
Introduzione:
Negli ultimi anni il mondo del Bodybuilding e del Fitness è stato letteralmente invaso da affermazioni sensazionalistiche di stampo pubblicitario su una classe di molecole steroidee provenienti dal mondo animale e vegetale: gli Ecdysteroidi. Tra questi sono emersi agli onori della cronaca il 20-hydroxyecdysone e il Turkesterone. Quest’ultimo, in particolare, è stato fortemente pubblicizzato da “influencer” con nozioni di biochimica pari o prossime allo 0 e da venditori con sete speculativa mista ad ignoranza e malafede.
In base a quanto sinteticamente esposto, ho deciso di riportare le cose come sono alla luce delle attuali evidenze scientifiche e della lucida osservazione empirica e dei dati aneddotici.
Introduzione agli Ecdysteroidi:
Gli Ecdysteroidi sono ormoni steroidei degli artropodi responsabili principalmente della muta, dello sviluppo e, in misura minore, della riproduzione;[1][2][3] esempi di Ecdysteroidi includono Ecdysone, Ecdysterone, Turkesterone e 2-deossiecdysone. [4] Questi composti sono sintetizzati negli artropodi a partire dal colesterolo alimentare attraverso la fase metabolica influenzata dalla famiglia dei citocromi P450.[5] I Fitodisteroidi sono presenti anche in molte piante, per lo più come agenti protettivi (tossine o antifeedanti) contro gli insetti erbivori.[6][7]
Infatti, gli Ecdysteroidi sono presenti in molte piante (circa il 6% delle piante esistenti)[8], anche se a livelli solitamente considerati insufficienti per l’estrazione o l’attività biologica. Alcune piante che presentano una maggiore quantità di Ecdysteroidi bioattivi sono:
- Asparagus Filicinus[9]
- Spinacia oleracea (Spinaci, fonte di 20-idrossidisone)[10]
- Quinoa, soprattutto nella crusca, che contiene principalmente 2-idrossidisone e makisteroni[11] e varia da 450-1300mcg/g di ecdysone equivalente[12].
- Ignami[13]
- Funghi a bottone bianco[11]
- Ajuga Turkestanica, fonte del “Turkesterone” idrossilato C-11[14]
- Rhaponticum carthamoides
- Silene Praemixta (2-deossiecdysterone e 2-deossi-alfa-ecdysone)
- Vitex Scabra, con l’1,8% di ecdisteroidi in peso[15] e altre specie di Vitex[16] come cymosa[17] e canescens[18].
Gli Ecdysteroidi prendono il loro nome dal fatto di avere uno scheletro carbossilico steroideo (sterone) e dall’essere associati al processo di muta, altrimenti noto come ecdisi. La ragione della loro esistenza nelle piante (essendo un ormone degli insetti), come precedentemente accennato, è che proteggono le piante dagli insetti non adattati alla loro tolleranza, e quindi sono una fitoalessina.[19-13]
Gli Ecdysteroidi sono composti ormonali coinvolti nel comportamento sessuale degli insetti, nella muta e nella metamorfosi. Gli Ecdysteroidi presentano una somiglianza strutturale con il Testosterone e sono considerati il composto simile al Testosterone più attivo negli insetti.
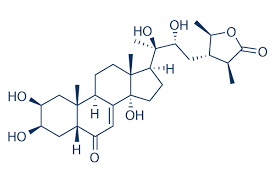
Di seguito è riportata la struttura generale della famiglia degli Ecdysteroidi:
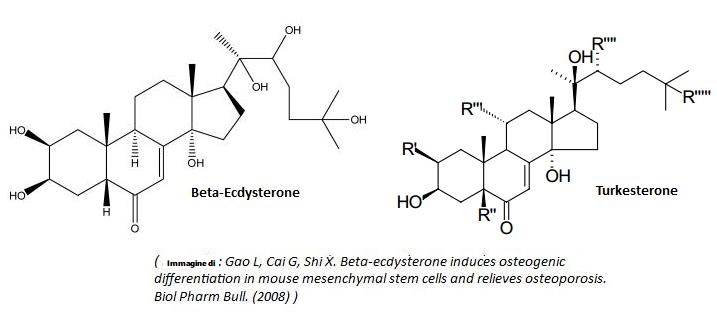
Sebbene dal 2001 siano noti oltre 200 Ecdysteroidi,[8] e ne siano stati registrati fino a 463[19], la maggior parte di essi non è bioattiva se ingerita per via orale.[20] Tra quelli più comuni, sia nella ricerca che nell’assunzione per via orale, vi sono:
- Ecdysone
- Ecdysterone e beta-ecdysterone
- 20-idrossiecdysone
- Turkesterone
- Integristerone A
- 24(28)-deidramakisterone A
- Viticosterone E
- Sileneoside A e C
- Ponasterone A
- Citasterone
Farmacologia [biodisponibilità, farmacocinetica, interazione cellulare/recettoriale e metabolismo]:
In uno studio, utilizzando Ecdysteroidi da 0,2mg/kg di peso corporeo (come ecdysone e 20-idrossidysone), l’Ecdysone sembrava avere un’emivita di eliminazione di 4 ore e il 20-idrossidysone un’emivita di eliminazione di 9 ore nell’uomo.[20] Non è nota un’emivita attiva nell’uomo.
Tuttavia, i modelli murini mostrano un’emivita di 8,15 minuti per il 20-idrossidysone quando viene iniettato alla dose di 50mg/kg di peso corporeo nella vena caudale[21] e risultati simili sono stati replicati con il 20-idrossidysone altrove.[22] È stata notata anche un’emivita di 48 minuti (per l’Ecdysteroide Ponasterone A) quando viene iniettato alla dose di 750g.[23]
Nella drosofilia è stato clonato un recettore citoplasmatico, denominato DopEcR, che si lega agli Ecdysteroni e alla Dopamina.[24] È stato teorizzato che alcuni dei meccanismi d’azione avvengano attraverso questo recettore e siano di natura non genomica (non influenzano il nucleo della cellula).[25][26] Tra i possibili effetti non genomici vi è l’afflusso di ioni calcio che inducono la fosforilazione di Akt, di cui parlerò più avanti.
