
Introduzione:
Nel dibattito scientifico l’origine e sviluppo della vita sulla terra è tutt’altro che arrivato ad un punto chiaro e incontestabile. Oggi, la maggior parte della comunità scientifica ha abbracciato la filosofia materialista e casualista, comunemente chiamata evoluzionismo. Ciò nonostante, con l’avanzare delle evidenze scientifiche in campo paleontologico, biochimico e genetico la crisi della fazione materialista/casualista si è aggravata specie con la nascita del movimento scientifico dell’Intelligence Design il quale, al contrario dei preponderanti e disperati evoluzionisti, applica rigorosamente il metodo scientifico e l’osservazione e comprensione oggettiva e lucida dei dati emergenti dalla ricerca.
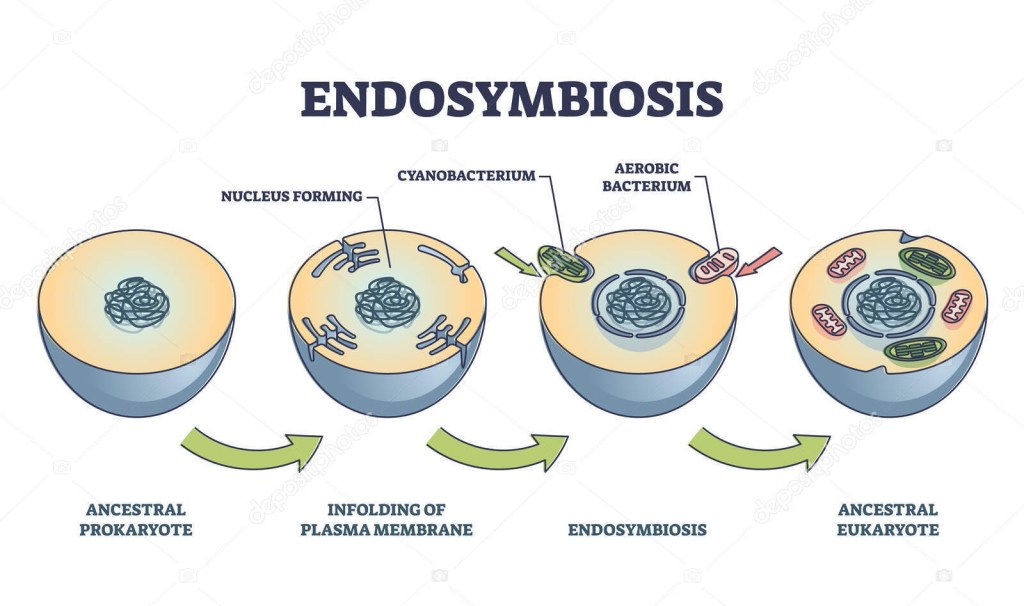
Tra le varie teorie (non dimostrate) del panorama evoluzionista ve ne è una che prende il nome di “endosimbiosi”. In breve, L’endosimbiosi (dal greco: ἔνδον = dentro; συν = insieme; βιος = vita) sarebbe una particolare forma di interazione biologica, una simbiosi mutualistica nella quale un organismo (di solito unicellulare) vive all’interno di un altro organismo.
L’endosimbiosi è un’interazione di carattere mutualistico: entrambe le specie che interagiscono traggono vantaggio dall’interazione; questo mutuo beneficio distingue la simbiosi mutualistica dal parassitismo e dal commensalismo. Stiamo praticamente parlando di una trasmissione di cooperatività mutualistica generativa o generazionale.
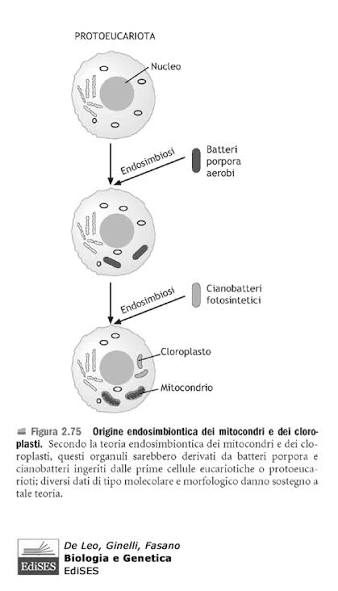
Alcuni scienziati hanno supposto che un’antica endosimbiosi abbia originato alcune caratteristiche permanenti degli organismi attuali, formulando la teoria endosimbiotica (chiamata anche teoria endosimbiontica o teoria dell’endosimbionte) riguardo alle origini di alcuni organismi.
L’ipotesi è che alcuni organismi biologici siano stati ingeriti da altri organismi e poiché ne trassero un vantaggio evoluzionistico di sopravvivenza reciproco, svilupparono una relazione simbiotica permanente che nelle generazioni è divenuta indissolubile e imprescindibile; come esempio viene postulato che, nel passato remoto del teorico Precambriano, un batterio aerobico (che richiede ossigeno) sia stato ingerito da un batterio anaerobio (per il quale forse l’ossigeno era tossico) acquisendo un vantaggio reciproco e che continuando la loro relazione mutualistica abbiano superato evoluzionisticamente gli altri organismi in quell’ambiente; nel tempo il batterio interno ha perso o spostato materiale genetico nel nucleo dell’ospitante, per la codifica di tutto ciò che non era più necessario o superfluo.
Ma stiamo parlando di una teoria stabilizzata su estese sperimentazioni, osservazioni scientifiche ed analisi genetiche?
Breve approfondimento sulla endosimbiosi

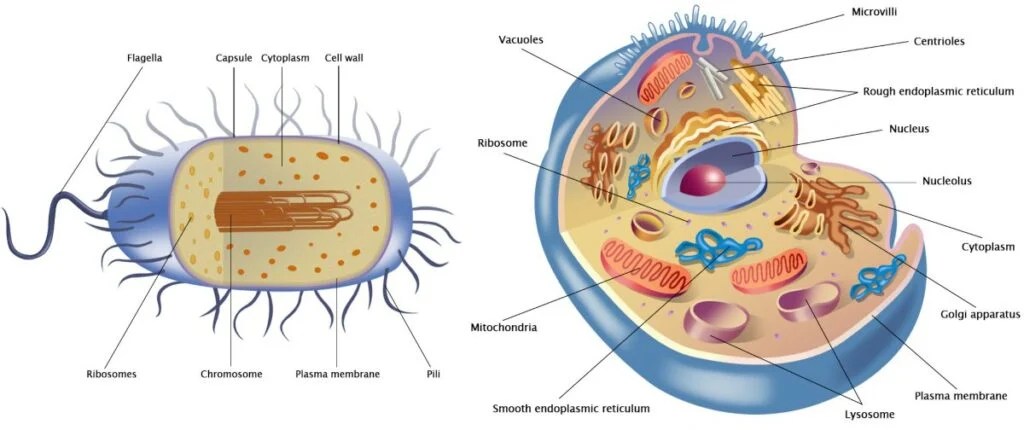
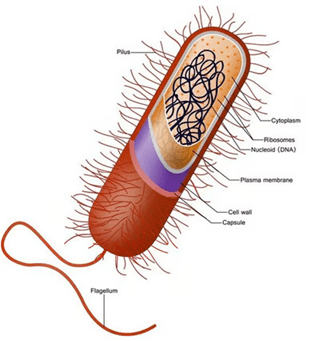
Come sappiamo, le cellule si dividono in due gruppi principali: procarioti (più piccola e semplice, ha il DNA sparso nel citoplasma (in una zona detta nucleoide) e non possiede organuli) ed eucarioti (hanno un nucleo ben definito che racchiude il DNA ed è ricca di organuli interni). La teoria dell’evoluzione postula che i procarioti si siano evoluti in eucarioti. Esiste un enorme divario tra questi due tipi di cellule che non avrebbe potuto essere colmato da forme di transizione inquanto avrebbero dovuto essere frutto di tentativi mutageni enormi e impossibilità o bassissima sopravvivenza. Il tentativo più diffuso di spiegare questo divario è la teoria dell’endosimbiosi di Lynn Margulis. I suoi sostenitori ipotizzano che alcune cellule proto-eucariotiche abbiano inglobato dei procarioti e che, successivamente, i proteobatteri inglobati si siano evoluti in organelli negli eucarioti primitivi. In questo articolo vengono esaminati i numerosi problemi principali di questa teoria, giungendo alla conclusione che essa è ampiamente accettata solo perché rappresenta l’ipotesi evolutiva più plausibile e non per via di prove empiriche. Infatti, come documentato in questo articolo, esistono numerose prove contrarie alla teoria dell’endosimbiosi.
Comprendere l’evoluzione della complessità cellulare eucariotica è una delle grandi sfide della biologia moderna. A differenza dei procarioti, le cellule eucariotiche sono altamente compartimentalizzate e contengono molti organelli delimitati da membrana, assenti nei batteri o negli archei (procarioti unicellulari non batterici). Oltre a numerose differenze genetiche e molecolari, nei procarioti non si trovano organelli compartimentalizzati complessi. Gli evoluzionisti generalmente tentano di spiegare come ciò sia avvenuto attraverso l’endosimbiosi, sostenendo che un antico archeo abbia inglobato un proteobatterio che alla fine ha dato origine al primo organello, il Mitocondrio.
