con l’erba di cui sopra. Tuttavia, gli effetti del Cromo erano promettenti, con una probabilità di attenuare l’aumento di peso da 5,76 kg a 2,7kg dopo sei mesi.(111)
Il Cromo può anche ridurre la perossidazione lipidica in alcune popolazioni. Sono necessari però ulteriori studi per determinarne la dose appropriata e chi potrebbe trarne reale beneficio.
Come accennato in precedenza, Il Cromo Picolinato, più di altre forme trivalenti di Cromo, ha la capacità di formare proossidanti che possono potenzialmente causare danni al DNA. La rilevanza per la supplementazione orale standard non è nota, poiché la concentrazione richiesta per danneggiare il DNA (livello alto di 50μM) è significativamente più alta di quella osservata nel sangue dopo l’ingestione orale di integratori. Inoltre, gli studi su soggetti umani non hanno notato danni al DNA con dosi supplementari standard (200-400mcg/die).
Il Cromo si accumula nei testicoli dei ratti quando iniettato, anche se i possibili benefici o danni nei testicoli con integrazione orale di Cromo non sono stati studiati. Il Cromo esavalente, la forma tossica non presente negli integratori, è noto per essere tossico per i testicoli.
Esistono diverse altre azioni potenziali legate all’uso del Cromo che sono state scientificamente documentate. Per ovvie ragioni, legate soprattutto all’argomento principale trattato in questa serie di articoli, ho omesso diversi studi di un certo interesse. Per chiunque volesse approfondire cliccare qui.
In seguito alle numerose informazioni riportate nel presente articolo, possiamo con una certa sicurezza concludere che una supplementazione di Cromo risulta maggiormente incisiva in caso di carenza del minerale a causa di una insufficiente assunzione con gli alimenti o in condizioni di insulino resistenza (sia “pre-diabetica” che nella condizione diabetica). Il potenziale anoressizzante del Cromo rappresenta sicuramente un elemento di vantaggio durante una dieta ipocalorico (specie se ipoglucidica). Il Cromo sembrerebbe avere anche una certa azione sul miglioramento dello stoccaggio del glicogeno e sulla sua preservazione, anche se la cosa, in realtà, non è mai stata riscontrata nell’uomo. Se tale azione fosse possibile o significativa, si potrebbe riflettere positivamente a livello prestativo ma che in un contesto di “scarico del glicogeno” potrebbe risultare limitante.
A questo punto la domanda che si ripresenta è “come si possono utilizzare queste informazioni per pianificare l’uso del Cromo”?
Vista la sua efficacia in soggetti patologici e/o in condizioni di insulino resistenza non patologica, l’uso temporalmente ridotto di 1mg (stand alone) o protratto di 400-600mcg (in combinazione con altri GDA; vedi possibile azione additiva con la Berberina e ALA) di Cromo Picolinato al giorno può portare a dei vantaggi in quei soggetti con una insulino-resistenza di base genetica, cioè individui con una tolleranza glucidica limitata rispetto alla media, o durante regimi ipercalorici (vedi peggioramento dell’insulino-resistenza durante regimi ipercalorici).
Dosi contenute di Cromo Picolinato (200-400mcg/die) assunte durante periodi ipocalorici possono aiutare il soggetto trattato a tollerare la riduzione calorica per via dell’effetto anoressizzante dato dall’uso di questo composto.
In combinazione con altri GDA durante e nel periodo successivo (periodo “protocollare” di 4 settimane) all’uso di Insulina esogena.
Alcuni effetti collaterali comuni riscontrati con l’uso del Cromo Picolinato (dose correlato) possono includere insonnia, cambiamenti di umore, irritabilità e mal di testa.
Altri effetti collaterali possibilmente riscontrabili con l’uso di alte dosi di Cromo Picolinato includono problemi di coordinamento o di equilibrio, problemi di concentrazione o difficoltà di pensiero, e sintomi legati a problemi epatici (che comprendono: nausea; mal di stomaco nella zona superiore; prurito; stanchezza; perdita di appetito; urina di colore scuro; ingiallimento della pelle o degli occhi (ittero)).
