
Introduzione:
Esiste una controversia sulla quantità massima di proteine che può essere utilizzata in un singolo pasto per la costruzione del tessuto magro da parte di coloro che sono impegnati in un allenamento contro-resistenza regolare. È stato proposto che la sintesi proteica muscolare sia massimizzata nei giovani adulti con un apporto di circa 20-25g di proteine di alta qualità; si ritiene che tutto ciò che supera questa quantità venga ossidato a scopo energetico o transaminato per formare urea e altri acidi organici. Tuttavia, questi risultati sono specifici per l’assunzione di proteine a rapida digestione senza l’aggiunta di altri macronutrienti in fisiologia. Il consumo di fonti proteiche ad azione più lenta, in particolare se consumate in combinazione con altri macronutrienti, ritarderebbe l’assorbimento e quindi potrebbe migliorare l’utilizzo degli aminoacidi che le compongono.
Un’idea sbagliata da tempo diffusa tra i non addetti ai lavori è che esista un limite alla quantità di proteine che può essere assorbita dall’organismo. Da un punto di vista nutrizionale, il termine “assorbimento” descrive il passaggio dei nutrienti dall’intestino alla circolazione sistemica. In base a questa definizione, la quantità di proteine che può essere assorbita è virtualmente illimitata. Dopo la digestione di una fonte proteica, gli aminoacidi (AA) che la compongono vengono trasportati attraverso gli enterociti della parete intestinale, entrano nella circolazione portale epatica e gli AA che non vengono utilizzati direttamente dal fegato entrano nel flusso sanguigno, dopo di che quasi tutti gli AA ingeriti diventano disponibili per l’utilizzo da parte dei tessuti. Mentre l’assorbimento non è un fattore limitante rispetto alle proteine intere, il consumo di singoli AA in forma libera può presentare problemi a questo proposito. In particolare, è stata dimostrata la potenziale competizione a livello della parete intestinale, con gli AA presenti in maggiore concentrazione assorbiti a scapito di quelli meno concentrati [1].
Come già accennato, è stato proposto che la sintesi proteica muscolare (MPS) sia massimizzata nei giovani adulti con un apporto di circa 20-25g di proteine di alta qualità, in linea con il concetto di “muscolo pieno”; si ritiene che tutto ciò che supera questa quantità venga ossidato a scopo energetico o transaminato per formare composti corporei alternativi [2].
Brad Schoenfeld e Alan Aragon hanno pubblicato una loro ricerca su questo argomento sul Journal of the International Society of Sports Nutrition volume 15, del 27 febbraio 2018. Lo scopo del loro articolo era duplice: 1) rivedere oggettivamente la letteratura nel tentativo di determinare una soglia anabolica superiore per l’assunzione di proteine per pasto; 2) trarre conclusioni rilevanti sulla base dei dati attuali, in modo da delucidare le linee guida per la distribuzione giornaliera di proteine per pasto, al fine di ottimizzare l’accrescimento del tessuto magro.
Grazie al loro eccellente lavoro andiamo ad analizzare la questione nel dettaglio al fine di poter conoscere il reale stato delle cose.
Impatto della velocità di digestione/assorbimento sull’anabolismo muscolare:
In uno studio spesso citato a sostegno dell’ipotesi che la MPS sia massimizzata con una dose proteica di ~ 20-25g, Areta et al. [3] hanno fornito quantità diverse di proteine a soggetti allenati contro-resistenza durante un periodo di recupero di 12 ore dopo l’esecuzione di un protocollo di esercizi di leg-extension a ripetizioni moderate e multi-set. Un totale di 80g di proteine del siero del latte è stato ingerito in una delle tre condizioni seguenti: 8 porzioni da 10g ogni 1,5 ore; 4 porzioni da 20g ogni 3 ore; o 2 porzioni da 40g ogni 6 ore. I risultati hanno mostrato che la MPS era maggiore in coloro che consumavano 4 porzioni da 20g di proteine, il che non suggerisce alcun beneficio aggiuntivo, e in realtà un aumento minore della MPS quando si consumava il dosaggio più alto (40 g) nelle condizioni imposte nello studio. Questi risultati ampliano quelli analoghi di Moore et al. [4] sul turnover dell’azoto nell’intero corpo.

