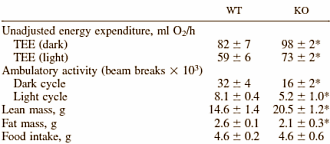Una delle singolarità che maggiormente caratterizzano l’Ormone della Crescita (GH) è rappresentata dall’aumento della sua secrezione correlata al sonno, che si verifica vicino all’inizio del così detto sonno a onde lente (fase tre) [1-2]. Questa secrezione notturna rappresenta quasi il 70% dell’intera sintesi giornaliera di GH secreta negli individui di sesso maschile. L’impulso notturno è di natura sessualmente dimorfica e significativamente meno pronunciato nelle donne [3-4].
Poiché questo è un evento secretorio endogeno così sostanziale, molte persone, sia atleti che preparatori, basandosi anche su speculazioni frutto di teorie più o meno plausibili esposte su alcuni libri del settore (errore in cui caddi anche io), nel tentativo di massimizzare l’efficacia dell’utilizzo di GH esogeno spesso si convincono del fatto che un particolare protocollo di temporizzazione possa essere usato per preservare questa secrezione notturna. Ma, questa ipotesi ha un reale riscontro nella pratica?
Come in tutti gli altri sistemi endocrini, la secrezione di GH è regolata da molteplici cicli di feedback negativo che sono tradizionalmente suddivisi in tre categorie: feedback ultracorto, corto, e lungo [5]. Il ciclo di feedback ultracorto è rappresentato dall’azione del GHRH (Growth hormone releasing hormone; Ormone di Rilascio della Somatotropina o Somatorelina) – che inibisce acutamente la secrezione di GH – e della Somatostatina (SRIF) – che sopprime il rilascio di GH [6-8].
Il ciclo di feedback corto è rappresentato dall’azione dei livelli sierici di GH elevati che agiscono direttamente sull’ipofisi per inibire l’ulteriore il rilascio di GH attraverso la soppressione del GHRH. Infatti, gli elevati livelli sierici di GH possono anche indurre l’inibizione del SRIF e del GHRH all’interno dell’ipotalamo. Sembra però che tale processo richieda un certo lasso di tempo per manifestarsi, poiché alcune prove hanno dimostrato che questo particolare circuito di feedback può richiedere da due a quattro ore per impostarsi [9-14]. Questo ciclo di feedback è anche quello sul quale prestiamo maggiore interesse.
Per completezza d’informazioni, il ciclo di feedback lungo si verifica quando elevati livelli sierici di IGF-1 agiscono sull’ipofisi riducendo consequenzialmente la secrezione di GH [15-16].
Fortunatamente, esistono due studi sull’uomo che possono fare ulteriore chiarezza in merito al quesito posto in questa sede. Il primo studio ha preso in esame soggetti sani di sesso maschile e femminile sottoponendoli ha somministrazioni di GH esogeno pari a 2UI/die divise in due iniezioni: una prima iniezione AM (08:00) e una seconda iniezione PM (17:00) [17]. Nonostante l’ultima somministrazione di GH sia avvenuta più di sei ore prima del sonno, il gruppo di ricercatori ha notato una completa soppressione delle secrezioni endogene notturna dell’ormone nonostante i livelli elevati di GH fossero stati eliminati da tempo dal sistema. Ciò sembrerebbe implicare l’azione di un meccanismo di feedback oltre a quello legato agli effetti diretti del GH.
Secrezione dell’Ormone della Crescita correlata al sonno dopo somministrazione di GH esogeno o soluzione salina – ore dopo l’inizio del sonno.
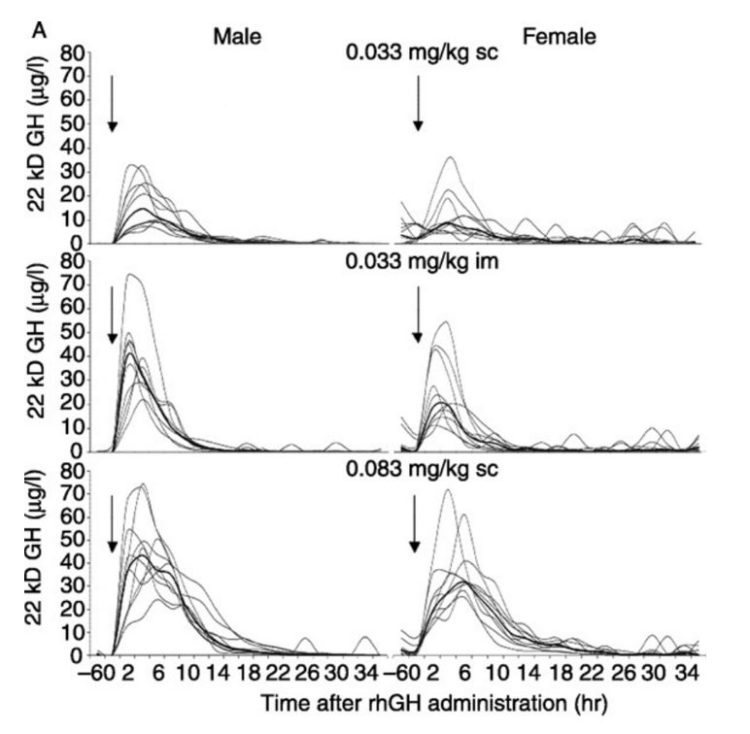
Il secondo studio [18] ha preso in esame soggetti sani di sesso maschile sottoponendoli alla somministrazione di una singola dose sottocutanea di 20kDa GH alle 21:00. La dose era variabile ed i soggetti sono stati divisi in quattro gruppi di dosaggio: 0.01, 0.025, 0.05 o 0.1mg/kg. Con uno scarto di alcune ore, tutti i gruppi di dosaggio hanno mostrato una completa soppressione delle secrezioni endogena di GH per un lasso di tempo pari a 16-24 ore. Infatti, eliminando due valori anomali, i soggetti rimanenti hanno mostrato una completa soppressione per tutte le 24 ore del monitoraggio. È stato interessante notare che uno dei valori anomali era nel gruppo ad alto dosaggio.
Secrezione dell’Ormone della Crescita correlata al sonno dopo somministrazione di GH esogeno o soluzione salina – tempo dopo la somministrazione.
Basandomi su quanto esposto, penso che il vero punto da tenere presente sia che gli utilizzatori di GH esogeno non dovrebbero concentrarsi sulla secrezione endogena di GH nelle ore notturne nel vano tentativo di preservare tale impulso. Come si è potuto vedere, sembrerebbe che qualsiasi dose significativa di GH esogeno aumenti la probabilità che tale impulso venga soppresso a prescindere. Infatti, a meno che non si utilizzi il GH esclusivamente per scopi lipolitici, l’impulso serale probabilmente non fornirà comunque un effetto additivo semplicemente a causa di come esso è strutturato.
Gabriel Bellizzi
Riferimenti:
Exogenous Growth Hormone and Nocturnal Secretions – By Chest Rockwell
Holl RW, Hartman ML, Veldhuis JD, Taylor WM, Thorner MO. Thirty-second sampling of plasma growth hormone in man: correlation with sleep stages. J Clin Endocrinol Metab. 1991 Apr;72(4):854-61
Obal F Jr, Krueger JM. GHRH and sleep. Sleep Med Rev. 2004 Oct;8(5):367-77. Review Van Cauter E, Plat L, Copinschi G. Interrelations between sleep and the somatotropic axis. Sleep. 1998 Sep 15;21(6):553-66. Review.
Jaffe CA, Ocampo-Lim B, Guo W, Krueger K, Sugahara I, DeMott-Friberg R, Bermann M, Barkan AL. Regulatory mechanisms of growth hormone secretion are sexually dimorphic. J Clin Invest. 1998 Jul 1;102(1):153-64.
Farhy LS, Straume M, Johnson ML, Kovatchev B, Veldhuis JD. Unequal autonegative feedback by GH models the sexual dimorphism in GH secretory dynamics. Am J Physiol Regul Integr Comp Physiol. 2002 Mar;282(3):R753-64.
Lumpkin MD, McDonald JK. Blockade of growth hormone-releasing factor (GRF) activity in the pituitary and hypothalamus of the conscious rat with a peptidic GRF antagonist. Endocrinology. 1989 Mar;124(3):1522-31.
Lumpkin MD, Mulroney SE, Haramati A. Inhibition of pulsatile growth hormone (GH) secretion and somatic growth in immature rats with a synthetic GH-releasing factor antagonist. Endocrinology. 1989 Mar;124(3):1154-9.
Berelowitz M, Firestone SL, Frohman LA. Effects of growth hormone excess and deficiency on hypothalamic somatostatin content and release and on tissue somatostatin distribution. Endocrinology. 1981 Sep;109(3):714-9.
Chomczynski P, Downs TR, Frohman LA. Feedback regulation of growth hormone (GH)-releasing hormone gene expression by GH in rat hypothalamus. Mol Endocrinol. 1988 Mar;2(3):236-41
Frohman MA, Downs TR, Chomczynski P, Frohman LA. Cloning and characterization of mouse growth hormone-releasing hormone (GRH) complementary DNA: increased GRH messenger RNA levels in the growth hormone-deficient lit/lit mouse. Mol Endocrinol. 1989 Oct;3(10):1529-36.
