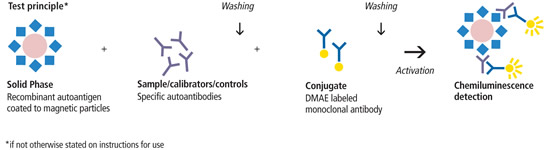
Corretta gestione estrogenica in TRT e nell'”Enhanced” [Bulk/Off-Season e Cut/Pre-Contest]
Introduzione agli aspetti pleiotropici degli estrogeni:
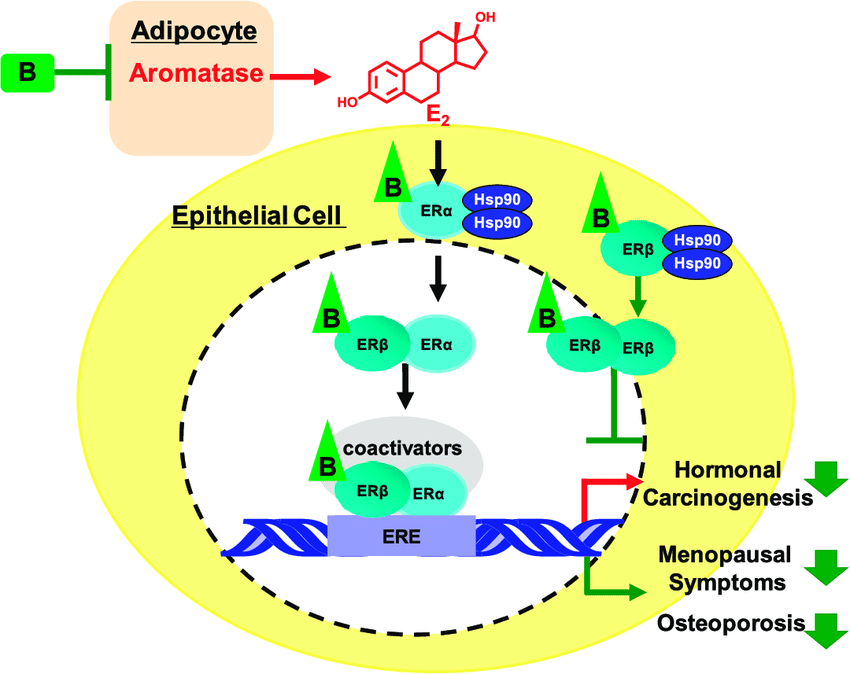
L’importanza della componente estrogenica, e nella fattispecie dell’estrogeno maggiormente attivo Estradiolo [E2], nel maschio è ormai nota e viene approfondita anche da studi di nuova pubblicazione.
Nei maschi, gli estrogeni esercitano effetti pleiotropici agendo su diversi tessuti e organi, tra cui il sistema riproduttivo. Nell’uomo, gli estrogeni sono in grado di esercitare la loro azione a diversi livelli attraverso il tratto riproduttivo e su diverse cellule riproduttive. Tuttavia, la regolazione della riproduzione maschile umana è complessa e il ruolo degli estrogeni è meno chiaro rispetto ai topi. Durante la vita fetale e perinatale, gli estrogeni agiscono sul sistema nervoso centrale modulando lo sviluppo di alcune aree cerebrali deputate al controllo del comportamento sessuale maschile in termini di definizione dell’identità di genere, sviluppo dell’orientamento sessuale ed evoluzione del normale comportamento sessuale maschile adulto. Questo effetto organizzativo e centrale degli estrogeni è particolarmente significativo in altre specie (soprattutto roditori e montoni), ma probabilmente meno importante negli uomini, dove i fattori psicosociali diventano più determinanti.[https://pmc.ncbi.nlm.nih.gov/]
L‘Estradiolo negli uomini è quindi essenziale per modulare la libido, la funzione erettile e la spermatogenesi. I recettori degli Estrogeni, così come l’Aromatasi, sono abbondanti nel cervello, nel pene e nei testicoli, organi importanti per la funzione sessuale. Nel cervello, la sintesi dell’Estradiolo è elevata nelle aree correlate all’attività sessuale. Inoltre, nel pene, i recettori degli Estrogeni si trovano in tutto il corpus cavernosum con elevata concentrazione intorno ai fasci neurovascolari. Un livello basso di Testosterone e elevato di Estrogeni aumenta l’incidenza della disfunzione erettile indipendentemente l’uno dall’altro. Nei testicoli, la spermatogenesi è modulata a tutti i livelli dagli Estrogeni, a partire dall’asse ipotalamo-ipofisi-gonadi, seguita dalle cellule di Leydig, Sertoli e germinali e terminando con l’epitelio ductale, l’epididimo e lo sperma maturo. La regolazione delle cellule testicolari mediante l’Estradiolo mostra sia un’influenza inibitoria che una stimolatoria, indicando un intricata sinfonia di modulazione dose-dipendente e temporalmente sensibile.[https://www.researchgate.net]
In genere si ritiene che gli estrogeni e il Testosterone siano i principali steroidi sessuali che regolano il metabolismo osseo rispettivamente nelle donne e negli uomini. Si sono infatti osservati uomini portatori di mutazioni omozigoti nel gene ER-alfa e uomini con mutazioni omozigoti nel gene dell’Aromatasi presentare osteopenia, epifisi non fuse e indici elevati di turnover osseo. Sebbene questi risultati indichino che gli estrogeni svolgono un ruolo nella regolazione dello scheletro maschile, hanno lasciato irrisolto il problema se gli estrogeni agiscano sullo scheletro maschile principalmente per migliorare l’acquisizione di massa ossea durante la crescita e la maturazione, o se agiscano anche per ritardare la perdita di massa ossea negli individui che invecchiano. Per risolvere questo problema, diversi studi osservazionali trasversali hanno messo in relazione la densità minerale ossea (BMD) con gli steroidi sessuali negli uomini anziani, scoprendo che gli estrogeni si correlavano meglio del Testosterone con la BMD. Inoltre, recenti studi longitudinali indicano che gli estrogeni biodisponibili si correlano meglio del Testosterone sia con l’aumento della BMD negli uomini giovani sia con la perdita di BMD negli uomini anziani. Tuttavia, questi studi osservazionali non dimostrano la causalità, che richiede studi interventistici diretti.[https://pubmed.ncbi.nlm.nih.gov/]
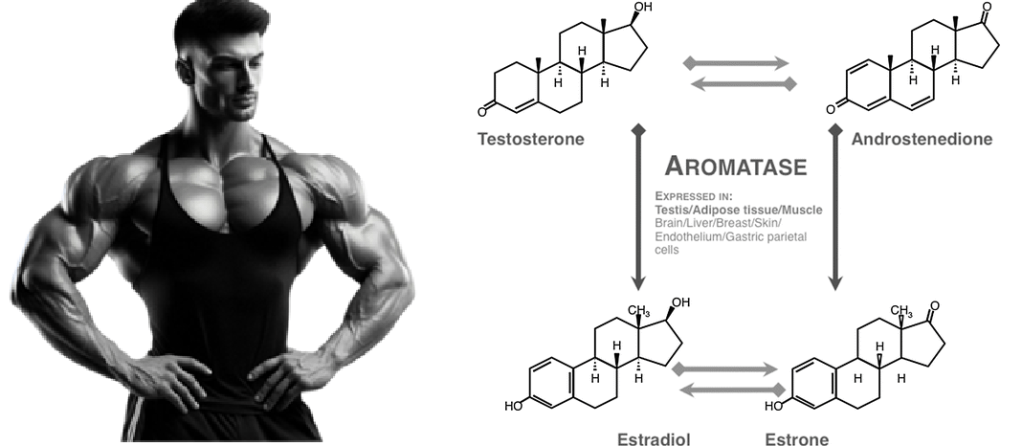
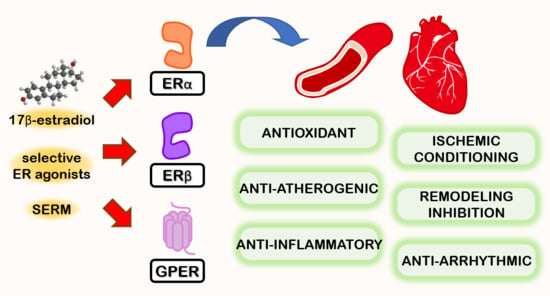
Sono stati osservati effetti favorevoli, oltre che a livello sessuale e sulla fisiologia ossea, sulla salute cerebrale e cardiovascolare correlata ad un adeguato livello di E2, mentre è stato seriamente sospettato un potenziale ruolo nella patologia prostatica dell’uomo che invecchia in seguito a dis-regolazione della T:E ratio. Gli estrogeni nell’uomo sono prevalentemente i prodotti dell’aromatizzazione periferica o in sede ghiandolare degli androgeni testicolari e surrenali. Gli estrogeni esercitano effetti sul cervello: sulla funzione cognitiva, sulla coordinazione dei movimenti, sul dolore e sullo stato affettivo, e sono forse protettivi nei confronti della malattia di Alzheimer. Gli effetti degli estrogeni sul sistema cardiovascolare includono quelli sui profili lipidici, sulla distribuzione del grasso, sui fattori endocrini/paracrini prodotti dalla parete vascolare (come endoteline, ossido nitrico), sulle piastrine, sui fattori infiammatori e sulla coagulazione.[https://pubmed.ncbi.nlm.nih.gov/]
Gli Estrogeni possono giocare un ruolo significativamente importante nella promozione di uno stato anabolico influenzando l’utilizzo del Glucosio nel tessuto muscolare. Ciò avviene attraverso un’alterazione del livello di glucosio 6-fosfato deidrogenasi disponibile, un enzima direttamente legato all’uso del glucosio per la crescita e il recupero del tessuto muscolare. Più specificamente, il G6PD è l’enzima che catalizza la prima reazione della via dei pentoso fosfati (definita anche Shunt dell’Esosomonofosfato [HMP shunt] o PPP da Pentose phosphate pathway), un processo metabolico citoplasmatico, parallelo alla glicolisi, in grado di generare NADPH e zuccheri pentosi (a 5 atomi di carbonio). Durante il periodo di rigenerazione tissutale seguente il danno muscolare, i livelli di G6PD aumentano considerevolmente, il che è ritenuto essere un meccanismo che il corpo attua per migliorare il recupero quando necessario. Sorprendentemente, si osserva che l’Estrogeno è direttamente legato al livello di G6PD che deve essere messo a disposizione delle cellule in questa fase di recupero. In sintesi, gli Estrogeni svolgono anche una azione metabolica accelerando la sintesi degli acidi nucleici, delle proteine e del glicogeno. Anche in questo caso, però, un livello eccessivo di E2 sembrerebbe poter causare alterazioni negative della sensibilità all’Insulina per via di una cronicità di stimolo e una sottoregolazione consequenziale.
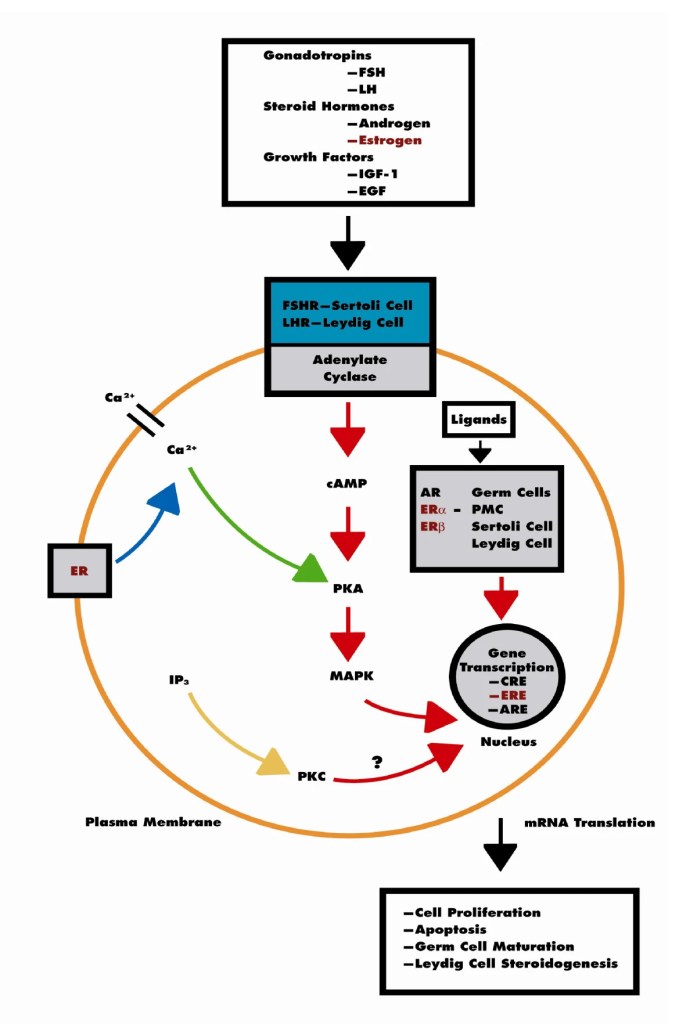
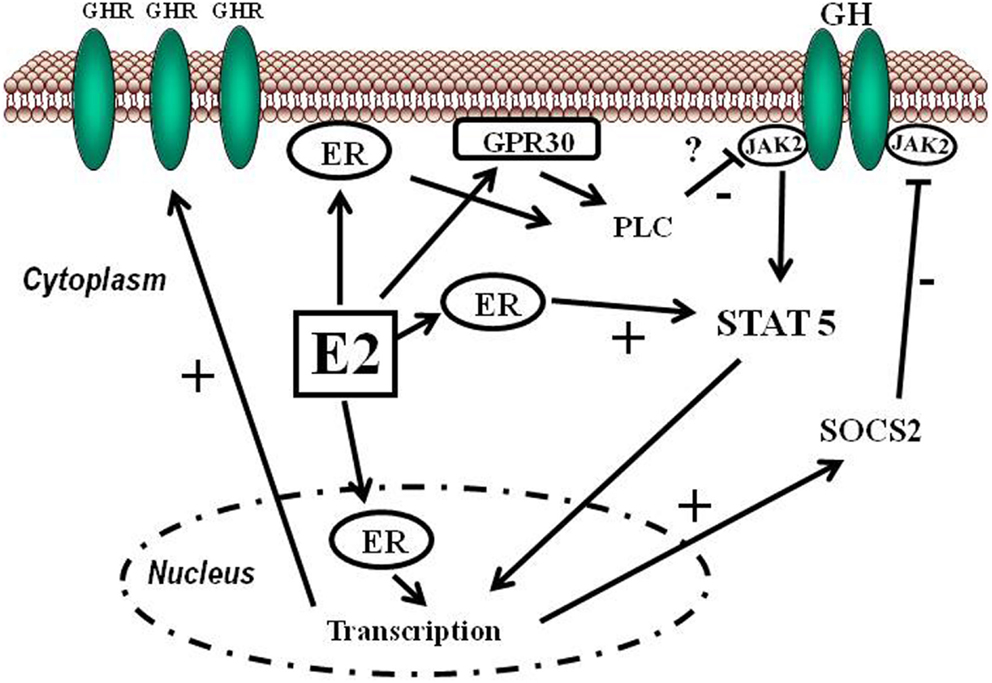
Sappiamo che gli estrogeni possono anche svolgere un ruolo importante nell’influenzare l’attività dell’Asse hGH/IGF-1. L’E2 ha un’affinità simile per l’ERα e l’ERβ e questi recettori sono attivati da un’ampia gamma di ligandi, tra cui i SERM (ad esempio, il Raloxifene e il Tamoxifene) e molti altri composti. L’ERβ è espresso nell’ovaio, nella prostata, nel polmone, nel tratto gastrointestinale, nella vescica, nel sistema ematopoietico e nel sistema nervoso centrale, mentre l’ERα è espresso principalmente nei tessuti riproduttivi, nel rene, nell’osso, nel tessuto adiposo bianco e nel fegato. Il fegato esprime ERα ma livelli quasi irrilevanti di ERβ, il che indica che le azioni specifiche degli estrogeni nel fegato possono essere imitate utilizzando agonisti selettivi di ERα come il Propilpirazolo-triolo (PPT) (Lundholm et al., 2008). Nel complesso, i dati sopra citati indicano che i meccanismi coinvolti nella segnalazione ER sono influenzati dal fenotipo cellulare, dal gene bersaglio e dall’attività o dal crosstalk con altre reti di segnalazione. Il fegato rappresenta un sito in cui si possono sviluppare interazioni fisiologicamente e terapeuticamente rilevanti tra estrogeni e hGH. Particolarmente rilevante è l’interazione degli estrogeni con la via di segnalazione GHR-JAK2-STAT5 nella regolazione della crescita somatica, del metabolismo dei lipidi e del glucosio e della “sessualità epatica”.