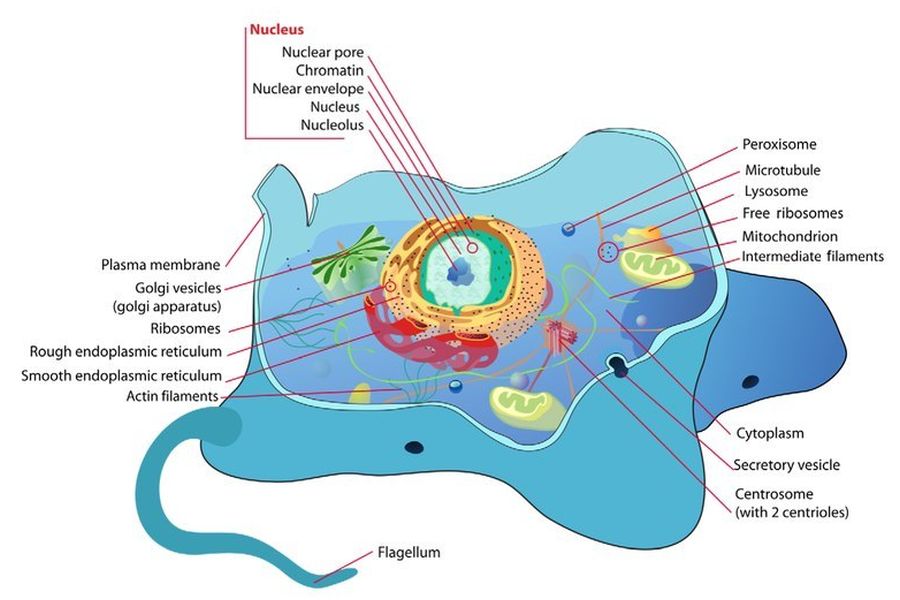
L’endosimbiosi è un fenomeno in cui un organismo vive all’interno di un altro in una relazione di mutuo beneficio, come si osserva nel caso di alcuni batteri che vivono all’interno delle termiti, ad esempio. In questo articolo, il termine “endosimbiosi” non si riferirà a tali fenomeni osservati, bensì all’ipotesi ampiamente accettata dal compianto Professor Lynn Margulis per spiegare l’origine dei Mitocondri, che è l’oggetto principale di questo lavoro (a volte indicata come “endosimbiosi primaria”). Essa viene invocata nel tentativo di colmare il divario esistente tra le cellule prive di organelli compartimentalizzati (procarioti) e quelle che li possiedono (eucarioti). Naturalmente, le differenze vanno ben oltre i Mitocondri. Alcuni degli altri organelli che un eucariote possiede, a differenza dei procarioti, sono il nucleo, il nucleolo, il reticolo endoplasmatico rugoso e liscio, l’apparato di Golgi, i centrioli, i perossisomi e i lisosomi.
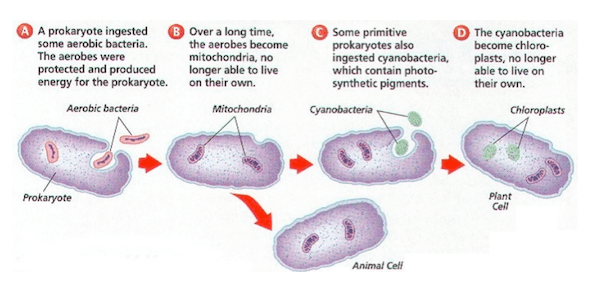
Secondo questa teoria dell’endosimbiosi, in un passato remoto un batterio procariote avrebbe inglobato un ipotetico proteobatterio, che sarebbe rimasto all’interno dell’ospite instaurando una relazione simbiotica. Ciò significa che centinaia di geni sarebbero stati in qualche modo modificati, assumendo funzioni completamente nuove. Inoltre, la teoria suggerisce che migliaia di geni del proteobatterio sarebbero stati trasferiti nel nucleo della cellula protoeucariote, mentre molti altri sarebbero stati scartati.
Il Mitocondrio come esempio di endosimbiosi(?)
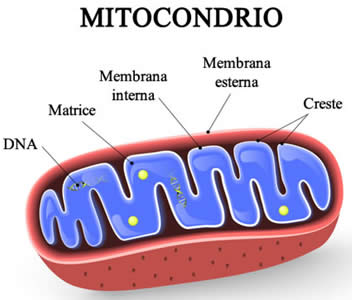
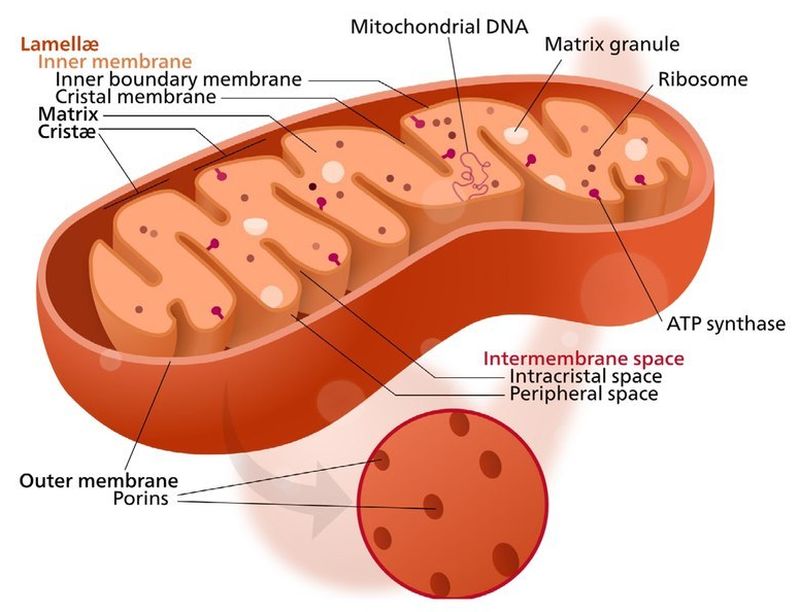
Il Mitocondrio (pl. mitocondri) è un organello presente nelle cellule della maggior parte degli eucarioti, come animali, piante e funghi. I Mitocondri hanno una struttura a doppia membrana e utilizzano la respirazione aerobica per generare adenosina trifosfato (ATP), che viene utilizzato in tutta la cellula come fonte di energia chimica. Furono scoperti da Albert von Kölliker nel 1857 nei muscoli volontari degli insetti. Il termine Mitocondrio, che significa “granulo filamentoso”, fu coniato da Carl Benda nel 1898. Il Mitocondrio è comunemente soprannominato “centrale energetica della cellula”, un’espressione resa popolare da Philip Siekevitz in un articolo del 1957 su Scientific American con lo stesso titolo.
I Mitocondri hanno generalmente una sezione trasversale compresa tra 0,75 e 3 μm², ma variano considerevolmente in dimensioni e struttura. A meno che non vengano colorati specificamente, non sono visibili. Il Mitocondrio è composto da compartimenti che svolgono funzioni specializzate. Questi compartimenti o regioni includono la membrana esterna, lo spazio intermembrana, la membrana interna, le creste e la matrice.
Oltre a fornire energia cellulare, i Mitocondri sono coinvolti in altre funzioni, come la segnalazione, la differenziazione cellulare e la morte cellulare, nonché nel mantenimento del controllo del ciclo cellulare e della crescita cellulare. La biogenesi mitocondriale è a sua volta coordinata temporalmente con questi processi cellulari.
I Mitocondri sono implicati in disturbi e condizioni umane come malattie mitocondriali,, disfunzioni cardiache, insufficienza cardiaca e autismo.
Il numero di Mitocondri in una cellula varia notevolmente a seconda dell’organismo, del tessuto e del tipo di cellula. Un globulo rosso maturo non ha mitocondri, mentre una cellula epatica può averne più di 2000.
Sebbene la maggior parte del DNA di una cellula eucariotica sia contenuta nel nucleo cellulare, il Mitocondrio ha un proprio genoma (“mitogenoma”) simile ai genomi batterici [sottolineiamo SIMILE]. Questa scoperta ha portato all’accettazione di una parte della comunità scientifica della simbiogenesi (teoria endosimbiotica) – secondo la quale gli antenati procarioti a vita libera dei Mitocondri moderni si sono fusi permanentemente con le cellule eucariotiche in un lontano passato, evolvendosi in modo tale che gli animali, le piante, i funghi e gli altri eucarioti moderni respirino per generare energia cellulare.
Seguendo la “logica” endosimbiontica, i Mitocondri, organuli delle cellule eucariotiche, si sarebbero quindi originati come organismi procarioti esterni, introdottisi nella cellula come endosimbionti, circa 1,5 miliardi di anni fa. I mitocondri si sarebbero sviluppati da proteobacteria (in particolare, Rickettsiales o affini e forse da un batterio molto vicino a Rickettsia prowazekii.)
Alcune ipotesi del fatto che i mitocondri si originarono da antiche endosimbiosi di batteri sono ad esempio:
- I mitocondri contengono DNA diverso da quello del nucleo cellulare e simile a quello dei bacteria; si nota la presenza di un DNA circolare a doppia elica e la presenza di ribosomi propri e di una doppia membrana. Come i batteri, i mitocondri non hanno istoni ed i loro ribosomi sono sensibili ad alcuni antibiotici (come il cloramfenicolo). In più i mitocondri sono organelli semiautonomi in quanto replicano, per scissione binaria, autonomamente rispetto alla cellula.
- Sono circondati da due o più membrane, la più interna delle quali mostra una composizione differente da quella delle altre membrane della cellula; si nota la presenza di molecole di cardiolipina ed assenza di colesterolo; la sua composizione è simile a quella di una membrana cellulare procariotica.
- Nuovi mitocondri si formano solamente attraverso un processo simile alla scissione binaria. In alcuni casi i mitocondri possono essere distrutti da alcuni agenti ambientali o disfunzioni patogenetiche senza tuttavia danneggiare la cellula che li ospita, e comunque non si rigenerano.
- La maggior parte della struttura interna e la biochimica dei mitocondri è molto simile a quella dei batteri. L’idea filogenetica, basata sui genomi di batteri, mitocondri ed eucarioti, suggerisce che i mitocondri siano strettamente derivati da bacteria.
- L’analisi della sequenza del DNA e la teoria filogenetica suggeriscono che il DNA nucleare probabilmente contiene geni che vennero dai batteri/mitocondri originali inglobati.
- Alcune proteine codificate nei nuclei sono trasportate agli organelli e i mitocondri hanno genomi piccoli se paragonati a quelli dei batteri. Ciò concorda con l’idea di un incremento della dipendenza sull’ospite eucariote dopo la formazione di un’endosimbiosi. La maggior parte dei geni dei batteri/organelli inglobati è andata perduti se inutile o si è spostata nel nucleo. La maggior parte dei geni necessari per le funzioni mitocondriali si trova nel nucleo. Molti hanno avuto origine dalla endosimbiosi batterica.
- Se i mitocondri si fossero originati ex novo, dovrebbero averlo fatto molteplici volte, nel qual caso la loro rispettiva somiglianza è difficilmente spiegabile. Molti protisti contengono batteri ospitati secondari che sono stati acquisiti da altri eucarioti contenenti mitocondri, e non direttamente.
- Tra gli eucarioti più antichi, i mitocondri sono maggiormente somiglianti ai batteri primigeni.
- I ribosomi dei mitocondri sono come quelli trovati nei batteri (70S).
- Le proteine originate dai mitocondri usano, come quelle dei batteri, N-formilmetionina come amminoacido iniziale.