Rosenthal SM, Kaplan SL, Grumbach MM. Short term continuous intravenous infusion of growth hormone (GH) inhibits GH-releasing hormone-induced GH secretion: a time-dependent effect. J Clin Endocrinol Metab. 1989 Jun;68(6):1101-5.
Yamauchi N, Shibasaki T, Ling N, Demura H. In vitro release of growth hormone-releasing factor (GRF) from the hypothalamus: somatostatin inhibits GRF release. Regul Pept. 1991 Mar 26;33(1):71-8.
Pontiroli AE, Lanzi R, Monti LD, Sandoli E, Pozza G. Growth hormone (GH) autofeedback on GH response to GH-releasing hormone. Role of free fatty acids and somatostatin. J Clin Endocrinol Metab. 1991 Feb;72(2):492-5.
Hashimoto Y, Kamioka T, Hosaka M, Mabuchi K, Mizuchi A, Shimazaki Y, Tsunoo M, Tanaka T. Exogenous 20K growth hormone (GH) suppresses endogenous 22K GH secretion in normal men. J Clin Endocrinol Metab. 2000 Feb;85(2):601-6.
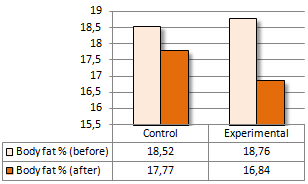
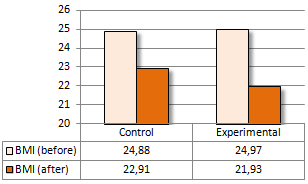
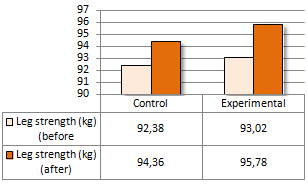
Una supplementazione di Colina pari a 2g al giorno sembra accelerare la perdita di peso in abbinamento ad una dieta ipocalorica. Gli scienziati della Zagazig University in Egitto sono giunti a questa conclusione dopo aver somministrato Colina a praticanti di arti marziali per una settimana. (1)
I ricercatori egiziani volevano constatare se la Colina potesse essere un supplemento alternativo alla Caffeina. La Caffeina, come risaputo, è un integratore usato efficacemente come coadiuvante per la perdita di peso, ma non tutti reagiscono bene alle dosi richieste per far si che la supplementazione sia efficace.
La Colina è una sostanza vitamina-simile che il corpo sintetizza in piccole quantità e di cui abbiamo necessità di assumerne circa 400-550 mg con la dieta. La Colina si trova nei tuorli, nei crostacei e nella carne. La Colina è un elemento fondamentale per le membrane cellulari e per la sintesi del neurotrasmettitore Acetilcolina e svolge un ruolo nel metabolismo lipidico.
I ricercatori hanno svolto l’esperimento utilizzando un campione di 22 donne praticanti arti marziali – judo o taekwondo. Una metà dei soggetti presi in esame è stata somministrata una capsula contenente 1g di Colina due volte al giorno, mentre all’altra metà è stato somministrato un placebo.
Le donne dello studio si stavano preparando per una competizione, quindi molto probabilmente stavano seguendo una dieta ipocalorica, sebbene i ricercatori non hanno divulgato alcuna informazione sulla dieta dei soggetti presi in esame.
La percentuale di grasso delle donne nel gruppo sperimentale è diminuita due volte più velocemente rispetto a quella delle donne nel gruppo di controllo. Questo è stato un effetto statisticamente significativo.
Il BMI delle donne prese in esame è calato in entrambi i gruppi. La diminuzione sembrava essere più veloce nel gruppo sperimentale, ma secondo i calcoli dei ricercatori l’effetto della supplementazione di Colina sulla riduzione del BMI non era statisticamente significativo.
Nella settimana in cui si è svolto l’esperimento, la forza muscolare delle donne prese in esame non è diminuita – in entrambi i gruppi.
I ricercatori hanno concluso affermando che, l’integrazione di Colina negli atleti di taekwondo e judo di sesso femminile può migliorare il metabolismo dei lipidi e favorire una rapida riduzione della massa corporea.
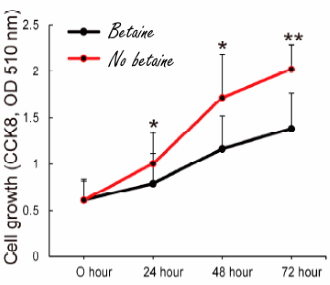
Una consistente supplementazione di Betaina sembra poter inibire la crescita delle cellule adipose. Secondo uno studio sugli animali, svolto dai ricercatori della Sichuan Agricultural University e pubblicato su Nutrients,(1) la Betaina consente al corpo di accumulare più massa magra mentre al tempo stesso esercita una sorta di controllo sulla crescita della massa grassa. Ciò è stato osservato quando il corpo riceve più energia di quella che consuma, ma anche quando l’apporto calorico e il dispendio calorico sono perfettamente bilanciati.
I ricercatori hanno svolto il presente sperimentato su diversi gruppi di topi. Alcuni gruppi sono stati alimentati con del grasso addizionale, altri gruppi sono stati alimentati con mangimi standard. Ad altri gruppi, invece, ad alcuni degli esemplari trattati è stata somministrata della Betaina attraverso l’acqua.
L’equivalente umano della dose utilizzata dai ricercatori è di 10-15 grammi di Betaina al giorno. Questo è un dosaggio decisamente più elevato rispetto ai 2-3 grammi raccomandati dalla maggior parte dei produttori di integratori. Non so con certezza se 10 grammi di Betaina al giorno siano dannosi. L’unico effetto collaterale da alte dosi di Betaina nel quale si potrebbe incorrere è un aumento della produzione di acido dello stomaco (non propriamente blando come possibile effetto).
Naturalmente, i topi sottoposti ad una alimentazione ipercalorica – con grasso aggiunto – [HFD] sono diventati più grassi dei topi che hanno ricevuto il normale mangime [NCW].
Nei topi alimentati con del normale mangime, l’integrazione con Betaina [HC-bet] ha avuto un effetto dimagrante. Nei topi i quali venivano alimentati con un surplus calorico in riferimento al loro dispendio energetico [H-BET], sembra che la supplementazione con Betaina abbia indotto questi animali a usare l’energia in eccesso per aumentare la loro massa corporea magra invece della loro massa grassa.
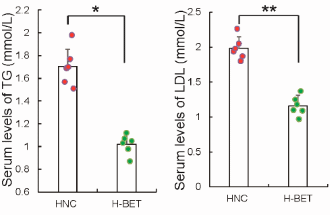
Negli animali sottoposti ad un regime alimentare ipercalorico (e che sono stati fatti ingrassare), la Betaina ha permesso che le concentrazioni di Trigliceridi [TG] e di colesterolo LDL rimanessero basse.
Quando i ricercatori hanno svolto esperimenti in vitro sulle cellule adipose 3T3-L1, hanno osservato che la Betaina ha un effetto inibente sulla loro crescita. La Betaina ha aumentato l’attività degli enzimi di crescita cyclin-D ed -E e ha attivato i geni inibitori della crescita P53 e P21. [Gli ultimi due geni inibiscono anche la crescita delle cellule tumorali e stimolano la produzione di mitocondri in numerosi tipi di cellule.]
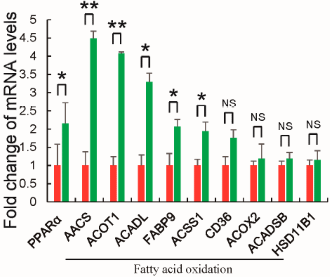
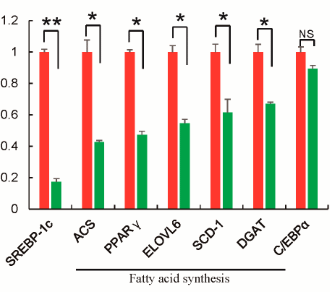
Nelle cellule adipose, la Betaina ha ridotto l’attività degli enzimi che accumulano calorie sotto forma di grasso. Nelle cellule adipose, tuttavia, l’attività degli enzimi che ossidano i grassi è aumentata. La Betaina sembra quindi presentare grosse similitudini d’effetto con la curcumina (2) e il tè verde (3).
Il presente studio ha mostrato che il trattamento con Betaina non solo ha la potenzialità di inibire significativamente la proliferazione e la differenziazione degli adipociti 3T3-L1 in vitro, ma ha anche notevolmente soppresso l’accumulo del tessuto adiposo biancho in vivo.
I ricercatori sottolineano il fatto che, dopo la supplementazione con Betaina, i topi nutriti con una dieta ricca di grassi mostravano una significativa riduzione dei livelli plasmatici di lipidi e lipoproteine, come Trigliceridi e l’LDL.