Chiedere prontamente assistenza medica di emergenza se si verificano segni di anafilassi, una reazione allergica grave che può includere orticaria, difficoltà di respirazione o gonfiore del viso, delle labbra, della lingua o della gola.
Esiste un caso studio di una donna che in seguito all’ingestione di 1200-2400mcg di Cromo (come Picolinato) per 4-5 mesi mostrava sintomi di danno renale. (112) In un altro caso studio, un Bodybuilder aveva sviluppato rabdomiolisi associata all’assunzione di 1.200mcg di Cromo Picolinato per due giorni.(113)
La possibile comparsa di questi effetti avversi può essere evitata con una attenta calibrazione della dose giornaliera di Cromo. Se ne sconsiglia quindi un assunzione superiore a 1mg/die (dose quest’ultima comunque relegabile a periodi d’uso brevi). Una supplementazione giornaliera di 200-400mcg di Cromo Picolinato è generalmente ben tollerata con una punta massima di dosaggio di 600mcg/die.
Per ottenere una migliore biodisponibilità del composto, il Cromo andrebbe assunto lontano dalla somministrazione di integratori di Zinco o con pasti contenenti fonti ricche di fitati (vedi cereali integrali e legumi; il cui contenuto di fitati può comunque essere ridotto con, ad esempio, l’ammollo e la adeguata cottura).
Come detto la volta scorsa per l’Acido Alfa Lipoico, anche con l’uso di Cromo Picolinato (o altra forma) è essenziale la cura del dosaggio e la ponderatezza nell’utilizzo.
Se non avete letto ancora la prima parte di questa serie di articoli vi invito a farlo.
Acido Alfa Lipoico: caratteristiche e possibili applicazioni.
Acido Alfa Lipoico
L’Acido Alfa Lipoico (ALA), denominato anche come AcidoLipoico (LA) o Acido Tiottico, fu isolato per la prima volta nel 1951 dal tessuto epatico ad opera dei biochimico americani L.J. Reed e I.C. Gunsalus i quali riuscirono ad ottenerne solo pochi milligrammi partendo da grandi quantità di tessuto.
L’ALA è un composto presente in natura con il nome chimico di acido 1,2-ditiolan-3-pentanoico, a volte indicato come Acido Tiottico.(1)
L’ALA si trova in diversi alimenti, principalmente nelle frattaglie e in alcuni frutti e verdure. (2)(3) Alcuni di questi alimenti con contenuto specifico includono:
Spinaci a 3.14 +/- 1.11mcg/g di peso secco come lipoillisina (4)(5)
Rene a 2,64 +/- 1,23mcg/g di peso secco come lipoillisina (4)(5)
Fegato a 1,51 +/- 0,75mcg/g di peso secco come lipoillisina (4)(5)
Broccoli a 0,94 +/- 0,25 mcg/g di peso secco come lipoillisina (4)(5)
Tessuto cardiaco a 0,86 +/- 0,33mcg/g di peso a secco come lipoillisina (4)(5)
Pomodori a 0,56 +/- 0,23mcg/g di peso secco come lipoillisina (4)(5)
La lipoillisina, un coenzima coinvolto nel funzionamento del complesso della piruvato deidrogenasi, è formata dalla coniugazione tra Acido Lipoico e un residuo di Lisina, che avviene grazie ad un legame di tipo ammidico, ed è una forma di conservazione dell’Acido Alfa Lipoico legato alle proteine.(6) È scisso in AL e Lisina attraverso l’azione dell’enzima glicoproteico lipoamidasi (a volte indicato come lipoillisina idrolasi), che circola nel siero umano.(7)
Anche se l’ALA è presente in varie fonti alimentari, come visto precedentemente, le sue concentrazioni negli alimenti tendono ad essere significativamente inferiori alle dosi standard per la supplementazione.
