
Breve introduzione all’argomento.

Era il 2014 quando lessi per la prima volta di questa metodica allenante denominata “Blood flow restriction training”(BFR training). Sebbene all’epoca suscitò in me un certo interesse, tale da spingermi a stendere qualche riga in un breve sunto che pubblicai sulla mia pagina Facebook, non dedicai molto tempo alla sperimentazione del metodo in questione, almeno fino a due anni fa. Merito delle evidenze scientifiche ed empiriche che mostravano come lo stress metabolico fosse un fattore di importanza non trascurabile nello stimolo ipertrofico, il rispolverare questa “bizzarra” tecnica di allenamento basata sull’acutizzazione dello stress metabolico muscolare fu per me naturalmente consequenziale.
Senza dilungarmi oltre, nel seguente articolo cercherò di esplicare in modo fruibile teoria e pratica di questa tecnica allenante, partendo dalle basi e terminando con le possibili applicazioni.
Stress Metabolico – il secondo tassello dello stimolo ipertrofico indotto da allenamenti contro resistenza –
E’ fuori da ogni ragionevole dubbio, ad oggi, che la Tensione Meccanica sia il fattore primario nella promozione della crescita muscolare indotta da allenamenti contro resistenza. In seconda posizione di importanza si attesta lo Sress Metabolico (1)(2)(3) il quale, in termini semplicistici, consiste in un accumulo di metaboliti indotto dall’esercizio, in modo particolare di lattato, fosfati inorganici e H+.(4)(5)

Lo Stress Metabolico è massimizzato durante le attività che si basano prevalentemente sulla glicolisi anaerobica per la produzione di energia. La glicolisi anaerobica è prevalente durante le attività che durano da circa 15 a 120 secondi e il corrispondente accumulo di metaboliti causa fatica indotta perifericamente (al contrario di quella indotta centralmente), vale a dire un affaticamento correlato a cambiamenti metabolici o biochimici, o entrambi, contrariamente ad una diminuzione dell’attività nervosa.(6) La ricerca indica che l’esecuzione di 1 set di 12 ripetizioni al cedimento (con un tempo sotto tensione totale di 37±3 sec) eleva i livelli di lattato muscolare a 91 mmol/Kg (peso secco) e i valori aumentano a 118mmol/Kg dopo 3 set.(7) Al contrario, nei protocolli che coinvolgono carichi molto pesanti (≥90% del 1RM) è stato osservato un accumulo di metaboliti minimo, a causa della breve durata dell’allenamento (generalmente <10 secondi per set) in cui si utilizza primariamente il sistema dei fosfati muscolari per fornire energia. Inoltre, durante gli allenamenti contro resistenza che si basano sulla glicolisi veloce l’ossigenazione del muscolo è compromessa. La persistente compressione del flusso circolatorio nel corso di un set a lunga durata causa un’ipossia acuta, aumentando così l’accumulo dei metaboliti.(8) La combinazione di questi fattori provoca il rapido accumulo di metaboliti intramuscolari insieme ad una concomitante diminuzione nei livelli di pH.(9)
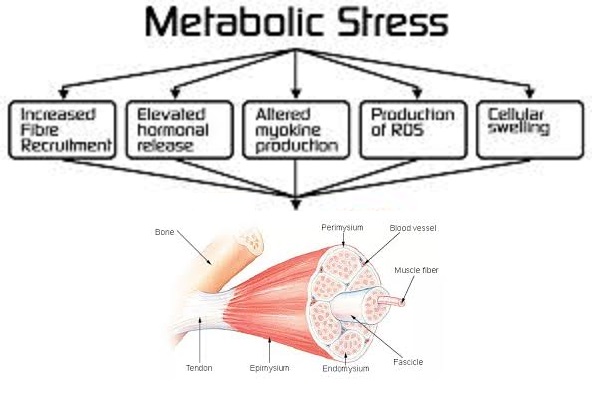
I classici allenamenti di Bodybuilding sono generalmente impostati in modo tale da sfruttare gli effetti ipertrofici dello stress metabolico a scapito di una più alta intensità di carico.(10, 11) Questi allenamenti, che comportano classicamente l’esecuzione di set multipli da 8 a 12 ripetizioni per set con intervalli di recupero fra un set e l’altro relativamente brevi (12), si sono mostrati in grado di aumentare lo stress metabolico a un livello maggiore dei regimi di allenamento ad alta intensità tipicamente impiegati dai Powerlifter.(13,14)E’ ben documentato che nonostante si allenino solitamente a intensità moderate di carico, i Bodybuilder mostrano fisici molto muscolosi e livelli di massa magra paragonabili, se non maggiori, a quelli raggiunti dai Powerlifter.(10,15) Infatti, vi sono prove che gli allenamenti tipici del Bodybuilding producono maggiori aumenti ipertrofici rispetto agli allenamenti a carico più elevato tipici degli allenamenti in stile Powerliftin (16,17,18), anche se i risultati non sono costanti in tutti gli esperimenti quando si equipara il volume di carico.(19, 20) Si pensa che una serie di fattori siano mediatori degli adattamenti ipertrofici derivati dallo stress metabolico indotto dall’esercizio, tra cui un aumentato reclutamento delle fibre, alterazioni nella sintesi di miochine, rigonfiamento cellulare, accumulo di specie reattive dell’ossigeno (ROS) e aumenti nella sintesi di ormoni sistemici.(21, 22, 23, 24)
Stress Metabolico e reclutamento delle fibre muscolari.
Un altro aspetto interessante dello stress metabolico è il suo impatto sul reclutamento delle fibre muscolari. Sebbene i carichi pesanti attivino fin dalle prime ripetizioni l’intero spettro di fibre, le ricerche mostrano che lo stress metabolico aumenta il reclutamento delle unità motorie a soglia più elevata anche con carichi leggeri. Gli studi mostrano che quando la fatica aumenta durante un esercizio sub-massimale prolungato, le soglie di reclutamento diminuiscono proporzionalmente.(25, 26, 27) Di conseguenza, l’attivazione delle fibre a contrazione rapida è alta, a condizione che il set sia portato al cedimento muscolare. Gli studi che hanno impiegato EMG (24-28), deplezione di glicogeno (29) e frazionamento dei fosfati organici (30, 4) hanno tutti dimostrato un aumentato reclutamento delle fibre a contrazione rapida nell’allenamento BFR, spingendo alcuni ricercatori a teorizzare che questo sia il fattore primario con cui l’occlusione del flusso ematico media l’anabolismo.(31, 32)
I meccanismi precisi attraverso i quali lo stress metabolico aumenta il reclutamento delle fibre muscolari non sono ancora del tutto chiari. E’ stato ipotizzato che l’accumulo di H+ svolga un ruolo determinante tramite l’inibizione della contrattilità delle fibre già attivate, favorendo così il reclutamento di ulteriori unità motorie di alta-soglia.(33, 34, 28)
MacDougal et al (7) hanno proposto che la fatica durante l’allenamento in un set singolo portato al cedimento sia dovuta a una combinazione di acidosi e deplezione di fosfocreatina, mentre l’acidosi sia la causa più probabile negli allenamenti contro resistenza a set multipli. Nonostatnte sembri che l’aumento del reclutamento delle fibre sia almeno parzialmente responsabile degli aumenti nell’ipertrofia associata a stress metabolico, sembra che anche altri fattori potrebbero avere un ruolo. Suga et al (4) hanno dimostrato che solo il 31% dei soggetti ha mostrato un reclutamento delle fibre a contrazione rapida durante un allenamento con occlusione al 20% del 1RM rispetto al 70% dei soggetti che hanno effettuato un allenamento non occlusivo al 65% del 1RM. Visto che l’allenamento BFR a questa intensità (20% del 1RM) ha dimostrato di aumentare la crescita muscolare in modo simile, o maggiore, all’allenamento contro resistenza ad alta intensità (35, 36), gli effetti anabolici non possono essere unicamente ricollegabili allo stesso reclutamento delle fibre. Questi risultati sono ulteriormente supportati dalle ricerche che mostrano ampiezze EMG significativamente più alte quando si effettua un allenamento tradizionale al 80% del 1RM rispetto all’allenamento occlusivo al 20% del 1RM, indicando una ridotta attivazione muscolare ad intensità più basse.(37) Anche in studi recenti nei quali è stato comparato l’allenamento a carico elevato contro quello a carico leggero è emerso una maggiore attivazione muscolare durante la sessione ad intensità più alta nonostante, durante la sessione a basso carico si sia verificato un accumulo di metaboliti molto più marcato.(38, 39, 40)
Stress Metabolico e sintesi di Miochine.