Inoltre, gli autori sottolineano nuovamente il fatto che, durante lo studio è stata osservata una ridotta espressione di alcuni geni che promuovono la sintesi lipidica nel tessuto adiposo bianco nei topi nutriti con una dieta ad alto contenuto di grassi.
Quello che mi accingo a riportare in questo articolo è il protocollo di GH per la perdita di grasso ideato da Chest Rockwell.
Storia del GH e perdita di grasso
Cos’è la lipolisi?
Relazione tra ormone dello stress / GH e digiuno
Meccanismi d’azione – Lipolisi GH-mediata
Tasso massimo di lipolisi
Farmacocinetica / Farmacodinamica
Sinergie tra i composti
Applicazione pratica e progettazione di uno stack di esempio
Storia del GH e perdita di grasso
Ipotalamo
L’Ormone della Crescita (GH) è un potente agente di mobilizzazione dei grassi i cui effetti, se bene all’epoca non ancora imputabili all’ormone in se, vennero osservati dai ricercatori fin dagli anni ’20, quando primitivi studi sugli animali dimostrarono che gli esemplari trattati con l’ipofisi erano più magri rispetto agli animali di controllo non trattati [1-2]. Tuttavia, si dovette attendere fino al 1945, quando il GH fu estratto per la prima volta dall’ipofisi [3], affinché gli scienziati iniziassero veramente a isolare gli effetti di mobilizzazione dei grassi dell’ipofisi su questo specifico polipeptide.
Cos’è la lipolisi?
Triacilglicerolo
La lipolisi è il processo fisiologico che fornisce all’organismo un substrato energetico sotto forma di acidi grassi attraverso il catabolismo del triacilglicerolo immagazzinato. Il triacilglicerolo immagazzinato all’interno delle goccioline lipidiche degli adipociti viene idrolizzato in acidi grassi e glicerolo e successivamente rilasciato nel circolo ematico per ossidazione e produzione di ATP [4-5]. Sarà importante capire le differenze tra mobilitazione e ossidazione dei grassi; tuttavia una revisione completa della lipolisi va oltre lo scopo di questo articolo.
Relazione tra ormone dello stress / GH e digiuno
Il GH è definibile come “ormone dello stress”, per sua natura, con l’aumento della sua secrezione durante eventi stressanti come il digiuno e l’esercizio fisico [6]. Per il resto di questo articolo, tuttavia, ci concentreremo maggiormente sul ruolo che il GH svolge durante il digiuno e su come sia applicabile a coloro che cercano di massimizzare il potenziale lipolitico dei loro protocolli di GH esogeno.
Il periodo di digiuno (post-riassorbimento) inizia circa sei ore dopo l’assunzione di cibo e gli obiettivi primari del corpo durante questa fase sono di fornire, convertire e conservare i substrati energetici. Durante questo periodo, vi è un aumento significativo del tasso di secrezione di GH che può durare per 48-72 ore [7-8]. L’innalzamento del GH secreto è direttamente correlato all’ampiezza dell’impulso, mentre la frequenza degli impulsi e i livelli di intervallo basali rimangono sostanzialmente invariati [9]. Durante il digiuno, il GH è l’unico ormone anabolico ad aumentare mentre aumentano i livelli degli ormoni catabolici (ad esempio Glucagone, Cortisolo, Epinefrina ecc.).
Meccanismi d’azione – Lipolisi GH-mediata
Le elevate percentuali di secrezione di GH portano con sé numerosi cambiamenti metabolici, che sono importanti da comprendere. La prima priorità del corpo durante il digiuno è quella di mantenere l’omeostasi del glucosio, in modo da fornire sufficiente glucosio al cervello e ad altri tessuti (ad esempio globuli rossi) che dipendono, principalmente o tassativamente (tessuti glucosio-prefereziali e glucosio-dipendenti), da questo substrato energetico [10]. Per raggiungere questo obiettivo, si verifica uno shift metabolico a favore del metabolismo lipidico in modo che vengano salvaguardate le preziose riserve glucidiche e aminoacidiche. Parallelamente a questo cambiamento shift metabolico a favore del metabolismo lipidico da parte del tessuto muscolare ed epatico, viene attuata la mobilizzazione del glicogeno in quanto non viene rilevato alcun apporto dietetico di glucosio. Grandi quantità di glucosio vengono anche rilasciate dal fegato nel flusso ematico per aiutare a mantenere i livelli di glucosio nel sangue in assenza di un apporto di glucosio con la dieta. Ciò è possibile, in gran parte, grazie al calo simultaneo dei livelli sierici di insulina che impedisce al glucosio rilasciato di entrare nei muscoli e nel tessuto adiposo.
Oltre a questo, con l’aumento del GH si verifica uno stato di insulino-resistenza, vitale per la conservazione delle preziose riserve di glucosio. Questi effetti antagonisti all’insulina che il GH porta con sé riducono l’ossidazione del glucosio e, al contrario, la necessità di precursori gluconeogenici delle riserve di proteine muscolari [11]. Ci sarebbero alcune riflessioni da fare sull’eventualità che sia lo stesso GH o l’aumento degli FFA i principali responsabili dell’instaurarsi di una accentuata insulino-resistenza, tuttavia, la questione potrà essere trattata in un altro articolo. Quindi, per legare tutto ciò che è stato fino ad ora detto, durante il digiuno l’aumento della secrezione di GH porta ad un aumento della mobilitazione degli FFA dagli adipociti, sotto-regola i GLUT-1 per inibire l’assorbimento di glucosio nei tessuti periferici, previene l’ossidazione del glucosio aumentando l’insulino-resistenza e preserva le riserve amminiche sia in modo diretto che indiretto.
È ben noto che il GH influenza la lipolisi, tuttavia i meccanismi esatti con cui lo fa rimangono in qualche modo elusivi. È stato ipotizzato che questo potrebbe essere multifattoriale con il GH il quale mostra la capacità di ridurre la lipoproteina lipasi del tessuto adiposo (LPL), stimolare la lipasi ormono-sensibile (HSL) e antagonizzare gli effetti antilipolitici dell’insulina. L’aumentata espressione del HSL negli adipociti aumenta il potenziale lipolitico in quanto l’HSL è intimamente coinvolto nel processo di idrolisi del triacilglicerolo. Una volta attivato, l’HSL viene trasportato alla periferia della goccia lipidica intracellulare dove idrolizza il triacilglicerolo a FFA e glicerolo. È anche riconosciuto da molti come l’enzima che determina la velocità della lipolisi [12]. Vale la pena notare che non tutti gli studi hanno universalmente dimostrato che il GH aumenta i livelli di mRNA del HSL negli adipociti [13-14].
Come già accennato, è stato anche dimostrato che il GH ha un impatto diretto sulla soppressione dell’attività della LPL nei tessuti adiposi umani, sebbene ciò non sia stato dimostrato nei tessuti muscoli scheletrici [15-17]. Il motivo per il quale ciò è potenzialmente rilevante per chi è interessato alla perdita di grasso è legato al fatto che la LPL è direttamente coinvolta nella liberazione degli acidi grassi nel flusso ematico, nel loro successivo immagazzinamento negli adipociti e / o renderli disponibili per la beta-ossidazione da parte dei tessuti muscolo scheletrici e del muscolo cardiaco. Quindi, se la LPL può essere soppressa nei tessuti adiposi, si potrebbe ipotizzare che meno acidi grassi potranno essere immagazzinati attivamente (riesterificati) mentre altri saranno disponibili per alimentare i processi metabolici.
Studi condotti su adipociti umani in coltura hanno dimostrato che il GH in realtà non ha effetti lipolitici diretti [18], ma aumenta significativamente la sensibilità alle catecolamine in queste cellule, suggerendo che il GH attivi la lipolisi in uno stadio successivo al coinvolgimento dei beta-adrenorecettori e / o Proteine-G. In effetti, è ragionevole supporre che il GH possa aumentare la densità dei beta-adrenorecettori, rendendo questo fattore di grande interesse per un protocollo volto a massimizzare la lipolisi. È stato precedentemente dimostrato che esistono beta-adrenocettori “di riserva” sugli adipociti umani e un aumento acuto del numero di recettori accoppiati aumenterebbe la sensibilità e, in definitiva, il potenziale lipolitico [19]. E, in modelli animali, è stato dimostrato che il GH aumenta specificamente l’espressione dei recettori β3-adrenergici negli adipociti, seguita dall’attivazione del HSL [20]. Quindi, oltre agli effetti direttamente mediati che il GH possiede, è ovvio che l’uso di un agonista beta-adrenergico potrebbe molto probabilmente creare un effetto additivo sul processo lipolitico.