L’ALA è un composto ditiolo presente in natura sintetizzato nei mitocondri a partire dall’Acido Ottanoico, con un buon livello di sintesi che si verifica nei mitocondri degli epatociti.(8)(3)
Il suo principale ruolo biologico è quello di cofattore di enzimi mitocondriali come l’Alfa-chetoglutarato deidrogenasi e il piruvato deidrogenasi.(9) L’ALA sembra essere coinvolto nella produzione di acetil-CoA, attraverso la decarbossilazione ossidativa del piruvato.(10)
È stato dimostrato che l’integrazione fornisce benefici protettivi contro l’ossidazione, l’infiammazione, il diabete e il declino cognitivo.(9)
L’ALA possiede un carbonio centro chirale e quindi può esistere in un isomero S o R. L’ALA non specificato è una soluzione ‘racemica’ di entrambi, mentre l’R-ALA, spesso venduto come integratore alimentare, è comunemente legato al sodio (Na-R-ALA). L’R-ALA è la forma naturale di Acido Lipoico biologicamente attiva.
Il legame disolfuro del ALA può essere scisso omoliticamente attraverso l’esposizione ai raggi UV e/o ad una fonte di calore (11)(12) durante la quale la struttura dell’anello ditiolano forma due radicali tiili e si auto-polimerizza in una catena lineare di disolfuri nota come PBCPD. Il nome completo è poli {3- (n-butano acido carbossilico) propil] disolfuro.(13) Questa polimerizzazione è vista come reversibile, con riconversione in ALA in soluzione alcalina o con coincubazione con agenti riducenti come il ditiotreitolo e il β-mercaptoetanolo.(14) È stato suggerito che l’ALA presente in natura può essere una miscela racemica che include un contenuto di PBCPD.(15)
In vivo, l’ALA può essere ridotto alla forma ditiol (dove la struttura dell’anello è rotta), denominata Acido Diidrolipoico (DHLA). (3) Nelle cellule provviste di mitocondri, questa riduzione è mediata dalla lipoamida deidrogenasi ed è una reazione NADH-dipendente. Nelle cellule sprovviste di mitocondri, questa riduzione avviene tramite NADPH con glutatione e tioredossina riduttasi.(16)
L’ALA ha un punto di fusione di 63 ° C quando in soluzione racemica e di 50 ° C come isomero R-ALA e l’associazione con sali con punti di ebollizione più elevati può migliorarne la stabilità.(17)(18)
L’ALA ingerito per via orale viene rapidamente assorbito nell’intestino in modo dipendente dal pH dei trasportatori dell’Acido Monocarbossilico (MCT). La sua coingestione con Acidi Monocarbossilici come i Trigliceridi a Catena Media o l’Acido Benzoico ne inibisce l’assorbimento.(19) Esiste anche la possibilità che l’ALA venga assorbito dal sistema di trasporto multivitaminico sodio-dipendente (SMVT). (20)(21) Nell’intestino, alcune molecole di ALA si convertono in Acido Diidrolipoico. La biodisponibilità complessiva della supplementazione di ALA è di circa il 30% (22)(9) a causa dell’elevata estrazione epatica.(23) La forma di ALA Na-R-ALA è completamente solubile in acqua.(24) Sebbene l’enantiomero R abbia tassi di assorbimento intestinale più elevati in vivo (25), l’enantiomero S può stabilizzare l’assorbimento prevenendo la polimerizzazione.(9)
La farmacocinetica sistemica dell’ALA è abbastanza rapida. Dopo un veloce assorbimento intestinale, l’ALA viene rapidamente suddiviso nei tessuti che lo assorbono (cervello, cuore e muscoli) , il che include un deposito transitorio nel fegato.(9)(26) L’ALA si accumula nel cervello dopo appena un’ora dall’ingestione (27) e viene immagazzinato in varie regioni dell’organo.(28)
Dopo ingestione orale di una miscela racemica di ALA da 600mg, la Cmax sembra essere di 6,86+/-1,29μg/mL con un Tmax di 50,8 minuti e una AUC complessiva di 8 ore di 5,65 ± 0,79μg/mL/h.(29)