Per colmare, o meglio, giustificare il fatto che non possono sopravvivere in ossigeno o fuori dalla cellula, si gioca la carta della perdita “funzionale” di molti geni necessari per la sopravvivenza, accaduto nel corso di un lungo intervallo di tempo in cui i mitocondri hanno co-abitato con i loro ospiti; i geni e i sistemi che non erano più necessari sarebbero stati semplicemente eliminati, o in molti casi trasferiti nel genoma ospite (infatti questi trasferimenti costituiscono un importante mezzo per la cellula ospite di regolare l’attività mitocondriale).
Secondo recenti osservazioni si è ipotizzato che gli endosimbionti mitocondriali possono sopravvivere almeno per un po’ a vita libera in fluidi corporei stabili, come il sangue, e che siano in grado di trasferirsi da cellula a cellula in caso di necessità, quindi sempre passando per uno stadio a vita libera, anche se breve.
Ma le cose non sono “liquidabili” in questo modo…
L’abisso del DNA procariotico-eucariotico
I procarioti contengono una singola molecola di DNA circolare che occupa la regione del nucleoide e spesso anche piccoli anelli di DNA extracromosomico a doppio filamento chiamati plasmidi. Il DNA dei procarioti è “profondamente diverso da quello degli eucarioti” che contengono “da due a quattro genomi di DNA nucleare, cloroplastico, microtubulare e mitocondriale separati e trasmessi indipendentemente”. Il contrasto procariotico-eucariotico è così grande da costituire “la più grande singola discontinuità evolutiva” della vita conosciuta, e “l’origine degli eucarioti è rimasta una delle questioni più enigmatiche, controverse e impegnative dell’evoluzione”. Rimangono solo due teorie naturalistiche sull’evoluzione degli organelli; tutte le altre sono state effettivamente confutate.
La prima teoria è l’ipotesi autogena, ovvero quella dell'”autogenerazione”, che postula che gli organelli si siano evoluti gradualmente da un organello precursore attraverso la selezione naturale delle mutazioni. Questa visione non trova riscontro in tutti gli organelli esistenti e, di conseguenza, è stata ampiamente respinta e sostituita da una qualche forma di endosimbiosi.
La seconda teoria è quella dell’endosimbiosi, detta anche teoria dell’endosimbiosi seriale (SET), simbiogenesi o ipotesi xenogena. I batteri inglobati si sono successivamente evoluti all’interno dei loro ospiti per acquisire funzioni specializzate, alcuni diventando mitocondri che alla fine hanno assunto il ruolo di fornire energia “caricando” l’ADP in ATP. Questo avviene tramite l’enzima ATP sintasi, una “macchina” molecolare presente sia negli eucarioti che nei procarioti. I procarioti sono organismi microscopici unicellulari che hanno esigenze energetiche molto inferiori rispetto agli eucarioti.
L’apparato di sintesi dell’ATP in un procariote è incorporato nella sua membrana cellulare, come sarebbe accaduto anche per il batterio inglobatore, che in questa fase della teoria è ancora un procariote. Al contrario, gli eucarioti utilizzano i mitocondri per caricare l’ADP, quindi quando l’antico archeo avrebbe presumibilmente inglobato un proteobatterio che sarebbe poi diventato un mitocondrio, doveva comunque soddisfare il proprio fabbisogno energetico durante l’evoluzione da procariote a eucariote. Inoltre, solo una “piccola frazione delle proteine necessarie per la propagazione e la funzione dei mitocondri è codificata dai loro genomi, mentre i geni nucleari codificano la stragrande maggioranza”. Il fulcro della spiegazione dell’endosimbiosi per l’abisso tra procarioti ed eucarioti è il DNA mitocondriale.
Dall’endosimbiosi agli organismi multicellulari
È noto da tempo che “il principio dell’endosimbiosi fu proposto più di un secolo fa, ma fu generalmente considerato una ‘fantasia divertente'”. Questa visione rimase sostanzialmente invariata fino al lavoro di Lynn Margulis, che sviluppò l’idea dell’endosimbiosi in modo molto dettagliato. Grazie al suo lavoro e alla sua influenza, l’endosimbiosi è passata da un’idea oscura e scarsamente accettata alla teoria più popolare sull’origine degli organelli oggi.
Lo scenario comune dell’endosimbiosi postula che le cellule eucariotiche a vita libera si siano infine unite in comunità oggi chiamate organismi multicellulari.11 I problemi emersero poco dopo la proposta della teoria dell’endosimbiosi. Ad esempio, ulteriori ricerche hanno scoperto che i mitocondri di funghi, piante e animali erano così diversi che l’endosimbiosi deve essersi verificata indipendentemente molte volte, il che non fa che moltiplicare le probabilità, già di per sé molto improbabili, che lo scenario endosimbiotico si verifichi anche solo una volta.
Margulis predisse che il DNA mitocondriale sarebbe stato diverso dal DNA nucleare, ma che sarebbe invece consistito in una miscela di sequenze provenienti da geni eubatterici e archeali. Nel 1963, si scoprì che i mitocondri possedevano DNA indipendente dal DNA nucleare. Un altro fattore importante a sostegno dell’endosimbiosi fu la scoperta, negli anni ’60, del DNA plastidiale (cpDNA) nei cloroplasti delle piante, chiamato plastoma, che secondo i sostenitori rendeva più plausibile il meccanismo dell’endosimbiosi per l’origine degli organelli.15
Si ritiene comunemente che gli organelli chiamati mitocondri siano comparsi una sola volta nella storia evolutiva, ma nonostante la loro comune origine, i DNA mitocondriali “L’architettura del genoma e il contenuto genico variano ampiamente tra gli eucarioti.” I ricercatori hanno scoperto che il mtDNA più grande si trova in un protozoo d’acqua dolce, Reclinomonas americana, che ha 69.034 nucleotidi e 97 geni che codificano 67 proteine, tra cui almeno 18 proteine non precedentemente note per essere codificate nei mitocondri. L’endosimbiosi deve quindi postulare che i batteri inglobati abbiano perso il 96-99% delle loro proteine, da circa 1.600 a meno di 67, a seconda dello specifico protobatterio inglobato. Per gli esseri umani, solo 37 geni sono essenziali per la respirazione cellulare.
Inoltre, i confronti tra proteine eucariotiche presenti negli organelli eucariotici hanno rivelato che non si tratta semplicemente di una miscela di sequenze provenienti da archei ed eubatteri, come previsto dall’endosimbiosi, ma che sono spesso uniche, in contrasto con l’idea che i genomi eucariotici siano una combinazione di geni eubatterici e archeali.
Anche tutti gli altri esempi esaminati mostrano notevoli differenze tra i genomi mitocondriali e batterici, come previsto date le differenze proteiche descritte da Kirkland in un paragrafo precedente. Persino per l’esempio più noto (mtDNA di R. americana), esistono enormi differenze tra i suoi 69 kbp (97 geni) e sia il genoma di 580 kbp (470 geni) di Mycoplasma genitalium, sia il genoma di 1.830 kbp (1.743 geni) di Haemophilus influenzae. Inoltre,
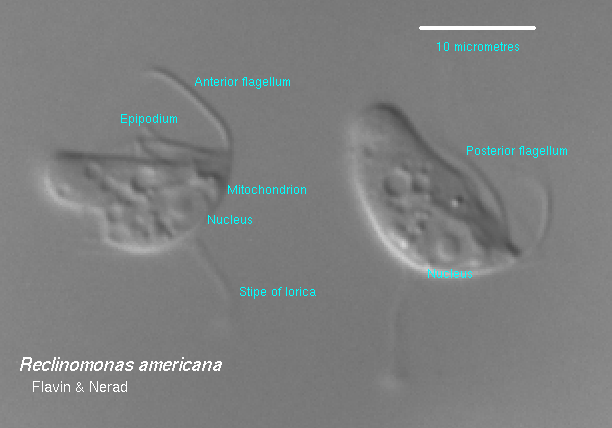
“Il confronto tra i genomi di Mycoplasma e Haemophilus ha suggerito che il loro diverso contenuto genico riflette ‘profonde differenze… tra questi due organismi’… In questo contesto, il genoma mitocondriale di Reclinomonas può essere considerato un esempio estremo di riduzione del genoma eubatterico, in cui gli unici geni rimanenti sono correlati all’espressione genica mitocondriale (trascrizione, elaborazione dell’RNA e traduzione) e alla biogenesi dei complessi proteici necessari per il trasporto di elettroni e la fosforilazione ossidativa accoppiata (inclusi i componenti implicati nel trasporto e nella biosintesi delle proteine mitocondriali).”¹⁹
In breve, questi pochi esempi illustrano l’abisso esistente tra il DNA batterico e il mtDNA, che è solo uno dei molti problemi dell’endosimbiosi.
Somiglianze e differenze tra mitocondri e batteri
La teoria dell’endosimbiosi si basa in gran parte sull’omologia tra organelli e batteri. Ad esempio, ogni mitocondrio possiede un genoma circolare come quello dei batteri, ma molto più piccolo e privo di proteine istoniche. Il DNA mitocondriale (mtDNA) si trova solitamente nella matrice mitocondriale, sebbene a volte sia attaccato alla membrana mitocondriale interna.