Lo stress metabolico può influenzare la crescita muscolare attraverso l’aumento delle miochine anaboliche, diminuendo le miochine cataboliche, o esercitando entrambe le azioni. Anche se questa affermazione ha una base logica, i risultati delle ricerche sull’argomento sono ambigue. Takarada et al (24) hanno mostrato un aumento graduale del IL-6 in seguito a vari set di estensione del ginocchio con BFR rispetto allo stesso volume di esercizio senza occlusione; i livelli sono rimasti elevati per 24 ore dopo la seduta allenante. Tuttavia, la dimensione dell’effetto è stata lieve, e la quantità assoluta dell’aumento è stata solo ¼ di quello riportato nell’allenamento eccentrico ad alto carico. Fujita et al (41) hanno osservato che sei giorni di allenamento occlusivo dell’estensore del ginocchio hanno aumentato l’area della sezione trasversale della coscia del 2,4% senza nessuna modifica nei livelli di IL-6.

Analogamente, altri studi hanno mostrato che i livelli di IL-6 sono rimasti invariati a seguito di protocolli di allenamento BFR noti per aumentare lo stress metabolico.(42, 43) La totalità di questi risultati sembra confutare il ruolo del IL-6 nell’ipertrofia indotta dallo stress metabolico. La correlazione tra stress metabolico e altri fattori di crescita locali non è stata studiata bene, e ciò impedisce di trarre una conclusione sulla loro reale importanza nell’induzione ipertrofica attraverso lo stress metabolico.
Le prove suggeriscono che lo stress metabolico può influenzare la crescita muscolare attraverso la riduzione di fattori catabolici locali. Kawada e Ishii (44) hanno segnalato una significativa diminuzione dei livelli di MSTN nel muscolo plantare di ratti Wistar in seguito ad allenamento BFR, se paragonati a un gruppo di controllo sottoposto a chirurgia fittizia. Al contrario, nessuna differenza nell’espressione genica MSTN è stata osservata negli esseri umani dopo 3 ore di esercizio a bassa intensità con e senza occlusione.(45) Un’altra prova sperimentale su esseri umani ha mostrato che sebbene l’allenamento BFR non abbia avuto effetto sul MSTN, esso ha diminuito diversi importanti fattori di trascrizione proteolitica (FOXO3A, atrogina-1 e MuRF-1) nelle 8 ore successive all’esercizio rispetto ad un gruppo di controllo sottoposto ad allenamento senza occlusione.(46) In uno studio su uomini fisicamente attivi, Laurentino et al (35) sono stati esaminati gli effetti dell’allenamento BFR sui livelli cronici di MSTN dopo 8 settimane di allenamento. I risultati hanno mostrato una significativa riduzione del 45% nell’espressine genica di MSTN nel caso di allenamenti BFR, rispetto ad una riduzione non significativa in caso di esercizio a bassa intensità senza occlusione. La natura conflittuale di questi risultati impedisce di stabilire se vi sia una correlazione significativa tra l’ipertrofia indotta dallo stress metabolico e la sintesi di miochine.
Stress Metabolico e osmosi cellulare.
Un altro meccanismo che si pensa possa mediare l’ipertrofia tramite lo stress metabolico è l’aumento dell’idratazione intracellulare (ovvero, rigonfiamento cellulare per osmosi). Si pensa che il rigonfiamento cellulare serva come regolatore fisiologico della funzione cellulare.(47, 48) Un vasto insieme di prove dimostra che un aumento dello stato di idratazione di una cellula allo stesso tempo aumenta la sintesi proteica e diminuisce il catabolismo. Questi risultati sono stati dimostrati in un ampia varietà di tipi cellulari inclusi osteociti, cellule del seno, epatociti e fibre muscolari.(49)
Le attuali teorie suggeriscono che un aumento dell’idratazione cellulare causa una pressione contro il citoscheletro e la membrana cellulare, e tale pressione viene percepita come una minaccia per l’integrità cellulare. In risposta la cellula aumenta l’attivazione a cascata di segnalazione anabolica che porta in fine al rafforzamento della sua struttura.(50, 51) La segnalazione sembra essere mediata attraverso osmocettori associati alle integrine presenti all’interno delle cellule.(52) Questi recettori attivano vie di trasduzione di proteine chinasi anaboliche, che pare siano mediate da fattori di crescita locali.(53, 54) Il PI3K sembra essere un importante componente del trasporto di amminoacidi nel muscolo a seguito di una maggiore idratazione cellulare.(52) Le ricerche suggeriscono che gli effetti anabolici siano anche indotti in modo indipendente dal mTOR (55), con prove di regolazione diretta da parte del MAPK.(56, 57). Inoltre, il rigonfiamento delle fibre muscolari può innescare la proliferazione delle cellule satellite e promuovere la loro fusione con le fibre interessate (58), fornendo un ulteriore impulso per la crescita.
Vi sono poche prove per quanto riguarda il gonfiore cellulare indotto da stress metabolico in seguito all’esercizio, e il suo eventuale ruolo nell’ipertrofia. Tuttavia, si può dare una spiegazione sensata a tale effetto. L’esercizio contro resistenza altera in modo acuto l’equilibrio indrico intra ed extra cellulare (59), e l’entità delle alterazioni dipende dal tipo di esercizio e dall’intensità dell’allenamento. Il rigonfiamento cellulare potrebbe essere aumentato da un tipo di allenamento contro resistenza che genera elevate quantità di acido lattico grazie alle proprietà osmolitiche del lattato (60, 61), anche se alcune ricerche hanno confutato questa ipotesi.(62) L’accumulo di lattato intramuscolare attiva meccanismi di regolazione del volume cellulare; gli effetti sembrano essere amplificati dall’aumento dell’acidosi associato.(49) Le fibre a contrazione rapida sono ritenute particolarmente sensibili ai cambiamenti osmotici, probabilmente perché contengono un’alta concentrazione di acquaporine-4 (AQP4), ovvero canali di trasporto dell’acqua.(60) Considerando che le fibre a contrazione rapida hanno dimostrato di avere il maggiore potenziale di crescita (63), un aumento del rigonfiamento in queste fibre potrebbe teoricamente migliorare il loro adattamento in modo significativo.
Stress Metabolico e sintesi di ormoni sistemici.