Nelle cellule, l’ALA viene principalmente metabolizzata attraverso la beta-ossidazione. (30) I principali metaboliti sono il Bisnorlipoato, il Tetranorlipoato, il β-idrossi-bisnorlipoato o i mercapto derivati bis-metilati di questi composti (26) e l’Acido Diidrolipoico, che subisce una rapida escrezione cellulare.(16)
L’ALA viene anche espulso rapidamente per filtrazione renale, con il 98% di ALA assorbito escreto entro 24 ore.(23) Tuttavia, la maggior parte dell’ALA ingerito per via orale viene persa nell’escrezione fecale prima che avvenga l’assorbimento intestinale.(30) Per questi motivi, l’ALA non è immagazzinato a lungo termine. Secondo Shay et al. (9) l’AUC media è di circa 160+/-35mcg/ml/min e la Cmax media è di 2,8+/-1,5 per una dose orale di 600mg di entrambi gli enantiomeri. Questi risultati rivaleggiano con il Na-R-ALA per la AUC (nonostante l’R-ALA mostri valori di picco più elevati ed escrezione più rapida), ma sono ben superiori rispetto all’S-ALA da solo.(25)
L’ALA può indurre l’espressione della lipasi trigliceridica nelle cellule epatiche (responsabile della diminuzione delle sequenze di trigliceridi in queste cellule (31)) secondarie all’attivazione dell’AMPK, che ha ridotto l’accumulo di lipidi in vitro. (32) L’AMPK è stato attivato in modo tempo e concentrazione-dipendente ed è stato in grado di farlo nonostante le concentrazioni elevate di glucosio (30 mM) e palmitato (0,1 mM) a concentrazioni di 0,25-1 mM.(32) Queste interazioni del AMPK sono indipendenti dalle proteine Sirtuine e sembrano aggirare le azioni dell’Insulina sul fattore di trascrizione nucleare FOXO1 prevenendo l’esclusione nucleare, che sembra essere secondaria anche all’AMPK.(32) Quando somministrato il 2,4% di ALA per 5 settimane attraverso la dieta a ratti geneticamente obesi, (circa 40mg/kg di peso corporeo in questo studio dopo il controllo della biodisponibilità del 20%) si è osservata una riduzione dell’accumulo di trigliceridi nel tessuto epatico (-26%) e un aumento del contenuto di glicogeno (+ 27%). Rispetto ai ratti trattati con una alimentazione caloricamente limitata, il gruppo dei ratti trattati con ALA presentava fegati più grandi ma senza biomarcatori anormali, probabilmente a causa del aumentato contenuto di glicogeno. (33)
Anche la produzione di anione superossido nel fegato dei ratti alimentati con l’1% di ALA sembra essere ridotta rispetto al gruppo di controllo e l’aumento della produzione di superossido in risposta all’aggiunta di glucosio alla dieta, per la maggior parte, è stato soppresso.(34)
L’AMPK non è l’unico meccanismo in grado di ridurre l’accumulo di grasso nel fegato, è infatti possibile farlo attraverso l’inibizione dell’azione genetica delle proteine pro-lipogeniche LXR e delle proteine specifiche 1.(35) L’ALA può aumentare il numero dei recettori PPARα quando viene inserito all’1% del contenuto della dieta per un periodo di 14 settimane ed è stato negativamente correlato (r = 0,8) con livelli di acidi grassi liberi nel sangue.(34)
Sebbene gli effetti benefici dell’ALA sulla fisiologia epatica nei modelli sopra citati siano ben stabiliti, uno studio che confronta gli effetti della supplementazione di ALA a lungo e breve termine in topi sani suggerisce che l’integrazione regolare a lungo termine può causare danni epatici.(36)
Topo “Black 6”
Per esaminare l’effetto della supplementazione di ALA a breve e lungo termine sul fegato, i topi “black 6” (C57BL6 / J, un ceppo comune di topi da laboratorio) sono stati trattati con ALA a 20mg/kg per 4 o 74 settimane.(36) Dopo il periodo di trattamento, i topi sono stati sottoposti ad eutanasia, seguita da analisi del tessuto epatico per il metabolismo dei lipidi e del colesterolo. L’integrazione di ALA a breve e a lungo termine ha causato un aumento della β-ossidazione e una diminuzione della lipogenesi. Al contrario, sia il trattamento a breve che a lungo termine aumentava il contenuto di colesterolo epatico del 70% e del 110%, rispettivamente, e aumentava i livelli di trigliceridi, inducendo trigliceridemia sistemica. Inoltre, nonostante il fatto che il trattamento a breve termine con ALA abbia diminuito l’espressione lipogenica, ha anche causato l’accumulo di grasso nel fegato. I topi trattati a lungo termine con ALA mostravano un fenotipo peggiore, con un ampio accumulo di grasso che portava alla steatosi epatica e ad un esteso danno epatico.(36)