L’E2 può regolare le azioni del hGH nel fegato modulando la reattività del hGH, che comprende cambiamenti nell’espressione del GHR epatico e il crosstalk con la via di segnalazione JAK2-STAT5 attivata dal hGH. Gli estrogeni possono indurre l’espressione di SOCS2, che a sua volta inibisce negativamente la via di segnalazione GHR-JAK2-STAT5.
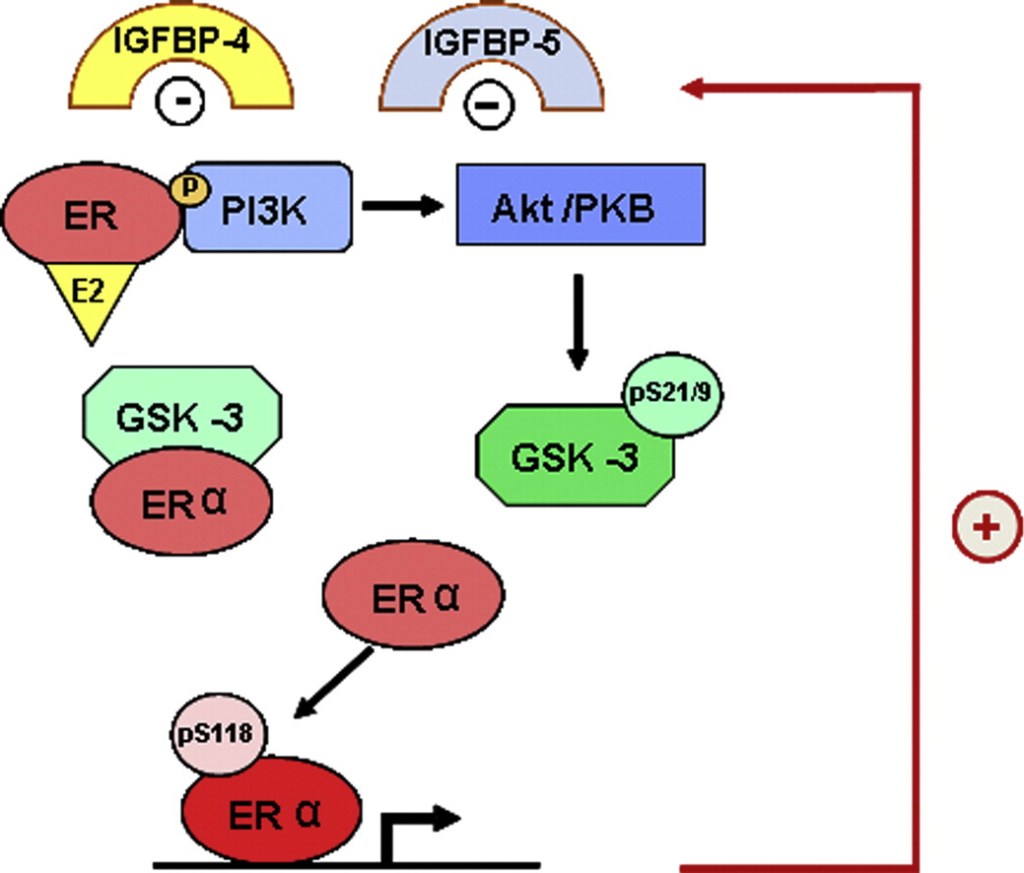
Inoltre, le differenze di composizione corporea legate al genere sono in parte mediate dagli steroidi sessuali che modulano l’Asse hGH/IGF-I (LeRoith, 2009; Rogol, 2010; Birzniece et al., 2011). Ciò è supportato dall’osservazione che le differenze di genere nella composizione corporea emergono al momento della crescita puberale. Inoltre, l’efficienza dell’attività del hGH è modulata anche dagli estrogeni in età adulta. Questo è esemplificato dal fatto che le donne sono meno reattive degli uomini al trattamento con hGH (Burman et al., 1997); il trattamento con hGH induce un maggiore aumento della massa magra e una diminuzione della massa grassa, o un maggiore aumento degli indici di turnover osseo e della massa ossea, nei pazienti GHD maschi rispetto alle femmine. È rilevante per la fisiologia del hGH l’alterazione della biodisponibilità dell’IGF-I dovuta alla somministrazione orale di dosi farmacologiche di estrogeni [rivisto da Leung et al., 2004]. La disponibilità e l’attività tissutale dell’IGF-I sono regolate dalle proteine leganti l’IGF (IGFBPs) (Kaplan e Cohen, 2007; LeRoith e Yakar, 2007; Ohlsson et al., 2009).
L’IGF-I circola quasi interamente come complesso ternario legato all’IGFBP-3 e all’ALS, entrambi fortemente regolati dal hGH nel fegato. Questo complesso ternario regola la biodisponibilità dell’IGF-I. Anche l’IGFBP-1 è una proteina di derivazione epatica che lega la piccola frazione di IGF-I libero e attenua l’effetto ipoglicemizzante del fattore di crescita (Lewitt et al., 1991). In contrasto con il suo effetto soppressivo sulla SLA e sull’IGF-I, la somministrazione orale di estrogeni aumenta l’IGFBP-1 circolante. Si può prevedere che l’effetto dell’aumento di IGFBP-1 riduca ulteriormente la frazione libera di IGF-I, con conseguente riduzione della sua attività. È interessante notare che l’attivazione della segnalazione GH-STAT5b induce l’espressione di ALS e IGF-I, ma inibisce IGFBP-1 (Ono et al., 2007). Pertanto, l’inibizione della via di segnalazione GHR-JAK2-STAT5 nel fegato (vedi sotto), molto probabilmente contribuisce agli effetti degli estrogeni su IGF-I, ALS e IGFBP-1. Pertanto, gli estrogeni esercitano effetti profondi sulle IGFPB derivate dal fegato quando vengono somministrati per via orale, che molto probabilmente modificano le azioni biologiche dell’IGF-I. Inoltre, la somministrazione orale di dosi farmacologiche di estrogeni può inibire gli effetti metabolici regolati dal GH (ad esempio, ossidazione lipidica, sintesi proteica) (Huang e O’Sullivan, 2009). Questi effetti sul metabolismo e sulla composizione corporea sono attenuati dalla somministrazione transdermica, suggerendo che il fegato è il principale sito di controllo regolatorio da parte degli estrogeni. Si rammenta, però, che in una condizione di iperestrogenemia [>45-60pg/dL] porta automaticamente ad un aumento delle concentrazioni di E2 a livello epatico con interferenze negative sull’Asse hGH/IGF1.
Gli estrogeni possono modulare le azioni del hGH sul fegato attraverso la modulazione della responsività al hGH, che include cambiamenti nell’espressione del GHR epatico e il crosstalk con la via di segnalazione JAK2-STAT5 attivata dal GH (Leung et al., 2004). In particolare, l’E2 può indurre l’espressione di SOCS2 e SOCS3, che a sua volta regola negativamente la via di segnalazione GHR-JAK2-STAT5, con conseguente riduzione dell’attività trascrizionale nel fegato. Pertanto, oltre alla regolazione da parte dell’E2 del modello di dimorfismo sessuale della secrezione ipofisaria di hGH, l’induzione dell’espressione di SOCS e l’inibizione della segnalazione JAK2-STAT5 è un meccanismo molto rilevante che, in parte, potrebbe spiegare come gli estrogeni inibiscano direttamente gli effetti del hGH in diverse azioni regolate da STAT5 (ad esempio, crescita somatica, composizione corporea, metabolismo e funzioni epatiche legate al sesso). Ipoteticamente, anche altri membri dei regolatori negativi della famiglia STAT possono contribuire all’interazione degli estrogeni con la segnalazione del hGH nel fegato. Ciò si spiega con la stimolazione da parte di ERα dell’espressione di PIAS3, che si lega e blocca l’attività di legame al DNA di STAT3. È interessante notare che l’attivazione di ER da parte di E2, seguita dall’interazione diretta di ER con STAT5, può anche inibire l’attività trascrizionale STAT5-dipendente (Faulds et al., 2001; Wang et al., 2004). D’altra parte, è stato dimostrato che l’attivazione di ERα o ERβ da parte di E2, attraverso meccanismi non genomici, induce un programma trascrizionale STAT5 (e STAT3) dipendente nelle cellule endoteliali (Bjornstrom e Sjoberg, 2005). Nel complesso, questi studi hanno dimostrato l’esistenza di un’interazione diretta tra la segnalazione di ER e STAT5, dimostrando inoltre che le conseguenze funzionali di questo crosstalk dipendono dal preciso contesto dell’ambiente intracellulare.
Se vi sentite confusi per ciò che è stato detto, dal momento che siete stati convinti da numerosi “top coach” che l’E2 deve rimanere alto per garantire una migliore risposta anabolica complessiva, e che i SERM causano una sottoregolazione dell’Asse hGH/IGF1 in quanto antagonizzano i ER, beh… le cose sono più complesse di così. Ad esempio, il Tamoxifene ha effetti prevalentemente antiestrogenici nelle mammelle, ma prevalentemente estrogenici nell’utero e nel fegato. Egualmente, il Raloxifene ha un’attività estrogenica in alcuni tessuti, come le ossa e il fegato, e un’attività antiestrogenica in altri tessuti, come il seno e l’utero. Ed è quindi l’aumento dell’attività recettoriale indotta da E2 o SERM ha causare alterazioni dell’Asse hGH/IGF1. In conclusione, la regolazione del E2 è essenziale per garantire ottimali risposte dell’Asse hGH/IGF1, evitando eccessi o cali eccessivamente prolungati e/o fuori contesto.
Un altro beneficio correlato a ottimali livelli di E2 riguarda la possibilità di indurre un aumento della concentrazione dei AR in alcuni tessuti. Ciò è stato dimostrato in studi svolti su ratti che hanno esaminato gli effetti degli Estrogeni sui AR cellulari in animali sottoposti ad orchiectomia (rimozione dei testicoli, spesso effettuata per diminuire la produzione endogena di Androgeni). Secondo lo studio, la somministrazione di Estrogeni ha determinato un aumento del legame recettoriale nel muscolo levator ani del Metribolone pari al 480%. E qui c’è il primo “inghippo” dal momento che il levator ani è parte dell’apparato sessuale dell’animale e non è paragonabile al muscolo-scheletrico umano. Infatti, lo studio ha esaminato l’effetto sui AR dato dagli Estrogeni nei tessuti musco scheletrici veloci (tibialis anterior e extensor digitorum longus), ma senza notare lo stesso aumento visto nel levator ani.[Modulation of the cytosolic androgen receptor in striated muscle by sex steroids. Endocrinology. 1984 Sep;115(3):862-6.] Se vi sia riscontro nell’uomo questo è comunque limitato alle possibilità di espressione genica del soggetto e, probabilmente, l’incremento degli AR non risulterebbe paragonabile a quello sperimentabile con dosi sovrafisiologiche di AAS.
Recettore degli Androgeni.
Sicuramente il mancato controllo del fattore estrogenico è tanto deleterio quanto può esserlo una sua soppressione marcata. Un’altra importante funzione degli estrogeni in ambo i sessi è la sua capacità di promuovere uno stato mentale di vigilanza. L’abuso di AI, con la marcata soppressione del E2 consequenziale, si manifestano stati di stanchezza. In tali condizioni, l’atleta, anche se sta seguendo un ciclo correttamente formulato, potrebbe non essere in grado di massimizzare i propri risultati di miglioramento della condizione fisica a causa di un’incapacità di allenarsi con pieno vigore. Questo effetto è talvolta anche soprannominato “letargia steroidea” o “Steroid Fatigue”. La ragione principale per cui ciò accade è legata all’importante azione di supporto all’attività della Serotonina data dall’E2. La Serotonina è uno dei principali neurotrasmettitori del corpo, di essenziale importanza per un adeguata lucidità mentale e un regolare ciclo sonno / veglia.[ Effect of estrogen-serotonin interactions on mood and cognition. Zenab Amin et al. Behav Cogn Neurosci Reviews 4(1) 2005:43-58] L’alterazione di questo neurotrasmettitore è associata anche alla sindrome da affaticamento cronico, e ciò ci fa comprendere quanto possa essere incisiva in particolare per la stanchezza. L’abbassamento dei livelli estrogenici nella menopausa è stata associata anche alla stanchezza, così come l’uso clinico di inibitori dell’aromatasi più recenti (e più potenti) come l’Anastrozolo, il Letrozolo, l’Exemestane, e il Fadrozolo in alcuni pazienti. L’uso di AAS non aromatizzabili e/o SARM non steroidei possono causare questo effetto, il quale, in questa circostanza, è dovuto alla soppressione/sottoregolazione della produzione endogena di Testosterone (riduzione del substrato principale nell’uomo per la sintesi di E2 per via della attività dell’enzima Aromatasi).
Gli estrogeni possono influire sui livelli di colesterolo, influenzando potenzialmente la salute cardiovascolare. In particolare, gli estrogeni possono influire sul colesterolo LDL (“cattivo”) e sul colesterolo HDL (“buono”); alcuni studi suggeriscono un legame tra livelli più elevati di estrogeni e una diminuzione del LDL e un aumento del HDL negli uomini. Il termine “elevati” non dovrebbe essere fuorviante dal momento che alterazioni metaboliche come quelle descritte in precedenza e correlate ad un eccessivo incremento del E2, potrebbero vedere sensibilmente ridotto questo effetto positivo.
In fine, sappiamo che se non si dispone di una quantità sufficiente di estrogeni rispetto ai livelli di androgeni nell’organismo, i livelli di cardiotossicità e neurotossicità saranno significativamente più alti di quelli che si avrebbero se si mantenessero livelli ottimali di estrogeni. Dal punto di vista del Bodybuilding, gli estrogeni a livello ottimale (tarato sul soggetto) sono necessari per ottimizzare la crescita muscolare, l’insulino-sensibilità e la sintesi di IGF-1 e fattori di crescita/segnalazione cellulare. Ricordiamoci, però, che un dosaggio fisiologico di Testosterone, e sua successiva aromatizzazione, risulta essere neuroprotettivo. Il Testosterone amplificava la neurotossicità solo a dosaggi sovrafisiologici anche senza AI. Sebbene l’aromatizzazione del Testosterone in E2 prevenga, seppur non marcatamente, una quantità significativa di danno neuronale, si può osservare chiaramente che le concentrazioni sovrafisiologiche di Testosterone esacerbano la neurotossicità in ogni caso e che i livelli sovrafisiologici di estrogeni non forniscono un aumento dose-dipendente della neuroprotezione.
Ci sono sempre dei limiti… sempre un fio da pagare. Sicuramente, come si può ben capire, un incremento spropositato di E2 in condizione di sovrafisiologia d’uso di AAS non garantirebbe altro che il sommarsi di altri effetti avversi.
Dopo questa “carrelata” di effetti positivi e limiti annessi al E2, e agli estrogeni in generale, possiamo entrare nel vivo della questione “gestione degli estrogeni” partendo dalla clinica, ossia dalla TRT.
Gestione estrogenica in TRT
Condizioni di bassi livelli di Testosterone si verificano nel 6-25% degli uomini. Secondo le linee guida dell’American Urological Association, il trattamento del Testosterone basso [vedi TRT] è indicato se i livelli sono inferiori alla soglia di 300ng/dL [3ng/mL] con segni o sintomi associati.2 Uno degli effetti collaterali più comuni riscontrabili nei pazienti in TRT, oltre alle variazioni dell’Ematocrito, è l’aumento dell’E2 attraverso l’attività dell’enzima Aromatasi (aromatizzazione), che converte il Testosterone in E2.
Sebbene la manifestazione di questa problematica può essere esacerbata o scatenata dalla cosomministrazione di hCG, le prime misure adottate per la sua gestione comprendo 1) modifiche nel dosaggio e/o nella cadenza di somministrazione e 2) modificando, se vi sono le circostanze a richiederlo, lo stile di vita del paziente portandolo ad una riduzione della massa grassa e, di conseguenza, dell’espressione dell’Aromatasi. Tuttavia, ciò potrebbe non essere sufficiente.
Si è notato, infatti, in diversi casi, che il tasso di aromatizzazione subiva un aumento nel corso del primo anno di trattamento clinico. Questo aumento non sembra essere correlato ad un aumento sensibile della massa grassa. Si è, di conseguenza, ipotizzato che tale risposta facesse parte di un adattamento epigenetico (o un tentativo in tal senso) in risposta a concentrazioni stabili di Testosterone in cronico. In definitiva, in questi casi, anche l’uso di ancillari steroidei [vedi Mesterolone e Drostanolone] non mostra miglioramenti assoluti nei livelli di E2, sebbene tissutalmente il Drostanolone a 100mg/settimana abbia mostrato una egregia attività controllo sull’azione estrogenica.