Si ipotizzano anche recettori nucleari (nei mammiferi) della superfamiglia dei recettori nucleari[27]. Il recettore dell’Ecdysterone si dimerizza con i recettori Ultraspiracle (USP) negli insetti per influenzare i geni, ma negli esseri umani deve dimerizzare con il recettore RXR.[20] Sebbene negli insetti l’USP possa dimerizzare con un’ampia varietà di recettori nucleari, nei mammiferi deve formarsi un complesso EcR:RXR perché si verifichino gli effetti. [28] Il legame dell’EcR con recettori non RXR non produce effetti genetici nei vertebrati.[29] È stato tuttavia osservato che l’RXR è un partner “riluttante” per l’EcR e che per la segnalazione genetica tramite EcR:RXR è necessario un relativo eccesso di RXR; questo è stato menzionato in uno studio[20] in relazione a un altro che ha indagato un modello in vitro su una linea cellulare di ovaio di criceto cinese.[30]

Gli Ecdysteroidi, in particolare il 20-idrossiecdysone e il Pinnatasterone, sono stati chiamati in causa come inibitori delle pompe di efflusso della P-glicoproteina e possono interagire con altri farmaci che vengono metabolizzati ampiamente dalla P-glicoproteina, come la Berberina o l’Icariina.[31]
Nei topi (e nell’uomo) l’escrezione avviene sia per via fecale che per via urinaria. Alcuni studi suggeriscono che la via fecale sia favorita, in quanto gli Ecdysteroidi vengono raccolti dal fegato e poi espulsi negli acidi biliari[32][33], tuttavia almeno uno studio osserva che entrambe le vie possono essere ugualmente importanti,[21] sebbene quest’ultimo studio abbia utilizzato un’iniezione di Ecdysteroidi da 50mg/kg di peso corporeo.
Nell’esaminare i metaboliti fecali, sono stati notati il 4-deossicedisone e composti con un anello B completamente ridotto.[34] In una review è stato osservato che questo metabolismo “ricorda la riduzione epatica del 4-en-3-one sull’anello-A degli ormoni steroidei dei vertebrati”.[20] Quando la scissione della catena laterale avviene tra il C20 e il C22, possono risultare i metaboliti Poststerone e 12-deossicedisone (dal 20-idrossicedysone). [Nei ratti è stato osservato anche un nuovo metabolita, il 2β,3β,6α,22R,25-pentaidrossi-5β-colest-8(14)-ene.[22] Infine, il metabolita 14-deossi-20-ecdysone (osservato come metabolita primario nelle urine umane) può avere interazioni con la microflora intestinale, poiché è noto che la microflora causa la deidrossilazione dei composti steroidei.[35]
Nell’uomo, l’escrezione urinaria di Ecdysterone tende a risultare in uno dei tre composti: l’Ecdysterone in forma invariata, il 2-deossiacidysterone o il Deossiacidysone. Il principale metabolita urinario, con una percentuale del 99,34%, è il Desossiacidysone a 21 ore dall’ingestione di 20mg di Ecdysterone.[36] Un’escrezione urinaria bifasica del composto progenitore è stata notata anche con l’analisi delle urine a 68 ore.[36] Questi metaboliti si trovano anche nelle urine dei ratti.[37]
In realtà non ci sono molte informazioni su questo argomento che possano essere considerate “conclusive”. Sembra che ci sia un’ampia varietà di metaboliti che non sono stati studiati e che il 20-HE persista più a lungo negli esseri umani che nei topi (4,1 ore contro 8,15 m) o che la sua risposta sia dipendente dalla dose. Al momento non si hanno risposte a proposito.
Interazioni neurologiche:
L’Ecdysterone è in grado di aumentare l’induzione enzimatica sia dell’acetilcolinesterasi[38] che della glutammato decarbossilasi.[39] Questi effetti sono a valle della capacità degli Ecdysteroidi di aumentare la sintesi proteica, in quanto l’aumento della sintesi proteica (attraverso l’aumento dell’efficacia dell’mRNA, come ipotizzato da Uchiyama & Otaka[40][41]) si applica ai tessuti proteici (muscolo scheletrico, proteine degli organi) e agli enzimi. L’aumento del glutammato decarbossilato è stato misurato al 25-30% in vivo dopo 2,5-50ug/kg di peso corporeo, anche se la dipendenza dalla dose non era chiara.[39]
Esercita inoltre alcuni effetti protettivi contro le tossine neurologiche.[42][43]
Salute cardiovascolare:
I Fitoecdysoni (la famiglia di cui fa parte l’Ecdysterone) sono promettenti come agenti di riduzione del Colesterolo[44], probabilmente attraverso una maggiore conversione del colesterolo in acidi biliari. Questi effetti sono stati riscontrati a 2,5mg/kg di peso corporeo.