Sebbene i risultati di Areta et al. [3] forniscano una visione interessante degli effetti dose-correlati dell’assunzione di proteine sullo sviluppo muscolare, è importante notare che una serie di fattori influenzano il metabolismo delle proteine alimentari, tra cui la composizione della fonte proteica, la composizione del pasto, la quantità di proteine ingerite e le specificità della routine di allenamento [5]. Inoltre, anche variabili individuali come l’età, lo stato di allenamento e la quantità di massa magra hanno un impatto sui risultati della costruzione muscolare. Un limite importante dello studio di Areta et al. [3] è che l’assunzione totale di proteine durante il periodo di studio di 12 ore è stata di soli 80g, corrispondenti a meno di 1g/kg di massa corporea. Questa quantità è di gran lunga inferiore a quella necessaria per massimizzare l’equilibrio proteico muscolare negli individui allenati contro-resistenza che hanno partecipato allo studio [6, 7]. Inoltre, la validità ecologica di questo lavoro è limitata, dal momento che l’assunzione abituale di proteine da parte di individui che puntano al guadagno o al mantenimento muscolare consumano abitualmente circa 2-4 volte questa quantità al giorno [8, 9].
Va inoltre notato che i soggetti di Areta et al. [3] hanno ingerito solo proteine del siero del latte per tutto il periodo post-esercizio. Il siero di latte è una proteina “ad azione rapida”; il suo tasso di assorbimento è stato stimato in ~ 10g all’ora [5]. A questo ritmo, basterebbero 2 ore per assorbire completamente una dose di 20g di siero di latte. Se da un lato la rapida disponibilità di AA tende a far aumentare la MPS, dall’altro una ricerca precedente che esaminava la cinetica delle proteine dell’intero organismo ha dimostrato che la concomitante ossidazione di alcuni degli AA può determinare un bilancio proteico netto inferiore rispetto a una fonte proteica che viene assorbita a un ritmo più lento [10]. Ad esempio, le proteine dell’uovo cotto hanno un tasso di assorbimento di circa 3g all’ora [5], il che significa che l’assorbimento completo di un’omelette contenente gli stessi 20g di proteine richiederebbe circa 7 ore, il che potrebbe contribuire ad attenuare l’ossidazione degli AA e quindi a promuovere un maggiore bilancio proteico netto positivo per tutto il corpo. Un’importante avvertenza è che questi risultati sono specifici per l’equilibrio proteico dell’intero corpo; la misura in cui ciò riflette l’equilibrio proteico del muscolo scheletrico rimane poco chiara.

Sebbene alcuni studi abbiano mostrato effetti simili delle proteine veloci e lente sul bilancio proteico muscolare netto [11] e sul tasso di sintesi frazionale [12,13,14], altri studi hanno dimostrato un maggiore effetto anabolico delle proteine del siero di latte rispetto alle fonti a più lenta digestione sia a riposo [15, 16], sia dopo l’esercizio contro-resistenza [16, 17]. Tuttavia, la maggior parte di questi risultati sono stati ottenuti durante periodi di prova più brevi (4 ore o meno), mentre periodi di prova più lunghi (5 ore o più) tendono a non mostrare differenze tra siero di latte e caseina sulla MPS o sul bilancio dell’azoto [18]. Inoltre, la maggior parte degli studi che hanno dimostrato un maggiore anabolismo con il siero di latte ha utilizzato una dose relativamente piccola di proteine (≤20g) [15,16,17]; rimane poco chiaro se dosi più elevate possano determinare una maggiore ossidazione delle fonti proteiche ad azione rapida rispetto a quelle ad azione lenta.
A completamento di questi risultati equivoci, una ricerca che ha esaminato il destino del siero di latte e della caseina intrinsecamente etichettati consumati all’interno del latte ha rilevato una maggiore incorporazione della caseina nel muscolo scheletrico [19]. Quest’ultimo risultato deve essere considerato con l’avvertenza che, sebbene si presuma che il turnover proteico nella gamba rifletta principalmente il muscolo scheletrico, è possibile che vi contribuiscano anche tessuti non muscolari. È interessante notare che la presenza o l’assenza di grassi del latte in concomitanza con la caseina micellare non ha ritardato il tasso di disponibilità di aminoacidi circolanti derivati dalle proteine o la sintesi proteica miofibrillare [20]. Inoltre, la coingestione di carboidrati con la caseina ha ritardato la digestione e l’assorbimento, ma non ha avuto alcun impatto sull’accrescimento delle proteine muscolari rispetto a una condizione di sole proteine [21]. L’implicazione è che il potenziale dei macronutrienti di accompagnamento di alterare i tassi di digestione non si traduce necessariamente in alterazioni dell’effetto anabolico dell’alimentazione proteica, almeno nel caso di proteine a lenta digestione come la caseina. Prima di trarre conclusioni definitive, è necessario effettuare ulteriori confronti sulla coingestione di grassi e/o carboidrati con altre proteine, profili di soggetti e relativa vicinanza all’allenamento.