Molecola di Anastrozolo
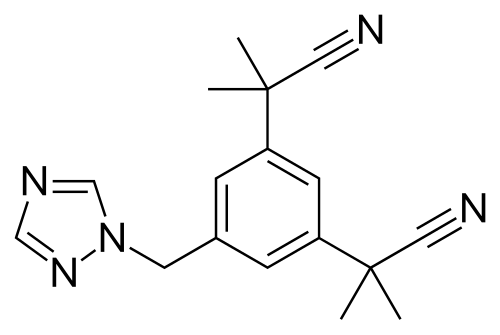
In questi casi la soluzione per il paziente è l’aggiunta di un inibitore dell’Aromatasi come l’Anastrozolo (AZ). L’Anastrozolo è un triazolo benzilico non steroideo. È noto anche come α,α,α’,α’-tetrametil-5-(1H-1,2,4-triazolo-1-ilmetil)-m-benziacetonitrile. L’Anastrozolo è strutturalmente correlato al Letrozolo, al Fadrozolo e al Vorozolo, tutti classificati come azoli.[Environmental Health Perspectives: Supplements.]
Struttura molecolare dell’Enzima Aromatasi
L’Anastrozolo agisce legandosi reversibilmente all’enzima Aromatasi e, attraverso un’inibizione competitiva, blocca la conversione degli androgeni in estrogeni nei tessuti periferici (extragonadici). È stato riscontrato che il farmaco raggiunge un’inibizione dell’Aromatasi compresa tra il 96,7% e il 97,3% al dosaggio di 1 mg/die e il 98,1% al dosaggio di 10mg/die nell’uomo. [Pertanto, 1 mg/die è considerato il dosaggio minimo necessario per ottenere la soppressione massima dell’Aromatasi con l’Anastrozolo. Questa diminuzione dell’attività dell’Aromatasi si traduce in una riduzione di almeno l’85% dei livelli di E2 nelle donne in postmenopausa. I livelli di corticosteroidi e altri steroidi surrenali non sono influenzati dall’Anastrozolo.
Bisogna specificare, però, che soggetti i quali mantengono una attività gonadale con l’uso di hCG durante la TRT risultano meno responsivi al tasso di inibizione del farmaco. Questo accade, con buona probabilità, perché una parte sostanziale dell’Estradiolo è prodotta dall’attività dell’Aromatasi nei testicoli. Nei testicoli, le concentrazioni di Testosterone [stimolate in questo caso dal hCG] arrivano a livelli circa 100 volte superiori a quelli presenti nel circolo ematico. Poiché gli AI devono inibire in modo competitivo l’Aromatasi, i dosaggi potrebbero dover essere più alti per portare a una significativa inibizione enzimatica nei testicoli. Infatti, in questi casi, la somministrazione giornaliera di Anastrozolo a 0,5 e 1mg porta ad una diminuzione dei livelli di Estradiolo di circa il 50% .
Molecola di Letrozolo
Interessante, a tal proposito, è il risultato osservato dopo 28 giorni di trattamento con Letrozolo alla dose di 2,5mg/die, dove i livelli di Estradiolo hanno subito una riduzione del 46% negli uomini giovani e del 62% negli uomini anziani. Uomini anziani con una ridotta attività gonadale.[https://pmc.ncbi.nlm.nih.gov/]
In generale, comunque, l’AZ è indicato per ritardare la maturazione epifisaria nei ragazzi adolescenti, ma è stato utilizzato nei maschi infertili con alterazioni dei livelli di Testosterone e livelli elevati di E2 per preservare la spermatogenesi. Il profilo degli effetti collaterali dell’AZ è stato ampiamente studiato nelle donne ed è un farmaco ben tollerato.4 L’elevato livello di E2 è più comunemente associato alla ginecomastia, ma non è l’unico meccanismo responsabile sebbene risulti il primario.5 L’AZ è stato utilizzato per trattare gli uomini con E2 elevato, ma attualmente esistono prove e/o linee guida limitate per la gestione ottimale degli elevati livelli di E2 negli uomini in TRT.
Nell’uomo vi sono due tipi principali di estrogeni: il potente Estradiolo e il meno potente, in senso di attività biologica, Estrone [E1]. Le quantità sono misurate in picogrammi per millilitro (pg/ml). Le medie tipiche di ciascuno di essi sono:
In uno studio pubblicato su Sex Med nel 2021, è stato riportato che su 1708 uomini in TRT, 51 (3%) sono stati trattati con AZ (AZ+). Di questi, 7 (14%) sono stati esclusi (3 precedentemente in trattamento con AZ e 4 con storia di cancro al seno). Un totale di 44 (2,6%) ha mostrato livelli elevati di estradiolo (range 40-165 pg/mL) ed è stato incluso. Il tempo mediano di somministrazione della TRT prima dell’inizio dell’AZ è stato di 11,96 mesi (IQR 4,63-31,44). I restanti 1657 uomini (97,0%) non hanno ricevuto il trattamento con AZ (AZ-).
Di conseguenza, in tale studio, solo il 2,6% degli uomini presentava un E2 tale da giustificare il trattamento con AZ. L’AZ è stato utilizzato negli uomini con una deficienza del Testosterone nel tentativo di ridurre la conversione del T in E2, pur mantenendo la fertilità.6 E’ stato utilizzato un cut-off per l’uso dell’AZ, ma la letteratura non supporta un cut-off chiaro o un’indicazione per l’inizio della terapia di abbassamento dell’E2. Similmente, in un campione nazionale, il 3,5% (1.200/34.016) degli uomini è stato trattato con un AI.
Sebbene gli uomini dello studio che ricevevano AZ, e quindi con un E2 più alto, avevano tassi maggiori, anche se non statisticamente significativi, di OSA e che tale condizione è correlato all’obesità e che gli uomini con obesità hanno livelli fisiologici maggiori di Aromatasi, aumentando così la conversione di T in E2, in corso di TRT anche in soggetti normopeso e con body fat contenute (se non basse) possono presentare condizioni di iperestrogenemia in specie dopo 12 mesi di trattamento. Probabilmente, questa risposta adattativa è la conseguenza di un tentativo di adattamento regolatorio della omeostasi ormonale in una condizione dove i livelli di T sono stabili e non caratterizzati [come nei soggetti in fisiologia funzionale] da fluttuazioni nelle 24h e nei mesi dell’anno [vedi variabili stagionali].
Un dato importante che andrebbe preso in considerazione, è che nello studio i soggetti non trattati con AZ avevano una terapia a base di Testosterone in soluzione ad uso topico, mentre gli uomini che utilizzavano AZ erano sottoposti ad una terapia a base di Testosterone somministrato per via intramuscolare. Ciò riflette alcuni dati che suggeriscono che aumenti statisticamente significativi di E2 sono stati osservati negli uomini in terapia iniettiva 3 mesi dopo l’inizio del TRT.
Quasi tutti gli uomini trattati con AZ hanno avuto un recupero dell’E2 entro i livelli normali (<40 pg/mL) con il contemporaneo mantenimento di un T sierico. Ciò evidenzia l’importanza di una regolazione appropriata, dati gli importanti processi fisiologici regolati dall’E2.5 Mentre l’AZ è comunemente usato come terapia aggiuntiva per aumentare i livelli di T negli uomini in cui è importante la conservazione della spermatogenesi e quindi evitare il Testosterone esogeno è fondamentale in diversi casi, nello studio discusso gli AI non hanno avuto un impatto sui livelli di Testosterone. Tuttavia, altri studi suggeriscono che la co-somministrazione di un’IA con Testosterone esogeno aumenta il T sierico. Tutti questi pazienti, però, hanno ricevuto un impianto di T, il che suggerisce una risposta differenziale in base al tipo di TRT.
I soggetti trattati con AZ hanno ricevuto una dose del AI pari a 0,5mg tre volte alla settimana (off-label). La regressione logistica è stata utilizzata per determinare i fattori predittivi di una maggiore probabilità di risposta al trattamento con Anastrozolo, definita come punteggio composito (riduzione dell’Estradiolo a meno di 60pg/mL e diminuzione di 20pg/mL dei livelli di Estradiolo).
Punti chiave per la gestione delE2 in TRT:
La modifica primaria è la riduzione della body fat;
Controllo più accurato del dosaggio terapeutico di Testosterone;
Controllo e/o modifica della dose e somministrazione di hCG [nonché valutarne i pro e i contro in funzione anche dell’età del paziente];
Aggiunta di un ancillare steroideo a dosi minime efficaci con attività di controllo del E2 [vedi, ad esempio, il Drostanolone o il Mesterolone];
L’ultimo intervento consiste nell’inserimento di un AI come l’Anastrozolo al dosaggio di 0,5mg per 3 volte a settimana.
Gestione estrogenica nell'”Enhanced”:
Trattato l’aspetto clinico della gestione estrogenica in contesto TRT/HRT, passiamo ora al contesto PEDs o, meglio, al contesto Enhanced BodyBuilding.
Il mondo della cultura fisica tout court non è mai stata immune alle mode generate da convinzioni rese “dogmatiche” perché affermate dal “guru” del settore. Questa tendenza ha e colpisce anche la questione della gestione estrogenica. Si passa dalla fobia e tendenza a mantenere i livelli di E2 cronicamente bassi all’esatto opposto con iperestrogenemia mantenuta con la convinzione che l’atleta ne gioverà. E’ superfluo dire che ambo le posizioni sono errate.
Come abbiamo potuto appurare all’inizio del presente articolo, gli estrogeni, ed in particolare l’E2, hanno una attività pleiotropica. Quindi? Quindi, ciò significa che può avere effetti multipli e, a volte, apparentemente non correlati, sullo sviluppo di determinate risposte organiche. Anche in questo caso, la frase mal tradotta e diffusa attribuita a Paracelso “E’ la dose che fa il veleno” può benissimo essere applicata anche al fattore estrogenico. E dal momento che ogni Preparatore dovrebbe avere una formazione di Biologia, Biochimica, Farmacologia, Andrologia e Endocrinologia, esso deve indirizzare tali conoscenze al fine di comprendere il soggetto interessato e operare al fine di trovare un settaggio ideale di E2. In poche parole, l’atleta dovrebbe essere un nuovo libro con informazioni aggiuntive e a se stanti.
Ricordiamoci, quindi, che negli uomini in fisiologia, i testicoli producono circa il 20% degli estrogeni circolanti. Il resto proviene dalla produzione locale da parte dei tessuti adiposi, cerebrali, cutanei e ossei, che esprimono l’Aromatasi (T ⇒ E₂).. Le concentrazioni di T nel sangue periferico degli uomini, pari a ~20nM, sono di almeno due ordini di grandezza superiori alle concentrazioni di E₂ (30-200pM). [Cooke PS, Nanjappa MK, Ko C, Prins GS, Hess RA. Estrogens in Male Physiology. Physiol Rev. 2017 Jul].
Mentre gli estrogeni esogeni e un loro eccesso endogeno causano patologie riproduttive maschili, gli estrogeni endogeni nel giusto assetto sono fondamentali per il funzionamento sessuale maschile. In parte, ciò è dovuto a un drastico aumento della SHBG (che riduce la biodisponibilità del T). [Damewood MD, Bellantoni JJ, Bachorik PS, Kimball AW Jr, Rock JA. Exogenous estrogen effect on lipid/lipoprotein cholesterol in transsexual males. J Endocrinol Invest. 1989 Jul-Aug;12(7):449-54.].
Un punto a sfavore della scelta deleteria di ridurre marcatamente l’E2 in cronico risiede senza dubbio nell’effetto positivo degli estrogeni nella lipidemia ematica.
Sappiamo che gli effetti degli AAS sui lipidi sono modulati attraverso:
(A) ↑ l’attività della trigliceride lipasi epatica (HTGLA), riducendo così le lipoproteine ad alta densità (↓HDL-C), e
(B) ↑Apo B, quindi ↑LDL-C.
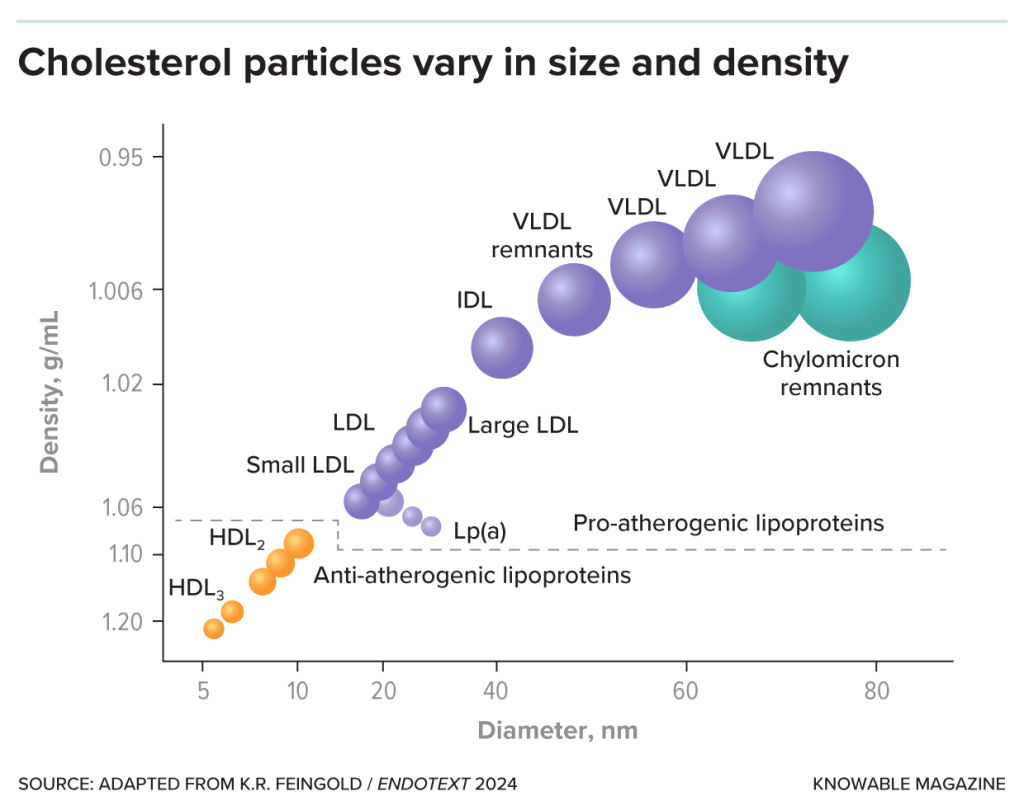
In primo luogo, per quanto riguarda (A): la lipasi epatica (HL) è un enzima secreto dal fegato che libera gli acidi grassi dal triacilglicerolo e dai fosfolipidi che fanno parte delle lipoproteine, comprese le lipoproteine ad alta densità (HDL). Gli AAS, aumentando la sua attività (HTGLA), inducono un passaggio da HDL₂ più grandi a HDL₃ più piccole, suscettibili di ulteriore degradazione, e quindi riducono le HDL. [Thompson, P. D. (1989).].
In secondo luogo, per quanto riguarda (B): l’Apo B è fortemente associata alle lipoproteine a bassissima densità (VLDL; una classe di particelle maggiormente aterosclerotica) e fornisce indicazioni sulle LDL effettivamente presenti nella circolazione sanguigna, forse perché gli AAS aumentano la secrezione epatica di queste lipoproteine. [Hartgens, F. (2004).].
Sezione della tabella presa da “Bond P, Smit DL, de Ronde W. Anabolic-androgenic steroids: How do they work and what are the risks? Front Endocrinol (Lausanne)”
La dislipidemia è caratterizzata da ↑LDL-C e ↓HDL-C.
Struttura molecolare della HTGLA
Gli AAS 17α-alchilati non aromatizzabili (metilati in C17), poiché non sono aromatizzabili o sono resistenti all’aromatizzazione e quindi non apportano benefici estrogenici ai lipidi (estrogeni ↓HTGLA), e poiché sono metabolizzati principalmente nel fegato, avendo maggiori effetti sulle proteine epatiche (ad es., HL), sono più dislipidemici e aterosclerotici degli androgeni parenterali (soprattutto Testosterone). [Friedl, K. E., Hannan, C. J., Jones, R. E., and Plymate, S. R. (1990).].
In particolare, i metilati in C-17 possono ↑Apo B, [Hartgens, F. (2004).] – che è associata a VLDL e LDL effettivamente in circolo – forse attraverso la ↑ secrezione epatica di queste lipoproteine.
Gli estrogeni prodotti dagli androgeni aromatizzanti, in particolare il T, favoriscono i lipidi riducendo l’HTGLA, provocando un flusso netto verso particelle HDL₂ più grandi, migliorando così la dislipidemia e l’aterosclerosi. Ovviamente vi è un “collo di bottiglia” determinato dal dosaggio totale di AAS utilizzati, dal protrarsi di una condizione metabolicamente alterata la quale si verifica in ipercalorica e in condizione di iperestrogenemia protratta.