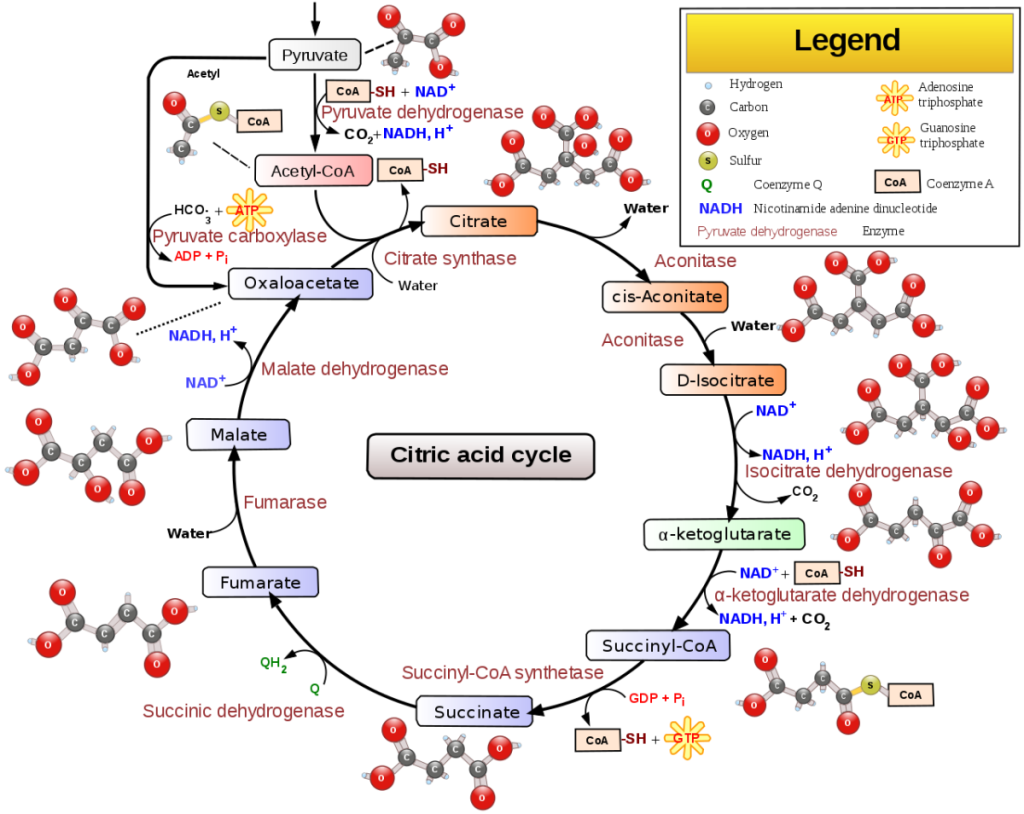
I mitocondri assomigliano molto ai batteri aerobici viola per dimensioni e forma. Entrambi utilizzano l’ossigeno per la produzione di ATP tramite il ciclo di Krebs. Alcuni antibiotici che uccidono i batteri inibiscono anche le funzioni mitocondriali. Queste somiglianze generali da sole non dimostrano l’endosimbiosi perché, come documentato di seguito, esistono molte differenze significative, spesso importanti e critiche.
Un fattore a favore dell’endosimbiosi è la composizione della membrana. La membrana esterna sia dei cloroplasti che dei mitocondri presenta somiglianze strutturali e chimiche con la membrana cellulare procariotica. Ricerche successive, tuttavia, hanno determinato che le membrane mitocondriali sono solo superficialmente simili alle membrane cellulari procariotiche. Una differenza risiede nel fatto che il proteobatterio che si presume sia penetrato nel protoeucariote avrebbe avuto una singola membrana, mentre i mitocondri moderni ne possiedono una doppia (interna ed esterna). La doppia membrana non è facoltativa, ma fondamentale per la sua funzione di caricamento dell’ADP. La membrana interna contiene numerose pieghe a forma di piastra chiamate creste, che presentano sacche membranose contenenti enzimi. Le creste possono essere esclusivamente lamellari o esclusivamente tubulari, ma alcuni mitocondri contengono entrambi i tipi. Un’altra differenza è che la membrana mitocondriale interna ha una composizione chimica diversa da quella dei procarioti, ma è identica a quella degli eucarioti, contrariamente a quanto previsto dalla teoria dell’endosimbiosi.

Margulis propose “che gli eucarioti si fossero formati a seguito di una graduale unione multi-endosimbiotica con i procarioti. Al contrario, altri, come de Duve e Stanier, proposero che la fagotrofia, che richiede un citoscheletro dinamico, un sistema endomembranoso e la perdita della parete cellulare rigida procariotica, si fosse evoluta prima dell’endosimbiosi”. In contrasto con questa proposta, la filogenetica basata sul mtDNA concluse in seguito che i mitocondri avrebbero potuto evolversi una sola volta da un α-proteobatterio. La visione dell’endosimbiosi come evoluzione degli organelli è ampiamente accettata non per via di prove empiriche, ma perché nessun’altra teoria è neanche lontanamente plausibile. Per questo motivo, Battley descrive l’endosimbiosi come “nella migliore delle ipotesi provvisoria”.
Poiché non esistono prove fisiche per la maggior parte delle fasi della transizione dalla cellula procariotica alla cellula eucariotica, si ricorre a ragionamenti astratti (ad esempio, i mitocondri e i cloroplasti possiedono un piccolo plasmide di DNA che superficialmente assomiglia al DNA procariotico) come supporto. In realtà, il DNA degli organelli è più simile ai geni nucleari eucariotici. Un esempio ben noto di alcuni geni degli organelli che assomigliano ai geni nucleari eucariotici è la presenza di introni, che sono raramente presenti nei geni procariotici.
L’endosimbiosi non risolve il problema dell’origine degli organelli.
Sebbene l’origine endosimbiotica dei mitocondri e dei cloroplasti sia ormai un dogma consolidato nei manuali, le proposte secondo cui la maggior parte degli “altri compartimenti cellulari siano il risultato della simbiosi… non sono altrettanto ampiamente accettate”. Per la maggior parte degli altri organelli, l’endosimbiosi è considerata da molti ricercatori una spiegazione implausibile per la loro origine. (È anche una spiegazione inadeguata per il mitocondrio e il cloroplasto, sebbene la teoria dell’evoluzione del cloroplasto sarà trattata in un articolo separato).
L’endosimbiosi non è una spiegazione adeguata per i mitocondri
Un grave problema scientifico dell’ipotesi dell’endosimbiosi è che, fin dalla sua prima formulazione, la teoria si è rivelata (e rimane tuttora) non verificabile.26 Il problema è che l’endosimbiosi “non propone un meccanismo reale e la maggior parte dei libri di testo presenta l’immagine semplicistica di una cellula che ingloba un’altra cellula che diventa un mitocondrio”. In realtà, oggi è molto meno plausibile di quando fu proposta per la prima volta, poiché oggi si sa molto di più sugli organelli (ad esempio i mitocondri) e sui batteri.
Tra gli altri problemi fondamentali della teoria vi sono: cosa ha impedito alla cellula ospite di digerire l’organismo invasore? e: da dove provengono le numerose altre strutture necessarie alla sopravvivenza di una cellula eucariotica? Ad esempio, i microtubuli non sono spiegati dalla teoria, sebbene siano necessari per la divisione cellulare e la motilità nelle cellule eucariotiche. De Duve osserva che non si sa nulla sull’evoluzione del sistema del citoscheletro cellulare, che richiede molte nuove innovazioni per funzionare. Simili prove citate a sostegno della teoria secondo cui i batteri spirocheti avrebbero dato origine ai flagelli sono problematiche. La tubulina, il componente principale dei microtubuli nelle cellule eucariotiche, non è stata trovata in nessun procariote. Per questi motivi, la maggior parte dei biologi evoluzionisti rifiuta l’idea che flagelli, tubulina e la maggior parte delle altre strutture cellulari abbiano avuto origine per endosimbiosi. Nella migliore delle ipotesi, l’endosimbiosi spiega l’origine di uno o due organelli. Ma affinché una cellula eucariotica funzioni, è necessario un insieme completamente nuovo di strutture, tutte le quali devono evolversi simultaneamente per garantire l’integrità funzionale.
Principali differenze tra i ribosomi e il mitoribosoma
Un argomento fondamentale a sostegno della teoria endosimbiotica sull’origine degli organelli era che la struttura dei ribosomi mitocondriali è “nettamente diversa” da quella dei ribosomi eucariotici, e che i ribosomi mitocondriali assomigliano a quelli dei procarioti. Contrariamente alla teoria endosimbiotica, tuttavia, i ribosomi mitocondriali dei mammiferi e le sequenze di amminoacidi che li compongono sono completamente diversi dalle corrispondenti caratteristiche nei procarioti. I ribosomi mitocondriali sono infatti così diversi dai ribosomi batterici da essere denominati mitoribosomi.
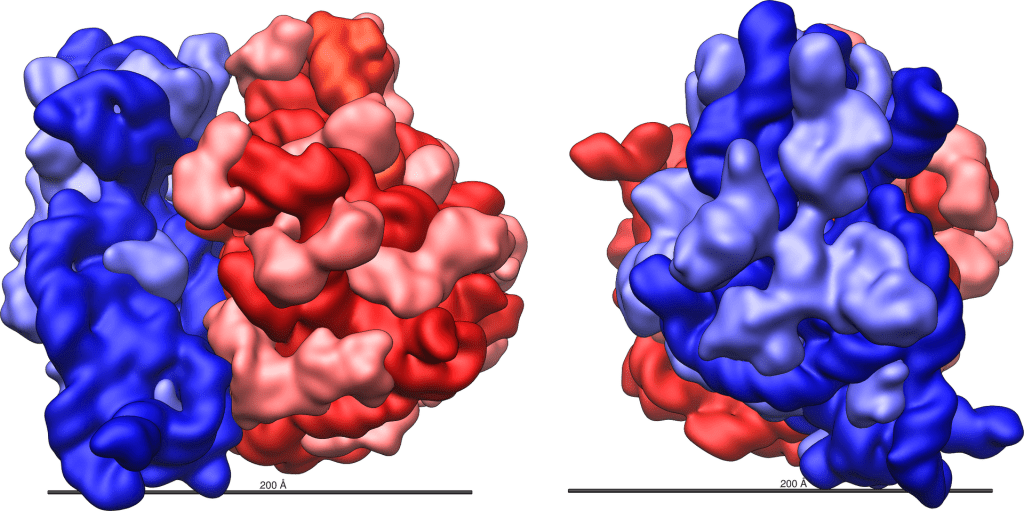
Quando l’endosimbiosi fu proposta per la prima volta, si presumeva che i ribosomi esistessero solo in due forme: una varietà più piccola, 70S, utilizzata nei procarioti, e un ribosoma più grande, 80S, utilizzato negli eucarioti. La S in 70S si riferisce all’unità di misura del coefficiente di sedimentazione. Il coefficiente di sedimentazione misura le differenze morfologiche di base, una quantità correlata alla dimensione della particella che è pari alla velocità terminale di avanzamento della particella quando centrifugata in un mezzo fluido standard, divisa per la forza centrifuga che agisce su di essa. Ci si aspettava che i ribosomi utilizzati nei mitocondri dei mammiferi assomigliassero ai ribosomi 70S dei procarioti, data la loro simile dimensione. Invece, i ricercatori hanno scoperto che “i ribosomi mitocondriali dei mammiferi (55S) differiscono inaspettatamente dai ribosomi batterici (70S) e citoplasmatici (80S), così come da altri tipi di ribosomi mitocondriali”.