Ratto “ZDF”
In particolare, lo studio di cui sopra risulta l’unico ad aver studiato gli effetti del trattamento con ALA a lungo termine in topi sani. Al contrario, altri studi che hanno mostrato un effetto benefico del trattamento a lungo termine con ALA sono stati condotti su modelli animali malati, inclusi ratti grassi nutriti con una alimentazione ricca di grassi (35) e ratti Zucker grassi e diabetici (ZDF).(37) Pertanto, il trattamento a lungo termine di ALA in modelli animali sani può essere tossico per il fegato, causando un fenotipo che ricorda da vicino quello della steatosi epatica non alcolica. (38) (39) Va notato che la dose di ALA utilizzata nello studio sui topi sani (36) (20mg/kg) è equivalente a circa 1,8g/giorno in un uomo di 90Kg. Sebbene si tratti di una dose relativamente alta di ALA, è abbastanza vicina alle dosi standard utilizzate nell’uomo e, per tale motivo, il loro utilizzo sul lungo termine richiede una certa cautela.
L’ALA ha effetti potenti sulla riduzione dell’appetito. Tuttavia, l’ALA sembra indurre qualche forma di perdita di peso oltre a quella legata alla semplice soppressione dell’appetito (sebbene la soppressione dell’appetito sembri essere il fattore di influenza più potente).(40) Almeno uno studio pair-fed (gruppo di controllo, gruppo supplementato con ALA, e gruppo alimentato con la quantità di calorie che gli animali del gruppo trattato con ALA ha voluto consumare) ha osservato che la statisticamente significativa perdita di peso diventava irrilevante tra il gruppo trattato con ALA e il gruppo nutrito con lo stesso monte calorico.(33)
Il fattore più significativo che influenza gli effetti di riduzione del peso dati dall’uso di ALA è la riduzione dell’appetito e gli studi pair-fed suggeriscono che ciò può rappresentare l’80-90% (stima approssimativa derivata dai grafici) degli effetti complessivi della riduzione del peso secondari all’uso dell’ALA.
In vitro, l’ALA è stato in grado di indurre la secrezione di apelina dalle cellule adipose (una adipokina che può regolare il metabolismo del glucosio), ma è stato ritenuto non correlato ai cambiamenti osservati in vivo.(41)
Per quanto riguarda l’AMPK, almeno uno studio ha riportato risultati che suggeriscono che l’inibizione del AMPK potrebbe verificarsi negli adipociti 3T3-L1, ma lo studio non è stato progettato per rispondere a queste domande.(41) Altri due studi sottolineano che nel tessuto adiposo bianco avviene sia una attivazione dell’AMPK che una sovraregolazione del mRNA.(42)(43)
Bomba Calorimetrica
Quando si studiano le interazioni dell’ALA con l’assorbimento dei nutrienti, la sua supplementazione per 56 giorni è stata in grado di ridurre l’assorbimento dei carboidrati secondario all’inibizione del trasportatore SGLT1 (trasportatore del glucosio sodio-dipendente) di circa un terzo, quando il jujenum è stato asportato e testato in vitro con alfa- metilglucosio. (44) Se testati in vivo allo 0,5% di ALA, tuttavia, non vi sono differenze significative nel contenuto calorico delle feci (valutato mediante bomba calorimetrica).(45)
Quindi, l’ALA ha il potenziale per inibire l’assorbimento dei nutrienti, ma non sembra abbastanza potente da creare un impatto significativo.