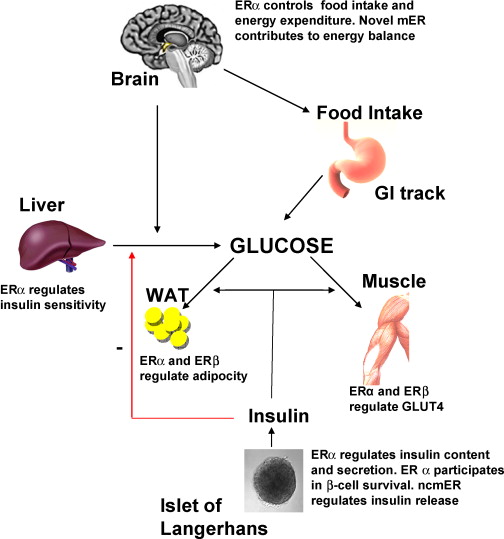
Per fare un altro esempio sulla necessità di mantenere una adeguata attività estrogenica sul lungo termine possiamo osservare l’effetto dell’E2 sul tessuto adiposo e il metabolismo energetico. I ricercatori utilizzano topi maschi knock-out per il recettore degli estrogeni (ERKO) per studiare gli effetti dell’E₂, senza alcuna influenza confondente da parte del Testosterone. I topi maschi ERKO possiedono depositi di tessuto adiposo (AT; tessuto grasso) che sono aumentati del 100% entro 9-12 mesi (approssimativamente la mezza età negli esseri umani). Questo aumento del tessuto adiposo riflette sia l’iperplasia che l’ipertrofia degli adipociti ed è accompagnato da intolleranza al glucosio e insulino-resistenza (IR). I topi maschi ERαKO, quelli con l’ER-α eliminato ma con l’ER-β intatto, hanno mostrato infiammazione ↑AT, dimensioni degli adipociti e alterata tolleranza al glucosio rispetto ai topi maschi normali. Parliamo, ovviamente, di effetti possibili in contesto di deficienze.
E’ interessante notare come il metabolismo del glucosio per kg di muscolo è del 45% più elevato nelle donne e questo è probabilmente mediato da ER-α. [Physiol Rev. 2017 Jul]. Negli uomini, gli effetti metabolici benefici del T sono mediati più dal suo prodotto aromatico (E₂) che dagli androgeni (E₂ > T ↓ di deposito nel AT). Circa il 20% degli estrogeni circolanti nell’uomo deriva dalla sintesi e dalla secrezione testicolare (nelle cellule di Leydig) e il resto dall’attività dell’Aromatasi periferica. [Adv Exp Med Biol. 2017].
In pratica, per gli uomini che utilizzano dosi sovrafisiologiche di androgeni aromatizzabili (ad esempio, T, Nandrolone [Deca, NPP], Boldenone [EQ], Trestolone [MENT], Metandienone [Dianabol]), ciò significa che i siti extra-testicolari dell’Aromatasi, in particolare le cellule adipose, sono prevalentemente responsabili degli effetti associati agli estrogeni (estrogenicità). I bodybuilder che associano gli estrogeni a modifiche sfavorevoli della massa grassa non hanno di per sé torto. Infatti, se da un lato gli estrogeni regolano l’aumento della massa grassa, dall’altro la ridistribuiscono in modo più femminile, in particolare sui glutei e sui fianchi, piuttosto che nella zona addominale, un modello di distribuzione del grasso più maschile… Nuovamente, si tratta di quantità e tempo…
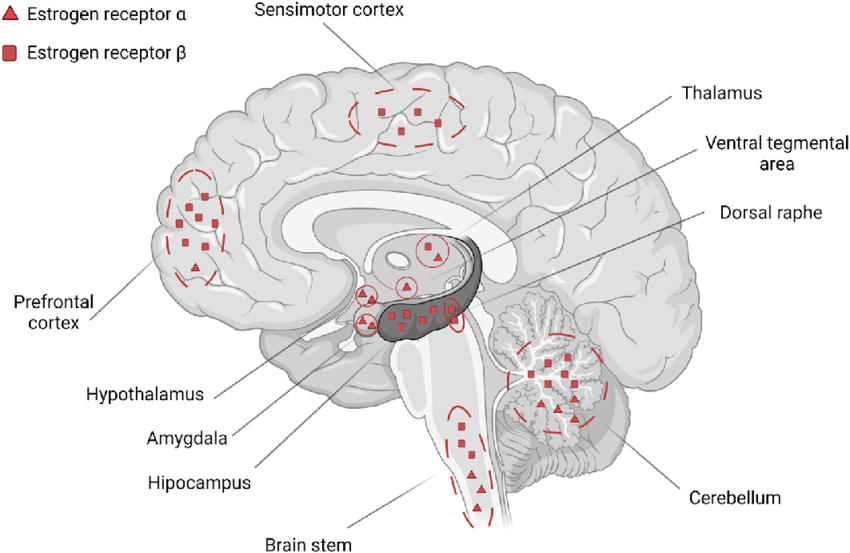
Inoltre, l’ER-α e -β sono espressi nel muscolo scheletrico umano in modo ubiquitario, nelle miofibre, nelle cellule endoteliali e nelle cellule satelliti. L’allenamento di contro-resistenza aumenta ER-α e -β nel muscolo scheletrico umano, suggerendo che la loro espressione è alterata dalle richieste funzionali del muscolo.
Mentre ER-α regola principalmente i lipidi del sangue e il rimodellamento osseo, si ritiene che ER-β medi il rimodellamento nel tessuto muscolare.
L’estradiolo (E₂) è un ligando “potente” per ER-β. Il suo tasso di dissociazione (κd) per questo recettore è di 2,08 nM nell’uomo. [Perkins, M. S., Louw-du Toit, R., and Africander, D. (2017). ]. Il suo tasso di dissociazione (κd) per ER-α, in confronto, è di soli 0,24 nM nell’uomo. [Perkins, M. S., Louw-du Toit, R., and Africander, D. (2017).]. Questo può spiegare gran parte del suo effetto anabolico nel muscolo.
Gli estrogeni sono componenti combinati di regimi steroidei somministrati a bovini maschi castrati per stimolare la crescita muscolare e migliorare la qualità della carne, il che indica che gli estrogeni hanno effetti anabolici sulla massa muscolare maschile. [Veldhuis, J. D., and Bowers, C. Y. (2003).]. Tuttavia, è importante notare le principali differenze tra i bovidi e l’uomo. Nei bovidi, l’E₂ esogeno aumenta in modo dose-dipendente l’IGF-I, ma nell’uomo l’effetto dell’E₂ sull’IGF-I è parabolico, soggetto a una forma a U inversa quando si traccia la dose (cioè la concentrazione ematica) sull’ascissa (asse delle ascisse) rispetto alla risposta (IGF-I) sull’ordinata (asse delle ordinate). Ciò è dovuto al fatto che gli estrogeni (ad esempio, l’E₂) diminuiscono la biodisponibilità dell’IGF-I aumentando la IGFBP-1. [Veldhuis, J. D., and Bowers, C. Y. (2003).]. Un altra volta emerge la necessità di un certo controllo estrogenico.
Nota: La terapia sostitutiva degli estrogeni (ERT; HRT) sembra facilitare la crescita del muscolo scheletrico nelle donne in postmenopausa, probabilmente attraverso effetti sulle cellule satelliti del muscolo. Dopo la menopausa, le donne subiscono un calo cronico dei livelli di estrogeni. Questo fenomeno è associato a una riduzione della sensibilità agli stimoli anabolici (ad esempio, l’allenamento di resistenza) che è reversibile con la ERT. Sebbene la ERT possa effettivamente diminuire la sintesi proteica muscolare al basale, aumenta la crescita dopo l’allenamento contro-resistenza, cioè la risposta anabolica. [Published 2019 Jan 15. ].
Gli estrogeni hanno anche un effetto drammatico sulla funzione muscolo-scheletrica, dato il loro ruolo nello sviluppo, nella maturazione e nell’invecchiamento di ossa, tendini e legamenti.
Gli estrogeni migliorano la massa e la forza muscolare e aumentano il contenuto di collagene nei tessuti connettivi. Tuttavia, a differenza di quanto accade nelle ossa e nei muscoli, dove gli estrogeni migliorano la funzione, nei tendini e nei legamenti gli estrogeni ↓rigidità, quindi ↓ prestazione e potenza e ↑ rischio di lesioni ai legamenti. [Published 2019 Jan 15.]
Mentre questo articolo si concentra sugli estrogeni, un articolo di prossima pubblicazione scritto da questo autore relativo agli AAS e al collagene, alle articolazioni e alle ossa (per-composto) approfondirà gli effetti di particolari androgeni aromatizzabili (ad esempio, Testosterone, Nandrolone, Metandienone) e non aromatizzabili (ad esempio, Stanozololo, Oxandrolone), del rhGH e di altri agenti anabolizzanti sui tessuti connettivi, compresi tendini, legamenti e ossa. Questi effetti sono diversi per classe e per composto. Mentre alcuni AAS e altri agenti anabolizzanti apportano benefici ai tessuti connettivi, altri li danneggiano.
Ricordiamoci che, sebbene gli androgeni abbiano effetti significativi sulle ossa maschili, gli estrogeni sono più importanti per la crescita e il mantenimento delle ossa. [Cauley, J. A. (2015).]. L’E₂ è essenziale per la mineralizzazione, la massa e il ricambio osseo normali negli uomini. [Cauley, J. A. (2015).].
L’aromatizzazione del T ⇒ E₂ è essenziale per gli effetti del T (T + E₂ > T) sull’osso (perdita + BMD). L’E₂ è chiaramente necessaria per la normale crescita e il mantenimento dell’osso negli uomini e l’E₂ media alcuni effetti del T sull’omeostasi ossea . L’azione finale degli estrogeni sullo scheletro è quella di diminuire il rimodellamento e il riassorbimento osseo, mantenendo la formazione ossea. [Cauley, J. A. (2015).].
Gli estrogeni aumentano anche in parte il contenuto di collagene del tendine attraverso un effetto indiretto sull’IGF-I. Gli estrogeni modulano direttamente sia l’IGF-I che le IGFBP (Hansen, et al., 2009b) e quindi possono, tramite l’IGF-I, influenzare il contenuto di collagene attraverso un aumento della sintesi di proteine del collagene tramite la produzione della LARP6 (La-related protein 6) (Blackstone, et al., 2014). LARP6 è una proteina legante aumentata dall’IGF-I, che si lega direttamente all’mRNA del collagene di tipo I e aumenta specificamente la traduzione del collagene di tipo I.
Sia nel tendine che nel legamento, la composizione in peso secco è costituita principalmente da collagene – tra il 60 e l’85% nel tendine e circa il 75% nel legamento. Di questo collagene, la maggior parte è di tipo I: 60% nel tendine e fino all’85% nel legamento.
La rigidità dei legamenti è una buona cosa (✓), in quanto è utile per mantenere la stabilità dell’articolazione e il rischio di lesioni in questi tendini che collegano le ossa alle altre ossa. Al contrario, poiché il tendine collega l’osso rigido al muscolo conforme, un tendine più rigido non è sempre vantaggioso.
La rigidità del tendine è mista: i tendini più rigidi favoriscono le prestazioni, ma aumentano anche il rischio di lesioni. In termini di prestazioni (ad esempio, forza, potenza, sprint), l’aumento della rigidità del tendine trasmette più velocemente le forze muscolari all’osso, aumentando così le prestazioni; tuttavia, questo interesse deve essere bilanciato dal potenziale di concentrazione delle deformazioni all’interno del muscolo, con conseguente rottura. Quando un muscolo collegato a un tendine lasso si contrae, il tendine si allunga in modo flessibile mentre il muscolo si accorcia. Un tendine rigido, invece, non si allunga, ma è costretto ad allungarsi durante la contrazione, causando forze eccentriche. Ciò significa che in un muscolo collegato a un tendine rigido, un carico eccentrico maggiore per un determinato movimento aumenta il rischio di lesioni.
Gli estrogeni diminuiscono la rigidità dei tendini e dei legamenti, riducendo le prestazioni e aumentando il rischio di rottura dei legamenti. Un altra questione di “dose”.
Nel contesto sessuale/riproduttivo, la segnalazione ER-α nell’uomo è di supporto:
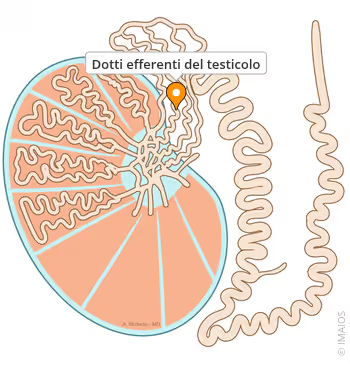
✓ Dotti efferenti e funzioni epididimali.
✓ Trasporto di ioni e riassorbimento di H₂O, necessari per sostenere il normale funzionamento dello sperma (riproduzione maschile).
✖ Concentrazioni di FSH e LH. [Bond, P. Article: Regulation of Testosterone Production. Aug 2021.].
Come l’E₂ nei confronti dell’IGF-I, l’effetto degli estrogeni sulla funzione sessuale è parabolico e soggetto a una forma a U inversa quando si traccia la dose (cioè la concentrazione nel sangue) sull’ascissa (asse delle ascisse) rispetto alla funzione sessuale (per esempio, libido, qualità erettile, fertilità) sull’asse delle ordinate (asse delle ordinate). Questo perché le concentrazioni alle quali l’E₂ sostiene i dotti efferenti, il funzionamento epididimale e la qualità e motilità degli spermatozoi sono basse, mentre quando gli estrogeni sono elevati (come nel caso di dosi sovrafisiologiche di T), le concentrazioni di FSH e LH e l’intera cascata del funzionamento ipotalamo-ipofisario gonadico iniziano a essere regolate negativamente. L’estradiolo è un ormone più fortemente soppressivo del T. [Veldhuis, J. D., and Bowers, C. Y. (2003).].
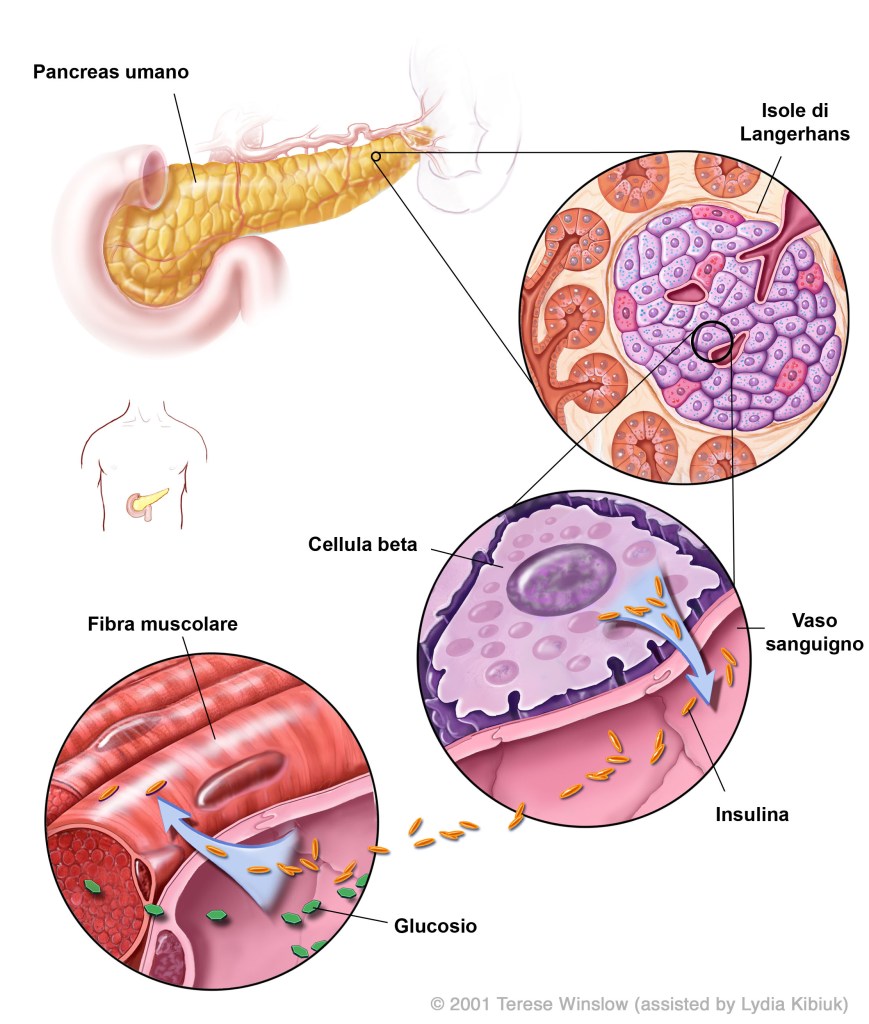
Gli estrogeni hanno effetti positivi sulla funzione delle β-cellule pancreatiche e sull’incidenza del diabete mellito, mediati in parte dagli effetti degli estrogeni sull’apoptosi delle β-cellule, sul contenuto di Insulina delle β-cellule, sull’espressione del gene dell’insulina e sul rilascio di insulina.