Tasso massimo di lipolisi
Ormone della Crescita (GH)
Quindi, mentre ci si adopera per creare uno stack che massimizzi il potenziale lipolitico del GH, esiste un limite effettivo (o teorico) alla velocità con cui ciò si verifichi? In realtà esiste una risposta; almeno per quanto riguarda esclusivamente il tasso massimo in cui il GH con somministrazione endovenosa possa provocare la lipolisi [21]. Si è constatata una dose di circa 3 mcg / kg (corrispondente a un picco medio di GH di 32,4 mcg / litro). La dose non era dipendente dall’età o dal sesso e si aggira intorno all’equivalente di 1,2-1,5 UI per un maschio magro di 100 kg. Una dose più elevata di questa non provoca in realtà un maggiore impatto sulla lipolisi. E’ interessante notare come questo sia anche essenzialmente il limite superiore dei picchi secretori endogeni naturali [22]. Si ipotizza che questo potrebbe essere un limite, o un collo di bottiglia, causato almeno in parte dai tassi di clearance extra-renale in concomitanza con i livelli di GHBP circolanti [23].
Ci sono alcune prove che questo sia un collo di bottiglia specifico del GH, e che i trattamenti combinati con varianti delle catecolamine producono un effetto additivo sulla lipolisi, maggiore di entrambi i singoli trattamenti [24-27] che supportano ulteriormente l’idea che il GH sia un mediatore lipolitico di mediazione. Alcuni hanno definito questo come un effetto permissivo sulla lipolisi indotta da catecolamine [28]. Aneddoticamente, sembra che un associazione di GH e composti catecolamine-stimolanti [29] abbia un effetto sinergico. Ma di questo se ne parlerà più approfonditamente proseguendo con questo articolo.
Farmacocinetica / Farmacodinamica
La farmacocinetica è una branca della farmacologia che studia gli effetti che i processi dell’organismo hanno sul farmaco (assorbimento, distribuzione, metabolismo, eliminazione). La farmacodinamica, invece, è lo studio degli effetti biochimici e fisiologici dei farmaci sull’organismo, ed il loro meccanismo d’azione [30]. È abbastanza importante capire entrambi, in relazione al GH, al fine di massimizzare il suo potenziale lipolitico e garantire che il GH somministrato sia ottimizzato così da prevenire la stesura di protocolli non corretti. Il GH lo sappiamo bene che è un composto costoso, quindi sarebbe nell’interesse dell’utilizzatore cercare di non sprecarlo.
Le vie di somministrazione modificano notevolmente la farmacocinetica del GH. Come discusso in precedenza, il GH endogeno viene secreto in maniera pulsatile e la sua presenza nel siero viene rapidamente eliminata attraverso i naturali feedback negativi di regolazione del corpo. Per simulare più strettamente questo comportamento secretorio, è necessario ricorrere alla somministrazione endovenosa una volta ogni 2-3 ore, che è il tempo necessario affinché il feedback ultra-corto (GHRH che inibisce la sua stessa secrezione) conduca al calo ematico dell’ormone. Tuttavia, a meno che non si è sotto la supervisione di un medico, questa non è una pratica che un qualsiasi preparatore degno di tale appellativo consiglierebbe e, infatti, in questo articolo ci si concentrerà invece sui due metodi di somministrazione del GH più popolari; sottocutaneo e intramuscolare.
A seguito di una iniezione SC o IM di GH, i livelli plasmatici di FFA e glicerolo aumentano dopo un breve periodo di latenza con un picco nei tassi di mobilizzazione dei grassi raggiunto intorno ai 150-160 minuti.
L’altro punto importante che vale la pena di sottolineare riguarda la tempistica d’iniezione del GH. Lo stato di digiuno è associato non solo a un aumento pronunciato della secrezione di GH endogeno, come precedentemente descritto, ma anche a un aumento della sensibilità lipolitica data dalla somministrazione di GH esogeno [33]. Infatti, tutti i marker lipolitici sono migliorati quando il GH viene somministrato a digiuno rispetto a quando viene somministrato in stato digestivo / postprandiale [34]. Anche il tasso di clearance globale è aumentato in modo significativo rimanendo nello stato di digiuno.
Vale anche la pena di notare che la somministrazione serale di GH ha dimostrato di avere una maggiore biodisponibilità, almeno quando somministrata a soggetti con deficit dell’Ormone della Crescita (GHD) [35]. Ciò potrebbe essere dovuto al fatto che tale somministrazioni mimi in modo ravvicinato il rilascio endogeno di GH nelle ore serali, ma questa è in gran parte solo una semplice speculazione.
Sinergie tra i composti
Recettore Beta-2 adrenergico
Quando si progetta uno stack finalizzato alla perdita di grasso che massimizza il potenziale lipolitico, bisogna scegliere con logica l’inserimento di composti con una potenziale sinergia, o con effetti additivi l’uno sull’altro. In precedenza, abbiamo discusso alcuni dei meccanismi mediante i quali il GH esercita i suoi effetti lipolitici. Uno dei suoi percorsi chiave coinvolge i recettori beta-adrenergici. Quindi sembrerebbe plausibile che se si è in grado di aumentare la sensibilità e / o l’espressione dei recettori beta-adrenergici, si possa ulteriormente ottimizzare la risposta lipolitica complessiva dello stack.
Recettore degli Androgeni
Gli androgeni inducono potenti effetti lipolitici direttamente attraverso i recettori degli androgeni (AR) espressi nel tessuto adiposo [36]. Questo è interessante, in sé e per sé, perché è un percorso diverso rispetto a quello utilizzato dal GH e, anche, perché è stato dimostrato che gli androgeni aumentano l’espressione dei recettore beta-adrenergici [37-38]. Si è già discusso del’aumento del numero dei recettori beta-adrenergici e della loro sensibilità e, in definitiva, come questo si rifletta positivamente sul potenziale lipolitico. Quindi questa potrebbe essere una potente sinergia nella metodologia di progettazione di uno stack.
L’asse tiroideo e l’asse GH / IGF hanno una relazione molto interessante, anche se complicata, l’una con l’altra. Di particolare interesse per il design di uno stack lipolitico sono i suoi impatti sull’mRNA del recettore beta-adrenergico e più specificamente i suoi impatti sull’espressione dei β3, che ha dimostrato di essere un passo critico nella lipolisi GH-mediata [39-41]. Inoltre gli ormoni tiroidei stimolano la sintesi, la degradazione e la mobilizzazione dei lipidi con conseguente aumento dei livelli degli FFA circolanti [42]. E l’ultima caratteristica sinergica che vale la pena di menzionare è che essi, proprio come il GH, sembrano regolare la sensibilità alle catecolamine dei processi metabolici.
E una volta aumentata la sensibilità e la densità dei recettori beta-adrenergici, è ovvio che un agonista beta-adrenergico sarebbe la ciliegina sulla torta per massimizzare il potenziale di mobilizzazione del grasso. Il Clenbuterolo [43] o il Salbutamolo sono entrambi candidati eccellenti per far parte di uno stack lipolitico sinergico; tuttavia una revisione completa degli agonisti beta-adrenergici va oltre lo scopo di questo articolo.
Applicazione pratica e progettazione di uno stack di esempio
Ora che si hanno le informazioni che stanno alla base di una progettazione oculata di uno stack lipolitico sinergico e avente come colonna portante l’uso del GH, come fare a progettare lo stack ed a gestire i composti facenti parte di esso?
Penso che sia abbastanza evidente il fatto che la somministrazione di GH (a fini lipolitici) debba essere fatta a digiuno. Sebbene gli effetti lipolitici del GH non siano completamente attenuati in presenza di cibo (diversamente dal GH endogeno), somministrandolo a stomaco pieno si eserciterebbe un serio antagonismo nei confronti dell’azione ricercata.
Utilizzando le informazioni esposte in precedenza riguardo al digiuno, insieme a una dose ben calibrata per ottenere la massima risposta lipolitica, si giunge all’applicazione di un’unica iniezione da 2UI di GH. Svolgere tale somministrazione al mattino appena svegli soddisfacerebbe molto bene i fini della pratica, poiché ci si troverà in uno stato di digiuno, più che probabile. Poiché le iniezioni sottocutanee hanno un tempo di eliminazione più lungo, questo sarebbe un buon metodo di somministrazione, soprattutto se si prevede di rimanere a digiuno per molte ore dopo l’iniezione. Per un potenziale effetto additivo sui tassi di mobilizzazione del grasso, vorrei anche prendere in considerazione l’esecuzione di attività strutturate (sia sotto forma di LISS che di allenamento di resistenza) durante questa finestra di digiuno [44].
Seguendo le stesse linee guida, si potrebbe molto probabilmente somministrare una seconda iniezione di 2UI di GH prima di andare a letto, sempre se si è in grado di consumare tutti i pasti della giornata in un lasso di tempo nel complesso ridotto ed entrare così nelle ore serali in uno stato di semi-digiuno. Come illustrato in precedenza, vi è anche evidenza che la somministrazione serale di GH per via sottocutanea fornisca una maggiore biodisponibilità. Quindi, se uno è costretto a scegliere dovrebbe farlo prendendo in considerazione una somministrazione AM o PM, in quanto questo sarebbe il tempo di somministrazione raccomandato. In caso contrario, l’utilizzo di una strategia che comprenda due iniezione da 2IU di GH, una somministrazione AM ed una PM, può teoricamente massimizzare il potenziale lipolitico nel corso delle 24 ore, assumendo che aderisca alle linee guida sul digiuno/semi-digiuno corrette.