L’Ecdysterone (termine intercambiabile con 20-idrossiecdysterone, o 20-HE) sembra essere in grado di sopprimere la formazione epatica di glucosio e quindi di abbassare i livelli di zucchero nel sangue indipendentemente dalla secrezione di Insulina e dai livelli sierici.[45] La soppressione del metabolismo del glucosio sembra provenire dalla fosfoenolpiruvato carbossichinasi e dalla glucosio-6-fosfatasi, oltre a indurre la fosforilazione di Akt nelle cellule epatiche.[46]
Quando viene somministrato ai ratti alla dose di 10mg/kg di peso corporeo, il composto correlato 20-idrossidysone è in grado di esercitare effetti antidiabetici e antiobesogeni in modelli di obesità animale, suggerendo che potrebbe esercitare questi stessi effetti nell’uomo.[46] Questi cambiamenti hanno anche portato a una maggiore secrezione di Adiponectina da parte degli adipociti di ratto.[46] È stato dimostrato che in altri modelli passati esercita proprietà antidiabetiche simili, indipendentemente dal metodo di ingestione/iniezione.[47][48]
Muscolo-scheletrico e prestazioni sportive:
La somministrazione di Ecdysterone (per via sottocutanea o endovenosa), a circa 5mg/kg di peso corporeo, sembra essere in grado di indurre la sintesi proteica in organi animali come il fegato[49][50] o il muscolo scheletrico.[51] Ciò è probabilmente dovuto a un aumento dell’efficienza di traduzione dell’mRNA piuttosto che a un aumento della sintesi dell’mRNA (trascrizione).[45] Inoltre, gli Ecdysteroidi possono essere in grado di aumentare l’incorporazione della leucina nelle cellule a una dose di 5mg/kg di peso corporeo (lo studio ha riguardato il fegato).[52]
Studi in vitro su cellule muscolari (con 20-idrossiecdysone e Turkesterone) hanno rilevato miglioramenti statisticamente significativi nella sintesi proteica in modo dipendente dalla dose a partire da 0,08nM, con un picco a 0,1nM con una sintesi proteica superiore del 110-120% rispetto al controllo e un plateau a concentrazioni comprese tra 1 e 10nM. [Il metabolita del 20-idrossisterone, il Rubrosterone, sembra essere altrettanto potente se si considera il fegato di topo.[49]
In studi comparativi diretti, l’Ecdysteroide chiamato “Turkesterone” sembra essere più potente rispetto agli altri Ecdysteroidi studiati[53][15], seguito dal Cyasterone e poi dal 20-idrossisterone.[54][22]
Rispetto al controllo, il Turkesterone ha aumentato la crescita dei ratti (sulla base di mg/die) del 63,5%, l’Ecdysterone del 51,9%, il 2-deossiecdysterone del 21,2% e l’alfa-ecdystone del 19,2%. Questo studio ha utilizzato il Metilandrostenediolo (51,9%) e il Nerobol (57,7%) come composti di confronto, anche se gli effetti del Nerobol [Methandienone] erano più localizzati al muscolo scheletrico, mentre gli Ecdysteroidi avevano un aumento della sintesi proteica sistemica (organo e muscolo).[15] Gli Ecdysteroidi in questo studio non hanno soppresso né causato lo sviluppo della prostata o delle vescicole seminali, e non hanno avuto effetti uterotropi nei ratti femmina; il peso del timo è aumentato del 23-35%, mentre è diminuito del 20% con il Nerobol. Le dosi utilizzate in questo studio sono state di 5mg/kg di peso corporeo per tutti gli Ecdysteroidi e di 10mg/kg di peso corporeo per il Metilandrostenediolo e il Nerobol.[15]
Per quanto riguarda i meccanismi d’azione, gli Ecdysteroidi sembrano in grado di provocare un rapido afflusso di Ca2+ nei miociti che porta alla fosforilazione di Akt e quindi alla sintesi proteica. [Questo effetto si verifica dopo 10 minuti di incubazione ed è inibito dagli inibitori della PI3K, come già visto in altri studi, ma anche dagli inibitori della GPRC e della PLC; inoltre, quando le cellule vengono private del calcio intracellulare, Akt non viene fosforilato e il legame del calcio libero con l’EGTA abbassa la sintesi proteica dal 16% all’8%.[55] Il calcio di per sé può essere un importante mediatore di Akt e della sintesi proteica[56-51], e gli Ecdysteroidi sembrano funzionare in modo vicario attraverso il Ca2+ e Akt.[57]
L’afflusso di calcio aumenta la fosforilazione di Akt di oltre 3 volte a una concentrazione di 0,1uM, con una dose-risposta decrescente fino a 5 volte a 1-10uM.[55] L’effetto di 1uM di 20-idrossiecdysterone su Akt ha raggiunto il picco a 2-4 ore, ma è rimasto superiore al valore di base fino a 24 ore.[55]
Nella discussione di uno studio[24] è stato anche notato che la “coda” degli Ecdysteroidi (γ-idrossi-γ-metilpentanoato), se separata dallo scheletro carbossilico steroideo, assomiglia al metabolita anabolizzante della Leucina HMB (beta-idrossi-metilbutirrato).

Studi in vivo hanno rilevato un aumento dell’anabolismo in un’ampia varietà di animali, come ratti e topi,[50][58][59] suini[60] e quaglie.[61] Gli effetti sul miglioramento della forza sembrano essere indipendenti dall’attività, come valutato dal tempo di nuoto forzato nei ratti che migliora senza un allenamento costante. Alcuni studi passati (precedenti al 2000) suggeriscono che possa aumentare la sintesi proteica anche negli esseri umani.[62] Anche nei ratti sono stati rilevati miglioramenti delle prestazioni.[63]
Nelle pecore, una dose orale di 0,02mcg/kg di Ecdysteroidi al giorno è stata in grado di aumentare il tasso di crescita corporea e la produzione di lana[22] ed è stata più evidente con un apporto nutritivo più scarso. Una dose altrettanto ridotta di 0,4 mg/kg di peso corporeo è stata in grado di aumentare la ritenzione di azoto e preservare la massa magra (al 112-116% del controllo) quando l’assunzione di cibo è stata ridotta dell’11-17% nei suini.[64]
L’aumento dell’attività della fosfatasi alcalina indotta dall’Ecdysterone sembra avvenire attraverso il recettore degli estrogeni.[65] Attraverso questo recettore, anche l’attività dei geni reporter degli estrogeni viene aumentata dall’Ecdysterone.