Negli animali alimentati con una dose di ALA del 0,5% dell’assunzione di cibo, quando l’apporto calorico era controllato (ricordiamoci che l’ALA può sopprimere l’appetito), il dispendio energetico valutato mediante calorimetria indiretta aumentava dal giorno 3 e continuava ad essere elevato per i 21 giorni del test.(46) Questi ratti hanno mostrato una maggiore espressione del UCP1 nel tessuto adiposo bruno e nell’espressione ectopica del UCP1 nel tessuto adiposo bianco, ritenuta una ragione dell’aumento del tasso metabolico.(46) Nei ratti più anziani trattati con ALA allo 0,75% per 4 settimane, è stato osservato un aumento del tasso metabolico attraverso un meccanismo AMPK / PGC-1° dipendente. Questo tasso metabolico (associato ad una riduzione del 18% dell’assunzione di cibo) ha comportato una perdita di peso totale del 15,8%.(47) Il consumo di ossigeno e la produzione di anidride carbonica in questi ratti anziani alimentati con ALA allo 0,75% sono aumentati rispettivamente del 27% e del 38%.(47)
In uno studio condotto su 228 persone (360 all’inizio con un alto tasso di abbandono), che erano obese o sovrappeso e con anomalie metaboliche (sindrome metabolica), trattate con 1.2g o 1.8g di ALA (divisi in tre dosi giornaliere prima dei pasti) per 20 settimane è stata osservata una significativa riduzione del peso nel gruppo trattato con 1.8g quando tutti i gruppi erano soggetti a un deficit di 600kcal.(48) La perdita di peso media è stata di 0,94+/-0,45kg nel gruppo placebo, 1,49+/-0,38kg nel gruppo trattato con 1,2g e di 2,76+/-0,53kg nel gruppo trattato con 1,8g.(48)
Nei ratti anziani, i miglioramenti nel contenuto del mRNA di GLUT4 e PGC-1a sono aumentati del 105% e dell’80% (rispettivamente) dopo 4 settimane di assunzione di ALA allo 0,75%. (47)
Uno studio sui ratti ha osservato che con iniezioni di ALA di 30mg/kg, le proteine da shock termico 72 e 25 sono state indotte nella dieta ad alto contenuto di grassi (60%) ma non in quella a basso contenuto di grassi (10%), condizione che è stata in grado di ridurre la segnalazione proinfiammatoria via JNK e NF-kB (riportata altrove (49) (50)) e di migliorare la resistenza all’insulina indotta dagli acidi grassi.(51) L’ALA è stato precedentemente implicato (insieme ad altri antiossidanti, Vitamina C e Vitamina E) nella riduzione dell’attività del IRS-1 e nel miglioramento della sensibilità all’insulina attraverso questo meccanismo.(52)
Sono stati osservati anche aumenti dei marker del metabolismo lipidico, cioè un aumento della fosforilazione di AMPK, ACC, FAS e ATGl. L’effetto di questa aumentata β-ossidazione è stata la diminuzione dell’accumulo di lipidi.(53) Un’aumentata espressione del SIRT1 è stata osservata nei miotubi in modo secondario all’AMPK e ad un aumento del rapporto NAD/NADH, tuttavia il knockdown del SIRT1 con siRNA riduce la β-ossidazione indotta dall’AMPK in queste cellule. (53) Questi effetti non erano significativamente più potenti del Resveratrolo, un noto inibitore della PDE4 che influenza l’AMPK.(53) L’ALA allo 0,5% della dieta ha dimostrato di ridurre l’accumulo di lipidi nelle cellule adipose, ma questo studio ha attribuito ciò agli effetti anoressizzanti (soppressione dell’appetito) piuttosto che attraverso l’azione dell’AMPK.(45)
Osservando il meccanismo della sovraregolazione dell’AMPK, è stato dimostrato che questo può verificarsi indipendentemente dal rapporto AMP: ATP (contrastando uno studio precedente che suggeriva come causa l’attivazione del LKB1 (53)) e secondario all’aumento della concentrazione di calcio intracellulare a 200 μM e 500um.(54) Il calcio intracellulare chelante può inibire gli effetti dell’ALA sull’AMPK, così come può inibire l’enzima CaMKK, che rilascia il calcio nei miociti.(54)