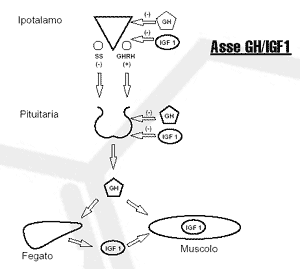
E’ stato teorizzato che gli aumenti acuti degli ormoni anabolici nel post-workout, derivati dall’accumulo di metaboliti durante allenamenti contro resistenza, possano aumentare la risposta ipertrofica. In particolare, lo stress metabolico indotto dall’esercizio è fortemente associato con un picco dei livelli di Ormone della Crescita in seguito all’allenamento.(64, 65, 66, 67, 68, 24) Anche se transitoria, la grandezza di questi aumenti è considerevole. Uno studio ha mostrato un aumento nei livelli di GH in seguito ad allenamento BFR di 10 volte maggiore rispetto a quello osservato con un allenamento non occlusivo di simile intensità (69); un’altra ricerca ha mostrato aumenti post-allenamento 290 volte maggiori rispetto al livello basale.(24) Si pensa che le elevazioni in seguito all’allenamento siano mediate da un maggiore accumulo di lattato o ioni H+.(64, 66) Soggetti deficitari di miofosforilasi, un enzima glicolitico responsabile della scomposizione del glicogeno e che quindi induce la produzione di lattato, mostrano una risposta attenuata del GH nel post-workout (70), fornendo forti prove per un collegamento tra produzione di lattato e rilasci di GH. Una diminuzione del pH indotta dai metaboliti può anche potenziare la sintesi e rilascio del GH tramite stimolazione del chemoriflesso regolata dai metaborecettori intramuscolari e le afferenze nervose di tipo III e IV.(71, 72)
Dal momento che il GH è in grado di potenziare la sintesi di IGF-1, sembra logico che un accumulo dei metaboliti sarebbe associato anche con un aumento post-workout dei livelli di IGF-1, noto peptide implicato nei processi anabolici del tessuto muscolare. Ciò è stato confermato in parte da studi che mostrano elevazioni significativamente maggiori di IGF-1 dopo il completamento di un allenamento mirato ad enfatizzare lo stress metabolico (73, 74, 75), anche se altre ricerche non sono riuscite a confermare tale associazione.(76) Inoltre, diversi studi (77, 69, 68), ma non tutti (45), hanno segnalato aumenti acuti dei livelli di IGF-1 post-workout a seguito di un allenamento BFR, e questo suggerisce che tali risultati siano stati mediati dallo stress metabolico. E’ importante notare che l’insieme delle ricerche riguarda l’isoforma sistemica di IGF-1, e i risultati non possono essere necessariamente estrapolati ai suoi effetti intramuscolari locali.
Per quello che vale in questo caso, l’effetto dello stress metabolico sugli aumenti in acuto di Testosterone rimane sconosciuto. Lu et al (78) hanno riferito che produzioni di lattato indotta dall’esercizio era correlata con l’aumento del Testosterone durante una sessione di nuoto ad alta intensità in ratti Sprague-Dawley. In una seconda parte dello studio, l’infusione diretta di lattato nei testicoli dei ratti è stata vista causare un aumento dose-dipendente dei livelli di Testosterone. D’altra parte, la ricerca controllata sugli esseri umani ha prodotto risultati disparati. Sebbene alcuni studi mostrino maggiore rilascio di Testosterone in seguito a protocolli di allenamento alto affaticamento metabolico rispetto a quelli che non causano accumulo significativo di metaboliti (79, 65, 66, 80, 81), altri non mostrano nessuna differenza significativa.(82, 83, 84) Inoltre, molti degli studi sull’allenamento BFR non sono riusciti a trovare aumenti acuti di Testosterone significativamente maggiori malgrado gli alti livelli di metaboliti (69, 83, 85), e questo mette in dubbio quanto l’ormone in questione sia influenzato dall’accumulo di metaboliti. Le incongruenze tra gli studi potrebbero riguardare fattori demografici come sesso, età ed anzianità di allenamento, e inoltre lo stato nutrizionale ha dimostrato di influenzare il rilascio di testosterone.(86) Comunque sia, l’ipotetico impatto dei picchi ormonali transitori in seguito all’allenamento sugli adattamenti ipertrofici rimane quanto meno discutibile. Se un tale ruolo è presente,il suo impatto sembrerebbe essere di poca importanza.
Tiriamo le somme…
Ci sono quindi forti evidenze che lo stress metabolico indotto dall’esercizio contribuisca alla risposta ipertrofica. Quello che resta da determinare è se tali effetti siano additivi allo stimolo derivato dalle forze meccaniche o se siano ridondanti dopo che una certa soglia di carico sia stata raggiunta. Un problema nel cercare di trarre inferenze dai disegni sperimentali di allenamento è che tensione meccanica e stress metabolico si presentano contemporaneamente, confondendo la capacità di distinguere gli effetti di uno da quelli dell’altro. Questo può portare a credere che i fattori metabolici siano la causa della crescita quando in realtà lo sono i fattori meccanici, o viceversa.
La capacità di derivare un rapporto di causa-effetto tra ipertrofia e stress metabolico è ulteriormente ridotta dal fatto che l’accumulo di metaboliti indotti dall’esercizio si verifica generalmente in tandem con il danno alle fibre muscolari. Data la credenza comune secondo cui l’esercizio che causa danno muscolare causi il maggiore stimolo anabolico (87), è difficile separare gli effetti di una variabile dall’altra per quanto riguarda gli adattamenti ipertrofici. Le ricerche che dimostrano che l’allenamento BFR aumenta la crescita muscolare senza danni significativi alle fibre suggeriscono che gli effetti ipertrofici di accumulo di metaboliti siano infatti separati da quelli del danno muscolare (88), anche se esistono prove contrastanti sull’argomento che impediscono di trarre una conclusione definitiva.(89)
Infine, e cosa più importante, i meccanismi responsabili di eventuali effetti anabolici dello stress metabolico non sono stati completamente chiariti. Sebbene l’aumentato reclutamento delle fibre muscolari sembra svolgere un ruolo importante, è improbabile che il reclutamento rappresenti il solo beneficio ipertrofico dell’accumulo di metaboliti. Piuttosto, le prove suggeriscono che interazione combinata di più fattori locali, e probabilmente anche sistemici, contribuisca alla crescita in maniera diretta o facilitate, o entrambe.(90) Il fatto che gli studi effettuati fino ad oggi riguardino principalmente soggetti non allenati lascia aperta la possibilità che i meccanismi possano differire in base all’anzianità di allenamento.
Le tecniche allenanti che possono enfatizzare lo stress metabolico vanno oltre il solo BFR (vedi, per esempio, flex between sets e peack of contrattion), ma questa, all’apparenza, “bizzarra” tecnica di allenamento ha mostrato di poter creare uno stress metabolico più accentuato. Per questo motivo e per via della sua scarsa diffusione nelle preparazioni dei Bodybuilder ho deciso di trattarlo in modo completo ed esauriente.