L’aumento dell’attività osservato nell’espressione del collagene di tipo I, dell’osteocalcina e di Runx2 non sembra avvenire attraverso il recettore degli estrogeni.[66]
Interazioni ormonali:
Al momento è stato condotto un solo studio sull’uomo con l’Ecdysterone. Dosato a 200 mg al giorno, non è stato osservato alcun risultato nei maschi che si allenavano alla resistenza per quanto riguarda il testosterone totale e libero o i cambiamenti nella composizione corporea rispetto al placebo.[64] Quando è stato testato il legame con il recettore degli androgeni, il 20-idrossiecdysterone non sembra avere alcuna affinità di legame e quindi non può attivare il recettore degli androgeni.[67]
Detto questo, nonostante l’assenza di influenza sul Testosterone stesso, l’Ecdysterone potrebbe essere in grado di esercitare effetti simili al testosterone attraverso le vie di trasduzione del segnale (anche se il meccanismo esatto non è ancora noto); un’azione che in definitiva ha lo stesso significato biologico del testosterone.[68]
Non ci sono molte prove, oltre a quelle in vitro, che suggeriscano l’utilità dell’Ecdysterone per la sintesi proteica muscolare o per l’aumento della forza.[69]
Quando è stato testato in vitro in C2C12, 1µM di 20-idrossiecdysone (20-HE) ha aumentato il diametro dei miotubi in modo indipendente dal recettore degli androgeni; sia i corticosteroidi che i bloccanti del recettore degli estrogeni hanno impedito al 20-HE di promuovere la crescita muscolare. [52] In seguito a ulteriori test, il 20-HE sembra attivare sia la variante alfa del recettore degli estrogeni (ERα; EC50 di 25,7nM) sia la variante beta (ERβ; EC50 13nM) e si è riscontrato che 10nM di 20-HE promuove la crescita muscolare attraverso ERβ.[52]

Quando è stato testato in vitro, il 20-idrossiecdysone sembra promuovere l’ipertrofia delle cellule muscolari agendo sul recettore beta degli estrogeni. Questa molecola sembra agire anche sul recettore alfa, e quando entrambi i recettori agiscono contemporaneamente la cellula muscolare sperimenta comunque l’ipertrofia.
Interazioni con i processi ossidativi:
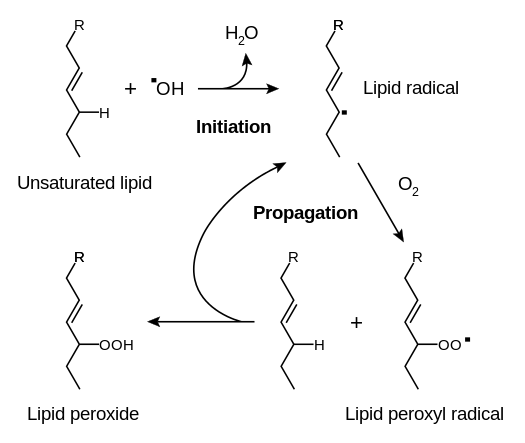
L’Ecdysterone esercita anche effetti protettivi contro la perossidazione lipidica da parte dei radicali liberi, ottenendo uno status di antiossidante.[70] Questo effetto è stato osservato a una dose molto bassa di 0,1mg/kg di peso corporeo ed è risultato più potente della vitamina D su base molecolare.
Sistemi organici periferici:
L’Ecdysterone, alla dose di 5mg/kg di peso corporeo, è in grado di ripristinare la normale velocità di filtrazione glomerulare e di sopprimere l’albuminuria nei ratti trattati con una miscela nefrotossica[46] e può alleviare i sintomi dell’uremia associata al danno epatico.[58]
Come discusso nella sezione sul metabolismo dei grassi, l’Ecdysterone è in grado di aumentare il tasso di secrezione biliare e di migliorare la rigenerazione del fegato dopo un danno da tossina (Eliotrina).[46]
Per quanto riguarda la pelle, l’Ecdysterone sembra essere in grado di promuovere la differenziazione dei cheratinociti e di accelerare piccole ferite e ustioni quando viene applicato esternamente.[46]
Longevità e aspettativa di vita:
Gli Ecdysteroidi sono uno dei due ormoni degli insetti (l’altro è l’ormone giovanile) che sembrano essere coinvolti nella durata della vita di questi animali, con l’Ecdysterone come agente che aumenta la durata della vita.[71][72] La trasfezione di Drosophilia con un recettore per l’Ecdysone aumenta la durata della vita.[72] Tuttavia, gli studi sull’uomo sono inesistenti e gli altri modelli animali molto preliminari.

Sicurezza e tossicità:
Gli Ecdysteroidi, nel complesso, sono abbastanza sicuri per l’ingestione. I benefici sembrano manifestarsi a dosi intorno ai 10mg/kg di peso corporeo, mentre la tossicità accertata nei mammiferi (roditori) è di 6400mg/kg di peso corporeo se iniettati e >9000mg/kg di peso corporeo se consumati per via orale.[42][41]
Tuttavia, i dati sull’uomo sono pochi e non sempre aventi un design ottimale al fine di poter fare una analisi oggettiva dei dati in essi riportati. Al momento però, sono maggiori le segnalazioni di mancanza totale di effetti piuttosto che di conseguenze negative dopo la loro somministrazione orale o transdermica. Gli effetti avversi negativi denunciati da alcuni utilizzatori sono stati dolori gastro-intestinali, reflusso e dissenteria comparse dopo l’inizio della somministrazione orale e cessate poco dopo l’interruzione della stessa. In alcuni casi, la somministrazione topica ha comportato la comparsa di rush cutanei nella zona di applicazione.