Un “tetto anabolico” acuto più alto di quanto si pensasse?
Più recentemente, Macnaughton et al. [22] hanno utilizzato un disegno randomizzato, in doppio cieco, all’interno del soggetto, in cui uomini allenati contro-resistenza hanno partecipato a due prove separate da circa 2 settimane. Durante uno studio i soggetti hanno ricevuto 20g di proteine del siero del latte subito dopo aver eseguito un allenamento contro-resistenza totale del corpo; durante l’altro studio è stato istituito lo stesso protocollo, ma i soggetti hanno ricevuto un bolo di 40g di proteine del siero del latte dopo l’allenamento. I risultati hanno mostrato che il tasso di sintesi frazionale miofibrillare era superiore di circa il 20% in seguito al consumo di 40g rispetto alla condizione di 20g. I ricercatori hanno ipotizzato che la grande quantità di massa muscolare attivata dall’allenamento contro-resistenza total body necessitasse di una maggiore richiesta di AA, soddisfatta da un consumo maggiore di proteine esogene. Va notato che i risultati di McNaughton et al. [22] sono in qualche modo in contrasto con un precedente lavoro di Moore et al. che non mostrava differenze statisticamente significative nella MPS tra l’assunzione di una dose di 20g e 40g di siero di latte in giovani uomini dopo un allenamento di leg extension, sebbene la dose più alta producesse un aumento assoluto dell’11% maggiore [23]. Se le differenze tra assunzioni superiori a ~ 20g per alimentazione siano significative dal punto di vista pratico rimangono speculative e probabilmente dipendono dagli obiettivi dell’individuo.

Dato che lo sviluppo muscolare è una funzione dell’equilibrio dinamico tra MPS e degradazione delle proteine muscolari (MPB), entrambe le variabili devono essere considerate in qualsiasi discussione sul dosaggio delle proteine nella dieta. Kim et al. [24] hanno cercato di indagare questo argomento fornendo 40 o 70g di proteine di manzo consumate come parte di un pasto misto in due occasioni distinte separate da un periodo di washout di circa 1 settimana. I risultati hanno mostrato che l’assunzione di proteine più elevate ha promosso una risposta anabolica significativamente maggiore per l’intero corpo, attribuita principalmente a una maggiore attenuazione della ripartizione delle proteine. Dato che i partecipanti hanno consumato pasti abbondanti e misti come alimenti interi contenenti non solo proteine, ma anche carboidrati e grassi alimentari, è logico ipotizzare che ciò abbia ritardato la digestione e l’assorbimento degli AA rispetto al consumo liquido di fonti proteiche isolate. Questo, a sua volta, avrebbe causato un rilascio più lento di AA in circolazione e quindi potrebbe aver contribuito alle differenze dose-dipendenti nella risposta anabolica all’assunzione di proteine. Un limite notevole dello studio è che le misure dell’equilibrio proteico sono state effettuate a livello di tutto il corpo e quindi non specifiche per i muscoli. Si può quindi ipotizzare che una parte, se non la maggior parte, dei benefici anticatabolici associati a una maggiore assunzione di proteine provenga da tessuti diversi dal muscolo, probabilmente dall’intestino. Tuttavia, il turnover proteico nell’intestino potrebbe fornire una via attraverso la quale gli aminoacidi accumulati possono essere rilasciati nella circolazione sistemica per essere utilizzati per le MPS, aumentando plausibilmente il potenziale anabolico [25]. Questa ipotesi rimane speculativa e merita ulteriori indagini. Si potrebbe essere tentati di attribuire queste marcate riduzioni della proteolisi a risposte insuliniche più elevate, considerando l’inclusione di una generosa quantità di carboidrati nei pasti consumati. Sebbene l’Insulina sia spesso considerata un ormone anabolico, il suo ruolo primario nell’equilibrio proteico muscolare è legato agli effetti anticatabolici [26]. Tuttavia, in presenza di AA plasmatici elevati, l’effetto dell’aumento dell’Insulina sul bilancio proteico muscolare netto si stabilizza entro un intervallo modesto di 15-30 mU/L [27, 28]. Dato che è stato dimostrato che una dose di 45g di proteine del siero del latte provoca un aumento dell’Insulina a livelli sufficienti per massimizzare il bilancio proteico muscolare netto [29], sembrerebbe che i macronutrienti aggiuntivi consumati nello studio di Kim et al. [24] abbiano avuto scarsa influenza sui risultati.