Il “Blood flow restriction training” (BFR)
Introduzione e studi correlati

Il “Blood flow restriction training” (abbreviato in “BFR training” e conosciuto anche con il nome di “Kaatsu Training”) o allenamento con occlusione è un approccio allenante che vede l’uso di lacci emostatici o apposite fasce posizionate all’altezza dell’inserzione articolare durante lo svolgimento di esercizi contro resistenza o aerobici.(91) Con questo metodo di allenamento, il flusso sanguigno verso e dagli arti viene limitato per l’appunto da una fascia durante il ciclo di contrazione e il periodo di riposo. Ciò comporta una parziale restrizione dell’afflusso arterioso ai muscoli, ma, in modo più significativo, limita il deflusso venoso dal muscolo.(92) Data la natura a carico leggero (generalmente pari a <40% del 1RM)e i benefici sulla forza derivanti dal BFR, esso può fornire un efficace stimolo riabilitativo clinico senza gli alti livelli di stress articolare associati all’allenamento a carico pesante.(93)
Diversi professionisti utilizzano una vasta gamma di protocolli BFR che non corrispondono a quanto riportato nella letteratura scientifica.(94) Gli attuali approcci che si concentrano sull’applicazione del BFR durante l’esercizio fisico consistono in sistemi di laccio emostatico pneumatico automatico o dispositivo gonfiabile portatile chiamato bracciale di occlusione. La ricerca che dimostra l’influenza della circonferenza della coscia e della larghezza della fascia (95) sulla pressione di occlusione ha accentuato la probabile necessità di un approccio individualizzato al BFR, in particolare per quanto riguarda l’impostazione della pressione della restrizione. Recentemente è in uso una tecnica per calcolare e prescrivere lo stimolo occlusivo in percentuale sulla pressione di occlusione totale negli arti, e rappresenta solo un esempio degli sforzi per tenere conto dei fattori di cui sopra e fornire un approccio individualizzato alla prescrizione dell’allenamento BFR relativamente rapido e sicuro. Mentre la relazione tra la pressione nel BFR e la compressione del tessuto sottostante durante l’esercizio non è ancora completamente compresa, l’allenamento BFR che utilizza il 40 -80% della pressione occlusiva negli arti è sicuro ed efficace se supervisionato da professionisti esperti (96);ciò nonostante, anche pressioni più basse possono fornire meno rischi senza la necessità di ricercare gradi pressori più elevati.
L’impatto dello stress metabolico sugli adattamenti ipertrofici è esemplificato negli studi sulla metodica BFR. La maggior parte della letteratura indica che l’allenamento BFR stimola la segnalazione anabolica e la sintesi proteica muscolare (43) e aumenta notevolmente la crescita muscolare (97) nonostante l’utilizzo di carichi spesso considerati troppo bassi per promuovere ipertrofia in modo significativo.(98, 99)
Si è ipotizzato che lo stress metabolico sia il fattore principale dell’ipertrofia indotta dal BFR. Sono stati notati forti accumuli di metaboliti durante questo tipo di allenamento (100), il che indica una associazione fra lo stress metabolico e la crescita muscolare osservata. A ulteriore sostegno di questa tesi, un aumento significativo nella sezione trasversale dei muscoli della coscia è stato notato in giovani soggetti maschi dopo 3 settimane di camminate con occlusione nelle gambe.(101)
Dato che giovani soggetti sani generalmente non subiscono aumenti della massa muscolare in seguito a sedute di allenamento aerobico a bassa intensità, lo studio fornisce una forte indicazione che esistono fattori diversi dalla tensione meccanica, responsabili degli adattamenti ipertrofici. Infatti, è stato osservato che gli aumenti nella sezione trasversale del muscolo sono significativamente correlati con i cambiamenti nei fosfati inorganici (r=.876) e nel pH intramuscolare (r=.601) durante l’allenamento BFR effettuato al 20% del 1RM. Ciò indica che lo stress metabolico generato durante l’allenamento contro resistenza è un regolatore chiave per la crescita muscolare.(102)
Gli studi che indagano gli effetti dell’allenamento contro resistenza in condizione di ipossia forniscono ulteriori prove della correlazione tra stress metabolico e crescita muscolare. Ko net al (103) hanno osservato che respirare solo il 13% di ossigeno durante un protocollo a set multipli e a basso carico ( 50% del 1RM), con intervalli di recupero piuttosto brevi (1 minuto) tra i set aumentava significativamente i livelli ematici di lattato rispetto allo stesso allenamento svolto in condizioni di normossia. Analogamente, Nishimura et al (104) hanno riferito aumenti significativamente maggiori nella sezione trasversale del flessore del gomito in seguito all’esecuzione di 4 set da 10 ripetizioni al 70% del 1RM effettuate in condizioni di ipossia acuta contro normossia. I meccanismi responsabili dell’aumento della risposta ipertrofica in seguito a sedute allenanti in ipossia devono ancora essere determinate, ma il maggiore accumulo di metaboliti è ritenuto svolgere un ruolo nel processo.(105)
Applicazioni pratiche
 Come già accennato, il BFR è una metodica allenante contro resistenza articolare applicabile anche ad esercizi cardio a bassa intensità.
Come già accennato, il BFR è una metodica allenante contro resistenza articolare applicabile anche ad esercizi cardio a bassa intensità.
Lo schema di ripetizione standard utilizzato nel BFR consiste nell’esecuzione di 30 ripetizioni seguite da una pausa di 30 secondi seguita a sua volta da altre 3 serie da 15 ripetizioni intervallate sempre da 30 secondi di pausa (30/15/15/15) con un carico del 50% o meno del 1RM generalmente nell’esercizio di chiusura della seduta allenante di un dato gruppo muscolare (vedi, ad esempio, bicipiti o tricipiti). Questa esecuzione ci dà un volume di 75 ripetizioni totali in BFR. Il primo set di 30 ripetizioni può essere visto come il carico di innesco per iniziare il ciclo Cori o il ciclo dell’acido lattico. Questo primo set è generalmente ben tollerato dal soggetto e spesso si percepisce come estremamente facile. Il laccio emostatico o fascia viene lasciato applicato durante il periodo di riposo, questo è molto importante per l’accumulo dei metaboliti nel muscolo target.
I seguenti 3 set e periodi di riposo sono molto più difficili da gestire a causa del successivo accumulo di lattato. L’RPE è strettamente correlato all’accumulo di lattato. Inoltre, i soggetti possono avvertire un leggero e ulteriore aumento della frequenza cardiaca durante l’esercizio. Ciò è normale a causa del ridotto ritorno venoso, del conseguente calo del volume ematico e dell’aumentata FC per mantenere la gittata cardiaca. Se in qualsiasi momento il soggetto dovesse avvertire vertigini o dolore da moderato a grave nella zona soggetta alla compressione con o senza intorpidimento o parestesia nell’arto deve essere immediatamente interrotta la sessione di allenamento.
Una volta che il soggetto ha terminato la sessione di allenamento, percepirà la riperfusione di sangue nell’arto e il “bruciore” dato dal lattato, i quali generalmente scompaiono in breve tempo. Spesso il soggetto si sente molto affaticati agli arti e studi sulla misurazione della produzione di forza immediatamente dopo il BFR anche a carichi bassi hanno dimostrato una forza significativamente ridotta. Ed è per questo motivo che questa metodica allenante viene diligentemente inserita in chiusura della seduta di allenamento e non prima dello svolgimento di esercizi con carichi elevati. Questi stessi esercizi non dovrebbero essere eseguiti in BFR. Tuttavia, ci saranno momenti in cui il soggetto non è in grado di raggiungere il volume target. Bisogna ricordare che il volume è la chiave per l’aumento della forza e l’ipertrofia nell’allenamento BFR.
Gli esercizi comunemente utilizzati con il BFR comprendono per la parte superiore del corpo: Curl (con bilanciere, manubri, macchine o cavi), French Press, Push Down ecc…
Gli esercizi comunemente utilizzati con il BFR comprendono per la parte inferiore del corpo: Leg-Extesion, Leg-Curl, Calf, Squat, Stacchi, camminata e Cyclette.
Di seguito sono riportate le linee guida da seguire per quanto riguarda la progressione dell’esercizio in BFR e le difficoltà con il raggiungimento del volume: Carico: 20-30% 1RM (determinato, stimato). Se il soggetto raggiunge:
- 75 ripetizioni = Continua con l’allenamento, rivaluta l’1RM entro 1-3 sessioni. Ristabilisci la nuova gamma del 20-30% man mano che la forza migliora.