Uno studio ha osservato che, nei ratti alimentati con una dieta al 10% di grassi l’ALA non era in grado di stimolare l’assorbimento del glucosio nelle cellule muscolari in vivo, ma era in grado di migliorare la riduzione del 54,7% dell’assorbimento di glucosio osservata nei ratti alimentati con una dieta ricca di grassi (60%) del 55,7%.(51)
La supplementazione orale di 1.8g di ALA per 2 settimane non sembra influenzare i tassi di secrezione di Insulina in uomini sani anche se sovrappeso o obesi.(55)
L’alterazione della sensibilità all’insulina osservata in condizioni di ipertrigliceridemia non sembra subire miglioramenti con l’uso giornaliero di 1,8g di ALA per 2 settimane.(55)
L’ALA è stato studiato per uso orale a dosi di 300, 600, 900 e 1.200mg di una miscela racemica per un periodo di assunzione di 6 mesi in soggetti con diabete di tipo II confermato (alcune in terapia anti-iperglicemica). I ricercatori hanno osservato una tendenza dose-dipendente nella riduzione della glicemia a digiuno e del HbA1c ad ogni dosaggio indicato, e una riduzioni significative quando tutti i gruppi sono stativagliati nell’insieme e quando i soggetti sono stati confrontati con il basale. (56) Uno studio più lungo durato 4 anni con una somministrazione giornaliera di 600mg di ALA ha mostrato una maggiore diminuzione del HbA1c associata all’ALA (0,67 ± 1,41%) rispetto al placebo (0,48 ± 1,46%), ma non ha raggiunto un impatto significativo.(57)
L’ALA, sopprimendo l’AMPK ipotalamico ma attivando l’AMPK periferico, presenta somiglianze meccanicistiche con l’ormone Leptina. Tuttavia, quando testato sui topi, sia gli animali con recettori della Leptina che quelli privi di tali recettori hanno mostrato questi effetti, suggerendo che l’ALA agisce come un Leptina-mimetico nei risultati (ma non nei meccanismi) e può aiutare a superare la resistenza alla Leptina bypassando il recettore.(46)
Quando somministrato a ratti soggetti a steatosi epatica non alcolica (NAFLD), l’ALA può sopprimerne la patogenesi e l’aumento previsto della Leptina (58) e aumentare i livelli di Leptina in un modello di diabete di tipo I, condizione quest’ultima che causa una riduzione della Leptina. (59) Se somministrato allo 0,25% della dieta in ratti sani, si osserva una diminuzione della Leptina circolante e dell’mRNA della Leptina dopo 8 settimane di assunzione ed è stata correlata (r = 0,908) con i livelli di tessuto adiposo bianco. (60) Adipociti isolati da questi ratti dopo 8 settimane soggetti a 250uM di ALA hanno aumentato la conversione del glucosio in lattato (con un significativo aumento del lattato del 44% a 500 μM) e questo aumento del lattato è stato correlato con una diminuzione della secrezione di Leptina.(60) L’ALA sembra essere associato ad una aumentata fosforilazione del Sp1, un fattore di trascrizione nucleare indotto dal glucosio che stimola la Leptina. La sua fosforilazione impedisce le sue azioni nel nucleo e le azioni dell’ALA sono state imitate dagli inibitori del PI3K.(60)
L’ALA è stato correlato ad un aumento dell’assorbimento di Creatina in seguito ad uno studio di Burke et al. (2003), nel quale venne stabilito che la cosomministrazione di ALA e Creatina migliorasse il trasporto e il contenuto muscolare totale di quest’ultima.(61) Purtroppo, lo studio presenta delle limitazioni che ne impediscono una valutazione concreta e che vanno dalle caratteristiche dei soggetti utilizzati (non allenati prima dello studio) al tipo di carboidrato assunto. In definitiva, sembra che l’ALA velocizzi soltanto l’uptake cellulare di Creatina ma non il contenuto totale di questa.