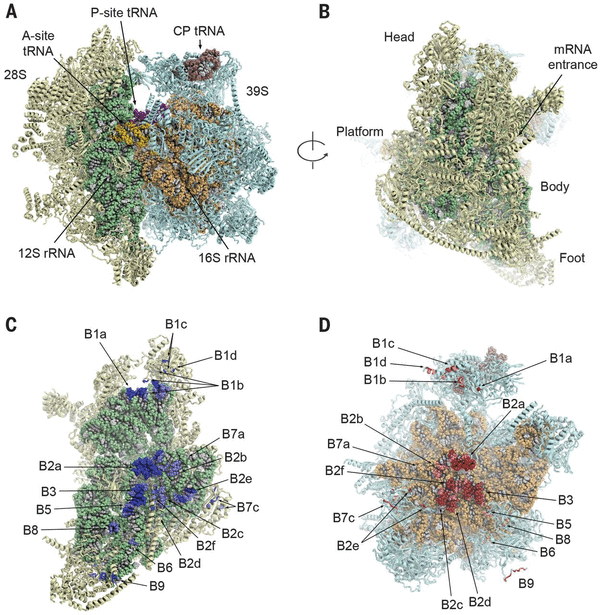
I mitocondri utilizzano un sistema necessario per la produzione di proteine che è anch’esso molto diverso da quello dei batteri. I ribosomi mitocondriali, chiamati mitoribosomi, sono descritti come mini-ribosomi “in miniatura”, contenenti una mini-RNA polimerasi e persino un mini-DNA. I mitoribosomi si differenziano dai ribosomi procariotici per il contenuto di RNA e proteine, nonché per la posizione e la funzione delle diverse componenti ribosomiali, e presentano differenze significative nella sequenza del DNA (in particolare nelle regioni che non entrano in contatto con il tRNA o con la catena polipeptidica in crescita). Tra le caratteristiche peculiari del ribosoma mitocondriale si annoverano nuovi mRNA che processano l’mRNA mitocondriale e un nuovo sito di legame per il guanosina trifosfato (GTP) utilizzato durante l’allungamento della catena polipeptidica.
Altre differenze tra i ribosomi batterici e i mitoribosomi riguardano la loro struttura e i loro processi di assemblaggio. Le proteine costituiscono una porzione maggiore dei mitoribosomi rispetto ai ribosomi procariotici. Alcune di queste proteine si trovano in posizioni inedite e svolgono funzioni diverse rispetto a quelle del ribosoma procariotico. Uno dei tanti esempi è rappresentato dal mitoribosoma 55S, tenuto insieme da 15 ponti intersubunitari, di cui solo sei simili a quelli utilizzati nei procarioti.33 Inoltre, 33 delle 81 proteine identificate finora nei mitoribosomi umani non hanno omologhi nei ribosomi procariotici.
Questi esempi delle numerose differenze tra il ribosoma procariotico e quello mitocondriale illustrano ulteriormente l’abisso che separa i due tipi di ribosoma. Si potrebbero documentare altri esempi e senza dubbio ne verranno scoperti altri con ulteriori ricerche.
Il problema della fagocitosi
Un altro problema è che la fagocitosi avrebbe presumibilmente introdotto un proteobatterio nei procarioti attraverso la “fagocitosi” di batteri, ma “la natura precisa della cellula ospite che si è associata a questo endosimbionte è, tuttavia, una questione ancora aperta”.¹ Il problema è che gli archei – in realtà tutti i procarioti – “non sono in grado di eseguire la fagocitosi e non c’è motivo di credere che abbiano mai avuto tali capacità”. Di conseguenza, “il modo in cui l’endosimbionte è entrato nel suo ospite è un enigma”. Il motivo per cui i procarioti non possiedono la capacità di fagocitosi è che si tratta di un processo complesso che richiede
“… una parete cellulare flessibile, un citoscheletro interno dinamico con proteine motrici che interagiscono con un complesso sistema endomembranoso, lisosomi che gemmano dal complesso di Golgi e sono diretti verso i vacuoli alimentari e particelle racchiuse in una coppa fagocitica basata sulla polimerizzazione spazialmente controllata dell’actina. “I caratteri sono assenti nei procarioti.”
Una soluzione teorica a questo problema suggerisce che l’organismo invasore “fosse un piccolo α-proteobatterio (facoltativo) aerobico, che penetrava e si replicava nel periplasma dell’ospite, diventando in seguito i mitocondri cellulari.” Questa storia, per ora, non trova riscontro empirico.
Altre differenze tra i mitocondri e i batteri
I mitocondri sono organelli importanti perché contengono i meccanismi e gli enzimi necessari per convertire il cibo in una molecola energetica chiamata adenosina trifosfato (ATP). Il processo enzimatico mediante il quale i mitocondri convertono il cibo in adenosina trifosfato (ATP) è chiamato fosforilazione ossidativa. Il processo finale prevede la conversione dell’adenosina difosfato (ADP) nella molecola ad alta energia ATP. I mitocondri producono il 90% dell’energia cellulare e la loro disfunzione causa diverse malattie che colpiscono il sistema nervoso centrale e, in seguito, altri sistemi.38 I mitocondri producono questa energia a partire da grassi, zuccheri e proteine; sono presenti in tutte le cellule umane, ad eccezione dei globuli rossi anucleati.
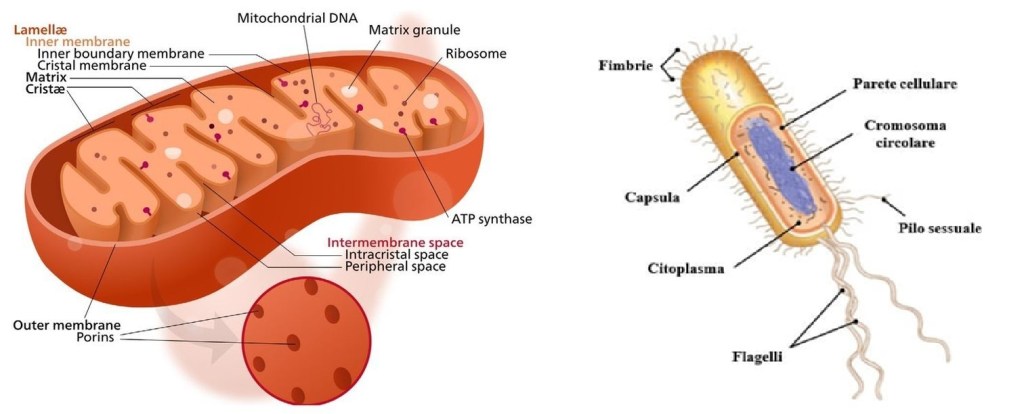
Definiti “le centrali energetiche della cellula”, sappiamo ora che le cellule più attive, come quelle muscolari, epatiche e dei tubuli renali, contengono un gran numero di mitocondri. Al contrario, le cellule meno attive, come quelle che secernono muco, contengono un numero relativamente basso di mitocondri. Altre funzioni dei mitocondri sono di tipo regolatorio, tra cui il controllo del livello di calcio citoplasmatico.39 Sono inoltre coinvolti in specifici processi di sintesi lipidica.
Le membrane interne contengono un ampio insieme di enzimi che convertono il cibo in ADP carico attraverso una serie di reazioni chiamate ciclo di Krebs o ciclo dell’acido citrico, che producono fosforilazione ossidativa. La regione interna del mitocondrio, chiamata matrice, è riempita di un gel contenente numerosi enzimi di diverso tipo. Le membrane interna ed esterna differiscono sia per attività enzimatica che per composizione lipidica. Alcuni enzimi, come l’ATPasi, sono permanentemente attaccati alla membrana mitocondriale. In breve, i mitocondri sono molto diversi dai batteri.
Geni che controllano i mitocondri
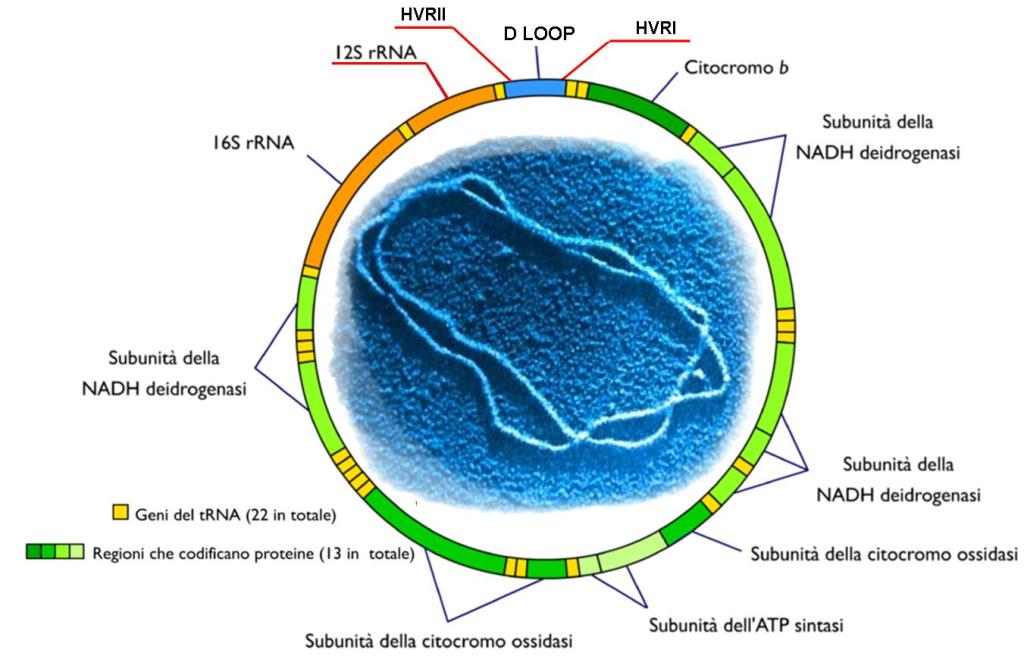
Come già accennato, il mitocondrio è un organello unico perché contiene il proprio DNA (mtDNA) sotto forma di plasmidi. Il DNA mitocondriale è utilizzato esclusivamente per le funzioni dell’organello stesso, in particolare per controllare, seppur non completamente, la propria replicazione.
Il mtDNA umano è stato completamente sequenziato. I suoi 16.569 paia di basi codificano per 37 geni, tra cui solo 13 geni che codificano per proteine, 22 geni per tRNA e due geni per rRNA, tutti essenziali. I sistemi respiratori non possono funzionare senza la presenza di tutte queste proteine e tRNA. Questi sono solo alcuni dei geni necessari ai mitocondri umani. Circa il 90% delle proteine importate dal citoplasma sono codificate nel nucleo, il che indica un elevato livello di integrazione che rende l’endosimbiosi insostenibile.