- 60-74 ripetizioni = Continua con l’allenamento, ma prolunga il periodo di riposo tra le serie 3 e 4 a 45 secondi. Fino al completamento di 75 ripetizioni.
- 45-59 Ripetizioni = Continua con l’allenamento, ma estendi il periodo di riposo tra tutte le serie a 45-60 secondi.
- <44 Ripetizioni = Ridurre il carico di circa il 10% fino a raggiungere 75 ripetizioni.
- Se costretto a fermarsi prima di 75 ripetizioni a causa di dolore, indolenzimento o sensazione di disagio generale nell’area di posizionamento del laccio emostatico = Ridurre la pressione del laccio emostatico del 10% ad ogni sessione di allenamento fino a raggiungere la tolleranza pressoria. Aumentare la pressione della cuffia del 10% per raggiungere la pressione di occlusione dell’arto fino alla massima tollerabilità soggettiva.
Nota: diversi atleti dividono gli allenamenti della settimana in due set: nel primo set svolgono allenamenti tipici del Powerlifting o ibridati (vedi Powerbuilding), nel secondo set si dedicano ad allenamenti prettamente metabolici con l’inserimento in chiusura della seduta di un esercizio in BFR.
Possibili effetti collaterali e controindicazioni
Gli effetti collaterali che possono essere riportati durante l’esecuzione di esercizi in BFR sono svenimento e vertigini, intorpidimento, dolore e disagio, indolenzimento muscolare ad insorgenza ritardata.(106)
Tutti i soggetti devono essere valutati per i rischi e le controindicazioni all’uso del laccio emostatico prima dell’applicazione del BFR. I soggetti a probabile rischio di reazioni avverse sono quelli con scarsi sistemi circolatori, obesità, diabete, calcificazione arteriosa, tratto falciforme, ipertensione grave o compromissione renale.(107) Potenziali controindicazioni da considerare sono tromboembolia venosa, compromissione vascolare periferica, anemia falciforme, infezione delle estremità, linfoadenectomia, cancro o tumore, estremità con accesso alla dialisi, acidosi, frattura aperta, aumento degli innesti vascolari a pressione intracranica o farmaci noti per aumentare il rischio di coagulazione.(108)
Quando si usano lacci o fasce emostatiche, la quantità di pressione sul sistema vascolare non può essere controllata e ci sono state segnalazioni di casi di rabdomiolisi dovuti al VOT.(109) Un sostenitore del metodo, il dottor Yoshiaki Sato, ha rischiato la vita nel 1967 per via della mancata regolazione del VOT, portandolo a concentrarsi maggiormente sul controllo e la regolazione della pressione applicata sulla vascolarizzazione.
Conclusione
Il BFR training è senza dubbio una metodica allenante interessante da poter inserire nella propria preparazione, in specie sui gruppi muscolari carenti e a conclusione di una seduta ad essi dedicata, o semplicemente come “richiamo”. L’atleta o il preparatore che desidera applicare in modo corretto il BFR deve essere in grado di gestirlo in maniera ottimale. A tal fine il presente articolo può offrire le basi necessarie affinché ciò possa avvenire.
Gabriel Bellizzi
Riferimenti:
- “Scienza e sviluppo dell’ipertrofia muscolare” di Brad Schoenfeld (pag. 36-41).
- Rooney, KJ, Herbert, RD, and Balnave, RJ. Fatigue contributes to the strength trining stimulus. Med Sci Sports Exerc. 26: 1160-1164, 1994.
- Schott, J, McCully, K, and Rutherford, OM. The role of metabolites in strength trining. II. Short versus long isometric contractions. Eur J Appl Physiol Occup Physiol. 71: 337-341, 1995.
- Smith, RC, and Rutherford. OM. The role of metabolites in strength trining. I. A comparison of eccentric and concentric contractions. Eur J Appl Physiol Occup Physiol. 71: 332-336, 1995.
- Suga T, Okita K, Morita N, Yokota T, Hirabayashi K, Horiuchi M, Takada S, Takahashi T, Omokawa M, Kinugawa S, Tsutsui H. Intramuscular metabolism during low-intensity resistance exercise with blood flow restriction. J Appl Physiol (1985). 2009 Apr;106(4):1119-24.
- Tesch PA, Colliander EB, Kaiser P. Muscle metabolism during intense, heavy-resistance exercise. Eur J Appl Physiol Occup Physiol. 1986;55(4):362-6.
- Robbins, DW, Goodale, TL, Docherty, D, Behm, DG, and Tran, QT. The effects of load and training pattern on acute neuromuscular responses in the upper body. J Strength Cond Res. 2010 Nov;24(11):2996-3007.
- MacDougall JD, Ray S, Sale DG, McCartney N, Lee P, Garner S. Muscle substrate utilization and lactate production. Can J Appl Physiol. 1999 Jun;24(3):209-15.
- Tamaki T, Uchiyama S, Tamura T, Nakano S. Changes in muscle oxygenation during weight-lifting exercise. Eur J Appl Physiol Occup Physiol. 1994;68(6):465-9.
- Street SF. Lateral transmission of tension in frog myofibers: a myofibrillar network and transverse cytoskeletal connections are possible transmitters.
J Cell Physiol. 1983 Mar;114(3):346-64.
- Fry AC. The role of resistance exercise intensity on muscle fibre adaptations. Sports Med. 2004;34(10):663-79.
- Schoenfeld BJ. The mechanisms of muscle hypertrophy and their application to resistance training. J Strength Cond Res. 2010 Oct;24(10):2857-72.
- Lambert CP, Flynn MG. Fatigue during high-intensity intermittent exercise: application to bodybuilding. Sports Med. 2002;32(8):511-22.
- Kraemer WJ, Marchitelli L, Gordon SE, Harman E, Dziados JE, Mello R, Frykman P, McCurry D, Fleck SJ. Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol (1985). 1990 Oct;69(4):1442-50.
- Kraemer WJ, Fleck SJ, Dziados JE, Harman EA, Marchitelli LJ, Gordon SE, Mello R, Frykman PN, Koziris LP, Triplett NT. Changes in hormonal concentrations after different heavy-resistance exercise protocols in women. J Appl Physiol (1985). 1993 Aug;75(2):594-604.
- Katch VL, Katch FI, Moffatt R, Gittleson M. Muscular development and lean body weight in body builders and weight lifters. Med Sci Sports Exerc. 1980;12(5):340-4.
- JOYEON CHOI, HIDEYUKI TAKAHASHI, YUJI ITAI, KAORU TAKAMATSU. THE DIFFERENCE BETWEEN EFFECTS OF “POWER-UP TYPE” AND “BULK-UP TYPE” STRENGTH TRAINING EXERCISES. Japanese Journal of Physical. Volume 47 (1998) Issue 1.
- Masuda K, Choi JY, Shimojo H, Katsuta S. Maintenance of myoglobin concentration in human skeletal muscle after heavy resistance training. Eur J Appl Physiol Occup Physiol. 1999 Mar;79(4):347-52.
- Schmidtbleicher et al.Neuronal adaptation and increase of cross-sectional area studying different strength training methods. In: Jonsson GB, ed. Biomechanics (vol. X-B, pp. 615-620). Champaign, IL: Human Kinetics, 1987.
- Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE, Ratamess NA, Kraemer WJ, Staron RS. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. Eur J Appl Physiol. 2002 Nov;88(1-2):50-60. Epub 2002 Aug 15.