Risultati longitudinali:
Sebbene gli studi discussi in precedenza offrano indicazioni sulla quantità di proteine che l’organismo può utilizzare in una determinata alimentazione, le risposte anaboliche acute non sono necessariamente associate a incrementi muscolari a lungo termine [30]. La risposta a questo argomento può essere data solo valutando i risultati di studi longitudinali che misurano direttamente le variazioni della massa magra con la somministrazione di dosaggi proteici diversi, nonché di proteine con velocità di digestione/assorbimento diverse.
Wilborn et al. [31] non hanno riscontrato differenze nell’aumento della massa magra dopo 8 settimane di integrazione pre e post esercizio contro-resistenza con siero di latte o caseina. Analogamente, Fabre et al. [32] non hanno riscontrato differenze tra i gruppi nell’aumento della massa magra, confrontando i seguenti rapporti di proteine del siero di latte/caseina consumate dopo l’esercizio: 100/0, 50/50, 20/80.
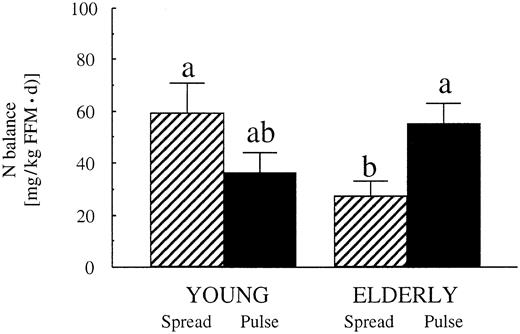
In uno studio di 14 giorni condotto su donne anziane, Arnal et al. [33] hanno dimostrato che l’apporto della maggior parte delle proteine giornaliere (79%) in un unico pasto (schema a impulsi) ha determinato una maggiore ritenzione di massa grassa rispetto a un apporto uniformemente distribuito in quattro pasti giornalieri (schema a spalmi). Uno studio successivo condotto dallo stesso laboratorio su giovani donne ha riportato effetti simili tra l’assunzione di proteine a impulsi e quella distribuita [34]. I risultati combinati di questi studi indicano che la massa muscolare non è influenzata negativamente dal consumo della maggior parte delle proteine giornaliere in un bolo abbondante. Tuttavia, nessuno dei due studi ha utilizzato un allenamento contro-resistenza regolato, limitando così la generalizzabilità a individui coinvolti in programmi di esercizio intenso.
Anche gli studi sul digiuno intermittente (IF) possono fornire indicazioni sugli effetti del dosaggio proteico. I protocolli tipici di IF richiedono l’assunzione di nutrienti giornalieri, comprese le proteine, in un arco di tempo ristretto – di solito meno di 8 ore – seguito da un digiuno prolungato. Una recente review sistematica ha concluso che l’IF ha effetti simili sulla massa grassa rispetto ai protocolli di alimentazione continua [35]. Tuttavia, gli studi esaminati nell’analisi riguardavano in genere assunzioni di proteine non ottimali consumate nell’ambito di una dieta a basso contenuto energetico senza una componente di allenamento contro-resistenza, limitando ancora una volta la possibilità di estrapolare i risultati da individui che svolgono tale tipo di routine allenante.
A colmare questa lacuna della letteratura contribuisce uno studio di 8 settimane condotto da Tinsley et al. [36], che ha messo a confronto un protocollo di alimentazione limitata nel tempo (TRF) con cicli di 20 ore di digiuno e 4 ore di alimentazione per 4 giorni alla settimana, con un gruppo a dieta normale (ND) in soggetti non allenati che praticavano allenamento di resistenza per 3 giorni alla settimana. Il gruppo TRF ha perso peso corporeo grazie a un minore apporto energetico (667 kcal in meno nei giorni di digiuno rispetto a quelli di non digiuno), ma non ha perso in modo significativo massa magra (0,2 kg); il gruppo ND ha guadagnato massa magra (2,3 kg), ma non in modo statisticamente significativo, anche se l’entità delle differenze fa pensare che questi risultati possano avere un significato pratico. L’aspetto forse più interessante è che l’area della sezione trasversale del bicipite brachiale e del retto femorale ha mostrato aumenti simili in entrambi i gruppi, nonostante i cicli di digiuno di 20 ore e i cicli di alimentazione concentrata nella TRF, suggerendo che l’utilizzo dell’apporto proteico nei cicli di alimentazione ad libitum di 4 ore non è stato ostacolato da un tetto acuto di anabolismo. Purtroppo, in questo studio le proteine e l’energia non sono state equiparate. Successivamente, uno studio di 8 settimane condotto da Moro et al. [37] su soggetti allenati contro-resistenza con un ciclo di 16 ore di digiuno/8 ore di TRF ha rilevato una perdita di grasso significativamente maggiore nei TRF rispetto agli ND (1,62 contro 0,31 kg), mentre la massa magra è rimasta invariata in entrambi i gruppi. Questi risultati mettono ulteriormente in discussione la preoccupazione di superare una certa soglia di assunzione di proteine per pasto per raggiungere l’obiettivo della ritenzione muscolare.