Viene sconsigliata un assunzione giornaliera superiore a 1.8g onde evitare possibili casi di tossicità. Viene consigliata la contemporanea somministrazione di vitamine del gruppo B insieme all’ALA dal momento che quest’ultimo ne può provocare la perdita.(62)
Esistono diverse altre azioni potenziali legate all’uso dell’ALA e che sono state scientificamente documentate. Per ovvie ragioni, legate soprattutto all’argomento principale trattato in questa serie di articoli, ho omesso diversi studi di un certo interesse. Per chiunque volesse approfondire cliccare qui.
Per concludere l’analisi delle potenzialità dell’Acido Alfa Lipoico sul piano metabolico (e prettamente glucidico), non si può che esserne affascinati ed in parte delusi quando si soppesano i possibili effetti con la loro limitata applicabilità. A differenza della Berberina (vista nel dettaglio nella prima parte di questa serie di articoli), che ha mostrato una versatilità estrema accompagnata da una efficacia ben documentata, l’ALA ha mostrato invece di avere un impatto significativo (soprattutto come GDA) solo in contesti patologici o in condizioni che potremmo definire “induttive alla patologia” (vedi diete con elevato contenuto lipidico e insulino resistenza correlata). Se da un lato l’ALA sembri avere un vantaggio sullo stoccaggio del glicogeno epatico dall’altro mostra (a dosaggi >1-1.2g) sul lungo periodo la possibilità di causare danno epatico in soggetti sani. Questo rappresenta sicuramente una limitazione al suo utilizzo. L’impatto dell’ALA sulla soppressione dell’appetito, sull’aumento della beta-ossidazione e sui meccanismi legati alla Leptina, risultano sicuramente interessanti per i soggetti sottoposti a diete ipocaloriche e/o ipoglucidiche.
A questo punto la domanda che si ripresenta è “come si possono utilizzare queste informazioni per pianificare l’uso dell’ALA”?
Vista la sua efficacia in soggetti patologici e/o in condizioni che in precedenza abbiamo classificato come “induttive alla patologia”, l’uso temporalmente ridotto di 1.2-1.8g (stand alone) o protratto di 600mg (in combinazione con altri GDA; vedi possibile azione additiva con la Berberina) di ALA al giorno può portare a dei vantaggi in quei soggetti con una insulino-resistenza di base genetica, cioè individui con una tolleranza glucidica limitata rispetto alla media, in condizioni nelle quali la percentuale dei macronutrienti pende in maniera marcata verso il consumo lipidico (vedi diete chetogeniche) o durante regimi ipercalorici (vedi peggioramento dell’insulino-resistenza durante regimi ipercalorici).
Dosi contenute di ALA (400-600mg/die) assunte durante periodi ipocalorici possono aiutare il soggetto trattato a tollerare la riduzione calorica per via dell’effetto anoressizzante dato dall’uso di questo composto (legato al suo effetto leptino-mimetico non recettore-dipendente a livello ipotalamico).
In combinazione con altri GDA durante e nel periodo successivo (periodo “protocollare” di 4 settimane) all’uso di Insulina esogena.
Gli effetti collaterali legati all’uso dell’ALA alle dosi e modalità di “sicurezza” sopra indicate sembrano essere rari e lievi, come rusch cutanei. I soggetti che decidono di assumerlo in concomitanza con l’uso di Insulina esogena devono prestare molta attenzione alla dose utilizzata del peptide in rapporto agli effetti indotti dalla dose di ALA utilizzata. È possibile, infatti, che possano aumentare i casi ipoglicemici (cosa possibile con tutti i GDA cosomministrati in tali circostanza). Un controllo regolare della glicemia ematica è una accortezza imprescindibile sia con l’uso di Insulina da sola che, in misura maggiore, cosomministrata con composti aventi effetti ipoglicemizzanti.
Quindi, cura del dosaggio e ponderatezza nell’utilizzo.