Bisogna comunque ricordare che ad ogni azione corrisponde una reazione uguale e contraria, nel mondo fisico e umano. Indi per cui, pretendere modifiche dell’omeostasi senza potenziali conseguenze indesiderate è da idioti illusi.
Conclusioni:
Se avete compreso bene quanto riportate, vi sarete resi conto che mancano interventi validi sull’uomo. Quindi, parlare di assenza di effetti collaterali o efficacia certa è solamente espressione di becera ignoranza venduta ad un pubblico in perenne ricerca della “pillola magica”. E, a dirla tutta, dopo anni di dati raccolti, vi posso dire che non è proprio il caso degli Ecdysteroidi l’essere la panacea di tutti i mali.
Vorrei che consideraste il fatto che il 20-idrossiecdysone (20HE), per fare un esempio, è risultato limitato nella segnalazione muscolo-scheletrica e epatica della protein chinasi B/Akt-mTOR1 nei topi. Anche la biodisponibilità del 20HE, consumato da solo o con la Leucina, è rimasta bassa a tutte le dosi ingerite.[https://pubmed.ncbi.nlm.nih.gov/26584207/]
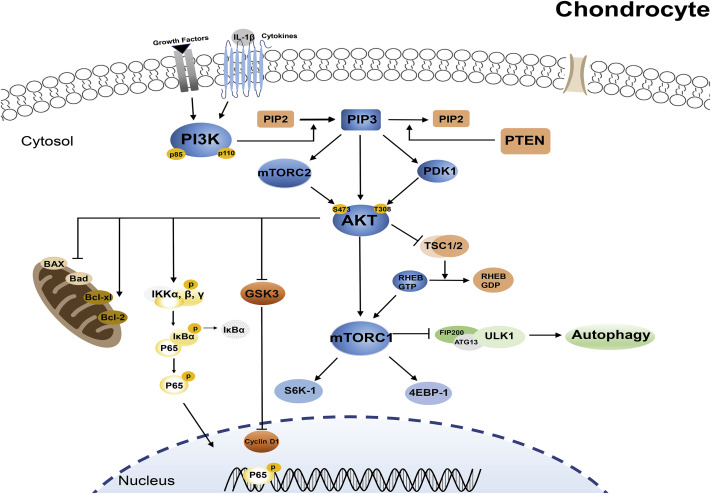
Sebbene esista uno studio svolto su 46 soggetti (principianti) affermi che la somministrazione di Ecdysterone ha portato ad osservare un aumento significativo della massa muscolare e aumenti significativamente più pronunciati nelle prestazioni di panca ad una ripetizione. Come se non bastasse, i ricercatori hanno affermato che non è stato rilevato alcun aumento dei biomarcatori di tossicità epatica o renale. La loro conclusione? Ché i dati raccolti sottolineano l’efficacia di un’integrazione di Ecdysterone rispetto alle prestazioni sportive, suggerendo fortemente l’inclusione dell’Ecdysterone nell’elenco delle sostanze e dei metodi proibiti nello sport nella classe S1.2 “altri agenti anabolizzanti”. Lo studio in questione è del 2019, ma rappresenta un eccezione in una letteratura contraria o neutra. Basta leggere bene lo studio per trovarne le “crepe di design” le quali lo rendono ottimo per i venditori ma pessimo per chiarire oggettivamente le cose.
La questione recettoriale mette ancora di più in difficoltà il valutare le possibili e reali potenzialità ipertrofiche negli esseri umani. Sebbene la struttura degli Ecdysteroidi sia in qualche modo simile a quella degli ormoni steroidei dei vertebrati, esistono diverse differenze strutturali tra i due gruppi di steroidi. Nonostante queste differenze strutturali essenziali, gli Ecdysteroidi esercitano nei vertebrati numerosi effetti simili a quelli degli steroidi ormonali dei vertebrati e possono servire come efficaci agenti anabolizzanti, epatoprotettivi, immunoprotettivi, antiossidanti e ipoglicemizzanti. Gli Ecdysteroidi non si legano ai recettori steroidei citosolici, ma è probabile che influenzino le vie di trasduzione del segnale, come gli steroidi anabolizzanti, probabilmente attraverso recettori legati alla membrana. L’applicazione di fitoecdisteroidi sarebbe anche una promettente alternativa all’uso di steroidi anabolizzanti-androgeni per l’apparente mancanza di effetti avversi, se non fosse che le conoscenze in nostro possesso sui reali effetti quando somministrati agli uomini siano molto limitate. Chiunque affermi con certezza che l’applicazione dei fitoecdisteroidi porti all’aumento delle dimensioni muscolari dovrebbe sapere che per poter affermare con certezza ciò è necessaria una ricerca rigorosa, poiché non è ancora disponibile una spiegazione citologica adeguata oltre a prove incontrovertibili.
E’ ormai risaputo che gli estrogeni hanno una serie di effetti metabolici sul muscolo scheletrico. Quando gli individui di sesso femminile perdono gli estrogeni attraverso l’ovariectomia, la funzione mitocondriale, la microviscosità della membrana e le attività del complesso I e I+III diminuiscono (Torres et al., 2018). La perdita di estrogeni provoca anche un aumento della produzione mitocondriale di H2O2 (Valencia et al., 2016), una diminuzione dei livelli di proteine antiossidanti come la glutatione perossidasi, la catalasi e la superossido dismutasi (Baltgalvis et al., 2010; Valencia et al., 2016) e un’alterata sensibilità all’insulina (Torres et al., 2018). Questi effetti sono dovuti alla perdita di estrogeni, poiché il ripristino di livelli normali di questi ormoni ripristina la redox cellulare e l’omeostasi del glucosio nel muscolo scheletrico (Spangenburg et al., 2012; Camporez et al., 2013; Torres et al., 2018).