Testosterone
L’uso di questo protocollo di dosaggio di GH, in combinazione con una co-somministrazione di androgeni, può aumentare il potenziale complessivo sulla perdita di grasso. Per questo motivo si dovrebbe prendere in considerazione l’uso di dosi sovra fisiologiche di AAS, oltre al fatto che tale inserimento previene il rischio di atrofia del tessuto magro durante periodi sostenuti di deficit calorico. Ci possono essere piccole differenze nel tasso di perdita di grasso prodotto dall’utilizzo di diversi tipi di androgeni, ed esso è strettamente legato al potenziale androgeno del composto, ma questo va oltre lo scopo di questo articolo. Anche il semplice inserimento di Testosterone nello stack funzionerebbe molto bene per lo scopo prefissato.
Aggiungendo sia composti tiroidei che Clenbuterolo, o Salbutamolo, nello stack si massimizzerà ulteriormente il potenziale lipolitico globale basato sulle loro proprietà sinergiche addizionate a quelle tra androgeni e GH spiegate in precedenza. A causa delle loro rispettive proprietà, il Clenbuterolo può essere somministrato una volta al giorno come pure il T4. Per quelli che usano il T3, si raccomanda un minimo di due dosi al giorno, distribuite a distanza di 12 ore. Chi sceglie di usare il Salbutamolo è raccomandabile distribuire la dose totale in più somministrazioni durante il giorno (4-5h di distanza l’una dall’atra).
Per riunire quanto detto fino ad ora, e al fine di renderlo comprensibile ai più, qui di seguito viene esposto uno schema breve ma decisamente esaustivo:
Cosomministrazione di AAS soprafisiologico
Cosomministrazione di Clenbuterolo o Salbutamolo + tiroidei
GH somministrato per via sottocutanea a una dose non superiore a 2 UI a digiuno due volte al giorno (AM / PM) o una volta al giorno (PM) se si è costretti a scegliere.
Attività strutturata eseguita durante lo stato di digiuno, post-iniezione per aumentare il potenziale di mobilizzazione del grasso.
Nota conclusiva
L’articolo ” The Most Effective Growth Hormone Protocol for Fat Loss”, il quale è stato utilizzato per la stesura del presente articolo, è un estratto sintetico ed abbreviato della sezione GH presente nel libro “The Growth Hormone Handbook” (titolo provvisorio) di Chester “Chest” Rockwell.
Gabriel Bellizzi
Riferimenti:
National Research Council (US) Committee on Technological Options to Improve the Nutritional Attributes of Animal Products. Designing Foods: Animal Product Options in the Marketplace. Washington (DC): National Academies Press (US); 1988. The Role of Growth Hormone in Fat Mobilization
Lee, M. O., and Schaffer, N. K.: Anterior Pituitary Growth Hormone and the Composition of Growth , J. Nutrition 7: 337 ( (March 10) ) 1934.
Li CH, Evans HM, Simpson ME. Isolation and properties of the anterior hypophyseal growth hormone. J Biol Chem. 1945;159:353–366.
Schoemaker RC, Buijs MM, Pijl H, Burggraaf J, Cohen AF. Modeling the influence of growth hormone on lipolysis. J Pharmacokinet Pharmacodyn. 2002 Apr;29(2):157-70
Lass A, Zimmermann R, Oberer M, Zechner R. Lipolysis – A highly regulated multi-enzyme complex mediates the catabolism of cellular fat stores. Progress in Lipid Research. 2011;50(1-4):14-27.
ROTH J, GLICK SM, YALOW RS, BERSON SA. Secretion of human growth hormone:physiologic and experimental modification. Metabolism. 1963 Jul;12:577-9
Hartman ML, Veldhuis JD, Johnson ML, Lee MM, Alberti KG, Samojlik E, Thorner MO. Augmented growth hormone (GH) secretory burst frequency and amplitude mediate enhanced GH secretion during a two-day fast in normal men. J Clin Endocrinol Metab. 1992 Apr;74(4):757-65.
Ho PJ, Friberg RD, Barkan AL. Regulation of pulsatile growth hormone secretion by fasting in normal subjects and patients with acromegaly. J Clin Endocrinol Metab. 1992 Sep;75(3):812-9.
Goldenberg N, Barkan A. Factors regulating growth hormone secretion in humans. Endocrinol Metab Clin North Am. 2007 Mar;36(1):37-55. Review
Berg JM, Tymoczko JL, Stryer L. Biochemistry. 5th edition. New York: W H Freeman; 2002. Section 30.3, Food Intake and Starvation Induce Metabolic Changes.
Nørrelund H. The metabolic role of growth hormone in humans with particular reference to fasting. Growth Horm IGF Res. 2005 Apr;15(2):95-122. Review.
Birnbaum MJ. Lipolysis: more than just a lipase. J Cell Biol. 2003 Jun 23;161(6):1011-2. Epub 2003 Jun 16. Review.
Slavin BG, Ong JM, Kern PA. Hormonal regulation of hormone-sensitive lipase activity and mRNA levels in isolated rat adipocytes. J Lipid Res. 1994 Sep;35(9):1535-41.
Ottosson M, Vikman-Adolfsson K, Enerbäck S, Elander A, Björntorp P, Edén S. Growth hormone inhibits lipoprotein lipase activity in human adipose tissue. J Clin Endocrinol Metab. 1995 Mar;80(3):936-41.
Richelsen B. Effect of growth hormone on adipose tissue and skeletal muscle lipoprotein lipase activity in humans. J Endocrinol Invest. 1999;22(5 Suppl):10-5. Review.
Richelsen B, Pedersen SB, Kristensen K, Børglum JD, Nørrelund H, Christiansen JS, Jørgensen JO. Regulation of lipoprotein lipase and hormone-sensitive lipase activity and gene expression in adipose and muscle tissue by growth hormone treatment during weight loss in obese patients. Metabolism. 2000 Jul;49(7):906-11.
Marcus C, Margery V, Kamel A, Brönnegård M. Effects of growth hormone on lipolysis in humans. Acta Paediatr Suppl. 1994 Dec;406:54-8; discussion 59. Review.
Arner P, Hellmér J, Wennlund A, Ostman J, Engfeldt P. Adrenoceptor occupancy in isolated human fat cells and its relationship with lipolysis rate. Eur J Pharmacol. 1988 Jan 27;146(1):45-56
Heffernan M, Summers RJ, Thorburn A, Ogru E, Gianello R, Jiang WJ, Ng FM. The effects of human GH and its lipolytic fragment (AOD9604) on lipid metabolism following chronic treatment in obese mice and beta(3)-AR knock-out mice. Endocrinology. 2001 Dec;142(12):5182-9.
Hansen TK, Gravholt CH, ØRskov H, Rasmussen MH, Christiansen JS, Jørgensen JO. Dose dependency of the pharmacokinetics and acute lipolytic actions of growth hormone. J Clin Endocrinol Metab. 2002 Oct;87(10):4691-8
Hartman ML, Faria AC, Vance ML, Johnson ML, Thorner MO, Veldhuis JD. Temporal structure of in vivo growth hormone secretory events in humans. Am J Physiol. 1991 Jan;260
Haffner D, Schaefer F, Girard J, Ritz E, Mehls O 1994 Metabolic clearance of recombinant human growth hormone in health and chronic renal failure. J Clin Invest 93:1163–1171
Fain JN. Effect of dibutyryl-3′,5′-AMP, theophylline and norepinephrine on lipolytic action of growth hormone and glucocorticoid in white fat cells. Endocrinology. 1968 Apr;82(4):825-30.
Goodman HM. Effects of growth hormone on the lipolytic response of adipose tissue to theophylline. Endocrinology. 1968 May;82(5):1027-34.
Yip RG, Goodman HM. Growth hormone and dexamethasone stimulate lipolysis and activate adenylyl cyclase in rat adipocytes by selectively shifting Gi alpha2 to lower density membrane fractions. Endocrinology. 1999 Mar;140(3):1219-27.
Fain JN, Cheema P, Tichansky DS, Madan AK. Stimulation of human omental adipose tissue lipolysis by growth hormone plus dexamethasone. Mol Cell Endocrinol. 2008 Nov 25;295(1-2):101-5.
Goodman HM 1970 Permissive effects of hormones on lipolysis. Endocrinology 86:1064–1074
Davis E, Loiacono R, Summers RJ. The rush to adrenaline: drugs in sport acting on the β-adrenergic system. British Journal of Pharmacology. 2008;154(3):584-597
Greenblatt, D. J. and Abourjaily, P. N. (2016), Pharmacokinetics and Pharmacodynamics for Medical Students: A Proposed Course Outline. The Journal of Clinical Pharmacology, 56: 1180–1195.
Keller A, Wu Z, Kratzsch J, Keller E, Blum WF, Kniess A, Preiss R, Teichert J, Strasburger CJ, Bidlingmaier M. Pharmacokinetics and pharmacodynamics of GH:dependence on route and dosage of administration. Eur J Endocrinol. 2007 Jun;156(6):647-53
Laursen T. Clinical pharmacological aspects of growth hormone administration. Growth Horm IGF Res. 2004 Feb;14(1):16-44. Review.