Poiché la maggior parte dei geni che controllano i mitocondri non si trova negli organelli stessi, ma nel nucleo cellulare, i sostenitori dell’endosimbiosi devono postulare un trasferimento di geni dagli organelli al nucleo della cellula ospite. Questo problema non è di poco conto: “la migrazione dei geni dagli endosimbionti al nucleo è notevole perché sembra aver sollevato più difficoltà di quante ne abbia risolte”. Un altro problema è:
“… in quale forma i geni trasferiti compiono fisicamente questo viaggio intracellulare: come RNA, come cDNA, come frammenti di DNA degli organelli o come interi cromosomi degli organelli? Le attuali teorie si concentrano sul cDNA come veicolo, basandosi su alcuni esempi tratti dalle piante. Ma altri meccanismi, che implicano il trasferimento diretto del DNA dai cromosomi degli organelli, potrebbero anche spiegare i dati disponibili”.
L’analogia non è dissimile dall’ipotizzare il trasferimento di una piccola casa in una casa più grande per spiegare le stanze di quest’ultima, quando queste possono essere spiegate più facilmente, anche da un punto di vista evolutivo, ipotizzando la loro costruzione individuale e separata. Questa preoccupazione è significativa in quanto i geni nei mitocondri rappresentavano una delle principali prove originali a sostegno della teoria dell’endosimbiosi. Da un punto di vista darwiniano, l’ipotesi che l’endosimbiosi ha sostituito, ovvero il processo di infiltrazione delle membrane all’interno delle cellule ospiti formando tutti gli organelli, incluso il nucleo, appare più plausibile e quindi ha ancora dei sostenitori. Man mano che i problemi con l’endosimbiosi si accumulano, l’ipotesi dell’infiltrazione delle membrane potrebbe tornare in auge.
In breve, la genetica suggerisce che questo trasferimento genico deve essersi verificato se l’endosimbiosi ha avuto luogo. I veri problemi sono: come sono sopravvissute le cellule pre-eucariotiche fino al trasferimento dei geni, come sono sopravvissute prima di questo trasferimento e perché e come sono stati trasferiti. La replicazione mitocondriale sembra richiedere il sistema di controllo nucleare perché, per quanto ne sappiamo, è universale. Tuttavia, il motivo per cui molti geni importanti per la funzione mitocondriale si trovino nel nucleo e come vi siano arrivati è oggetto di molte speculazioni evoluzionistiche.
Una delle scoperte più inaspettate è stata la scarsità di geni che potrebbero supportare l’endosimbiosi. Uno studio ha scoperto, contrariamente alle aspettative della teoria, che i confronti con l’organismo, chiamato endosimbionte α-proteobatterico, ampiamente ritenuto essere l’endosimbionte batterico
“… hanno identificato un nucleo conservato di proteine derivate dall’endosimbionte α-proteobatterico che ha dato origine al mitocondrio ed è stata la fonte del genoma mitocondriale negli eucarioti contemporanei. Un risultato sorprendente delle analisi filogenetiche è la proporzione relativamente piccola (10-20%) del proteoma mitocondriale che mostra una chiara ascendenza α-proteobatterica. Una grande frazione di proteine mitocondriali ha tipicamente omologhi rilevabili [similarità di sequenza] solo in altri eucarioti e si presume rappresenti proteine emerse specificamente all’interno degli eucarioti.”
I ricercatori hanno concluso:
“Comprendere l’origine e l’evoluzione del mitocondrio rimane una sfida, nonostante le Un’ondata di dati e intuizioni biochimiche, cellulari, di biologia molecolare e filogenetica, accumulate nei quasi cinque decenni trascorsi dalla moderna rinascita della consolidata ipotesi dell’endosimbiosi; l’idea che questo organello sia un batterio endosimbionte addomesticato e altamente rielaborato. L’abbondanza di informazioni relative all’evoluzione delle cellule eucariotiche (in particolare e più recentemente i dati di sequenza) e le divergenze su come i dati vengono analizzati e interpretati hanno generato una pletora di idee, spesso contrastanti, su quando e come, in un contesto endosimbiontico, si sia originato il mitocondrio.
Mitocondri dipendenti dal genoma nucleare
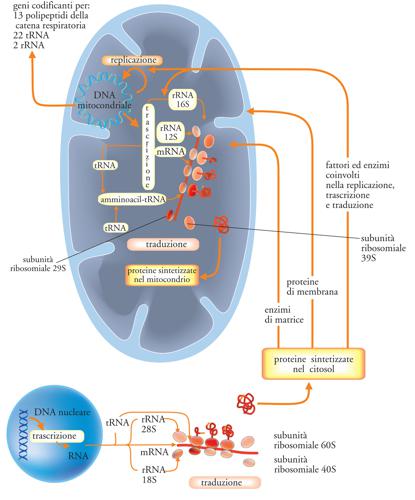
Un altro problema importante dell’endosimbiosi è che il genoma del mtDNA non è indipendente, ma è funzionalmente integrato con il genoma nucleare. Sono necessarie oltre 90 proteine per produrre i ribosomi mitocondriali, e quasi tutte vengono fornite all’organello dal nucleo dell’ospite. (Tra le fonti consultate, nessuna è stata in grado di fornire un numero specifico che, al momento della stesura di questo testo, rimane sconosciuto). Questi geni sono codificati dal DNA nucleare; le proteine risultanti vengono sintetizzate nel citosol cellulare e poi trasportate singolarmente all’interno dell’organello.
I mitocondri umani “devono importare il 99% delle loro proteine dal citoplasma”.51 Sarebbe molto più semplice (e quindi, secondo la “legge della parsimonia”, una spiegazione migliore) far evolvere i mitocondri da zero piuttosto che incorporare un organismo indipendente che richiederebbe: 1) la perdita della maggior parte dei suoi geni; 2) l’evoluzione di molti nuovi geni, il che implica: 3) Il fatto che la maggior parte dei geni necessari al funzionamento debba essersi evoluta originariamente nel nucleo e che, senza questi geni, il mitocondrio non potrebbe funzionare. Inoltre, l’utilizzo del sistema a due geni, nucleare e mitocondriale, richiede l’evoluzione di un macchinario di importazione estremamente complesso che coinvolge recettori di superficie complessi, sistemi di legame e un sistema di segnalazione mirato.
Un altro problema complesso è che alcuni mitocondri utilizzano codoni genetici diversi da tutti i codici batterici ed eucariotici. Ad esempio, il codone CUA normalmente codifica per la leucina, ma nei mitocondri del lievito codifica per la treonina. In realtà, esistono molteplici codici per codificare la treonina, e sono presenti modelli di riconoscimento dei codoni più complessi per altri amminoacidi. Per questi motivi, alcuni ipotizzano che il codice mitocondriale si sia evoluto in alcuni mitocondri di lievito, poiché, per quanto ne sappiamo, tutti i mitocondri di lievito presentano le stesse differenze di codice rispetto ai batteri. Man mano che verranno completati ulteriori sequenziamenti genetici su diversi mitocondri di altri organismi, è probabile che vengano scoperte altre differenze di codifica.
Un codice mitocondriale identico a quello utilizzato nei batteri, ma diverso da quello utilizzato negli eucarioti, se così fosse, potrebbe avvalorare l’ipotesi dell’endosimbiosi. Tuttavia, la differenza effettivamente esistente ha spinto alcuni evoluzionisti a ipotizzare che il codice mitocondriale sia più “primitivo” e quello batterico più evoluto. Contrariamente a questa visione, la differenza è meglio spiegata dall’ipotesi che il codice mitocondriale sia progettato per le esigenze specifiche dei mitocondri, in particolare per impedire lo scambio di geni del mtDNA con geni nucleari.
Spostare il problema altrove
Un altro problema importante della teoria dell’endosimbiosi è che non risolve il problema dell’evoluzione degli organelli. Al contrario, evita il problema perché parte dall’esistenza di un sistema complesso e funzionante che non è in grado di spiegare. A fini argomentativi,
«… supponiamo che la simbiosi immaginata da Margulis sia stata in realtà un evento comune nel corso della storia della vita. La domanda importante per noi biochimici è: la simbiosi può spiegare l’origine di sistemi biochimici complessi?
«Chiaramente no. L’essenza della simbiosi è l’unione di due cellule separate, o due sistemi separati, entrambi già funzionanti. Nel caso del mitocondrio, una cellula vitale preesistente è entrata in una relazione simbiotica con un’altra cellula simile. Né Margulis né nessun altro ha offerto una spiegazione dettagliata di come si siano originate le cellule preesistenti.»
Inoltre, i sostenitori della teoria simbiotica devono «presupporre che le cellule invasive fossero già in grado di produrre energia da sostanze nutritive; Essi [presuppongono] esplicitamente che la cellula ospite fosse già in grado di mantenere un ambiente interno stabile che avrebbe avvantaggiato il simbionte.”
Margulis e Sagan proposero che le prime cellule eucariotiche fossero i protottisti: le amebe, le diatomee, le alghe giganti e le alghe rosse. Queste creature eucariotiche, tuttavia, sono per molti aspetti più simili agli eucarioti di livello “superiore” che ai procarioti. Anche se la teoria dell’endosimbiosi non si adatta ai fatti esaminati in questo articolo, essa viene comunque periodicamente riproposta quando le teorie alternative si rivelano errate.

Un’altra indicazione dei problemi dell’endosimbiosi è l’ampio disaccordo tra i ricercatori sui meccanismi alla base del concetto. Ad esempio, Margulis e Sagan notano che alcuni batteri sono stati rinominati “archei” da Carl Woese, una terminologia ora ampiamente accettata.
“Questa classificazione rifiuta l’endosimbiosi ed è una “negazione della loro natura batterica” perché Ciò comporta l’elevazione del gruppo degli “archei” a uno status paritario rispetto agli altri batteri e a tutti gli eucarioti. Il risultato è la formazione di tre gruppi fondamentali chiamati domini, o superregni, il che contraddice la teoria dell’endosimbiosi.58
Un’altra preoccupazione è che l’endosimbiosi implichi un’evoluzione relativamente rapida, in contrasto con l’evoluzione graduale darwiniana. Dato che non esiste una spiegazione gradualista plausibile per l’evoluzione degli eucarioti, gli evoluzionisti sono stati spinti a combinare posizioni gradualiste e non gradualiste darwiniane. O’Malley scrive che, come sosteneva Maynard Smith, l’endosimbiosi è spiegata dalle mutazioni standard e dalle macromutazioni.59 Altri ricercatori sostengono che si debbano tenere in considerazione altri fattori, rendendo l’evoluzione degli eucarioti ancora più complessa (e di conseguenza meno probabile). Ad esempio, Edgar aggiunge che “l’evoluzione concomitante del sistema redox dell’acido L-ascorbico dovrebbe essere considerata un fattore chiave che ha portato all’evoluzione degli eucarioti multicellulari e rimane coinvolta nel mantenimento della multicellularità e di molte altre caratteristiche eucariotiche”.