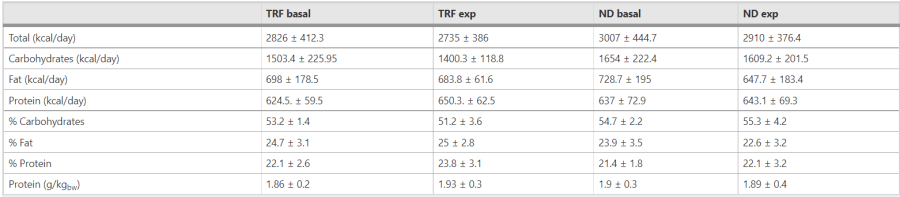
In contrasto con i risultati di cui sopra, che mostrano effetti da neutri a positivi dell’assunzione di un pasto concentrato nel tempo, Arciero et al. [38] hanno confrontato 3 diete: 2 diete ad alto contenuto proteico (35% dell’energia totale) composte da 3 (HP3) e 6 pasti al giorno (HP6), e un apporto proteico tradizionale (15% dell’energia totale) consumato in 3 pasti al giorno (TD3). Durante la fase eucalorica iniziale di 28 giorni, HP3 e HP6 hanno consumato rispettivamente 2,27 e 2,15g/kg di proteine, mentre TD3 ne ha consumate 0,9g/kg. Il HP6 è stato l’unico gruppo ad aver guadagnato significativamente massa magra. Durante la successiva fase eucalorica di 28 giorni, HP3 e HP6 hanno consumato proteine rispettivamente a 1,71 e 1,65g/kg, mentre TD3 ha consumato 0,75g/kg. Il HP6 ha mantenuto il suo aumento di massa magra, superando gli altri due trattamenti in questo senso (HP in realtà ha mostrato una perdita significativa di massa magra rispetto al controllo). La discrepanza tra questi ultimi risultati e quelli degli studi IF/TRF rimane da riconciliare. In ogni caso, è degno di nota il fatto che in questo filone manchino confronti specificamente orientati all’obiettivo del guadagno muscolare, in particolare confronti ipercalorici.

(a) significativamente diverso da CON, P < 0,05; (b) significativamente diverso da BAL, P < 0,05; (y) il grasso corporeo totale e addominale (Figura 3B e C) era significativamente più basso in BAL e NEG in HP6 rispetto a TD3, P < 0,05. I valori tra parentesi () indicano la variazione media delta per ciascun gruppo.[38]
Conclusioni:
È necessario fare un’importante distinzione tra le sfide acute con i pasti che confrontano diverse quantità di proteine (comprese le somministrazioni seriali nella fase acuta dopo l’allenamento di resistenza) e le somministrazioni croniche con i pasti che confrontano diverse distribuzioni di proteine durante il giorno, nel corso di diverse settimane o mesi. Gli studi longitudinali che esaminano la composizione corporea non hanno corroborato in modo coerente i risultati degli studi acuti che esaminano il flusso proteico muscolare. Quantificare la quantità massima di proteine per pasto che può essere utilizzata per l’anabolismo muscolare è stata una sfida a causa della moltitudine di variabili da indagare. Forse la sintesi più completa dei risultati in questo campo è stata fatta da Morton et al. [2], che hanno concluso che 0,4g/kg/pasto stimolerebbero in modo ottimale le MPS. Ciò si basava sull’aggiunta di due deviazioni standard alla loro scoperta che 0,25g/kg/pasto stimolano al massimo la MPS nei giovani uomini. In linea con questa ipotesi, Moore et al. [39] hanno sottolineato che i loro risultati sono mezzi stimati per massimizzare la MPS e che i massimali di dosaggio possono essere di ~ 0,60g/kg per alcuni uomini anziani e di ~ 0,40g/kg per alcuni uomini giovani. È importante notare che queste stime si basano sulla sola fornitura di una fonte proteica a rapida digestione, che potrebbe aumentare il potenziale di ossidazione degli AA se consumata in boli più grandi. Sembra logico che una fonte proteica ad azione più lenta, soprattutto se consumata in combinazione con altri macronutrienti, ritardi l’assorbimento e quindi migliori l’utilizzo degli AA costituenti. Tuttavia, le implicazioni pratiche di questo fenomeno rimangono speculative e discutibili [21].