Møller N, Pørksen N, Ovesen P, Alberti KG. Evidence for increased sensitivity of fuel mobilization to growth hormone during short-term fasting in humans. Horm Metab Res. 1993 Mar;25(3):175-9.
Moller L, Dalman L, Norrelund H, Billestrup N, Frystyk J, Moller N, Jorgensen JO. Impact of fasting on growth hormone signaling and action in muscle and fat. J Clin Endocrinol Metab. 2009 Mar;94(3):965-72
Jørgensen JO, Møller N, Lauritzen T, Alberti KG, Orskov H, Christiansen JS. Evening versus morning injections of growth hormone (GH) in GH-deficient patients: effects on 24-hour patterns of circulating hormones and metabolites. J Clin Endocrinol Metab. 1990 Jan;70(1):207-14.
Blouin K, Veilleux A, Luu-The V, Tchernof A. Androgen metabolism in adipose tissue: recent advances. Mol Cell Endocrinol. 2009 Mar 25;301(1-2):97-103
Xu X, De Pergola G, Björntorp P. The effects of androgens on the regulation of lipolysis in adipose precursor cells. Endocrinology. 1990 Feb;126(2):1229-34
De Pergola G. The adipose tissue metabolism: role of testosterone and dehydroepiandrosterone. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S59-63. Review
Viguerie N, Millet L, Avizou S, Vidal H, Larrouy D, Langin D. Regulation of human adipocyte gene expression by thyroid hormone. J Clin Endocrinol Metab. 2002 Feb;87(2):630-4
Rubio A, Raasmaja A, Silva JE. Thyroid hormone and norepinephrine signaling in brown adipose tissue. II: Differential effects of thyroid hormone on beta 3-adrenergic receptors in brown and white adipose tissue. Endocrinology. 1995 Aug;136(8):3277-84.
Ghosh M, Das S. Increased beta(2)-adrenergic receptor activity by thyroid hormone possibly leads to differentiation and maturation of astrocytes in culture. Cell Mol Neurobiol. 2007 Dec;27(8):1007-21. Epub 2007 Sep 8
Pucci E, Chiovato L, Pinchera A. Thyroid and lipid metabolism. Int J Obes Relat Metab Disord. 2000 Jun;24 Suppl 2:S109-12. Review
Kim HK, Della-Fera MA, Hausman DB, Baile CA. Effect of clenbuterol on apoptosis, adipogenesis, and lipolysis in adipocytes. J Physiol Biochem. 2010 Sep;66(3):197-203
Taaffe DR, Thompson JL, Butterfield GE, Hoffman AR, Marcus R. Recombinant human growth hormone, but not insulin-like growth factor-I, enhances central fat loss in postmenopausal women undergoing a diet and exercise program. Horm Metab Res. 2001 Mar;33(3):156-62.
Il Dinitrofenolo (DNP) ha un parente chimico che, come il DNP, è stato commercializzato negli anni Trenta come aiuto legale per il dimagrimento. Il Dinitrocresolo è stato osservato essere cinque volte più potente del DNP, come riportato da Eric Colman della FDA in un articolo sulla sua storia pubblicato su Regulatory Toxicology and Pharmacology. (1)
Era diventato evidente all’inizio del XX secolo che il DNP accelerasse il dimagrimento, quando i ricercatori hanno analizzato le morti misteriose avvenute tra i lavoratori delle fabbriche di dinamite durante la Grande Guerra che era deceduti a causa di una febbre mortale. La causa di ciò era il composto DNP, che era stato rilasciato durante il processo produttivo. I ricercatori scoprirono che il DNP aveva aumentato il tasso metabolico, portando ad una estrema perdita di peso.
Questa scoperta ispirò il medico americano Maurice Tainter, negli anni ’30, a svolgere esperimenti sulle persone in sovrappeso. Nei test il DNP aveva aumentato il metabolismo dei soggetti fino al 50%, come si legge nell’articolo di revisione “Use of Dinitrophenol in Nutritional Disorders: A Critical Survey of Clinical Results”, pubblicato nel 1934 da Tainter. (2) Nello studio di Tainter i soggetti in sovrappeso avevano perso fino a 1,5kg di grasso a settimana con dosi giornaliere di circa 350 mg di DNP. Il DNP si è rivelato un aiuto più efficace per il dimagrimento rispetto alla Tiroxina (T4), che i medici avevano sperimentato in quel momento. Il DNP ha aumentato la produzione di calore del corpo del 1200%.
Il DNP inattiva l’enzima che immagazzina l’energia, che viene generata nella cellula dagli acidi grassi e dal glucosio, in ATP. Invece l’energia viene dissipata sotto forma di calore.
Tainter pubblicò i risultati della sua ricerca nel 1933. Il 7 febbraio 1934 Isabella Laboratories, di proprietà dell’inserzionista Harry Goroy, produsse il primo lotto di integratori che contenevano come principio attivo il DNP. Il prodotto venne chiamato Formula 281 e una capsula conteneva 100 mg di DNP. “Infine un aiuto dimagrante che ti darà una figura che gli uomini amano e della quale le donne saranno gelose. Nessun pericolo per la tua salute! Non c’è bisogno di cambiare il tuo stile di vita!”, Affermavano gli annunci pubblicitari dell’epoca.
Il DNP prese piede. Alcuni degli aiuti dimagranti a base di DNP sono riportati qui sopra. Nello stesso anno Tainter stimò che centomila americani avevano già utilizzato il DNP entro pochi mesi dal suo lancio. I prodotti contenenti il composto “miracoloso” apparvero in tutto il mondo.
Dinitrocresolo
Un anno dopo, nel 1935, Goroy decise che era giunto il momento di lanciare un prodotto ancora più potente: Improved Formula 281. Una capsula conteneva 35 mg di Dinitrocresolo.
Il Dinitrocresolo era risultato essere cinque volte più potente del DNP. Proprio prima che la Isabella Laboratories iniziasse l’aggiunta della sostanza ai suoi integratori, un articolo apparve sul The Lancet in cui i ricercatori britannici dimostrarono che il Dinitrocresolo era uno stimolatore metabolico ancora più efficace del DNP. Secondo gli inglesi, una dose giornaliera di 240mg aumentava il metabolismo del 70% in pochi giorni. (3)
“Si brucia letteralmente il grasso”, dichiarava un annuncio pubblicitario. “Assolutamente senza effetti collaterali”. A parte un rash cutaneo, o il cambiamento di colore della pelle o degli occhi. Ma se si verificava uno di questi effetti tutto quello che si doveva fare era bere un bicchiere d’acqua con bicarbonato di sodio e voila! Problema risolto.
Il Dinitrocresolo ebbe successo analogo al DNP. Nel 1935 la Isabella Labs vendette 550.000 capsule. Cominciarono a comparire prodotti contenenti entrambe le sostanze come principi attivi.
Nel frattempo stava divenendo chiaro che il DNP e il Dinitrocresolo non erano effettivamente composti sicuri. Vi furono un paio di morti. I sostenitori del DNP pensavano che i decessi dovessero essere ricollegati a casi di overdose, dosi eccessive assunte da utilizzatori testardi e impazienti che non seguivano le istruzioni di dosaggio. Warner Horner, un ottico, sconvolse queste speranze quando scoprì che gli utilizzatori che erano diventati ciechi dopo aver assunto il DNP, l’avevano fatto sotto la guida di un medico. Horner ha stimato che nel 1936 ben 2500 americani erano diventati ciechi in seguito all’assunzione di DNP. (4)
La FDA contattò la Isabella Laboratories nel 1936 chiedendo alla società di interrompere la produzione di DNP. Legalmente, la Formula 281 non era un medicinale. Era un supplemento per combattere l’obesità e, secondo i medici dell’epoca, l’obesità non era una malattia. La FDA ha avuto poteri per regolamentare le vendite dei farmaci, ma non le vendite di integratori. Fu solo nel 1937, da quando la Isabella Laboratories aveva fatto i suoi reclami pubblicitari così stravaganti che la Formula 281 poteva essere legalmente considerata una medicina, e la FDA era in grado di fare qualcosa a riguardo. La Formula 281 e la Improved Formula 281 sono scomparsi dal mercato e nel 1938 Harry Goroy pagò una multa di 50.000 dollari.
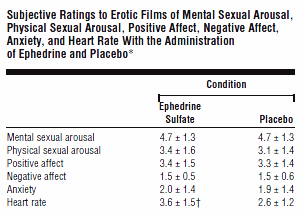
Se le donne assumono una dose di 50mg di Efedrina, il loro corpo reagisce maggiormente agli stimoli sessuali dopo tre quarti d’ora. I sessuologi dell’Università di Washington hanno scoperto ciò nel 1998. (1)
L’Efedrina è un alcaloide stimolante contenuto nella pianta Ephedra sinica. Il composto è stato scoperto nel 1885 dal farmacologo giapponese Nagai Nagayoshi mentre stava studiando piante medicinali cinesi e giapponesi. Negli anni ’90 l’Efedrina divenne un ingrediente popolare nei supplementi per la perdita di peso quando il guru del doping Dan Duchaine mescolava l’Efedrina con Aspirina e Caffeina nel suo Ultimate Orange.