Non è assolutamente chiaro se gli estrogeni abbiano un effetto positivo o negativo sul sistema cardiovascolare. Mentre alcuni effetti protettivi del T sono mediati indirettamente attraverso l’E₂, la diminuzione del T sierico è più fortemente associata a rischi più elevati di morte per malattie cardiovascolari negli uomini rispetto alle variazioni dell’E₂ sierico. Al contrario, livelli sierici di E₂ più elevati sono stati riportati in uomini con malattia coronarica e arresto cardiaco improvviso.
A differenza delle femmine, l’E₂ inibisce la guarigione delle ferite nei maschi attraverso l’ER-α.
Risaputo è il fatto che l’E₂ influenza la ritenzione di liquidi agendo sul SNC (regolando la sete e l’assunzione di liquidi e sodio) e sul RAAS (agendo principalmente sull’aldosterone). [Curtis, K. S. (2015).].
Adesso la questione dovrebbe essere più chiara:
L’E2 deve essere mantenuto entro range di concentrazioni e attività/sensibilità ottimali per il soggetto al fine di sfruttare i benefici additivi e diretti di adeguati livelli di E2;
Ogni eccesso di E2, più si allontana dal punto 0 (ottimale) pende verso la comparsa di effetti collaterali maggiori degli eventuali benefici residui;
Il punto chiave primario nella modulazione funzionale dell’E2 per il miglioramento della composizione corporea risulta essere la manipolazione in difetto (abbassamento significativo) per previ periodi e livelli mantenuti regolari secondo sensibilità individuale (mantenimento livelli ottimali) nel lungo termine.
Gestione estrogenica in Bulk/Off-Season:
Abbiamo passato in rassegna i lati positivi e potenzialmente additivi al miglioramento della composizione corporea mantenendo livelli adeguati di E2. Abbiamo altresì visto, che vi è sempre un “collo di bottiglia”, un limite oltre il quale un aumento di E2 causa esclusivamente, o maggiormente, effetti collaterali controproducenti al miglioramento della condizione dell’atleta.
E’ corretto quindi partire esaminando le migliori possibilità di gestione estrogenica in una fase della preparazione annuale dove, solitamente, i bodybuilder, o i loro preparatori, trascurano il fattore estrogenico convinti di poter ottenere vantaggi soprattutto sulla ipertrofia ottenibile.
In questo caso, ovviamente, vanno tenute in considerazione le seguenti variabili soggettive:
Body Fat [variabile genetica-alimentare];
Tasso di aromatizzazione [variabile genetica];
Utilizzo di hCG [variabile iatrogena];
Uso di AAS con attività estrogenica intrinseca (quindi non secondaria alla aromatizzazione) come l’Oxymetholone [variabile iatrogena];
Presenza di un “accenno” di ginecomastia o sua precedente comparsa e successiva regressione [variabile genetica];
Nel caso del primo punto, la questione si risolve generalmente con un break diet/mini Cut della lunghezza dipendente dalla condizione della body fat. Per quanto riguarda, invece, il tasso di aromatizzazione, il quale è connesso all’espressione genica dell’enzima Aromatasi, essa può subire anche adattamenti epigenetici sul lungo termine anche, come accennato nella sezione dedicata alla TRT, con dosi di Testosterone entro il limite massimo fisiologico. L’atleta che presenta questa caratteristica predominante tra i punti sopradetti, e manifesta gli effetti tipici di una iperestrogenemia [ritenzione idrica con massivo aumento pressorio, sviluppo ginecomastico, aumento della massa grassa soprattutto con distribuzione a modello femminile ecc…] dovrà avere molta più accortezza nel dosaggio degli AAS aromatizzabili. Se il soggetto, non presenta un accenno di ginecomastia o non ha presentato tale accenno in passato [fatto regredire farmacologicamente] potrà intervenire con una minima dose efficace di AI [es. 0.5mg di Anastrozolo da 3 a 4 volte a settimana] in misura dipendente dalla dose di AAS aromatizzabili utilizzati e al monitoraggio dei livelli di E2. Se invece il soggetto presenta un accenno di ginecomastia [ovviamente non operabile] o ha presentato tale accenno in passato [fatto regredire farmacologicamente] potrà iniziare con l’uso di un SERM calibrato secondo dosaggio degli AAS aromatizzabili [generalmente da 10 a 30mg di Tamoxifene/die e 60mg/die di Raloxifene]. Ovviamente, anche la presenza di hCG può esacerbare tale situazione. In questo caso le scelte possono essere 1) interruzione d’uso dell’hCG per il tempo di durata della preparazione 2) riduzione del dosaggio e/o dilatazione della tempistica di somministrazione 3) aumentare la dose di AI in funzione dell’alterazione della risposta data. E’ inutile dire che la miglior scelta ricade spesso nel punto 2. Nel caso in cui sia presente un AAS con attività estrogenica intrinseca [quindi indipendente dall’attività della Aromatasi, come l’Oxymetholone] il soggetto, specie se presenta una certa sensibilità recettoriale, dovrebbe preventivamente assumere un SERM al dosaggio minimo efficace [es. 10-20mg di Tamoxifene] al fine di evitare eventuali comparse di ginecomastia legata alla suddetta molecola.
E per quanto riguarda le donne in Off-Season/Bulk?
Ormai sappiamo che la manipolazione della attività estrogenica mostra maggiore impatto sul miglioramento della composizione corporea nelle donne. L’Estradiolo, come estrogeno dominante, attraverso il legame con le subunità recettoriali influenza altre vie ormonali come l’Asse GH/IGF1. Infatti, nonostante la produzione giornaliera di hGH è circa 2 volte superiore nelle giovani donne rispetto agli uomini e varia di 20 volte in base allo sviluppo sessuale e all’età, la sua azione (compresa quella del IGF1) è strettamente influenzata dall’attività estrogenica (in senso inversamente proporzionale per concentrazione epatica di E2). Gli estrogeni inibiscono l’attivazione della via JAK/STAT da parte del hGH. L’inibizione è dose-dipendente e deriva dalla soppressione della fosforilazione di JAK2 indotta dal hGH, con conseguente riduzione dell’attività trascrizionale di STAT3 e STAT5. Gli estrogeni stimolano l’espressione di SOCS-2, che a sua volta inibisce l’azione di JAK2. L’espressione di SOCS-2 è sovra-regolata dagli estrogeni in modo ERα-dipendente. Dato il ruolo centrale di JAK2 nell’attivazione di molteplici cascate di segnalazione del GHR, gli estrogeni possono influenzare anche altre vie a valle per esercitare un impatto più ampio sull’azione del hGH e sulla riduzione del IGF-1 libero per via anche di un aumento delle IGFBP.
Gestione estrogenica in Cut/Pre-Contest:
Quando si parla di Cut e/o Pre-Contest, la riduzione marcata del fattore estrogenico per un breve periodo di tempo può risultare favorevole al miglioramento marcato della condizione.
“Spessore della pelle” e ritenzione idrica:
Quando si tratta di raggiungere la condizione da gara, l’eliminazione del grasso e dell’acqua sottocutanei sono i due obiettivi più importanti, in quanto hanno il più grande effetto complessivo sul aspetto. Questi non sono gli unici fattori, però. Variabili come la pienezza muscolare, la durezza e la densità muscolare, e le striature giocano un ruolo nel determinare il giudizio degli osservatori. Un altro fattore, anche se molto meno frequentemente esposto rispetto ai precedenti, è lo spessore della pelle.
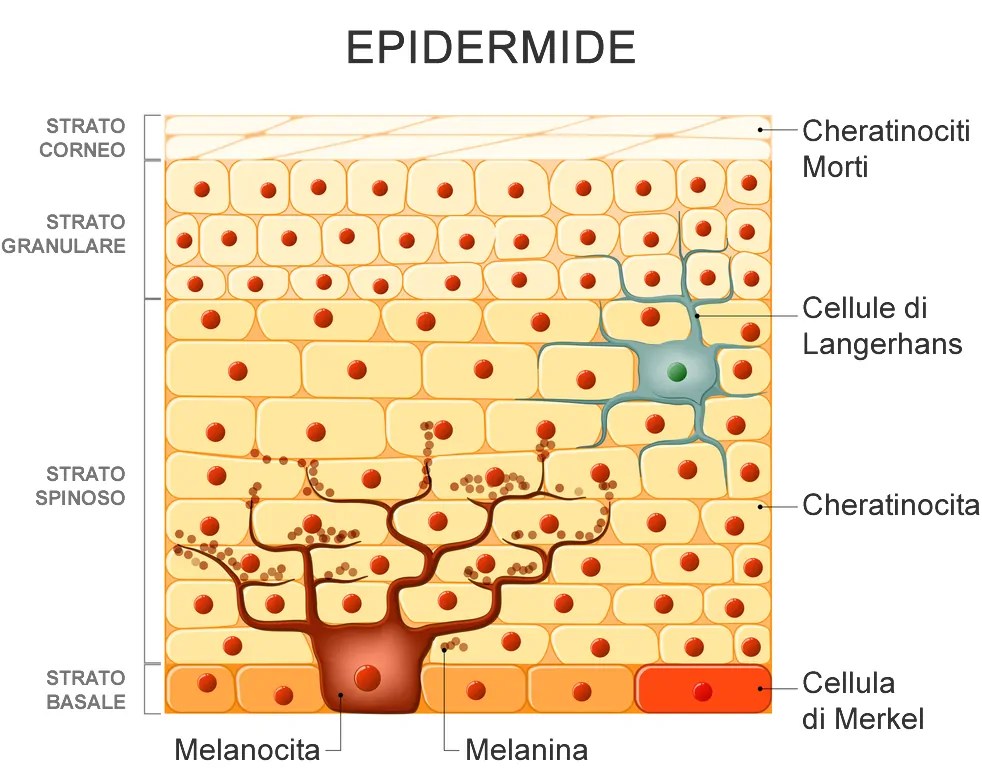
La pelle è composta da tre strati; l’epidermide (strato più esterno), il derma (lo strato centrale), e il tessuto sottocutaneo (lo strato più interno). Anche se lo strato sottocutaneo è tecnicamente considerato parte della pelle, è li dove è conservato il grasso sottocutaneo ed è quindi molto variabile in termini di contenuto totale ed è distinto dagli altri due strati, che sono ciò che noi di solito pensiamo quando sentiamo la parola “pelle”. Il derma è facilmente il più spesso, dal momento che costituisce circa il 90% dello spessore totale della pelle e di conseguenza, è molto più adatto a sfocare la definizione muscolare che l’epidermide. Tuttavia, entrambi gli strati contribuiscono a questo effetto, rendendo necessaria la loro riduzione al minimo essenziale per la visualizzazione massima dei dettagli muscolari. Purtroppo, sia l’rhGH che gli estrogeni possono avere un effetto profondo sullo spessore della pelle, rendendo la loro cattiva gestione potenzialmente controproducente per raggiungere le condizioni ottimali per la gara.
Tralasciando l’effetto del rhGH, che al momento ci interessa “poco”, gli estrogeni sulla sintesi di collagene non sono di certo migliori, dal momento che interessano sia il derma e l’epidermide attraverso percorsi multipli.[https://pubmed.ncbi.nlm.nih.gov/] Nel valutare i suoi effetti sul derma, troviamo che gli estrogeni operano attraverso uno degli stessi meccanismi del hGH sul aumento della sintesi del collagene, ma il modo in cui si compie questo processo è un po’ diverso. In questo caso, gli estrogeni stimolano i fibroblasti dermici (cellule all’interno dello strato del derma che generano il tessuto connettivo), una funzione primaria è produrre collagene. In uno studio, gli estrogeni hanno dimostrato di aumentare la produzione di collagene di tipo I del 76%. Anche se non è così drammatica come con il rhGH, questo è ancora un aumento di tutto rispetto, soprattutto alla luce della capacità degli estrogeni di promuovere la sintesi di acido ialuronico. Direttamente coinvolto nella idratazione cutanea, un aumento dei livelli di acido ialuronico si traduce in un aumento del contenuto di acqua dermica e una successiva espansione del volume della pelle. In uno studio, la somministrazione di estrogeni ha mostrato un aumento della sintesi di acido ialuronico a un pieno 70% nel giro di due settimane.
Gli Estrogeni hanno anche pronunciati effetti sull’epidermide, aumentando lo spessore della pelle attraverso tre meccanismi distinti. Il primo è la stimolazione dell’attività mitotica nei cheratinociti; il tipo cellulare principale trovato nell’epidermide (i cheratinociti costituiscono circa il 90% di tutte le cellule epidermiche). In parole povere, questo significa che l’estrogeno induce la proliferazione dei cheratinociti tramite scissione cellulare, portando a un aumento complessivo del numero di cheratinociti presenti nella pelle. Il secondo è inibendo direttamente l’apoptosi (morte cellulare) dei cheratinociti e l’ottundimento della produzione di chemochine; molecole infiammatorie che possono potenzialmente contribuire alla distruzione della cellula. Infine, gli estrogeni svolgono un ruolo nella idratazione epidermica, volumizzando questo strato di pelle.
Eliminare gli effetti negativi degli estrogeni sullo spessore della pelle è fortunatamente un compito tutto sommato facilmente gestibile, in quanto richiede il mantenimento dei propri livelli di estrogeni entro l’intervallo massimo (come indicato sopra). E’ importante notare che gli effetti negativi degli estrogeni sullo spessore della pelle possono richiedere diversi mesi affinché vangano eliminati completamente, quindi il mantenimento di un elevato livello di estrogeni durante i precedenti mesi di preparazione alla gara per poi farli calare fino al livello minimo solo un paio di settimane prima dell’esibizione non è l’ideale. Per tutti coloro che usano grandi dosi di AAS aromatizzabili, o per i soggetti particolarmente sensibili, per la maggior parte della preparazione, è buona cosa tenere questo concetto in mente.
Basandomi sui dati raccolti sul campo da diversi atleti, e con la conferma anche di esperti del settore come Mike Arnold, la cosa migliore sarebbe optare per il mantenimento di base un livello di estrogeni normale (<60ng/dL con range ideale 30-40ng/dL) per poi calare i livelli a 10-20ng nell’ultimo paio di settimane prima della gara. Oltre a ciò, facendo calare i livelli verso il basso a circa 10ng nelle ultime settimane, si elimina quasi completamente la ritenzione idrica persistente che potrebbe ancora essere un problema, lasciando un livello di estrogeni sufficiente a garantire un ottimale accumulo di glicogeno in concerto additivo con altri PEDs cosomministrati [i dettagli su questo ultimo punto saranno trattati più avanti].
Effetto sui recettori α2-AR e “grasso testardo”:
Nel tessuto adiposo sottocutaneo (sc), gli estrogeni possono aumentare il numero di recettori α2A-AR, che sono coinvolti nell’inibizione della lipolisi. Ciò può contribuire al tipico modello di distribuzione del grasso femminile, in cui il grasso viene immagazzinato maggiormente a livello sottocutaneo piuttosto che viscerale. Inoltre, una condizione estrogenica non ben manipolata può rendere , dato questo effetto, più difficile raggiungere definizioni estremizzate.
E’ stato infatti osservato che l’aumento del numero di recettori α2A-AR causa una risposta lipolitica attenuata dell’epinefrina negli adipociti sc; al contrario, non è stato osservato alcun effetto degli estrogeni sull’espressione dell’mRNA dei recettori α2A-AR negli adipociti del deposito di grasso intra-addominale. Dai risultati ci viene mostrato che gli estrogeni abbassano la risposta lipolitica nel deposito di grasso sc aumentando il numero di recettori α2A-AR, mentre gli estrogeni non sembrano influenzare la lipolisi negli adipociti del deposito di grasso intra-addominale. Questi risultati dimostrano che gli estrogeni attenuano la risposta lipolitica attraverso la sovraregolazione del numero di recettori α2A-AR antilipolitici solo nel sc e non nei depositi di grasso viscerale. Pertanto, questi risultati offrono una spiegazione del modo in cui gli estrogeni, se non ben modulati, rendono molto più ostica la mobilitazione del grasso sc.[https://pubmed.ncbi.nlm.nih.gov/15070958/]
Quindi, per avere un ottimale controllo sulla manipolazione del numero e densità dei α2A-AR al fine di facilitare la mobilitazione e l’uso dei depositi adiposi sc nelle aree particolarmente ostiche, oltre all’uso concomitante di Yohimbina e ACE II inibitori, il mantenimento per circa 8-12 settimane di un range del E2 pari a 20-30pg/dL risulta particolarmente funzionale.