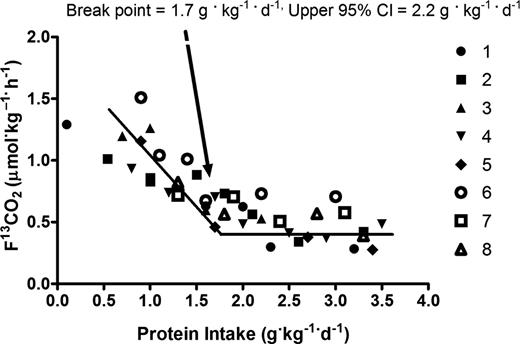
L’insieme delle prove indica che l’apporto proteico totale giornaliero per massimizzare l’aumento di massa e forza muscolare indotto dall’allenamento contro-resistenza è di circa 1,6g/kg, almeno in condizioni di non dieta (eucalorica o ipercalorica) [6]. Tuttavia, 1,6g/kg/giorno non deve essere considerato un limite ferreo o universale oltre il quale l’apporto proteico sarà sprecato o utilizzato per esigenze fisiologiche diverse dalla crescita muscolare. Una recente meta-analisi sull’integrazione proteica in soggetti che praticano allenamenti contro-resistenza ha riportato un intervallo di confidenza superiore del 95% (CI) di 2,2g/kg/giorno [6]. Anche Bandegan et al. [7] hanno mostrato un CI superiore di 2,2g/kg/die in una coorte di giovani bodybuilder maschi, sebbene il metodo di valutazione (tecnica di ossidazione degli aminoacidi con indicatore) utilizzato in questo studio non sia stato accettato universalmente per determinare il fabbisogno proteico ottimale. Ciò rafforza la necessità pratica di personalizzare la programmazione dietetica e di rimanere aperti al superamento delle medie stimate. È quindi una soluzione relativamente semplice ed elegante consumare proteine con un apporto target di 0,4g/kg/pasto in un minimo di quattro pasti per raggiungere un minimo di 1,6g/kg/giorno, se l’obiettivo primario è la costruzione di muscoli. Utilizzando l’assunzione giornaliera del CI superiore di 2,2g/kg/giorno negli stessi quattro pasti, sarebbe necessario un massimo di 0,55g/kg/pasto. Questa tattica applicherebbe ciò che è attualmente noto per massimizzare le risposte anaboliche acute e gli adattamenti anabolici cronici. Sebbene le ricerche dimostrino che il consumo di dosi maggiori di proteine (>20g) comporta una maggiore ossidazione degli AA [40], le prove indicano che questo non è il destino di tutti gli AA ingeriti in più, poiché alcuni vengono utilizzati per la costruzione dei tessuti. Sono comunque necessarie ulteriori ricerche per quantificare una soglia superiore specifica per l’assunzione di proteine per pasto.
In conclusione, possiamo indentificare tre punti chiave per la gestione proteica:
- Totale proteico giornaliero funzionale nell’atleta contro-resistenza da 1,5 a 3g/Kg/die, da calibrare a seconda dell’apporto glucidico e la risposta individuale;
- una media proteica per pasto tra 0,4 e 0,5g/Kg;
- evitare il consumo di proteine idrolizzate o aminoacidi liberi oltre i 10-15g [ovviamente in assunzione senza altro alimento o macronutriente] in stato fisiologico, discorso differente in contesto “doped” con protocollo GH/Insulina.
Gabriel Bellizzi [CEO BioGenTech]
Riferimento:
- Gropper SS, Smith JL, Groff JL: Advanced Nutrition and Human Metabolism. Belmont, CA: Wadsworth Cengage Learning; 2009.
- Morton RW, McGlory C, Phillips SM. Nutritional interventions to augment resistance training-induced skeletal muscle hypertrophy. Front Physiol. 2015;6:245.
- Areta JL, Burke LM, Ross ML, Camera DM, West DW, Broad EM, Jeacocke NA, Moore DR, Stellingwerff T, Phillips SM, Hawley JA, Coffey VG. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J Physiol. 2013;591(Pt 9):2319–31.
- Moore DR, Areta J, Coffey VG, Stellingwerff T, Phillips SM, Burke LM, Cleroux M, Godin JP, Hawley JA. Daytime pattern of post-exercise protein intake affects whole-body protein turnover in resistance-trained males. Nutr Metab (Lond). 2012;9(1):91. -7075-9-91
- Bilsborough S, Mann N. A review of issues of dietary protein intake in humans. Int J Sport Nutr Exerc Metab. 2006;16(2):129–52.