Perché l’evoluzione degli organelli è [praticamente] impossibile

Behe sostiene che il divario esistente tra cellule eucariotiche e procariotiche non potrà mai essere colmato a causa della complessità irriducibile. La complessità anche di una macchina semplice può essere ridotta solo fino a un certo punto: al di sotto di tale limite, la macchina non può funzionare. L’esempio classico è una trappola per topi standard, che deve avere almeno cinque componenti principali per funzionare: una piattaforma, una barra di supporto, un martello, un fermo e una molla.
Una trappola per topi non funzionerà finché non saranno presenti tutte le sue parti necessarie, ognuna delle quali deve essere progettata correttamente per articolarsi con le altre. Un’ipotesi contraria è quella di proporre che alcune di queste parti possano essere eliminate con vari metodi, come ad esempio inchiodando la trappola al pavimento. Questo approccio non elimina una parte, ma la sostituisce con un’altra; il pavimento viene utilizzato come base. Allo stesso modo, gli organelli non funzioneranno se non esiste ogni parte necessaria, correttamente progettata e prodotta, e tutte queste parti devono essere assemblate correttamente per formare un sistema operativo.
Gli organelli sono strutture molto complesse, costituite da migliaia di parti complesse più piccole, e il problema della complessità irriducibile è molto probabilmente valido anche per ogni singola parte di ciascun organello. Una cellula non può sopravvivere senza i ribosomi, ognuno dei quali contiene migliaia o addirittura decine di migliaia di molecole, ognuna delle quali deve essere assemblata secondo specifiche precise. Pertanto, la vita cellulare è impossibile finché tutte le sue parti necessarie non vengono prodotte e assemblate correttamente. Sebbene il DNA sia descritto come rappresentante di “un’intelligenza massiccia… [esso] non ha di per sé né futuro né presente. Il DNA senza una cellula che lo sostenga ed esprima non ha alcun significato fisiologico”.
Pochi scienziati si sono persino cimentati nello speculare sui dettagli delle forme di transizione tra gli ipotetici pre-organelli, per non parlare della presentazione di prove delle migliaia di forme di transizione necessarie a creare uno scenario plausibile che possa collegare le cellule a vita libera con le cellule dotate di organelli utilizzate negli organismi multicellulari.
Il sistema di trasporto cellulare è un altro esempio che illustra perché il concetto di complessità irriducibile renda impossibile l’evoluzione degli organelli. Dopo essere state prodotte, le proteine non fluttuano liberamente all’interno della cellula, ma devono essere trasportate da un meccanismo appropriato ovunque siano necessarie. Due meccanismi comuni per il trasporto delle proteine sono il trasporto controllato e il trasporto vescicolare.
Il trasporto controllato richiede la costruzione di una porta tra il citoplasma e la membrana nucleare e un sensore chimico (una proteina dotata del tag di identificazione corretto). Quando il pacchetto proteico si avvicina al sensore, questo apre la porta, permettendo alla proteina di passare. Questo meccanismo di controllo richiede che la proteina abbia il tag di identificazione appropriato e una porta programmata per aprirsi in risposta. La porta stessa contiene a sua volta molte parti, introducendo così un ulteriore livello di complessità irriducibile. Ciascuno di questi componenti del trasporto controllato è complesso e consiste di migliaia di parti a livello molecolare, tutte necessarie affinché il sistema di trasporto controllato funzioni.
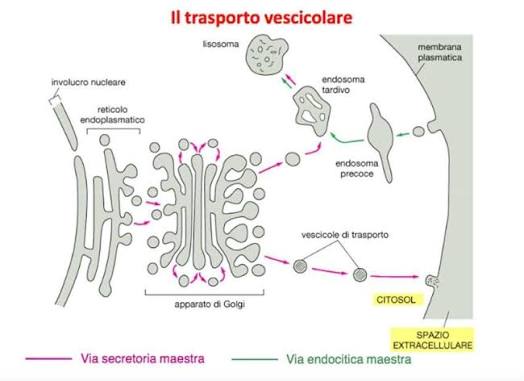
Anche il sistema di trasporto vescicolare utilizza una serie di sensori appositamente progettati. Invece di una porta, il tag di identificazione appropriato fa sì che la membrana del compartimento si prolunghi verso l’esterno, distaccandosi e formando una vescicola che circonda completamente la proteina. La vescicola di trasporto si sposta quindi verso una destinazione predeterminata dal suo tag di identificazione. Se il tag della vescicola e il sensore di identificazione corrispondono, un altro sensore riconosce la vescicola, che si fonde con il compartimento.
Successivamente, il processo di distacco viene invertito per consentire alle proteine di essere trasportate all’interno del nuovo compartimento. La complessità quasi certamente irriducibile del sistema deve includere due complessi sistemi di sensori, due tag di identificazione e il recipiente stesso. A un livello superiore, ogni tag di identificazione del sensore e i recipienti di trasporto sono, a livello molecolare, anch’essi costituiti da migliaia di componenti, ognuna delle quali è probabilmente un esempio di complessità irriducibile. La vescicola deve contenere tutte le strutture che le consentono di gemmare dal compartimento originale e poi di unirsi a un altro compartimento.
Altri problemi dell’endosimbiosi
La teoria standard dell’endosimbiosi è stata recentemente oggetto di critiche da più fronti e alcuni ricercatori stanno ora proponendo una nuova teoria per spiegare l’evoluzione degli organelli. Alcuni di questi scienziati ritengono che una nuova teoria “potrebbe risolvere alcuni problemi persistenti della teoria prevalente” dell’endosimbiosi. I dettagli di questa nuova teoria sono ancora vaghi. Ammettono che, anche se una nuova teoria fosse argomentata in modo elegante, probabilmente ci saranno molte difficoltà che una nuova ipotesi non prenderà in considerazione. Gli evoluzionisti concludono anche che potremmo dover ammettere che
“… il mitocondrio è stato un fortunato incidente. In primo luogo, la cellula ancestrale – probabilmente un archeobatterio, come suggeriscono recenti analisi genetiche – acquisì la capacità di inglobare e digerire molecole complesse. Iniziò a predare i suoi compagni microbici. A un certo punto, tuttavia, questa cellula predatrice non digeriva completamente la sua preda, e ne risultò una cellula ancora più efficiente quando un pasto destinato a diventare preda si stabilì permanentemente e divenne il mitocondrio.”
Inoltre, per decenni gli scienziati hanno creduto che
“… avessero esempi dei diretti discendenti di quegli eucarioti primitivi: alcuni protisti privi di mitocondri. Ma recenti analisi dei geni in questi organismi suggeriscono che anche loro un tempo possedevano mitocondri, ma li persero in seguito (Science, 12 settembre 1997, p. 1604). Questi risultati suggeriscono che gli eucarioti potrebbero in qualche modo aver acquisito i loro mitocondri prima di aver sviluppato la capacità di inglobare e digerire altre cellule.”
Un altro problema dell’endosimbiosi è l’ampio disaccordo tra i ricercatori sul meccanismo. Una sintesi ha concluso:
“… che i modelli ‘mitocondriali precoci’, che postulano l’acquisizione del protomitocondrio da parte di un ospite archeale, sono più plausibili dei modelli ‘mitocondriali tardivi’. Tuttavia, poiché i procarioti non sono in grado di effettuare la fagocitosi, tali modelli non sono riusciti a suggerire un meccanismo ragionevole attraverso il quale l’endosimbionte abbia avuto accesso al suo ospite.”
Una soluzione a tutti questi problemi è dare più tempo all’evoluzione. Come spiega il professor Edgar, ci sono voluti 0,5 miliardi di anni perché i procarioti (batteri e archei) sviluppassero “una biochimica piuttosto complessa e alcune caratteristiche eucariotiche”, ma
“… la transizione dai procarioti unicellulari agli eucarioti multicellulari e aerobici ha richiesto altri 2,5 miliardi di anni per iniziare. Il fattore o i fattori chiave che alla fine hanno causato questa transizione a lungo ritardata sono una questione che è stata al centro di numerose ricerche e un argomento di discussione per molti anni.”
Conclusioni:

Esistono due gruppi principali di organismi: procarioti ed eucarioti. Non sono mai stati rinvenuti organismi intermedi tra di loro, con organelli parzialmente sviluppati. Salvo possibili eccezioni minori, ciò che si trova è o l’assenza di organelli, oppure organelli completamente funzionali e sviluppati. “Non esistono anelli mancanti tra eucarioti e batteri, né nei fossili né negli organismi viventi.” Inoltre, è persino difficile ipotizzare, attraverso la costruzione di storie fantasiose, come possano esistere i legami tra procarioti ed eucarioti.
L’endosimbiosi postula che i mitocondri fossero un tempo batteri a vita libera e che “nelle prime fasi dell’evoluzione le cellule eucariotiche ancestrali si nutrissero semplicemente dei loro futuri partner.” Sia la teoria della conversione graduale che quella dell’endosimbiosi richiedono migliaia di forme di transizione, ognuna delle quali conferisce alla cellula un vantaggio competitivo rispetto alle cellule non modificate.
L’idea dell’endosimbiosi è popolare non per via delle prove empiriche, ma perché nessun’altra ipotesi è minimamente plausibile. La completa assenza di prove fossili e di altro tipo rappresenta un ulteriore problema. Pertanto, il professor Battley descrive la nozione di endosimbiosi come “provvisoria nella migliore delle ipotesi”. Un problema principale dell’endosimbiosi è che è sempre stata impossibile da verificare. Ulteriori ricerche e conoscenze hanno spinto un ricercatore all’avanguardia in questo campo a concludere nel 1998 che gli studi
“… pubblicati negli ultimi due o tre anni, molti dei quali derivanti da progetti di sequenziamento del genoma, suggeriscono che sia giunto il momento di una nuova teoria. In particolare, sta emergendo che i genomi nucleari eucariotici contengono molti geni di origine batterica (a volte α-proteobatterica) che non hanno nulla a che fare con le funzioni mitocondriali. Inoltre, gli eucarioti privi di mitocondri, che eravamo giunti a considerare diretti discendenti degli antichi proto-eucarioti, presentano geni mitocondriali nei loro genomi nucleari.”