- Schoenfeld BJ, Ratamess NA, Peterson MD, Contreras B, Sonmez GT, Alvar BA. Effects of different volume-equated resistance training loading strategies on muscular adaptations in well-trained men. J Strength Cond Res. 2014 Oct;28(10):2909-18.
- Gordon SE, Kraemer WJ, Vos NH, Lynch JM, Knuttgen HG. Effect of acid-base balance on the growth hormone response to acute high-intensity cycle exercise. J Appl Physiol (1985). 1994 Feb;76(2):821-9.
- Goto K, Ishii N, Kizuka T, Takamatsu K. The impact of metabolic stress on hormonal responses and muscular adaptations. Med Sci Sports Exerc. 2005 Jun;37(6):955-63.
- Nishimura A, Sugita M, Kato K, Fukuda A, Sudo A, Uchida A. Hypoxia increases muscle hypertrophy induced by resistance training.
Int J Sports Physiol Perform. 2010 Dec;5(4):497-508.
- Takarada Y, Nakamura Y, Aruga S, Onda T, Miyazaki S, Ishii N. Rapid increase in plasma growth hormone after low-intensity resistance exercise with vascular occlusion. J Appl Physiol (1985). 2000 Jan;88(1):61-5.
- Houtman CJ, Stegeman DF, Van Dijk JP, Zwarts MJ. Changes in muscle fiber conduction velocity indicate recruitment of distinct motor unit populations. J Appl Physiol (1985). 2003 Sep;95(3):1045-54. Epub 2003 May 23.
- Sahlin K, Söderlund K, Tonkonogi M, Hirakoba K. Phosphocreatine content in single fibers of human muscle after sustained submaximal exercise.
Am J Physiol. 1997 Jul;273(1 Pt 1):C172-8.
- Vøllestad NK, Vaage O, Hermansen L. Muscle glycogen depletion patterns in type I and subgroups of type II fibres during prolonged severe exercise in man. Acta Physiol Scand. 1984 Dec;122(4):433-41.
- Takarada Y, Takazawa H, Sato Y, Takebayashi S, Tanaka Y, Ishii N. Effects of resistance exercise combined with moderate vascular occlusion on muscular function in humans. J Appl Physiol (1985). 2000 Jun;88(6):2097-106.
- Ingemann-Hansen T, Halkjaer-Kristensen J, Halskov O. Skeletal muscle phosphagen and lactate concentrations in ischaemic dynamic exercise. Eur J Appl Physiol Occup Physiol. 1981;46(3):261-70.
- Suga T, Okita K, Morita N, Yokota T, Hirabayashi K, Horiuchi M, Takada S, Omokawa M, Kinugawa S, Tsutsui H. Dose effect on intramuscular metabolic stress during low-intensity resistance exercise with blood flow restriction. J Appl Physiol (1985). 2010 Jun;108(6):1563-7. doi: 10.1152/japplphysiol.00504.2009. Epub 2010 Apr 1.
- Loenneke JP, Fahs CA, Wilson JM, Bemben MG. Blood flow restriction: the metabolite/volume threshold theory. Med Hypotheses. 2011 Nov;77(5):748-52. doi: 10.1016/j.mehy.2011.07.029. Epub 2011 Aug 12.
- Meyer RA. Does blood flow restriction enhance hypertrophic signaling in skeletal muscle? J Appl Physiol (1985). 2006 May;100(5):1443-4.
- Debold, EP. Recent insights into the molecular basis of muscular fatigue.
Med Sci Sports Exerc. 44(8): 1440-1452, 2012.
- Miller, KJ, Garland, SJ, Ivanova, T, and Ohtsuki, T. Motor-unit behavior in humans during fatiguing arm movements. J Neurophysiol. 75: 1629-1636, 1996.
- Laurentino, GC, Ugrinowitsch, C, Roschel, H, Aoki, MS, Soares, AG, Neves, M, Jr, Aihara, AY , Fernandes Ada, R, and Tricoli, V . Strength training with blood flow restriction diminishes myostatin gene expression. Med Sci Sports Exerc. 44: 406-412, 2012.
- Yasuda, T, Abe, T, Sato, Y , Midorikawa, T, Kearns, CF, Inoue, K, Ryushi, T, and Ishii, N. Muscle fiber cross-sectional area is increased after two weeks of twice daily KAATSU-resistance training. Int J KAATSU Train Res. 1: 65-70, 2005.
- Manini, TM, and Clark, BC. Blood flow restricted exercise and skeletal muscle health. Exerc Sport Sci Rev. 37: 78-85, 2009.
- Akima, H, and Saito, A. Activation of quadriceps femoris including vastus intermedius during fatiguing dynamic knee extensions. Eur J Appl Physiol. 113: 2829-2840, 2013.
- Cook, SB, Murphy, BG, and Labarbera, KE. Neuromuscular function after a bout of low-load blood flow-restricted exercise. Med Sci Sports Exerc. 45: 67-74, 2013.
- Schoenfeld, BJ, Contreras, B, Willardson, JM, Fontana, F, and TiryakiSonmez, G. Muscle activation during low- versus high-load resistance training in well-trained men. Eur J Appl Physiol. 114: 2491-2497, 2014.
- Fujita, T, Brechue, WF, Kurita, K, Sato, Y , and Abe, T. Increased muscle volume and strength following six days of low-intensity resistance training with restricted muscle blood flow. Int J KAATSU Training Res. 4: 1-8, 2008.
- Abe , T, Beekley, MD, Hinata, S, Koizumi, K, and Sato, Y . Day-to-day change in muscle strength and MRI-measured skeletal muscle size during 7 days KAATSU resistance training: A case study. Int J Kaatsu Training Res. 1: 71-76, 2005.
- Fry, CS, Glynn, EL, Drummond, MJ, Timmerman, KL, Fujita, S, Abe, T, Dhanani, S, Volpi, E, and Rasmussen, BB. Blood flow restriction exercise stimulates mTORC1 signaling and muscle protein synthesis in older men. J Appl Physiol. 108: 1199-1209, 2010.
- Kawada, S, and Ishii, N. Skeletal muscle hypertrophy after chronic restriction of venous blood flow in rats. Med Sci Sports Exerc. 37: 11441150, 2005.
- Drummond, MJ, Fujita, S, Abe, T, Dreyer, HC, Volpi, E, and Rasmussen, BB. Human muscle gene expression following resistance exercise and blood flow restriction. Med Sci Sports Exerc. 40: 691-698, 2008.
- Manini, TM, Vincent, KR, Leeuwenburgh, CL, Lees, HA, Kavazis, AN, Borst, SE, and Clark, BC. Myogenic and proteolytic mRNA expression following blood flow restricted exercise. Acta Physiol. (Oxf.) 201: 255263, 2011.
- Haussinger, D, Lang, F, and Gerok, W . Regulation of cell function by the cellular hydration state. Am J Physiol. 267: E343-E355, 1994.
- Haussinger, D. The role of cellular hydration in the regulation of cell function. Biochem J. 313 (Pt 3): 697-710, 1996.
- Lang, F, Busch, GL, Ritter, M, Volkl, H, Waldegger, S, Gulbins, E, and Haussinger, D. Functional significance of cell volume regulatory mechanisms. Physiol Rev. 78: 247-306, 1998.
- Lang, F. Mechanisms and significance of cell volume regulation. J Am Coll Nutr. 26: 613S-623S, 2007.
- Schoenfeld, BJ. The mechanisms of muscle hypertrophy and their application to resistance training. J Strength Cond Res. 24: 2857-2872, 2010.
- Low, SY , Rennie, MJ, and Taylor, PM. Signaling elements involved in amino acid transport responses to altered muscle cell volume. FASEB J. 11: 1111-1117, 1997.