- Morton RW, Murphy KT, McKellar SR, Schoenfeld BJ, Henselmans M, Helms E, Aragon AA, Devries MC, Banfield L, Krieger JW, Phillips SM. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. Br J Sports Med. 2017;
- Bandegan A, Courtney-Martin G, Rafii M, Pencharz PB, Lemon PW. Indicator amino acid-derived estimate of dietary protein requirement for male bodybuilders on a nontraining day is several-fold greater than the current recommended dietary allowance. J Nutr. 2017;147(5):850–7.
- Spendlove J, Mitchell L, Gifford J, Hackett D, Slater G, Cobley S, O’Connor H. Dietary intake of competitive bodybuilders. Sports Med. 2015;45(7):1041–63.
- Antonio J, Ellerbroek A, Silver T, Vargas L, Peacock C: The effects of a high protein diet on indices of health and body composition–a crossover trial in resistance-trained men. J Int Soc Sports Nutr 2016, 13:3–016–0114-2. eCollection 2016.
- Dangin M, Boirie Y, Guillet C, Beaufrere B: Influence of the protein digestion rate on protein turnover in young and elderly subjects. J Nutr 2002, 132(10):3228S–33S.
- Tipton KD, Elliott TA, Cree MG, Wolf SE, Sanford AP, Wolfe RR. Ingestion of casein and whey proteins result in muscle anabolism after resistance exercise. Med Sci Sports Exerc. 2004;36(12):2073–81.
- Mitchell CJ, McGregor RA, D’Souza RF, Thorstensen EB, Markworth JF, Fanning AC, Poppitt SD, Cameron-Smith D. Consumption of milk protein or whey protein results in a similar increase in muscle protein synthesis in middle aged men. Nutrients. 2015;7(10):8685–99.
- Reitelseder S, Agergaard J, Doessing S, Helmark IC, Lund P, Kristensen NB, Frystyk J, Flyvbjerg A, Schjerling P, van Hall G, Kjaer M, Holm L. Whey and casein labeled with L-[1-13C]leucine and muscle protein synthesis: effect of resistance exercise and protein ingestion. Am J Physiol Endocrinol Metab. 2011;300(1):E231–42.
- Dideriksen KJ, Reitelseder S, Petersen SG, Hjort M, Helmark IC, Kjaer M, Holm L. Stimulation of muscle protein synthesis by whey and caseinate ingestion after resistance exercise in elderly individuals. Scand J Med Sci Sports. 2011;21(6):e372–83.
- Pennings B, Boirie Y, Senden JM, Gijsen AP, Kuipers H, van Loon LJ. Whey protein stimulates postprandial muscle protein accretion more effectively than do casein and casein hydrolysate in older men. Am J Clin Nutr. 2011;93(5):997–1005.
- Burd NA, Yang Y, Moore DR, Tang JE, Tarnopolsky MA, Phillips SM. Greater stimulation of myofibrillar protein synthesis with ingestion of whey protein isolate v. Micellar casein at rest and after resistance exercise in elderly men. Br J Nutr. 2012;108(6):958–62.
- Tang JE, Moore DR, Kujbida GW, Tarnopolsky MA, Phillips SM. Ingestion of whey hydrolysate, casein, or soy protein isolate: effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J Appl Physiol (1985). 2009;107(3):987–92.
- Witard OC, Wardle SL, Macnaughton LS, Hodgson AB, Tipton KD. Protein considerations for Optimising skeletal muscle mass in healthy young and older adults. Nutrients. 2016;8(4):181.
- Soop M, Nehra V, Henderson GC, Boirie Y, Ford GC, Nair KS. Coingestion of whey protein and casein in a mixed meal: demonstration of a more sustained anabolic effect of casein. Am J Physiol Endocrinol Metab. 2012;303(1):E152–62.
- Gorissen SHM, Burd NA, Kramer IF, van Kranenburg J, Gijsen AP, Rooyackers O, van Loon LJC. Co-ingesting milk fat with micellar casein does not affect postprandial protein handling in healthy older men. Clin Nutr. 2017;36(2):429–37.
- Gorissen SH, Burd NA, Hamer HM, Gijsen AP, Groen BB, van Loon LJ. Carbohydrate coingestion delays dietary protein digestion and absorption but does not modulate postprandial muscle protein accretion. J Clin Endocrinol Metab. 2014;99(6):2250–8.
- Macnaughton LS, Wardle SL, Witard OC, McGlory C, Hamilton DL, Jeromson S, Lawrence CE, Wallis GA, Tipton KD. The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Physiol Rep. 2016;4(15) https://doi.org/10.14814/phy2.12893.
- Moore DR, Robinson MJ, Fry JL, Tang JE, Glover EI, Wilkinson SB, Prior T, Tarnopolsky MA, Phillips SM. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am J Clin Nutr. 2009;89(1):161–8.