Al di là dei ruoli metabolici, gli estrogeni sono chiaramente benefici per la massa e la forza muscolare, almeno nei modelli animali (McClung et al., 2006; Kitajima e Ono, 2016). Si ipotizza, quindi, che in assenza di estrogeni, il muscolo è più soggetto a lesioni e questo limita la ricrescita (McClung et al., 2006). Sulla base di questi e altri dati, Enns e Tiidus (2010) hanno proposto che gli estrogeni potrebbero stabilizzare la matrice extracellulare o agire come antiossidanti per ridurre le lesioni muscolari; tuttavia, l’effetto degli estrogeni sul muscolo umano non è stato definito con altrettanta chiarezza perché le variazioni di estrogeni sono transitorie o associate a differenze confondenti di età, livello di forma fisica e tipo e intensità di esercizio (Enns e Tiidus, 2010). Infine, molti studi che cercano di capire il ruolo degli estrogeni sulla funzione muscolare si concentrano sulle differenze di sesso, che vanno ben oltre le semplici variazioni dei livelli ormonali.
Una delle differenze muscolo-scheletriche meglio caratterizzate tra uomini e donne è il tasso di rottura del legamento crociato anteriore (ACL). Le rotture del legamento crociato anteriore si verificano con una frequenza da 2 a 8 volte maggiore tra le atlete rispetto agli uomini (Arendt e Dick, 1995; Adachi et al., 2008). Quando la concentrazione di estrogeni aumenta durante il ciclo mestruale, aumenta anche la lassità del ginocchio (Shultz et al., 2010, 2011, 2012a). Infatti, è stato riscontrato che la lassità del ginocchio è aumentata tra 1 e 5 mm tra il primo giorno delle mestruazioni e il giorno successivo all’ovulazione, a seconda dei livelli di estrogeni. Infine, Park et al. hanno riscontrato una diminuzione del 17% della rigidità del ginocchio durante la fase ovulatoria, con una variazione della lassità del ginocchio da 13,35 ± 2,53 mm durante la fase follicolare a 14,43 ± 2,60 mm durante l’ovulazione (Park et al., 2009). Al contrario, Carcia et al. (2004) non hanno riscontrato variazioni nello spostamento del ginocchio in relazione al ciclo; tuttavia, è importante notare che questi autori hanno utilizzato la lunghezza del ciclo auto-riferita per stimare la fase mestruale, mentre gli altri studi hanno misurato direttamente i livelli di estrogeni in concerto con la lassità del ginocchio. Poiché Myer et al. (2008) hanno dimostrato che per ogni aumento di 1,3 mm dello spostamento del ginocchio, il rischio di lesione del legamento crociato anteriore aumenta di 4 volte, l’aumento della lassità del ginocchio riportato da Deie, Park e Shultz potrebbe spiegare il tasso di rottura del legamento crociato anteriore da 2 a 8 volte superiore nelle donne (Arendt e Dick, 1995; Adachi et al., 2008). Quindi, se gli estrogeni diminuiscono l’attività della lisil-ossidasi nei tendini, ci si aspetterebbe che questo diminuisca la rigidità dei tendini e quindi l’incidenza di lesioni ai muscoli associati. In effetti, come già detto, le donne subiscono meno lesioni muscolari degli uomini (Hägglund et al., 2009; Edouard et al., 2016).
È chiaro che gli estrogeni hanno un effetto significativo sulla funzione muscolo-scheletrica. In passato, gran parte della ricerca si è concentrata sulla forte connessione tra estrogeni e ossa. Tuttavia, recentemente l’effetto degli estrogeni su altri tessuti muscolo-scheletrici, come muscoli, tendini e legamenti, è diventato oggetto di maggiori ricerche. Questi studi chiariscono che gli estrogeni migliorano la proteostasi muscolare e aumentano il contenuto di collagene dei tendini; tuttavia, i benefici sull’osso e sul muscolo avvengono al prezzo di una minore rigidità del tessuto connettivo. Tuttavia, con l’aumento delle donne che praticano sport, è chiaro che questi effetti fisiologici degli estrogeni contribuiscono a diminuire la potenza e le prestazioni e rendono le donne più inclini a subire infortuni catastrofici ai legamenti.
Ma perchè ho riportato tutto ciò? Semplicemente per farvi capire alcuni punti essenziali:
- La mancanza di interazione con i AR da parte degli Ecdysteroidi e la loro affinità nei mammiferi a carico dei ERα e ERβ;
- tale affinità ha il potenziale di promuove la crescita muscolare attraverso il ERβ;
- tale interazione, però, non è mai stata accuratamente monitorata e quantificata nell’uomo, di conseguenza non si ha alcuna certezza che il potenziale sia statisticamente significativo;
- date tali caratteristiche sembrerebbe più probabile che l’assunzione di Ecdysteroidi possa avere un qualche effetto protettivo su tendini e legamenti oltre che sulla matrice ossea;
- la scarsa biodisponibilità che caratterizza gli Ecdysteroidi assunti per via orale renderebbero molto difficile il mantenimento di una soglia ematica efficace;
- notando le caratteristiche strutturali degli Ecdysteroidi, sebbene abbiano mostrato una emivita di 8h circa in media, non suggerisce un potenziale di legame ormone-recettore elevato con conseguente ulteriore dubbio del grado di efficacia;
- l’affinità con i ER potrebbe, se si dovessero raggiungere soglie ematiche sufficienti, indurre un feedback negativo a livello ipotalamico-ipofisario con conseguente calo della secrezione di LH e FSH e alterazioni consequenziali di Testosterone, DHT ed Estradiolo [vedi alterazioni nel comportamento sessuale, depressione, letargia ecc…];
- si è proposta l’ipotesi che l’interazione con i ER a livello ipotalamico-ipofisario possa dare un effetto simile a quello riscontrato con i SERM. Tuttavia tale ipotesi risulta ad oggi indimostrata;
- l’affinità con i ER potrebbe, se si dovessero raggiungere soglie ematiche sufficienti, causare la comparsa di effetti collaterali estrogeno-dipendenti, sebbene l’iterazione con il ERβ, in specie l’isoforma ERβ1, abbia mostrato un attività protettiva antitumorale che, secondo alcuni, potrebbe avere una valenza preventiva sullo sviluppo di ginecomastia. Anche questa rimane una ipotesi senza dimostrazione;
- possibile interazione ad esito sconosciuto con l’asse GH/IGF-1 mediata dal legame con il ERα e ERβ a livello epatico.