La teoria dell’endosimbiosi è stata attaccata da molte altre parti, e senza dubbio questi attacchi continueranno. Il direttore del Laboratorio di Genetica dell’Università di Clemson ha definito l’endosimbiosi una teoria in crisi che, nella migliore delle ipotesi, spiega ben poco dell’evoluzione degli eucarioti dai procarioti. L’origine dell’intero nuovo sistema cellulare, gli eucarioti, rimane ancora da spiegare. Ha concluso:
“… le sequenze di molti genomi eucariotici mostrano ora chiaramente che i repertori genici necessari al funzionamento dei mitocondri non derivano dai batteri, ma sono straordinariamente unici per il tipo di organismo in cui si trovano. Sebbene esistano alcune somiglianze genetiche, queste corrispondenze sono plausibilmente spiegate dal concetto ingegneristico standard di riutilizzo del codice: codice comune per risolvere problemi simili. La grande quantità di dati genomici sta ora distruggendo completamente l’idea dell’evoluzione su tutti i fronti, persino nell’ambito dell’endosimbiosi, una delle teorie preferite dai laicisti.”
La teoria dell’endosimbiosi presenta seri problemi. I mitocondri differiscono dai batteri: geneticamente, strutturalmente, funzionalmente e operativamente. Inoltre, il tempo non si è rivelato utile per spiegare queste differenze all’interno dei parametri della teoria, ma ha solo accentuato il contrasto tra le due.
Questa analisi può solo delineare alcuni dei principali problemi relativi al tentativo di colmare il divario tra procarioti ed eucarioti attraverso il concetto di endosimbiosi. Per criticare efficacemente questa idea problematica, sarebbe necessario un intero articolo per ciascuna delle aree problematiche in questione.
Gabriel Bellizzi [CEO BioGenTech]
Riferimenti:
- Archibald, J., Endosymbiosis and eukaryotic cell evolution, Current Biology 25(19):R911–R921, 5 October 2015; R911.
- Helder, M., Endosymbiotic theory, Creation Dialogue 24(4):6, December 1997.
- Spuhler, J.N., Evolution of mitochondrial DNA in monkeys, apes, and humans, Yearbook of Physical Anthropology 31:15–48, 1988; p. 16.
- Davidov, Y. and Jurkevitch, E., Predation between prokaryotes and the origin of eukaryotes, Bioessays 31(7):748–757, July 31, 2009; p. 748.
- Spuhler, ref. 3, p. 18.
- Palmer, J. et al., It takes teamwork: how endosymbiosis changed life on earth, evolution.berkeley.edu/evolibrary/article/endosymbiosis_01, 2007.
- Berg, O. and Kirkland, C.G., Why mitochondrial genes are most often found in nuclei, Molecular Biology and Evolution 17(6):951–961, June 2000; p. 95, academic.oup.com/mbe/article/17/6/951/1037844.
- Davidov and Jurkevitch, ref. 4, pp. 748–749.
- Cavalier-Smith, T., Our symbiotic origins? The Sciences 37(4):46–47, 1997; p. 46.
- Margulis, L., Origin of eukaryotic cells: evidence and research implications for a theory of the origin and evolution of microbial, plant, and animal cells on the Precambrian earth, Yale University Press, New Haven, CT, 1970.
- Hoagland, M. et al., Exploring the Way Life Works: The science of biology, Random House, New York, p. 72, 1995.
- Spuhler, ref. 3, p. 19.
- Gray, M.W., Mitochondrial evolution, Cold Spring Harbor Perspectives in Biology 4(9):a011403, 2012.
- Spuhler, ref. 3, p. 15.
- Davidov and Jurkevitch, ref. 4, p. 748–749.
- Burger, G. and Lang, B.F., Parallels in genome evolution in mitochondria and bacterial symbionts, IUBMB Life 55(4–5):205–212, April–May 2003.
- Lang, B. et al., An ancestral mitochondrial DNA resembling a eubacterial genome in miniature, Nature 387:493–497, May 1997; p. 493.
- Kurland, C.G. et al., Genomics and the irreducible nature of eukaryote cells, Science, 312:1011–1014, 2006.
- Lang et al., ref. 17, p. 496.
- Davidov and Jurkevitch, ref. 4, p. 748.
- Lewin, B., Genes VI, Oxford University Press, Oxford, UK, 1997.
- Battley, E.H., Book review of Dexter et al., Tracing the History of Eukaryotic Cells: The enigmatic smile, Columbia University Press, New York, in The Quarterly Review of Biology 71(2):275–276, June 1994, p. 276.
- Vallès, Y. et al., Group II introns break new boundaries: presence in a bilaterian’s genome, PloS ONE 3(1):e1488, February 2008 ǀ doi:10.1371/journal.pone.0001488.
- See for example Bergstrom, C.T. and Dugatkin, L.A., Evolution, Norton, New York, pp. 442–445, 2016.
- Behe, M., Darwin’s Black Box: The biochemical challenge to evolution, Basic Books, New York, p. 189, 1996.
- Margulis, L., The origin of plant and animal cells, American Scientist 59(2):230–235, March–April 1971, p. 230.
- de Roos, A.A., Critique on the endosymbiotic theory for the origin of mitochondria, Telic Thoughts, 17 October 2007; researchgate.net/publication/322228552_A_critique_on_the_endosymbiotic_theory_for_the_origin_of_mitochondria.
- Mears, J.A., et al., A structural model for the large subunit of the mammalian mitochondrial ribosome, J. Molecular Biology 358(1):193–212, April 2006.
- O’Brien, T.W., Evolution of a protein-rich mitochondrial ribosome: implications for human genetic disease, Gene 286(1):73–79, 6 March 2002.
- O’Brien, T., Properties of human mitochondrial ribosomes, IUBMB Life 55(9):505–513. 2003.
- Mears et al., ref. 28, p. 193.
- Frank-Kamenetskii, M.D., Unraveling DNA: The most important molecule of life, Perseus Books, Reading, MA, p. 68, 1997.
- Sharma, M.R., et al., Structure of the mammalian mitochondrial ribosome reveals an expanded functional role for its component proteins, Cell 115(1):97–108, 2003.
- Smits, P., et al., Reconstructing the evolution of the mitochondrial ribosomal proteome, Nucleic Acids Research 35(14):4686–4703, July 2007.
- Davidov and Jurevitch, ref. 4, pp. 748, 750.
- Davidov and Jurevitch, ref. 4, p. 750.
- Davidov and Jurevitch, ref. 4, pp. 748–749.
- Wallace, D.C., Mitochondrial DNA in aging and disease, Scientific American, New York, 1997.
- Rizzuto, R., Mitochondria as sensors and regulators of calcium signaling, Nature Reviews Molecular Cell Biology 13:566–578, 2012.
- Hoffmeister, M. et al., Mitochondrial trans-2-Enoyl-CoA reductase of wax ester fermentation from Euglena gracilis defines a new family of enzymes involved in lipid synthesis, J. Biological Chemistry 280:4329–4338, 2004.
- Frank-Kamenetskii, ref. 32, p. 68–69.
- Bogenhagen, D.F. et al., Kinetics and mechanism of mammalian mitochondrial ribosome assembly, Cell Replication 22(7):1935–1944, February 13, 2018.
- Bogenhagen, ref. 42, p. 1935.
- Helder, ref. 2, pp. 6–7.
- de Duve, C., The birth of complex cells, Scientific American 274(4):50–57, April 1996, p. 57.
- Henze, K. and Martin, W., How do mitochondrial genes get into the nucleus? TRENDS in Genetics 17(7):383–387, July 2001, p. 383.
- Helder, ref. 2, p. 7.
- Henze and Martin, ref. 46, pp. 286–387.
- Gray, M.W., Mosaic nature of the mitochondrial proteome: implications for the origin and evolution of mitochondria, PNAS 112(33):10133–10138, August 2015, p. 10133.
- Gray, ref. 49, p. 10133–10134.
- Schatz, G., Just follow the acid chain, Nature 388(6638):121–122, July 1997, p. 121.
- Dietmeier, K. et al., Tom5 functionally links mitochondrial preprotein receptors to the general import pore, Nature 388:195–200, July 1997.
- Su, D. et al., An unusual tRNAThr derived from tRNAHis reassigns in yeast mitochondria the CUN codons to threonine, Nucleic Acid Research 39(11):4866–4874, 2011: ǀ doi:10.1093/nar/gkr073; and Gray, M. and Doolittle, W.F., Has the endosymbiont hypothesis been proven? Microbiological Reviews 46(1):1–42, 1982.
- Miranda, I. et al., Evolution of the genetic code in yeasts, Yeast 23(3):203–213, 2006.
- Behe, ref. 25, p. 189.
- Margulis, L. and Sagan, D., What is Life? Simon & Schuster, New York, p. 154, 1995.
- Margulis and Sagan, ref. 56, p. 91.
- Margulis, L. and Sagan, D., Acquiring Genomes: A theory of the origins of species, Basic Books, p. 154, 2002.
- O’Malley, M.A., Endosymbiosis and its implications for evolutionary theory, PNAS 112(33):10270–10277, 2015; p. 10270.
- Edgar, J.A., L-ascorbic acid and the evolution of multicellular eukaryotes, J. Theoretical Biology 476:62–73, September 2019.
- Huang, L., Massa, L., and Karle, J., Drug target interaction energies by the kernel energy method in aminoglycoside drugs and ribosomal A site RNA targets, PNAS 104(11):4261–4266. 2006.
- Kornberg, A., For the Love of Enzymes: The odyssey of a biochemist, Harvard University Press, Cambridge, MA, p. 316, 1989.
- Vogel, G., Did the first complex cell eat hydrogen? Science 279(5357):1633–1634, March 1998, p. 1633.
- Davidov and Jurkevitch, ref. 4, p. 755.
- Brown, C., Are there organelles in bacteria? CRSQ 42(2):129–130, September 2005.
- Margulis and Sagan, ref. 58, p. 139.
- Tuszynski J.A., Molecular and Cellular Biophysics, CRC Press, Boca Raton, FL, p. 207, 2007.
- Margulis, ref. 26, p. 230.
- Doolittle, W.F., A paradigm gets shifty, Nature 392:15–16, March 1998; p. 15.
- Mathieu, L.G. and Sonea, S., A powerful bacterial world. ScienceDirect (Endeavour) 19(3):112–117, 1995; sciencedirect.com/science/article/abs/pii/016093279597496U.
- Tomkins, J.P., Endosymbiosis: a theory in crisis, Acts & Facts 44(11):13, November 30, 2015.