- Kim IY, Schutzler S, Schrader A, Spencer HJ, Azhar G, Ferrando AA, Wolfe RR. The anabolic response to a meal containing different amounts of protein is not limited by the maximal stimulation of protein synthesis in healthy young adults. Am J Physiol Endocrinol Metab. 2016;310(1):E73–80.
- Deutz NE, Wolfe RR. Is there a maximal anabolic response to protein intake with a meal? Clin Nutr. 2013;32(2):309–13.
- Abdulla H, Smith K, Atherton PJ, Idris I. Role of insulin in the regulation of human skeletal muscle protein synthesis and breakdown: a systematic review and meta-analysis. Diabetologia. 2016;59(1):44–55.
- Greenhaff PL, Karagounis LG, Peirce N, Simpson EJ, Hazell M, Layfield R, Wackerhage H, Smith K, Atherton P, Selby A, Rennie MJ. Disassociation between the effects of amino acids and insulin on signaling, ubiquitin ligases, and protein turnover in human muscle. Am J Physiol Endocrinol Metab. 2008;295(3):E595–604.
- Rennie MJ, Bohe J, Smith K, Wackerhage H, Greenhaff P. Branched-chain amino acids as fuels and anabolic signals in human muscle. J Nutr. 2006;136(1 Suppl):264S–8S.
- Power O, Hallihan A, Jakeman P. Human insulinotropic response to oral ingestion of native and hydrolysed whey protein. Amino Acids. 2009;37(2):333–9.
- Mitchell CJ, Churchward-Venne TA, Parise G, Bellamy L, Baker SK, Smith K, Atherton PJ, Phillips SM. Acute post-exercise myofibrillar protein synthesis is not correlated with resistance training-induced muscle hypertrophy in young men. PLoS One. 2014;9(2):e89431.
- Wilborn CD, Taylor LW, Outlaw J, Williams L, Campbell B, Foster CA, Smith-Ryan A, Urbina S, Hayward S. The effects of pre- and post-exercise whey vs. casein protein consumption on body composition and performance measures in collegiate female athletes. J Sports Sci Med. 2013;12(1):74–9.
- Fabre M, Hausswirth C, Tiollier E, Molle O, Louis J, Durguerian A, Neveux N, Bigard X. Effects of Postexercise protein intake on muscle mass and strength during resistance training: is there an optimal ratio between fast and slow proteins? Int J Sport Nutr Exerc Metab. 2017;27(5):448–57.
- Arnal MA, Mosoni L, Boirie Y, Houlier ML, Morin L, Verdier E, Ritz P, Antoine JM, Prugnaud J, Beaufrere B, Mirand PP. Protein pulse feeding improves protein retention in elderly women. Am J Clin Nutr. 1999;69(6):1202–8.
- Arnal MA, Mosoni L, Boirie Y, Houlier ML, Morin L, Verdier E, Ritz P, Antoine JM, Prugnaud J, Beaufrere B, Mirand PP. Protein feeding pattern does not affect protein retention in young women. J Nutr. 2000;130(7):1700–4.
- Seimon RV, Roekenes JA, Zibellini J, Zhu B, Gibson AA, Hills AP, Wood RE, King NA, Byrne NM, Sainsbury A. Do intermittent diets provide physiological benefits over continuous diets for weight loss? A systematic review of clinical trials. Mol Cell Endocrinol. 2015;418(Pt 2):153–72.
- Tinsley GM, Forsse JS, Butler NK, Paoli A, Bane AA, La Bounty PM, Morgan GB, Grandjean PW. Time-restricted feeding in young men performing resistance training: a randomized controlled trial. Eur J Sport Sci. 2017;17(2):200–7.
- Moro T, Tinsley G, Bianco A, Marcolin G, Pacelli QF, Battaglia G, Palma A, Gentil P, Neri M, Paoli A. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. J Transl Med. 2016;14(1):290.
- Arciero PJ, Ormsbee MJ, Gentile CL, Nindl BC, Brestoff JR, Ruby M. Increased protein intake and meal frequency reduces abdominal fat during energy balance and energy deficit. Obesity (Silver Spring). 2013;21(7):1357–66.
- Moore DR, Churchward-Venne TA, Witard O, Breen L, Burd NA, Tipton KD, Phillips SM. Protein ingestion to stimulate myofibrillar protein synthesis requires greater relative protein intakes in healthy older versus younger men. J Gerontol A Biol Sci Med Sci. 2015;70(1):57–62.
- Witard OC, Jackman SR, Breen L, Smith K, Selby A, Tipton KD. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am J Clin Nutr. 2014;99(1):86–95.