Siete confusi? Non dovreste. Infondo, adesso, avete gli strumenti di logica per valutare quanto possa valere la pena acquistare un integratore contenente Ecdysteroidi. Si tratta di un azzardo che potrebbe non limitarsi solo alla componente economica visti i dosaggi necessari per poter sperare (forse) di osservare miglioramenti nella composizione corporea e/o nelle prestazioni atletiche.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
- de Loof A (2006). “Ecdysteroids: the overlooked sex steroids of insects? Males: the black box”. Insect Science.
- Krishnakumaran A, Schneiderman HA (December 1970). “Control of molting in mandibulate and chelicerate arthropods by ecdysones”. The Biological Bulletin.
- Margam VM, Gelman DB, Palli SR (June 2006). “Ecdysteroid titers and developmental expression of ecdysteroid-regulated genes during metamorphosis of the yellow fever mosquito, Aedes aegypti (Diptera: Culicidae)”. Journal of Insect Physiology.
- “Ecdysteroids Information”. Examine.com. Retrieved 27 May 2015.
- Mykles DL (November 2011). “Ecdysteroid metabolism in crustaceans”. The Journal of Steroid Biochemistry and Molecular Biology.
- Dinan L (June 2001). “Phytoecdysteroids: biological aspects”. Phytochemistry.
- Dinan L, Savchenko T, Whiting P (July 2001). “On the distribution of phytoecdysteroids in plants”. Cellular and Molecular Life Sciences.
- Dinan L. Phytoecdysteroids: biological aspects. Phytochemistry. (2001)
- Wu JJ, et al. Steroidal saponins and ecdysterone from Asparagus filicinus and their cytotoxic activities. Steroids. (2010)
- Schmelz EA, et al. Interactions between Spinacia oleracea and Bradysia impatiens: a role for phytoecdysteroids. Arch Insect Biochem Physiol. (2002)
- Ecdysteroids from Chenopodium quinoa Willd., an ancient Andean crop of high nutritional value.
- Taxonomic distribution of phytoecdysteroids in seeds of members of the chenopodiaceae.
- A new ecdysteroid and other constituents from two Dioscorea species.
- Comparative experimental investigation of the anabolic activity of phytoecdysteroids and steranabols.
- Suksamrarn A, Kumpun S, Yingyongnarongkul BE. Ecdysteroids of Vitex scabra stem bark. J Nat Prod. (2002)
- Sena Filho JG, et al. Ecdysteroids from Vitex species: distribution and compilation of their 13C-NMR spectral data. Chem Biodivers. (2008)
- dos Santos TC, Delle Monache F, Leitão SG. Ecdysteroids from two Brazilian Vitex species. Fitoterapia. (2001)
- Suksamrarn A, Promrangsan N, Jintasirikul A. Highly oxygenated ecdysteroids from Vitex canescens root bark. Phytochemistry. (2000)
- Ecdybase: The Ecdysone Database.
- Practical uses for ecdysteroids in mammals including humans: an update.
- Pharmacokinetics of ecdysterone in experiments.
- Kumpun S, et al. The metabolism of 20-hydroxyecdysone in mice: relevance to pharmacological effects and gene switch applications of ecdysteroids. J Steroid Biochem Mol Biol. (2011)
- Albanese C, et al. Sustained mammary gland-directed, ponasterone A-inducible expression in transgenic mice. FASEB J. (2000)
- Rapid, Nongenomic Responses to Ecdysteroids and Catecholamines Mediated by a Novel Drosophila G-Protein-Coupled Receptor.
- Iga M, Iwami M, Sakurai S. Nongenomic action of an insect steroid hormone in steroid-induced programmed cell death. Mol Cell Endocrinol. (2007)
- Schlattner U, et al. Non-genomic ecdysone effects and the invertebrate nuclear steroid hormone receptor EcR–new role for an “old” receptor. Mol Cell Endocrinol. (2006)
- Laudet V. Evolution of the nuclear receptor superfamily: early diversification from an ancestral orphan receptor. J Mol Endocrinol. (1997)
- Thomas HE, Stunnenberg HG, Stewart AF. Heterodimerization of the Drosophila ecdysone receptor with retinoid X receptor and ultraspiracle. Nature. (1993)
- Yao TP, et al. Drosophila ultraspiracle modulates ecdysone receptor function via heterodimer formation. Cell. (1992)
- Functional transfer of an elementary ecdysone gene regulatory system to mammalian cells: Transient transfections and stable cell lines.
- Nguyen VT, et al. Selective modulation of P-glycoprotein activity by steroidal saponines from Paris polyphylla. Fitoterapia. (2009)
- Hikino H, Oizumi Y, Takemoto T. Absorption, distribution, metabolism, and excretion of insect-metamorphosing hormone ecdysterone in mice. Yakugaku Zasshi. (1972)
- Lafont R, Girault JP, Kerb U. Excretion and metabolism of injected ecdysone in the white mouse. Biochem Pharmacol. (1988)
- Girault JP, Lafont R, Kerb U. Ecdysone catabolism in the white mouse. Drug Metab Dispos. (1988)
- Winter J, Bokkenheuser VD. Bacterial metabolism of natural and synthetic sex hormones undergoing enterohepatic circulation. J Steroid Biochem. (1987)
- Tsitsimpikou C, et al. Study of excretion of ecdysterone in human urine. Rapid Commun Mass Spectrom. (2001)
- Study of ecdysterone metabolites isolated from rat urine.