Successivamente decine, se non centinaia di aziende fondarono i loro stack dimagranti sul progetto di Duchaine, anche se la maggior parte di loro non usava analoghi sintetici dell’Efedrina, ma estratti della pianta Ephedra. Ciò andò avanti fino a quando gli stack diventarono così forti che alcuni utilizzatori abusandone andarono in contro a importanti effetti avversi, così che il governo americano bandì l’uso degli estratti di Ephedra negli integratori. I governi europei seguirono.
Nello studio qui riportato i ricercatori hanno esaminato l’Efedrina solfato perché volevano conoscere l’effetto degli ormoni stimolanti come l’Adrenalina e Noradrenalina sulla sessualità femminile. I farmaci che bloccano gli effetti di questi ormoni, come i beta-bloccanti, riducono l’interesse sessuale delle donne. Pertanto, hanno motivato i ricercatori, una sostanza come l’Efedrina solfato dovrebbe aumentare l’interesse sessuale. Dopo tutto, l’Efedrina mima l’effetto dell’Adrenalina.
I ricercatori hanno somministrato a venti donne di età compresa tra i 19 ei 44 anni una dose di 50mg di Efedrina solfato. Tre quarti d’ora dopo l’assunzione i soggetti dello studio hanno visionato un film pornografico che mostrava “una coppia eterosessuale impegnarsi in preliminari, nel rapporto sessuale, e nel sesso orale”. ” In un’altra sessione le donne hanno visionato un film simile, ma gli era stato somministrato un placebo.
L’Efedrina non ha avuto alcun effetto sull’eccitazione mentale delle donne, anche se hanno riferito di sentire che il loro battito cardiaco era aumentato. Vedere la tabella seguente.
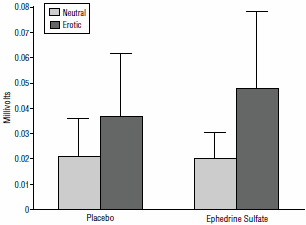
L’Efedrina ha avuto un effetto sulla circolazione sanguinea attraverso la vagina. Utilizzando un fotopletismografo i ricercatori sono stati in grado di osservare che l’Efedrina aumentava l’afflusso di sangue alla vagina nelle donne mentre guardavano il film porno. La figura seguente mostra questo. Neutral = durante la visione di immagini non erotiche; erotico = guardando il porno.
I ricercatori non erano del tutto sicuri di come interpretare i loro risultati. Secondo quanto riportato da loro, è possibile che l’Efedrina aumenti la risposta sessuale fisiologica fornendo un impulso ai già crescenti livelli di eccitazione sessuale. Se l’Efedrina possa fornire o meno un ‘imput iniziale’ alle donne che presentano difficoltà nell’eccitamento sessuale ha bisogno di ulteriori indagini.
Durante lo studio le donne non hanno avuto problemi con gli effetti collaterali. Tuttavia, questi possono sorgere se l’Efedrina viene utilizzata per un periodo di tempo più lungo o a dosi più alte. Dosi più elevate di Efedrina sono state correlate alla comparsa di una serie di effetti collaterali, ad esempio ansia, irrequietezza, insonnia, vertigini, mal di testa, e ipertensione.
I Bodybuilder che stanno pensando di utilizzare inibitori della Miostatina nel prossimo futuro non solo vedranno aumentate la loro massa muscolare, ma probabilmente diventeranno più definiti. Secondo i ricercatori del Diabetes and Obesity Center of Excellence americano, inibendo l’azione della Miostatina aumenta la sensibilità del corpo all’ormone Leptina. (1)
La Miostatina è una proteina con cui le cellule muscolari inibiscono la propria crescita. La Miostatina ha certamente altre funzioni importanti, ma in organismi adulti, per quanto ne sappiamo, l’inibizione dell’effetto della Miostatina non ha effetti collaterali notevoli. A parte un drammatico aumento della massa muscolare. Ed è per questo che i Bodybuilder stanno seguendo i risultati degli esperimenti con inibitori della Miostatina con interesse.
Tuttavia, gli esperimenti non vengono svolti pensando agli atleti. I ricercatori sperano che gli inibitori della Miostatina potranno aiutare a combattere la debolezza legata all’età e alle malattie muscolari degenerative. Nel caso dello studio che i ricercatori del Diabetes and Obesity Center of Excellence hanno pubblicato sul AJP – Endocrinology and Metabolism, gli autori sperano che il controllo della Miostatina possa essere utile anche per combattere l’obesità.
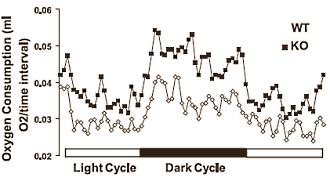
I ricercatori hanno svolto esperimenti con topi nei quali il gene della Miostatina era stato disattivato [KO] e con topi normali. Secondo la tabella sottostante, i topi ai quali era stato deattivato il gene della Miostatina non solo erano diventati più muscolosi, ma avevano anche una percentuale di grasso anormalmente bassa.
A causa della loro massa muscolare aumentata, i topi ai quali era stato deattivato il gene della Miostatina consumavano più energia. Si può vedere ciò nelle figure per il loro consumo di ossigeno. Ciò che è strano è che disattivare il gene della Miostatina non porta ad un aumento dell’assunzione di cibo. Ci si aspetterebbe che questo accada, ma non avviene. I topi con il gene della Miostatina inattivo mangiano la stessa quantità dei topi normali.
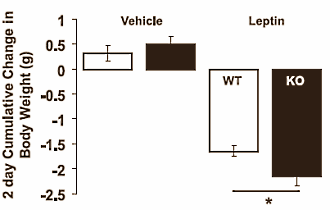
Quando i ricercatori hanno iniettato la Leptina ai topi, notarono che i topi senza Miostatina perdevano più peso rispetto ai topi normali. La Leptina è un ormone che le cellule adipose producono se aumentano di volume. Questo sopprime l’appetito e aumenta il metabolismo. I ricercatori concludono che la disattivazione della Miostatina aumenta la sensibilità alla Leptina e di conseguenza riduce l’appetito.
I ricercatori sospettano che gli inibitori della Miostatina possono essere quindi interessanti candidati per la medicina anti obesità, anche se sono attenti nella loro formulazione. Questi risultati suggeriscono che la Miostatina segnali direttamente o indirettamente attraverso l’aumento della massa muscolare scheletrica effetti di vasta portata sull’omeostasi energetica aumentando la possibilità che un fattore sconosciuto associato ai muscoli indotto dalla carenza di Miostatina possa conferire un beneficio metabolico.
I ricercatori della Amylin Pharmaceuticals hanno svolto degli esperimenti con una combinazione di Metionillettina [Metreleptina] e di una versione modificata dell’ormone Amilina. Le persone in sovrappeso possono perdere fino a mezzo chilogrammo alla settimana con una combinazione di queste due sostanze. (1)
Quando il pancreas secerne insulina, le cellule beta sintetizzano anche l’ormone Amilina. L’Amilina fa si che lo stomaco digerisca il cibo più lentamente, e invia al cervello un segnale di sazietà. Un perfetto aiuto dimagrante, pensarono alcuni farmacologi intelligenti – in modo tale che venne istituita la Amylin Pharmaceuticals. L’azienda ha lanciato una versione dell’amilina che differisce dalla versione naturale da un paio di aminoacidi e, di conseguenza,il suo tempo di attività aumenta. L’analogo è chiamato Pramlintide, ma la confezione contenente la sostanza è denominata Symlin.
Symlin agisce, secondo prove effettuate dalla Amilyn Pharmaceuticals, (2)(3) Le iniezioni di questo ormone aumentano l’efficacia di una dieta per la perdita di peso, anche se i risultati non sono sconvolgenti.
Un problema nel quale incappano molte persone in sovrappeso è l’aumento dell’appetito mano a mano che la dieta prosegue. Ciò è dovuto al fatto che le cellule adipose sovra-alimentate sintetizzano livelli elevati dell’ormone soppressore dell’appetito Leptina. La riduzione della Leptina sembra essere ciò che più ostacola i dietisti.
Anche se gli studi con la Leptina svolti alla fine degli anni novanta non hanno fornito risultati convincenti, i ricercatori hanno deciso di dare un’altra possibilità a questo composto. Hanno combinato la metreleptina, versione sintetica della leptina, con pramlintide.
I ricercatori hanno reclutato 177 soggetti in sovrappeso, ma non obesi, ai quali per prima cosa è stata data una dieta con il 40% di calorie in meno rispetto al basale per 4 settimane. I ricercatori hanno combinato questo con 2 iniezioni da 180 e poi 360 microgrammi di pramlintide al giorno.