Composizione strategica di AAS/SERM/AI per il maggior controllo estrogenico nel Pre-Contest/Cut:
Un ulteriore dettaglio di design della preparazione alla gara o Cut risiede nel corpo dei PEDs componenti la stessa. Vi sono, infatti, AAS che sono particolarmente funzionali al massimo controllo della attività estrogenica e che trovano il loro migliore inserimento proprio in quei momenti della preparazione dove la riduzione marcata della componente estrogenica è essenziale per il “tocco finale” per il palco, lo shooting fotografico o per un semplice obbiettivo personale da amatore.
Boldenone Undecylenato:
Molecola di Boldenone
Il Boldenone [1,4-androstadiene-3-one,17b-ol] è uno steroide anabolizzante-androgeno spesso legato all’estere Undecylenato. Strutturalmente molto simile al Testosterone, il Boldenone differisce da questo per il doppio legame tra C1 e C2. Tale modifica rende la molecola un substrato molto meno affine all’enzima 5-α reduttasi rispetto al Testosterone, ed era teoria comune che riducesse anche il tasso di aromatizzazione della molecola.
Ed è per questo ultimo punto che molti ritengono che il Boldenone possa essere utilizzato come base “Mix” con il Testosterone o in sostituzione ad esso per coloro che sono inclini agli effetti collaterali estrogenici. E, in teoria, utilizzando il Boldenone, si potrebbe ridurre il rischio di sviluppare effetti collaterali correlati ad un livello elevato di estrogeni poiché dovrebbe aromatizzare circa la metà del Testosterone. Vista la riduzione marcata del E2 si era ipotizzato che uno (o più) dei suoi metaboliti agisca come un Inibitore dell’Aromatasi (AI). Secondo questa ipotesi, i metaboliti del Boldenone sono in realtà la causa del ridotto impatto dell’Enzima Aromatasi su questa molecola e su altri substrati soggetti come il Testosterone. Ma, i limiti di verifica hanno reso questa ipotesi piuttosto traballante; i livelli sierici di Estradiolo sono stati spesso determinati utilizzando kit di test immunologico per elettrochemiluminescenza (ECLIA). Parliamo di uno dei peggiori metodi di test ematico dal momento che è soggetto a influenze ormonali non ricercate o a limitazione di precisione della conta ormonale.
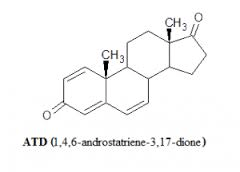
In letteratura, passando al vaglio i vari metaboliti del Boldenone, il famoso “AI” del mercato grigio ATD non era elencato nello studio sui metaboliti umani condotto sul Boldenone [https://pubmed.ncbi.nlm.nih.gov/]. Proprio come con l’ADD, l’ATD ha dimostrato di essere sia un metabolita del Boldenone che di metabolizzare in Boldenone [https://pubmed.ncbi.nlm.nih.gov/]. In uno studio in vitro è stato dimostrato che l’ATD riduce significativamente la biosintesi degli estrogeni.[https://pubmed.ncbi.nlm.nih.gov/] Il problema, però, è che l’ATD non risulta essere un metabolita significativamente espresso nell’uomo e, nel pratico, la sua potenzialità AI è spesso stata deludente o ad un tasso blando. La risultante della ricerca, quindi, non solo non chiarisce se il Boldenone agisca come un AI ma anche se esso aromatizza in Estradiolo ad un dato tasso.
Rappresentazione dei passaggi componenti il test immunologico per elettrochemiluminescenza (ECLIA).
Le cose diventano più chiare quando i campioni ematici vengono analizzati con test LC/MS-MS ultra sensibile e non l’ECLIA. Analizzando i campioni di un utilizzatore sottoposto ad unciclo comprendente circa 850mg di Boldenone Undecylenato e 250mg di Testosterone Enantato a settimana, i risultati mostravano un significativo aumento dei livelli di Estrone, con un risultato di 662pg/mL, con il limite massimo dell’intervallo di riferimento pari a 65pg/mL. Il livello di Estradiolo non era rilevabile con meno di 2,5pg/mL. Sappiamo, inoltre, che il Testosterone utilizzato dal soggetto in questione era realmente Testosterone (1431 ng/dL), poiché, per l’appunto, i campioni ematici sono stati sottoposti ad un test specifico LC/MS-MS, che è il gold standard per verificare il totale esatto del Testosterone evitando il rilevamento incrociato di altri anabolizzanti. La quantità di Testosterone rilevata, normalmente, porterebbe ad un livello di Estradiolo medio-alto, non di certo così basso come è risultato. Fortunatamente, con il risultato del esame del sangue comprendente l’Estrone [E1], possiamo finalmente risolvere l’enigma su ciò che con molta probabilità accede realmente.
E’ stato osservato come anche con uno schema di dosaggio quasi identico tra Testosterone e Boldenone (vedi esempio 300mg di Boldenone e 400mg di Testosterone) i livelli di Estradiolo risultano generalmente bassi.
Quindi, a questo punto, si potrebbe ipotizzare che il Boldenone non aromatizza effettivamente in Estradiolo, né inibisca l’enzima Aromatasi ma, piuttosto, esso potrebbe competere con il Testosterone nell’interazione con l’Aromatasi dando come prodotto aromatico l’E1.
Il Boldenone sembra avere una maggiore tendenza alla conversione in Estrone e una forza di legame all’enzima Aromatasi dose-dipendente
L’E1 è un estrogeno molto meno potente dell’estradiolo e, in quanto tale, è un estrogeno relativamente debole.[Kuhl H (August 2005).] Secondo uno studio, le affinità di legame relative dell’estrone per l’ERα e l’ERβ umani erano rispettivamente il 4,0% e il 3,5% di quelle dell’Estradiolo. Secondo uno studio, l’affinità di legame relativa dell’E1 per l’ERα e l’ERβ umani è pari al 4,0% e al 3,5% di quella dell’E2, rispettivamente, e la capacità transattiva relativa dell’E1 per l’ERα e l’ERβ è pari al 2,6% e al 4,3% di quella dell’E2, rispettivamente.[Escande A, Pillon A, Servant N, Cravedi JP, Larrea F, Muhn P, et al. (May 2006).] In accordo, l’attività estrogenica dell’estrone è stata riportata a circa il 4% di quella dell’Estradiolo.
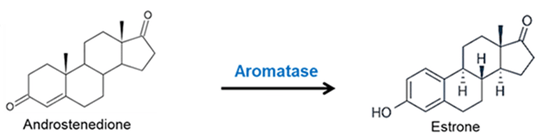
La via principale attraverso la quale l’Estrone viene biosintetizzato coinvolge l’Androstenedione come intermedio, con quest’ultimo che viene convertito in Estrone dall’enzima Aromatasi. Questo è il punto chiave da ricordare nel contesto di questa anamnesi di design dei componenti PEDs della Preparazione alla gara o Cut.
In definitiva, il Boldenone, con tutta probabilità, ha una funzione di “ormone esca” per l’enzima Aromatasi. Sappiamo però che, probabilmente, la sua conversione in estrogeno lo vede convertirsi prevalentemente in Estrone [E1] e non in Estradiolo [E2]. Sappiamo che l’Estrone può convertirsi in Estradiolo (e viceversa) ma che il tasso in cui ciò avviene è molto basso. Siamo a conoscenza del fatto che l’E1 è un estrogeno molto meno potente dell’E2 e, come tale, è un estrogeno relativamente debole.[Kuhl H (August 2005), Escande A et al. (May 2006), Ruggiero RJ, Likis FE (2002)]. Ciò, darà come risultante, una attività estrogenica tissutale [genomica e non genomica] nettamente ridotta anche con un dosaggio di Boldenone non particolarmente elevato [es. 300-500mg/week].
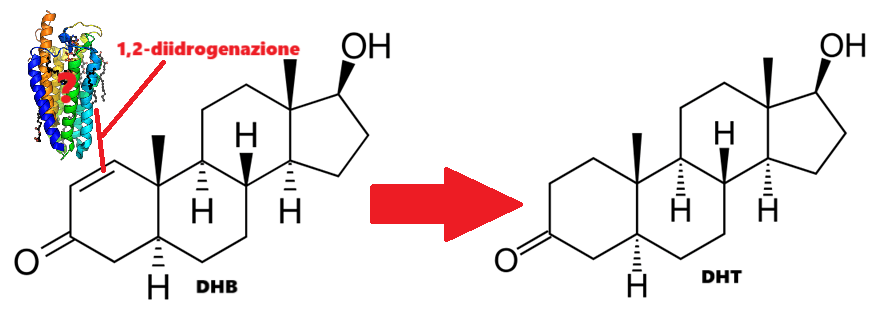
DHB:
Molecola di DHB
L’1-Testosterone (maggiormente noto come Dihydroboldenone, abbreviato in DHB), è uno steroide anabolizzante-androgeno sintetico (AAS) e un derivato 5α-ridotto del Boldenone (Δ1-testosterone). Si differenzia dal prodotto 5α-ridotto del Testosterone, il DHT, per la presenza di un doppio legame C1-C2 nell’anello A dello scheletro carbossilico.[William Llewellyn (2009).] Il DHB possiede una potente affinità di legame con il AR ma non presenta particolare selettività tissutale e ha un’elevata capacità di stimolare la transattivazione AR-dipendente. In vivo, a differenza del Testosterone Propionato, il DHB aumenta anche il peso del fegato con possibile disfunzionalità d’organo [dose e tempo dipendente].[Friedel A et al. (August 2006).]
Il DHB è strutturalmente simile al Methenolone (Primobolan/Rimobolan) e allo Stenbolone. In quanto 1-ene, il DHB è un AAS le cui caratteristiche anabolizzanti predominano su quelle androgene (mascolinizzanti, calvizie, ecc.).
Differenze e similitudini molecolari tra Methenolone e DHB
È interessante notare che il DHB metabolizza principalmente a DHT (5α-diidrotestosterone; DHT), pur non essendo soggetto a nuove interazione con la 5α-reduttasi, ed è particolarmente epatotossico – cosa insolita per un AAS iniettabile – sia direttamente, con prove dell’aumento delle dimensioni del fegato (visto prevalentemente negli animali), sia indirettamente, dato che i produttori e i chimici usano tipicamente solventi industriali come il guaiacolo, una sostanza chimica pericolosa e probabilmente cancerogena, per mantenere il DHB in soluzione.
Struttura Proteine C-reattiva
La particolare intollerabilità e i controversi effetti anabolizzanti del DHB, considerato da una fazione di fedelissimi come un “Tren lite” e da un’altra fazione di detrattori che lo considerano fondamentalmente un Methenolone meno selettivo [della stessa classe di 1-eni] ma con dolore post-iniezione (PIP) e innalzamento della Proteina C-reattiva (CRP), lo classificano tra gli AAS meno utili per i bodybuilder. Eppure, c’è una caratteristica che, a “piccole dosi” e “strategicamente” può rendere il DHB una molecola opzionale da inserire nella preparazione per il controllo estrogenico.
Per introdurre questa potenzialità del DHB, vorrei sottolineare quanto accennato in precedenza sul principale metabolita di questo AAS: il DHT. Il DHB metabolizza principalmente in DHT, pur non essendo suscettibile alla attività della 5α-reduttasi [Fragkaki et al. C. (2009).]. Come ben sappiamo, specialmente in passato, il DHT veniva utilizzato per trattare le pazienti con cancro al seno estrogeno dipendente. L’aumento di questa molecola in risposta ai processi metabolici ai quali è soggetto il DHB causa un controllo estrogenico a livello dell’attività tissutale.
Esistono solo spiegazioni imperfette per la sintesi di DHT come metabolita del DHB. Peter Bond ha comunicato via e-mail di essere rimasto sorpreso dall’identificazione da parte di Fragkaki del DHT come metabolita primario dell’1-testosterone, poiché suggerisce che qualche enzima finora non identificato è responsabile della 1,2-diidrogenazione di questo androst-1-ene-3-one. [Bond, Peter 2 April 2022].
Processo ipotetico dato dall’enzima “X” che interagendo con il DHB avvia la 1,2-diidrogenazione dando come metabolita principale il DHT.
Ma l’attività propria del DHB sulla componente estrogenica dipende da una capacità di aumentare l’E1.
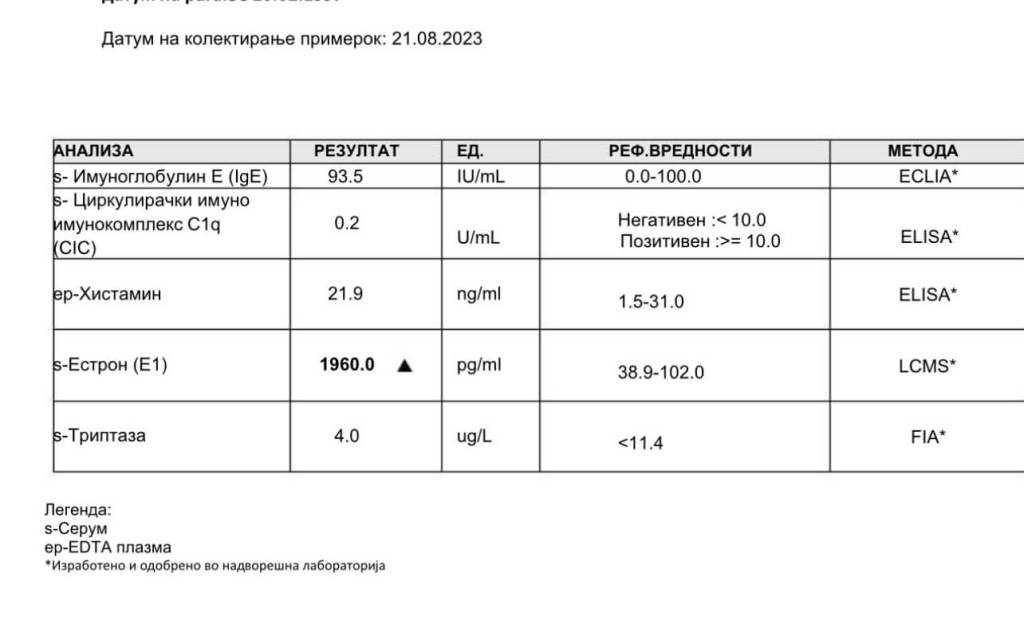
Come si può vedere dalla sopraesposta griglia, la LCMS sensibile per l’Estrone (E1) mostra un valore notevole di 1.960,0 pg/mL per l’E1 in un individuo che utilizza dosi fino a 700mg di DHB i.m. q.w. con una costante di 100mg di Testosterone i.m. q.w. Tali alterazioni sono state viste anche con dosi di DHB inferiori [es. 200-400mg/week] anche se l’impatto è ovviamente a grado dose-dipendente.
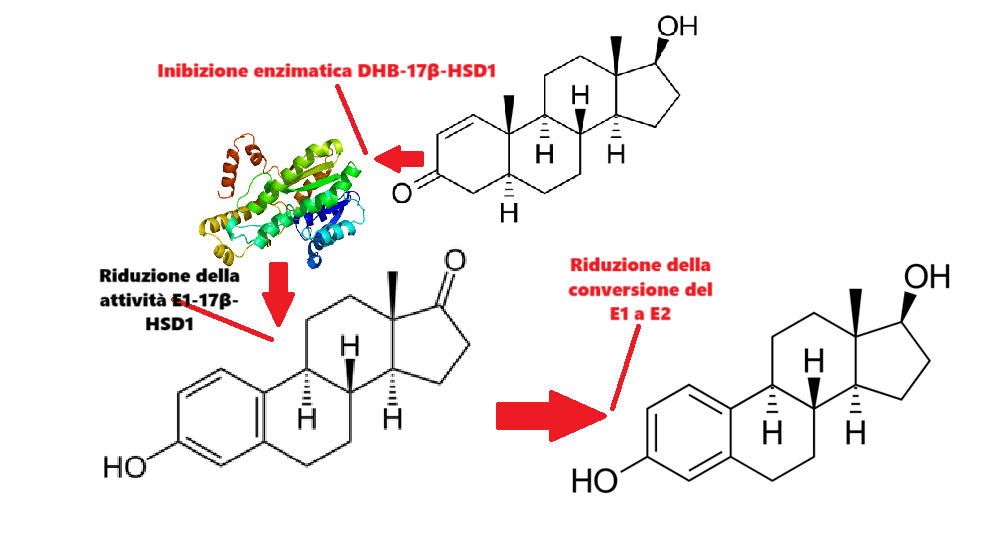
L’ipotesi è che il DHB possa agire come inibitore della 17β-HSD1, aumentando così il DHT e l’E1 e diminuendo l’E2. L’E2 stimola, mentre il DHT inibisce la crescita del cancro al seno e lo sviluppo della ginecomastia (modulando l’attività tissutale e sistemica del E2).