- Catalan RE, et al. Ecdysterone induces acetylcholinesterase in mammalian brain. Comp Biochem Physiol C. (1984)
- Chaudhary KD, Lupien PJ, Hinse C. Effect of ecdysone on glutamic decarboxylase in rat brain. Experientia. (1969)
- Otaka T, et al. Stimulatory effect of insect-metamorphosing steroids from ferns on protein synthesis in mouse liver. Chem Pharm Bull (Tokyo). (1969)
- Otaka T, Okui S, Uchiyama M. Stimulation of protein synthesis in mouse liver by ecdysterone. Chem Pharm Bull (Tokyo). (1969)
- Protective effect of ecdysterone on amnesia induced by diazepam and alcohol 1.
- Dinan L, Lafont R. Effects and applications of arthropod steroid hormones (ecdysteroids) in mammals. J Endocrinol. (2006)
- Mironova VN, et al. Hypocholesterolemic effect of phytoecdysones during experimental hypercholesterolemia in rats. Vopr Med Khim. (1982)
- Chen Q, Xia Y, Qiu Z. Effect of ecdysterone on glucose metabolism in vitro. Life Sci. (2006)
- Kizelsztein P, et al. 20-Hydroxyecdysone decreases weight and hyperglycemia in a diet-induced obesity mice model. Am J Physiol Endocrinol Metab. (2009)
- Matsuda H, Kawaba T, Yamamoto Y. Pharmacological studies of insect metamorphotic steroids. Nihon Yakurigaku Zasshi. (1970)
- Hypoglycemic Activity of the Total Ecdysteroid Extract from Ajuga turkestanica.
- Otaka T, et al. Stimulatory effect of insect-metamorphosing steroids from Achyranthes and Cyathula on protein synthesis in mouse liver. Chem Pharm Bull (Tokyo). (1968)
- The Effect of Ecdysterone on the Biosynthesis of Proteins and Nucleic Acids in Mice.
- Parr MK1, et al. Estrogen receptor beta is involved in skeletal muscle hypertrophy induced by the phytoecdysteroid ecdysterone. Mol Nutr Food Res. (2014)
- Syrov VN, Kurmukov AG, Sakhibov AD. Effect of turkesterone and nerobol on the activity of the protein synthesizing system of mouse liver. Vopr Med Khim. (1978)
- Syrov VN. Mechanism of the anabolic action of phytoecdisteroids in mammals. Nauchnye Doki Vyss Shkoly Biol Nauki. (1984)
- Syrov VN, Khushbaktova ZA. Experimental study of pharmacotherapeutic effect of phytoecdisteroids and nerobol in toxic liver damage. Eksp Klin Farmakol. (2001)
- Gorelick-Feldman J, Cohick W, Raskin I. Ecdysteroids elicit a rapid Ca2+ flux leading to Akt activation and increased protein synthesis in skeletal muscle cells. Steroids. (2010)
- Constantino S, et al. The ecdysone inducible gene expression system: unexpected effects of muristerone A and ponasterone A on cytokine signaling in mammalian cells. Eur Cytokine Netw. (2001)
- Syrov VN, Kurmukov AG. Anabolic activity of phytoecdysone-ecdysterone isolated from Rhaponticum carthamoides (Willd.) Iljin. Farmakol Toksikol. (1976)
- 20-Hydroxyecdysone increases fiber size in a muscle-specific fashion in rat.
- Effect of 20-hydroxyecdysone on the protein synthesis in pigs.
- Stimulation of growth and development in Japanese quails after oral administration of ecdysteroid-containing diet.
- Chermnykh NS, et al. The action of methandrostenolone and ecdysterone on the physical endurance of animals and on protein metabolism in the skeletal muscles. Farmakol Toksikol. (1988)
- Azizov AP, Seĭfulla RD. The effect of elton, leveton, fitoton and adapton on the work capacity of experimental animals. Eksp Klin Farmakol. (1998)
- Wilborn CD, et al. Effects of methoxyisoflavone, ecdysterone, and sulfo-polysaccharide supplementation on training adaptations in resistance-trained males. J Int Soc Sports Nutr. (2006)
- Gao L, Cai G, Shi X. Beta-ecdysterone induces osteogenic differentiation in mouse mesenchymal stem cells and relieves osteoporosis. Biol Pharm Bull. (2008)
- Spindler KD, et al. Ecdysteroid hormone action. Cell Mol Life Sci. (2009)
- Gorelick-Feldman J, et al. Phytoecdysteroids increase protein synthesis in skeletal muscle cells. J Agric Food Chem. (2008)
- Báthori M, et al. Phytoecdysteroids and anabolic-androgenic steroids–structure and effects on humans. Curr Med Chem. (2008)
- Bucci LR. Selected herbals and human exercise performance. Am J Clin Nutr. (2000)
- Kuzmenko AI, et al. Effects of vitamin D3 and ecdysterone on free-radical lipid peroxidation. Biochemistry (Mosc). (1997)
- Spindler KD, et al. Ecdysteroid hormone action. Cell Mol Life Sci. (2009)
- Tatar M, Yin C. Slow aging during insect reproductive diapause: why butterflies, grasshoppers and flies are like worms. Exp Gerontol. (2001)
- Simon AF, et al. Steroid control of longevity in Drosophila melanogaster. Science. (2003)