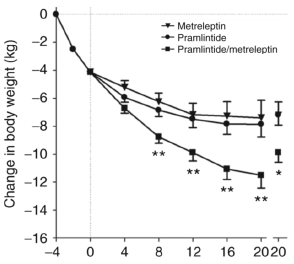
Poi è iniziato il vero percorso, che è durato 20 settimane. I ricercatori hanno consigliato ai soggetti in esame di seguire una dieta contenente il 20% di calorie in meno di quanto consumassero e hanno diviso i soggetti in tre gruppi. Ad un gruppo sono stati somministrate due iniezioni al giorno contenenti 5 mg di metreleptina, ad un altro gruppo è stato somministrato due iniezioni giornaliere di pramlintide da 360 microgrammi e ad un altro gruppo è stato somministrato entrambi i protocolli.
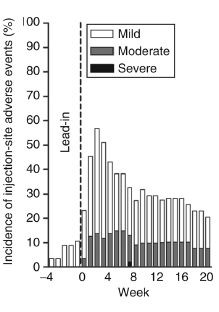
L’effetto collaterale peggiore sperimentato dai soggetti dello studio è stata l’irritazione nel punto di iniezioni. La figura seguente mostra i risultati del gruppo di “combinazione”. Le iniezioni di metreleptina probabilmente irritano più delle iniezioni di pramlintide.
Un altro effetto collaterale era la nausea, ma questo era temporaneo.
I ricercatori scrivono che il loro trattamento è promettente. “I risultati di questo studio forniscono una dimostrazione clinica iniziale di un approccio neurochormonale integrato alla farmacoterapia dell’obesità e garantisce un ulteriore sviluppo della pramlintide / metreleptina come regime per la perdita di peso”.
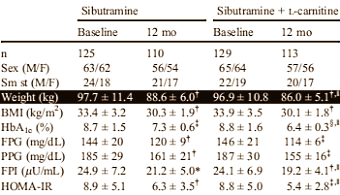
Come ben sappiamo la Sibutramina è fuori commercio, ma, nonostante ciò, questo coadiuvante per la perdita di perso, un distante parente dell’anfetamina, è ancora disponibile sul mercato nero. I ricercatori dell’Università di Pavia hanno riportato su Metabolism che la Sibutramina funziona meglio quando combinata con la L-Carnitina. (1)
Della Sibutramina ne ho già parlato in passato, ne ho riportato gli effetti sul aumento del metabolismo e sulla soppressione dell’appetito. La Sibutramina è un composto discretamente efficace, ma è stata vietata dopo che degli studi hanno indicato che il suo utilizzo poteva portare allo sviluppo di malattie cardiovascolari. Quando i ricercatori italiani hanno svolto il loro esperimento, nel quale hanno somministrato a 254 uomini e donne diabetici di tipo II una dose di 10mg/die di Sibutramina per un anno, la Sibutramina era ancora una sostanza legale.
Poiché la L-Carnitina ha effetti benefici teorici per i malati di diabete, i ricercatori hanno voluto verificare gli effetti di una combinazione di Sibutramina e Carnitina. Così hanno somministrato a metà dei soggetti presi in esame solo Sibutramina, e all’altra metà hanno aggiunto 2g di L-Carnitina al giorno.
Nel corso dell’anno il gruppo Sibutramina aveva perso 9,1 kg; Il gruppo Carnitina-Sibutramina aveva perso 10,9 kg. Il gruppo Carnitina-Sibutramina non solo aveva perso più peso, ma sembrava anche aver migliorato la sensibilità all’insulina.
Vedi immagine seguente: HbA1c = emoglobina glicata, un marker per il diabete; FPG = glucosio plasmatico a digiuno; PPG = glucosio plasmatico postprandiale; HOMA-IR = valutazione del modello dell’omeostasi dell’insulina plasmatica a digiuno dell’indice di insulino-resistenza.
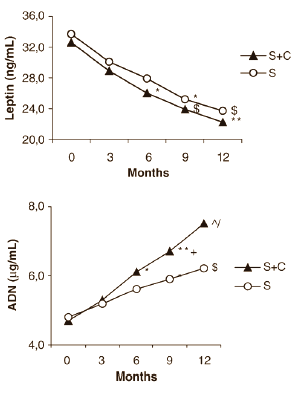
L’integrazione con L-Carnitina ha portato ad una maggiore produzione dell’ormone adiponectina e ha ridotto la produzione del fattore infiammatorio TNF-alpha. Ciò suggerisce che la L-Carnitina ha ridotto le reazioni infiammatorie provocate dal diabete di tipo II.
Non sono registrati effetti collaterali nel gruppo al quale era stata somministrata L-Carnitina.
Nota: la ricerca non è stata finanziata da un produttore di L-Carnitina, ma dall’università nella quale è stato svolto lo studio.
I mirtilli, le ciliegie e le uve contengono fenoli noti ai chimici come gli antociani. Secondo i nutrizionisti del dipartimento americano dell’agricoltura, questi composti hanno un effetto positivo sulla ricomposizione corporea. Secondo i ricercatori, riducono la quantità di massa grassa e aumentano la massa magra. (1)
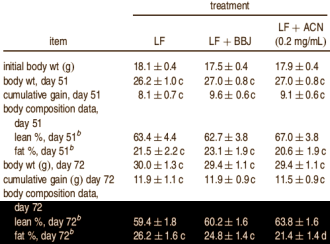
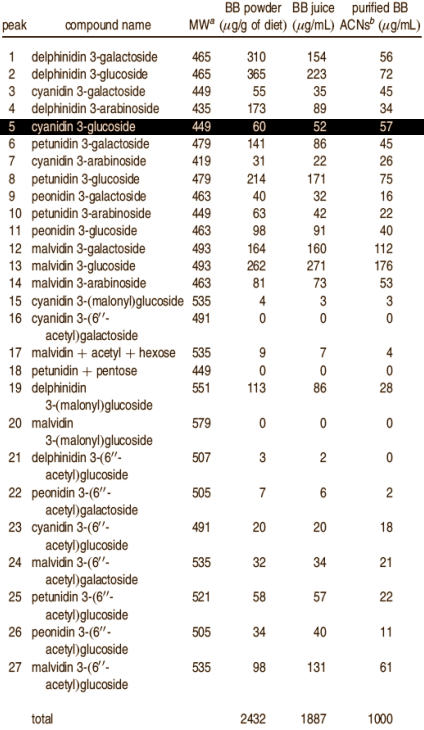
I ricercatori hanno svolto un esperimento su topi della durata di 72 giorni. Gli animali sono stati alimentati con una dieta a bassa percentuale di grassi (LF) o ad alto contenuto di grassi (HF). Ad alcuni degli animali è stato somministrato un supplemento di succo di mirtillo (BBJ), ad altri sono stati somministrati antociani puri estratti dai mirtilli e disciolti nell’acqua potabile (ACN). Al gruppo “ACN” sono stati somministrati 0,2mg di antociani per ml di acqua potabile.
I ricercatori volevano constatare se tali sostanze avessero effetti “anti obesità”, e osservarono questo effetto in entrambi i gruppi. Le figura seguenti mostrano che l’integrazione con antociani in particolare ha avuto un effetto qualitativo sul guadagno di peso dei topi. L’estratto di mirtillo ha aiutato i topi a guadagnare più massa magra e meno grasso.
I ricercatori sono stati in grado di studiare e comprendere le dinamiche d’azione delle sostanze contenute nel mirtillo esaminando il sangue dei topi. Gli antociani contenuti nelle bacche hanno migliorato l’attività dell’Insulina. I topi trattati necessitavano di meno Insulina e le loro cellule adipose producevano meno Leptina (quest’ultimo è un effetto non proprio positivo metabolicamente parlando). Le cellule adipose producono Leptina quando aumentano di volume. Sembra che gli antociani migliorino l’effetto dell’insulina nelle cellule muscolari piuttosto che nelle cellule adipose. [Ci si chiede se gli antociani dovrebbero essere considerati come degli AMPK-boosters]
Gli antociani assicurano un migliore assorbimento del glucosio ematico da parte delle cellule muscolari, impedendo così la crescita delle cellule adipose. Ecco perché l’integrazione ha avuto maggiore effetto nel gruppo sottoposto a una dieta con una bassa percentuale di grassi. L’effetto è stato ancora più evidente quando i ricercatori hanno aggiunto 1 mg di antociani invece di 0,2 mg in un millilitro di acqua potabile.
Sì, i risultati riportati sono corretti. Nel gruppo dei topi sottoposti ad una dieta ad alto contenuto di grassi la dose più elevata di antociani non ha avuto alcun effetto sulla percentuale di grasso corporeo, ma nel gruppo sottoposto ad una dieta a basso contenuto di grassi la percentuale di grasso corporeo si è dimezzata.
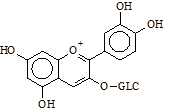
La composizione degli antociani che i ricercatori hanno utilizzato è mostrata nella figura seguente . La cianidina-3-glucoside, il composto 5 evidenziato nella figura seguente, è il principio attivo contenuto nel Indigo-3G, integratore alimentare prodotto dalla Biotest. (2)