Struttura della 17β-HSD1
La 17β-idrossisteroide deidrogenasi 1 (17β-HSD1) è un enzima che nell’uomo è codificato dal gene HSD17B1.[Luu-The V et al. Feb 1990] Questo enzima ossida o riduce il gruppo C17 idrossi/cheto di androgeni ed estrogeni ed è quindi in grado di regolare la potenza di questi steroidi sessuali.
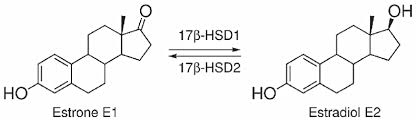
Questo enzima è responsabile dell’interconversione di Estrone (E1) ad Estradiolo (E2) e dell’interconversione di Androstenedione a Testosterone:
L’isozima 17β-HSD1 umano è altamente specifico per gli estrogeni rispetto agli androgeni, mentre l’isozima dei roditori è meno specifico.[Saloniemi T et al. 2012]
Il DHT, strutturalmente simile al DHB, è un noto, anche se debole, inibitore della 17β-HSD1. Affinché gli androgeni DHT o Testosterone (T) diventino un buon ligando per questo enzima, devono imitare più da vicino la forma planare dell’anello A dell’E2 e fornire un ampio nucleo idrofobico. [He, W., Gauri, M., Li, T., Wang, R., & Lin, S.-X. (2016).]. Il DHB soddisfa entrambi i requisiti, differenziandosi dal DHT per un doppio legame C-1- C-2, che gli conferisce un anello A più simile a quello degli estrogeni, e dal Testosterone per la sua ampia spina dorsale idrofobica di 5α-androstene. Questi 5α-androsteni hanno un’affinità per l’AR 173 volte maggiore rispetto alla loro controparte orientata verso il β; ad esempio, il DHT ha una maggiore planarità ed è meno ingombrante rispetto al T per inserirsi nella cavità idrofobica del sito di legame sull’AR. [Liao, S., Hung, S. C., Tymoczko, J. L., & Liang, T. (1976)].
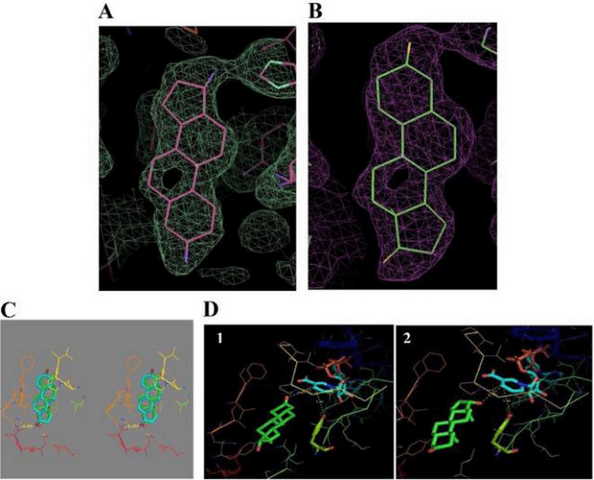
Legame 17β-HSD1 da parte del DHT: (A) – (E) Cristallizzazione 3D del complesso DHT/17β-HSD1 e dati di docking.
L’enzima 17β-HSD1 è altamente espresso nel tessuto adiposo e nella prostata degli uomini. La sua funzione è quella di catalizzare la riduzione dell’estrogeno debole E1 all’estrogeno forte E2 e la sua espressione è correlata positivamente all’attivazione dell’E1 e ai livelli di E2. L’inibizione della 17β-HSD1 riduce l’E2, anche se l’abolizione (a 0,00pg/mL) richiederebbe l’inibizione anche dell’Aromatasi e della Solfatasi [Thomas MP et al. J Steroid Biochem Mol Biol. 2013].
La 17β-HSD1 catalizza l’ultima fase della biosintesi degli estrogeni attivi (E2 e 5-androstene-3β,17β-diolo), mentre l’E1 è il suo substrato primario, l’enzima ha una certa attività per substrati non estrogenici. Il DHT è un substrato non riconosciuto dalla 17β-HSD1 e si lega in modo simile all’E2, con un’orientazione simile, ma con alcune differenze per accogliere un’ulteriore massa (per esempio, il gruppo metilico C-19). Il complesso (17β-HSD1-DHT) ha siti di legame con gli steroidi ben definiti e il valore Km noto per il DHT è 17 volte superiore a quello dell’E2 (quindi ci vuole più DHT per saturare l’enzima) e l’attività specifica 26 volte inferiore. [Lin, S. X., et al. 1999].
Azione schematizzata del DHB come inibitore della 17β-HSD1
In conclusione, il DHB, se inibisce la 17β-HSD1, lo fa in modo contenuto – certamente non con una specificità sufficiente per essere commercializzato per uso clinico nel trattamento del cancro al seno. Sebbene la letteratura citata suggerisca che gli androgeni C19 non sono inibitori della 17β-HSD1 altamente potenti e selettivi – in modo da consentire un buon legame e un’elevata velocità di reazione – gli androgeni sono ligandi attivi per l’enzima. Per massimizzare il legame, un inibitore progettato per essere specifico per la 17β-HSD1 deve imitare la forma planare dell’anello A dell’E2 e mantenere un ampio nucleo idrofobico che possa interagire con il sito di legame steroideo dell’enzima. [Lin, S. X., et al. 1999]. L’anello A del DHB, più simile a quello degli estrogeni, e l’ampio backbone idrofobico del 5α-androstene soddisfano questi requisiti, rendendolo un androgeno C19 teoricamente più potente del DHT nell’inibire la 17β-HSD1.
Anastrozolo e Exemestane:
Dell’Anastrozolo abbiamo già parlato nel paragrafo in riferimento alla gestione estrogenica durante una TRT. Abbiamo ribadito che in condizione di attività testicolare mantenuta per via d’uso dell’hCG, l’Anastrozolo darà una soppressione del E2 al dosaggi tra 0,5 e 1mg/die di circa il 50% . Data questa variabile, generalmente si interrompe la somministrazione di hCG 21-14 giorni prima del contest in concomitanza con la sospensione d’uso a 14 giorni dal contest del Testosterone. L’ambiente creato, in calo di substrati aromatizzabili a E2 e stimolanti l’espressione del Aromatasi [vedi hCG], il potenziale dell’Anastrozolo non solo aumenta in % di rapporto con i substrati con cui competere ma anche in termini di efficacia per concentrazione.
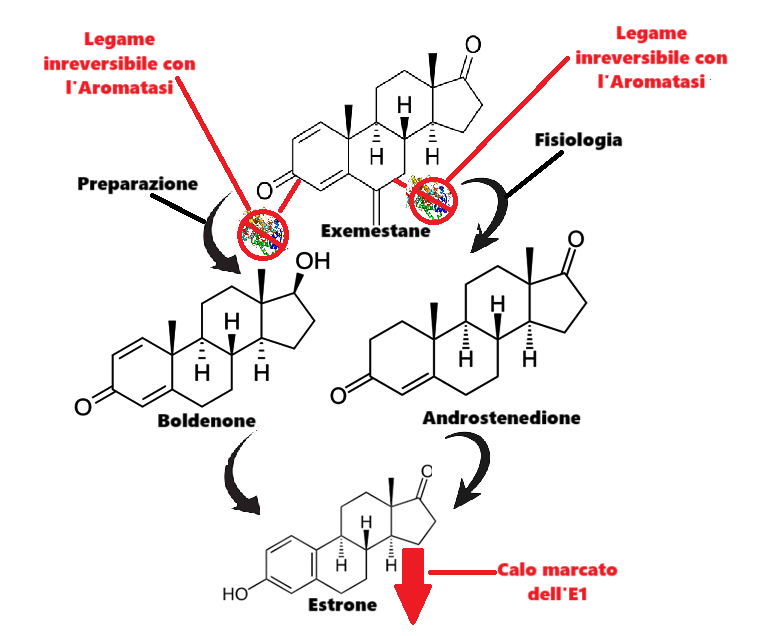
Come abbiamo visto in precedenza, il Boldenone sembra essere particolarmente soggetto alla conversione nel debole E1. Sebbene questo estrogeno sia di debole potenza biologica, con dosi di 500mg> di Boldenone potrebbe giocare un ruolo di “fino” la sua riduzione. Sicuramente starete pensando che basta l’Anastrozolo anche per questo scopo, e non è del tutto sbagliato come ragionamento. Però, e c’è un però, esiste un AI che potrebbe lavorare in sinergia con l’Anastrozolo nelle ultime settimane pre-contest e “occuparsi” maggiormente della componente E1. Sto parlando del Exemestane, comunemente conosciuto come Aromasin.
Molecola di Exemestane
L’Exemestane è un inibitore orale steroideo dell’Aromatasi. L’Exemestane è un inattivatore irreversibile steroideo di tipo I dell’Aromatasi, strutturalmente correlato al substrato naturale 4-androstenedione. Agisce come falso substrato per l’enzima Aromatasi e viene trasformato in un intermedio che si lega irreversibilmente al sito attivo dell’enzima causandone l’inattivazione, un effetto noto anche come “inibizione suicida”. Essendo strutturalmente simile ai bersagli enzimatici, l’Exemestane si lega in modo permanente agli enzimi, impedendo loro di convertire gli androgeni in estrogeni.[Jasek, W, ed. (2007)]
Uno studio condotto su giovani maschi adulti ha rilevato che il tasso di soppressione degli estrogeni per l’Exemestane variava dal 35% per l’E2 al 70% per l’E1.[Mauras N et al. December 2003] Ed è proprio questa la caratteristica che rende sensato l’uso del Exemestane in combinazione, e in questa fase, con l’Anastrozolo.
Ma perchè questa caratteristica?
L’Exemestane è noto chimicamente come 6-metilideneandrosta-1,4-diene-3,17-dione. Come gli inibitori dell’Aromatasi Formestano e Atamestano, l’Exemestane è uno steroide strutturalmente simile al 4-androstenedione, il substrato naturale dell’Aromatasi. Si distingue dalla molecola A4 solo per il gruppo metilidenico in posizione 6 e per un ulteriore doppio legame in posizione C1-2. Si è quindi ipotizzato che la sua similitudine al A4 sia una possibile risposta alla maggiore soppressione del E1 rispetto al E2.
Molecola di Androstenedione
L’Androstenedione, o 4-androstenedione (abbreviato in A4 o Δ4-dione), noto anche come androst-4-ene-3,17-dione, è un ormone steroide endogeno debole e intermedio nella biosintesi dell’Estrone e del Testosterone a partire dal DHEA. È strettamente correlato all’Androstenediolo (androst-5-ene-3β,17β-diolo). Esso è un substrato diretto per l’E1 che, a sua volta, può essere convertito in E2 per via della attività della 17β-HSD1 [vedi sezione dedicata al DHB]; tuttavia il tasso di conversione è generalmente basso.
Questa similitudine nella struttura steroidea sembra indicare una ipotetica competizione maggiore e migliore con il 4A piuttosto che con il Testosterone, con la risultante della maggiore soppressione del E1 rispetto all’E2.
Oltre alla similitudine conformazionale con il 4A, l’Exemestane condivide una similitudine tipica degli “1-ene” e del Boldenone: il doppio legame in C-1-C-2.
Unendo i due “punti di similitudine molecolare” possiamo ipotizzare una maggior competizione di legame specifica con il Boldenone e una ulteriore e addizionale diminuzione del livello estrogenico assoluto anche a carico del E1. I test fatti per verificare la teoria sono stati soddisfacenti, e il “tocco dry” raggiunto dai soggetti sperimentali era generalmente maggiore rispetto a coloro che non hanno applicato questa strategia.
Competizione schematizzata del Exemestane per il legame con l’enzima Aromatasi nei confronti dei due substrati “similari”, Boldenone e Androstenedione.
Un possibile approccio con questo “AI mix” prevede l’uso alternato di 0,5mg di Anastrozolo e 12,5-25mg di Exemestane per le ultime due settimane di preparazione alla gara.
Un altra ipotesi che si unisce a quella sopra esposta, vede una attività inibitoria maggiore sulla Solfatasi steroidea da parte del Exemestane e una conseguente riduzione dell’Estrone attivo.
La proteina codificata da questo gene catalizza la conversione dei precursori steroidei solfatati in steroidi liberi. Ciò include il DHEA solfato, l’Estrone solfato, il Pregnenolone solfato e il Colesterolo solfato, tutti nelle loro forme non coniugate (DHEA, Estrone, pregnenolone e colesterolo, rispettivamente).[4][5] La proteina codificata si trova nel reticolo endoplasmatico, dove è presente come omodimero.
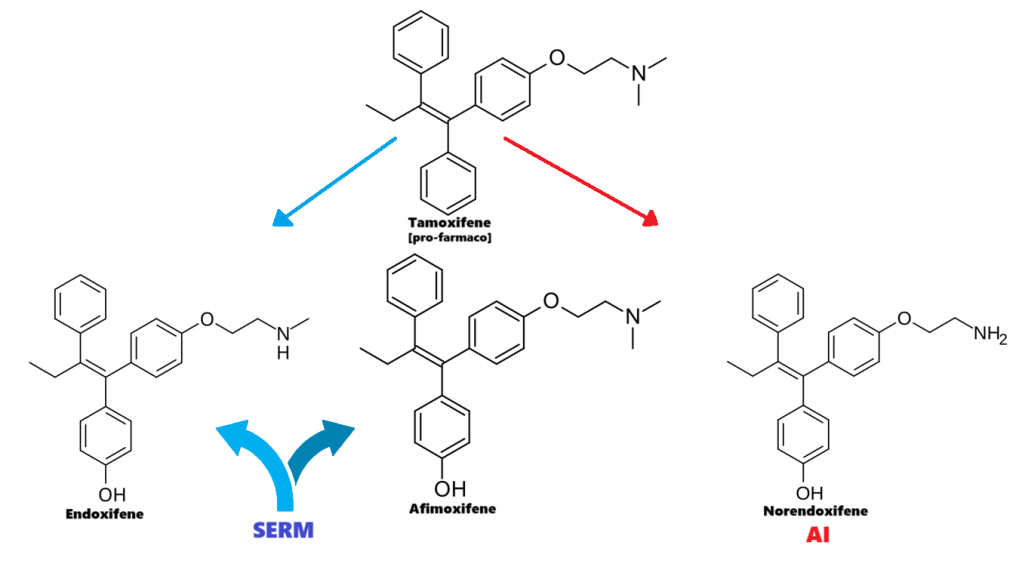
Tamoxifene:
Molecola di Tamoxifene
Il Tamoxifene è un pro-farmaco i cui metaboliti agiscono come modulatori selettivi dei recettori degli estrogeni (SERM), ovvero come agonista parziale dei recettori degli estrogeni (ER), e come Inibitori della Aromatasi (AI). Ha un’attività mista estrogenica e antiestrogenica, con un profilo di effetti diverso a seconda dei tessuti. Ad esempio, il prodotto metabolico del Tamoxifene ha effetti prevalentemente antiestrogenici nel seno, ma prevalentemente estrogenici nell’utero e nel fegato. Nel tessuto mammario, agisce come antagonista ER, inibendo la trascrizione dei geni che rispondono agli estrogeni.[Wang DYet al. (February 2004)] Un effetto collaterale benefico del Tamoxifene è che previene la perdita di massa ossea agendo come agonista ER (cioè imitando gli effetti degli estrogeni) in questo tipo di cellule. Pertanto, inibendo gli osteoclasti, previene l’osteoporosi.
Come accennato, il Tamoxifene agisce come pro-farmaco di metaboliti attivi come l’Endoxifene (4-idrossi-N-desmetiltamoxifene) e l’Afimoxifene (4-idrossitammoxifene; 4-OHT).[Klein DJ et al. (November 2013).] Questi metaboliti hanno un’affinità per gli ER da 30 a 100 volte superiore a quella del Tamoxifene stesso.[Ahmad A et al. (December 2010).] L’Afimoxifene ha mostrato di avere il 178% e il 338% dell’affinità dell’estradiolo per l’ERα e l’ERβ, rispettivamente. [Kuhl H (August 2005).] Le potenze antiestrogeniche dell’Endoxifene e dell’Afimoxifene sono molto simili. Tuttavia, l’Endoxifene è presente in concentrazioni molto più elevate rispetto all’Afimoxifene ed è ora ritenuto la principale forma attiva del Tamoxifene nell’organismo. Il Norendoxifene (4-idrossi-N,N-didesmetiltamoxifene), un altro metabolita attivo del Tamoxifene, è risultato agire come un potente inibitore competitivo dell’Aromatasi (IC50 = 90 nM) ed è con tutta probabilità coinvolto nell’attività antiestrogenica complessiva del Tamoxifene.