- Clarke, MS, and Feeback, DL. Mechanical load induces sarcoplasmic wounding and FGF release in differentiated human skeletal muscle cultures. FASEB J. 10: 502-509, 1996.
- Lambert, IH, Hoffmann, EK, and Pedersen, SF. Cell volume regulation: Physiology and pathophysiology. Acta Physiol. (Oxf.) 194: 255-282,
2008. - Schliess, F, Richter, L, vom Dahl, S, and Haussinger, D. Cell hydration and mTOR-dependent signalling. Acta Physiol. (Oxf.) 187: 223-229, 2006.
- Finkenzeller, G, Newsome, W , Lang, F, and Haussinger, D. Increase of cjun mRNA upon hypo-osmotic cell swelling of rat hepatoma cells. FEBS Lett. 340: 163-166, 1994.
- Schoenfeld, B, Kolber, MJ, and Haimes, JE. The upright row: Implications for preventing subacromial impingement. Strength Cond J. 33: 25-28, 2011.
- Dangott, B, Schultz, E, and Mozdziak, PE. Dietary creatine monohydrate supplementation increases satellite cell mitotic activity during compensatory hypertrophy. Int J Sports Med. 21: 13-16, 2000.
- Sjogaard, G. Water and electrolyte fluxes during exercise and their relation to muscle fatigue. Acta Physiol Scand Suppl. 556: 129-136, 1986.
- Frigeri, A, Nicchia, GP, Verbavatz, JM, Valenti, G, and Svelto, M. Expression of aquaporin-4 in fast-twitch fibers of mammalian skeletal muscle. J Clin Invest. 102: 695-703, 1998.
- Sjogaard, G, Adams, RP, and Saltin, B. Water and ion shifts in skeletal muscle of humans with intense dynamic knee extension. Am J Physiol. 248: R190-6, 1985.
- Usher-Smith, JA, Fraser, JA, Bailey, PS, Griffin, JL, and Huang, CL. The influence of intracellular lactate and H+ on cell volume in amphibian skeletal muscle. J Physiol. 573: 799-818, 2006.
- Kosek, DJ, Kim, JS, Petrella, JK, Cross, JM, and Bamman, MM. Efficacy of 3 days/wk resistance training on myofiber hypertrophy and myogenic mechanisms in young vs. older adults. J Appl Physiol. 101: 531-544, 2006.
- Gordon, SE, Kraemer, WJ, Vos, NH, Lynch, JM, and Knuttgen, HG. Effect of acid-base balance on the growth hormone response to acute highintensity cycle exercise. J Appl Physiol. 76: 821-829, 1994.
- Gotshalk, LA, Loebel, CC, Nindl, BC, Putukian, M, Sebastianelli, WJ, Newton, RU, Hakkinen, K, and Kraemer, WJ. Hormonal responses of multiset versus single-set heavy-resistance exercise protocols. Can J Appl Physiol. 22: 244-255, 1997.
- Hakkinen, K, and Pakarinen, A. Acute hormonal responses to two different fatiguing heavy-resistance protocols in male athletes. J Appl Physiol. 74: 882-887, 1993.
- Pierce, JR, Clark, BC, Ploutz-Snyder, LL, and Kanaley, JA. Growth
hormone and muscle function responses to skeletal muscle ischemia. J Appl Physiol. 101: 1588-1595, 2006. - Takano, H, Morita, T, Iida, H, Asada, K, Kato, M, Uno, K, Hirose, K, Matsumoto, A, Takenaka, K, Hirata, Y , Eto, F, Nagai, R, Sato, Y , and Nakajima, T. Hemodynamic and hormonal responses to a short-term lowintensity resistance exercise with the reduction of muscle blood flow. Eur J Appl Physiol. 95: 65-73, 2005.
- Fujita, S, Abe, T, Drummond, MJ, Cadenas, JG, Dreyer, HC, Sato, Y , Volpi, E, and Rasmussen, BB. Blood flow restriction during low-intensity resistance exercise increases S6K1 phosphorylation and muscle protein synthesis. J Appl Physiol. 103: 903-910, 2007.
- Godfrey, RJ, Whyte, GP, Buckley, J, and Quinlivan, R. The role of lactate in the exercise-induced human growth hormone response: Evidence from McArdle disease. Br J Sports Med. 43: 521-525, 2009.
- Loenneke, JP, Wilson, GJ, and Wilson, JM. A mechanistic approach to blood flow occlusion. Int J Sports Med. 31: 1-4, 2010.
- Viru, M, Jansson, E, Viru, A, and Sundberg, CJ. Effect of restricted blood flow on exercise-induced hormone changes in healthy men. Eur J Appl Physiol Occup Physiol. 77: 517-522, 1998.
- Kraemer, WJ, Marchitelli, L, Gordon, SE, Harman, E, Dziados, JE, Mello, R, Frykman, P, McCurry, D, and Fleck, SJ. Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol. 69: 1442-1450, 1990.
- Kraemer, WJ, Gordon, SE, Fleck, SJ, Marchitelli, LJ, Mello, R, Dziados, JE, Friedl, K, Harman, E, Maresh, C, and Fry, AC. Endogenous anabolic hormonal and growth factor responses to heavy resistance exercise in males and females. Int J Sports Med. 12: 228-235, 1991.
- Rubin, MR, Kraemer, WJ, Maresh, CM, Volek, JS, Ratamess, NA, Vanheest, JL, Silvestre, R, French, DN, Sharman, MJ, Judelson, DA, Gomez, AL, Vescovi, JD, and Hymer, WC. High-affinity growth hormone binding protein and acute heavy resistance exercise. Med Sci Sports Exerc. 37: 395-403, 2005.
- Kraemer, WJ, Aguilera, BA, Terada, M, Newton, RU, Lynch, JM, Rosendaal, G, McBride, JM, Gordon, SE, and Hakkinen, K. Responses of IGF-I to endogenous increases in growth hormone after heavy-resistance exercise. J Appl Physiol. 79: 1310-1315, 1995.
- Abe, T, Yasuda, T, Midorikawa, T, Sato, Y , Kearns, C, Inoue, K, Koizumi, K, and Ishii, N. Skeletal muscle size and circulating IGF-1 are increased after two weeks of twice daily KAATSU resistance training. Int J Kaatsu Training Res. 1: 6-12, 2005.
- Lu, SS, Lau, CP, Tung, YF, Huang, SW , Chen, YH, Shih, HC, Tsai, SC, Lu, CC, Wang, SW , Chen, JJ, Chien, EJ, Chien, CH, and Wang, PS. Lactate and the effects of exercise on testosterone secretion: Evidence for the involvement of a cAMP-mediated mechanism. Med Sci Sports Exerc. 29: 1048-1054, 1997.
- Buresh, R, Berg, K, and French, J. The effect of resistive exercise rest interval on hormonal response, strength, and hypertrophy with training. J Strength Cond Res. 23: 62-71, 2009.
- McCaulley, GO, McBride, JM, Cormie, P, Hudson, MB, Nuzzo, JL, Quindry, JC, and Travis Triplett, N. Acute hormonal and neuromuscular responses to hypertrophy, strength and power type resistance exercise. Eur J Appl Physiol. 105: 695-704, 2009.
- Smilios, I, Pilianidis, T, Karamouzis, M, and Tokmakidis, SP. Hormonal
responses after various resistance exercise protocols. Med Sci Sports Exerc. 35: 644-654, 2003. - Kraemer, WJ, Marchitelli, L, Gordon, SE, Harman, E, Dziados, JE, Mello, R, Frykman, P, McCurry, D, and Fleck, SJ. Hormonal and growth factor responses to heavy resistance exercise protocols. J Appl Physiol. 69: 1442-1450, 1990.