Metaboliti epatici principali del Tamoxifene
Il suo inserimento è “preliminare”, ovvero avviene fin dal principio della preparazione ad un dosaggio contenuto e comunemente assestato sui 10-20mg/die. La sua azione è in funzione “preparatoria” al tocco finale delle fasi conclusive della preparazione al contest. Come già detto, inserendolo con tempistiche adeguate il Tamoxifene contribuisce alla riduzione dei α2A-AR adipocitari facilitando la mobilitazione e l’uso (dipendente da dieta e fattori termogenici) del “grasso testardo” in combinazione potenziale con Yohimbina e ACE II inibitori.
Un altra cosa particolarmente interessante riguardante il Tamoxifene, o, meglio i suoi metaboliti attivi, è che può influenzare positivamente il metabolismo del glucosio, in particolare nel muscolo scheletrico. Alcuni studi suggeriscono che il Tamoxifene può avere un impatto sulla tolleranza al glucosio e sulla sensibilità all’insulina, con alcune evidenze che indicano un aumento della resistenza all’insulina in determinati contesti, soprattutto in presenza di obesità. Tuttavia, altre ricerche indicano che il Tamoxifene può migliorare la tolleranza al glucosio, evidenziando la complessità dei suoi effetti sul metabolismo del glucosio. In pratica, l’Afimoxifene agisce come un agonista del ER nel muscolo-scheletrico sopperendo al drastico calo del E2 nella fase Cut/Pre-Contest mantenendo i vantaggi dell’estrogeno la dove maggiormente servono all’atleta. Inoltre, alcuni studi su animali hanno dimostrato che il Tamoxifene può proteggere dai danni muscolari causati dalle contrazioni migliorando il recupero post esercizio, migliorare la forza muscolare e persino migliorare la struttura muscolare.
Ricordiamoci che la maggior parte degli effetti dati dai metaboliti del Tamoxifene deriva dal loro legame ad alta affinità con ERα ed Erβ, che sono espressi nei tessuti estrogeno-responsivi di maschi e femmine, compreso il muscolo scheletrico. Esistono diverse isoforme di ERβ. L’ERβ1 è considerata l’isoforma fisiologicamente attiva, mentre l’ERβ2, un’isoforma più lunga con un’affinità molto ridotta per gli estrogeni, agisce in modo dominante negativo per gli altri ER. L’espressione di ERβ è stata riscontrata nelle fibre muscolari e nei capillari dell’uomo. I principali effetti fisiologici degli estrogeni nei muscoli scheletrici sono, come abbiamo già detto, coadiuvanti l’aumento della forza e il recupero muscolare.
La presenza dei recettori ERβ nelle fibre muscolari e nei capillari è cruciale nella risposta fisiologica all’attività trascrizionale ER-mediata e ai conseguenti effetti biologici. Si ritiene che un meccanismo d’azione sia costituito, nel muscolo, da un potenziamento della miogenesi e dell’angiogenesi mediata dal fattore di crescita endoteliale vascolare (VEGF). Questi effetti ER-mediati possono favorire la riparazione e l’adattamento del tessuto muscolare dopo l’allenamento (Wiik et al., 2005). I recettori degli estrogeni regolano la trascrizione genica attraverso i classici elementi di risposta agli estrogeni (ERE) e i geni AP-1 responsivi. Il Tamoxifene agisce come antagonista ER sugli ERE ma come agonista ERβ/agonista parziale sui geni responsivi della proteina attivatrice 1 (AP-1), responsabili di diverse funzioni nella crescita e nella proliferazione cellulare (Jain e Koh 2010). In condizione di ipoestrogenemia, la componente di parzialità agonista diventa predominante. Infatti, clinicamente, gli agonisti parziali possono essere utilizzati per attivare i recettori in modo da dare una risposta submassimale desiderata quando sono presenti quantità inadeguate del ligando endogeno, oppure possono ridurre la sovrastimolazione dei recettori quando sono presenti quantità eccessive del ligando endogeno.[Zhu BT (April 2005).]
Relazione tra attività percentuale e concentrazione di agonisti completi e parziali
* In caso nella preparazione siano presenti AAS sintetizzati e usati in clinica per il trattamento del cancro al seno estrogeno dipendente, quali Drostanolone e/o Methenolone, l’uso di SERM e AI andrebbe con ulteriore attenzione calibrato per ovvie attività riduttive sulla funzione estrogenica connesse a queste due molecole. Nel caso in cui venga utilizzato il Methylepitiostanolo, il quale è la forma metilata di un AAS utilizzato in medicina per il trattamento del cancro al seno (Epitiostanolo), e che possiede una marcata attività AI, l’uso di Anasttrozolo risulta superfluo mentre (per i motivi sopra citati) l’uso di Exemestane rimane funzionale.
Punti importanti:
Se un soggetto non presenta particolare sensibilità all’attività dell’E2, l’uso del Boldenone mixato al Testosterone [rapporto da tarare sul soggetto] e l’aggiunta terminale di solo Anastrozolo EOD sono di gran lunga sufficienti;
Eliminare i substrati altamente aromatizzabili nelle ultime due settimane pre-contest, nel caso sopra detto, possono limitarsi alla sola omissione d’uso del hCG;
E’ consigliabile eseguire monitoraggi con esami ematici per E2 e E1 durante il percorso preparatorio onde poter valutare aggiustamenti dello stesso;
Gli effetti collaterali connessi ad un marcato abbassamento estrogenico comprendono peggioramento del profilo lipidico, affaticamento, umore basso, sbalzi di umore, letargia, dolori articolari e diminuzione della funzione sessuale;
Questo tipo di pratiche NON SONO PER PRINCIPIANTI O INTERMEDI MA PER ATLETI ESPERTI E DI ALTO LIVELLO!
Nulla di ciò che è stato scritto rappresenta una indicazione o un incitamento all’uso di farmaci dopanti e illegali!
E per quanto riguarda le donne in Cut/Pre-contest?
Di quanto sopra elencato, per un efficace controllo estrogenico in una atleta è sufficiente l’uso combinato di un SERM e un AI. Il Tamoxifene può essere sostituito con il Raloxifene anche se di minore potenza d’impatto. Il Letrozolo risulta invece una potenziale sostituzione al Anastrozolo, diversamente dagli uomini. In caso l’atleta abbia tra gli AAS in uso Drostanolone e/o Methenolone, il discorso fatto pocanzi continua ad essere valido e andranno calibrate con maggiore accuratezza, mgXmg, il SERM e l’AI inseriti. Anche in questo caso occorre dare particolarmente attenzione ai tempi di soppressione e allo scalo graduale delle molecole regolatrici al termine della preparatzione.
Conclusioni:
Abbiamo visto come gestire un eventuale iperestrogenemia sia per variabili di condizione della composizione-corporea e sia adattativa in TRT e come poter gestire al meglio in termini di vantaggi sulla composizione corporea il fattore estrogenico in Bulk/Off-Season e in Cut/Pre-Contest.
Quindi, i bodybuilder in Off-Season/Bulk beneficiano di un aumento controllato dell’E₂ grazie a una migliore risposta agli stimoli anabolici e alla riparazione (effetti sul danno muscolare). Un adeguato controllo del E2 anche in questa fase rende più facile la riduzione/gestione dei sides legati ad un aumento della attività estrogenica come la ritenzione di liquidi e una distribuzione del grasso più femminile, sui fianchi e sui glutei.
L’Aromatizzazione dei substrati soggetti porta ad un aumento dell’IGF-I e quindi delle possibilità di crescita muscolare, ma i livelli di E₂ nel sangue devono essere ottimizzati piuttosto che massimizzati a causa della forma parabolica, a U inversa, della curva E₂/IGF-I. Occorre tuttavia tenere conto del carico di allenamento, a causa degli effetti deleteri degli estrogeni sulla meccanica di tendini e legamenti (riduzione della rigidità). L’uso eccessivo di farmaci e non strategico di inibitori dell’aromatasi (AI) può avere conseguenze indesiderate attraverso la riduzione degli estrogeni, in particolare sul metabolismo osseo, sulla gestione dei lipidi e sulle potenzialità ipertrofiche muscolari, in particolare sulla risposta anabolica all’allenamento. Ecco perchè l’unico momento funzionale ad una maggiore soppressione della attività estrogenica, come abbiamo visto, risulta essere limitata al Cut/Pre-Contest. E’ consigliabile, quindi, cercare di mantenere un range ottimale del E2 per la maggior parte dell’anno. Tale range dovrebbe attestarsi tra i 40-60pg/dL ± 30pg [ossia con una punta massima di tolleranza pari a 90pg/ml] dipendente dalla sensibilità soggettiva.
Gabriel Bellizzi [CEO BioGenTech]
Approfondimenti bibliografici:
Cooke PS, Nanjappa MK, Ko C, Prins GS, Hess RA. Estrogens in Male Physiology. Physiol Rev. 2017 Jul 1;97(3):995-1043. doi:10.1152/physrev.00018.2016.
Chidi-Ogbolu N, Baar K. Effect of Estrogen on Musculoskeletal Performance and Injury Risk. Front Physiol. 2019;9:1834. Published 2019 Jan 15. doi:10.3389/fphys.2018.01834
Zhang Y, Stefanovic B. mTORC1 phosphorylates LARP6 to stimulate type I collagen expression. Sci Rep. 2017 Jan 23;7:41173. doi:10.1038/srep41173.
Kim, J. H., Cho, H. T., and Kim, Y. J. (2014). The role of estrogen in adipose tissue metabolism: insights into glucose homeostasis regulation [Review]. Endocrine Journal, 61(11), 1055–1067. doi:10.1507/endocrj.ej14-0262
Eyster, K. M. (Ed.). (2016). Estrogen Receptors. Methods in Molecular Biology. doi:10.1007/978-1-4939-3127-9
Cauley, J. A. (2015). Estrogen and bone health in men and women. Steroids, 99, 11–15. doi:10.1 016/j.steroids.2014.12.010
Veldhuis, J. D., and Bowers, C. Y. (2003). Human GH pulsatility: An ensemble property regulated by age and gender. Journal of Endocrinological Investigation, 26(9), 799–813. doi:10.1007/bf03345229
Damewood MD, Bellantoni JJ, Bachorik PS, Kimball AW Jr, Rock JA. Exogenous estrogen effect on lipid/lipoprotein cholesterol in transsexual males. J Endocrinol Invest. 1989 Jul-Aug;12(7):449-54. doi:10.1007/BF03350728.
Rubinow KB. Estrogens and Body Weight Regulation in Men. Adv Exp Med Biol. 2017;1043:285-313. doi:10.1007/978-3-319-70178-3_14
Stachenfeld, N. S. (2014). Hormonal Changes During Menopause and the Impact on Fluid Regulation. Reproductive Sciences, 21(5), 555–561. doi:10.1177/1933719113518992
Curtis, K. S. (2015). Estradiol and osmolality: Behavioral responses and central pathways. Physiology and Behavior, 152, 422–430. doi:10.1016/j.physbeh.2015.06.017
Warner M, Fan X, Strom A, Wu W, Gustafsson JÅ. 25 years of ERβ: a personal journey. J Mol Endocrinol. 2021 Oct 15;68(1):R1-R9. doi: 10.1530/JME-21-0121
Kley, H. K., Deselaers, T., Peerenboom, H., and Kruskemer, H. L. (1980). Enhanced Conversion of Androstenedione to Estrogens in Obese Males*. The Journal of Clinical Endocrinology and Metabolism, 51(5), 1128–1132. doi:10.1210/jcem-51-5-1128
Thompson, P. D. (1989). Contrasting Effects of Testosterone and Stanozolol on Serum Lipoprotein Levels. JAMA: The Journal of the American Medical Association, 261(8), 1165. doi:10.1001/jama.1989.03420080085036
Friedl, K. E., Hannan, C. J., Jones, R. E., and Plymate, S. R. (1990). High-density lipoprotein cholesterol is not decreased if an aromatizable androgen is administered. Metabolism, 39(1), 69–74. doi:10.1016/0026-0495(90)90150-b
Hartgens, F. (2004). Effects of androgenic-anabolic steroids on apolipoproteins and lipoprotein (a). British Journal of Sports Medicine, 38(3), 253–259. doi:10.1136/bjsm.2003.000199
Perkins, M. S., Louw-du Toit, R., and Africander, D. (2017). A comparative characterization of estrogens used in hormone therapy via estrogen receptor (ER)-α and -β. The Journal of Steroid Biochemistry and Molecular Biology, 174, 27–39. doi:10.1016/j.jsbmb.2017.07.022
Bond P, Smit DL, de Ronde W. Anabolic-androgenic steroids: How do they work and what are the risks? Front Endocrinol (Lausanne). 2022 Dec 19;13:1059473. doi: 10.3389/fendo.2022.1059473. PMID: 36644692; PMCID: PMC9837614.
Lin, S. X., Han, Q., Azzi, A., Zhu, D.-W., Gongloff, A., & Campbell, R. L. (1999). 3D-structure of human estrogenic 17β-HSD1: binding with various steroids. The Journal of Steroid Biochemistry and Molecular Biology, 69(1-6), 425–429. doi:10.1016/s0960-0760(99)00062-x
Pubblicato da Gabriel Bellizzi [also known as Ružička, The Biochemist] - CEO BioGenTech -
Negli anni trenta del ventesimo secolo si è verificata una febbre dell’oro scientifica di proporzioni inaudite nel campo della nascente endocrinologia. Questa impresa è stata portata avanti con tanta celerità grazie al pionieristico lavoro di biochimici Adolf Friedrich Johann Butenandt e Lavoslav Stjepan Ružička, entrambi premi Nobel per la chimica nel 1939 grazie proprio alla pubblicazione dell’articolo “Sulla preparazione artificiale dell’ormone testicolare testosterone (androstene-3-one--17-olio)”.
Il potenziale del Testosterone e dei suoi primi derivati che videro la luce nella seconda metà degli anni trenta del 900, arrivo’ all’orecchio degli sportivi d’élite tanto che nel 1938 vi fu una prima pubblicazione che parlava del potenziale uso del Testosterone nel Bodybuilding.
Grazie agli abbattimenti dei costi di produzione delle molecole di sintesi, resi possibili dal genio della chimica Russell Earl Marker e dalla sua “Marker degradation”, nella seconda metà degli anni quaranta l’uso di AAS si è diffuso nelle squadre olimpiche di molti paesi. Successivamente tocco’ al pubblico amatoriale. E' nel 1976 che vi fu una nuova svolta, cioè la nascita della società di biotecnologie “Genetech” nata dall’incontro tra l’imprenditore Robert Swanson e Herbert Boyer, biochimico dell’Università della California. I due decisero di fondare questa società per lo sfruttamento commerciale delle tecniche del DNA ricombinante messe a punto da Boyer. Insulina e hGH divennero parte del corollario di farmaci utilizzati dai bodybuilder, e l’era dei “Freak” venne inaugurata.
Purtroppo, lo “scandalo DOPING” negli anni 80’, e le successive restrizioni di “facciata” hanno smantellato massivamente quella nicchia di ricercatori che lavoravano a stretto contatto con gli atleti e facevano ricerca sul campo. Essi non sono “estinti” ma sono obliati da una certa narrativa di comodo. Da qui il problema presente: l’atleta è in balia di leggende e metodiche partorite da menti non avvezze alla complessità della farmacologia partendo dalle basi della biochimica.
La BioGenTech è un laboratorio di ricerca che opera direttamente sul campo dapprima della sua fondazione grazie al lavoro del CEO Amedeo Gabriel Bellizzi. Nel 2021, ha visto la luce e ha preso concretezza un idea: fornire informazioni valide e affidabili su una scienza multidisciplinare. Nessun circo delle pulci, ma qualcosa che si può vedere e constatare.
Noi alla BioGenTech, la quale è una realtà collaborativa sebbene diretta da una mente, siamo scienziati puri con un atteggiamento snobistico nei confronti dei soli affari. Riteniamo la sola corsa al denaro una cosa da bottegai, poco stimolante dal punto di vista intellettuale. E la ricerca al servizio del commerciale, quindi resa scientismo, può andare bene solo per chi non e’ dotato di etica o è limitato nella materia.
Quindi il nostro atteggiamento nei confronti di chi e’ impegnato nello scientismo speculativo, e’ essenzialmente di critica e avversione. Il Nostro tradizionale antagonismo fa sì che non subiamo la contaminazione del marketing e, ogniqualvolta si scatena un dibattito su questioni biotecnoiogiche, non manchiamo di porci al di sopra delle parti discutendo dei problemi ai massimi livelli.
Contattaci per informazioni su coaching, anti-aging e TRT/HRT sulle piattaforme Instagram, Telegram, Whatsapp o all’indirizzo mail teamympus86@outlook.it
CEO Amedeo Gabriel Bellizzi [Biochimico, esperto in nutrizione sportiva, coach di BodyBuilding, PEDs consulter, esperto in tecniche Anti-Aging, TRT e HRT, ricercatore e divulgatore scientifico indipendente]
Vedi tutti gli articoli di Gabriel Bellizzi [also known as Ružička, The Biochemist] - CEO BioGenTech -