- Reeves, GV , Kraemer, RR, Hollander, DB, Clavier, J, Thomas, C, Francois, M, and Castracane, VD. Comparison of hormone responses following light resistance exercise with partial vascular occlusion and moderately difficult resistance exercise without occlusion. J Appl Physiol. 101: 1616-1622, 2006.
- Suga, T, Okita, K, Morita, N, Yokota, T, Hirabayashi, K, Horiuchi, M, Takada, S, Omokawa, M, Kinugawa, S, and Tsutsui, H. Dose effect on intramuscular metabolic stress during low-intensity resistance exercise with blood flow restriction. J Appl Physiol. 108: 1563-1567, 2010.
- Viru, M, Jansson, E, Viru, A, and Sundberg, CJ. Effect of restricted blood flow on exercise-induced hormone changes in healthy men. Eur J Appl Physiol Occup Physiol. 77: 517-522, 1998.
- Kraemer, WJ, and Ratamess, NA. Hormonal responses and adaptations to resistance exercise and training. Sports Med. 35: 339-361, 2005.
- Schoenfeld, BJ. Does exercise-induced muscle damage play a role in skeletal muscle hypertrophy? J Strength Cond Res. 26: 1441-1453, 2012.
- Loenneke, JP, Thiebaud, RS, and Abe, T. Does blood flow restriction result in skeletal muscle damage? A critical review of available evidence. Scand J Med Sci Sports 24: e415-e422, 2014.
- Wernbom, M, Paulsen, G, Nilsen, TS, Hisdal, J, and Raastad, T. Contractile function and sarcolemmal permeability after acute low-load resistance exercise with blood flow restriction. Eur J Appl Physiol. 112: 2051-2063, 2012.
- Widegren, U, Ryder, JW , and Zierath, JR. Mitogen-activated protein kinase signal transduction in skeletal muscle: Effects of exercise and muscle contraction. Acta Physiol Scand. 172: 227-238, 2001.
- Scott, Brendan R.; Loenneke, Jeremy P.; Slattery, Katie M.; Dascombe, Ben J. (March 2015). “Exercise with blood flow restriction: an updated evidence-based approach for enhanced muscular development”. Sports Medicine (Auckland, N.Z.). 45 (3): 313–325.
- Patterson, Stephen D.; Hughes, Luke; Head, Paul; Warmington, Stuart; Brandner, Christopher (2017-06-22). “Blood flow restriction training: a novel approach to augment clinical rehabilitation: how to do it”. Br J Sports Med. 51 (23): bjsports–2017–097738.
- Hughes, Luke; Paton, Bruce; Rosenblatt, Ben; Gissane, Conor; Patterson, Stephen David (2017-07-01). “Blood flow restriction training in clinical musculoskeletal rehabilitation: a systematic review and meta-analysis”. Br J Sports Med. 51 (13): 1003–1011.
- Patterson, Stephen D.; Brandner, Christopher R. (February 2017). “The role of blood flow restriction training for applied practitioners: A questionnaire-based survey” (PDF). Journal of Sports Sciences. 36 (2): 123–130.
- Loenneke, Jeremy P.; Fahs, Christopher A.; Rossow, Lindy M.; Sherk, Vanessa D.; Thiebaud, Robert S.; Abe, Takashi; Bemben, Debra A.; Bemben, Michael G. (2011-12-06). “Effects of cuff width on arterial occlusion: implications for blood flow restricted exercise”. European Journal of Applied Physiology. 112 (8): 2903–2912.
- Soligon, SD; Lixandrão, ME; Biazon, TMPC; Angleri, V; Roschel, H; Libardi, CA (September 2018). “Lower occlusion pressure during resistance exercise with blood-flow restriction promotes lower pain and perception of exercise compared to higher occlusion pressure when the total training volume is equalized”. Physiology International. 105 (3): 276–284.
- Loenneke, JP, Wilson, JM, Marin, PJ, Zourdos, MC, and Bemben, MG. Low intensity blood flow restriction training: A meta-analysis. Eur J Appl Physiol. 112(5): 1849-1859, 2011.
- Campos, GER, Luecke, TJ, Wendeln, HK, Toma, K, Hagerman, FC, Murray, TF, Ragg, KE, Ratamess, NA, Kraemer, WJ, and Staron, RS.
Muscular adaptations in response to three different resistance-training regimens: Specificity of repetition maximum training zones. Eur J Appl Physiol. 88: 50-60, 2002. - Kraemer, WJ, Adams, K, Cafarelli, E, Dudley, GA, Dooly, C, Feigenbaum, MS, Fleck, SJ, Franklin, B, Fry, AC, Hoffman, JR, Newton,
RU, Potteiger, J, Stone, MH, Ratamess, NA, and Triplett-McBride, T. American College of Sports Medicine position stand. Progression models in resistance training for healthy adults. Med Sci Sports Exerc. 34: 364380, 2002. - Loenneke, JP, Wilson, GJ, and Wilson, JM. A mechanistic approach to blood flow occlusion. Int J Sports Med. 31: 1-4, 2010.
- Abe, T, Kearns, CF, and Sato, Y . Muscle size and strength are increased following walk training with restricted venous blood flow from the leg muscle, Kaatsu-walk training. J Appl Physiol. 100: 1460-1466, 2006.
- Takada, S, Okita, K, Suga, T, Omokawa, M, Kadoguchi, T, Sato, T, Takahashi, M, Yokota, T, Hirabayashi, K, Morita, N, Horiuchi, M, Kinugawa, S, and Tsutsui, H. Low-intensity exercise can increase muscle mass and strength proportionally to enhanced metabolic stress under ischemic conditions. J Appl Physiol. 1985(113): 199-205, 2012.
- Kon, M, Ikeda, T, Homma, T, and Suzuki, Y . Effects of low-intensity resistance exercise under acute systemic hypoxia on hormonal responses. J Strength Cond Res. 26: 611-617, 2012.
- Nishimura, A, Sugita, M, Kato, K, Fukuda, A, Sudo, A, and Uchida, A. Hypoxia increases muscle hypertrophy induced by resistance training. Int J Sports Physiol. Perform. 5: 497-508, 2010.
- Schliess, F, Richter, L, vom Dahl, S, and Haussinger, D. Cell hydration and mTOR-dependent signalling. Acta Physiol. (Oxf.) 187: 223-229, 2006.
- Brandner, Christopher & May, Anthony & Clarkson, Matthew & Warmington, Stuart. (2018). Reported Side-effects and Safety Considerations for the Use of Blood Flow Restriction During Exercise in Practice and Research. Techniques in Orthopaedics. 33. 1. 10.1097/BTO.0000000000000259.
- Jump up ↑ DePhillipo NN, Kennedy MI, Aman ZS, Bernhardson AS, O’Brien L, LaPrade RF. Blood Flow Restriction Therapy After Knee Surgery: Indications, Safety Considerations, and Postoperative Protocol. Arthroscopy techniques. 2018 Oct 1;7(10):e1037-43.
- Johnny Owens. Owens Recovery Science. Blood Flow Restriction Rehabilitation Accessed from http://www.owensrecoveryscience.com
- Spranger MD, Krishnan AC, Levy PD, O’Leary DS, Smith SA. Blood flow restriction training and the exercise pressor reflex: a call for concern. American Journal of Physiology-Heart and Circulatory Physiology. 2015 Sep 4;309(9):H1440-52.
